Embed Size (px)
Citation preview

Limfjordsprojektet. Rapport nr. 8, 1998, pp. 223-234.
Limfjordensnaturforhold i fortidenfaunacendringer belyst vedsubfossiler fra Bj0rnsholm Bugt
Jens Tang Christensen, Tomas Cedhagen og J0rgen Hylleberg
Limfjorden har f0rt en omtumlet tilvcerelse fra sin dannelse som smeltevandsdaltil et arkipelag og sin senere tilstand vekslende,mellem en grenet fjord og et sundfra Nords0en til Kattegat. Disse omskiftelser har uundgaeligt sat sig spor i faunaen. En analyse af dennes subfossile rester, hvor de er bevarede i en ubrudtstratigrafi, kan derfor danne grundlag for en beskrivelse af forholdene og, i forbindelse med datering af materialet, for en tidsfcestelse af indtradte cendringer.Hvor havbundens sediment er tilstrcekkeligt bl0dt, er det muligt at udtage uforstyrrede pmver af denne ned til forholdsvis stor dybde.
Faunaens fordeling i sedimentkernen giver mulighed for at inddele denne i etantal zoner, idet der er nogle ret skarpt markerede cendringer i faunaens sammenscetning, tydende pa hurtigt indtradte milj0forandringer. Den vcesentligstearsag til disse faunaforandringer synes at vcere cendringer i saltholdigheden,hvilket igen er et udtryk for graden af vandudveksling mellem Bj0rnsholm Bugtog Nords0en eller Skagerrak.
lEndringer i vandudvekslingen er et resultat af samspillet mellem den isostatiske landhcevning betinget af den skandinaviske iskappes bortsmeltning,eustatiske processer (cendringer i havniveauet) samt materialetransport (dannelse af landtanger og strandvolde) (K. S. Petersen 1985). I det tidsrum, sommaterialet spcender over, er havoverfladens niveau i omradet faldet et par meter(K. S. Petersen 1992).
I sommeren 1993 blev 10 sedimentkerner, hvoraf nogle var pa 5,5 m's lcengde,udtaget fra L0gst0r-Liv0 Bredning. Analyser af faunaen i en af disse kerner(LIM1002) fra Bj0rnsholm Bugt danner grundlag for de f0lgende betragtningerover naturforholdene i Limfjorden i de sidste ca. 3000 ar. Fra samme bugt har dertidligere vceret udtaget og analyseret et antal 2 m lange borekerner (J. Hylleberg1992; P. Kristensen et al. 1995).
Materiale og metoder
Den analyserede sedimentkerne blev optaget i Bj0rnsholm Bugt pa positionen56°50'3 N, 9°08'96 0 (fig. 1). Kernen blev taget med en 'piston-corer' fra BiologiskInstituts forskningskutter Genetica II i juni 1993. Princippet er, at et mr (stal ellerPVC, 8 cm i diam.) trykkes ned i sedimentet ved blybelastning, hvorefter det halesop, idet det lukkes foroven af et stempel. Det anvendte mr var 6 m langt. Pa landmaltes lcengden af kernen i mret, hvorefter den blev presset ud med et stempel i
223

CJ lIzvcdc Iiolo.:~nc ffiJrlnC Jtlq,m~cr
[3£J GIJcl.ll-l~nJskJb
----50....."-_'.' • '" III'" I~
Fig. 1. Geomorfologisk kort over Limfjorden med lmdersfJgelsespositionen angivet ved en stjerne.
en dertil indrettet bakke, hvor den kunne skceres i 3 em sektioner. Ved udpresningen sker der et tab af sediment og vand mellem rorvceg og stempel. Denudpressede lcengde var saledes 531 em, mens lcengden i roret maltes til 552 em.Sektionerne blev med stor forsigtighed vadsigtet gennem en 500 ~m sigte, og allefauna- og florarester overf0rtes til papstykker med eelluloseklister ved hjcelp afpensel eller pineet.
Materialet, som tilbageholdes pa. en 500 ~m sigte, bestar for hovedpartensvedkommende af skallerne fra bl0ddyr (snegle og muslinger) samt skeletrester afpighude (sostjerner, slangestjerner og s0pindsvin). Ogsa skeletdele af krebsdyrog rester fra ledor.me, som kceber eller rester af mr, forekommer hyppigt. Endeligindeholder materialet en del knogler fra fisk, skaller af foraminiferer samt resteraf insekter. Plantemateriale optrceder i form af alegrces og alger samt terrestriskmateriale som fm og mos. Fisk, muslingekrebs, foraminiferer samt materiale afterrestrisk oprindelse er dog ikke behandlet ncermere her. Den marine makroinvertebratfauna omfatter i alt 78 arter. For sa vidt angar bl0ddyrfaunaen har vifulgt nomenklaturen i K. R. Jensen & J. Knudsen (1995).
Datering af materiale blev foretaget ved aeeelerator-massespektrometri paAMS-Iaboratoriet, Aarhus Universitet. Konventionelle 14C-aldre (felf 1950) erkorrigerede for reservoireffekt med o<;eanreservoiralderen 400 ar, for kalibreringi kalenderar efter M. Stuiver & P. J. Reimer (1993).
Faunaens fordeling
Faunaen i de nederste 330 em af sedimentkernen har et tydeligt marint prceg.AIle 3 em sektioner fra denne zone indeholder skalrester og pigge fra S0musen
224
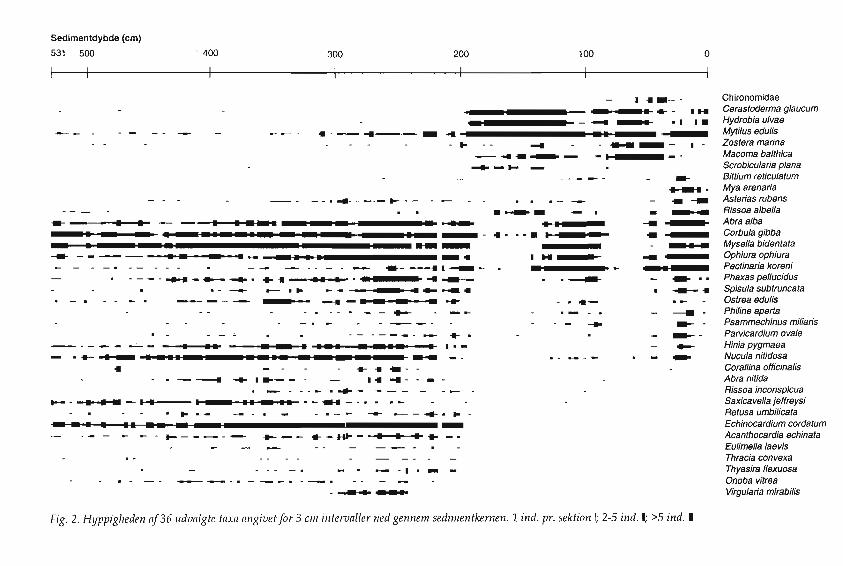
Sedimentdybde (em)
531 500 ·400 300 200 100 o
I I I I I I I
- __ - 1-
I .. __ -
_ ... - IH- ·11-
•••
__ •• - ••• -I • _.. _ .......
• - _. ••__._ - - • - ...... I _-.. ---- :~::::__ --------_. - .- _... _.. I'"- - - - - I _ -. ._-.t _-------_. ...... .. . .. ...--- .. -----.- ~ ....-. .. -- -- ..... --- ..------ . --- -.-- ,..... - . ..----
- -1·-
ChironomidaeCerastoderma g/aucumHydrobia ulvaeMytilus edulisZostera marinaMacoma balthicaScrobicularia planaBittium reticula tumMya arenariaAsterias rubensRissoa albellaAbra albaCorbula gibbaMysella bidentataOphiura ophiuraPectinaria koreniPhaxas pellucidusSplsula subtruncataOstrea edulisPhiline apertaPsammechinus miliarisParvicardium ova/eHinia pygmaeaNucula nit/dosaCorallina officinalisAbranitidaRissoa inconspicuaSaxicavella jeffreysiRetusa umbilicataEchinocardium cordatumAcanthocardia echinataEulimella laevisThracia convexaThyasira flexuosaOnoba vitreaVirgularla mirabilis
-..... -----_..---........--. ---..
-eo
••I· -• - • 1.1 • ••• •
-----
_._-- ~-
----- ....... -- .... --
....--- ....- _... ---- ...... - -• -_ .... 1_- - 1 ._-
~ .•••• -1" I ••• 1. -- ... -- ..... - --. - -.-- .... -- ...... -_ - -... ------ _-- .. -Hl-- ......
tvD:: Fig. 2. Hyppigheden af36 lIdvlllgte taxa angivet for 3 elll intervaller ned gennem sedimentkemen. 1 indo pro sektion I; 2-5 indo I; >5 indo I

Echinocardium cordatum. Dette irregulcere s0pindsvin forekomrner op til 198 em'sdybde. Ogsa den piggede hjertemusling (Acanthocardia echinata) forekomrnerudelukkende i denne zone op til 201 em's dybde (fig. 2).
Blandt muslingeme er Thracia convexa, Thyasira flexuosa, Modiolarca tumida,Mysia undata, Tellimya ferruginosa, Angulus tenuis, Chamelea striatula, Fabulinafabula, Nuculoma tenuis, Hiatella arctica, Mya truncata samt Saxicavella jeffreysiudelukkende fundet i denne nedre marine zone, sidstncevnte art dog med enenkelt undtagelse i intervallet 114-117 em. Blandt sneglene er f0lgende udelukkende fundet i den nedre marine zone: Retusa umbilicata, Aclis ascaris, Aclis minor,Eulimella laevis, Vitreolina philippii, Epitonium clathratulum, Onoba vitrea ogChrysallida indistincta. Muslingen Tellimya ferruginosa (= Montacuta ferruginosa) erdelvis komrnensal pa S0musen Echinocardium, idet den ofte sidder hceftet tildennes analpigge. Flere af sneglene er knyttede til pighude som parasitter (Aclis,Vitreolina).
Et interessant element er forekomsten af s0fjeren eller s0straet Virgulariamirabilis fra 306 op til 243 em. Dette koraldyr forekomrner i dag kun pa dyberevand ind i Kattegat, hvor saltholdigheden er passende h0j. Den nedre marinezone indeholder ogsa arter, som forsvinder ved overgangen til den f0lgendezone, men som komrner igen i h0jere lag. Det gcelder bl.a. muslingeme Abra alba,
~~_. -" -
~ ,~1
.. . . . . .
Fig. 3. Repr<esentanter for den bundlevende fauna i Limfjordens marine periode. 1, Corallinaofficinalis; 2, Asterias rubens; 3, Ostrea edulis; 4, Abra nitida; 5, Abra alba; 6, SaxicavellajeJfreysi; 7, Retusa umbilicata; 8, Onoba vitrea; 9, Corbula gibba; 10, Hinia pygmaea; 11, Ophiuraophiura; 12, Nucztla nitidosa; 13, Pectinaria koreni; 14, Parvicardiulll ovale; 15 Mysella bidentata;16, Echinocardium chordarnm; 17, Rissoa inconspicua; 18, Spisula subtnmcata; 19, Tlzyasiraflexuosa; 20, Phaxas pellucidus; 21, Virgularia mirabilis; 22, Acanthocardia echinata.
226

Fig. 4. Arter som dorninerer bundfaunaen i brakvandsperioderne. 1, Chironomidae; 2, Mytilusedulis; 3, Macoma balthica; 4, Cerastoderma glaucum; 5, Hydrobia ulvae; 6, Scrobicularia plana;7, Rissoa albella.
Corbula gibba, Mysella bidentata, n.0ddemuslingen Nucula nitidosa, den lilleknivmusling Phaxas pellucidus og osters Ostrea edulis, borsteormen Pectinariakoreni og slangestjernen Ophiura ophiura (fig. 3).
Overgangen i ea. 2 m's dybde fra den nederste marine zone til den folgende erret skarpt markeret i faunaen, dels ved de ovenfor mevnte arters forsvinden, delsved tilkornsten af brakvandstolerante arter som hjertemuslingen Cerastodennaglaucum (fra 198 em), ostersomuslingen Macoma balthica (fra 189 em), Scrobiculariaplana (fra 192 em) og den store dyndsnegl Hydrobia ulvae (fra 195 em). Hjertemuslingen Parvicardium hauniense forekommer i en enkelt horisont (189-192).Denne musling har en ret sn~ver salinitetstoleranee meUem 5 og 12 %0. Nar viser disse arters forekornst i sammenh~ng med de arter, som forsvinder i sammeperiode, tegner der sig billedet af et brakvandsrniljo: Abra alba er v~k fra zonen135-189, Mysella bidentata fra 135-192, den liUe knivmusling Phaxas pellucidus fra135-192, slangestjernen Ophiura ophiura fra 135-192, bortset fra horisonten 141144. Vandet bliver gennem denne brakvandsperiode gradvist mere salt, saledesat vi i 135 em's dybde finder forhold, der rninder om omradets nuv~rende tilstand.
En ny brakvandsperiode indtr~der omkring horisonten 87-90 og str~kker sigop til 54 em's dybde i sedimentet. I denne brakvandsperiode optr~der ostersomuslingen (Macoma balthica) i stort tal, mens Scrobicularia plana, som varalrnindelig i den tidligere brakvandsperiode, forsvinder.
Fra 54 em og op til 42 em er der en vis saltpavirkning at spore i faunaen, idetOphiura ophiura igen optr~der. Herefter vender fjorden tilbage til brakvandstilstand op til 36 em's dybde. Denne brakvandsperiode er karakteriseret vedstore m~ngder af Chironornider i sedimentet. Der rna have v~ret en sandmyggeplage ved fjorden i denne periode (fig. 4).
227

Zonen fra 30 til 36 em er prceget af stigende tilgang af saltvandsfauna, og fra30 em og op til overfladen er faunaen som i dag (fig. 5). I den nuvcerende fjordtilstand optrceder ogsa arter, der ikke har vceret til stede siden den nedre marinezone, sasom trugmuslingen Spisula subtruncata og dvcergkonken Hinia pygmaea.Her optrceder ligeledes den almindelige sandmusling (Mya arenaria), som nceppehar haft mulighed for at leve her f0r, selvom den maske allerede er indf0rt fraNordamerika til Europa i sen vikingetid (A. C. Johansen 1918:92; K. S. Petersen etal. 1992).
14C-datering
Datering af marine skalrester frembyder en del problemer, i scerdeleshed nar detdrejer sig om lavvandede ornrader med ringe vandudveksling med oeeanet, ogmed tilfersel af grundvand med gammelt kulstof. Den langsomme udveksling afCO2 mellem atrnosfceren og det oeeaniske reservoir bevirker, at organismeropbygget af marint kulstof far en hejere tilsyneladende alder. Bestemmelse afreservoiralderen ud fra skaller med kendt alder har vist, at mens Nords0en ogSkagerrak kan betragtes sam en del af det nordatlantiske reservoir med entilsyneladende alder pa omkring 400 ar, sa er Limfjorden ikke en del af etensartet 14C-reservoir (S. Heier-Nielsen et al. 1995). Skaller med kendt alder fraLimfjorden har h0jere og mere variable tilsyneladende aldre, hvilket kantilskrives tilf0rsel af gammelt karbonat via grundvandsti1l0b, den sakaldtehardtvandseffekt. Variationer i grundvandstill0b og i graden af udveksling medNords0en/Skagerrak kan saledes give en betydelig usikkerhed i dateringen af
Fig. 5. Arter som er almindeligt forekommende i Bjernsholm Bugt under de n1lVcerende ogtidligere Jjordprcegede forhold. 1, Corbula gibba; 2, Ophiura ophiura; 3, Mya arenaria; 4, Abraalba; 5, Phaxas pellucidus; 6, Bittium reticulatum; 7, Pectinaria koreni; 8, Mytillls edulis; 9,Asterias rubens.
228
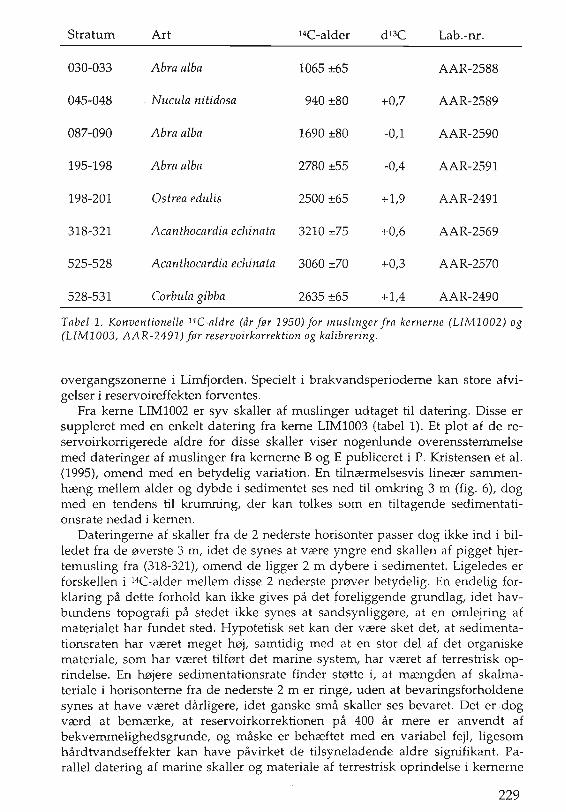
Stratum Art HC-alder d 13C Lab.-m.
030-033 Abra alba 1065 ±65 AAR-2588
045-048 . Nucula nitidosa 940 ±80 +0,7 AAR-2589
087-090 Abra alba 1690 ±80 -0,1 AAR-2590
195-198 Abra alba 2780 ±55 -0,4 AAR-2591
198-201 Ostrea edulis 2500 ±65 +1,9 AAR-2491
318-321 Acanthocardia echinata 3210 ±75 +0,6 AAR-2569
525-528 Acanthocardia echinata 3060 ±70 +0,3 AAR-2570
528-531 Corbula gibba 2635 ±65 +1,4 AAR-2490
Tabel 1. Konventionelle 14C-aldre (ar fer 1950) for muslinger fra kernerne (LIM1002) og(LIM1003, AAR-2491) fM reservoirkorrektion og kalibrering.
overgangszonerne i Limfjorden. Specielt i brakvandsperioderne kan store afvigelser i reservoireffekten forventes.
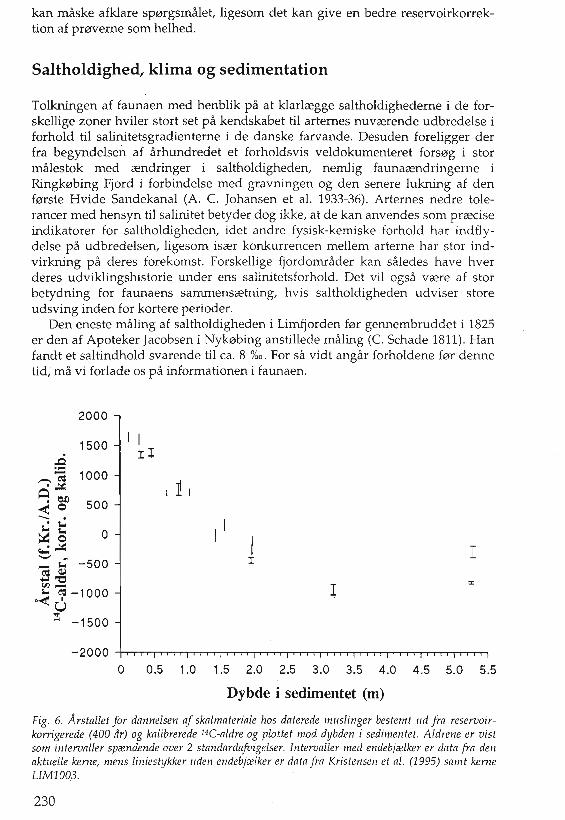
Fra kerne LIM1002 er syv skaller af muslinger udtaget til datering. Disse ersuppleret med en enkelt datering fra kerne LIMI003 (tabel 1). Et plot af de reservoirkorrigerede aldre for disse skaller viser nogenlunde overensstemmelsemed dateringer af muslinger fra kernerne B og E publiceret i P. Kristensen et al.(1995), omend med en betydelig variation. En tilncermelsesvis linecer sammenhceng mellem alder og dybde i sedimentet ses ned til omkring 3 m (fig. 6), dogmed en tendens til krumning, der kan tolkes som en tiltagende sedimentationsrate nedad i kernen.
Dateringerne af skaller fra de 2 nederste horisonter passer dog ikke ind i billedet fra de 0verste 3 m, idet de synes at vcere yngre end skallen af pigget hjertemusling fra (318-321), omend de ligger 2 m dybere i sedimentet. Ligeledes erforskellen i 14C-alder mellem disse 2 nederste pmver betydelig. En endelig forklaring pa dette forhold kan ikke gives pa det foreliggende grundlag, idet havbundens topografi pa stedet ikke synes at sandsynligg0re, at en omlejring afmaterialet har fundet sted. Hypotetisk set kan der vcere sket det, at sedimentationsraten har vceret meget h0j, samtidig med at en stor del af det organiskemateriale, som har vceret tilfmt det marine system, har vceret af terrestrisk oprindelse. En h0jere sedimentationsrate finder st0tte i, at mcengden af skalmateriale i horisonterne fra de nederste 2 m er ringe, uden at bevaringsforholdenesynes at have vceret darligere, idet ganske sma skaller ses bevaret. Det erdogvcerd at bemcerke, at reservoirkorrektionen pa 400 ar mere er anvendt afbekvemmelighedsgrunde, og maske er behceftet med en variabel fejl, ligesomhardtvandseffekter kan have pavirket de tilsyneladende aldre signifikant. Parallel datering af marine skaller og materiale af terrestrisk oprindelse i kernerne
229
kan maske afklare sp0rgsmalet, ligesom det kan give en bedre reservoirkorrektion af pr0veme som helhed.
Saltholdighed, klima og sedimentation
Tolkningen af faunaen med henblik pa at klarlCEgge saltholdighedeme i de forskellige zoner hviler stort set pa kendskabet til artemes nuvCErende udbredelse iforhold til salinitetsgradienteme i de danske farvande. Desuden foreligger derfra begyndelsen af arhundredet et forholdsvis veldokumenteret fors0g i stormalestok med CEndringer i saltholdigheden, nemlig faunaCEndringeme iRingk0bing Fjord i forbindelse med gravningen og den senere lukning af denfmste Hvide Sandekanal (A. C. Johansen et al. 1933-36). Artemes nedre tolerancer med hensyn til salinitet betyder dog ikke, at de kan anvendes som prCEciseindikatorer for saltholdigheden, idet andre fysisk-kemiske forhold har indflydelse pa udbredelsen, ligesom iSCEr konkurrencen mellem arteme har stor indvirkning pa deres forekomst. Forskellige fjordomrader kan saledes have hverderes udviklingshistorie under ens salinitetsforhold. Det viI ogsa VCEre af storbetydning for faunaens sammensCEtning, hvis saltholdigheden udviser storeudsving inden for kortere perioder.
Den eneste rnaling af saltholdigheden i Limfjorden f0r gennembruddet i 1825er den af Apoteker Jacobsen i Nyk0bing anstillede mating (c. Schade 1811). Hanfandt et saltindhold svarende til ca. 8 %0. For sa vidt angar forholdene f0r dennetid, rna vi forlade os pa informationen i faunaen.
2000
I IIl1500
..t:2..~ ea 1000.~
8etl< 0 500-- .s.: t= 0~o~~'-' ~__ -500
eo= a.l
'tl:5-eo=-10000< U...
0-< -1500
I 11
II
I
I
x
o 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5 5.0 5.5
Dybde i sedimentet (m)
Fig. 6. Arstallet for dannelsen af skalmateriale hos daterede m;lslinger bestemt ud fra reservoirkorrigerede (400 dr) og kalibrerede 14C-aldre og plottet mod dybden i sedimentet. Aldrene er vistsom intervllller spi£ndende over 2 standllrdllfuigelser. Intervaller med endebji£lker er dllta frll denIlktuelle kerne, mens liniestlJkker uden endebji£lker er dlltll frll Kristensen et Ill. (1995) sll1nt kerneLIM1003.
230
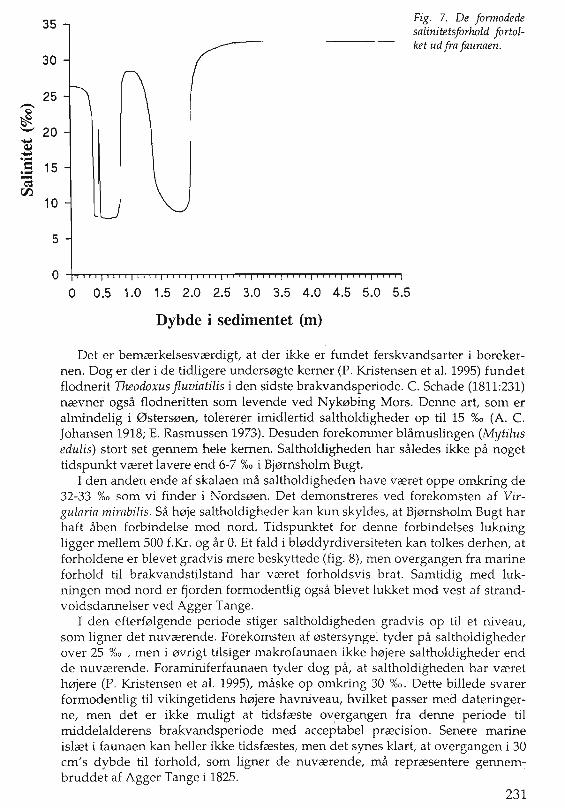
35
30
25..-Q
*-- 20~...-...= 15...-c=
00-10
5
Fig. 7. De fonnodedesalinitetsforhold fortolket ud fra faunaen.
o 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5 5.0 5.5
Dybde i sedimentet (m)
Det er bemCErkelsesvCErdigt, at der ikke er fundet ferskvandsarter i borekernen. Dog er der ide tidligere unders0gte kerner (P. Kristensen et al. 1995) fundetfIodnerit Theodoxus fluviatilis i den sidste brakvandsperiode. C. Schade (1811:231)nCEvner ogsa fIodneritten som levende ved Nyk0bing Mors. Denne art, som eralmindelig i 0sters0en, tolererer imidlertid saltholdigheder op til 15 0/00 (A. C.Johansen 1918; E. Rasmussen 1973). Desuden forekommer blamuslingen (Mytilusedulis) stort set gennem hele kernen. Saltholdigheden har saIedes ikke pa nogettidspunkt VCEret lavere end 6-7 %0 i Bj0rnsholm Bugt.
I den anden ende af skalaen rna saltholdigheden have VCEret oppe omkring de32-33 %0 som vi finder i Nords0en. Det demonstreres ved forekomsten af Virgularia mirabilis. Sa h0je saltholdigheder kan kun skyIdes, at Bj0fnsholm Bugt harhaft aben forbindelse mod nord. Tidspunktet for denne forbindelses lukningligger mellem 500 f.Kr. og ar O. Et fald i bl0ddyrdiversiteten kan tolkes derhen, atforholdene er blevet gradvis mere beskyttede (fig. 8), men overgangen fra marineforhold til brakvandstilstand har VCEret forholdsvis brat. Samtidig med lukningen mod nord er fjorden formodentlig ogsa blevet lukket mod vest af strandvoldsdannelser ved Agger Tange.
I den efterf0lgende periode stiger saltholdigheden gradvis op til et niveau,som ligner det nuvCErende. Forekomsten af 0stersyngel tyder pa saltholdighederover 25 %0 , men i 0vrigt tilsiger makrofaunaen ikke h0jere saltholdigheder endde nuvCErende. Foraminiferfaunaen tyder dog pa, at saltholdigheden har VCEreth0jere (P. Kristensen et al. 1995), maske op omkring 30 %0. Dette billede svarerformodentlig til vikingetidens h0jere havniveau, hvilket passer med dateringerne, men det er ikke muligt at tidsfCEste overgangen fra denne periode tilmiddelalderens brakvandsperiode med acceptabel prCEcision. Senere marineislCEt i faunaen kan heller ikke tidsfCEstes, 'men det synes klart, at overgangen i 30em's dybde til forhold, som ligner de nuvCErende, rna reprCEsentere gennembruddet af Agger Tange i 1825.
231
Med den usikkerhed, som i 0jeblikket er knyttet til dateringen af marineskaller fra fjordomrader, kan 14C-dateringerne ikke bidrage til en afklaring afsammenhrengen mellem faunaovergangene og beretninger om abninger oghavgennembrud i de historiske kilder. Det er muligt at beskrive salinitetsforl0beti forhold til dybden (fig. 7), men sammenhrengen mellem dette og de historiskebegivenheder viI stadig vrere aben for fortolkning.
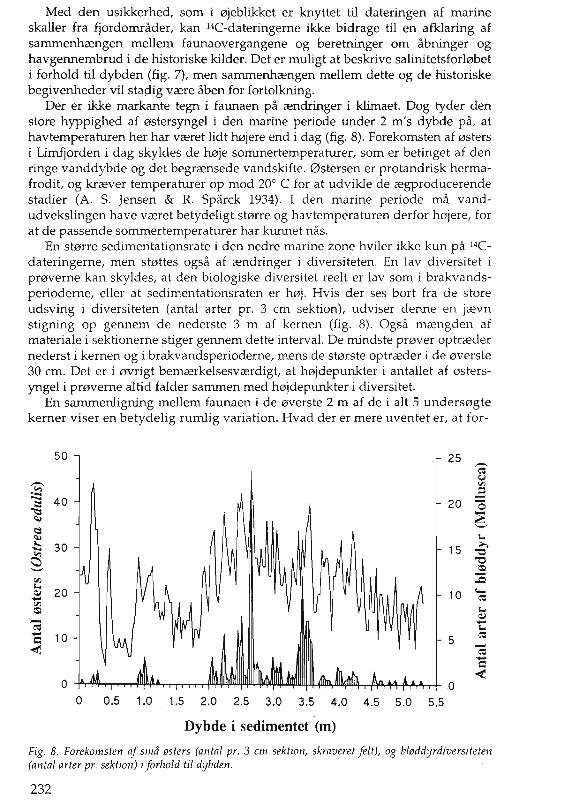
Der er ikke markante tegn i faunaen pa rendringer i klimaet. Dog tyder denstore hyppighed af 0stersyngel i den marine periode under 2 m's dybde pa, athavtemperaturen her har vreret lidt Mjere end i dag (fig. 8). Forekomsten af 0stersi Limfjorden i dag skyldes de ooje sommertemperaturer, som er betinget af denringe vanddybde og det begrrensede vandskifte. 0stersen er protandrisk hermafrodit, og krrever temperaturer op mod 20° C for at udvikle de regprodueerendestadier (A. S. Jensen & R. Sparek 1934). I den marine periode rna vandudvekslingen have vreret betydeligt st0rre og havtemperaturen derfor h0jere, forat de passende sommertemperaturer har kunnet nas.
En stmre sedimentationsrate i den nedre marine zone hviler ikke kun pa 14Cdateringerne, men st0ttes ogsa af rendringer i diversiteten. En lay diversitet ipmverne kan skyldes, at den biologiske diversitet reelt er lay som i brakvandsperioderne, eller at sedimentationsraten er h0j. Hvis der ses bort fra de storeudsving i diversiteten (antal arter pro 3 em sektion), udviser denne en jrevnstigning op gennem de nederste 3 m af kernen (fig. 8). Ogsa mrengden afmateriale i sektionerne stiger gennem dette interval. De mindste pmver optrredernederst i kernen og i brakvandsperioderne, mens de st0rste optrreder i de 0verste30 em. Det er i 0vrigt bemrerkelsesvrerdigt, at h0jdepunkter i antallet af 0stersyngel i pmverne altid falder sammen med h0jdepunkter i diversitet.
En sammenligning mellem faunaen i de 0verste 2 m af de i alt 5 unders0gtekerner viser en betydelig rurnlig variation. Hvad der er mere uventet er, at for-
50 25 -~(,J- '".=:l .2- 40.§ 20 '0
~ ~I::S ---~
10.
30 15,.,
C "0"0
--- IS:!
'" :c10.
20 ....~ 10.... ~
'"IS:! 10.~- ....
~ 10..... 10 ~c 5< -;....
C
<0 00 0.5 1.0 _1.5 2.0 2.5 3.0 3.5 4.0 4.5 5.0 5.5
-Dybde i sedimentet (m)
Fig. 8. Forekomsten af sma lJsters (antal pro 3 em sektion, skraveret felt), og bllJddyrdiversiteten(an tal arter pro sektion) i forhold til dybden.
232

233
skellen mellem kernerne mht. den kvantitative forekomst af visse arter, somLeks. den store borsteorm Neanthes virens, synes at holde sig.
En detaljeret analyse af faunaen i sedimentkerner kan bidrage viEsentligt tilforstaelsen af miljoforholdene til forskellige tider. Disse undersogelser kan ogsa~ene til at stotte fortolkningen af det historiske kildemateriale. En uafhiEngigtidsfiEstelse af indtradte forandringer opnar vi dog forst, nar problemerne medfysisk datering af materiale fra kystniEre omrader er blevet lost.
Tak
En stor tak skal rettes til Jan Heinemeier, Susanne Heier-Nielsen og Vibeke Jensen, AMS-Iaboratoriet i Arhus, for velvillighed og hurtighed i forbindelse med14C-datering af materiale fra kernen, samt til Kurt Ockelmann, MarinbiologiskLaboratorium, Helsingor, for hjiElp med bestemmelse af sma, vanskelige muslinger.
Litteratur
Heier-Nielsen, S., J. Heinemeier, H. L. Nielsen & N. Rud1995: Recent reservoir ages for Danish fiords and marine waters. (Radiocarbon 37).
pp. 875-882.
Hylleberg, J.1992: iEndringer i Limfjordens dyreliv i historisk tid.
(Limfjordsprojektet. Rapport nr. 4). pp. 37-61.
Jensen, A. S. & R. Sparck1934: Danmarks Fauna. B10ddyr II. Saltvandsmuslinger. K0benhavn. 208 p.
Jensen, K. R. & J. Knudsen1995: Annotated checklist of recent marine molluscs of Danish waters.
Zoological Museum, Copenhagen. 73 p.
Johansen, A. C. (red.)1918: Randers Fjords Naturhistorie. K0benhavn. 520 p.
Johansen, A. c., H. Blegvad & R. Sparck (red.)1933-36: Ringk0bing Fjords naturhistorie i brakvandsperioden 1915-1931. K0benhavn.
252 p.
Kristensen, P., S. Heier-Nielsen & J. Hylleberg1995: Late-Holocene salinity fluctuations in Bj0rnsholm Bay, Limfjorden, Denmark, as
deduced from micro- and macrofossil analysis(The Holocene, 5). pp. 313-322.

Petersen, K. S.1985: The late Quaternary history of Denmark. The Weichselian icesheets and
land/sea configuration in the late Pleistocene and Holocene.Qoumal of Danish Archaeology, 4). pp. 7-22.
1992: Om st0rre geologiske/rnilj0historiske rendringer i Limfjorden i Holocren.(Limfjordsprojektet. Rapport ill. 4). pp. 17-28.
Petersen, K. S., J. Heinemeier & N. Rud1992: Clams before Columbus? (Nature 359). p. 679.
Rasmussen, E.1973: Systematics and ecology of the Isefjord marine fauna (Denmark).
(Ophelia 11). pp. 1-507.
Schade, C.1811: Beskrivelse over 0en Mors. Aalborg. 372 p.
Stuiver, M. & P. J. Reimer.1993: Extended 14C data base revised CALIB 3.0 HC age calibration.
(Radiocarbon 35). pp. 215-230.
234





























