Embed Size (px)
Citation preview

Management and Conservation Article
Lesser Scaup Breeding Probability and Female Survival onthe Yukon Flats, Alaska
KATE H. MARTIN,1,2 University of Alaska Fairbanks, 211 Irving I Building, Fairbanks, AK 99775, USA
MARK S. LINDBERG, University of Alaska Fairbanks, 311 Irving I Building, Fairbanks, AK 99775, USA
JOEL A. SCHMUTZ, United States Geological Survey, Alaska Science Center, 1011 E Tudor Road, Anchorage, AK 99503, USA
MARK R. BERTRAM, United States Fish and Wildlife Service, Yukon Flats National Wildlife Refuge, 101 12th Avenue, Room 264, Fairbanks, AK99701, USA
ABSTRACT Information on the ecology of waterfowl breeding in the boreal forest is lacking, despite the boreal region’s importance to
continental waterfowl populations and to duck species that are currently declining, such as lesser scaup (Aythya affinis). We estimated breeding
probability and breeding season survival of female lesser scaup on the Yukon Flats National Wildlife Refuge, Alaska, USA, in 2005 and 2006.
We captured and marked 93 lesser scaup with radiotransmitters during prelaying and nesting periods. Although all marked lesser scaup females
were paired throughout prelaying and incubation periods, we estimated breeding probability over both years as 0.12 (SE 5 0.05, n 5 67) using
telemetry. Proportion of lesser scaup females undergoing rapid follicle growth at capture in 2006 was 0.46 (SE 5 0.11, n 5 37), based on
concentration of yolk precursors in blood plasma. By combining methods based on telemetry, yolk precursors, and postovulatory follicles, we
estimated maximum breeding probability as 0.68 (SE 5 0.08, n 5 37) in 2006. Notably, breeding probability was positively related to female
body mass. Survival of female lesser scaup during the nesting and brood-rearing periods was 0.92 (SE 5 0.05) in 2005 and 0.86 (SE 5 0.08) in
2006. Our results suggest that breeding probability is lower than expected for lesser scaup. In addition, the implicit assumption of continental
duck-monitoring programs that all paired females attempt to breed should be reevaluated. Recruitment estimates based on annual breeding-pair
surveys may overestimate productivity of scaup pairs in the boreal forest. (JOURNAL OF WILDLIFE MANAGEMENT 73(6):914–923;
2009)
DOI: 10.2193/2007-587
KEY WORDS Alaska, Aythya affinis, boreal forest, breeding probability, female survival, lesser scaup, Yukon Flats.
Extensive research has provided managers with informationon population dynamics of many waterfowl species nestingin the prairie and parkland regions of North America (e.g.,Johnson et al. 1992, Rohwer 1992, Beauchamp et al. 1996),but data on ducks breeding in the boreal-forest region arelacking even though the region is used by .34% of thecontinental waterfowl population (Wilkins et al. 2006).Breeding ecology of boreal ducks is poorly described withonly a few studies of nest survival (Grand 1995, Fournierand Hines 1998, Safine 2005, Walker et al. 2005, Corcoranet al. 2007) and female survival (Brook and Clark 2005).Furthermore, the boreal forest is the most critical breedingarea for some species that are currently declining, such aslesser scaup (Aythya affinis; Austin et al. 2000, Afton andAnderson 2001). In 2008, the combined populations oflesser and greater scaup were .40% below the average of thecontinental population from 1970 to 1979 (Zimpfer et al.2008). Seventy percent of lesser scaup breed in the westernboreal forest of Alaska (USA) and Canada, which is wherescaup populations have seen the steepest declines (Bellrose1980, Afton and Anderson 2001).
Waterfowl management in North America is largely basedon spring surveys of breeding duck populations, which haveseveral untested assumptions, including 1) all observedpaired females attempt to nest, and 2) the proportion ofnon-breeders (unpaired F) in a population is the same acrossall regions of the continent (Cowardin and Blohm 1992,
Nichols et al. 1995). However, for many species, theproportion of non-breeding females in the population andhow these non-breeders are distributed across the continentis unknown. Because regional differences in breeding seasonlength, nesting habitat, food sources, and predator commu-nities may influence duck production rates (Johnson et al.1992), it is likely that breeding probability differs amongspecies and across the landscape. Information on thedifferences between vital rates in the boreal forest and otherregions could improve management of the continental scauppopulation (Koons et al. 2006).
Population models predict that changes in female survivalwill affect scaup population growth more than proportionalchanges in productivity (Flint et al. 2006, Koons et al.2006); however, these predictions are based on limited datafor seasonal patterns in mortality and other populationparameters (e.g., breeding probability), particularly for lesserscaup in the boreal forest. Because a substantial portion offemale mortality in other duck species occurs during thebreeding season, the breeding portion of the annual cyclecould be more crucial to population dynamics than theremainder of the year (Sargeant and Raveling 1992,Hoekman et al. 2002).
We predicted that most female scaup migrating to theboreal forest would attempt to breed, resulting in low femalesurvival during the breeding season due to increased risk ofpredation during incubation and brood rearing (Sargeantand Raveling 1992, Walker and Lindberg 2005); thus, ourinitial focus was female breeding-season survival. However,data collected in our first field season led us to expand our
1 E-mail: [email protected] Present address: Oregon Natural Desert Association, 201 SEFairview Street, Prineville, OR 97754, USA
914 The Journal of Wildlife Management N 73(6)

objectives to include breeding probability, defined as theprobability that a female will lay
L
1 egg during thebreeding season. Non-breeding or delayed breeding hasbeen documented in tribe Aythyini, in species such ascanvasbacks (Aythya valisineria; Anderson et al. 2001),tufted ducks (A. fuligula; Blums et al. 2002), and redheads(A. americana; Arnold et al. 2002). Non-breeding has alsobeen documented for lesser (Afton 1984, Esler et al. 2001)and greater scaup (Gorman et al. 2008), although rates ofnon-breeding are not quantified for most species of ducks,especially in the boreal-forest region. Thus, our objectiveswere to examine factors affecting breeding probability andfemale breeding-season survival of lesser scaup on theYukon Flats in interior Alaska.
STUDY AREA
Our study area was located on the Western Yukon Flats ineastern interior Alaska. The Yukon Flats was encompassedby the 3.3-million-ha Yukon Flats National Wildlife Refugeand comprised the largest interior wetland basin in Alaska(Heglund 1992). Bisected by the Yukon River, .48% of thelandscape was classified as wetland and riparian areas, withas many as 40,000 lakes occurring on the refuge (UnitedStates Fish and Wildlife Service [USFWS] 1987). TheYukon Flats supported about 800,000 duck pairs annually,the second-largest concentration of breeding ducks inAlaska (USFWS 1987), and was regarded as a principalarea of production in the North American WaterfowlManagement Plan (USFWS 1986). Scaup were the mostabundant waterfowl species in this area, with .100,000pairs surveyed/year (Conant and Groves 2005). Waterfowlnested in forested areas and wet graminoid meadows(Heglund 1992), but the region included a varied landscapethat was heavily influenced by frequent fire disturbance andflooding (USFWS 1987). The Yukon Flats had a conti-nental subarctic climate characterized by great seasonalextremes of temperature and daylight including summertemperatures that exceed 38u C (USFWS 1987).
Our study site was located on the Long Lake wetlandcomplex (66u209N, 147u589W) approximately 25 km westof the village of Beaver, Alaska (Fig. 1). This site wasrepresentative of wetland habitats in the region and includedlakes surrounded by emergent vegetation, wet meadows,black spruce bogs (Picea mariana), and mixed-forest uplandscharacterized by stands of aspen (Populus tremuloides), paperbirch (Betula papyrifera), and white spruce (Picea glauca).
METHODS
Field Data and Lab MethodsWe captured females on breeding grounds using decoy trapsbaited with female scaup, northern shovelers (Anas clypeata),and American wigeon (A. americana; Anderson et al. 1980,Sharp and Lokemoen 1987). We focused on scaup but alsocaptured and collected data on some shoveler and wigeon(see Martin 2007). We used 10–14 decoy traps throughoutthe study area for 24 hours per day from 20 May to 10 June2005 and 12 May to 10 June 2006. We attached a USFWSleg band, coded nasal disks, and 9-g transmitter equipped
with a mortality switch (Advanced Telemetry Systems,Isanti, MN) to each captured female. We attachedtransmitters using a prong-and-glue attachment method(Pietz et al. 1995). We weighed each female and measuredwing chord, culmen, tarsus, and head length. We deter-mined age in 2005 by comparing eye color with charts fromTrauger (1974). In 2006, we compared eye color withMunsell soil color charts (GretagMacbeth, New Windsor,NY; Trauger 1974) to examine eye color with moreaccuracy, which we then compared with age determinationsin Trauger (1974). In 2006, to examine the proportion offemales undergoing rapid follicle growth, we collected bloodfrom the brachial vein and placed it into heparinizedvacutainers using blood collection kits (23 gauge needleswith 30.5-cm tubing; BD Vacutainer, Franklin Lakes, NJ).We kept blood in coolers until centrifuged, usually within2 hours of collection. We extracted plasma from blood andfroze it until lab analysis. The University of Alaska-Fairbanks Institutional Animal Care and Use Committeereviewed and approved capture and handling methods(Protocol no. 04-13).
After we radiomarked a female we located it once or twiceper day until 15 July to determine its fate (alive or dead) andbreeding status (i.e., nesting, renesting, or apparently notnesting) using ground telemetry. During this intensivesearch period we also radiotracked females 1–2 times perweek, using a floatplane (Husky A-1B, Aviat Aircraft Inc.,Afton, WY; Bush Hawk XP, Found Aircraft, Inc., ParrySound, ON, Canada) equipped with a telemetry receiver, toensure that we located all females on the study area. If wesuspected that a female was on a nest during an aerialtelemetry session, we confirmed its location by ground
Figure 1. Long Lake complex on the Yukon Flats National WildlifeRefuge, Alaska, USA. Lakes where lesser scaup females were captured andmonitored in 2005–2006 are in black, with the inset showing the location ofthe study area in Alaska.
Martin et al. N Demography of Lesser Scaup 915

telemetry as soon as possible. After 15 July, we locatedfemales every 3–5 days using both ground and aerialtelemetry. When we detected a mortality signal and founda carcass at the site, we collected data (e.g., hair, predatortrack, disturbance level) to determine apparent cause ofmortality (Sargeant et al. 1998). In addition, after 15 July2006, we shot 10 marked female scaup that had notapparently nested. We froze carcasses immediately aftercollection and later extracted the ovaries, which we stored in10% formalin.
We located nests of unmarked females by dragging ropesthrough meadow habitats and used nest traps to capturefemales between 10 days and 25 days of incubation tosupplement the sample size for estimating adult survivalprobability (Weller 1957, Petrula 1994). We marked nestingfemales with radiotransmitters and processed them in thesame way as decoy-trapped females, except we lightlyanesthetized the bird with methoxyflurane (Metofane;Schering-Plough Animal Health Corporation, Union, NJ)before placing it back on the nest to reduce the chance ofnest abandonment (Rotella and Ratti 1990).
We analyzed blood plasma from 2006 females for evidenceof yolk precursors (YP), including vitilogenin (VTG) andvery low density lipoprotein (VLDL) that indicate rapidfollicle growth (RFG; Challenger et al. 2001). Yolkprecursor concentration in the blood increases to detectablelevels when the first follicle begins RFG and remains highthroughout the laying period (Gorman 2005). Plasmasamples were assayed for VTG and total VLDL usingdiagnostic kits for vitellogenic zinc and total triglycerides atSimon Fraser University following the methods of Mitchelland Carlisle (1991). We classified females undergoing RFGas those with VLDL levels
L
5.3 mmol/L and VTG levels
L
1.4 ug/mL based on prior work with greater scaup(Gorman 2005). In addition, we macroscopically examinedthe ovaries of shot females for postovulatory follicles, usingvisual cues as described in Lindstrom et al. (2006).
Statistical AnalysisWe used general linear models in Program MARK (Whiteand Burnham 1999) to generate maximum-likelihoodestimates and associated sampling variances of breedingprobability and breeding season survival. We used the logitlink function for all models to accommodate the binomialnature of survival and breeding probability data (Ramseyand Schafer 1997). We began by developing a set ofbiologically relevant models prior to each analysis and usedAkaike’s Information Criterion (AICc) to select the best-approximating model (Burnham and Anderson 1998).There was likely some overdispersion created by assumptionviolations; however, we chose to use models that may beoverparameterized instead of using biased estimates ofoverdispersion (Dinsmore et al. 2002). We estimatedsampling variance of products using the delta method(Seber 1982).
We estimated 2 parameters related to breeding effort:breeding probability using telemetry data in 2005 and 2006and probability that a female was undergoing RFG at the
time of capture (RFG probability), as determined from YPanalysis in 2006. For both analyses, we used the known-fatemodule in Program MARK by treating breeding and RFGprobability as a single-occasion trial (i.e., 11 5 breeding),which allows probabilities to be modeled as a function ofmultiple covariates, such as time, age, and morphologicalcharacteristics (Dinsmore et al. 2002). We hypothesizedthat older females and those with larger body reserves wouldbe more likely to nest (Johnson et al. 1992). In addition, wehypothesized that annual variation in food resources orother environmental conditions could lead to annualvariation in breeding probability. We considered both 2-way additive models with age, body mass, and year. Weincluded additive models that controlled for date of capturebecause body mass probably increases after arrival to thebreeding area (Esler et al. 2001). To control for potentialeffects of variation in structural size (Alisauskas and Ankney1987, Sedinger et al. 1997) we performed a principalcomponents analysis (PROC FACTOR; SAS Instituteversion 9.1, SAS Institute Inc., Cary, NC) on 4 structuralmeasurements: tarsus, culmen, wing chord, and head length.We used the first principal component (PC1), whichaccounted for 43% of the variation in the morphologicalmeasurements in additive models with body mass.
To define uncertainty associated with using telemetry toestimate breeding probability, McPherson et al. (2003)developed a method to estimate the number of missednesting attempts. Because of a limited sample size, we used asimilar ad hoc equation based on the Mayfield method(Miller and Johnson 1978, Johnson and Shaffer 1990) toestimate the number of nesting attempts we would expectfrom radiomarked females (NMJ) in both 2005 and 2006:
NMJ ~xJ
PJ
i~1SJ
where J is the date the oldest nest was found, xJ is thenumber of nests that survived to day J, and SJ is the dailysurvival rate (DSR) for each day up to day J. We estimatedDSR from nests of marked and unmarked scaup, whichwere monitored as part of an ongoing study of nest fate(Martin 2007). Nest age ( J ) included the laying period andwe calculated it as the sum of clutch size (assuming 1 egg/day) and the number of days incubated when we found thenest. Using the delta method (Seber 1982), we estimatedsampling variance of NMJ as
Var NMJ
� �~
xJ
� �2
Var S1S2 � � � SJ
� �
Because there was uncertainty about classification offemales in nonnesting and non-RFG groups, we calculatedthe maximum breeding probability of scaup in 2006 bycombining all methods: telemetry, YP, and examination ofpostovulatory follicles (POF) of 10 collected females. Wecombined data in 3 increments: 1) combining telemetry dataand YP data, 2) combining telemetry and YP data with POFdata, and 3) combining telemetry, YP data, and POF data
916 The Journal of Wildlife Management N 73(6)

and inflating the number of RFG females to compensate foran unknown amount of detection error due to early captureprior to the initiation of RFG. We combined data bydetermining for each data source the classification of anindividual. For example, telemetry data may not indicatethat an individual was a breeder, but examination of YP mayindicate that the individual initiated RFG. Breedingprobability increased as we added each data source becausewe had a higher probability of detecting breeding from .1source; however, we treated detection of breeding as aprobabilistic event and determined misclassification ratesbased on combined data sources (i.e., double-sampling;Bond 2005). Total proportion of misclassified breeders isthe product of the misclassification rates from YP andtelemetry methods, which we then added to the totalproportion of females classified as breeders with eithermethod to attain the overall estimate of breeding probabil-ity. For example, if 5 of 20 individuals were classified asbreeders based on telemetry, 10 of the 20 individuals wereclassified as breeders based on YP data, and 3 individualswere classified as breeders based on both data sources, theproportion of females classified as breeder by either methodwould be 0.60 (12/20). If 2 of the birds classified as breedersbased on telemetry were classified as non-breeders with YP,and 5 classified as breeders with YP were classified as non-breeders with telemetry, the overall misclassification ratewould be 0.04 (2/20 3 5/20), and breeding probabilitywould be 0.64 (0.60 + 0.04). We could not calculate amisclassification error for POF data because we examinedonly 10 of 37 females for POFs. Thus, when we includedPOF data (second and third approaches), we incorporatedthe misclassification rate for telemetry and YP data only andsimply added individuals that were also classified as breedersbased on POF to the proportion of breeders. In the thirdapproach, we also assumed that all females captured on orbefore 31 May had the same RFG probability as femalescaptured after 31 May. 31 May was the latest date that afemale was classified as non-RFG but actually laid
L
1 eggas indicated by POF methods. For all analyses we assumedthat all females characterized as undergoing RFG laid
L
1egg.
To estimate female survival probability we used the nestsurvival module in Program MARK, which is a form ofknown-fate model that allows for irregular samplingintervals. For period-specific estimates of survival probabil-ity, we raised the daily survival probability by 82, which isthe number of days per year that we monitored survival,calculated from the first day of capture (15 May) to the lastday of monitoring (4 Aug). We considered the followingcovariates of female daily survival: year, temporal trendacross the breeding season, nesting behavior (i.e., nesting ornot nesting), and body mass at capture. We were notconfident in our estimates of female age based on eye colorand, therefore, did not include age as a variable in ouranalysis. We hypothesized that average female survivalprobability would differ by year because of variation in theenvironment (e.g., weather, food resources, predator abun-dance) and in proportion of females in the population that
attempt to nest (Rotella et al. 2003). In addition, femalesurvival probability could vary across the breeding season,because females may be at greater risk of mortality duringincubation or brood-rearing periods than during theprelaying period. Nesting behavior may put females athigher risk of predation than other activities (Sargeant andRaveling 1992). Thus, we considered nesters and nonnestersas separate groups, where we defined nesters as those birdswe observed on nests using radiotelemetry. Likewise, weincluded body mass as a covariate because females withlarger body mass are undoubtedly in better condition butparadoxically may have lower survival because they expendmore effort on reproduction (Rotella et al. 2003).
RESULTS
Breeding ProbabilityAll marked female scaup remained paired after capture andthroughout the nest-initiation period. In 2005, 7 out of 31radiomarked scaup nested (one nested twice), and in 2006, 4out of 38 radiomarked scaup nested. In both years one scaupleft the study area directly after capture and we censored itfrom the analysis of breeding probability.
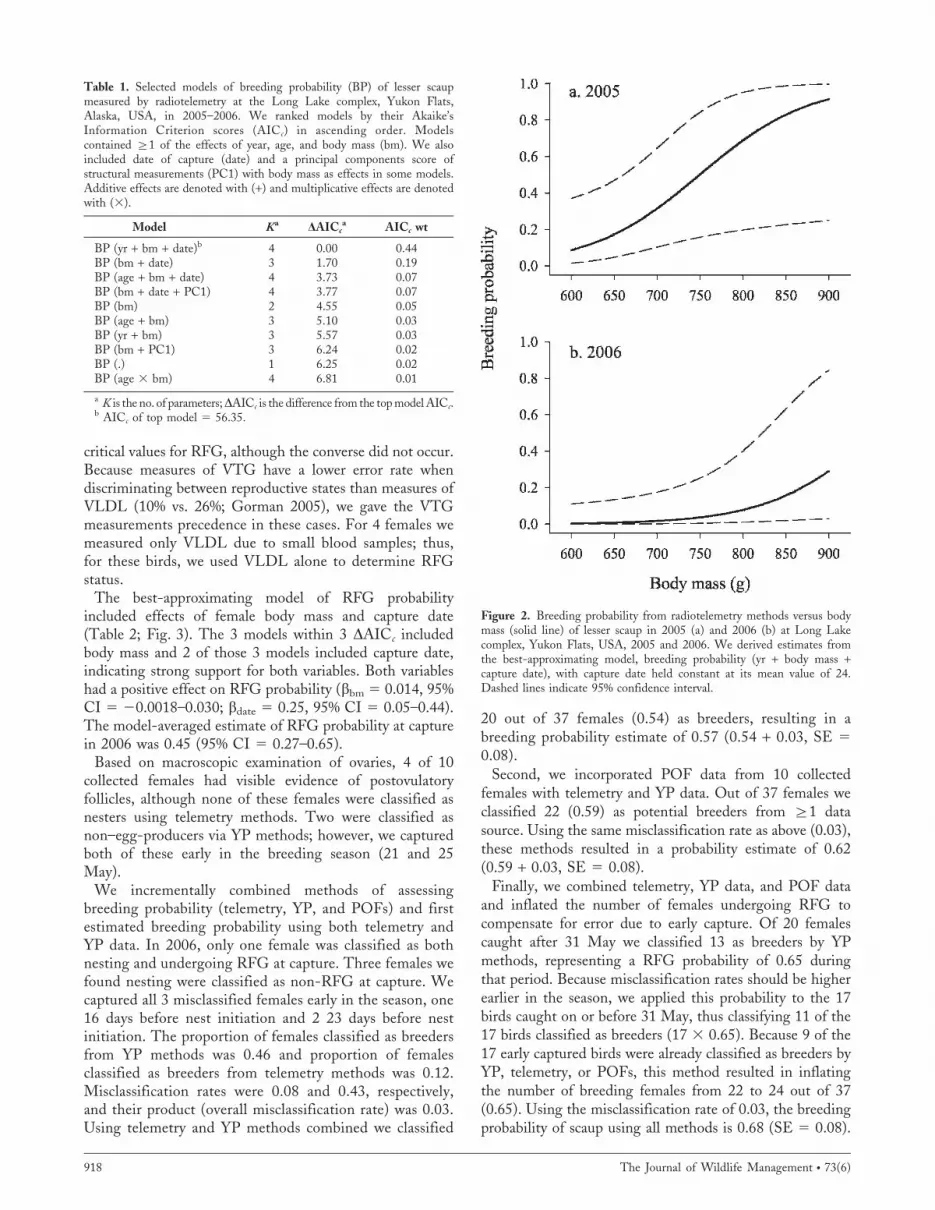
The best-approximating model suggested that yearlyvariation, female body mass, and capture date wereimportant predictors of breeding probability (Table 1;Fig. 2). All models within 4 DAICc included body massand capture date. Breeding probability was higher in 2005than 2006 (b2005 5 3.27, 95% CI 5 20.16–6.70; b2006 5
29.12, 95% CI 5 218.90–0.65) and there was evidence of apositive effect of body mass (bbm 5 0.016, 95% CI 5 0.001–0.031; Fig. 2) and a negative effect of capture date (bdate 5
20.25, 95% CI 5 20.45 to 20.05) on breeding probabilityin the top model. We chose to use a model-averagedestimate because of high model-selection uncertainty(Table 1). Based on telemetry methods, breeding probabilitywas 0.11 (95% CI 5 0.05–0.24).
Average age of telemetry nests at discovery was 8 days (SE5 1.1, n 5 12). In 2005, we found 8 nests from markedfemales and the oldest nest was 16 days old. The nestsurvival probability during this interval was 0.27 (SE 5
0.08). Out of 8 nests 3 survived to day 16 and one failed in atime interval that included day 16. Assuming that the fourthnest survived to day 16, the estimated number of nestsinitiated by marked lesser scaup was 15 (SE 5 39). If weassumed that the fourth nest did not survive to day 16, theestimated number of nests initiated was 11 (SE 5 52) in2005. In 2006, we found the oldest nest of a marked femalescaup at 13 days old, and the nest survival probability duringthis period was 0.21 (SE 5 0.07). We located 4 nests ofmarked females, one of which survived to 13 days. Based onthese data, we would expect that 5 scaup nests (SE 5 15)were initiated by marked females in 2006. These estimatessuggest that we found 0.60–0.75 of nests initiated bymarked scaup (12 out of either 16 or 20 nests).
Yolk precursor analysis indicated that 17 out of 37 lesserscaup were undergoing RFG when captured in 2006. Of thefemales categorized as RFG 7 had VLDL plasma levels,5.3 mmol/L but VTG plasma levels .1.4 ug/mL, the
Martin et al. N Demography of Lesser Scaup 917
critical values for RFG, although the converse did not occur.Because measures of VTG have a lower error rate whendiscriminating between reproductive states than measures ofVLDL (10% vs. 26%; Gorman 2005), we gave the VTGmeasurements precedence in these cases. For 4 females wemeasured only VLDL due to small blood samples; thus,for these birds, we used VLDL alone to determine RFGstatus.
The best-approximating model of RFG probabilityincluded effects of female body mass and capture date(Table 2; Fig. 3). The 3 models within 3 DAICc includedbody mass and 2 of those 3 models included capture date,indicating strong support for both variables. Both variableshad a positive effect on RFG probability (bbm 5 0.014, 95%CI 5 20.0018–0.030; bdate 5 0.25, 95% CI 5 0.05–0.44).The model-averaged estimate of RFG probability at capturein 2006 was 0.45 (95% CI 5 0.27–0.65).
Based on macroscopic examination of ovaries, 4 of 10collected females had visible evidence of postovulatoryfollicles, although none of these females were classified asnesters using telemetry methods. Two were classified asnon–egg-producers via YP methods; however, we capturedboth of these early in the breeding season (21 and 25May).
We incrementally combined methods of assessingbreeding probability (telemetry, YP, and POFs) and firstestimated breeding probability using both telemetry andYP data. In 2006, only one female was classified as bothnesting and undergoing RFG at capture. Three females wefound nesting were classified as non-RFG at capture. Wecaptured all 3 misclassified females early in the season, one16 days before nest initiation and 2 23 days before nestinitiation. The proportion of females classified as breedersfrom YP methods was 0.46 and proportion of femalesclassified as breeders from telemetry methods was 0.12.Misclassification rates were 0.08 and 0.43, respectively,and their product (overall misclassification rate) was 0.03.Using telemetry and YP methods combined we classified
20 out of 37 females (0.54) as breeders, resulting in abreeding probability estimate of 0.57 (0.54 + 0.03, SE 5
0.08).Second, we incorporated POF data from 10 collected
females with telemetry and YP data. Out of 37 females weclassified 22 (0.59) as potential breeders from
L
1 datasource. Using the same misclassification rate as above (0.03),these methods resulted in a probability estimate of 0.62(0.59 + 0.03, SE 5 0.08).
Finally, we combined telemetry, YP data, and POF dataand inflated the number of females undergoing RFG tocompensate for error due to early capture. Of 20 femalescaught after 31 May we classified 13 as breeders by YPmethods, representing a RFG probability of 0.65 duringthat period. Because misclassification rates should be higherearlier in the season, we applied this probability to the 17birds caught on or before 31 May, thus classifying 11 of the17 birds classified as breeders (17 3 0.65). Because 9 of the17 early captured birds were already classified as breeders byYP, telemetry, or POFs, this method resulted in inflatingthe number of breeding females from 22 to 24 out of 37(0.65). Using the misclassification rate of 0.03, the breedingprobability of scaup using all methods is 0.68 (SE 5 0.08).
Figure 2. Breeding probability from radiotelemetry methods versus bodymass (solid line) of lesser scaup in 2005 (a) and 2006 (b) at Long Lakecomplex, Yukon Flats, USA, 2005 and 2006. We derived estimates fromthe best-approximating model, breeding probability (yr + body mass +capture date), with capture date held constant at its mean value of 24.Dashed lines indicate 95% confidence interval.
Table 1. Selected models of breeding probability (BP) of lesser scaupmeasured by radiotelemetry at the Long Lake complex, Yukon Flats,Alaska, USA, in 2005–2006. We ranked models by their Akaike’sInformation Criterion scores (AICc) in ascending order. Modelscontained
L
1 of the effects of year, age, and body mass (bm). We alsoincluded date of capture (date) and a principal components score ofstructural measurements (PC1) with body mass as effects in some models.Additive effects are denoted with (+) and multiplicative effects are denotedwith (3).
Model Ka DAICca AICc wt
BP (yr + bm + date)b 4 0.00 0.44BP (bm + date) 3 1.70 0.19BP (age + bm + date) 4 3.73 0.07BP (bm + date + PC1) 4 3.77 0.07BP (bm) 2 4.55 0.05BP (age + bm) 3 5.10 0.03BP (yr + bm) 3 5.57 0.03BP (bm + PC1) 3 6.24 0.02BP (.) 1 6.25 0.02BP (age 3 bm) 4 6.81 0.01
a K is the no. of parameters;DAICc is the difference from the top model AICc.b AICc of top model 5 56.35.
918 The Journal of Wildlife Management N 73(6)

Female SurvivalIn addition to 69 females captured prebreeding we includedsurvival data from 24 scaup captured during nesting (14 in2005 and 10 in 2006). We right-censored data when femalesleft the study area for .14 days because we could notdetermine their fates during those intervals. There were 9mortalities during the study: 2 in 2005 and 7 in 2006. Causeof mortality for females was usually unknown but we found2 females partially consumed by predators in the vicinity oftheir nests.
The best-approximating model indicated that femalesurvival varied by year, and estimated female scaup survivalwas 0.92 (95% CI 5 0.86–0.98) in 2005 and 0.76 (95% CI5 0.61–0.92) in 2006. The second-best model indicated aneffect of nesting behavior on survival, where survival ofnonnesters was 0.86 (95% CI 5 0.68–1.04) and survival ofnesters was 0.55 (CI 5 0.10–1.07). Body mass and datewere not supported (Table 3).
DISCUSSION
Breeding ProbabilityWe used several methods to investigate breeding probabil-ity, which resulted in a range of estimates. However, allmethods indicated that breeding probability of scaup wassubstantially ,1.0 at our study area because even our mostliberal estimates indicated breeding probability was ,0.65.This suggests that the boreal-forest scaup population mayconsist of a larger portion of non-breeders than is currentlyassumed.
Our telemetry-based estimate of 0.11 breeding probabilitywas likely negatively biased. Nest-detection probability canbe low during the egg-laying period, when females spendless time on their nests than during incubation and dailysurvival rates are generally lower than for older nests (Klettand Johnson 1982, Afton and Paulus 1992). However, webelieve that the apparent breeding probability estimate wasnot driven by poor nest-detection probability because ourcalculations suggest that we found approximately 60–75% ofnests that had been initiated, similar to results for prairie-nesting mallards (McPherson et al. 2003).
Although we found a higher RFG probability thanbreeding probability, measures of RFG using YP areprobably still negatively biased. In previously studied duckspecies, levels of YP were only elevated up to 12–16 daysprior to laying (Gorman 2005, Safine 2005). Across the setof observed nests in our study area, initiation dates differedby .30 days for what we assumed to be first nest attempts.Despite timing our capture period to minimize timebetween capture and mean nest-initiation date of scaup,we may have captured some individuals on the breeding areaprior to initiation of RFG. Collections of prairie-nestingducks before and during nest initiation have also includedsome non-RFG birds (Alisauskas et al. 1990, Afton andAnkney 1991, Ankney and Alisauskas 1991), indicating thatnesting is either asynchronous or some portion of thepopulation is not breeding.
Conversely, the estimate of breeding probability usingcombined methods (0.68) is probably positively biased. Iftrue breeding probability was actually 0.68, then our nest-detection probability was only 0.16, which is unlikelyconsidering our methodology. Additionally, in combiningmethods we assumed that if a female initiated RFG, then
Table 2. Models of probability of undergoing rapid follicle growth(RFGP) by lesser scaup at Long Lake complex, Yukon Flats, Alaska,USA, in 2006. We ranked models by their Akaike’s Information Criterionscores (AICc) in ascending order. Models contained
L
1 of the effects of ageand body mass (wt). We also included date of capture (date) and a principalcomponents score of the structural measurements (PC1) with body mass(bm) as effects in some models. Additive effects are denoted with (+).
Model Ka DAICca AICc wt
RFGP (bm + date)b 3 0.00 0.37RFGP (bm + date + PC1) 4 0.04 0.36RFGP (age + bm + date) 4 2.32 0.11RFGP (bm + PC1) 3 2.58 0.10RFGP (age + bm + PC1) 4 5.10 0.03RFGP (bm) 2 5.90 0.02RFGP (age + bm) 3 8.26 0.01RFGP (.) 1 9.11 0.00RFGP (age) 2 11.16 0.00
a K is the no. of parameters;DAICc is the difference from the top model AICc.b AICc from top model 5 44.05.
Table 3. Models of daily survival rate (DSR) of female lesser scaup atLong Lake complex, Yukon Flats, Alaska, USA, in 2005–2006. We rankedmodels by their Akaike’s Information Criterion (AICc) scores in ascendingorder. Models contained one of the effects of year, season day (linear trendacross the season), body mass (bm), and whether the female was nesting ornot nesting (nest).
Model Ka DAICca AICc wt
DSR (yr)b 2 0.00 0.32DSR (nest) 2 0.46 0.26DSR (.) 1 0.67 0.23DSR (season day) 2 2.30 0.10DSR (bm) 2 2.52 0.09
a K is the no. of parameters;DAICc is the difference from the top model AICc.b AICc for top model 5 115.74.
Figure 3. Probability of undergoing rapid follicle growth (RFG) versusbody mass (solid line) of lesser scaup at Long Lake complex, Yukon Flats,USA, in 2006. We derived estimates from the best-approximating model,RFG probability (body mass + capture date), with capture date heldconstant at its mean value of 31. Dashed lines indicate 95%confidence interval.
Martin et al. N Demography of Lesser Scaup 919

she always laid L1 egg. However, follicular atresia andsubsequent reabsorption of nutrients from developedfollicles has been documented in arctic-nesting geese (Barry1962) and other birds (Gilbert et al. 1983, Thomas 1988).For species that occasionally forego breeding in a given year,females should continually assess environmental cues thataffect their chances of reproductive success and survivalbecause egg development and incubation have highenergetic costs and incubation incurs high predation risk(Afton and Paulus 1992, Alisauskas and Ankney 1992,Sargeant and Raveling 1992). If conditions becomeunfavorable (e.g., poor food availability, adverse weather,high predator abundance), the ability to abort eggdevelopment at any stage and retain the lipid and proteinin follicles would be advantageous to females (Barry 1962).Some of the inconsistency between breeding probabilityestimates using radiotelemetry and yolk precursor data couldbe explained by follicular atresia. Conversely, examiningPOFs from shot birds, instead of telemetry or YP methods,could eliminate the problem with inconsistent resultsbetween methods (Lindstrom et al. 2006). However, oursmall sample of marked females collected late in the nestingseason is probably biased because rapid follicle regressionmay obstruct identification.
Although we did not observe any negative effects ofcapture, handling, nasal marker, or transmitter attachmenton female ducks, we cannot discount a marker effect.Marker effects could induce follicular atresia in birdsundergoing RFG, or if females are captured prior to RFG,it may reduce the number of birds that begin RFG. To ourknowledge, there are no studies that directly measure theeffect of handling and transmitter attachment on breedingeffort of female ducks in a field setting because of thedifficulty of including a true control group. However, bothPaquette et al. (1997) and Rotella et al. (1993) comparedsutured back-mounted transmitters and implanted trans-mitters on mallards and found lower renesting probabilityand number of days devoted to nesting activity for femaleswith back-mounted transmitters but no difference in thenumber of females that initiated nests. In addition, nasalmarkers have been found to induce slight behavior changesin scaup (Brook and Clark 2002) and they reduce renestingprobability, but not breeding probability, in mallards(Howerter et al. 1997). Scaup clearly differ from mallardsin many aspects of their life history and morphology, so wewould expect markers to affect scaup differently. However,even if the methods we employed induced subtle effects onscaup breeding probability, it is unlikely that they couldinduce the high rates of non-breeding that we found (nearly0.90 with radiotelemetry methods).
There is evidence that other diving duck populationscontain non-breeders (Anderson et al. 2001, Arnold et al.2002, Blums et al. 2002). Although there are no availableestimates of scaup breeding probability in the boreal forest,breeding probability of lesser scaup in Manitoba, Canada,was between 0.72 and 0.98 over 4 years (Afton 1984), andnon-breeding has been observed in boreal-forest populationsof lesser scaup and in greater scaup on the Yukon Delta
(Esler et al. 2001, Gorman et al. 2008). Estimates ofbreeding probability for lesser scaup in the prairies arehigher than our estimates from the Long Lake complex in2005 and 2006, but it still suggests that a proportion offemale scaup do not breed each year. Furthermore, we wouldexpect estimates of breeding probability to be lower forducks in the boreal forest because they migrate farther thanprairie ducks and scaup are experiencing deterioratingconditions on spring staging areas (Anteau and Afton2004). Interestingly, all marked females in our study, eventhose that indicated no evidence of breeding usingtelemetry, YP, or POF data, remained paired after captureand throughout the nest-incubation period. Therefore,paired females may not necessarily be breeders andbreeding-pair surveys may overestimate the number ofbreeding scaup.
Anteau and Afton (2004) have asserted that foodlimitation on migratory routes has led to a reduction offemale body mass and lipid reserves in spring, which hasreduced reproductive success and contributed to the scauppopulation decline. Anteau and Afton (2004) found thatfemales migrating through the upper Midwest were about60 g lighter in 2000 and 2001 than females in the late1970s, when food resources were thought to be moreabundant. Similarly, mean body mass of female scaupcaptured by decoy trapping during the prelaying period in2005 and 2006 at the Long Lake complex was 700 g (n 5
71, SE 5 7.1), approximately 70 g less than mean bodymass of prenesting females shot in 1991 on the Yukon Flats(D. Esler, Simon Fraser University, unpublished data; x 5
768 g, n 5 34, SE 5 13.7). Our comparison of body massesdoes not span the same time frame as Anteau and Afton(2004), but the low body mass of captured females, as well astheir low breeding probability at the Long Lake complex,supports the portion of the spring condition hypothesis thatfemale body condition affects breeding probability. How-ever, in a comparison of collected scaup between 2003 and2004 and years prior to the scaup population decline (1968–1970), DeVink (2007) found that spring scaup body massdid not decline in another boreal-forest breeding site. Thus,it is possible that our decoy trapping provided a biasedsample toward birds in poorer condition (Greenwood et al.1986, Reinecke and Shaiffer 1988), though this bias indecoy trapping has not been tested formally.
Even if scaup are arriving in poorer body condition than inthe past, female scaup can quickly recover if food isabundant. Martin (2007) found that captive scaup cancompletely restore body mass losses of 65 g (11% of bodymass) in ,4 days, with no observed changes in bodycomposition. Further, rates at which modeled body mass isregained is dependent on quantity and quality of foodavailable to scaup. Because lesser scaup spend 3–4 weeks onthe breeding grounds before initiating a nest (Afton 1984),birds presumably have time to make up for any mass lost enroute if adequate food resources are available. Scaup dependin part on exogenous resources for the energy to supportreproduction (Esler et al. 2001, Gorman et al. 2008); thus,food availability could also act as a cue in activating the
920 The Journal of Wildlife Management N 73(6)

physiological process leading to reproduction (Barboza andJorde 2002). We are unaware of any direct data on scaupfood availability in the boreal forest during prebreeding.However, landscape-level changes in the boreal forestassociated with a warming climate, including reduced watersurface area of ponds and lakes (Riordan et al. 2006), mayhave caused shifts in aquatic invertebrate abundance orphenology that in turn affect scaup reproductive effort(Corcoran 2005).
Female SurvivalScaup breeding-season survival was higher than we expectedand could be due to a low proportion of breeding females inthe population and, thus, less risk of predation duringnesting activities. Marker retention was high, because only 5females lost transmitters before the last day of monitoring.We located all missing transmitters in deep water and couldnot retrieve them; thus, they probably fell off duringpreening. We believe that this was not a result of mortalitybecause all occurred in the last month of monitoring, whenthe attachment was likely less secure.
Survival rates of female lesser scaup at the Long Lakecomplex differed between years (0.92 and 0.76). In 2005,both mortalities were due to predation while females werenesting, whereas in 2006, 3 females died after the nestingand brood-rearing periods were largely over (after 25 Jul).Previous studies found that most female mortality duringthe breeding season occurred while the female wasincubating or rearing broods (Cowardin et al. 1985, Kirbyand Cowardin 1986, DeVries et al. 2003, Richkus et al.2005). Exclusion of mortalities that occurred after nestingand brood-rearing periods resulted in female survivalprobability of 0.86 (SE 5 0.07) in 2006. Although weobserved no direct effect of handling or markers on scaupsurvival, subtle, indirect effects cannot be completelydismissed.
Previous studies in the boreal-forest report that scaupfemale survival probability was 0.80 during the first 42 daysof the breeding season, prior to brood rearing (Brook andClark 2005), and 0.80 during the brood-rearing period (44days; Walker and Lindberg 2005) for females that obviouslybred. Therefore mortality during the nesting and brood-rearing periods (0.36) constitutes a substantial portion ofannual mortality of breeding scaup. In contrast, survival ofscaup at the Long Lake complex during nesting and brood-rearing periods averaged 0.89 (x 5 0.92 and 0.86) over bothyears. The relatively high survival may have been a result oflow breeding effort at our study area. Brook and Clark(2005) did not determine reproductive status of females, butSafine (2005) found that survival probability of egg-producing white-winged scoters (Melanitta fusca) was lowerthan that of non–egg-producing females. Survival of femalescaup may vary spatially across the continent (Lake et al.2006), and temporally, based on reproductive effort.
Life history theory predicts trade-offs between compo-nents of fitness; namely, that reproductive effort in one yearmay impair subsequent survival and reproduction (Stearns1989, Cam et al. 1998). Some evidence supports this idea of
a tradeoff for diving ducks (Rotella et al. 2003) and forprairie-nesting dabbling duck species (Arnold and Clark1996, Dufour and Clark 2002). Duck species may exhibitthis tradeoff to varying degrees along a continuum from r-selected to K-selected life history strategies (Johnson andGrier 1988). A strongly r-selected species attempts toreproduce each year and consequently has a low survivalprobability. If the survival cost of breeding varies by age orenvironmental conditions, a K-selected species may deferbreeding when conditions are unfavorable, which couldincrease their probability of survival and reproduction infollowing years. Species in tribe Aythyini have beenregarded as more K-selected than species in Anatini(Vickery and Nudds 1984, Johnson and Grier 1988), butour results suggest that boreal-forest scaup are even closer tothe K-selected extreme than previously thought.
MANAGEMENT IMPLICATIONS
Spring breeding-pair surveys assume that all pairs attempt tobreed (Cowardin and Blohm 1992, Nichols et al. 1995);however, we found that
L
33% of lesser scaup pairs on ourstudy site were not breeding. Recruitment estimates basedon annual breeding-pair surveys may be overestimatingproductivity of scaup pairs in the boreal forest, which in turnmay have implications for harvest management of thesebirds (Trauger 1971). Managers should reevaluate thedefinition of a breeding pair and the assumption that allscaup pairs, no matter where they are counted during annualspring surveys, have equal potential productivity. Inaddition, estimates of breeding effort are virtually nonexis-tent for most duck populations. We lack sufficientmethodology to quantify the bias associated with currenttelemetry and physiological methods of estimating breedingprobability. Development of novel methods examiningbreeding probability and additional research on howenvironmental and physiological cues influence RFGinitiation and completion are clearly needed.
ACKNOWLEDGMENTS
Financial support for this project was provided by the YukonFlats National Wildlife Refuge, the Delta WaterfowlFoundation, the University of Alaska Fairbanks (UAF)Angus Gavin Migratory Bird Research Fund, the AlaskaCooperative Fish and Wildlife Research Unit, the UAFDepartment of Biology, and the UAF Institute of ArcticBiology. The staff of Yukon Flats National Wildlife Refugeassisted with all aspects of field logistics; we especially thankN. Guldager, M. Hinkes, and M. Vivion. We are indebtedto all of the hard-working field assistants and volunteers,including C. VanStratt, J. Wells, D. Safine, C. Hisaoka, J.Kopanke, E. Smith, J. Minerva, T. Heuer, B. Whitehill, J.Fox, P. Williams, J. Akaran, S. Dufford, L. Devaney, A.Martin, T. Tanner, P. Barboza, T. Welsh, J. Schmidt, andM. Petrula. We also thank the staff at the UAF AnimalQuarters for maintenance of captive birds, and M.MacCluskie, D. Esler, and T. Williams for lab analyses.Comments from 2 anonymous reviewers greatly improvedthis manuscript.
Martin et al. N Demography of Lesser Scaup 921

LITERATURE CITED
Afton, A. D. 1984. Influence of age and time on reproductive performanceof lesser scaup. Auk 101:255–265.
Afton, A. D., and M. G. Anderson. 2001. Declining scaup populations: aretrospective analysis of long-term population and harvest survey data.Journal of Wildlife Management 65:781–796.
Afton, A. D., and C. D. Ankney. 1991. Nutrient-reserve dynamics ofbreeding lesser scaup: a test of competing hypotheses. Condor 93:89–97.
Afton, A. D., and S. L. Paulus. 1992. Incubation and brood care. Pages 62–108 in B. D. J. Batt, A. D. Afton, M. G. Anderson, C. D. Ankney, D. H.Johnson, J. A. Kadlec, and G. L. Krapu, editors. Ecology andmanagement of breeding waterfowl. University of Minnesota Press,Minneapolis, USA.
Alisauskas, R. T., and C. D. Ankney. 1987. Age-related variation innutrient reserves of breeding American Coots. Canadian Journal ofZoology 65:2417–2420.
Alisauskas, R. T., and C. D. Ankney. 1992. The cost of egg laying and itsrelationship to nutrient reserves in waterfowl. Pages 30–61 in B. D. J.Batt, A. D. Afton, M. G. Anderson, C. D. Ankney, D. H. Johnson, J. A.Kadlec, and G. L. Krapu, editors. Ecology and management of breedingwaterfowl. University of Minnesota Press, Minneapolis, USA.
Alisauskas, R. T., R. T. Eberhardt, and C. D. Ankney. 1990. Nutrientreserves of breeding ring-necked ducks. Canadian Journal of Zoology68:2524–2530.
Anderson, M. G., M. S. Lindberg, and R. B. Emery. 2001. Probability ofsurvival and breeding for juvenile female canvasbacks. Journal of WildlifeManagement 65:385–397.
Anderson, M. G., R. D. Sayler, and A. D. Afton. 1980. A decoy trap fordiving ducks. Journal of Wildlife Management 44:217–219.
Ankney, C. D., and R. T. Alisauskas. 1991. Nutrient-reserve dynamics anddiet of breeding female gadwalls. Condor 93:799–810.
Anteau, M. J., and A. D. Afton. 2004. Nutrient reserves of lesser scaup(Aythya affinis) during spring migration in the Mississippi Flyway: a testof the spring condition hypothesis. Auk 121:917–929.
Arnold, T. W., M. G. Anderson, M. D. Sorenson, and R. B. Emery. 2002.Survival and philopatry of female redheads breeding in SouthwesternManitoba. Journal of Wildlife Management 66:162–169.
Arnold, T. W., and R. G. Clark. 1996. Survival and philopatry of femaledabbling ducks in south central Saskatchewan. Journal of WildlifeManagement 60:560–568.
Austin, J. E., A. D. Afton, M. G. Anderson, R. G. Clark, C. M. Custer, J.S. Lawrence, J. B. Pollard, and J. K. Ringelman. 2000. Declining scauppopulations: issues, hypotheses, and research needs. Wildlife SocietyBulletin 28:254–263.
Barboza, P. S., and D. G. Jorde. 2002. Intermittent fasting during winterand spring affects body composition and reproduction of a migratoryduck. Journal of Comparative Physiology B 172:419–434.
Barry, T. W. 1962. Effects of late seasons on Atlantic brant reproduction.Journal of Wildlife Management 26:19–26.
Beauchamp, W. D., R. R. Koford, T. D. Nudds, R. G. Clark, and D. H.Johnson. 1996. Long-term declines in nest success of prairie ducks.Journal of Wildlife Management 60:247–257.
Bellrose, F. C. 1980. Ducks, geese and swans of North America. Stackpole,Harrisburg, Pennsylvania, USA.
Blums, P., R. G. Clark, and A. Mednis. 2002. Patterns of reproductiveeffort and success in birds: path analyses of long-term data fromEuropean ducks. Journal of Animal Ecology 71:280–295.
Bond, J. C. 2005. Nutrient acquisition and allocation strategies forreproduction by female harlequin ducks. Thesis, Simon Fraser University,Burnaby, British Columbia, Canada.
Brook, R. W., and R. G. Clark. 2002. Retention and effects of nasalmarkers and subcutaneous implanted radio transmitters on breedingfemale scaup. Journal of Field Ornithology 73:206–212.
Brook, R. W., and R. G. Clark. 2005. Breeding season survival of femalelesser scaup in the northern boreal forest. Arctic 58:16–20.
Burnham, K. P., and D. R. Anderson. 1998. Model selection and inference:a practical information-theoretic approach. Springer-Verlag, New York,New York, USA.
Cam, E., J. E. Hines, J. Monnat, J. D. Nichols, and E. Danchin. 1998. Areadult nonbreeders prudent parents? The kittiwake model. Ecology79:2917–2930.
Challenger, W. O., T. D. Williams, J. K. Christians, and F. Vezina. 2001.Follicular development and plasma yolk precursor dynamics through thelaying cycle in the European starling. Physiological and BiochemicalZoology 74:356–365.
Conant, B., and D. J. Groves. 2005. Waterfowl breeding population survey:Alaska–Yukon (Crew area 1). U.S. Fish and Wildlife Service, Juneau,Alaska, USA.
Corcoran, R. 2005. Lesser scaup nesting ecology in relation to waterchemistry and macroinvertebrates on the Yukon Flats, Alaska. Thesis,University of Wyoming, Laramie, USA.
Corcoran, R. M., J. R. Lovvorn, M. R. Bertram, and M. T. Vivion. 2007.Lesser scaup nest success and duckling survival on the Yukon Flats,Alaska. Journal of Wildlife Management 71:127–134.
Cowardin, L. M., and R. J. Blohm. 1992. Breeding population inventoriesand measures of recruitment. Pages 423–445 in B. D. J. Batt, A. D.Afton, M. G. Anderson, C. D. Ankney, D. H. Johnson, J. A. Kadlec, andG. L. Krapu, editors. Ecology and management of breeding waterfowl.University of Minnesota Press, Minneapolis, USA.
Cowardin, L. M., D. S. Gilmer, and C. W. Shaiffer. 1985. Mallardrecruitment in the agricultural environment of North Dakota. WildlifeMonographs 92.
DeVink, J. M. 2007. Comparative reproductive energetics and seleniumecotoxicology in three boreal-breeding waterfowl species. Dissertation.University of Saskatchewan, Saskatoon, Canada.
DeVries, J. H., J. J. Citta, M. S. Lindberg, D. W. Howerter, and M. G.Anderson. 2003. Breeding-season survival of mallard females in theprairie pothole region of Canada. Journal of Wildlife Management67:551–563.
Dinsmore, S. J., G. C. White, and F. L. Knopf. 2002. Advanced techniquesfor modeling avian nest survival. Ecology 83:3476–3488.
Dufour, K. W., and R. G. Clark. 2002. Differential survival of yearling andadult female mallards and its relation to breeding habitat conditions.Condor 104:297–308.
Esler, D., J. B. Grand, and A. D. Afton. 2001. Intraspecific variation innutrient reserve use during clutch formation by lesser scaup. Condor130:810–820.
Flint, P.L., J.B. Grand, T.F. Fondell, J.A. Morse. 2006. Populationdynamics of greater scaup breeding on the Yukon–Kuskokwim Delta,Alaska. Wildlife Monographs 162.
Fournier, M. A., and J. E. Hines. 1998. Productivity and populationincrease of subarctic breeding canvasbacks. Journal of Wildlife Manage-ment 62:179–184.
Gilbert, A. B., M. M. Perry, D. Waddington, and M. A. Hardie. 1983.Role of atresia in estimating the follicular hierarchy in the ovary of thedomestic hen. Journal of Reproduction and Fertility 69:221–227.
Gorman, K. B. 2005. Reproductive energetics of female greater scaup:nutritional and physiological correlates of timing and state of reproduc-tion. Thesis, Simon Fraser University, Burnaby, British Columbia,Canada.
Gorman, K. B., D. Esler, P. L. Flint, and T. D. Williams. 2008. Nutrientreserve dynamics during egg production by female greater scaup:relationships with timing of reproduction. Auk 125:384–394.
Grand, J. B. 1995. Nesting success of ducks on the central Yukon Flats,Alaska. Canadian Journal of Zoology 73:260–265.
Greenwood, H., R. G. Clark, and P. J. Weatherhead. 1986. Condition biasof hunter-shot mallards (Anas platyrhynchos). Canadian Journal ofZoology 64:599–601.
Heglund, P. J. 1992. Patterns of wetland use among aquatic birds in theinterior boreal forest region of Alaska. Dissertation, University ofMissouri–Columbia, USA.
Hoekman, S. T., L. S. Mills, D. W. Howerter, J. H. DeVries, and I. J. Ball.2002. Sensitivity analyses of the life cycle of mid-continent mallards.Journal of Wildlife Management 66:883–900.
Howerter, D. W., B. L. Joynt, R. B. Emery, and T. P. Sankowski. 1997.Effects of nasal discs on nesting by mallards. Journal of Field Ornithology68:1–6.
Johnson, D. H., and J. W. Grier. 1988. Determinants of breedingdistributions of ducks. Wildlife Monographs 100.
Johnson, D. H., J. D. Nichols, and M. D. Schwartz. 1992. Populationdynamics of breeding waterfowl. Pages 446–485 in B. D. J. Batt, A. D.Afton, M. G. Anderson, C. D. Ankney, D. H. Johnson, J. A. Kadlec, andG. L. Krapu, editors. Ecology and management of breeding waterfowl.University of Minnesota Press, Minneapolis, USA.
922 The Journal of Wildlife Management N 73(6)

Johnson, D. H., and T. L. Shaffer. 1990. Estimating nest success: whenMayfield wins. Auk 107:595–600.
Kirby, R. E., and L. M. Cowardin. 1986. Spring and summer survival offemale mallards from north-central Minnesota. Journal of WildlifeManagement 50:38–43.
Klett, A. T., and D. H. Johnson. 1982. Variability in nest survival rates andits importance to nesting studies. Auk 99:77–87.
Koons, D. N., J. J. Rotella, D. W. Willey, M. Taper, R. G. Clark, S.Slattery, R. W. Brook, R. M. Corcoran, and J. R. Lovvorn. 2006. Lesserscaup population dynamics: what can be learned from available data?Avian Conservation and Ecology 1:6.
Lake, B., J. Walker, and M. S. Lindberg. 2006. Survival of ducks banded inthe boreal forest of Alaska. Journal of Wildlife Management 70:443–449.
Lindstrom, E. B., M. W. Eichholz, and J. M. Eadie. 2006. Postovulatoryfollicles in mallards: implications for estimates of breeding propensity.Condor 108:925–935.
Martin, K. H. 2007. Breeding ecology and fasting tolerance of scaup andother ducks in the boreal forest of Alaska. Thesis, University of Alaska–Fairbanks, USA.
McPherson, R., T. W. Arnold, L. Armstrong, and C. J. Schwarz. 2003.Estimating the number of nests initiated by radiomarked mallards.Journal of Wildlife Management 67:843–851.
Miller, H. W., and D. H. Johnson. 1978. Interpreting the results of nestingstudies. Journal of Wildlife Management 42:471–478.
Mitchell, M. A., and A. J. Carlisle. 1991. Plasma zinc as an index ofvitellogenin production and reproductive status in the domestic fowl.Comparative Biochemistry and Physiology 100:719–724.
Nichols, J. D., F. A. Johnson, and B. K. Williams. 1995. Managing NorthAmerican waterfowl in the face of uncertainty. Annual Review of Ecologyand Systematics 26:177–199.
Paquette, G. A., J. H. DeVries, R. B. Emery, D. W. Howerter, B. L. Joynt,and T. P. Sankowski. 1997. Effects of transmitters on reproduction andsurvival of wild mallards. Journal of Wildlife Management 61:953–961.
Petrula, M. J. 1994. Nesting ecology of ducks in interior Alaska. Thesis,University of Alaska–Fairbanks, USA.
Pietz, P. L., D. A. Brandt, G. L. Krapu, and D. A. Buhl. 1995. Modifiedtransmitter attachment method for adult ducks. Journal of FieldOrnithology 66:408–417.
Ramsey, F. L., and D. W. Schafer. 1997. The statistical sleuth: a course inmethods of data analysis. Wadsworth, Belmont, California, USA.
Reinecke, K. J., and C. W. Shaiffer. 1988. A field test for differences incondition among trapped and shot mallards. Journal of WildlifeManagement 52:227–232.
Richkus, K. D., F. C. Rohwer, and M. J. Chamberlain. 2005. Survival andcause-specific mortality of female northern pintail in southern Saskatch-ewan. Journal of Wildlife Management 69:574–581.
Riordan, B., D. Verbyla, and A. D. McGuire. 2006. Shrinking ponds insubarctic Alaska based on 1950–2002 remotely sensed images. Journal ofGeophysical Research 111:1–11.
Rohwer, F. C. 1992. The evolution of reproductive patterns in waterfowl.Pages 486–539 in B. D. J. Batt, A. D. Afton, M. G. Anderson, C. D.Ankney, D. H. Johnson, J. A. Kadlec, and G. L. Krapu, editors. Ecologyand management of breeding waterfowl. University of Minnesota Press,Minneapolis, USA.
Rotella, J. J., R. G. Clark, and A. D. Afton. 2003. Survival of female lesserscaup: effects of body size, age and reproductive effort. Condor 105:336–347.
Rotella, J. J., D. W. Howerter, T. P. Sankowski, and J. H. DeVries. 1993.Nesting effort by wild mallards with 3 types of radio transmitters. Journalof Wildlife Management 57:690–695.
Rotella, J. J., and J. T. Ratti. 1990. Use of methoxyflurane to reduce nestabandonment of mallards. Journal of Wildlife Management 54:627–628.
Safine, D. E. 2005. Breeding ecology of white-winged scoters on the YukonFlats, Alaska. Thesis, University of Alaska–Fairbanks, USA.
Sargeant, A. B., and D. G. Raveling. 1992. Mortality during the breedingseason. Pages 396–422 in B. D. J. Batt, A. D. Afton, M. G. Anderson, C.D. Ankney, D. H. Johnson, J. A. Kadlec, and G. L. Krapu, editors.Ecology and management of breeding waterfowl. University ofMinnesota Press, Minneapolis, USA.
Sargeant, A. B., M. A. Sovada, and R. J. Greenwood. 1998. Interpretingevidence of depredation of duck nests in the prairie pothole region. U.S.Geological Survey, Jamestown, North Dakota, USA.
Seber, G. F. 1982. The estimation of animal abundance and relatedparameters. Second edition. Macmillan, New York, New York, USA.
Sedinger, J. S., C. D. Ankney, and R. T. Alisauskas. 1997. Refinedmethods for assessment of nutrient reserve use and regulation of clutchsize. Condor 99:836–840.
Sharp, D. E., and J. T. Lokemoen. 1987. A decoy trap for breeding-seasonmallards in North Dakota. Journal of Wildlife Management 51:711–715.
Stearns, S. C. 1989. Trade-offs in life-history evolution. FunctionalEcology 3:259–268.
Thomas, V. G. 1988. Body condition, ovarian hierarchies, and their relationto egg formation in Anseriform and Galliform species. Proceedings ofInternational Ornithological Congress 19:353–363.
Trauger, D. L. 1971. Population ecology of lesser scaup in subarctic taiga.Dissertation, Iowa State University, Ames, USA.
Trauger, D. L. 1974. Eye color of female lesser scaup in relation to age. Auk91:243–254.
United States Fish and Wildlife Service [USFWS]. 1986. North Americanwaterfowl management plan. U.S. Department of the Interior, Wash-ington, D.C., USA.
United States Fish and Wildlife Service [USFWS]. 1987. Yukon FlatsNational Wildlife refuge comprehensive conservation plan, environmen-tal impact statement and wilderness review. U.S. Fish and WildlifeService, Final Draft, Anchorage, Alaska, USA.
Vickery, W. T., and T. D. Nudds. 1984. Detection of density-dependenteffects in annual duck censuses. Ecology 65:96–104.
Walker, J., and M. S. Lindberg. 2005. Survival of scaup ducklings in theboreal forest of Alaska. Journal of Wildlife Management 69:592–600.
Walker, J., M. S. Lindberg, M. C. MacCluskie, M. J. Petrula, and J. S.Sedinger. 2005. Nest survival of scaup and other ducks in the boreal forestof Alaska. Journal of Wildlife Management 69:582–591.
Weller, M. W. 1957. An automatic nest-trap for waterfowl. Journal ofWildlife Management 21:456–458.
White, G. C., and K. P. Burnham. 1999. Program MARK: survivalestimation from populations of marked animals. Bird Study 46(Supple-ment):120–138.
Wilkins, K. A., M. C. Otto, and M. D. Koneff. 2006. Waterfowlpopulation status 2006. U.S. Department of Interior, Fish and WildlifeService, Laurel, Maryland, USA.
Zimpfer, N. L., G. S. Zimmerman, E. D. Silverman, and M. D. Koneff.2008. Trends in duck breeding populations 1955–2008. U.S. Departmentof Interior, Fish and Wildlife Service, Laurel, Maryland, USA.
Associate Editor: Arnold.
Martin et al. N Demography of Lesser Scaup 923





























