Embed Size (px)
Citation preview

Measurement of Strain-DependentToxicity in the Indene BioconversionUsing Multiparameter Flow Cytometry
A. Amanullah,1* C. J. Hewitt,2 A. W. Nienow,2 C. Lee,3 M. Chartrain,3
B. C. Buckland,3 S. W. Drew,3 J. M. Woodley1
1Advanced Centre for Biochemical Engineering, Department ofBiochemical Engineering, University College London, Torrington Place,London W1CE 7JE, United Kingdom2Centre for Bioprocess Engineering, School of Chemical Engineering, TheUniversity of Birmingham, Edgbaston, Birmingham B15 2TT,United Kingdom3Department of Bioprocess Research and Development, Merck ResearchLaboratories, Merck & Co. Inc., Rahway, New Jersey 07065; e-mail:[email protected]
Received 29 August 2001; accepted 2 July 2002
DOI: 10.1002/bit.10479
Abstract: The bionconversion of indene to cis-(1S,2R)-indandiol, a potential key intermediate in the synthesis ofMerck’s HIV protease inhibitor, CRIXIVAN�, can beachieved using Rhodococcus, Pseudomonas putida, andEscherichia coli strains. This study reports on the appli-cation of multiparameter flow cytometry for the mea-surement of cytoplasmic membrane integrity and mem-brane depolarization as indicators of toxic effects of thesubstrate, product, and by-products using each of thesestrains. Measurements of oxygen uptake rate (OUR) andoptical density (OD) as indicators of metabolic activityand biomass growth, respectively, were also made. Mea-surements of the cytoplasmic membrane potential, cellviability, and respiratory activity provided a sensitive setof parameters to assess toxicity in the indene bioconver-sion and provided the basis for process improvementsand strain selection. The toxic concentrations of the sub-strate, product, and by-products for each strain havebeen determined. The results show that it is possible toaccumulate cis-(1S,2R)-indandiol and cis-1-amino-2-indanol up to 20 g/L without significant negative effectson cell physiology using any of the strains tested. TheGram-negative P. putida (421-5 and GM 730) and E. colistrains were more resistant to indene and the isolatedchemicals of the biotransformation than the Gram-positive Rhodoccoccus I24 strain, possibly due to thepresence of the outer membrane and efflux pumpmechanisms. P. putida GM 730 and the E. coli TDO 123strains responded similarly to toxic effects, and the E.coli TDO 123 strain was more resistant than the P. putida421-5 strain. In addition to the recommendations forstrain selection, the identified targets for bioprocess im-provement include a combination of genetic as well as
process engineering approaches. © 2003 Wiley Periodicals,Inc. Biotechnol Bioeng 81: 405–420, 2003.Keywords: flow cytometry; strain selection; substrate,product, and by-product toxicity; indene bioconversion;HIV protease inhibitor
INTRODUCTION
The synthesis of chiral optically pure compounds by asym-metric synthesis or via precise resolution of enantiomersoften provides an attractive manufacturing route of opticallypure drugs that exhibit highly specific pharmacologicalactivity (Buckland et al., 1999). One such exampleis the manufacture of Merck’s HIV protease inhibitor,CRIXIVAN�. The production of the key intermediate forthis drug, cis-(1S,2R)-1-aminoindan-2-ol [(−)-CAI] in enan-tiomerically pure form, is highly technically demanding,requiring a complicated chemical synthesis to the racematefollowed by a crystallization/resolution step (Buckland etal., 1999). Alternatively, (−)-CAI can be directly synthe-sized using either cis-(1S,2R)- or trans-(1R,2R)-indandiolmade using a bioprocess based on the bioconversion ofindene to cis-(1S,2R)-indandiol and trans-(1R,2R)-indandiol using Rhodococcus I24 (Chartrain et al., 1998;Buckland et al., 1999), Pseudomonas putida 421-5 (Con-nors et al., 1997), P. putida GM 730, and Escherichia coliTDO 123 strains (Reddy et al., 1999). The characteristics ofthe strains are described later.
Figure 1 shows the currently understood indene biocon-version pathway for Rhodococcus I24 (Stafford et al.,2001). This strain exhibits both monooxygenase as well asdioxygenase activity. The biotransformation of indene tocis-(1S,2R)-indandiol is characterized by low yields withthe production of various undesirable monooxygenation by-products such as 1-indenol and 1-indanone. The dioxygen-ated products are cis-(1S,2R)-indandiol and trans-(1R,2R)-
Correspondence to: Dr. Ashraf Amanullah*Present address: Fermentation and Cell Culture, Bioprocess R&D,
Merck Research Laboratories, Merck & Co. Inc., WP26C1-101, P.O. Box4, Westpoint, Pennsylvania; telephone: 215-652-0423; fax: 215-652-8691;e-mail: [email protected]
Contract grant sponsor: Merck & Co. Inc.
© 2003 Wiley Periodicals, Inc.

indandiol. Stafford et al. (2001) showed that up to 97.5% ofindene is converted to indan oxide via a monooxygenaseand that 47% and 37% of indan oxide is hydrolyzed totrans-(1R,2R)-indandiol and cis-(1S,2R)-indandiol, respec-tively (13.5% accumulated in the medium). Only 2.5% ofindene is directly converted to cis-(1R,2S)-indandiol. Ad-ditionally, cis-(1S,2R)-indandiol was further oxidized to1-keto-2-hydroxy indan, while trans-(1R,2R)-indandiol wasnot degraded. Furthermore, product concentrations and pro-ductivity are also low, and the reasons for this are not fullyunderstood, although it is believed that the substrate, prod-uct, and by-products inhibit their own synthesis and mayexhibit toxic effects on the culture (Connors et al., 1997;Chartrain et al., 1998) above critical concentrations. Evenafter considerable process development, the maximum con-centration of indandiol reported was typically 4 g/L (usinga Rhodococcus B264-1 strain) and the yield (g/g basis) ofindandiol on indene was 0.22 (Chartrain et al., 1998; Buck-land et al., 1999). These performance indices were lowerusing Pseudomonas (Connors et al. 1997) and recombinantE. coli strains (Buckland et al. 1999). This low titre andproductivity render the current bioprocess economically un-viable.
In terms of process improvement, one objective may be toconstruct metabolic pathways which allow the conversionof indene to either cis-(1S,2R)-indandiol at high yields and
prevent its further degradation to 1-keto-2-hydroxy indan orto engineer pathways directly from indene to (−)-CAI.Firstly, our interest was to obtain a better understanding ofthe toxic effects of the substrate, product, and by-productson host organism and metabolic activity. This is essential inaddition to knowledge of the effects of the chemicals onenzyme activities if a suitable metabolic pathway is to bedesigned. This information can also be used to decidewhether implementation of in situ product removal tech-niques (Lye and Woodley, 1999) will improve process per-formance. Secondly, our interest was to provide a basis forhost selection given that the bioconversion of indene toindandiol can be conducted using a number of differentstrains. Finally, we aimed to quantify the limits of cis-(1S,2R)-indandiol and cis-(1S,2R)-1-aminoindan-2-ol accu-mulation in order to see whether it was possible for the cellsto tolerate enhanced levels of the desired product after theimprovements in the bioprocess.
Previously we have reported the use of multiparameterflow cytometry to assess the physiological state of a Rho-dococcus I24 strain in response to indene toxicity (Aman-ullah et al., 2002). In the current study we report on theapplication of flow cytometry for the measurement of thetoxic effects of indene and the metabolites of the biocon-version using one Gram-positive and three Gram-negativebacteria, all capable of converting indene to cis-indandiol.The overall goal was to construct a metabolic pathway thatallows the conversion of indene to indandiol at high yieldsand with high enantiomeric specificity. An appropriate hostthat can grow to high cell densities and can function in anaqueous environment with high levels of indandiol (andby-products) or cis-(1S,2R)-1-aminoindan-2-ol is requiredfor this purpose.
Strain Characteristics
The characteristics of all four strains capable of convertingindene to indandiol are shown in Table I. Rhodococcus I24was a wild-type, Gram-positive, rod-shaped organism andwas isolated from toluene-contaminated soil in a screeningprogram by Merck scientists (Chartrain et al., 1998). Thisstrain was thought to be more resistant to indene comparedto the P. putida strains and did not require toluene for in-duction of dioxygenase activity (Buckland et al., 1999).Rhodococcus I24 converted indene to cis-(1S-2R)-indandioland trans-(1S,2R)-indandiol at concentrations of 0.4 and 1.4g/L, respectively, in approximately 200 h (Chartrain et al.,
Figure 1. Metabolic pathways of the indene bioconversion (Stafford etal., 2001).
Table I. Strain characteristics.
Straincis-(1S, 2R)-Indandiol
(g/L)trans-(1S, 2R)-Indandiol
(g/L)Enantiomeric
excess Yield (g/g)Space–time
yield (mg/L/h)
Rhodococcus I24 0.40 1.40 >98% 0.20 20Pseudomonas 421-5 1.80 — >98% 0.20 30Pseudomonas GM 730 1.80 — >98% 0.20 30Escherichia coli TDO 123 1.0 — ∼33% 0.20 90
406 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 81, NO. 4, FEBRUARY 20, 2003

1998). Both P. putida 421-5 (Connors et al., 1997) and P.putida GM 730 were wild-type, Gram-negative, rod-shapedorganisms, which converted indene to cis-indandiol only. P.putida 421-5 was derived from P. putida F1 in a screeningprogram and did not require toluene as an inducer of thedioxygenase activity and produced 20-fold higher concen-trations of indandiol compared to the parent strain (Connorset al., 1997). The use of strains of P. putida as hosts offersthe advantages of rapid growth, well-studied aromatic me-tabolism, and well-developed systems for genetic manipu-lations. Its disadvantage is that the intrinsic stereoselectivityof toluene dioxygenase is limited to an enantiomeric excess(ee) of 30%, with significant levels of undesirable by-product formation (Connors et al., 1997). However, afterselective degradation of one of the isomers, the ee increasesto 98%. There are no published data available with respectto indene bioconversion using the P. putida GM 730 strain,although unpublished results (personal communication,Merck & Co.) suggest that its performance is similar to theP. putida 421-5 strain. E. coli TDO 123 was a GMO withthe cloned toluene dioxygenase (TDO) genes from P. putidaand was capable of converting indene to cis-indandiol up to1 g/L (Buckland et al., 1999). E. coli offers rapid growthand well-studied metabolic and genetic systems. The yieldof cis-indandiol from indene was 0.20 with each straintested, and the enantiomeric excess of the desired (1S,2R)enantiomer was >98% in all cases except with the E. coliTDO 123 strain, where it was ∼33% (Buckland et al., 1999).
Cytoplasmic Membrane AssociatedSolvent Toxicity
The cytoplasmic membrane in both types of bacteria con-sists of a phospholipid bilayer that serves as a selectivebarrier for protons and ions, regulation of the intracellularenvironment, maintenance of turgor pressure, and as a ma-trix for embedded proteins that are required for energytransduction. In most solvent toxicity studies using bacteriaand yeast (Ingram, 1976; Uribe et al., 1985; Harrop et al.,1989; Osborne et al., 1990; Inoue and Horikoshi, 1991;McDonald and Goldfine, 1991; Sikemma et al., 1992; Ver-mue et al., 1993; Weber and de Bont, 1994; Isken and deBont, 1996; Kieboom et al., 1998), the cytoplasmic mem-brane is mentioned as a possible target, but there is verylittle known about the mode of action of such solvents.Sikemma and co-workers (1992) demonstrated that the in-teraction of tetralin with the membrane of both Gram-positive and Gram-negative bacteria as well as on modelliposome systems resulted in increased permeability of themembrane and loss of the cytoplasmic proton motive force.The importance of the cell membrane in solvent toxicity isalso demonstrated by the differences in the toxicity behaviorof Gram-positive and Gram-negative bacteria, with the lat-ter generally found to be slightly more resistant (Harrop etal., 1989; Inoue and Horikoshi, 1991). Osborne and co-workers (1990) investigated the effects of toxicity using abroad range of solvents and found a correlation between the
11�-hydroxylase activity of Rhizopus nigricans and the sol-vent concentration in the cell membrane.
Flow Cytometry
Flow cytometry is a powerful technique for characterizingcell populations using scattered light. With the introductionof fluorescent dyes that bind to specific intracellular com-ponents it is possible to obtain valuable information on cellphysiology and cellular metabolism. The ability of flowcytometers to make multiparameter measurements sets themapart from most other cell analytical techniques. Anothermajor advantage of this technique is that it is possible toconduct quantitative analysis on a large heterogenous cellpopulation rapidly and on a single-cell basis. It is routinelyused with animal cells and in the medical field, but use withbacterial cells is less well substantiated due to their muchsmaller size, hence the requirement for greater sensitivity ofthe measurement technique (Davey and Kell, 1996). How-ever, recently, significant progress has been made (Nebe-von Caron and Badley, 1995; Shapiro, 1995; Davey andKell, 1996; Hewitt et al., 1998, 1999a,b) and is the basis forthe methodology described here. The application of flowcytometry to quantify intact cell and membrane potentialand to characterize efflux pump systems will be brieflydescribed.
Fluorescent Probes
Intact (Viable) Cells
The measurement of membrane integrity is either based ondye exclusion or dye retention (Shapiro, 1995; Davey andKell, 1996). Propidium iodide (PI; Sigma, Poole, Dorset,U.K.) is a vital fluorescent probe that binds to DNA butcannot cross an intact cytoplasmic membrane (membrane)(Nebe-Von Caron and Badley, 1995; Davey and Kell, 1996;Hewitt et al., 1999a,b). Intact cells show selective perme-ability of the membrane. However, once the membrane in-tegrity is lost and both metabolic activity and cell divisionare absent, detected events are classified as dead cells.Therefore, PI-negative cell populations represent those cellswith an intact membrane and are generally regarded as vi-able cells, whereas PI-positive cells represent dead cells.
Cytoplasmic Membrane Potential
Passive and active transport systems across the membraneof healthy bacterial cells generate an electrochemical gra-dient (polarized membranes), giving a measure of cellularmetabolic activity (Shapiro, 1995). Dissipation of the mem-brane potential (depolarized membranes) leads to the situ-ation where the cell is less able to protect its cytoplasm fromthe surrounding medium and can severely impair normalfunctioning of the cell. Bis-Oxonol (BOX) (MolecularProbes, Eugene, OR) is a lipophilic, non-toxic anionic fluo-
AMANULLAH ET AL.: MEASUREMENT OF STRAIN DEPENDENT TOXICITY USING MULTIPARAMETER FLOW CYTOMETRY 407

rescent stain that binds to the membrane provided it is de-polarized. One advantage of using BOX is that it is insen-sitive to the presence of efflux pump systems in the mem-brane (Davey and Kell, 1996). Thus dual staining with PIand BOX can give quantitative information on cell popula-tions with intact membranes, intact polarized membranes,intact depolarized membranes, and dead cells. Rhodamine123 (Rh 123) (Molecular Probes) has also been used tomeasure membrane potential. It is a lipophilic, cationicprobe that binds to the membrane provided that it is polar-ized, and the measurements of PI/Rh 123 in principle shouldbe similar to that with PI/BOX.
MATERIALS AND METHODS
Strains
Rhodococcus I24 (MA7205), P. putida 421-5, P. putida GM730, and E. coli TDO 123 strains were kindly donated byMerck & Co. (Rahway, NJ). Lyophilized cultures were re-generated in 25 mL of 30 g/L Tryptic Soy Broth (TSB) for20 h at 200 rpm and 30°C (except for E. coli which wasregenerated at 37°C). A 50% solution of glycerol/water wasadded in a 1:1 v/v ratio, and 1-mL aliquots were dispensedin cryovials and stored at −70°C.
Shake Flasks
Three 250-mL shake flask cultures were each inoculatedwith the contents of a thawed vial and incubated at 200 rpmand 30°C (except for E. coli which was regenerated at 37°C)for 24 h. The medium used was 30 g/L TSB and 1 mL/Lpolypropylene glycol antifoam. Two flasks were used toinoculate a 14-L bioreactor (10-L working volume).
Fermentations
Fed-batch cultures were conducted using a 14-L fermenter(Infors, Bottmingen, Switzerland), and operational detailscan be found in Amanullah et al., 2002).
Medium
The composition of the fermentation medium (in g/L) forthe Rhodococcus and Pseudomonas strains (medium A),which was successfully developed to support high biomassconcentrations (>30 g dry cell weight/L), was as follows:glucose, 45.0; citric acid, 1.0; MgSO4 � 7H2O, 2.0;KH2PO4, 2.0; (NH4)2SO4, 6.0; Fe2SO4, 0.1; CaCl2, 0.2;yeast extract (Becton Dickinson, Oxford, U.K.), 5.0; and 2.5mL/L trace metal solution (citric acid, 0.5 g; H3BO4, 0.30 g;ZnCl2, 0.05 g; CuCl2 � 2H2O, 0.01 g; MnCl2 � 4H2O, 0.03g; NiCl � 6H2O, 0.20 g; NaMoO4 � 2H2O, 0.03 g, dissolvedin 1 L). Polypropylene glycol was used as antifoam at aconcentration of 1.0 mL/L. Glucose, yeast extract, and thia-mine solutions were sterilized separately and added post-
sterilization to the salts solution. Glucose and (NH)4SO4
solutions (50%) were fed to the culture from 1 h after in-oculation at rates of 0.38 and 0.17 g/L/h, respectively. Thisresulted in an initial accumulation of both components be-fore eventual depletion at higher biomass concentrations.Medium B, used for broth dilution in some toxicity experi-ments, was medium A with glucose excluded.
The batch medium used for the E. coli strain was similaras described above except that the initial glucose concen-tration was reduced to 5 g/L in order to avoid overflowmetabolism in the presence of high glucose levels leading toacetate formation, which in turn can result in toxicity to theculture (Meyer et al., 1984). In addition, thiamine wasadded to the E. coli cultures at a concentration of 0.01 g/L.Glucose at a concentration of 500 g/L with 2× concentrationof medium A was fed at a rate of 2.3 mL/L/h at 1 h afterinoculation. This feeding rate resulted in a specific growthrate of 0.1 h−1 which was well below the 0.35 h−1 reportedfor acetate toxicity (Meyer et al., 1984).
Analyses
Biomass and Glucose
Biomass concentration was determined by measuring theoptical density (OD) at 660 nm by means of a double-beamspectrophotometer (Kontron, Watford, Hertfordshire, U.K.)and using a previously established OD versus cell dryweight (100°C for 24 h) calibration. Repeated measure-ments showed variations of ±3%.
Glucose analysis was performed using a Dionex ion chro-matography system (Dionex Corp., Sunnydale, CA) (Aman-ullah et al., 2002).
Indene and Its Metabolites
Indene and its metabolites were extracted from the brothusing propan-2-ol in a 1:1 ratio, and reverse-phase chroma-tography was used with a Dionex HPLC (Dionex Corp.)using a Zorbax RX-C8 column as reported previously(Amanullah et al., 2002).
Flow Cytometry
Flow cytometric analyses were conducted using a CoulterEPICS ELITE analyser (Beckman Coulter Ltd., High Wy-combe, Buckinghamshire, U.K.). All solutions were filteredusing a 0.2-�m filter to remove particulates that could in-terfere with the analysis. Culture samples were diluted withDulbecco’s buffered saline (DBS, pH 7.2) and stained withPI/BOX. The working concentration of PI (Sigma, Poole,Dorset, U.K.) and BOX (Molecular Probes) in DBS were 5and 10 mg/mL, respectively. PI fluorescence was measuredat 630 nm, while BOX fluorescence was measured at 525nm. The total number of cells analyzed per sample was50,000, resulting in a standard error of <0.33% in the mea-
408 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 81, NO. 4, FEBRUARY 20, 2003

surements. Cell physiology was quantified using fluorescentprobes in terms of cell populations with intact membranes(regarded as viable cells), intact polarized membrane, intactdepolarized membrane, and dead cells. Ethidium bromide(EB) (Molecular Probes) was also used to investigatewhether the strains possessed efflux pump systems capableof expelling EB (see later). The working concentration ofEB in DBS was 10 mg/L, and its fluorescence was measuredat 575 nm. Since there was considerable spectral overlapbetween EB and PI emitted fluorescence, the systems’ com-pensation was set up such that PI-emitted fluorescence waseliminated from the EB-emitted fluorescence detector(Hewitt et al., 1999b).
Toxicity Measurements in Bioreactors
Toxicity measurements were conducted under fermentationconditions in a SIXFORS system (Infors AG, Bottmingen,Switzerland). The SIXFORS system consisted of six iden-tical mini-bioreactors constructed of borosilicate glass.Each bioreactor was equipped with dissolved oxygen ten-sion (DOT), pH and temperature probes, a ring sparger, twomagnetically driven turbine impellers, exit air condenser,and sampling line. The agitation speed, aeration rate, pH,and temperature could be independently controlled in eachfermenter. In this study, identical conditions were main-tained in each bioreactor: working volume � 400 mL(maximum working volume � 500 mL), temperature �30°C (37°C for E. coli), pH � 7.2 ± 0.1, agitation speed �650 rpm (the maximum possible), and an aeration rate of1.5 vvm.
Typically 2.5 L of fermentation broth from the exponen-tial growth phase (4.6 dry cell weight/L) was removed fromthe 14-L bioreactor and 400 mL was aseptically transferredundiluted to each of the previously sterilized mini-bioreactors. A smaller broth volume was used when dilutionwith medium B was necessary to achieve a biomass con-centration of 4.6 g/L. The oxygen uptake rate of the cultureswas measured according to Amanullah et al., 2002, fol-lowed by the addition of indene and the other chemicals tothe desired concentrations. Indene and 1-indanone were ob-tained from Fluka (Buchs, Switzerland), and 1-indenol, cis-indandiol, keto-1-hydroxy-2-indan, and cis-(1S,2R)-1-aminoindan-2-ol were kindly donated by Merck & Co., Inc.Indene was poorly water-soluble (≈100 mg/L at 30°C), andsince the added concentrations exceeded the solubility limit,indene existed as a second phase. All other chemicals weresoluble in the fermentation medium at the concentrationsused. The oxygen uptake rate of the cultures was measuredagain after 1 h of incubation with the chemicals. One bio-reactor where no chemical was added was used as the con-trol. Samples were taken after 1 h of incubation for mea-surement of optical density (OD) at 660 nm (measured witha standard error of 3%), flow cytometric analysis and HPLCanalysis of residual indene and its metabolites.
Toxicity Measurements in Shake Flasks
Experiments were also performed in shake flasks cultures inorder to study the influence of high concentrations ofchemicals while minimizing the amounts used. Brothsamples of 20 mL were aseptically removed from the 14-Lbioreactor and placed in separate 200-mL shake flasks. Thebiomass concentration was measured as 4.5 g dry cellweight/L. The chemical of interest was added at the desiredconcentration and incubated for 1 h at 200 rpm and 30°C(37°C for E. coli). Samples were removed from each flaskfor physiological measurements using flow cytometry andcompared to a control.
RESULTS
Staining Controls
Controls were established in order to differentiate betweencells with intact membranes (viable) and cells with non-intact membranes (dead cells). Figure 2a shows, by way ofexample, the light scattering properties (plot of forwardangle scatter against side angle scatter) using E. coli. Thischaracteristic light scattering property of E. coli is discussedlater. Figure 2b shows results with dual staining of an E. coliculture in late-exponential phase using PI/BOX to differen-tiate between viable and non-viable cells and between po-larized and depolarized cells. PI-negative cells (92.4%) withan intact membrane are depicted on the left hand portion ofthe figure (lower and upper left quadrants). PI-positive cellswith a non-intact membrane, which were considered to benon-viable, are depicted on the right-hand portion of thefigure (upper and lower right quadrants). These two popu-lations of cells are also shown in the PI histogram (Fig. 2d).Lower left quadrant in Fig. 2 shows PI-negative/BOX-negative cells (86.1%) representing polarized cells with anintact membrane, and these were considered to be healthyviable cells. Quadrant 2 shows PI-negative/BOX-positivecells (6.3%) representing depolarized cells with an intactmembrane, and these were considered to be viable but meta-bolically stressed (with a disrupted electron transport sys-tem across the membrane) cells. These two populations ofcells are also shown in the BOX histogram (Fig. 2c). Upperright quadrant shows PI-positive cells (6.0%) representingcells with a non-intact membrane, and these were consid-ered to be dead.
Light Scattering Properties
Figure 3 shows the light scattering properties of the fourstrains with forward-angle scatter (FALS) plotted againstthe orthogonal (right angle or RALS) scatter and can beused to detect bacteria from the background signal (Robert-son et al., 1998 Hewitt et al., 1999b). FALS can be used tomeasure cell size provided the refractive index of the me-dium does not change and RALS can be used as a measureof cell granularity (Nebe-von Caron and Badley, 1995).Analysis of Rhodococcus I24 (Fig. 3a), P. putida 421-5
AMANULLAH ET AL.: MEASUREMENT OF STRAIN DEPENDENT TOXICITY USING MULTIPARAMETER FLOW CYTOMETRY 409

(Fig. 3b), and P. putida GM 730 (Fig. 3c) strains revealed aunimodal distribution with respect to both FALS andRALS. The FALS signal for both Rhodococcus I24 (Fig. 3a)and P. putida 421-5 (Fig. 3b) resulted in similar diffuseclusters, while the signal was considerably lower for the P.putida GM 730 strain, indicating the much smaller size ofthe latter. This was also confirmed microscopically. It ap-
pears that in spite of the advances in flow cytometric bac-terial detection, we may be at the limits of detection withthis strain. E. coli TDO 123 generated a unimodal distribu-tion with respect to FALS and a bimodal distribution withrespect to RALS (Fig. 3c). It is thought that this ratherdiffuse cluster may arise because cells which are elongatedcell can vary in range from the major to the minor axis as
Figure 2. (a) Light scattering properties (plot of forward-angle scatter against side-angle scatter) using E. coli. (b) Results of dual staining of an E. coliculture in late-exponential phase using PI/BOX to differentiate between viable and non-viable cells and between polarized and depolarized cells. PI-negativecells (92.4%) with an intact membrane are depicted on the left-hand portion of the figure (lower and upper left quadrants). PI-positive cells with a non-intactmembrane, which were considered to be non-viable, are depicted on the right-hand portion of the figure (upper and lower right quadrants). These twopopulations of cells are also shown in the PI histogram (d). The lower left quadrant shows PI-negative/BOX-negative cells (86.1%) representing polarizedcells with an intact membrane, and these were considered to be healthy viable cells. The upper left quadrant shows PI-negative/BOX-positive cells (6.3%)representing depolarized cells with an intact membrane, and these were considered to be viable but metabolically stressed (with a disrupted electrontransport system across the membrane) cells. These two populations of cells are also shown in the BOX histogram (c). The upper right quadrant showsPI-positive cells (6.0%) representing cells with a non-intact membrane and which were considered to be dead.
410 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 81, NO. 4, FEBRUARY 20, 2003

they travel through the LASER beam (Hewitt et al., 1999b).Importantly it can be seen that each strain displays a distinct“fingerprint” light scattering property.
Toxicity Studies with Rhodococcus I24
We have chosen to define toxicity in this study as an effectwhere a measured parameter is 10% lower compared to thevalue of the control during a test duration of 1 h. Othershave used a less stringent criterion. For instance Sikemmaand co-workers (1992) and Vermue and co-workers (1993)have used a value of 50% relative to the control. Figure 4shows, by way of example, indene toxicity results using
Rhodococcus I24. The concentrations of the chemicals cho-sen in these tests ranged from those typically measured inthe bioconversion of indene to much higher levels. Figure 4shows the measurements of intact cell membrane, polarizedcells with intact membranes, optical density (OD), and oxy-gen uptake rate (OUR) normalized to a control using Rho-dococcus I24. Indene concentrations of up to 0.25 g/L(0.037 g indene/g dry cell weight) did not significantly af-fect intact membrane, membrane polarization, respiratoryactivity, or biomass growth. Dissipation of the membranepotential of a significant cell population occurred at 0.50g/L (0.074 g indene/g dry cell weight). Between this valueand 1.5 g/L (0.221 g indene/g dry cell weight), the changes
Figure 3. Strain-dependent light scattering properties.
AMANULLAH ET AL.: MEASUREMENT OF STRAIN DEPENDENT TOXICITY USING MULTIPARAMETER FLOW CYTOMETRY 411

in these parameters were relatively insignificant with smalldecreases compared to the control, although the OUR de-creased substantially at 1.5 g/L. At 2.5 g/L (0.368 g in-dene/g dry cell weight) there was a significant increase inthe dead cell population. Since a 10% decrease compared tothe control has been defined as a toxic effect in this study,an indene concentration of 0.5 g/L or greater is consideredas toxic to the culture.
The effects of adding 1-indenol in cultures of Rhodococ-cus I24 showed that membrane polarization decreased by19% compared to the control at a concentration of 1.5 g/L.A substantial decrease in the OUR was measured at 3 g/Land was accompanied by membrane depolarization of al-most the entire cell population, whereas the changes in ODand intact cell population only decreased by ∼10% even upto concentrations of 6 g/L of 1-indenol. The measurementsof cell physiological parameters and metabolic activity inresponse to 1-indanone exposure showed that significantdecreases in the polarized cell population, OUR and ODoccurred at 2.5 g/L compared to the control, whereas theintact cell membrane population only decreased at a con-centration of 6 g/L, which was also sufficient to cause mem-brane depolarization of the entire cell population. Nochanges relative to the control could be measured up to 1g/L of 1-keto-2-hydroxy-indan. Membrane polarization andOUR decreased by 16% and 20%, respectively, at 1.8 g/L,whereas the intact cell membrane population and OD re-mained unaffected until 3 g/L. Data using >3 g/L of 1-keto-2-hydroxy-indan were from shake flask tests, which wereemployed to minimize the quantities of the chemical used,and as a result OUR measurements could not be made. Theoverlapping data points at 3 g/L suggested that the results ofthe toxicity experiments in shake flasks were comparable tothose in the bioreactors.
Toxic effects could not be measured using either I24cis-(1S,2R)-indandiol and cis-(1S,2R)-1-aminoindan-2-ol[(−)-CAI] even with concentrations as high as 10 g/L. Tox-icity as measured by a ∼15% decrease in the polarized mem-brane population and OUR was only evident at 20 g/L (datanot shown).
Comparison of Strains with Respect toIndene Toxicity
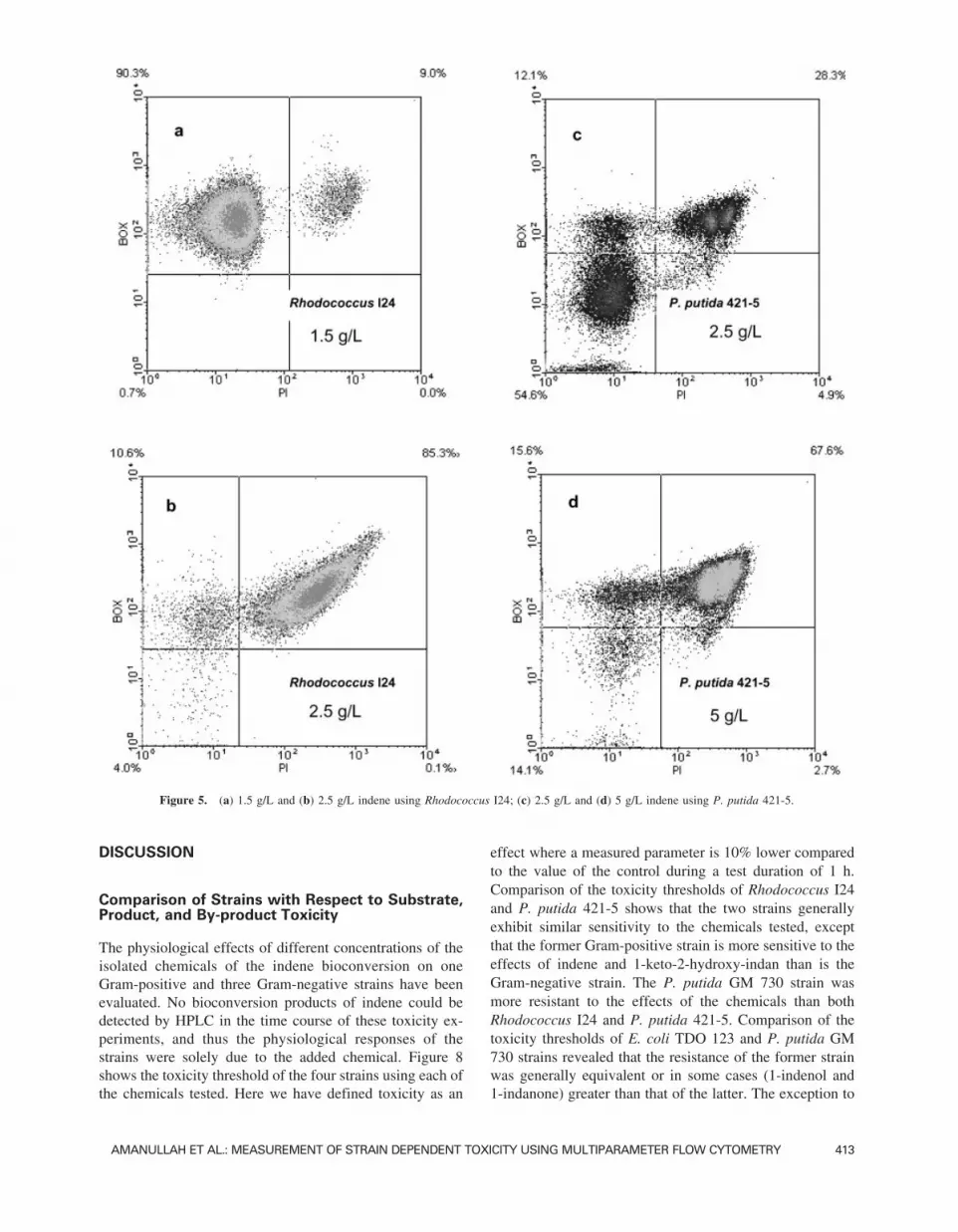
The effects of indene toxicity on the four strains are shownby way of an example in Figs. 5 and 6. The biomass con-centration of each culture was held constant at ∼4.6 g drycell weight/L. The choice of indene concentrations shown inthese figures was based on demonstrating both non-toxic aswell as toxic effects with respect to the intact cell membranepopulation. The results of the non-toxic concentrations weresimilar to the controls in all cases. Figures 5a and b show theeffects of indene exposure at concentrations of 1.5 and 2.5g/L with Rhodococcus I24. The dead cell population in-creased from 9% to 85.3% using indene at 1.5 and 2.5 g/L,respectively, and clearly the latter concentration was toxicto the culture. With the P. putida 421-5 strain, the use of 2.5g/L of indene (Fig. 5c) did not result in significant changescompared to the control (data not shown) with ∼55% and67% of the cell population remaining polarized and with anintact cell membrane, respectively. A significant change oc-curred at 5 g/L (Fig. 5d) with an increase in the in the deadcell population to ∼70%, whereas at 5 g/L with E. coli TDO123 very little change was measured with respect to thecontrol (data not shown), with nearly 85% and 92% remain-ing polarized and with an intact cell membrane, respectively(Fig. 6a). Toxic effects with this strain were only measuredat an indene concentration of 7.5 g/L (Fig. 6b). The indenetoxicity threshold was even higher with the P. putida GM730 strain, where 15 g/L of indene was required to causesignificant changes in the polarized and dead cell popula-tions (Fig. 6d).
Comparison of Strains with Respect toProduct Toxicity
Figures 7a and b show by way of an example the results ofexposure to 20 g/L cis-(1S,2R)-1-aminoindan-2-ol and 20g/L cis-(1S,2R)-indandiol, respectively, using P. putida GM730. It is clear that this strain was able to tolerate at least 20g/L of either chemical. The increase in the depolarized cellmembrane population with the use of cis-(1S,2R)-indandiolcompared to the use of cis-(1S,2R)-1-aminoindan-2-ol wasprobably due to the presence of impurities in cis-(1S,2R)-indandiol, which was isolated and purified via bioconver-sion, whereas the latter chemical was made via a chemicalsynthesis, with possibly lower impurities. Figures 7c and dshow the results of exposure of E. coli TDO 123 to 20 g/Lcis-(1S,2R)-1-aminoindan-2-ol and 20 g/L cis-(1S,2R)-indandiol, respectively. Similar results were obtained withboth chemicals, suggesting that this strain was more tolerantto the impurities discussed earlier. As with the P. putidaGM 730 strain, the E. coli TDO 123 strain was also capableof tolerating at least 20 g/L of the desired products. Theseproducts manifested toxic effects on Rhodococcus I24 andP. putida 421-5 strains at concentrations in excess of 15 g/L(data not shown).
Figure 4. Indene toxicity using Rhodococcus I24.
412 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 81, NO. 4, FEBRUARY 20, 2003
DISCUSSION
Comparison of Strains with Respect to Substrate,Product, and By-product Toxicity
The physiological effects of different concentrations of theisolated chemicals of the indene bioconversion on oneGram-positive and three Gram-negative strains have beenevaluated. No bioconversion products of indene could bedetected by HPLC in the time course of these toxicity ex-periments, and thus the physiological responses of thestrains were solely due to the added chemical. Figure 8shows the toxicity threshold of the four strains using each ofthe chemicals tested. Here we have defined toxicity as an
effect where a measured parameter is 10% lower comparedto the value of the control during a test duration of 1 h.Comparison of the toxicity thresholds of Rhodococcus I24and P. putida 421-5 shows that the two strains generallyexhibit similar sensitivity to the chemicals tested, exceptthat the former Gram-positive strain is more sensitive to theeffects of indene and 1-keto-2-hydroxy-indan than is theGram-negative strain. The P. putida GM 730 strain wasmore resistant to the effects of the chemicals than bothRhodococcus I24 and P. putida 421-5. Comparison of thetoxicity thresholds of E. coli TDO 123 and P. putida GM730 strains revealed that the resistance of the former strainwas generally equivalent or in some cases (1-indenol and1-indanone) greater than that of the latter. The exception to
Figure 5. (a) 1.5 g/L and (b) 2.5 g/L indene using Rhodococcus I24; (c) 2.5 g/L and (d) 5 g/L indene using P. putida 421-5.
AMANULLAH ET AL.: MEASUREMENT OF STRAIN DEPENDENT TOXICITY USING MULTIPARAMETER FLOW CYTOMETRY 413

this finding was for indene, where the toxic concentrationwas 50% higher at 7.5 g/L for P. putida GM 730 comparedto E. coli TDO 123. In general, the resistance of the strainsto the chemicals tested decreased in the following order: E.coli TDO 123; P. putida GM 730; P. putida 421-5; Rho-dococcus I24, with the Gram-negative strains exhibitinghigher tolerance to the chemicals than the Gram-positivestrain. In addition, 1-indanone was found to be the mosttoxic of the chemicals tested followed by indene. The timescale used in the toxicity studies allows primarily for pri-mary protective mechanisms, and it can be argued that it isnot representative of conditions on a bioprocessing scale,where secondary response mechanisms can play a role.
However, as shown here, the toxic effects of the chemicalsused on the biocatalysts are lethal and irreversible even ona primary response scale, and it is unlikely that protectivemechanisms over a longer time scale will have an impact onchemical tolerance. The latter is only likely to be effectiveif the primary effects are sublethal and reversible.
Relationship between Cytoplasmic MembraneDepolarization and Oxygen Uptake Rate
Figure 9, where the oxygen uptake rate (normalized to acontrol) is plotted against the fraction of polarized cellpopulation (normalized to a control) for each of the strains
Figure 6. (a) 5 g/L and (b) 7.5 g/L indene using E. coli TDO 123; (c) 10 g/L and (d) 15 g/L indene using P. putida GM 730.
414 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 81, NO. 4, FEBRUARY 20, 2003

tested in response to indene and 1-indanone exposure. Thisis the first time such results have been reported and are alsoimportant since it is possible to relate the measurement ofmembrane potential on a single-cell basis via flow cytom-etry to a more commonly measured parameter such as oxy-gen uptake rate. The decrease in respiratory activity withmembrane depolarization shown in Fig. 9 may be expectedsince the respiratory enzymes are located within the cyto-plasmic membrane and impairment of the functioning of themembrane is likely to influence the respiratory activity ofthe cell. Enzymes known to be located in the cytoplasmicmembrane include ATPases, transport proteins, transfer-ases, and various oxidoreductases. Osborne and co-workers(1990) found a correlation between 11-�-hydroxylase ac-tivity of R. nigricans and the solvent concentration in the
cell membrane. In addition, Sikkema et al. (1992) alsoshowed that tetralin inhibited the activity of cytochrome coxidase in strains of Arthrobacter.
Mode of Action of the Toxic Chemicals
The results presented in this study show that dissipation ofthe membrane potential and loss of respiratory activity oc-curred in a concentration-dependent manner and provided asensitive measurement of physiological stress. In compari-son to the decreases in membrane potential and respiratoryactivity, the changes in optical density and cytoplasmicmembrane integrity were disproportionately smaller at lowconcentrations of the chemicals in all tests, suggesting that
Figure 7. (a) 20 g/L cis-(1S,2R)-1-aminoindan-2-ol using P. putida GM 730; (b) 20 g/L cis-(1S,2R)-indandiol using P. putida GM 730; (c) 20 g/Lcis-(1S,2R)-1-aminoindan-2-ol using E. coli TDO 123; and (d) 20 g/L cis-(1S,2R)-indandiol using E. coli TDO 123.
AMANULLAH ET AL.: MEASUREMENT OF STRAIN DEPENDENT TOXICITY USING MULTIPARAMETER FLOW CYTOMETRY 415

the primary mode of action of the chemicals was the per-turbation of the membrane-associated energy transductionprocess in the cells rather than membrane disruption. Theresults presented here also highlight the danger in relyingsolely on optical density as a measure of bacterial growthwhen assessing the effects of chemical toxicity. It is pos-sible that pore formation in the membrane due to the actionof the chemicals may affect energy generation by an effluxof glycolysis intermediates, ions, and/or reduced enzymeactivities after intracellular acidification (Bennik et al.,1997). Boucabeille et al. (1998) also reported an immediateperturbation of the permeability properties of the cytoplas-mic membrane of the Gram-positive Listeria innocua due tothe formation of pore like structures, resulting in a loss ofpotassium, membrane depolarization, hydrolysis of internalATP, and efflux of inorganic phosphate in response to lowdoses of the bacteriocin, linenscin OC2. Furthermore, in areview of bacteriocins of Gram-positive bacteria, Jack andco-workers (1995) reported that treatment of differentstrains to the toxin, nisin, resulted in membrane depolariza-tion and efflux of radiolabeled compounds and of ATP,whereas changes in the concentrations of larger molecules,such as sugars and proteins, were negligible, promptingspeculation that the toxin formed pores in the cytoplasmic
membrane rather than producing complete membrane dis-ruption. We have also previously found that extracellularmeasurements of cations by HPLC revealed that exposureof the Rhodococcus I24 strain to indene resulted in thenonselective permeability of the cytoplasmic membranewith a cellular efflux of both NH4
+ and K+ of up to 60 and70 mg/L, respectively, at an indene concentration of 2.5 g/L,accompanied by an influx of Na+ of up to 390 mg/L (Aman-ullah et al., 2002). At higher concentrations of the chemi-cals, the pores formed in the cytoplasmic membrane may beof sufficiently large diameters to cause complete loss ofmembrane integrity, resulting in cell death, which may beexacerbated by osmoinduced influx of water through thepores (Jack et al., 1995).
Mechanisms of Bacterial Tolerance toToxic Chemicals
The accumulation of lipophilic compounds in the cytoplas-mic membrane can have significant effects on their struc-tural and functional properties as shown in this study. How-ever, bacteria can counter-balance these effects in severalways. Probably the most important factor responsible for theincreased tolerance of the Gram-negative strains to thechemicals of the indene bioconversion is the presence of theouter membrane, a structure lacking in Gram-positivestrains. Inoue and Horikoshi (1989) reported that Gram-negative bacteria were able tolerate solvents with log P(logarithm of the octanol–water partition coefficient) as lowas 2.4 while Gram-positive strains could not grow in thepresence of solvents with log P values under 4.9. Theseauthors used 32 Gram-negative and 28 Gram-positivestrains in their study. Similar differences in solvent toler-ance between Gram-positive and Gram-negative bacteriawere reported by Harrop and co-workers (1989). Vermueand co-workers (1993) distinguished between two forms oftoxicity (molecular and phase). Molecular toxicity was at-tributed to molecules of the chemicals dissolved in the aque-ous phase, resulting in enzyme inhibition, protein denatur-ation and/or membrane modification. Phase toxicity, on theother hand, was caused by the presence of a second phase,and it was postulated to result in the extraction of nutrientsand cell wall components. They also reported that in gen-eral, the tolerance of the Gram-negative Acinetobacter andPseudomonas strains was higher than that of the Gram-positive Arthrobacter and Nocardia strains. Ming and Dae-schel (1993) suggested membrane fluidity as an alternativemechanism to explain the differences in bacterial sensitivityto toxic chemicals, where a less compact membrane mayfacilitate pore formation. Membrane fluidity can be regu-lated in cells by alteration of their membrane compositionfrom cis- to trans-unsaturated fatty acids (Weber et al.,1994). This isomerization occurs rapidly (<15 min) anddoes not require an energy input (de Bont, 1998). However,at best the membrane structure and mechanism cited abovecan counteract the effects of toxic chemicals temporarily.The active removal of chemicals from the cytoplasmic
Figure 8. Strain-dependent substrate, by-product, and product toxicity.Toxicity is defined as a 10% decrease in measured parameter compared tocontrol.
416 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 81, NO. 4, FEBRUARY 20, 2003

membrane would be a dynamic and longer-lasting mecha-nism of tolerance.
Active efflux systems in bacteria have been extensivelystudied (refer to reviews by Levy, 1992; Sikkema et al.,1995; Nikaido, 1994). It is well known that multi-drug ef-flux pumps in bacteria can export a wide range of structur-ally dissimilar compounds (Levy, 1992; Li et al., 1998;Ramos et al., 1998). Solvent tolerance in E. coli has alsobeen reported to be linked to the expression of multi-drugresistant genes (Aono et al., 1991). Isken and de Bont(1996) using a solvent-tolerant strain of P. putida S12showed that the accumulation of 14C-labeled toluene dem-onstrated an energy-dependent efflux system. This strainwas also found to be resistant to a number of antibiotics.Subsequently Kieboom and co-workers (1998) cloned thegenes responsible for the efflux pumps in P. putida S12.The amino acid sequences encoded by these three geneswere significantly similar to the proton dependent multi-drug efflux pumps. These pumps are composed of threeproteins that are thought to span the inner and outer mem-branes of Gram-negative bacteria. Many studies (Li et al.,
1991; Molenaar et al., 1992; de Bont, 1998; Ramos et al.,1998) have reported that efflux pump systems are energydependent (either the generation of ATP or the presence ofa cytoplasmic membrane potential).
One way of demonstrating the presence of an effluxpump system in cells with a membrane potential, which maybe responsible for efflux of toxic chemicals as one possiblemethod to avoid toxicity, was via the use of the cationicfluorescent probe ethidium bromide (EB; MolecularProbes). EB binds to DNA and can cross the cytoplasmicmembrane, but it is actively pumped out in most cells via anon-specific proton/antiport (Nebe-von Caron et al., 2000).Ethidium bromide was used with PI/BOX as a triple stain(PI/BOX/EB) to investigate whether efflux pump systemscapable of expelling EB were present in the strains studied.Figure 10a shows EB/PI/BOX staining of the Gram-positiveRhodococcus I24 taken from the exponential phase of afermentation. Quadrant 1 represents unstained cells, whichare polarized cells actively pumping out EB. The resultsindicate that there are virtually no pumping cells with thisstrain. Quadrant 2 represents non-pumping, polarized cells,
Figure 9. Relationship between oxygen uptake rate and membrane polarization for the four strains tested. Both parameters are normalized to the control.
AMANULLAH ET AL.: MEASUREMENT OF STRAIN DEPENDENT TOXICITY USING MULTIPARAMETER FLOW CYTOMETRY 417

which stained with EB. Over 68% of the cell population fellin this category. Cells stained with BOX and EB (non-pumping depolarized cells with an intact cytoplasmic mem-brane) are shown in quadrant 3 and represent 28% of thecell population. Cells stained with PI, BOX, and EB areshown in quadrant 4 and are considered to be dead cells.Figure 10b shows EB/PI/BOX staining of an exponential-
phase Gram-negative P. putida GM 730 strain. With thisstrain, nearly 75% of the cell population possessed pumpsystems capable of expelling EB (quadrant 1), while only6.4% of the cell population were non-pumping polarizedcells (quadrant 2). Non-pumping, depolarized cells with anintact cytoplasmic membrane constituted 10.8% of thepopulation, while the dead cell population accounted for7.9%. Similar results were obtained with the Gram-negativeE. coli TDO 123 strain. Thus it appears that the Gram-negative strains tested here have efflux pump systems thatare capable of exporting EB, while the Gram-positive straindoes not. This characteristic may be a contributory factor tothe higher tolerance shown by the Gram-negative strains inresponse to the chemicals of the indene bioconversion. Themechanism by which these pumps function is unclear, al-though a diffusion barrier must be present in order to allowthe pump to function in an effective manner (de Bont,1998). The outer membrane of Gram-negative bacteria iscommonly viewed as a barrier to toxic compounds, and theefflux pump systems would be expected to function in tan-dem with this barrier.
Targets for Strain/Bioprocess Improvement
Based solely on the results of tolerance in response to ex-posure of the chemicals of the indene biotransformation, themost appropriate hosts to conduct this biotransformationappear to be either the E. coli TDO 123 or P. putida GM730 strains. The toluene dioxygenase activity in P. putidahas been shown to exhibit poor stereoselectivity, with enan-tiomerically pure cis-(1S,2R)-indandiol obtained only atlong bioconversion times due to the resolution catalyzed bya cis-(1R, 2S)-indandiol dehydrogenase (Connors et al.,1997). Although Rhodococcus I24 offers advantages interms of high stereoselectivity in relatively short times, ithas been shown here to be the least-tolerant strain whenexposed to the chemicals of the indene biotransformation.The tolerance of E. coli TDO 123 strain has been shownhere to be nearly equal to that of P. putida GM 730 strain.In addition, E. coli offers the advantages of a well-studiedmetabolism and methods have already been established togrow high cell density cultures for enhanced productivity.Based on the results obtained in this study, there are severalpossibilities for enhancing the performance of the strainsand/or bioprocess. The first possibility is to engineer thestrains by deleting the dehydrogenase activity that is respon-sible for converting cis-(1S,2R)-indandiol to 1-keto-2-hydroxy indan (Fig. 1), allowing the accumulation of cis-(1S,2R)-indandiol. This study has demonstrated that thiswould be an attractive option since the cells can toleratelevels as high as 20 g/L of cis-(1S,2R)-indandiol withoutcausing changes in cell physiology. The E. coli TDO 123strain is in fact such a strain and comparison to another E.coli D-160-1 strain which does have the dehydrogenase ac-tivity showed that the ee of cis-indandiol was reduced by66% in the former strain (Buckland et al., 1999). The de-crease in enantiomeric excess is offset by an increase in
Figure 10. (a) EB/PI/BOX staining of Rhodococcus I24. Quadrant 1:unstained cells (polarized cells actively pumping out EB). Quadrant 2: cellsstained with EB (non-pumping polarized cells). Quadrant 3: cells stainedwith BOX and EB (non-pumping depolarized cells with an intact cytoplas-mic membrane). Quadrant 4: cells stained with PI, BOX, and EB (deadcells). (b) EB/PI/BOX staining of P. putida GM 730. The legends for thequadrants are as above.
418 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 81, NO. 4, FEBRUARY 20, 2003

yield due the absence of 1-keto-2-hydroxy indan. The netresult of this would be a ∼20% increase in the yield ofcis-indandiol from indene. This strategy would also requirethe removal of the toxic mono-oxygenated by-products atconcentrations >2.5 g/L via in situ product removal tech-niques (Lye and Woodley, 1999).
An alternative strategy would be to reduce the dioxygen-ase activity that converts indene to the monooxygenatedby-products (i.e., increase the ratio of the concentrations ofcis-indandiol to the monooxygenated by-products) via di-rected evolution strategies, combined with in situ removalof cis-(1S,2R)-indandiol or with the deletion of the dehy-drogenase activity preventing the formation of 1-keto-2-hydroxy indan (the implications for reduced enantiomericexcess and increased yield have been discussed earlier).Removal of indandiols from a biotransformation involvingPseudomonas strains has been demonstrated by Barbirato etal. (1998). The use of transaminases that can convert cis-indandiol to cis-1-amino-2-indanol is also an interesting op-tion since it has also been demonstrated here that it is pos-sible to accumulate the latter up to 20 g/L without signifi-cant negative effects on cell physiology using all of thestrains tested.
Stafford et al. (2001) using a mutant strain of Rhodococ-cus I24 (Rhodococcus KY1) suggested that a promisingapproach to increase the yield of the (1R,2R)-indandiols wasto hydrolyze selectively indan oxide to trans-(1R,2R)-indandiol by introducing a limonene-1,2-epoxide hydrolase.The activity of the expressed enzyme was estimated to besufficient to support the amount of indan oxide produced.This approach would nullify the need for the dehydrogenaseknockout as little or no cis-(1S,2R)-indandiol would be pro-duced. However, accumulation of trans-(1R,2R)-indandiolcan still be problematic since it has also been shown thataccumulation of this compound even to levels of 1.5 g/L caninhibit the monooxygenase activity by as much as 50%(Stafford et al., 2000). Thus in order to benefit from thisgenetic modification approach, in situ removal of the trans-(1R,2R)-indandiol should also be incorporated into thisstrategy.
CONCLUSION
Multiparameter flow cytometry has been used to evaluatethe effects of exposure to chemicals of the indene biotrans-formation on cell physiology using one Gram-positive andthree Gram-negative strains, all of which were capable oftransforming indene to cis-indandiol. It has been shown hereto be a rapid and powerful technique for the measurement oftoxicity in whole cell biocatalysis. Measurements of thecytoplasmic membrane potential, respiratory activity, mem-brane integrity, and biomass growth provided a sensitive setof parameters to evaluate toxicity. The primary mode ofaction at low concentrations of the chemicals was probablythe formation of pores resulting in the perturbation of themembrane-associated energy transduction process in thecells rather than total membrane disruption. At higher con-
centrations of the chemicals, the pores formed in the cyto-plasmic membrane may be of sufficiently large diameters tocause complete loss of membrane integrity, resulting in celldeath. The decrease in respiratory activity with a dissipationof the cytoplasmic membrane potential was measured in allfour strains. The toxic limits of the substrate, by-products,and product have been determined, and the information ob-tained may be used to implement strain/bioprocess improve-ments as well to provide guidelines in the operation of thebiotransformation. It has been demonstrated that it is pos-sible to accumulate cis-(1S,2R)-indandiol and cis-1-amino-2-indanol up to 20 g/L without significant negative effectson cell physiology using all of the strains tested. The Gram-negative P. putida (421-5 and GM 730) and E. coli TDO123 strains were more resistant to indene and the isolatedchemicals of the biotransformation than the Gram-positiveRhodoccoccus I24. In general, the P. putida GM 730 and theE. coli TDO 123 strains responded similarly to toxic effectsof the chemicals, whilst the E. coli TDO 123 strain wasmore resistant than the 421-5 strain. Based on the results oftolerance in response to exposure of the chemicals of theindene biotransformation, the most appropriate hosts to con-duct this biotransformation appear to be either the E. coliTDO 123 or P. putida GM 730 strains. Probably the mostimportant factor responsible for the increased tolerance ofthe Gram-negative strains was the presence of the outermembrane, a structure lacking in Gram-positive strains. Atriple stain was used to demonstrate the presence of effluxpump systems in the Gram-negative strains whilst it wasshown to be absent in the Gram-positive strain. The pres-ence of the efflux mechanisms may also be a contributoryfactor for the higher tolerance shown by the Gram-negativestrains in response to exposure to the chemicals of the in-dene bioconversion. The targets for bioprocess improve-ment includes a combination of genetic as well as engineer-ing approaches.
REFERENCES
Allman R, Manchee R, Lloyd D. 1993. Flow cytometric analysis of het-erogeneous bacterial populations. In: Lloyd D, editor. Flow cytometryin microbiology. London: Springer-Verlag. p 27–47.
Amanullah A, Hewitt CJ, Nienow AW, Lee C, Chartrain M, Buckland BC,Drew SW, Woodley JM. Application of flow cytometry using fluo-rescent probes to study substrate and product toxicity in the indenebioconversion. Biotechnol Bioeng 83:239–249.
Aono R, Aibe K, Inoue A, Horikoshi K. 1991. Preparation of organicsolvent-tolerant mutants of Escherichia coli. Agric Biol Chem 55:1935–1938.
Barbirato F, Verdoes JC, de Bont JAM, Werf MJ. 1998. The Rhodococcuserythroplois DCL14 limonene-1,2-epoxide hydrolase gene encodes anenzyme belonging to a novel class of epoxide hydrolases. FEBS Lett438:293–296.
Bennik MHJ, Verhuel A, Abee T, Naaktgeboren-Stoffels G, Gorris GM,Smid EJ. 1997. Interactions of nisin and pediocin PA-1 with closelyrelated lactic acid bacteria that manifest over 100-fold differences inbacteriocin sensitivity. Appl Environ Microbiol 63:3628–3636.
Boucabeille C, Letellier L, Simonet J, Henckes G. 1998. Mode of action oflinenscin OC2 against Listeria innocua. Appl Environ Microbiol 64:3416–3421.
AMANULLAH ET AL.: MEASUREMENT OF STRAIN DEPENDENT TOXICITY USING MULTIPARAMETER FLOW CYTOMETRY 419

Boucabeille C, Letellier L, Simonet J, Henckes G. 1998. Mode of action oflinenscin OC2 against Listeria innocua. Appl Environ Microbiol 64:3416–3421.
Buckland BC, Drew SW, Connors NC, Chartrain MC, Lee C, Salmon PM,Gbewonyo K, Zhou W, Gailliot P, Singhvi R, Olewinski RC, Sun W,Reddy J, Zhang J, Jackey BA, Taylor C, Goklen KE, Junker B,Greasham RL. 1999. Microbial conversion of indene to indandiol; akey intermediate in the synthesis of CRIXIVAN. J Metab Eng 1:63–74.
Chartrain M, Jackey B, Taylor C, Sandford V, Gbewonyo K, Lister L,Dimichele HC, Heimbach B, Maxwell C, Pascoe, Buckland BC,Greasham R. 1998. Bioconversion of indene to cis-(1S,2R)-indandioland trans-(1R,2R)-indandiol by Rhodococcus species. J Ferment Bio-eng 86:550–558.
Connors N, Prevoznak R, Chartrain M, Reddy J, Singvi R, Patel Z, Olewin-ski R, Salmon P, Wilson J, Greasham R. 1997. Conversion of indeneto cis-(1S),(2R)-indandiol by mutants of Pseudomonas putida F1. J IndMicrobiol Biotechnol 18:353–359.
Davey HM, Kell DB. 1996. Flow cytometry and cell sorting of heterog-enous microbial populations: the importance of single cell analyses.Microbiol Rev 60:641–696.
de Bont JAM. 1998. Solvent-tolerant bacteria in biocatalysis. Trends Bio-technol 16:493–499.
Harrop AJ, Hocknull MD, Lilly MD. 1989. Biotransformations in organicsolvents: a difference between Gram-positive and Gram-negative bac-teria. Biotechnol Lett 11:807–810.
Hewitt CJ, Lotte AB, McFarlane CM, Nienow AW. 1998. The use of flowcytometry to study the impact of fluid mechanical stress on Esch-erichia coli W3110 during continuous cultivation in an agitated bio-reactor. Biotechnol Bioeng 63:705–711.
Hewitt CJ, Nebe-von Caron G, Nienow AW, McFarlane CM. 1999a. Useof multistaining flow cytometry to characterize the physiological stateof Escherichia coli W3110 in high cell density fed-batch cultures.Biotechnol Bioeng 63:705–711.
Hewitt CJ, Nebe-von Caron G, Nienow AW, McFarlane CM. 1999b. Theuse of multi-parameter flow cytometry to compare the physiologicalresponse of Escherichia coli W3110 to glucose limitation duringbatch, fed-batch and continuous culture cultivation. J Biotechnol 75:251–254.
Ingram LO. 1976. Adaptation of membrane lipids to alcohols. J Bacteriol125:670–678.
Inoue A, Horikoshi K. 1989. A Pseudomonas thrives in high concentra-tions of toluene. Nature 338:264–266.
Isken S, de Bont JAM. 1996. Active efflux of toluene in a solvent-resistantbacterium. J Bacteriol 178:6056–6058.
Jack RW, Tagg JR, Ray B. 1995. Bacteriocins of Gram-positive bacteria.Microbiol Rev 59:171–200.
Kieboom J, Dennis JJ, de Bont JAM, Zylstra GJ. 1998. Identification andmolecular characterisation of an efflux pump involved in Pseudomo-nas putida S12 solvent tolerance. J Biol Chem 273:85–91.
Levy SB. 1992. Active efflux mechanisms for antimicrobial resistance.Microb Agents Chemother 36:695–703.
Li X, Zhang L, Poole K. 1998. Role of multidrug efflux systems of Pseu-domonas aeruginosa in organic solvent tolerance. J Bacteriol180:2987–2991.
Lye GJ, Woodley JM. 1999. Application of in situ product-removal tech-niques to biocatalytic processes. Trends Biotechnol 17:395–402.
McDonald DL, Goldfine H. 1991. Effects of solvents and alcohols on the
polar lipid composition of Clostridium butyricum under conditions ofcontrolled lipid chain composition. Appl Environ Microbiol 57:3517–3521.
Meyer HP, List C, Fiechter A. 1984. Acetate formation in continuouscultures of Escherichia coli K12 D1. J Biotechnol 1:355–358.
Ming X, Daeschel MA. 1993. Nisin resistance of foodborne bacteria andthe specific resistance responses of Listeria monocytogenes Scott-A. JFood Prot 56:944–948.
Molenaar D, Bolhuis H, Abee T, Poolman B, Konings WN. 1992. Theefflux of a fluorescent probe is catalysed by an ATP-driven extrusionsystem in Lactococcus lactis. J Bacteriol 174:3118–3124.
Nebe-von Caron G, Badley RA. 1995. Viability assessment of bacteria inmixed populations using flow cytometry. J Microsc 179:55–66.
Nebe-von Caron G, Stephens PJ, Hewitt CJ, Powell JR, Badley RA. 2000.Analysis of bacterial function by multi-colour fluorescence flow cy-tometry and single cell sorting. J Microbiol Methods 42:97–114.
Nikaido H. 1996. Multidrug efflux pumps of Gram-negative bacteria. JBacteriol 178:5853–5859.
Nikaido H. 1994. Prevention of drug access to bacterial targets: perme-ability barriers and active efflux. Science 264:382–388.
Osborne SJ, Leaver J, Turner MK, Dunnill P. 1990. Correlation of bio-catalytic activity in an organic–aqueous two liquid phase system withsolvent concentration in the cell membrane. Enzyme Microb Technol12:281–291.
Ramos JL, Duque E, Godoy P, Segura A. 1998. Efflux pumps in toluenetolerance in Pseudomonas putida DOT-T1E. J Bacteriol 180:3323–3329.
Robertson BR, Button DK, Koch AL. 1998. Determination of the bio-masses of small bacteria at low concentrations in a mixture of specieswith forward light scatter measurements by flow cytometry. Appl En-viron Microbiol 64:3900–3909.
Sikkema J, Poolman B, Konings WN, de Bont JAM. 1992. Effects of themembrane action of tetralin on the functional and structural propertiesof artificial and bacterial membranes. J Bacteriol 174:2986–2992.
Sikkema J, de Bont JAM, Poolman B. 1995. Mechanisms of membranetoxicity of hydrocarbons. Microbiol Rev 59:201–222.
Shapiro HM. 1995. Practical flow cytometry, 3rd edition. New York:Wiley-Liss Inc. (ISBN 0-471-30376-3).
Stafford DE, Yanagimachi KS, Amanullah A, Drew SW, Buckland BC,Sinskey AJ, Stephanopoulos G. 2000. Characterization of indene bio-conversion in Rhodococcus sp. KY1 continuous cultures using radio-active tracers and multi-channel flow cytometry. Paper 219(1–2):BIOT 69. 219th ACS Spring Meeting, San Francisco, CA, 26–30March 2000.
Stafford DE, Yanagimachi KS, Stephanopoulos G. 2001. Metabolic engi-neering of indene bioconversion in Rhodococcus sp. Adv BiochemEng/Biotechnol 73:85–101.
Uribe SJ, Ramirez J, Pena A. 1985. Effects of �-pinene on yeast membranefunctions. J Bacteriol 161:1196–1200. Washington, DC: AmericanChemical Society.
Vermue M, Sikkema J, Verheul A, Bakker R, Trampers J. 1993. Toxicityof homologous series of organic solvents for the Gram-positive bac-teria Arthrobacter and Nocardia sp. and the Gram-negative bacteriaAcinetobacter and Pseudomonas sp. Biotechnol Bioeng 42:747–758.
Weber FJ, Isken S, de Bont JAM. 1994. Cis–trans isomerization of fattyacids as a defense mechanism of Pseudomonas putida strains to toxicconcentrations of toluene. Microbiology 140:2013–2017.
420 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 81, NO. 4, FEBRUARY 20, 2003





























