Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Mapping of NPR-B immunoreactivity in the brainstemof Macaca fascicularis
Essam M. Abdelalim • Ikuo Tooyama
Received: 17 November 2010 / Accepted: 21 March 2011 / Published online: 1 April 2011
� Springer-Verlag 2011
Abstract C-type natriuretic peptide (CNP), the most
abundant natriuretic peptide hormone in the brain, plays an
important role in neuroendocrine function. The physio-
logical effects of CNP are mediated by the natriuretic
peptide receptor-B (NPR-B). Although CNP and NPR-B
have been detected in several brain regions, little is known
about the neuroanatomical localization of NPR-B protein
in the brainstem. In the present study, we investigated the
topographical distribution of NPR-B immunoreactivity in
the monkey brainstem. The data demonstrate widespread
NPR-B immunoreactivity throughout the brainstem. NPR-B
immunoreactivity was located in the superior colliculus,
inferior colliculus, periaqueductal gray, oculomotor
nucleus, red nucleus, ventral tegmental area, substantia
nigra, and cerebral peduncle of the midbrain, as well as in
the abducens nucleus, medial vestibular nucleus, lateral
vestibular nucleus, parabrachial nucleus, locus coeruleus,
trigeminal motor nucleus, pontine reticular nucleus, facial
nucleus, oral part of the spinal trigeminal nucleus, cochlear
nucleus, raphe magnus nucleus, raphe pallidus nucleus,
pontine nucleus of the pons, the dorsal motor nucleus of the
vagus, hypoglossal nucleus, nucleus tractus solitarius,
gracile nucleus, cuneate nucleus, medial vestibular nucleus,
spinal trigeminal nucleus, nucleus ambiguus, lateral para-
gigantocellular nucleus, lateral reticular nucleus, and the
inferior olivary nucleus of the medulla oblongata. The
widespread distribution of NPR-B-immunoreactive struc-
tures throughout the monkey brainstem indicates that CNP
may be involved in several physiological mechanisms,
acting as a neurotransmitter and/or neuromodulator.
Keywords Neuropeptide � NPR-B � Midbrain � Pons �Medulla � Primates
Introduction
The natriuretic peptide receptor-B (NPR-B), also known as
guanylyl cyclase (GC)-B, is one of the natriuretic peptide
receptors that bind natriuretic peptide hormones. The
natriuretic peptides are a family of three structurally related
hormones, namely atrial natriuretic peptide (ANP), B-type
natriuretic peptide (BNP), and C-type natriuretic peptide
(CNP) (Nakao et al. 1992; Anand-Srivastava and Trachte
1993; Espiner et al. 1995). The actions of these natriuretic
peptides are mediated by binding to the natriuretic peptide
receptors. ANP and BNP elicit their effects by binding and
activating the cell surface NPR-A receptor, whereas CNP
activates the homologous NPR-B receptor (Koller et al.
1991; Kuhun 2003; Garbers et al. 2006; Potter et al. 2006).
All three natriuretic peptides also bind the natriuretic
peptide clearance receptor NPR-C, which chiefly regulates
local concentrations of the natriuretic peptides through
receptor-mediated internalization and degradation (Maack
1992; Matsukawa et al. 1999).
In addition to established roles for natriuretic peptides in
regulating neuroendocrine and cardiovascular function (see
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00429-011-0313-1) contains supplementarymaterial, which is available to authorized users.
E. M. Abdelalim (&) � I. Tooyama
Molecular Neuroscience Research Center,
Shiga University of Medical Science, Setatsukinowa-cho,
Otsu, Shiga 520-2192, Japan
e-mail: [email protected]
E. M. Abdelalim
Department of Cytology and Histology,
Faculty of Veterinary Medicine, Suez Canal University,
Ismailia, Egypt
123
Brain Struct Funct (2011) 216:387–402
DOI 10.1007/s00429-011-0313-1

Imura et al. 1992; Gutkowska et al. 1997; Levin et al.
1998), these peptides have other functions. For example,
natriuretic peptides enhance the survival of PC12 and
embryonic basal forebrain cells (Fiscus et al. 2001). In
addition, activation of the CNP/NPR-B system in chon-
drocytes of the growth plate can be used as a therapy
for achondroplasia in humans (Yasoda et al. 2004) and
Sabbatini et al. (2005) found that centrally applied CNP
enhances pancreatic secretion by activating its receptor.
Recently, the CNP/NPR-B system was found to be
involved in the regulation of bidirectional plasticity in the
CA1 area of the hippocampus (Decker et al. 2010).
CNP predominates in the central nervous system, with
highest levels of CNP mRNA found in the olfactory nuclei,
limbic cortex, hippocampus, amygdala, hypothalamus, and
brainstem (Garbers 1992). The presence of natriuretic
peptides and their receptors has been reported in monkey
brain (Wilcox et al. 1991; Abdelalim et al. 2006, 2007),
indicating a central role for natriuretic peptides in this
species. A previous study in the monkey reported that
NPR-B mRNA expression was confined to the adrenal
medulla, pituitary, and cerebellum (Wilcox et al. 1991).
In the rat, NPR-B mRNA is expressed in the limbic
cortex, neocortex, olfactory bulb, hippocampus, amygdala,
preoptic–hypothalamic neuroendocrine circuits, ventral
tegmental area (VTA), substantia nigra, and in the motor
nuclei of the cranial nerves. Intermediate expression of
NPR-B mRNA has been found in brainstem nuclei con-
trolling autonomic function (Langub et al. 1995; Herman
et al. 1996; Thiriet et al. 2001). Although NPR-B mRNA
has been demonstrated in astrocyte cultures (Deschepper
and Picard 1994), an in situ hybridization study in the rat
brain failed to detect NPR-B signals in the glia (Herman
et al. 1996). In the rat brain, NPR-B protein immunore-
activity has been found in the VTA, substantia nigra,
caudate–putman, nucleus accumbens, frontal cortex, hip-
pocampus, cortex, and cerebellum (Thiriet et al. 2001).
The physiological effects of CNP in the brain depend on
the distribution of its receptors. To our knowledge, the
expression of NPR-B protein has not been investigated in
detail in the brainstem of any species. To further under-
stand the central actions of CNP in the brainstem, we
examined NPR-B immunoreactivity in the brainstem of the
cynomologus monkey (Macaca fascicularis) in the present
study using immunohistochemical techniques.
Materials and methods
Tissue preparation
Samples were obtained from the brains of cynomologus
monkeys (M. fascicularis), after animals that had been used
for other research purposes by other investigators had
been killed. The animal use protocols in the present study
were approved by the Institutional Animal Care and Use
Committee (IACUC) of Shiga University of Medical
Science.
The brains were collected at different times and handled
individually. All brains from the three monkeys were
subjected to the same procedures. The brain was removed
from two males and one female of cynomologus monkeys
(3, 10, and 12 years) and fixed immediately in 4% para-
formaldehyde in 0.1 M phosphate buffer (pH 7.4) for 2
days at 4�C before being immersed in 15% sucrose in
0.1 M phosphate buffer (pH 7.4) with 0.1% sodium azide
for cryoprotection. The sucrose solution was changed every
day for 4 days, after which time the brains were stored in
sucrose at 4�C until sectioning. Fixed tissues were sub-
jected to cryostat sectioning at –20�C.
Western blotting
Western blot analysis was performed to confirm the spec-
ificity of the NPR-B antibody. Briefly, rat brain and mon-
key heart samples were homogenized in ice-cold 50 mM
Tris–HCl buffer (pH 7.4) containing protease inhibitors
(Complete Mini; Roche Diagnostics, Mannheim, Ger-
many). After initial centrifugation at 800g for 5 min at 4�C
to remove tissue fragments, the supernatant fraction was
centrifuged at 100,000g for 1 h at 4�C. The supernatant
was collected and considered as the cytosolic fraction,
whereas the pellets (i.e. the membrane fraction) were
resuspended in RIPA buffer (50 mM Tris–HCl, 150 mM
NaCl, 1% Nonidet P-40, 0.25% Na-deoxycholate, 1 mM
phenylmethylsulfonyl fluoride, and 19 Complete Mini
protease inhibitor cocktail).
Protein samples (40 lg) from the cytosolic and mem-
brane fractions were electrophoresed by sodium dodecyl
sulfate–polyacrylamide gel electrophoresis (SDS–PAGE;
7% gels) and transferred onto polyvinylidene difluoride
membranes. Non-specific protein binding sites were
blocked by incubating membranes for 2 h at room tem-
perature in 10% skim milk in 25 mM Tris-buffered saline
containing 0.1% Tween-20 (TBST; pH 7.4), followed by
4 h incubation at 4�C with a rabbit polyclonal antibody
against NPR-B (Santa Cruz Biotechnology) at a dilution of
1:1000 in TBST. Blots were washed twice for 10 min each
time with 25 mM TBST before being incubated for 1 h at
room temperature with peroxidase-conjugated anti-rabbit
IgG (1:10000 dilution; Jackson ImmunoResearch Labora-
tories, Inc.). After extensive washing with 25 mM TBST,
blots were developed using SuperSignal West Pico
Chemiluminescent substrate (Pierce) and visualized using
an LAS-3000 FujiFilm Lumino-Image Analyzer (FujiFilm,
Tokyo, Japan).
388 Brain Struct Funct (2011) 216:387–402
123

Immunohistochemistry
Serial cryostat sections (20 lm) were floated in 0.1 M
phosphate-buffered saline with 0.3% Triton X-100, pH 7.4
(PBST), for 4 days at 4�C and treated in a free-floating state.
Endogenous peroxidase activity was blocked by incubating
the sections in 0.5% H2O2 in PBST for 30 min. After sections
had been rinsed three times with PBST for 10 min each time,
they were incubated in 4% normal horse serum (Invitrogen)
for 40 min at room temperature, and then they were incu-
bated for 72 h at 4�C with a rabbit polyclonal antibody
against human NPR-B (1:400 dilution; Santa Cruz Bio-
technology) or a polyclonal rabbit anti-NPR-B antibody
(1:300 dilution; Abgent, Sant Diego, CA, USA). The primary
antibody was diluted in 1% normal horse serum in PBST.
After sections had been rinsed three times with PBST
(10 min each time), they were incubated for 1 h at room
temperature with biotinylated goat anti-rabbit IgG (1:3000
dilution; Vector Laboratories, Burlingame, CA, USA)
before being rinsed another three time with PBST. The
sections were finally incubated for 1 h at room temperature
with an avidin–biotin–peroxidase complex (1:4000 dilution;
ABC Elite; Vector Laboratories) and the peroxidase-labeled
sections were developed in 0.02% 3,3-diamine-benzidine
tetrahydrochloride with 0.07% nickel ammonium sulfate in
50 mM Tris–HCl (pH 7.6), with 0.005% hydrogen peroxide.
Double immunohistochemistry
For double immunostaining we used 4% normal horse
serum as a blocking solution. After the first NPR-B
immunostaining which produced a purple color, the sec-
tions were treated for 30 min at room temperature with
0.5% H2O2 in PBST to eliminate residual horseradish
peroxidase activity. The sections were then washed in
PBST and incubated with mouse monoclonal anti-tyrosine
hydroxylase (TH) antibody (1:3000; Chemicon). After
three rinses with PBST, the sections were incubated for 2 h
at room temperature with histofine anti-mouse IgG com-
plex (1:100; Nichirei, Japan). After thee further rinses with
PBST, the sections were developed in 0.02% 3,3-diamine-
benzidine tetrahydrochloride in 50 mM Tris–HCl (pH 7.6),
with 0.005% hydrogen peroxide, to yield a brown color.
Mapping
Mapping was performed using a camera lucida, and diagrams
of the midbrain, pons, and medulla oblongata were prepared.
Data analysis
The major divisions and nuclei of the brainstem were
identified according to Szabo and Cowan (1984). Images of
selected immunostained sections were captured and orga-
nized into photographic panels. Adobe Photoshop was used
to view the images. To improve visualization of the results,
only the brightness and contrast of the images were
adjusted using Adobe Photograph, without any further
manipulation of the images.
Results
Specificity of the NPR-B antibody
Since the Western blotting has been reported as a strong
tool to determine the antibody specificity (Burry 2000), the
specificity of the NPR-B antibody was examined by wes-
tern blotting. The antibody recognized a single band at
approximately 120 kDa in the membrane fractions of the
rat brain and monkey heart which corresponded to NPR-B
protein (Fig. 1a, b).
Also, we compared the sequence of the immunogen used
to produce the NPR-B antibody with the corresponding
sequences of the monkey and rat (Fig. 1c). The antibody
used in this study is a polyclonal antibody raised against
the amino acids 171–250 of NPR-B of human origin. The
comparison of these 80 amino acids sequences of NPR-B
between human and monkey showed that only one amino
acid is different between human and monkey (98.75%
identity). Also, comparing this region between human and
rat showed 98.75% identity, indicating that the antibody
used in the current study can recognize NPR-B in monkey
and rat tissues (Fig. 1c).
Additionally, we examined the immunolabeling of NPR-
B in monkey brainstem using another antibody (from dif-
ferent company) raised against NPR-B. This antibody has
been previously used in other studies (Abdelalim et al.
2008b). The antibody showed identical staining patterns in
the brainstem of monkey (supplementary Fig. 1). Collec-
tively, these findings suggest that the protein recognized by
the NPR-B antibody in the present study is indeed, NPR-B.
Distribution of NPR-B-immunoreactive structures
in the monkey brainstem
NPR-B immunoreactivity was distributed extensively in
neuronal structures throughout the brainstem of the cyno-
mologus monkey. Examination of the immunostained
sections from the three monkeys showed no difference in
the pattern of the distribution of NPR-B immunoreactivity.
Figure 2 summarizes the distribution of NPR-B immuno-
reactivity in the midbrain (Fig. 2a, b), pons (Fig. 2c, d),
and medulla oblongata (Fig. 2e–g), as drawn using the
camera lucida. As shown in Fig. 2, NPR-B-positive neu-
rons were observed in specific brain regions. The highest
Brain Struct Funct (2011) 216:387–402 389
123

density of NPR-B-immunoreactive structures was observed
in the pons, followed by the medulla oblongata.
Distribution of NPR-B-immunoreactive structures
in the midbrain
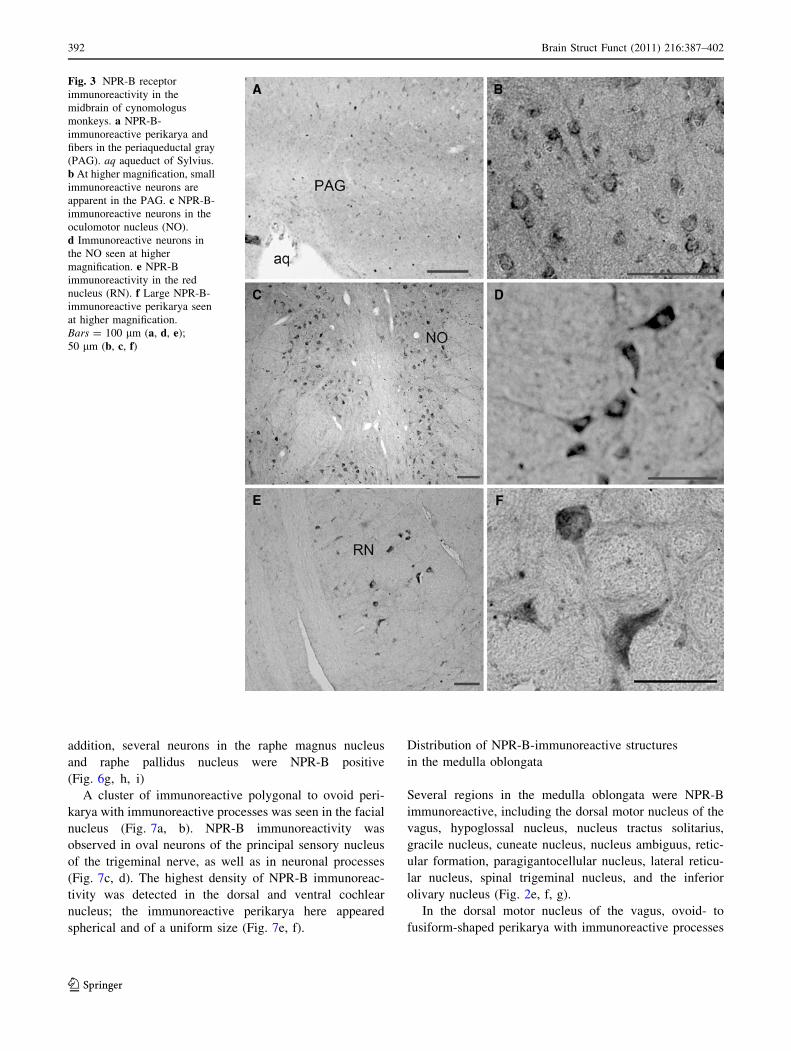
In the midbrain, NPR-B immunoreactivity was observed in
the superior and inferior colliculi, periaqueductal gray
(PAG), medial lemniscus, oculomotor nucleus, red nucleus,
cerebral peduncle, VTA, and substantia nigra (Fig. 2a, b).
In the PAG, several NPR-B-immunoreactive neurons
were seen surrounding the aqueduct of Sylvius (Fig. 3a, b).
High-intensity NPR-B immunoreactivity was observed in
different-shaped neurons in the oculomotor nucleus, with
several neurons having immunoreactive processes (Fig. 3c,
d). The highest intensity of NPR-B immunoreactivity in the
midbrain was seen in the large neurons of the red nucleus
(Fig. 3e, f).
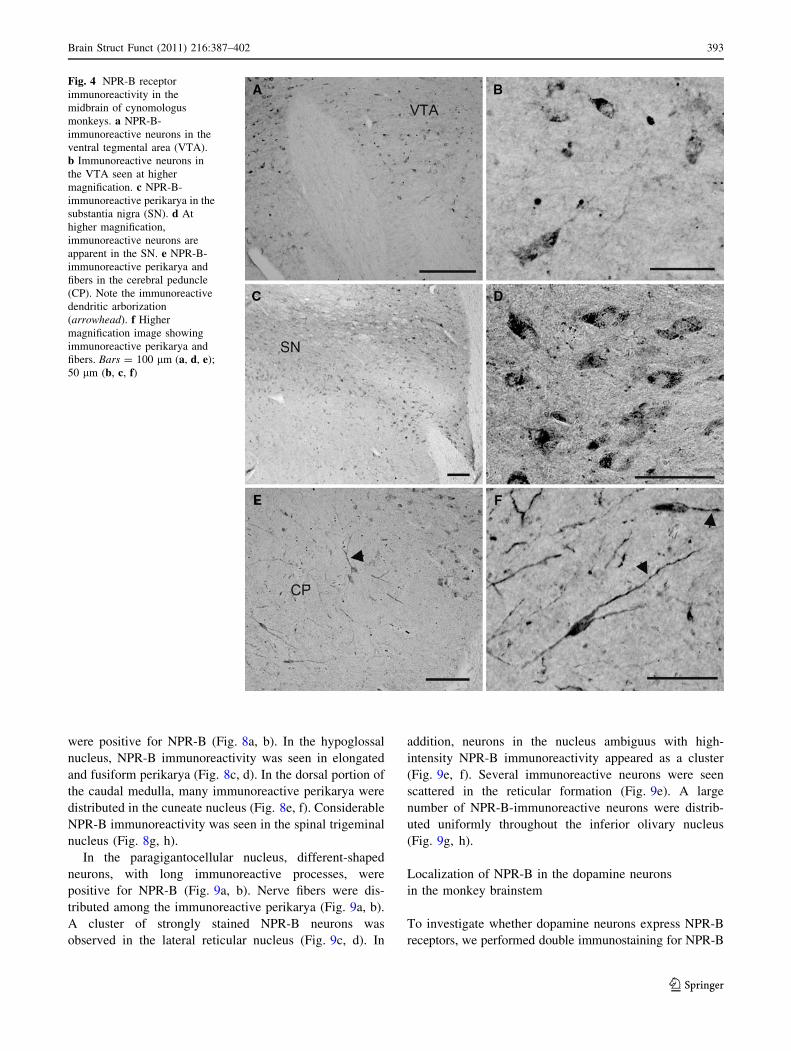
NPR-B immunoreactivity was observed in the VTA
(Fig. 4a, b). At the level of the substantia nigra, there was a
large number of closely packed, moderately NPR-B-
immunoreactive neurons (Fig. 4c, d). In the cerebral
peduncle, intense NPR-B immunoreactivity appeared in
elongated perikarya and their processes (Fig. 4e, f). Some
immunoreactive primary dendrites divided into secondary,
then tertiary dendrites (Fig. 4e). Intensely NPR-B-immu-
noreactive axons were found in the cerebral peduncle area
(Fig. 4e, f).
Distribution of NPR-B-immunoreactive structures
in the pons
The highest density of NPR-B immunoreactivity was found
in the pons. NPR-B immunoreactivity was seen in neurons
in several regions, including the abducens nucleus, medial
vestibular nucleus, parabrachial nucleus, lateral vestibular
nucleus, facial nucleus, oral part of the spinal trigeminal
nucleus, cochlear nucleus, locus coeruleus, trigeminal
motor nucleus, pontine reticular formation, pontine
nuclei, raphe magnus nucleus, and raphe pallidus nucleus
(Fig. 2c, d).
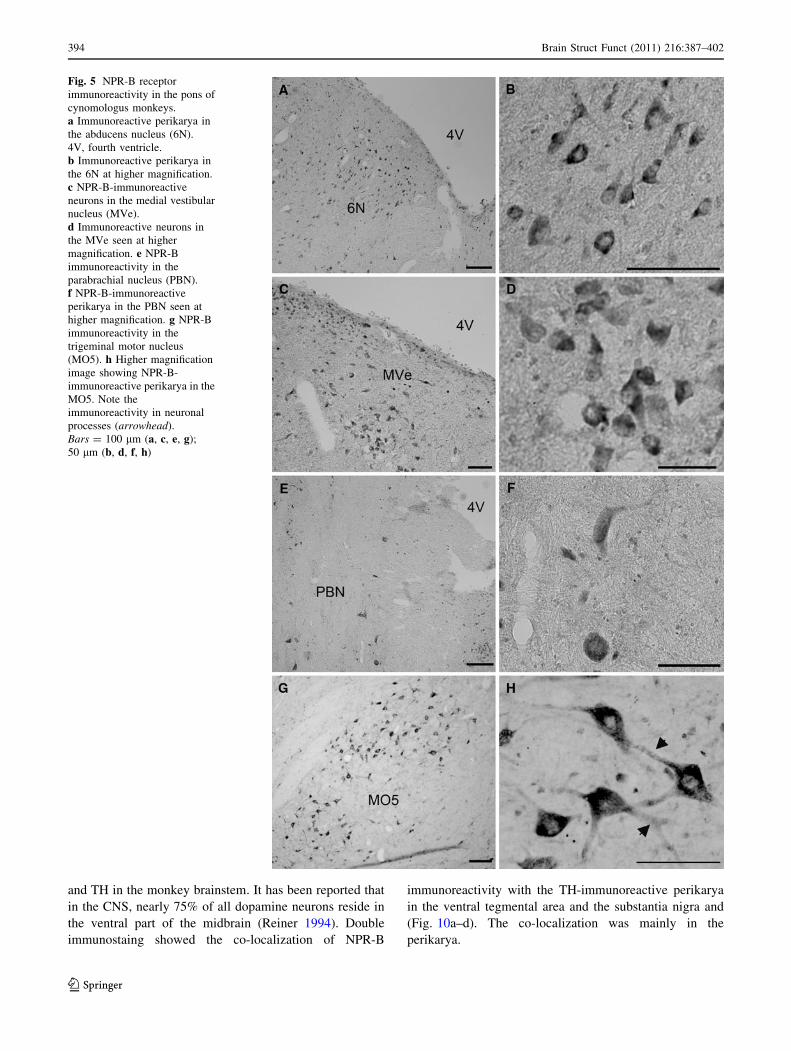
Strongly stained NPR-B-immunoreactive neurons were
detected just under the fourth ventricle in the abducens
nucleus (Fig. 5a, b). In the medial vestibular nucleus, oval
to fusiform immunoreactive neurons were observed
(Fig. 5c, d). Immunoreactive neurons with different shapes
were observed scattered throughout the parabrachial
nucleus (Fig. 5e, f) and immunoreactive neuronal pro-
cesses were seen in some neurons (Fig. 5f). High-intensity
NPR-B immunoreactivity was observed in the trigeminal
motor nucleus (Fig. 5g, h).
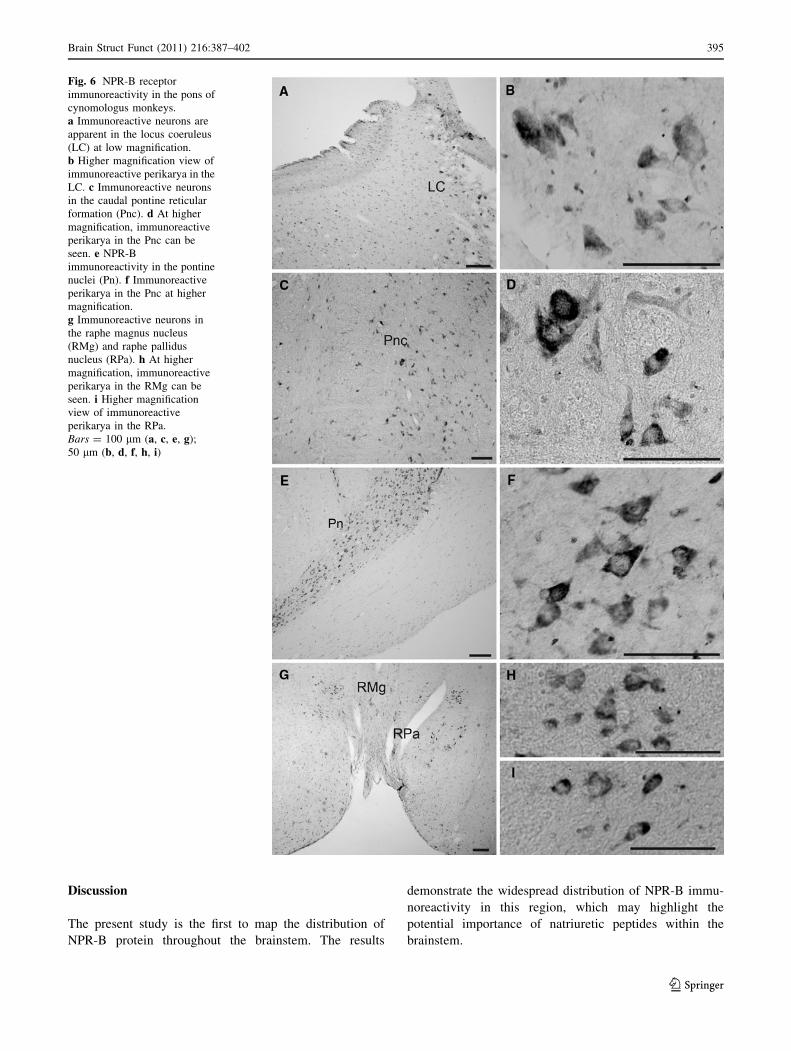
NPR-B-immunoreactive perikarya were detected in the
locus coeruleus (Fig. 6a, b). Diffuse distribution of dif-
ferent-sized, intensely NPR-B-immunoreactive neurons
were observed in the pontine reticular nucleus (Fig. 6c, d).
In the pontine nucleus, a high density of NPR-B-
immunoreactive neurons was observed (Fig. 6e, f). In
Fig. 1 Antibody specificity for
the NPR-B receptor. Western
blots were performed on
extracts from rat brain (a) and
monkey heart (b). Adjacent
lanes were loaded with
equivalent amounts of protein
from membrane and cytosolic
fractions. The antibody
recognized a single band at
approximately 120 kDa in the
membrane fraction only in rat
brain and monkey heart.
c Comparison of the amino acid
sequences of the human
immunogen (from 171 to 250)
used to produce NPR-B
antibody with those of monkey
and rat.The GeneBank
accession numbers are human
NPR-B: NM_003995, monkey
NPR-B: XM_001085221, and
rat (NM_053838). Dots (.) show
identical amino acids
390 Brain Struct Funct (2011) 216:387–402
123
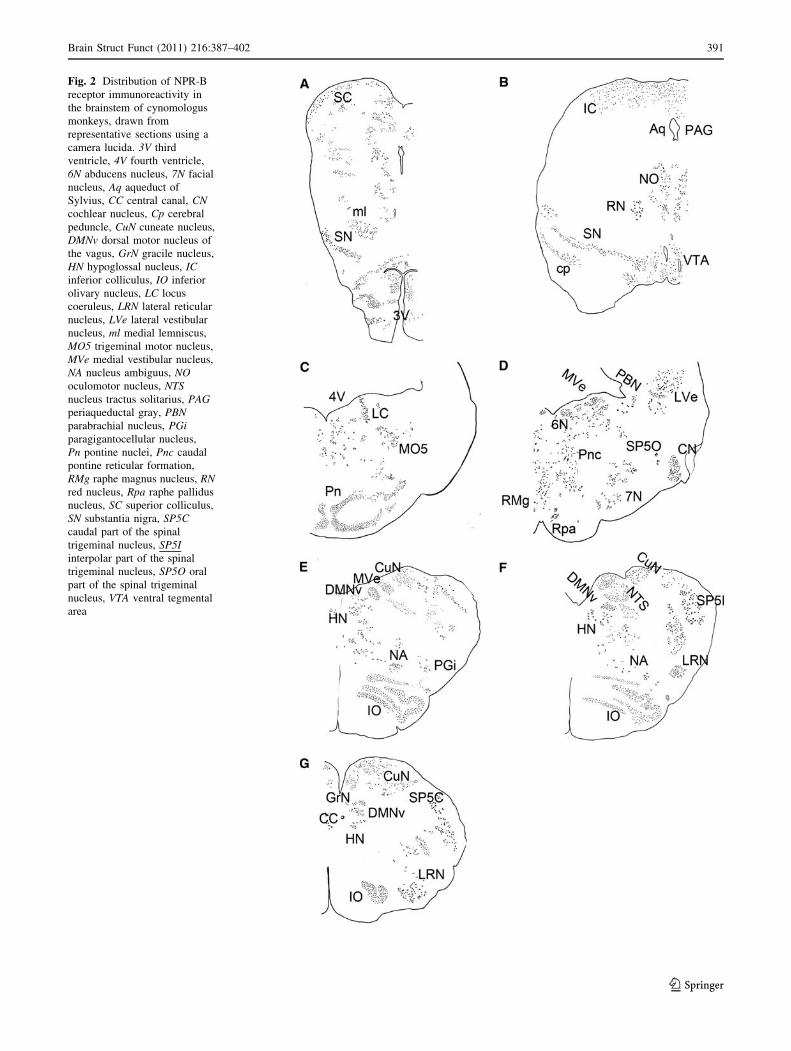
Fig. 2 Distribution of NPR-B
receptor immunoreactivity in
the brainstem of cynomologus
monkeys, drawn from
representative sections using a
camera lucida. 3V third
ventricle, 4V fourth ventricle,
6N abducens nucleus, 7N facial
nucleus, Aq aqueduct of
Sylvius, CC central canal, CNcochlear nucleus, Cp cerebral
peduncle, CuN cuneate nucleus,
DMNv dorsal motor nucleus of
the vagus, GrN gracile nucleus,
HN hypoglossal nucleus, ICinferior colliculus, IO inferior
olivary nucleus, LC locus
coeruleus, LRN lateral reticular
nucleus, LVe lateral vestibular
nucleus, ml medial lemniscus,
MO5 trigeminal motor nucleus,
MVe medial vestibular nucleus,
NA nucleus ambiguus, NOoculomotor nucleus, NTSnucleus tractus solitarius, PAGperiaqueductal gray, PBNparabrachial nucleus, PGiparagigantocellular nucleus,
Pn pontine nuclei, Pnc caudal
pontine reticular formation,
RMg raphe magnus nucleus, RNred nucleus, Rpa raphe pallidus
nucleus, SC superior colliculus,
SN substantia nigra, SP5Ccaudal part of the spinal
trigeminal nucleus, SP5Iinterpolar part of the spinal
trigeminal nucleus, SP5O oral
part of the spinal trigeminal
nucleus, VTA ventral tegmental
area
Brain Struct Funct (2011) 216:387–402 391
123
addition, several neurons in the raphe magnus nucleus
and raphe pallidus nucleus were NPR-B positive
(Fig. 6g, h, i)
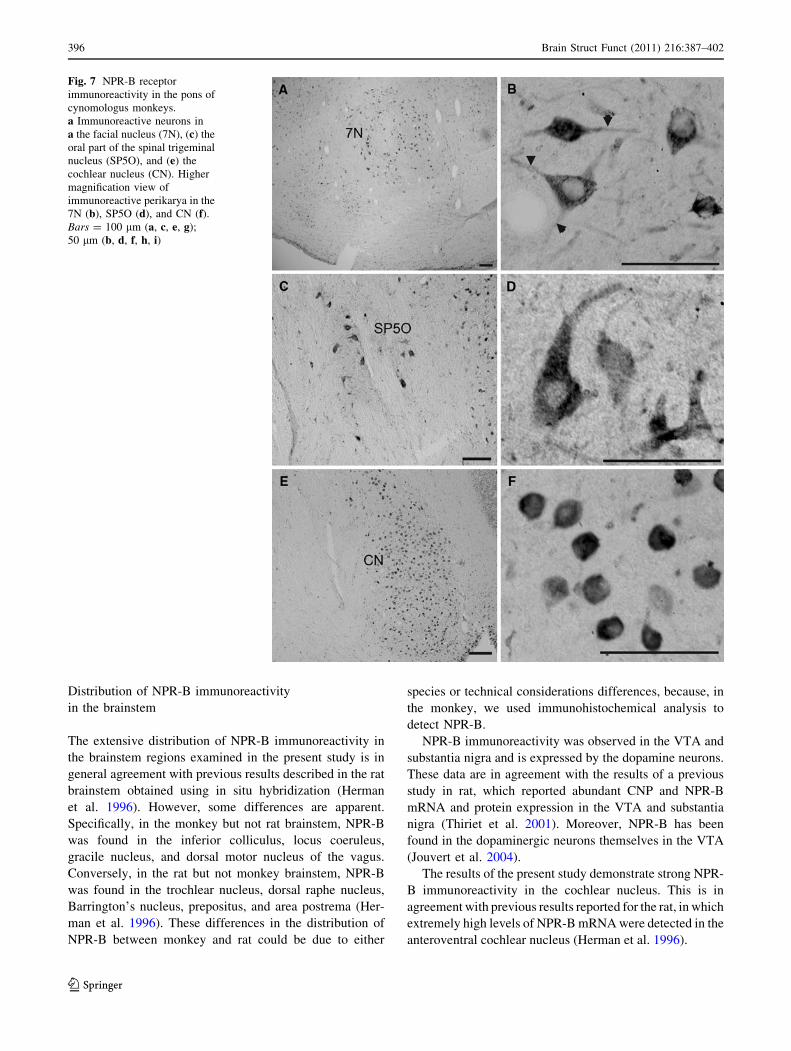
A cluster of immunoreactive polygonal to ovoid peri-
karya with immunoreactive processes was seen in the facial
nucleus (Fig. 7a, b). NPR-B immunoreactivity was
observed in oval neurons of the principal sensory nucleus
of the trigeminal nerve, as well as in neuronal processes
(Fig. 7c, d). The highest density of NPR-B immunoreac-
tivity was detected in the dorsal and ventral cochlear
nucleus; the immunoreactive perikarya here appeared
spherical and of a uniform size (Fig. 7e, f).
Distribution of NPR-B-immunoreactive structures
in the medulla oblongata
Several regions in the medulla oblongata were NPR-B
immunoreactive, including the dorsal motor nucleus of the
vagus, hypoglossal nucleus, nucleus tractus solitarius,
gracile nucleus, cuneate nucleus, nucleus ambiguus, retic-
ular formation, paragigantocellular nucleus, lateral reticu-
lar nucleus, spinal trigeminal nucleus, and the inferior
olivary nucleus (Fig. 2e, f, g).
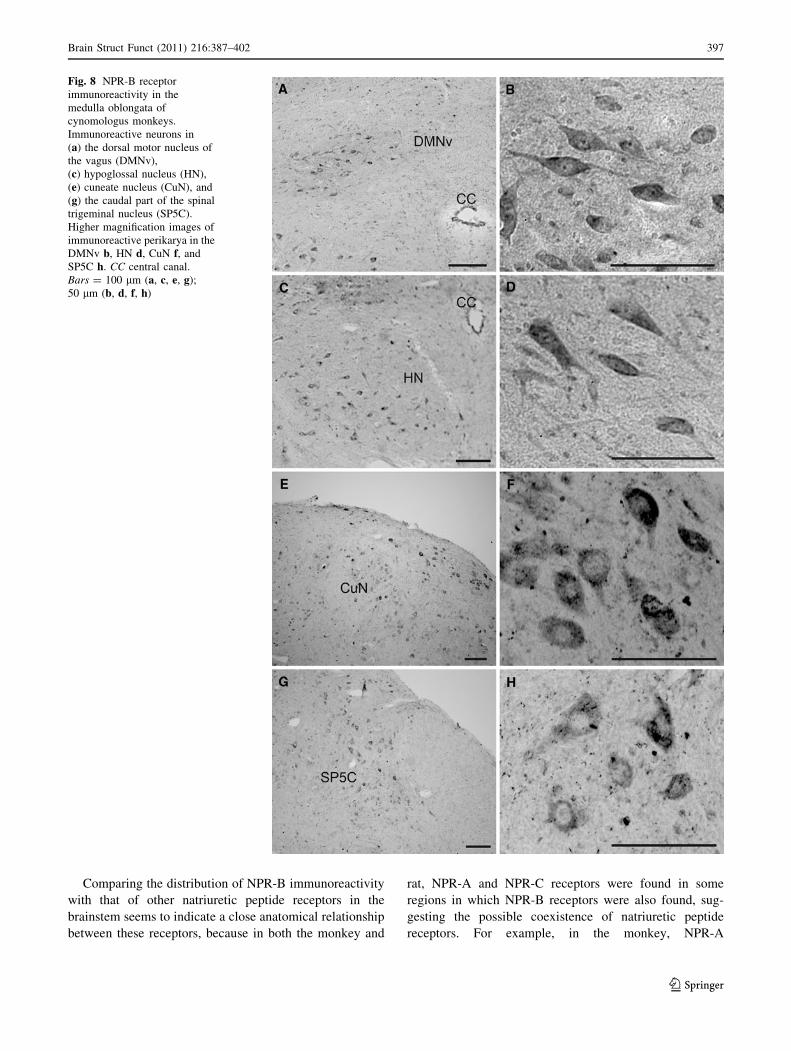
In the dorsal motor nucleus of the vagus, ovoid- to
fusiform-shaped perikarya with immunoreactive processes
Fig. 3 NPR-B receptor
immunoreactivity in the
midbrain of cynomologus
monkeys. a NPR-B-
immunoreactive perikarya and
fibers in the periaqueductal gray
(PAG). aq aqueduct of Sylvius.
b At higher magnification, small
immunoreactive neurons are
apparent in the PAG. c NPR-B-
immunoreactive neurons in the
oculomotor nucleus (NO).
d Immunoreactive neurons in
the NO seen at higher
magnification. e NPR-B
immunoreactivity in the red
nucleus (RN). f Large NPR-B-
immunoreactive perikarya seen
at higher magnification.
Bars = 100 lm (a, d, e);
50 lm (b, c, f)
392 Brain Struct Funct (2011) 216:387–402
123
were positive for NPR-B (Fig. 8a, b). In the hypoglossal
nucleus, NPR-B immunoreactivity was seen in elongated
and fusiform perikarya (Fig. 8c, d). In the dorsal portion of
the caudal medulla, many immunoreactive perikarya were
distributed in the cuneate nucleus (Fig. 8e, f). Considerable
NPR-B immunoreactivity was seen in the spinal trigeminal
nucleus (Fig. 8g, h).
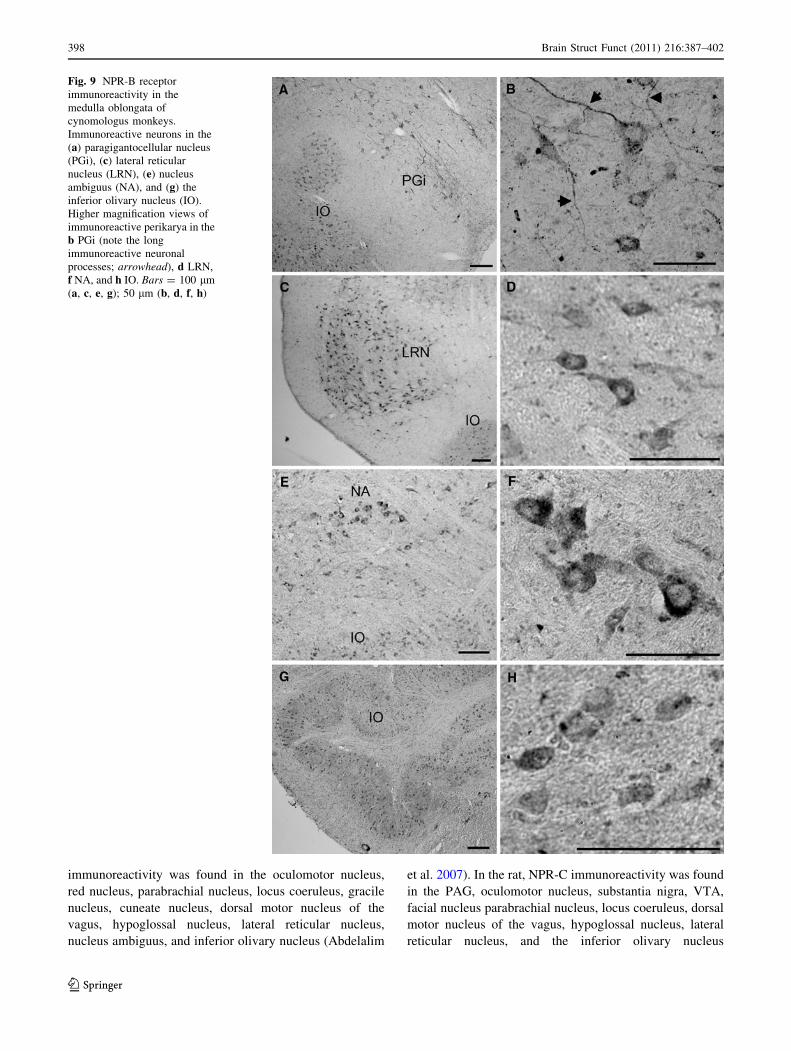
In the paragigantocellular nucleus, different-shaped
neurons, with long immunoreactive processes, were
positive for NPR-B (Fig. 9a, b). Nerve fibers were dis-
tributed among the immunoreactive perikarya (Fig. 9a, b).
A cluster of strongly stained NPR-B neurons was
observed in the lateral reticular nucleus (Fig. 9c, d). In
addition, neurons in the nucleus ambiguus with high-
intensity NPR-B immunoreactivity appeared as a cluster
(Fig. 9e, f). Several immunoreactive neurons were seen
scattered in the reticular formation (Fig. 9e). A large
number of NPR-B-immunoreactive neurons were distrib-
uted uniformly throughout the inferior olivary nucleus
(Fig. 9g, h).
Localization of NPR-B in the dopamine neurons
in the monkey brainstem
To investigate whether dopamine neurons express NPR-B
receptors, we performed double immunostaining for NPR-B
Fig. 4 NPR-B receptor
immunoreactivity in the
midbrain of cynomologus
monkeys. a NPR-B-
immunoreactive neurons in the
ventral tegmental area (VTA).
b Immunoreactive neurons in
the VTA seen at higher
magnification. c NPR-B-
immunoreactive perikarya in the
substantia nigra (SN). d At
higher magnification,
immunoreactive neurons are
apparent in the SN. e NPR-B-
immunoreactive perikarya and
fibers in the cerebral peduncle
(CP). Note the immunoreactive
dendritic arborization
(arrowhead). f Higher
magnification image showing
immunoreactive perikarya and
fibers. Bars = 100 lm (a, d, e);
50 lm (b, c, f)
Brain Struct Funct (2011) 216:387–402 393
123
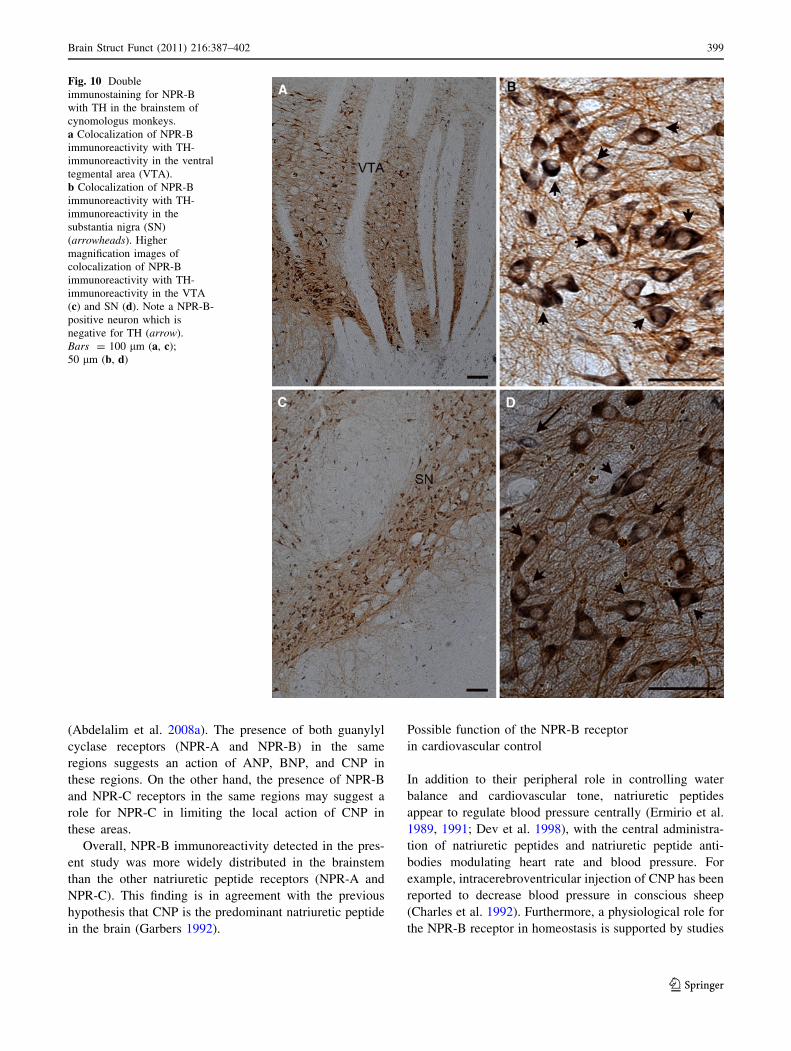
and TH in the monkey brainstem. It has been reported that
in the CNS, nearly 75% of all dopamine neurons reside in
the ventral part of the midbrain (Reiner 1994). Double
immunostaing showed the co-localization of NPR-B
immunoreactivity with the TH-immunoreactive perikarya
in the ventral tegmental area and the substantia nigra and
(Fig. 10a–d). The co-localization was mainly in the
perikarya.
Fig. 5 NPR-B receptor
immunoreactivity in the pons of
cynomologus monkeys.
a Immunoreactive perikarya in
the abducens nucleus (6N).
4V, fourth ventricle.
b Immunoreactive perikarya in
the 6N at higher magnification.
c NPR-B-immunoreactive
neurons in the medial vestibular
nucleus (MVe).
d Immunoreactive neurons in
the MVe seen at higher
magnification. e NPR-B
immunoreactivity in the
parabrachial nucleus (PBN).
f NPR-B-immunoreactive
perikarya in the PBN seen at
higher magnification. g NPR-B
immunoreactivity in the
trigeminal motor nucleus
(MO5). h Higher magnification
image showing NPR-B-
immunoreactive perikarya in the
MO5. Note the
immunoreactivity in neuronal
processes (arrowhead).
Bars = 100 lm (a, c, e, g);
50 lm (b, d, f, h)
394 Brain Struct Funct (2011) 216:387–402
123
Discussion
The present study is the first to map the distribution of
NPR-B protein throughout the brainstem. The results
demonstrate the widespread distribution of NPR-B immu-
noreactivity in this region, which may highlight the
potential importance of natriuretic peptides within the
brainstem.
Fig. 6 NPR-B receptor
immunoreactivity in the pons of
cynomologus monkeys.
a Immunoreactive neurons are
apparent in the locus coeruleus
(LC) at low magnification.
b Higher magnification view of
immunoreactive perikarya in the
LC. c Immunoreactive neurons
in the caudal pontine reticular
formation (Pnc). d At higher
magnification, immunoreactive
perikarya in the Pnc can be
seen. e NPR-B
immunoreactivity in the pontine
nuclei (Pn). f Immunoreactive
perikarya in the Pnc at higher
magnification.
g Immunoreactive neurons in
the raphe magnus nucleus
(RMg) and raphe pallidus
nucleus (RPa). h At higher
magnification, immunoreactive
perikarya in the RMg can be
seen. i Higher magnification
view of immunoreactive
perikarya in the RPa.
Bars = 100 lm (a, c, e, g);
50 lm (b, d, f, h, i)
Brain Struct Funct (2011) 216:387–402 395
123
Distribution of NPR-B immunoreactivity
in the brainstem
The extensive distribution of NPR-B immunoreactivity in
the brainstem regions examined in the present study is in
general agreement with previous results described in the rat
brainstem obtained using in situ hybridization (Herman
et al. 1996). However, some differences are apparent.
Specifically, in the monkey but not rat brainstem, NPR-B
was found in the inferior colliculus, locus coeruleus,
gracile nucleus, and dorsal motor nucleus of the vagus.
Conversely, in the rat but not monkey brainstem, NPR-B
was found in the trochlear nucleus, dorsal raphe nucleus,
Barrington’s nucleus, prepositus, and area postrema (Her-
man et al. 1996). These differences in the distribution of
NPR-B between monkey and rat could be due to either
species or technical considerations differences, because, in
the monkey, we used immunohistochemical analysis to
detect NPR-B.
NPR-B immunoreactivity was observed in the VTA and
substantia nigra and is expressed by the dopamine neurons.
These data are in agreement with the results of a previous
study in rat, which reported abundant CNP and NPR-B
mRNA and protein expression in the VTA and substantia
nigra (Thiriet et al. 2001). Moreover, NPR-B has been
found in the dopaminergic neurons themselves in the VTA
(Jouvert et al. 2004).
The results of the present study demonstrate strong NPR-
B immunoreactivity in the cochlear nucleus. This is in
agreement with previous results reported for the rat, in which
extremely high levels of NPR-B mRNA were detected in the
anteroventral cochlear nucleus (Herman et al. 1996).
Fig. 7 NPR-B receptor
immunoreactivity in the pons of
cynomologus monkeys.
a Immunoreactive neurons in
a the facial nucleus (7N), (c) the
oral part of the spinal trigeminal
nucleus (SP5O), and (e) the
cochlear nucleus (CN). Higher
magnification view of
immunoreactive perikarya in the
7N (b), SP5O (d), and CN (f).Bars = 100 lm (a, c, e, g);
50 lm (b, d, f, h, i)
396 Brain Struct Funct (2011) 216:387–402
123
Comparing the distribution of NPR-B immunoreactivity
with that of other natriuretic peptide receptors in the
brainstem seems to indicate a close anatomical relationship
between these receptors, because in both the monkey and
rat, NPR-A and NPR-C receptors were found in some
regions in which NPR-B receptors were also found, sug-
gesting the possible coexistence of natriuretic peptide
receptors. For example, in the monkey, NPR-A
Fig. 8 NPR-B receptor
immunoreactivity in the
medulla oblongata of
cynomologus monkeys.
Immunoreactive neurons in
(a) the dorsal motor nucleus of
the vagus (DMNv),
(c) hypoglossal nucleus (HN),
(e) cuneate nucleus (CuN), and
(g) the caudal part of the spinal
trigeminal nucleus (SP5C).
Higher magnification images of
immunoreactive perikarya in the
DMNv b, HN d, CuN f, and
SP5C h. CC central canal.
Bars = 100 lm (a, c, e, g);
50 lm (b, d, f, h)
Brain Struct Funct (2011) 216:387–402 397
123
immunoreactivity was found in the oculomotor nucleus,
red nucleus, parabrachial nucleus, locus coeruleus, gracile
nucleus, cuneate nucleus, dorsal motor nucleus of the
vagus, hypoglossal nucleus, lateral reticular nucleus,
nucleus ambiguus, and inferior olivary nucleus (Abdelalim
et al. 2007). In the rat, NPR-C immunoreactivity was found
in the PAG, oculomotor nucleus, substantia nigra, VTA,
facial nucleus parabrachial nucleus, locus coeruleus, dorsal
motor nucleus of the vagus, hypoglossal nucleus, lateral
reticular nucleus, and the inferior olivary nucleus
Fig. 9 NPR-B receptor
immunoreactivity in the
medulla oblongata of
cynomologus monkeys.
Immunoreactive neurons in the
(a) paragigantocellular nucleus
(PGi), (c) lateral reticular
nucleus (LRN), (e) nucleus
ambiguus (NA), and (g) the
inferior olivary nucleus (IO).
Higher magnification views of
immunoreactive perikarya in the
b PGi (note the long
immunoreactive neuronal
processes; arrowhead), d LRN,
f NA, and h IO. Bars = 100 lm
(a, c, e, g); 50 lm (b, d, f, h)
398 Brain Struct Funct (2011) 216:387–402
123
(Abdelalim et al. 2008a). The presence of both guanylyl
cyclase receptors (NPR-A and NPR-B) in the same
regions suggests an action of ANP, BNP, and CNP in
these regions. On the other hand, the presence of NPR-B
and NPR-C receptors in the same regions may suggest a
role for NPR-C in limiting the local action of CNP in
these areas.
Overall, NPR-B immunoreactivity detected in the pres-
ent study was more widely distributed in the brainstem
than the other natriuretic peptide receptors (NPR-A and
NPR-C). This finding is in agreement with the previous
hypothesis that CNP is the predominant natriuretic peptide
in the brain (Garbers 1992).
Possible function of the NPR-B receptor
in cardiovascular control
In addition to their peripheral role in controlling water
balance and cardiovascular tone, natriuretic peptides
appear to regulate blood pressure centrally (Ermirio et al.
1989, 1991; Dev et al. 1998), with the central administra-
tion of natriuretic peptides and natriuretic peptide anti-
bodies modulating heart rate and blood pressure. For
example, intracerebroventricular injection of CNP has been
reported to decrease blood pressure in conscious sheep
(Charles et al. 1992). Furthermore, a physiological role for
the NPR-B receptor in homeostasis is supported by studies
Fig. 10 Double
immunostaining for NPR-B
with TH in the brainstem of
cynomologus monkeys.
a Colocalization of NPR-B
immunoreactivity with TH-
immunoreactivity in the ventral
tegmental area (VTA).
b Colocalization of NPR-B
immunoreactivity with TH-
immunoreactivity in the
substantia nigra (SN)
(arrowheads). Higher
magnification images of
colocalization of NPR-B
immunoreactivity with TH-
immunoreactivity in the VTA
(c) and SN (d). Note a NPR-B-
positive neuron which is
negative for TH (arrow).
Bars = 100 lm (a, c);
50 lm (b, d)
Brain Struct Funct (2011) 216:387–402 399
123

demonstrating that CNP expression in the olfactory region
is modulated by water and salt balance (Cameron et al.
2001). These effects may be mediated through receptors
located in the brainstem because the PAG (Carrive et al.
1989; Inui et al. 1994), parabrachial nucleus (Chamberlin
and Saper 1992), locus coeruleus (Anselmo-Francil et al.
1999), dorsal motor nucleus of the vagus, nucleus tractus
solitarius (Paton 1999), paragigantocellular nucleus
(Brown and Guyenet 1984; Schreihofer and Guyenet
1997), and nucleus ambiguus (Machado and Brody 1988)
are thought to be involved in the central control of car-
diovascular homeostasis. Consistent with this, we found
abundant NPR-B immunoreactivity in these regions in the
monkey brainstem, which is highly indicative of an
important role for CNP in central cardiovascular
regulation.
Other possible functions of the NPR-B receptor
in the brainstem
CNP has been postulated to have diverse functional roles in
different regions of the central nervous system (Thiriet
et al. 2001; Jouvert et al. 2004; Sabbatini et al. 2005),
including many of the brainstem structures examined in the
present study. The presence of the NPR-B receptor in many
regions of the monkey brainstem implies that CNP may
have different functions.
The presence of NPR-B immunoreactivity in the supe-
rior colliculus, oculomotor nucleus, vestibular nuclei,
abducens nucleus, facial nucleus, and hypoglossal nucleus
indicates that CNP could be involved in the control of eye
and head movements. The neural organization of the
pathways from the superior colliculus to motoneurons in
the horizontal oculomotor system has been analyzed
extensively (Sparks 1999; Scudder et al. 2002). Further-
more, the natriuretic peptides and their receptors have been
detected in eye structures (Fernandez-Durango et al. 1995;
Kuribayashi et al. 2006; Abdelalim et al. 2008b).
Centrally applied CNP enhances pancreatic secretion
through a vagal pathway by activating central natriuretic
peptide guanylyl cyclase-coupled receptors (Sabbatini et al.
2005). mRNA expression for the natriuretic peptide
receptor and CNP has been localized in the paraventricular
nucleus and the dorsal motor nucleus of the vagus, both of
which are important sites for the regulation of gastroin-
testinal function (Langub et al. 1995; Herman et al. 1996).
Together with the results of the present study, these find-
ings suggest a role for NPR-B receptors located in the
dorsal motor nucleus of the vagus of the monkey brainstem
in the central control of pancreatic function.
In the present study, NPR-B immunoreactivity was
detected in the dopamine neurons in the substantia nigra
and VTA, which suggests the involvement of NPR-B in
dopamine neurons functions. Recently, we found that the
NPR-B receptor is expressed by dopaminergic cells in the
retina (Abdelalim et al. 2008b). It has been reported pre-
viously that CNP inhibits dopamine release by stimulating
NPR-B receptors and increasing intracellular GMP con-
centrations (Jouvert et al. 2004). In addition, CNP has been
shown to regulate cocaine-induced dopamine release and
the expression of immediate early genes in brain neurons
(Thiriet et al. 2001). CNP has been found to improve
learning and consolidation of learning in a passive avoid-
ance paradigm (Telegdy et al. 1999). Interestingly, dopa-
mine has been shown to be one of the mediating
neurotransmitters in the effect of CNP on learning effects
(Telegdy et al. 1999).
In addition, in the present study NPR-B immunoreac-
tivity was observed in brain regions related to general
somatic afferents, including the principal sensory trigemi-
nal nucleus, gracile nucleus, and cuneate nucleus. Although
previous studies have reported the involvement of natri-
uretic peptides in the baroreflex (Thomas et al. 1997), little
information is available regarding the role of CNP/NPR-B
system in general somatic afferents. Further studies are
required to identify the potential physiological relevance of
the NPR-B receptor.
Also, the presence of the NPR-B receptor in the locus
coeruleus suggests that CNP could be involved in the
control of neurons in the dorsal horn in the monkey,
because neurons in the locus coeruleus project to the spinal
cord.
The widespread distribution of NPR-B immunoreactiv-
ity observed in the current study may help establish the
possible functions of the NPR-B receptor in brainstem
nuclei. Further studies are needed to establish the physio-
logical functions of the NPR-B receptor in the brainstem
because the role of CNP as a neurotransmitter and/or
neuromodulator in these nuclei remains to be elucidated.
Conclusion
The present study provides the first detailed map of NPR-B
immunoreactivity in the monkey brainstem. Combined
with previous functional studies, our data support a role for
natriuretic peptides in the central control of cardiovascular
homeostasis. Furthermore, we demonstrated that the NPR-B
receptor is widely distributed throughout brainstem regions
related to other functions, suggesting the involvement of
CNP and the NPR-B receptor in mediating a wide range of
brainstem functions.
Acknowledgments This work was supported by a Grant-in-Aid for
Scientific Research (no. 21-09133) from the Japan Society for the
Promotion of Science.
400 Brain Struct Funct (2011) 216:387–402
123

References
Abdelalim EM, Takada T, Torii R, Tooyama I (2006) Molecular
cloning of BNP from heart and its immunohistochemical
localization in the hypothalamus of monkey. Peptides
27:1886–1893
Abdelalim EM, Osman AHK, Takada T, Torii R, Tooyama I (2007)
Immunohistochemical mapping of NPR-A in the brainstem of
Macaca fascicularis. Neuroscience 145:1087–1096
Abdelalim EM, Masuda C, Bellier JP, Saito A, Yamamoto S, Mori N,
Tooyama I (2008a) Distribution of natriuretic peptide receptor-C
immunoreactivity in the rat brainstem and its relationship to
cholinergic and catecholaminergic neurons. Neuroscience
155:192–202
Abdelalim EM, Masuda C, Tooyama I (2008b) Expression of
natriuretic peptide-activated guanylate cyclases by cholinergic
and dopaminergic amacrine cells of the rat retina. Peptides
29:622–628
Anand-Srivastava MB, Trachte GT (1993) Atrial natriuretic factor
receptors and signal transduction mechanisms. Pharmacol Rev
45:455–497
Anselmo-Francil JA, Rocha MJA, Peres-Polon VL, Moreira ER,
Antunes-Rodrigues J, Franci CR (1999) Role of the locus
coeruleus on blood pressure response and atrial natriuretic
peptide secretion following extracellular volume expansion.
Brain Res Bull 50:173–177
Brown DL, Guyenet PG (1984) Cardiovascular neurons of brainstem
with projections to spinal cord. Am J Physiol 247:R1009–R1016
Burry RW (2000) Specificity controls for immunocytochemical
methods. J Histochem Cytochem 48:163–165
Cameron VA, Cumming SA, Espiner EA, Nicholls G, Richards M
(2001) C-type natriuretic peptide expression in olfactory regions
of rat brain is modulated by acute water deprivation, salt loading
and central angiotensin II. Neuroendocrinology 73:46–53
Carrive P, Bandler R, Dampney RA (1989) Viscerotopic control of
regional vascular beds by discrete groups of neurons within the
midbrain periaqueductal gray. Brain Res 493:385–390
Chamberlin NL, Saper CB (1992) Topographic organization of
cardiovascular responses to electrical and glutamate microsti-
mulation of the parabrachial nucleus of the rat. J Comp Neurol
326:245–262
Charles CJ, Richards AM, Espiner EA (1992) Central C-type
natriuretic peptide but not atrial natriuretic factor lowers blood
pressure and adrenocortical secretion in normal conscious sheep.
Endocrinology 131:1721–1726
Decker JM, Wojtowicz AM, Liotta BA, Braunewell KH, Heinemann
U, Behrens CJ (2010) C-type natriuretic peptide modulates
bidirectional plasticity in hippocampal area in CA1 in vitro.
Neuroscience 169:8–22
Deschepper CF, Picard S (1994) Effects of C-type natriuretic peptide
on rat astrocytes: regional differences and characterization of
receptors. J Neurochem 62:1974–1982
Dev BR, Nandakumaran M, Philip L, John SJ (1998) Brain natriuretic
peptide-mediated changes in the extracellular neurotransmitter
turnover in the rostral ventrolateral medulla. Neuroscience
84:255–262
Ermirio R, Ruggeri P, Cogo CE, Molinari C, Calaresu FR (1989)
Neuronal and cardiovascular responses to ANF microinjected
into the solitary nucleus. Am J Physiol 256:R577–R582
Ermirio R, Ruggeri P, Cogo CE, Molinari C, Calaresu FR (1991)
Neuronal and cardiovascular responses to ANF microinjected
into nucleus ambiguus. Am J Physiol 260:R1089–R1094
Espiner EA, Richards AM, Yandle TG, Nicholls MG (1995)
Natriuretic hormones. Endocrinol Metab Clin North Am
24:481–509
Fernandez-Durango R, Nunez DJ, Brown MJ (1995) Messenger
RNAs encoding the natriuretic peptides and their receptors are
expressed in the eye. Exp Eye Res 61:723–729
Fiscus RR, Tu AW, Chew SB (2001) Natriuretic peptides inhibit
apoptosis and prolong the survival of serum-deprived PC12
cells. Neuroreport 12:185–189
Garbers DL (1992) Guanylyl cyclase receptors and their endocrine,
paracrine and autocrine ligands. Cell 71:1–4
Garbers DL, Chrisman TD, Wiegn P, Katafuchi T, Albanesi JP,
Bielinski V, Barylko B, Redfield MM, Burnett JC Jr (2006)
Membrane guanylyl cyclase receptors: an update. Trends
Endocrinol Metab 17:251–258
Gutkowska J, Antunes-Rodrigues J, McCann SM (1997) Atrial
natriuretic peptide in brain and pituitary gland. Physiol Rev
77:465–515
Herman JP, Doglas CM, Rucker D, Langub MC Jr (1996) Locali-
zation of natriuretic peptide-activated guanylate cyclase mRNA
in the rat brain. J Comp Neurol 369:165–187
Imura H, Nakao N, Itoh H (1992) The natriuretic peptide system in the
brain: implications in the central control of cardiovascular and
neuroendocrine functions. Front Neuroendocrinol 13:217–249
Inui K, Murase S, Nosaka S (1994) Facilitation of the arterial
baroreflex by the ventrolateral part of the midbrain periaqu-
eductal gray matter in rats. J Physiol 477(Pt 1):89–101
Jouvert P, Revel MO, Lazaris A, Aunis D, Langley K, Zwiller J
(2004) Activation of the cGMP pathway in dopaminergic
structures reduces cocaine-induced EGR-1 expression and loco-
motor activity. J Neurosci 24:10716–10725
Koller KJ, Lowe DG, Bennett GL, Minamino N, Kangawa K, Matsuo
H, Goeddel DV (1991) Selective activation of the B natriuretic
peptide receptor by C-type natriuretic peptide (CNP). Science
252:120–123
Kuhun M (2003) Structure, regulation, and function of mammalian
membrane guanylyl cyclase receptors, with a focus on guanylyl
cyclase-A. Circ Res 93:700–709
Kuribayashi K, Kitaoka Y, Kumai T, Munemasa Y, Kitaoka Y,
Isenoumi K, Motoki M, Kogo J, Hayashi Y, Kobayashi D, Ueno
S (2006) Neuroprotective effect of atrial natriuretic peptide
against NMDA-induced neurotoxicity in the rat retina. Brain Res
1071:34–41
Langub MC Jr, Warson RE Jr, Herman JP (1995) Distribution of
natriuretic peptide precursor mRNA in the rat brain. J Comp
Neurol 356:183–199
Levin ER, Gardner DG, Samson WK (1998) Natriuretic peptides.
N Engl J Med 339:321–328
Maack T (1992) Receptors of atrial natriuretic factors. Annu Rev
Physiol 54:11–27
Machado BH, Brody MJ (1988) Role of the nucleus ambiguus in the
regulation of heart rate and arterial pressure. Hypertension
11:602–607
Matsukawa N, Grzesik WJ, Takahashi N, Pandey KN, Pang S,
Yamauchi M, Smithies O (1999) The natriuretic peptide clearance
receptor locally modulates the physiological effects of the
natriuretic peptide system. Proc Nalt Acad Sci USA 96:7403–7408
Nakao K, Ogawa Y, Suga SI, Imura H (1992) Molecular biology and
biochemistry of the natriuretic peptide system: II. Natriureticpeptide receptors. J Hypertens 10:1111–1114
Paton JFR (1999) Nucleus tractus solitarii: integrating structures. Exp
Physiol 84:815–833
Potter LR, Abbey-Hosch S, Dickey DM (2006) Natriuretic peptides,
their receptors, and cyclic guanosine monophosphate-dependent
signaling functions. Endocr Rev 27:47–72
Reiner A (1994) Catecholaminergic innervation of the basal ganglia
in mammals: anatomy and function In: Smeets WJAJ, Reiner A
(eds) Phylogeny and development of catecholamine systems in
Brain Struct Funct (2011) 216:387–402 401
123

the CNS of vertebrates. Cambridge University Press, Cambridge,
pp 247–272
Sabbatini ME, Rodriguez MR, Corbo NS, Vatta MS, Bianciotti LG
(2005) C-type natriuretic peptide applied to the brain enhances
exocrine pancreatic secretion through a vagal pathway. Eur J
Pharmacol 524:67–74
Schreihofer AM, Guyenet PG (1997) Identification of C1 presympa-
thetic neurons in rat rostral ventrolateral medulla by juxtacellular
labeling in vivo. J Comp Neurol 387:524–536
Scudder CA, Kaneko CRS, Fuchs AF (2002) The brainstem burst
generator for saccadic eye movements. Exp Brain Res
142:439–462
Sparks DL (1999) Conceptual issues related to the role of the superior
colliculus in the control of gaze. Curr Opin Neurobiol 9:698–707
Szabo J, Cowan WM (1984) A stereotaxic atlas of the brain of the
cynomologus monkey (Macaca fascicularis). J Comp Neurol
222:265–300
Telegdy G, Kokavszky K, Nyerges A (1999) Action of C-type
natriuretic peptide (CNP) on passive avoidance learning in rats:
involvement of transmitters. Eur J Neurosci 11:3302–3306
Thiriet N, Jouvert P, Gobaille S, Solov’eva O, Gough B, Aunis D, Ali
S, Zwiller J (2001) C-type natriuretic peptide (CNP) regulates
cocaine-induced dopamine increase and immediate early gene
expression in rat brain. Eur J Neurosci 14:1702–1708
Thomas CJ, Rankin AJ, Head GA, Woods RL (1997) ANP enhances
bradycardic reflexes in normotensive but not spontaneously
hypertensive rats. Hypertension 29:1126–1132
Wilcox JN, Augustine A, Goeddel DV, Lowe DG (1991) Differential
regional expression of three natriuretic peptide receptor genes
within primate tissue. Mol Cell Biol 11:3454–3462
Yasoda A, Komatsu Y, Chusho H, Miyazawa T, Ozasa A, Miura M,
Kurihara T, Rogi T, Tanaka S, Suda M, Tamura N, Ogawa Y,
Nakao K (2004) Overexpression of CNP in chondrocytes rescues
achondroplasia through a MAPK-dependent pathway. Nat Med
10:80–86
402 Brain Struct Funct (2011) 216:387–402
123





























