Embed Size (px)
Citation preview

Ld
MDV
R
timtHtssiMttP
c
da(Tattmtbmatspsutc
E
Biochemical and Biophysical Research Communications 287, 1093–1098 (2001)
doi:10.1006/bbrc.2001.5503, available online at http://www.idealibrary.com on
ocalization of Mitochondrial HSP56 Chaperoninuring Sea Urchin Development
. C. Roccheri, M. Patti, M. Agnello, F. Gianguzza, E. Carra, and A. M. Rinaldi1
ipartimento di Biologia Cellulare e dello Sviluppo “A. Monroy,” Universita di Palermo,iale delle Scienze, Parco D’Orleans, 90128 Palermo, Italy
eceived July 31, 2001
partially unfolded state (3–9). In addition, it seemsnfdaa
tpiItmrmsg
la5lfP
aio
M
PEoowoems
g
We have previously demonstrated that Paracentro-us lividus nuclear genome encodes for the heat shocknducible chaperonin homolog Hsp 56 (1) and that the
ature protein is localized in the mitochondrial ma-rix (2). In this paper we report that constitutivesp56 is maternally inherited, in fact it is present in
he in unfertilized eggs, and that it has a perinuclearpecific localization during cleavage. In the latertages both the constitutive and the heat shock induc-ble chaperonin has a specific territorial distribution.
oreover following heat shock, the Hsp56 appears inhe cytoplasm and in the postmitochondrial superna-ant beside the mitochondrial fraction. © 2001 Academic
ress
Key Words: mitochondria; sea urchin; development;haperonine; heat shock.
In the eukaryotic systems the majority of mitochon-rial proteins are nuclearly coded, and are synthesizeds preproteins with an additional N-terminal segmentpresequences) of about 20–60 amino acid residues.hey are then translocated to mitochondria with thessistance of cytosolic factors or chaperones and sortedo the different organelle compartments. The recogni-ion of the preproteins and the transport across the twoembranes are facilitated by translocases present in
he mitochondrial outer (TOM) and inner (TIM) mem-ranes. Once preproteins get to the matrix compart-ent, specific peptidases remove the presequences
nd, since the preproteins are translocated in an ex-ended state, they are folded to the native state as-isted by molecular chaperones. The 60 kDa heat shockrotein (Hsp60) has essential functions both in un-tressed and stressed cells, by binding and stabilizingnstable conformations of precursor proteins, in ordero allow the correct folding, assembly into the definitiveomplexes or the transport across membranes in a
1 To whom correspondence should be addressed. Fax: 091-420897.-mail: [email protected].
1093
ow clear, that the Hsp60 is essential not only forolding of imported proteins but also in protein degra-ation process, regulating both the protein turnovernd the degradation of incorrectly folded proteins orggregates.Since this chaperonin is very highly conserved
hroughout evolution from bacteria to man (10) itsathway and its function have been extensively stud-ed in many organisms from bacteria to yeast and man.t is coded by the nucleus, and after synthesis andranslocation, it usually remains in the mitochondrialatrix. It seems extremely interesting to study the
egulation of the synthesis and activity in a develop-ental system, sea urchin, in which we have demon-
trated the uncoupling of nuclear and mitochondrialene expression and replication (11–13).We have already demonstrated that Paracentrotus
ividus mitochondrial matrix contains a constitutivend heat shock inducible Hsp60 chaperonin homolog of6 kDa molecular mass (2), and more recently we iso-ated and characterized a full-length cDNA encodingor the heat shock inducible chaperonin homolog ofaracentrotus lividus embryos (1).In this paper, we present data aimed to better char-
cterize and localize both the constitutive and heatnduced Hsp56 in eggs and embryos at different devel-pmental stages.
ATERIALS AND METHODS
Embryo culture. Adult sea urchins of the Mediterranean speciesaracentrotus lividus were collected along Sicily’s western coasts.ggs were fertilized and embryos grown at 18°C, at a concentrationf 5000/ml in Millipore filtered sea water (mfsw) containing antibi-tics, in a beaker with a rotating propeller, until the desired stagesere reached. Fertilization was carried out in mfsw containing 3 mMf p-aminobenzoic acid (PABA, Sigma) in order to avoid fertilizationnvelop hardening. Fertilized eggs were soon passed through a 75m mesh to remove fertilization envelopes and then reared as de-cribed above.
Treatments. Cultures of Paracentrotus lividus embryos at theastrula stage, were subjected to different treatments as follows.
0006-291X/01 $35.00Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.

Litium chloride (LiCl) treatment: eggs from fertilization to gastrulasfitg
cMipccas
ltawr
(aaea
ut
bw2bpw((aicwB
ws4
ewaslw(ibcaaa(msp
u1
R
msk
esdpmoukheesrSmp
oswa
Vol. 287, No. 5, 2001 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
tage were incubated with 20 mM LiCl; then washed twice withltered sea water and cultured in the same medium until the con-rols had achieved the pluteus stage. Heat treatment: embryos atastrula stage were cultured at 31°C for 2 h.
Cell fractionation. The embryos were suspended in 0.25M su-rose in TKV buffer (50 mM Tris–HCl buffer, pH 7.6 containing 0.4
KCl and 1 mM EDTA) with the addition of the following proteasenhibitors: 1 mM Phenylmethylsulfonylfluoride (PMSF), 5 mMaraminobenzamidine, 1 mg/ml trypsin inhibitor, 10 mM amino-apronic acid, and homogenized in Downce. The homogenate wasentrifuged at 700g for 10 min. From the supernatant mitochondriand of post mitochondrial fraction were prepared as previously de-cribed (12).
Sample preparation and electrophoretic analysis. Embryos wereysed by Dounce homogenization in O’Farrel urea lysis buffer withhe addition of protease inhibitor cocktail (Boehringer). SDS–PAGEnalysis was performed as previously reported (14): aliquots (100 mg)ere run at 80 V on 10% SDS–polyacrylamide slab gels for 4.5 h at
oom temperature.Two-dimensional gel electrophoresis was carried out as described
15). Cylindrical gels of the first dimension, containing 1.6% pH 5–7nd 0.4% pH 3–10 ampholytes (LKB), were loaded with the samemount of samples (150 mg) and run overnight at 300 V. The secondlectrophoretic dimension was performed on 10% SDS–polyacryl-mide slab gels and run for 5 h at 100 V.Molecular weights for mono- and two-dimensional gels were eval-
ated by comparison with a set of standard proteins (Rainbow pro-ein molecular weight markers, Amersham).
Western blotting. After electroforesis the proteins were electro-lotted onto nitrocellulose paper (Hybond C, or ECL, Amersham)ith a semidry apparatus (Novablot Pharmacia), at 0.8 mA/cm2 forh. The filters were then incubated with the blocking solution (3%
ovine serum albumin, 5% horse serum, 0.02% sodium azide inhosphate buffered saline solution, PBS) for 3 h, washed 3 timesith PBS and incubated overnight with the following antibodies
1:500 dilution): monoclonal anti human hsp-60 raised in miceSIGMA), or mouse monoclonal anti-actin raised in mice (SIGMA), ornti-Mn superoxide dismutase (Mn-SOD) polyclonal antibody raisedn rabbit (StressGene Biotech. Corp.). After removal of unspecificomplexes by several washings with PBS, the filters were reactedith anti mouse IgG alkaline phosphatase conjugate and stained byCIP/NBT (Sigma).
Embryo fixation. Embryos, at different developmental stages,ere fixed for 12–16 h with Bouin fixative (15:5:1 of picric acid
olution, 37% formaldehyde, glacial acetic acid, respectively), kept at°C in 70% ethanol after serial dehydratation.
Immunocytochemistry. Samples of about one to two thousandmbryos of each considered stage were analyzed for each assay. Afterashing with slow waving in 100% ethanol and then in 100% meth-nol containing 2 mM EDTA, the embryos were rehydrated witherial ethanol, washed 3 times with PBST (phosphate-buffered sa-ine containing 0.1% Tween 20) and allowed to incubate overnightith monoclonal anti-hsp60 (1:250 dilution) or monoclonal anti-actin
1:200 dilution) or with the polyclonal anti-Mn-SOD (1:250 dilution)n PBST containing 3% BSA. In control embryos the primary anti-odies were omitted, anti mouse anti IgG alkaline phosphatase (AP)onjugated (1:1000 dilution; Promega), as secondary antibody, wasdded to the samples for 1 h after 3 washes with PBST. The second-ry antibody surplus was eliminated by washing 3 times with PBSTnd then with TBS. Staining was developed for 1–3 h by NBT/BICPBoehringer Mannheim) in a buffer, containing 100 mM NaCl, 100M Tris–HCl, pH 9.5, 50 mM MgCl2. The staining reaction was
topped by two washes with 50% ethanol. Whole embryos resus-ended in 80% glycerol were finally mounted on coverslips, observed
1094
nder an Axioskop MC80 microscope (Zeiss) and photographed with00 Asa Kodak Gold film.
ESULTS AND DISCUSSION
We have previously demonstrated that sea urchinitochondrial matrix contains a constitutive and a
tress inducible protein belonging to the family of 60Da proteins, very well conserved in evolution.At first, in the aim to understand if the Hsp56 is
xpressed in the same way in both control and heathocked gastrula embryos, we compared the mitochon-rial protein patterns by two-dimensional gel electro-horesis and by Western blot analysis reacting with aonoclonal serum anti-human Hsp60 which, as previ-
usly demonstrated (2), specifically recognizes the searchin hsp56 band in Western blot. The protein of 56Da is present in both samples but apparently inigher amount in the mithocondria of heat shockedmbryos (Figs. 1a and 1b) as control, the same proteicxtracts have been reacted with the antibody anti Mn-uperoxide-dismutase (Mn-SOD) which specificallyecognizes the mitochondrial and not the cytoplasmicOD, and so far it can be considered a mitochondrialarker. Figure 1 (c and d) shows that Mn-SOD is
resent in same amount in both mitochondrial ex-
FIG. 1. Immunoblot analysis of two-dimensional electrophoresisf mitochondrial proteins purified from control (18°C, a, c) and heathocked (31°C, b, d) gastrulae reacted with the anti-Hsp60 (a, b) andith the anti- SOD (c, d) antisera. Molecular weight markers in kDare indicated on the right side of the figure.

tstssadttcpattc
pdtesrHTcguni
ferent embryonic stages. During the early developmen-twr
h4Sbo
m4garsp
Vol. 287, No. 5, 2001 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
racts. On the other hand, we have previously demon-trated that following the heat shock the synthesis ofhe hsp56 increases. We can then suppose that in re-ponse to heat shock, the hsp56 is not only largelyynthesized but soon imported into the organelles. Inddition, the protein is present in the postmitochon-rial supernatant of gastrula shocked embryos, besidehe mitochondrial compartment; whereas it is only athe mitochondrial level and not in the cytoplasm of theontrol gastrulae (Fig. 2a). On the contrary SOD isresent in the mitochondria in both extracts but it isbsolutely absent from the postmitochondrial superna-ant (Fig. 2b), suggesting that the different distribu-ion of the hsp56 is not due to some change of mito-hondria permeability induced by the shock.
As far as the constitutive hsp is concerned, 56 kDa isresent in the same amount in eggs and embryos atifferent developmental stages (Fig. 3a). On the con-rary, the amount of actin increases during embryogen-sis, as demonstrated by the reaction with the anti-erum anti-actin (Fig. 3b). Figure 3c summarizes theesults of the present experiments, highlighting thatsp56 is constant while actin concentration increases.hese data can be explained considering that mito-hondria are synthesized and accumulated during oo-enesis, and no new synthesis has been demonstratedntil later stages of embryo development (13) when cellumber increases. On the basis of the these results, we
nvestigated the chaperonin localization in situ in dif-
FIG. 2. Western blot analysis of mitochondrial, (control, 1, andeat shocked, 2) and postmitochondrial (control, 3, and heat shocked,) extracts reacted with the anti-Hsp60 (a) and with the anti-Mn-OD (b) antibodies; the positions of the hsp56 and SOD are indicatedy the arrows. The molecular weight markers in kDa are indicatedn the right.
1095
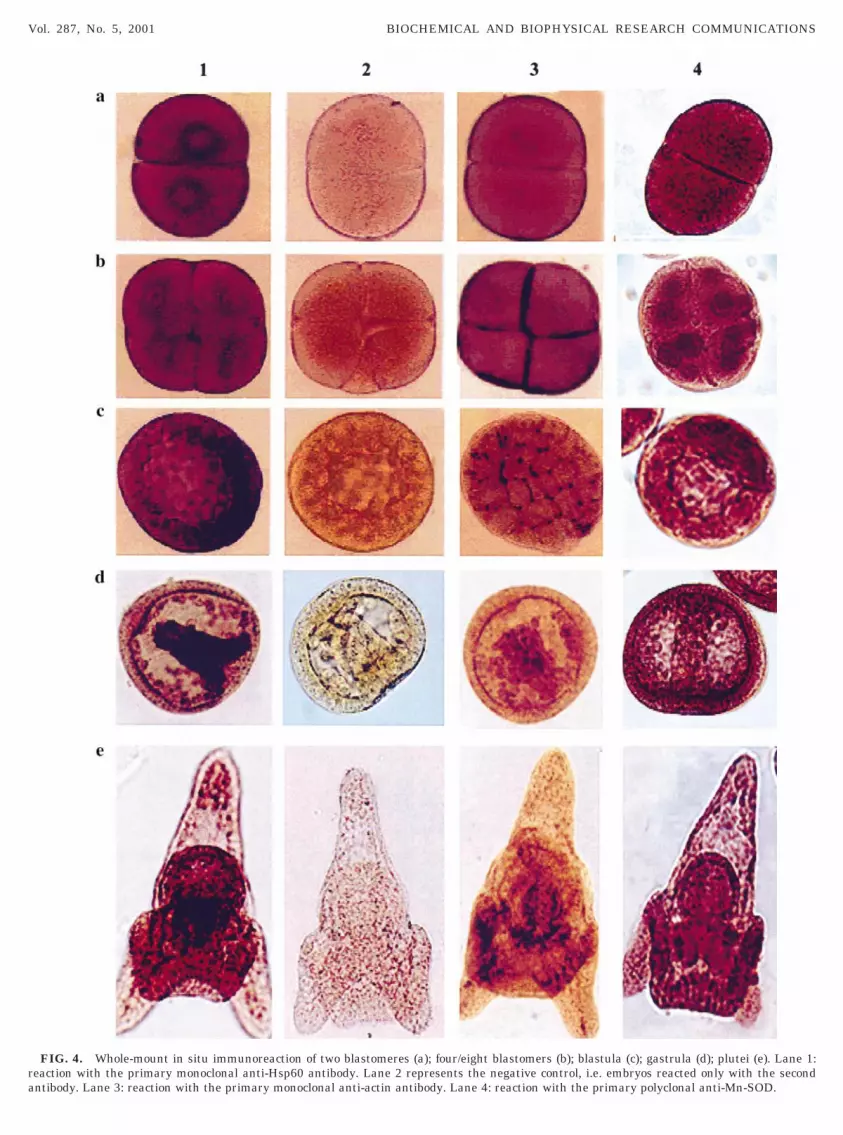
al stages, the chaperonin has a uniform distributionith a preferential accumulation in the perinuclear
egion (Fig. 4, lane 1, a, b, c). At later stages, i.e.,
FIG. 3. Western blot analysis of proteins from different develop-ental stages (lane 2, unfertilized eggs; lane 3, two blastomers; lane
, four/eight blastomers; lane 5, morulae; lane 6, blastulae; lane 7,astrulae; lane 8, plutei), reacted with anti-Hsp60 antibody (a); withnti-actin antibody (b). The histogram shown in c summarizes theesults (normalized per number of cells) obtained by densitometriccanning of the filters shown in a and b (Jandel Sigma Scanrogram).
ra
Vol. 287, No. 5, 2001 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 4. Whole-mount in situ immunoreaction of two blastomeres (a); four/eight blastomers (b); blastula (c); gastrula (d); plutei (e). Lane 1:eaction with the primary monoclonal anti-Hsp60 antibody. Lane 2 represents the negative control, i.e. embryos reacted only with the secondntibody. Lane 3: reaction with the primary monoclonal anti-actin antibody. Lane 4: reaction with the primary polyclonal anti-Mn-SOD.

gcadae(bilosdTte
lithium ions, an efficient inducer of vegetalization ofsclsha6Ssb
ttIaTdephtabe
tohacp
rt
s
Vol. 287, No. 5, 2001 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
astrula and plutei (Fig. 4, lane 1, d, e) it is moreoncentrated in some territories, like mesenchime andrchenteron. The distribution of actin is completelyifferent: in fact it is localized in the cleavage planes,nd therefore in the intercellular boundaries, duringarly development, and diffused in the later stagesFig. 4, lane 3); while SOD is distributed in all thelastomers, more concentrated around the nuclei dur-ng cleavage (Fig. 4, lane 4, a, b, c) and quite uniform inater stages, with a preferential accumulation in theral arms and intestine of pluteus (Fig. 4, lane 4, d, e),uggesting the specificity of the antibodies and in ad-ition a quite uniform distribution of mitochondria.he fact that the Hsp56 is preferentially localized inhe endoderm is confirmed by the observation of thexogastrulae, obtained incubating early embryos with
FIG. 5. Whole-mount in situ immunoreaction of an exogastrulaeacted with the primary monoclonal anti-Hsp60 (a). (b) The nega-ive control.
FIG. 6. In situ immunoreaction of heat shocked embryos reactedtages.
1097
ea urchin embryos (16), in which the majority of thehaperonin is in the endoderm (Fig. 5). As concerns theocalization of the Hsp56 during the heat shock re-ponse, the observations of whole mounts show that ineat shocked embryos the protein increases and alsoppears in other embryo territories, like ectoderm (Fig.a; see also Fig. 4, lane 1, d, e as control) while Mn-OD (Fig. 6b) is not redistributed remaining in theame amount and localization as in the control em-ryos (compare to Fig. 5, lane 4, d, e).On the basis of these results we can conclude that
he constitutive Hsp56 chaperonin is synthesized andranslocated into mitochondria, where it accumulates.t shows a precise cellular localization during cleavage,nd a territorial distribution in later development.his could be explained by admitting that mitochon-ria are distributed in homogeneous way inside thembryos, but that the protein is differentially ex-ressed in different embryo territories. Following theeat shock, the Hsp56 is newly synthesized, it appearso be accumulated in all embryo territories, and itppears in the cytoplasm possibly because it has noteen yet imported or because it has been specificallyxported from the organelles.The implication of the subcellular localization and
he involvement of mitochondrial molecular chaper-nes in diverse cellular processes, have been recentlyypothesized (17, 18), suggesting important functionst specific extramitochondrial sites, particularly at theell surface. Therefore, in our conditions, we may hy-othesize that the protein is synthesized in higher
th anti-Hsp60 (a) and anti-Mn-SOD (b). 1, gastrula, and 2, pluteus
wi
amount and distributed de novo also in the ectoderm inrf
A
Mw9
R
raad, N. J. (1997) The role of molecular chaperones in mitochon-
1
1
1
1
1
1
1
1
1
Vol. 287, No. 5, 2001 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
esponse to heat shock, in order to protect the embryosrom environmental insults.
CKNOWLEDGMENTS
We thank G. Giudice for critical reading of the manuscript and G.orici for the computing assistance and data elaboration. This workas supported by grants from MURST and CNR (60% and Stress No.9.02499).
EFERENCES
1. Gianguzza, F., Ragusa, M. A., Roccheri, M. C., Di Liegro, I., andRinaldi, A. M. (2000) Isolation and caracterization of a Paracen-trotus lividus cDna encoding a stress-inducible chaperonin. CellStress Chaperones 5, 87–89.
2. Roccheri, M. C., Bosco, L., Ristuccia, M. E., Cascino, D., Giudice,G., Oliva, A. O., and Rinaldi, A. M. (1997) Sea urchin mitochon-drial matrix containins a 56-kDa chaperonine-like protein. Bio-chem. Biophys. Res. Commun. 234, 646–650.
3. Ellis, R. J., and van der Vies, S. M. (1991) Molecular chaperones.Ann. Rev. Biochem. 60, 321–347.
4. Craig, E. A., Gambill, B. D., and Nelson, R. J. (1993) Heat shockproteins: Molecular chaperones of protein biogenesis. Microbiol.Rev. 57, 402–414
5. Ellis, R. J., and Hartl, F. U. (1999) Priciples of protein folding inthe cellular environment. Curr. Opin. Struct. Biol. 9, 102–110.
6. Hartl, F. U. (1996) Molecular chaperones in protein folding.Nature 381, 571–580.
7. Soltys, B. J., and Gupta, R. S. (1999) Mitochondrial molecularchaperones hsp60 and msh70. In Stress Proteins (Latchman,D. S., Ed.), pp. 68–100, Springer.
8. Ryan, M. T., Naylor, D. J., Hoj, P. B., Clark, M. S., and Hoogen-
1098
drial protein import and folding. Int. Rev. Cytol. 174, 127–193.9. Pfanner, N., Craig, E. A., and Maijer, M. (1994) The protein
import machinery of the mitochondrial inner membrane. Tibs.19, 368–372.
0. Gupta, R. S. (1995) Evolution of the chaperonin families (hsp60,hsp10 and Tcp-) of proteins and the origin of eukaryotic cells.Mol. Microbiol. 15, 1–11.
1. Rinaldi, A. M., De Leo, G., Arzone, A., Salcher-Cillari, I., andMutolo, V. (1979) Biochemical and electron microscopic evidencethat cell nucleus negatively controls genomic activivty in earlysea urchin development. Proc. Natl. Acad. Sci. USA 76, 1916.
2. Rinaldi, A. M., Carra, E., Salcher-Cillari, I., and Oliva, O. A.(1983) The nucleus negatively controls the synthesis of mito-chondrial proteins in the sea urchin egg. Cell Biol. Int. Rep. 7,211–218.
3. Rinaldi, A. M., and Giudice, G (1985) Nuclear Interaction inEarly Development (Metz and Monroy, A., Eds.), pp.267–377,Academic Press Inc.
4. Casano, C., Roccheri, M. C., Onorato, K., Cascino, D., and Gian-guzza, F. (1998) Deciliation: A stressful event for Paracentrotuslividus embryos. Biochem. Biophys. Res. Commun. 248, 628–634.
5. Roccheri, M. C., Cascino, M., and Giudice, G. (1993) Two dimen-sional electrophoretic analisis of stress protein in P. lividus. J.Submicrosc. Cytol. Pathol. 25, 173–179.
6. Giudice, G. (1986) The sea urchin embryo, Springer-Verlag, NewYork.
7. Poccia, F., Piselli, P., Vendetti, S., Bach, S., Amendola, A.,Placido, R., and Colizzi, V. (1996) Heat-shock protein expressionon the membrane of T cells undergoing apoptosis. Immunology88, 6–12.
8. Soltys, B. J., and Gupta, R. S. (1997) Cell surface localization ofthe 60-kDa heat shock chaperonin protein (hsp60) in mamma-lian cells. Cell Biol. Int. 21, 315–320.





























