Embed Size (px)
Citation preview

Journal of Experimental Psychology:Animal Behavior Processes1992, Vol. 18, No. 2,115-125
Copyright 1992 by the American Psychological Association, Inc.0097-7403/9243.00
Latent Inhibition in Humans: Single-Cue Conditioning RevisitedOttmar V. Lipp and David A. T. Siddle
University of Queensland, Brisbane, Queensland, AustraliaDieter Vaitl
Department of Clinical Psychology, University of GiessenGiessen, Federal Republic of Germany
Latent inhibition in human Pavlovian conditioning was assessed by way of autonomic responses.In Experiment 1 (N = 72), three pairs of conditioning and control groups were preexposed to 0,10, or 20 to-be-conditioned stimuli (to-be-CSs), respectively. Acquisition of electrodermal first-interval and heart rate response conditioning were detectable only in the zero preexposurecondition. However, 20 preexposures were needed for latent inhibition of vasomotor responseconditioning. In Experiment 2 (N = 48), preexposure to the to-be-CS was compared withpreexposure to a stimulus that was not presented during subsequent acquisition. CS preexposurecompletely abolished electrodermal first-interval and heart rate response conditioning. Althoughvasomotor conditioning was not affected by preexposure, latent inhibition of second-intervalelectrodermal response conditioning was obtained. Taken together, the data from both experi-ments provide clear evidence for latent inhibition in human Pavlovian conditioning.
The retardation of Pavlovian conditioning following con-ditioned stimulus (CS) preexposure is a phenomenon welldocumented in animal research. The CS preexposure effect,also termed latent inhibition, has been demonstrated in sev-eral experimental designs in a variety of species (see Lubow,1973, 1989 for reviews). The phenomenon is of particularinterest because it offers an opportunity to test predictionsderived from recent models of Pavlovian conditioning (e.g.,Pearce & Hall, 1980; Wagner & Brandon, 1989). In researchon human Pavlovian conditioning, however, only a few stud-ies of latent inhibition have been performed. This is surprisingfor two reasons. First, in human research, as compared toanimal research, there is a paucity of theoretical accounts ofthe processing of simple stimuli (Siddle & Remington, 1987).Second, and from a very different perspective, research inlatent inhibition seems to be promising with respect to per-sonality and clinical psychology. Based on experiments withamphetamine-treated animals (Lubow, 1989; Weiner, 1990)and a few studies with humans (Baruch, Hemsley, & Gray,1988a, 1988b), there are good reasons to believe that latentinhibition might be an appropriate paradigm with which toassess attentional deficits in schizophrenics and "psychoti-cism-prone" normals. Each of these topics should be sufficientto motivate increased interest in this phenomenon.
Nevertheless, there is only sparse evidence for the existenceof latent inhibition in humans. This is especially true for the
A preliminary report of these data was presented at the 30th AnnualMeeting of the Society for Psychophysiological Research, held in NewOrleans, Louisiana, October 1989.
We thank Sabine Hirth for assistance with data collection inExperiment 2 and Rudolf Stark and Harald Gruppe for providingsoftware for acquisition and scoring of physiological data. Thanks aredue Robert Lubow for his critical comments on an earlier draft ofthis article.
Correspondence concerning this article should be addressed toOttmar V. Lipp, Department of Psychology, University of Queens-land, Brisbane, Queensland, Australia 4072. Electronic mail may besent to Ottmar @ psych.psy.uq.oz.au.
most frequently used preparation in human Pavlovian con-ditioning research, conditioning indexed by autonomic re-sponses. Eight studies have attempted to establish latent in-hibition in skin conductance conditioning. One of them failedto detect retardation of learning after to-be-CS preexposure(Zeiner, 1970), three discovered a retardation but only duringthe initial trials of acquisition (Bjorkstrand, 1990; Booth,Siddle, & Bond, 1989; Siddle, Remington, & Churchill, 1985),and one detected facilitation of conditioning after to-be-CSpreexposure (Silver, 1973). The retardation of learning re-ported in the remaining three studies (Brandeis, 1974; Maltz-man, Raskin, & Wolff, 1979; Surwit & Poser, 1974) is difficultto interpret because of shortcomings in design or procedure.In comparison to the clear-cut results in animal research, thispattern of results is not convincing. These data, together withthe difficulties in obtaining latent inhibition in other para-digms such as eyelid conditioning (Schnur & Ksir, 1969),prompted Lubow, Alek, and Arzy (1975) to propose thatmasking tasks are necessary to establish latent inhibition inadult humans. Masking tasks are applied to distract subjects'attention from stimulus preexposure or conditioning. Subjectsare instructed to perform these masking tasks as primary taskswhile the to-be-CS is presented or conditioning is adminis-tered. Such procedures are said to prevent the recognition ofthe change that occurs between preexposure and acquisitionand that Lubow et al. (1975) hypothesized results in reducedlatent inhibition. On the basis of these suggestions, Ginton,Urea, and Lubow (1975) developed an associative learningtask capable of producing latent inhibition, a retardation inthe ability to recognize the contingency between two stimuliafter preexposure of the target stimulus under conditions ofmasking. In this particular procedure, subjects were instructedto count the number of syllables read to them during preex-posure while the to-be-CS, a tone, was presented intermit-tently. Other masking tasks required subjects to perform aprobability learning task concurrent with preexposure andconditioning (Schnur & Ksir, 1969) or to memorize andrepeat the sequence of stimuli, the CS plus several others,presented on each trial (Booth et al., 1989).
115

116 O. LIPP, D. SIDDLE, AND D. VAITL
However, the necessity for masking in human autonomicconditioning remains questionable (Lubow, 1989). As shownby Booth et al. (1989), masking increased latent inhibition ofelectrodermal conditioning after CS preexposure, which was,although small, also present without masking. These resultssuggest a different approach to the function of masking.Besides the shift from preexposure to acquisition, maskingalso influences the speed of learning during acquisition itself,possibly by increasing task difficulty. This conclusion seemsreasonable if the data obtained from Booth et al.'s measureof unconditioned stimulus (US) expectancy during CS pre-sentations are taken into account. US expectancy was assessedcontinuously by asking the subjects to adjust a pointer on adial ranging from "certain that the US is about to occur" to"certain that the US is not about to occur." Whereas differ-ential learning, as indexed by this measure, was completedwithin four trials under no-masking conditions, a comparabledegree of stimulus differentiation was not obtained through-out the whole acquisition phase under masking conditions. Areliable CS preexposure effect, however, was evident with thismeasure only under masking conditions. Thus, whether ornot preexposure occurs during masking may be relativelyunimportant compared to the use of an appropriate testprocedure during acquisition.
Most of the recent experiments in human autonomic con-ditioning have used aversive USs (e.g., electric shock or loudnoise) and differential conditioning designs that allow within-subject control of associative processes. Differential condi-tioning is usually evident within a few trials. At least withrespect to first-interval electrodermal response conditioning,these designs are more sensitive to Pavlovian learning thanare single-cue conditioning procedures (Ohman, 1983). Onthe other hand, the rapidity of conditioning may minimize oreven neutralize latent inhibition effects induced by CS preex-posure. To circumvent such an obstacle, single-cue condition-ing designs may be preferable. In addition, single-cue condi-tioning designs, which involve between-subject controls forconditioning, allow one to disentangle the processes of inhib-itory and excitatory learning that are confounded in differ-ential conditioning. By use of single-cue conditioning, be-tween-subject control of associative learning, which seems tobe a disadvantage at first glance, offers an opportunity tomeasure the impact of CS preexposure solely on excitatoryconditioning. Although it is clear that control groups thatreceive noncontingent presentations of CS and US are nec-essary, such groups were not included in some of the earlierstudies (Surwit & Poser, 1974). Especially if physiologicalmeasures are recorded during conditioning, control groupsare the only means of separating changes in responding thatare due to nonassociative processes such as habituation orsensitization from changes in responding that are due toassociative learning. An additional manipulation to reducethe rapidity of conditioning might be to use a nonaversiveUS. Instead of an aversive US, a nonaversive reaction-timetask served as US in the present study. Such a US, althoughproposed (Ohman, 1983), has been used rarely because ofseveral weaknesses in comparison to aversive USs (Siddle etal., 1985). In two recent experiments, however, a reaction
time US capable of coping with these disadvantages wasintroduced (Lipp & Vaitl, 1988, 1990).
Given that a single-cue conditioning paradigm was chosento assess conditioning of physiological responses, two issuesconcerning the learning curves to be expected in the nopreexposure groups deserve mention. First, learning in theconditioning group of such a design occurs in most instancesafter one or two pairings of CS and US. This implies that themaximum conditioned response is likely to occur during theearly trials of acquisition and that a negatively acceleratedlearning curve is not expected (Maltzman, 1979). Second,orienting responses will occur during the initial trials of ac-quisition in the no preexposure conditioning and controlgroups because there is no habituation prior to conditioning.This implies that learning curves derived from measures thatalso reflect orienting (e.g., skin conductance responses to CSonset) differ from learning curves derived from measures thatdo not reflect orienting (e.g., skeletal reflexes). The latterusually show an increase in responding in the conditioninggroup and a stable, low level of responding in the controlgroup. Learning curves derived from physiological measuresthat reflect orienting activity display only a small increase inresponding during the initial part of acquisition and a subse-quent decline even in the presence of further CS-US pairings(Dawson & Schell, 1987; Maltzman, 1979). Responding incontrol groups, on the other hand, decreases rapidly becauseof habituation. However, the level of responding should besimilar for both conditioning and control groups during thefirst acquisition trial where only orienting causes responses.Moreover, learning curves derived from physiological meas-ures that are not that much influenced by orienting (e.g., skinconductance responses late during long-lasting CSs; Ohman,1983) should display the same features as learning curvesbased on skeletal muscle reflexes.
Aside from the questionable suitability of the experimentaldesign, another shortcoming of previous studies that assessedlatent inhibition in human autonomic conditioning shouldbe mentioned. In all of these studies, responses from only oneresponse system, electrodermal activity, were recorded. Al-though electrodermal activity is the most sensitive measurein human conditioning research, it seems appropriate for tworeasons to enlarge the scope of the present research by record-ing dependent measures from different response systems.First, there is evidence that experimental procedures differ-entially affect different response systems. This holds especiallyfor digital pulse amplitude responses in comparison to skinconductance responses. Differential susceptibility of thesemeasures has been found in several experimental designs,including habituation (e.g., Furedy, 1968) and Pavlovian con-ditioning (e.g., Hamm, Vaitl, & Lang, 1989). Second, in thefew experiments that have attempted to demonstrate latentinhibition of autonomic conditioning in animal research,researchers have exclusively used conditioning of heart ratechange. Although the results of these studies are equivocal(Lubow, 1989), heart rate response conditioning seems to bea sensitive index of latent inhibition. Thus, skin conductance,heart rate, and digital pulse amplitude responses served asdependent measures in the present experiments.

LATENT INHIBITION IN HUMANS 117
Experiment 1
In Experiment I, the number of CS preexposures was variedbetween groups. As animal research has indicated, the amountof subsequent latent inhibition covaries with the number ofpreexposures (Lubow, 1965). No guideline is available fromthe literature on the appropriate number of preexposure trialsrequired for reliable latent inhibition in human single-cueconditioning. However, a review of previous experimentsusing differential conditioning in humans suggests that 20preexposures are sufficient to produce latent inhibition. Tocomplete the design, 10 and 0 preexposures were administeredto separate groups. On the basis of the animal literature, weexpected latent inhibition to occur following 20 CS preexpo-sures, whereas we expected no retardation after a preexposureperiod without any stimulation. We expected that after 10preexposures, retardation of Pavlovian conditioning, if any,would be evident only during the initial trials of acquisition.
Method
Subjects. A total of 90 students, aged 18-39 years, participated;76 subjects received 10 deutsche marks (DM) for participation and14 volunteered to fulfill their course requirements. The data from 18subjects were rejected from statistical analyses because more than 5%of their data was missing. In these cases, either more than five skinconductance responses or more than three digital pulse amplituderesponses could not be scored. Missing data were due to eithermovement or respiration artifacts. Subjects were assigned randomlyto one of six groups, with the restriction that there were equal numbersof women and men in each group.
Apparatus. Skin conductance was recorded by means of a stand-ard 0.5-V circuit (Coulbourne S 71-22). Two silver/silver chlorideelectrodes (0.8 cm in diameter) were filled with a 0.05-M sodiumchloride paste and attached to the thenar and hypothenar eminencesof the subject's right hand. An electrocardiogram (ECG; lead II) wasrecorded using blank steel plate ECG electrodes (Siemens) and SYN-APSE electrode paste. Digital pulse amplitude was measured by aphotoplethysmographic device (Siemens E2031) placed on the tip ofsubject's right index finger. Respiration was recorded using a cheststrain gauge (Coulbourne S 72-25).
Yellow light from a bulb (Osram, 100 W) served as the to-be-conditioned stimulus. To achieve an indirect colored lighting of thesubject's room, the bulb was placed behind a monochrome TVmonitor (Loewe Electronic) that was used to display reaction timefeedback. The TV monitor was placed 150 cm in front of the subjectat eye level. A tone (1000 Hz, 70 dB) served as the "go" stimulus forthe reaction time task. Tone duration was 3 s, if it was not terminatedby the appropriate response, which was pressing a small buttonunderneath the left index finger. The tone was presented throughfour speakers mounted on the ceiling of the subject room. Reactiontime was displayed on the TV monitor for 4 s after each presentation.After the first five trials, additional feedback was provided wheneverthe actual reaction time was faster than the average reaction time ofthe previous five trials. As well as the actual difference score (d =average - actual reaction time), additional praise was displayedcorresponding to the magnitude of the differences (0 < d < 21 ms:"good"; 20 ms < d < 41 ms: "very good"; d > 40 ms: "excellent").Subjects were not informed with respect to the scaling of the feedback.
A microcomputer (Commodore C64) controlled the stimulus se-quence and timing, produced the tone stimulus, and also recorded
reaction times. Physiological data and events were recorded by aphysiopolygraph (Siemens Mingograf) with paper speed of 6 mm/s.In addition, the data were continuously recorded and stored on anIBM-compatible AT (Creussen AT), with different sampling rates foreach channel (events, 10 Hz; skin conductance and digital pulseamplitude, 25 Hz; ECG, 250 Hz).
Procedure. Subjects were seated in a comfortable reclining chairin a separate room. To mask sounds from outdoors and from theadjacent control room, white noise (55 dB) was delivered into thesubjects' room.
After attaching the measurement devices, all subjects were in-formed that the purpose of the experiment was to measure physiolog-ical responses elicited by simple light stimuli and during a reactiontime task. Subjects were asked to press the button as quickly aspossible whenever the tone was presented, but were told that theremight be some periods when no tones or lights would occur. Finally,they were asked to make themselves comfortable, to refrain frommoving, to breath regularly, and to pay attention to the stimuli.
Half of the subjects from each of the three preexposure conditions(0, 10, or 20 preexposure trials) were assigned to the conditioninggroup (Group E), whereas the other half of each preexposure condi-tion was assigned to the control group (Group C). During preexpo-sure, one pair of conditioning and control groups (E-PRE20/C-PRE20) received 20 presentations of the to-be-CS (light stimulus witha duration of 8 s). The intertrial interval (onset to onset) variedrandomly between 32 and 38 s. The second pair of groups (E-PRE10/C-PRE10) was exposed to no stimuli for a period of time equivalentto 10 preexposure trials, and then received 10 to-be-CS presentations.The third (E-PREO/C-PREO) was exposed to a period without anystimulation which lasted as long as 20 preexposure trials. Thus, theoverall duration of the preexposure period was the same in eachgroup.
During acquisition, conditioning groups received 20 pairings of CSand reaction time task US. The onset of the go stimulus coincidedwith CS offset (8-s delay conditioning). The intertrial interval (CSonset to CS onset) was the same as during the preexposure phase.Control groups also received 20 presentations of CS and US, but inan explicitly unpaired order. Stimuli were presented in a randomsequence, with the restriction that no more than two consecutivestimuli were the same. The first stimulus in this sequence was a USfor half of the subjects in each control group. The presentation of thestimuli (either CS or US) was separated by an interstimulus interval(stimulus onset to stimulus onset), varying randomly between 16 and19 s. The overall duration of the acquisition session was the same forthe conditioning and control groups. During extinction, 12 CSs werepresented to each group. The intertrial interval was the same as duringthe preexposure period. After the extinction phase, subjects wereasked to answer a few questions to assess their ability to verbalize thecontingencies.
Data reduction and response definitions. We inspected respirationcurves to assess respiration-related artifacts in physiological measures.Skin conductance responses (SCRs) and digital pulse amplitude re-sponses (DPARs) coinciding with coughs, sneezing, or sighing wereexcluded and replaced by estimates for missing data calculated bymeans of multiple regression using only the responses of the respectivesubject as predictors.
The SCRs elicited by CS presentations were calculated in threelatency windows, as recommended by Prokasy and Kumpfer (1973).First-interval responses (FIRs) began within 1 to 4 s after CS onset;second-interval responses (SIRs) began within 4 to 9 s after CS onsetand third-interval responses (unconditioned responses during acqui-sition in conditioning groups or omission responses during extinction)began within 9 to 13s after CS onset. In control groups, uncondi-tioned responses were measured from the 1-4 s period after US onset.

118 O. LIPP, D. SIDDLE, AND D. VAITL
Only FIRs were calculated during the preexposure phase. Changes inskin conductance were log transformed [log (1 + SCR)] and rangecorrected by dividing each subject's SCR by her or his maximumresponse (Lykken & Venables, 1971).
We calculated DPARs in a latency window of 2 to 10s after CSonset, as recommended by Furedy (1968). Within this period, thefirst of a sequence of three successive vasoconstrictive pulse ampli-tudes marked response onset (I > J > K or I > J > L), whereasresponse offset was indicated by the first of three consecutive vaso-dilative pulse amplitudes (T > U > V). To circumvent the problemof individual differences in responding because of differences inbaseline DPAR, we range corrected pulse amplitudes marking re-sponse onset (I) and offset (T). We used a covariance-analytic ap-proach to calculate change scores (Cronbach & Furby, 1970). Thepulse amplitude indicating the onset of vasoconstriction (I) served ascovariate. The adjusted amplitude of the offset pulse (T) was definedas the DPAR magnitude measure. The more pronounced the vaso-constriction, the smaller is the adjusted pulse amplitude score.
We calculated heart rate (HR) responses from raw ECG. The Rwaves of the ECG were triggered with offline software. Interbeatintervals were converted to HR on a half-second time base. HRchange scores were calculated for each trial within the CS interval of8 s by subtracting the mean HR during the last 2 s prior to CS onset.
Analyses. Responses during the preexposure phase were analyzedby means of separate analyses of variance (ANOVA)/analyses ofcovariance (ANCOVA) for groups preexposed to different numbersof stimuli (2 x 10 and 2 x 20, respectively). The first factor wasbetween-groups (Conditioning) and the second was within-subject(Trials). With respect to HR, a third factor (Half-Seconds), alsowithin-subject, was included.
A 3 x 2 X 20 factorial design was applied for acquisition and a 3x 2 x 12 factorial design was applied for extinction to analyze SCRand DPAR data. The first (Preexposure) and second (Conditioning)factors were between groups, whereas the third (Trials: 20 duringacquisition and 12 during extinction) was a within-subject factor. Inanalyses of HR responses, an additional within-subject factor (Half-Seconds) was included to complete a 3 x 2 x (20/12) x 17 factorialdesign. A 3 X 2 x 20 (Preexposure x Conditioning x Trials) factorialANOVA was calculated to assess differences in reaction time duringacquisition.
Main effects and interactions involving repeated measures weretested using degrees of freedom adjusted in terms of the degree ofasymmetry in the covariance matrices (Greenhouse & Geisser, 1959).Post hoc multiple comparison of means were calculated by means ofTukey tests. Level of significance was set at .05 in all analyses.
Results
Preexposure. Skin conductance FIRs decreased with stim-ulus repetition in both preexposure conditions. This reductionin electrodermal orienting was substantiated by main effectsfor Trials in both cases for PRE10, F(9, 198) = 25.6, MS, =.027; for PRE20, F(\9,418) = 17.3, MS, = .019. Comparableresults were obtained for DPAR. Again, responses declinedwith stimulus repetition: for PRE10, f\9, 197) = 2.4, MSe =.019; for PRE20, F(19, 417) = 2.8, MS, = .01. Reliable HRresponses during preperiod were obtained in PRE10 groupsonly: for main-effect Half-Seconds, F(16, 352) = 3.9, MS. =9.99, but not in Groups PRE20. There was no differencebetween conditioning and control groups during preexposure.
Acquisition. Retardation of Pavlovian conditioning afterCS preexposure was evident in electrodermal FIR, DPAR,
and HR responses. As depicted in Figure 1, electrodermalFIR in the conditioning group exceeded responding in thecontrol group if no stimuli were presented during preperiod.Following 10 or 20 preexposure trials, however, the perfor-mance of conditioning and control groups did not differreliably. This effect was substantiated by a Preexposure xConditioning interaction in the overall analysis, F(2, 66) =9.12, MS, = .24, and by separate analyses at each level ofPreexposure: for Main effect Conditioning PREO, F ( l , 22) =18.59, MS, = .36; for PRE10, F(l, 22) < 1; for PRE20, F(l,22) < 1. Responding in the conditioning group that receivedno stimuli during preperiod (E-PREO) did not exceed respond-ing in the respective control group (C-PREO) on the first trial,as can be seen in Table 1.
Analyses of HR responses revealed a comparable patternof results. The impact of CS preexposure, which can be seenin Figure 2, was substantiated by a Preexposure x Condition-ing x Half-Seconds interaction, F(32, 1056) = 2.34, MS, =18.7. Conditioning and control groups displayed different HRresponses only if no stimuli were presented during preperiod:for interaction Conditioning x Half-Seconds, F(16, 352) =6.9, MS, = 18.89. As separate analyses revealed, HR changedreliably in both groups: for main-effect trials E-PREO, F(16,176) = 7.7, MS, = 19.33; for C-PREO, F(16, 176) = 9.13,MS, = 18.4, but this change differed in direction. HR signif-icantly increased in group C-PREO, exceeding base-levelwithin Seconds 3 to 6 after CS onset (critical difference =2.09 beats per minute, bpm). Although Figure 2 seems tosupport such a conclusion, the change in HR displayed inGroup C-PREO did not reliably differ from the HR responsesdisplayed in the other control groups (F < I). In Group E-PREO, HR reliably decreased, falling below base level fromsecond 6 to US onset (t = 2.14). The minimum of the averageheart rate response displayed in Group E-PREO occurred 7 safter CS onset, whereas in Group C-PREO, the minimum HRwithin the period of reliable HR slowing in Group E-PREO,it occurred at second 7.5 (see Figure 2). Table 1 displays themean HR change scores during the first 3 trials of acquisition,which were obtained in Groups E-PREO/C-PREO at the re-spective times of minimum average HR. As can be seen, therewas no clear evidence for an enhanced HR deceleration inGroup E-PREO, as compared to Group C-PREO, during theinitial trials of acquisition.
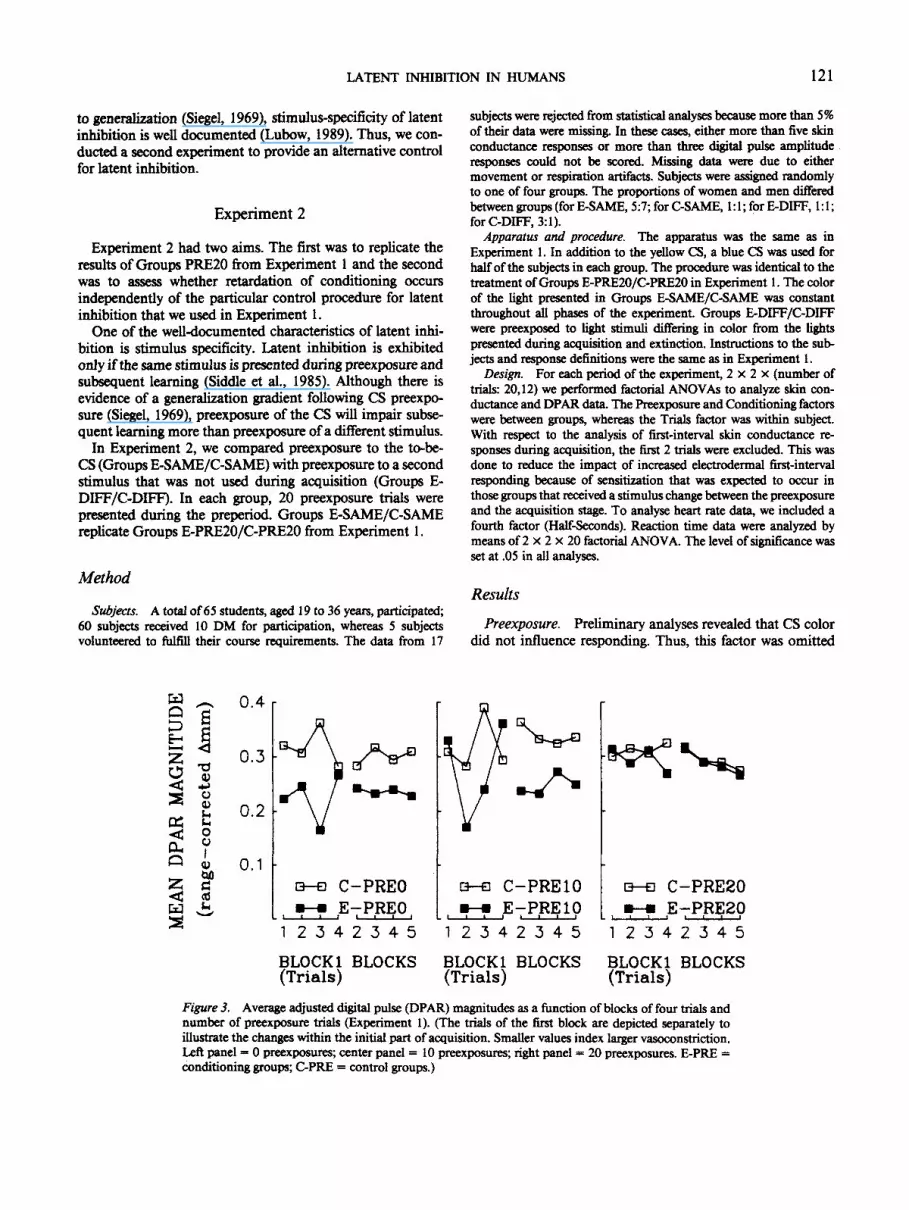
With respect to DPAR, the retarding effects of CS preex-posure was less marked. As can be seen in Figure 3, respondingin conditioning groups exceeded responding in the respectivecontrol groups after 0 and 10 preexposures to the CS, but notafter 20. This effect was substantiated by a Preexposure xConditioning interaction, F(2, 65) = 4.46, MS, = .05. Inseparate analyses at each level of preexposure, conditioningwas evident after 0 and 10 preexposures: for main-effectconditioning PREO, F(l,2l) = 8.24, MS, = .08; for PRE10,F(l, 21) = 11.85, MS, = .05, but not after 20. AlthoughFigure 3, which displays adjusted DPAR magnitudes, seemsto support such a conclusion, the differences between condi-tioning and control groups were not present during the initialtrials of acquisition, as the raw values displayed in Table 1indicate.

LATENT INHIBITION IN HUMANS 119
Number of CS preexposures did not significantly affectsecond-interval SCR conditioning during acquisition (Figure1). Conditioning was detected regardless of the number ofpreexposure trials. A main effect of Conditioning was revealedin the overall analysis, F(l, 66) = 8.44, MS, = .1, but nointeraction involving the preexposure factor reached signifi-cance.
Reaction time performance did not differ as a function ofeither group (conditioning or control) or preexposure condi-tion. In each group, reaction times decreased across trials,F(19, 1,254) = 59.41, MSC = .01.
Extinction. Third-interval electrodermal response condi-tioning was displayed irrespective of the number of preexpo-sure trials used. This was evidenced by a main effect ofConditioning, F(l, 66) = 12.47, MS, = .08, and a Condition-ing x Trials interaction, F(ll, 726) = 4.96, MSC = .02.Conditioning groups displayed larger overall third-intervalresponses than did control groups. Responses in conditioninggroups declined with stimulus repetition, F(\ 1, 363) = 10.8,MS, = .03, whereas responding in controls did not changesignificantly. A similar pattern was found for HR conditioningduring extinction. HR responses displayed in conditioninggroups differed from responses of controls, as indicated by a
Conditioning x Half-Seconds interaction, F(\6, 352) = 8.21,MSC = 14.6. In control groups, HR increased, F(16, 528) =5.74, MSe = 12.49, exceeding base level reliably 4 s after CSonset (critical difference = 1.22 bpm). The average responsein conditioning groups, however, was an HR decrease, F(\6,528) = 9.56, MSe = 16.72, peaking 6 to 8 s after CS onset(critical difference = 1.39 bpm). Stimulation during the preex-posure period did not affect the difference between groups.
During extinction, CS preexposure effects were evident onlyin electrodermal FIR and SIR. Conditioning groups displayedlarger responses than did control groups if no stimuli werepresented during the preperiod, but not after 10 or 20 preex-posures. This effect was confirmed by Preexposure x Condi-tioning interactions: for FIR, F(2, 66) = 7.08, MS, = .14; forSIR, F\2, 66) = 4.14, MS, = .02. In separate analyses at eachlevel of Preexposure, a main effect of Conditioning was de-tected only in Groups PREO: for FIR, F(l, 22) = 8.84, MS,= .21; for SIR, ^1,22) = 8.3, MS, = .01.
DPARs during extinction were not reliably affected eitherby conditioning or the number of preexposures.
Verbal report. Subjects' ability to verbalize the contingen-cies was assessed by the question "Did you detect any regu-larities in the occurrence of the reaction time task?", which
CO
a -5 -oS <u
1.0
0.8
0.6
0.4
0.2
E-PREOB-B C-PREO
E-PRE10B-O C-PRE10
•*»,
E-PRE20B-B C-PRE20
1 2 3 4 2 3 4 5 1 2 3 4 2 3 4 5 1 2 3 4 2 3 4 5
0.3
0.1
•-• E-PREOB-B C-PREO
•-• E-PRE10B-B C-PRE10
m-m E-PRE20B—B C-PRE20
1 2 3 4 2 3 4 5 1 2 3 4 2 3 4 5 1 2 3 4 2 3 4 5
BLOCK 1 BLOCKS(Trials)
BLOCK 1 BLOCKS(Trials)
BLOCK1 BLOCKS(Trials)
Figure 1. Average first- (upper panels) and second-interval electrodermal responses (lower panels)during acquisition as a function of blocks of four trials and number of preexposure trials (Experiment1). (The trials of the first block are depicted separately to illustrate the changes within the initial part ofacquisition. Left panels = 0 preexposures; center panels = 10 preexposures; right panels = 20 preexpo-sures. E-PRE = conditioning groups; C-PRE = control groups; SCR = skin conductance response.)

120 O. LIPP, D. SIDDLE, AND D. VAITL
was embedded in several other questions. Only 2 subjects (1in group E-PRE10 and 1 in Group C-PREO) were unable toanswer appropriately (correct answer for Group E: "Yes, thetone occurred after the light was switched off"; correct answerfor Group C: "No").
Discussion
Pronounced CS preexposure effects were detected duringacquisition of Pavlovian SCR, DPAR and HR conditioning.Among these measures, vasomotor responses seemed to bethe least susceptible to latent inhibition. Although latentinhibition was evident in first-interval SCR and HR responseconditioning following only 10 CS preexposures, 20 preex-posure trials were necessary to retard DPAR conditioning.Electrodermal SIR conditioning, however, was not affectedby CS preexposure and was present in all preexposure condi-tions. As expected, conditioning of physiological responsesthat reflect orienting (electrodermal FIR, DPAR, and HR)was established very early during acquisition in Groups PREO.The learning curves (e.g., electrodermal FIR) show the ex-pected pattern, a slight increase in responding followed by adecrease in Group E-PREO, as compared to a rapid decreasein Group C-PREO. In contrast to electrodermal FIR condi-tioning, conditioning of electrodermal SIR was not evidentduring the first three trials of acquisition (Table 1). The impactof CS preexposure was evident also during extinction. Bothfirst- and second-interval SCR conditioning were evident onlyif there were no CSs presented during preperiod.
In comparison with previous data, the extent of latentinhibition observed here is somewhat surprising. Whereas
Table 1Average Responses During First 3 Trials of Acquisition inGroups Displaying Conditioning in Experiment 1
Group Trial 1 Trial 2 Trial 3
Electrodermal FIRE-PREOC-PREO
Electrodermal SIRE-PREOC-PREOE-PRE10C-PRE10E-PRE20C-PRE20
Digital pulse amplitude*E-PREOC-PREOE-PRE10C-PRE10
Change in heart rateE-PREO (Sec 7)C-PREO (Sec 7.5)
0.650.58
0.110.130.050.090.060.12
0.330.370.350.28
-3.22-2.2
0.690.28
0.160.040.090.140.070.03
0.290.370.230.27
-4.72-0.91
0.680.28
0.120.040.050.050.090.02
0.240.450.300.42
-1.21-2.67
Note. FIR = first-interval responses; E-PREO and C-PREO = exper-imental and control groups receiving no presentations of stimulus; E-PRE10 and C-PRE10 = experimental and control groups receiving10 presentations of stimulus; E-PRE20 and C-PRE20 = experimentaland control groups receiving 20 presentations of stimulus. Sec =second.* Raw values.
3210•*
B -2a I,0 -3
PREO
W
COw
ffio
3210
-1-2-3
3210
-1-2-3
PRE10
Q-Q ControlConditipning
PRE20
0 1 2 3 4 5 6 7 8
SECONDS AFTER CS ONSETFigure 2. Heart rate responses (in beats per minute; bpm) duringacquisition averaged over 20 trials as a function of conditioned-stimulus (CS) duration and number of preexposure trials. (Experi-ment 1: upper panel = 0 preexposures; center panel = 10 preexpo-sures; lower panel = 20 preexposures.)
earlier studies demonstrated retardation of conditioning onlyduring the initial trials of acquisition, if any, the retardationof learning induced with the present procedure is rather clear-cut. Latent inhibition was detected after only 10 CS preex-posures. Furthermore, the effect was still present after 20pairings of CS and US during acquisition.
This pattern of results might not be due merely to CSpreexposure, but might have resulted from the procedure usedto control for latent inhibition. Although the application ofdifferent numbers of preexposure trials is a frequently usedcontrol procedure (Lubow, 1989), Siddle and Remington(1987) recommend preexposure to a different stimulus, onewhich is not used as the CS during acquisition, as an appro-priate control. Although the CS preexposure effect is subject
LATENT INHIBITION IN HUMANS 121
to generalization (Siegel, 1969), stimulus-specificity oflatentinhibition is well documented (Lubow, 1989). Thus, we con-ducted a second experiment to provide an alternative controlfor latent inhibition.
Experiment 2
Experiment 2 had two aims. The first was to replicate theresults of Groups PRE20 from Experiment 1 and the secondwas to assess whether retardation of conditioning occursindependently of the particular control procedure for latentinhibition that we used in Experiment 1.
One of the well-documented characteristics of latent inhi-bition is stimulus specificity. Latent inhibition is exhibitedonly if the same stimulus is presented during preexposure andsubsequent learning (Siddle et al., 1985). Although there isevidence of a generalization gradient following CS preexpo-sure (Siegel, 1969), preexposure of the CS will impair subse-quent learning more than preexposure of a different stimulus.
In Experiment 2, we compared preexposure to the to-be-CS (Groups E-SAME/C-SAME) with preexposure to a secondstimulus that was not used during acquisition (Groups E-DIFF/C-DIFF). In each group, 20 preexposure trials werepresented during the preperiod. Groups E-SAME/C-SAMEreplicate Groups E-PRE20/C-PRE20 from Experiment 1.
Method
Subjects. A total of 65 students, aged 19 to 36 years, participated;60 subjects received 10 DM for participation, whereas 5 subjectsvolunteered to fulfill their course requirements. The data from 17
subjects were rejected from statistical analyses because more than 5%of their data were missing. In these cases, either more than five skinconductance responses or more than three digital pulse amplituderesponses could not be scored. Missing data were due to eithermovement or respiration artifacts. Subjects were assigned randomlyto one of four groups. The proportions of women and men differedbetween groups (for E-SAME, 5:7; for C-SAME, 1:1; for E-DIFF, 1:1;forC-DIFF, 3:1).
Apparatus and procedure. The apparatus was the same as inExperiment 1. In addition to the yellow CS, a blue CS was used forhalf of the subjects in each group. The procedure was identical to thetreatment of Groups E-PRE20/C-PRE20 in Experiment 1. The colorof the light presented in Groups E-SAME/C-SAME was constantthroughout all phases of the experiment. Groups E-DIFF/C-DIFFwere preexposed to light stimuli differing in color from the lightspresented during acquisition and extinction. Instructions to the sub-jects and response definitions were the same as in Experiment 1.
Design. For each period of the experiment, 2 x 2 x (number oftrials: 20,12) we performed factorial ANOVAs to analyze skin con-ductance and DPAR data. The Preexposure and Conditioning factorswere between groups, whereas the Trials factor was within subject.With respect to the analysis of first-interval skin conductance re-sponses during acquisition, the first 2 trials were excluded. This wasdone to reduce the impact of increased electrodermal fust-intervalresponding because of sensitization that was expected to occur inthose groups that received a stimulus change between the preexposureand the acquisition stage. To analyse heart rate data, we included afourth factor (Half-Seconds). Reaction time data were analyzed bymeans of 2 x 2 x 20 factorial ANOVA. The level of significance wasset at .05 in all analyses.
Results
Preexposure. Preliminary analyses revealed that CS colordid not influence responding. Thus, this factor was omitted
ca _g ?E- J
1 *2 .25 «05 i•<! OCX o
Q iZ fl"<rj *CJ »-c
0.4
0.3
0.2
0.1B-e C-PREO
E-PREOB-O C-PRE10
E-PRE101 2 3 4 2 3 4 5 1 2 3 4 2 3 4 5 1
B-e C-PRE20m-m ETPRE202 3 4 2 3 4 5
BLOCK1 BLOCKS(Trials)
BLOCK1 BLOCKS(Trials)
BLOCK1 BLOCKS(Trials)
Figure 3. Average adjusted digital pulse (DPAR) magnitudes as a function of blocks of four trials andnumber of preexposure trials (Experiment 1). (The trials of the first block are depicted separately toillustrate the changes within the initial part of acquisition. Smaller values index larger vasoconstriction.Left panel = 0 preexposures; center panel = 10 preexposures; right panel = 20 preexposures. E-PRE =conditioning groups; C-PRE = control groups.)

122 O. LIPP, D. SIDDLE, AND D. VAITL
in all subsequent analyses. Skin conductance FIR decreasedwith stimulus repetition in each group, as indicated by a maineffect for Trials, F(19, 836) = 22.47, MSC = .02. Habituationof the electrodermal orienting response did not differ betweengroups. Light presentations elicited reliable HR responsesthroughout the preperiod: for main-effect Half-Seconds, F( 16,704) = 3.18, MSC = 13.6. This response did not change withstimulus repetition. With respect to DPAR, no changes weredetected during preperiod.
Acquisition. CS preexposure effects were evident in first-and second-interval SCR and in HR response conditioning.As displayed in Figure 4, first-interval electrodermal responsesin the conditioning group exceeded responses in controls onlyif a different stimulus was presented during preperiod. Thisfinding was confirmed by the overall analysis, which revealeda significant Preexposure x Conditioning interaction, F( 1,44)= 4.56, MS, = .31. In separate analyses at each level ofpreexposure, conditioning was detected only if a different
m
HQD
O
S
KOCO
w
1.0
0.8
0.6
0.4
0.2
E-DIFFo-£) C-DIFF
•-• E-SAMEo-Q C-SAME
1 2 3 4 2 3 4 5 1 2 3 4 2 3 4 5
uuOOI
cu(10C(0
0.3
0.2
0.1
E-DIFFe-e C-DIFF
E-SAMEQ-Q C-SAME
1 2 3 4 2 3 4 5 1 2 3 4 2 3 4 5
BLOCK1 BLOCKS(Trials)
BLOCK1 BLOCKS(Trials)
Figure 4. Average first- (upper panels) and second-interval electro-dermal responses (lower panels) during acquisition as a function ofblocks of four trials and preexposed stimulus (Experiment 2). (Thetrials of the first block are depicted separately to illustrate the changeswithin the initial part of acquisition. Left panel = preexposure of adifferent [DIFF] stimulus; right panels = conditioned-stimulus preex-posure. E = conditioning groups; C = control groups; SCR = skinconductance response.)
stimulus was presented during preexposure: For main-effectconditioning, F(l, 22) = 8.89, MS, = .12. A similar patternof result was evident in SIR conditioning (see Figure 4). Again,there was a Preexposure x Conditioning interaction, F(1, 44)= 5.16, MSe = .08. The conditioning group displayed largerSIRs than the control group after preexposure to a differentstimulus, F([, 22) = 18.87, MS, = .08, but there was nodifference between conditioning and control groups when theCSs during preexposure and acquisition were the same. Elec-trodermal first- and second-interval responding in Group E-DIFF did not exceed responding in Group C-DIFF on thefirst trial of acquisition (Table 2).
Mean HR responses elicited during acquisition are depictedin Figure 5. As this figure indicates, responses displayed inthe respective groups were affected by the different preexpo-sure conditions. Statistical analysis substantiated this findingby revealing a Preexposure x Conditioning x Half-Secondsinteraction, F(16, 704) = 2.64, MS, = 16.4. Conditioning andcontrol groups displayed different HR responses only follow-ing preexposure to different stimuli—for Interaction Condi-tioning x Half-Seconds, F(16, 352) = 9.19, MS, = 12.1—which was not the case after preexposure to the CS usedduring acquisition. The HR in Group C-DIFF increased,F(16, 176) = 6.77, MS, = 11.4, exceeding base level withinSeconds 3-7 after CS onset (critical difference = 1.65 bpm).In Group E-DIFF, HR decreased, F(16, 176) = 12.24, MS, =12.7, falling below base level within the last 2 s prior to USonset (critical difference =1.74 bpm). The minimum of theaverage heart response displayed in Group E-DIFF occurred8 s after CS onset. The minimum HR that was displayed inGroup C-DIFF during the period of reliable HR slowing inGroup E-DIFF occurred at the same time. Table 2 shows themean HR change scores 8 s after CS onset that were obtainedin groups E-DIFF/C-DIFF during the first 3 trials of acquisi-tion. As can be seen, changes in HR varied widely, and theoverall pattern of enhanced HR slowing in Group E-DIFFwas not clearly evident within these initial trials of acquisition.
Contrary to Experiment 1, CS preexposure did not influ-ence DPAR conditioning. Irrespective of the stimuli presentedduring preexposure, conditioning groups displayed larger va-soconstrictive responses than did controls: for main-effectConditioning, F(l, 43) = 9.56, MS, = .07. In addition, reac-tion times did not differ between groups. However, reactiontimes became faster across trials, F(\9, 836) = 50.05, MS, =.01.
Extinction. Results obtained from the extinction phaseclosely mirrored the pattern obtained during acquisition. Dif-ferences in conditioning due to the kind of stimuli presentedduring preexposure emerged for skin conductance and heartrate conditioning, but not for the DPAR.
Although first-interval skin conductance responding in theconditioning group exceeded control performance after preex-posure to a different stimulus, this was not the case after CSpreexposure. The difference in conditioning due to the stim-ulation during preperiod was indicated by a Preexposure xConditioning interaction, F( 1,44) = 4.56, MS, = .31. Separateanalyses, however, revealed only a marginally significant dif-ference between Groups E-DIFF and C-DIFF: for main-effectConditioning, F(l, 22) = 3.8, MS, = .16.

LATENT INHIBITION IN HUMANS 123
Electrodermal SIR and TIR were also clearly affected bythe respective preexposure conditions. The differences be-tween preexposure conditions were substantiated by signifi-cant Preexposure x Conditioning interactions for SIR, F(l,44) = 4.28, MS, = .02, and TIR, F(l,44) = 5.87, MS, = .066.In both intervals, responses in the conditioning group werelarger than responses in the control group after preexposureto a different stimulus: for SIR, P(l, 22) = 7.77, MSC = .02;for TIR, F( 1,22) = 9.71, MSt = .06, but not after preexposureto the CS.
The same pattern of result was obtained for heart rateconditioning. The type of stimulus presented during preperiodreliably affected heart rate responding during extinction, asconfirmed by a Preexposure x Conditioning x Half-Secondsinteraction, F(16, 704) = 2.96, MS, = 12.7. Different heartrate responses in conditioning and control groups were evi-dent only if different stimuli were presented during preexpo-sure: for Interaction Conditioning x Half-Seconds, F(16, 352)= 4.69, MSe = 12.93, but not between groups preexposed tothe CS. In Group C-DIFF, HR did not change reliably, asrevealed by separate analyses. In Group E-DIFF, HR de-creased, F(16, 176) = 4.63, MS, = 13.18, falling below baselevel within Seconds 6-7 after CS onset (critical difference =2.28 bpm).
Verbal report. Seven subjects from Group E-SAME wereunable to verbalize the contingencies. All subjects in theremaining three groups correctly identified the relation be-tween CS and US.
Supplementary Analyses
Experiments 1 and 2 yielded similar results for electroder-mal first-interval response and heart rate conditioning after20 to-be-CS preexposures. No conditioning was evidentthroughout the acquisition phase. Less straightforward con-clusions can be drawn from electrodermal second-interval
Table 2Average Responses During First 3 Trials of Acquisition inGroups Displaying Conditioning in Experiment 2
Group
Electrodermal FIRE-DIFFC-DIFF
Electrodermal SIRE-DIFFC-DIFF
Digital pulse ampli-tude
E-DIFFC-DIFFE-SAMEC-SAME
Change in heart rateE-DIFF (Sec 8)C-SAME (Sec 8)
Trial 1
0.510.51
0.060.04
0.280.290.270.39
-2.381.25
Trial 2
0.450.35
0.120.05
0.280.260.220.30
-0.68-4.07
Trial3
0.440.08
0.120.05
0.240.340.240.35
-4.711.94
B—Q Controli Conditioning
Note. FIR = first-interval response; SIR = second interval response;E-DIFF and C-DIFF = experimental and control groups exposed toa second stimulus; E-SAME and C-SAME = experimental and controlgroups exposed to to-be-conditioned stimulus. Sec = second.
SECONDS AFTER CS ONSETFigure 5. Heart rate responses (in beats per minute; bpm) duringacquisition averaged over 20 trials as a function of conditioned-stimulus (CS) duration and preexposed stimulus. (Experiment 2:upper panel = preexposure of a different [DIFF] stimulus; lowerpanel = CS preexposure.)
and vasomotor response data. Electrodermal SIR condition-ing did not reliably differ between preexposure conditions inExperiment 1, whereas differences were evident in Experi-ment 2. The reverse result was obtained in DPAR condition-ing. Supplementary analyses were performed to clarify theseequivocal results and to test whether there was conditioningafter 20 to-be-CS preexposures.
A 2 x 2 X 2 (Experiment x Conditioning x Half ofAcquisition) factorial ANOVA was calculated to assess theimpact of 20 to-be-CS preexposures on subsequent electro-dermal SIR conditioning. This analysis incorporated datafrom Groups E-PRE20/C-PRE20 from Experiment 1 and E-DIFF/C-DIFF from Experiment 2. A 2 X 2 x 2 ANCOVAwas calculated to assess DPAR conditioning. The pulse am-plitude indicating the onset of vasoconstriction served ascovariate.
As depicted in Figure 6, electrodermal second-interval re-sponding in the combined conditioning groups reliably ex-ceeded responding in controls during the second half ofacquisition, but not during the first. This was substantiatedby a Conditioning x Half of Acquisition interaction, F(l, 44)= 4.13, MS, = .01. Separate ANOVAs for each half ofacquisition revealed no difference during the first half, F(l,47) < 1, but a main effect for Conditioning during the secondhalf, F( 1,47) = 5.32, MS, = .01. No main effect or interactioninvolving the Experiment factor reached significance (all Fs< 2). Analysis of DPAR revealed a main effect for Condition-

124 O. LIPP, D. SIDDLE, AND D. VAITL
0.12
1-10 11-20TRIALS
Figure 6. Average second-interval electrodermal responses duringacquisition as a function of blocks of 10 trials in groups that received20 preexposures of the to-be-conditioned stimulus (Experiment 1: E-PRE20/C-PRE20; Experiment 2: E-SAME/C-SAME; SCR = skinconductance response.)
ing, F(\, 43) = 5.25, MSC = .01 and an Experiment xConditioning interaction, F(\, 43) = 3.5, MSC = .01, which,although not significant (p = .068), was analyzed further inseparate ANCOVAs. Responding in the conditioning groupreliably exceeded responding in controls in Experiment 2,F(l, 21) = 6.16, MS, = .01. There was no difference betweengroups in Experiment 1, F(l, 21) < 1.
Discussion
The purpose of Experiment 2 was to replicate the mainfindings of Experiment 1 and to demonstrate the stimulusspecificity of the CS preexposure effect. Regarding the latterissue, there is clear evidence that latent inhibition as assessedin the present procedure is stimulus specific. No retardationof conditioning was detected for any measure after preexpo-sure to stimuli that were not used as CS during subsequentacquisition and extinction of Pavlovian conditioning.
The replication of the results obtained in Groups E-PRE20/C-PRE20 from Experiment 1 was successful. With the excep-tion of DPAR conditioning, which was also less susceptibleto latent inhibition in Experiment 1, no conditioning wasevident during acquisition following 20 preexposures of theto-be-CS. Thus, both experiments yielded the same resultswith respect to the acquisition of first-interval skin conduc-tance and heart rate response conditioning. In Experiment 2,latent inhibition was also reliably evident in second-intervalskin conductance response conditioning during acquisition,which was not the case in Experiment 1. This differencebetween experiments is due mainly to the conditioning per-formance in the respective groups, which were not preexposedto the CS. After preexposure to 20 different stimuli, second-interval SCR conditioning was more pronounced than follow-ing a preperiod without any stimulation (see Figures 1 and4).
Both experiments yielded similar results with respect tofirst- and second-interval SCR and DPAR conditioned re-sponding during extinction. In Experiment 2, impairment of
conditioning after CS preexposure was also evident in third-interval SCR and in heart rate conditioning, which was notthe case in Experiment 1. This additional evidence of latentinhibition may be due to an even more pronounced effect ofCS preexposure in Experiment 2, which was characterized bya greater number of subjects who were unable to verbalize thecontingencies.
General Discussion
Two separate experiments demonstrated retardation of hu-man Pavlovian autonomic conditioning following CS preex-posure. Retardation of single-cue conditioning was evident ifeither different numbers of preexposures to the CS or preex-posure to a different stimulus were used as control conditions.In both experiments, pronounced latent inhibition was ob-tained in conditioning of first-interval skin conductance andheart rate responses. Evidence for latent inhibition in digitalpulse amplitude response was detected only in Experiment 1and evidence for latent inhibition in electrodermal second-interval response conditioning was detected only in Experi-ment 2.
The extent of latent inhibition with the present preparationis in contrast to results reported in the literature (Siddle &Remington, 1987). In previous experiments, reduction ofconditioning following CS preexposure occurred only early inthe acquisition series. The present results may be due to thecombination of two features of the experimental procedure:the use of single-cue conditioning as opposed to differentialconditioning, and the application of an effective US that isnot aversive. This combination seems to be more appropriatefor eliciting latent inhibition than the use of single-cue con-ditioning with aversive USs (Brandeis, 1974; Silver, 1973;Surwit & Poser, 1974), or differential conditioning with areaction time task US (Siddle et al., 1985). Although it ispremature to propose a preferred experimental paradigm forresearch on latent inhibition in humans, the conclusion canbe drawn that masking is not necessary in such a paradigm.As documented by Booth et al. (1989), masking tasks canincrease CS preexposure effects in human electrodermal con-ditioning, but they are not prerequisites for its occurrence.The present results, however, suggest that preexposure to adifferent stimulus is a better control procedure for latentinhibition than is variation in the number of preexposures.The former procedure, as used in Experiment 2, was morelikely to reveal effects of to-be-CS preexposure, at least ifconditioning of electrodermal and heart rate responses areconsidered.
Besides discrepancies regarding previous experiments inhuman Pavlovian conditioning, the present results are alsodifficult to interpret within the framework of animal modelsof Pavlovian conditioning. Although these models (e.g.,Pearce & Hall, 1980; Lubow, Weiner, & Schnur, 1981; Wag-ner & Brandon, 1989) differ widely with respect to the basicprocesses assumed to govern latent inhibition, each modelsuggests that conditioning is eventually obtained when thepreexposed stimulus is repeatedly paired with the US. Thus,these models find it difficult to accommodate the completesuppression of electrodermal first-interval response and heartrate response conditioning throughout 20 pairings of CS and

LATENT INHIBITION IN HUMANS 125
US. However, that the latent inhibition produced in thepresent paradigm is not a question of all or nothing can beseen in the second-interval electrodermal data. Conditioningof electrodermal second-interval responses was reliably ob-tained during the second half of acquisition after 20 to-be-CSpreexposures.
The present data clearly demonstrate latent inhibition inhuman Pavlovian conditioning of autonomic responses. Asmentioned earlier, latent inhibition is a promising tool withseveral applications. Besides its impact in the developmentand testing of models of human Pavlovian conditioning (Sid-die & Remington, 1987), it may be quite useful in researchin personality and clinical psychology (Baruch et al., 1988a,1988b; Lubow, 1989). The experimental procedure intro-duced in the present study may help to encourage furtherresearch in this area, and thus enlarge our understanding ofstimulus processing in humans and its relationship to suchprocesses in animals.
References
Baruch, I., Hemsley, D. B., & Gray, J. A. (1988a). Latent inhibitionand "psychotic proneness" in normal subjects. Personality andIndividual Differences, 9, 777-783.
Baruch, I., Hemsley, D. B., & Gray, J. A. (1988b). Differentialperformance of acute and chronic schizophrenics in a latent inhi-bition task. Journal of Nervous and Mental Disease, 176, 598-606.
Bjorkstrand, P.-A. (1990). Effects of conditioned stimulus pre-expo-sure on human electrodermal conditioning to fear-relevant andfear-irrelevant stimuli. Biological Psychology, 30, 35-50.
Booth, M. L., Siddle, D. A. T., & Bond, N. W. (1989). Effects ofconditioned stimulus fear-relevance and preexposure on expect-ancy and electrodermal measures of human Pavlovian condition-ing. Psychophysiology, 26, 281-291.
Brandeis, R. (1974). The influence of number of preexposures to onestimulus and to two stimuli on the latent inhibition effect in humanGSR conditioning. Unpublished master's thesis, Bar Ilan Univer-sity, Ramat Gan, Israel.
Cronbach, L. J., & Furby, L. (1970). How we should measure"change"—or should we? Psychological Bulletin, 74, 68-80.
Dawson, M. E., & Schell, A. M. (1987). Human autonomic andskeletal classical conditioning: The role of conscious cognitivefactors. In G. Davey (Ed.), Cognitive processes and Pavlovianconditioning in humans (pp. 27-55). Chichester, England: JohnWiley & Sons.
Furedy, J. J. (1968). Human orienting reaction as a function ofelectrodermal versus plethysmographic response modes and singleversus alternating stimulus series. Journal of Experimental Psy-chology, 77, 70-78.
Ginton, A., Urea, G., & Lubow, R. E. (1975). The effect of preex-posure to a nonattended stimulus on subsequent learning: Latentinhibition in adults. Bulletin of the Psychonomic Society, 5, 5-8.
Greenhouse, S. W., & Geisser, S. (1959). On methods in the analysisof profile data. Psychometrika, 24, 95-112.
Hamm, A. O., Vaitl, D., & Lang, P. J. (1989). Fear conditioning,meaning, and belongingness: A selective association analysis. Jour-nal of Abnormal Psychology, 98, 395-406.
Lipp, O. V., & Vaitl, D. (1988). Reaction time task as unconditionalstimulus: On conditioning skin conductance responses and heartrate, using a nonaversive unconditional stimulus. Pavlovian Journalof Biological Science, 23, 165-172.
Lipp, O. V., & Vaitl, D. (1990). Reaction time task as unconditionalstimulus: Comparing aversive and nonaversive unconditional stim-uli. Pavlovian Journal of Biological Science, 25, 77-83.
Lubow, R. E. (1965). Latent inhibition: Effects of frequency ofnonreinforced preexposure of the CS. Journal of Comparative andPhysiological Psychology, 60, 454-457.
Lubow, R. E. (1973). Latent inhibition. Psychological Bulletin, 79,398-407.
Lubow, R. E. (1989). Latent inhibition and conditioned attentiontheory. Cambridge, England: Cambridge University Press.
Lubow, R. E., Alek, M., & Arzy, J. (1975). Behavioral decrementfollowing stimulus preexposure: Effects of number of preexposures,presence of a second stimulus, and interstimulus interval in chil-dren and adults. Journal of Experimental Psychology: AnimalBehavior Processes, 104, 178-188.
Lubow, R. E., Weiner, I., & Schnur, P. (1981). Conditioned attentiontheory. In G. Bower (Ed.), The Psychology of Learning and Moti-vation (Vol. 15, pp. 1-49). San Diego, CA: Academic Press.
Lykken, D. T., & Venables, P. H. (1971). Direct measurement ofskin conductance: A proposal for standardization. Psychophysiol-ogy, 8, 656-672.
Maltzman, I. (1979). Orienting reflexes and classical conditioning inhumans. In H. D. Kimmel, E. H. van Olst, & J. F. Orlebeke (Eds.),The orienting reflex in humans (pp. 323-353). Hillsdale, NJ: Erl-baum.
Maltzman, I., Raskin, D. C., & Wolff, C. (1979). Latent inhibition ofthe GSR conditioned to words. Physiological Psychology, 7, 193-203.
Ohman, A. (1983). The orienting response during Pavlovian condi-tioning. In D. Siddle (Ed.), Orienting and habituation: Perspectivesin human research (pp. 315-369). Chichester, England: John Wiley& Sons.
Pearce, J. M., & Hall, G. (1980). A model for Pavlovian learning:Variations in the effectiveness of conditioned but not of uncondi-tioned stimuli. Psychological Review, 87, 532-552.
Prokasy, W. F., & Kumpfer, K. L. (1973). Classical conditioning. InW. F. Prokasy & D. C. Raskin (Eds.), Electrodermal Activity inPsychological Research (pp. 157-202). San Diego, CA: AcademicPress.
Schnur, P., & Ksir, C. J. (1969). Latent inhibition in human eyelidconditioning. Journal of Experimental Psychology, 80, 388-389.
Siddle, D. A. T., & Remington, B. (1987). Latent inhibition andhuman Pavlovian conditioning: Research and relevance. In G.Davey (Ed.), Cognitive processes and Pavlovian conditioning inhumans (pp. 115-146). Chichester, England: John Wiley & Sons.
Siddle, D. A. T., Remington, B., & Churchill, M. (1985). Effects ofconditioned stimulus preexposure on human electrodermal con-ditioning. Biological Psychology, 20, 113-127.
Siegel, S. (1969). Generalization of latent inhibition. Journal of Com-parative and Physiological Psychology, 69, 157-159.
Silver, A. I. (1973). Effects of prior CS presentations on classicalconditioning of the skin conductance response. Psychophysiology,10, 583-588.
Surwit, R. S., & Poser, E. G. (1974). Latent inhibition in the condi-tioned electrodermal response. Journal of Comparative and Phys-iological Psychology, 86, 543-548.
Wagner, A. R., & Brandon, S. E. (1989). Evolution of a structuredmodel of Pavlovian conditioning (AESOP). In S. B. Klein & R. R.Mowrer (Eds.), Contemporary learning theories: Pavlovian condi-tioning (pp. 149-189). Hillsdale, NJ: Erlbaum.
Weiner, I. (1990). Neural substrates of latent inhibition: The switchingmodel. Psychological Bulletin, 108, 442-461.
Zeiner, A. R. (1970). Orienting response and discrimination condi-tioning. Physiology and Behavior, 5, 641-646.
Received February 14, 1991Revision received June 14, 1991
Accepted August 8, 1991





























