Embed Size (px)
Citation preview

This article was downloaded by: [117.174.231.191]On: 25 March 2014, At: 10:51Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Bolletino di zoologiaPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tizo19
Microarchitecture and mechanics of the sea‐urchinperistomial membraneIain C. Wilkie a , M.Daniela Candia Carnevali b & Francesco Andrietti ba Department of Biological Sciences , Glasgow Caledonian University , 70 CowcaddensRoad, Glasgow, G4 0BA, Scotlandb Dipartimento di Biologia ≪Luigi Gorini≫ , Università degli Studi di Milano , viaCeloria 26, Milano, I‐20133, ItalyPublished online: 28 Jan 2009.
To cite this article: Iain C. Wilkie , M.Daniela Candia Carnevali & Francesco Andrietti (1994) Microarchitectureand mechanics of the sea‐urchin peristomial membrane, Bolletino di zoologia, 61:1, 39-51, DOI:10.1080/11250009409355857
To link to this article: http://dx.doi.org/10.1080/11250009409355857
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”)contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensorsmake no representations or warranties whatsoever as to the accuracy, completeness, or suitability for anypurpose of the Content. Any opinions and views expressed in this publication are the opinions and viewsof the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sources of information. Taylorand Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses,damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connectionwith, in relation to or arising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Boll. Zool. 61: 39-51 (1994)
Microarchitecture and mechanics ofthe sea-urchin peristomial membrane
IAIN C. WILKIEDepartment of Biological Sciences, Glasgow Caledonian University,70 Cowcaddens Road, Glasgow G4 0BA, Scotland
M. DANIELA CANDIA CARNEVALIFRANCESCO ANDRIETTIDipartimento di Biologia «Luigi Gorini»,Università degli Studi di Milano,via Celoria 26, I-20133 Milano (Italy)
ABSTRACT
The peristomial membrane is an area of flexible body wall whichconnects the test to the lantern or masticatory apparatus of regularsea-urchins. The peristomial membrane of Paracentrotus lividuswas examined by various light microscopical techniques and byscanning and transmission electron microscopy, and its mechanicalproperties were investigated by load-deformation tests. Attentionwas focused on the collagenous dermis which consists mainly ofcrossed-fibre arrays arranged in three histologically distinctsublayers. This microarchitectural arrangement gives rise to non-linear load-deformation characteristics: when subjected to verticaldeformation such as would occur during lantern protraction, theperistomial membrane shows firstly low stiffness and then high stiff-ness phases. The response of the peristomial membrane to chemicalagents suggests that the tensile properties of the dermis are underphysiological control. Our results indicate that the collagenous fibresof the dermis belong to separate populations whose stiffness can beadjusted differentially to meet changing functional demands on theperistomial membrane.
KEY WORDS: Sea-urchin - Peristomial membrane - Collagenoustissue - Microarchitecture - Mechanical properties.
ACKNOWLEDGEMENTS
The research reported in this paper was funded by the ConsiglioNazionale delle Ricerche, Rome and by the Royal Society of London.It benefited from the assistance of F. Bonasoro, E. Lucca and G.Melone.
(Received 7 September 1993 - Accepted 17 September 1993)
INTRODUCTION
Collagenous connective tissues are ubiquitous in thebodies of animals where they function primarily as struc-tural materials providing, amongst other functions,protection, support, elastic energy storage and stresstransfer. It is widely recognized that the collagenoustissues of vertebrates show a broad diversity oforganization at the light microscope level which can becorrelated with their mechanical properties and specificfunctions: dermis, tendon, ligament, and arterial wall arewell-studied examples (see e.g., Wilkes et al., 1973;Evans & Barbenel, 1975; Nordin & Frankel, 1980; Gib-bons & Shadwick, 1991). It is also true but less a matter ofcommon knowledge that echinoderm collagenoustissues exhibit just as great a range of microarchitectures:parallel fibre arrays, crossed-helical arrays, orthogonalnetworks and random feltworks have all been observed(Wilkie, 1984). This heterogeneity may be related to thepresence in echinoderms, as in vertebrates, of an exten-sive endoskeleton.
Most collagenous structures have as their basic struc-tural units submicroscopic cross-striated collagen fibrilswhich are believed to show the same properties of hightensile strength and stiffness across the Animal Kingdom.However, partly as a result of variations in microarchitec-ture, i.e., in the grouping of fibrils into larger fibres andin the conformation and spatial arrangement of thefibres, different collagenous structures can exhibit verydifferent mechanical properties and therefore fulfil dif-ferent functional requirements (compare, for example,mammalian tendon and dermis, and see Fung, 1984).Despite the considerable interest that echinodermcollagenous tissues have attracted over the past twentyyears, the relationship between their microscopicorganization and their «conventional» biomechanics hasbeen investigated only rarely (Woodley, 1967; O'Neill,1989). The reason for this neglect has been the distrac-tion caused by the apparently unique capacity of someechinoderm collagenous tissues to undergo drastic,neurally modulated changes in mechanical propertieswithin a timescale of seconds or less, a phenomenonknown as variable tensility. The biochemistry, phar-macology and mechanics of these mutable collagenoustissues (MCTs) have been the subject of extensive resear-ch during the course of which their structural diversityhas been revealed almost incidentally (see reviews byMotokawa, 1984a; Wilkie, 1984; Wilkie & Emson, 1988;Candia Carnevali & Wilkie, 1992). The presence of MCTshas been confirmed in all echinoderm classes except therecently discovered Concentricycloidea, for which nodata are as yet available. With regard to sea-urchin MCTs,most research has focused on the capsular ligament (orcatch apparatus) of the spine-test joint (Hidaka &Takahashi, 1983; Diab & Gilly, 1984; Morales etal, 1989;Trotter & Koobi 1989), and within the specific context ofthe sea-urchin lantern, or masticatory apparatus, Wilkieet al. (1992) have shown that the compass depressor
Dow
nloa
ded
by [
117.
174.
231.
191]
at 1
0:51
25
Mar
ch 2
014

40 I. C. WILKIE, M. D. CANDIA CARNEVALI, F. ANDRIETTI
ligaments consist of MCT and Birenheide (1990) hassuggested that the ligaments connecting the teeth to thejaws are mutable.
The peristomial membrane (PM) is another mainlycollagenous structure that is linked physically and func-tionally to the sea-urchin lantern. It is an area of flexiblebody wall surrounding the mouth and connecting thelantern to the edge of the heavily calcified test (Fig. 1).We have already given a detailed account of theultrastructure of the PM (Candia Carnevali et al., 1990)and in a study of its mechanical properties we provided
cdP9
PP ap
Fig. 1 - Semidiagrammatic drawing of the lantern and associatedstructures of P. lividus. Two jaws, part of the perignathic girdle (pg)and part of the peristomial membrane (pm) have been removed; themuscles and compass depressors have been omitted from the leftside, ap, ambulacral plate (ligamentous attachment to jaw notshown); cd, compass depressor; ce, compass elevator; ja, jaw; lr, lipregion; pp, pedicellarial base-plate; pr, protractor muscle; re, retrac-tor muscle; t, tooth.
evidence that its thick dermal layer consists of MCT(Wilkie et al., 1993). The latter investigation showed thatas well as behaving like a typical non-linear viscoelasticbiomaterial, the PM can also undergo reversible stiffeningor softening, which serves to lock the lantern in positionor enhance its mobility. Apart from work by Burke et al.(1989) on the ultrastructure and immunological charac-terization of sea-urchin collagen which includes data onthe PM, and speculations by Ellers & Telford (1992) onthe relationship between internal test pressure and PMgeometry, there is little other mention of the PM in therecent literature.
In the present paper we describe and attempt tocorrelate the microarchitecture of the PM and its basicmechanical properties both when it is acting like a nor-mal biomaterial and when it is displaying variable ten-sility.
MATERIALS AND METHODS
All of the morphological and experimental studies described inthis paper were conducted on Paracentrotus lividus (Lam.) from theLigurian coast of Italy. Observations on the behaviour of intactanimals were made using both this species and the relatedechinacean sea-urchins Echinus esculentus L. and Psammechinusmiliaris (Gmelin) from the Firth of Clyde, Scotland.
Morphological investigations
The microarchitecture of the collagenous dermis of theperistomial membrane (PM) was investigated by means of fourtechniques.
(1) Brightfield light microscopyMaterial was fixed in 70% ethanol alone, Bouin's solution or
neutral buffered formalin and embedded in paraffin wax directly orafter décalcification in saturated EDTA (pH adjusted to 7.0 with KOHpellets). Sections 8 urn thick were stained by Milligan's trichromemethod (Humason, 1979). Two fixed specimens were not embeddedbut stained whole in 1% aniline blue for 15 min before beingdehydrated, cleared in xylene and mounted in DPX.
(2) Polarized light microscopyWhole specimens of the PM were fixed in 70% ethanol,
dehydrated, cleared in xylene and mounted in DPX. They wereexamined between crossed polarizers in an Olympus BH-2microscope equipped with Nomarski optics for differential inter-ference contrast and with a PM-10ADS photomicrographic system.
(3) Scanning electron microscopyA whole specimen was partly digested using 0.1 N NaOH solution,
fixed in a 2% glutaraldehyde - 4% paraformaldehyde mixture in 0.1M cacodylate buffer and post-fixed in 1 % osmic acid in the same buf-fer. It was critical point dried, gold-coated and observed in a Cam-bridge Stereoscan 250 MK2.
(4) Transmission electron microscopyUndigested specimens were fixed as in (3) above, prestained with
1% uranyl acetate in 25% ethanol for 2 h, dehydrated in an ethanolseries and embedded in an Epon-Araldite mixture. Ultrathin sectionswere stained with uranyl acetate and lead citrate, and observed in aJeol 100SX electron microscope.
Mechanical investigations
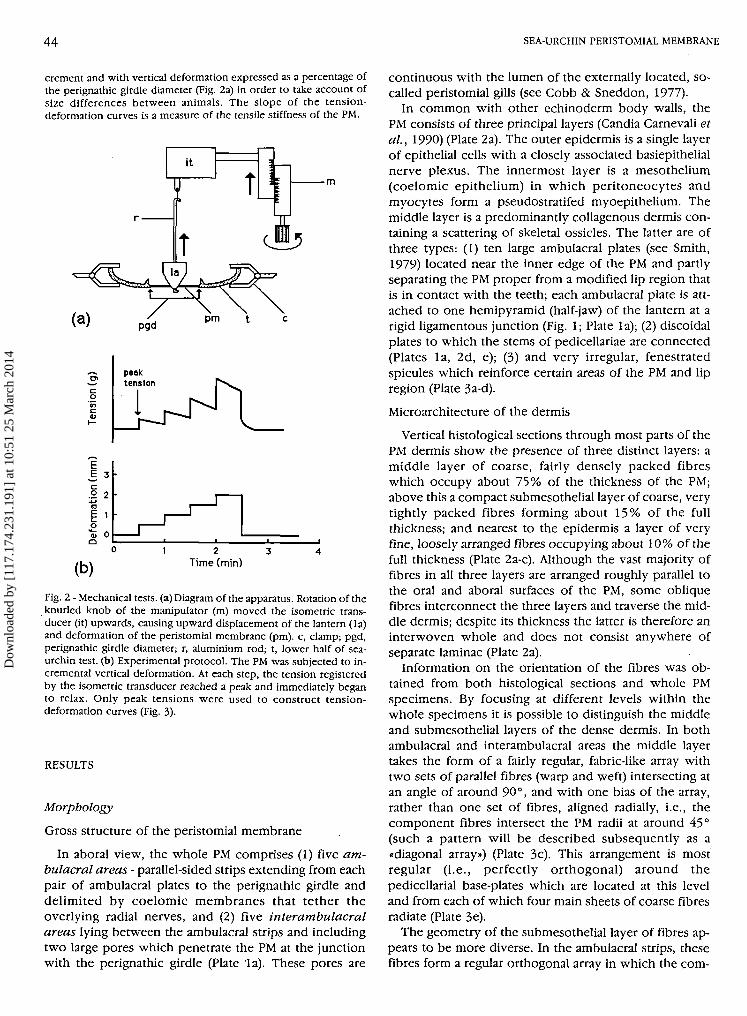
For mechanical testing, whole PM preparations were made byremoving the top half of the test and cutting all compass depressorsand retractor, protractor and compass elevator muscles (Fig. 1). Thisleft the lantern connected to the PM only at strong ligamentous junc-tions between the jaws and the ambulacral plates which are embed-ded in the PM close to the mouth. The half-test was clamped rigidlyin the horizontal position and the PM was linked to an isometricforce transducer via an aluminium rod inserted down the pharynx ofthe lantern and between the teeth (Fig. 2a). In such preparations theinterpyramidal muscles which interconnect the jaws were left intact.To avoid artefacts caused by contraction of these muscles, chemicalagents were tested on preparations from which the lantern had beenremoved and in which the PM was connected directly to the trans-ducer, as described in detail by Wilkie et al. (1993). The isometrictransducer was mounted on the moving section of a manipulatorwhich was used to raise it by 0.5 or 1.0 mm increments every 15, 30,or 60 sec, thus subjecting the PM to vertical deformation (Fig. 2b).Depending on whether the preparation was orientated with its lowersurface facing downwards (as in vivo and as shown in Fig. 2a) or up-wards, this procedure subjected the PM to retraction or protractionrespectively. The output from the transducer, which represented theresistance of the PM to imposed deformation, was recorded on anoscillograph. Tension-deformation curves were constructed usingvalues for the peak tension generated by each deformational in-
Dow
nloa
ded
by [
117.
174.
231.
191]
at 1
0:51
25
Mar
ch 2
014
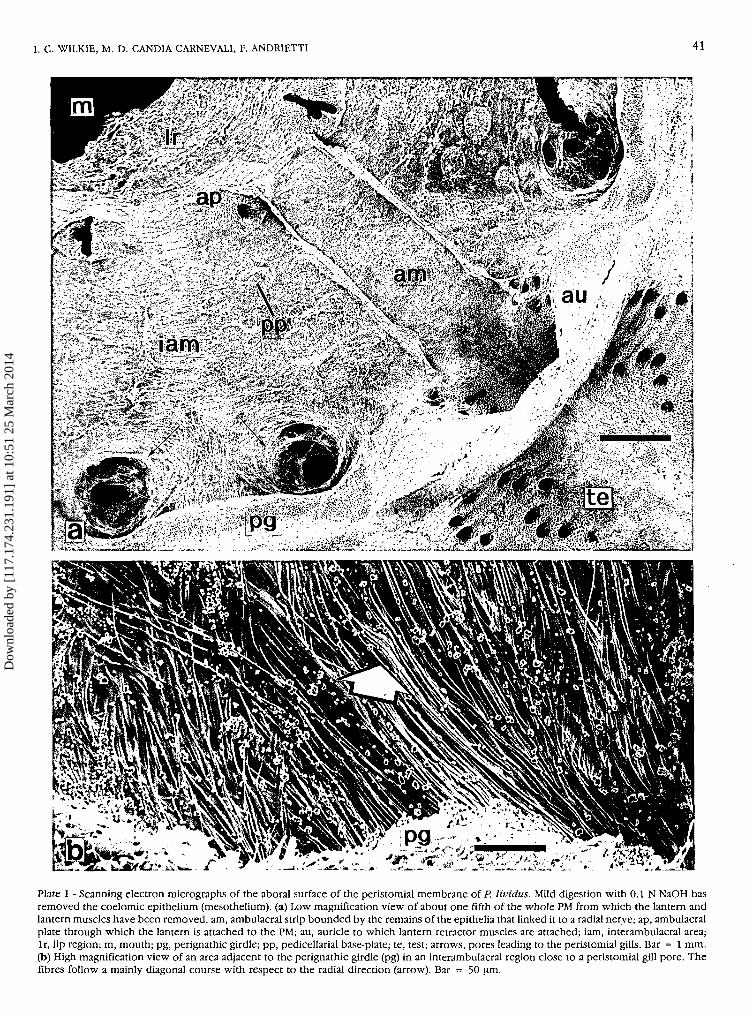
I. C. WILKIE, M. D. CANDIA CARNEVALI, F. ANDRIETTI 41
Wmm^«zC£* ^ '
Plate 1 - Scanning electron micrographs of the aboral surface of the peristomial membrane of P. lividus. Mild digestion with 0.1 N NaOH hasremoved the coelomic epithelium (mesothelium). (a) Low magnification view of about one fifth of the whole PM from which the lantern andlantern muscles have been removed, am, ambulacral strip bounded by the remains of the epithelia that linked it to a radial nerve; ap, ambulacralplate through which the lantern is attached to the PM; au, auricle to which lantern retractor muscles are attached; iam, interambulacral area;lr, lip region; m, mouth; pg, perignathic girdle; pp, pedicellarial base-plate; te, test; arrows, pores leading to the peristomial gills. Bar = 1 mm.(b) High magnification view of an area adjacent to the perignathic girdle (pg) in an interambulacral region close to a peristomial gill pore. Thefibres follow a mainly diagonal course with respect to the radial direction (arrow). Bar = 50 um.
Dow
nloa
ded
by [
117.
174.
231.
191]
at 1
0:51
25
Mar
ch 2
014
r~
• • • • • ' • • • • • ^
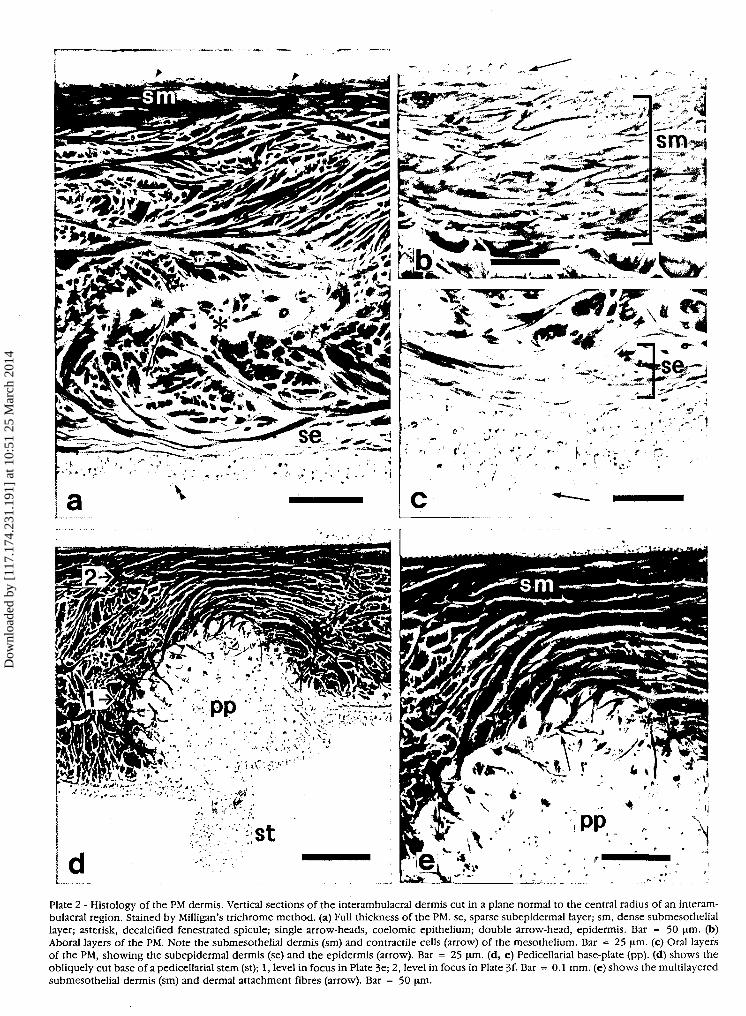
Plate 2 - Histology of the PM dermis. Vertical sections of the interambulacral dermis cut in a plane normal to the central radius of an interam-bulacral region. Stained by Milligan's trichrome method, (a) Full thickness of the PM. se, sparse subepidermal layer; sm, dense submesotheliallayer; asterisk, decalcified fenestrated spicule; single arrow-heads, coelomic epithelium; double arrow-head, epidermis. Bar = 50 um. (b)Aboral layers of the PM. Note the submesothelial dermis (sm) and contractile cells (arrow) of the mesothelium. Bar = 25 um. (c) Oral layersof the PM, showing the subepidermal dermis (se) and the epidermis (arrow). Bar = 25 um. (d, e) Pedicellarial base-plate (pp). (d) shows theobliquely cut base of a pedicellarial stem (st); 1, level in focus in Plate 3e; 2, level in focus in Plate 3f- Bar = 0.1 mm. (e) shows the multilayeredsubmesothelial dermis (sm) and dermal attachment fibres (arrow). Bar = 50 \im.
Dow
nloa
ded
by [
117.
174.
231.
191]
at 1
0:51
25
Mar
ch 2
014
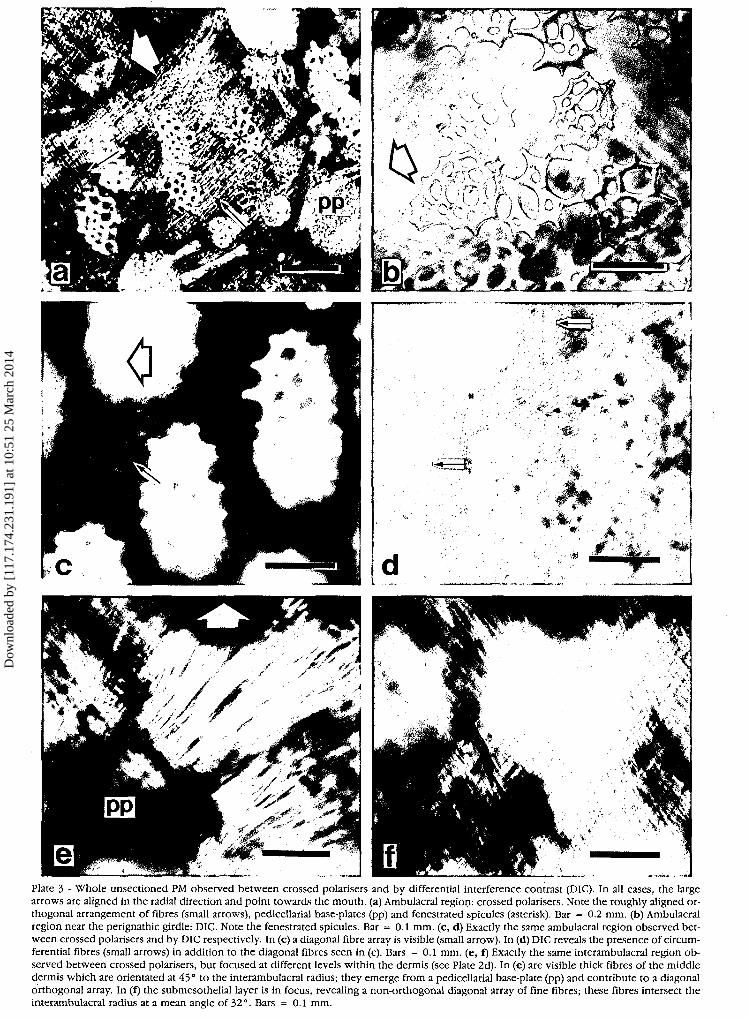
Plate 3 - Whole unsectioned PM observed between crossed polarisers and by differential interference contrast (DIC). In all cases, the largearrows are aligned in the radial direction and point towards the mouth, (a) Ambulacral region: crossed polarisers. Note the roughly aligned or-thogonal arrangement of fibres (small arrows), pedicellarial base-plates (pp) and fenestrated spicules (asterisk). Bar = 0.2 mm. (b) Ambulacralregion near the perignathic girdle: DIC. Note the fenestrated spicules. Bar = 0.1 mm. (c, d) Exactly the same ambulacral region observed bet-ween crossed polarisers and by DIC respectively. In (c) a diagonal fibre array is visible (small arrow). In (d) DIC reveals the presence of circum-ferential fibres (small arrows) in addition to the diagonal fibres seen in (c). Bars = 0.1 mm. (e, f) Exactly the same interambulacral region ob-served between crossed polarisers, but focused at different levels within the dermis (see Plate 2d). In (e) are visible thick fibres of the middledermis which are orientated at 45° to the interambulacral radius; they emerge from a pedicellarial base-plate (pp) and contribute to a diagonalorthogonal array. In (f) the submesothelial layer is in focus, revealing a non-orthogonal diagonal array of fine fibres; these fibres intersect theinterambulacral radius at a mean angle of 32°. Bars = 0.1 mm.
Dow
nloa
ded
by [
117.
174.
231.
191]
at 1
0:51
25
Mar
ch 2
014
44 SEA-URCHIN PERISTOMIAL MEMBRANE
crement and with vertical deformation expressed as a percentage ofthe perignathic girdle diameter (Fig. 2a) in order to take account ofsize differences between animals. The slope of the tension-deformation curves is a measure of the tensile stiffness of the PM.
E 3
§2«3E ioto 0
(b)1 2 3
Time (min)
Fig. 2 - Mechanical tests, (a) Diagram of the apparatus. Rotation of theknurled knob of the manipulator (m) moved the isometric trans-ducer (it) upwards, causing upward displacement of the lantern (la)and deformation of the peristomial membrane (pm). c, clamp; pgd,perignathic girdle diameter; r, aluminium rod; t, lower half of sea-urchin test, (b) Experimental protocol. The PM was subjected to in-cremental vertical deformation. At each step, the tension registeredby the isometric transducer reached a peak and immediately beganto relax. Only peak tensions were used to construct tension-deformation curves (Fig. 3).
RESULTS
Morphology
Gross structure of the peristomial membrane
In aboral view, the whole PM comprises (1) five am-bulacral areas - parallel-sided strips extending from eachpair of ambulacral plates to the perignathic girdle anddelimited by coelomic membranes that tether theoverlying radial nerves, and (2) five interambulacralareas lying between the ambulacral strips and includingtwo large pores which penetrate the PM at the junctionwith the perignathic girdle (Plate la). These pores are
continuous with the lumen of the externally located, so-called peristomial gills (see Cobb & Sneddon, 1977).
In common with other echinoderm body walls, thePM consists of three principal layers (Candia Carnevali etal., 1990) (Plate 2a). The outer epidermis is a single layerof epithelial cells with a closely associated basiepithelialnerve plexus. The innermost layer is a mesothelium(coelomic epithelium) in which peritoneocytes andmyocytes form a pseudostratifed myoepithelium. Themiddle layer is a predominantly collagenous dermis con-taining a scattering of skeletal ossicles. The latter are ofthree types: (1) ten large ambulacral plates (see Smith,1979) located near the inner edge of the PM and partlyseparating the PM proper from a modified lip region thatis in contact with the teeth; each ambulacral plate is att-ached to one hemipyramid (half-jaw) of the lantern at arigid ligamentous junction (Fig. 1; Plate la); (2) discoidalplates to which the stems of pedicellariae are connected(Plates la, 2d, e); (3) and very irregular, fenestratedspicules which reinforce certain areas of the PM and lipregion (Plate 3a-d).
Microarchitecture of the dermis
Vertical histological sections through most parts of thePM dermis show the presence of three distinct layers: amiddle layer of coarse, fairly densely packed fibreswhich occupy about 75% of the thickness of the PM;above this a compact submesothelial layer of coarse, verytightly packed fibres forming about 15% of the fullthickness; and nearest to the epidermis a layer of veryfine, loosely arranged fibres occupying about 10% of thefull thickness (Plate 2a-c). Although the vast majority offibres in all three layers are arranged roughly parallel tothe oral and aboral surfaces of the PM, some obliquefibres interconnect the three layers and traverse the mid-dle dermis; despite its thickness the latter is therefore aninterwoven whole and does not consist anywhere ofseparate laminae (Plate 2a).
Information, on the orientation of the fibres was ob-tained from both histological sections and whole PMspecimens. By focusing at different levels within thewhole specimens it is possible to distinguish the middleand submesothelial layers of the dense dermis. In bothambulacral and interambulacral areas the middle layertakes the form of a fairly regular, fabric-like array withtwo sets of parallel fibres (warp and weft) intersecting atan angle of around 90°, and with one bias of the array,rather than one set of fibres, aligned radially, i.e., thecomponent fibres intersect the PM radii at around 45°(such a pattern will be described subsequently as a«diagonal array») (Plate 3c). This arrangement is mostregular (i.e., perfectly orthogonal) around thepedicellarial base-plates which are located at this leveland from each of which four main sheets of coarse fibresradiate (Plate 3e).
The geometry of the submesothelial layer of fibres ap-pears to be more diverse. In the ambulacral strips, thesefibres form a regular orthogonal array in which the com-
Dow
nloa
ded
by [
117.
174.
231.
191]
at 1
0:51
25
Mar
ch 2
014
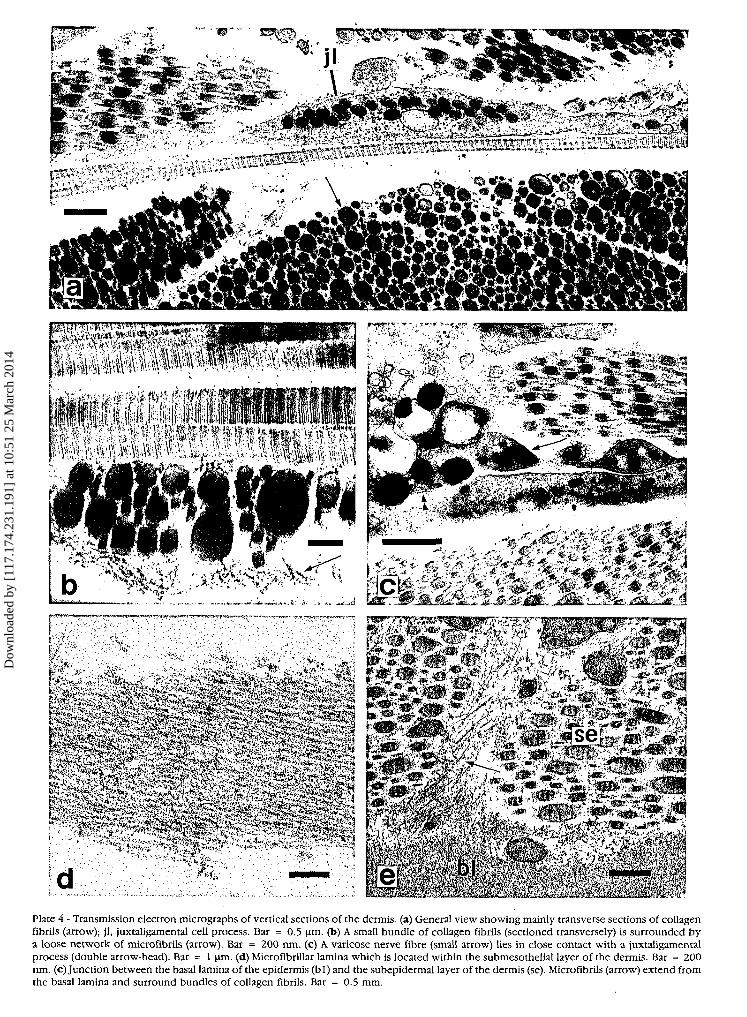
Plate 4 - Transmission electron micrographs of vertical sections of the dermis. (a) General view showing mainly transverse sections of collagenfibrils (arrow); jl, juxtaligamental cell process. Bar = 0.5 \im. (b) A small bundle of collagen fibrils (sectioned transversely) is surrounded bya loose network of microfibrils (arrow). Bar = 200 nm. (c) A varicose nerve fibre (small arrow) lies in close contact with a juxtaligamentalprocess (double arrow-head). Bar = 1 nm. (d) MicrofibriUar lamina which is located within the submesothelial layer of the dermis. Bar = 200nm. (e)Junction between the basal lamina of the epidermis (bl) and the subepidermal layer of the dermis (se). Microfibrils (arrow) extend fromthe basal lamina and surround bundles of collagen fibrils. Bar = 0.5 mm.
Dow
nloa
ded
by [
117.
174.
231.
191]
at 1
0:51
25
Mar
ch 2
014

46 I. C. WILKIE, M. D. CANDIA CARNEVALI, F. ANDRIETTI
ponent fibres are aligned radially and circumferentially(which will be described subsequently as an «alignedarray») (Plate 3a, d, f). This also seems to be thepredominant orientation of the submesothelial fibres inthe interambulacral regions near the ambulacral strips,but not elsewhere in the interambulacra where diagonalarrays are apparent, as revealed by scanning electronmicroscopy of the partly digested specimen (Plate lb).
Vertical and horizontal histological sections suggest thatthroughout the PM the subepidermal fibres are arranged ina regular orthogonal, diagonal array (Plate 2a, c).
The whole DPX-mounted PM preparations which hadnot been decalcified or stained and the SEM preparationalso provided information on the distribution of ossiclesin the PM. The pedicellarial base-plates (Plate 2d, e) arescattered evenly throughout the interambulacral areasand along the edges of the ambulacral strips (Plates la,3a). It is noticeable, however, that the irregularfenestrated spicules (Plate 2a) are most abundant in theambulacral areas and at the outer edge of the PM near theperignathic girdle (Plate 3a, b). Few fenestrated spiculesare observed in the interambulacral areas away from theouter edge.
In the transmission electron microscope the fibres atall levels of the dermis appear as parallel aggregations ofcross-banded fibrils with a periodicity of ca. 65 nm.Transverse sections of the fibres reveal that the diameterof their component fibrils is always very variable,reaching a maximum of about 380 nm, and that the fibrilsare usually tightly packed leaving little interfibrillar space(Plate 4a). Frequently the fibrils appear to be interconnect-ed by fine microfibrils (diameter ca. 12 nm) which alsosometimes form networks which surround and separatethe aggregations (Plate 4b). However, these microfibrilsare not always visible.
Within the submesothelial layer there is a distinct sheetof densely packed microfibrils which are also ca. 12 nmin diameter (Plate 4d). Semithin araldite sections indicatethat this sheet is highly fenestrated and that it is restrictedto the interambulacral regions.
Cellular elements are very sparse in the middle dermisbut more abundant in the subepidermal layer and in thesubmesothelial region above the microfibrillar lamina;they include possible fibroblasts, varicose cell processeswith small granules (maximum diameter 190 nm) whichare presumably axons, and cell bodies and processes con-taining large electron-dense granules (maximumdiameters 850 and 300 nm in the cell bodies andprocesses respectively) (Plate 4a) which are found oc-casionally in close contact with the varicose axon-likeprocesses (Plate 4c). The large granule containing cellbodies seem to be restricted to the loose subepidermaland submesothelial layers near their respective junctionswith the dense dermis, whereas the large granule con-taining processes are distributed sparsely throughout thedense dermis in the electron-lucent regions between thefibril aggregations (i.e., not inside the fibril aggregations).
The TEM also reveals the nature of the junctions be-
tween the dermis and two cellular layers that cover it.Networks of microfibrils that surround collagen fibresclose to the epidermis or mesothelium extend towards,and penetrate, the basal lamina of the cellular layers(Plate 4e).
Mechanical properties
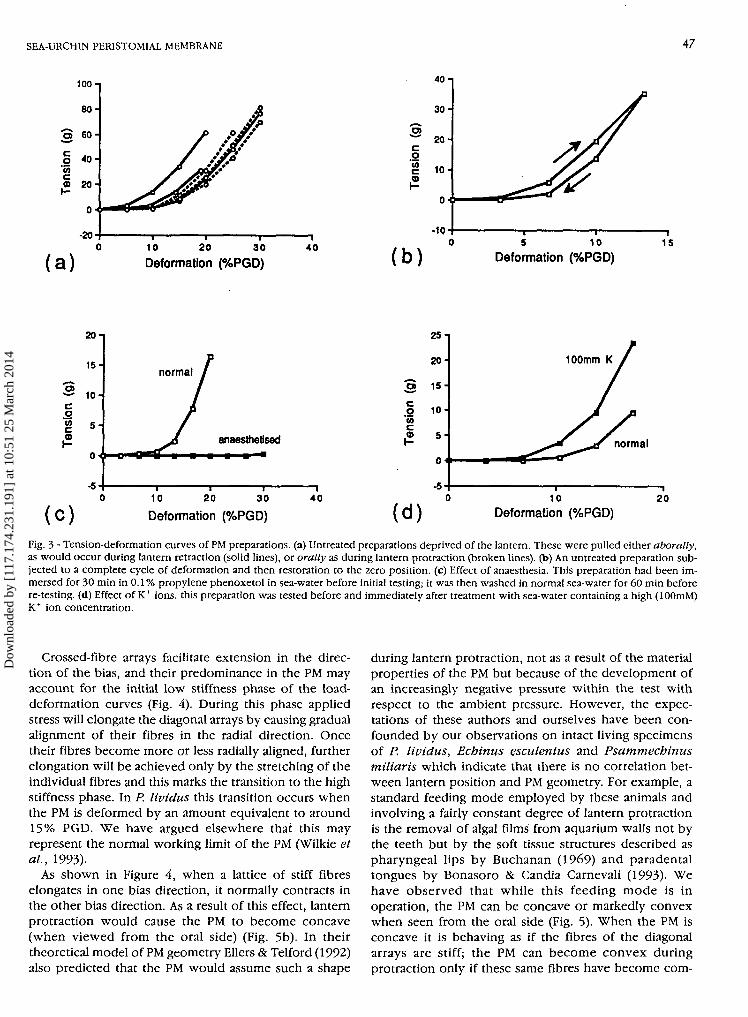
When whole PM preparations were subjected to step-wise vertical deformation, the resulting tension-deformation curves were J-shaped (Fig. 3a), indicatingthat the PM was compliant at low levels of deformationbut as deformation progressed became stiff. In mostcases the transition from low to high stiffness was fairlyabrupt and took place at a vertical deformationequivalent to around 15% perignathic girdle diameter(PGD).
Tests on preparations without the lantern, in which thePM was stretched incrementally and then returned to thezero position by décrémentai reductions, provided infor-mation on the elasticity of the PM. In Figure 3b the areaunder the upper (loading) curve represents the workdone in deforming the PM, and the area under the lower(unloading) curve represents the energy recovered bypassive elastic recoil. The latter expressed as a proportionof the former is a measure of elasticity known as the«resilience». In Figure 3b the resilience is 71%.
The effects of certain chemical agents on the tension-deformation behaviour of the PM were determined in or-der to assess whether or not it shows variable tensility, i.e.,the capacity to undergo inherent changes in passivemechanical properties. Immersion for 30 min in sea-watercontaining the anaesthetic propylene phenoxetol caused aprofound but reversible «softening» of the PM (Fig. 3c).Anaesthetization in effect prolonged in the low stiffnessphase of the tension-deformation curve so that the shift tohigh stiffness occurred at a much greater level of defor-mation than usual. On the other hand elevation of the ex-ternal potassium ion concentration had the opposite effect.Sea-water containing 100 mM K+ reversibly stiffened thePM, i.e., shortened the low stiffness phase so that the shiftto high stiffness occurred at a lower level of deformationthan usual (Fig. 3d). Acetylcholine as a 1 mM solution in sea-water also stiffened the PM reversibly (not illustrated).
DISCUSSION
PM dermis as a conventional biomaterial
In P. lividus the dermis of the PM consists of three in-terconnected layers of fibres: a submesothelial layerwhich includes a microfibrillar lamina, a middle layer,and a sparse subepidermal layer. The fibres in each ofthese layers lie mainly parallel to the external and internalsurfaces of the PM and are arranged in crossed-fibrearrays which are mainly diagonal, i.e., their constituentfibres are aligned at an angle to the radial and circum-ferential directions.
Dow
nloa
ded
by [
117.
174.
231.
191]
at 1
0:51
25
Mar
ch 2
014
SEA-URCHIN PER1STOM1AL MEMBRANE 47
(a)
S
.1
10 20 30Deformation (%PGD)
40(b)
-105 10
Deformation (%PGD)15
3co
(c )10 20 30
Deformation (%PGD)
40
Deformation (%PGD)
Fig. 3 - Tension-deformation curves of PM preparations, (a) Untreated preparations deprived of the lantern. These were pulled either aborally,as would occur during lantern retraction (solid lines), or orally as during lantern protraction (broken lines), (b) An untreated preparation sub-jected to a complete cycle of deformation and then restoration to the zero position, (c) Effect of anaesthesia. This preparation had been im-mersed for 30 min in 0.1% propylene phenoxetol in sea-water before initial testing; it was then washed in normal sea-water for 60 min beforere-testing, (d) Effect of K+ ions, this preparation was tested before and immediately after treatment with sea-water containing a high (lOOmM)K+ ion concentration.
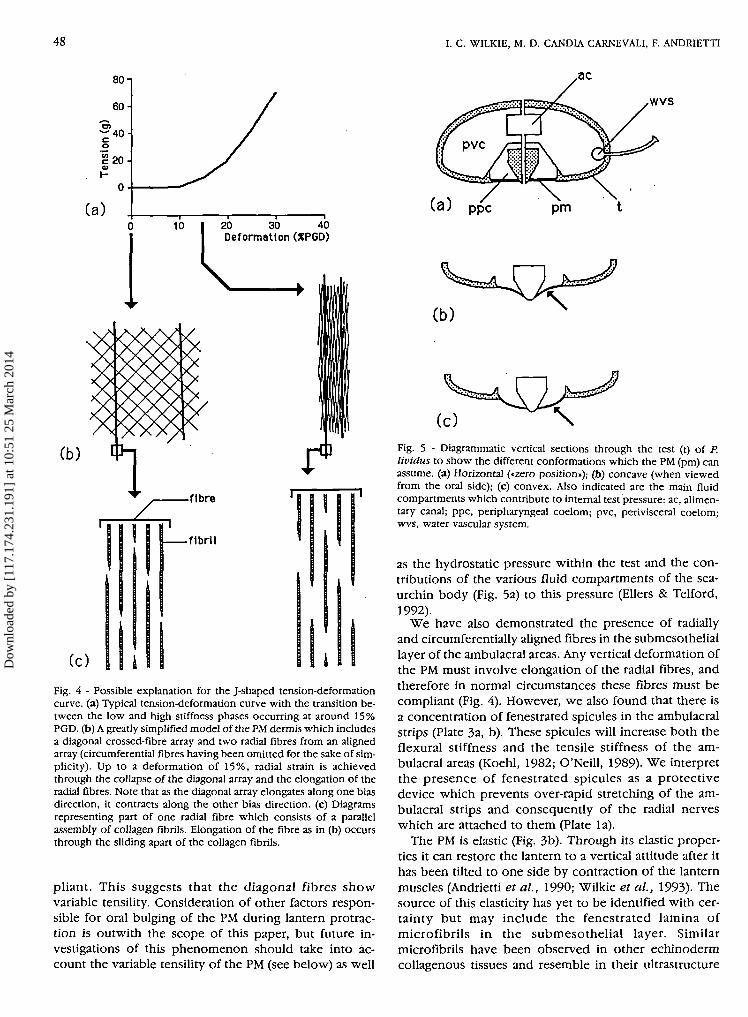
Crossed-fibre arrays facilitate extension in the direc-tion of the bias, and their predominance in the PM mayaccount for the initial low stiffness phase of the load-deformation curves (Fig. 4). During this phase appliedstress will elongate the diagonal arrays by causing gradualalignment of their fibres in the radial direction. Oncetheir fibres become more or less radially aligned, furtherelongation will be achieved only by the stretching of theindividual fibres and this marks the transition to the highstiffness phase. In P. lividus this transition occurs whenthe PM is deformed by an amount equivalent to around15% PGD. We have argued elsewhere that this mayrepresent the normal working limit of the PM (Wilkie etal., 1993).
As shown in Figure 4, when a lattice of stiff fibreselongates in one bias direction, it normally contracts inthe other bias direction. As a result of this effect, lanternprotraction would cause the PM to become concave(when viewed from the oral side) (Fig. 5b). In theirtheoretical model of PM geometry Ellers & Telford (1992)also predicted that the PM would assume such a shape
during lantern protraction, not as a result of the materialproperties of the PM but because of the development ofan increasingly negative pressure within the test withrespect to the ambient pressure. However, the expec-tations of these authors and ourselves have been con-founded by our observations on intact living specimensof P. lividus, Echinus esculentus and Psammechinusmiliaris which indicate that there is no correlation bet-ween lantern position and PM geometry. For example, astandard feeding mode employed by these animals andinvolving a fairly constant degree of lantern protractionis the removal of algal films from aquarium walls not bythe teeth but by the soft tissue structures described aspharyngeal lips by Buchanan (1969) and paradentaltongues by Bonasoro & Candia Carnevali (1993). Wehave observed that while this feeding mode is inoperation, the PM can be concave or markedly convexwhen seen from the oral side (Fig. 5). When the PM isconcave it is behaving as if the fibres of the diagonalarrays are stiff; the PM can become convex duringprotraction only if these same fibres have become com-
Dow
nloa
ded
by [
117.
174.
231.
191]
at 1
0:51
25
Mar
ch 2
014
48 I. C. WILKIE, M. D. CANDIA CARNEVALI, F. ANDRIETTI
10 20 30 40Deformation {%PGD)
fibre
fibri l
(c) iFig. 4 - Possible explanation for the J-shaped tension-deformationcurve, (a) Typical tension-deformation curve with the transition be-tween the low and high stiffness phases occurring at around 15%PGD. (b) A greatly simplified model of the PM dermis which includesa diagonal crossed-fibre array and two radial fibres from an alignedarray (circumferential fibres having been omitted for the sake of sim-plicity). Up to a deformation of 15%, radial strain is achievedthrough the collapse of the diagonal array and the elongation of theradial fibres. Note that as the diagonal array elongates along one biasdirection, it contracts along the other bias direction, (c) Diagramsrepresenting part of one radial fibre which consists of a parallelassembly of collagen fibrils. Elongation of the fibre as in (b) occursthrough the sliding apart of the collagen fibrils.
pliant. This suggests that the diagonal fibres showvariable tensility. Consideration of other factors respon-sible for oral bulging of the PM during lantern protrac-tion is outwith the scope of this paper, but future in-vestigations of this phenomenon should take into ac-count the variable tensility of the PM (see below) as well
wvs
ppc pm t
Fig. 5 - Diagrammatic vertical sections through the test (t) of P.lividus to show the different conformations which the PM (pm) canassume, (a) Horizontal («zero position»); (b) concave (when viewedfrom the oral side); (c) convex. Also indicated are the main fluidcompartments which contribute to internal test pressure: ac, alimen-tary canal; ppc, peripharyngeal coelom; pvc, perivisceral coelom;wvs, water vascular system.
as the hydrostatic pressure within the test and the con-tributions of the various fluid compartments of the sea-urchin body (Fig. 5a) to this pressure (Ellérs & Telford,1992).
We have also demonstrated the presence of radiallyand circumferentially aligned fibres in the submesotheliallayer of the ambulacral areas. Any vertical deformation ofthe PM must involve elongation of the radial fibres, andtherefore in normal circumstances these fibres must becompliant (Fig. 4). However, we also found that there isa concentration of fenestrated spicules in the ambulacralstrips (Plate 3a, b). These spicules will increase both theflexural stiffness and the tensile stiffness of the am-bulacral areas (Koehl, 1982; O'Neill, 1989). We interpretthe presence of fenestrated spicules as a protectivedevice which prevents over-rapid stretching of the am-bulacral strips and consequently of the radial nerveswhich are attached to them (Plate la).
The PM is elastic (Fig. 3b). Through its elastic proper-ties it can restore the lantern to a vertical attitude after ithas been tilted to one side by contraction of the lanternmuscles (Andrietti et al., 1990; Wilkie et al., 1993). Thesource of this elasticity has yet to be identified with cer-tainty but may include the fenestrated lamina ofmicrofibrils in the submesothelial layer. Similarmicrofibrils have been observed in other echinodermcollagenous tissues and resemble in their ultrastructure
Dow
nloa
ded
by [
117.
174.
231.
191]
at 1
0:51
25
Mar
ch 2
014

SEA-URCHIN PERISTOMIAL MEMBRANE 49
and solubility characteristics the fibrillin-containingmicrofibrils associated with vertebrate elastin (Keene etal, 1991; Thurmond & Trotter, 1992).
At present the significance of the three sublayers in thedermis is not clear. This investigation has shown thatapart from their relative thickness they differ in terms of(1) the mean diameter of their constituent fibres, (2) thepacking density of the fibres, (3) the orientation of thefibres, and (4) their relation to the ossicles. Burke et al.(1989) found significant differences between the meanperiodicity of collagen fibrils from the three sublayers inthe PM dermis of Strongylocentrotus franciscanus: 47Anm in the submesothelial layer; 44.4 nm in the middlelayer; and 50.8 nm in the subepidermal layer. Such dif-ferences may reflect variations in the proportion or com-position of interfibrillar proteoglycans in the threesublayers (Ruggeri & Benazzo, 1984), which in turn couldhave implications for their mechanical properties andtherefore make it possible that there is a certain degree offunctional differentiation between the sublayers. Thedenser submesothelial and middle sublayers must exertby far the greatest influence on the overall mechanicalproperties of the PM, the former perhaps being primarilyresponsible for setting a dimensional limit and thuspreventing overextension of the PM and especially of themicrofibrillar lamina and contractile mesothelium. Asparse subepidermal layer is found in other echinoderminteguments (holothurian: Menton & Eisen, 1970;asteroid: Wilkie et al, 1990) and is perhaps functionallyequivalent to the papillary layer of mammalian dermis(Wilkes et al., 1973) and the loose subepithelial stroma ofmammalian uterine cervix (Ferenczy, 1980). It may actsimply as a compliant force-dissipating junctional zonebetween the cellular epidermis and the dermis.
PM dermis as a mutable collagenous tissue
The reasonably close clustering of the tension-deformation curves for different PM specimens in Figure3a suggests that they represent a particular physiologicalstate: a normal baseline condition in which load-deformation properties are determined by diagonalarrays of stiff fibres and aligned arrays of compliantfibres. In this normal working condition, vertical deform-ation of the PM involves straightforward radial alignmentof the diagonal fibres and elongation of the radial fibres(Fig. 4).
We have demonstrated that this baseline behaviour canbe modified by chemical factors. Anaesthetizationreduces the stiffness of the PM (Fig. 3 c) to an extent thatmay mimic its compliant mechanical state when lanternprotraction is accompanied by oral bulging of the PM(Fig. 5c). This «softening» effect implies that the normalbaseline stiffness of the PM is maintained by a certainlevel of nervous activity which can be abolished byanaesthetization. On the other hand acetylcholine or ex-cess K+ increases the stiffness of the PM (Fig. 3d) possiblydue to the stimulation of nervous pathways: acetyl-choline may act at post-synaptic sites and excess K+ may
have a non-specific depolarizing action on the neuralcomponents of such pathways. The same pattern ofresponses to anaesthetization, acetylcholine and K+ hasbeen observed in the largely ligamentous compassdepressors of P. lividus (Wilkie et al, 1992) and is con-sistent with the view that the mechanical properties ofthe PM dermis are under nervous control, i.e., that it is amutable collagenous tissue (MCT) (Wilkie et al, 1993).The PM dermis contains cellular elements which aresimilar to those of other echinoderm MCTs: the largegranule containing cells resemble the juxtaligamentalcells whose processes permeate all MCTs and which mayeffect directly the tensile changes in these tissues (Wilkie,1979; Candia Carnevali & Wilkie, 1992); the small granulecontaining varicose processes are probably axons andtheir close proximity to the large granule containing cellsprovides further morphological evidence for a functionallink between juxtaligamental cells and the motor nervoussystem, a relationship which is most obvious inophiuroids (Wilkie, 1979; Cobb, 1985).
By what mechanism is the tensility of the PM dermismodulated? The fibres of the dermis are bundles ofdiscontinuous collagen fibrils. It is likely that, as theirultrastructural appearance suggests, the interstitialcollagen fibrils of echinoderms have a molecularorganization closely similar to that of those mammalianfibrils that are dominated by type I collagen (D'Alessio etal, 1989; Shimizu et al, 1990) and therefore that theyhave the same physical characteristics of extreme stiff-ness and high tensile strength (Viidik et al., 1982). A con-siderable body of evidence indicates that the variabletensility of echinoderm MCTs depends on changes not inthe collagen fibrils but in the ground substancemolecules that interconnect the fibrils. As in mammalianconnective tissues these interfibrillar molecules areproteoglycans (Trotter & Koob, 1989; Kariya et al,1990a, b; Scott, 1991) and it is likely that the stiffness ofMCTs is adjusted through changes in the type or densityof interactions between adjacent proteoglycan moleculesor between proteoglycans and collagen fibrils(Motokawa, 1984a; Wilkie, 1984; Wilkie & Emson, 1988).Anaesthetic-induced softening may thus result from thesuppression of proteoglycan-mediated cohesion be-tween fibrils in the diagonal fibres which allows inter-fibrillar slippage to occur.
As well as operating between fibrils, such a molecularmechanism may also operate between fibres. Some ofthe 12 nm microfibrils seen in ultrathin sections are likelyto be condensed proteoglycans and it has been observedthat these appear to link together both adjacent fibrilsand adjacent bundles of fibrils (i.e., fibres) (Plate 4b).Acetylcholine- and K+-induced stiffening could resultfrom changes in proteoglycan-mediated interfibre (as op-posed to interfibrillar) interactions. The collapse of thediagonal fibre arrays that is responsible for the low stiff-ness phase of the load-deformation curve must be ac-companied by distortion of, and perhaps intermolecularshear within, the proteoglycan networks that link the
Dow
nloa
ded
by [
117.
174.
231.
191]
at 1
0:51
25
Mar
ch 2
014

50 I. C. WILKIE, M. D. CANDIA CARNEVALI, F. ANDRIETTI
fibres together. It is conceivable that fibre arrays can bestiffened by the strengthening of intermolecular interac-tions (e.g. through an increase in crosslink density)which prevents such proteoglycan distortion or shear.
CONCLUSIONS
The PM dermis has a complex micro-organization:although at one level it can be regarded as a single three-dimensional meshwork of collagen fibres, this meshworkhas three histologically distinct but structurally intercon-nected sublayers; the sublayers consist largely of latticesof fibres having either a diagonal or an aligned orien-tation with respect to the radial and circumferentialdirections; and in what we believe is the normal workingstate the collagen fibres of those lattices are likely tobelong to separate populations with differing tensileproperties (stiff fibres in the diagonal arrays and com-pliant fibres in the aligned arrays).
The simple mechanical tests employed in our ex-periments have given only a glimpse of the tensileproperties of the PM and how these properties are relatedto its structure. They have provided information on theelasticity of the PM and on its non-linear load-deformation characteristics when undergoing radialstrain. We have not investigated its creep behaviour,time-dependence, anisotropy, or any aspect of its flexuralstiffness as opposed to its tensile stiffness. However, allof these must be important for the mechanical func-tioning of the PM, and their quantification could providea fuller explanation of its structural complexity.
Similarly, whilst our results have demonstrated that thePM dermis consists of MCT whose mechanical propertiesare under physiological control, a more detailed analysisof nervously mediated influences on its biomechanicalparameters is required. As has been shown for otherMCTs (Hidaka & Takahashi, 1983; Motokawa, 1984), thiswould provide more insight into the molecularmechanisms underlying variable tensility.
REFERENCES
Andrietti F., Candia Carnevali M. D., Wilkie I. C, Lanzavecchia G.,Melone G., Celentano F. C, 1990 - Mechanical analysis of the sea-urchin lantern: the overall system in Paracentrotus lividus. J.Zool., Lond., 200: 345-366.
Birenheide R., 1990 - Functional analysis of the tooth support inechinoids. In: C. De Ridder, P. Dubois, M. LaHaye & M. Jangoux(eds), Echinoderm research. Balkema, Rotterdam, pp. 203-206.
Bonasoro F., Candia Carnevali M. D., 1993 - Atypical chordoid struc-tures in the Aristotle's lantern of regular echinoids. Acta zool., (inpress).
Buchanan J. B., 1969 - Feeding and the control of volume within thetests of regular sea-urchins. J. Zool., Lond., 159: 51-64.
Burke R. D., Bouland C, Sanderson A. I., 1989 - Collagen diversity inthe sea urchin, Strongylocentrotus purpuratus. Comp. Biochem.Physiol., 94B: 41-44.
Candia Carnevali M. D., Bonasoro F., Andrietti F., Melone G., WilkieI. C, 1990 - Functional morphology of the peristomial mem-
brane of regular sea-urchins: general organization andmechanical properties in Paracentrotus lividus. In: C. De Rid-der, P. Dubois, M. LaHaye & M. Jangoux (eds), Echinodermresearch. Balkema, Rotterdam, pp. 207-216.
Candia Carnevali M. D., Wilkie I. C, 1992 - Gli straordinari tessuticonnettivi degli echinodermi. Le Scienze, 286: 58-70.
Cobb J. L. S., 1985 - The motor innervation of the oral plate ligament inthe brittlestar Ophiura ophiura (L.). Cell Tissue Res., 242: 685-688.
Cobb J. L. S., Sneddon E., 1977 - An ultrastructural study of the gillsof Echinus esculentus. Cell Tissue Res., 182: 265-274.
D'Alessio M., Ramirez F., Suzuki H. R., Solursh M., Gambino R., 1989- Structure and expression of a sea-urchin fibrillar collagen gene.Proc. natl. Acad. Sci., 86: 9303-9307.
Diab M., Gilly W. F., 1984 - Mechanical properties and control ofnonmuscular catch in spine ligaments of the sea urchin,Strongylocentrotus franciscanus. J. exp. Biol., 111: 155-170.
Ellers O., Telford M., 1992 - Causes and consequences of fluctuatingcoelomic pressure in sea-urchins. Biol. Bull., 182: 424-434.
Evans J. H., Barbenel J. C, 1975 - Structural and mechanical proper-ties of tendon related to function. Equine Vet. Journal, 7: 1-8.
Ferenczy A., 1980 - The ultrastructure of the human cervix. In: F.Naftolin & P. G. Srubblefield (eds), Dilatation of the uterine cer-vix. Raven Press, New York, pp. 27-44.
Fung Y. C, 1984 - Structure and stress-strain relationship of softtissues. Am. Zool., 24: 13-22.
Gibbons C. A., Shadwick R. E., 1991 - Circulatory mechanics in thetoad Bufo marinus I. Structure and mechanical design of the aor-ta. J. exp. Biol., 158: 275-289.
Hidaka M., Takahashi K., 1983 - Fine structure and mechanicalproperties of the catch apparatus of the sea urchin spine, acollagenous connective tissue with muscle-like holding capacity.J. exp. Biol., 103: 1-14.
Humason G. L., 1979 - Animal tissue techniques. 4th edn. Freeman,San Francisco.
Kariya Y., Watabe S., Ochiai Y., Murata K., Hashimoto K., 1990a -Glycosaminoglycan involved in the cation-induced change ofbody wall structure of sea cucumber Stichopus japonicus. Con-nect. Tiss. Res., 25: 149-159.
Kariya Y, Watabe S., Ochiai Y., Hashimoto K., Murata K., 1990b -Glycosaminoglycan from the body wall of the sea cucumberStichopus japonicus. Comp. Biochem. Physiol., 95B: 387-392.
Keene D. R., Maddox B. K., Kuo H.-J., Sakai L. Y., Glanville R. W.,1991 - Extraction of extendable beaded structures and their iden-tification as fibrillin-containing extracellular matrix microfibrils.J. Histochem. Cytochem., 39: 441-449.
Koehl M. A. R., 1982 - Mechanical design of spicule-reinforced con-nective tissue: stiffness. J. exp. Biol., 98: 239-267.
Menton D. N., Eisen A. Z., 1970 - The structure of the integument ofthe sea cucumber, Thyone briareus. J. Morphol., 131: 17-36.
Morales M., Del Castillo J., Smith D. S., 1989a - Acetylcholine sen-sitivity of the spine-test articular capsule of the sea-urchinEucidaris tribuloides. Comp. Biochem. Physiol., 94C: 547-554.
Motokawa T., 1984a - Connective tissue catch in echinoderms. Biol.Rev., 59: 255-270.
Motokawa t., 1984b - Viscoelasticity of holothurian body wall. J.exp. Biol., 109: 63-75.
Nordin M., Frankel V. H., 1980 - Biomechanics of collagenoustissues. In: M. Nordin & V. H. Frankel (eds), Basic biomechanicsof the skeletal system. Lea & Febiger, Philadelphia, pp 87-110.
O'Neill P., 1989 - Structure and mechanics of starfish body wall. J.exp. Biol., 147: 53-89.
Ruggeri A., Benazzo F., 1984 - Collagen-proteoglycan interaction. In:A. Ruggeri & P. M. Motta (eds), Ultrastructure of the connectivetissue matrix. Martinus Nijhoff, Boston, pp. 113-125.
Scott J. E., 1991 - Proteoglycan: collagen interactions in connectivetissues. Ultrastructural, biochemical, functional and evolutionaryaspects. Int. J. biol. Macromol., 13: 157-161.
Shimizu K., Amemiya S., Yoshizato K., 1990 - Biochemical and im-munological characterization of collagen molecules fromechinothurioid sea urchin Astbenosoma ijimai. Biochim.Biophys. Acta, 1038: 39-46.
Dow
nloa
ded
by [
117.
174.
231.
191]
at 1
0:51
25
Mar
ch 2
014

SEA-URCHIN PERISTOMIAL MEMBRANE 51
Smith A. B., 1979 - Peristomial tube feet and plates of regularechinoids. Zoomorphology, 94: 67-80.
Thurmond F., Trotter J. A., 1992 - Microfibrils from sea cucumberbody wall. Am. Zool., 32: 44A.
Trotter J. A., Koob T. J., 1989 - Collagen and proteoglycan in a sea-urchin ligament with mutable mechanical properties. Cell TissueRes., 258: 527-539.
Viidik A., Danielsen C. C., Oxlund H., 1982 - On fundamental andphenomenological models, structure and mechanical propertiesof collagen, elastin and glycosaminoglycan complexes.Biorheology, 19: 437-451.
Wilkes G. L., Brown I. A., Wildnauer R. H., 1973 - Thebiomechanical properties of skin. CRC Reviews inBioengineering, August 1973: 453-495.
Wilkie I. C., 1979 - The juxtaligamental cells of Ophiocomina nigra(Abildgaard) (Echinodermata: Ophiuroidea) and their possiblerole in mechano-effector function of collagenous tissue. CellTissue Res., 197: 515-530.
Wilkie I. C., 1984 - Variable tensility in echinoderm collagenoustissues: a review. Mar. Behav. Physiol., 11: 1-34.
Wilkie I. C., Emson R. H., 1988 - Mutable collagenous tissues andtheir significance for echinoderm palaeontology and phylogeny.In: C.R.C. Paul & A.B. Smith (eds), Echinoderm phylogeny andevolutionary biology. Clarendon Press, Oxford, pp. 311-330.
Wilkie I. C., Candia Carnevali M. D., Andrietti F., 1993 - Variable ten-sility of the peristomial membrane of the sea-urchin Paracen-trotus lividus (Lam.). Comp. Biochem. Physiol., 105A: 493-501.
Wilkie I. C., Candia Carnevali M. D., Bonasoro F., 1992 - The com-pass depressors of Paracentrotus lividus (Echinodermata,Echinoida): ultrastructural and mechanical aspects of theirvariable tensility and contractility. Zoomorphology, 112:143-153.
Wilkie I. C., Griffiths G. V. R., Glennie S. F., 1990 - Morphologicaland physiological aspects of the autotomy plane in the aboral in-tegument of Asterias rubens L. (Echinodermata). In: C. De Rid-der, P. Dubois, M. LaHaye & M. Jangoux (eds), Echinodermresearch. Balkema, Rotterdam, pp. 301-313.
Woodley J. D., 1967 - Problems in the ophiuroid water vascularsystem. Symp. zool. Soc. Lond., 20: 75-104.
Dow
nloa
ded
by [
117.
174.
231.
191]
at 1
0:51
25
Mar
ch 2
014





























