Embed Size (px)
Citation preview

INFOMUSAINFOMUSAThe International Magazine on Banana and Plantain
INFOMUSA is published with the support of the Technical Centerfor Agricultural and RuralCooperation (CTA)
CTA
Vol. 10 N° 2December 2001
IN THIS ISSUEIntegrated management ofSigatoka diseases in cv. Africa
Organic fertilization vsinorganic fertilization in “Cachaco” plantain in Colombia
Influence of leaf removal on shoot and root growth in banana
Effect of hail and wind on thefruits of Dominico hartón andFHIA-21
Polyphenol oxidase activityand gene expression in thefruit of Goldfinger
In vitro multiplication of FHIA-18 plantain in thepresence of paclobutrazol
Preliminary results of a virulence test of the populations of Focfor different banana cultivars
Evolution of the nematofaunaof plantain in Côte d’Ivoire
Parasite nematodes of bananain western Burkina Faso
Genetic and genomic diversityin banana based on moleculartools
Banana in images
Classification andcharacterization of Musaexotica, M. alinsanaya and M. acuminata ssp. errans
Thesis
MusaNews
MusaForum
INIBAP News
Books etc.
Announcements
PROMUSA News

Vol. 10, N° 2Cover photo:Fruits stalls in Tagaytay,Philippines.(D. Mowbray, Baobab Productions)
Publisher: International Network for the Impro-vement of Banana and PlantainManaging editor: Claudine PicqEditorial Committee: Emile Frison, Jean-Vincent Escalant,Suzanne Sharrock, Charlotte LustyPrinted in FranceISSN 1023-0076Editorial Office: INFOMUSA, INIBAP, Parc ScientifiqueAgropolis II, 34397 Montpellier Cedex 5,France. Telephone + 33-(0)4 67 61 1302; Telefax: + 33-(0)4 67 61 03 34; E-mail: [email protected]//:www.inibap.orgSubscriptions are free for developingcountries readers. Article contributionsand letters to the editor are welcomed.Articles accepted for publication may beedited for length and clarity. INFOMUSAis not responsible for unsolicited mater-ial, however, every effort will be made torespond to queries. Please allow threemonths for replies. Unless accompaniedby a copyright notice, articles appearingin INFOMUSA may be quoted or repro-duced without charge, provided acknowl-edgement is given of the source.French-language and Spanish-languageeditions of INFOMUSA are also pub-lished.To avoid missing issues of INFOMUSA,notify the editorial office at least sixweeks in advance of a change of address.
Views expressed in articles are thoseof the authors and do not necessarilyreflect those of INIBAP.
INFOMUSA Vol. 10, N° 2
CONTENTS
Integrated management of black and yellow Sigatoka diseases in plantain cv.
Africa ................................................................................................................. 3
Organic fertilization vs inorganic fertilization in ‘Cachaco’ plantain
in Colombia ....................................................................................................... 7
Influence of leaf removal on shoot and root growth in banana
(Musa spp.) ...................................................................................................... 10
The effect of hail and wind on the development and quality of the fruits
of Dominico hartón and FHIA-21 plantain.................................................... 13
Polyphenol oxidase activity and gene expression in the fruit of Goldfinger
bananas (AAAB, FHIA-01)............................................................................... 17
In vitro multiplication of FHIA-18 plantain in the presence
of paclobutrazol.............................................................................................. 22
Preliminary results of a virulence test of the populations of Fusarium
oxysporum f.sp. cubense for different banana cultivars
in the greenhouse........................................................................................... 24
Evolution of the nematofauna of plantain, Musa AAB, in Côte d’Ivoire.......... 26
Parasite nematodes of banana in western Burkina Faso ................................... 28
Genetic and genomic diversity in banana (Musa species and cultivars)
based on D2 analysis and RAPD markers ....................................................... 29
Banana in images.................................................................................................. 34
Classification and characterization of Musa exotica, M. alinsanaya
and M. acuminata ssp. errans......................................................................... 35
Thesis ..................................................................................................................... 39
MusaNews ............................................................................................................. 40
MusaForum ........................................................................................................... 46
INIBAP News.......................................................................................................... 46
Books etc. .............................................................................................................. 51
Announcements.................................................................................................... 52
PROMUSA News............................................................................................. I to XII
The mission of the International Network for the Improvement of Banana and Plantainis to sustainably increase the productivity of banana and plantain grown on smallhold-ings for domestic consumption and for local and export markets.The Programme has four specific objectives:• To organize and coordinate a global research effort on banana and plantain, aimed at
the development, evaluation and dissemination of improved cultivars and at the con-servation and use of Musa diversity
• To promote and strengthen collaboration and partnerships in banana-relatedresearch activities at the national, regional and global levels
• To strengthen the ability of NARS to conduct research and development activities onbananas and plantains
• To coordinate, facilitate and support the production, collection and exchange ofinformation and documentation related to banana and plantain.
INIBAP is a programme of the International Plant Genetic Resources Institute(IPGRI), a Future Harvest center.
INFOMUSAINFOMUSAThe International Magazine on Banana and Plantain
INFOMUSA is published with the support of the Technical Centerfor Agricultural and RuralCooperation (CTA)
CTA
Vol. 10 N° 2December 2001
IN THIS ISSUEIntegrated management ofSigatoka diseases in cv. Africa
Organic fertilization vsinorganic fertilization in “Cachaco” plantain in Colombia
Influence of leaf removal on shoot and root growth in banana
Effect of hail and wind on thefruits of Dominico hartón andFHIA-21
Polyphenol oxidase activityand gene expression in thefruit of Goldfinger
In vitro multiplication of FHIA-18 plantain in thepresence of paclobutrazol
Preliminary results of a virulence test of the populations of Focfor different banana cultivars
Evolution of the nematofaunaof plantain in Côte d’Ivoire
Parasite nematodes of bananain western Burkina Faso
Genetic and genomic diversityin banana based on moleculartools
Banana in images
Classification andcharacterization of Musaexotica, M. alinsanaya and M. acuminata ssp. errans
Thesis
MusaNews
MusaForum
INIBAP News
Books etc.
Announcements
PROMUSA News

L.A. Gómez Balbín and J. Castaño Zapata
The diseases known as yellow(Mycosphaerella musicola Leach)and black (Mycosphaerella fijiensis
Morelet) Sigatoka are plant pathologicalproblems which constrain Musa produc-tion all over the world (Merchán 1996).
Yellow Sigatoka, first reported inColombia in 1937, is the disease with thehighest incidence in the AndeanColombian zone where it is generally notcontrolled (Belalcázar et al. 1991).Although economic losses of more than50% due to yellow Sigatoka are reportedworldwide (Burt et al. 1997), in theColombian coffee zone, the damagingeffects of the disease seem to be compen-sated by the favourable environment forMusa cultivation (Merchán 1996).However, in the central coffee zone, whereplantain Dominico hartón (Musa AAB) iswidely cultivated, yellow Sigatoka diseasecontinues to prevail. Therefore it is impor-tant to consider substituting this variety byothers such as cv. Africa, a selection of theAfrican clone ‘M. bouroukou 1’, Hartóntype, that is well adapted in areas rangingfrom the sea level up to 1700 masl, with abunch that can reach 26 kg and with thefruit quality similar to that of Dominicohartón (Corpoica 2000). Moreover, this cul-tivar possesses excellent qualities withrespect to the central finger on the thirdhand, which has an average weight of510 g, a circumference of 18 cm and exter-nal and internal fruit length of 32 and 26 cm , respectively (Belalcázar et al.1998). The plant cycle is approximately10 months, depending on application offertilizer, which allows one or two func-tional leaves to be maintained until har-vest (Corpoica 2000).
Black Sigatoka is the main disease inbanana and plantain cultivation inCentral America, Panama, Colombia andEcuador, as well as in many other coun-tries. This disease attacks the leaves,resulting in the reduction of the leaf area,thus decreasing their photosyntheticcapacity and affecting the growth andproductivity of the plants. The fruit qual-ity is also reduced due to prematurebunch maturation (Corbana 1996).
The difficulties linked to the manage-ment of both Sigatoka diseases increasewhen the pathogen develops geneticresistance to the main fungicides, par-ticularly to the systemic fungicides(Merchán 1996). Nevertheless, diseasemanagement based only on the use ofchemical products is possible, but at avery high cost, especially for the smalland medium producers who are the mostaffected (Maciel Cordero et al. 1998).Due to this, the possibility of integratingdifferent management practices wasoutlined, taking into account that chem-ical management represents a veryimportant tool in the control of theSigatoka diseases. However, it was recog-nized that the competitive capacity ofthe fungus and its ability to reproduceand persist on the leaf surface could becompensated by optimum managementof nutritional conditions and by farmingpractices that do not favour the infec-tion process of the causal agents.Therefore, farming practices of chemicalor cultural nature, added to the use of aresistant or tolerant variety, representthe fundamental part of an integratedstrategy for the management of theSigatoka diseases.
This study was performed in order todetermine the effect of different agro-nomic practices on the behaviour of theSigatoka diseases in the plantain Africaand to establish an integrated manage-ment system for both diseases.
Materials and methodsThis research was carried out at theMontelindo farm, property of theUniversity of Caldas, located in Palestinadistrict, Santágueda region at 1050 masl,with average temperature of 24ºC, annualrainfall of 2200 mm, relative humidity of86% and 5.3 hours of daily sunshine.Plantain plants were cv. Africa from invitro tissue cultures, and Dominicohartón suckers as a source of inoculumplanted along the edges surroundingeach treatment, at 2 x 3 m distance.
The plants were disinfected for onehour in Mancozeb WP (5 g L-1 of water)solution, and Carbofuran GR (15 g/sitearound each plant) was added. Duringthe production cycle, cultural practiceswere applied including optimum fertiliza-
tion, considering that the bunch qualityand the shelf-life of the fruit are closelyrelated to the organic matter content ofthe soil and to the nutrition of the plant(Arcila et al. 1999). Based on the soilanalysis which indicated potassiumdeficit (0.15 meq/100 g), and consideringthe importance of an adequate nutrientbalance for the proper management ofthe Sigatoka diseases (Pérez 1998), wedetermined an optimum fertilization planwith applications of 1 kg of poultry dung,1 kg of ashes and 200 g dolomitic lime atplanting and three periodical applica-tions of Kumba (15-4-24-2) of 200 g persite every 2 months from planting to flow-ering; after which the same product wasapplied to the follower determined forthe next cycle.
Equally, chemical and mechanicalmethods were integrated using scythesand Glyphosate application at a dose of1.6 L ha-1 for the management of grasses,especially gramineous species such asRottboellia exaltata L. F., Paspalum vir-gatum L. and Panicum maximum Jacq.,at 3, 6 and 9 months after planting.
During the total cropping cycle,36 deleafings were carried out to elimi-nate the necrotic area due to Sigatokadiseases, considering that the systematicpruning (each 7-10 days) of the leaves orfractions of the leaves with maturelesions reduces the inoculum production(Pérez 1998).
As for the chemical management, appli-cations of Propiconazol EC (systemic) ata dose of 0.4 L ha-1 and/or Mancozeb SC(contact) at a dose of 1.75 L ha-1 (Rohm &Haas Colombia S.A. 1998) were madeusing a 18-L sprayer EFCO AT 2080 withlarge range nozzle capacity, depending onthe treatment and according to a biologi-cal warning system developed by Fagesand Jiménez (1995), cited by Merchán(1996). This consists of weekly diseaseassessments on 10 plants selected at thejuvenile stage, from the third month afterplanting, and these plants are replaced atflowering or when they become too tall.The observations are made on the adaxialside and at the end of the fourth leaf,where the presence of the developmentalstages of Sigatoka diseases is recordedand quantified. In the examined area themost advanced stage is established as
INFOMUSA — Vol 10, N° 2 3
Integrated management of black and yellowSigatoka diseases in plantain cv. Africa
Diseases Control of Sigatoka

determined by a coefficient according tothe following scale:
The grade of the dominant symptom isdetermined as a minus (-) when up to 50necrotic lesions per leaf are observed anda plus (+) when there are more than 50lesions. After adding the values which aredetermined on 10 plants, a level of infec-tion on the leaf No. 4 (ILL4) is obtainedfor the respective week. These values areplotted on a graph in order to determinethe date of spraying based on the curvebehaviour. When the ILL4 increases by200 units with respect to the previousweek, fungicides are applied.
Twelve treatments distributed in a ran-domized block design with three replica-tions were applied. The treatments were: 1. Optimum fertilization + Deleafing
(OF+D)2. Optimum fertilization + Contact
chemical (OF +CC)3. Optimum fertilization + Systemic
chemical (OF +SC)4. Deleafing + Contact chemical (D +
CC)5. Deleafing + Systemic chemical (D +
SC)6. Contact chemical + Systemic chemi-
cal (CC + SC)7. Optimum fertilization + Deleafing +
Contact chemical (OF + D + CC)8. Optimum fertilization + Deleafing +
Systemic chemical (OF + D + SC)9. Optimum fertilization + Contact
chemical + Systemic chemical (OF +CC + SC)
10. Deleafing + Contact chemical +Systemic chemical (D + CC + SC)
11. Optimum fertilization + Deleafing +Contact chemical + Systemic chemi-cal (OF + D + CC + SC)
12. Absolute control.The following variables were evaluated:
the youngest diseased leaf (YDL), whichis the youngest leaf with streaks clearlyvisible from the ground (Orjeda 1998);the youngest leaf spotted (YLS), which isthe first open leaf presenting 10 or more
discrete necrotic and mature lesions or abig necrotic area with 10 clear centers,counting the leaves from the top down-wards (Stover and Dickson 1970); and theseverity index (SI), calculated for eachplant in each replicate (Craenen 1998)according to the following formula:
Severity index = ∑nb x 100(N-1) T
Where:n = number of leaves in each grade.b = grade (0 = symptomless, 1 = less
than 1% of leaf area affected (only streaksand/or up to 10 spots), 2 = 1-5% of leafarea affected, 3 = 6-15% of leaf areaaffected, 4 = 16-33% of leaf area affected,5 = 34-50% of leaf area affected,6 = 51-100% of leaf area affected).
N= total number of grades used in thescale (7).
T= total number of leaves assessed.The following parameters were also
measured: number of functional leaves atflowering, i.e. number of leaves with morethan 50% of green area (Orjeda 1998);number of functional leaves at harvest;bunch weight (kg) at harvest; secondhand weight (g); weight (g) of the centralfinger on the second hand; length (cm) ofthe central finger on the second hand;plant height (m) at flowering; and pseu-dostem diameter (cm) at flowering at 1 mfrom the ground level.
Marginal analysis was performed whichallowed to identify the most viable treat-ments from an economical point of view,comparing the investment for each ofthem compared to the control treatment.
Statistical analysis was performed bymeans of the statistical software SAS(Statistical Analysis System for DOS ver-sion 6.0), complementing it with Duncan’smultiple comparison tests at 5%.
Results and discussionThe plant height (PHF) and the pseu-dostem diameter (PDF) at flowering pre-sented significant and highly significantdifferences respectively, allowing to con-clude that this behaviour would be deter-mined by the combined effect of theapplied treatments. The number of func-tional leaves at flowering and harvest(FLF and FLH) presented highly signifi-cant differences according to the treat-ments. The treatment OF + D + CC + SCtreatment resulted in an average of 12.1and 4.8 leaves at flowering and harvest,respectively, compared with the controlwhich presented 8.8 and 2.3 leaves atthese stages (Table 1). Likewise, thetreatments OF+CC and OF+D+CCallowed the flowering stage to be reachedwith 2.9 more leaves than the control.Taking into account that the plantain veg-etative growth stops when the male budemerges (Champion 1968), and that theplant should have a minimum of eightfunctional leaves (Meredith 1970, Stover1972, Martínez 1984), and consideringthat the leaf area available to the plantfrom the flowering stage is a determiningfactor in crop production (Aristizábal andLandinez 1993), the notable effect of thetreatment based on OF+D+CC+SC onthe bunch weight can be appreciated(17.7 kg, compared to 4.1 kg for the con-trol) (Table 2).
The control reached the floweringstage with fewer leaves and with a sever-ity index (SI) similar to the treatmentsD+CC, CC+SC and D+CC+SC (Table 3).Even so it presented values which werenot different statistically than those pro-duced by these treatments for each vari-able of the yield: CFW, CFL, SHW and BW(Table 2). Although the studies per-formed by Grisales (1995) in the same
4 INFOMUSA — Vol 10, N° 2
Table 1. Effect of different agronomic practices for the management of black andyellow Sigatoka diseases on some growth parameters in plantain Africa.
Treatment 1) PHF PDF FLF FLH
OF + D 3.3 2) ab 3) 27.1 bcd 8.9 d 1.5 b
OF + CC 3.6 a 27.9 ab 11.7 ab 2.0 b
OF + SC 3.1 b 27.7 abc 10.4 abcd 1.8 b
D + CC 3.2 b 26.2 d 10.5 abcd 1.5 b
D + SC 3.3 ab 27.3 bcd 10.8 abc 2.1 b
CC + SC 3.5 ab 28.8 a 9.0 cd 1.6 b
OF + D + CC 3.5 ab 28.3 ab 11.7 ab 1.6 b
OF + D + SC 3.1 b 28.1 abc 9.5 cd 2.1 b
OF + CC + SC 3.4 ab 28.6 a 10.2 bcd 2.3 b
D + CC + SC 3.2 ab 26.9 cd 9.5 cd 1.8 b
OF + D + CC + SC 3.5 ab 28.0 abc 12.1 a 4.8 a
Control 3.4 ab 26.9 cd 8.8 d 2.3 b1) OF: Optimum fertilization; D: Deleafing; CC: Contact chemical; SC: Systemic chemical; PHF: Plant height (m) at flowering; PDF:Pseudostem diameter (cm) at flowering; FLF: Number of leaves at flowering; FLH: Number of leaves at harvest. 2) Mean of three replications and five plants per repetition.3) Means within each column accompanied by different letters denote significant statistical differences according to the multiplecomparison range of Duncan at 5% of probability.
Stage Grade Coefficient
1 - 20
+ 40
2 - 60
+ 80
3 - 100
+ 120
4 - 140
+ 160
5 - 180
+ 200
6 - 220
+ 240
zone show that the leaf necrosis causedby Sigatoka diseases is usually severe andintensifies after flowering, the controlsimply displayed its genetic characteris-tics showing a high level of tolerance toSigatoka diseases. The plants reachedfruiting with practically 50% of leaf tissuephotosynthetically active despite a highindex of severity. This phenomenon,according to the physiological studies byGanry (1989), could be explained as aneffect of early accumulation of assimi-lates in the rhizome which could compen-sate for the loss of leaf activity during theperiod of fruit filling. This process isfavoured by the exceptional climatic andsoil conditions which prevailed duringthe experiment, which gave particularadvantage to the control treatment.
During the assessment of the Sigatokadiseases behaviour in terms of severity,the results obtained indicate that alltreatments behave as the control treat-ment, except the treatmentOF+D+CC+SC which was less affected bydisease attack (Table 3). Consideringthat in this treatment an optimum fertil-ization plan has been included, the effectof the nutritional component as a sourceof resistance of the plants to the attacksby Sigatoka diseases in plantain Africa isclear. This theory was presented previ-ously by Huber (1996) and Craenen(1998), suggesting that soil fertility andplantain and banana growth could have adirect influence on the severity of blackSigatoka. In fact, in soils with a properorganic matter content, such as reportedin the soil analysis of this study (4.88%),the proliferation of the root system isstimulated, with the resulting increase ofwater and nutrient uptake. As a result,plant growth is more vigorous with ahigher number of functional leaves beingproduced, which are thus less damagedby diseases. At the same time, theadverse effect of different agronomicpractices, such as OF, D, CC and SC, onthe diseases is observed (Table 3), sinceother treatments with optimum fertiliza-tion presented severity indices higherthan this treatment.
It is important to mention that thetreatments OF+CC and OF+D+CC, alsoincluding the optimum fertilization plan,resulted in the highest values of severityindex (SI) with 58.4% and 55.1%, respec-tively, greater than the control. However,in these cases, the final yield expressedas BW, was 15.5% higher than that of thecontrol, confirming once again the toler-ance of the plantain Africa to Sigatokadiseases.
As shown in Table 3, there were no sig-nificant statistical differences in the vari-
ables YDL and YLS between the treat-ments with respect to the control, whichcould be explained by the high inoculumpressure in the crop that severelyaffected the plants in all treatments.
After performing the analysis of data, itcan be seen that the deleafing practicefollowed by optimum fertilization had thesame effect on the severity of the diseasesthat the combination of this practice withthe application of a contact or a systemicchemical.
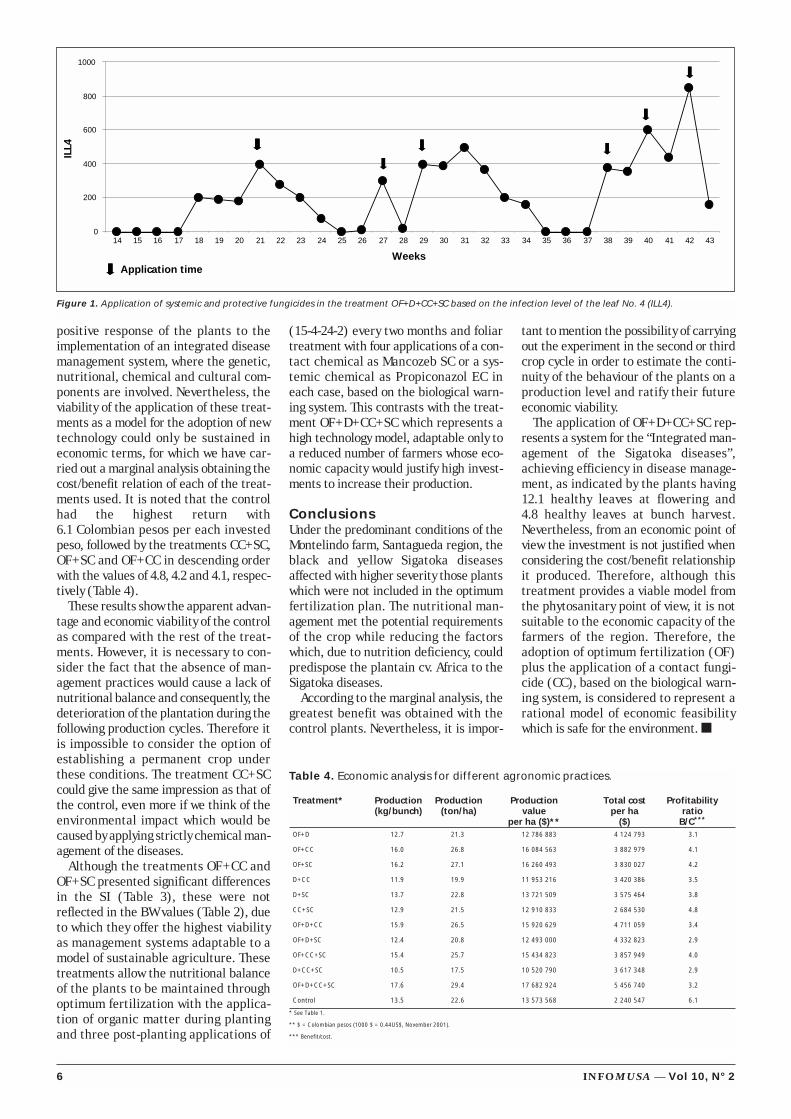
On the other hand, six applications ofthe Mancozeb SC and Propiconazol ECcombination in the treatmentOF+D+CC+SC, based on the biologicalwarning system (Figure 1) stopped signif-icantly the infective process of the fungi,thus confirming the effectiveness of theapplications based on the infection levelof the leaf No. 4. In this system, chemicalproducts are applied particularly whenthe disease severity reaches critical
levels. However, strictly chemical diseasemanagement is considered unfavourablesocioeconomically and environmentallyin developing countries, given the highcontamination levels that can result ifchemicals are applied indiscriminately orin inadequate doses, which can alsoaggravate the possible development ofchemical resistance in the causal agentsof Sigatoka diseases, particularly due tosystemic products (Craenen 1998).
The best results obtained in terms ofyield were recorded with the treatmentsOF+CC, OF+SC and OF+D+CC+SC(Table 2). The first two represent aneffective strategy for the management ofthe Sigatoka diseases, integrating thenutritional component plus the applica-tion of a preventive or curative fungicide,respectively, based on a biological warn-ing system. The third treatment, in whichthe maximum values for each yield para-meter were obtained, would indicate the
INFOMUSA — Vol 10, N° 2 5
Table 2. Effect of different agronomic practices for the Sigatoka diseasesmanagement on some production parameters.
Treatment* CFW CFL SHW BW
OF + D 510.1 1) cde 2) 26.1 abc 3.5 bcd 12.8 de
OF + CC 618.5 ab 27.6 a 4 abc 16.1 ab
OF + SC 562.7 abcd 26.4 abc 4.2 ab 16.3 a
D + CC 458.9 de 24.5 c 2.9 de 12 de
D + SC 516 abcde 25 abc 3.2 de 13.7 bcd
CC + SC 418.5 e 24.5 c 2.9 de 12.9 de
OF + D + CC 535 abcd 27.2 abc 4.4 a 15.9 abc
OF + D + SC 497.5 cde 25.8 abc 3.3 cde 12.5 de
OF + CC + SC 571 abc 27.4 ab 4.1 ab 15.4 abc
D + CC + SC 417.3 e 24.5 c 2.6 e 10.5 e
OF + D + CC + SC 630.3 a 27.4 ab 4.4 a 17.7 a
Control 481.4 cde 24.7 bc 3.1 de 13.6 cd* See Table 1.
CFW: Central finger weight (g) on the second hand; CFL: Central finger length (cm) on the second hand; SHW: Second handweight (kg); BW: Bunch weight (kg). 1) Mean of three replications and five plants per replication.2) Means within each column accompanied by different letters denote significant statistical differences according to the multiplecomparison range of Duncan at 5% of probability.
Table 3. Effect of different agronomic practices for the Sigatoka diseasesmanagement on the behaviour of plantain Africa.
Treatment* YDL YLS SI
OF + D 4 a* 5 a* 45.8 bc**
OF + CC 5 a 5 a 58.4 a
OF + SC 5 a 6 a 46.3 bc
D + CC 4 a 5 a 49.6 abc
D + SC 4 a 5 a 48.7 abc
CC + SC 4 a 5 a 49.3 abc
OF + D + CC 5 a 5 a 55.1 ab
OF + D + SC 4 a 5 a 43.3 c
OF + CC + SC 4 a 5 a 45.0 bc
D + CC + SC 4 a 5 a 47.8 abc
OF + D + CC + SC 5 a 5 a 39.5 c
Control 4 a 5 a 48.9 abc* See Table 1.
YDL: The youngest diseased leaf at flowering; YLS: The youngest leaf spotted at flowering; SI: Severity index (%) at flowering.
** Means within each column accompanied by different letters denote significant statistical differences according to the multiplecomparison range of Duncan at 5% of probability.
positive response of the plants to theimplementation of an integrated diseasemanagement system, where the genetic,nutritional, chemical and cultural com-ponents are involved. Nevertheless, theviability of the application of these treat-ments as a model for the adoption of newtechnology could only be sustained ineconomic terms, for which we have car-ried out a marginal analysis obtaining thecost/benefit relation of each of the treat-ments used. It is noted that the controlhad the highest return with6.1 Colombian pesos per each investedpeso, followed by the treatments CC+SC,OF+SC and OF+CC in descending orderwith the values of 4.8, 4.2 and 4.1, respec-tively (Table 4).
These results show the apparent advan-tage and economic viability of the controlas compared with the rest of the treat-ments. However, it is necessary to con-sider the fact that the absence of man-agement practices would cause a lack ofnutritional balance and consequently, thedeterioration of the plantation during thefollowing production cycles. Therefore itis impossible to consider the option ofestablishing a permanent crop underthese conditions. The treatment CC+SCcould give the same impression as that ofthe control, even more if we think of theenvironmental impact which would becaused by applying strictly chemical man-agement of the diseases.
Although the treatments OF+CC andOF+SC presented significant differencesin the SI (Table 3), these were notreflected in the BW values (Table 2), dueto which they offer the highest viabilityas management systems adaptable to amodel of sustainable agriculture. Thesetreatments allow the nutritional balanceof the plants to be maintained throughoptimum fertilization with the applica-tion of organic matter during plantingand three post-planting applications of
(15-4-24-2) every two months and foliartreatment with four applications of a con-tact chemical as Mancozeb SC or a sys-temic chemical as Propiconazol EC ineach case, based on the biological warn-ing system. This contrasts with the treat-ment OF+D+CC+SC which represents ahigh technology model, adaptable only toa reduced number of farmers whose eco-nomic capacity would justify high invest-ments to increase their production.
ConclusionsUnder the predominant conditions of theMontelindo farm, Santagueda region, theblack and yellow Sigatoka diseasesaffected with higher severity those plantswhich were not included in the optimumfertilization plan. The nutritional man-agement met the potential requirementsof the crop while reducing the factorswhich, due to nutrition deficiency, couldpredispose the plantain cv. Africa to theSigatoka diseases.
According to the marginal analysis, thegreatest benefit was obtained with thecontrol plants. Nevertheless, it is impor-
tant to mention the possibility of carryingout the experiment in the second or thirdcrop cycle in order to estimate the conti-nuity of the behaviour of the plants on aproduction level and ratify their futureeconomic viability.
The application of OF+D+CC+SC rep-resents a system for the “Integrated man-agement of the Sigatoka diseases”,achieving efficiency in disease manage-ment, as indicated by the plants having12.1 healthy leaves at flowering and4.8 healthy leaves at bunch harvest.Nevertheless, from an economic point ofview the investment is not justified whenconsidering the cost/benefit relationshipit produced. Therefore, although thistreatment provides a viable model fromthe phytosanitary point of view, it is notsuitable to the economic capacity of thefarmers of the region. Therefore, theadoption of optimum fertilization (OF)plus the application of a contact fungi-cide (CC), based on the biological warn-ing system, is considered to represent arational model of economic feasibilitywhich is safe for the environment. ■
6 INFOMUSA — Vol 10, N° 2
0
200
400
600
800
1000
14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43
Weeks
ILL4
Application time
Figure 1. Application of systemic and protective fungicides in the treatment OF+D+CC+SC based on the infection level of the leaf No. 4 (ILL4).
Table 4. Economic analysis for different agronomic practices.
Treatment* Production Production Production Total cost Profitability (kg/bunch) (ton/ha) value per ha ratio
per ha ($)** ($) B/C***
OF+D 12.7 21.3 12 786 883 4 124 793 3.1
OF+CC 16.0 26.8 16 084 563 3 882 979 4.1
OF+SC 16.2 27.1 16 260 493 3 830 027 4.2
D+CC 11.9 19.9 11 953 216 3 420 386 3.5
D+SC 13.7 22.8 13 721 509 3 575 464 3.8
CC+SC 12.9 21.5 12 910 833 2 684 530 4.8
OF+D+CC 15.9 26.5 15 920 629 4 711 059 3.4
OF+D+SC 12.4 20.8 12 493 000 4 332 823 2.9
OF+CC+SC 15.4 25.7 15 434 823 3 857 949 4.0
D+CC+SC 10.5 17.5 10 520 790 3 617 348 2.9
OF+D+CC+SC 17.6 29.4 17 682 924 5 456 740 3.2
Control 13.5 22.6 13 573 568 2 240 547 6.1
* See Table 1.
** $ = Colombian pesos (1000 $ = 0.44US$, November 2001).
*** Benefit/cost.

E. Echeverry Navarro
It is considered that the use of organicfertilizer contributes to restoring bio-diversity, biological dynamics and the
soil fertility lost under the pressure ofcontinuous agricultural use.
Large quantities of nutrients are takenup during each crop cycle and the soilbecomes more impoverished than if itremained fallow. A cultivated soil loses alarge proportion of its biodynamicsbecause of the salt residues from inor-ganic fertilizers and the large quantitiesof irrigation water than cause the leach-ing and deep percolation of nutrients andthe leaching of bases, acidifying the soiland making it toxic. All these factors con-tribute to loss of fertility and soil biodi-versity (Orozco 1999).
Organic farming is a system that incor-porates agronomic, economic and socialaspects. In this type of agriculture, pref-erence goes to the use of organic factorsof production (manure or compost) asfertilizer; insect pests and diseases arecontrolled using plant extracts acting asrepellents or bioregulators; natural orinduced biological control is used, as iscrop rotation for protection of the latterand to make better use of soils
The use of organic fertilizers is per-haps the oldest and most common prac-tice in organic farming. Farmers havelong used manure, compost, biologicalfertilizers and agroindustrial wastes assources of organic fertilization (Lapradeand Riz 1999).
Organic matter of plant and animalorigin affects various soil properties andso maintaining its level is one of the key
factors in rational farming. The decom-position of plant and animal residues inthe soil forms a biological system inwhich carbon is returned to the atmos-phere in the form of carbon dioxide andin which nitrogen becomes available inammonia and nitrate form. Elementssuch as phosphorus, sulphur and othernutrients are present in forms that canbe taken up by plants.
It is difficult to achieve a permanentincrease in soil organic content under theusual tropical conditions because of thehigh temperatures and moisture thatcause the rapid oxidation of the plant andanimal residues in the soil. However, it isvery useful for physical, chemical and bio-logical reasons to conserve existingorganic matter by spreading plantresidues or well-decomposed manure orby using green manure.
INFOMUSA — Vol 10, N° 2 7
Organic fertilization vs inorganic fertilization in ‘Cachaco’ plantain in Colombia
Agronomy Organic farming
ReferencesArcila M., F. Aranzazu, C. Castrillon, J. Valencia, M.
Bolaños & P. Castellanos. 1999. Laborales cultur-ales. Pp. 19-20 in El cultivo del plátano.CORPOICA, Manizales, Comité Departamental deCafeteros del Quindio. FUDESCO. Armenia.Colombia.
Aristizabal L.M & C.R. Landinez. 1993. Contribuciónde las hojas al llenado del racimo en plátano(Musa AAB cv. Hartón Enano). RevistaUniversidad de Caldas 13 (1-3): 77-89.
Belalcázar S., J. Valencia & M. Arcila. 1998.Conservación y evaluación de la colección colom-biana de musáceas. Pp. 1-20 in Memorias:Seminario Internacional sobre Producción dePlátano (M.J. Giraldo Cardona, S. BelalcázarCarvajal, D.G. Cayón Salinas & R.G. Botero Isaza,eds). CORPOICA, Armenia, Colombia.
Belalcázar S. 1991. El cultivo del plátano en eltrópico: Manual de asistencia técnica No. 50. (S.L.Belacázar, J.C. Toro & R. Jaramillo, eds). ICA,CIID, Comité de Cafeteros de Colombia, INIBAP.Cali, Colombia. 376pp.
Burt J.A., J. Rutter & H. González. 1997. Short dis-tance wind dispersal of the fungal pathogens caus-ing Sigatoka diseases in banana and plantain.Plant Pathology 40(4): 451-458.
CORBANA. 1996. El combate de la Sigatoka negra.Departamento de Investigaciones, CorporaciónBananera Nacional. Boletín No. 4. CORBANA, SanJosé, Costa Rica.
CORPOICA. 2000. http://www.corpoica.org.coCraenen K. 1998. Technical manual on black
Sigatoka disease of banana and plantain.International Institute of Tropical Agriculture,Ibadan, Nigeria.
Champion J. 1968. El plátano. Pp. 48-70 in
Agricultura Tropical. Blume, Barcelona.Fages O. & F. Jiménez. 1995. El control de la Sigatoka
negra en el cultivo de plátano. Hoja técnica MIPNo. 4. CATIE-Cirad. Turrialba, Costa Rica. 4pp.
Ganry J. 1992. Mejoramiento del cultivo del plátanoen la zona cafetera de Colombia. 1989-1992.Informe final. Santafé de Bogotá (Colombia).Federacafé-ICA-Cirad.
Grisales L.F. 1995. La Sigatoka amarilla y la produc-ción de plátano en la zona cafetera central deColombia. Cenicafé (Colombia) 46(3): 176-178.
Huber D.M. 1996. Manejo de la nutrición para el combate de patógenos de plantas. San José,Costa Rica.
Maciel Cordeiro Z., A. Pires de Matos, S. de Oliveirada Silva. 1998. Black Sigatoka confirmed in Brazil.INFOMUSA 7(1):31.
Martínez G.A. 1984. Determinación del área mínimafoliar en plátano en el trópico húmedo. RevistaICA 19(2): 183-187.
Merchán V. 1996. Prevención y manejo de la Sigatokanegra. ICA seccional Caldas. Boletin informativo.Manizales. 30pp.
Meredith D.S. 1970. Banana leaf spot disease(Sigatoka) caused by Mycosphaerella musicola
Leach. Phytopath. Paper 11. CommonwealthMycological Institute, Kew, UK.
Orjeda G. 1998. Evaluation of Musa germplasm forresistance to Sigatoka diseases and Fusarium wilt.INIBAP Technical Guidelines 3. INIBAP,Montpellier, France.
Pérez L. 1998. Black Sigatoka disease control inbanana and plantain plantations in Cuba. INFO-MUSA 7(1):26-30.
Rohm & Haas Colombia S.A. 1998. Evaluación delfungicida Dithane 60 SC (Mancozeb) bajo un pro-grama de control de Sigatoka negra(Mycosphaerella fijiensis Morelet), en el cultivode banano - zona del Magdalena - Colombia. SantaFe de Bogotá. Rohm and Haas Colombia S.A.
Stover R.H. 1972. Banana, plantain and abaca dis-eases. 1st ed. Commonwealth MycologicalInstitute, Kew, UK. 316pp.
Stover R.H. & J.D. Dickson. 1970. Leaf spot ofbananas caused by Mycosphaerella musicola:methods of measuring spotting prevalence and severity. Tropical Agriculture (Trinidad) 47:289-302.
Luz Adriana Gómez Balbín is studying Agriculturalengineering at the Facultad de CienciasAgropecuarias, Universidad de Caldas and JairoCastaño Zapata is a Professor, Departamento deFilotecnía, Facultad de Ciencias Agropecuarias,Universidad de Caldas, Apartado Aéreo 275,Manizales, Colombia.

The mixing of easily decomposedorganic material with the soil enrichesboth the fauna and flora in the environ-ment and especially the bacterial popula-tion. It thus stimulates the decomposi-tion of the organic material alreadypresent in the soil and that of addedmaterial and is beneficial for plant nutri-tion (Muñoz 1994).
It is not rare in the central southernpart of Tolima department (Colombia),for small and medium scale farmers tocombine crop farming and small-scaleanimal husbandry. A few heads of cattlegraze during the day in the green areasalong the roads, tracks and irrigationcanals. The animals are enclosed in smallpens at night, where manure accumu-lates and is then stored nearby, becominga pollution factor through the breeding ofinsects, and especially dipterans.
Farms are devoted mainly to foodcrops. The most common cultivatedspecies in the zone are maize, cassava,beans and of course ‘Cachaco’ plantain,the staple rural foodstuff in the region.
The bunches of ‘Cachaco’ plantainsharvested are generally very small andtherefore not very suitable for sale on thelocal market. This is why several farmersdecided to apply inorganic fertilizer toincrease bunch size. However, this proce-dure is very expensive for them, if onlybecause of the fertilizer transport costs.
In order to find solutions that areviable from the economic and environ-mental points of view, we developed theexperiments described below to improvethe production and productivity of‘Cachaco’ plantain while reducing pro-duction costs and making appropriate useof the organic matter produced on thefarm. In the process, we tried to solve theproblem of the pollution caused by theaccumulation of manure around the cat-tle pens, which are almost always tooclose to small farmers’ dwellings.
It was also sought to generate andtransfer these techniques within theframework of participative learning bythe communities concerned.
Material and methodsThe experiment lasted for 14 months(1998-1999) at two locations in the cen-tral southern part of Tolima department:
In the Espinal municipal area, at VillaMercedes farm at Aguablanca, the soilcorresponds to the range of soils observedin the Espinal zone, average annual tem-perature is 28°C, altitude 400 m, averageannual precipitation in two rainy seasonsis 1400 mm and relative humidity is 70%.
According to the soil analyses per-formed in the laboratory, the soil is loamy-
clayey, pH 6.0 (slightly acid), low organiccontent (1%), average P (15.9 ppm) andS (6.7 ppm) levels, high Fe (73.0 ppm)level, medium to low Cu (0.60 ppm) con-tents and low Zn (1.4 ppm), Mn (7.1 ppm)and B (0.10 ppm) contents. The Ca con-tent is relatively high (2.7 meq/100 g soil),and the Mg (0.63 meq/100 g soil) andK (0.32 meq/100 g soil) are low. TheCa:Mg ratio is high and the cationexchange capacity (CEC) is low.
In the Suárez municipal area, TierraMía farm at Limonal on the bank of theriver Magdalena in the alluvial plain ofthis river at an altitude of 400 m, averageannual temperature 28°C, averageannual precipitation 1300 to 1700 mm intwo seasons and relative humidity 75%.
Analyses showed that the soil hasloamy-clayey texture, pH 6.9 (practicallyneutral), a low organic content (1.86%),high S (15.7 ppm), P (39.8 ppm), Zn(30 ppm) and Mn (374 ppm) contents, amedium to high B (1.06 ppm) content,medium Fe (15.0 ppm.) and Cu (1.2 ppm)levels, medium to high Ca level(17.8 meq/100 g soil) and high Mg(2.6 meq/100 g soil) and K (0.45 meq/100 gsoil) levels. This soil displays a highCa:Mg ratio and a high cation exchangecapacity.
MaterialThe plantain cultivar known in the regionas ‘Cachaco’ (ABB Simmonds) was used.This clone is most frequently planted andappreciated by consumers in the zone.
The inorganic fertilizer used was ‘triple18’ (N-P-K); urea containing 46% N was
applied as a source of nitrogen and potas-sium chloride containing 60% potassiumas a source of K2O.
The organic fertilizer used was dry,well-decomposed cattle manure (themost abundant type in the zone) col-lected from the pens in which the cattleare stabled for the night.
Treatments
T1: organic fertilization (organic matter)Cattle manure was applied at 2 kg perplant when the suckers were planted.Five further monthly applications of 2 kgeach were subsequently applied, makinga total of 12 kg.
T2: inorganic fertilization (chemical)100 g ‘triple 18’ was applied at plantingand then a further 100 g 30 days after theplanting of suckers in the field.
This was followed by the application of100 g urea and 100 g potassium chlorideto each plant in the form of four monthlyapplications from Day 60 after the plant-ing of the suckers.
T3: 50% organic fertilizer plus 50%inorganic fertilizerApplied on the same dates as in treat-ments T1 and T2.
T4: Control, with no fertilization
MethodsPlanting out was performed in early 1998with the 2.50 x 2.50 m spacing generallyused in the zone. Healthy suckers fromthe region were first cleaned and disin-fected with a mixture of fungicide(Manzate) and insecticide (liquidCarbofuran) sprayed directly on theplants in the planting hole.
Fertilization was performed as in thetreatments above.
The necessary agricultural tasks wereperformed during plant growth, in partic-ular mechanical weed mowing and some-times weed cutting with a ‘machete’. Thesmall secondary suckers were alsoremoved with a special tool, leaving onlythe mother plant and two suckers at mostat each plant position. Deleafing was onlynecessary twice in the trial in Suárezmunicipal area as a phytosanitary mea-sure necessitated by black Sigatokaattacks. No insecticide or fungicide wasapplied to the plantation.
Experimental protocolAn experimental protocol of completelyrandomized blocks was used, with fourtreatments and three repetitions.
8 INFOMUSA — Vol 10, N° 2
Cachaco plantain bunches, without/withinflorescence.
Results and discussionThe data were compiled and processedand combined statistical analysis wasperformed for the two locations, the fourtreatments and the three repetitions, tak-ing development and production parame-ters into account.
Development was evaluated using thefollowing parameters:• plant height in cm,• pseudostem girth 1 m above the
ground, measured in cm,• total number of functional leaves at
harvesting.Three parameters were used for the pro-
duction variable:• total number of hands in the bunches
harvested,• total number of fingers in the bunches
at harvesting,• bunch weight in g at harvesting.
Analysis of variance (ANOVA) is pre-sented in Table 1 for the six measure-ments of the two parameters consideredfor the two locations and the four treat-ments and for the interactions betweenlocations and treatments.
Very significant differences in the threedevelopment parameters and the threeproduction parameters can be observedbetween Espinal and Suárez. The resultsfor Suárez are better, especially for bunchweight at harvesting, which interestsfarmers most. This may be the result ofbetter soil conditions, as can be seen inthe results of the laboratory analyses.
There are no significant differencesbetween treatments with regard to plantheight, pseudostem girth and bunchweight at harvesting.
However, there are significant differ-ences between treatments with regard tothe number of functional leaves at har-vesting and very significant differences innumbers of hands and fingers per bunch,which can be ascribed to the soil condi-tions described above.
No statistically significant differencewas found for interaction between loca-tions and treatments.
The effect of organic fertilization on thecultivation of ‘Cachaco’ is shown in
Table 2, with a comparison of the two loca-tions: Suárez with fertile soils containinggood nutrients and Espinal with highlandplateau soils with less nutrients. Theorganic content was low in both cases.
It was noticed in this experiment withmultiple combinations that the measure-ments concerning the growth parameter -that is to say plant height and pseu-dostem girth - are greater in Suárezwhere the ‘Cachaco’ plants were tallerand thicker than those in Espinal.
The number of functional leaves at har-vesting was greater in Espinal than inSuárez. This is possibly because relativehumidity is higher and the plants arestronger in Suárez. Fungal attacks arefacilitated there, leading to a decrease inthe number of functional leaves, whereasthe risks are smaller in Espinal becausethe atmosphere is drier and plant growthless strong.
The three measurements of productionparameters were higher in Suárez, andespecially bunch weight at harvesting.This may be the result of better soil con-ditions (fertile valley soil) at the plantinglocation.
The effect of organic and inorganic fer-tilization of ‘Cachaco’ plantain in relationto the development and production vari-ables measured during the experiment isanalyzed in Table 3. Statistically signifi-cant differences are observed in the num-bers of hands and fingers. The results oftreatment T4 (no fertilization) are lessgood than those of T1 and T2. Significantdifferences were also found in the num-ber of functional leaves at harvesting; thiswas smaller in treatment T4 than in T3.
No statistical differences wereobserved in bunch weight at harvesting,the most important variable for growers,although the bunches grown with no fer-tilization (T4) weighed up to 1600 g lessthan those to which organic (cattlemanure), inorganic or mixed (inorganicfertilizer + manure) fertilizer had beenapplied.
Economic analysisThe costs of organic and inorganic fertil-izer (T1 versus T2) are compared inTable 4. It can be noted that organic fer-tilization costs Colombian pesos (COP)160 000 (approximately US$ 80), i.e. 33%of the cost of chemical fertilization ofCOP 480 000 (US$ 240). Production costsare thus clearly lower with organic fertil-ization.
Allowing for the fact that in terms ofbunch weight there are no statistical dif-ferences between production usingchemical fertilization or organic fertiliza-tion, it is clear that it is better to useorganic fertilization that is cheaper andalso contributes to improving the physi-cal properties of the soil and conservesfertility better in the long term.
It is considered that one day’s labour isrequired to gather and transport a tonne ofmanure, given the short distances betweencattle pens and plantations. The dailywage is currently COP 10 000 (= US$5).
ConclusionsIt is recommended from the economic,ecological and production point of viewthat decomposed cattle manure recov-ered from pens should be used to fertilize
INFOMUSA — Vol 10, N° 2 9
Table 1. Analysis of variance of the development and production variables in ‘Cachaco’ plantain at two locations in Tolimadepartment. Espinal, 1999.
Development Location F2 Treatment Location by treatment
and production variables DF1 Square mean Pr > F2 DF Square mean Pr > F DF Square mean Pr > F
Plant height 1 349619.620 0.0001** 3 1050.843 0.2177 NS 3 536.399 0.4819 NS
Pseudostem girth 1 2618.770 0.0001** 3 27.112 0.2116 NS 3 12.356 05206 NS
Functional leaves at harvesting 1 4.950 0.0107* 3 2.393 0.0261 * 3 1.718 0.0639 NS
Hands per bunch 1 5.802 0.0003** 3 1.466 0.0085** 3 0.113 0.7017 NS
Fingers per bunch 1 3639.575 0.0001** 3 94.375 0.0090** 3 54.822 0.0473 NS
Bunch weight 1 764310493.500 0.0001** 3 3597893.220 0.0684 NS 3 1372386.280 0.3603 NS1 DF = degree of freedom; 2 Pr>F =% of probability; NS = Not significant; * = Significant; ** = Highly significant
Table 2. The effect of organic and mineral fertilization on the cultivation of‘Cachaco’ plantain in two locations in Tolima department. Espinal, 1999.
Location Variables
Plant Pseudostem Leaves Hands Fingers Bunch height girth at per per weight (cm) (cm) harvesting bunch bunch (kg)
Suárez 443.72 A 57.14 A 6.25 B 5.36 A 65.30 A 19.588 A
Espinal 202.33 B 36.25 B 7.15 A 4.37 B 40.67 B 8.302 BValues with different letters in the same column are statistically different (Tukey P ≤ 0.05).
crops of ‘Cachaco’ plantain. It also helpsto prevent this manure from being a fac-tor in pollution.
When cattle manure of this type ofsource is used for fertilization, pollutionand fertilization costs decrease withoutreducing bunch production. This experi-ment shows that organic fertilization isas effective for the productivity of‘Cachaco’ plantain as inorganic fertiliza-tion, which is more expensive
Cost comparison showed that the costof organic fertilization was a third (33%)of that of inorganic fertilization. It wasobserved that bunch weight was equiva-lent in both cases. The lower cost oforganic fertilization thus results ingreater crop profitability.
AcknowledgementsThe author thanks Antonio MaríaCaicedo (MS), agricultural engineer, ofCORPOICA C.I.N. Nataima, for valuablehelp in the statistical analysis of the workpresented. ■
ReferencesBelalcázar S. (ed.). 1991. El cultivo del plátano en el
trópico. Instituto Colombiano Agropecuario ICA.Armenia, Quindío, Colombia. 491pp.
Castro F.H. 1996. Bases técnicas para elconocimiento y manejo de los suelos del vallecálido del Alto Magdalena. CORPOICA Regional 6: 25-45.
Echeverry N.E. 1996. Costos de producción en el cul-tivo del plátano en el CRECED Centro Tolima.Recopilación de información UMATA, ICARegional 6. Espinal. 8pp.
Laprade C.S. & R. Ruiz B. 1999. Productive behaviourof FHIA-01 (AAAB) and FHIA-02 (AAAB), underinorganic and organic fertilization programmes.Pp. 171-176 in Organic/environmentally friendlybanana production. Proceedings of a workshopheld at EARTH, Guácimo, Costa Rica, 27-29 July1998 (F.E. Rosales, S.C. Tripon & J. Cerna, eds).INIBAP, Montpellier, France.
Muñoz A.R. 1994. Los abonos orgánicos y su uso en laagricultura. Pp. 293-304 in Fertilidad de suelos,diagnóstico y control (M.F. Silva, ed.). SociedadColombiana de la ciencia del suelo. Santafé deBogotá. Colombia.
Orozco R. J. 1999. Organic fertilizers and their appli-cation in banana cropping. Pp. 79-85 in
Organic/environmentally friendly banana produc-tion. Proceedings of a workshop held at EARTH,Guácimo, Costa Rica, 27-29 July 1998 (F.E.Rosales, S.C. Tripon and J. Cerna, eds). INIBAP,Montpellier, France.
Steel R. & Torrie. 1988. Bioestadística: Principios yprocedimientos. 2nd ed. (1rst ed. in Spanish).McGraw-Hill, México. 622pp.
The author works at CORPOICA, C.I. Nataima,Apartado Postal 064, Espinal (Tolima) Colombia.
10 INFOMUSA — Vol 10, N° 2
Table 3. The effect of organic and inorganic fertilization on the cultivation of ‘Cachaco’ plantain at two locations in Tolimadepartment. Espinal, 1999.
Treatment Variables
Plant Pseudostem Leaves Hands Fingers Bunch height girth at per per weight (cm) (cm) harvesting bunch bunch (kg)
T1 organic fertilization 330.55 A 48.12 A 7.08 AB 5.01 A 56.28 A 14.309 A
T2 inorganic fertilization 320.47 A 45.85 A 6.55 AB 5.17 A 55.51 A 14.313 A
T3 mixed fertilization 335.55 A 48.72 A 7.30 A 5.16 A 52.63 AB 14.375 A
T4 control 305.55 A 44.10 A 5.88 B 4.13 B 47.53 B 12.785 AValues with different letters in the same column are statistically different (Tukey P ≤ 0.05).
Table 4. Comparison of the cost of chemical and organic fertilization of ‘Cachaco’plantain (Musa ABB) in central Tolima. Espinal, 1999.
Type of Source of Dose Dose Cost of Cost of fertilization inorganic per plant in kg/ha for fertilizer in fertilizer in
fertilizer in grams 1333 plants US$ per kg COP per ha
Inorganic Triple 18 200 266.6 520 168 632
Urea 46 400 533.2 340 181 288
KCl 400 533.2 320 170 624
Sub-total 480544
Organic Cattle manure 12 000 16 000 10 160000* Costs in 1999 were: Triple 18 compound fertilizer COP 26 000/50 kg bag (=$US13)
Urea, 46% N COP 17 000/50 kg bag (=$US8.50)
KCl, 60% K2O COP 16 000/50 kg bag (=$US8)
Daily wage in the zone COP 10 000 (=$US5)
Influence of leaf removal on shoot and root growthin banana (Musa spp.)
Physiology Crop development
G. Blomme, A. Tenkouano and R. Swennen
Plant size and bunch weight ofMusa spp. directly depends on thenumber and size of functional
leaves (Martinez Garnica 1984). Thus,leaf area can be used to estimate thephotosynthetic capacity and to predictthe performance of a banana crop(Turner 1980, Swennen and De Langhe1985, Stover and Simmonds 1987). Leafarea reduction by pests or diseases, e.g.
black Sigatoka (Mycosphaerella fijiensisMorelet), reduces the yield (Jaramillo1987, Stover and Simmonds 1987, Jegeret al. 1995). According to Robinson(1996), pruning of banana leaves is usu-ally carried out for two main reasons.Firstly, the spread of the leaf spot dis-

ease, black Sigatoka, can be reduced byexcising leaves with more than 50%necrotic area. Secondly, one or two younghealthy leaves which are rubbing andscarring fingers on the developing bunchmay be sacrificed to improve the fruitquality, if bunch covers are not used.
Several studies have been conductedto estimate the smallest number ofleaves needed for maximum yield.Satyanarayana (1986) concluded that nofewer than 12 leaves were required dur-ing the vegetative stage to achieve maxi-mum yields in the dessert banana ‘DwarfCavendish’. However, Robinson et al.(1992) found that maximum yield andfinger length could be achieved in thedessert banana ‘Williams’ with eightleaves retained at flower emergence, dueto a compensatory increase in CO2uptake by up to 35% on the prunedplants. Martinez Garnica (1984) alsofound that at least eight leaves at flower-ing were required to ensure normal plantdevelopment of plantains.
The relationships between root andshoot growth have been described forseveral crops (Pearsall 1927, Brouwer1966, Broschat 1998, Fort and Shaw1998). Marler and Stushnoff (1999)reported a significant and negativeeffect of defoliation of papaya plants onroot densities. For example, fine rootscompletely disappeared one week post-defoliation. Brown and Don Scott (1984)reported that changes in soybean shootgrowth also affect root growth and vice-versa. For example, excessive tempera-ture and evaporative demand or inade-quate light reduce photosyntheticactivity, which not only slows shootgrowth but also limits photosynthatetranslocation to the roots, therebyreducing root growth. The decline in theroot growth subsequently decreases theflow of water and nutrients to the shoot.Thus a stress originating in the canopy istransmitted at varying degrees of sever-ity throughout the entire plant (Brownand Don Scott 1984). Hodgkinson andBecking (1977) reported for differentarid zone plants that defoliation or shootremoval caused a decrease in rootgrowth and branching and an increasein root mortality.
Like in other crop species, there is astrong relationship between root andshoot growth in Musa spp. (Swennen1984, Blomme and Ortiz 1996). However,the influence of leaf removal on root sys-tem development of Musa spp. has notbeen investigated. The objective of thisstudy was to assess the influence of areduced leaf area on the root system sizeof banana (Musa spp.) plants.
Materials and methodsThis study was carried out at the IITAHigh Rainfall station at Onne (4º42’N,7º10’E, 5 masl) in southeastern Nigeriawhich represents the humid forest eco-logical zone. The soil is an ultisol derivedfrom coastal sediments, well drained butpoor in nutrients and with a pH of 4.3 in1:1 H2O. The average annual rainfall is2400 mm distributed monomodally fromFebruary until November. The genotypesevaluated were ‘Valery’ (Musa AAA,Cavendish sub-group) and ‘Obino l’Ewai’(Musa AAB, medium French plantain).In vitro-derived plants were used asplanting material. Planting was done on16 June 1998 and evaluation was done at33 weeks after planting (WAP), duringthe pre-flowering stage, in earlyFebruary 1999. Leaf removal started at15 WAP. The field layout was a factorialrandomized complete block design withtwo replications. There were three leafpruning treatments: control (no prun-ing), mild pruning (seven leaves left),and severe pruning (four leaves left).There were four plants for each genotypeand treatment. Plant spacing was2 m x 2 m. Only the biggest sucker wasallowed to grow. The field was irrigatedduring the dry season at a rate of 100 mmper month. The experimental area wastreated with the nematicide Nemacur(a.i. fenamiphos) at a rate of 15 g plant-1
(3 treatments year-1). Fertilization wasdone with muriate of potassium (60% K)at a rate of 600 g plant-1 year-1, and urea(47% N) at a rate of 300 g plant-1 year-1,split over 6 equal applications during therainy season, i.e. March until November.No mulch was applied.
Root system and shoot characteristicswere measured for the plant crop and thesucker. The height of the plant (PH, cm),circumference of the pseudostem at soillevel (PC, cm), number of leaves (NL),number of leaves produced from plantinguntil assessment (TNL) and leaf area(LA, cm2) were measured. Leaf lengthand leaf widest width were measured andLA was calculated according to Obiefunaand Ndubizu (1979). In addition, lengthof the tallest sucker (LS, cm) was mea-sured. The root system of the motherplant and the suckers was completely dugout. The following root characteristicswere assessed for the mother plant: num-ber of adventitious roots or cord roots(NR), average diameter at the base of thecord roots (AD, mm), cord root length(LR, cm) and root dry weight (DR, g). Inaddition, root characteristics measuredfor the whole mat (i.e. mother plant andsuckers) included: total root dry weight(TD, g) and total cord root length
(TL, cm). The average diameter of thecord roots was measured with a VernierCalliper, while the cord root length wasmeasured using the line intersect methodaccording to Newman (1966) andTennant (1975). The line intersectmethod consists of spreading the cordroots on a grid, where after the interac-tion points root-gridline are counted. Thenumber of interaction points were thenmultiplied by the length conversion fac-tor 2.3571 appropriate to the used gridsize of 3 cm by 3 cm.
Significant differences of the growthcharacteristics were established usingthe least significant difference method(LSD) of PROC GLM by SAS (SAS 1989).
Results and discussionIncreased leaf pruning resulted in a sig-nificant reduction of most growth para-meters of the plant crop, except for thenumber of cord roots (Table 1). This indi-cates that leaf area influences the overallplant growth and confirms observationsmade by Brown and Don Scott (1984). Nosignificant differences were observed forthe total number of leaves produced, indi-cating that the leaf production rate wasnot affected by leaf removal.
The leaf area reduction in ‘Obinol’Ewai’ was 25 and 61% for mild andsevere pruning, respectively (Table 1).However, the percentage reduction inpseudostem growth, corm and root devel-opment characteristics was less andranged, for example for the plant crop,from 4 to 21% and 17 to 54% for mild andsevere pruning, respectively (Table 1).This points to a compensatory increase inphotosynthetic efficiency in the remain-ing leaves as shown by Robinson et al.(1992). Similarly, Myers and Ferree(1983) reported that defoliation of youngapple trees caused an increased photo-synthetic rate in the remaining leaves.
However, the percentage reduction inroot dry weight and cord root length in‘Valery’ was higher than the percentagereduction in leaf area under mild prun-ing, while the percentage reduction inleaf area and root dry weight was similarunder severe pruning (Table 1).
Sucker height in ‘Obino l’Ewai’ wasreduced by 33% with mild pruning and81% with severe pruning (Table 2). Incontrast, pruning did not significantlyaffect sucker growth in ‘Valery’ (Table 2).This could be explained by the differencein suckering behaviour between the twogenotypes. Eckstein et al. (1995) andShanmugavelu et al. (1992) reported ahigher independence of the maiden typesuckers, i.e. large suckers with foliageleaves and a large root system, compared
INFOMUSA — Vol 10, N° 2 11
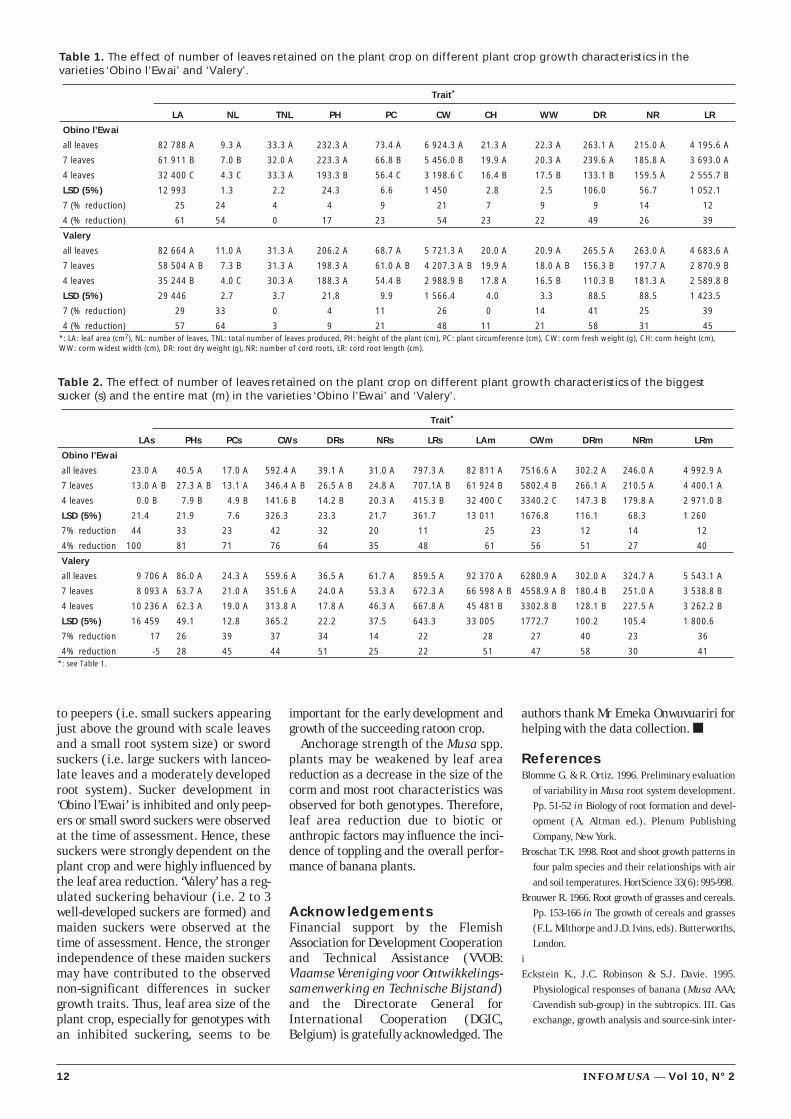
to peepers (i.e. small suckers appearingjust above the ground with scale leavesand a small root system size) or swordsuckers (i.e. large suckers with lanceo-late leaves and a moderately developedroot system). Sucker development in‘Obino l’Ewai’ is inhibited and only peep-ers or small sword suckers were observedat the time of assessment. Hence, thesesuckers were strongly dependent on theplant crop and were highly influenced bythe leaf area reduction. ‘Valery’ has a reg-ulated suckering behaviour (i.e. 2 to 3well-developed suckers are formed) andmaiden suckers were observed at thetime of assessment. Hence, the strongerindependence of these maiden suckersmay have contributed to the observednon-significant differences in suckergrowth traits. Thus, leaf area size of theplant crop, especially for genotypes withan inhibited suckering, seems to be
important for the early development andgrowth of the succeeding ratoon crop.
Anchorage strength of the Musa spp.plants may be weakened by leaf areareduction as a decrease in the size of thecorm and most root characteristics wasobserved for both genotypes. Therefore,leaf area reduction due to biotic oranthropic factors may influence the inci-dence of toppling and the overall perfor-mance of banana plants.
AcknowledgementsFinancial support by the FlemishAssociation for Development Cooperationand Technical Assistance (VVOB:Vlaamse Vereniging voor Ontwikkelings-samenwerking en Technische Bijstand)and the Directorate General forInternational Cooperation (DGIC,Belgium) is gratefully acknowledged. The
authors thank Mr Emeka Onwuvuariri forhelping with the data collection. ■
ReferencesBlomme G. & R. Ortiz. 1996. Preliminary evaluation
of variability in Musa root system development.Pp. 51-52 in Biology of root formation and devel-opment (A. Altman ed.). Plenum PublishingCompany, New York.
Broschat T.K. 1998. Root and shoot growth patterns infour palm species and their relationships with airand soil temperatures. HortScience 33(6): 995-998.
Brouwer R. 1966. Root growth of grasses and cereals.Pp. 153-166 in The growth of cereals and grasses(F.L. Milthorpe and J.D. Ivins, eds). Butterworths,London.
iEckstein K., J.C. Robinson & S.J. Davie. 1995.
Physiological responses of banana (Musa AAA;Cavendish sub-group) in the subtropics. III. Gasexchange, growth analysis and source-sink inter-
12 INFOMUSA — Vol 10, N° 2
Table 1. The effect of number of leaves retained on the plant crop on different plant crop growth characteristics in thevarieties ‘Obino l’Ewai’ and ‘Valery’.
Trait*
LA NL TNL PH PC CW CH WW DR NR LR
Obino l’Ewai
all leaves 82 788 A 9.3 A 33.3 A 232.3 A 73.4 A 6 924.3 A 21.3 A 22.3 A 263.1 A 215.0 A 4 195.6 A
7 leaves 61 911 B 7.0 B 32.0 A 223.3 A 66.8 B 5 456.0 B 19.9 A 20.3 A 239.6 A 185.8 A 3 693.0 A
4 leaves 32 400 C 4.3 C 33.3 A 193.3 B 56.4 C 3 198.6 C 16.4 B 17.5 B 133.1 B 159.5 A 2 555.7 B
LSD (5%) 12 993 1.3 2.2 24.3 6.6 1 450 2.8 2.5 106.0 56.7 1 052.1
7 (% reduction) 25 24 4 4 9 21 7 9 9 14 12
4 (% reduction) 61 54 0 17 23 54 23 22 49 26 39
Valery
all leaves 82 664 A 11.0 A 31.3 A 206.2 A 68.7 A 5 721.3 A 20.0 A 20.9 A 265.5 A 263.0 A 4 683.6 A
7 leaves 58 504 A B 7.3 B 31.3 A 198.3 A 61.0 A B 4 207.3 A B 19.9 A 18.0 A B 156.3 B 197.7 A 2 870.9 B
4 leaves 35 244 B 4.0 C 30.3 A 188.3 A 54.4 B 2 988.9 B 17.8 A 16.5 B 110.3 B 181.3 A 2 589.8 B
LSD (5%) 29 446 2.7 3.7 21.8 9.9 1 566.4 4.0 3.3 88.5 88.5 1 423.5
7 (% reduction) 29 33 0 4 11 26 0 14 41 25 39
4 (% reduction) 57 64 3 9 21 48 11 21 58 31 45*: LA: leaf area (cm2), NL: number of leaves, TNL: total number of leaves produced, PH: height of the plant (cm), PC: plant circumference (cm), CW: corm fresh weight (g), CH: corm height (cm),WW: corm widest width (cm), DR: root dry weight (g), NR: number of cord roots, LR: cord root length (cm).
Table 2. The effect of number of leaves retained on the plant crop on different plant growth characteristics of the biggestsucker (s) and the entire mat (m) in the varieties ‘Obino l’Ewai’ and ‘Valery’.
Trait*
LAs PHs PCs CWs DRs NRs LRs LAm CWm DRm NRm LRm
Obino l’Ewai
all leaves 23.0 A 40.5 A 17.0 A 592.4 A 39.1 A 31.0 A 797.3 A 82 811 A 7516.6 A 302.2 A 246.0 A 4 992.9 A
7 leaves 13.0 A B 27.3 A B 13.1 A 346.4 A B 26.5 A B 24.8 A 707.1A B 61 924 B 5802.4 B 266.1 A 210.5 A 4 400.1 A
4 leaves 0.0 B 7.9 B 4.9 B 141.6 B 14.2 B 20.3 A 415.3 B 32 400 C 3340.2 C 147.3 B 179.8 A 2 971.0 B
LSD (5%) 21.4 21.9 7.6 326.3 23.3 21.7 361.7 13 011 1676.8 116.1 68.3 1 260
7% reduction 44 33 23 42 32 20 11 25 23 12 14 12
4% reduction 100 81 71 76 64 35 48 61 56 51 27 40
Valery
all leaves 9 706 A 86.0 A 24.3 A 559.6 A 36.5 A 61.7 A 859.5 A 92 370 A 6280.9 A 302.0 A 324.7 A 5 543.1 A
7 leaves 8 093 A 63.7 A 21.0 A 351.6 A 24.0 A 53.3 A 672.3 A 66 598 A B 4558.9 A B 180.4 B 251.0 A 3 538.8 B
4 leaves 10 236 A 62.3 A 19.0 A 313.8 A 17.8 A 46.3 A 667.8 A 45 481 B 3302.8 B 128.1 B 227.5 A 3 262.2 B
LSD (5%) 16 459 49.1 12.8 365.2 22.2 37.5 643.3 33 005 1772.7 100.2 105.4 1 800.6
7% reduction 17 26 39 37 34 14 22 28 27 40 23 36
4% reduction -5 28 45 44 51 25 22 51 47 58 30 41*: see Table 1.

action over a complete crop cycle. J. Hortic. Sci.70(1):169-180.
Fort S.B. & D.V. Shaw. 1998. Phenotypic correlationsbetween root and shoot traits of strawberry infumigated and non-fumigated soils. HortScience33: 222-224.
Hodgkinson K.C. & H.G.B. Becking. 1977. Effect ofdefoliation on root growth of some arid zoneperennial plants. Aust. J. Agric. Res. 29: 31-42.
Jaramillo R. 1987. Banana and plantain productionin Latin America and the Caribbean. Pp. 39-43 in Banana and plantain breeding strategies (G.J. Persley and E.A. De Langhe, eds). ACIARProceedings No. 21. ACIAR, Canberra.
Jeger M.J., S. Eden-Green, J.M. Thresh, A. Johanson,J.M. Waller & A.E. Brown. 1995. Banana diseases.Pp. 317-381 in Bananas and plantains (S. Gowen,ed.). World crop series. Chapman & Hall. London.
Marler T.E. & C. Stushnoff. 1999. Root, stem, andfruit growth of ‘Tainung 1’ papaya plants followingdefoliation (abstract). HortScience 34(3): 488-489
Martinez Garnica A. 1984. Effecto del número dehojas sobre la producción de plátano en el trópicohúmedo Colombiano. Revista ICA 19: 357-359.
Myers S.C. & D.C. Ferree. 1983. Influence of summerpruning and tree orientation on net photosynthe-sis, transpiration, shoot growth, and dry weight
distribution in young apple trees. J. Am. Soc.Hortic. Sci. 108: 4-9.
Newman E.I. 1966. A method of estimating the totallength of root in a sample. J. appl. Ecol. 3: 139-145.
Obiefuna J.C. & T.O.C. Ndubizu. 1979. Estimating leafarea of plantain. Sci. Hortic. 11: 31-36.
Pearsall W.H. 1927. Growth studies. VI. On the rela-tive size of plant organs. Ann. Bot. 41: 449-556.
Robinson J.C. 1996. Bananas and Plantains. CABInternational. Wallingford, Oxon, UK. 238 pp.
Robinson J.C., T. Anderson & K. Eckstein. 1992. Theinfluence of functional leaf removal at floweremergence on components of yield and photosyn-thetic compensation in banana. J. Hortic. Sci.67(3): 403-410.
SAS Institute Inc. 1989. SAS/STAT user’s guide, ver-sion 6, 4th edition, volume 1. Cary, N.C.: SASInstitute Inc.
Satyanarayana M. 1986. Effect of number of func-tional leaves on growth and yield of ‘DwarfCavendish’ banana (AAA). Newsletter Int. GroupHortic. Physiol. Banana 9: 34-36.
Shanmugavelu K.G., K. Aravindakshan & S. Sathiamoorthy. 1992. Banana Taxonomy,Breeding and Production Technology. 1st ed.,Metropolitan Book Co. Pvt. Ltd. New Delhi-110002, India. pp. 266-293.
Stover R.H. & N.W. Simmonds. 1987. Bananas, 3rd ed.Longman, UK. 468pp.
Swennen R. 1984. A physiological study of the suck-ering behaviour in plantain (Musa cv. AAB). Ph.D.thesis, Dissertationes de Agricultura n°132,Faculty of Agriculture, Katholieke UniversiteitLeuven, 180pp.
Swennen R. & E.A. De Langhe. 1985. Growth parame-ters of yield of plantain (Musa cv. AAB). Ann. Bot.56: 197-204.
Tennant D. 1975. A test of a modified line intersectmethod of estimating root length. J. Ecol. 63: 995-1001.
Turner D.W. 1980. Some factors related to yield com-ponents of bananas in relation to sampling toassess nutrient status. Fruits 35: 19-23.
Guy Blomme* and Abdou Tenkouano work at theCrop Improvement Division, International Institute ofTropical Agriculture (IITA), c/o L.W. Lambourn & Co.,Carolyn House, 26 Dingwall Road, Croydon CR9 3EE,England and Rony Swennen at the Laboratory ofTropical Crop Improvement, Catholic University ofLeuven (K.U.Leuven), Kasteelpark Arenberg 13, 3001Heverlee, Belgium.
* Presently working as Associate Scientist at the INIBAPOffice for eastern and southern Africa
INFOMUSA — Vol 10, N° 2 13
The effect of hail and wind on the development and quality of the fruits of Dominico hartón and FHIA-21 plantain
Fruit physiology Climatic effects
C.R. Parra, L.M. Lara Sarmiento, G. Cayón Salinas
and G. Giraldo Giraldo
Growth, development and agricul-tural production are the result ofclose interactions between the cli-
matic factors in the production zone andthe genotype of the species cultivated.However, the production cycle of theplants is affected if, during certain pre-cise stages of cultivation, one of these fac-tors reaches proportions exceeding theirtolerance (Cayón et al. 1998a). Morespecifically, modification of the aerialparts of the plants by leaf removal oruncontrollable physical damage affectsthe usual conditions of the plant and dis-turbs leaf area growth and fruiting(Decoteau 1990).
The central coffee zone of Colombia isoften exposed to high winds accompa-nied by heavy rainfall and hail, causing
serious damage to crops. One such cli-matic event on 20 March 1998 causedlosses of more than US$ 6 million forplantain growers in the Quindío adminis-trative department. One of the most com-mon types of damage in all plantain pro-ducing regions is the tearing of leaflaminae by wind. However, if this doesnot cause abscission and a loss of activeleaf area, the plant vegetative and pro-duction cycle is satisfactory.
Observations of coffee crops after agale concern the most visible effects ofthe impact of hailstones, with woundingand bark stripping of branches, theremoval, perforation and tearing of leaflaminae, laceration and wounding ofberries, which turn an almost black darkgreen colour. This type of damageappears to be all the more serious whenthe coffee berries are soft (Valencia andArcila 1976).
The aim of this study is the evaluationof the impact of a hailstorm with strong
wind on Dominico hartón and FHIA-21hybrid plants at three stages of develop-ment and the gauging of the conse-quences for the physicochemical proper-ties of plantain.
Material and methodsThe study was performed at the El
Agrado research centre in theMontenegro municipal area (Quindío) at4°28’N and 75°49’W, an altitude of1320 m, average annual temperature21°C, relative humidity 80% and averageannual precipitation 1985 mm. Theseagroecological conditions are those ofvery humid tropical forest. The soil of theexperimental plot was loamy-sandy withpH 6.2 and contained 2.9% organic mat-ter. A very local storm occurred on 20March 1998 in the Montenegro municipalarea in which the El Agrado research sta-tion is located, with heavy rain (34 mm),strong wind (>100 km.h-1) and hail, with-out affecting neighbouring areas.

Three days later, all the plots of dam-aged Dominico hartón and FHIA-21 wereinspected to select plants that had suf-fered the gale at different stages in theirproduction cycle. Plants at the followingstages were identified and marked: 1) starting of flowering; 2) 15th day afterflowering (15 DAF) and 3) between 30 and 45 DAF. The control plants (notaffected by the phenomenon) wereselected and marked in plots close to theexperimental centre that had not suf-fered strong wind or hail. A completelyrandomized design was used with fourtreatments (stages of development),three repetitions and six plants per rep-etition.
The weight, number of hands and fin-gers of the bunches harvested when thefruits were physiologically ripe (120 DAF)were recorded. Two fruits were takenfrom each of hands 1, 3 and 5 and theweight, external length and circumfer-ence were noted. Fruits with necroticspots on peel and pulp were observed oneach bunch. The spots were excised witha scalpel and weighed to calculate thepercentage of necrotic pulp and peel perfruit. The fruits were peeled to measurepulp and peel weight. The texture of freshpulp was measured with a penetrometer,the pH was measured by titration and sol-uble solids were determined by refrac-tometry. Several fruits were then slicedcrosswise and placed in a sodiummetabisulphite solution 0.5% to preventenzymatic browning. Once dried in a fanoven at 80°C for 24 hours, these sampleswere reduced to powder and the mois-ture, mineral content (ash), fibre, starch(enzymatic hydrolysis) and total sugarswere analyzed (Antrone method). Thedata obtained were compared usingTukey’s analysis of variance method(P<0.05) using the MSTAT-C programme(Michigan State University).
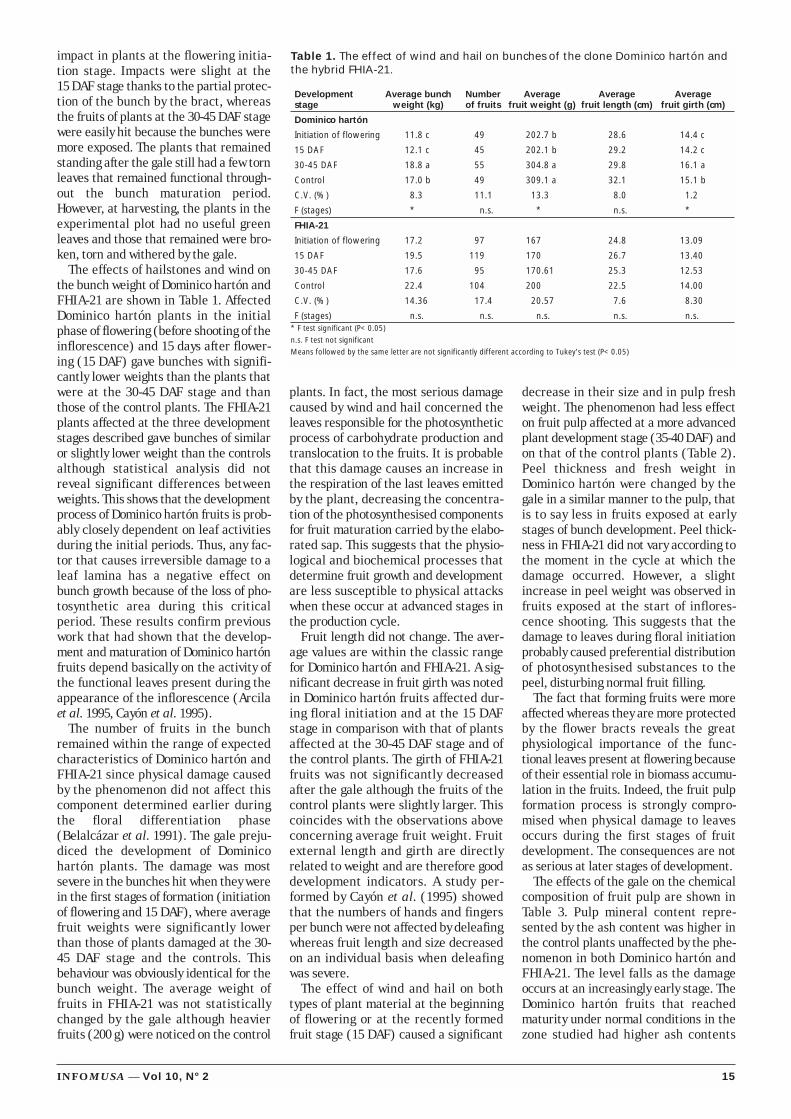
Results and discussionThe strong wind caused the toppling of
many plants by fissuring of the pseu-dostem and uprooting. It also caused thetearing and removal of whole leaf laminasections (Figure 1A). Hailstones hit thepseudostem, leaves, inflorescences andbunches hard (Figure 1B) causing dam-age that was initially in the form ofcreased brown chlorotic points that soonturned dark brown or black (Figure 1C).Peel necrotic areas spread to the fruits,thus affecting their quality.
The direct effect of hailstorm impacton the fruits of bunches in course of for-mation was different according to thedevelopment stage. The position of theinflorescence prevented hailstorm
14 INFOMUSA — Vol 10, N° 2
Figure 1. Damage to the plantation caused by wind (A) and by the impact of hailstones to bunches,pseudostems (B) and leaves (C).
A
B
C
impact in plants at the flowering initia-tion stage. Impacts were slight at the15 DAF stage thanks to the partial protec-tion of the bunch by the bract, whereasthe fruits of plants at the 30-45 DAF stagewere easily hit because the bunches weremore exposed. The plants that remainedstanding after the gale still had a few tornleaves that remained functional through-out the bunch maturation period.However, at harvesting, the plants in theexperimental plot had no useful greenleaves and those that remained were bro-ken, torn and withered by the gale.
The effects of hailstones and wind onthe bunch weight of Dominico hartón andFHIA-21 are shown in Table 1. AffectedDominico hartón plants in the initialphase of flowering (before shooting of theinflorescence) and 15 days after flower-ing (15 DAF) gave bunches with signifi-cantly lower weights than the plants thatwere at the 30-45 DAF stage and thanthose of the control plants. The FHIA-21plants affected at the three developmentstages described gave bunches of similaror slightly lower weight than the controlsalthough statistical analysis did notreveal significant differences betweenweights. This shows that the developmentprocess of Dominico hartón fruits is prob-ably closely dependent on leaf activitiesduring the initial periods. Thus, any fac-tor that causes irreversible damage to aleaf lamina has a negative effect onbunch growth because of the loss of pho-tosynthetic area during this criticalperiod. These results confirm previouswork that had shown that the develop-ment and maturation of Dominico hartónfruits depend basically on the activity ofthe functional leaves present during theappearance of the inflorescence (Arcilaet al. 1995, Cayón et al. 1995).
The number of fruits in the bunchremained within the range of expectedcharacteristics of Dominico hartón andFHIA-21 since physical damage causedby the phenomenon did not affect thiscomponent determined earlier duringthe floral differentiation phase(Belalcázar et al. 1991). The gale preju-diced the development of Dominicohartón plants. The damage was mostsevere in the bunches hit when they werein the first stages of formation (initiationof flowering and 15 DAF), where averagefruit weights were significantly lowerthan those of plants damaged at the 30-45 DAF stage and the controls. Thisbehaviour was obviously identical for thebunch weight. The average weight offruits in FHIA-21 was not statisticallychanged by the gale although heavierfruits (200 g) were noticed on the control
plants. In fact, the most serious damagecaused by wind and hail concerned theleaves responsible for the photosyntheticprocess of carbohydrate production andtranslocation to the fruits. It is probablethat this damage causes an increase inthe respiration of the last leaves emittedby the plant, decreasing the concentra-tion of the photosynthesised componentsfor fruit maturation carried by the elabo-rated sap. This suggests that the physio-logical and biochemical processes thatdetermine fruit growth and developmentare less susceptible to physical attackswhen these occur at advanced stages inthe production cycle.
Fruit length did not change. The aver-age values are within the classic rangefor Dominico hartón and FHIA-21. A sig-nificant decrease in fruit girth was notedin Dominico hartón fruits affected dur-ing floral initiation and at the 15 DAFstage in comparison with that of plantsaffected at the 30-45 DAF stage and ofthe control plants. The girth of FHIA-21fruits was not significantly decreasedafter the gale although the fruits of thecontrol plants were slightly larger. Thiscoincides with the observations aboveconcerning average fruit weight. Fruitexternal length and girth are directlyrelated to weight and are therefore gooddevelopment indicators. A study per-formed by Cayón et al. (1995) showedthat the numbers of hands and fingersper bunch were not affected by deleafingwhereas fruit length and size decreasedon an individual basis when deleafingwas severe.
The effect of wind and hail on bothtypes of plant material at the beginningof flowering or at the recently formedfruit stage (15 DAF) caused a significant
decrease in their size and in pulp freshweight. The phenomenon had less effecton fruit pulp affected at a more advancedplant development stage (35-40 DAF) andon that of the control plants (Table 2).Peel thickness and fresh weight inDominico hartón were changed by thegale in a similar manner to the pulp, thatis to say less in fruits exposed at earlystages of bunch development. Peel thick-ness in FHIA-21 did not vary according tothe moment in the cycle at which thedamage occurred. However, a slightincrease in peel weight was observed infruits exposed at the start of inflores-cence shooting. This suggests that thedamage to leaves during floral initiationprobably caused preferential distributionof photosynthesised substances to thepeel, disturbing normal fruit filling.
The fact that forming fruits were moreaffected whereas they are more protectedby the flower bracts reveals the greatphysiological importance of the func-tional leaves present at flowering becauseof their essential role in biomass accumu-lation in the fruits. Indeed, the fruit pulpformation process is strongly compro-mised when physical damage to leavesoccurs during the first stages of fruitdevelopment. The consequences are notas serious at later stages of development.
The effects of the gale on the chemicalcomposition of fruit pulp are shown inTable 3. Pulp mineral content repre-sented by the ash content was higher inthe control plants unaffected by the phe-nomenon in both Dominico hartón andFHIA-21. The level falls as the damageoccurs at an increasingly early stage. TheDominico hartón fruits that reachedmaturity under normal conditions in thezone studied had higher ash contents
INFOMUSA — Vol 10, N° 2 15
Table 1. The effect of wind and hail on bunches of the clone Dominico hartón andthe hybrid FHIA-21.
Development Average bunch Number Average Average Average stage weight (kg) of fruits fruit weight (g) fruit length (cm) fruit girth (cm)
Dominico hartón
Initiation of flowering 11.8 c 49 202.7 b 28.6 14.4 c
15 DAF 12.1 c 45 202.1 b 29.2 14.2 c
30-45 DAF 18.8 a 55 304.8 a 29.8 16.1 a
Control 17.0 b 49 309.1 a 32.1 15.1 b
C.V. (%) 8.3 11.1 13.3 8.0 1.2
F (stages) * n.s. * n.s. *
FHIA-21
Initiation of flowering 17.2 97 167 24.8 13.09
15 DAF 19.5 119 170 26.7 13.40
30-45 DAF 17.6 95 170.61 25.3 12.53
Control 22.4 104 200 22.5 14.00
C.V. (%) 14.36 17.4 20.57 7.6 8.30
F (stages) n.s. n.s. n.s. n.s. n.s.* F test significant (P< 0.05)
n.s. F test not significant
Means followed by the same letter are not significantly different according to Tukey’s test (P< 0.05)
than those found in the present study(Belalcázar et al. 1991, Morales et al.1998). This may show that damage toleaves, pseudostem and bunches may becritical for the normal translocation ofminerals to forming fruits when such
damage occurs between flower initiationand the 45 DAF stage.
The starch concentration in the pulp ofDominico hartón fruits did not decreaseafter damage to the plants during the var-ious stages of development. A small
decrease in the starch content was notedin FHIA-21 fruits at the 15 DAF and 30-45 DAF stages. The starch concentrationwas thus not markedly changed, in spiteof peel damage. It would appear thatmost of the sucrose required for starchsynthesis during fruit formation is fromleaf photosynthesis, with very little fromthe peel. According to Simmonds (1973)and Burdon et al. (1993), this is becausethe photosynthetic activity of plantainleaves is much higher than that of form-ing fruits because stomata density ismuch lower in the latter.
The damage to FHIA-21 plants did notsignificantly increase the total sugarsconcentration in the pulp of fruits har-vested at physiological maturity. Nomarked change in the total sugars con-tent was observed in Dominico hartónplants affected at early developmentstages (flower initiation and 15 DAF) incontrast with fruits at the 30-45 DAFstage in which the levels displayed amarked increase (5.1%). Such a figure isprobably a direct result of the effect ofhailstones on the fruits, which are moreexposed at this stage. Indeed, thenecrotic areas that formed may havecaused an increase in fruit respirationand hence premature ethylene produc-tion resulting in the breaking down ofstarch into sugars. This did not takeplace at earlier stages because thebunch was protected by the inflores-cence bracts. This is in agreement with astudy on Dominico hartón plants sub-jected to severe deleafing and in whichhigher total sugars levels were observed,indicating that drastic defoliationcauses an increase in the sugar synthe-sis process or that the sugars weretranslocated from the peel to the fruitpulp (Cayón et al. 1998b).
Evaluation of the physical damagecaused to Dominico hartón and FHIA-21bunches by the impact of hailstones isshown in Table 4. The bunches of both cul-tivars at the 30-45 DAF stage displayed ahigher percentage of fruits with necroticspots since they were more exposed tohailstone impact than the less developedfruits (15 DAF) that were sheltered by thebracts of the inflorescence. The weightand percentage of necrotic peel werelower in the fruits affected at the mostadvanced stage (30-45 DAF) while theweight and percentage of necrotic peelincreased, indicating that the initialnecrotic spots on fruit peel spread rapidlyinward, damaging the fruit pulp. It shouldbe underlined that although FHIA-21 hadfewer fruits, the percentage of fruits withnecrotic spotting was lower for the twostages considered. This can be attributed
16 INFOMUSA — Vol 10, N° 2
Table 3. The effects of the impact of wind and hail on the chemical composition ofthe pulp of fruits of Dominico hartón and FHIA-21.
Development stage Average ash Average starch Average total content (%) content (%) sugars content (%)
Dominico hartón
Initiation of flowering 1.3 c 83.3 1.2 b
15 DAF 1.4 b 82.7 1.6 b
30-45 DAF 1.4 b 80.4 5.1 a
Control 1.7 a 80.0 1.4 b
C.V. (%) 4.9 2.7 42.0
F (stages) * n.s. *
FHIA-21
Initiation of flowering 1.5 b 83.0 a 1.6
15 DAF 1.4 c 80.0 b 1.4
30-45 DAF 1.3 c 82.3 b 1.6
Control 1.8 a 84.0 a 1.3
C.V. (%) 6.0 1.4 16.9
F (Stages) * * n.s.* F test significant (P<0.05)
n.s. F test not significant
Means followed by the same letter are not significantly different according to Tukey’s test (P< 0.05)
Table 4. Damage caused by hailstone impact at two stages in the development ofbunches of Dominico hartón and FHIA-21.
Dominico hartón FHIA-21
Damage caused by hailstones 15 DAF 30-45 DAF 15 DAF 30-45 DAF
Fruits with necrotic spots (%) 96 100 79 81
Necrotic peel:fruit weight (g) 1.13 0.91 1.38 0.74
Necrotic peel:fruit (%) 1.1 0.6 1.4 0.9
Necrotic pulp:fruit weight (g) 0.72 1.5 0.13 0.16
Necrotic pulp:fruit (%) 1.1 1.6 0.5 0.3
Table 2. The effects of wind and hail on the pulp and peel of Dominico hartón andFHIA-21 fruits.
Development stage Pulp Peel
Average Average Average Averagethickness (cm) fresh weight(g) thickness (cm) fresh weight (g)
Dominico hartón
Initiation of flowering 11.4 b 126.1 c 0.38 c 76.6 b
15 DAF 11.0 c 121.1 c 0.38 c 81.0 b
30-45 DAF 12.5 a 186.2 b 0.41 b 118.6 a
Control 12.3 a 191.4 a 0.44 a 117.7 a
C.V. (%) 2.0 5.9 5.0 8.6
F (Stages) * * * *
FHIA-21
Initiation of flowering 7.2 85.7 c 3.7 81.3 a
15 DAF 10.1 101.1 b 4.1 68.9 c
30-45 DAF 10.0 95.7 b 3.8 62.2 c
Control 11.1 122.4 a 4.0 77.6 b
C.V. (%) 7.9 7.9 7.1 4.9
F (Stages) n.s. * n.s. ** F test significant (P< 0.05)
n.s. F test not significant
Means followed by the same letter are not significantly different according to Tukey’s test (P< 0.05)

to the more voluminous and compactFHIA-21 bunches that protect the inner-most fruits.
ConclusionsDominico hartón and FHIA-21 fruits
were most seriously damaged when windand hail affected plants at the flower ini-tiation and 15 days after flowering stages.
Hail damage to Dominico hartón andFHIA-21 fruits affected quality because ofthe appearance of necrotic spotting ofthe peel; this necrosis then spread to thepulp of the plantain.
The impact of hailstones on Dominicohartón and FHIA-21 plantain caused aslight decrease in the mineral (ash) con-tent of fruit pulp.
A significant increase in the total sug-ars content of the pulp of Dominicohartón fruits affected at an advancedstage of development (30-45 DAF) wasobserved.
AcknowledgementsThe authors thank Corporación
Colombiana de Investigación Agro-pecuaria CORPOICA and Universidaddel Quindío for their technical and finan-cial support and Mrs Gloria Inés López,secretary, for her patient and accuratetranscription work. ■
ReferencesArcila M.I., S. Belalcázar, J.A. Valencia & G. Cayón.
1995. Influencia del número de hojas en post-flo-ración sobre el llenado de los frutos del clon de
plátano Dominico hartón (Musa AAB Simmonds).Pp. 90-93 in Mejoramiento de la producción delcultivo del plátano. Segundo informe técnico1984-1994. Región 9. ICA, COMITECAFE Quindío,CIID, CORPOICA, INIBAP, INPOFOS. Armenia,Colombia.
Belalcázar S., J. Valencia & E. Lozada. 1991. Laplanta y el fruto. Pp. 43-89 in El cultivo del plá-tano en el trópico (S. Belalcázar, ed.). ICA, INI-BAP, CIID, COMITECAFE Quindío. Feriva,Colombia.
Burdon J.N., K.G. Moore & H. Wainwright. 1993. Thepeel of plantain and cooking banana fruits. Ann.Appl. Biol. 123 (2): 391-402.
Cayón G., J.E. Lozada & S. Belalcázar. 1995.Contribución fisiológica de las hojas funcionalesdel plátano (Musa AAB Simmonds) durante elllenado del racimo. Pp. 725-739 in ACORBAT.Memorias XI Reunión de la Asociación para laCooperación en Investigaciones de Banano en elCaribe y en América Tropical (V. Morales Soto,ed.). ACORBAT, San José, Costa Rica.
Cayón G., S. Belalcázar & J.E. Lozada. 1998a.Ecofisiología del plátano (Musa AAB Simmonds).Pp. 221-236 in Seminario internacional sobre pro-ducción de plátano. Memorias (M. Giraldo, S.Belalcázar, G. Cayón & R. Botero, eds). COR-POICA, Universidad del Quindío, COMITECAFÉQuindío, SENA. Armenia, Colombia.
Cayón G., M.I. Arcila, S. Belalcázar & L.D. Celis.1998b. Efecto de la remoción de hojas en floraciónsobre la partición de materia seca, carbohidratosy proteína en el racimo del plátano Dominicohartón (Musa AAB Simmonds). Pp. 153-164 in
ACORBAT. Memorias XIII Reunión de laAsociación para la Cooperación enInvestigaciones de Banano en el Caribe y en
América Tropical (L. Hidalgo Arizaga, ed.). CONA-BAN, Guayaquil, Equateur.
Decoteau D.R. 1990. Tomato leaf development anddistribution as influenced by leaf removal anddecapitation. HortScience 25: 681-684.
Morales H., S. Belalcázar & G. Cayón. 1998. Efecto dela época de cosecha sobre la composición físico-química de los frutos en cuatro clones comercialesde musáceas. Pp. 237-246 in Seminario interna-cional sobre producción de plátano. Memorias (M.Giraldo, S. Belalcázar, G. Cayón & R. Botero, eds).CORPOICA, Universidad del Quindío,COMITECAFÉ Quindío, SENA. Armenia,Colombia.
Simmonds N.W. 1973. Los plátanos. Blume,Barcelone. 539pp.
Valencia G. & J. Arcila. 1976. Daños por granizo encafetales. Avances Técnicos No. 53. CENICAFE,Colombia. 2pp.
Charli Rose Parra, Luz Mery Lara Sarmiento andGermán Giraldo Giraldo work at Universidad delQuindío, apartado aéreo 360, Tel. (576) 7460100;Gerardo Cayón Salinas at Corporación Colombianade Investigación Agropecuaria, CORPOICA, apartadoaéreo 1807. Tel. (576) 7493498. E-mail :[email protected]
INFOMUSA — Vol 10, N° 2 17
Polyphenol oxidase activity and gene expression in the fruit of Goldfinger bananas (AAAB, FHIA-01)
Fruit physiology oxidative browning
P.S. Gooding, C. Bird and S.P. Robinson
Banana is an important crop world-wide and one of the most impor-tant food sources in the developing
world (May et al. 1995). Annual produc-tion of bananas and plantains is esti-mated to be approximately 88 milliontonnes (FAO 1999), making them thelargest food crop in the world after thecereals (INIBAP 1997).
Variety SH-3481 (FHIA-01) was intro-duced into Australia from the breedingprogramme in Honduras (Daniells et al.1995). Popularly known as Goldfinger, it
was produced by a cross of a dwarf LadyFinger type (Dwarf Prata = SantaCatarina Prata AAB ‘Pome’) with SH-3142 which has some resistance toburrowing nematodes, Sigatoka leaf spotdiseases and Fusarium wilt (Rowe andRosales 1993). Goldfinger is a strongplant with good architecture, yielding alarge bunch size and fruit with goodflavour and postharvest green life. Thediced fruit shows less oxidation andbrowning than many commercial culti-vars (Rowe and Rosales 1993, Smith et al. 1998).
The oxidation of phenolic substrates bythe enzyme polyphenol oxidase (PPO; EC1.10.3.2) is thought to be the major cause
of the brown discolouration of bananafruit during handling, storage and pro-cessing (Mayer and Harel 1979, Vamos-Vigyazo 1981, Vaughn et al. 1988, Walkerand Ferrar 1998). This problem is of con-siderable importance to the food industryas it affects nutritional quality andappearance, reduces consumer accept-ability and therefore causes significanteconomic impact, both to primary pro-ducers and the food processing industry(Bachem et al. 1994). The damagecaused by harvesting and transport canmake the fruit unfit for sale and the useof banana in part processed fresh foodslike fruit salad and in processed foods,such as cakes and gateaux, could become
more viable if the fruit itself remainedpale yellow and attractive to the con-sumer after being peeled and sliced.
Enzymatic browning is caused by theproduction of complex polyphenolics, areaction catalyzed predominantly byPPO, which can oxidize a wide range ofphenolic substrates to produce reactivequinones (Vamos-Vigyazo 1981, Walkerand Ferrar 1998). These o-quinones canpolymerize and covalently bind tonucleophilic aminoacids to produce darkbrown or black pigments in postharvestfruits and vegetables (Mayer and Harel1979, Mayer and Harel 1991). PPOenzymes which are encoded by nucleargenes (Lax et al. 1984) are localized inthe plastid, where they are associatedwith the internal thylakoid membranes(Vaughn et al. 1988) and remain physi-cally separated from their phenolic sub-strates found in the vacuole (Robinsonand Dry 1992). Therefore, the browningreaction is usually only initiated upon tis-sue damage, where a loss of cellular com-partmentation allows PPO and phenolicsubstrates to mix (Vamos-Vigyazo 1981,Walker and Ferrar 1998).
Isoforms of banana PPO have been par-tially purified from pulp, peel and budsand characterized with respect to theirsubstrate specificities, optimum pH, pI,Km and the effect of inhibitors(Montgomery and Sgarbieri 1975,Thomas and Janave 1986, Oba et al. 1992,Sojo et al. 1998). Between 9 and 14 iso-enzymes have been reported from par-tially purified preparations of bananapulp separated by polyacrylamide gelelectrophoresis or isoelectric focusing(Montgomery and Sgarbieri 1975,Thomas and Janave 1986) and 10 werefound in peel (Montgomery and Sgarbieri1975). Two isozymes were reported fromextracts of banana buds separated bychromatography (Oba et al. 1992) butonly one was reported in banana pulppurified using an aqueous two-phase sys-tem (Sojo et al. 1998). The multiplicityobserved in the earlier reports may becaused by covalent modifications to thePPO enzyme during purification, as itreacted with (poly)phenols and tannins(Sojo et al. 1998). Using the most effi-cient enzyme purification system (Sojo etal. 1998) the pI value for banana PPOwas 6.2, the pH optimum was around pH5.5, and the Km was 0.57 mM in the pres-ence of dopamine, the natural substrateof PPO in banana (Palmer 1963).
Genes encoding PPO have been clonedand characterized from a wide range ofplants including banana (Gooding et al.2001) and generally comprise multi-genefamilies (Cary et al. 1992, Newman et al.
18 INFOMUSA — Vol 10, N° 2
Figure 1. PPO activity and fresh weight of Goldfinger fruit at different stages of development.A : Fresh weight gain of banana fruit through development from young inflorescence buds to full sizegreen fruit. B : PPO activity (units per gram fresh weight) of fruit samples through development.C : PPO activity (units per fruit) of fruit samples through development. Values are plotted as the meanof three independent samples and bars represent standard errors of the mean.

1993, Boss et al. 1995, Thygesen et al.1995). Interestingly, banana PPO con-tains a small (85 base pair) intron(Gooding et al. 2001), whereas all otherPPO genes characterized to date (mostlyfrom dicotyledonous plants) do not.
In banana fruit PPO activity is highestin young immature tissues and decreasestowards maturity (Gooding et al. 2001).It is known that Goldfinger fruit is moreresistant to oxidative browning thanother dessert varieties (Rowe andRosales 1993, Smith et al. 1998), but nodata on PPO activity and expression havebeen presented. Here we describe theexpression of four previously character-ized PPO cDNAs from banana cv. Williams(AAA group, Cavendish subgroup;Gooding et al. 2001), as well as the rela-tive PPO activity in Goldfinger fruit andother tissues compared to Williams.
Materials and methods
Plant materialBanana fruit, cultivars ‘Goldfinger’(AAAB, FHIA-01) and ‘Williams’ (AAA)were obtained from Nambour,Queensland, Australia. Fruit was har-vested at different stages, ranging fromyoung flower buds to full-sized green fruit(stage A-F, Figure 1B-C and Figure 2). Aflower head was dissected before itopened and young flower bud materialwas dissected into petals, stamens andstyle, which were removed and calledflowers (FL). The remaining material,which ultimately forms the fruit, was des-ignated stage A. A range of different sizedfruit was collected from post-floweringhands (stages B-F) and the pulp was sep-arated from the peel. All tissues were cutinto small pieces, frozen in liquid nitro-gen, ground to a fine powder in a coffeegrinder and stored at –70oC.
PPO activityA modification of the procedure ofRobinson and Dry (1992) was used todetermine PPO activity. Frozen tissuesamples were ground in a coffee grinderto a fine powder then approximately 0.1 gwas homogenized with a glasshomogenizer in 2 ml of buffer containing100 mM sodium phosphate, pH 7.0, 1%Triton X-100, 5 mM ascorbic acid. Analiquot of the whole homogenate wasused to determine PPO activity in an oxy-gen electrode. The initial rate of oxygenuptake was measured at 25o C, in 50 mMsodium phosphate, pH 7.0. The reactionwas started by the addition of 4-methylcatechol to a final concentration of2.5 mM, with one unit of enzyme activitydefined as that which resulted in the
uptake of 1µmol of molecular oxygen perminute under these assay conditions. Theaddition of SDS (0.1-1%) to the assay,which activates PPO in some plants(Vaughn and Duke 1988), had no effecton PPO activity of banana samples so itwas not included in the standard assay.
Total RNA extractionThe protocol was adapted from Chang etal . (1993) and was suitable for extract-ing RNA from banana pulp, peel, flower,stem, leaf and root tissue. Approximately3 g of powdered tissue was added to 15 mlof extraction buffer (containing 2% CTAB,2% PVP, 100 mM Tris-HCl, pH 8.0, 25 mMEDTA, 2 M NaCl, 0.5 g l-1 spermidine and 2% 2-mercaptoethanol) previouslywarmed to 65o C. The mixture wasextracted twice using chloroform:isoamy-lalcohol and the RNA precipitated with2 M lithium chloride. After centrifuga-tion, the RNA pellet was resuspended inbuffer containing 1 M NaCl, 0.5% SDS, 10 mM Tris-HCl, pH 8.0, 1 mM EDTA,extracted once with an equal volume ofchloroform:isoamylalcohol and precipi-tated with ethanol.
Total DNA extractionYoung banana leaves were harvested,frozen in liquid nitrogen and ground to afine powder. To 4 g of material, 2 ml ofCTAB buffer (containing 100 mM Tris-HCl, pH 8.0, 20 mM EDTA, pH 8.0, 2%CTAB, 1.4 M NaCl, 4% PVP-40, 0.1% ascor-bic acid, 0.1% diethyldithiocarbamic acidand 2% 2-mercaptoethanol) was added tomake a smooth paste. The paste wasadded to a centrifuge tube containing13 ml of 100 mM Tris-HCl, pH 8.0, 50 mMEDTA, pH 8.0, 100 mM NaCl, 2% 2-mer-
captoethanol and 1 ml of 20% SDS. Aftervigorous mixing, the tube was incubatedat 65o C for 20 minutes, then 5 ml of 5 Mpotassium acetate was added and thetube was centrifuged at 18 000 rpm for20 minutes. The supernatant was recov-ered, 15 ml of cold propan-2-ol was addedand the mixture was incubated at –20o Cfor 20 minutes. After centrifugation at 18 000 rpm for 20 minutes, the super-natant was discarded, the pellet washedin 70% ethanol, air dried and resus-pended in 500 µl of water.
Northern blottingTotal RNA (4 µg) from samples at differ-ent developmental and ripening stageswas separated on a 1.2% agarose gel con-taining 10% formaldehyde to maintaindenaturing conditions. The RNA wastransferred to Zeta-Probe membrane(Bio-Rad) by overnight capillary transferin 0.05 N NaOH. Membranes were prehy-bridized at 65o C for two hours inhybridizing solution containing 0.25 Msodium phosphate, pH 7.0, 1 mM EDTA,7% SDS. Hybridization was performedovernight at 65o C in fresh hybridisingsolution plus the [α-32P]dATP-labelledprobe. Hybridized membranes were incu-bated four times at 65o C in a high strin-gency wash solution containing 0.1 x SSC,0.1% SDS for 15 minutes, before beingautoradiographed for 16-72 hours at –80o
C using X-OMAT film (Kodak) with inten-sifying screens.
Southern blottingTotal genomic DNA (5 µg) was digestedwith appropriate restriction enzymes orleft uncut, separated on a 1.0% (w/v)agarose Tris-borate gel at 2.4 V cm-1 for
INFOMUSA — Vol 10, N° 2 19
Figure 2. Northern blots showing PPO gene expression in Goldfinger tissues. Each track represents 4 µg total RNA extracted from Goldfinger tissues, probed with either BPO1 (top row), BPO11(second row), BPO34 (third row) or BPO35 (bottom row). Samples were taken from younginflorescences (A), a developmental series of pulp (BF-FF) and peel (BP-FP), flowers (FL), and inner leaf roll (LR).

16 hours as previously described(Gooding et al . 1999) and transferred toZeta-Probe membrane (Bio-Rad) byovernight capillary blotting in 0.4 NNaOH. Membranes were prehybridized,hybridized, washed and examined byautoradiography as described above.
ResultsTissue samples were collected fromGoldfinger fruit at six different develop-mental stages designated A (young inflo-rescence, preflowering), B (15 g fruit),C (30 g fruit), D (60 g fruit), E (90 g fruit)and F (full size green fruit; 120 g) as wellas from flowers and young leaf roll mater-ial. Samples were assayed for PPO activ-ity and used for total RNA extraction.
PPO activity in Goldfinger andWilliams tissuesFor PPO activity measurements, samplesfrom three different fruits, flowers andleaf rolls were taken from Goldfinger andWilliams, ground up and assayed individ-ually and the mean PPO activity calcu-lated (Table 1). In both cultivars the PPOactivity was highest in flowers and lowestin mature fruit. In all tissues assayed thePPO activity in Goldfinger was signifi-cantly lower than in Williams (Table 1).
Goldfinger fruit samples were analyzedat six stages through fruit developmentfrom young inflorescence buds to full sizegreen fruit. It should be noted that the
young inflorescence sample was a mix-ture of pulp and peel, as the tissues werenot clearly differentiated enough to beadequately separated at this stage. Themean fresh weight of young inflorescencebuds (stage A) was 4.3 g and fruit weightincreased to around 120 g when full sizebut still green (stage F). The pulp madeup less than 20% of the total fresh weightat stage B but increased steadily, making
up 40% of the total fresh weight of the fullsize green fruit (stage F; Figure 1A).
On a gram per fresh weight basis, PPOactivity in pulp was considerably lowerthan in peel and decreased as fruitweight increased from about 40 U/g F.W.at stage B to 15 U/g F.W. at stage F (Figure1B). In peel, PPO activity rose to a peakof almost 150 U/g F.W. at stage C, thendecreased dramatically to less than halfof this value at stages D-F (Figure 1B).Total activity per gram fresh weight inGoldfinger fruit, calculated by determin-ing the total activity in pulp and peel anddividing by total fruit weight, mirroredthe curve of the peel sample, which con-tributed the majority of PPO activity(Figure 1B).
PPO activity was also calculated on aper fruit basis by multiplying the unit pergram value with the weight of the pulpand peel at each developmental stage(Figure 1C). These data give a clear indi-cation of when during development thePPO enzyme is being actively synthesizedand an insight into the total amount ofPPO protein present in the fruit. PPOactivity in pulp increased slowly as fruitsize increased, from 120 units per fruit atstage B to 750 units per fruit at stage F(Figure 1C). In the peel samples however,PPO activity increased considerably from1500 units per fruit at stage B to around3500 units per fruit at stage C and itremained at this level through the rest offruit development (Figure 1C). Again,due to the higher PPO activity in the peeltissue on a fresh weight basis, the graphof total activity closely followed that ofthe peel (Figure 1C).
PPO gene expression in GoldfingertissuesFour different partial length PPO clonesof approximately 900 base pairs were pre-viously cloned from banana (cv. Williams)and characterized (Gooding et al. 2001).These four PPO clones were used as mol-ecular probes against total RNAextracted from Goldfinger flowers, youngleaf rolls and fruit at different develop-mental stages. Total RNA extracted fromflowers, inner leaf rolls and pulp and peelsamples at each developmental stage wasprobed with each of the four PPO cDNAs(BPO1, BPO11, BPO34 and BPO35;Gooding et al. 2001). The specific activityof each probe was measured and found tobe of the same order of magnitude(around 2x105 cpm) and autoradiographexposures were identical. BPO1 wasexpressed in flowers and the inner leafroll (Figure 2, tracks FL and LR) and inpulp samples taken from fruit up to 60 g(Figure 2, tracks BF, CF and DF) with the
peak of signal occurring in track CF takenfrom 30 g fruit. Some BPO1 message wasalso detected in young inflorescence tis-sue (Figure 2, track A) and in peel fromup to 30 g fruit (Figure 2, tracks BP andCP). There was no detectable expressionof BPO11, BPO34 or BPO35 in any of thedeveloping fruit samples, nor in the flow-ers and leaf roll (Figure 2), even at longerexposures (data not shown).
Southern analysis of genomic DNAfrom GoldfingerTo compare the genomic organizationand copy number of PPO genes in the twocultivars, genomic DNA was extractedfrom young leaf roll tissue from bothGoldfinger and Williams (AAA group,Cavendish subgroup), digested with suit-able restriction enzymes or left undi-gested. The genomic DNA was run onagarose gels, blotted and probed witheach of the four banana PPO clones(Figure 3). Southern analysis revealedthe presence of multiple copies of BPO1 ,BPO11 , BPO34 and BPO35 within thegenome of both Goldfinger and Williams(Figure 3). Though the banding patternswere very similar between the two vari-eties, restriction length fragment poly-morphisms (RFLPs) were detected,notably for BPO1 and BPO34 whendigested with EcoRI and HindIII, forBPO35 when restricted with BamHI andEcoRI and for BPO11 cut with HindIII(Figure 3).
DiscussionIt has been reported that PPO activity ishigh in developing tissues and in meris-tematic tissues, but decreases as cellsmature (Cary et al. 1992, Shahar et al.1992, Hunt et al. 1993, Boss et al. 1995,Gooding et al. 2001). In banana (cv.Williams), PPO activity was found to behighest in flowers, young leaf roll and inyoung fruit samples (Gooding et al.2001). In the Goldfinger tissues assayed,PPO activity was also highest in flowers,young unexpanded leaf roll and youngpeel (Table 1 and Figure 1B). However,Goldfinger tissues were conspicuouslylower in PPO than the correspondingWilliams samples. PPO activities inWilliams flowers and young leaf roll wereapproximately 500 U/g F.W. (Gooding etal. 2001) compared to around 250 and100 U/g F.W. respectively in Goldfinger(Table 1). In fruit tissues, a similar lowerlevel of PPO activity was observed inGoldfinger compared to Williams. PulpPPO activity in Goldfinger was less thanhalf that of Williams at equivalent devel-opmental stages, ranging from about 40 U/g F.W. in young pulp to 20 U/g F.W.
20 INFOMUSA — Vol 10, N° 2
Table 1. Comparison of PPO activity(units per gram fresh weight) inflowers, young leaf roll, prefloweringinflorescence, immature fruit andmature fruit from banana cv.Goldfinger and cv. Williams. Meanvalues of PPO activity were calculatedfrom three separate samples of eachtissue and are given + standard error.
Goldfinger Williams
Flowers 259 + 32 502 + 81
Young leaf roll 108 + 11 507 + 85
Young 107 + 14 166 + 13inflorescence (stage A)
Immature 122 + 11 287 + 9fruit (30-40 g)
Mature fruit 35 + 3 63 + 1
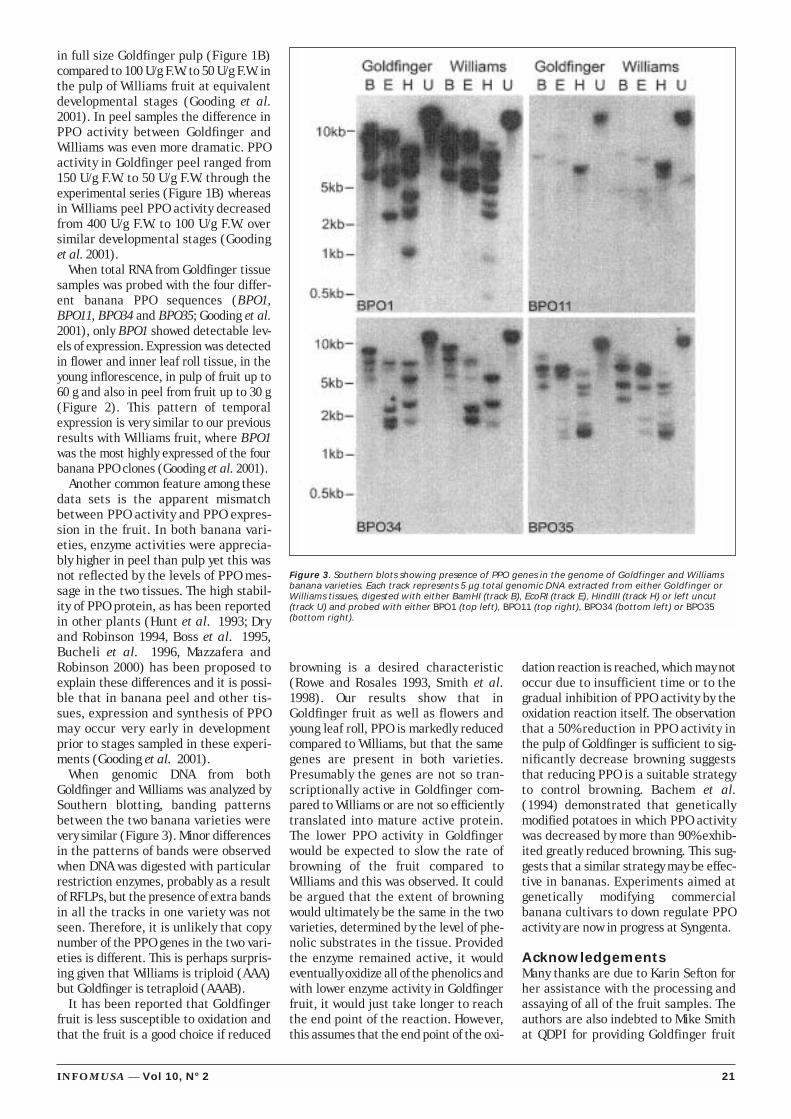
in full size Goldfinger pulp (Figure 1B)compared to 100 U/g F.W. to 50 U/g F.W. inthe pulp of Williams fruit at equivalentdevelopmental stages (Gooding et al.2001). In peel samples the difference inPPO activity between Goldfinger andWilliams was even more dramatic. PPOactivity in Goldfinger peel ranged from150 U/g F.W. to 50 U/g F.W. through theexperimental series (Figure 1B) whereasin Williams peel PPO activity decreasedfrom 400 U/g F.W. to 100 U/g F.W. over similar developmental stages (Gooding et al. 2001).
When total RNA from Goldfinger tissuesamples was probed with the four differ-ent banana PPO sequences (BPO1,BPO11, BPO34 and BPO35; Gooding et al.2001), only BPO1 showed detectable lev-els of expression. Expression was detectedin flower and inner leaf roll tissue, in theyoung inflorescence, in pulp of fruit up to60 g and also in peel from fruit up to 30 g(Figure 2). This pattern of temporalexpression is very similar to our previousresults with Williams fruit, where BPO1was the most highly expressed of the fourbanana PPO clones (Gooding et al. 2001).
Another common feature among thesedata sets is the apparent mismatchbetween PPO activity and PPO expres-sion in the fruit. In both banana vari-eties, enzyme activities were apprecia-bly higher in peel than pulp yet this wasnot reflected by the levels of PPO mes-sage in the two tissues. The high stabil-ity of PPO protein, as has been reportedin other plants (Hunt et al. 1993; Dryand Robinson 1994, Boss et al. 1995,Bucheli et al. 1996, Mazzafera andRobinson 2000) has been proposed toexplain these differences and it is possi-ble that in banana peel and other tis-sues, expression and synthesis of PPOmay occur very early in developmentprior to stages sampled in these experi-ments (Gooding et al. 2001).
When genomic DNA from bothGoldfinger and Williams was analyzed bySouthern blotting, banding patternsbetween the two banana varieties werevery similar (Figure 3). Minor differencesin the patterns of bands were observedwhen DNA was digested with particularrestriction enzymes, probably as a resultof RFLPs, but the presence of extra bandsin all the tracks in one variety was notseen. Therefore, it is unlikely that copynumber of the PPO genes in the two vari-eties is different. This is perhaps surpris-ing given that Williams is triploid (AAA)but Goldfinger is tetraploid (AAAB).
It has been reported that Goldfingerfruit is less susceptible to oxidation andthat the fruit is a good choice if reduced
browning is a desired characteristic(Rowe and Rosales 1993, Smith et al.1998). Our results show that inGoldfinger fruit as well as flowers andyoung leaf roll, PPO is markedly reducedcompared to Williams, but that the samegenes are present in both varieties.Presumably the genes are not so tran-scriptionally active in Goldfinger com-pared to Williams or are not so efficientlytranslated into mature active protein.The lower PPO activity in Goldfingerwould be expected to slow the rate ofbrowning of the fruit compared toWilliams and this was observed. It couldbe argued that the extent of browningwould ultimately be the same in the twovarieties, determined by the level of phe-nolic substrates in the tissue. Providedthe enzyme remained active, it wouldeventually oxidize all of the phenolics andwith lower enzyme activity in Goldfingerfruit, it would just take longer to reachthe end point of the reaction. However,this assumes that the end point of the oxi-
dation reaction is reached, which may notoccur due to insufficient time or to thegradual inhibition of PPO activity by theoxidation reaction itself. The observationthat a 50% reduction in PPO activity inthe pulp of Goldfinger is sufficient to sig-nificantly decrease browning suggeststhat reducing PPO is a suitable strategyto control browning. Bachem et al.(1994) demonstrated that geneticallymodified potatoes in which PPO activitywas decreased by more than 90% exhib-ited greatly reduced browning. This sug-gests that a similar strategy may be effec-tive in bananas. Experiments aimed atgenetically modifying commercialbanana cultivars to down regulate PPOactivity are now in progress at Syngenta.
AcknowledgementsMany thanks are due to Karin Sefton forher assistance with the processing andassaying of all of the fruit samples. Theauthors are also indebted to Mike Smithat QDPI for providing Goldfinger fruit
INFOMUSA — Vol 10, N° 2 21
Figure 3. Southern blots showing presence of PPO genes in the genome of Goldfinger and Williamsbanana varieties. Each track represents 5 µg total genomic DNA extracted from either Goldfinger orWilliams tissues, digested with either BamHI (track B), EcoRI (track E), HindIII (track H) or left uncut(track U) and probed with either BPO1 (top left), BPO11 (top right), BPO34 (bottom left) or BPO35(bottom right).

samples at many different developmentalstages and for helpful dialogue through-out this project. ■
ReferencesBachem C.W.B, G.J. Speckmann, P.C.G. van der
Linde, F.T.M. Verheggen, M.D. Hunt, J.C. Steffens & M. Zabeau. 1994. Antisense expression ofpolyphenol oxidase genes inhibits enzymaticbrowning in potato tubers. Bio/Tech 12: 1101-1105.
Boss P.K., R.C. Gardner, B.J. Janssen & G.S. Ross.1995. An apple polyphenol oxidase cDNA is up-regulated in wounded tissues. Plant Mol. Biol. 27: 429-433.
Bucheli C.S., I.B. Dry & S.P. Robinson. 1996. Isolationof a full-length cDNA encoding polyphenol oxidasefrom sugarcane, a C4 grass. Plant Mol. Biol. 31: 1233-1238.
Cary J.W., A.R. Lax & W.H. Flurkey. 1992. Cloning andcharacterization of cDNAs coding for Vicia faba
polyphenol oxidase. Plant Mol. Biol. 20: 245-253.Chang S.J., J. Puryear & J. Cairney. 1993. A simple
and efficient method for isolating RNA from pinetrees. Plant Mol. Biol. Rep. 11: 113-116.
Daniells J., K. Pegg, C. Searle, M. Smith, T. Whiley, P. Langdon, N. Bryde & T. O’Hare. 1995.Goldfinger in Australia : a banana variety withpotential. INFOMUSA 4(1):5-6.
Dry I.B. & S.P. Robinson. 1994. Molecular cloningand characterization of grape berry polyphenoloxidase. Plant Mol. Biol. 26:495-502.
FAO. 1999. Boletín trimestral de Estadística 12: 34.Gooding P.S., N.P. Batty, A.P. Goldsbrough
& P.M. Mullineaux. 1999. Plant cell-directed con-trol of virion sense gene expression in wheat dwarfvirus. Nucleic Acids Res. 27: 1709-1718.
Gooding P.S., C. Bird & S.P. Robinson. 2001.Molecular cloning and characterization of bananafruit polyphenol oxidase. Planta (in press ).
Hunt M.D., N.T. Eannetta,Y. Haifeng, S.M. Newman & J.C. Steffens. 1993. cDNA cloning and expres-sion of potato polyphenol oxidase. Plant Mol. Biol.21: 59-68.
INIBAP. 1997. Annual Report. International Networkfor the Improvement of Banana and Plantain,Montpellier, France.
Lax A.R., K.C. Vaughn & G.E. Templeton. 1984.Nuclear inheritance of polyphenol oxidase inNicotiana . J. Hered. 75: 285-287.
May G.D., R. Afza, H.S. Mason, A. Wiecko, F.J. Novak& C.J. Arntzen. 1995. Generation of transgenicbanana (Musa acuminata ) plants viaAgrobacterium -mediated transformation.Bio/Tech 13: 486-492.
Mayer A.M. & E. Harel. 1979. Polyphenol oxidases inplants. Phytochemistry 18: 193-215.
Mayer A.M. & E. Harel. 1991. Polyphenol oxidasesand their significance in fruit and vegetables. Pp.373-398 in Food Enzymology (P.F. Fox, ed.).Elsevier Science Publishers, New York.
Mazzafera P. & S.P. Robinson. 2000. Characterizationof polyphenol oxidase in coffee. Phytochemistry55: 285-296.
Montgomery N.W. & V.C. Sgarbieri. 1975. Isoenzymesof banana polyphenol oxidase. Phytochemistry 14: 1245-1249.
Newman S.M., N.T. Eannetta, H. Yu, J.P. Prince, M.C. de Vincente, S.D. Tanksley & J.C. Steffens.1993. Organization of the tomato polyphenol oxi-dase gene family. Plant Mol. Biol. 21: 1035-1051.
Oba K., N. Iwatsuki, I. Uritani, A.M. Alvarez & V.V. Garcia. 1992. Partial purification and char-acterization of polyphenol oxidase isozymes in banana bud. Biosci. Biotech. Biochem. 56: 1027-1030.
Palmer J.K. 1963. Banana polyphenol oxidase.Preparation and properties. Plant Physiol. 38: 508-513.
Robinson S.P. & I.B. Dry. 1992. Broad bean leafpolyphenol oxidase is a 60-kilodalton protein sus-ceptible to proteolytic cleavage. Plant Physiol. 99: 317-323.
Rowe P. & F. Rosales. 1993. Diploid breeding at FHIAand the development of Goldfinger (FHIA-01).INFOMUSA 2(2): 9-11.
Sambrook J., E.F. Fritsch & T. Maniatis. 1989.Molecular cloning: a laboratory manual, 2nd ed.
Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY.
Shahar T., N. Hennig, T. Guttinger, D. Hareven & E. Lifschitz. 1992. The tomato 66.3-kD polyphe-nol oxidase gene: molecular identification anddevelopmental expression. Plant Cell 4: 135-147.
Smith M.K., S.D. Hamill & P.W. Langdon. 1998.Selection of new banana varieties for the cool sub-tropics in Australia. Acta Hort. 490: 49-56.
Sojo M.M., E. Nunez-Delicado, F. Garcia-Carmona &A. Sanchez-Ferrer. 1998. Partial purification of abanana polyphenol oxidase using Triton X-114 andPEG 8000 for removal of polyphenols. J. Agric.Food Chem. 46: 4924-4930.
Thomas R. & M.T. Janave. 1986. Isoelectric focusingevidence for banana isoenzymes with mono anddiphenolase activity. Journal of Food Science 51: 384-387.
Thygesen P.W., I.B. Dry & S.P. Robinson. 1995.Polyphenol oxidase in potato. Plant Physiol. 109:525-531.
Vamos-Vigyazo L. 1981. Polyphenol oxidase and per-oxidase in fruits and vegetables. CRC CriticalReviews in Food Science 15: 49-127.
Van Gelder C.W.G., W.H. Flurkey & H.J. Wichers.1997. Sequence and structural features of plant andfungal tyrosinases. Phytochemistry 45: 1309-1323.
Vaughn K.C., A.R. Lax & S.O. Duke. 1988. Polyphenoloxidase: the chloroplast oxidase with no estab-lished function. Physiol. Plant. 72: 659-665.
Walker J.R.L. & P.H. Ferrar. 1998. Diphenol oxidases,enzyme-catalysed browning and plant diseaseresistance. Biotechnology and GeneticEngineering Reviews 15: 457-498.
Paul Gooding and Simon Robinson work at CSIRO– Plant Industry, PO Box 350, Glen Osmond, SouthAustralia 5064, Australia. Email:[email protected]; Fax. (+61) 8 8303 8601;Colin Bird works at Syngenta, Jealott’s Hill ResearchStation, Bracknell, Berkshire, RG12 6EY, UK.
22 INFOMUSA — Vol 10, N° 2
In vitro multiplication of FHIA-18 plantain in the presence of paclobutrazol
Tissue culture Shoot regulation in liquid medium
M. Daquinta, Y. Lezcano, M. Escalonaand R. Santos.
In vitro culture techniques usingshoot tips or meristems have beenused since 1972 for obtaining clean
material and performing rapid multipli-cation of different clones of the genusMusa (Berg and Bustamante 1974, Dore
Swamy et al. 1983). These techniqueshave enhanced national and interna-tional exchange of tissue culture materialwith the assurance that this does notspread diseases. In vitro conservationand cryopreservation (Villalobos andAbdelnour 1992) developed, as did tissueculture techniques supporting geneticimprovement programmes (Escalant andTeisson 1991).
In practice, these techniques have dis-played various advantages over tradi-tional multiplication methods includingthe obtaining of pathogen-free plants, thesmaller volume of seed material to betransported, a decrease in the areasdevoted to seed multiplication, continu-ous seedling production all the yearround and the rapid multiplication ofrecent or new varieties. All this con-

tributes to accelerating the developmentof new plantations.
Paclobutrazol (PBZ) is a growthinhibitor very commonly used in agricul-ture but whose effect on plantain in vitrohad not been reported. The present workon FHIA-18 plantain concerns the effectof PBZ on the budding of shoots in differ-ent types of culture.
Material and methodsShoot tip buds from nurseries were usedfor the in vitro culture of FHIA-18. Theplant material was washed with deter-gent before two disinfection operationswith calcium hypochlorite 3%. The buds,1 cm high with a base of approximately0.5 cm2 were subcultured on MS medium(Murashige & Skoog 1962) enriched with4 mg/L BAP. The pH of the culture mediawas adjusted to 5.8 before autoclave ster-ilization at 1.2 kg/cm2 and 121°C for 15minutes. All the cultures were main-tained under the same photoperiod con-ditions, in natural light, at 25° ± 2°C andhigh relative humidity (>80%).
Part of the pseudostem of the micro-propagated plants was incised to themeristem and cut transversally. The twoparts obtained were subjected to theexperiments described below.
Evaluation of the effect of PBZ onthe number and height of shoots ofFHIA-18
The buds were cultured on MS mediumenriched with 4 mg/L BAP, alone or com-bined with paclobutrazol (1 or 2 mg/L)using various culture methods: solidmedium, stationary liquid medium andtemporary immersion. Seven explantswere used per flask with 10 repetitionsper treatment. Three repetitions wereperformed for temporary immersion.
Temporary immersion is a new tissueculture technique in which the explantsare bathed in the culture medium for avery short time (3 minutes) every threehours. The medium returns to the cultureflask storage. These operations arerepeated for three weeks (Teisson andAlvard 1995).
Results and discussionVariations in the number of buds wereobserved according to the medium andthe type of culture used (Table 1). MSmedium with 4 mg/L BAP induced asmaller number of buds in stationaryliquid medium. However, the addition of2 mg/L PBZ improved this parameter. Theaddition of PBZ to the medium alwaysincreased the number of buds in solidmedium and in stationary liquid medium.This was not the case in the temporary
immersion system in which the mediumenriched with 4 mg/L BAP displayed bet-ter results, even though there were nosignificant differences with the mediumenriched with 2 mg/L PBZ. The synergiceffect of PBZ has been reported in orna-mental Araceae (Werbrouck and Debergh1996). This stimulant effect did notappear to occur with the concentrationsused in this work on plantain. However,Ziv et al. (1998) achieved the stimulationof banana budding with Ancymidol,another growth inhibitor. Temporaryimmersion was found to be the culturemethod that increases the number ofshoots. Teisson and Alvard (1995)stressed the advantages of this type ofculture. A temporary immersion systemflask is shown in Figure 1. The smaller
shoot size can be seen; this enables theproduction of a larger number of plantsper culture unit.
Shoot growth behaviour is shown inTable 2. The addition of PBZ to the cul-ture medium reduced shoot height what-ever the type of culture. A greater reduc-tion in shoot growth according to theincrease in PBZ concentration can beseen in the liquid culture medium.However, greater shoot height was alwaysobtained in the liquid media (stationaryand temporary immersion) than in cul-ture on solid medium whatever the treat-ment used. This would appear to showthat the type of culture has considerableeffect on bud growth. Although noinhibiting effect of PBZ on banana andplantain shoot growth has been observed,
INFOMUSA — Vol 10, N° 2 23
Figure 1. A temporary immersion system flask containing plantain explants cultured in the presenceof PBZ.
Table 1. The effect of PBZ on the number of shoots of FHIA-18 in different types of culture. Ciego de Avila, 1997.
Types of culture
Treatment Solid Liquid Temporary immersion
MS + 4 BAP 3.8 b 3.4 b 7.4 a
MS + 4BAP + 1PBZ 5.2 b 4.7 c 6.2 a
MS + 4BAP + 2PBZ 4.0 c 6.0 b 7.3 a
C.V. 11.43 13.87 10.92The values with different letters in the same column are different in Duncan’s test (p<0.05).
Table 2. The influence of PBZ on the height( (in cm) of FHIA-18 shoots in thedifferent types of culture. Ciego de Avila, 1997.
Type of culture
Treatment Solid Liquid Temporary immersion
MS + 4 BAP 2.10 b 3.30 a 3.69 a
MS + 4BAP + 1PBZ 1.68 b 2.16 a 2.51 a
MS + 4BAP + 2PBZ 1.58 b 1.70 b 2.32 a
C.V. 9.03 10.34 8.46The values with different letters in the same column are different in Duncan’s test (p<0.05).

this has been reported for numerousplants, such as pineapple (Daquinta et al.1994) and apple (Steffens and Wang1986). Wang found that inhibition ofshoot extension corresponded to achange in cell wall composition in whichPBZ degraded polysaccharides. Thexylem:phloem ratio also decreased.
ConclusionsIt was verified in this work that PBZ
has an inhibitory effect on the shootgrowth of FHIA-18, although the coeffi-cient of multiplication for PBZ treat-ments did not differ from the control inthe case of temporary immersion. Boxuset al. (1991) reported that the develop-ment of very vigorous leaves on bananaplants reduces the number of explantsper flask to a maximum of seven or eight.This also makes subcultures difficultbecause the phenolized parts must be cutback and cleaned, delaying micropropa-gation and making the work more labori-ous. With PBZ, the size of tissue cultureplants is reduced, making the techniquemore effective for banana micropropaga-tion. The use of this growth inhibitor inculture media enables us to increase thenumber of explants per culture unit andthus make better use of the media. Itseems that synergy between PBZ andcytokinins can only be obtained at higherconcentrations or by using otherinhibitors, as mentioned by Ziv et al.(1998). ■
References Berg L.A. & M. Bustamante. 1974. Heat treatment
and meristem culture for the production of virus-free bananas. Phytopathology 64: 320-322.
Boxus P., J.M. Terzi, C. Lievens, M. Pylyser, P. Ngaboyamahina & K. Duhem. 1991.Improvement and perspectives of micropropaga-tion techniques applied to some hot climateplants. Acta Horticulturae 289: 55-64.
Daquinta M.,T. Martinez, R. Castillo & R. Benega.1994. Estimulación del ahijamiento en vitroplan-tas de piña con paclobutrazol. Centro Agrícola21(2): 75-82.
Dore Swamy R., N.K. Sirinasa & E. Chacko. 1983.Tissue-culture propagation of banana. ScientiaHorticulturae 18: 247-253.
Escalant J.V. & C. Teisson. 1988. Embryogenèsesomatique chez Musa sp. CR Acad. Sc. Paris 306: 277-281.
Murashige T. & F. Skoog. 1962. A revised medium forrapid growth and bioassays with tobacco tissuecultures. Physiol. Plant. 15: 473-497.
Steffens G.L. & S.Y. Wang. 1986. Biochimical andphysiological alterations in apple trees caused by agibberellin biosynthesis inhibitor, Paclobutrazol.Acta Horticulturae (ISHS) 179: 433-442.
Teisson C. & D. Alvard. 1995. A new concept of plantin vitro cultivation liquid medium: Temporary
24 INFOMUSA — Vol 10, N° 2
immersion. Pp. 105-110 in Current issues in Plant Molecular and Cellular Biology (M. Terzi, R. Cella & A. Falavigna, eds). Kluwer Acad. Pub.,Dordrecht, Pays-Bas.
Villalobos V. & A. Abdelnour. 1992. Cryoconservationof Musa spp. and its potential for long-term stor-age of other tropical crops. Pp. 197-209 inConservation of plant genes, Academic Press, Inc.
Werbrouck S.P.O. & P.C. Debergh. 1996. Imidazolefungicides and paclobutrazol enhance cytokinin-
induced adventitious shoot proliferation inaraceas. Plant Growth Regulation 15: 81-85.
Ziv M., G. Ronen & M. Raviv. 1998. Proliferation ofmeristematic clusters in disposable presterilizedplastic bioreactors for the large-scale micropropa-gation of plants. In Vitro Cell. Dev. Biol. Plant 34: 152-158.
The authors work at Centro de Bioplantas, Carreteraa Morón Km. 9, Ciego de Avila, CP 69450. Cuba. E-mail: [email protected]
Preliminary results of a virulencetest of the populations ofFusarium oxysporum f.sp. cubensefor different banana cultivars in the greenhouse
Diseases Early screening of Foc pathogenicity
Do Nang Vinh, Nguyen Van Khiemand Le Huy Ham
Fusarium oxysporum f.sp. cubense(Foc) is considered a highly com-plex pathogen. Up to now,
researches on banana have shown rela-tionships between races of Foc and theirhosts in many parts of the world: races 1and 2 attack not only Gros Michel (AAA),Silk, Lady Finger (AAB), but also Bluggoeand Pisang Awak (ABB); race 4 attacksthe Cavendish group (AAA) and cultivarssusceptible to races 1 and 2 (Waite 1977,Bentley et al., 1998).
In Vietnam, race 1 (VCGs 0124, 0124/5,0125, 0124/5-0125, 0124-0124/5-0125)attacks Chuoi Tay (Pisang Awak), ChuoiNgop (Bluggoe), Chuoi Com La (Silk,AAB) (Khiem et al. 2000) and Tay Tia(Silk, AAB) (Pegg and Moore 1995).However, the response of Vietnamese cul-tivars in collections to Foc has not previ-ously been thoroughly studied.
The objective of this study was to char-acterize different isolates of Foc usingthe greenhouse virulence test.
Material and methodsThree-month-old tissue culture bananaplants of eight different cultivars (ChuoiTay (Pisang Awak, ABB), Ngop (Bluggoe,ABB), Tay Tia (Silk, AAB), Com La (Silk,AAB), Man (Mysore, AAB), Tieu Trung(Cavendish, AAA), Tay Nang (AAA) andBom (AA) were grown in 20-L pots con-taining Pasteur-sterilized soil in a green-house at 25-35oC.
Samples of 250 g banana tissue affectedby isolates belonging to VCGs 0124,VCG 0124/5, VCG 0124-0124/5-0125 wereburied in a 5 cm-deep trench, at a dis-tance of 7-10 cm from the plant pseu-dostem. Non-inoculated banana tissuewas added to control pots. Each test iso-late was used to inoculate four plants ofdifferent banana cultivars. Disease sever-ity was assessed after 4 months accordingto Moore et al. (1993). Plants were scoredfor external symptoms on a 1-5 scale:1, no yellow discolouration of leaves;2, yellowing of lower leaves; 3, yellowingof all lower leaves with some discoloura-tion of younger leaves; 4, all leaves yellow; 5, plant dead. Rhizomes and pseudostemsof plants were then cut longitudinally todetermine the extent of invasion of vascu-lar tissue. Rhizome discolouration wasrated on a 1-7 scale: 1, no discolouration;2, discolouration confined to root rhizomeplexus; 3, trace to 5% of stellar region dis-coloured; 4, 6-20% of stellar region dis-coloured; 5, 21-50% of stellar region dis-coloured; 6, 51-99% of stellar regiondiscoloured; 7, 100% discoloured.
Pseudostems were scored for invasion ofvascular tissue on a 1-4 scale: 1, no inva-sion of pseudostem; 2, invasion of outerleaf sheaths only; 3, invasion of moststands except for central leaf cylinder;4, invasion of all vascular strands. Ratingswere summed to give a total disease rating.
ResultsAll isolates of race 1 (VCGs 0124, VCG0124/5, and ‘VCG 0124-0124/5-0125’)caused the first symptoms of Fusarium wilt

disease observed on young leaves after 47-65 days in Com La, and 62-75 days inChuoi Tay. Typical disease symptoms werealso expressed in Chuoi Tay, and Com Laafter 4 months of inoculation in the green-house. These isolates did not significantlydiffer in virulence in either of these culti-vars (total rating ranged from 11.5-13.0)(Table 1). The pathogen was readily re-iso-lated from the discoloured tissue and theVCGs of the isolates confirmed. Non-inocu-lated control plants of both Chuoi Tay andCom La remained healthy and free fromexternal or internal disease symptoms(total rating was about 3.25-3.50).
Race 1 isolates (VCGs 0124, VCG 0124/5,and ‘VCG 0124-0124/5-0125’) did not causeexternal or internal symptoms in ChuoiTieu Trung (Cavendish, AAA), Tay Nang(AAA), Tay Tia (AAB), Man (Mysore, AAB),Bom (AA) and Ngop (Bluggoe, ABB) in thesame conditions (total rating ranged 3.5-4.5) (Table 1). However, internal exam-ination of the rhizomes of plants of thesecultivars revealed a light discolouration ofthe roots up to, but not beyond, the junc-tion of the root and the stellar region of therhizome. All non-inoculated control plantsof six of these cultivars remained healthyand free from external or internal symp-toms (total rating about 3.0-4.0).
DiscussionThe greenhouse pathogenicity test demon-strated that only Chuoi Tay (Pisang Awak,ABB) and Chuoi Com La (AAB) wereattacked by race 1 (VCGs 0124, 0124/5,0124-0124/5-0125). Chuoi Tieu Trung(Cavendish, AAA), Chuoi Tay Nang (AAA),Tay Tia (AAB), Man (Mysore, AAB), Ngop(Bluggoe, ABB), and Chuoi Bom (AA) werenot affected by these VCGs of race 1.However, in another study (Khiem et al.2000), we indicated that Chuoi Tay, ChuoiCom La and Chuoi Ngop cultivars wereinfected by Fusarium wilt in the field. Peggand Moore (1995) also reported Tay Tia(AAB) to be susceptible to Fusarium wiltin Vietnam. In our opinion, Chuoi Ngop andTay Tia become infected only in severeenvironmental conditions. Therefore, thestudy indicates that Chuoi Tay and Com Lacultivars are susceptible to Fusarium wiltdisease, but Tay Tia cultivar is not suscep-tible to Fusarium wilt disease as originallyreported by Pegg and Moore (1995).
Results from the greenhouse patho-genicity test also support the evolutionarymodel proposed by Puhalla (1985), whichsuggests that the genes governing vegeta-tive compatibility and those for patho-genicity in formae speciales of Fusariumoxysporum are fixed in the same thallus.Virulence for different banana cultivarswas also found to vary within a VCG.
Beckam et al. (1962) and Buddenhagen(1990) indicated that all strains of thepathogen are able to colonize the roots ofboth resistant and susceptible cultivars.Therefore, it is not unexpected that resis-tant banana cultivars (e.g. Chuoi Tieu, TayNang, etc.) inoculated with the race 1VCGs in the greenhouse pathogenicitytest were only able to block the pathogenat the root-rhizome plexus. ■
ReferencesBeckman C.H., S. Halmos & M.E. Mace. 1962. The
interaction of host, pathogen and soil tempera-ture in relation to susceptibility of Fusarium wiltof bananas. Phythopathology 51: 507-515.
Bentley S., K.G. Pegg, N.Y. Moore, R.D. Davis & I.W.Buddenhagen. 1998. Genetic variation among veg-etative compatibility groups of Fusarium oxyspo-
rum f.sp. cubense by DNA fingerprinting. Ecologyand Population Biology 88(12): 1283-1293.
Buddenhagen I.W. 1990. Banana breeding andFusarium wilt. Pp. 107-113 in Fusarium wilt ofbanana. First International Conference onFusarial Wilt of Banana, Miami, USA, 27-30 August1989 (R.C. Ploetz, ed.). APS, St Paul, USA.
Khiem N.V., N.D. Vinh, C.B. Phuc & L.H. Ham. 2000.Study on vegetative compatibility and distributionof Fusarium oxysporum f.sp. cubense in NorthVietnam. Journal of Agriculture and FoodIndustry: 416-418.
Moore N.Y., K.G. Pegg, R.N. Allen & J.A.G. Irwin.1993. Vegetative compatibility and distribution ofFusarium oxysporum f.sp. cubense in Australia.Australian Journal of Experimental Agriculture33: 797-802.
Pegg K.G. & N.Y. Moore. 1995. Fusarium wilt(Panama wilt) in the Asia and Pacific area. Pp. 42-44 in Banana Production, Biotechnology andBiodiversity. Workshop held in Hanoi, VietnamOct. 1995.
Puhalla J.E. 1985. Classification of strains ofFusarium oxysporum on the basis of vegetativecompatibility. Canadian Journal of Botany 63: 179-183.
Waite B.H. 1997. Inoculation studies and naturalinjection of banana varieties with races 1 andrace 2 of Fusarium oxysporum f.sp. cubense.Plant Disease Reporter 61: 15-19.
The authors work at the Institute of AgriculturalGenetics, Tu liem, Hanoi, Vietnam.
INFOMUSA — Vol 10, N° 2 25
Table 1. Disease severity ratings for different banana cultivars inoculated withisolates of Foc in the greenhouse (values are means of 4 plants).
Cultivar VCG Leaf Rhizome Pseudostem Total symptoms discolouration discolouration rating
(1-5) (1-7) (1-4) (1-16)
Chuoi Tay 0124 3.5 6.0 3.0 12.5
(ABB) 0124/5 3.5 5.0 3.0 11.5
0124-0124/5-0125 3.25 6.0 3.25 12.5
Control - 1.25 1.0 1.0 3.25
Chuoi 0124 3.25 6.25 3.5 13.0
Com La 0124/5 3.75 6.0 3.25 13.0
(AAB) 0124-0124/5-0125 3.25 5.75 3.25 12.25
Control - 1.25 1.25 1.0 3.5
Chuoi Tieu 0124 1.0 1.5 1.0 3.5
(AAA) 0124/5 1.25 1.25 1.25 3.5
0124-0124/5-0125 1.5 1.25 1.0 3.75
Control - 1.0 1.0 1.0 3.0
Chuoi 0124 1.5 1.0 1.0 3.5
Tay Nang 0124/5 1.5 1.25 1.0 3.75
(AAA) 0124-0124/5-0125 1.25 1.75 1.0 4.0
Control - 1.25 1.0 1.0 3.25
Chuoi 0124 1.5 2.0 1.0 4.5
Tay Tia 0124/5 1.5 1.25 1.0 3.75
(AAB) 0124-0124/5-0125 1.5 1.5 1.0 4.0
Control - 1.25 1.0 1.0 3.25
Chuoi Man 0124 1.0 1.5 1.0 3.5
(AAB) 0124/5 1.5 1.5 1.0 4.0
0124-0124/5-125. 1.5 1.5 1.0 4.0
Control - 1.25 1.0 1.0 3.25
Chuoi 0124 1.0 1.5 1.0 3.5
Bom 0124/5 1.75 1.25 1.0 4.0
(AA) 0124-0124/5-0125 1.5 1.75 1.0 4.25
Control - 1.25 1.0 1.0 3.25
Chuoi 0124 1.5 1.5 1.0 4.0
Ngop 0124/5 1.25 1.75 1.0 4.0
(ABB) 0124-0124/5-0125 1.5 1.75 1.0 4.25
Control - 1.5 1.5 1.0 4.0

A. Adiko and A. Badou N’Guessan
Plantain is a major staple food intropical and subtropical regions,particularly in West and Central
Africa, which account for 44% of globalproduction (Frison and Sharrock 1988).Cultivated by smallholders using tradi-tional farming systems, plantain is gener-ally mixed-cropped with either perennialcrops or other food crops. Consequentlythe yield is low and varies between 5 and10 t.ha-1.
At present, there is a noticeable trendtowards pure stands of plantain, particu-larly in Latin America and the Caribbean,following Belalcázar’s work relative to theuse of high-density planting (Belalcázarand Cayón 1998).
Besides the inadequacies of the tradi-tional cropping systems, the productionof plantain is hindered by several pestsand diseases. Among these biotic factors,plant parasitic nematodes are one of the main constraints (Gowen andQuénéhervé 1990). In Côte d’Ivoire, a large-scale survey was conducted in1985-1986 to determine the fauna ofnematodes that parasitize the crop andthe distribution of the banana nematode,Radopholus similis (Adiko 1988). Owingto the lack of regulatory control measuresenforcement in the country, it appearednecessary to assess the changes thatmight have occurred within the nema-tode populations associated with the crop12 years later.
Materials and methodsThe survey was conducted in 1997 and1998. Surveyed fields included all types ofplantain production systems, compoundproduction, plantain associated with foodcrops, cash crops and plantain in purestands. One hundred and thirteen (113)samples were collected (63 in theSoutheast and 50 in the Midwest).Sampling consisted of digging with ashovel a hole about 50 cm long from thebase of the mother plant, 30 cm wide and30-40 cm deep. All the roots thus removedwere collected. At each sampling site, 5 root subsamples were taken, thoroughlymixed and a composite sample (800-1000 g) was pooled. Samples were sealed
in polyethylene bags. From one field tothe other, the shovel was rid of soil parti-cles. In the laboratory, roots were rinsedwith tap water and chopped in 0.5 cm sec-tions. Nematodes were extracted accord-ing to the mistifier extraction technique(Seinhorst 1950). They were identified togenus level and counted under a stereobinocular microscope. For identificationto species level, specimens of each genuswere gently killed by immersion in hotwater and processed in glycerine accord-ing to Seinhorst’s (1959) rapid method.
Nematode populations were definedaccording to their frequency and abun-dance. Fortuner and Merny (1973)defined the frequency as the number ofsamples where a species is isolated. Theyfurther considered as frequent a speciesfound in at least 30% of the samples. As tothe abundance, it was determined as thelog average number of the species in thefield. Thus, on the basis of the thresholdof banana tolerance to R. similis set byGuérout (1972), we considered a speciesabundant when its mean number wasgreater than 3 (log 1000 individuals/100 gof roots).
Results and discussionsContrary to the first survey that recordedplant parasitic nematodes in both soil androots of plantain (Adiko 1988), the studyherein reported deals only with nematodesin the roots. Six species of endoparasiticnematodes were found associated withplantain, including Helicotylenchus mul-ticinctus, Hoplolaimus pararobustus,Meloidogyne spp., Pratylenchus coffeae,Radopholus similis and Rotylenchulusreniformis. The fields in the traditionalplantain-producing regions of theSoutheast harboured all the species
whereas in the Midwest, the recent plan-tain-producing area, only four species wereisolated (Table 1). Based on Fortuner andMerny’s classification parameters (1973),these species fall into three and twogroups in the Southeast and the Midwestrespectively (Figure 1):• frequent and abundant species (upper
right area of the diagrams): H. multi-cinctus, Meloidogyne spp. and P. cof-feae in the Southeast; H. multicinctus,Meloidogyne spp. in the Midwest;
• frequent but non-abundant species(lower right area of the diagrams): R. similis in the Southeast;
• non-frequent and non-abundantspecies (lower left area of the dia-grams): H. pararobustus and R. reni-formis in the Southeast; R. similis andP. coffeae in the Midwest.As was the case already in 1985-1986
(Adiko 1988), H. multicinctus andMeloidogyne remain the most wide-spread and numerous nematodes infect-ing plantain roots. The superficialnecrotic lesions and galls caused respec-tively by the two species are known not tocause significant impediment to the pro-ductivity of Musa in tropical regions(Stover 1972, Adiko 1989, Gowen andQuénéhervé 1990).
Hardly detectable in the samples dur-ing the first survey (Adiko 1988), H. pararobustus was still absent in theMidwest but became relatively frequentand abundant in the Southeast. As to theinfestation of R. reniformis, it remainedunchanged, i.e. absent in the Midwestand relatively frequent and abundant inthe Southeast.
The main features of the fauna ofnematodes associated with plantainreside in the infestations of P. coffeae and
Evolution of the nematofauna of plantain, Musa AAB, in Côte d’Ivoire
Pests Survey on nematode diversity
Table 1. Occurrence and population densities of nematode species associated with plantain in Côte d’Ivoire.
Southeast Midwest
Nematode species Nematodes/100g % infested Nematodes/100g % infested roots fields roots fields
Helicotylenchus multicinctus 53 314 96 41 525 100
Hoplolaimus pararobustus 613 19 - -
Meloidogyne spp. 1 940 90 1 851 93
Pratylenchus coffeae 2 704 35 4 2
Radopholus similis 460 43 48 2
Rotylenchulus reniformis 556 12 - -
26 INFOMUSA — Vol 10, N° 2

R. similis. Indeed, P. coffeae, which wasisolated for the first time on plantain inCôte d’Ivoire 12 years ago in only onefield in Aboisso (Adiko 1988), hasbecome prevalent in the whole Southeastregion. As a matter of fact, its populationsize superseded that of R. similis, for-merly one of the most frequent and abun-dant nematodes in the traditional plan-tain-growing areas. The fields of theMidwest were still relatively free of thesetwo damaging nematodes (Sarah et al.1996, Bridge et al. 1997). The occurrenceof P. coffeae throughout the Southeast issymptomatic of the dangers resultingfrom the unregulated movement of plant-ing material. In fact, the infestations of P. coffeae and R. similis in the Midwestwere recorded in a single field estab-lished with suckers taken from theSoutheast. Undoubtedly, P. coffeae will bedisseminated to the 5500 ha of commer-cial banana orchards in the south of thecountry.
Both P. coffeae and R. similis aremigratory endoparasitic nematodesinvading the root cortex and the corm ofMusa. Furthermore, their life cycle fromegg to egg is of 20-27 days at 25-30ºC(Loos 1962, Pinochet 1978). Therefore,the recent prevalence of P. coffeae over R. similis, previously well established,could suggest that the former is probablymore pathogenic than the latter. The con-comitant presence of these nematodeswarrants further research to elucidatetheir interactions on both plantain andbanana. ■
ReferencesAdiko A. 1988. Plant-parasitic nematodes associated
with plantain, Musa parasidiaca (AAB), in theIvory Coast. Revue de Nématologie 11(1): 109-113.
Adiko A. 1989. Effect of Meloidogyne incognita onplantain, Musa AAB. International NematologyNetwork Newsletter 6(3): 27-30.
Belalcázar S.C. & G.S. Cayón. 1998. High-densityplanting. Pp. 147-154 in Proceedings of the
International seminar on plantain production,Armenia, Quindío, Columbia, 4-8 May 1998 (M.J. Gerardo Cardona, S.L. Belalcazár Carvajal,D.G. Cayón Salinas & R.G. Botero Isaza, eds).
Bridge J., R. Fogain & P.R. Speijer. 1997. The rootlesion nematodes of banana. Musa Pest FactSheets No. 2. INIBAP, Montpellier, France.
Fortuner R. & G. Merny. 1973. Les nématodes phy-toparasites des racines associés au riz en Basse-Casamance (Sénégal) et en Gambie. Cahiers del’ORSTOM, série Biologie 21: 3-18.
Frison E. & S. Sharrock. 1998. The economic, socialand nutritional importance of banana in theworld. Pp. 21-35 in Bananas and food security/Lesproductions bananières: un enjeu économiquemajeur pour la sécurité alimentaire. Internationalsymposium, Douala, Cameroon, 10-14 November1988 (C. Picq, E. Fouré & E.A. Frison, eds). INIBAP, Montpellier, France.
Gowen S. & P. Quénéhervé. 1990. Nematode para-sites of banana, plantains and abaca. Pp. 431-460in Plant parasitic nematodes in subtropical and tropical agriculture (M. Luc, R.A. Sikora & J. Bridge, eds). CAB International,Wallingford, UK.
Guérout R. 1972. Relations entre les populations deRadopholus similis Cobb et la croissance dubananier. Fruits 27: 331-337.
Loos C.A. 1962. Studies on the life history and habitsof the burrowing nematode, Radopholus similis,the cause of blackhead disease of banana.Proceedings of the Helminthological Society ofWashington 29: 43-52.
Pinochet J. 1978. Histopathology of the root lesionnematode, Pratylenchus coffeae on plantains,Musa AAB. Nematologica 24: 337-340.
Sarah J.L., J. Pinochet & J. Stanton. 1996. The bur-rowing nematode of bananas, Radopholus similis
Cobb, 1913. Musa Pest Fact Sheets No.1. INIBAP,Montpellier, France.
Seinhorst J.W. 1950. De betekenis van de toestandvan de grond voor het optreden van aanstastingdoor het stengelaatje (Ditylenchus dipsaci
(Kühn) Filipjev). Tijdschrift Plantenziekten 56: 291-349.
Seinhorst J.W. 1959. A rapid method for the transferof nematodes from fixative to anhydrous glycerin.Nematologica 4: 67-69.
Stover R.H. 1972. Banana, plantain and abaca dis-eases. Commonwealth Mycological Inst., Kew,Surrey, England. 316pp.
The authors work at the Centre National de RechercheAgronomique (CNRA), 01 BP 1536 Abidjan 01, Côte d’Ivoire.
INFOMUSA — Vol 10, N° 2 27
Southeast
0
1
2
36 5
4
3
2
14
5
0 10 20 30 40 50 60 70 80 90 100
Frequency (%)
Ab
un
dan
ce
Midwest
0
1
2
3
3
1
2
4
4
5
0 10 20 30 40 50 60 70 80 90 100
Frequency (%)
Ab
un
dan
ce
1. Helicotylenchus multicinctus2. Meloidogyne spp.3. Radopholus similis
4. Pratylenchus coffeae5. Hoplolaimus pararobustus6. Rotylenchulus reniformis
Figure 1. Occurrence frequency and abundance indexes of nematode species associated with plantain in Côte d’Ivoire.
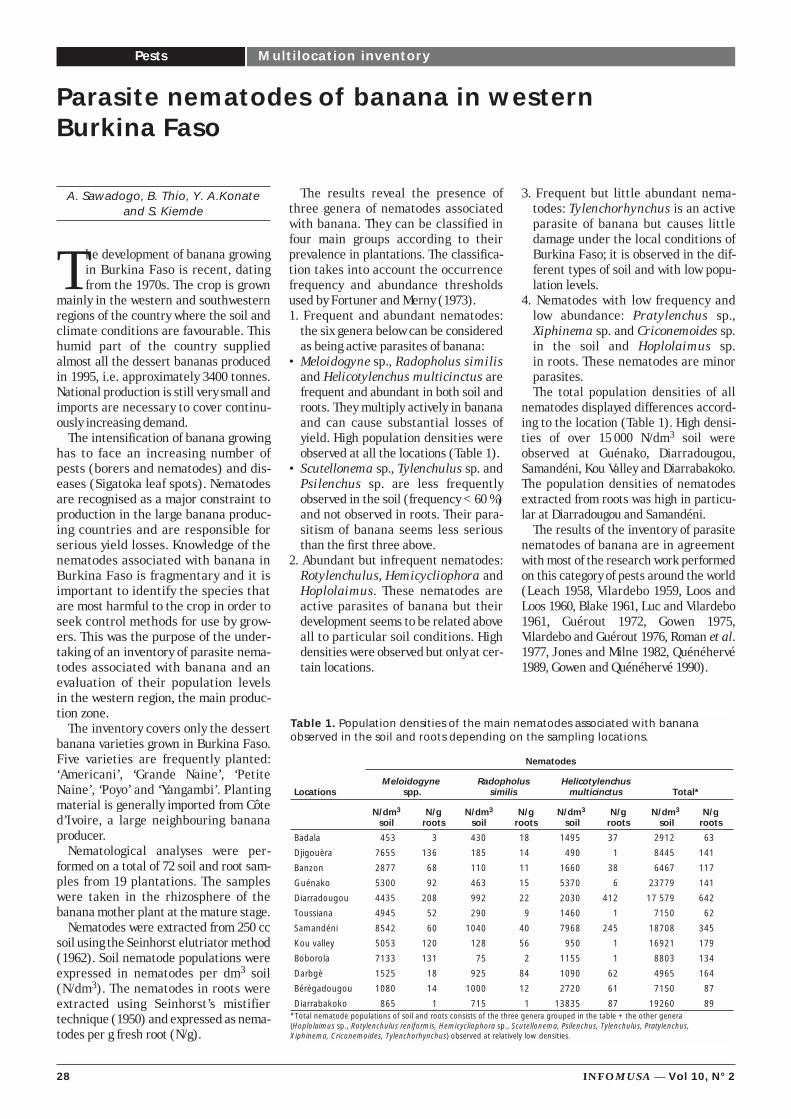
A. Sawadogo, B. Thio, Y. A.Konateand S. Kiemde
The development of banana growingin Burkina Faso is recent, datingfrom the 1970s. The crop is grown
mainly in the western and southwesternregions of the country where the soil andclimate conditions are favourable. Thishumid part of the country suppliedalmost all the dessert bananas producedin 1995, i.e. approximately 3400 tonnes.National production is still very small andimports are necessary to cover continu-ously increasing demand.
The intensification of banana growinghas to face an increasing number ofpests (borers and nematodes) and dis-eases (Sigatoka leaf spots). Nematodesare recognised as a major constraint toproduction in the large banana produc-ing countries and are responsible forserious yield losses. Knowledge of thenematodes associated with banana inBurkina Faso is fragmentary and it isimportant to identify the species thatare most harmful to the crop in order toseek control methods for use by grow-ers. This was the purpose of the under-taking of an inventory of parasite nema-todes associated with banana and anevaluation of their population levels in the western region, the main produc-tion zone.
The inventory covers only the dessertbanana varieties grown in Burkina Faso.Five varieties are frequently planted:‘Americani’, ‘Grande Naine’, ‘PetiteNaine’, ‘Poyo’ and ‘Yangambi’. Plantingmaterial is generally imported from Côted’Ivoire, a large neighbouring bananaproducer.
Nematological analyses were per-formed on a total of 72 soil and root sam-ples from 19 plantations. The sampleswere taken in the rhizosphere of thebanana mother plant at the mature stage.
Nematodes were extracted from 250 ccsoil using the Seinhorst elutriator method(1962). Soil nematode populations wereexpressed in nematodes per dm3 soil(N/dm3). The nematodes in roots wereextracted using Seinhorst’s mistifiertechnique (1950) and expressed as nema-todes per g fresh root (N/g).
The results reveal the presence ofthree genera of nematodes associatedwith banana. They can be classified infour main groups according to theirprevalence in plantations. The classifica-tion takes into account the occurrencefrequency and abundance thresholdsused by Fortuner and Merny (1973).1. Frequent and abundant nematodes:
the six genera below can be consideredas being active parasites of banana:
• Meloidogyne sp., Radopholus similisand Helicotylenchus multicinctus arefrequent and abundant in both soil androots. They multiply actively in bananaand can cause substantial losses ofyield. High population densities wereobserved at all the locations (Table 1).
• Scutellonema sp., Tylenchulus sp. andPsilenchus sp. are less frequentlyobserved in the soil (frequency < 60 %)and not observed in roots. Their para-sitism of banana seems less seriousthan the first three above.
2. Abundant but infrequent nematodes:Rotylenchulus, Hemicycliophora andHoplolaimus. These nematodes areactive parasites of banana but theirdevelopment seems to be related aboveall to particular soil conditions. Highdensities were observed but only at cer-tain locations.
3. Frequent but little abundant nema-todes: Tylenchorhynchus is an activeparasite of banana but causes littledamage under the local conditions ofBurkina Faso; it is observed in the dif-ferent types of soil and with low popu-lation levels.
4. Nematodes with low frequency and low abundance: Pratylenchus sp.,Xiphinema sp. and Criconemoides sp.in the soil and Hoplolaimus sp. in roots. These nematodes are minorparasites.The total population densities of all
nematodes displayed differences accord-ing to the location (Table 1). High densi-ties of over 15 000 N/dm3 soil wereobserved at Guénako, Diarradougou,Samandéni, Kou Valley and Diarrabakoko.The population densities of nematodesextracted from roots was high in particu-lar at Diarradougou and Samandéni.
The results of the inventory of parasitenematodes of banana are in agreementwith most of the research work performedon this category of pests around the world(Leach 1958, Vilardebo 1959, Loos andLoos 1960, Blake 1961, Luc and Vilardebo1961, Guérout 1972, Gowen 1975,Vilardebo and Guérout 1976, Roman et al.1977, Jones and Milne 1982, Quénéhervé1989, Gowen and Quénéhervé 1990).
28 INFOMUSA — Vol 10, N° 2
Parasite nematodes of banana in western Burkina Faso
Pests Multilocation inventory
Table 1. Population densities of the main nematodes associated with bananaobserved in the soil and roots depending on the sampling locations.
Nematodes
Meloidogyne Radopholus Helicotylenchus Locations spp. similis multicinctus Total*
N/dm3 N/g N/dm3 N/g N/dm3 N/g N/dm3 N/g soil roots soil roots soil roots soil roots
Badala 453 3 430 18 1495 37 2912 63
Djigouèra 7655 136 185 14 490 1 8445 141
Banzon 2877 68 110 11 1660 38 6467 117
Guénako 5300 92 463 15 5370 6 23779 141
Diarradougou 4435 208 992 22 2030 412 17 579 642
Toussiana 4945 52 290 9 1460 1 7150 62
Samandéni 8542 60 1040 40 7968 245 18708 345
Kou valley 5053 120 128 56 950 1 16921 179
Boborola 7133 131 75 2 1155 1 8803 134
Darbgè 1525 18 925 84 1090 62 4965 164
Bérégadougou 1080 14 1000 12 2720 61 7150 87
Diarrabakoko 865 1 715 1 13835 87 19260 89*Total nematode populations of soil and roots consists of the three genera grouped in the table + the other genera (Hoplolaimus sp., Rotylenchulus reniformis, Hemicycliophora sp., Scutellonema, Psilenchus, Tylenchulus, Pratylenchus,Xiphinema, Criconemoides, Tylenchorhynchus) observed at relatively low densities.

This first inventory of parasite nema-todes of banana in Burkina Faso revealsthat the nematode population densitiesat the different locations are higher thanthe threshold of economic damage. Thehigh profitability of the crop is leading toan increase in the area under banana inBurkina Faso. Studies in greater depth onthe damage caused to banana by nema-todes under the soil and climate condi-tions of Burkina Faso and which aim ateffective control methods easily used bygrowers are therefore necessary. ■
ReferencesBlake C.D. 1961. Root rot of bananas caused by
Radopholus similis (Cobb) and its control in NewSouth Wales. Nematologica 6: 295-310.
Fortuner R. & G. Merny. 1973. Les nématodes para-sites des racines associés au riz en BasseCasamance (Sénégal) et en Gambie. CahiersORSTOM Série Biologie 21: 3-20.
Gowen S.S. 1975. Improvement of banana yields withnematicides. Pp. 121-125 in Proceedings of the
8th British Insecticide and Fungicide Conference,1975, Brighton, United Kingdom.
Jones R.K. & D.L. Milne. 1982. Nematode pests ofbananas. Pp. 30-37 in Nematology in SouthernAfrica (D.P. Keetch & J. Heyns, eds). Dept. Agric.and Fisheries, Republic of South Africa. Sci. Bull.No. 400.
Guérout R. 1972. Relations entre les populations deRadopholus similis et la croissance du bananier.Fruits 127: 331-337.
Leach R. 1958. Blackhead toppling disease ofbananas. Nature 181: 204-205.
Loos C.A. & S.B. Loos. 1960. Preparing free bananaseed. Phytopathology 50: 383-386.
Luc M. & A. Vilardebo. 1961. Les nématodes associésau bananier dans l’Ouest Africain. Fruits 16: 205-219.
Pinochet J. 1987. La variabilidad de Radopholus
similis en banano en las diferentes regiones pro-ductoras del mundo. Pp. 175-182 in Memorias VIIReunion ACORBAT 23-27 Sept. 1985, San José,Costa Rica (J.J. Galindo & R. Jaramillo Celis,eds). Boletin Tecnico 121. CATIE, Turrialba,Costa Rica.
Roman J., X. Rivas, D. Oromas & J. Rodriguez. 1977.Further experiments on the chemical control ofnematodes in plantains (Musa acuminata x M. balbisiana, AAB). Journal of Agriculture ofUniversity of Puerto Rico 61: 192-199.
Quénéhervé P. 1989. Population of nematodes insoils under banana, cv. Poyo, in the Ivory Coast. 3.Seasonal dynamics of populations in mineral soil.Revue de Nématologie 12(2): 149-160.
Vilardebo A. & R. Guérout. 1976. Nematodes speciesof West Africa, Madagascar and Réunion, withsome comments on their biology. Nematropica 6: 53-54 (Abstr.).
Seinhorst J.W. 1950. De betekenis van de toestandvan de grond voor het optreden van aanstastingdoor het stengelaatje (Ditylenchus dipsaci
(Kühn) Filipjev). Tijdschrift Plantenziekten 56: 291-349.
Seinhorst J.W. 1962. Modification of elutriationmethod for extracting nematodes from soil.Nematologica 8: 117-128.
The authors work at the INERA research laboratories,Farako-Bâ Station, BP 403, Bobo-Dioulasso, BurkinaFaso. E-mail: [email protected]
INFOMUSA — Vol 10, N° 2 29
A. Rekha, K.V. Ravishankar, L. Anandand S.C. Hiremath
Bananas are one of the oldest culti-vated fruits. Being a highly evolvedcrop, many banana cultivars have a
number of synonyms resulting in a some-what confused taxonomical status ofmany cultivars. Simmonds (1962) con-cluded that the present day cultivarshave evolved by the hybridization of twomain species, M. acuminata and M. bal-bisiana, which were considered the maincontributors of A and B genomes respec-tively. All cultivars were classified intovarious genomic groups such as AA, AAA,AB, AAB and ABB respectively based on amorphological scoring method (Stoverand Simmonds 1987). The primary centreof diversity of wild M. acuminata is theMalayan area extending to Assam,Burma, Siam, Indochina and Philippines.The wild M. balbisiana are found alongwith M. acuminata and in peninsularIndia and western Ghats (Simmonds1962). Recently Shepherd and Ferreira(1992) identified cultivars derived fromhybridization with M. schizocarpa among
the Musa germplasm of Papua NewGuinea. A Philippine clone was consid-ered to be the result of an ancienthybridization between M. balbisiana andM. textilis. D’Hont et al. (2000) observedthe occurrence of M. schizocarpa and M. textilis genomes among the chromo-some complement of Musa cultivars.These observations reveal the complexityof the genomic structure of present daybanana cultivars and their evolution.
Morphological characterization hasbeen the major tool for classifying bananacultivars into different genomic groups.Efforts have been made to classifybananas and plantains using quantitativetraits. Swennen et al. (1995) studied thepattern of variability among African plan-tains. The variability and genetic diver-gence studies among Indian bananaswere conducted by Valsalakumari et al.(1985, 1993). Simmonds et al. (1990)reported the relationship among culti-vated bananas based on numerical taxon-omy. The results of these studies revealedthe relationships and helped to eliminatesome synonyms. Molecular techniqueshave proved to be a powerful tool inexploiting such situations and under-
standing the genetic relationships amongbanana cultivars and species. Such stud-ies were reported by Gawel and Jarret(1991), Gawel et al. (1992), Jarret et al.(1992, 1993), Kaemmer et al. (1992),Howell et al. (1994), and Bhat et al.(1994) among Musa species and culti-vars. In the present study, an effort hasbeen made to understand morphological,quantitative relationships and genomicrelations among the bananas cultivars ofSouth India along with the introducedcultivars of banana using D2 analysis andRAPD markers.
Materials and methods
Plant materialBanana accessions were selected fromthe field germplasm plot at the IndianInstitute of Horticultural Research,Hessaraghatta, Bangalore. Thirty-threeaccessions of banana were used for mor-phological studies and 28 accessions forthe RAPD studies.
D2 analysisMorphological observations wererecorded from the selected 33 accessions
Genetic and genomic diversity in banana (Musa species and cultivars) based on D2 analysis and RAPD markers
Genetic resources Classifying using molecular tools
from a field germplasm plot whichincluded 5 Musa species, 25 cultivarsbelonging to different genomic groups(Simmonds 1987) and 3 synthetichybrids. The plantation included 3 plants
per accessions with a planting distance of1.75 x 1.75 m and regular cultural prac-tices. The data on plant morphology,quantitative yield and fruit parameterswere recorded (Table 1). D2 statistics, a
measure for a group distance based onmultiple characters as proposed byMahalanobis, were calculated using SPARsoftware (Doshi and Gupta 1991).Grouping of varieties was done following
Table 1. Growth and yield parameters (mean).
Character*
Musa accessions 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
M. acuminata 153.30 1.70 13.70 7.00 3.00 1.40 30.70 21.00 7.00 116.70 2.30 6.20 1.50 5.60 23.70
M. a. ssp. malaccensis 123.30 2.70 15.30 11.00 2.30 1.50 20.30 28.00 7.30 97.90 3.00 7.50 2.10 6.00 21.70
Annara Kunnan 121.00 2.10 11.30 6.30 7.00 1.50 30.30 30.30 10.00 98.70 3.00 8.20 2.10 14.00 21.10
Pisang Lilin 122.30 1.40 11.70 10.00 5.70 1.60 48.00 31.30 6.00 70.70 4.40 13.10 2.30 35.20 24.70
Tongat 146.00 1.70 11.00 2.00 5.00 1.30 36.70 40.00 8.30 100.70 2.80 9.10 2.20 27.00 21.10
M. balbisiana (1) 144.00 3.80 22.00 7.00 11.30 1.80 60.00 50.00 9.70 125.00 15.20 13.40 4.20 104.10 20.10
M. balbisiana (2) 211.70 5.20 31.30 11.00 14.00 2.60 73.70 81.70 12.70 186.00 24.00 18.80 4.90 155.60 17.30
Ney Poovan 127.30 2.90 20.00 11.70 5.30 1.80 66.70 56.70 10.70 134.70 9.20 12.30 3.20 62.20 28.60
Elakki Bale 118.30 2.70 17.30 8.30 6.00 1.90 61.30 49.00 10.30 144.70 9.10 11.60 3.00 55.30 28.50
Kunnan 119.00 2.30 16.00 6.30 6.30 1.70 45.00 30.30 7.70 79.00 4.30 11.30 3.30 57.70 23.70
Rasa Kadali 112.30 2.90 19.00 5.70 5.30 1.70 41.70 52.00 11.00 138.70 5.30 9.10 2.70 36.80 24.20
Dwarf Cavendish 131.30 1.30 17.00 6.30 5.70 1.20 57.70 39.30 6.70 64.70 10.80 17.70 3.80 138.40 22.60
Gros Michel 141.00 2.90 20.30 6.00 6.30 1.70 58.30 41.00 6.00 71.00 8.80 16.30 4.30 129.30 24.20
Grande Naine 123.00 1.40 16.30 5.30 6.30 1.70 71.30 36.30 5.70 48.00 5.50 17.40 3.60 123.40 21.30
Robusta 132.30 2.40 19.30 7.00 7.00 1.90 65.00 58.70 7.30 83.70 16.20 20.40 4.20 168.40 23.70
Red Banana 144.70 2.90 23.00 2.70 4.30 2.00 65.30 33.70 4.30 45.00 7.90 15.30 4.40 149.50 23.00
Rasthali 116.00 3.00 19.00 7.30 6.70 1.90 61.70 36.00 6.30 78.00 6.60 14.10 3.60 83.80 26.40
N. Rasabale 119.30 2.90 20.70 8.00 4.30 1.20 40.00 46.00 7.30 79.00 6.30 13.00 3.30 71.00 27.50
Nendran 116.00 2.20 16.70 2.70 2.30 1.40 46.70 33.00 5.00 33.70 4.00 16.80 3.70 105.30 30.30
Velethan 124.30 2.90 20.70 3.30 2.00 1.60 58.00 41.70 5.30 55.70 8.60 20.10 3.90 159.80 30.30
Palayan Kodan 133.00 2.80 18.30 5.00 6.00 1.70 60.00 46.70 12.00 129.70 12.00 11.40 3.20 60.00 22.00
Ladan Small 174.00 2.70 20.00 4.70 4.70 1.80 62.00 40.00 6.30 73.70 6.50 12.70 4.00 87.70 25.30
Kullan 179.70 1.90 19.70 4.30 3.70 1.50 62.70 33.00 6.30 66.00 6.20 12.20 3.40 67.90 24.00
Virupakshi 173.00 2.70 18.70 3.70 2.30 1.90 61.30 37.00 6.30 75.30 8.50 11.40 3.30 63.10 27.00
Rajapuri 177.20 1.50 18.00 4.70 4.00 1.30 55.70 36.70 6.70 60.70 6.30 12.80 3.50 62.30 23.80
Muthia 128.70 3.00 22.70 6.00 6.30 1.80 65.00 49.30 7.30 83.30 12.90 15.90 4.80 151.20 21.70
Klue Tiparot 127.30 3.00 22.00 8.70 7.00 1.90 70.70 48.30 7.30 80.30 12.10 15.00 4.80 126.60 25.00
Cuba 117.30 3.00 20.00 4.70 8.00 1.80 65.70 38.70 6.00 61.70 13.00 15.90 5.30 142.20 22.90
Monthan 92.00 3.10 23.30 4.70 6.20 2.00 66.70 61.70 6.30 65.70 13.30 19.90 5.00 222.20 19.30
Karpooravally 130.70 3.30 25.30 7.70 8.00 2.00 65.20 53.00 10.70 170.00 18.30 14.10 3.50 97.10 26.70
M. a. x M. r. 200.30 1.50 11.30 7.00 7.30 1.20 43.30 27.00 8.30 78.30 1.80 1.30 1.90 2.20 0.00
M. a. x M. b. 201.30 1.50 11.30 2.00 6.70 1.10 47.70 27.70 7.70 86.00 1.40 1.30 2.50 2.10 0.00
M. a. x Kadali 140.70 1.60 14.70 7.30 5.00 1.40 51.30 28.30 6.30 70.70 4.50 9.70 2.60 32.80 28.30
Grand mean 140.10 2.50 18.40 6.20 5.80 1.60 54.10 41.60 7.70 89.50 8.30 12.90 3.50 85.00 22.70*Characters: 1. No. days from flowering to harvest; 2. Plant height (m); 3. Pseudostem girth (cm); 4. No. suckers; 5. No. leaves; 6. Leaf length (m); 7. Leaf width (cm); 8. Bunch length (cm); 9. No. hands; 10. No. fingers; 11. Bunch weight (kg); 12. Finger length( cm); 13. Finger girth (cm); 14. Finger weight ( g); 15. TSS (oBrix)
Table 2. Character means in different clusters of banana accessions.
Cluster
Character I II III IV V
1. No. days from flowering to harvest 139.31 200.83 130.28 112.41 211.67
2. Plant height (m) 2.29 1.53 2.76 2.79 5.20
3. Pseudostem girth (cm) 18.19 11.33 18.09 20.17 31.33
4. No. suckers 5.98 4.50 8.05 3.84 11.00
5. No. leaves 5.40 7.00 6.04 4.66 14.00
6. Leaf length (m) 6.65 1.14 1.67 1.46 2.63
7. Leaf width (cm) 57.84 45.50 45.00 51.75 73.67
8. Bunch length (cm) 40.58 27.33 41.57 48.75 81.67
9. No. hands 7.09 8.00 9.48 5.66 12.67
10. No. fingers 79.16 82.16 125.91 54.17 186.00
11. Bunch weight (kg) 7.97 1.57 8.06 9.73 24.00
12. Finger length (cm) 13.67 1.30 10.12 18.44 18.18
13. Finger girth (cm) 3.57 2.21 2.75 4.45 4.91
14. Finger weight (g) 88.97 2.13 46.54 157.38 155.56
15. TSS (oBrix) 24.41 0.0 23.72 25.73 17.33
30 INFOMUSA — Vol 10, N° 2

Tocher’s method as described by Singhand Chaudhary (1979). Average intra-cluster distances were calculated usingthe formula ∑Di
2/n where Di2 is the sum
of distances between possible combina-tions (n) of the populations included inthe cluster. Average inter-cluster dis-tances were calculated by taking eachcluster and their distances from the othercluster. The cluster diagram was drawnwith the help of square roots of D2 valuesshowing relationships within andbetween clusters (Figure 1). The charac-ter means of different clusters are shownin Table 2.
RAPD analysisTwenty-eight genotypes were selected forthe study, including 6 Musa species, 3 synthetic hybrids, introduced cultivarsand important Indian cultivars. The geno-types involved and their genomic classifi-cation are represented in Table 3. TheDNA was extracted from a mature leaf bythe modified CTAB method (Ravishankaret al. 2000). DNA concentrations were cal-culated by measuring the absorbance at260 nm with a UV spectrophotometer. PCR amplifications were made using 10 different random primers: OPA-03, OPA-04, OPA-10, OPA-13, OPE-01, OPE-08,OPE-18, OPE-20, OPQ-11 and OPQ-18(Operon Technologies, USA). The reactionvolume (25 ml) consisted of 0.3 mMprimer, 2.5 µM MgCl2, 0.5 u Taq-polymerase100 µM dNTPs and 50 ng of template DNA.The PCR was carried out with initialdenaturation at 93oC for 3 minutes and 40 cycles of 93oC for 2 sec., 35oC for 2 sec.,72oC for 1 min., final extension at 72oC for 8 minutes. The PCR products were sep-arated by electrophoresis on 1.5% agarosegel containing 0.5 µg/ml of ethidium bro-mide. RAPD bands were visualized underUV light and photographed. The RAPDbands were analyzed for each primer andscored as ‘0’ for absence of the band at aparticular position and as ‘1’ for presenceof the band (Table 4). The data obtainedwere analyzed using a cluster analysis pro-gram to form a dendrogram based on min-imum variance algorithm (Ward 1963)(Figure 2). Further, principal componentanalysis was done to group the genotypes(Figure 3).
Results and discussion
D2 analysisD2 analysis revealed 5 clusters (Table 5).The first cluster involved 19 cultivars.They were the edible fruit types of AA,AAA, AB, AAB groups and cultivar Muthiaof group ABB, which has smaller fruitscompared to the fruits of other ABB culti-
vars. Cluster II had only two genotypes,which were the synthetic hybrids ofcrosses M. acuminata x M. balbisianaand M. acuminata x M. rubra. Both haddwarf plant types with small inediblefruits with no pulp. Cluster III had 7 geno-types. The three wild M. acuminata typeswere included in this group with M. balbisiana (BB), Ney Poovan,Rasakadali (AB) and Karpooravally (ABB) .
Plant height and TSS seem to be majorfactors in this cluster. The fourth clusterinvolved four genotypes: Velethan,Nendran (AAB plantain), Cuba andMonthan (ABB). This group mainly represents large fruits. Cluster V had only one genotype, Bhimaithia, a wild M. balbisiana (BB) type collected fromAssam. They are highly robust plants withheavy bunches and large-seeded fruits.
Figure 1. Cluster diagram showing inter- and intra-distances.
Table 3. Musa species and cultivars for RAPD studies.
Sl No. Name Genomic group Geographical area Fruit type Variety No.
1 Musa acuminata AA IIHR, Bangalore wild V17
2 M. a. ssp. malaccensis AA ITC0609 wild V14
3 Annara Kunnan AA IIHR, Bangalore(Kerala) wild V21
4 Musa balbisiana (1) BB IIHR, Bangalore wild V26
5 Musa balbisiana (2) BB IIHR, Bangalore (N.E. India) wild V27
6 Musa rubra - N.E. India (Arunachal Pradesh) wild V2
7 Pisang Lilin AA IIHR, Bangalore dessert V15
8 Elakki Bale AB IIHR, Bangalore (Karnataka) dessert V18
9 Ney Poovan AB IIHR, Bangalore Kerala/Tamil Nadu dessert V19
10 Dwarf Cavendish AAA IIHR, Bangalore dessert V20
11 Robusta AAA IIHR, Bangalore dessert V28
12 Gros Michel AAA ITC1122 dessert V13
13 Grande Naine AAA IIHR, Bangalore dessert V22
14 Red Banana AAA IIHR, Bangalore (Kerala) dessert V23
15 Palayan Kodan AAB IIHR, Bangalore Kerala/Tamil Nadu dessert V3
16 Rasthali AAB IIHR, Bangalore (Tamil Nadu) dessert V7
17 Ladan Small AAB IIHR, Bangalore (Bihar) dessert V5
18 Kullan AAB IIHR, Bangalore (S. India) dessert V4
19 Virupakshi AAB IIHR, Bangalore (Tamil Nadu) dessert V6
20 Velethan AAB IIHR, Bangalore (Kerala) dessert V1
21 Cuba ABB IIHR, Bangalore (Cuba) cooking V9
22 Monthan ABB IIHR, Bangalore (Tamil Nadu) cooking V8
23 Muthia ABB IIHR, Bangalore (N. India) cooking V11
24 Karpooravally ABB IIHR, Bangalore (Tamil Nadu) cooking V10
25 Klue Tiparot ABB ITC0652 cooking V12
26 M. a. x M. r. - IIHR, Bangalore (synthetic hybrid) - V25
27 M. a. x M. b. - IIHR, Bangalore (synthetic hybrid) - V24
28 M. a. x Kadali - IIHR, Bangalore (synthetic hybrid) - V16
INFOMUSA — Vol 10, N° 2 31
The D2 clusters were based on thequantitative characters, the major con-tributory characters being finger length,TSS (oBrix), plant height, finger girth,number of days from the date of flower-ing to harvest, and pseudostem girth (Table 6). These characters differ widelyfrom AA wild to BB wild genotypes. Themajor group consisted of cultivars withaverage fruit size and moderate TSS indi-cating the preference for the taste andpalatability, which contributed to selec-tion and domestication. Two types of M. balbisiana were distinguished. The M. balbisiana (1) or fruitless variety or
Elovazhai of South India had smallerfruits and bunches, while the M. bal-bisiana (2) had larger fruits and muchtaller plants with large pseudostem girth.These two types were distinctly separatedinto two different groups.
The mean for parameters of variouscharacters calculated among differentclusters was highest in cluster V for allcharacteristics except TSS. It had onlyone accession, the wild M. balbisianaand the fruits are inedible, as they are fullof seeds. Cluster III had the highest num-ber of fingers. Cluster IV had the longestfinger size and also highest finger weight.
Clusters I, II and IV had medium-sizedplants with a good TSS.
Inter-cluster distances showed thatclusters II and IV were distant from allother clusters. Among the clusters involv-ing edible cultivars, clusters III and IVshowed the highest distance from theothers. Cluster III showed highest diver-gence with highest intra-cluster distanceof 12.0. Involving cultivars with desirablecharacters, disease resistance and highinter-cluster distances would result inhighly segregating generations in breed-ing programmes.
RAPD analysisGenomic relationship studies were con-ducted among 28 genotypes using RAPDmarkers. Among the 10 primers used for amplification, 6 primers (OPA-03, OPA-04, OPA-10, OPA-13, OPE-18 andOPQ-18) gave scorable polymorphicbands. The number of bands produced byeach primer with different genotypes var-ied. A total number of 60 bands were ana-lyzed using Squared Euclidian distanceand Ward’s minimum variance algorithm.The dendrogram (Figure 2) showed twomain clusters that differentiated all AAgenotypes from those of BB types. The BBgroup in turn had two nodes with bothwild M. balbisiana types in one groupand the cultivars Cuba, Monthan,Karpooravally and Muthia (ABB group)in another group that showed very closerelationships. The hybrid M. acuminatax M. balbisiana was placed betweenthese two groups. The cultivar GrosMichel (AAA) was placed along with thecultivars of group ABB. A similar observa-tion was also made by Bhat et al. (1994).
The larger cluster consisted of 20 geno-types, divided into two subgroups ofdiploids and triploids. The cultivarVelethan (AAB) showed a close relation-ship with wild M. acuminata. The culti-var Rasthali (AAB) was placed close toAB group cultivars Ney Poovan and ElakkiBale, which showed similar fruit types.
32 INFOMUSA — Vol 10, N° 2
Figure 3. Principal component analysis of RAPD-generated markers for banana cultivars (for equivalence names/numbers of varieties, see Table 3).
V1V17
V7V18V19V14V15V16
V2V25V12V23V20V22V28V21
V3V4V5V6V8V9
V10V11V13V24V26V27
0 20 40 60 80 100 120 140
Linkage Distance
Figure 2. Dendogram of banana cultivars constructed by cluster analysis of RAPD markers (for equivalence names/numbers of varieties, see Table 3).
Table 4. Number of polymorphic andmonomorphic bands obtained with various primers.
Primer No. polymorphic No. monomorphic bands bands
OPA-03 7 2
OPA-04 7 -
OPA-10 12 -
OPA-13 10 -
OPE-18 10 1
OPQ-18 11 -
Total 57 3

The two cultivars Elakki Bale and NeyPoovan of AB group were very close toeach other. They have similar morpholog-ical characters such as pseudostemcolour, height, fruit type and quality.Thus, it appears that they are confirmedas synonyms. The wild M. acuminata ssp.malaccensis (AA) was closer to the culti-var Pisang Lilin (AA). They were close toAAA group cultivars. The hybrid M. acuminata Kadali (AA) was alsocloser to Pisang Lilin of this cluster,showing its affinity to AA group cultivars.
The second part of this clusterincluded cultivars of groups AAA andAAB. The cultivars Dwarf Cavendish andGrande Naine (AAA) were highly related,with similar morphological characterssuch as pseudostem height, colour, fruittype and quality. They were also closer toRobusta (AAA), which represents theCavendish group. The cultivar KlueTiparot (= Klue Tiparad) (ABB) (Jennyet al. 1997) was found to be placed alongwith the cultivar Red Banana (AAA). Thereasons for this relationship needs fur-ther investigations. The cultivarsVirupakshi, Ladan Small and Kullan(AAB Pome subgroup) were very closelyrelated. They also had similar fruit andplant morphology except for Kullan, adwarf plant. The cultivar Palayan Kodan(AAB Mysore subgroup) was placedbetween AAA and AAB group cultivars.All the cultivars of ABB group were foundto be in one cluster. The cultivarsMonthan and Cuba (ABB) were close toeach other though they differed in theirfruit types: Monthan had large fruits,while Cuba had shorter and stout fruits.Karpooravally and Muthia were closelyrelated. The two M. balbisiana (BB)types were morphologically differentthough placed near each other in thedendrogram. M. balbisiana (1) had ashorter pseudostem and smaller fruitsthan M. balbisiana (2). The hybrid M. acuminata x M. rubra was placedalong with M. rubra, which was betweenAA and AAA groups. The wild M. rubra ofsection Rhodochlamys showed a closerrelation to M. acuminata group.
Principal component analysis alsorevealed a similar pattern of clustering ofAAB and AB cultivars in one cluster andalso cultivars of AA and AAA along withM. rubra, M. acuminata and M. acumi-nata ssp. malaccensis (Eigen value 58.5percent). The cultivars of ABB were in athird group and the M. balbisiana clones(BB) were in a separate group. The wildAnnara Kunnan (AA) was distinctly sepa-rated and the hybrid M. acuminata x M.balbisiana was between the ABB groupcultivars and wild M. balbisiana types.
The above studies showed the associa-tion of genotypes based on the genomicclassification of Simmonds. Some aspectsneed further investigations, as there wasconfusion regarding the grouping of culti-vars such as Velethan, Ney Poovan andRasthali from South India. They are saidto be AAB and AB group but are placedfar away from the BB group. It might bedue to the involvement of other Musaspecies in their genomes as reported byD’Hont et al. (2000) who showed theinvolvement of M. textilis and M. schizo-carpa genomes among the chromosomesof Musa cultivars. The three wild M. acuminata showed different affinitiesto some cultivars. M. acuminata(Aduthurai) was reported as M. acumi-nata ssp. burmanica by Simmonds andwas found in Western Ghats of South Indiaas per Bhaktavatsalu and Sathiamoorthy(1979). This was closer to Velethan (AAB)whereas M. acuminata ssp. malaccensiswas closer to Pisang Lilin. The wildAnnara Kunnan of Kerala was found to becloser to Cavendish cultivars. Closer rela-tionships were observed between culti-vars Kullan, Virupakshi, Rajapuri andLadan Small. Similar relationships were
observed between the cultivars of theCavendish group (Dwarf Cavendish, GrosMichel, Robusta) and also between Cubaand Monthan of ABB cooking types inboth methods of analysis. Irrespectivelyof the place of collection, the Cavendish(AAA group) cultivars and cookingbananas (ABB group) showed closer rela-tions. Pome and Mysore (AAB group) sub-groups were in one cluster.
D2 analysis using morphological char-acters revealed 5 clusters. These clustershad genotypes from different genomicgroups. This differentiated cultivarsmainly according to fruit size. RAPDanalysis grouped cultivars based on theirgenomic constitution. However, thegrouping of some cultivars such asVelethan, Ney Poovan and Rasthali needsfurther studies. Both D2 analysis andRAPD were able to differentiate two wildtypes of M. balbisiana, Bhimaithia andElovazhai.
AcknowledgementWe are thankful to Director IIHR, for
providing the facilities to carry out theabove work and to Mr G.K. Naidu whohelped in D2 analysis. ■
INFOMUSA — Vol 10, N° 2 33
Table 5. D2 clusters.
Cluster Number of genotypes List of of genotypes
I 19 Kullan (AAB), Virupakshi (AAB), Rajapuri (AAB), Ladan Small (AAB),M. acuminata ¥ Kadali (-), Dwarf Cavendish (AAA), Gros Michel (AAA),Kunnan (?), Robusta (AAA), Nanjangud Rasabale (AAB), Pisang Lilin (AA),Red banana (AAA), KlueTiparot (ABB), Grande Naine (AAA),Palayan Kodan (AAB), Tongat (AA), Rasthali (AAB), Muthia (ABB),Elakkibale (AB)
II 2 M. acuminata ¥ M. balbisiana, M. acuminata ¥ M. rubra
III 7 Karpooravally (ABB), Ney Poovan (AB), Rasa Kadali (AB), M. balbisiana (1)(BB), M. acuminata ssp. malaccensis (AA), M. acuminata (AA),Annara Kunnan (AA)
IV 4 Velethan (AAB), Nendran (AAB), Cuba (ABB), Monthan (ABB)
V 1 M. balbisiana (2) (BB)
Table 6. Contribution of various characteristics to divergence.
Sl. No. Character Frequency % contribution
1 No. days from flowering to harvest 45 8.5
2 Plant height 89 16.9
3 Pseudostem girth 33 6.2
4 No. suckers 5 0.9
5 No. leaves 2 0.4
6 Leaf length 3 0.5
7 Leaf breadth 3 0.5
8 Bunch length 8 1.5
9 No. hands 8 1.5
10 No. fingers 3 0.5
11 Bunch weight 12 2.7
12 Finger length 138 26.1
13 Finger girth 79 14.9
14 Finger weight 4 0.7
15 TSS 96 18.2

ReferencesBhaktavatsalu C.M. & S. Sathiamoorthy. 1979.
Banana clonal situation in India - A résumé. Fruits34(2): 99-105.
Bhat K.V, R.L. Jarret & Z.W. Liu. 1994. RFLP charac-terization of Indian Musa germplasm for clonalidentification and classification. Euphytica 80 (1-2): 95-103.
D’Hont A., A.Paget-Goy, J. Escoute & F. Carreel. 2000.The interspecific genome structure of cultivatedbanana, Musa spp. revealed by genomic DNA in
situ hybridization. Theor. Appl. Genet. 100: 177-183.Doshi S.P. & K.C. Gupta. 1991. Statistical package of
Agricultural Research Data Analysis. IARI, ICAR,New Delhi.
Gawel N. & R.L. Jarret. 1991. Cytoplasmic geneticdiversity in bananas and plantains. Euphytica52(1): 19-23.
Gawel N., R.L. Jarret & A.D. Whittemore. 1992.Restriction fragment length polymorphism(RFLP) based on phylogenetic analysis of Musa.Theor. Appl. Genet. 84(3-4): 286-290.
Howell E.C., H.J. Newbury, R.L. Swennen, L.A. Withers& B.V. Fort-Lloyd. 1994. The use of RAPD for identi-fying and classifying Musa germplasm. Genome 37: 328-332.
Jarret R.L., N. Gawel, A.D. Whittemore & S. Sharrock.1992. RFLP-based phylogeny of Musa species in
Papua New Guinea. Theor. Appl. Genet. 84(5-6):579-584.
Jarret R.L., D.R. Vuylsteke, N.J. Gawel, R.B. Pimental& L.J. Dunbar. 1993. Detecting genetic diversity indiploid banana using PCR and primers from a highly repetitive DNA sequence. Euphytica 62(1-2): 69-76.
Jenny C., F. Carreel & F. Bakry. 1997. Revision onbanana taxonomy: Klue Tiparot (Musa spp.)reclassified as a triploid. Fruits 52(2): 83-91.
Kaemmer D., R. Afrza, K. Weising, G. Khal & F.J. Novak.1992. Oligonucleotide and amplification fingerprinting of wild species and cultivars of banana(Musa spp.) Biotechnology 10(19): 1030-1035.
Ravishankar K.V., Lalitha Anand & M.R. Dinesh. 2000.Assessment of genetic relatedness among mangocultivars of India using RAPD markers. Journal of Horticultural Science and Biotechnology 75(2): 198-201.
Simmonds N.W. 1962. The evolution of bananas.Longman, London.
Simmonds N.W. & S.T.C. Weatherup. 1990. Numericaltaxonomy of the cultivated bananas. TropicalAgriculture (Trinidad) 67(1): 90-92.
Stover R.H. & N.W. Simmonds. 1987. Bananas (3rd
ed.). Longman, London.Swennen R., D. Vuylsteke & R. Ortiz. 1995.
Phenotypic diversity and patterns of variation in
West and Central African plantains (Musa spp.,AAB Group, Musaceae). Economic Botany 49(3):320-327.
Singh R. & B.D. Chudhary. 1979. Biometrical meth-ods in quantitative genetics analysis. KalyaniPublishers, New Delhi.
Shepherd K. & F.R. Ferreira. 1984. The PNG biologicalfoundation’s banana collection at Laloki PortMoresby, Papua New Guinea. Newsletter-RegionalCommittee for Southeast Asia (IBPGR) 8(4): 28-29.
Valsala Kumari P.K. & P.C. Sivaraman Nair. 1993.Genomic classification of Indian banana cultivars.Tropical Agriculture (Trinidad) 70(2): 162-164.
Valsala Kumari P.K., P.C. Sivaraman Nair & P.V. Prabhakaran. 1984. Genetic divergence inbanana. Agricultural Research Journal of Kerala23(2): 146-149.
Ward J.H. 1963. Hierarchic grouping to optimize anobjective function. Journal of the AmericanStatistical Association 58: 236-239.
A. Rekha, K.V. Ravishankar and Lalitha Anandwork at the Indian Institute of Horticultural Research(IIHR), Hessaraghatta, Bangalore 560 089, India. S.C. Hiremath works at the Department of Botany,Karnatak University, Dharwad 580 003, India.
34 INFOMUSA — Vol 10, N° 2
Banana in images
Information Iconographic database
G. Boussou
The assembling of iconographic col-lections on banana is a very oldapproach for specialists. P.H. Allen
already possessed a large collection ofphotographs taken from 1959 onwardsduring prospection missions in PapuaNew Guinea (Rosales et al. 1999). Otherresearchers such as R. Swennen (KUL,Leuven, Belgium) possess large quanti-ties of images gathered during theircareers. These iconographic collectionsare of inestimable value since they tracenot only the professional career of theauthors but also the evolution of researchtechniques, the biodiversity and variety ofbananas (Figure 1), the symptoms ofpests and diseases, the damage they causeand cultural practices in the differentregions of the world. Finally, these imagesare sometimes the only witness to rare orendangered cultivars (Touré 2000).
Certain researchers have given numer-ous images to the INIBAP Informationand Communication Unit so that theseresources can be used and illustrate pub-lications. The stock of images has grown
over the years and has become difficult tohandle without suitable classification.
ObjectivesSeveral objectives are involved in the cre-ation and updating of an iconographicdatabase: the illustration of publications(annual reports, INFOMUSA, etc.), therecording of history and the researchactivities of the various member bodies ofthe INIBAP network, the use of new tech-nologies and promotion targeting donorsand the general public on the occasion ofsymposia and exhibitions. Finally, thestorage of these images in an appropriateformat on an electronic storage mediumenables better archiving and enhancesexchanges. Classification and access tothe data is facilitated by computerizedquerying (Dollin Du Fresnel 2000).
The collection and processing of imagesThe slides, photographs or digital images(CD-ROM) are sent by the authors. Eachimage is accompanied by the followinginformation: the date and place of thetaking of the photo, the author and hisaffiliation, the title and comments. The
images area digitized in a high-resolutionformat (TIFF format) for subsequentprinting and publication. This format isused for the storage of the images onmagnetic tape and CD-ROM. Copies arethen made in a format with a lower reso-lution (JPEG format) for display usingthe Internet and for sending to users bye-mail for a preliminary selection.
The images are organized in 10 themes:Biodiversity, Plant improvement, Eventsand meetings, Markets, Postharvest, Fieldproduction, Pests, Diseases, Laboratorytechniques and Use. Processing the imagesincludes cataloguing using a Microsoft®Access data entry form. The content of the image is described with a choice ofkeywords (indexing) using the AGROVOCthesaurus and INIBAP’s own list ofdescriptors.
Classification makes access to informa-tion much easier. Thematic access toinformation enables preliminary sortingand prevents incoherent responses(Degez and Masse 2000). In contrast withtext documents in which search can beperformed in the reference, the textand/or the abstract, it is impossible toquery an iconographic document using a

portion of the image. It is thus importantto complement the images by a title, acomment and good indexing. Cooperationbetween researchers and informationprofessionals is essential in order togather a maximum amount of informationabout the real message carried by theimage and thus avoid subjectivity inindexing and hence in interpretation(Lissalde 1998).
The recorded data is stored in a centralbase (SQL server), thus keeping the prob-lem of duplicates and multiple versions ofthe base to a minimum. Display on theInternet is by means of the InternetInformation Service (IIS, Microsoft®)using Active Server Page technology(ASP, Microsoft®).
Searching and displaying of resultsThe queryable fields are the country inwhich the photograph was taken, theauthor, the cultivar and the keywords. Asingle word can also be sought in theentire reference. Work is in progress torefine searches and to improve the rele-vance of responses.
Search responses are displayed in twocolumns with a thumbnail version of theimage on one side and the title, theauthor and the reference of the image onthe other. A double-click displays a largerversion of a photo (in JPEG format) orthe entire description. The images cho-sen for subsequent use are available in TIFF format from the INIBAPInformation and Communication Unit.
The authors of the images referencedin the photo library do not receive any
payment or reward but mention of thename of the author and his affiliationmust accompany the images published.Readers who wish to contribute to theiconographic base managed by INIBAPcan send their images to Gaston Boussou,Information and Communication Unit.
AcknowledgementsINIBAP thanks all the researchers whohave contributed images to theInformation and Communication Unit.Special thanks go to R. Swennen and histeam for their availability and for thefinancial and technical resources mobi-lized for this project. ■
ReferencesDegez D. & C. Masse. 2000. L’indexation à l’ère
d’Internet. Documentaliste 37(2): 118-120.Dollin Du Fresnel M. 2000. Archivage électronique.
Documentaliste 37(2): 89-90.Lissalde C. 1998. Indigo base: la base d’images fixes
numérisées de l’Orstom. Documentaliste 35(1):43-50.
Rosales F., E. Arnaud & J. Coto (eds). 1999. A tributeto the work of Paul A. Allen: a catalogue of wild andcultivated bananas. INIBAP, Montpellier, France.
Touré K.K. 2000. Pour valoriser les fonds de la pho-tothèque de l’IFAN, mémoire visuelle de l’Afrique.Documentaliste 37(3/4): 174-181.
Gaston Boussou manages the image database at INIBAP headquarters.
INFOMUSA — Vol 10, N° 2 35
Figure 1. This is the only banana I know with very very slanting behaviour. This is the normal statureat maturity of the plant. (Comments from Rony Swennen, author of the picture).
Classification and characterization of Musa exotica,M. alinsanaya and M. acuminata ssp. errans
Genetic resources Collecting and discovering
R.V. Valmayor
Banana is native to Southeast Asiaand is the region’s greatest contri-bution to the tropical fruit industry.
From its center of origin, banana wasintroduced to Africa, Latin America, andthe South Pacific where it gained popular-ity and great economic importance. In thedeveloping countries of the world, cookingbananas serve as a staple food whiledessert bananas are the most affordable
fruit. Vast plantations have been estab-lished in the tropics and banana is shippedto distant temperate markets where thefruit enjoys strong and consistent demand.
But the commercial cultivars of bananasuccumb to serious diseases. For almost acentury, Gros Michel was the leadingexport variety until it had to be replacedby Cavendish clones in the mid-1960sbecause it was susceptible to Fusariumwilt. Cavendish banana later proved to besusceptible to the Sigatoka disease, forwhich existing control measures were
economically prohibitive and environ-mentally hazardous.
Every time a serious epidemic occurs, abanana breeding programme is launched.A vital part of any plant improvement pro-gramme is the assemblage of germplasmmaterials that provide sources for resis-tance genes. The favourite donors are thewild relatives that thrive under adverseconditions. The popular destination ofbanana collection missions is SoutheastAsia, the center of origin and diversity ofbananas.

The earlier banana improvement pro-gramme was based in Trinidad andJamaica in the former British WestIndies. The exploration missions pri-marily covered Malaysia, Thailand (for-merly known as Siam) and Myanmar(formerly Burma). The banana collec-tors concentrated on wild Musa acumi-nata Colla, the progenitor with closeaffinity to both Gros Michel andCavendish cultivars. The large assem-blage of the polymorphic M. acuminatawas segregated into various forms.These forms were later classified bySimmonds (1956) into five subspecies:(1) malaccensis (Ridl) Simmonds, (2) siamea Simmonds, (3) burmanicaSimmonds, (4) microcarpa (Beccari)Simmonds, and (5) banksii (F.v. Muell.)Simmonds. The characteristic that dis-tinguishes banksii is the presence ofhermaphrodite basal flowers ratherthan female flowers as commonlyobserved in the other Musa species.
The United Fruit Company and itsbanana collection teams sponsored thelater banana improvement programme,concentrating on Indonesia and thePhilippines. The germplasm materialsthey gathered were sent to the LancetillaPlant Introduction Garden in La Lima,Honduras. Wild M. acuminata and itsedible derivatives were again the centerof interest because wild M. balbisianaand its hybrids with M. acuminata intro-duce the gene responsible for starchinessin their hybrid progenies. Starchiness isan undesirable characteristic of bananasin the export market.
The banana germplasm resources ofVietnam had not been systematicallyexplored due to the country’s extendedstruggle for freedom. But in 1994,Vietnam’s Agricultural Science Institute(VASI) received a grant from theInternational Network for theImprovement of Banana and Plantain(INIBAP) to collect, conserve, and eval-uate indigenous Musa germplasm. Fiveprospection missions were launchedand a large collection was established atPhu Ho Fruit Research Center in VinhPhu province. Under the leadership ofMr. Le Dinh Danh, the author joined theexploration of the Cuc Phuong ForestReservation, collected and introducedM. exotica R. Valmayor sp. nov. to LosBaños, Laguna, Philippines. Much ear-lier (1959-60), the author was appointedas the Filipino counterpart of PaulAllen, leader of the United Fruit Co.banana prospection mission to thePhilippines. Together, they explored thewhole country, gathered all sorts ofMusa germplasm, and cooperated to
reestablish the national banana varietycollection of the University of thePhilippines Los Baños.
The common M. acuminata in thePhilippines with its perfect basal flowerclusters had earlier been associated withthe subspecies banksii (Brewbaker andGorrez 1956). But detailed characteriza-tion studies revealed major differences inplant, fruit and flower bract characteris-tics from the banksii found in Papua New Guinea, Samoa and northernQueensland, Australia. This finding ledAllen to suggest the reclassification of the Philippine acuminata under thesubspecies errans (Allen 1965). Theadoption of errans conforms with the pri-ority of usage principle as Fr. Blanco first applied the term errans for theacuminata of the Philippines in 1837whereas F.v. Muell. used banksii in 1863(Cheesman 1948). The adoption of M. acuminata Colla ssp. errans (Blanco)R. Valmayor, comb. nov., is the first effortto formalize the classification and regis-tration of the new subspecies.
Another major discovery of the bananaprospection missions in the Philippineswas that alinsanay was a distinct species.Peasant farmers had formerly mistakenalinsanay with wild abaca (M. textilisNee). Brewbaker and Gorrez (1956) latersurmised it to be a hybrid between abacaand the local saging matsing (M. acumi-nata). However, based on the results ofdetailed morphological and cytogeneticstudies, alinsanay significantly differsfrom abaca in plant, fruit, and flowerbract characteristics. Neither can alin-sanay be accepted as a hybrid of abacawhich has 10 pairs of chromosomes andsaging matsing which has 11 pairs ofchromosomes, because the expected ster-ile progenies are fully seeded during nor-mal meiosis. Simmonds rejected thehybrid theory, whereas Allen recom-mended elevation of alinsanay to a dis-tinct species. The author agrees withAllen, and this report is the first effort todescribe M. alinsanaya R. Valmayor sp.nov. as an indigenous Musa species in thePhilippines.
The botanical description of M. exoticais original while descriptions of alinsanayand saging matsing have been adaptedfrom Brewbaker and Gorrez (1956). Theaccompanying diagnostic descriptions(in Latin and English) rationalize theirnew status as species and subspecies andexplain why the given names wereselected. Photographs provide strong evi-dences of the uniqueness of the threewild banana species. A note on the cytol-ogy of M. exotica by Mendioro (2001)indicating 10 pairs of chromosomes sup-
ports its classification under sectionCallimusa.
Botanical descriptions of new Musa species and subspecies
Musa exotica R. Valmayor, sp. nov.(Figure 1)Plant stooling profusely; suckers vertical,originating close to parent plant; pseu-dostem slender 0.5-2 m high, 3-6 cm indiameter, light green with purplishblotches.
Leaf petioles green with sparse purpleblotches, 20-42 cm long, petiole canalopen with erect margins, slightly wingednot clasping the pseudostem, petiolemargins with purple lines. Leaf bladesoblong, up to 1.5 m long, 25-40 cm wide,upper and lower surface light green, dull,leaf base pointed, asymmetric, hardly anywax on surface, midrib green, leaf corru-gation striped, cigar leaf green. Bractleaves 2, basal portion orange-red, tipgreen, leaflike.
Inflorescence erect, its peduncle 10-15 cm long, smooth, yellow-green;flower bud ovoid, 10 cm long, 4-6 cm wide,bract colour orange-red on the outside,paler on the inside, apex yellow, bractsgreatly overlap, lifting 2 or more at a time,not revolute, smooth, not waxy, stronglygrooved.
Basal flowers female, 2-3 per bract,compound tepal yellow, 2.5-2.8 cm long,tip greenish, free tepal cream, closelyappressed to the large orange stigma.Stamens infertile, 5, short, inserted at thebase of the style. Ovary yellow, smooth,about 5 mm long, pedicel almost indistin-guishable from ovary.
Male bud in advanced flowering erect,10 cm long, 4 cm wide. Bracts orange-redon the external face, paler on the inter-nal face, tip yellow, greatly overlapping,several bracts lifted at any one time, per-sistent, turning brown and falling off at alater stage.
Male flowers 3 per bract; compoundtepal yellow-orange, lobe green, longitu-dinally grooved; free tepal yellow-orange, rectangular, boat-shaped,slightly longer than compound tepal.Fertile stamens 5 as long as compoundtepal, filaments creamy, translucent.Anthers yellowish orange; style yellow-ish; stigma orange. Mature male flowersbrown, persistent at its early stages,dehiscent at maturity.
Fruits yellow, few, 10-22, uniseriate,perpendicular to stalk, 2-3 per hand,small, 2-3 cm long, 3-4 cm wide, straight,seedless, angular, apex blunt; style per-sistent, brown at maturity.
36 INFOMUSA — Vol 10, N° 2

Original accession collected from theCuc Phuong Forest Reservation, NinhBinh province, Vietnam on February 17,1995. Important physiographic data:Latitude, 20°14’- 20°24’ North; Longitude,105°29’-105°44’ East; Elevation, 200-650 m; Average rainfall, 2157 mm;Temperature, 24.7°C (max 39°C, min16.7°C); Soil type, Argillite – Quartz, yel-low feralite.
Holotypus: Living accession now grow-ing in the garden of the author in LosBaños and at the Horticulture Nursery,University of the Philippines Los Baños.Herbarium specimen held at theDepartment of Botany, College of Artsand Sciences, University of thePhilippines Los Baños, CAHUP.
Musa alinsanaya R. Valmayor, sp.nov. (Figure 2)Plant stooling; pseudostems 2.5-4.0 mhigh, 20-30 cm in diameter at base, deepreddish purple; leaf sheath and petiolesglaucous, green.
Leaf blades oblong, 3-4 m long, 40-55 cm wide, truncate at apex, rounded atbase, with unequal basal lamina, green,glaucous, midribs green; petioles 50-60 cm long, their margin almost erect,leaving an open adaxial channel, ratherupright, closely appressed to the pseu-dostem.
Inflorescence subhorizontal, its pedun-cle and rachis thickly pubescent withbrown hairs, flowers in the 10-16 basalhands female, upper hands male.
Female flowers from 12-23 per hand(average 16) in two rows; compound tepalwhite, tips green with filiformappendages; free tepal white, about halfas long as compound tepal.
Male bud in advanced blooming ovoid,more or less acute, the bracts imbricate,outer surface green with purple edges,about two-thirds of the length of the bud,not conspicuously ribbed, subacute attips, glaucous, inner surface deep reddishpurple at tip and gradually becominglighter at basal end. One bract lifted at atime, deciduous.
Male flowers per bract from 8-12 (aver-age 10) in two rows 5-6 cm long; com-pound tepal 4.0-4.8 cm long, with an aver-age of 4.5 cm; white, tips green with 3filiform appendages about 3-4 mm long,the center filiform usually a little bitshorter; free tepal 2.5-3.0 cm long with anaverage of 2.7 cm, 1.0-1.3 cm wide with anaverage of 1.1 cm, boat-shaped, white,apiculate.
Fruit bunch asymmetrical, fruits andpedicels exhibiting marked geotropicreflex. Individual fruits very seedy, 10-12 cm long, 2.0-2.3 cm in diameter, con-spicuously 4-5 angled, abruptly narrowingat the base into a pedicel of 1.5 cm long,and at the apex into a conspicuous acu-
men; pericarp about 2 mm thick, green atripening, pulp nearly absent, white; fruitsheavily seeded.
Seeds black, flattened, more or lesssmooth, 3-4 mm in diameter and 5-7 mmhigh.
The description of alinsanay was basedon plants growing along roadsides in theBicol National Park between Daet,Camarines Norte Province and Sipocot,Camarines Sur Province, Philippines.Original description by Dr James L.Brewbaker and David D. Gorrez, formerlyof the University of the Philippines LosBaños.
Holotypus: Living accession now main-tained at the Abaca Research Center,Leyte State University in Baybay, LeyteProvince, Philippines. Herbarium speci-men held at the same institution.
Musa acuminata Colla ssp. errans(Blanco) R. Valmayor, comb. nov.(Figure 3) Basionym: Musa troglodytarum L. var.errans Blanco, Fl. Filip. 247. 1837.
INFOMUSA — Vol 10, N° 2 37
Figure 1. Musa exotica R. Valmayor sp. nov.
Figure 2. Musa alinsanaya R. Valmayor sp. nov. (Original photographs from Brewbaker and Gorrez).
Figure 3. Musa acuminata Colla ssp. errans (Blanco) R. Valmayor ssp. nov.

Plant stooling freely, habitat sylvan;pseudostems 3.0-4.5 m high, 25-30 cm indiameter at height of 50 cm, green ormore or less purplish, black-brown withage; leaf sheaths and petioles, glaucous,green. Fiber strips poorly, longitudinallyweak, stems transversely very tough.
Leaf blades oblong, 2.5-4.0 m long, 50-70 cm wide, truncate at tip, rounded atbase, with basal lamina equal, green,glaucous, mid-ribs green; petioles 50-90 cm long, their margins almost erect,leaving an open adaxial channel, ratherlax, closely appressed to pseudostemwhere margins flare and become more orless blackened and scarious.
Inflorescence subhorizontal or pen-dent, its peduncle and rachis thicklypubescent with short brown hairs, basalflowers hermaphrodite, the number ofhands 15-17, upper hands male. Spadixacute, green, 50-60 cm long.
Perfect flowers 20-28 per bract in tworows; 2 or 3 stamens perfectly developedin flowers of all basal bracts, sheddingabundant pollen, remaining stamensmore or less rudimentary (staminodal);development of staminodes into well-formed stamens taking place in the api-cal, 5-7 hands preceding the male flow-ers. Subtending bracts long (30-35 cm)narrow (9-11 cm), green, conspicuouslyribbed longitudinally, transversely corru-gated between the ribs, compound tepalabout 3.4 cm long, 1.2 cm wide, translu-cent, yellow-tipped, deeply lobed, theouter two lobes acute and 10-11 mm long,central three lobes fused with outer twoof these appearing as serrations about 4 mm from tip of central lobe; free tepaltranslucent, boat-shaped, rounded at tipwith faint shoulders, about half as long ascompound tepal; stigma broad, flattened,slightly bifurcate, upper part of style red-speckled; ovary pale green, distinctly 3-5 angled, glabrous above, minutely pubes-cent at base.
Male bud in advanced blooming ovoidto top-shaped, acute, the bracts convo-lute, reaching the tip of bud; bracts paleyellowish, green turning red-brown onfading, longitudinally ribbed, trans-versely corrugated between the ribs,acute at tips, glaucous, inner surface lightgreen. One bract lifted at a time, not con-spicuously revolute on fading, deciduous.
Male flowers about 20 per bract in tworows; tepals similar in length and colourto those of female flowers; compoundtepal yellow at tip, lobed, with outer lobesonly 4-6 mm long tipped with a short fila-ment, inner 3 lobes equal in length, 3-4 mm long; free tepal boat-shaped, acutelytipped with more or less prominent shoul-ders; stamens at first about as long as com-
pound tepal, later exserted, colourless,stigma and ovary greatly reduced.
Fruit bunch asymmetrical if borne sub-horizontally, regularly symmetric whenpendent; the 300-350 fruits and pedicelsexhibiting marked geotropic curvature.Individual fruits very seedy, 8-12 cm long,1.3-1.6 cm in diameter, conspicuously 4-5angled when immature but only obso-letely angulate when mature, abruptlynarrowed at the base into a pedicel of 1-1.5 cm (minutely pubescent) and at theapex into a conspicuous acumen 6-8 mmlong; pericarp about 2 mm thick, yellow-green at ripeness, pulp nearly absent,whitish.
Seeds dull black, commonly more orless warty, irregularly angulate, stronglyflattened, 6-8 mm broad and 3-4 mm high.
The description of saging matsing wasbased on plants growing on the slopes ofMt. Makiling, Los Baños, Laguna,Philippines. Original description by Dr James Brewbaker and David D. Gorrez,formerly of the University of thePhilippines Los Baños.
Neotypus: M. acuminata ssp. errans isnatural to the forest of Mt. Makiling.Since the original material used by Fr. Blanco is no longer available, freshspecimens were collected from Mt.Makiling. Herbarium specimens held atthe Department of Botany, College of Artsand Sciences, University of thePhilippines Los Baños, CAHUP.
Diagnostic characters of thethree wild bananas in Latinand English
(In Latin). M. exotica R. Valmayor, sp.nov. A musis speciebus popularis orna-mentalis aliis differt alabastro masculinovalde imbricato pulchro aurantiorubro.Hoc alabastrum masculinum ab illomusarum pictarum aliarum bracteis sin-gulis ad apicem flavissimis et distaleviridis discedent. Tepalum compositumfloris masculi colore vivide flavo-auranto,ad apicem lobis viridibus notat. Exspecibus aliis seriei Callimusae fructibusjuvenibus flavissimus ad pedunculumerectum perpendicularibus Musa exoticapraeterea distinguit. Epitheton exoticapulchritudinem et singularitatem specieinovae indicat.
(In English). M. exotica R. Valmayor isdifferentiated from other popular orna-mental bananas by its attractive, orange-red, male bud. The markedly imbricateorange-red, male bud is distinguishedfrom other colourful bananas by thebright yellow apices of individual bractsthat are lined with a streak of green atthe distal end. The compound tepal of themale flower is characterized by a bright,
yellow-orange colour, punctuated at itstip by green lobes. M. exotica can furtherbe differentiated from other species inthe Callimusa series by the bright yellowcolour of immature fruits that areattached perpendicular to the erect stalk.The term exotica was selected todescribe the beauty and uniqueness ofthe new species.
(In Latin). Musa alinsanaya R.Valmayor sp. nov. A Musa textilis Nee(nomen commune: abaca) et ceterospeciebus seriei Australimusae coloribusbractearum extus viridibus autem intusatrorubropurpureis distinguenda. Colorruber basin versus bractearum gradatimdecrescens. Ex abaca, quae fructus com-parate perpauciores grandioresqueinfructescentibus subhorizontales exori-entes ferens, alinsanay fructibusnumerosibus parvis geotropis infructes-centibus pendentibus etiam recedit.Musa fehi Bert. ex Viell. (nomen com-mune: Fe’i banana) fructibus grandibusesculentis infructescentibus erectibusvalde distinguit. Musa peekeliiLauterbach (nomen commune: pakel)differt coloribus bractearum et bracteismultis (singillatim in alinsanay) simul-taneis elevatis. Ephitheton alinsanayanomen vernaculum alinsanay perpetuat.
(In English). M. alinsanaya R.Valmayor sp. nov. is distinguished fromabaca, M. textilis Nee and other speciesin the Australimusa series by the greenouter surface but deep reddish purpleinner surface of its bracts. The red col-oration gradually becomes lighter towardthe base. The species can also be differ-entiated from abaca by the large numberof small, geotropically reflexed fruitsborne on pendent bunches in contrast tothe fewer but larger fruits arising fromsubhorizontal bunches of abaca. It is dis-tinctly different from Fe’i bananas, M. fehi Bert. ex Viell., which bears largeand edible fruits on erect, uprightbunches. M. alinsanaya also differs frompakel, M. peekelii Lauterbach, because ofits bract color and the habit of lifting onlyone bract at a time in contrast to pakelwhere many bracts lift simultaneously.The term alinsanaya was adapted to per-petuate its vernacular name.
(In Latin). Musa acuminata Colla ssp.errans (Blanco) R. Valmayor, comb. nov.
Basionym: Musa trogloditarum L. var.errans Blanco, Fl. Filip. 247, 1837
Differt a subspeciebus aliis Musaeacuminatae floribus hermaphroditis infasciculis florum basilibus contentis.Subspecies banksii Papua New Guineae,Samoae, et Queenslandiae borealis hunccharactere insolitem etiam praebentem,autem subspeciem errans infructescen-
38 INFOMUSA — Vol 10, N° 2

tibus usque ad 26 manis unaquaequeusque ad 25 fructibus parvis congestispraeditis. Ex contrario, infructescenssubspeciei banksii 10-14 manisunaquaeque 13-17 fructibus relativegrandioribusque laxioribus praedita.
(In English). M. acuminata Colla ssp.errans (Blanco) R. Valmayor comb. nov.
Basionym: Musa troglodytarum L. var.errans Blanco, Fl. Filip. 247, 1837.
This subspecies is differentiated fromother acuminata subspecies by the pres-ence of hermaphrodite flowers in itsbasal flower clusters. This characteristicis unique also in one other acuminatasubspecies, namely, banksii of PapuaNew Guinea, Samoa and northernQueensland, Australia. However, the sub-species errans differs from banksii inhaving an extended fruit bunch bearingup to 26 hands with as many as 25 small,densely packed fruits per hand. On theother hand, the fruit bunch of banksiionly produces 10 to 14 hands with each
hand bearing 13 to 17 fruits that arelarger and less compact than those oferrans. ■
ReferencesAllen P.H. 1965. Annotated index of Philippine
Musaceae. Philipp Agric 49: 320-411.Brewbaker J.L.& D.D. Gorrez. 1956. Classification of
Philippine Musae III. (a) Saguing matsing (Musa
banksii F.v.M.); (b) Alinsanay, a putative hybridof M. textilis and M. banksii. Philipp Agric 40: 258-268.
Cheesman E.E. 1948. Classification of the bananas.III Critical notes on species. (b) Musa Colla. KewBull. No. 1, pp. 17-28. (d) Musa banksii F.v. Muell.Kew Bull. No. 2, pp. 154-157.
Mendioro M.S. 2001. Cytology of Musa exotica
R. Valmayor sp. nov. Phil Agric Scientist 84(3): 245.Simmonds N.W. 1956. Botanical results of the banana
collecting expedition 1954-1955. Kew Bull 3: 463-489.
This article has already been publishedin THE PHILIPPINE AGRICULTURAL
SCIENTIST Vol. 84 No. 3, 325 – 331,September 2001 - ISSN 0031-7454
Classification and Characterization ofNew Musa sp. and ssp.
R.V. ValmayorTHE PHILIPPINE AGRICULTURAL
SCIENTIST kindly authorized its full-textpublication in this issue of INFOMUSA.
Ramon V. Valmayor is President, PhilippineAgriculture and Resources Research Foundation,Incorporated (PARRFI), c/o the Philippine Council forAgriculture, Forestry, and Natural Resources Researchand Development (PCARRD), Paseo de Valmayor, Los Baños, Laguna, Philippines (e-mail: [email protected]). Dr Valmayor is also former Professor, Department ofHorticulture, College of Agriculture, University of thePhilippines Los Baños (UPLB), 4031 College, Laguna,Philippines, former Director General of PCARRD andformer Regional Coordinator of the Asia and PacificNetwork (ASPNET) of INIBAP.
INFOMUSA — Vol 10, N° 2 39
Diversity of Radopholus similis (Cobb, 1893)(Nematoda : Tylenchida)Dept. of Biology, Faculty of Sciences, University of Gent, June 2000
Gamal Abdalla Elbadri
The variability of 19 populations ofRadopholus similis collectedworldwide was studied from various
angles. Morphological and morphometri-cal variations were found within andbetween populations. Variations occurrede.g. in total body length, number of lipannules, tail shape and length.Characters separating R. citrophilusfrom R. similis were overlapping betweenthe species. HinfI, NdeII and TaqI of ITSregions did not separate the populations.A group composed of populations fromKarkoug, Sennar, Chendi (Sudan),Calathea and Indonesia was visualized by AluI, RsaI and Tru9I restrictions.Sequencing of the ITS separated thesame group by 10-24 substitutions. Thesame five populations were clustered sep-arately for all combinations of alignmentsand tree construction methods. RAPDanalysis resulted in 179 scored bandsfrom 10 decamer primers. OPA3 distin-guished the five populations. The dendro-gram constructed on the basis of the
RAPD results placed the five populationsin one main cluster.
Reproductive fitness was found to varyamong populations and was controlled by the temperature. At 15°C only threepopulations originating from ornamen-tals reproduced. Maximum reproductionwas observed at 25°C. At this tempera-ture populations from Chendi andCalathea showed the highest reproduc-tion; the lowest reproduction wasobserved for populations from Indonesiaand Australia. At 30°C the reproductiondeclined. Juvenile reproduction obtainedin single female tests ranked the popula-tions in the same way. Populations dif-fered also in number of reproducingfemales and in the male:female ratio.
When inoculated on banana,Anthurium andreanum, or Marantaamabilis the populations showed a spe-cific relationship to their host of origin.Populations isolated from ornamentalsand black pepper multiplied well on M. amabilis and A. andreanum but noton banana. Populations isolated frombanana (with exception of populationsfrom Chendi, Sennar, Kamlin and
Karkoug) multiplied poorly on the orna-mentals. Four banana cultivars weretested for five populations. The cv.Yangambi Km5 proved its resistance to allof the populations; cv. Gros Michel wasfound to be moderately resistant. PisangJari Buaya was found to be as susceptibleas Grande Naine. None of the populationsdid multiply and inflict damage to Citrussinensis. The results obtained from themolecular studies correlate with the bio-logical observations. ■
Estimation of root developmentfrom shoot traits in plantain andbanana (Musa sp.) p. 15. We apolo-gize for the error in the list of authors which should read : G. Blomme, R. Swennen, A. Tenkouano, R. Ortizand D. Vuylsteke.
Erratum in previous issue(INFOMUSA Vol. 10, No. 1)
Thesis

Musanews
Africa
CRBP becomes CARBAP In February 2001, the Centre africain derecherches sur bananiers et plantains(African centre for banana and plantainresearch - CARBAP) was created throughan intergovernmental agreement signedafter the 2nd West and Central AfricanConference of Ministers of Research andDevelopment (COMRED/AOC).
Following the declaration made at thefirst Conference of the COMRED/AOC inYaoundé (12-16 January 1999), announc-ing the regional status of the Centrerégional de recherches sur bananiers etplantains (CRBP) for the benefit of allresearch institutions within West andCentral Africa adhering to its objectivesand its missions, the Governments ofCameroon, Central African Republic,Democratic Republic of Congo, Gabonand Equatorial Guinea agreed to trans-form CRBP into an African ResearchCentre for Bananas and Plantains(Centre africain de recherches surbananiers et plantains - CARBAP).
CRBP was established at Njombé(Cameroon) by a convention signed onthe 25 October 1989 between the Ministryof Scientific Research and Technology(MINREST) of the Republic of Cameroonand the Centre de coopération interna-tionale en recherche agronomique pourle développement (CIRAD, France). Thenew centre, CARBAP, will automaticallyinherit the overall CRBP patrimony.
CARBAP is mandated to improve theproduction of plantains and other locallyconsumed bananas to meet the needs of agrowing population through the develop-ment of a sustainable and environmen-tally friendly agriculture. In order toachieve this aim, CARBAP has a triplefunction of research, development andtraining.
This centre which has a regional man-date, will develop its activities taking intoaccount the different aspects of the com-modity chain (production, commercial-ization and transformation) as well aswith the various participants in this chain(producers, operators and consumers).CARBAP is therefore mandated toaddress the various regional problems aswell as agronomic and economic con-straints which limit the production ofbananas and plantains in the countrieswithin the two sub-regions of West andCentral Africa.
On 11 June, 2001, Dr Kodjo Tomekpe,Musa geneticist, was nominated asDirector of the new centre during the
transition period which should concludewith the setting up of the centre’s newmanagement structure in 2002.
As part of his new function, Dr Tomekpewill strengthen links with the nationalAfrican agricultural research systems andinternational organizations as well asenhancing contacts with donors.
Musa collecting expedition in TanzaniaA Musa germplasm collecting expedition,involving Dr Deborah Karamura (INI-BAP), Dr Edmond De Langhe (KULeuvenUniversity, Belgium) and Dr AlliyMbwana (ARI, Tanzania) took place inTanzania in July. The expedition wasorganized by INIBAP, with support fromIITA and the Rockefeller Foundation. Theexpedition was motivated by (1) a seriouslack of knowledge about banana cultivarsin the highlands of East-Africa other thanthe Great Lakes region, and (2) the grow-ing risk that unexplored yet potentiallyimportant material for genetic improve-ment could be lost due to changing farmmanagement practices in the region. Theexpedition covered the slopes of theKilimanjaro and the Usambara Hills and,to a lesser extent, the South Pare Hills.
The expedition proved to be most suc-cessful. Twenty-one interesting cultivarswere collected, of which 10 are mostprobably newly acquired diploids of greatpotential interest for genetic improve-ment of the African Highland Bananas.An entirely new group of AAA-triploidswas identified, which was called the‘Ilalyi’-group by the team. No componentof this group has ever been detectedbefore in the Great Lakes region.Furthermore as a result of linguistic stud-
ies carried out by the team, the hithertorather obscure nomenclature/synonymyof the cultivars in this region has beensubstantially clarified. No wild (seedy)diploids were found, and the chances thatsuch varieties exist on the continent arenow considered extremely remote. Foreach accession collected, three suckerswere obtained and these have beenplanted at the Tengeru HorticulturalResearch Station, under the supervisionof Dr A.S.S. Mbwana.
As a result of the collecting work, theteam proposed a theory of how AfricanHighland bananas in general may havebeen generated, with significant implica-tions for genetic, as well as cultural andhistorical aspects. The team was also ableto collect much information regardingspecific banana management practicesand utilization processes. Several produc-tion practices and many attractive prepa-rations –of immature and/or mature fruits- that do not exist in the Great Lakesregion were encountered. On the otherhand, many of the preparations typical forthe latter region are apparently not knownin the visited zones. The team noted theconsiderable cultural enrichment whichcould be achieved through mutual intro-duction of these practices and uses.
A number of recommendations weremade by the collecting team. These areas follows:• That the term ‘AAA-EA’ sensu stricto
(AAA-EA s.s.) be applied only to theAAA-EA of the Great Lakes region, aspreviously identified and described(Karamura 1999), and that the term‘AAA-EA’ sensu lato (AAA-EA s.l.) beapplied to cover all of the AAA-EA cul-tivars in the broadest sense.
40 INFOMUSA — Vol 10, N° 2
Drs D. Karamura and M. Mbwana learning from a farmer during their collecting mission in Tanzania.

• That the same team organize a briefvisit to the field collection at fruitingstage (about August 2002), in order tofinalize the identification and classifi-cation of the accessions. The results ofthis task would form the base for dupli-cation at international level of thedefinitively identified new accessions.
• That a second, similar expedition beeffected in the Taita Hills and theGikuyu area in Kenya, and a third oneon the highlands in Mbeya andMorogoro in Tanzania
• That local agronomy officers explorefurther for new cultivars in the zonevisited by the team, after getting dulyacquainted with the basic diversity asit can be studied at the Tengeru stationfield collection.
• That the ploidy of all the presumeddiploids collected be assessed throughflow cytometry/chromosome countingat the earliest possible convenience.
ReferenceKaramura, D.A. 1999. Numerical taxonomic studies of
the East African Highland bananas (Musa AAA-East Africa) in Uganda. (PhD Thesis, Dept. ofAgricultural Botany, University of Reading, January1998). INIBAP, Montpellier, France. 192 pp.
Copies of the full report of the collecting
expedition are available on request from INIBAP
headquarters.
Evidence of banana cultivation incentral Africa 2500 years agoBanana phytoliths found in refuse pits inarchaeological sites around Yaoundé,Cameroon, have been dated to 2500 BPaccording to researchers at theUniversity of Yaoundé and the RoyalMuseum of Central Africa in Belgium.This is the earliest unequivocal proof ofagriculture in central Africa, indicatingthat Musa species moved out of Asia, thecentre of origin of the genus, and acrossthe Indian Ocean a millennium earlierthan previously thought.
The phytoliths, microscopic particlesof silicon found on plant leaves, werefound in Nkang, one of the large rainfor-est settlements under study, along withpotsherds, polished stone tools, iron slag,charcoal, charred Canarium and Elaeisnuts and occasional animal bones. Theywere present at the bottom of a refuse pitand encrusted on the base of a vessel. Thepit was dated to 2490-2400 BP accordingto the 14C content of three charcoal sam-ples, showing strong correspondencewith the dates of other similar sites in thearea.
Several species of Ensete, the onlyother genus in Musaceae, are endemic totropical Africa. A comparative study of
the phytoliths of Musa and Ensete wascarried out with material from the RoyalBotanical Garden of Meise andKatholieke Universiteit Leuven both inBelgium. Both genera have phytolithswith a characteristic volcano shape.However, Musa species are distinguishedby the concave nature of the slope, protu-berances from the basal part and asmooth but sometimes crenated rim. Thephytoliths from the refuse pit showed fea-tures characteristic of Musa.
Edible bananas are seedless and theirpropagation outside their native area isnecessarily anthropogenic. This is thefirst substantial evidence to supposethat rainforest-dwellers were practisingcultivation in humid tropical Africa morethan 2000 years ago. This early farmingof plantains provides context to the pre-sent-day existence of a secondary centreof diversity of plantain in West Africa andmay explain the increase in village den-sity in the forest environment at thistime and the early expansion of Bantupeople. It also potentially agrees with DeLanghe et al. (1996) hypothesis, thatplantains reached the east African coastby 3000 BP.
ReferenceDe Langhe E.A.L., R. Swennen & D. Vuylsteke. 1996.
Plantain in early Bantu world. Azania (29-30).Based on: Mbida Mindzie C., H. Doutrelepont, L. Vrydaghs, R. Swennen, R.J. Swennen, H. Beeckman, E. De Langhe & P. De Maret. 2001.First archaeological evidence of banana cultivationin central Africa during the third millennium beforepresent. Vegetation History and Archaeobotany 10: 1-6.
Revision of BARNESA’s strategyBARNESA was established as theregional banana research network forEastern and Southern Africa in 1994 andINIBAP has been providing coordinationand secretariat support to the networksince 1997. BARNESA is one of severalnetworks operating in the region underthe auspices of ASARECA (Associationfor Strengthening Agricultural Researchin Eastern and Central Africa). Thestrategic objective of ASARECA is “to pro-mote regional economic growth by devel-oping, introducing and disseminatingagricultural technologies which bothcreate markets and respond to prevail-ing and future economic opportunitiesfor new technologies as well as main-taining the long-term sustainability ofthe agricultural resource base”. With theadoption of the ASARECA strategy in1997, it became necessary to review thegoals and objectives of networks and pro-grammes operating under the umbrella
of ASARECA in order to align their activi-ties with the objectives and purpose ofASARECA. In the case of BARNESA, thisresulted in a series of consultations andworkshops involving various stakehold-ers, with the view of realigning BAR-NESA’s original technology-focused strat-egy with that of the market-orientedapproach of ASARECA.
Following the 2001 BARNESA SteeringCommittee meeting, a Select Committeewas appointed to review and finalize theBARNESA strategy. As a result of thework of the Committee, the goal, purpose,and outputs of BARNESA have beendefined as follows:
Super goalIncreased contribution of bananas tofood security and economic growthwithin Eastern and Central Africa (ECA).
GoalTo contribute to increased and sustainableagricultural productivity in the region.
PurposeAppropriate market-oriented technolo-gies for the production of bananas andtheir products adopted and utilized.
Outputs• BARNESA consolidated as a regional
network in ECA region;• The establishment of a sustainable
commercialized banana sector in ECAfacilitated;
• Technologies for sustainable produc-tion and postharvest utilization thataddress users’ needs disseminated;
• National Agricultural ResearchSystems (NARS) capacity and linkageswith relevant partners strengthened;
• Banana research and development inthe ECA region effectively coordinated.
CommercializationBARNESA will assist in the developmentof a commercialized banana sector bydocumenting and analyzing the existingand potential production and marketopportunities for bananas and bananaproducts in the region, taking into con-sideration the changes that have takenplace in the recent past. These changesinclude economic restructuring atnational and regional levels, liberaliza-tion of trade, and formation of economicblocks such as COMESA (CommonMarket for Eastern and Southern Africa)and the East African Community. Thediverse roles of development organiza-tions, such as non-governmental organi-zations (NGOs) and community-basedorganizations (CBOs), Women Groups,
INFOMUSA — Vol 10, N° 2 41

etc., will also be documented. Analysis ofmarket opportunities will include assess-ment of the impact of demographictrends such as population growth andstructure and urbanization on bananaconsumption. Experience with commer-cial banana farming in countries such asSouth Africa will be used in laying out ofstrategies for market-oriented bananaproduction in the region.
Since both table and green-cookingbananas depend largely on urban mar-kets, understanding urban consumermarket requirements will be pivotal to amarket-oriented production of the cropin the region. Another emerging marketfor banana is banana-based processedproducts (chips, crisps, figs, juice blends,and liquor) and the flour products (cake,bread and biscuits). Processed productsprobably constitute the single mostimportant but yet to be exploited aspectof commercial banana production andmarketing. Market opportunities for thevegetative parts of the banana plant arevery limited. Probably the most promis-ing use of the pseudostem is fibre produc-tion, which is the foundation for the fast-growing handicraft industry in allbanana-growing countries of the region.
Technology development and capacitystrengtheningThe focus in this area will be to promoteregional collaboration as a means ofenhancing cost-efficiency in identifyingand addressing the priority constraints toan integrated approach to banana pro-duction, marketing and utilization withinthe region. As part of this objective, BAR-NESA will also catalyze and supportresearch on critical gaps in the produc-tion and marketing chain that are identi-fied as a priority at regional level. BAR-NESA will also work with NARS to buildup human capacity through regionaltraining courses and, where feasible,exchange visits and attachment of per-sonnel to commercial production, pro-cessing and marketing institutions withinand outside the region.
Coordination of banana research and development BARNESA activities are directed by aSteering Committee, members of whichguide participating national teams andrepresent them at regional planningmeetings. The secretariat will organizebiennial meetings of stakeholders toreview progress and suggest new regionalinitiatives. The secretariat will also playkey role in establishing and maintaininglinkages with other networks and institu-tions and will publish a regular newslet-
ter to keep the stakeholders informed ofnew developments in the banana indus-try within and outside the region. At thenational level BARNESA will encourageand assist where necessary the respec-tive NARS to put in place or strengthen anational banana research and develop-ment coordination mechanism. The keyobjective here is to ensure participationand ownership of the banana researchand development agenda by all stake-holders, including representatives ofresearch and extension services, farmersand women organizations, processors,transport/marketing associations, privatesector, NGOs, University researchers, andplanning officials in GovernmentMinistries. At the regional level, member-ship of the Steering Committee will bebroadened to include representatives ofstakeholders in the private sector such ascommercial farming, industrial process-ing and marketing. Linkages will bedeveloped with NGOs, CBOs and govern-ment institutions associated with policy-making and technology transfer, and withadvanced research institutions for accessto genetic materials and technologies.
Within the family of ASARECA net-works and programmes, BARNESA willcollaborate with: • Eastern and Central African
Programme for Agricultural PolicyAnalysis (ECAPAPA) on socioeconomicand policy related research issues;
• FOODNET (an ASARECA postharvestnetwork funded by USAID) on productdevelopment;
• African Highland Initiative (AHI),Agroforestry Research Networks forAfrica (AFRENA) and the Soil andWater Management Network (SWM-NET) on natural resources manage-ment;
• Eastern African Plant GeneticResources Network (EAPGREN) ongenetic resources; and with
• East and Central Africa Bean ResearchNetwork (ECABREN) and ASARECA-Animal Agriculture Research Network(A-AARNET) on baseline surveys andcapacity building activities.
Principles of implementationIn fulfilling the above objectives, BAR-
NESA will be guided by the followingprinciples:
• Within the concept of a holisticapproach to banana research anddevelopment, including crop manage-ment technologies that enhance sus-tainability of the natural resource base,BARNESA will concentrate its effortson the critical areas that are mostlikely to catalyze change;
• Emphasis on products that create mar-kets or respond to existing marketneeds;
• Ensuring congruence of regional activi-ties to national priorities;
• Participation of stakeholders in theplanning and execution of activities atthe local level;
• Gender consciousness in all activitiesof the network;
• The special needs of disadvantagedgroups will be specifically addressed inall aspects of project implementation;
• Timely execution of the agreedresearch agenda with due considera-tions to stated milestones, outputs andmeasurable indicators.
The scope of BARNESAUnlike other networks and pro-
grammes of ASARECA, BARNESA wasformed as a research network servingcountries in Southern Africa as well asEastern and Central Africa. It is felt thatBARNESA has still a lot to gain fromdirect association with countries insouthern Africa. BARNESA will therefore,while serving primarily the countries ofECA region, maintain strong linkageswith countries of Southern Africa espe-cially in its activities of capacity-building,institutional linkages and sourcing oftechnological information.
Asia and the Pacific
Managing banana bunchy topdisease in PakistanBananas are an important fruit crop inPakistan, where around 100 000 tonnesare produced annually from an area ofmore than 26 000 hectares. The majorityof the crop is produced in the SindhProvince which lies between 67-71°N and24-28°E. A number of varieties are grownin Pakistan, but the most common (90%of production) is Dwarf Cavendish,known locally as Basrai.
During the early 1990s, banana bunchytop virus spread rapidly in Pakistan. Insome districts, more than 60% of the cropwas infected and production declined by90%. To overcome this problem, effortshave been directed towards the produc-tion of clean (virus-free) planting mater-ial of variety Basrai using tissue culture.Experiments showed that the best resultswere obtained using explants of 4-6 mmwhich were initiated in liquid medium.For multiplication, explants were trans-ferred to solid medium containing 5.0 mg/l BAP and an average of 6-8 shootswere produced every four weeks. After invitro rooting using IBA, the plants werehardened in a greenhouse for 8 weeks.
42 INFOMUSA — Vol 10, N° 2

Once the plants had 6-8 leaves they wereplanted in the field. The survival rate ofplants using this procedure was almost100%.More information is available from AishMohammad et al. Agricultural BiotechnologyInstitute, National Agricultural Research Centre,Park Road, Islamabad, Pakistan.
Biochemical analysis of Musa cv.Red (AAA) and its green variantThe banana cultivar ‘Red’ (AAA) is widelycultivated and can be found in mostbanana-growing countries of the world.In India, it is particularly important inthe states of Maharashtra, Tamil Naduand Kerala. Because of the attractive sizeand colour, the fruits always fetch a highprice in the markets in India. During themultiplication of Red banana by micro-propagation, a small number of greenvariant plants were produced. In theseplants, both the vegetative parts and thefruit bunch remained green throughoutthe life of the plant, with the fruits devel-oping a yellowish tint during ripening. Inthe normal plants, the characteristic redcolour started developing on the leavesand petioles within one week of trans-planting tissue cultured plants into potsand the plant and fruit remained reduntil maturity. Green variants were pro-duced when explants were inoculated onMS medium supplemented with 0.1mg/lNAA and 8mg/l BA, whereas explantsinoculated on MS medium supplementedwith 0.1mg/l NAA and 8mg/l kinetin didnot produce green variants. The additionof kinetin instead of BA also had theeffect of reducing the multiplication rateof the plants in tissue culture.
Biochemical analyses of total protein,reducing sugars and total carbohydratewere carried out on the ripened fruit pulpof Red banana, its green variant andother locally available cultivars –Rasthali (Silk AAB), Robusta (AAA) andNendran (Plantain AAB). The results arepresented in Table 1. It can be seen thatthe protein content was higher in Redbanana compared to the others, and thetotal carbohydrate content was lower.Reducing sugar content was high inRasthali, while the total carbohydratecontent was greatest in Nendran. Littledifference in biochemical analysis wasfound between Red banana and the greenvariant. Further information is available from Asalatha S.Nair, Department of Botany, University ofKerala, Kariavattom, Thiruvananthapuram 695581, Kerala, India.
New pests of banana reported in Tamil Nadu, IndiaBanana, the most important fruit crop ofIndia, is prone to attack by different
pathogens such as fungi, bacteria,viruses, insects and nematodes. The shortnotes below report on recent observa-tions carried out in Tamil Nadu by scien-tists of NRCB, the Indian NationalResearch Centre for Banana.
Small banana weevil: Polytusmellerborgii Boheman (Dryophthoridae:Curculionoidea)B. Padmanaban, M. Kandasamy and S. Sathiamoorthy
Among the 28 borer pests associated withbananas and plantains worldwide (SeshuReddy et al. 1994), banana weevil borers(banana rhizome weevil, Cosmopolitessordidus Germar and banana stem wee-vil, Odoiporus longicollis Oliver) arewidely distributed and cause importanteconomic losses (Dutt and Maiti 1972,Gold 1994, Padmanaban and Sundararaju1999).
During a field visit to Coimbatore, amajor banana-growing area in TamilNadu, a very small weevil was noticed atthe top of the rhizome as well as in theleaf sheaths of the leading commercialcultivars French Plantain (NendranAAB) and Poovan (Mysore AAB). Theweevil was found to feed and survive onrotting rhizomes. Symptoms of damagedue to weevil feeding were also noticedon leaf sheaths. The weevil was alsorecorded in the cut rhizomes removedafter the harvest. On average 2-3 weevilsper rhizome and 2-4 weevils per plant werecollected. The weevil looks like a minia-ture banana rhizome weevil (Figure 1).The morphometry of the new insect incomparison to the banana weevil borersis presented in Table 2. The NaturalHistory Museum, London has identified itas Polytus mellerborgii Boheman, also
commonly known as small banana weevil.This weevil was first recorded in China(Zhou and Wu 1988), though this is thefirst record of its occurrence in India.
Thanks are due to Dr C. Lyal, InsectInformation Services, EntomologyDepartment, The Natural HistoryMuseum, London, for identification of theinsect.
Banana beetle: Sybra praeusta Pascoe(Cerambycidae: Coleoptera)B. Padmanaban, M. Kandasamy, P. Sundararaju and S. Sathiamoorthy
Bananas and plantains in India are depre-dated by 30 species of insects (Wadhi andBatra 1964 ). The banana weevil borers(banana stem weevil, Odoiporus longi-collis Oliver and banana rhizome weevil,Cosmopolites sordidus Germar) are themost important pests of banana causingserious economic damage.
Surveys undertaken during March 2001in the banana plantations nearJeyapuram (Tiruchirapalli district) andthe NRCB farm revealed that in patches
INFOMUSA — Vol 10, N° 2 43
Table 1. Biochemical analysis of fruit from Red, green and other locally availablecultivars.
Total sugar Protein
Name of the cultivar Reducing sugar (%) Total carbohydrate (%)
Red Banana (AAA) 35.21 36.98 32.90
Green variant (AAA) 34.54 35.14 32.84
Rasthali (AAB) 45.31 41.35 23.54
Robusta (AAA) 38.39 41.72 21.64
Nendran (AAB) 40.38 51.67 25.21
Table 2. Morphometry of Polytus mellerborgii in relation to banana weevil borers.
Body length Body width Length of rostrum Insect (mm) (mm) (mm)
Banana rhizome weevil, Cosmopolites sordidus 11.0 3.5 2.5
Banana stem weevil, Odoiporus longicollis 16.0 5.5 5.0
Small banana weevil, Polytus mellerborgii 4.0 1.5 1.5
Figure 1. Magnified view of (a) Banana rhizomeweevil and (b) Small banana weevil.

banana plants were exhibiting unusualyellowing and consequent drying of leafsheaths. Further examination of the leafsheath showed extensive gallery forma-tion along with deposition of frass mater-ial (Figures 2 f-g). Grubs were presentinside the gallery and their feeding wasrestricted to the two outermost leafsheaths. The grubs were reared on theoriginal leaf sheath in special containersin the NRCB laboratory. The yellowishwhite larva is typical of Cerambycid grub(Figures 2 b, 2e). The larval duration was16 days under laboratory conditions. Thegrubs pupated as a yellowish orangeexarate pupa (Figures 2c, 2d). Beetleswith yellowish grey markings and twoblack spots on the elytra (Figure 2a)emerged from the pupa. The beetle wasidentified as Sybra sp. following Duffy(1968). With the help of the NaturalHistory Museum, London, the exact iden-tity of the beetle was found as Sybrapraeusta Pascoe. Infestation of thisCerambycid beetle was noticed on 7-month-old cultivars such as Nendran(French plantain AAB), Pisang Awak(ABB), Agnisagar (AAA) and Dudh Sagar(AAB).
It is reported that Sybra species are dis-tributed in India and Sri Lanka. They arepolyphagous beetles developing on fruits,pods, fleshy stems and tubers of a widerange of plant species. Sybra praeustaresembles Sybra ceylonensis Breu., whichis found infesting Calotropis procera. S. praeusta, at present, is found affectingbananas and plantains sporadically anddoes not appear to be distributed in other
parts of Tamil Nadu. Affected plants, how-ever, show sudden yellowing of outerleaves resulting in a reduction of func-tional leaves as the plant ages.
Thanks are due to Dr Sharon Shute,Insect Information Services, Departmentof Entomology, The Natural HistoryMuseum, London, for identifying theinsect.
Cyst nematode: Heterodera oryzicolaRao & Jayaprakash, 1978P. Sundararaju, I. Cannayane and S. Sathiamoorthy
The cyst nematode Heterodera oryzicola,reported for the first time in Kerala onbanana cv. Nendran, was found to causesevere root damage resulting in seriouseconomic losses (Charles and Venkitesan1984). Koshy et al. (1987) reported itswidespread occurrence on banana cv.Njalipoovan from Goa, India. Charles andVenkitesan (1994) reported that an ini-tial inoculum of 100 to 1000 viable cystsper plant at planting time could reducethe bunch weight by 20.5 to 56.6 per cent.So far this nematode had been found tobe confined in reclaimed paddy lands ofKerala, with reports of stray occurrenceson banana from Goa. No information wasavailable on the occurrence and distribu-tion of the cyst nematode on banana inTamil Nadu, where banana is grownextensively.
During the routine examination of fieldsoil samples in banana experimentalplots at the NRCB Farm, Podavur, a largenumber of cysts were observed in cv.
Robusta (AAA). Detailed examination ofthe roots revealed the presence of about86-105 matured cysts and 12-15 juvenilesfrom 250 g soil, and 22-29 white femalesper 10 g of roots. Each matured cyst con-tained about 18-47 viable eggs.
Inoculation of banana roots with secondstage larvae resulted in the penetration ofroots and development of white femaleswith egg sacs, filled with viable eggs. Aftercloser observation of their morphologicalparameters such as vulval cone, whitefemales and juveniles, the species wasidentified as Heterodera oryzicola Rao &Jayaprakash, 1978. The white femalesattach and feed on the fibrous secondaryand tertiary roots by modifying the hostcells (syncytia). The primary roots weredevoid of white females.
The finding clearly indicates the needfor an intensive survey programmethroughout the banana-growing belts ofTamil Nadu for cost-effective manage-ment of the cyst nematode.
References related to the sectionon banana pests in Tamil NaduCharles J.S.K & T.S. Venkitesan. 1984. New hosts of
Heterodera oryzicola Rao & Jayaprakash, 1978, inKerala, India. Indian J. Nematol. 14: 181-182.
Charles J.S.K & T.S. Venkitesan. 1994.Histopathological changes in banana cv. Nendranroots parasitized with cyst nematode, Heterodera
oryzicola. Pp. 216-217 in Proc. Kerala Sci. Congr.,January, 1994, Thiruvananthapuram, India.
Duffy E.A.J. 1968. A monograph of the immaturestages of Oriental timber beetles (Cerambycidae).British Museum Natural History, London.
Dutt N. & B.B. Maiti. 1972. Bionomics of the bananapseudostem borer Odoiporus longicollis Oliver(Coleoptera: Curculionidae). Indian J.Entomology 34(1): 20-30.
Gold C.S. 1994. Banana weevil: Ecology, pest statusand prospects for integrated control with empha-sis on East Africa. Pp. 49-74 in Proceedings of aSymposium on biological control in tropical crophabitats. Third International Conference onTropical Entomology, 30 Oct.-4 Nov. 1994, Nairobi,Kenya (S.K. Saini, ed.). ICIPE, Nairobi, Kenya.
Koshy P.K., V.K. Sosamma & J.R. Faleiro. 1987.Occurrence of Heterodera oryzicola on banana inGoa. Indian J. Nematol. 17: 334.
Padmanaban B & P. Sundararaju. 1999. Occurrenceof banana weevil borers. (Curculionidae:Coleoptera). Insect Environment 5: 135.
Seshu Reddy K.V., J.S. Prasad & R.A. Sikora. 1994.Biointensive management of crop borers ofbanana in Proceedings of a Symposium on biolog-ical control in tropical crop habitats. ThirdInternational Conference on TropicalEntomology, 30 Oct.-4 Nov. 1994, Nairobi, Kenya(S.K. Saini, ed.). ICIPE, Nairobi, Kenya.
Wadhi S.R. & H.N. Batra. 1964. Tropical and subtrop-ical fruit pests in Entomology in India (N.C. Pant,ed.). Entomological Society of India, New Delhi.
44 INFOMUSA — Vol 10, N° 2
Figure 2. Cerambycid beetle, Sybra praeusta Pascoe.a. Adult beetle; b. Grub; c. Pupa - ventral view; d. Pupa – dorso-lateral view; e. Grub feeding inside thegalleries; f-g. Leaf sheath showing feeding damage at outer and inner side (see gallery and frassmaterial).

Zhou S & X. Wu. 1988. A species of Curculionidae firstrecorded in China. J. Zhong Kai. Agrotech. Coll.,China 1(1): 33-34.
For further information, please contact therelevant authors at: National Research Centre forBanana (NRCB), Tiruchirapalli 620017, TamilNadu, India.
Formosana: a new Cavendishvariety released in TaiwanThe breeding programme of the TaiwanBanana Research Institute has created anew banana variety, ‘Formosana’, whichis resistant to Fusarium wilt. This dis-ease, caused by Fusarium oxysporumf.sp. cubense (Foc) race 4, is a majorproblem in Taiwan and the new varietyrepresents a breakthrough in bananabreeding. Derived from the wilt-suscepti-ble variety ‘Giant Cavendish’, itssomaclone ‘Formosana’ has a higher levelof resistance to Foc than ‘Tai-Chiao No.1’, the wilt-tolerant variety released forcommercial production in 1992, and bet-ter yield characteristics than ‘GiantCavendish’. ‘Formosana’ is scheduled forrelease for commercial planting as ofJanuary 2002, to replace both ‘Tai-ChiaoNo. 1’ and ‘Tai-Chiao No. 3’, two resistantvarieties already released and planted onabout 1600 ha of infested farms in south-ern Taiwan. The commercialization ofFormosana is expected to have a greatimpact on Taiwan banana industrythrough reduction of losses due to Focattacks, yield increase, and more evenripening than with ‘Giant Cavendish’,thus leading to better competitiveness ofTaiwan bananas on the export market.For further information, contact: C. Hwang,Taiwan Banana Research Institute, PO Box 18,Chiuju, Pingtung 90403, Taiwan.
Third Australian QBAN workshopThe Quality Banana Approved Nursery(QBAN) scheme is an importantAustralian national initiative to ensurethat the banana industry is provided withuniform, disease and pest-free plantingmaterial. Its aim is to prevent the intro-duction, movement, and spread of notifi-able pests and diseases of bananas. QBANplantlets are grown only from virus testedand registered tissue culture materialfrom QBAN accredited facilities.
The QBAN scheme was conceived in1993 and is now a growing alliance of 20laboratories and nurseries providing tis-sue culture plantlets or hardened nurseryplants. Technical and regulatory supportis provided principally by the QueenslandDepartment of Primary Industries(QDPI). In June 2001 a QBAN workshopwas held in North Queensland at theCentre for Wet Tropics Agriculture.Researchers from QDPI and invited
speaker Gus Molina, Regional Coordinatorfor INIBAP - Asia and Pacific, presentedpapers covering a range of technicalissues concerning the management ofbanana germplasm and the production ofclean planting material. The QBAN guide-lines were reviewed and the impact of therecent black Sigatoka incursion inAustralia was discussed.Source: Sharon Hamill, QDPI, Maroochy ResearchStation, PO Box 5083, SCMC, Nambour, 4560Australia, Phone +61 07 54449639 Fax +61 0754412235. Email: [email protected]
Latin America and the Caribbean
Launching of MusacolThe constitutive agreement of the RedColombiana de Investigación yDesarrollo de Plátano y Banano(Colombian Network for Plantain andBanana Research and Development -Musacol) was signed on 3rd August 2001in Armenia, Colombia. Members of thenetwork include national entities such asCorpoica, Fedeplátano and severalColombian universities.
The objective of Musacol is to orga-nize, coordinate and catalyze institu-tional efforts in research, developmentand technology transfer in a dynamic,systematic and concerted way, with theaim of solving the technological limita-tions of the plantain and banana cultiva-tion. Through this mechanism, Musacolwill generate practical and scientificrecommendations for improving the pro-duction, processing and commercializa-tion of the crop, garanteeing the sus-tainibility and equity of the productionsystems.
The Board of Musacol designatedAlfredo Rodríguez Saavedra to act as thefirst Executive Director of Musacol for atwo-year period.More information on Musacol website at:www.musacol.org.co
In Memoriam
Ken Shepherd INIBAP is very sad to inform readers ofINFOMUSA of the death on 16 October2001 of Kenneth Shepherd, a Britishresearcher well known in Musa geneticimprovement circles, and wishes to pre-sent its most sincere codolences to hiswife, Eileen, and children.
Having worked on the same plant forover 40 years, Ken made an immensecontribution not only in cytogenetics(Shepherd 1999) and improvement butalso in taxonomy. In 1994, in recognitionof his scientific contribution, INIBAPpresented Ken with a plaque of apprecia-
tion in the name of the world bananacommunity.
Here is a word by his colleagues atEmbrapa Mandioca e Fruticultura, withwhom he worked for a large part of hiscareer.
Homage to Kenneth Shepherd byEmbrapa Mandioca e Fruticultura
“Pacovan Ken” is the name of a newbanana variety launched by EmbrapaMandioca e Fruticultura in homage tothe eminent international scientistKenneth Shepherd, one of the breedersof the variety.
Kenneth Shepherd was born inEngland in 1927. After gaining a brilliantdegree in agriculture and botany atDurham University, England, in 1947,Ken started his career by working on thegenus Solanum for two years inCambridge. After this initial phase, hededicated the rest of his career to thebanana, which became his favourite‘muse’. He worked from 1950 to 1960 inthe banana breeding programme at theImperial College of Tropical Agriculturein Trinidad, and then from 1960 to 1980in the Jamaican Banana Board’s breed-ing programme, where he created sev-eral genotypes including the hybrid M53.The latter was then added to theEmbrapa genebank and was the originof the variety ‘Pacovan Ken’. The programme also developed other hybrids such as “Calypso’, ‘Bucanier”and “Ambrosia”, all of which were bredby this great scientist.
From December 1981 onwards, Kencontinued his career in Brazil as a
INFOMUSA — Vol 10, N° 2 45
Ken Shepherd in the FHIA Musa collection at the occasion of the first meeting of the Musabreeders’ network held at FHIA, Honduras inMay 1994.

consultant with Embrapa, where he setup a breeding programme.
Ken then played a true role of leader atthe head of a team of young researchersand technicians by passing on, with hisnatural modesty, all the knowledge thathe possessed. His most preciousmoments were his field trips to accom-pany the installation of a trial, when hehelped with planting, cleaning and trans-planting, or to attentively monitor thegrowth of the new genotypes. He wasthen as happy as when he smoke a goodBahia cigar, one of his weaknesses.
Throughout his time spent atEmbrapa, Ken played a vital role in thebreeding programme, in particular byintroducing genetic material from othercountries, a complex, difficult task thatwas only possible because of his con-tacts and his credibility with the inter-national bodies working on Musa. Brazil thus possesses today the secondactive genebank in the world including the most important parents—anunequalled treasure.
Kenneth Shepherd published more than50 books that are references for the worldbanana community. Unfortunately, muchother work has remained unpublished.
The first concrete result of his work inBrazil was the launching of the ‘Pioneira’cultivar in 1992.
Difficulties arose for the renewal ofhis contract when he was working asconsultant for Embrapa. In the early1990s, he even remained for two yearswithout being paid his fees. On hisdeparture from Brazil in 1994, he leftbehind him a well trained team capableof continuing the work on promisinggenotypes. One of these was “PacovanKen”, launched in November 2001 as anational crop plant. The new variety isnot only more productive that the‘Pacovan’ variety traditionally grown inthe Nordeste in Brazil, but it is alsoresistant to yellow and black Sigatokasand to Panama disease, the three cursesof banana around the world.
We are sure that this sincere homage toKenneth Shepherd, using the short formof his name (Ken) in the name of the newvariety, is not much in comparison withthe immense heritage that he has left to the Brazilian and world banana com-munities.
When we chose to name this hybrid inhis honour, we tried to contact him inPortugal, where he was spending hisretirement, to invite him to the launchingof the new variety on 30 November 2001.Unfortunately, we then received the sadnews that he had died on 16 October 2001at the age of 74.
We shall always remember KennethShepherd as a model of competence,modesty, abnegation, impartiality andaltruism. Embrapa Mandioca e FruticulturaNovember 2001
ReferenceShepherd K. 1999. Cytogenetics of the genus Musa.
INIBAP, Montpellier, France.
Norman W. SimmondsNorman Simmonds, author of everybanana researchers reference book,“Bananas” in the Longman TropicalAgriculture Series (1959, 1966 and withR.H. Stover in 1987) and also “The evolu-
tion of bananas”,has died onJanuary 4, at theage of 79 years.Simmond’s careerspanned morethan five decades,focussing on plantbreeding, taxon-omy, economicbotany and tropi-
cal farming systems. His expertise onbananas, sugar cane and potatoes wasparticularly well recognized. Major col-lecting trips in East Africa and Asia andthe Pacific during the 1940s and 50sallowed him to develop a deep under-standing of the origin and spread ofbananas. In 1955, together with KenShepherd, he created the genome-basedsystem of nomenclature for cultivatedbananas (using A and B for acuminataand balbisiana genomes) as an alterna-tive to the Linnaean system to reflectmore effectively levels of ploidy andhybridization. In his lifetime, he pub-lished 48 scientific papers on bananaalone, covering many subjects fromgenetics to the development of fruit andgermination of seeds. Right until his lastweeks Simmonds actively participated inagricultural research, regularly interact-ing with researchers and responding tonew developments through his frequentletters published in journals; one of hislast letters is published in theMusaForum section of this issue ofINFOMUSA (see below). He was a strongsupporter of the concept of INIBAP, andfollowed avidly our activities throughpublications such as INFOMUSA, fre-quently providing eloquent criticism. Hemade his disapproval of the investmentin research on biotechnology well known.This outspoken voice will be missed. Butit is his ground-paving and thorough workon banana and in agricultural botany as awhole which renders him unforgettable
and quite irreplaceable. It was a sombremoment when this great career ended.
MusaForum
Last letter from Prof. N.W. Simmonds to INFOMUSAIn a recent article in this periodical,Manzur Macias presented some neatdrawings and evidence to the effect thatpoorly suckering bananas could some-times at least be provoked into bettersuckering by mutilating the growing areasof the corms. He worked on FHIA-20 andfound that mutilation worked for up to 4cycles. He cites examples from otherbananas showing the same effect but notfrom the really striking example of EnseteThis genus is normally non-suckering and,though it bears plenty of buds, cannot bevegetatively propagated. However, thepeople of the Sidamo area have invented amethod whereby elderly corms are hol-lowed out and the cavities packed with soiland dung, whereupon buds grow wellenough for practical propagation. TheSidamo people eat the starchy foods thatare produced and recognize clones.Indeed, Ensete, with a perennial cabbage,is their staple food, but they also keepdiverse stocks. It will be interesting to seewhether many new cooking bananasrequire this kind of trick to facilitate prop-agation. It would be well worth study.
ReferenceManzur Macias D. 2001. In situ mass propagation of
the FHIA-20 banana hybrid using benzylaminop-urine. INFOMUSA 10(1):3-4.
Further reading:Simmonds N.W. 1958. Ensete cultivation in the south-
ern highlands of Ethiopia. Tropical Agriculture(Trinidad) 35:303-307.
Smeds H. 1955. The Ensete planting culture of east-ern Sidamo, Ethiopia. Acta geogr. Paris 13(4): 40.
INIBAP news
Catching up with relativelynew facesThree members of staff have joined INIBAP regional offices in the past yearand have escaped introduction in INFO-MUSA. We take the chance now to rectifythe situation.
Helen Namatovu Byarugaba is a pro-gramme assistant at the INIBAP office inKampala, Uganda. She took up the post inFebruary this year, coming with a back-ground in business and marketing. Helenis Ugandan. She graduated from MakerereUniversity with a degree in Commerce andhas gained experience in banking and cus-tomer management through a trainingcourse run by Barclays Bank in Uganda.
46 INFOMUSA — Vol 10, N° 2

Together with Siifa Balinda Lwasa, sheforms the backbone of programme supportin what is now the largest INIBAP officeoutside of Montpellier.
Maria Angeli Maghuyop has beentechnical assistant at the INIBAP officein Los Baños, the Philippines, since July2000. Her role involves providing supportin the implementation of theInternational Musa Testing Programme(IMTP) Phase III and providing programme support in organizing meet-
ings and produc-ing reports andother materials.Angie is Filipinaand has a degreein agriculture, spe-cializing in plantpathology, fromthe University ofthe PhilippinesLos Baños (UPLB)
and worked previously as a researcher atUPLB, the Entomology and PlantPathology Division of the InternationalRice Research Institute (IRRI-EPPD)and as programme assistant in thePhilippine Council for Agriculture,Forestry and Natural Resources Researchand Development (PCARRD).
Dr Robert Nkendah is a researcherunder the Young ProfessionalsProgramme of the Food and AgricultureOrganization of the United Nations(FAO), seconded for a period of 11 months to the INIBAP office in Douala,Cameroon, starting in January this year.Robert is a Cameroonian Agroeconomistwith a doctorate in rural economy fromthe University of Abidjan, Côte d’Ivoire.Banana and plantain were the subject ofhis doctoral studies, which he carried outwhile based at the Centre Ivoirien deRecherches Economiques et Sociales(CIRES). Whilst at INIBAP, he has been
working under the joint-supervision of the INIBAP regional coordinator forWest and Central Africa and the
Agriculture PlantProduction andP r o t e c t i o nDivision (AGPC)of the (FAO). His chief taskshave included col-laborating withMUSACO mem-bers to gatherbaseline informa-
tion on banana production into a spe-cially-developed database, and to con-duct a bibliographic study of periurbanagriculture around Yaounde and Douala,the two major cities of Cameroon.
Better knowledge of information management toolsIn recent years, INIBAP’s Informationand Communication Unit has developednew information dissemination aid tools.INIBAP held a training session from 8 to12 October 2001 to initiate regional officeprogramme assistants in the use of thesetools and had the pleasure of welcomingas participants Versalynn Roa (Asia andthe Pacific), Lisette Vega (Latin Americaand the Caribbean), Helen NamatovuByarugaba and Siifa Lwasa (eastern andsouthern Africa). Marie-MadeleineMbakop Ngamy (western and centralAfrica) was unfortunately not able toattend.
The participants were able to explorethe INIBAP Web site, the MUSADOC 2001CD-ROM and the “Les Bananes/Bananas”multimedia CD-ROM. They attended pre-sentations of the MGIS programme (Musagermplasm information system) and theimage database (see the article on p. 34 ofthis issue). Their new knowledge will make
it easier for them to reply to questionsfrom banana information users at theregional level and will give them increasedeffectiveness in querying the MUSALITdatabase (bibliographical references) andBRIS (Banana research information sys-tem). They were also trained to use theinternal database managing the distribu-tion list, an essential tool for the sendingof the publications produced by INIBAP.
The week spent in Montpellier was theoccasion for useful exchange betweenregional office and INIBAP headquartersstaff.
Meeting banana farmers on the big screenINIBAP’s first venture with the movingimage has resulted in a swift-moving 18 minute narrated film, providing abroad introduction to the importance ofbananas in world agriculture, problemsfaced by smallholder farmers and howINIBAP is trying to address them.
The film was produced by a Canadiancompany, called Baobab Productions,which has worked with several FutureHarvest centres. Their philosophy issomewhat unique in that they do notemploy a large camera and productioncrew, but work as a one-man band whoshoots, interviews, produces and directsall in one. The pressure may be intense,but the budget is low and the effect onthose being filmed is far less intrusive.David Mowbray, the one-man band andpresident of Baobab has travelled withhis small digital video camera throughoutAsia, Africa, Europe, Central Americaand the Caribbean on an excruciatingtimetable to interview banana farmers,researchers and INIBAP partners.
The result is a beautiful sequence ofimages, with music and narration, illus-trating the diversity of people involved inbanana production and marketingaround the world, from huge banana auc-tions in India to large plantations in Cuba
INFOMUSA — Vol 10, N° 2 47
INIBAP Regional programme assistants at INIBAP HQ (from left to right: Siifa Lwasa, Lisette Vega,Helen Namatovu Byarugaba and Versalynn Roa).

and smallholdings and women’s coopera-tives in Africa. Launching the film at theAnnual General Meeting of the CGIAR atthe World Bank in Washington, INIBAPwas able to introduce the INIBAP/PRO-MUSA support group to some of the pro-jects and individuals they have beenfinancing. The film is being distributed asa CD-ROM for viewing on a computer.
For more information contactCharlotte Lusty or Claudine Picq at INIBAP HQ.
MGIS NewsFollowing the MGIS training course heldin India, in May 2001, new sets of datahave been added to the MGIS database byDr Uma, National Research Centre onBanana (NRCB), Trichy and Dr A. Rekha,Indian Institute of HorticulturalAgriculture (IIHR), Bangalore, India.This update brings the number of acces-sions recorded for the NRCB collection to813, along with 290 morphotaxonomicdescriptions, 45 photos and 54 agronomicevaluations. Dr Rekha has sent 26 pass-part data as a starting set.
To date, 4153 accessions have beenrecorded in MGIS by 13 institutions, withinformation on 809 shipments of mater-ial, 1619 morphotaxonomic descriptions,1495 agronomic evaluations, 300 evalua-tions to stress and 687 photos.
IITA-INIBAP Joint Programmefor Africa – Annual planningmeetingOn 29 September, the annual planningmeeting between IITA and INIBAP washeld in Kampala, Uganda. This meetingwas organized in order to review progressin the joint programme which had beeninitiated in 2000 and to identify any con-straints to collaboration. All participantsagreed that the joint programme was awelcome initiative and noted thatprogress had been made during the lastyear in achieving greater integrationbetween the INIBAP and IITA pro-grammes in Africa. It was emphasizedthat collaborative work programmesshould be discussed and developed at theannual meetings of the SteeringCommittees of the regional networks(MUSACO and BARNESA). In this way,regional research agenda can be devel-oped involving all players, national,regional and international. Specific areasof discussion included IITA’s participa-tion in the Uganda Biotechnology Projectbeing implemented by INIBAP, the com-plementary roles of INIBAP and IITA withregard to the conservation of germplasmand the roles of IITA and INIBAP in thepublication of MusAfrica.
IPGRI Board of Trusteesvisited UgandaThe IPGRI Board of Trustees for 2001Session II, met in Nairobi, Kenya,September 17-21, 2001. As part of theirprogramme it was arranged for them tovisit Uganda to meet stakeholders andpartners in the country and view first-hand the field activities under way. Thenon-stop programme took the visitors 400 km to the south-west of Uganda, aswell as to research facilities in and aroundKampala. Members of the BOT had theopportunity to exchange ideas not onlywith researchers, research managers andpolicy makers, but also with farmers par-ticipating in IPGRI’s research programme.
Visit to banana in situ conservationsite in BushenyiBushenyi is one of the five benchmarksites which INIBAP and partners haveestablished in Eastern Africa. It islocated nearly 400 km to the south-westof Kampala, in a highland agro-ecosys-tem that is completely dominated bybananas grown in subsistence holdings.Here banana is the way of life and peo-ple’s lives are about bananas. Peoplehere work in banana fields and eat‘matooke’ (a banana dish); cows andgoats feed on banana leaves and pseu-dostems and every vehicle that drives byis carrying bunches of bananas. Farmerswho can not afford iron roofs use bananaleaf base fibre as thatch for their houses.Most gardens have been under continu-
ous banana cultivation for more than 200years or more.
The importance of bananas in thisregion of Uganda became quickly appar-ent to the BOT as the farmers launchedan intense programme of activities toshow the visitors what they do withbananas and what bananas do for them.The demonstrations focused on bananautilization, including the cultural as wellas the nutritional aspects and the BOTwas treated to a break time buffet of vari-ous dishes and beverages. Other demon-strations were focused on illustrating fac-tors affecting genetic erosion, includingsoil erosion, soil nutrient deficiencies,pests and diseases.
The key partner at this site is a commu-nity-based organization (CBO) called theAssociation for the Advancement ofSustainable Rural Development (ASASU-RUDE). Through funding from a USAIDgrant ASASURUDE has been able to pro-vide a cow to its members to help addressthe nutritional needs of the region. Bycoincidence, baseline studies carried outas part of the in situ conservation projecthad revealed that serious soil nutrientproblems in banana gardens was con-tributing to cultivar loss. Arrangementswere made to link the two projects so thatproject farmers received a zero-grazedcow. With the support of the governmentextension department, project farmerswere taught how to compost manure usingcow dung and urine as the base. Themanure has been applied to banana
48 INFOMUSA — Vol 10, N° 2
Marcio de Miranda Dos Santos, IPGRI BOT Chair and Geoff Hawtin, IPGRI Director General, looking atmaking banana compost in Bushenyi (C. Hoogendoorn, IPGRI).

gardens with resounding success. Farmersin impressive style demonstrated to theBOT members their skills in preparingnutrient-rich manure for their bananas.
At the end of the visit, farmers treatedthe BOT members to a ‘matooke’ meal,during which the chair of ASASURUDErequested IPGRI to look into a number ofconstraints the farmers and the CBO werefacing. In response, the IPGRI DirectorGeneral (DG), Dr Geoffrey Hawtin, offeredto provide a motorcycle to the CBO, toalleviate the transport problem. DrHawtin also thanked the farmers for themeal and for being effective partners inIPGRI’s research programmes. The visitwas crowned by the planting of bananatrees by the BOT chair and the IPGRI DG.
Meeting with partners in and around KampalaBack in Kampala, the BOT had discus-sions with the Association forStrengthening Agricultural Research inEast and Central Africa (ASARECA) andofficials of the National AgriculturalResearch Organization (NARO). The dis-cussions centred on the two IPGRI-coordinated networks (BARNESA andEAPGREN) operating in the region andhow their activities could be strength-ened. In addition the party met with theUganda Minister of State for Agriculture,Hon. Israel Kibirige Sebunya, who laterhosted a Minister’s lunch. Prof. JosephMukiibi, the Director General of NARO-Uganda, also attended the meeting. MrJohn Wasswa Mulumba conducted theBOT through Entebbe Botanical Gardenand provided an outline of the PGR activ-ities of NARO.
The BOT also visited NARO’s ForestryResearch Institute (FORI) field sites atNamanve and Kifu, where NARO andICRAF have collaborative projects on for-est conservation. The BOT visited thefield station of the Eastern and SouthernAfrican Research Centre (ESARC) of theInternational Institute of TropicalAgriculture (IITA) and attended demon-strations presented by a joint team ofNARO and IITA scientists on cassava,potatoes, yams and banana. Later theparty visited the tissue culture facility atNamulonge Agricultural and AnimalResearch Institute (NAARI) and atMakerere University AgriculturalResearch Institute, Kabanyolo, where theproblems of genebank operations inUganda were discussed. The four-day visitwas concluded with a trip to MakerereUniversity and a meeting with the Deanof the Faculty of Agriculture, who hailedthe institutional linkages betweenMakerere University and IPGRI.
Relaunching of the networkfor Asia and the Pacific:BAPNET (Banana Asia-PacificNetwork)A planning workshop of the Asia-PacificRegional Banana Research andDevelopment network was held onSeptember 11-14, 2001 in Kalutara, SriLanka in place of the traditional annualRegional Advisory Committee meeting.The workshop was very significant as itaimed to relaunch a new and revitalizedbanana network that is responsive to thebanana R&D needs in the region. WhileASPNET has achieved significant accom-plishments in the region since its cre-ation in 1991 as an INIBAP initiative, thenetwork members felt that there is now aneed to review the goals and objectives ofthe network, including its modusoperandi, structure, role of its membersas well as that of INIBAP, so that it willbecome a sustainable, relevant and trulyNARS-based banana network.
Heads of member NARS in the regionor their representatives, an APAARI (AsiaPacific Association of AgriculturalResearch Institutions ) representative,INIBAP staff including the Director ofINIBAP participated in the planningworkshop. A professional facilitatorensured an efficient workshop. As an out-put, a new constitution of the networkwas formulated, together with the net-work logical framework. The participantschose a new name of the network, theBanana Asia Pacific Network (BAPNET).The full relaunching and operationaliza-tion of the network is expected in 2002 asthe BAPNET constitution will be fullyofficially approved by all heads of NARSas well as the complete SteeringCommittee members shall be finalizedand strategic planning completed.
The workshop was organized by theAsia Pacific Office of INIBAP in collabo-ration with the Department ofAgriculture (DOA) of Sri Lanka as the
local host through the active support ofDr S.S.B.D.G. Jayawardena, DirectorGeneral of DOA.
The meeting was attended by 15 repre-sentatives of 12 country and institutionalmembers:• Australia – Mr Robert Williams
(Programme Leader - Tropical TreeFruit, Queensland Horticulture Ins-titute, Department of PrimaryIndustries)
• Bangladesh – Dr Syed Md. MonowarHossain (Director, HorticultureResearch Centre/Bangladesh Agric-ultural Research Institute)
• Cambodia – Mr Pith Khon Hel(Germplasm Specialist and PlantBreeder, Cambodian AgriculturalResearch and Development Institute)
• China – Prof. Tang Xiaolang (ViceDirector, Guangdong Academy ofAgricultural Science)
• India – Dr R.N. Pal (Deputy DirectorGeneral for Horticulture, IndianCouncil of Agricultural Research)
• Indonesia – Dr Ahmad Dimyati(Director, Central Research Institutefor Horticulture)
• Malaysia – Dr Nik Hassan Nik Masdek(Senior Research Officer, MalaysianAgricultural Research and Devel-opment Institute)
• Philippines – Dr Patricio Faylon(Executive Director, Philippine Councilfor Agriculture, Forestry and NaturalResources Research and DevelopmentInstitute and also representingAPAARI) and Nicomedes Eleazar(Assistant Director, Bureau ofAgricultural Research)
• Sri Lanka – Dr S.S.B.D.G. Jayawardena(Director General, Dept. of Agriculture),Dr C. Kudagamage (Director, Horti-culture Research and DevelopmentInstitute, DA), Dr Sujatha Weerasinghe(Banana Coordinator, HORDI, DA)
• Thailand – Dr Wasan Pongsomboon(Agricultural Scientist, Horticulture
INFOMUSA — Vol 10, N° 2 49
Participants of the Asia/Pacific Musa Network relaunching workshop.

Research Institute/Dept. of Agric-ulture)
• Taiwan Banana Research Institute – Dr S.C. Hwang (Director, TBRI)
• Secretariat of the Pacific Community –represented by Mrs Valérie Kagy-Cargnelli (Plant Pathology andPostharvest Scientist, IAC/CIRAD)
Dr Emile Frison (Director, INIBAP) and Ms Suzanne Sharrock (GermplasmConservation Scientist) of INIBAP HQprovided guidance while Dr AgustinMolina (Regional Coordinator) andVersalynn Roa (Technical/AdministrativeAssistant) of INIBAP for Asia/Pacificserved as secretariat. A facilitator, Mr Michael Carter of the University ofWolverhampton (UK) was present toguide the participants on the use of logi-cal frameworks in the organization andprogramme planning of BAPNET. Therepresentatives of Vietnam and Pakistanwere not able to attend but still remainas members of the network.
IMTP III training conducted in MalaysiaA training session, jointly organized byINIBAP Headquarters and BAPNET (for-merly INIBAP-ASPNET), in collaborationwith the Malaysian Agricultural Researchand Development Institute (MARDI),International Plant Genetic ResourcesInstitute – Asia, Pacific and Oceania(IPGRI-APO) office at Serdang, Malaysia,and the Centre de coopération interna-tionale en recherche agronomique pourle développement (CIRAD) was con-ducted on June 18-22, 2001 at theRenaissance Palm Garden Hotel inPutrajaya and at MARDI Headquarters inSerdang, Malaysia. A total of 28 partici-pants from 15 countries attended thecourse: seven came from Latin Americaand the Caribbean, two from West andCentral Africa and 19 from Asia.
The training focused on the evaluationof banana leaf spot diseases and methodsof collecting data. It was conducted priorto the launch of Phase III of theInternational Musa Testing Programme(IMTP) so as to allow the standardizationof methods of collecting data on agron-omy, pathology and other areas. It alsoaimed to train research personnel in leafspot disease evaluation, including therecognition of different fungus species.
As part of the course there were lec-tures, practical laboratory work, visits tobanana plantations of various capacitiesand to the MARDI banana germplasm col-lection, where some of the training indata and sample collection and symptomrecognition took place. The trainers wereDrs Jean Carlier and Marie-Françoise
Zapater from CIRAD and Jean-VincentEscalant from INIBAP, France. Dr SitiHawa Jamaluddin of MARDI and formerrepresentative of Malaysia to theRegional Advisory Committee, served asthe local organizer.
COGENT-INIBAP collaboration:Intercropping banana under coconut-basedfarming systems Banana has been chosen as an intercropto increase the income and food supply ofresource-poor coconut growers. INIBAP-Asia/Pacific and the InternationalCoconut Genetic Resources Network(COGENT) launched a collaborative pro-ject in the Philippines aiming toenhance the livelihood of farmers whosemajor income is from producing coconut.Banana is a high value year-round crop,which may be viably grown inside thecoconut groves. COGENT’s belief is thatthrough increasing the income ofcoconut farmers they will effectively pro-mote the conservation of coconutgermplasm.
The project is largely funded by anAsian Development Bank (ADB) grant toCOGENT. Together with INIBAP-Asia/Pacific, COGENT will study thetechnical feasibility and socioeconomicviability of intercropping banana in newand existing coconut plantations in 14 project sites in the Philippines. Threesites involve establishing new coconutplantings, while 11 sites are located inexisting coconut plantations. Thebanana crop will generate income in theformer sites whilst the coconut grow intofruit-bearing trees, but immediately aug-ment the incomes of coconut farmers inthe latter sites.
The Philippine Coconut Authority(PCA) is responsible for implementingthe work and INIBAP will provide assis-tance through the provision of disease-free tissue-cultured planting materialsand training of farmer-cooperators inestablishing nurseries, planting, inte-grated pest management and other cul-tural practices concerning banana. Atpresent, tissue-cultured bananaseedlings are growing in the project siteof Dolores, Quezon; Sto Niño, Cagayan;San Jose, Nueva Ecija; and Ligao, Albay.
Visiting scientist onmolecular characterization in GuadeloupeINIBAP sponsored the visit of Dr RachelSotto, a Filipino scientist working onMusa balbisiana in the University of thePhilippines, to Guadeloupe for a trainingon molecular characterization usingmicrosatellites for Musa with emphasison Musa balbisiana, on August 7-25, 2001under the supervision of Dr FrançoiseCarreel of CIRAD. The training focusedmainly on DNA extraction. Other activi-ties included DNA quantification, prepa-ration of agarose gel, PCR, electrophore-sis and visualization of bands. Themolecular technique taught was onCleaved Amplified PolymorphicSequence (CAPS) using chloroplast DNA.The knowledge and skills acquired duringthe training would later be used to fur-ther characterize the variation observedin the Philippine Musa balbisianagermplasm collection.
Training activities in LatinAmerica and the Caribbean In the framework of the project “Trainingand research for integrated management
50 INFOMUSA — Vol 10, N° 2
Dr Jean Carlier from CIRAD shows how to recognize Mycosphaerella spp. to the participants of the IMTP III training course (A. Molina, INIBAP).

of black Sigatoka on plantain in LatinAmerica and the Caribbean”, funded bythe Fondo Regional de TecnologíaAgropecuaria (FONTAGRO), two train-ing courses took place:
From 17 to 25 July 2001. “Workshop ontraining and research for integrated man-agement of black Sigatoka on plantain inLatin America and the Caribbean, to whichparticipated 25 people from 10 countries:Brazil, Colombia, Costa Rica, DominicanRepublic, Ecuador, Honduras, Mexico,Nicaragua, Panama and Venezuela.
From 3 to 7 September 2001.“International course on methods for theanalysis of the sensibility of M. fijiensisto fungicides and the establishment ofmonitoring methods”. Fifteen partici-pants came from nine countries of theregion: Brazil, Colombia, Costa Rica,Dominican Republic, Ecuador, Mexico,Nicaragua, Panama and Venezuela.
From 29 October to 3 November, INIBAP coorganized with the CubanMinistry of Agriculture a training courseon Plantain production.
MUSALAC meeting in HondurasThe 2nd Meeting of the Plantain andBanana Research and DevelopmentNetwork for Latin America and theCaribbean (MUSALAC) was held fromAugust 6 to 10, 2001 at the FundaciónHondureña de Investigación Agrícola(FHIA) in San Pedro Sula, Honduras.Thirty-five scientists from 13 countries ofthe region participated in this meeting,representing Bolivia, Brazil, Colombia,Costa Rica, Cuba, Dominican Republic,Ecuador, Honduras, Mexico, Nicaragua,Panama, Puerto Rico and Venezuela andthree international and regional institu-tions (CATIE, IICA and INIBAP).
Drs Altagracia Rivera de Castillo(President of MUSALAC), Miguel AngelBonilla (Vice Minister of Agriculture,Honduras), Adolfo Martínez (DirectorGeneral of FHIA, Honduras) and FranklinRosales (INIBAP-LAC’s RegionalCoordinator and MUSALAC’s executivesecretariat) opened the meeting. Themain topic of the meeting was the tech-nological offer and all the country andinstitutional representatives had theopportunity to present technologies, ser-vices or products available through theirinstitutions. Special guests made presen-tations on topics of general interest suchas nematodes, socioeconomics, organicbanana production. The Network’sSteering Committee also had its annualmeeting and Dominican Republic wasdesignated as the venue for the 2002MUSALAC meeting.
INFOMUSA — Vol 10, N° 2 51
MusaDoc CD-RomThe third edition of the MusaDoc CD-Rom, MusaDoc 2001 is now available.Updated versions of the INI-BAP databases, MUSALIT,containing over 6000 bib-liographic records andabstracts on Musa,and BRIS, the data-base on bananaresearchers, are bothsearchable on the CD-Rom. All recent publi-cations are alsoincluded in MusaDoc2001 - new factsheets onpests and diseases, INIBAPannual report 2000, two new publications
from the Asia and Pacific region on“Advancing banana and plantain R & D
in Asia and the Pacific” and“Managing banana and
citrus diseases”, newtechnical guidelines on“Cryoconservation ofMusa germplasm”,and the second edi-tion of Musalogueentitled “Diversity of
the genus Musa”. TheCD-Rom also provides an
updated summary of INI-BAP’s activities.
The CD-Rom is available fromINIBAP Headquarters.
Books etc.
Participants of the training workshop on integrated management of black Sigatoka (CATIE, Costa Rica,17-25 July 2001)
Participants to the 2nd MUSALAC meeting (F. Rosales, INIBAP).

Advancing banana andplantain R & D in Asia and the Pacific, Vol. 10Proceedings of the 10th INIBAP-ASPNET Regional AdvisoryCommittee meeting held inBangkok, Thailand, 10-11November 2000
Edited by A.B. Molina, V.N. Roa and M.A. Maghuyop
For the second time, the proceedings ofthe 10th INIBAP-ASPNET RegionalAdvisory Committee meeting are pub-lished in a 162-page-book. This publica-tion provides most up-to-date informa-tion on Musa research, including threetechnical presentations on breeding,nematodes and integrated pest control.It also includes an update on currentresearch and perspectives in severalcountries of the region, among whichAustralia, China, India, Indonesia,Pacific Islands, Malaysia, thePhilippines, Sri Lanka and Taiwan, and areport on the ASPNET network’s activi-ties since its last meeting.
Copies are available from the INIBAPRegional Office in the Philippines.
Factsheets on Musa diseasesand pestsAs announced in the last issue of INFOMUSA, factsheets on Musa dis-eases No. 9 and 10, on “False Panamadisorder on banana” and “Wilt-like dis-order of bananas in Uganda” are nowavailable. The fifth factsheet on Musapests is dedicated to “The banana stem
weevil Odoiporus longicollis” and was prepared by B. Padmanaban and S. Sathiamoorthy. It describes Odoiporuslongicollis Oliver (Coleoptera: Cur-culionidae) and provides information on
its biology, distribution and biologicalcycle, the damages caused by the weeviland the control methods (cultural,chemical and biological).
Copies of all factsheets are availablefrom INIBAP Headquarters.
52 INFOMUSA — Vol 10, N° 2
Announcements
The Colombian Association of BananaGrowers, AUGURA, and the Associationfor the Cooperation on Banana Researchin the Caribbean and Tropical America,ACORBAT international, are pleased toannounce the preparation of the XVth
ACORBAT World meeting to be held inCartagena de Indias, Colombia, from 27 October to 2 November 2002, accord-ing to the commitment made during theXIVth Association meeting in Puerto Ricoin 2000.
ACORBAT 2002 will focus on the sus-tainability of the banana and plantainsector and will discuss the following sub-jects: Agronomy; Plant protection; Plantgenetics, improvement and biotechnol-ogy; Physiology; Postharvest manage-ment; Agroindustry; Economics and mar-keting; Environmental impact; Regionalanalysis and Soil biochemistry.
The fascinating city of Cartagena deIndias, founded in 1533, was declared a“World Heritage Site” by UNESCO and has
hosted many international meetings. Thecity boasts scenery of great naturalbeauty, as well as a very special historyand charm, offering museums, colonialstreets and historic buildings, all with adistinctive Caribbean flavour. Full secu-rity measures are ensured. Visit the Website at:http://www.cartagenadeindias.com/.
The XVth meeting of ACORBAT is fore-seen as one of the most important eventsfor the banana and plantain sector in2002 and an exceptional platform forinteraction for all stakeholders from thebanana and plantain production sectorthroughout the world.
For more information, visit AUGURA’s Web siteat: www.augura.com.coContact: Sabina Alvarez E. E-mail: [email protected]: (574) 321 13 33 – Fax: (574) 321 41 90
Other important meetingsare also announced on p. 12of the PROMUSA section.
XVth meeting of ACORBAT(1st announcement)27 October-2 November 2002,Cartagena de Indias, Colombia
HAPPY NEWYEAR
from all INIBAPstaff
2002

The Instituto de Biotecnología de lasPlantas (IBP) is organizing the VIth
International symposium on plantbiotechnology, to be held from 17 to 21June 2002. The programme includes con-ferences, round tables, workshops andposters sessions, as well as commercialexhibits and an important meeting of
PROMUSA. The events will take place atthe hotel “Villa La Granjita”, 3 km fromSanta Clara.
Major themes include: Genetic trans-formation and molecular biology; Celland tissue culture; Somatic embryogene-sis and artificial seeds; Mass propaga-tion; Plant breeding based on somaclonal
variation, mutagenesis and in vitroselection; Decontamination and diagno-sis of pathogenic microorganisms;Microbial contamination in in vitro tissue cultures; Production of secondarymetabolites; Information, marketing andintellectual property in the area of plantbiotechnology.
Registration feesDelegates: 150 US$ Students: 75US$ Accompanying person: 50US$ For more information, contact:Lic. Orlando Gregorio ChavianoSecretariado ejecutivo IPBCarretera a Camajuani Km 5Santa Clara CP 54 830 Villa Clara, CubaTel.: 53 42 281257Fax: 53 42 281329Email: [email protected]@yahoo.comhttp://www.cuba.cu/ciencia/ibp/home.htm
INFOMUSA — Vol 10, N° 2 53
All you want to know about
BANANASon a multimedia
CD-ROMThis multimedia CD-ROM on bananashas been produced as a joint effort ofthe International Network for theImprovement of Bananas and Plantain(INIBAP) and the Centre de coopérationinternationale en recherche agronomiquepour le développement (CIRAD).
Organized in seven chapters, this very attractive CD-ROM presents information on all the aspects of the croplavishly illustrated with films, photographs, maps, comics, etc.
Demonstrated during the EXPO 2000 in Hanover, Germany (August 2000) and during the Banana and Pineapple professional meeting organized by CIRAD-FLHOR in Montpellier, France (September 2000).
Limited edition.Buy it now hot from thepress !
Rates :Price for one copy 25 US $5 copies or more 15 US $ per copy
Please return your orderand payment to: INIBAPParc Scientifique Agropolis II, 34 397 Montpellier Cedex 5, FranceTel.: 33 - (0)4 67 61 13 02 Fax: 33 - (0)4 67 61 03 34Email: [email protected]
• The world’s largest herb
• The much-travelled
banana
• Banana – a basic food
• The industrial chain of
dessert banana
• Watch that plant grow!
• Protecting the banana
and its future
• Banana as veg or
as dessert?
VIth Internationalsymposium on plantbiotechnology (2nd announcement)17-21 June 2002, Santa Clara, Cuba
The Instituto de Investigaciones deSanidad Vegetal (INISAV), the majorCuban research centre in the area of plantprotection, offers in 2002 a cycle of train-ing activities that are specifically targetedat researchers, teachers and all thoseinterested in the phytosanitary aspects intropical agriculture. This cycle includesconferences, workshops, discussions andscientific and cultural trips. Part of the
time will be dedicated to hands-on train-ing so as to allow each participant toreceive a practical training. At the end ofthe cycle, participants will receive a cer-tificate from INISAV, which is accreditedby the Ministry of Higher Studies.
International courses (40-hour each)include the following themes: Advancedtechnologies for the control of horticul-tural crops: impact and perspectives;
Breakthroughs in phytopathology;Phytosanitary surveillance; Managementof resistance to fungicides: impact on thereduction of fungicides application oneconomically important crops; Biologicalcontrol of crops diseases; Internationalcourse on plant sanitary status and tax-onomy of microorganisms potentially use-ful in biological control.
For more information, contact:Dr Emilio Fernández GonzálvezSubdirector División de MicrobiologíaCalle 110 No 514, e/ 5ta B y 5ta F.Miramar, Playa, Ciudad de La HabanaFax: (537)240535, 229366Tel.: (537)294753Email : [email protected]://www.ceniai.inf.cu/ciencia/inisav
INISAV training activities in 2002

The International Institute of TropicalAgriculture (IITA) invites applicationsfor two Postdoctoral Fellows for workon Musa. (1) a Molecular Geneticist to be based at its Headquarters in Ibadan, Nigeria and (2) aBreeder/Pathologist to be based atIITA’s Eastern and Southern AfricaRegional Center, in Kampala, Uganda.
IITA is one of the 16 internationalagricultural research institutes of theConsultative Group on InternationalAgricultural Research (CGIAR),which is co-sponsored by the WorldBank, the Food and AgricultureOrganization of the United Nations,the United Nations DevelopmentProgramme and the InternationalFund for Agricultural Development.The Institute has programmes inmany countries of sub-Saharan Africa.IITA scientists from over 30 countrieswork in partnership with nationalagricultural agencies and farmers toincrease agricultural production intropical Africa, and ultimately to raisethe productivity and income of small-scale farmers in an ecologically sus-tainable way. For more information onIITA, visit our Web site at:http://www.iita.org
Banana and plantain constitutemajor food sources in Africa, grownmainly by small-scale farmers withincomplex cropping systems. Major con-straints to production include suscep-tibility of most traditional cultivars toblack Sigatoka disease, Fusarium wilt,various nematode species, and viruses.IITA has developed a strong breedingprogramme to produce resistant culti-vars using a combination of conven-tional cross-breeding and molecular
breeding techniques. Banana and plan-tain improvement is a relatively under-researched area that involves interspe-cific hybridization, genome and ploidymanipulation.
(1) Molecular GeneticistThe emphasis of the position will be onidentifying molecular markers inbanana and plantains for importantagronomic traits including partheno-carpy, apical dominance, and resis-tance to black Sigatoka and nema-todes. The individual will also beresponsible for genome analysis,germplasm characterization, finger-printing and detection of somaticinstability of hybrids.
The ideal candidate for the positionof Musa Molecular Geneticist will havethe following: a recent PhD degree(obtained within the last two years) inmolecular genetics including demon-strated skills in DNA markers (RAPD,AFLP, SSRs among others), a goodunderstanding of polyploid genetics;an ability to work as a member of amulti-disciplinary team; and the abilityto work well in a cross-cultural setting.Fluency in English is required, while aworking knowledge of French, wouldbe a distinct advantage.
(2) Breeder/PathologistThe individual appointed to this posi-tion will assist and provide support to asenior breeder in the development ofnew improved banana cultivars andgermplasm with primary emphasis onhigh yield potentials, improved diseaseand pest resistance and adaptation tobiotic and abiotic stresses. In additionto assisting with on-going bananabreeding activities, the individual will
carry out independent research to sup-port the overall cultivar developmentgoals with increased emphasis on dis-ease and pest resistance breeding.
The ideal candidate for the positionof Banana Breeder/Plant Pathologistwill have the following: a recent PhDdegree (obtained within the last twoyears) in plant breeding with a minorin plant pathology (or vice versa), withsome knowledge in cytology and newtools of molecular breeding; an abilityto work as a member of a multi-disci-plinary team; the ability to work wellin a cross-cultural setting. Fluency inEnglish is required while a workingknowledge of French, would be a dis-tinct advantage. Field experience inthe humid tropics, particularly inAfrica would be highly desirable.
Salary and benefits are internation-ally competitive and include housing,personal car allowance, educationallowance for up to two children,annual home leave travel, and healthservices. Initial appointment will be fortwo years, extendable to three years.
Applicants for this position areinvited to send their curriculum vitae,date of availability, and names andaddresses of three professional refer-ees by email to Frances McDonald,Manager of Human Resources [email protected] or by fax toF. McDonald at 234-2-241-2221. Hardcopies may also be sent to her atten-tion through IITA’s international mail-ing address – Lambourn (UK) Ltd.,Carolyn House, 26 Dingwall Road,Croydon CR9 3EE, UK. Applicationswill be received until 28 February 2002.
IITA is an equal opportunity employerand would particularly welcome appli-cations from women candidates.
54 INFOMUSA — Vol 10, N° 2
INTERNATIONALINSTITUTE OF TROPICALAGRICULTUREPostdoctoral Fellows for work on Musa

INFOMUSA — Vol 10, N° 2 55
• HeadquartersParc Scientifique Agropolis II34397 Montpellier Cedex 5 – FRANCEe-mail: [email protected]://www.inibap.orgDirectorDr Emile FRISONe-mail: [email protected] Genetic Resources ScientistDr Jean-Vincent ESCALANTe-mail: [email protected] Conservation ScientistMs Suzanne SHARROCKe-mail: [email protected] Information/CommunicationsMs Claudine PICQe-mail: [email protected] in charge MGISMs Elizabeth ARNAUDe-mail: [email protected] ManagerMr Thomas THORNTONe-mail: [email protected]
• Regional Office for Latin America and the CaribbeanRegional CoordinatorDr Franklin E. ROSALESAssociate Scientist, Musa technology transferLuis POCASANGRE
C/o CATIEApdo 60-7170 Turrialba, COSTA RICATel/Fax: (506) 556 2431e-mail: [email protected]
• Regional Office for Asia and thePacificRegional CoordinatorDr Agustín MOLINAC/o IRRI Collaborators Center3rd Floor, Los Baños, Laguna 4031PHILIPPINESFax: (63-49) 536 05 32e-mail: [email protected]
• Regional Office for West andCentral AfricaRegional CoordinatorDr Ekow AKYEAMPONGAssociate Scientist, Technology transferKim JACOBSENC/o CRBP – BP 12438Douala, CAMEROONFax: (237) 342 91 56e-mail: [email protected]
• Regional Office for Eastern andSouthern AfricaRegional CoordinatorDr Eldad KARAMURA
Associate Scientist, Technology transferGuy BLOMMEPO Box 24384KampalaUGANDAFax: (256-41) 28 69 49e-mail: [email protected]
• INIBAP Transit Center (ITC)Officer in chargeMs Ines VAN DEN HOUWEKatholieke Universiteit LeuvenLaboratory of Tropical Crop ImprovementKasteelpark Arenberg 13,B-3001 LeuvenBELGIUMFax: (32-16) 32 19 93e-mail: [email protected]
• Associate Expert, NematologyMr Thomas MOENSC/o CORBANALa Rita Research StationApdo 390-7210Guápiles, COSTA RICAFax: (506) 763 30 55e-mail: [email protected]
Typescripts should be prepared inEnglish, French or Spanish and submit-ted in duplicate to the Managing Editor.They should be double-spaced through-out. All pages (including tables figures,legends and references) should be num-bered consecutively. Include the fullname of all the authors of the paper,together with the addresses of theauthors at the time of the work reportedin the paper. Indicate also the authornominated to receive correspondenceregarding the paper.
If the typescript was prepared on acomputer, please send a copy on diskette(or by e-mail) along with the printedones, indicating the name and version ofthe wordprocessor used. • Abstracts: An abstract not exceeding
200-250 words should be sent in thesame language as the typescript, aswell as translations (including thetitle) into the two other languages, ifthis is possible.
• Acronyms: These should be written infull the first time they appear in the
text, followed by the acronym inparenthesis.
• References: All literature referencesmade in the text should be referred toby author(s) and year of publication(e.g.: Sarah et al. 1992, Rowe 1995). Alist of references, in alphabeticalorder, should be provided at the end ofthe text.
Please follow the style shown below:Periodicals: Sarah J.L., C. Blavignac &
M. Boisseau. 1992. Une méthode delaboratoire pour le criblage variétaldes bananiers vis-à-vis de la résistanceaux nématodes. Fruits 47(5): 559-564.
Books: Stover R.H. & N.W. Simmonds.1987. Bananas (3rd edition). Longman,London, United Kingdom.
Articles (or chapters) in books: Bakry F.& J.P. Horry. 1994. Musa breeding atCIRAD-FLHOR. Pp. 169-175 in TheImprovement and Testing of Musa: aGlobal Partnership (D.R. Jones, ed.).INIBAP, Montpellier, France.
Tables: These should be numbered con-secutively and referred to by these num-
ber in the text. Each table shouldinclude a title.Illustrations: These should be numberedconsecutively and referred to by thesenumbers in the text. Each illustrationshould include a clear and simple caption.Graphs: provide the corresponding rawdata with the graphs.Drawings: provide originals if this ispossible.Black and white photographs: providethem on bright paper and with good con-trast.Colour photographs: provide good qual-ity proofs and films or original slides.Note: When plant material used for theexperiments reported originates or is reg-istered in the INIBAP genebank, its acces-sion number (ITC code) should be indi-cated within the text or in a tabular form.
Thank you in advance for followingthese instructions
This will facilitate and acceleratethe editing work.
Instructions to authors
INIBAP addresses

The following publications are available fromheadquarters:INIBAP/CTA/CIRAD 2001. J. Daniells, C. Jenny, D. Karamura & K. Tomekpe. Musalogue: a
catalogue of Musa germplasm. Diversity in the genus Musa (E. Arnaud & S. Sharrock,compil.).
INIBAP/CTA 2001. B. Panis & N.T. Thinh. Cryopreservation of Musa germplasm (J.V. Escalant et S. Sharrock, eds). INIBAP Technical Guidelines 5.
INIBAP 2001. Networking bananas and plantains: Annual Report 2000.CIRAD/INIBAP 2000. Bananas.INIBAP 2000. M. Holderness, S. Sharrock, E. Frison & M. Kairo (eds). Organic banana
2000: Towards an organic banana initiative in the Caribbean. Report of the interna-tional workshop on the production and marketing of organic bananas by smallholderfarmers. 31 October-4 November 1999, Santo Domingo, Dominican Republic.
INIBAP 2000. G. Orjeda (compil.). Evaluating bananas: a global partnership. Results ofIMTP Phase II.
INIBAP/EARTH/IDRC 1999. F.E. Rosales, S.C. Tripon & J. Cerna (eds). Organic/environ-mentally friendly banana production. Proceedings of a workshop held at EARTH,Guácimo, Costa Rica, 27-29 July 1998.
INIBAP/CRBP/CTA/CF 1999. C. Picq, E. Fouré & E.A. Frison (eds). Bananas and foodsecurity/Les productions bananières: un enjeu économique majeur pour la sécurité ali-mentaire. Proceedings of an International Symposium held in Douala, Cameroon, 10-14 November 1998.
INIBAP/FHIA 1999. F.E. Rosales, E. Arnaud & J. Coto (eds). A tribute to the work of PaulH. Allen: a catalogue of wild and cultivated bananas.
INIBAP/RF/SDC 1999. E.A. Frison, C.S. Gold, E.B. Karamura & R.A. Sikora (eds). MobilizingIPM for sustainable banana production in Africa. Proceedings of a workshop on bananaIPM held in Nelspruit, South Africa, 23-28 November 1998.
INIBAP 1999. E. Akyeampong (ed.). Musa Network for West and Central Africa. Report ofthe second Steering Committee meeting held at Douala, Cameroon, 15-16 November1998.
INIBAP 1999. K. Shepherd. Cytogenetics of the genus Musa.INIBAP 1998. E. Akyeampong (ed.). Musa Network for West and Central Africa. Report of
the first Steering Committee meeting held at Douala, Cameroon, 8-10 December 1997.INIBAP 1998. E.A. Frison & S.L. Sharrock (eds). Banana streak virus: a unique virus-Musa
interaction? Proceedings of a workshop of the PROMUSAvirology working group heldin Montpellier, France, 19-21 January 1998.
INIBAP 1998. C. Picq (ed.). Segundo seminario/taller de la Red regional de informaciónsobre banano y plátano de America Latina y el Caribe. San José, Costa Rica, 10-11 July1997.
INIBAP 1998. B.K. Dadzie. Post-harvest characteristics of black Sigatoka resistantbanana, cooking banana and plants hybrids. INIBAP Technical Guidelines 4.
INIBAP 1998. G. Orjeda in collaboration with the PROMUSA working groups on Sigatokaand Fusarium. Evaluation of Musa germplasm for resistance to Sigatoka diseases andFusarium wilt. INIBAP Technical Guidelines 3.
CIRAD/INIBAP 1998. Les bananes.
INIBAP/ACIAR 1997. E. Arnaud & J.P. Horry (eds). Musalogue, a catalogue of Musagermplasm: Papua New Guinea collecting missions 1988-1989.
INIBAP/CTA/FHIA/NRI/ODA 1997. B.K. Dadzie & J.E. Orchard. Post-harvest RoutineScreening of Banana and Plantain Hybrids: Criteria and Methods. INIBAP TechnicalGuidelines 2.
INIBAP/CTA 1997. P.R. Speijer & D. De Waele. Screening of Musa Germplasm forResistance and Tolerance to Nematodes. INIBAP Technical Guidelines 1.
INIBAP/The World Bank 1997. E.A. Frison, G. Orjeda & S. Sharrock (eds). PROMUSA: AGlobal Programme for Musa Improvement. Proceedings of a meeting held in Gosier,Guadeloupe, March 5 and 9, 1997.
INIBAP-IPGRI/CIRAD. 1996. Descriptors for Banana (Musa spp.).
The following publications are available from Asia and the Pacific office:INIBAP-ASPNET 2001. A.B. Molina, V.N. Roa & M.A.G. Maghuyop (eds). Advancing banana
and plantain R & D in Asia and the Pacific Vol. 10. Proceeding of the 10th INIBAP-ASP-NET Regional Advisory Committee (RAC) meeting held at Bangkok, Thailand, 10-11November 2000.
INIBAP-ASPNET/MARDI 2001. A.B. Molina, N.H. Nik Masdek & K.W. Liew (eds). BananaFusarium wilt management: towards sustainable cultivation. Proceedings of the inter-national workshop on the management of Fusarium wilt disease held in Genting,Malaysia, 18-20 October 1999.
INIBAP-ASPNET 2000. A.B. Molina & V.N. Roa (eds). Advancing banana and plantain R & D in Asia and the Pacific. Proceedings of the 9th INIBAP-ASPNET RegionalAdvisory Committee meeting held at South China Agricultural University, Guangzhou,China, 2-5 November 1999.
INIBAP-ASPNET/FFTC 2000. A.B. Molina, V.N. Roa, J. Bay-Petersen, A.T. Carpio & J.E.A.Joven (eds). Managing banana and citrus diseases. Proceedings of a regional workshopon disease management of banana and citrus through the use of disease-free plantingmaterials held in Davao City, Philippines, 14-16 October 1998.
INIBAP-ASPNET 2000. R.V. Valmayor, S.H. Jamaluddin, B. Silayoi, S. Kusumo, L.D. Danh,O.C. Pascua & R.R.C. Espino. Banana cultivar names and synonyms in Southeast Asia.
INIBAP-ASPNET 1999. V.N. Roa & A.B. Molina (eds). Minutes: Eighth meeting of INIBAP-ASPNET Regional Advisory Committee (RAC) hosted by the Queensland HorticultureInstitute (DPI) in Brisbane, Australia, 21-23 October 1998.
INIBAP-ASPNET 1998. Minutes: Seventh meeting of INIBAP/ASPNET Regional AdvisoryCommittee (RAC) hosted by the Vietnam Agricultural Science Institute (VASI) inHanoi, Vietnam, 21-23 October 1997.
INIBAP-ASPNET 1997. V. N. Roa & R. V. Valmayor (eds). Minutes: Sixth meeting of INI-BAP-ASPNET Regional Advisory Committee (RAC) hosted by National ResearchCenter on Banana (ICAR) in Tiruchirapalli, India, 26-28 September 1996.
INIBAP-ASPNET 1996. R. V. Valmayor, V. N. Roa & V. F. Cabangbang (eds). RegionalInformation System for Banana and Plantain - Asia and the Pacific (RISBAP):Proceedings of a consultation/workshop held at Los Baños, Philippines, 1-3 April 1996.(ASPNET Book Series No. 6).
Publications from INIBAP
ww
w.inib
ap.o
rg

The scientists taking part in the collabora-tion to advance Musa genome studies metat the National Science Foundation (NSF)in Arlington, USA, last July to launch theformally-named Global Musa GenomicsConsortium. This follows a series of plan-ning meetings and responds to the recom-mendations arising out of the PROMUSAmeeting, which took place in Thailand inNovember 2000. The Consortium is madeup of 26 public-funded institutes from 13 countries, including developed and lessdeveloped nations. NSF provided the fun-ding for scientists from developing coun-tries to attend the meeting.
The purpose of the 3.5-day gatheringwas to discuss the strategy, functioning,rules and division of tasks of the group.Agreement was made in the setting up of amanagement committee and a secretariat,which will be performed by INIBAP. It isalso planned that an external advisorycommittee will be established. The focusof discussions on research centred on pro-moting and advancing the work on map-ping, cDNA and BAC libraries, genomesequencing, use of mutants and bioinfor-matics. Funding sources were discussedand a number of research proposals areunder development.
Since the meeting took place many part-ners have been highly active. In particular,institutes in Brazil have met and formedMusagene-BR, a national network forMusa genomics, under the leadership of Empresa Brasiliera de PesquisaAgropecuaria (EMBRAPA) and theUniversidade Católica de Brasilia (UCB).
During their discussions, they were able toconsolidate the main research areas,where they will contribute to the GlobalMusa Genomics Consortium, to identifysources of funding and develop proposals.European partners have also developed aproposal for funding by the EuropeanCommission to support their research andthe setting up of an information portalwhich will provide access to up-to-dateinformation and results on the Web.
The press response to the launching ofthe initiative has been exceptional.Through Future Harvest, tens of journalistswere able to interview members of theconsortium, resulting in well over 100articles and broadcasts being made in atleast nine languages in newspapers, onthe Web, television and radio throughoutthe world. Their interest concentrated onthe importance of banana in world foodsecurity and the fact that the Consortiumintends to ensure all results and products,no matter their commercial value, will bemade freely available for the use of thesmallholder farmer.
PROMUSA I
PROMUSAA global Programme for Musa Improvment
INFOMUSA — Vol 10, N° 2
PROMUSA N° 8
ContentsGenome busting - Launching the Global
Musa Genomics Consortium . . . . . . . .p. IUganda Biotechnology project –
2nd annual planning meeting . . . . . . . .p. IIPROMUSA Nematology working group
meeting . . . . . . . . . . . . . . . . . . . . . . . . .p. II• Abstracts of papers and posters . . . . . .p. VISince the 2nd PROMUSA global meeting
in 1998 . . . . . . . . . . . . . . . . . . . . . . . . .p. IXMeeting announcements . . . . . . . . . . .p. XII• 2nd International workshop on Sigatoka
leaf spot diseases of bananas . . . . . . .p. XII• Meeting of the PROMUSA Sigatoka
working group . . . . . . . . . . . . . . . . . . .p. XII• Meeting of the PROMUSA Nematology
working group . . . . . . . . . . . . . . . . . . .p. XII• 3rd International symposium on the
Molecular and cellular biology of bananas . . . . . . . . . . . . . . . . . . . . . .p. XII
• Meeting of the PROMUSA Genetic improvement and Virology working groups . . . . . . . . . . . . . . . . . . . . . . . . .p. XII
What is PROMUSA ?The Global Programme for Musa
Improvement (PROMUSA) is a broad-basedprogramme which aims at involving all themajor players in Musa improvement. It wasdeveloped as a means to link the work carriedout towards addressing the problems of export banana producers, with those initiatives directed towards improving bananaand plantain production at the subsistence and smallholder level. The global programmebuilds upon existing achievements and isbased upon ongoing research initiatives. PROMUSA is therefore a mechanism to further maximize the outputs and acceleratethe impact of the overall Musa improvementeffort. The programme is an innovative mechanism to bring together research carriedout both within and outside the CGIAR, creating new partnerships between NationalAgricultural Research Systems (NARS) andresearch institutes in both developing anddeveloped countries. The formation of suchpartnerships will also contribute to strengthening the capacity of NARS to conduct Musa-related research.The major thrust of PROMUSA is to develop a wide range of improved banana varietiesfrom which growers worldwide can selectthose most suited to their needs. The programme brings together conventionalbreeding based on hybridization techniqueswith genetic engineering and biotechnological breeding approaches. Thisbroad-based genetic improvement effort issupported by research being carried out onspecific pests and diseases within the various PROMUSA working groups. An efficient mechanism for evaluating new varieties produced within the framework ofPROMUSA is also an essential component ofthe programme.
Genome busting - Launching the Global MusaGenomics ConsortiumArlington, USA, 17-20 July 2001

The second planning meeting of theUganda Biotechnology project was held inKampala from 26-28 September 2001. Thismeeting brought together representatives ofthe various project partners, includingNARO, IITA, INIBAP, KUL and theUniversity of Makerere. The meeting focu-sed on reviewing the progress made in thefirst year of the project and discussing workplans for the second year.
It was generally agreed that the project ismaking good progress technically, with acti-vities related to both the production of cellsuspension and genetic transformationmoving forward rapidly. A range of eight dif-ferent varieties on East African HighlandBananas (EAHB) of local importance havebeen selected and established in the field inUganda. These varieties are used to providemale flowers as starting material for cell sus-pension cultures that are the starting pointfor genetic transformation work. Proliferatingcultures, scalps and male flowers are beinginoculated in tissue culture routinely.Capacity for the establishment and mainte-nance of cell suspensions has been reinfor-ced at the NARO research station inKawanda, where the existing facilities havebeen upgraded and the necessary chemi-cals and equipment purchased and installed.
Development of a genetic transformationsystem on EAHB started in January 2001 atKULeuven in Belgium, where an UgandanPhD student is carrying out the work usingthe Agrobacterium-mediated transformation(A-MT) system. Transformation experimentshave been carried out using four differentcultivars, and transient and stable expres-sion of gfp and gusA reporter genes hasbeen achieved.
On the administrative side, as well as theproject personnel that have been appointed(See INFOMUSA 10(1):45), a TechnicalCoordinating Committee has been set up atthe national level in Uganda. This commit-tee, which aims to provide guidance to theproject, includes representatives of theUganda Consumer Protection Association,the Uganda National Council for Science &Technology and the National EnvironmentalManagement Agency. In addition, privatesector research and the different projectpartners are also represented on the com-mittee.
During the planning meeting, biosafetyissues were discussed at length and theneeds of the project in this respect were
II PROMUSA INFOMUSA — Vol 10, N° 2
Participants to the 2nd planning meeting of the Ugandan Biotechnology project.
Uganda Biotechnology project – 2nd annual planning meeting26-28 September 2001, Kampala
The third meeting of the Nematology wor-king group (NWG) was held on 24-25 May,2001 in Hulala Lakeside Lodge, Hazyview,South Africa, on the occasion of the 15th
Symposium of the Nematological Society ofSouthern Africa (NSSA).
ParticipantsDirk De Waele (KU Leuven, Belgium, NWGconvenor) and Jean-Vincent Escalant(INIBAP/PROMUSA), chairmen.
Anne-Marie Auwerkerken (IITA, Nigeria),Danny Coyne (IITA, Nigeria),Jose Sancho Cumbi (INIA, Mozambique),Mieke Daneel (ARC-ITSC, South Africa),Karen de Jager (ARC-ITSC, South Africa),Carine Dochez (IITA, Uganda),Emilio Fernandez (INISAV, Cuba),Svetlana Gaidashova (ISAR, Rwanda),Philippe Gnonhouri (CNRA, Côte d’Ivoire),Liao Jinling (SCAU, China),Eldad Karamura (INIBAP/BARNESA,Uganda),
taken into account in the development ofYear 2 work-plans. The need to initiate theapplication for the introduction of geneticallytransformed plants was considered urgent.In this respect, a containment facility will berequired as a priority. Considering the levelof funds presently available for Year 2 of theproject, it was agreed that the equipping ofthe molecular biology laboratory would bedelayed until Year 3. However, the renova-tion of the building that will be used formolecular biology research will continue asplanned.
Regarding the characterization of resis-tance to weevils, a PhD student has recentlybeen recruited and this work will be carriedout at Makerere University, in collaborationwith IITA. Also in the framework of the pro-ject, a PhD student is due to start researchshortly at the University of Pretoria, working
on the identification of sources of weevilresistance. Once this research commences,the University of Pretoria will automaticallybecome a new project partner.
With regard to the technical work for Year 2, it was agreed that the cell suspen-sion work and genetic transformation forblack Sigatoka resistance should continue asplanned. In addition, the work on the poten-tial of lectins in respect to nematode resis-tance at KULeuven will continue, but if positi-ve results are obtained by the University ofLeeds on proteinase inhibitors, efforts will bemade to incorporate this approach into theproject. The potential of these and otherapproaches to nematode resistance, inclu-ding the use of secondary metabolites beingstudied at KUL, will be assessed at the endof the second year and a strategy for thispart of the project developed at this time.
PROMUSA Nematology working group meeting 24–25 May 2001, South Africa

Thomas Moens (CORBANA, Costa Rica),Josephine Namaganda (NARO, Uganda),Luis Pocasangre (INIBAP/CATIE, CostaRica),Patrick Quénéhervé (IRD, Martinique),
Khadija Rajab (Department of Agriculture,Zanzibar),Jean-Louis Sarah (CIRAD-AMIS, France),Anita Severn-Ellis (ARC-ITSC, South Africa),Nguyen Thi Tuyet (VASI, Vietnam).
Status of nematology researchwithin NWGAll participants presented an outline of theirnematological research with regard to thethree research priorities: a) nematode com-munities and biodiversity, b) resistancescreening (including methods, sources andmechanisms), and c) damage and yield losspotential of nematode populations.
Improvement of workinggroup interactions
Mailing list
In PROMUSA, a general mailing list andspecific working groups’ mailing lists exist.Persons on the general mailing list will notnecessarily receive mail from the workinggroups’ mailing lists. Interaction willdepend on the members themselves, whe-ther they choose to be involved or not. Asa consequence, the success of the list will
INFOMUSA — Vol 10, N° 2 PROMUSA III
PROMUSA Nematology working group participants in South Africa.
Nematode communities and biodiversity
Country/RegionEmilio Fernandez • Main nematode species identified were Radopholus similis, Meloidogyne incognita and Pratylenchus coffeae. Present only in some areas were (Cuba) Helicotylenchus multicinctus and Rotylenchulus reniformis. Thomas Moens • Variation observed in the reproductive fitness of R. similis populations from different production areas in Costa Rica. Population dynamics of (Costa Rica) M. incognita (on Grande Naine and Gros Michel) are studied as well.Luis Pocasangre • Pratylenchus spp. are a bigger problem in plantain while R. similis is more a problem in banana. Apparently, Pratylenchus spp. cause more (Latin America) problems in Central America and the Caribbean than elsewhere.
• In the Dominican Republic, R. similis was found to be the major problem on banana. Pratylenchus spp. were more serious on plantain although R. similis, Helicotylenchus and Meloidogyne spp. were also found.
Patrick Quénéhervé • Musa nematode (mainly Meloidogyne) biodiversity studied as part of a project on Integrated Pest Management in collaboration with several Martinique/ national and international research groups (INRA, EMBRAPA, CIRAD-AMIS, CARBAP, PPRI).GuadeloupePhilippe Gnonhouri • Banana and plantain research has been neglected before. Nematodes are an important problem with P. coffeae now found widely spread in the (Côte d’Ivoire) Côte d’Ivoire. Svetlana Gaidashova • In Rwanda, Pratylenchus goodeyi and R. similis are found, however R. similis is never dominant. A baseline study of nematodes (and diseases) (Rwanda) on Musa is needed. Khadija Rajab • Nematodes recorded on banana are P. coffeae, Meloidogyne spp., R. similis, H. multicinctus and Helicotylenchus dihystera.(Zanzibar)Mieke Daneel • R. similis is found only in three areas. There is a distinct difference in the pathogenicity between R. similis populations from Natal and (South Africa) Mpumalanga.
• Other species found are Pratylenchus spp., Meloidogyne javanica and M. incognita, R. reniformis and H. multicinctus, with Helicotylenchusand Meloidogyne spp. found in almost every sample.
• A small-farmer survey showed the presence of Meloidogyne and Helicotylenchus spp. in all areas. R. similis is restricted to the Eastern Cape and Natal.
Nuygen Thi Tuyet • R. similis does not occur on banana in Vietnam even though the banana cultivars grown in Vietnam are susceptible to R. similis. R. similis(Vietnam) is however found on ginger, durian, groundnut, and coffee in the highlands. The biodiversity of these R. similis populations will be investigated.
Surveys will be undertaken in the central Vietnam highlands to collect more populations. • P. coffeae causes serious damage to banana in north and central Vietnam. Further studies will focus on the biodiversity of P. coffeae in Vietnam.
A survey is planned in north and central Vietnam to determine the occurrence of this nematode on various crops including banana.Jean-Louis Sarah • Studies on R. similis diversity continue. Approaches on regional diversity are now tested (i.e. Cameroon with CARBAP, Brazil with (France) EMBRAPA, etc.)
• A project on diversity among Pratylenchus coffeae has been undertaken.Liao Jinling • Main nematode species isolated from banana are R. reniformis (63%), Meloidogyne spp. (60%), Pratylenchus spp. (43%) and (China) Helicotylenchus spp. (45%).
• R. similis has been found on flower plants and on citrus but a larger scale survey to determine the occurrence of this nematode is necessary.

depend on the use thereof. It is importantto remember that being communicative ispart of being a member of the workinggroup.
The following suggestions for the use ofthe working group mailing list were made:• Make sure your name is on the correct
mailing list.• Current members and new members
should introduce themselves with a shortdescription (1/2 page) summarising theiractivities. Members will also be requestedto give a regular update of their activities.
• Names of researchers that will be able toactively contribute can be nominated andadded to the mailing list.
• Abstracts of publications can be sent tothe mailing list (by preference after thepublications have been accepted for publi-cation). This will give everyone the oppor-tunity to have access to the publication.
• MSc and PhD documents can also bemade available in this way.
IV PROMUSA INFOMUSA — Vol 10, N° 2
Screening for nematode resistance at IITA, Namulonge, Uganda (Photo: J. V. Escalant, INIBAP).
Resistance screening
Country/RegionEmilio Fernandez • Cultivars Giant Cavendish, Grande Naine, CEMSA 3/4, Burro CEMSA and Zanzibar were found to be susceptible to R. similis while SH-3436 (Cuba) and FHIA-18 were resistant. Luis Pocasangre • FHIA-01 is resistant to R. similis in the field. This is in agreement with findings in South Africa where low numbers of R similis were found (Latin America) in the roots of FHIA-01. FHIA-01 was, however, attacked by Helicotylenchus and Meloidogyne spp. This shows the importance of evaluating
a cultivar against more than one nematode species.• Plantain has become very important, even for export. Horn Plantain is planted in general but is susceptible to R. similis, FHIA-20 is tolerant:
good yields are obtained, even in the presence of nematodes.• In the Dominican Republic, 3 million plants of FHIA-20 will be planted to replace Horn Plantain.• In Bolivia, funding has been made available for the cultivation of alternative crops to coca. Banana and plantain cultivars (FHIA-01, FHIA-17,
FHIA-18, FHIA-23 and FHIA-25) will be planted. In fact, FHIA-01 and FHIA-18 are already being cultivated with success.Patrick Quénéhervé • The resistance of Musa genotypes to Meloidogyne spp. is being examined.(Martinique/ • Musa diploids and improved hybrids are being screened for resistance to R. similis, P. coffeae and Meloidogyne spp. Guadeloupe) • A similar programme has been initiated for plantain in Guadeloupe in collaboration with CARBAP.
• A study with various Musa accessions and cultivars for in vivo root infection and reproduction of Meloidogyne arenaria and M. incognitahas been started. This study is also done with Pratylenchus spp.
Anne-Marie Auwerkerken • Early screening assistance is provided to the Musa breeding programme of IITA in Nigeria for the evaluation of parent and hybrid lines. (IITA Nigeria) A simple cost-effective screening method based on single root inoculation is being used. based on single root inoculation is being used. Carine Dochez • Early screening assistance is provided to the Musa breeding programme of IITA in Uganda for the evaluation of landraces, wild banana, (IITA Uganda) hybrids and improved hybrids. A simple cost-effective screening method based on single root inoculation is being used. Josephine Namaganda • Involved in routine screening of cultivars for the national programme and IITA.(Uganda)Sveta Gaidashova • The old Musa genebank in Rwanda should be rejuvenated. (Rwanda)Eldad Karamura • A new genetic engineering programme has been initiated coordinated by INIBAP. NARO, IITA, CIRAD, KULeuven, JIC and Leeds University (Uganda) are also involved. The programme focuses on the introduction of foreign genes for resistance to weevils, nematodes and black Sigatoka. Kadija Rajab • Zanzibar has 26 local Musa varieties that have not yet been screened for resistance to nematodes. (Zanzibar)Nuygen Thi Tuyet • Greenhouse screening of Vietnam Musa germplasm for resistance/tolerance to P. coffeae and Meloidogyne spp. has shown differences (Vietnam) in resistance to P. coffeae but not to Meloidogyne spp.
• It was found that SH-3477 as well as Yangambi Km5 show resistance to P. coffeae.Mieke Daneel • FHIA-01 was found to be more tolerant to R. similis. Chinese Cavendish was more susceptible to R. similis, compared to Williams and (South Africa) Grand Nain. Grand Nain was more susceptible to Meloidogyne spp. compared with Williams and Chinese Cavendish.
• Pisang Awak (commonly grown in rural areas) is more tolerant to nematode damage.• Aeroponics (with roots suspended in the air) will be tested as a possible method for nematode resistance screening.
Liao Jinling • South China Agricultural University has a collection of about 70 Musa genotypes. This collection has not yet been evaluated for nematode (China) resistance sources.Dirk de Waele • An overview is presented of all the Musa genotypes screened under greenhouse and in vitro conditions at KULeuven.(Belgium)

• Students, training and collaborators canbe requested through the list.
• Available jobs can be advertised. • Regional activities or meetings can be
announced through the list.
Web siteItems for the web site should first be send tothe convenor who will ensure that the contri-butions are placed on the web site.
Suggestions for the web site:• Short introduction of each researcher of
the working group. This could be linked tothe researcher’s own web site or that ofher/his institution.
• References, abstracts of publications orlinks to publications and electronic publi-cations should be listed.
• A core collection of papers could be esta-blished in this way.
• A list of activities and meetings can beplaced on the web site.During the NWG meeting in Bangkok,
Thailand it was decided to develop threedatabases that will eventually be linked tothe web site: a compilation of the databaseon nematode communities and biodiversitywill be facilitated by J.-L. Sarah; damageand yield loss potential database by R.Fogain with the assistance of M. Daneeland D. Coyne, while the database onsources and mechanisms of resistance willbe facilitated by D. De Waele. A deadlinefor the completion of the databases hasbeen set for the 2002 Nematology meetingto be held in Tenerife.
A suggestion was made that a genebankor nematode culture reference bank shouldbe compiled similar to the ITC genebank forMusa. Accession numbers will be given tonematode populations maintained.
Future work plan for the NWG
Activities in the three main areasidentified
It was suggested that future activitiesshould focus on the three main areas thathave been identified.
• More emphasis should be placed on stu-dies on the influence of nematodes onyield loss.
• Members are encouraged to investigatethe biodiversity of nematode populationson banana in their own countries orregions.
• Various groups are currently carrying outscreening for resistance. It was sugges-ted that the results should be combined.
PROMUSA regional activitiesThe need for basic training for nematodeidentification was expressed by the NARSin preparation for IMTP III. Mozambiqueneeds training on all basic nematologyaspects for two persons. Inge van den Bergcan provide practical nematology training inSouth Africa. Mieke Daneel can be contac-ted in this regard.
IITA is in the early stages of setting uptraining facilities in Nigeria.
Angola has showed interest in bananacultivation.
In Costa Rica screening of material willbe carried out by CORBANA.
A new PROMUSA regional initiative hasbeen launched in the Philippines for thescreening of Musa germplasm. A trainingcomponent has been included for nematolo-gists from the region for participation inIMTP III.
Members of the NWG should bringresearchers in their region together viasmall groups.
Links with the Genetic improvement working group(GIWG)
Several aspects were identified duringthe previous NWG meeting in Bangkok thatwere of importance to GIWG. Activities thatcould contribute towards genetic improve-ment were identified.
a) Sources of resistanceA comment was made that there were notenough sources of resistant material andthe following was suggested:
• Southeast Asia as center of origin of Musashould be examined for new sources ofresistance, by preference in combinationwith the Fusarium working group.
• The area where Pisang Jari Buaya origi-nated from should be sourced for relatedresistant banana cultivars.
b) Molecular markersToo early as a segregating population isneeded to identify molecular markers. Sucha population is not yet available.
c) Mechanisms of resistanceThe mechanism of resistance might dependon the type of banana. Little is known howe-ver about the mechanism of resistance andit might be important to look at.
d) Genetic basis of inheritabilityA segregating population is also needed.
A PROMUSA project should perhaps beinitiated. Possible partners identified areIITA-Uganda, CIRAD- Martinique, etc.
e) Nematicidal genesThe collaborative biotechnology project ini-tiated in Uganda will cover these aspects.
f) ScreeningIn vitro screening is being carried out by KULeuven.
Several members carry out greenhouseand field screening trials (see above).
The PROMUSA regional initiative in thePhilippines for the collection and screening ofgermplasm could be used as a start-off for a complementary project, for example inAfrica for the screening of the African high-land banana and Zanzibar germplasm forresistance.
All screening results are currently frag-mented and need to be collected and com-bined to provide accessible source for thebreeders, i.e. database.
g) Strategy for breedersScreening of diploids for resistance tonematodes will be carried out.
More input with regards to the needs of thebreeders is requested, i.e. type of resistance.
Breeders need to release plants forscreening purposes.
Not all diploid plants are available from ITC.
INFOMUSA — Vol 10, N° 2 PROMUSA V
Damage and yield loss potential of nematodes
Country/RegionLuis Pocasangre • In Costa Rica, nematode damage causes a loss of on average 306 bunches/ha. At US$ 1.8 per bunch the loss is approximately of US$ 551/ha. (Costa Rica) • Due to the growing importance of plantain, a new project has been launched in Panama to determine the loss of production caused by
nematodes, nematode diversity and biocontrol.Mieke Daneel • R. similis has a severe impact on banana production.(South Africa) • Meloidogyne spp. have little effect on banana production but banana is important as a host plant from which these nematodes can infect other
crops in rural areas. This group of nematodes is also linked to False Panama disease.• Study on Helicotylenchus spp. is needed to determine the pathogenicity of this group of nematodes to bananas. Meloidogyne spp. are a problem
because vegetables are highly susceptible to these nematodes and bananas are a good host.

IMTP IIIScreening for nematode resistance will bepart of the International Musa TestingProgramme (IMTP) for the first time. Thisactivity can involve all members.
a) Musa genotypes to be screened (see list)The plants for the programme have beenindexed and are available from ITC.
Several genotypes were included fromdifferent groups.
Additional plants more relevant for nema-tode resistance screening were added tothe official IMTP III list by D. De Waele.These plants were selected on the basis ofresearch data.
It is not necessary to include all the geno-types on the list.
b) InvolvementThe IMTP trials are not funded and partici-pants will have to contribute to the trialthrough their own funding.
The level of involvement will depend onthe member and could involve the standardfield screening or an in-depth study.
c) Parameters to be recorded for fieldscreening (minimum package) • A site/sites should be selected that is/are
ecologically relevant to the region.• The nematode population of the site
should be representative of the popula-tion on banana in the region. More thanone site could be used to evaluate diffe-rent nematode populations.
• A survey of the site is needed, as it isnecessary to determine the nematodepopulation that you are working with. Asample of the population could be sent toJ.-L. Sarah at Montpellier for a comparati-ve analysis.
• An history of the site should be compiled. • Nematodes in the roots should be counted
at flowering and at harvest. If seasoninfluences the nematode populations in thesoil, additional root sampling is suggestedduring the season depending on the area.
• Root necrosis needs to be recorded and amount of living and dead roots in a20 x 20 x 20 cm box.
• All inputs/treatments should be documented.• Flowering and harvest time should be
recorded as well as bunch weight. Otheryield data can be collected and can be found in the INIBAP TechnicalGuidelines written by Speijer and De Waele (1997).
d) Planting of the trial8-15 plants should be planted as singleplant treatments in a complete randomizeddesign. Plants should be treated againstblack Sigatoka, if necessary.
e) Nematode isolation/sampling techniqueThe use of a standard extraction techniquewas suggested. It was however indicatedthat the type of extraction would not influen-ce end results as long as the standardcontrols are included in the IMTP trial.
The maceration and sieving technique issuggested for the training of nematologistsand equipment can be donated/supplied.
f) Research possibilities for the in-depthstudies• Host-pathogen relationships.• Pot trials with single species infestations.• Yield loss assessment by replicating the
trial with no nematodes. Nematicides canbe used to ‘create’ the replicated trialwithout nematodes.
• Study of differences in resistance bet-ween tissue culture- and sucker-derivedplant material.
• Study of influence of environment onplant resistance.
• Relation between root damage (smallplant) and resistance.
• Cause of variation in resistance betweensites.
Feeling about participation in the PROMUSA working group• Members got to know each other on a
personal level, which promoted a lessconstrained environment for interaction.
• Several members felt that they were nowless isolated and found that people werewilling to help.
• Interaction with the other members pro-moted the sharing of information andtechniques.
• Opportunities for collaboration and assis-tance were created.
• Backing of PROMUSA could promotefund raising.
• Members felt more motivated as theynow contribute to a much wider cause.
• Progress was noticed over the last tenyears.
• By working together a global researchsystem could emerge.
• Research can be done more efficientlyand duplication reduced.
• The advantages of PROMUSA are beingrecognized and now duplicated in othercrops.
Abstracts of papers and posters presented duringthe PROMUSA Nematologyworking group meeting
Screening Musa germplasm forresistance to Radopholus similis
C. Dochez1, D. Makumbi1, A. Tenkouano2
and D. De Waele3
1International Institute of Tropical Agriculture (IITA) – Easternand Southern Africa Regional Centre (ESARC), P O Box 7878, Kampala, Uganda2International Institute of Tropical Agriculture (IITA) – HighRainfall Station Onne, Nigeria, c/o Lambourn & Co., CarolynHouse, 26 Dingwall Road, Croydon CR9 3EE, England3Katholieke Universiteit Leuven, Laboratory of Tropical CropImprovement, Kasteelpark Arenberg 13, 3001 Leuven,Belgium
Worldwide, plant-parasitic nematodes areknown as one of the most important bioticconstraints to sustainable Musa production.In the lowlands of central Uganda, the mostdamaging nematode species is Radopholussimilis, while at higher elevationsPratylenchus goodeyi is the most prevalentspecies. Use of nematicides to controlnematodes has adverse environmentaleffects and is too costly for subsistence far-mers. A promising alternative is the use ofnematode resistant varieties. Such varietiescan be obtained either by evaluating thehost response of existing genotypes or byclassical breeding. However, for screeningall available germplasm a rapid and reliablescreening method is needed. An earlyscreening method was developed at IITA,which is based on the inoculation of indivi-dual roots and needs less nematode inocu-lum and less plant material per genotypecompared with classic pot trials. By usingsingle roots for inoculation, the host response
VI PROMUSA INFOMUSA — Vol 10, N° 2
IMTP III: list of genotypes for nematologicalevaluation
On official IMTP III listPisang Jari Buaya AA ITC0312Calcutta 4 AA ITC0249Pisang Mas AA-Sucrier ITC0653Pisang Ceylan AAB-Mysore ITC1441Cachaco ABB ITC0643Saba ABB ITC1138FHIA-03 FHIA hybrid ITC0506FHIA-18 FHIA hybrid ITC1319FHIA-23 FHIA hybrid ITC1265FHIA-25 FHIA hybrid ITC1418TMB2x9128-3 IITA hybrid ITC1437TM3x15108-6 (Pita 16) IITA hybrid ITC1417Not on official IMTP III listGrande Naine AAA ITC1256Yangambi Km5 AAA ITC1123Paka AA ITC0320Selangor (AA) acuminata ITC1060Kunnan AB (Ney Poovan) ITC1034M. balbisiana BB (“Honduras”) ITC0247M. balbisiana BB (“Tani”) ITC1120Foconah AAB (Pome-Prata) ITC0649Pisang Lemak Manis ABB ITC1183FHIA-01 FHIA-hybrid ITC0504

to nematode attack is not affected by diffe-rences in root growth rates between Musagenotypes. Currently germplasm is beingscreened for resistance to R. similis at IITA-ESARC in Uganda. The germplasm includeEast African highland bananas (EAHB),hybrids derived from those and hybrids deri-ved from diallel crosses, including a segre-gating population. Each genotype is testedin comparison with a resistant cultivar(Yangambi Km5) and a susceptible one(Valery). Nineteen hybrids, of which fiveEAHB, have been identified as resistant toR. similis with the early screening method.Pot trials are in progress for confirmation ofthis resistance.
In vitro and in vivo screening for resistance to Radopholus similisin Musa
N.T. TuyetVietnam Agricultural Science Institute, Van Dien, Thanh Tri,Hanoi, Vietnam
Two greenhouse experiments were carriedout with five Musa cultivars belonging toMysore group (AAB) and five Musa cultivarsfrom the Philippines (AA) to determinesources of resistance and tolerance toRadopholus similis. The susceptible‘Grande Naine’ was chosen as the referen-ce cultivar. Each in vitro micropropagatedplantlet was transplanted to a 12 cm-diame-ter plastic pot containing about 1000 cm3
autoclaved substrate and maintained at anambient temperature of 25-30°C with a 12-hour photoperiod. Eight weeks (forMysore cultivars) and ten weeks (for culti-vars from the Philippines) after inoculationwith 1000 vermiforms of R. similis per plant,the plants were harvested and the numberof nematodes per 10 g of root, root nemato-de populations, fresh root weight and per-centage of root necrosis were assessed.
Variability was seen between cultivarsbelonging to Mysore group in terms of rootnematode populations. ‘Gorolo’ was consi-dered the least susceptible to R. similis withthe total number of nematodes recoveredwas 32 562 and the percentage of rootnecrosis was 22.81 %. ‘Lady Finger(Nelson)’ was shown to be highly suscep-tible to R. similis damage with final popula-tion was 99 009 nematodes and 33.13 % ofroot necrosis. This susceptibility was notsignificantly different from ‘Grande Naine’.
The results of five cultivars from thePhilippines demonstrated that most of themwere as susceptible to R. similis as ‘GrandeNaine’.
An in vitro experiment was conducted toinvestigate if resistance to R. similis isexpressed in plantlets growing in rootingmedium under in vitro conditions. The resis-tant and susceptible sources such as‘Yangambi Km5’, ‘SH-3142’, ‘Pisang JariBuaya’, ‘Gros Michel’, ‘Grande Naine’ and‘Cachaco’ were tested in this study. Themicropropagated plantlets were rooted inglass jars, which were maintained in agrowth chamber at 26°C with a 16-hourphotoperiod (one replicate = 1 jar with 2plantlets). After four weeks, when plantletshad formed enough roots, each one wasinoculated with 25 females. Eight weeksafter inoculation the total nematode popula-tions in the roots and in the medium weredetermined. The preliminary results under invitro conditions showed the resistance to R.similis of ‘Pisang Jari Buaya’ and ‘SH-3142’and susceptibility to R. similis of ‘GrandeNaine’. However, ‘Yangambi Km5’ wasconfirmed resistant in vivo or in the fieldtests, but showed less resistance in the invitro test. This study shows that the in vitroprocedure has several advantages compa-red to greenhouse or field screening.
Current status of the nematological situation on plantain and banana in Latin America
L. Pocasangre1, R.A. Sikora2 and M. Araya3
1International Network for the Improvement of Banana andPlantain (INIBAP)/Centre Agronomico Tropical deInvestigación Y Enseñanza (CATIE), Unidad de Biotecnologia7170, Turrialba, Costa Rica2 Institut für Pflanzenkrankheiten, University of Bonn,Nussallee 9, 53115 Bonn, Germany3 Corporación Bananera Nacional (CORBANA S.A.), Apdo390, 7210 Guápiles, Costa Rica
In Latin America nematodes are responsiblefor yield reduction of up to 50% in commer-cial banana plantations. Typical nematodecontrol consists of two or three applicationof nematicides per year. The majority ofnematological research in Latin Americahas been conducted in large commercialbanana plantations and information is spar-se about the nematodes species attackingplantain. In this survey, nematological stu-dies were conducted in commercial bananaand plantain plantations in several countriesin Latin America viz. Bolivia, Costa Rica,Guatemala, Honduras and DominicanRepublic. The results of this survey showthat the nematode species of major concernin commercial banana plantation isRadopholus similis. Densities of R. similisup to 45 000/100 g of fresh roots werefound in Guatemala and Costa Rica. In
plantain on the contrary, several species ofPratylenchus were the most frequentlyfound nematodes in the root systems.Densities of Pratylenchus spp. up to 5400nematodes/100 g of fresh roots were foundin the Dominican Republic, Honduras andCosta Rica. R. similis was also found inplantain but in lower densities than in bana-na roots. More nematological studies areneeded in plantain in order to design anadequate strategy of control for the smallplantations in Latin America.
Correlation between nematodenumbers and damage to banana(Musa AAA) roots under commercial conditions
T. Moens1, M. Araya2 and D. De Waele3
1 Agreement VVOB-INIBAP-CORBANA S.A., Apdo 390, 7210Guápiles, Costa Rica2 CORBANA S.A., Apdo 390, 7210 Guápiles, Costa Rica3 Laboratory of Tropical Crop Improvement, KULeuven, Kasteelpark Arenberg 13, 3001 Leuven, Belgium
The correlation between Radopholus similisand total nematode numbers, and grade ofroot necrosis and root damage in commer-cial plantations with mixed Musa AAA cvsGrand Naine and Valery was investigated ina series of four experiments. Three differentroot damage indices, the Australian index,the Bridge and Gowen index and a Localindex, were compared assessing 100 ran-domly selected root samples. Correlationsfor the three indices were highly significantand varied between 0.38 and 0.40. In a sub-sequent experiment with the Local index,the correlation between R. similis and totalnematodes, and root necrosis and damagewas not improved when measuring one orboth root halves. Using the local index, theabove mentioned correlation was evaluatedin randomly selected root samples frommother plants and their respective followersuckers in a third experiment. Mean rootnecrosis and damage grade in mother rootswere double those of follower sucker roots.Correlation coefficients between R. similisand total nematodes, and root necrosis anddamage in follower suckers ranged between0.41 and 0.48, and were always significant.In contrast, these correlations using motherroots were never significant. In the lastexperiment, roots from mother plants andtheir respective follower suckers werearranged in 10 different damage classes. R. similis and total nematode numbers werecorrelated with root necrosis and damagefor 0-5, 6-10 and 0-10 damage groups.Correlations were highest for root necrosisand R. similis in 0-5 interval, ranging bet-
INFOMUSA — Vol 10, N° 2 PROMUSA VII

ween 0.62 (P # 0.0002) for mother rootsand 0.75 (P # 0.0001) for sucker roots.
Management of nematodes of banana systems in Uganda
J.M. Namaganda1, S. Gowen2, P. Namanya1 and I. Kashaija3
1Kawanda Agricultural Research Institute, P.O Box 7065,Kampala, Uganda2University of Reading, Department of Agriculture, EarleyGate, P.O Box 236, Reading, Berkshire, RG6 2AT, UnitedKingdom3Kachwekano Agricultural Research and Development Centre,Kabale, Uganda
The host status of weed and crop speciescommonly associated with banana fields tothe banana nematodes Radopholus similis,Helicotylenchus multicinctus andPratylenchus goodeyi was studied inscreenhouse experiments. The weed spe-cies Ageratum conyzoides, Bidens pilosa,Commelina benghalensis, Crassocephalumcrepidioides, Cyperus esculentus, Digitariascalarum, Eleusine indica, Galinsoga parvi-flora, Senecio disfolius and Tagetes minutawere non-hosts of the three banana nema-todes. Amaranthus sp., Digitaria velutinaand Solanum nigrum were poor hosts of H.multicinctus while D. velutina was also apoor host of P. goodeyi. Cassava and sweetpotato were non-hosts of the three nema-todes, groundnut a non-host of P. goodeyiand the phaseolus bean and maize poorhosts of P. goodeyi.
In on-station and on-farm field experi-ments aimed at devising croppingsequences that would minimize nematodedamage and enable acceptable levels ofbanana production to be maintained, bana-na nematode population densities declinedto negligible levels after 13 months with abreak crop of cassava or sweet potato plan-ted immediately after a nematode infestedbanana crop.
Enhanced biodegradation potential of banana soils
T. MoensAgreement VVOB-INIBAP-CORBANA S.A., Apdo 390, 7210Guápiles, Costa Rica
The phenomenon that regular application ofthe same nematicide to the soil decreasesthe efficiency of the used product is widelyknown. This is probably due to a selectionfor microorganisms able to decompose theactive compounds. In this series of experi-ments in a commercial banana plantation,enhanced biodegradation was induced byapplying the same nematicide every fourmonths for at least five times. Tested
nematicides were fenamiphos (Nemacur®), ethoprophos (Mocap®) and carbofuran(Furadan®). Grade of biodegradation wasmeasured through a biological test, develo-ped by Pattison et al. After sampling the soilaround various banana plants, 3 subsamplesof 5 kg were taken and received a differenttreatment. One batch served as untreatedcontrol, the second was sterilized andapplied with nematicide at 10 µg a.i./g of soiland the third was only applied with nematici-de at 10 µg/g of soil. Starting from the day oftreatment, maize was planted in little reci-pients filled with around 60 g of soil of thesethree batches, and inoculated with 500Radopholus similis. Plantlets were harvestedone week later and the number of nema-todes penetrated in the maize root over thisperiod was counted. This procedure wasrepeated every 2 weeks up to 8 weeks.
The existence of enhanced biodegrada-tion was clearly observed for the threenematicides used. Already two weeks afterinitiation of the evaluation, nematode num-bers in the nematicide-treated batches werethe same as in the untreated control foreach of the three nematicides. This meansthat two weeks after application the activeingredient completely lost its effect. Theseresults show the importance of nematiciderotation. Further investigation on other fre-quently used nematicides and under diffe-rent soil and climate conditions is needed.
New Musa hybrids with resistance to Radopholus similis(poster)
A. Auwerkerken1, I. Rotifa1, D. De Waele2
and A. Tenkouano1
1International Institute of Tropical Agriculture (IITA), c/oLambourn & Co., Carolyn House, 26 Dingwall Road, CroydonCR9 3EE, England2Laboratory of Tropical Crop Improvement, KU Leuven,Kasteelpark, Arenberg 13, 3001 Leuven, Belgium
The burrowing nematode Radopholus similisis one of the major nematode species attac-king banana and plantain. Incorporation ofresistance through plant breeding is anadvantageous form of nematode control. Thescreening experiment was established in ashade house at the IITA High Rainfall Stationat Onne in Southern Nigeria (7°E, 5°N, 10 masl). Three suckers from each hybridand two reference cultivars, Yangambi Km5(resistant) and Valery (susceptible) wereplanted together in sawdust in woodenboxes. Four weeks after planting, a smallplastic cup was placed on three of the emer-ged roots at about 5 cm from the corm. A
suspension of 50 females of R. similis waspoured on each individual root. Eight weekslater, the number of nematodes was countedand a Dunnett’s T-test was done to comparethe square root transformed mean reproduc-tion ratio. Fourteen hybrids showed a repro-duction rate not significantly different fromYangambi Km5. These hybrids included fourhybrids from 2x-4x crosses, seven hybridsthat have the resistant hybrid TMP2x9128-3in their pedigree and the tetraploid hybridsA10-SPS-548-9 and 7152-2. The latter isknown as the promising hybrid PITA 14. Theselected hybrids supported a significantlylower nematode reproduction rate and aretherefore interesting for further evaluation.
Preliminary results of treatmentsfor replanting in bananas (poster)
M. Daneel, D. Mdluli, K. De Jager, J. Husselman and C. NeethlingARC-Institute for Tropical and Subtropical Crops, Private BagX11208, Nelspruit 1200, South Africa
Replanting of bananas has always been aproblem, especially where Radopholussimilis is present. Nematode infestation oftissue-cultured plants is a huge problembecause of the low tolerance levels of thesmall plantlets. Several replant treatmentsfor both commercial and rural farmers arebeing tested. Treatments used are methylbromide (MB), metam-sodium (Herbifume),plastic cover, sugarcane, kraal manure,deep hole, PL Plus, aldicarb, water pasteu-rization and fallow. Treatments began inSeptember 1999 with planting of sugarcanefollowed by plastic cover in November andthe application of kraal manure inDecember. Herbifume was applied at theend February and MB in April. PL Plus andTemik were applied at planting, which wasdone in June 2000. Preliminary results sho-wed huge differences in initial growth ratebetween the different treatments, with MBthe tallest plants followed by plastic cover,Herbifume and kraal manure. The lowestnematode numbers in roots were found inMB followed by Temik, plastic cover, kraalmanure and Herbifume.
Pest and disease complexes of Musa in rural farming systems(poster)
M. Daneel1, N. Dillen2, J. Husselman1, K. DeJager1 and D. De Waele2
1ARC-Institute for Tropical and Subtropical Crops, Private BagX11208, Nelspruit 1200, South Africa2Laboratory of Tropical Crop Improvement, KU Leuven,Kasteelpark Arenberg 13, 3001 Leuven, Belgium
VIII PROMUSA INFOMUSA — Vol 10, N° 2

A survey was carried out in the rural areasof South Africa to determine production, cul-tivars and the pest and disease complexeson Musa spp. Bananas are found from thenorthern part of the Northern Province(Venda), downwards in the eastern part ofthe Northern Province and Mpumalanga, inSwaziland as well as the coastal areas ofKwaZulu-Natal to as far as Eastern Cape(old Transkei). However, bananas are neverfound in more than two to five mats per hou-sehold. Nematodes were present in allsamples, with Meloidogyne spp. being themost abundant. Other species found werespiral nematodes and in a few casesRadopholus similis and Pratylenchus cof-feae. The most commonly planted cultivar isPisang Awak (ABB), followed by Cavendish(AAA), but in much lower numbers.Plantains were found at five sites. The lesssweet tasting Pisang Awak seems to betolerant to nematodes and leaf diseasesand is also more drought resistant. It there-fore appears to be more adapted to the cli-matic and environmental conditions as wellas the specific cultural practices followed bycommunities in rural areas.
Is a growth stimulant an effectiveagent to reduce nematode damage on banana plants? (poster)
M. Daneel, A. De la Bretesche, J. Husselman, K. De Jager and C. NeethlingARC-Institute for Tropical and Subtropical Crops, Private BagX11208, Nelspruit 1200, South Africa
Several products have been tested forcontrol of nematodes on banana plantlets
in the glasshouse. Plants were infestedwith a mixed population of Radopholussimilis, Meloidogyne spp. andHelicotylenchus spp. One month later,applications were made at two intervals, 14days apart and plants were evaluated twomonths later. Treatments were Agent Xcombined with PL Plus, PL Plus, Agent Xcombined with PL251, Agent X, NemacurGR and an untreated control. Results sho-wed that plants treated with Nemacur GRhad the lowest numbers of nematodes inthe roots followed by PL Plus, Agent X &PL Plus and Agent X. Plants treated withAgent X alone or in combination had thehighest plant mass. However, a root necro-sis index confirmed that the combination ofAgent X and PL Plus is a better option forbananas as it reduced nematode numbersand increased growth potential, making theplant more tolerant against nematodes.
Effect of different cleaning techniques for planting materialon banana plant growth andnematode damage (poster)
S.V. Gaidashova, C.M. Gatarayiha, A. Nsabimana and B. UwimpuhweISAR (Institut des Sciences Agronomiques du Rwanda),B.P.138, Butaré, Rwanda
Nematodes are recognized as a majorbanana pest usually transmitted in newfields through infested suckers. Differenttechniques for planting material cleaningwere suggested by scientists but, actually,they are applied rarely and by a limitednumber of small-scale farmers becausethey are costly and labour/resource consu-
ming. The objective of this study was tocompare different cleaning techniques forbanana planting material in terms of theirinfluence on plant growth parameters andnematode damage. On-station experimentwas established in Rubona with highlandcultivar Mbwaziruma and comprised fourtreatments: 1) paring of suckers beforeplanting; 2) paring and sun drying for threedays at 25-30°C; 3) hot water treatment for20 min at 55°C and 4) normal farmer’s plan-ting method. Attendance of suckers wasobserved twice: at two and four weeks afterplanting. Plant growth parameters andnematode damage were observed 5, 10and 14 months after planting (MAP). Sun-dried suckers showed significantly (p<0.05)lower level of attendance two weeks afterplanting. However, no difference was obser-ved between treatments two weeks later. Asa consequence, the same treatment (sunexposure) showed significantly lower growthparameters (p<0.05) 5 MAP. Later no diffe-rence was observed. Nematode damagewas significantly (p<0.05) higher withcontrol than any other treatments, 10 and14 MAP. Preliminary results indicate thateffects of all tested cleaning techniqueswere comparable and show significantreduction in nematode damage. Therefore,the cheapest and less labour/resourceconsuming technique (1st and 2nd treat-ments) could be recommended for small-scale farmers.
INFOMUSA — Vol 10, N° 2 PROMUSA IX
Since the second global PROMUSA mee-ting, held in Douala, Cameroon, inNovember 1998, INIBAP, in its role asPROMUSA secretariat, has begun to deve-lop different mechanisms whose aims are toestablish and ensure an information flowbetween members of the various PROMU-SA working groups (Fusarium, GeneticImprovement, Nematology, Sigatoka,Virology). The nomination of Core Groupswith respective Chairpersons, within theindividual working groups has been comple-ted, and once again the aim is to facilitate
the exchange of information and work bothwithin and between the working groups.
Steering Committee of PROMUSAIn consultation with the respective regionaland sub-regional organizations, it has beenagreed that the banana regional networks(MUSALAC, MUSACO, BAPNET and BAR-NESA) could nominate representatives for NARS in the steering committee ofPROMUSA.
Convenors of the Workinggroups of PROMUSADuring the 3rd Global Meeting ofPROMUSA, the participants proceeded withthe election of the new convenors of the dif-ferent working groups of PROMUSA.• Genetic improvement:
Dr Jaroslav Dolezel, IEB, and Czech Rep.Dr Sathiamoorthy, NRCB, INDIA
• Sigatoka: Dr. Jean Carlier, CIRAD, France
• Nematodes: Prof. Dirk de Waele,KULeuven, Belgium
• Virology: Dr Jackie Hughes, IITA, Nigeria• Fusarium: Dr Suzy Bentley,
QDPI, Australia
Since the 2nd PROMUSA global meeting in 1998

Progress within the PROMUSA global programme
The spirit of collaboration and partnershipshas developed considerably both duringand since the last global meeting. Specificareas of collaboration include:
Genetic improvement working group
Good interactions have taken place bet-ween the Sigatoka and Genetic improve-ment working groups in the framework ofthe International Musa Testing Programme(IMTP). The IMTP phase II is finished andthe results are published. In order to facilitateaccess and visibility of the results, a databa-se will soon be available on the PROMUSAweb site and as a CD-ROM.
Phase III of IMTP has been initiated andmore than 30 institutions have alreadyagreed to participate. A catalogue of 27candidates (hybrids, landraces and soma-clonal variants) has been developed, wherepeople can choose from those that they areinterested in.
The contacts between all the PROMUSAworking groups has enabled the Geneticimprovement working group to establishlists of priorities within each researchtheme: Sigatoka, virus, nematology andFusarium.
Close collaboration has been initiatedbetween CIRAD-FLHOR, CRBP (now CARBAP) and IEB with the development ofa new technology to purify chromosomefractions. This would be very useful forMusa genome analysis and mapping.
Discussions are being held between diffe-rent members of the group in order to getmore funds so as to develop one or moreproposals.
DNA markers to improve plantains andstarchy bananas have been submitted to theEuropean Union in the framework of thePCRDT/INCO-DEV. The project was develo-ped between different partners ofPROMUSA: University of Reading in UK,KULeuven in Belgium, Frankfurt University inGermany, IEB in Czech republic, CRBP inCameroon, NARO in Uganda and INIBAP.
A project has been developed and hasbeen accepted to be funded by the FrenchGovernment on “Mapping of the Musa acu-minata translocation break points throughmolecular cytogenetics”. This project, sub-mitted in the framework of the “AgropolisAdvanced Research Platform” involves anInternational Agricultural Research Centre
(IPGRI/INIBAP), an Advanced ResearchInstitute from the Montpellier platform(CIRAD) and a Research Institute from theBrazilian National Agricultural ResearchSystem (EMBRAPA). A researcher fromEMBRAPA, who is on secondment toINIBAP, is carrying out the work in the labo-ratories of CIRAD in Montpellier, France.
A new project has been accepted to befunded by the French Government on“Structural and functional genomics ofbanana streak virus sequence integrationinto the Musa genome”. This project, sub-mitted in the framework of the “AgropolisAdvanced Research Platform” involves anInternational Agricultural Research Centre(IPGRI/INIBAP), an Advanced ResearchInstitute from the Montpellier platform(CIRAD) and two Research Institutes: CINVESTAV, Mexico and IIHR, India.Researchers from both institutions will carryout the work in the laboratories of CIRAD inMontpellier, France.
The project novel approaches to theimprovement of banana production inEastern Africa implemented by INIBAP incollaboration with different local, regionaland international institutions (NARO,Makerere University and IITA, Uganda,KULeuven University, Belgium and CIRAD,France) aims to develop biotechnologycapacities to facilitate the improvement ofEast African Highland Bananas.
Genomics on bananaFollowing a suggestion made during the firstsymposium on “Molecular and CellularBiology of Banana” carried out at CornellUniversity in 1999, a PROMUSA workshopon Musa genomics was organized on 6-8April 2000 in Montpellier, France. Toward theend of the meeting a remarkable degree ofconsensus for the Banana Genomics initiati-ve was reached. All parties agreed to form aBanana Genomic consortium. PROMUSAoffers a good framework in which to assumeworldwide leadership in the new initiative onbanana genomics public research, throughthe development and implementation of avisionary strategy and whose activities willbe developed within the Banana genomicsconsortium. The consortium will bring toge-ther and enhance combined expertise (fromboth the public and private sector).
The 2nd international symposium on“Molecular and Cellular Biology of Banana”was organized in Byron Bay, Australia fromthe 31st of October to the 3rd of November2000.
The Banana genomics consortium hasbeen extended with several new partners
who met in Washington DC from 17-20 July,2001. During this meeting, all the partnersworked together in the fine-tuning of thestrategy, focusing on the activities to bedeveloped and how to share their develop-ment between the different partners. Themeeting has also been an important oppor-tunity for the partners to develop differentconcept notes to further develop full projectproposals to be submitted to donors.European partners within the consortiumagreed to develop a project proposal to besubmitted before the 15th of October 2001 inthe framework of the 5th PCRD programme of the EU.
Different segregating populations alreadyexist (or soon will exist) with different levelsof accessibility. INIBAP, in collaborationwith CIRAD-FLHOR in Guadeloupe, FWI,will facilitate the development of thesepopulations and their availability to all thePROMUSA participants.
Sigatoka working groupDifferent activities have been initiated to develop a detailed understanding ofpopulation structures of the pathogensMycosphaerella sp.
Priority has been given to Asia: a projectwas developed to be presented in the fra-mework of the 5th PCRDT/INCO-DEV:“Breeding of bananas for Southeast Asianproduction: Identification of resistant wildgermplasm to the leaf spot diseases”.
A collaborative initiative has been set upat a regional level in Asia involving severalcountries in to characterize the “Sigatoka”diseases present in their country and to par-ticipate in the study of the geographical dis-tribution of the different pathogens:Mycosphaerella musicola, M. fijiensis andthe new disease: M. eumusae.
This initiative is part of other different stu-dies already ongoing in Latin America andAfrica in collaboration with CIRAD-AMIS,France.
Efficiency, durability and management ofresistance have also been identified as stra-tegic research to be developed. In thiscontext, studies have been initiated on theanalysis of resistance (CIRAD/CARBAP, IITAand FHIA). Collaboration between breedingprogrammes would allow a better understan-ding on what genes are involved, and whichmechanisms are inherited (PR, HR).
Development of early screening methods:An artificial inoculation method undercontrolled conditions is being developedbetween CIRAD and CARBAP. Leaf frag-ments are maintained through in vitro condi-tions after inoculation with conidia.
X PROMUSA INFOMUSA — Vol 10, N° 2

Nematology working groupInitiative for collaborative work:Screening/evaluation experiments andtrials through a “modular” type global pro-ject, which could contain different projectcomponents (different sources of funding).
The third meeting of the Nematologyworking group (NWG) was held 24–25May 2001 in Hulala Lakeside Lodge,Hazyview, South Africa (see report in thisissue) on the occasion of the 15th
Symposium of the Nematological Societyof Southern Africa (NSSA). During thismeeting participants had the opportunity todefine all complementary research activi-ties with the Genetic improvement workinggroup.
Different results obtained in the frame-work of PROMUSA have been publishedwith a special acknowledgement and reco-gnition of PROMUSA.1) Host plant response of banana (Musa
spp.) cultivars from Southeast Asia tonematodes by R. Stoffelen, Vu Thi ThanhTam, R. L. Swennen and D. De Waele.International Journal of Nematology(1999) Vol. 9(2): 130-136.
2) Host plant response of Fusarium wilt resis-tant Musa genotypes to Radopholus similisand Pratylenchus coffeae by R. Stoffelen,R. Verlinden, J. Pinochet, R. L. Swennenand D. De Waele. International Journal ofPest Management (accepted).
Virology working group
This group focussed on activities identi-fied as priorities within PROMUSA and dis-tributed tasks between the different teams.
Development of reliable virus detectionmethods to prevent dispersal of virusinfected germplasm, to define and limitgeographical distribution and to improvedetection methods reflecting those invirus indexing procedures
QDPI and CIRAD will work together toestablish that Onne strain of BSV is a majorstrain present as episomal infection ingermplasm.
FUSAGx and QDPI will develop IC-PCRfor BSV + Potex.
FUSAGx and JIC will work on PCR pro-cedures to differentiate integrated and epi-somal BSV sequences to detect low or nosymptoms in plants.
QDPI and CIRAD will revise detectionprocedures for Virus Indexing Centers.
BSV: To eliminate particular viruses as aconstraint to production.
FUSAGx will investigate the reduction inthe level of activation of episomal BSV intissue culture.
John Innes Centre, Univ. of Minnesota,FUSAGx, KULeuven and CIRAD will colla-borate in the investigation of differencesbetween Musa varieties in the activation ofepisomal BSV during tissue culture.
CIRAD, Univ. of Minnesota and JohnInnes Centre will work on the understandingof the mechanism of activation.
CIRAD and Univ. of Minnesota will inves-tigate the linkage of integrated sequences(e.g. to B genome) and the genetic mecha-nisms involved in activation and multiplica-tion of integrated BSV in Musa sp.
A project proposal had been developed to be presented in the framework of the 5th PCRDT/INCO-DEV.: “Thematic: BSVvirus, new molecules as inhibitor of retrovi-rus; serological tools; clean up plant fromPotex; Potex diagnostic tools; geneticaspect of the activitation of the inserted viralsequency.”
The project is being modified so that it canbe represented in the framework of the newcall for proposals of the PCRDT/INCO-DEV.
A collaborative project has been develo-ped between IITA, PPRI and QDPI on theBanana Die Back Virus (BDBV)
Fusarium working groupThe third meeting of the Fusarium workinggroup (FWG) was held from 21-22 October,1999 in Kuala Lumpur, Malaysia, on theoccasion of the International Seminar“Fusarium wilt of banana: towards sustai-nable cultivation”.
Full details of recommendations foraction, research opportunities and possiblepartners for collaboration have been establi-shed and published in the PROMUSA sec-tion of INFOMUSA 8(2).
The discussions on Fusarium wilt mana-gement issues involved issues related topathogen diversity, disease managementstrategies, epidemiology and other relevantresearch issues. It was noted that there is aneed for training in the use of DAF andVCG techniques, and that further collectionand analysis of samples is still required insome parts of the world. It was recommen-ded that a centralized database of Fusariumoxysporum f.sp. cubense (Foc) geneticdiversity be developed and that this be lin-ked with information regarding the reactionof landraces/hybrids to Foc (e.g. MGISdatabase).
In relation to disease management strate-gies, the need for education and farmer-awareness was highlighted, especially
regarding the use of clean planting materialand resistant varieties.
The FWG reviewed and updated thereport of the Fusarium wilt working groupprepared after the first global PROMUSAmeeting (Guadeloupe, March 1997). Therevised document is now available via thePROMUSA website.
PublicationsMajor activities and progress within the framework of PROMUSA are published twicea year in a specific section of INFOMUSA.Abstracts, publications, meeting reports,visits, congress information etc.
Seven PROMUSA sections have nowbeen published in INFOMUSA and the enti-re collection is available on the PROMUSAwebsite: http://www.promusa.org/publica-tions/publication.htm
Technical guidelines facilitate methodolo-gies for our partners and are also very help-ful for training purposes: The INIBAP techni-cal guidelines No. 5, “Cryopreservation ofMusa germplasm” published in 2001, havebeen developed between different partici-pants from PROMUSA.
Different proceedings of Banana scientificevents have also been published in the PROMUSA section of INFOMUSA: • International symposium on the molecular
and cellular biology of bananas, 22-25March 1999 in Ithaca, NY, USA.
• FAO/IAEA 3rd Research coordinationmeeting of the collaborative research pro-ject on “Cellular biology and biotechnolo-gy, including mutation techniques forcreation of new useful banana geno-types”. This meeting was organized from4-8 October 1999 in Colombo, Sri Lanka,by the International Atomic EnergyAgency (IAEA).
A PROMUSA web site has been set upand is now available at the followingaddress: http://www.promusa.org/
PROMUSA Listservers One general list called PROMUSA ([email protected]) and five sub-lists have beenset up: • PROMUSAGEN([email protected]) • PROMUSASIG ([email protected])
for Sigatoka• PROMUSANEM([email protected])
for Nematology• PROMUSAVIR ([email protected])
for Virology and • PROMUSAFUS ([email protected])
for Fusarium.
INFOMUSA — Vol 10, N° 2 PROMUSA XI

XII PROMUSA INFOMUSA — Vol 10, N° 2
2nd International workshop on Sigatoka leaf spot diseases of bananasCosta Rica, 20-23 May 2002
Until the 1970s, plantain, as well as other popu-lar banana cultivars grown for local consump-tion, were not seriously affected by majordiseases. However, the rapid expansion of theblack leaf streak/black Sigatoka disease(Mycosphaerella fijiensis) in the 1980s causedenough damage for small producers to encoura-ge INIBAP to organize the 1st International work-shop on Sigatoka leaf spot diseases of bananasin San José, Costa Rica in March 1989.
The organization of the 2nd internationalworkshop on Sigatoka leaf spot diseases ofbananas, including mainly black leaf streak,Sigatoka and Septoria diseases, is planned totake place in San José, Costa Rica from 20 to23 May 2002.
The workshop should facilitate the analysisof the current situation on the banana leaf spotdiseases (Sigatoka, black Sigatoka andSeptoria) all around the world.
In order to maximize the outputs of the mee-ting and to guarantee the development of newstrategies to help in the control of the differentdiseases mentioned above, the participation tothis workshop will be only on invitation, andwill be extended by the organizing committeeafter consulting with the scientific committee.
Meeting of the PROMUSASigatoka working groupGuápiles, Costa Rica,24-25 May 2002
We would like to take advantage of the organi-zation of the 2nd International Workshop onSigatoka leaf spot diseases on banana toorganize back-to-back the meeting of theSigatoka working group of PROMUSA on 24-25 May 2002.
This meeting will take place at the Escuelade Agricultura para el Trópico Húmedo(EARTH), Guápiles, Costa Rica.
The meeting will not only facilitate the analy-sis of the current situation on the different activi-ties developed in the framework of PROMUSA,but will also refine the orientation for the groupand help develop new concept notes in order tofacilitate the writing up of project proposals.
The participation to this meeting will be asusual only on invitation, and will be extendedby the Secretariat (INIBAP) in agreement with
the convenor of the working group. However,as you may already know, INIBAP cannotassume the participation costs for all the parti-cipants and we would appreciate very much if you could use the opportunity of the 2nd International workshop on Sigatoka leafspot diseases on banana to find any grant orsupport to participate in sharing the costs.
We will rely on the convenor of the workinggroup to communicate further details on theworkplan.Contacts:Jean-Vincent Escalant, Secretary of PROMUSA:[email protected] Carlier, convenor of the Sigatoka working group:[email protected] Moffatt, Programme Assistant INIBAP: [email protected]
Meeting of the PROMUSANematology working group7-13 June 2002During the 4th International Congress ofNematology in Tenerife 2002 (June 7-13), themembers of the PROMUSA Nematology wor-king group present at the congress will havethe opportunity to meet under the coordinationof the convenor of the working group Prof. Dirkde Waele.
For further details, please contact: [email protected].
3rd International symposiumon the Molecular and cellularbiology of bananasLeuven, Belgium,9-11 September 2002The first international symposium on theMolecular and cellular biology of bananas, heldin March 1999 at Cornell University in Ithaca,USA, was organized to create a forum whereall those involved in the molecular and cellularbiology of bananas would have an opportunityto meet and exchange ideas about theirresearch activities. The meeting was a resoun-ding success and the concept continued the-reafter under the auspices of PROMUSA.
During the 2nd symposium, held in October2001, in Byron Bay, Australia, participants pre-sented their research activities on differenttopics including Genomics, Gene expressionin transgenic plants, Plant pathology anddisease resistance, Biodiversity and evolution,Biochemistry and fruit ripening, Intellectualproperty and genetically modified organisms.
INIBAP and KULeuven are pleased toannounce the organization of the 3rd
International symposium on the Molecular and cellular biology of bananas to be held in Leuven, Belgium from 9 to 11 September2002.Contacts:Prof. Rony Swennen, KULeuven, Belgium [email protected] Escalant, INIBAP [email protected]
Meeting of the PROMUSAGenetic improvement andVirology working groupsLeuven, Belgium,13-14 September 2002Taking the opportunity of the 3rd Internationalsymposium on Molecular and cellular biologyon banana, a meeting of both the GeneticImprovement and Virology working groups ofPROMUSA will be organized back-to-back tothe symposium on 13 and 14 September 2002.
This meeting will take place in Leuven,Belgium.
The meeting will not only facilitate the analy-sis of the current situation on the different activi-ties developed in the framework of PROMUSA,but will also refine the orientation for the groupsand help develop new concept notes in order tofacilitate the writing up of project proposals.
During these two days, the working groupswill meet separately in two different rooms.However, it has been suggested by both bree-ders and virologists to have also a joint meetingto discuss the Banana Streak Virus (BSV) pro-blematics faced by the breeders in the creationof new hybrids. Details on the organization ofthis meeting will be communicated later.
The participation to the meeting will be asusual only on invitation, and will be extendedby the Secretariat (INIBAP) in agreement withthe convenors of the working groups.However, as you may already know, INIBAPcannot assume the participation costs for allthe participants and would appreciate verymuch if you could use the opportunity of the 3rd
International symposium on Molecular and cel-lular biology on banana to find any grant orsupport to participate in sharing the costs.
The convenors of the working groups willcommunicate further details on the workplan.
Contacts: Jean-Vincent Escalant, Secretary of PROMUSA: [email protected] Hughes, Convenor Virology working group: [email protected] Dolezel, Convenor Genetic improvement working group: [email protected] Moffatt, Programme Assistant INIBAP: [email protected]
Meeting announcements





























