Embed Size (px)
Citation preview

Person. individ. Difl Vol. 13, No. 9, PP. 1003-1012, 1992 0191-8869/92 $5.00 + 0.00 Printed in Great Britain. All rights reserved Copyright 0 1992 Pergamon Press Ltd
LATENT INHIBITION IN HUMAN PAVLOVIAN DIFFERENTIAL CONDITIONING: EFFECT OF
ADDITIONAL STIMULATION AFTER PREEXPOSURE AND RELATION TO SCHIZOTYPAL TRAITS*
OTTMAR V. LIPP’~. and DIETER VAITL’
‘Department of Psychology, University of Queensland, Brisbane QLD 4072, Australia and *Department of Clinical Psychology, University of Giessen, Otto Behaghel Str. 10,
6300 Giessen, Germany
(Received 13 September 1991)
Summary-Latent inhibition has been defined as the retardation of Pavlovian conditioning due to conditioned stimulus (CS) preexposure. The present study investigated (1) the effect of additional stimulation signalling the shift from preexposure to acquisition, and (2) the relationship between “schizotypal traits” and latent inhibition in human electrodermal conditioning. Three groups (48 subjects) were presented with 20 preexposure, 8 acquisition, and 8 extinction trials in a differential conditioning paradigm. One group received different stimuli during preexposure and acquisition, whereas the remaining groups (SAME, SAME + S) received the same stimuli throughout the experiment. In group SAME + S, an additional signal was presented at the end of the preexposure phase. Latent inhibition was evident in electrodermal first interval response conditioning during acquisition and second interval response conditioning during acquisition and extinction. Contrary to results from animal research, latent inhibition was not disrupted by the additional signal in group SAME + S. A covariation of schizotypal traits and latent inhibition was detected in both groups preexposed to the CSs. During acquisition, latent inhibition of electrodermal first interval response conditioning was evident in subjects scoring low in “schizotypy”, but not in high scorers. The latter results replicate previous findings obtained from a different latent inhibition paradigm.
Latent inhibition, the retardation of Pavlovian conditioning following to-be-conditioned stimulus (CS) preexposure, is viewed as a promising paradigm for research in human Pavlovian conditioning. Siddle and Remington (1987) have argued that latent inhibition may prove to be a key phenomenon to develop and test models of human stimulus processing, comparable to the impact of latent inhibition in animal research (see Lubow, 1989 for a review). Besides its relevance for theory testing, latent inhibition also seems of relevance in applied clinical research.
Lubow (1973) proposed the use of CS preexposure as a behavioural prophylactic measure in order to immunize against potentially phobic stimuli. Although two recent studies (Bjdrkstrand, 1990; Booth, Siddle & Bond, 1989) failed to demonstrate that the fear relevance of the preexposed CSs affects the extent of latent inhibition, this notion deserves further elaboration. Cannon, Best, Batson and Feldman (1983) demonstrated latent inhibition in a taste aversion paradigm in humans and pointed to the relevance of their results with respect to the prevention of nausea induced during cancer chemotherapy. In addition, latent inhibition paradigms seem to be appropriate for monitoring attentional malfunction in schizophrenia (Lubow, 1989). As Baruch, Hemsley and Gray (1988a) reported, symptom-free chronic schizophrenics and normals, but not acute schizophrenics, displayed latent inhibition as assessed in an associative learning task (Ginton, Urea & Lubow, 1975). In this particular preparation, the task was to learn the contingency between a white noise stimulus and a change on a scoreboard. Prior to the learning task, all Ss had to work on a different task, counting the number of syllables presented. The experimental group was preexposed to the noise stimulus during this task whereas no noise was presented in the control groups. Normals and chronic schizophrenics required more trials to reach the learning criterion after preexposure than did the respective no preexposure controls. There was no difference between experimental and control group in the subsample of acute schizophrenics. Comparable results were obtained in
*Portions of this paper were presented at the 1990 Meeting of rhe Society for Psychophysiological Research, Boston, MA. tTo whom correspondence should be addressed.
PAID 1319-D 1003

1004 OTTMAR V. LIPP and DIETER VAITL
normals who differed in measures of schizotypal traits, psychoticism and schizotypy (Baruch, Hemsley & Gray, 1988b). Low scorers displayed latent inhibition which was not the case in high scorers. Keeping in mind the caution which is necessary in interpreting these results, the methodological elegance of these experiments is striking. The presumed attentional malfunctioning in “psychosis prone” and schizophrenic Ss is tracked by improved performance in the learning task as assessed by reduced latent inhibition.
Given the variety of potential applications of latent inhibition, the number of studies performed with human Ss is remarkably low. This may be due to the problems involved in establishing the CS-preexposure effect in adult humans (Lubow, Alek & Arzy, 1975; Siddle & Remington, 1987). The results of the early studies are, at best, equivocal (Zeiner, 1970; Silver, 1973; Surwit & Poser, 1974). Summarizing the lack of empirical evidence for latent inhibition in adult humans, Lubow er al. (1975) concluded that additional experimental effort might be necessary to produce the phenomenon. Lubow et aE. argued that adults might recognize the shift from preexposure to acquisition and thus respond only to the new situation, with the result that latent inhibition is attenuated. To reduce Ss’ attention to the task characteristics, the authors proposed the use of masking tasks as a necessary prerequisite for obtaining latent inhibition in adults. This suggestion is supported by data from research in human eyelid conditioning and animal learning. Latent inhibition of eyelid conditioning in adults could be demonstrated only by use of a masking task (Lubow, 1989). In animals, latent inhibition was shown to be disrupted if the shift from preexposure to acquisition is accentuated by additional signals (Lantz, 1973; Rudy, Rosenberg & Sandell, 1977). However, more recent experiments in human autonomic conditioning have questioned the necessity for a masking task. Clear evidence of latent inhibition was obtained in both, differential (Siddle, Remington & Churchill, 1985; Booth et al., 1989; Bjorkstrand, 1990) and single cue conditioning (Lipp, Siddle & Vaitl, 1992) without any masking procedure.
To increase further the understanding of latent inhibition in human Pavlovian conditioning of autonomic responses, the present experiment was designed to answer the following questions: Does additional stimulation after a preexposure period and prior to acquisition reduce latent inhibition? As proposed by Lubow et al. (1975), an additional signal separating preexposure and acquisition should restore Ss’ attention and increase the distinctiveness of the different experimental phases. Do measures of schizotypal traits covary with the extent of latent inhibition displayed during Pavlovian conditioning indexed by autonomic responses? That is to say, are the results obtained by Baruch et al. (1988b) replicable in differential Pavlovian conditioning of autonomic measures (Siddle et al., 1985)?
Subjects METHOD
Fifty-two paid volunteers participated. The data from four Ss were rejected from subsequent analyses due to experimenter errors. The remaining 48 Ss (mean age = 23.3 years; range = 16-32 years) were allocated randomly to three groups with the restriction of the same proportion of males and females (1: 3) across groups. Ss were students at the University of Giessen or a neighbouring high school. All Ss were healthy and free of psychiatric symptoms.
Design
Three groups of Ss were included in the present experiment. Group DIFF received presentations of stimuli during preperiod that differed from the CSs employed during acquisition. Group DIFF was expected to display reliable conditioning and served as a control for latent inhibition effects (Siddle & Remington, 1987). Ss in group SAME were presented with the same stimuli during preperiod which were employed as CSs during acquisition. Latent inhibition was expected to occur in this group. To assess the effect of additional stimulation after CS-preexposure group SAME + S was included. Similar to group SAME, group SAME + S was exposed to the to-be-CSs during preperiod. However, an additional stimulus was presented in group SAME + S between preperiod and acquisition. It was hypothesized that the additional stimulation would disrupt latent inhibition, i.e. the conditioning performance in group SAME + S was expected to exceed conditioning performance in group SAME.

Latent inhibition and schizotypal traits 1005
In addition to appropriate controls for latent inhibition, studies which assess Pavlovian conditioning by means of autonomic responses have to employ a control for non-associative effects which might result in increased responses. For this purpose two procedures are available, single cue conditioning which requires separate control groups (Lipp et al., 1992) or differential conditioning which offers a within S control for conditioning (Booth et al., 1989). The latter procedure, which is more frequently used in contemporary research, was adopted in the present experiment. In a differential paradigm, conditioning is assessed by comparison of responding to a CS paired with the unconditioned stimulus (US) with that to a CS which is never paired with the US.
The relationship between schizotypal traits and latent inhibition was assessed in each of the three groups separately. It was hypothesized that the extent of latent inhibition will differ between high and low scorers in groups which overall display latent inhibition (e.g. group SAME). In groups which do not display latent inhibition (e.g. DIFF) no differences in conditioning between high and low scores were expected.
Apparatus
Skin conductance was recorded by a standard 0.5 V constant voltage circuit (Coulbourn S 71-22) using two Ag/AgCl electrodes of 0.8 cm dia. Electrodes were placed at the thenar and hypothenar eminences of the Ss right hand. A 0.05 molar NaCl paste served as electrolytic medium. Respiration was monitored with a chest strain gauge (Coulbourn S 72-25).
Lights (blue/yellow; Osram Concentra 100 W) and white graphics (square/cross) were used as CSs. The graphics (11 x 11 cm) were displayed on a monochrome TV monitor (Loewe Opta) which was placed 150 cm in front of the Ss at eye level. The bulbs delivering the light stimuli were placed behind the TV monitor, producing an indirect coloured illumination of the subject room. A tone (1000 Hz, 70 dB) was used as the “go” signal for the reaction time (RT) task employed as US. The tone was delivered through 4 speakers mounted underneath the ceiling of the subject room. The tone lasted for 3 set or was terminated by Ss’ response which consisted of pressing a small button underneath her/his left index finger. RTs were displayed on the TV screen. Beginning with the third RT trial, additional feedback was displayed whenever the actual RT was shorter than the average RT of the preceding two trials. Together with the RT, the difference (d = average RT - actual RT) was displayed with additional praise varying according to the magnitude of differences (0 msec < d < 21 msec: “good”; 20 msec < d < 41 msec: “very good”; d > 40 msec: “excellent”). Ss were not informed with respect to the relation of phrases to differences. This US procedure has repeatedly been shown to be a reliable US for differential conditioning (Lipp & Vaitl, 1988, 1990).
Physiological data were stored on an IBM-compatible AT (Creussen AT) which also managed trial sequences and intertrial intervals. Skin conductance and respiration were recorded with 10 Hz. Stimulus timing and RT registration was controlled by a microcomputer (Commodore C64) which also produced tones and graphics.
Procedure
Ss were seated in a reclining chair in a separate room. White noise (55 dB) was delivered into the subject room throughout the experiment in order to mask sounds from outdoors and from the adjacent control room. Prior to the experiment, all Ss were instructed in the same manner. They were informed that the purpose of the experiment was to record physiological responses elicited by simple stimuli like graphics or lights and by a RT task. After attachment of the recording devices, the tone stimulus was presented to familiarize the Ss with the imperative stimulus. Ss were instructed to press the button underneath their left index fingers immediately after they heard the tone. They were asked to sit in a comfortable manner, to refrain from moving, and to breath regularly. After completion of the experiment, Ss were asked to fill out some questionnaires. Similar to Baruch et al. (1988b), the Launay and Slade (198 1) Hallucination Scale, the Schizotypal Personality Questionnaire (Claridge & Broks, 1984) and Eysenck’s Personality Questionnaire (Eysenck, Eysenck & Barrett, 1985) were administered to assess schizotypal traits.
Treatment during acquisition and extinction was the same for each group. During acquisition, termination of one CS (CS+) coincided with the onset of the US whereas the second CS (CS)

1006 OTTMAR V. LIPP and DIETER VAITL
was presented unpaired with the US. During extinction, no USs were presented. In both phases there were eight presentations each of CS+ and CS-, each lasting 8 sec. Half the Ss in each group received lights during acquisition and extinction whereas the remaining received graphics. The use of a particular colour/shape as CS+ was counterbalanced within groups. The first stimulus during acquisition was a CS+ for half of the Ss.
During preexposure each S received 20 presentations of each stimulus from one category (graphics or lights). Group DIFF Ss were preexposed to stimuli from one category (e.g. lights) whereas stimuli from the second category were displayed during acquisition and extinction (e.g. graphics). In group SAME, stimuli from the same category were presented throughout the entire experiment. Group SAME + S was treated the same way as group SAME, but an additional signal was displayed 10 set after the offset of the 40th preexposure trial. The signal stimulus consisted of a multiple display of the word “ATTENTION” on various locations of the TV screen, lasting for 4 sec. Within this period each graphic and each light was presented once for 1 sec. After a break (1 set), this stimulus complex was repeated.
Stimuli throughout the whole experiment were presented in a random sequence with the restriction that no more than 2 consecutive stimuli were the same. Intertrial intervals (CS offset to CS onset) varied at randomly ordered intervals of 15,20,25,30, and 35 sec. The intertrial interval following the last stimulus during preperiod was 35 set for each S.
Scoring and statistical analyses
Respiration curves were inspected to detect irregularities in breathing. Skin conductance responses which coincided with coughs, sneezing, or sighing were replaced by estimates for missing data, calculated by means of multiple regression analysis (BMDPAM). Overall, 85 out of 3456 responses were replaced (2.45%). Skin conductance responses which exceeded 0.05 PSiemens (pS) were scored in three latency windows: first interval responses began within l-4 set after CS onset, second interval responses began within 4-9 set following CS onset, and third interval responses began 9-13 set after CS onset (Prokasy & Kumpfer, 1973). Only first interval responses were calculated during the preperiod.
RT data were subjected to a two factorial ANOVA (Group x Trials). Although the RT data are reported, they were not expected to be affected by the latent inhibition manipulation. Skin conductance data were analysed by means of three factorial ANOVAs (Group x Conditioning x Trials) for each experimental period separately to determine the impact of the additional signal after CS-preexposure. The first factor (Group, i.e. manipulation of latent inhibition) was between-S whereas the remaining (Conditioning, i.e. responding to CS+ vs CS; Trials of preperiod, acquisition, and extinction, respectively) were within-S factors. Latent inhibition, that is differences in conditioning, is evidenced by a Group x Conditioning x Trial or a Group x Conditioning interaction during acquisition or extinction in such a design. Corrected degrees of freedom, adjusted in terms of the degree of asymmetry in the covariance matrices (Greenhouse & Geisser, 1959), were used in all analyses to test main effects and interactions involving repeated measures. Post hoc multiple comparisons of means were calculated by means of Tukey tests. Level of significance was set at 0.05.
RESULTS
Additional signals and latent inhibition
Preexposure. Preliminary analyses revealed that the factor stimulus category (light/graph) did not influence responding. Thus, this factor was omitted in all subsequent analyses. Groups differed with respect to the overall electrodermal responding displayed during preexposure phase [main effect group: F(2/45) = 8.52, MS, = 2.12, P c O.OOl]. Average responses in group DIFF (mean = 0.42 PS, SD = 0.56) reliably exceeded responses in group SAME (mean = 0.22 PLS, SD = 0.24) and group SAME + S (mean = 0.08 PS, SD = 0.06). The electrodermal orienting responses decreased with stimulus repetition in all groups [F(19/855) = 4.23, MS, = 0.25, P < O.OOl]. The course of habituation did not differ between groups.
Acquisition. Analyses of electrodermal first interval responses were performed only on trials 2-8. Trial 1 was excluded in order to avoid the influence of increased responding elicited by the stimulus
Latent inhibition and schizotypal traits 1007
Table I. Summary of primary analyses of electrodermal data durine acauisition
Effect df MS. F
Group (G) Conditioning (C) Trials (T) CXG CxT CxGxT
Group (G) Conditioning(C) Trials (T) CxG CxT CxGxT
‘P < 0.05; **p < 0.01.
Firsf interval response 2145 3.43 10.08” l/45 1.22 I 1.451. 61270 0.49 6.20” 2145 1.22 4.10’ 61270 0.36 2.38
121270 0.36 0.44 Second interval response
2145 0.70 5.88** l/45 0.10 I .65 71315 0.17 1.58 2145 0.10 5.18” 71315 0.16 1.64
14/315 0.16 I .08
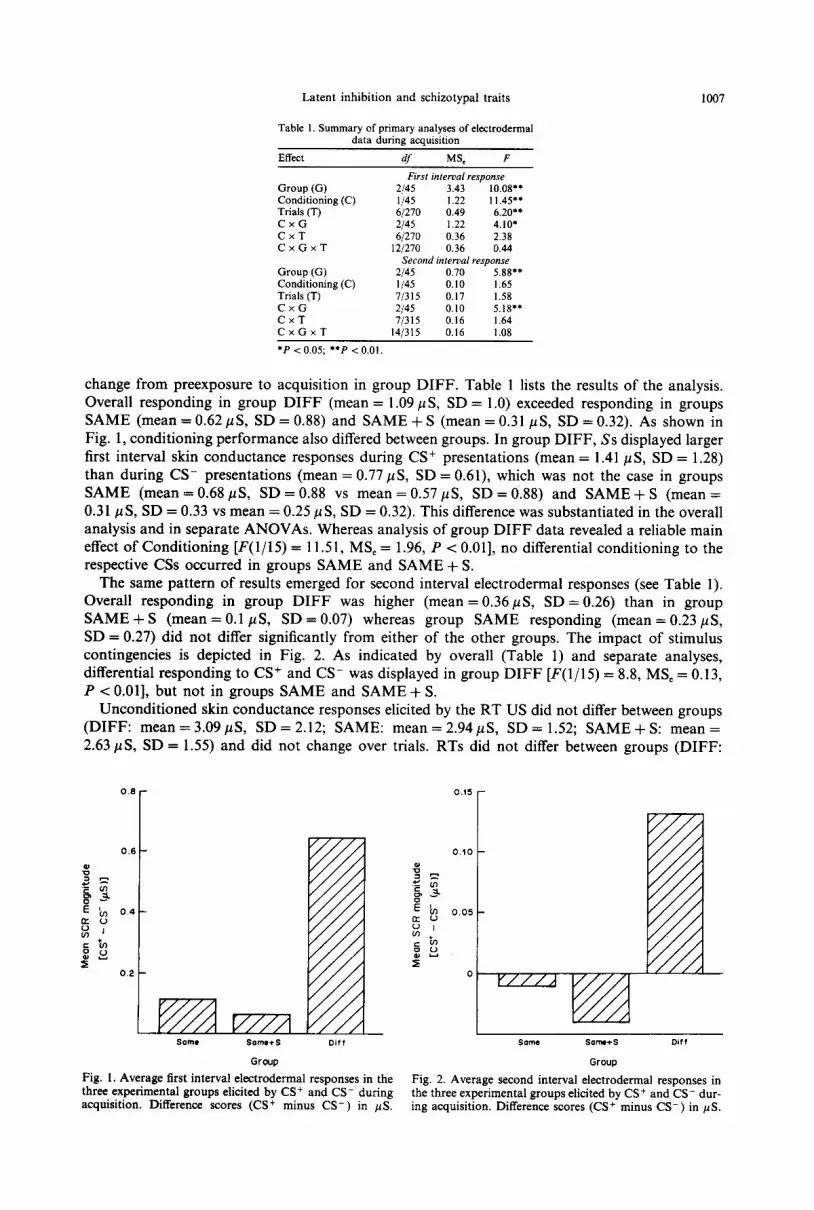
change from preexposure to acquisition in group DIFF. Table 1 lists the results of the analysis. Overall responding in group DIFF (mean = 1.09 pLs, SD = 1 .O) exceeded responding in groups SAME (mean = 0.62 pS, SD = 0.88) and SAME + S (mean = 0.31 PS, SD = 0.32). As shown in Fig. 1, conditioning performance also differed between groups. In group DIFF, Ss displayed larger first interval skin conductance responses during CS+ presentations (mean = 1.41 PLS, SD = 1.28) than during CS- presentations (mean = 0.77 pts, SD = 0.61), which was not the case in groups SAME (mean=0.68@, SD=0.88 vs mean=OS7pS, SD =0.88) and SAME+ S (mean = 0.31 ,u S, SD = 0.33 vs mean = 0.25 ~1 S, SD = 0.32). This difference was substantiated in the overall analysis and in separate ANOVAs. Whereas analysis of group DIFF data revealed a reliable main effect of Conditioning [F(1/15) = 11.51, MS, = 1.96, P < 0.011, no differential conditioning to the respective CSs occurred in groups SAME and SAME + S.
The same pattern of results emerged for second interval electrodermal responses (see Table 1). Overall responding in group DIFF was higher (mean = 0.36 PUS, SD = 0.26) than in group SAME + S (mean = 0.1 PS, SD = 0.07) whereas group SAME responding (mean = 0.23 PS, SD = 0.27) did not differ significantly from either of the other groups. The impact of stimulus contingencies is depicted in Fig. 2. As indicated by overall (Table 1) and separate analyses, differential responding to CS+ and CS- was displayed in group DIFF [F(1/15) = 8.8, MS, = 0.13, P < 0.011, but not in groups SAME and SAME + S.
Unconditioned skin conductance responses elicited by the RT US did not differ between groups (DIFF: mean = 3.09 PS, SD = 2.12; SAME: mean = 2.94 PS, SD = 1.52; SAME + S: mean = 2.63 PS, SD = 1.55) and did not change over trials. RTs did not differ between groups (DIFF:
Same Oiff Salt-W+!3
Group
Fig. 1. Average first interval electrodermal responses in the three experimental groups elicited by CS+ and CS- during
Fig. 2. Average second interval electrodermal responses in
acquisition. Difference scores (CS+ minus CS-) in pS. the three experimental groups elicited by CS+ and CS- dur- ing acquisition. Difference scores (CS+ minus CS-) in PS.

1008 OTTMAR V. LIPP and DIETER VAITL
Table 2. Summary of primary analyses of electrodemal data during acquisition
Effect df MS. F
Group (G) Conditioning (C) Trials (T) CxG CxT CxGxT
Group (G) Conditioning (C) Trials (T) CxG‘ CxT CxGxT
Group (G) Conditioning(C) Trials (T) CxG CxT CxGxT
First interual response 2145 1.96 4.43* l/45 0.64 7.10’ 71315 0.29 8.81” 2145 0.64 2.60 71315 0.23 1.80
14/315 0.23 1.31 Second interval response
2145 0.31 10.42” l/45 0.13 4.27’ 71315 0.10 1.70 2145 0.13 4.13’ 71315 0.11 1.74
14/315 0.11 1.21 Third interval response
2145 0.46 6.17** l/45 0.22 9.19” 71315 0.14 7.55” 2145 0.22 2.52 71315 0.15 1.01
141315 0.15 0.54
‘P < 0.05: **p < 0.01
mean = 344 msec, SD = 101; SAME: mean = 363 msec, SD = 57; SAME + S: mean = 361 msec, SD = 75). However, RTs became faster across trials [F(7/315) = 69.86, MS, = 0.01, P c O.OOl].
Extinction. Table 2 summarizes the results of the overall analyses of electrodermal responses during extinction. Group DIFF displayed larger first (mean = 0.54 pS, SD = 0.51) and third (mean = 0.27 ,uS, SD = 0.37) interval skin conductance responses than did group SAME + S (mean = 0.18 pS, SD = 0.12 and mean = 0.06 pS, SD = 0.05). Group SAME responses (mean = 0.36 pS, SD = 0.64 and mean = 0.18 pS, SD = 0.17) did not differ significantly from either of the other groups. Irrespective of the preexposure treatment, CS+ presentations elicited larger responses in both latency windows (mean = 0.44 pS, SD = 0.55 and mean = 0.22 pS, SD = 0.32) than did CS presentations (mean = 0.28 pS, SD = 0.4 and mean = 0.12 pS, SD = 0.1). Thus, there was no latent inhibition due to CS-preexposure evident in either first or third interval electrodermal responses. With respect to second interval electrodermal responding, however, CS-preexposure did affect conditioning performance during extinction (Table 2). As substantiated by separate ANOVAs, differential second interval responding during extinction was displayed in group DIFF only. Following preexposure to a different stimulus, larger second interval responses [F(1/15) = 5.22, MS, = 0.3, P < 0.051 were elicited during CS+ (mean = 0.34 pS, SD = 0.37) than during CS- presentations (mean = 0.18 pS, SD = 0.16) which was not the case in groups preexposed to the CSs. Overall, group DIFF exhibited larger responses (mean = 0.26 pS, SD = 0.29) than group SAME (mean = 0.11 pS, SD = 0.09) or group SAME + S (mean = 0.04 pS, SD = 0.02).
Latent inhibition as indexed by superior conditioning performance in group DIFF was evident in first interval electrodermal responses during acquisition as well as in second interval electrodermal responses during acquisition and extinction. However, latent inhibition was not disrupted by the additional stimulus displayed in group SAME + S which did not reliably differ in conditioning performance from group SAME.
Schizotypal traits and latent inhibition
The correlations between the psychoticism subscale (EPQ-P) of the EPQ and the hallucination scale (LSH), and the EPQ-P and the schizotypy subscale (STA) of the STQ were low (0.15 and 0.14, respectively), however, STA and LSH were significantly correlated (0.63). Each experimental group was divided into low and high scorers by median split with respect to the three measures of psychosis proneness to assess the relationship between personality traits and latent inhibition. Three factorial ANOVAs (Score x Conditioning x Trials) were calculated in each experimental group separately for those dependent measures which indexed latent inhibition (first interval electrodermal responses during acquisition, second interval electrodermal responses during acquisition and extinction). The additional stimulus employed in group SAME + S did not attenuate latent inhibition which indicates that group SAME + S can be treated as a replication
Latent inhibition and schizotypal traits 1009
0.6 c m STA low
0.6
Same Some+S
Group
Dlff
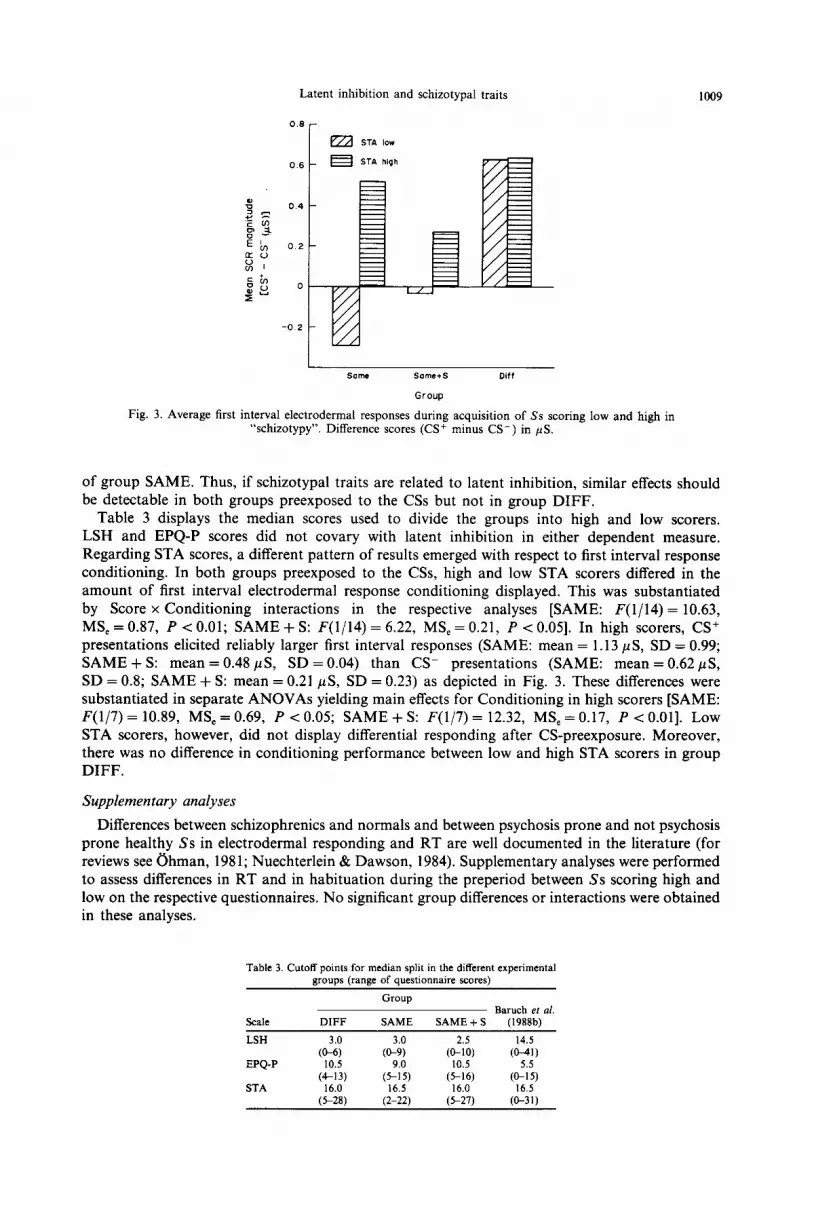
Fig. 3. Average first interval electrodermal responses during acquisition of Ss scoring low and high in “schizotypy”. Difference scores (CS+ minus CS-) in PCS.
of group SAME. Thus, if schizotypal traits are related to latent inhibition, similar effects should be detectable in both groups preexposed to the CSs but not in group DIFF.
Table 3 displays the median scores used to divide the groups into high and low scorers. LSH and EPQ-P scores did not covary with latent inhibition in either dependent measure. Regarding STA scores, a different pattern of results emerged with respect to first interval response conditioning. In both groups preexposed to the CSs, high and low STA scorers differed in the amount of first interval electrodermal response conditioning displayed. This was substantiated by Score x Conditioning interactions in the respective analyses [SAME: F(1/14) = 10.63, MS, = 0.87, P < 0.01; SAME + S: F(1/14) = 6.22, MS, = 0.21, P < 0.051. In high scorers, CS+ presentations elicited reliably larger first interval responses (SAME: mean = 1.13 pS, SD = 0.99; SAME + S: mean = 0.48 pS, SD = 0.04) than CS- presentations (SAME: mean = 0.62 pS, SD = 0.8; SAME + S: mean = 0.21 pS, SD = 0.23) as depicted in Fig. 3. These differences were substantiated in separate ANOVAs yielding main effects for Conditioning in high scorers [SAME: F(1/7) = 10.89, MS, = 0.69, P < 0.05; SAME + S: P(1/7) = 12.32, MS, = 0.17, P < 0.011. Low STA scorers, however, did not display differential responding after CS-preexposure. Moreover, there was no difference in conditioning performance between low and high STA scorers in group DTFF.
Supplementary analyses
Differences between schizophrenics and normals and between psychosis prone and not psychosis prone healthy Ss in electrodermal responding and RT are well documented in the literature (for reviews see ohman, 198 1; Nuechterlein & Dawson, 1984). Supplementary analyses were performed to assess differences in RT and in habituation during the preperiod between Ss scoring high and low on the respective questionnaires. No significant group differences or interactions were obtained in these analyses.
Table 3. Cutoff points for median split in the different experimental groups (range of questionnaire scores)
Scale DIFF
Group
SAME Baruch er al.
SAME + S (1988b)
LSH 3.0 3.0 2.5 14.5 (O-6) w-9) (O-10) (O-41)
EPQ-P 10.5 9.0 10.5 5.5 (413) (5-15) (s16) (O-15)
STA 16.0 16.5 16.0 16.5 (5-28) (2-22) (~27) (O-31)

1010 OTTMAR V. LIPP and DIETER VAITL
DISCUSSION
The present study had two aims. The first was to determine whether the retarding effect exerted by CS-preexposure on subsequent Pavlovian conditioning could be prevented by an additional stimulus signalling the shift from preexposure to acquisition (Lubow et al., 1975) and the second was to assess the relationship between schizotypal traits and latent inhibition in normals. Latent inhibition was detected in electrodermal responses. Differential responding in first and second interval responses during acquisition and in second interval responses during extinction was retarded following CS-preexposure.
It is evident that the additional stimulus presented in group SAME + S did not disrupt latent inhibition. Conditioning performance in groups SAME and SAME + S did not differ in either of the measures in which latent inhibition was obtained. These results are in contrast to findings obtained in animal research (Lantz, 1973; Rudy et al., 1977). Although the functional relation of habituation and latent inhibition is not completely understood at present (Swan & Pearce, 1988; Hall & Honey, 1989), the failure to disrupt latent inhibition may be due to the lack of dishabituation of the orienting response elicited by the CSs after presentation of the additional signal. Whereas the signal itself elicited a marked response (mean = 1.35 p S, SD = 1.12), the first interval responses elicited by the first CS during acquisition (mean = 0.07 ,uS, SD = 0.04) did not exceed responses elicited by the last CS during preperiod (mean = 0.03 pS, SD = 0.01). Further research is needed to specify conditions which prevent the occurrence of latent inhibition.
With respect to the relationship between schizotypal traits and latent inhibition, the conclusion is clear. EPQ-P scores did not covary with latent inhibition in the present study. This finding is deviant from Baruch et al’s (1988b) results and may be due to the use of different versions of the EPQ. Whereas Baruch et al. (1988b) used the first edition of the EPQ (Eysenck & Eysenck, 1975), a revised version (Eysenck et al., 1985), comprising a fairly new P-scale with more items, was employed in the present study. The discrepancy between the LSH median scores (14.5) reported by Baruch et al. (1988b) and the LSH median scores obtained in the present study (3, 3, 2.5) is even more obvious. As it seems, Baruch et al. (1988b) did not employ the final version of the Launey Slade Hallucination Scale (Launey & Slade, 1981) which has 12 items, but a longer, unpublished version. However, consistent with Baruch et d’s (1988b) findings, the LSH score did not covary with latent inhibition. Norma1 Ss differing in schizotypy as measured by the STA-scale (Claridge & Broks, 1984) clearly displayed differences in the extent of latent inhibition in Pavlovian conditioning. Whereas low scorers displayed latent inhibition after CS-preexposure, this was not the case in high scorers as was shown in two separate groups. The present data can be viewed as a multi-method validation of the results obtained by Baruch et al. (1988b).
In recent theories of associative learning, latent inhibition is viewed as resulting from a reduction in attention to the preexposed stimulus with a consequent loss of associability (for a review see Lubow, 1989). As the present data demonstrate, stimulus associability is not reduced after preexposure in Ss scoring high on STA. Taking into account the relationship between habituation and latent inhibition mentioned above, and the fact that high and low scorers did not differ in the speed of habituation, it is tempting to speculate that two processes take place during extended preexposure: (a) habituation which is completed within a few trials and is not context specific (Churchill, Remington & Siddle, 1987; Hall & Honey, 1989), and (b) learning that the preexposed stimulus is irrelevant and thus can be ignored. The latter process is context specific (Hall & Honey, 1989) and may require more trials to be completed. Habituation seems to be a necessary (Swan & Pearce, 1988), but not sufficient prerequisite to establish latent inhibition. High STA scorers did not display latent inhibition despite the fact that they habituated to the preexposed stimuli in the same manner as did the low scorers. Learning that a stimulus is irrelevant and can be ignored seems to be impaired in high STA Ss. The same seems to hold for acute schizophrenics (Baruch et al., 1988a). The notion of differences in stimulus processing between Ss scoring high and low in schizotypy is supported by studies of the negative priming effect (Beech & Claridge, 1987; Beech, Baylis, Smithson & Claridge, 1989). In the negative priming paradigm, which was developed to investigate selective attention, high STA scorers displayed less inhibitory priming by an irrelevant object than low scorers. Whereas the RTs to a test stimulus were prolonged in low scorers after presentation of a negative priming stimulus, this was not the case in high scorers. Further research

Latent inhibition and schizotypal traits 1011
is needed to clarify whether both phenomena (reduced latent inhibition and negative priming) are due to the same mode of stimulus processing.
In summary, the present data, together with findings from preliminary studies with clinical samples (Baruch et al., 1988a; Beech, Powell, McWilliam & Claridge, 1989) support the conclusion that the application of procedures like latent inhibition, with their unique feature of indexing presumed deviances in certain subgroups by increased performance, is a promising approach for research in personality and psychopathology.
Acknowledgements-We wish to thank Sabine Hirth, Susanne Rohman and Christiane Gerhardt for assistance in the data collection, and Rudolf Stark for providing software for acquisition of physiological data. Thanks are due to Robert E. Lubow and David Siddle for their critical comments on earlier drafts of this paper.
REFERENCES
Baruch, I., Hemsley, D. B. & Gray, J. A. (1988a). Differential performance of acute and chronic schizophrenics in a latent inhibition task. Journal of Nervous and Mental Disease, 176, 5988606.
Baruch, I., Hemsley, D. B. & Gray, J. A. (1988b). Latent inhibition and “Psychotic proneness” in normal subjects. Personality and Individual Differences, 9, 171-783.
Beech, A. & Claridge, G. (1987). Individual differences in negative priming: Relations with schizotypal personality traits. British Journal of Psychology, 78, 349-356.
Beech, A., Baylis, G. C., Smithson, P. & Claridge, G. (1989). Individual differences in schizotypy as reflected in measures of cognitive inhibition. British Journal of Clinical Psychology, 28, 117-129.
Beech, A., Powell, T., McWilliam, J. & Claridge, G. (1989). Evidence of reduced “cognitive inhibition” in schizophrenia. British Journal of Clinical Psychology, 28, 109-l 16.
Bjorkstrand, P.-A. (1990). Effects of conditioned stimulus pre-exposure on human electrodermal conditioning to fear-relevant and fear-irrelevant stimuli. Biological Psychology, 30, 35-50.
Booth, M. L., Siddle, D. A. T. & Bond, N. W. (1989). Effects of conditioned stimulus fear-relevance and preexposure on expectancy and electrodermal measures of human Pavlovian conditioning. Psychophysiology, 26, 28 l-29 1,
Cannon, D. S., Best, M. R., Batson, J. D. & Feldman, M. (1983). Taste familiarity and apomorphine-induced taste aversion in humans. Behavior Research and Therapy, 21, 669-673.
Churchill, M., Remington, B. & Siddle, D. A. T. (1987). The effect of context on long-term habituation of the orienting response in humans. Quarterly Journal of Experimental Psychology, 39B, 315-338.
Claridge, G. & Broks, P. (1984). Schizotypy and hemisphere function I: Theoretical considerations and the measurement of schizotypy. Personality and Individual Differences, 5, 633-648.
Eysenck, H. J. & Eysenck, S. B. G. (1975). Manual of the Eysenck Personality Questionnaire. London: Hodder & Stoughton. Eysenck, S. B. G., Eysenck, H. J. & Barrett, P. (1985). A revised version of the psychoticism scale. Personality and Individual
DtFerences, 6, 2 l-29. Ginton, A., Urea, G. & Lubow, R. E. (1975). The effect of preexposure to a non-attended stimulus on subsequent learning:
Latent inhibition in adults. Bulletin of the Psychonomic Society, 5, 5-8. Greenhouse, S. W. & Geisser, S. (1959). On methods in the analysis of profile data. Psychometrika, 24, 95-l 12. Hall, G. & Honey, R. C. (1989). Contextual effects in conditioning, latent inhibition and habituation: Associative and
retrieval functions of contextual cues. Journal of Experimental Psychology: Animal Behavior Processes, 15, 232-241. Lantz, A. E. (1973). Effect of number of trials, interstimulus interval, and dishabituation durine CS habituation on
subsequent.conditioning in a CER paradigm. Animal Learning and Behavior, I, 213-271. - Launey, G. & Slade, P. (1981). The measurement of hallucinatory predisposition in male and female prisoners.
Personality and Individual Dtyerences, 2, 221-234. Lipp, 0. V. & Vaitl, D. (1988). Reaction time task as unconditional stimulus: On conditioning skin conductance responses
and heart rate using a nonaversive unconditional stimulus. Pavlovian Journal of Biological Science, 23, 165-172. Lipp, 0. V. & Vaitl, D. (1990). Reaction time task as unconditional stimulus: Comparing aversive and nonaversive
unconditional stimuli. Pavlovian Journal of Biological Science, 25, 77-83. Lipp, 0. V., Siddle, D. A. T. & Vaitl, D. (1992). Latent inhibition in humans: Single cue conditioning revisited.
Journal of Experimental Psychology: Animal Behavior Processes 18, 115-126. Lubow, R. E. (1973). Latent inhibition. Psychological Bulletin, 79, 398-407. Lubow, R. E. (1989). Latent inhibition and conditioned attention theory. Cambridge: Cambridge University Press, Lubow, R. E., Alek, M. & Arzy, J. (1975). Behavioral decrement following stimulus preexposure: Effects of number of
preexposures, presence of a second stimulus, and interstimulus interval in children and adults. Journal of Experimental Psychology: Animal Behavior Processes, 104, 178-188.
Nuechterlein, K. H. & Dawson, M. E. (1984). Information processing and attentional functioning in the developmental course of schizophrenic disorders. Schizophrenia Bulletin, 16, 160-203.
ohman, A. (1981). Electrodermal activity and vulnerability to schizophrenia: A review. Biological Psychology, 12, 87-145. Prokasy, W. F. & Kumpfer, K. L. (1973). Classical conditioning. In Prokasy, W. F. & Raskin, D. C. (Eds), Electrodermal
activity in psychological research @p. 157-202). New York: Academic Press, Rudy, J. Q., Rosenberg, L. & Sandell, J. H. (1977). Disruption of a taste familiaritv effect bv a novel exterocentive
stimulation. Journal-of Experimental Psychoiogy:‘AnimalBehavior Processes, 3, 26-36. . Siddle, D. A. T. & Remington, B. (1987). Latent inhibition and human Pavlovian conditionine: Research and relevance.
In Davey, G. (Ed.), CGgnitive processes and Pavlovian conditioning in humans (pp. 115-14:). Chichester: John Wiley & Sons.
Siddle, D. A. T., Remington, B. & Churchill, M. (1985). Effects of conditioned stimulus preexposure on human electrodermal conditioning. Biological Psychology, 20, 113-127.

1012 OTTMAR V. LIPP and DIETER VAITL
Silver, A. I. (1973). Effects of prior CS presentations on classical conditioning of the skin conductance response. Psychophysiology, IO, 583-588.
Surwit, R. S. & Poser, E. G. (1974). Latent inhibition in the conditioned electrodermal response. Journal of Comparutiue and Physiological Psychology, 86, 543-548.
Swan, J. A. & Pearce, J. M. (1988). The orienting response as an index of stimulus associability in rats. Journal of Experimental Psychology: Animal Behavior Processes, 14, 292-301.
Zeiner, A. R. (1970). Orienting response and discrimination conditioning. Physiology and Behavior, 5, 641-646.

















![Striatal amphetamine-induced dopamine release in patients with schizotypal personality disorder studied with single photon emission computed tomography and [123I]iodobenzamide](https://img.pdfslide.net/doc/110x75/635eb6ef2dc9f5f89b06e3e1/striatal-amphetamine-induced-dopamine-release-in-patients-with-schizotypal-personality-1701677425.jpg)











