Embed Size (px)
Citation preview

EXPERIMENTAL NEUROLOGY 62, ZOO-213 (1978)
Laryngeal Reflex Regulation: Peripheral and Central Neural Analyses
G. E. LUCIER, J. DAYNES, AND B. J. SESSLE
Divisio+t of Biological Sciences, Far&y of Dentistry, University of Torofzto, Tororzto M5G lG6, Canada
Received February 13, 1978; rezisio,t received JlLne 9, 1978
Because of the many functions (e.g., glottic closure, gagging, coughing, swallowing, speech) in which the internal laryngeal muscles are active, complex regulatory mechanisms must exist to modify and coordinate these activities. Little information is available, however, on these neural mech- anisms, and the present study attempted to provide more information by investigating, in the adult cat, the effects of stimuli to various orofacial and upper respiratory tract sites and nerves on 110 single units recorded in individual laryngeal muscles, in the recurrent laryngeal nerve (RLN), and in the brain stem nucleus ambiguus. A particularly powerful excitatory influ- ence on unitary activity related to laryngeal adduction was produced by laryngeal stimuli; in many instances this influence could not be modified by other sensory inputs (e.g., from vagus or glossopharyngeal nerves). Laryn- geal stimulation also produced a profound cessation of respiration and suppression of the inspiratory-related activity of laryngeal abductor units. Excitatory and inhibitory effects as a result of interacting various stimuli were also seen in recordings from motoneurons of the RLN and nucleus ambiguus. The effects described show characteristics similar to those noted in other oral-facial muscles and indicate common integrative and regulatory mechanisms. This study in the adult cat provides a basis of comparison for future studies in the kitten where the effects of laryngeal stimulation are augmented to such an extent that they have been implicated in disorders such as the sudden infant death syndrome (SIDS).
INTRODUCTION
The internal laryngeal musculature is involved in a multiplicity of func- tions ranging from relatively simple reflex events such as glottic closure, gagging, and coughing to more complex motor activities such as swallow-
Abbreviations : RLN, SLN-recurrent, superior laryngeal nerves ; SIDS-sudden infant death syndrome; IX, X, XII-ninth, tenth, twelfth cranial nerves.
200
0014-4886/78/0621-0200$02.00/O Copyright @ 1978 by Academic Press, Inc. All rights of reproduction in any form reserved.

REGULATION OF LARYNGEAL REFLEXES 201
ing. Most of the laryngeal muscles, many of which show respiratory-related activity, receive their motor supply from nucleus ambiguus motoneurons in the caudal brain stem (3, 4). A particularly powerful reflex excitant of the recurrent laryngeal nerve (RLN) and laryngeal adductor muscle activity is sensory information passing in the superior laryngeal nerve (SLN) which concomitantly suppresses laryngeal abductor muscle activity ; stimulation of other nerves (e.g., trigeminal, intercostal) or areas supplied by them tends to evoke weaker effects in both groups of muscles (3, 15, 18, 27, 34). Little information is available, however, on the interaction of these various stimuli in the control of the laryngeal musculature. Even less is known of the maturation and development of laryngeal reflex activity, and yet upper respiratory tract stimuli in a number of animal species can reflexly induce a powerful cessation of respiration which is especially pro- longed, and sometimes fatal, in the neonate and which has been implicated in disorders such as the sudden infant death syndrome (SIDS) (3, 6, 21, 24, 25, 30). It was also suggested, however, that spasm of the laryngeal muscles ( 11, 14, 33)) or lack of adequate laryngeal reflex activity (IS), may contribute to the etiology of this disorder. The purpose of this study was to determine what effect particular neural and chemical stimuli that we have previously shown produce apnea in the adult cat (25) and kitten (S) have on laryngeal muscle activity in the adult cat and thereby provide a basis for future comparison with such activity in the developing animal. Some results were briefly reported previously (7, 23).
METHODS
The study was carried out on 18 adult cats (2.5 to 4.0 kg) anesthetized with chloralose (60 mg/kg). Each animal’s blood pressure, expired per- centage COa, and rectal temperature were continuously monitored. The following nerves were dissected and carefully isolated on the right side for subsequent electrical stimulation : infraorbital nerve, internal branch of the SLN, glossopharyngeal nerve (IX), cervical vagus (X), and the RLN which was exposed about 5 mm below the cricoid cartilage ; X was also exposed more laterally at this same level. In some animals the contralateral SLN, IX, and X, and ipsilateral hypoglossal and phrenic nerves were also exposed. A cannula was inserted into the trachea and directed up toward the larynx so that fluids (isotonic saline, distilled water, saturated sodium bicarbonate) might be delivered into the larynx by using a syringe attached to the cannula. A saturated sodium bicarbonate solution was used because we had previously noted its infusion to be particularly effective in inducing apnea (8, 25) ant1 exciting ST,N afferents (32). Another tracheal cannula was inserted in a cautlal direction to ensure a patent airway and to prevent fluids from entering the lungs, and to allow artificial

202 LUCIER, DAYNES, AND SESSLE
respiration, if necessary, by connections to a ventilator (Model 607, Harvard Apparatus, Millis, Massachusetts). Respiration was continuously monitored by sampling (Medical Gas Analyzer LB-l, Beckman Instru- ments, Palo Alto, California) expired CO2 from this cannula ; in some animals, respiration was also monitored by recording phrenic neural activity.
In experiments in which recordings were made from internal laryngeal muscles, fluids were not applied through a cannula but delivered directly to the laryngeal mucosa exposed by a ventral midline incision. Bipolar recordings were made from the ipsilateral thyroarytenoid or posterior cricoarytenoid muscles by using fine wire electrodes (1) inserted 1 to 2 mm apart into the muscle. The activity of the cricothyroid muscle was similarly recorded except that no laryngeal exposure was required. Re- sponses were amplified (preamplifier 113, PAR, Princeton, New Jersey), displayed on an oscilloscope (D13/5AlSN Tektronix, Beaverton, Oregon), and selected responses were photographed (utilizing camera C4P, Grass Instruments, Quincy, Massachusetts, and Polaroid Camera C5, Tektronix, Beaverton, Oregon). Activity of units in the RLN supplying the laryngeal muscles was also recorded, usually in paralyzed cats that were artificially ventilated. A single-fiber dissection recording technique (29, 30) was used to record bipolarly the neural activity from the central cut end of the RLN after the nerve had been sectioned just before it entered the larynx.
During studies of brain stem respiratory neurons (24, 25) in cats paralyzed with gallamine triethiodide and artificially ventilated, units were also recorded in nucleus ambiguus, the site of the cell bodies of moto- neurons supplying the laryngeal muscles (3, 4). Recording sites were con- firmed histologically (19) and single motoneurons were identified by their antidromic response (22) to RLN or X stimulation (0.01 to 5.0 mA, 0.1 ms).
To test for synaptically evoked excitatory and inhibitory influences on ambiguus motoneurons, RLN units, or laryngeal muscle motor units, bipolar electrical stimuli were applied to SLN, IX, X, infraorbital, and hypoglossal nerves. Stimulus spread was minimized by covering each nerve-electrode interface with plastic sleeves and low-melting-point wax. Facilitatory and inhibitory influences were tested on tonic or respiratory- related activity of single motoneurons and units, and on their reflexly or antidromically evoked discharges. For the latter tests, conditioning pro- cedures were used similar to those described previously (19, 22). BriefIy, this involved the application of a conditioning stimulus to a nerve at various intervals preceding the test stimulus that reflexly or antidromically acti- vated the cell. The time course of any change in excitability produced by the conditioning was obtained by comparing, at various conditioning-test intervals, the total number of spikes evoked by 10 to 20 test (viz., control)

REGULATION OF LARYNGEAL REFLEXES 203
I-
IPSI SLN
B
SPONTANEOUS'
C-L SLN
f--z-- m II- Lb n-
IPSI SLN
C-L SLN
FIG. 1. Effects of peripheral stimuli on single units in thyroarytenoid muscle. A- the top left record shows the responses of a unit to ipsilateral (IPSI) IX stimulation; this reflex response was quite reproducible as illustrated in the top record in the middle column which shows its responses to five successive ipsilateral IX stimuli. Each of the other records in the left and middle columns illustrate the presence or not of responses to five successive stimuli to ipsilateral SLN, X, and IO, and contra- lateral (C-L) SLN and X stimuli. The bottom two records in the middle column show the respiratory enhancement of the unit’s reflex responsiveness to ipsilateral SLN stimulation when the SLN stimuli were delivered outside the expiratory-related burst of the unit (upper) and inside the burst (lower). The right column of records shows the unit’s (CONTROL) respiratory rhythmic activity related to expiration which is upward in the accompanying trace of percentage expired CO,. and the enhancement of this activity with concomitant apnea produced by SLN stimulation at 10 Hz delivered for the period indicated between the arrows. B-this unit fired to ipsilateral SLN, IX, IO, X, and contralateral SLN stimuli; the reproducibility of such effects is indicated by the top middle record showing five superimposed responses to five successive ipsilateral SLN stimuli. The unit fired spontaneously as shown, with an expiratory-related rhythm that was unaffected by saline infusion into the larynx. Sodium bicarbonate (BICARB.) infusion, however, caused sustained activity of the unit and disrupted respiration as illustrated. Calibrations : voltage, 0.2 mV; CO,, 4.5%; time, 10 ms for left and middle columns of records in A and B, and 10 s for right column of records except 4 s in A, CONTROL.

204 LUCIER, DAYNES, AND SESSLE
stimuli with the total number elicited by a similar number of test stimuli applied after the conditioning stimuli.
RESULTS
The findings are based on multiunit recordings from the laryngeal muscles and RLN, and on recordings of 110 single units from the muscles, RLN, and nucleus ambiguus. A total of 60 single units were recorded from the laryngeal muscles : 26 from the thyroarytenoid, 13 from the crico- thyroid, and 21 from the posterior cricoarytenoid muscles. In addition, 27 units were recorded from RLN, and 23 single motoneurons from the nucleus ambiguus.
Thyroarytenoid MuscEc. Multiunit recordings from this adductor muscle of the larynx revealed phasic activity related to the expiratory phase of respiration. At the single-unit level, expiratory-related rhythmic activity was noted in 16 of the 26 units recorded; the remaining 10 units showed no respiratory-related or tonic activity. This expiratory-related activity could be enhanced and transformed into a continuous firing pattern by the application of sodium bicarbonate to the larynx or stimulation of SLN. SLN stimulation of 10 Hz or greater appeared most effective, but often SLN stimuli at 1 Hz could produce this sustained discharge. The discharge was often maintained as long as the stimulus (SLN or sodium bicarbonate) was applied, and was accompanied by a period of prolonged apnea (Fig. 1). Although less effective, water and tactile stimulation of the larynx also caused excitation in some units, but saline was without effect. Stimulation of IX also sometimes caused some enhancement of activity, but X stimuli or stimulation of other sites such as the infraorbital nerve had little or no effect on rhythmic activity.
The powerful excitatory effect on the thyroarytenoid by upper respi- ratory tract stimuli was also apparent when short-latency effects of nerve stimulation were examined (Figs. 1, 2A). Very low-intensity (0.02 to 0.2 mA, 0.1 ms) SLN stimulation reflexly excited 25 of the 26 units (latency 8 to 20 ms), IX and X stimuli excited 17 and 16, respectively (latency 8 to 25 ms), and infraorbital nerve stimulation activated only four (latency 10 to 20 ms). The effects of contralateral stimuli were not routinely or systematically examined, but it was apparent that stimulation of the contralateral IX, X, or SLN could be effective in exciting some units (Fig. 1). The latency of the responses evoked was 1 to 2 ms longer than that of responses elicited by ipsilateral stimuli, and higher stimulus inten- sities were generally required, as others have reported (3, 17, 31).
These reflex effects, in particular those produced by SLN stimuli, were accompanied by visible glottic adduction occurring with each stimulus delivered every 1 or 2 s. If the frequency of the SLN stimulus was
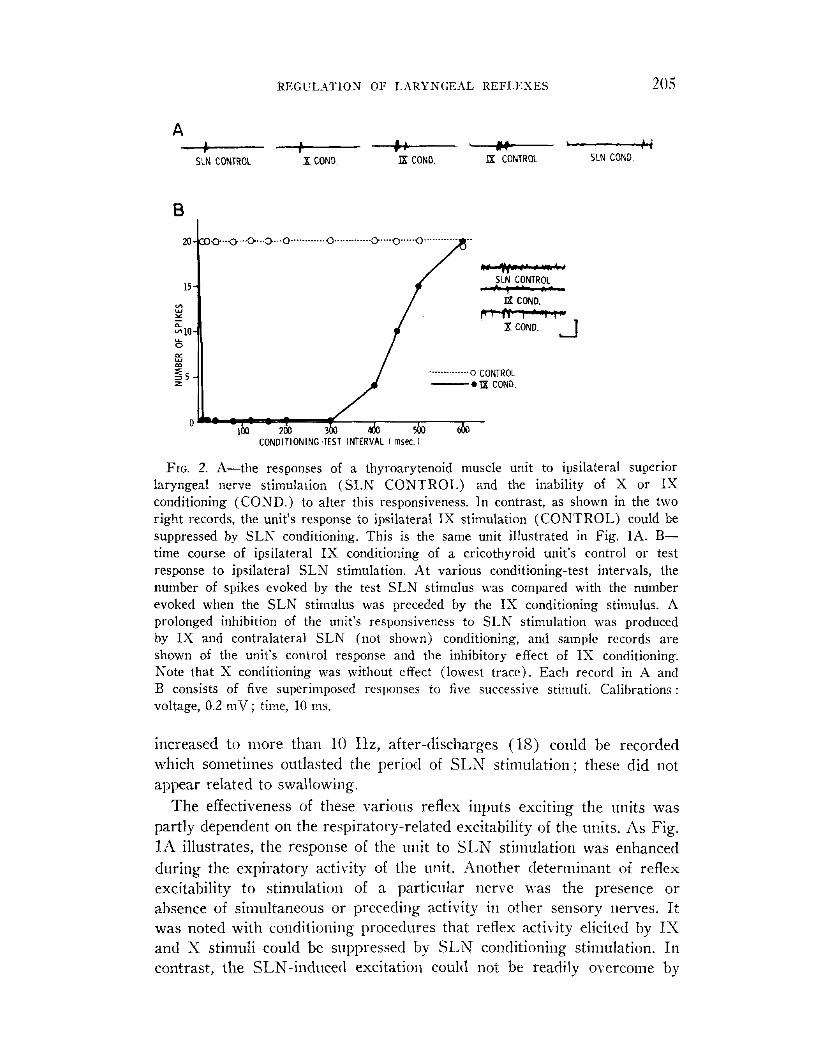
REGI.:LATION OF L.AKYNGEAL KEFLI~xES 205
A ----t------L ci
SLN CONTROL X COND. IX CON& IX CONTROL SLN COND.
m
15 IX CDND.
CONDITIONING-TEST INTERVAL f msec. 1
FIG. 2. A-the responses of a thyroarytenoid muscle unit to ipsilateral superior laryngeal nerve stimulation (SLN CONTROL) and the inability of X or IX conditioning (COND.) to alter this responsiveness. In contrast, as shown in the two right records, the unit’s response to ipsilateral IX stimulation (CONTROL) could be suppressed by SLN conditioning. This is the same unit illustrated in Fig. 1A. B- time course of ipsilateral IX conditioning of a cricothyroid unit’s control or test response to ipsilateral SLN stimulation. At various conditioning-test intervals, the number of spikes evoked by the test SLN stimulus was compared with the number evoked when the SLN stimulus was preceded by the IX conditioning stimulus. A prolonged inhibition of the unit’s responsiveness to SLN stimulation was produced by IX and contralateral SLN (not shown) conditioning, and sample records are shown of the unit’s control response and the inhibitory effect of IX conditioning. Note that X conditioning was without effect (lowest trace). Each record in A and B consists of five superimposed responses to five successive stimuli. Calibrations: voltage, 0.2 mV; time, 10 ms.
increased to more than 10 Hz, after-discharges (IS) could be recorded which sometimes outlasted the period of SLN stimulation ; these did not appear related to swallowing.
The effectiveness of these Tarious reflex inputs exciting the units was partly dependent on the respiratory-related excitability of the units. As Fig. 1A illustrates, the response of the unit to SLN stimulation was enhanced during the expiratory acti\+y of the unit. Another deterniinant of reflex excitability to stimulation of 3 particular nerve was the presence or absence of simultaneous or precedin g activity in other sensory nerves. It was noted with conditioning procedures that reflex activity elicited by IX and ,‘i stimuli could be suppressed by SIB conditioning stimulation. In contrast, the SLN-induced excitation could not be readily overcome by

206 LUCIER, DAYNES, AND SESSLE
0 I .*
d
FIG. 3. Activity of a single unit recorded in the posterior cricoarytenoid muscle. a-Part of the unit’s spontaneous activity that was related to inspiration. b-The spikes were passed through a window discriminator as shown in a, so that a dot display of instantaneous frequency could be obtained. The traces show the unit’s inspiratory-related discharge, the dot display of its discharge frequency, and the percentage expired CO% c-Infraorbital nerve (IO) stimulation for the period indi- cated by arrows affected neither the unit’s rhythmic activity nor respiration. d-In contrast, SLN stimulation (10 Hz) suppressed both unitary activity and respiration. Calibrations : voltage, 0.2 mV; frequency, 30 impulses/s ; CO?, 6.0% ; time 10 ms (a), 2 s (b), 10 s (c, d).
IX, X, and infraorbital nerve conditioning stimuli (Fig. ZA), e.g., in 8 of 13 units tested with IX and X conditioning, no change occurred in their reflex excitability to SLN stimulation. An early period of short (10 to 20 ms) facilitation occurred in four units, and X conditioning had a short inhibitory effect on the SLN-induced response in the remaining unit.
Cricothyroid ~~zzlscle. The expiratory-related activity and lack of suscepti- bility of SLN-induced reflex activity to other influences, characteristics noted for the thyroarytenoid, were not a feature of the cricothyroid recorded in the same animals. Multiunit and single-unit recordings revealed tonic or respiratory-related activity in only one unit. SLN stimulation evoked a reflex discharge in the cricothyroid, and 10 of the 13 single units recorded were excited (latency 8 to 20 ms) by this stimulus ; in contrast, IX, X, or infraorbital nerve stimuli excited only two units. Nonetheless, the SLN- induced activity was influenced by these other stimuli, and a prolonged suppression (as long as 400 ms or more), sometimes preceded by a short- lasting facilitation, was characteristic with IX and X conditioning stimula-

REGULATION OF LARYNGEAL REFLEXES 207
tion (Fig. ZB) . A prolonged inhibition often preceded by facilitation was typical of conditioning interactions between SIX, IS, S, and iufraorbital nerve stimuli on multiunit and single-unit activity.
Posterior CricoarJltmoid Mz~clc. Inspiratory-related activity was com- monly seen in multiunit recordings from this adbuctor muscle. Of the 21 single units recorded, 16 showed this characteristic. The inspiratory-related activity could be suppressed by upper respiratory tract stimuli, especially by low-intensity SLN stimulation (10 Hz or greater) and sodium bicarb- onate infusion which, as pointed out above, also could produce apnea and adductor excitation. Eleven of 13 neurons with a respiratory-related rhythm and tested with these stimuli exhibited suppression. Figure 3 shows a typical effect, although not all units showed such complete suppression of their inspiratory-related activity for the stimulus duration as shown. Other stimuli such as those to the infraorbital nerve (Fig. 3) or forepaw caused in contrast little modification of the inspiratory-related pattern of activity. Water infusion into the larynx was sometimes effective, but saline infusion caused no change in activity.
Only six of the 21 units could be excited by SLN, IS, or S stimuli. However, 10 units were activated by infraorbital nerve stimulation (latency 14 to 20 ms) ; conditioning effects on these evoked responses were not tested.
These various excitatory and inhibitory effects were also frequently noted in single-unit recordings from RLN and from nucleus ambiguus.
Recurrent Laryngeal Ncme. Respiratory-related activity ( 10 to 15 bursts/min) was noted in 13 of the 27 single units recorded in RLN. SLN stimulation and water or sodium bicarbonate infusion were effective in suppressing this activity in some of these units, but in five units showing respiratory-related activity, SLN stimulation produced a clear-cut short- latency activation. This excitatory effect (latency 6 to 15 ms) was also noted in the remaining 14 units in which respiratory-related activity was not a feature. Infraorbital nerve and IX stimuli were effective as excitatory or conditioning stimuli in only five units, but X stimulation reflexly excited 18 units (latency 9 to 20 ms), 10 of which showed respiratory- related activity. Stimulation of S also produced a prolonged inhibition of SLN-evoked activity in seven of 10 units tested, and in two of these the inhibition was preceded by a short-lasting facilitation. When the SLK was used for conditioning stimulation, suppression of X-evoked activity was the typical effect.
Nucleus Anzbigzlzis. Recordings of single-unit activity from this nucleus in the paralyzed animal allowed examination of the excitatory-inhibitory interactions in a situation free of movements that might have conceivably altered motor unit excitability. A respiratory-related rhythm was noted
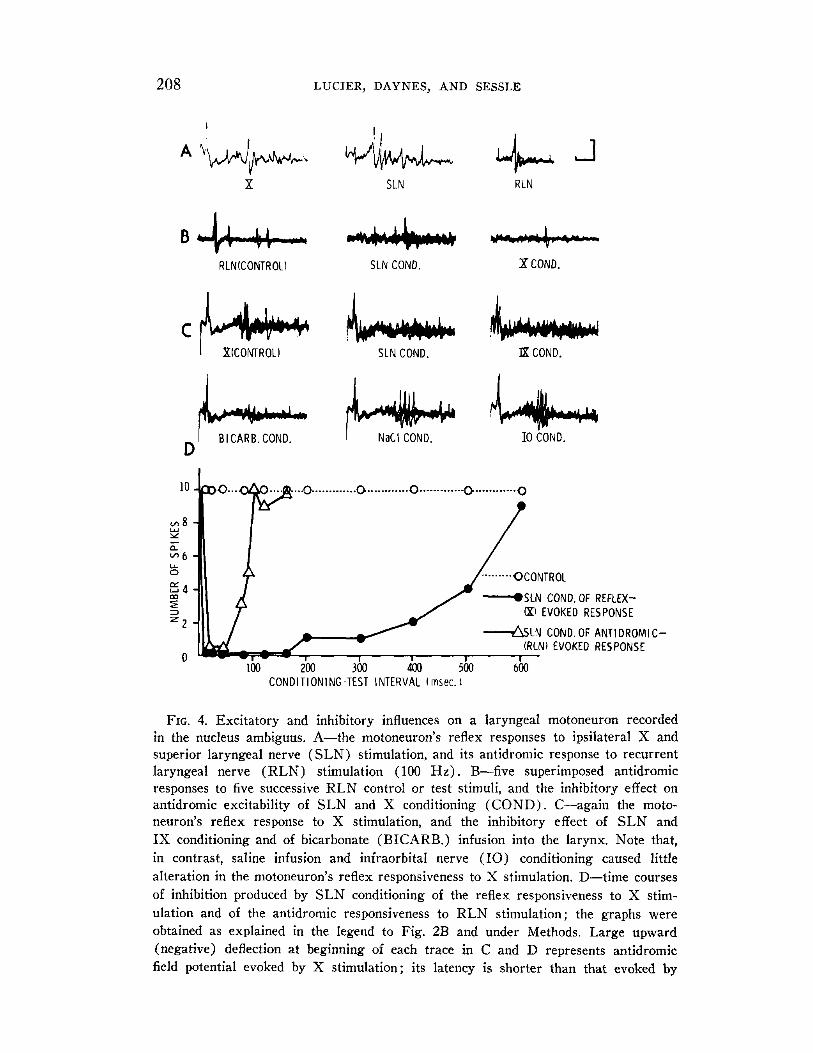
208 LUCIER, DAYNES, AND SESSLE
x SIN RLN
RLNfCONTROLI SLNCOND. XCOND.
D IO COND.
WSLN CONkOF REFLEX- (x) EVOKED RESPONSE
dLN COND.OF ANTIDROMIC- (RLNI EVOKED RESPONSE
100 200 300 400 500 600 CONDITIONING-TEST INTERVAL Imsec.1
FIG. 4. Excitatory and inhibitory influences on a laryngeal motoneuron recorded in the nucleus ambiguus. A-the motoneuron’s reflex responses to ipsilateral X and superior laryngeal nerve (SLN) stimulation, and its antidromic response to recurrent laryngeal nerve (RLN) stimulation (100 Hz). B-five superimposed antidromic responses to five successive RLN control or test stimuli, and the inhibitory effect on antidromic excitability of SLN and X conditioning (COND). C-again the moto- neuron’s reflex response to X stimulation, and the inhibitory effect of SLN and IX conditioning and of bicarbonate (BICARB.) infusion into the larynx. Note that, in contrast, saline infusion and infraorbital nerve (IO) conditioning caused little alteration in the motoneuron’s reflex responsiveness to X stimulation. D-time courses of inhibition produced by SLN conditioning of the reflex responsiveness to X stim- ulation and of the antidromic responsiveness to RLN stimulation; the graphs were obtained as explained in the legend to Fig. 2B and under Methods. Large upward (negative) deflection at beginning of each trace in C and D represents antidromic field potential evoked by X stimulation; its latency is shorter than that evoked by

REGULATION OF LARYKGEAL REFLEXES 209
in five of the motoneurons recorded in the nucleus as well as in 12 inter- neurons recorded in or adjacent (possibly the retroambiguus region (9) ) to the nucleus ; these interneurons, in contrast to the motoneurons, could not be antidromically excited by _X or RLN stimulation. As noted for single units in the RLN and the laryngeal muscle, both excitatory or sup- pressive effects on the rhythmic activity of the motoneurons and inter- neurons were noted after sodium bicarbonate or SLiY stimulation.
Of the 23 units identified as motoneurons by their antidromic response (mean latency + standard error : 2.37 + 0.25 ms), 15 could be reflexly activated by SLN (latency 5 to 10 ms), five by X (latency 6 to 10 ms), 10 by IX (latency 5 to 18 ms), and three units by high-intensity hypo- glossal nerve (latency 12 to 15 ms) stimuli (Fig. 4). SLN, X, and IX conditioning stimulation also resulted in a short period of inhibition, last- ing about 10 to 90 ms, of the antidromic responses of the motoneurons (Fig. 4) ; infraorbital or hypoglossal nerve conditioning was ineffective in seven motoneurons tested. A longer inhibition was noted when reflexly evoked activity was conditioned, e.g., single SLN conditioning stimuli were particularly effective in suppressing IS- and S-evoked reflex activity, and this suppression, sometimes preceded by facilitation, could last for 500 ms or more (Fig. 4).
DISCUSSION
Respiratory-related activity was a particular feature of the laryngeal muscles, and was also noted in some of the RLN nerve fibers and ambiguus motoneurons supplying them. Our findings of expiratory-related activity in the thyroarytenoid muscle during quiet respiration in the anesthetized cat, and of inspiratory-related activity in the posterior cricoarytenoid but little or no tonic activity in the cricothyroid muscle, are in agreement with earlier reports [e.g., see (12, 27, 34) 1. However, as pointed out pre- viously (9, 12, 27, 34), these activities can be enhanced or suppressed by a number of factors (e.g., depth of anesthesia, degree of postural activity in the muscles, ventilatory resistance, hypoventilation, and hyperventilation, breathing through larynx or tracheotomy tube) which undoubtedly account for the conflicting findings referred to earlier.
The primary drive for this respiratory-related laryngeal activity appears to come from brain stem respiratory centers, but superimposed on this drive may be modifying sensory influences from areas including lungs and chest wall and upper respiratory tract. These sensory influences can affect not only the respiratory rhythm of the laryngeal muscles, but eveu
RLN stimulation (B) especially because of the shorter conduction ‘ ath to brain stem from the X stimulation site. Each record in B-D represents five superimposed responses to five successive stimuli. Calibrations: voltage, 0.2 mV; time 4 ms.

210 LUCIER, DAYNES, AND SESSLE
respiration itself. A single low-intensity SLN stimulus in particular elicited a short-latency reflex discharge in the RLN and the adductors, the thy- roarytenoid and cricothyroid muscles ; the reflex excitability was enhanced when the stimulus was delivered during the expiratory activity of the muscles. Repetitive electrical stimulation of the SLN was especially effective in causing a continuous excitation of the thyroarytenoid muscle and visible laryngeal adduction, accompanied by cessation of respiration and suppression of the inspiratory-related activity of the abductor, the posterior cricoarytenoid muscle. These observations of the reflex effects of SLN electrical stimulation are in agreement with previous reports (14, 15, 18, 27, 31, 34) and appear related to glottic closure to protect the lower airway. The infusion into the larynx of fluids (water and sodium bicarbonate) previously observed to produce apnea also causes powerful excitation of adductor muscle and suppression of abductor muscle activity. This response apparently is initiated by receptors in the laryngeal mucosa which are particularly sensitive to certain chemical substances. Factors such as pH, temperature, osmolarity, and tactile stimulation appear to have been ruled out as prerequisites for the response (2, 29, 30)) and we (32) and Shingai (28) have noted that SLN “water” units (29), presumed responsible for water-induced apnea (30), are also powerfully excited by sodium bicarbonate.
Stimulation of other nerves or sites was not as effective as SLN stimuli. Nonetheless, many units could be excited by X, IX, and infraorbital nerve stimuli, in agreement with previous findings in RLN and laryngeal muscles (15, 27, 31). Conditioning of the reflex excitability of the laryngeal muscles and of the RLN fibers supplying them revealed that profound inhibitory effects as well as excitatory influences can modify the excitability of the laryngeal muscles. Such prolonged inhibitory effects may contribute for example to the suppressive effects of peripheral stimuli on laryngeal abductor activity. It is noteworthy that SLN-induced excitation of thy- roarytenoid activity is not readily modifiable by competing sensory influ- ences; this further emphasizes the powerful excitatory nature of this influence on laryngeal function.
These reflex inputs to laryngeal muscles and the complex interactions between them were also apparent in recordings of ambiguus motoneuron activity in the paralyzed cat. These motoneurons could be antidromically excited by X or RLN stimulation at latencies comparable to those of earlier studies (9, 10, 13). Their activation at short-latency by SLN and X stim- ulation was also reported (10, 13), but the present study has in addition revealed reflex excitation of some by XII stimulation, and, in conformity with the findings in RLN and laryngeal muscles, by IX stimulation.

REGULATION OF LARYNGEAL REFLEXES 211
Postsynaptic inhibitory effects of SLN stimuli in ambiguus motoneurons (10) probably contributed to the prolonged suppressive effects of con- ditioning noted in the present study. However, presynaptic depolarization of SLN, X, and IX afferent endings has been described in the solitary tract nucleus (5, 16, 19, 20), the site of interneurons for many reflex influences on laryngeal motoneurones (3). This, plus the long time course of the inhibition, suggests that presynaptic mechanisms in addition may be in- volved in the prolonged inhibition we noted in laryngeal motoneurons, as it appears to be for the reflexly induced suppression of jaw, facial, and hypoglossal motoneurons (22, 23, 26). Moreover, as ambiguus moto- neurons show many reflex interactions similar to those noted in these other oral-facial motoneurons, common integrative and regulatory mechanisms are suggested for these various motoneuron pools that participate in many of the same oral-facial motor functions, e.g., swallowing, suckling, and coughing.
The powerful reflex inputs and interactions described also likely come into play in functional disorders of the region. For example, laryngeal adductor activity may be especially sustained with laryngeal stimuli, may even outlast the stimulation, and cannot readily be overcome by competing sensory stimuli (e.g., IX, X). Such reflex effects may be involved in the etiology of laryngospasm (11, 14, 33). Their lack has been invoked ( 18) as a possible mechanism contributing to neonatal clinical problems such as aspiration during feeding and SIDS, although RLN-induced laryngeal closure may prove fatal in young animals, and this too has been implicated in SIDS (33). Because of such conflicting view points, and because pro- longed apnea may accompany the laryngeal adductor activity produced by upper respiratory tract stimuli and may even prove fatal in the neonatal animal (6, 21, 30), further studies are indicated of maturational changes in the reflex controls of the laryngeal muscles demonstrated here in the adult animal.
REFERENCES
1. BASMAJIAN, J. V., AND G. STECKO. 1962. A new bipolar electrode for electro- myography. J. Appl. Physiol. 17: 849.
2. BOUSHEY, H. A., P. S. RICHARDSON, J. G. WIDDICOMBE, AND J. C. M. WISE. 1974. The response of laryngeal afferent fibres to mechanical and chemical stimuli. J. Physiol. (Lo12don) 240: 153-175.
3. DOTY, R. W. 1968. Neural organization of deglutition. Pages 1861-1902 in C. F. CODE, Ed., Handbook of Physiology, Sect. 6, Alimentary Canal, Vol. 4. Amer- ican Physiological Society, Bethesda, Maryland.
4. GACEK, R. R. 1975. Localization of laryngeal motor neurons in the kitten. LaryizgoscoPe 85 : 1841-1861.

212 LUCIER, DAYNES, AND SESSLE
5. KENNY, D. J. 1976. Excitatory and Inhibitory Zltfluences front the Upper Respira- tory Tract on Neurons in the Solitary Tract Nucleus aped Adjacent Reticular Formation in the Cat, Ph.D. Thesis, University of Toronto, Toronto.
6. LEE, J. C., B. J. STOLL, AND S. E. DOWNING. 1977. Properties of the laryngeal chemoreflex in neonatal piglets. Am. J. Physiol. 233: R30-R36.
7. LUCIER, G. E., AND B. J. SESSLE. 1977. Effects of upper respiratory tract stimuli on the laryngeal musculature. Sot. Neurosci. Abstr. 3 : 869.
8. LUCIER, G. E., A. T. STOREY, AND B. J. SESSLE. 1978. Effects of upper respira- tory tract stimuli on neonatal respiration: reflex and single neuron analyses in the kitten. Biol. Neonate, In press.
9. MERRILL, E. G. 1970. The lateral respiratory neurones of the medulla: Their association with nucleus ambiguus, nucleus retroambigualis, the spinal accessory nucleus and the spinal cord. Brain Res. 24: 11-28.
10. MORI, M. 1973. Intracellular potentials of ambiguus motoneurons. Jap. J. Physiol. 23 : 25 l-259.
11. MURAKAMI, Y., AND J. A. KIRCHNER. 1972. Mechanical and physiological prop- erties of reflex laryngeal closure. Am. Otol. Rhihol. Larylrgol. 81 : 1-13.
12. MURAKAMI, Y., AND J. A. KIRCHNER. 1972. Respiratory movements of the vocal cords. An electromyographic study in the cat. Laryugoscope 82 : 454-467.
13. PORTER, R. 1963. Unit responses evoked in the medulla oblongata by vagus nerve stimulation. J. Physiol. (London) 168 : 717-735.
14. REX, M. A. E. 1970. Studies on the nerve pathways involved in the laryngeal reflex (laryngospasm) in the cat. Br. J. Anaesth. 42 : 1067-1071.
1.5. RUDOMIN, P. 1966. The electrical activity of the cricothyroid muscles of the cat. Arch. Znt. Physiol. Biochem. 74: 135-153.
16. RUDOMIN, P. 1967. Presynaptic inhibition induced by vagal afferent volleys. 1. Physiol. (London) 30: 964981.
17. SASAKI, C. T., AND M. SUZUKI. 1976. Laryngeal reflexes in cat, dog, and man. Arch. 0 tolaryngol. 102 : 400-402.
18. SASAKI, C. T., M. SUZUKI, AND M. HORIUCHI. 1977. Postnatal development of laryngeal reflexes in the dog. Arch. Otolaryngol. 103 : 138143.
19. SESSLE, B. J. 1973. Excitatory and inhibitory inputs to single neurones in the solitary tract nucleus and adjacent reticular formation. Brain Res. 53: 319-331.
20. SESSLE, B. J. 1973. Presynaptic excitability changes induced in single laryngeal primary afferent fibers. Brain Res. 53 : 333-342.
21. SESSLE, B. J. 1975. Sensory inputs and interactions in single brain stem neurones involved-in function of upper and lower respiratory tracts. Pages 131-144 in J. F. BOSMA AND J. SHOWACRE, Eds., Development of Upper Respiratory Anatomy and Function, U.S. Government Printing Office, Washington, D.C.
22. SESSLE, B. J. 1977. Modulation of alpha and gamma trigeminal motoneurons by various peripheral stimuli. Exp. Neurol. 54: 323-339.
23. SESSLE, B. J. 1978. Sensory regulation of brain stem motoneurones: physiological and clinical implications. In J. H. PERRYMAN AND L. VAN KIRK, Eds., Oral Physiology and Occlusion. Pergamon, New York. In press.
24. SESSLE, B. J., L. F. GREENWOOD, AND D. J. KENNY. 1974. Influences from upper respiratory tract on caudal brainstem neurones of cat. Can. J. Physiol. Pharm- acol. 52: 895498.
25. SESSLE, B. J., L. F. GREENWOOD, J. P. LUND, AND G. E. LUCIER. 1978. Effects of upper respiratory tract stimuli on respiration and single respiratory neurones in the adult cat. Exp. Ncurol. 61 : 245-259,

REGULATION OF LARYNGEAL REFLEXES 213
26. SESSLE, B. J,, AND D. J. KENNY. 1973. Control of tongue and facial motility: Neural mechanisms that may contribute to movements such as swallowing and sucking. Pages 222-231 in J. F. BOSMA, Ed., Fourth Sylrtposizkm olt Oral Serz- sation and Perception: Dcvelopmcut irt the Fetus aud Isfa?lt. National Institutes of Health, Bethesda, Maryland.
27. SHERREY, J. H., AND D. MEGIRIAN. 1974. Spontaneous and reflexly evoked laryn- geal abductor and adductor muscle activity of cat. Exp. Ncwol. 43 : 487-498.
28. SHINGAI, T. 1977. Ionic mechanism of water receptors in the laryngeal mucosa of the rabbit. Jap. J. Physiol. 27 : 27-42.
29. STOREY, A. T. 1968. A functional analysis of sensory units innervating epiglottis and larynx. Exp. Newel. 20 : 366-383.
30. STOREY, A. T., AXD P. JOHNSON. 1975. Laryngeal water receptors initiating apnea in the lamb. Exp. NcrGrol. 47: 42-55.
31. SUZUKI, M., AND C. T. SASAKI. 1977. Effect of various sensory stimuli on reflex laryngeal adduction. Am. Otol. Rhitrol. Larylzgol. 86 : 30-36.
32. SZOSE, L., G. E. LUCIER, AND B. J. SESSLE. 1978. Laryngeal receptor afferents in the adult cat responding to water and bicarbonate solutions. 21st Canadian Federation of Biological Societies Proceedings 21 : Abstr. 80, p. 20.
33. TAYLOR, E. M., D. SUTTON, C. R. LARSON, 0. A. SMITH, AND R. C. LINDEMAN. 1976. Sudden death in infant primates from induced laryngeal occlusion. Arch. 0 tolaryngol. 102 : 291-296.
34. WYKE, B. 1974. Respiratory activity of intrinsic laryngeal muscles; an experi- mental study. Pages 408-429 in B. WYKE, Ed., Ventilatory and Phonatory Control Systems, Oxford Univ. Press, London.





























