Embed Size (px)
Citation preview

Lysophosphatidic acid stimulates cell migration ofsatellite cells. A role for the sphingosine kinase/sphingosine 1-phosphate axisFrancesca Cencetti1,2, Gennaro Bruno1, Sabrina Blescia1, Caterina Bernacchioni1, Paola Bruni1,2
and Chiara Donati1,2
1 Dipartimento di Scienze Biomediche Sperimentali e Cliniche ‘Mario Serio’, Universit�a di Firenze, Italy
2 Istituto Interuniversitario di Miologia, Italy
Keywords
lysophosphatidic acid; satellite cells; skeletal
muscle; sphingosine 1-phosphate;
sphingosine kinase
Correspondence
P. Bruni, Dipartimento di Scienze
Biomediche Sperimentali e Cliniche ‘Mario
Serio’, Universit�a di Firenze, Viale G.B.
Morgagni 50, Firenze 50134, Italy
Fax: +39 05527511351
Tel: +39 0552751204
E-mail: [email protected]
(Received 13 March 2014, revised 4 July
2014, accepted 29 July 2014)
doi:10.1111/febs.12955
Regulation of the motility of skeletal muscle precursor cells, such as satel-
lite cells, is critically important for their proper recruitment at the site of
tissue damage, and ultimately for its correct repair. Here we show that
lysophosphatidic acid (LPA), which is well-recognized as a powerful bioac-
tive agent, strongly stimulates cell migration of activated murine satellite
cells. The biological effect exerted by LPA was found to be induced via
activation of LPA1 and LPA3, being abolished by cell treatment with the
antagonist Ki16425, and severely impaired by siRNA-mediated down-regu-
lation of the two receptor isoforms. In contrast, silencing of LPA2 potenti-
ated the stimulation of cell motility by LPA, suggesting that it is negatively
coupled to cell migration. Pharmacological inhibition of both sphingosine
kinase (SK) isoforms using VPC96047, or the selective blocking of SK1
using VPC96091, abolished cell responsiveness to LPA; in agreement, gene
silencing of SK1 or SK2 significantly reduced the biological effect of LPA.
Moreover, the LPA-dependent stimulation of cell chemotaxis was found to
be impaired by down-regulation of the sphingosine 1-phosphate (S1P)
receptors S1P1 or S1P4 by specific siRNAs. In summary, the results
obtained support the notion that the sphingosine kinase/sphingosine
1-phosphate (SK/S1P) axis is critically involved in the mechanism by which
LPA elicits its pro-migratory action. This study provides compelling new
information on the regulatory mechanisms of satellite cell motility, and
reinforces the view that the SK/S1P signaling pathway plays a crucial role
in the control of skeletal muscle precursor cell biology.
Introduction
Lysophosphatidic acid (LPA) and sphingosine 1-phos-
phate (S1P) are two potent bioactive lysophospholip-
ids that, mainly acting via engagement of multiple
G protein-coupled receptors, are able to regulate key
biological processes in mammalian cells [1]. Impor-
tantly, among the wide variety of cell types respon-
sive to LPA and S1P, a range of adult and
embryonic stem cells and progenitors are included,
suggesting that, by means of regulating their survival,
proliferation, differentiation and motility, these
lysophospholipids play a significant role in the
maintenance, renewal and trafficking of various
populations of stem cells and progenitors in the body
[2].
Abbreviations
BSA, bovine serum albumin; DMEM, Dulbecco’s modified Eagle’s medium; LPA, lysophosphatidic acid; S1P, sphingosine 1-phosphate; SCR,
scrambled; SK, sphingosine kinase.
4467FEBS Journal 281 (2014) 4467–4478 ª 2014 FEBS

There is a wealth of experimental results in favor of
a key role of S1P in regulation of the regenerative
potential of skeletal muscle, a tissue that has the ability
to self-repair following trauma or disease due to the
existence of precursor cells, which reside beneath the
basal lamina of each myofiber and are named satellite
cells [3,4]. Indeed, S1P has been shown to act as power-
ful mitogenic cue in quiescent and activated mouse
satellite cells [5,6]. Moreover, this lysosphingolipid has
been reported to be capable of stimulating the motility
of activated satellite cells, which is critical for their cor-
rect recruitment at the site of tissue lesions [6]. Interest-
ingly, these biological effects induced by S1P appear to
be transmitted by ligation to S1P receptors: S1P2 and
S1P3 are involved in transmission of the mitogenic
effect of the sphingolysolipid [6,7], whereas S1P1 and
S1P4 appear to be implicated in the migratory response
elicited by S1P [6]. Interestingly, recent studies per-
formed in mice in which S1P2 was genetically ablated
or pharmacologically blocked have provided experi-
mental evidence that this receptor subtype promotes
skeletal muscle regeneration [8], while altered cell-cycle
progression has been observed in satellite cells of S1P3
knockout mice [9]. Furthermore, comprehensive studies
performed in the C2C12 myoblast cell line, which is
largely used to investigate the biological properties of
skeletal muscle precursor cells, have clearly demon-
strated that S1P and its metabolism play a critical role
in myogenic differentiation, and that the S1P signaling
pathway is used by various growth factors and cyto-
kines to elicit specific biological effects [10–16].Present knowledge of the biological role of LPA in
skeletal muscle precursor cells is more limited. An
early report described the stimulatory effect of LPA
on cell proliferation of C2C12 myoblasts, accompanied
by its inhibitory action on myogenic differentiation
[17]. The mitogenic effect of LPA in myoblasts was
further confirmed in a subsequent study, in which the
LPA action was found to be exerted via ligation of
LPA1/LPA3 receptors and activation of the phosphati-
dylinositol-4,5-bisphosphate 3-kinase pathway [18].
Additionally, a recent study has shown that LPA, act-
ing via Gai2, robustly stimulated hypertrophy of myo-
tubes in a protein kinase C dependent manner [19].
In a previous study, we observed that LPA acts as
mitogenic cue in activated satellite cells [6]. This
prompted us to better characterize the biological
action of LPA in these skeletal muscle precursor cells
and to explore its molecular mechanism of action.
Here we report that LPA significantly enhances migra-
tion of activated satellite cells, and that cross-talk with
sphingosine kinase (SK) and S1P receptors plays a key
role in transmission of this biological effect.
Results
In order to obtain more information on the biological
role performed by LPA in skeletal muscle precursors,
taking into consideration the crucial importance of
regulation of cell motility for skeletal muscle regenera-
tion, we assessed whether it affects this biological
parameter by measuring chemotaxis of activated satel-
lite cells toward LPA using the Boyden chamber assay.
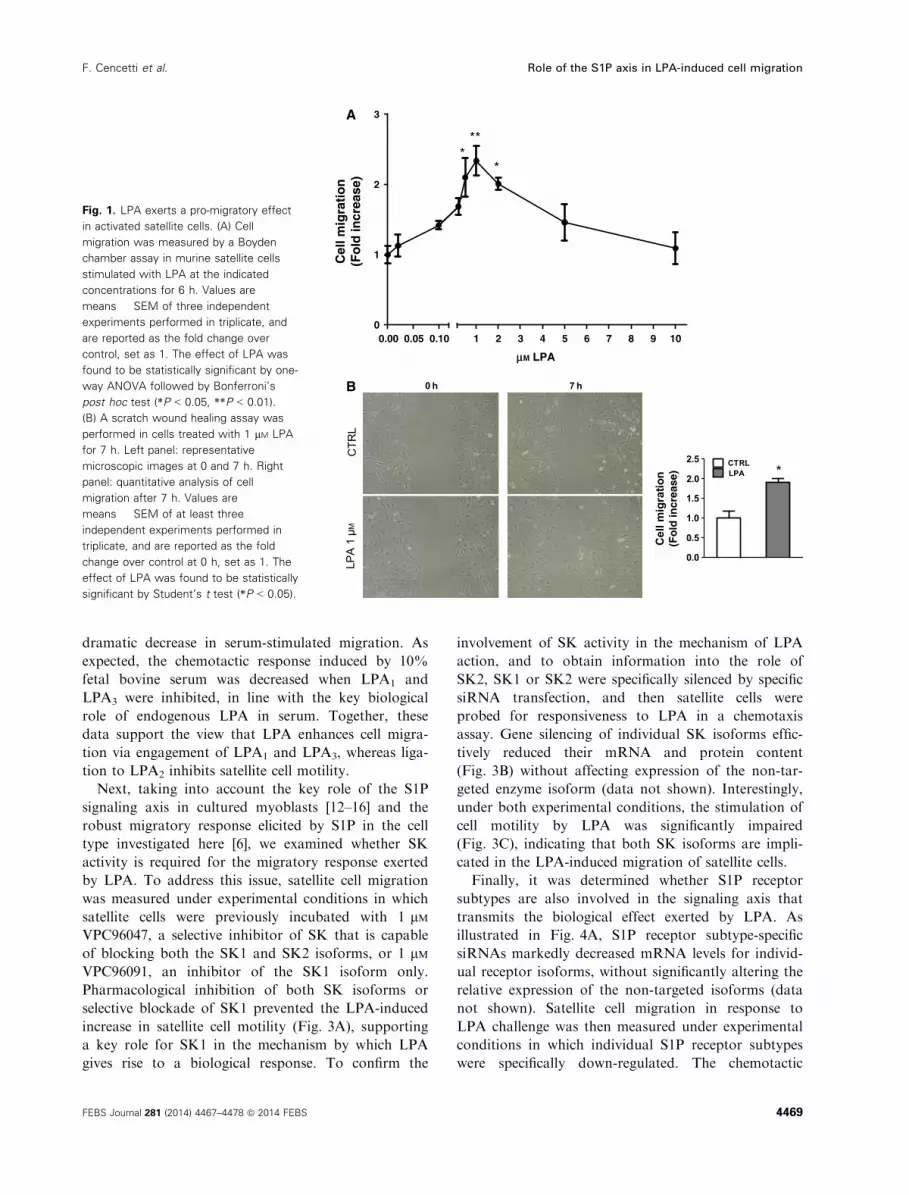
The results shown in Fig. 1A indicate that LPA influ-
enced chemotactic movement, producing a typical
bell-shaped curve. The maximal increase in the number
of migrated cells after 6 h of incubation was observed
using 1 lM LPA. This concentration was therefore
used in subsequent experiments. A scratch wound
healing assay performed at 7 h after cell injury and
treatment with 1 lM LPA confirmed that this lyso-
phospholipid is capable of strongly stimulating cell
motility (Fig. 1B).
To investigate the role of LPA receptor subtypes in
transmitting the biological effect of LPA, their relative
expression at the mRNA level was assessed by real-
time PCR. As shown in Fig. 2A, LPA1 is the domi-
nant receptor subtype expressed, with LPA2 and LPA3
being clearly detectable but significantly less repre-
sented. Cell migration experiments were then per-
formed using cultured satellite cells treated with the
LPA1/LPA3 antagonist Ki16425 (5 lM) 40 min prior
incubation with LPA. The pharmacological blockade
of LPA receptors fully prevented the enhancement
of cell migration exerted by 1 lM LPA (Fig. 2B). To
corroborate the experimental evidence regarding the
involvement of LPA receptor subtypes in the biologi-
cal response elicited by LPA, RNA interference tech-
nology was used. Cell transfection with specific
siRNAs strongly down-regulated individual LPA
receptor subtypes (Fig. 2C), but did not significantly
affect expression levels of the non-targeted LPA recep-
tor subtypes (data not shown). As shown in Fig. 2D,
reduced expression of LPA1 completely abolished the
stimulation of the migratory properties of satellite cells
exerted by 1 lM LPA; similarly, gene silencing of
LPA3 attenuated the biological action of LPA. In
contrast, when LPA2 expression was diminished, the
stimulatory effect of LPA on cell migration was poten-
tiated (Fig. 2D). To further demonstrate the anti-
migratory action mediated by LPA2, the chemotactic
response elicited by LPA was evaluated in cells chal-
lenged with serum in the presence or absence of the
LPA1/LPA3 antagonist Ki16425 (5 lM). As shown in
Fig. 2E, addition of 1 lM LPA strongly enhanced the
cell motility induced by 10% fetal bovine serum,
whereas blockade of LPA1 and LPA3 resulted out in a
4468 FEBS Journal 281 (2014) 4467–4478 ª 2014 FEBS
Role of the S1P axis in LPA-induced cell migration F. Cencetti et al.
dramatic decrease in serum-stimulated migration. As
expected, the chemotactic response induced by 10%
fetal bovine serum was decreased when LPA1 and
LPA3 were inhibited, in line with the key biological
role of endogenous LPA in serum. Together, these
data support the view that LPA enhances cell migra-
tion via engagement of LPA1 and LPA3, whereas liga-
tion to LPA2 inhibits satellite cell motility.
Next, taking into account the key role of the S1P
signaling axis in cultured myoblasts [12–16] and the
robust migratory response elicited by S1P in the cell
type investigated here [6], we examined whether SK
activity is required for the migratory response exerted
by LPA. To address this issue, satellite cell migration
was measured under experimental conditions in which
satellite cells were previously incubated with 1 lMVPC96047, a selective inhibitor of SK that is capable
of blocking both the SK1 and SK2 isoforms, or 1 lMVPC96091, an inhibitor of the SK1 isoform only.
Pharmacological inhibition of both SK isoforms or
selective blockade of SK1 prevented the LPA-induced
increase in satellite cell motility (Fig. 3A), supporting
a key role for SK1 in the mechanism by which LPA
gives rise to a biological response. To confirm the
involvement of SK activity in the mechanism of LPA
action, and to obtain information into the role of
SK2, SK1 or SK2 were specifically silenced by specific
siRNA transfection, and then satellite cells were
probed for responsiveness to LPA in a chemotaxis
assay. Gene silencing of individual SK isoforms effic-
tively reduced their mRNA and protein content
(Fig. 3B) without affecting expression of the non-tar-
geted enzyme isoform (data not shown). Interestingly,
under both experimental conditions, the stimulation of
cell motility by LPA was significantly impaired
(Fig. 3C), indicating that both SK isoforms are impli-
cated in the LPA-induced migration of satellite cells.
Finally, it was determined whether S1P receptor
subtypes are also involved in the signaling axis that
transmits the biological effect exerted by LPA. As
illustrated in Fig. 4A, S1P receptor subtype-specific
siRNAs markedly decreased mRNA levels for individ-
ual receptor isoforms, without significantly altering the
relative expression of the non-targeted isoforms (data
not shown). Satellite cell migration in response to
LPA challenge was then measured under experimental
conditions in which individual S1P receptor subtypes
were specifically down-regulated. The chemotactic
0.00 0.05 0.100
1
2
3
1 2 3 4 5 6 7 8 9 10
μμM LPA
**
Cel
l mig
rati
on
(Fo
ld in
crea
se)
**
A
B
Fig. 1. LPA exerts a pro-migratory effect
in activated satellite cells. (A) Cell
migration was measured by a Boyden
chamber assay in murine satellite cells
stimulated with LPA at the indicated
concentrations for 6 h. Values are
means � SEM of three independent
experiments performed in triplicate, and
are reported as the fold change over
control, set as 1. The effect of LPA was
found to be statistically significant by one-
way ANOVA followed by Bonferroni’s
post hoc test (*P < 0.05, **P < 0.01).
(B) A scratch wound healing assay was
performed in cells treated with 1 lM LPA
for 7 h. Left panel: representative
microscopic images at 0 and 7 h. Right
panel: quantitative analysis of cell
migration after 7 h. Values are
means � SEM of at least three
independent experiments performed in
triplicate, and are reported as the fold
change over control at 0 h, set as 1. The
effect of LPA was found to be statistically
significant by Student’s t test (*P < 0.05).
4469FEBS Journal 281 (2014) 4467–4478 ª 2014 FEBS
F. Cencetti et al. Role of the S1P axis in LPA-induced cell migration

action of LPA was significantly attenuated when S1P1
or S1P4 were silenced, suggesting that these two recep-
tors are the main transducers of the LPA biological
effect (Fig. 4B). In contrast, down-regulation of S1P2,
which is known to transmit an anti-migratory
response in several cell types, including satellite cells
[1,6,20], did not affect the action exerted by LPA.
Similarly, specific silencing of S1P3 did not influence
the migratory effect of LPA. In agreement with the
results obtained, the chemotactic effect exerted by
LPA was unaltered in satellite cells isolated from S1P3
knockout mice (Fig. 4C). A diagram illustrating the
signaling mechanisms downstream of LPA is provided
in Fig. 5.
Discussion
Directed cell motility of satellite cells is a prerequisite
for proper skeletal muscle repair as these precursor
cells are located at the peripheral region of the skeletal
muscle fibers and must migrate to the site of muscle
damage in order to be recruited for tissue repair.
LPA1 LPA2 LPA3
0.00
0.05
0.10
0.8
1.0
1.2
mR
NA
exp
ress
ion
leve
l2^
(-ΔΔΔΔ
Ct)
0.0
0.5
1.0
1.5
2.0
2.5
CTRLLPA
Vehicle
5 mM Ki16425
Cel
l mig
rati
on
(Fo
ld in
crea
se)
*
0.0
0.5
1.0
1.5 SCR-siRNA
LPA1-siRNA
LP
A1
mR
NA
exp
ress
ion
2^(
-ΔΔΔΔ ΔΔΔΔ
Ct)
*
0.0
0.5
1.0
1.5SCR-siRNA
LPA3-siRNA
LP
A3
mR
NA
exp
ress
ion
2^(
-ΔΔΔΔ ΔΔΔΔ
Ct)
**
0.0
0.5
1.0
1.5 SCR-siRNA
LPA2-siRNA
LP
A2
mR
NA
exp
ress
ion
2^(
- ΔΔΔΔ ΔΔΔΔ
Ct)
**
0
1
2
3
4
CTRL
LPA
SCR-siRNA
LPA1-siRNA
LPA3-siRNA
Cel
l mig
rati
on
(Fo
ld in
crea
se) #
#
LPA2-siRNA
**
–+++ +
+–+
+––
–
1 µM LPA
+––5 µM Ki16425
10% FCS
0.0
0.5
1.0
1.5
2.0
2.55
10
15
Cel
l mig
rati
on
(Fo
ld in
crea
se)
#
**
§§
A B
C
D E
4470 FEBS Journal 281 (2014) 4467–4478 ª 2014 FEBS
Role of the S1P axis in LPA-induced cell migration F. Cencetti et al.

Despite its crucial importance, the regulation of
satellite cell motility at present is not clearly character-
ized, and the potential physiological modulators of the
process have not been fully identified.
Here we report that LPA, a lysophospholipid with
powerful biological activity, markedly stimulates migra-
tion of satellite cells isolated from mouse tibialis ante-
rior muscle fibers and maintained in culture. The
described biological action of LPA may be physiologi-
cally relevant, as LPA is present in extracellular fluids
[21] and may become available to skeletal muscle pre-
cursor cells during tissue damage. In this regard, it is
interesting to note that most of the circulating LPA is
generated by the action of autotaxin, a secreted lyso-
phospholipase D that cleaves the choline headgroup
from lysophosphatidylcholine, an abundant plasma
phospholipid [22]. Although there are presently no
reports describing the production of autotaxin by skele-
tal muscle cells, this enzyme shows a broad tissue distri-
bution, and its transcript has been detected in human
skeletal muscle [23]. In addition, it is tempting to specu-
late that the pro-inflammatory environment that favors
skeletal muscle repair may enhance local LPA forma-
tion, in analogy with what has been demonstrated to
occur in lungs [24]. LPA appears to robustly enhance
the chemotaxis of satellite cells acting via LPA1 and
LPA3. This was demonstrated by blockade of the bio-
logical effect by the selective LPA1/LPA3 antagonist
Ki16425, as well as the abolition or reduction of LPA
efficacy following gene silencing of LPA1 or LPA3. The
finding that LPA1 and LPA3 transmit the biological
effect of LPA is in agreement with a recent study in
which these two receptor subtypes were implicated
in transmitting the increase in intracellular calcium in
C2C12 myoblasts [18].
Interestingly, it was found that down-regulation of
LPA2 expression by gene silencing was responsible for
an increased migratory response induced by LPA, sug-
gesting that LPA2 is coupled to anti-migratory signals.
In this respect, previous studies have described anti-
migratory action transmitted by distinct LPA receptors
[25,26], and LPA2 has already been implicated in the
inhibition of migration and invasion of pancreatic
cancer cells [27].
Another notable finding of this study is that the S1P
signaling axis is required for the biological response
elicited by LPA in these cells, consistent with the
notion that a complex cross-talk between LPA and S1P
occurs in these cells. In cultured myoblasts, the SK/S1P
signaling axis was previously implicated in transmission
of important biological effects evoked by growth
factors and cytokines, such as the pro-myogenic effect
of insulin growth factor-1 [15] and tumor necrosis
factor-a at low dose [12], the pro-fibrotic [13] and pro-
apoptotic [16] effects of transforming growth factor-b,and, more pertinent to this study, the pro-migratory
Fig. 2. Role of LPA receptors in the migratory action of LPA in activated satellite cells. (A) Expression of LPA receptors at the mRNA level.
Quantitative mRNA analysis was performed by real-time PCR by concurrent amplification of the target sequence of murine LPA1, LPA2 and
LPA3 genes together with that of 18S rRNA. The results are expressed as fold changes according to the 2�ΔΔCT method, using LPA1 as the
calibrator. Values are means � SEM of three independent experiments performed in triplicate. (B) Effect of LPA1 and LPA3 antagonists on
the migratory action of LPA. Cell migration was assessed using the Boyden chamber apparatus in cells pre-incubated in the presence or not
of 5 lM Ki16425 for 40 min, before being treated with 1 lM LPA, as described in Experimental procedures. Values are means � SEM of
three independent experiments, each performed in triplicate, and are reported as the fold change over the control, set as 1. The effect of
Ki16245 was found to be statistically significant by two-way ANOVA followed by Bonferroni’s post hoc test (*P < 0.05). (C) LPA receptor
down-regulation by gene silencing. Real-time PCR was performed in cells transfected with scrambled siRNA (SCR-siRNA) or with siRNAs
specific for murine LPA1, LPA2 and LPA3, by amplification of the target sequence of murine LPA receptors together with that of 18S rRNA.
The results are expressed as fold changes according to the 2�ΔΔCT method, utilizing as the calibrator each receptor subtype in SCR-siRNA-
transfected myoblasts. Values are means � SEM at least of three independent experiments performed in triplicate. The effect of siRNA
transfection on the mRNA levels of LPA receptors was found to be statistically significant by Student’s t test (*P < 0.05, **P < 0.01). (D)
Effect of down-regulation of LPA receptors on the migratory action of LPA. Cells treated with control SCR-siRNA or with specific siRNA for
individual LPA receptors were used to measure cell migration toward 1 lM LPA for 6 h in a Boyden chamber apparatus. Values are
means � SEM of at least three independent experiments performed in triplicate. The inhibitory effect of LPA1 and LPA3 down-regulation on
LPA-induced cell motility was found to be statistically significant by two-way ANOVA followed by Bonferroni’s post hoc test (#P < 0.05).
The positive effect of LPA2 down-regulation on LPA-induced cell migration was found to be statistically significant by two-way ANOVA
followed by Bonferroni’s post hoc test (**P < 0.01). (E) Effect of LPA1/LPA3 blockade on the migratory action of LPA in the presence of
serum. Cell migration was assessed using a Boyden chamber apparatus in cells pre-incubated with or without 5 lM Ki16425 for 40 min,
before being incubated with 10% fetal bovine serum (FCS) in the lower chamber, in the presence or absence of 1 lM LPA. Values are
means � SEM of three independent experiments, each performed in triplicate, and the results are reported as the fold change over
untreated cells, set as 1. The migration elicited by LPA was found to be statistically significant compared to serum alone by Student’s t test
(**P < 0.01). The reduction of serum-induced cell motility by Ki16425 was found to be statistically significant by Student’s t test
(§§P < 0.01). The inhibitory effect of LPA in the presence of Ki16245 was found to be statistically significant by Student’s t test (#P < 0.05).
4471FEBS Journal 281 (2014) 4467–4478 ª 2014 FEBS
F. Cencetti et al. Role of the S1P axis in LPA-induced cell migration

effect of platelet-derived growth factor [14]. The pres-
ent results support the notion that transmission of bio-
logical responses evoked by extracellular cues also
relies on S1P inside-out signaling in satellite cells. Inter-
estingly, LPA, here identified as new agonist capable of
stimulating the migratory properties of satellite cells by
exploiting the S1P signaling axis, is an agonist that
shares many biological properties with S1P as well as
an analogous mechanism of action involving ligation to
a panel of G protein-coupled receptors [1,28]. Among
the multiple agents recognized as exerting their biologi-
cal action at least in part via regulation of the S1P sig-
naling pathway, LPA had already been reported to
stimulate migration of gastric cancer cells by means of
SK1 and S1P3 up-regulation through a mechanism that
involves LPA1 engagement and epidermal growth fac-
tor receptor transactivation [29]. In agreement, LPA1
was found to be implicated in this study in the regula-
tion of S1P signaling pathway; however, no significant
change was observed in the mRNA expression or pro-
tein content of SK1 after incubation with LPA within
the time frame adopted for Boyden chamber assays.
0.0
0.5
1.0
1.5
2.0
2.5CTRL
LPA
Cel
l mig
rati
on
(Fo
ld in
crea
se)
Vehicle
VPC96091
VPC96047
*
*
0.0
0.5
1.0
1.5 SCR-siRNASK1-siRNA
SK
1 m
RN
A e
xpre
ssio
n2^
(-ΔΔΔΔ ΔΔ
Ct)
*
0.0
0.5
1.0
1.5 SCR-siRNASK2-siRNA
SK
2 m
RN
A e
xpre
ssio
n 2
^(- Δ
ΔΔΔ ΔΔC
t)
*
SK1
β-actin
β-actin
SCR-siRNA
SK1-siRNA
SK2
SCR-siRNA
SK2-siRNA
0.0
0.5
1.0
1.5 SCR-siRNASK2-siRNA
SK
2 re
lati
ve d
ensi
ty(F
old
ch
ang
e)
0.0
0.5
1.0
1.5 SCR-siRNASK1-siRNA
SK
1 re
lati
ve d
ensi
ty(F
old
ch
ang
e)
*
**
0.0
0.5
1.0
1.5
2.0
2.5CTRL
LPA
SCR-siRNA
SK1-siRNA
SK2-siRNA
Cel
l mig
rati
on
(Fo
ld c
han
ge)
**
*
A
B
C
Fig. 3. Involvement of sphingosine kinase in cell migration induced
by LPA. (A) Effect of SK inhibitors on the migratory action of LPA.
Cell migration across Matrigel-coated membranes was performed
by a Boyden chamber assay in satellite cells treated or not with
1 lM VPC96091 or 1 lM VPC96047, to measure migration toward
1 lM LPA, as described in Experimental procedures. Values are
means � SEM of three independent experiments, each performed
in triplicate, and the results are reported as the fold change over
control, set as 1. The effect of VPC96091 and VPC96047 in LPA-
treated satellite cells was found to be statistically significant by
two-way ANOVA followed by Bonferroni’s post hoc test
(*P < 0.05). (B) SK1 and SK2 down-regulation by gene silencing.
Satellite cells were transfected with non-targeting SCR-siRNA or
with siRNAs specific for SK1 or SK2 as described in Experimental
procedures. Upper panels: real-time PCR was performed by
amplification of the target sequence of murine SK1 or SK2
together with that of 18S rRNA. The results are expressed as fold
changes according to the 2�ΔΔCT method, utilizing as the calibrator
each SK isoform in SCR-siRNA transfected myoblasts. Values are
means � SEM at least of three independent experiments
performed in triplicate. The effect of siRNA transfection on SK1 or
SK2 mRNA levels was found to be statistically significant by
Student’s t test (*P < 0.05). Lower and middle panels: western
blot analysis was performed by using specific antibodies against
SK1 or SK2. The results of densitometric analysis are reported as
means � SEM of three independent experiments. The effect of
siRNA transfection on SK1 or SK2 protein levels was found to be
statistically significant by Student’s t test (*P < 0.05, **P < 0.01).
(C) Effect of SK1 or SK2 down-regulation on the migratory action
of LPA. Cells transfected with control SCR-siRNA or with specific
siRNAs for each SK isoform were used to measure cell migration
toward 1 lM LPA for 6 h in a Boyden chamber apparatus. Values
are means � SEM of at least three independent experiments
performed in triplicate. The inhibitory effect of SK1 and SK2 down-
regulation on LPA-induced cell motility was found to be statistically
significant by two-way ANOVA followed by Bonferroni’s post hoc
test (*P < 0.05; **P < 0.01).
4472 FEBS Journal 281 (2014) 4467–4478 ª 2014 FEBS
Role of the S1P axis in LPA-induced cell migration F. Cencetti et al.

Moreover, siRNA experiments involving SK isoforms
support a role not only for SK1 but also for SK2 in
the stimulatory effect of LPA. This enzyme isoform,
although reported to be involved in biological effects
distinct from those mediated by SK1 [30], in some cases
acts in parallel with SK1, transmitting inside the cell
the biological action exerted by extracellular cues.
Intriguingly, SK2 had already been shown to be
involved in agonist-stimulated cell migration [31], in
agreement with the present results. Another interesting
0.0
0.5
1.0
1.5SCR-siRNA
S1P1-siRNA
S1P
1m
RN
A e
xpre
ssio
n2^
(-ΔΔΔΔ ΔΔΔΔ
Ct)
**
0.0
0.5
1.0
1.5SCR-siRNA
S1P3-siRNA
S1P
3 m
RN
A e
xpre
ssio
n2^
(-ΔΔΔΔ ΔΔΔΔ
Ct)
*
0.0
0.5
1.0
1.5SCR-siRNA
S1P2-siRNA
S1P
2 m
RN
A e
xpre
ssio
n2^
(-ΔΔΔΔ ΔΔΔΔ
Ct)
0.0
0.5
1.0
1.5 SCR-siRNA
S1P4-siRNA
S1P
4 m
RN
A e
xpre
ssio
n2^
(-ΔΔΔΔ ΔΔΔΔ
Ct)
*
*
0.0
0.5
1.0
1.5
2.0
2.5 CTRLLPA
SCR-siRNA
S1P1-siRNA
S1P2-siRNA
S1P3-siRNA
S1P4-siRNA
Cel
l mig
rati
on
(Fo
ld in
crea
se)
**
**
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5 CTRLLPA
Cel
l mig
rati
on
(Fo
ld in
crea
se)
S1P3KOwt
A
B C
Fig. 4. Role of S1P receptors in satellite cell migration induced by LPA. (A) Down-regulation of S1P receptors by gene silencing. Cells
transfected with control SCR-siRNA or with siRNAs specific for murine S1P1, S1P2, S1P3 and S1P4 were subjected to real-time PCR by
amplification of the target sequence of murine S1P receptors together with that of 18S rRNA. Results are expressed as fold changes
according to the 2�ΔΔCT method, utilizing as the calibrator each receptor subtype in SCR-siRNA-transfected myoblasts. Values are
means � SEM at least of three independent experiments performed in triplicate. The effect of siRNA transfection on S1P receptor mRNA
levels was found to be statistically significant by Student’s t test (*P < 0.05; **P < 0.01). (B) Effect of S1P receptor down-regulation on the
migratory action of LPA. Cells transfected with non-targeting SCR-siRNA or with specific siRNAs for individual murine S1P receptors were
used to measure cell migration toward 1 lM LPA for 6 h in a Boyden chamber apparatus. Values are means � SEM of at least three
independent experiments performed in triplicate. The inhibitory effect of S1P1 and S1P4 down-regulation on LPA-induced cell motility was
found to be statistically significant by two-way ANOVA followed by Bonferroni’s post hoc test (**P < 0.01). (C) Chemotactic response to
LPA in S1P3 knockout (KO) satellite cells. Satellite cells isolated from wild-type (wt) and S1P3 KO mice were used to measure cell migration
toward 1 lM LPA for 6 h in a Boyden chamber apparatus. Values are means � SEM of at least three independent experiments performed
in triplicate.
4473FEBS Journal 281 (2014) 4467–4478 ª 2014 FEBS
F. Cencetti et al. Role of the S1P axis in LPA-induced cell migration

result of this study is the identification of S1P1 and
S1P4 as molecular transducers of the migratory
response exerted by LPA. These two S1P receptor sub-
types were previously identified as being involved in
stimulation of satellite cell migration by exogenous S1P
[6]. Thus, it appears that exogenous as well as endoge-
nous S1P is capable of driving satellite cell chemotaxis
through these two receptor subtypes. In contrast, S1P2
down-regulation by gene silencing did not affect the
migration induced by LPA, whereas it was previously
found to be responsible for the enhancement of satellite
cell motility stimulated by exogenous S1P [6]. This find-
ing supports the view that, in contrast to exogenous
S1P, which is freely available to all the S1P receptors
exposed at plasma membrane, endogenous S1P gener-
ated inside the cells following LPA challenge is avail-
able only at specific membrane domains where S1P2 is
absent, thus making the S1P inside-out signaling selec-
tive for certain receptor subtypes.
Overall, these results shed new light on the molecu-
lar mechanisms that underlie the regulation of directed
cell motility in satellite cells, identifying LPA as a new
player in this process and reinforcing the concept that
the S1P signaling axis is critically involved in the regu-
lation of key biological events in skeletal muscle pre-
cursor cells.
Experimental procedures
Materials
Biochemicals, TRI Reagent, cell culture reagents,
Dulbecco’s modified Eagle’s medium (DMEM), fetal
bovine serum, horse serum, protease inhibitor cocktail,
bovine serum albumin (BSA), collagenase and Ki16425
were obtained from Sigma-Aldrich (Woodlands, TX,
USA). siRNA duplexes corresponding to DNA target
sequences of mouse S1P1 (SASI_Mm02_00312691 and
SASI_Mm02_00312692), mouse S1P2 (SASI_Mm01_00082
880 and SASI_Mm01_00082881), mouse S1P3 (SASI_
Mm01_00145233 and SASI_Mm01_00145234), mouse S1P4
(SASI_Mm01_00094192 and SASI_Mm01_00094193),
mouse LPA1 (SASI_Mm02_00318340), mouse LPA2 (SASI
_Mm01_00165280), mouse LPA3 (SASI_Mm01_00022765),
mouse SK1 (SASI_Mm01_00033983 and SASI_Mm01_
00033984), mouse SK2 (SASI_Mm01_00050883 and SA
SI_Mm01_00050884) and scrambled siRNA (Mission
Universal Negative control #1) were obtained from Sigma-
Proligo (Woodlands, TX, USA). Lipofectamine RNAi-
MAXTM, all reagents required to perform real-time PCR,
and the TaqMan gene expression assay probes for LPA1
(Mm00439145_m1), LPA2 (Mm00469562_m1), LPA3 (Mm0
0469562_m1), S1P1 (Mm02619656_s1), S1P3 (Mm026
20181_s1), S1P4 (Mm00468695_s1), SK1 (Mm00448841_g1)
and SK2 (Mm00445021_m1) were purchased from Life
Technologies (Carlsbad, CA, USA). The gene expression
assay probe for mouse S1P2 (Mm.PT.47.12362698) was
purchased from Integrated DNA Technologies Inc. (Coral-
ville, IA, USA). The enhanced chemiluminescence reagent
was purchased from GE Healthcare Europe (Milan, Italy).
Coomassie Brilliant Blue reagent was obtained from
Bio-Rad (Hercules, CA, USA). Chick embryo extract was
obtained from Sera Laboratories International Ltd
(Haywards Heath, UK). Collagen type I was purchased
from Millipore (Billerica, MA, USA). Matrigel basement
membrane matrix was obtained from BD Biosciences (Bed-
ford, MA, USA). The LPA used in the study was 1-oleoyl
lysophosphatidic acid sodium salt, purchased from Cayman
Chemical (Ann Arbor, MI, USA). Restore western blot
stripping buffer was purchased from ThermoFisher
Scientific (Rockford, IL, USA). Polycarbonate filters (8 lm
Fig. 5. Diagram of the signaling
mechanisms downstream of LPA.
4474 FEBS Journal 281 (2014) 4467–4478 ª 2014 FEBS
Role of the S1P axis in LPA-induced cell migration F. Cencetti et al.

pores) and modified Boyden chambers were obtained from
Neuroprobe (Gaithersburg, MD, USA). Diff-Quick stain-
ing solution was purchased from Dade-Behring (Lieder-
bach, Germany). Pharmacological inhibitors of SK
(VPC96091 and VPC96047) were kindly provided by K.
Lynch and T.L. MacDonald (Department of Pharmacol-
ogy, University of Virginia, Charlottesville, VA, USA).
SK2 (N-terminal region) rabbit polyclonal and SK1 (cen-
tral region) rabbit polyclonal antibodies were purchased
from ECM Biosciences LLC (Versailles, KY, USA). Sec-
ondary antibodies conjugated to horseradish peroxidase
were obtained from Santa Cruz Biotechnology, Inc. (Santa
Cruz, CA, USA).
Animals
Male C57BL/6 mice were obtained from Charles River
Laboratories (Calco, Italy). S1P3 knockout mice were
kindly provided by B. Levkau (Institute of Pathophysiol-
ogy, University of Essen, Germany), with the consent of
Prof. J. Chun, Department of Molecular and Cellular Neu-
roscience, The Scripps Research Institute, La Jolla, CA,
USA. Animals were housed in a specific pathogen-free
facility at a temperature range of 21–24 °C with a light/
dark cycle of 12 h/12 h. Food and water were constantly
available.
Mice (2–7 months old) were killed by rapid cervical dis-
location, following the procedure approved by the Ethical
Committee for Animal Experiments of the University of
Florence and EEC Guidelines for Animal Care (Directive
86/609/EEC). All efforts were made to minimize the num-
ber of the animals used in the study and reduce their suf-
fering.
Primary satellite cell isolation and culture
Individual muscle fibers containing satellite cells were iso-
lated from the tibialis anterior muscle as described previ-
ously [6,32,33]. The muscles were remove from the
hindlimb by microdissection, handling them by their ten-
dons to diminish damage to the fibers, and incubated in
0.2% w/v collagenase type I/DMEM for 1–2 h at 37 °C.Following enzymatic digestion, single myofibers were
detached by repetitively triturating the muscle with a wide-
mouth Pasteur pipette. The digested muscle was subse-
quently pre-plated into 100 mm Petri dishes for 1 h at
37 °C to remove fibroblasts, which adhere more rapidly to
the dish than satellite cells do [34]. The resulting suspension
was then plated into 24-well plates pre-coated with
1 mg�mL�1 Matrigel in a medium comprising DMEM sup-
plemented with 10% horse serum, 0.5% chick embryo
extract, 100 U�mL�1 penicillin, 100 lg�mL�1 streptomycin,
100 lg�mL�1 kanamycin, 2.5 lg�mL�1 amphotericin B and
2 mM L-glutamine (plating medium) at 37 °C in 5% CO2.
Satellite cells were released from the fibers ~ 12–24 h after
plating.
After 3–4 days, the plating medium was replaced with a
medium comprising DMEM supplemented with 20% fetal
bovine serum, 10% horse serum, 1% chick embryo extract,
100 U�mL�1 penicillin, 100 lg�mL�1 streptomycin,
100 lg�mL�1 kanamycin, 2.5 lg�mL�1 amphotericin B and
2 mM L-glutamine), which favors cell proliferation. Satellite
cells were periodically checked for expression of Pax7, a
paired-box transcription factor that is expressed by resting
satellite cells and is involved in the myogenic program [35].
Cell cultures at passage number 1-4 were used.
Cell transfection
Transfection of siRNA duplexes was performed in cells
grown in p60 Petri dishes (45 000 cells per dish), using
Lipofectamine RNAiMAX, as described previously [13].
Lipofectamine RNAiMAX was incubated with siRNA in
DMEM without serum and antibiotics at room tempera-
ture for 20 min, then the lipid/RNA complexes were com-
bined with cells with gentle agitation to a final
concentration of 85 nM in DMEM containing fetal bovine
serum. After 24 h, cells were incubated with DMEM with-
out serum but containing 1 mg�mL�1 BSA, and then used
for experiments within 72 h from the start of transfection.
The efficacy of transfection in down-regulating the expres-
sion of specific molecular targets was evaluated by real-time
RT-PCR and occasionally confirmed by western blot analy-
sis.
Western blot analysis
Activated satellite cells were lysed for 30 min at 4 °C in a
buffer containing 50 mM Tris pH 7.5, 120 mM NaCl, 1 mM
EDTA, 6 mM EGTA, 15 mM Na4P2O7, 20 mM NaF, 1%
Nonidet P-40 and protease inhibitor cocktail (1.04 mM 4-
(2-aminoethyl) benzenesulfonyl fluoride hydrochloride F,
0.08 lM aprotinin, 0.02 mM leupeptin, 0.04 mM bestatin,
15 lM pepstatin A and 14 lM N-[N-(L-3-trans-carboxyir-
ane-2-carbonyl)-L-leucyl]-agmatine; [1-[N-[(L-3-trans-carb
oxyoxirane-2-carbonyl)-L-leucyl]amino]-4-guanidinobutane]).
Cell lysates were prepared by centrifugation of cell extracts
for 15 min at 10 000 g at 4 °C. Total proteins (20 lg) fromlysates were resuspended in Laemmli’s SDS sample buffer.
Samples were subjected to SDS/PAGE for 90 min at
120 mA, before transfer of proteins to poly(vinylidene di-
fluoride) membranes. Then the membranes were washed
(10 min at room temperature in Tris-buffered saline with
0.1% Tween 20 for three times), probed overnight with the
primary antibodies at 4 °C, and subsequently with specific
secondary antibodies for 1 h at room temperature. The
presence of bound antibodies was revealed by chemilumi-
nescence.
4475FEBS Journal 281 (2014) 4467–4478 ª 2014 FEBS
F. Cencetti et al. Role of the S1P axis in LPA-induced cell migration

Real-time PCR
Total RNA (1 lg), extracted from satellite cells using TRI
Reagent, was reverse-transcribed using a high-capacity
cDNA reverse transcription kit (Applied Biosystems, Foster
City, CA, USA), according to the manufacturer’s instruc-
tions. Quantification of the mRNA level of LPA receptors,
S1P receptors and SK1/SK2 was performed by real-time
PCR using TaqMan gene expression assays with the auto-
mated ABI Prism 7700 sequence detector system (Applied
Biosystems, Foster City, CA, USA). Measurements was
performed in triplicate, as previously described [36], by con-
current amplification of the target sequence together with
that of 18S rRNA, which was chosen as a housekeeping
gene. Relative quantification of mRNA expression was per-
formed using the 2�DDCT method, and data were normal-
ized to 18S rRNA expression [37].
Cell migration
Cell migration was measured using a modified Boyden
chamber apparatus as previously described [20]. Polycar-
bonate filters with 8 lm pores were pre-coated with
Matrigel (250 lg�mL�1) for 60 min at 37 °C. Cells were
detached using 0.05% trypsin containing 0.02% EDTA,
and resuspended in DMEM containing 250 lg�mL�1 heat-
inactivated BSA. An aliquot of the cell suspension
(1.5 9 104) was loaded into the upper chamber. LPA was
dissolved in 2 mL ethanol/water (1 : 1 v/v), and divided
in aliquots into sterile Pyrex tubes, dried under nitrogen
and stored at �20 °C. When necessary, samples were
resuspended by pipetting to a concentration of 1 mM in
sterile NaCl/Pi before sonication (three times for 10 s in a
bath sonicator) and vortexing. Further dilutions were per-
formed in the medium used for the cell migration assay
(DMEM containing 250 lg�mL�1 heat-inactivated BSA).
LPA at the indicated concentrations was added to the
lower chamber, and samples were kept in an incubator at
37 °C with a 5% CO2 atmosphere for 6 h. Polycarbonate
filters were fixed overnight using methanol at 4 °C and
stained using Diff-Quick staining solution. Migration was
evaluated by counting the number of migrated cells in six
random fields per filter. When indicated, satellite cells
were pre-treated with 5 lM Ki16425 for 40 min before
being trypsinized. Treatment with SK inhibitors was
accomplished incubating 1.5 9 104 cells in the presence of
1 lM VPC96047 or 1 lM VPC96091 in the upper well of
a Boyden chamber.
The scratch wound healing assay was performed as
described previously [6]. Satellite cells were seeded in 12-
well tissue culture plates and incubated at 37 °C with 5%
CO2 for 24 h until confluence. Cells were then serum-
starved overnight, and wounded by scratching with a
200 lL standard sterile pipette tip. The cell monolayer was
washed twice with 5 ml DMEM containing 0.1 % BSA at
room temperature for 2 min in order to remove cell debris,
and incubated with or without 1 lM LPA. At least six
scratched areas for each sample were photographed using a
Nikon (Amsterdam, The Netherlands) digital camera con-
nected to a Nikon phase-contrast extra long working
distance 0.3. Migration of satellite cells was quantified by
counting the number of cells that had migrated into the
scratched area after 7 h compared to the number present in
the initial wound.
Statistical analysis
Densitometric analysis of the western blot bands was per-
formed using IMAGEJ software (http://imagej.nih.gov/ij/) and
QUANTITY ONE analysis software (Bio-Rad). Graphical repre-
sentations were created using GRAPHPAD PRISM 5.0 (Graph-
Pad Software, San Diego, CA, USA). Statistical analysis
was performed using Student’s t test, one-way ANOVA
and two-way ANOVA followed by Bonferroni’s post hoc
test. Asterisks and other symbols indicate statistical signifi-
cance at the levels indicated in the legends.
Acknowledgements
This research was supported by grants from the
University of Florence to C.D., and from the Fond-
azione Cassa di Risparmio di Lucca (BRUNICRL12)
to P.B. The authors are indebted to K.L. Lynch and
T.L. MacDonald (University of Virginia, Charlottes-
ville, VA), who kindly provided VPC96047 and
VPC96091, and to J. Chun (Scripps Research Insti-
tute, La Jolla, CA) and B. Levkau (Institute of Path-
ophysiology, University of Essen, Germany), for
generating and providing S1P3 knockout mice,
respectively.
Author contributions
F.C., C.D. and P.B. planned the experiments; F.C.,
G.B., C.B. and S.B. performed the experiments; F.C.,
G.B. and S.B. analyzed the data; P.B. and C.D. wrote
the paper.
References
1 Ishii I, Fukushima N, Ye X & Chun J (2004)
Lysophospholipid receptors: signaling and biology.
Annu Rev Biochem 73, 321–354.
2 Pebay A, Bonder CS & Pitson SM (2007) Stem cell
regulation by lysophospholipids. Prostaglandins Other
Lipid Mediat 84, 83–97.
3 Bruni P & Donati C (2013) Role of sphingosine
1-phosphate in skeletal muscle cell biology. Handb
Exp Pharmacol 216, 457–467.
4476 FEBS Journal 281 (2014) 4467–4478 ª 2014 FEBS
Role of the S1P axis in LPA-induced cell migration F. Cencetti et al.

4 Donati C, Cencetti F & Bruni P (2013) Sphingosine
1-phosphate axis: a new leader actor in skeletal muscle
biology. Front Physiol 4, 338.
5 Nagata Y, Partridge TA, Matsuda R & Zammit PS
(2006) Entry of muscle satellite cells into the cell cycle
requires sphingolipid signaling. J Cell Biol 174, 245–253.
6 Calise S, Blescia S, Cencetti F, Bernacchioni C, Donati
C & Bruni P (2012) Sphingosine 1-phosphate stimulates
proliferation and migration of satellite cells: role of S1P
receptors. Biochim Biophys Acta 1823, 439–450.
7 Loh KC, Leong WI, Carlson ME, Oskouian B, Kumar
A, Fyrst H, Zhang M, Proia RL, Hoffman EP & Saba
JD (2012) Sphingosine-1-phosphate enhances satellite
cell activation in dystrophic muscles through a S1PR2/
STAT3 signaling pathway. PLoS ONE 7, e37218.
8 Germinario E, Peron S, Toniolo L, Betto R, Cencetti F,
Donati C, Bruni P & Danieli-Betto D (2012) S1P2
receptor promotes mouse skeletal muscle regeneration.
J Appl Physiol 113, 707–713.
9 Fortier M, Figeac N, White RB, Knopp P & Zammit
PS (2013) Sphingosine-1-phosphate receptor 3 influences
cell cycle progression in muscle satellite cells. Dev Biol
382, 504–516.
10 Donati C, Meacci E, Nuti F, Becciolini L, Farnararo
M & Bruni P (2005) Sphingosine 1-phosphate regulates
myogenic differentiation: a major role for S1P2
receptor. FASEB J 19, 449–451.
11 Meacci E, Nuti F, Donati C, Cencetti F, Farnararo M
& Bruni P (2008) Sphingosine kinase activity is required
for myogenic differentiation of C2C12 myoblasts. J Cell
Physiol 214, 210–220.
12 Donati C, Nincheri P, Cencetti F, Rapizzi E, Farnararo
M & Bruni P (2007) Tumor necrosis factor-a exerts
pro-myogenic action in C2C12 myoblasts via
sphingosine kinase/S1P2 signaling. FEBS Lett 581,
4384–4388.
13 Cencetti F, Bernacchioni C, Nincheri P, Donati C &
Bruni P (2010) Transforming growth factor-b1 induces
transdifferentiation of myoblasts into myofibroblasts
via up-regulation of sphingosine kinase-1/S1P3 axis.
Mol Biol Cell 21, 1111–1124.
14 Nincheri P, Bernacchioni C, Cencetti F, Donati C &
Bruni P (2010) Sphingosine kinase-1/S1P1 signalling
axis negatively regulates mitogenic response elicited by
PDGF in mouse myoblasts. Cell Signal 22, 1688–1699.
15 Bernacchioni C, Cencetti F, Blescia S, Donati C &
Bruni P (2012) Sphingosine kinase/sphingosine 1-
phosphate axis: a new player for insulin-like growth
factor-1-induced myoblast differentiation. Skelet Muscle
2, 15.
16 Cencetti F, Bernacchioni C, Tonelli F, Roberts E,
Donati C & Bruni P (2013) TGFb1 evokes myoblast
apoptotic response via a novel signaling pathway
involving S1P4 transactivation upstream of Rho-kinase-
2 activation. FASEB J 27, 4532–4546.
17 Yoshida S, Fujisawa-Sehara A, Taki T, Arai K &
Nabeshima Y (1996) Lysophosphatidic acid and bFGF
control different modes in proliferating myoblasts.
J Cell Biol 132, 181–193.
18 Xu YJ, Tappia PS, Goyal RK & Dhalla NS (2008)
Mechanisms of the lysophosphatidic acid-induced
increase in [Ca2+]i in skeletal muscle cells. J Cell Mol
Med 12, 942–954.
19 Minetti GC, Feige JN, Rosenstiel A, Bombard F,
Meier V, Werner A, Bassilana F, Sailer AW, Kahle P,
Lambert C et al. (2011) Gai2 signaling promotes
skeletal muscle hypertrophy, myoblast differentiation,
and muscle regeneration. Sci Signal 4, ra80.
20 Becciolini L, Meacci E, Donati C, Cencetti F, Rapizzi
E & Bruni P (2006) Sphingosine 1-phosphate inhibits
cell migration in C2C12 myoblasts. Biochim Biophys
Acta 1761, 43–51.
21 Aoki J, Taira A, Takanezawa Y, Kishi Y, Hama K,
Kishimoto T, Mizuno K, Saku K, Taguchi R & Arai H
(2002) Serum lysophosphatidic acid is produced
through diverse phospholipase pathways. J Biol Chem
277, 48737–48744.
22 Moolenaar WH & Perrakis A (2011) Insights into
autotaxin: how to produce and present a lipid
mediator. Nat Rev 12, 674–679.
23 Giganti A, Rodriguez M, Fould B, Moulharat N, Coge
F, Chomarat P, Galizzi JP, Valet P, Saulnier-Blache JS,
Boutin JA et al. (2008) Murine and human autotaxin a,b and c isoforms: gene organization, tissue distribution,
and biochemical characterization. J Biol Chem 283,
7776–7789.
24 Oikonomou N, Mouratis MA, Tzouvelekis A, Kaffe E,
Valavanis C, Vilaras G, Karameris A, Prestwich GD,
Bouros D & Aidinis V (2012) Pulmonary autotaxin
expression contributes to the pathogenesis of
pulmonary fibrosis. Am J Respir Cell Mol Biol 47,
566–574.
25 Lee Z, Cheng CT, Zhang H, Subler MA, Wu J,
Mukherjee A, Windle JJ, Chen CK & Fang X (2008)
Role of LPA4/p2y9/GPR23 in negative regulation of
cell motility. Mol Biol Cell 19, 5435–5445.
26 Jongsma M, Matas-Rico E, Rzadkowski A, Jalink K &
Moolenaar WH (2011) LPA is a chemorepellent for
B16 melanoma cells: action through the
cAMP-elevating LPA5 receptor. PLoS One 6, e29260.
27 Komachi M, Tomura H, Malchinkhuu E, Tobo M,
Mogi C, Yamada T, Kimura T, Kuwabara A, Ohta H,
Im DS et al. (2009) LPA1 receptors mediate
stimulation, whereas LPA2 receptors mediate
inhibition, of migration of pancreatic cancer cells in
response to lysophosphatidic acid and malignant
ascites. Carcinogenesis 30, 457–465.
28 Choi JW & Chun J (2013) Lysophospholipids and their
receptors in the central nervous system. Biochim
Biophys Acta 1831, 20–32.
4477FEBS Journal 281 (2014) 4467–4478 ª 2014 FEBS
F. Cencetti et al. Role of the S1P axis in LPA-induced cell migration

29 Shida D, Fang X, Kordula T, Takabe K, Lepine S,
Alvarez SE, Milstien S & Spiegel S (2008) Cross-talk
between LPA1 and epidermal growth factor receptors
mediates up-regulation of sphingosine kinase 1 to
promote gastric cancer cell motility and invasion.
Cancer Res 68, 6569–6577.
30 Igarashi N, Okada T, Hayashi S, Fujita T, Jahangeer S
& Nakamura S (2003) Sphingosine kinase 2 is a nuclear
protein and inhibits DNA synthesis. J Biol Chem 278,
46832–46839.
31 Hait NC, Sarkar S, Le Stunff H, Mikami A, Maceyka
M, Milstien S & Spiegel S (2005) Role of sphingosine
kinase 2 in cell migration toward epidermal growth
factor. J Biol Chem 280, 29462–29469.
32 Rosenblatt JD, Lunt AI, Parry DJ & Partridge TA
(1995) Culturing satellite cells from living single
muscle fiber explants. In vitro Cell Dev Biol 31,
773–779.
33 Cossu G, Zani B, Coletta M, Bouche M, Pacifici M &
Molinaro M (1980) In vitro differentiation of satellite
cells isolated from normal and dystrophic mammalian
muscles. A comparison with embryonic myogenic cells.
Cell Differ 9, 357–368.
34 Musaro A & Barberi L (2010) Isolation and culture
of mouse satellite cells. Methods Mol Biol 633,
101–111.
35 Seale P, Sabourin LA, Girgis-Gabardo A, Mansouri A,
Gruss P & Rudnicki MA (2000) Pax7 is required for
the specification of myogenic satellite cells. Cell 102,
777–786.
36 Donati C, Cencetti F, Nincheri P, Bernacchioni C,
Brunelli S, Clementi E, Cossu G & Bruni P (2007)
Sphingosine 1-phosphate mediates proliferation and
survival of mesoangioblasts. Stem Cells 25,
1713–1719.
37 Livak KJ & Schmittgen TD (2001) Analysis of relative
gene expression data using real-time quantitative PCR
and the 2�DDCT method. Methods 25, 402–408.
4478 FEBS Journal 281 (2014) 4467–4478 ª 2014 FEBS
Role of the S1P axis in LPA-induced cell migration F. Cencetti et al.





























