Embed Size (px)
Citation preview

1
Title: Mesophyll diffusion conductance to CO2: an unappreciated central player in photosynthesis
Authors: Jaume Flexas1*, Margaret M. Barbour2, Oliver Brendel3, H. M. Cabrera1,4, Marc
Carriquí1, Antonio Diaz-Espejo5, Cyril Douthe3,9, Erwin Dreyer3, Juan P. Ferrio JP6, Alexander
Galle1, Jeroni Galmés1, Naomi Kodama7, Hipolito Medrano1, Ülo Niinemets8, Jose J. Peguero-
Pina1, Alicia Pou1, Miquel Ribas-Carbo1, Magdalena Tomas1, Tiina Tosens8, Charles R. Warren9
1. Research Group in Plant Biology under Mediterranean Conditions, Universitat de les Illes Balears; Carretera de
Valldemossa Km 7.5; 07122 Palma de Mallorca, Illes Balears, Spain
2. Faculty of Agriculture, Food and Natural Resources, The University of Sydney, Private Bag 4011, Narellan NSW
2567, Australia
3. INRA, UMR 1137 “Ecologie et Ecophysiologie Forestières”, F 54280 Champenoux
3. Université de Lorraine, UMR 1137 “Ecologie et Ecophysiologie Forestières”, Faculté des Sciences, F54500
Vandoeuvre, France
4. Centro de Ecología Aplicada Ltda., Av. Suecia 3304, Ñuñoa, Santiago, Chile
5. Instituto de Recursos Naturales y Agrobiología, IRNAS-CSIC, Apartado 1052, 41080, Sevilla, Spain
6. Department of Crop and Forest Sciences, ETSEA, Universitat de Lleida, Avda. Rovira Roure 191; 25198 Lleida,
Spain
7. Agro-Meteorology Division, National Institute for Agro-Environmental Sciences, 3-1-3 Kannondai, Tsukuba, 305-
8604 Japan
8. Institute of Agricultural and Environmental Sciences, Estonian University of Life Sciences, Kreutzwaldi 1, Tartu
51014, Estonia
9. School of Biological Sciences, The University of Sydney, NSW 2006, Australia
*Corresponding author: Jaume Flexas
e-mail [email protected] / fax 34 / 971 / 173184 / tel 34 / 971 / 172365
Grup de Recerca en Biologia de les Plantes en Condicions Mediterrànies, Departament de
Biologia, Universitat de les Illes Balears. Carretera de Valldemossa Km 7.5, 07122 Palma de
Mallorca, Balears, SPAIN.

2
Keywords
aquaporins, climate change, conductance, diffusion, leaf anatomy, plant hydraulics, photosynthesis
Table of contents
Abstract
1. Introduction
2. How different is gm among phylogenetically distant groups and how has gm evolved?
3. New evidence as for the main determinants of gm
3.1. Changing the nature of the diffusing molecule: carbonic anhydrases
3.2. Changing the nature of the diffusing medium: aquaporins
3.3. Changing the diffusing distance: anatomical properties of leaves and cells
4. Which environmental conditions does gm respond to?
4.1. Acclimation to and recovery after drought and salinity
4.2. Nutrient stress
4.3. Changing CO2 concentration around leaves
5. How important is gm in limiting photosynthesis?
5.1. Photosynthesis limitations in response to environmental variables
5.2. Photosynthesis limitations within complex canopies
6. Modeling and including gm in photosynthesis models
7. Can gm be uncoupled from the water path regulation?
7.1. Co-regulation of gm and stomatal conductance
7.2. Co-regulation of gm and hydraulic conductance in the mesophyll
8. Concluding remarks and future prospects
Acknowledgements
References

3
Abstract
Mesophyll diffusion conductance CO2 is a key photosynthetic trait that has been studied
intensively in the past years. The intention of the present review is to update the knowledge on gm,
and highlight the important unknown and controversial aspects that require future work. The
photosynthetic limitation imposed by mesophyll conductance is large, and under certain conditions
it can be the most significant photosynthetic limitation. New evidence shows that anatomical traits,
such as cell wall thickness and chloroplast distribution are amongst the stronger determinants of
mesophyll conductance, although rapid variations in response to environmental changes might be
regulated by other factors such as aquaporin conductance.
There are a number of gaps in knowledge that should be major research priorities for the
near future include, how different is mesophyll conductance among phylogenetically distant
groups and how has it evolved? can it be uncoupled from the water path regulation? what are the
main drivers of mesophyll conductance? The need for mechanistic and phenomenological models
of mesophyll conductance and its incorporation in process-based photosynthesis models is also
highlighted.

4
1. Introduction
Photosynthesis in plants has been considered for decades to be limited only by two factors:
the velocity of diffusion of CO2 through stomata and the capacity of photosynthetic machinery in
leaves to convert light energy to biochemical energy and fix CO2 into sugars. While diffusion is a
passive physical process, it can be subject to large regulation in plants. According to Fick's law,
diffusion depends on substance (e.g. CO2) diffusivity, which differs depending on temperature but
also on the nature (mainly viscosity) of the media in which diffusion occurs (e.g. water, air, etc.),
and the distance of diffusion. Since the mesophyll pathway consists of a complex array of
‘physical barriers’ to CO2 diffusion, including air, cell walls, lipid membranes and liquid
cytoplasm and stroma, differing both in nature and size (i.e. 'distance'), there is a large variation
among leaves in diffusion conductance to CO2 in the mesophyll (gm), and the latter can be
regulated by changing either the distance or the nature of (part of) the diffusion pathway inside
leaves.
Early studies already suggested that the diffusion of CO2 from sub-stomatal cavities to the sites of
carboxylation inside chloroplasts could limit photosynthesis (e.g., [1-3]), which has become more
evident after several methods for the estimation of gm have become more available – including a
method based on 13C-discrimination during photosynthesis[4], a method combining chlorophyll
fluorescence and gas exchange measurements [5, 6] and model-based methods [6-8]. For details on
methods for gm estimation, the required precautions when using them and specific strategies of
adjustment, see refs [9], [10], [11]. Thus, although during decades the majority of researchers
considered gm large enough not to significantly limit photosynthesis, it has become now evident
that that gm is the third major player in the process of photosynthesis, together with stomatal
conductance and biochemical capacity.
The current understanding on gm has been recently reviewed [12]. In addition, specific
reviews on the mechanisms regulating gm [13], and on the ecophysiological and ecological

5
significance of gm [14-16] have been published. These papers are recommended as the best
introduction to the importance of gm in plant physiology. As there has been rapid gain in
understanding of gm, the aims of the present paper are: (1) to update information of current
knowledge accumulated after the recent reviews; (2) to discuss the most obscure / controversial
aspects on gm function and regulation, such as its response to CO2, or how much it limits
photosynthesis; and (3) to highlight the obvious gaps in knowledge on this subject and the future
research needs.
2. How different is gm among phylogenetically distant groups and how have
mechanisms controlling gm evolved?
The rate of diffusion conductance to CO2 in the mesophyll (gm) has now been estimated for
more than 100 species, and it is now possible to search for phylogenetic / evolutionary patterns.
The vast majority of estimates of gm are for Spermatophytes [14] (angiosperms and
gymnosperms), with only very few data for liverworts and hornworts [17]. Most surprisingly there
are no measurements available for phylogenetically intermediate groups such as mosses,
lycophytes, equisetophytes, or ferns. This constitutes a serious gap in our knowledge that
precludes driving any broad conclusion as for the evolution of mechanisms controlling gm.
Some valuable information can nevertheless be obtained by comparing the existing data for
liverworts and hornwort gametophytes with those for Spermatophytes belonging to different
phylogenetic and/or functional groups (Fig.1). At first sight, it is evident that there are variations in
the average rate of gm among different plant groups, and that these variations are more closely
correlated with evolutionary advancements than stomatal conductance (gs). The largest values for
both conductances are found among non-woody angiosperms, whereas grasses present somewhat
higher gm values than annual dicots (Fig. 1). The lowest values are found in liverworts and

6
hornworts for which gs is set as zero as they lack stomata, and CO2 has to diffuse through the
cuticle and epidermis. Among Spermatophytes, conifers show the lowest values. Since
angiosperms are evolutionary more recent than gymnosperms and non-Spermatophytes (the
earliest fossil records for conifers dating back to 290 Myr as compared to 200 Myr for
angiosperms) and grasses represent a relatively recent evolutionary event (ca. 70 Myr), an
evolutionary trend towards larger gm than gs values is plausible. Accordingly, gas exchange
capacity of Angiosperms was greatly increased following the Cretaceous period in association
with leaf morphological changes [18].
A closer inspection reveals that significant differences also appear among angiosperms as a
function of their functional type or growth form. Hence, herbaceous plants show the highest
values, followed by semi-deciduous and deciduous shrubs and trees, while the lowest values are
found in evergreen shrubs and trees, similar to those displayed by gymnosperms. Therefore, at
least, part of the observed variation may not reflect evolutionary trends but is simply the result of
adaptations to particular growth forms and/or environments, e.g., thicker leaves having lower gm
(see next section). Conversely, in some of the groups displayed in Fig. 1, the number of species
included is low and biased. For instance, there are only three genera within grasses (monocots) for
which gm has been determined (Triticum, Oryza and Phragmites), while herbaceous genera are
dicots. Hence, currently available data do not allow distinguishing patterns between monocots and
dicots, or separating the effects of life form and phylogenetic position.
In the case of hornworts and liverworts, estimates of gm are five orders of magnitude
smaller than for Spermatophytes (Fig. 1 inset). Despite the fact that hornworts and liverworts lack
stomata and show a low degree of cuticularization, they may contain some kind of cuticle or
procuticle whose conductance could affect gm estimates. However, the conductance to CO2 of the
thicker and more developed cuticle of higher plants is already around 2.5 10-4 mol m-2 s-1 [19], i.e.,
still an order of magnitude larger than gm estimated in Bryophytes [17]. This evidence suggests

7
that hornworts and liverworts present a truly restricted gm, which may explain their slow growth.
Again, this strongly suggests the occurrence of an evolutionary trend towards higher gm, although
knowledge of gm values of intermediate groups – i.e. mosses and ferns – may be necessary to
confirm such a trend. It is remarkable that values found for bryophytes are similar to the lowest
values of CO2 permeabilities reported for biological membranes [13]. It is not known whether this
means that internal CO2 diffusion in tissues of early land plant forms depends on simple diffusion
through membranes without facilitating agents such as aquaporins. Of the genes encoding for
proteins possibly facilitating gm in higher plants (see section 3), no aquaporin (searched as putative
PIP1 family - i.e. Plasma membrane Integral Protein - gene) or carbonic anhydrase has been
described for hornworts, liverworts, lycophytes, equisetophytes or ferns, and only two putative
genes for PIP1 but up to 13 for carbonic anhydrase have been described in mosses
(www.ncbi.nlm.nih.gov/genbank/). However, this does not necessarily mean that these
genes/proteins do not exist is these groups, as very few data are available in databases for the
genome sequences of these species groups. Moreover, in these groups the importance of carbon
concentrating mechanisms involving carbonic anhydrases, pyrenoids, carboxysomes, etc. probably
mask the importance of gm -related components.
In summary, there is a significant gap in knowledge concerning phylogenetic/evolutionary
trends in gm. To overcome this gap it may be necessary to compile data for monocot species of
families other than Poaceae as well as for lower forms such as lycophytes, equisetophytes, ferns
and mosses. These data should be measured under the same environmental and developmental
conditions to derive phylogenetic implications from a trait like gm, which is under strong
environmental control. This knowledge will help understanding the evolution of gm, the
mechanisms involved and the extent of co-variation of gm and gs.

8
3. New evidence as for the main determinants of gm
3.1. Changing the nature of the diffusing molecule: carbonic anhydrases
CO2 molecules passing from sub-stomatal cavities to chloroplasts diffuse through gas-
phase in leaf intercellular air spaces, liquid phase in cell walls, cytosol and chloroplast stroma and
lipid phase in plasmalemma and chloroplast envelope membranes (Fig. 2). The rate of diffusion
through the composite segments of the diffusion pathway depends on the effective thickness and
diffusivity of each component section [16]. “Effective” denotes the circumstance that the diffusion
path length is generally longer than the linear distance from sub-stomatal cavities to chloroplast
due to tortuosity and/or limited porosity of the diffusion pathway [20]. Diffusion coefficients for
CO2 in free water, tabulated in physical chemistry handbooks cannot be directly applied to leaves
due to presence of solutes and macromolecules in liquid-phase components of the diffusion
pathway, changes in pH, temperature, etc. [20]. Moreover, CO2 can interconvert with HCO3- inside
leaf cells in a reversible reaction catalyzed by carbonic anhydrases. Since the diffusivities of CO2
and HCO3- as well as their pH and temperature dependencies differ [21] carbonic anhydrases could
play a role on the regulation of gm by means of changing the nature of the diffusing molecule.
Early work [22, 23] showed that extremely high reduction in carbonic anhydrase activity
did not result in major photosynthetic limitation. However, it has been also shown that the
contribution of carbonic anhydrase to gm is species dependent, and their role may become more
important when gm is low as in sclerophyllous species [24]. Moreover many different carbonic
anhydrases, with different cellular locations have been characterized [25] and it has been
suggested that carbonic anhydrases can account of 1% of total protein, and those located in
chloroplasts (βCA1 and βCA5) could potentially contribute to increase gm. Up to now, however,
genetic modification of different forms of carbonic anhydrases in Arabidopsis, either chloroplastic
or not, have resulted in differently phenotypes differing in growth but with no measurable
difference in gm (Genty, personal communication). Despite of these results, a role of carbonic

9
anhydrases in the regulation of gm in some species and/or under certain conditions cannot be ruled
out.
3.2. Changing the nature of the diffusing medium: aquaporins
Besides the nature of the diffusing molecule (i.e., CO2 or HCO3-), carbon dioxide diffusion can be
altered either by the nature or the distance of the diffusion media. Concerning the nature of the
diffusion media, the lipid phase is presumably more limiting for CO2 diffusion than the aqueous
phase and this in turn more limiting than the air phase. Membranes (cell, chloroplast) constitute the
physical components of the lipid phase. Although the rate of diffusion of CO2 through membranes
has been often considered very large and the lipid phase assumed to have a negligible effect on
CO2 diffusion, there is still a debate over the degree to which biological membranes are permeable
to CO2, and estimated permeation coefficients vary over several orders of magnitude [26-28]. With
the discovery of aquaporins, it has been suggested that apart from their function in facilitating
water diffusion across membranes they constitute a key means for regulating CO2 diffusion
through membranes [29-31]. Altered expression of aquaporins has been shown to result in changes
in membrane permeability to CO2 in plants [32, 33] and in animals [30, 34]. Experimental
evidence in favor of an important role for aquaporins in CO2 diffusion in intact leaves comes from
studies demonstrating enhanced photosynthesis in plants over-expressing aquaporins, and reduced
photosynthesis in genetically modified plants with impaired aquaporin function (Fig. 3). These
changes are driven at least in part by changes in gm [32-36]. Similarly, reduced photosynthesis and
gm are observed in plants in which aquaporins have been inhibited by HgCl2 [37-39]. However,
genetic transformation to change aquaporin expression and HgCl2 inhibition also lead to
simultaneous changes in net CO2 assimilation rates (An) and stomatal conductance (gs) (Fig. 3, 4)
[40]. Indeed, the relative diffusion limitation of photosynthesis due to gm is directly related to the
CO2 drawdown from sub-stomatal cavities to chloroplasts (Ci-Cc = An/gm) that depends on gm, An

10
and gs (Ci = f(An, gs)) [41, 42]. Therefore, demonstrating changes in gm is not sufficient to confirm
an alteration in the degree to which mesophyll diffusion limits photosynthesis.
Inherent limitation of drawing broad conclusions from such studies is the lack of biological
replication, even if multiple transformed lines have been used in specific studies. So far, no study
with transformants has looked into the changes in quantitative limitations between biochemistry,
stomata and mesophyll diffusion. We calculated the limitations of photosynthesis due to stomata
(ls), mesophyll diffusion conductance (lm) and biochemistry (lb) according to Grassi and
Magnani[43]:
( )
( )
kg
gl
kgg
kgl
kgg
kgl
+=
+=
+=
tot
totb
totm
totm
tots,CO2
tots
(1)
where gs,CO2 is the stomatal conductance to CO2, gtot is the total diffusion conductance for CO2
determined as:
ms,CO2
tot 111
gg
g+
= , (2)
and k is the first derivative of net assimilation rate An with respect to chloroplastic CO2 and is
given as [44]:
( )( )2
mc
mcmax *
KC
KVk
++Γ= , (3)
where Vcmax is the maximum carboxylase activity of Rubisco, Γ*is the hypothetical CO2
compensation point in the absence of dark respiration, and Km is the effective Michaelis-Menten
constant for CO2 that considers the competitive inhibition by O2. Rubisco kinetic characteristics
were taken from Niinemets and Tenhunen [45] and Cc was the value reported in the given study.

11
Due to lack of biological replication, we had to pool different aquaporins atpip1;2-1,
atpip2;3-1, Nt AQP1, HvPIP2;1 in the quantitative limitation analysis. Nevertheless, the analysis
with pooled data for photosynthesis, stomatal conductance and mesophyll conductance broadly
agreed with individual published studies, i.e., that overexpressed lines generally had higher values
of these traits, while antisense transformants had lower values (Fig. 3). Analysis of the quantitative
limitations further demonstrated that the degree of gm-limitation of photosynthesis is larger in
plants with genetically reduced aquaporin levels (Fig. 3). However, in modified lines with over-
expressed aquaporins, gm limited photosynthesis to a similar degree as in control plants due to the
parallel occurrence of reduced stomatal limitation (Fig. 3). In addition, genetic modifications in
aquaporins also led to alterations in the degree of the limitation by photosynthetic capacity per se
(Fig. 3) and also can lead to changes in foliage anatomy and photosynthesis [32-36].
Therefore, there is evidence for the involvement of aquaporins in limiting photosynthetic
CO2 assimilation rate, but it is unclear whether this role is only due to their influence on gm. As for
the thermodynamics of their involvement in gm, simulations based on molecular models have
further indicated high activation energy for CO2 passage through aquaporin monomers, suggesting
that CO2 movement through aquaporins is energetically unfavorable compared with passive
diffusion through membranes, except for highly polar membranes with low CO2 solubility [46-48].
However, the aquaporin family contains a large number of homologs [49, 50], and molecular
simulation studies indicate that the energetic barrier against CO2 movement can be lowered by
only a few point mutations [47, 48], suggesting that in principle, aquaporins most probably exist
with widely varying permeability for dissolved gases. Arabidopsis aquaporin AtPIP1;2-1 has a
much higher permeability for CO2 than AtPIP2;3-1 [32], supporting this concept. It is possible that
having different aquaporins with different activation energies allows plants to regulate CO2
diffusivity and H2O diffusivity more or less independently, although water and CO2 conductances
are often co-regulated (see Section 7). In addition, aquaporin tetramers rather than monomers may

12
be functionally active in CO2 conductance in tobacco [51]. This suggestion is also corroborated by
molecular simulations suggesting that the central cavity of the tetramer requires almost 50% less
activation energy for facilitating CO2 diffusion than the aquaporin monomer [48]. However, this
barrier is still relatively large compared with direct diffusion through some highly hydrophobic
membranes [48], although the theoretical physical parameters of a lipid bilayer are not necessarily
equal to those of a biological membrane [52]. It is also important to consider that all studies on
aquaporins have been conducted in herbaceous species with mesophytic leaves that have
intrinsically high gm due to thin cell walls and high surfaces of chloroplasts exposed to
intercellular air species [14, 15]. In such species, the relative contribution of lipid phase
conductance to total diffusion conductance is expected to be larger than in species with thick cell
walls and low surfaces of chloroplasts exposed such as trees, in particular evergreen trees (Fig. 2).
The bulk of diffusion conductance in evergreens is expected to be in the liquid phase, for which
the diffusion distance can be largely variable.
3.3. Changing the diffusing distance: anatomical properties of leaves and cells
The diffusion distance is given by the size of the different anatomical components of the diffusion
path. In early studies, leaf anatomical characteristics were considered to constitute the chief
limitation of gm [1, 53-57], and anatomical traits were used to infer gm [54, 56]. At the time of
these early studies, the methods to estimate gm had not yet been established, and a separation of
physical diffusion conductance (known as gm today) and biochemical conductance, a variable
quantifying the photosynthetic capacity (chiefly the carboxylase activity of Rubisco) under a
conductance formalism, could not be reliably achieved. On the basis of anatomical measurements,
it was concluded that the physical diffusion conductance inside the leaves was large compared
with the stomatal and biochemical conductances [56, 58]. However, these early studies did not
precisely characterize the thickness of cell walls and cytosol. In addition, they assumed that the

13
diffusion flux in aqueous and lipid phases varies with the ratio of exposed mesophyll area (Smes) to
total leaf area (S) ratio (Smes/S) that characterizes the number of parallel diffusion pathways [54,
55]. This assumes that the exposed surface of mesophyll cells is covered by chloroplasts. Yet,
recent studies have shown that the surface of exposed chloroplasts (Sc) to leaf area ratio (Sc/S) is
generally smaller than Smes/S [15, 59-61], implying that the use of Smes/S as a substitute of Sc/S
underestimates the anatomical constraints on diffusion.
So far, strong negative correlations between gm and cell wall thickness have been demonstrated,
especially when differences in Sc/S are considered by calculating gm per unit exposed chloroplast
surface area [16]. However, there have been surprisingly few quantitative analyses linking gm to
leaf anatomy. The few studies available demonstrate that gm can be quantitatively estimated from
leaf anatomical measurements, although certain assumptions on the tortuosity of diffusion path
length in gas phase, porosity of cell wall, effective diffusion coefficient in cytosol and chloroplast
and permeability of plasmalemma and chloroplast envelope are needed [15, 62]. These
quantitative analyses have corroborated the correlative findings of the role of cell wall thickness
and Sc/S as the main determinants of gm across species with widely varying anatomy [16]. Linking
gm to detailed anatomical measurements is promising, but tedious. Conversely, gm has also been
correlated with integrated leaf traits such as leaf dry mass per unit area, thickness and density [12,
14, 63]. Although strong relationships have been observed when pooling data from multiple
studies, especially for non-stressed plants [14], significant outliers such as Australian sclerophylls
have been denoted in other studies, reflecting the circumstance that high dry mass per unit leaf
area, thickness and density are not always associated with thick cell walls [63, 64]. The
correlations between dry mass per unit leaf area and gm can also vary across environmental
gradients in water availability, that can alter cell wall thickness [15], and in light availability that
can alter Sc/S [59, 65] at given values of leaf traits such as dry mass per unit area, thickness and
density.

14
4. Which environmental conditions does gm respond to?
Mesophyll conductance to CO2 responds to environmental factors either in the long term or
rapidly, i.e. in minutes-hours [10]. Recent reviews have already highlighted the incidence of
varying environmental conditions such as soil water availability, salinity, growth irradiance and
temperature on gm [12, 14]. In the recent years, the important contribution of gm in limiting
photosynthesis during drought and salinity has been emphasized, knowledge has improved as for
nutrient stress effects on gm, and many more data have been obtained regarding the controversial
effects of rapid changes in CO2 concentration on gm.
4.1. Acclimation to and recovery after drought and salinity
Under drought and salinity, the degree of photosynthesis limitation by gs and gm is similar
(see Section 5), but gm can respond to water deficit and subsequent re-watering at different
velocities than gs [66-69]. Most remarkably, gm acclimates and recovers during prolonged drought
in tobacco and grapevine plants [66, 67], while in stressed soybeans it was hardly affected [70]
despite a significant reduction of gs in all cases. In a study on tobacco [67], gm and gs first
decreased during the onset of drought, whereas during sustained drought, gm recovered to control
values despite maintenance of low gs. Moreover, drought interacted with environmental
conditions, since gm did not decrease when the experiment was repeated at lower light intensities
and milder temperatures [59]. Thus, gm does not necessarily respond in the same manner as gs
when stress is prolonged and factors other than hydraulics or water status impact on gm.
Furthermore, gm exceeded control levels after re-watering in the Mediterranean semi-deciduous
shrub Cistus albidus [71]. In C. albidus, consecutive drought and re-watering cycles resulted in a
drought cycle-dependent adjustment of leaf gas exchange towards reduced limitation by gm and
enhanced water use efficiency with each consecutive cycle, while the evergreen oak Quercus ilex

15
had a very stable response during successive cycles, lowering gm and gs in parallel [71].
Acclimation of gm during prolonged drought and facilitated recovery after re-watering represent an
effective way of optimizing CO2 diffusion transiently under limited CO2 supply, thereby boosting
photosynthetic activity and water use efficiency after relief of stress. On the other hand, gm and
photosynthetic activity can vary among ecotypes of the same species under similar water
availability due to differences in e.g. photosynthetic capacity, osmotic adjustment and leaf
structure [69, 72]. Thus, plant growth form alone cannot explain the differences in gm response to
limited water availability, but may allow for contrasting general trends in photosynthesis among
evergreens and deciduous species [73]. Whole plant structure also affects leaf diffusion
components and hence the capacity to recover from and survive drought, as observed in pre-
conditioned nursery plants of different age and size [74] and in the woody legume Prosopis
velutina [75]. Low root to shoot ratios [74] as well as lasting effects of hydraulic failure [75]
resulted in a delayed or only partial recovery of gm and leaf gas exchange after re-watering.
4.2. Nutrient stress
Compared with responses to drought, little is known about the influence of plant nutrition
on gm, and only recently have the effects of nutrition been addressed. In line with some earlier
reports, leaf nitrogen content correlates positively with photosynthetic activity and gm across
several woody and herbaceous species [76-78], whereas a negative relationship of leaf nitrogen
and gm was observed when related to tree height in Pinus densiflora [79, 80]. Such a decline of gm
with tree height can be related to decreasing water potential affecting leaf expansion and structure
[80, 81].
Photosynthetic activity and gm correlated well with the supply of K in hickory seedlings
[82], whereas biochemical modifications and/or structural changes seemed to primarily limit
photosynthesis. More research is needed to gain further insight into the K - gm relationship.

16
Changes in P nutrition revealed no direct effect on gm [69, 83]. Excess of Zn in Beta vulgaris and
excess nickel Ni in Populus nigra strongly decreased leaf conductance (gm and gs) [84, 85]. This
was presumably caused by changes in mesophyll structure, which affected leaves developing
during stress more than mature ones [85]. Therefore, photosynthetic activity under excess Zn and
Ni was primarily limited through impaired leaf conductance.
4.3. Changing CO2 concentration around leaves
The effects of varying CO2 concentrations on gm display different trends for long-term
(acclimation) and for short term responses. No general trend has been observed for plants grown
under elevated CO2 (i.e. 500-600 ppm) with no change, decreased or increased gm being reported,
possibly depending on the species and time [86, 87]. In contrast, changes in gm under short-term
exposure to different CO2 concentrations seem to follow a general trend across many species [88-
94], with a negative correlation between gm and sub-stomatal and/or ambient CO2 concentration
(Fig. 5). However, some exceptions have been published [95], leading to a controversy as to
whether the observed responses are real or a simple measurement artefact. The data recorded under
low CO2 (< 100 ppm) have to be taken with caution due to errors with the available techniques
[10, 89] and it seems very likely that gm declines with rising CO2 levels. The same trend was
observed with very different techniques, each relying on very different assumptions [see e.g. 9]
[11] [86] [96] [97]. As a general pattern, gm largely differs at the ends of a Ci gradient (0 to 2000
µmol mol-1) (Fig. 5). Still, variation of gm within a smaller range of CO2 concentrations might be
small. This is particularly true when referring to CO2 changes projected for the next few decades
(e.g., the range from 300 to 800 µmol mol-1 in Fig. 5, which reflects a range of ambient CO2
concentrations of about 500 to 1000 µmol mol-1).
Several explanations for the decline of gm with increasing CO2 have been proposed,
including fine adjustments for balancing Ci and Cc, and avoiding large decreases of cell pH [12]

17
and increased cell leakiness under high pCO2 [98]. However, these adjustments might be restricted
by structural conditions at the mesophyll/chloroplast level, allowing gm to change within a
relatively narrow range (see Section 3 on the possible limit of the contribution of aquaporins to
gm). Moreover, other internal factors such as respiration may also affect the determination of gm
under varying CO2 concentrations, e.g. if a constant value is assumed. The true rate varies with
CO2, but activities of respiratory pathways can change the resistance of the diffusion pathway, e.g.
by altering the mean path length [98]. In fact several reports, either theoretical [98] or empirical
[92, 99], but not others (Flexas), have described an effect of O2 concentration on gm and its
dependency on CO2 concentration. The reasons for this effect remain unclear, although the
influence of photorespiration and of changes in the spatial distribution of CO2 emission (in
mitochondria) and carboxylation (in the chloroplasts) have been claimed. Clearly, more detailed
studies are required to fully understand the effect of O2 concentration on gm.
5. How important is gm in limiting photosynthesis?
5.1. Photosynthesis limitations in response to environmental variables
Once it was demonstrated and accepted by most of the scientific community that gm is finite, and
possibly dynamically regulated, it became important to quantify how much mesophyll diffusion
limits photosynthesis. In the 90’s and beginning of this century, photosynthesis limitation by gm
was ignored - for simplicity and because of the difficulty to estimate gm with methods available -
despite the early warnings that gm was finite, variable and limiting photosynthesis ([3] [100]).
Recently, a comprehensive analysis was described where total photosynthesis limitations were
estimated and disentangled into its three components: stomatal, mesophyll and biochemical
limitation [43]. This is a relative analysis because the percentage of reduction of the net
assimilation rate for each limiting component is estimated on the basis of a control value without
any of these limitations. As the severity of the stress progresses total limitation increases and so

18
potentially does each of the three components. Up to now, such analysis has been applied mainly
to quantify water stress-induced limitations in herbaceous [90, 101] and woody species [102-105],
as well as during recovery from water stress [67, 71, 101]. The limitation of photosynthesis by gm
was followed as well during seasonal changes [106, 107], leaf ontogeny [81, 90, 108], temperature
acclimation [109], Zn contamination [84] and nitrogen and phosphorus fertilization [83]. Data for
limitation analysis from all these studies are pooled in Figure 6, in which the Total limitation was
considered 0 for the ‘control’ plants (i.e., non stress conditions), and increasingly higher for
stressed plants up to a maximum of 1. It can be observed that, as total limitation increases with
increasing stress, stomatal mesophyll conductance and, to a lesser extend, biochemical limitations
increase (Fig. 6). The scattered results are a consequence of including a number of species with
different growth forms with a range of structural and anatomical characteristics, different types of
stress, and varying severities of the same type of stress from mild to severe stress in the same
analysis. Yet, it can be observed that up to a total limitation of 25%, limitations are mainly
represented by stomata and mesophyll. From this point to larger total limitation, all three
limitations increase, but the relative contribution of stomata becomes generally the larger,
followed by that of mesophyll and, finally biochemical limitations. Cases in which mesophyll
limitations account for more than 75% of TL are under mild water stress [101], first stages of re-
watering [101], where atmospheric demand was impaired from soil water availability [105], or in
the response of photosynthesis to temperature [109]. All the data available point out that
mesophyll conductance limitations to photosynthesis are of similar magnitude as stomatal
limitations, and generally greater than biochemical limitations. Besides limiting photosynthesis in
response to environmental clues, the spatial variability of gm within canopies, especially in relation
to height in tall trees, has also an impact in limiting photosynthesis.
5.2. Photosynthesis limitations within complex canopies

19
As mentioned in Section 3, gm is highly determined by leaf structure, as suggested by the
negative relationship between gm and leaf mass per unit area. Additionally, gm scales positively
with leaf photosynthetic capacity. However, both variables, leaf mass per unit area and
photosynthetic capacity, are positively correlated with local irradiance in canopies, so that opposite
forces operate when increasing height, controlling the gm of leaves. The situation is further
complicated by the fact that in evergreens leaves of different age are found at different tree
heights. Hydraulic limitations in very tall trees exert a limitation on gs especially in the upper
leaves which, in turn, may exert a negative feedback on gm but at the same time older leaves with
expectedly reduced gm might be found at the lower parts of the canopy. As expected, different
results have been obtained when analyzing tree height / leaf canopy position on gm.
For instance, sun leaves at the top of a 34 m tall conifer Pseudotsuga menziesii were
compared with leaves collected at the bottom (10% incident PPFD) [110]. Despite a significant
difference in leaf mass per unit area there was a strong correlation between An and gm. The
variability of gm was high within each irradiance level or leaf type, and the differences in gm
between the locations were not significant. Mesophyll conductance was also studied across the
canopy profile in different-aged leaves of the oak Quercus ilex. Here, in contrast to Pseudotsuga,
strong positive curvilinear relationships between gm and mean irradiance were found for all leaf
age classes except in oldest leaves [111]. However, the degree of limitation of photosynthesis by
gm was actually slightly larger at higher irradiance, suggesting increasing photosynthetic limitation
by gm at the top of the canopy [111]. The spatial distribution of structural (leaf N and chlorophyll
content and leaf mass per unit area), and functional leaf traits (maximum velocity of carboxylation,
maximum capacity for electron transport and gm) were studied along the canopy of the deciduous
Fagus sylvatica in relation to irradiance and leaf age [112]. A multivariate approach was used
based on path analysis to disentangle the relationship among these variables. The primary role of
structural adjustment was confirmed i.e., the plastic response of leaf mass per unit area which was

20
in this case negatively related to gm, for the acclimation of leaves to the local irradiance in a
canopy. In much taller trees, however, hydraulic limitations could exert a larger role than local
irradiance in setting photosynthetic characteristics of the top leaves. For instance the response of
leaf mass per unit area to local irradiance was altered in Sequoia sempervirens (113 m) as height
increased [113]. There was a transition region in the canopy where the primary determinants of
leaf morphology and structure switched from local irradiance to hydraulics. It was concluded that
structural changes due to hydraulics indirectly reduced net CO2 assimilation rates via increased
respiration rates and decreased gs and gm.
The other question is whether total tree height itself (i.e., not the height of a given leaf
inside a tree, but the total height of the tree itself) can alter gm and its role in constraining
photosynthesis. Specifically, leaf mass per unit area increases as trees increase in height and this is
associated with reductions of net CO2 assimilation [114-116], gs [117] and, perhaps, gm [114, 118,
119]. Lower gm in taller trees has recently been confirmed experimentally. There was a decrease of
gm with total tree height in the conifer Pseudotsuga menziesii [120], suggesting that gravity and the
water path length were likely the main determinants of trends in foliar characteristics via their
effects on leaf water potential during leaf expansion. Also, there was a tight relationship between
leaf mass per unit area and gm in Pinus densiflora that was the inevitable consequence of the
morphological acclimation to height [79]. Similarly, the age effect on gm in leaves of the
deciduous Nothofagus solandri was only evident in tall trees (15 m tall), while in shorter trees (2
m tall) no differences were found [81]. It seems that changes in gm with tree height occur in
proportion to changes in gs and photosynthetic capacity, such that photosynthesis is limited to a
similar degree by gm in different-size trees [14, 116].
To sum up, most studies conclude that the main determinant of gm modifications with tree
height is leaf structure represented by leaf mass per unit area, which can be modulated by local

21
irradiance, leaf age and hydraulic gradients. The complexity of their interactions does not allow
yet a clear prediction of gm changes with tree height.
6. Modeling and including gm in photosynthesis models
Mesophyll diffusion of CO2 must be taken into account in leaf gas exchange models, since
considering an infinite gm is not correct. The difficulty rises when deciding a value of gm to be
applied in each specific scenario, and as a function of how it varies in space and time. Currently,
we are not able to incorporate gm in models with a mechanistic basis due to the lack of sufficient
knowledge on the mechanisms involved in the regulation of gm. This being said, there have been
several attempts to empirically include gm in models. One of the first examples was proposed
modeling the soil-plant-atmosphere continuum in a Quercus-Acer forest [121]. These authors, as
[122], concluded that Vcmax and Jmax were underestimated in most studies due to neglecting gm.
They included a different constant value of gm for each species. Later on, two approaches were
used in rice to include a variable gm as a function of N or as a function of gs [123]. The models
were based on previous work where gm usually scaled with gs. The model based on gs explained
more variation in measured An than the model based on N. It was argued that gm changes during
different developmental stages of the crop, and the apparent coordination between gm and gs
allowed the use of gs as a scaling factor. The same approach was used in modeling the response of
C3 and C4 plants to water stress [124]. The inclusion of gm in a photosynthesis model was further
justified by the response of gm to temperature [11]. None of these approaches allows for a flexible
dependency between gm, gs and photosynthetic capacity like the one that occurs, for instance,
under water stress.
One of the remaining challenges in leaf gas exchange models is to take into account the
effects of seasonal water stress. Water stress, as mentioned above, affects the relative importance
of each component in limiting photosynthesis, depending on the degree of stress. In this sense,

22
some have used a limitation analysis [43] to infer what must be included in the models that mimic
the observed behavior of An. Three values of gm were used to simulate An assuming that stomatal
limitation is equal, smaller or larger than mesophyll limitation [125]. It was concluded that
diffusive limitations can explain water flux responses to seasonal changes in soil water availability
only if gm was included in the models. The use of either stomatal or biochemical limitation alone
did not mimic the observed data. The same conclusion was reached about the importance of
combining several components in the limitation during acclimation to stress [108]. They concluded
that to reproduce the general pattern of C3 photosynthesis during water stress, the highest
limitation strength must be imposed by gm, then by gs, and finally by the biochemical capacity.
The impact of gm on model predictions of carbon isotope discrimination has been also
assessed by testing whether a fixed or a variable gm depending on gs or time of day improved
model predictions in mature juniper trees [126]. The incorporation of gm in the model did not
consistently improve carbon isotope discrimination. These results contrast with those were the
inclusion of a variable gm (as a function of gs), improved the model predictions of isotope
composition of respired carbon from a coastal Douglas-fir forest in comparison with a model with
a fixed gm [124]. Recently, [127] have warned about the need of including the ternary effect of
transpiration rate in the equations for carbon isotope discrimination. The effect is greatest when the
leaf-to-air vapor mole fraction difference is greatest, which could explain some of the
contradictory results commented above.
Although it is obvious that a realistic model for predicting An should incorporate gm, Oliver
et al. [128] concluded that the use of gm did not improved the performance of the An model, as long
as Vcmax was seasonally tuned. Effectively, a similar prediction of An can be obtained either
reducing gm or overreducing Vcmax, i.e., Cc or Ci-basis. However, if we are interested in using a
mechanistic model, the actual regulation of gm and Vcmax as a function of the degree of stress
should be taken into account. In this sense, Niinemets et al. [14] showed how the inclusion of gm in

23
models results in a description of leaf acclimation to changing environmental conditions, and in a
more realistic description of daily photosynthesis, especially in leaves under stress. While An-Ci
parameterization predicted a negative carbon balance at midday in plants under water stress, actual
measurements and simulations with the An-Cc approach yielded a carbon gain. Similarly, a
biochemical photosynthesis model on a Cc-basis was used to explain the potentially favorable
response of evergreens plants to climate change due to their robust leaves and low gm [15].
Currently, the determination of the seasonal evolution of gm or its dynamic in cycles of stress and
recovery, and even distribution within canopies, is seen as a huge drawback in the process of
incorporation in models of process-based land-surface schemes [128]. Definitely, more
information is needed to make possible the inclusion of gm in leaf-gas exchange models. This
information is hard to be obtained at a large scale of space and time due to the limitations in the
use of most used techniques of gm determination [10]. However, more efficient and
straightforward methods for determining average canopy gm, like that proposed by Ubierna and
Marshall [129] based on δ13C of phloem content, can be useful in the future for ecophysiological
and ecosystem model applications.
7. Can mesophyll conductance be uncoupled from the water path regulation? 7.1. Co-regulation of gm and stomatal conductance
The previous sections have demonstrated that gm and gs are very often co-regulated,
although not under all instances. Some degree of co-regulation has been suggested between gm and
plant hydraulics. Water vapor and CO2 share at least a part of their pathways in leaves. Both gases
are exchanged with the atmosphere through stomata. In addition, both water vapor and CO2 must
cross the aerial sub-stomatal cavity. Additionally, after leaving the leaf xylem, liquid water not
only moves along apoplastic pathways but also (partly mediated by aquaporins) crosses cell
membranes and flows through the plasmalemma (symplastic pathway) and cell vacuoles

24
(transcellular pathway) to the sites of evaporation [130, 131]. Accordingly, liquid water and CO2
diffusion share partly common diffusion pathways in the mesophyll [13, 16]. The involvement of
aquaporins in the diffusion of water and possibly CO2 also suggests at least partly common
pathways for both molecules, although it is also possible that different aquaporins could be
involved in each case.
Because of overlapping transport pathways, some degree of co-regulation is expected
between gm and water transport in leaves. Indeed, variations of gm are generally closely related to
those in gs, e.g., variations among species (see Section 2), induced by water stress (see Section 4),
etc, but not necessarily in transgenic plants with different levels of aquaporins (see Section 3) or
under combined water stress and low irradiance (see Section 4). From a purely photosynthetic
perspective, this co-regulation is expected and may optimize photosynthesis. For instance, energy
and water-consuming stomatal opening does not translate into effective photosynthesis if gm equals
zero, but requires higher gm.
However, from the perspective of leaf water use efficiency, theoretical considerations
suggest that some uncoupling between the two conductances may be advantageous. Under steady-
state, net photosynthesis is:
An = gs (Ca – Ci) = gm (Ci – Cc),
where Ca, Ci and Cc are the atmospheric, sub-stomatal and chloroplastic CO2
concentrations, respectively. An /gs is of the intrinsic water use efficiency at the leaf level, as gs
controls transpiration in a constant environment. From the equation, at constant photosynthetic
activity (i.e., ‘demand’ for Cc), increased An/gs can be achieved by increasing the ratio of gm to gs.
Some degree of uncoupling between the two conductances, i.e. variation in gm/gs has been
observed. There is a progressive increase in gm along the leaf of the monocot Triticale, but little
variation in gs [132]. In this case, gm is the main driver for changes in observed carbon isotope
discrimination (∆13C), an indicator of Cc / Ca. Dry climate populations of the deciduous trees Picea

25
[133] and Populus [134] have larger gm/gs than their relatives from milder climates. Genetic- and
drought-induced variability in gm/gs in grapes [135] and tomato [136] is significantly and
positively correlated with water use efficiency. In all these examples, increased gm/gs and water
use efficiency was accompanied by decreased An, suggesting that manipulating water use
efficiency by means of gm regulation may always result in decreased production capacity.
However, a simultaneously higher An, gm/gs and water use efficiency was observed in the
evergreen conifer Abies pinsapo than in its close relative A. alba (Peguero-Pina, unpublished).
Similarly, a close relationship was found between An and gm along a range of Pseudotsuga
menziesii trees varying in total height, but virtually no change in gs i.e., a higher gm/gs as total
height declined and An and water use efficiency increased [80]. Understanding the mechanisms
regulating gm in such a way that it can uncouple from gs to some extent may be an essential step
for future genetic manipulation of plants aiming simultaneous increases in photosynthesis and
water use efficiency.
7.2. Co-regulation of gm and hydraulic conductance in the mesophyll
At least in part, tight co-regulation of gm and gs may arise from the close relationship often
found between gs and the conductance of water within the mesophyll. Up to now, it has not been
possible to determine hydraulic conductance of the mesophyll directly, but several methods to
measure whole leaf hydraulic conductance (Kleaf) are available, mainly based on the measurement
of water flow relative to a water potential gradient[137]. A general positive relationship occurs
across species between gm and Kleaf, with fast growing species showing the highest values for both
variables and conifers falling in a group with the lowest values (Fig. 7a). The underlying reason
for such relationship might be the existence of anatomical limitations to CO2 and water
conductances, which are likely to be higher in species with thicker mesophyll layers and greater
surface of mesophyll cells [80, 138-142]. Nevertheless, this does not seem to be a universal

26
relationship, and herbaceous monocots with high gm, around 0.5 mol m-2 s-1 [36, 132, 143], may
have Kleaf values below 5 mmol H2O s-1 MPa-1 m-2 [144, 145]. Whether this is due to anatomical or
biochemical particularities of these species is still a matter of debate. In this sense, at least for
some species the pattern of change in response to drought and tree height for gm and Kleaf is
comparable to the general pattern observed across species (Fig. 7b), suggesting that not only
anatomical but also differences in biochemical regulation are involved in interspecific differences
for both variables.
The main limitation of Kleaf as a surrogate for the hydraulic conductance of the mesophyll
is that Kleaf involves both mesophyll and xylem resistances, and although the former plays a
significant role in whole Kleaf, its relative contribution may vary with species and experimental
conditions [130, 137, 146, 147]. Studies on the environmental response of leaf water isotopic
enrichment offer a new way to assess short-term changes in mesophyll hydraulic resistance [148-
151]. During transpiration, leaf water becomes enriched in the heavier isotopes, 18O and 2H. The
enrichment at the sites of evaporation can be modeled from environmental variables [152, 153].
(see Appendix I for details). However, the observed enrichment in the leaf lamina does not
generally agree with modeled values at the site of evaporation, since back diffusion of enriched
water from the sites of evaporation to the rest of the leaf is counteracted by a mass flow of non-
enriched water driven by transpiration (Péclet effect [154]). The magnitude of this effect is
proportional to the transpiration rate, the distance from the xylem to the evaporative surface, and a
scaling factor, which accounts for the higher velocity of water through a porous media than if it
were moving through the leaf as a slab (i.e. as derived from transpiration). From these models, a
“scaled effective path length” (Leff, the product of the actual distance and the scaling factor) can be
determined by comparing modeled enrichment at the site of evaporation with observed values
[154-156]. Since Leff accounts both for the length of the water pathway and its tortuosity, it is
theoretically related to mesophyll hydraulic resistance to water flow. In a recent work, Ferrio et al.

27
[149] showed a tight link between Leff and both Kleaf and gm in response to experimental treatments
(Fig. 8), although the relationship between Leff and gm reached an asymptote at higher values of gm
(gm >200 mmol CO2 m-2 s-1; Fig. 8b). These findings provided empirical evidence in support of the
theoretical link between Leff and hydraulic conductance of the mesophyll. Most interestingly, they
also showed that at least in response to certain environmental variables, changes in gm and
hydraulic conductance of the mesophyll are closely and positively related, implying that the gm / gs
ratio can vary but over a limited range. Nevertheless, the lack of relationship between gm and Leff
at higher values of gm suggests that diffusion of CO2 can be enhanced beyond the common
limitations for water and CO2. It is likely, for example, that well-watered plants may have reached
their maximum values of hydraulic conductance of the mesophyll, determined by anatomical
limitations, while gm may still respond to other regulations that maximize photosynthesis. One
possible explanation for the uncoupling between CO2 and water conductances has been proposed
[51]: the aquaporin NtAQP1, from the PIP1 family, did not increase water transfer, but enhanced
CO2 diffusion. Conversely, NtPIP2;1 from the PIP2 family, favored water transport but did not
affect CO2 diffusion. The changing of the proportion of the two different aquaporins in a tetramer
progressively varied the water and CO2-related functions. As a consequence, even though both leaf
hydraulics and CO2 diffusion respond to changes in aquaporin conductivity, different
combinations of aquaporin subunits in aquaporin tetramers may promote either water or CO2
transfer, or both, depending on the proportion of PIP1 or PIP2. Thus, regulation of the function of
aquaporins would take place as a result of a competition among subunits for the formation of
tetramers, in a way that would allow enhancing CO2 fixation and at the same time reducing water
use. Indirect evidence in vivo in support to this hypothesis was obtained by comparing the values
of gm and effective path length in a wild type and two tobacco mutants and antisense and
overexpressing NtAQP1lines (Fig. 9) (Kodama, unpublished, plants courtesy of Dr. R.
Kaldenhoff). Plants with increased gm had increased effective path length (i.e., decreased hydraulic

28
conductance of the mesophyll). The overexpressing line (with a greater expression of PIP1s) had
the highest gm and effective path length, and the antisense line had the lowest values. We speculate
that this is the result of different aquaporin forms proportion in tetramers following altered
expression of one of the two forms (PIP1 and PIP2). According to [51], one water conducting
aquaporin (PIP2) is enough to facilitate water transport to a level close to maximum, while 3 or 4
CO2 conducting aquaporins (PIP1) are required to reach maximum CO2 diffusion. We suggest that,
in antisense lines, the proportion of PIP1 aquaporin in most aquaporin tetramers falls well below 3,
while in overexpressing lines the proportion is between 3 and 4. In contrast, the effective path
length and gm were negatively correlated when comparing individual plants within each mutant
type, suggesting a common trend for CO2 and water conductance. In this case, we suggest that
variations in the total expression of aquaporins among individuals, while keeping identical
proportions among subunits, results in a concomitant increase in conductance of CO2 and of water
vapor.
8. Concluding remarks and future prospects
We have stressed that the share of overall photosynthetic limitation by mesophyll
conductance is large and can be the most significant factor limiting photosynthesis under certain
conditions and certain plant functional types. This statement is backed up by ample evidence, and
we argue that gm should be included in any study analyzing limitations to photosynthesis, as well
as in models for predicting rates of photosynthesis.
Significant progress has recently been made in quantitatively linking gm to foliage
anatomical and structural traits. Cell wall thickness and chloroplast distribution seem to play a
dominant role in determining the upper limit of gm. However, rapid variations in response to
environmental cues might not be regulated by anatomical traits. Aquaporins seem to be only partly
responsible, although their mechanistic bases remain unclear.

29
We conclude that further developments in the field require more advanced understanding
of the currently most obscure points, which include:
1. The role of aquaporins in diffusion conductance, especially in species growing in
stressful environments and having particularly low values of gm.
2. To what extent water and CO2 transport processes are coordinated and how does this
affect photosynthesis? It seems that the coordination is not necessarily maintained across
environmental gradients and gradients of tree height.
3. What is the genetic basis of gm and its genetic variability? Data on genetic variability
within species, the degree of heritability of gm, as well as for entire phylogenetic groups, notably
ferns and mosses are far too scarce.
Acknowledgements
We thank Ichiro Terashima and Yusuke Mizokami for insightful suggestions on the MS
and editor plus five additional anonymous reviewers for numerous useful improvements. The
study was financially supported by the Estonian Ministry of Science and Education (grant
SF1090065s07), the Spanish Ministry of Science and Innovation through projects BFU2008-
01072 (MEFORE), AGL2009-11310/AGR, BFU2011-23294 (MECOME) and CGL2009-13079-
C02-01 (PALEOISOTREE), and the European Commission through European Regional Fund (the
Estonian Center of Excellence in Environmental Adaptation), and the Marie Curie project MC-
ERG-246725 (FP7). J.P.F. is supported by the Ramón y Cajal program (RYC-2008-02050). A.G.
had a Swiss National Science Fellowship (PA00P3_126259). M.M.B. is supported by an
Australian Research Council Future Fellowship (FT0992063). CD was supported by a grant from
the French government and by the cooperation project Tranzfor (Transferring Research between
EU and Australia–New Zealand on Forestry and Climate Change, PIRSES-GA-2008-230793)
funded by the European Union.

30
References
[1] P.S. Nobel, Internal leaf area and cellular CO2 resistance: photosynthetic implications of variations with growth conditions and plant species, Physiol. Plant., 40 (1977) 137-144. [2] P.S. Nobel, Biophysical plant physiology and ecology, W. H. Freeman and Company, San Francisco, 1983. [3] P. Gaastra, Photosynthesis of crop plants as influenced by light, carbon dioxide, temperature, and stomatal diffusion resistance. , Mededelingen van Landbouwhogeschool te Wageningen, Nederland 59 (1959) 1-68. [4] J.R. Evans, T.D. Sharkey, J.A. Berry, G.D. Farquhar, Carbon Isotope Discrimination measured Concurrently with Gas Exchange to Investigate CO2 Diffusion in Leaves of Higher Plants, Funct. Plant. Biol., 13 (1986) 281-292. [5] P.C. Harley, F. Loreto, G.D. Marco, T.D. Sharkey, Theoretical Considerations when Estimating the Mesophyll Conductance to CO2 Flux by Analysis of the Response of Photosynthesis to CO2, Plant Physiol., 98 (1992) 1429-1436. [6] G. Bongi, F. Loreto, Gas-exchange properties of salt-stressed olive (Olea europaea L.) leaves., Plant. Physiol., 90 (1989) 1408-1416. [7] G.J. Ethier, N.J. Livingston, On the need to incorporate sensitivity to CO2 transfer conductance into the Farquhar–von Caemmerer–Berry leaf photosynthesis model, Plant Cell Environ., 27 (2004) 137-153. [8] Ü. Niinemets, A. Cescatti, M. Rodeghiero, T. Tosens, Leaf internal diffusion conductance limits photosynthesis more strongly in older leaves of Mediterranean evergreen broad-leaved species, Plant Cell Environ., 28 (2005) 1552-1566. [9] C. Warren, Estimating the internal conductance to CO2 movement, Funct. Plant Biol., 33 (2006) 431-442. [10] T.L. Pons, J. Flexas, S. von Caemmerer, J.R. Evans, B. Genty, M. Ribas-Carbo, E. Brugnoli, Estimating mesophyll conductance to CO2: methodology, potential errors, and recommendations, J. Exp. Bot., 60 (2009) 2217-2234. [11] X. Yin, P.C. Struik, Theoretical reconsiderations when estimating the mesophyll conductance to CO2 diffusion in leaves of C3 plants by analysis of combined gas exchange and chlorophyll fluorescence measurements, Plant Cell Environ., 32 (2009) 1513-1524. [12] J. Flexas, M. Ribas-Carbó, A. Diaz-Espejo, J. Galmés, H. Medrano, Mesophyll conductance to CO2: current knowledge and future prospects, Plant Cell Environ., 31 (2008) 602-621. [13] J.R. Evans, R. Kaldenhoff, B. Genty, I. Terashima, Resistances along the CO2 diffusion pathway inside leaves, J. Exp. Bot., 60 (2009) 2235-2248. [14] U. Niinemets, A. Diaz-Espejo, J. Flexas, J. Galmes, C.R. Warren, Role of mesophyll diffusion conductance in constraining potential photosynthetic productivity in the field, J. Exp. Bot., 60 (2009a) 2249-2270. [15] T. Tosens, Ü. Niinemets, V. Vislap, H. Eichelmann, P. Castro-Díez, Developmental changes in mesophyll diffusion conductance and photosynthetic capacity under different light and water availabilities in Populus tremula: how structure constrains function, Plant Cell Environ., (2011) In Press. [16] I. Terashima, Y.T. Hanba, D. Tholen, Ü. Niinemets, Leaf functional anatomy in relation to photosynthesis, Plant Physiol., 155 (2011) 108-116. [17] M. Meyer, U. Seibt, H. Griffiths, To concentrate or ventilate? Carbon acquisition, isotope discrimination and physiological ecology of early land plant life forms, Philos. T. R. Soc. B., 363 (2008) 2767-2778.

31
[18] T.S. Feild, T.J. Brodribb, A. Iglesias, D.S. Chatelet, A. Baresch, G.R.J. Upchurch, B. Gomez, B.A. Mohr, C. Coiffard, J. Kvacek, C. Jaramillo, Fossil evidence for Cretaceous escalation in angiosperm leaf vein evolution., P. Natl. Acad. Sci. USA., 108 (2011a) 8363-8366. [19] J.S. Boyer, S. Chin Wong, G.D. Farquhar, CO2 and water vapor exchange across leaf cuticle (epidermis) at various water potentials, Plant Physiol., 114 (1997) 185-191. [20] Ü. Niinemets, M. Reichstein, Controls on the emission of plant volatiles through stomata: a sensitivity analysis, J. Geophys. Res. Atm., 108 (2003) 4211, doi:4210.1029/2002JD002626.
[21] R.E. Zeebe, On the molecular diffusion coefficients of dissolved CO2; HCO3- and CO32- and
their dependence on isotopic mass. , Geochim. Cosmochim. Ac., 75 (2011) 2483-2498. [22] D. Price, S. von Caemmerer, J.R. Evans, J.W. Yu, J. Lloyd, V. Oja, P. Kell, K. Harrison, A. Gallagher, M. Badger, Specific reduction of chloroplast carbonic anhydrase activity by antisense RNA in transgenic tobacco plants has a minor effect on photosynthetic CO2 assimilation., Planta, 193 (1994) 331-340. [23] T.G. Williams, L.B. Flanagan, J.R. Coleman, Photosynthetic gas exchange and discrimination against 13CO2, and C18O16O in tobacco plants modified by an antisense construct to have low chloroplastic carbonic anhydrase. , Plant Physiol., 112 (1996) 319-326. [24] J.S. Gillon, D. Yakir, Internal conductance to CO2 diffusion and C18OO discrimination in C3 leaves. , Plant Physiol., 123 (2000) 201-213. [25] N. Fabre, I.M. Reiter, N. Becuwe-Linka, B. Genty, D. Rumeau, Characterization and expression analysis of genes encoding α and β carbonic anhydrases in Arabidopsis., Plant Cell Environ., 30 (2007) 617-629. [26] W.F. Boron, V. Endeward, G. Gros, R. Musa-Aziz, P. Pohl, Intrinsic CO2 permeability of cell membranes and potential biological relevance of CO2 channels, ChemPhysChem, 12 (2011) 1017-1019. [27] B.L. de Groot, J.S. Hub, A Decade of Debate: Significance of CO2 Permeation through Membrane Channels still Controversial, ChemPhysChem, 12 (2011) 1021-1022. [28] A. Missner, P. Kügler, S.M. Saparov, K. Sommer, J.C. Mathai, M.L. Zeidel, P. Pohl, Carbon dioxide transport through membranes, J. Biol. Chem., 283 (2008) 25340-25347. [29] G.J. Cooper, W.F. Boron, Effect of PCMBS on CO2 permeability of Xenopus oocytes expressing aquaporin 1 or its C189S mutant, Am. J. Physiol., 275 (1998) 1481-1486. [30] V. Endeward, R. Musa-Aziz, G.J. Cooper, L.-M. Chen, M.F. Pelletier, L.V. Virkki, C.T. Supuran, L.S. King, W.F. Boron, G. Gros, Evidence that aquaporin 1 is a major pathway for CO2 transport across the human erythrocyte membrane, FASEB J., 20 (2006) 1974-1981. [31] N.L. Nakhoul, B.A. Davis, M.F. Romero, W.F. Boron, Effect of expressing the water channel aquaporin-1 on the CO2 permeability of Xenopus oocytes, Am. J. Physiol., 274 (1998) C543-C548. [32] M. Heckwolf, D. Pater, D.T. Hanson, R. Kaldenhoff, The Arabidopsis thaliana aquaporin AtPIP1;2 is a physiologically relevant CO2 transport facilitator, Plant J., 67 (2011) 795-804. [33] N. Uehlein, B. Otto, D.T. Hanson, M. Fischer, N. McDowell, R. Kaldenhoff, Function of Nicotiana tabacum aquaporins as chloroplast gas pores challenges the concept of membrane CO2 permeability, Plant Cell, 20 (2008) 648-657. [34] N. Uehlein, C. Lovisio, F. Siefritz, R. Kaldenhoff, The tobacco aquaporin NtAQP1 is a membrane CO2 pore with physiological functions, Nature, 425 (2003) 734-737. [35] J. Flexas, M. Ribas-Carbó, D.T. Hanson, J. Bota, B. Otto, J. Cifre, N. McDowell, H. Medrano, R. Kaldenhoff, Tobacco aquaporin NtAQP1 is involved in mesophyll conductance to CO2 in vivo, Plant J., 48 (2006) 427-439.

32
[36] Y.T. Hanba, M. Shibasaka, Y. Hayashi, T. Hayakawa, K. Kasamo, I. Terashima, M. Katsuhara, Overexpression of the barley aquaporin HvPIP2;1 increases internal CO2 conductance and CO2 assimilation in the leaves of transgenic rice plants, Plant Cell Physiol., 45 (2004) 521-529. [37] S.-I. Miyazawa, S. Yoshimura, Y. Shinzaki, M. Maeshima, C. Miyake, Relationship between mesophyll conductance to CO2 diffusion and contents of aquaporin localized at plasma membrane in tobacco plants grown under drought conditions, in: J.F. Allen, E. Gantt, J.H. Golbeck, B. Osmond (Eds.) Photosynthesis. Energy from the sun: 14th International Congress on Photosynthesis, Springer, Berlin, 2008a, pp. 805-808. [38] S.-I. Miyazawa, S. Yoshimura, Y. Shinzaki, M. Maeshima, C. Miyake, Deactivation of aquaporins decreases internal conductance to CO2 diffusion in tobacco leaves grown under long-term drought, Funct. Plant Biol., 35 (2008b) 553-564. [39] I. Terashima, K. Ono, Effects of HgCl2 on CO2 dependence of leaf photosynthesis: evidence indicating involvement of aquaporins in CO2 diffusion across the plasma membrane, Plant and Cell Physiol., 43 (2002) 70-78. [40] R. Aharon, Y. Shahak, S. Wininger, R. Bendov, Y. Kapulnik, G. Galili, Overexpression of a plasma membrane aquaporin in transgenic tobacco improves plant vigor under favorable growth conditions but not under drought or salt stress, Plant Cell, 15 (2003) 439-447. [41] U. Niinemets, A. Diaz-Espejo, J. Flexas, J. Galmes, C.R. Warren, Role of mesophyll diffusion conductance in constraining potential photosynthetic productivity in the field, Journal of Experimental Botany, 60 (2009a) 2249-2270. [42] U. Niinemets, A. Diaz-Espejo, J. Flexas, J. Galmes, C.R. Warren, Importance of mesophyll diffusion conductance in estimation of plant photosynthesis in the field, Journal of Experimental Botany, 60 (2009b) 2271-2282. [43] G. Grassi, F. Magnani, Stomatal, mesophyll conductance and biochemical limitations to photosynthesis as affected by drought and leaf ontogeny in ash and oak trees, Plant Cell Environ., 28 (2005) 834-849. [44] Ü. Niinemets, J.D. Tenhunen, N.R. Canta, M.M. Chaves, T. Faria, J.S. Pereira, J.F. Reynolds, Interactive effects of nitrogen and phosphorus on the acclimation potential of foliage photosynthetic properties of cork oak, Quercus suber, to elevated atmospheric CO2 concentrations, Global Change Biology, 5 (1999) 455-470. [45] Ü. Niinemets, J.D. Tenhunen, A model separating leaf structural and physiological effects on carbon gain along light gradients for the shade-tolerant species Acer Saccharum., Plant Cell Environ., 20 (1997) 845-866. [46] J.S. Hub, B.L. de Groot, Does CO2 permeate through aquaporin-1?, Biophysical Journal, 91 (2006) 842-848. [47] J.S. Hub, B.L. de Groot, Mechanism of selectivity in aquaporins and aquaglyceroporins, Proceedings of the National Academy of Sciences of the United States of America, 105 (2008) 1198-1203. [48] J.S. Hub, H. Grubmüller, B.L. Groot, E. Beitz, Dynamics and Energetics of Permeation Through Aquaporins. What Do We Learn from Molecular Dynamics Simulations?, in: Handbook of Experimental Pharmacology, Springer Berlin Heidelberg, 2009, pp. 57-76. [49] C. Maurel, Plant aquaporins: Novel functions and regulation properties, FEBS Lett., 581 (2007) 2227-2236. [50] C. Maurel, L. Verdoucq, D.-T. Luu, V. Santoni, Plant aquaporins: membrane channels with multiple integrated functions, Annu. Rev. Plant Biol., 59 (2008) 595-624. [51] B. Otto, N. Uehlein, S. Sdorra, M. Fischer, M. Ayaz, X. Belastegui-Macadam, M. Heckwolf, M. Lachnit, N. Pede, N. Priem, A. Reinhard, S. Siegfart, M. Urban, R. Kaldenhoff, Aquaporin tetramer composition modifies the function of tobacco aquaporins, J. Biol. Chem., 285 (2010) 31253-31260.

33
[52] N. Uehlein, H. Sperling, M. Heckwolf, R. Kaldenhoff, The Arabidopsis aquaporin PIP1;2 rules cellular CO2 uptake., Plant Cell Environ., 35 (2012) 1077-1083. [53] A. Laisk, V. Oja, M. Rahi, Diffusion resistances as related to the leaf anatomy, Fiziol. Rast., 17 (1970) 40-48. [54] P.S. Nobel, Physicochemical and environmental plant physiology, 4th ed., Academic Press, Inc., San Diego - New York - Boston - London - Sydney - Tokyo - Toronto, 1991. [55] P.S. Nobel, L.T. Zaragoza, W.K. Smith, Relation between mesophyll surface area, photosynthetic rate, and illumination level during development for leaves of Plectranthus parviflorus Henckel, Plant Physiol., 55 (1975) 1067-1070. [56] M. Rahi, Ob anatomicheskih parametrah lista v svyazi s diffuzionnymi soprotivleniyami. (The characteristics of leaf anatomy and diffusion resistances), P. Acad. Sci. Estonian SSR. B., 20 (1971) 85-94. [57] P.S. Nobel, L.T. Zaragoza, W.K. Smith, Relation between mesophyll surface area, photosynthetic rate, and illumination level during development for leaves of Plectranthus parviflorus Henckel, Plant Physiol., 55 (1975) 1067-1070. [58] A. Laisk, Kinetika fotosinteza i fotodyhaniya C3-rastenii. (Kinetics of photosynthesis and photorespiration in C3-plants), Nauka, Moscow, 1977. [59] R. Oguchi, K. Hikosaka, T. Hirose, Leaf anatomy as a constraint for photosynthetic acclimation: differential responses in leaf anatomy to increasing growth irradiance among three deciduous trees, Plant Cell Environ., 28 (2005) 916-927. [60] I. Terashima, T. Araya, S.-I. Miyazawa, K. Sone, S. Yano, Construction and maintenance of the optimal photosynthetic systems of the leaf, herbaceous plant and tree: an eco-developmental treatise, Ann. Bot.-London, 95 (2005) 507-519. [61] I. Terashima, Y.T. Hanba, Y. Tazoe, P. Vyas, S. Yano, Irradiance and phenotype: comparative eco-development of sun and shade leaves in relation to photosynthetic CO2 diffusion, J. Exp. Bot., 57 (2006) 343-354. [62] J.J. Peguero-Pina, J. Flexas, J. Galmés, D. Sancho-Knapik, G. Barredo, D. Villarroya, E. Gil-Pelegrín, Leaf anatomical properties in relation to differences in mesophyll conductance to CO2 and photosynthesis in two related Mediterranean Abies species, under review. [63] Ü. Niinemets, I.J. Wright, J.R. Evans, Leaf mesophyll diffusion conductance in 35 Australian sclerophylls covering a broad range of foliage structural and physiological variation, J. Exp. Bot., 60 (2009c) 2433-2449. [64] F. Hassiotou, M. Renton, M. Ludwig, J.R. Evans, E.J. Veneklaas, Photosynthesis at an extreme end of the leaf trait spectrum: how does it relate to high leaf dry mass per area and associated structural parameters?, J. Exp. Bot., 61 (2010) 3015-3028. [65] R. Oguchi, K. Hikosaka, T. Hiura, T. Hirose, Leaf anatomy and light acclimation in woody seedlings after gap formation in a cool-temperate forest, Oecologia, 149 (2006) 571-582. [66] J. Flexas, M. Baron, J. Bota, J.-M. Ducruet, A. Galle, J. Galmes, M. Jimenez, A. Pou, M. Ribas-Carbo, C. Sajnani, M. Tomas, H. Medrano, Photosynthesis limitations during water stress acclimation and recovery in the drought-adapted Vitis hybrid Richter-110 (V. berlandierixV. rupestris), J. Exp. Bot., 60 (2009) 2361-2377. [67] A. Galle, I. Florez-Sarasa, M. Tomas, A. Pou, H. Medrano, M. Ribas-Carbo, J. Flexas, The role of mesophyll conductance during water stress and recovery in tobacco (Nicotiana sylvestris): acclimation or limitation?, J. Exp. Bot., 60 (2009) 2379-2390. [68] L.X. Hu, Z.L. Wang, B.R. Huang, Diffusion limitations and metabolic factors associated with inhibition and recovery of photosynthesis from drought stress in a C-3 perennial grass species, Physiol. Plantarum, 139 (2010) 93-106. [69] C.R. Warren, I. Aranda, F.J. Cano, Responses to water stress of gas exchange and metabolites in Eucalyptus and Acacia spp., Plant Cell Environ., 34 (2011) 1609-1629.

34
[70] J.A. Bunce, Use of the response of photosynthesis to oxygen to estimate mesophyll conductance to carbon dioxide in water-stressed soybean leaves, Plant Cell Environ., 32 (2009) 875-881. [71] A. Galle, I. Florez-Sarasa, H.E. Aououad, J. Flexas, The Mediterranean evergreen Quercus ilex and the semi-deciduous Cistus albidus differ in their leaf gas exchange regulation and acclimation to repeated drought and re-watering cycles, J. Exp. Bot., (2011). [72] M. Centritto, M. Lauteri, M.C. Monteverdi, R. Serraj, Leaf gas exchange, carbon isotope discrimination, and grain yield in contrasting rice genotypes subjected to water deficits during the reproductive stage, J. Exp. Bot., 60 (2009) 2325-2339. [73] J. Gulias, J. Cifre, S. Jonasson, H. Medrano, J. Flexas, Seasonal and inter-annual variations of gas exchange in thirteen woody species along a climatic gradient in the Mediterranean island of Mallorca, Flora, 204 (2009) 169-181. [74] L. Varone, M. Ribas-Carbo, C. Cardona, A. Galle, H. Medrano, L. Gratani, J. Flexas, Stomatal and non-stomatal limitations to photosynthesis in seedlings and saplings of Mediterranean species pre-conditioned and aged in nurseries: Different response to water stress, Environ. Exp. Bot., 75 (2012) 235-247. [75] V. Resco, B.E. Ewers, W. Sun, T.E. Huxman, J.F. Weltzin, D.G. Williams, Drought-induced hydraulic limitations constrain leaf gas exchange recovery after precipitation pulses in the C3 woody legume, Prosopis velutina, New Phytol., 181 (2009) 672-682. [76] H.E. Bown, M.S. Watt, E.G. Mason, P.W. Clinton, D. Whitehead, The influence of nitrogen and phosphorus supply and genotype on mesophyll conductance limitations to photosynthesis in Pinus radiata, Tree Physiology, 29 (2009) 1143-1151. [77] Y. Li, Y.X. Gao, X.M. Xu, Q.R. Shen, S.W. Guo, Light-saturated photosynthetic rate in high-nitrogen rice (Oryza sativa L.) leaves is related to chloroplastic CO2 concentration, Journal of Experimental Botany, 60 (2009) 2351-2360. [78] W. Yamori, T. Nagai, A. Makino, The rate-limiting step for CO2 assimilation at different temperatures is influenced by the leaf nitrogen content in several C3 crop species, Plant, Cell & Environment, 34 (2011) 764-777. [79] Q. Han, Height-related decreases in mesophyll conductance, leaf photosynthesis and compensating adjustments associated with leaf nitrogen concentrations in Pinus densiflora, Tree Physiol., 31 (2011) 976-984. [80] D.R. Woodruff, F.C. Meinzer, B. Lachenbruch, D.M. Johnson, Coordination of leaf structure and gas exchange along a height gradient in a tall conifer, Tree Physiol., 29 (2009) 261-272. [81] D. Whitehead, M.M. Barbour, K.L. Griffin, M.H. Turnbull, D.T. Tissue, Effects of leaf age and tree size on stomatal and mesophyll limitations to photosynthesis in mountain beech (Nothofagus solandrii var. cliffortiodes), Tree Physiol., 31 (2011) 985-996. [82] S.H. Jin, J.Q. Huang, X.Q. Li, B.S. Zheng, J.S. Wu, Z.J. Wang, G.H. Liu, M. Chen, Effects of potassium supply on limitations of photosynthesis by mesophyll diffusion conductance in Carya cathayensis, Tree Physiol., 31 (2011) 1142-1151. [83] H.E. Bown, M.S. Watt, E.G. Mason, P.W. Clinton, D. Whitehead, The influence of nitrogen and phosphorus supply and genotype on mesophyll conductance limitations to photosynthesis in Pinus radiata, Tree Physiol., 29 (2009) 1143-1151. [84] R. Sagardoy, S. Vázquez, I.D. Florez-Sarasa, A. Albacete, M. Ribas-Carbó, J. Flexas, J. Abadía, F. Morales, Stomatal and mesophyll conductances to CO2 are the main limitations to photosynthesis in sugar beet (Beta vulgaris) plants grown with excess zinc, New Phytol., 187 (2010) 145-158. [85] V. Velikova, T. Tsonev, F. Loreto, M. Centritto, Changes in photosynthesis, mesophyll conductance to CO2, and isoprenoid emissions in Populus nigra plants exposed to excess nickel, Environ. Pollut., 159 (2011) 1058-1066.

35
[86] C.J. Bernacchi, P.B. Morgan, D.R. Ort, S.P. Long, The growth of soybean under free air CO2 enrichment (FACE) stimulates photosynthesis while decreasing in vivo Rubisco capacity, Planta, 220 (2005) 434-446. [87] E.L. Singsaas, D.R. Ort, E.H. Delucia, Elevated CO2 effects on mesophyll conductance and its consequences for interpreting photosynthetic physiology, Plant Cell Environ., 27 (2003) 41-50. [88] J.A. Bunce, Variable responses of mesophyll conductance to substomatal carbon dioxide concentration in common bean and soybean, Photosynthetica, 48 (2010) 507-512. [89] C. Douthe, E. Dreyer, D. Epron, C.R. Warren, Mesophyll conductance to CO2, assessed from online TDL-AS records of 13CO2 discrimination, displays small but significant short-term responses to CO2 and irradiance in Eucalyptus seedlings, J. Exp. Bot., 62 (2011) 5335-5346. [90] J. Flexas, A. Diaz-Espejo, J. Galmes, R. Kaldenhoff, H. Medrano, M. Ribas-Carbo, Rapid variations of mesophyll conductance in response to changes in CO2 concentration around leaves, Plant Cell Environ., 30 (2007) 1284-1298. [91] F. Hassiotou, M. Ludwig, M. Renton, E.J. Veneklaas, J.R. Evans, Influence of leaf dry mass per area, CO2, and irradiance on mesophyll conductance in sclerophylls, J. Exp. Bot., 60 (2009) 2303-2314. [92] Y. Tazoe, S. Von Caemmerer, G.M. Estavillo, J.R. Evans, Using tunable diode laser spectroscopy to measure carbon isotope discrimination and mesophyll conductance to CO2 diffusion dynamically at different CO2 concentrations, Plant Cell Environ., 34 (2011) 580-591. [93] V. Velikova, T. Tsonev, C. Barta, M. Centritto, D. Koleva, M. Stefanova, M. Busheva, F. Loreto, BVOC emissions, photosynthetic characteristics and changes in chloroplast ultrastructure of Platanus orientalis L. exposed to elevated CO2 and high temperature, Environ. Pollut., 157 (2009) 2629-2637. [94] D. Vrabl, M. Vaskov, M. Hronkova, J. Flexas, J. ŠantrČek, Mesophyll conductance to CO2 transport estimated by two independent methods: Effect of variable CO2 concentration and abscisic acid, J. Exp. Bot., 60 (2009) 2315-2323. [95] Y. Tazoe, S. Von Caemmerer, M.R. Badger, J.R. Evans, Light and CO2 do not affect the mesophyll conductance to CO2 diffusion in wheat leaves, J. Exp. Bot., 60 (2009) 2291-2301. [96] H. During, Stomatal and mesophyll conductances control CO2 transfer to chloroplasts in leaves of grapevine (Vitis vinifera L.), Vitis, 42 (2003) 65-68. [97] F. Loreto, P.C. Harley, G.D. Marco, T.D. Sharkey, Estimation of mesophyll conductance to CO2 flux by three different methods, Plant Physiol., 98 (1992) 1437-1443. [98] D. Tholen, X.G. Zhu, The mechanistic basis of internal conductance: A theoretical analysis of mesophyll cell photosynthesis and CO2 diffusion, Plant Physiol., 156 (2011) 90-105. [99] C. Douthe, Relationship between leaf gas exchange and 13C discrimination during photosynthesis: Estimations and rapid variations of mesophyll conductance to CO2, PhD Disseratation, (2011). [100] H.G. Jones, Moderate-term water stresses and associated changes in some photosynthetic parameters in cotton., New Phytol., 72 (1973) 1095-1105. [101] J. Galmes, H. Medrano, J. Flexas, Photosynthetic limitations in response to water stress and recovery in Mediterranean plants with different growth forms, New Phytol., 175 (2007) 81-93. [102] A. Diaz-Espejo, E. Nicolas, J.E. Fernandez, Seasonal evolution of diffusional limitations and photosynthetic capacity in olive under drought, Plant Cell Environ., 30 (2007) 922-933. [103] G. Grassi, E. Vicinelli, F. Ponti, L. Cantoni, F. Magnani, Seasonal and interannual variability of photosynthetic capacity in relation to leaf nitrogen in a deciduous forest plantation in northern Italy, Tree Physiol., 25 (2005) 349-360. [104] J.-M. Limousin, L. Misson, A.-V. Lavoir, N.K. Martin, S. Rambal, Do photosynthetic limitations of evergreen Quercus ilex leaves change with long-term increased drought severity?, Plant Cell Environ., 33 (2010) 863-875.

36
[105] A. Perez-Martin, J. Flexas, M. Ribas-Carbo, J. Bota, M. Tomas, J.M. Infante, A. Diaz-Espejo, Interactive effects of soil water deficit and air vapour pressure deficit on mesophyll conductance to CO2 in Vitis vinifera and Olea europaea, J. Exp. Bot., 60 (2009) 2391-2405. [106] G. Egea, A. Verhoef, P.L. Vidale, Towards an improved and more flexible representation of water stress in coupled photosynthesis-stomatal conductance models, Agricultural and Forest Meteorology, 151 (2011) 1370-1384. [107] L. Misson, J.-M. Limousin, R. Rodriguez, M.G. Letts, Leaf physiological responses to extreme droughts in Mediterranean Quercus ilex forest, Plant, Cell & Environment, 33 (2010) 1898-1910. [108] G. Egea, A. Verhoef, P.L. Vidale, Towards an improved and more flexible representation of water stress in coupled photosynthesis-stomatal conductance models, Agr. Forest Meteorol., 151 (2011) 1370-1384. [109] S.B. Zhang, Temperature acclimation of photosynthesis in Meconopsis horridula var. racemosa Prain, Chinese Source, 51 (2010) 457-464. [110] C.R. Warren, G.J. Ethier, N.J. Livingston, N.J. Grant, D.H. Turpin, D.L. Harrison, T.A. Black, Transfer conductance in second growth Douglas-fir (Pseudotsuga menziesii (Mirb.)Franco) canopies, Plant Cell Environ., 26 (2003) 1215-1227. [111] Ü. Niinemets, A. Cescatti, M. Rodeghiero, T. Tosens, Complex adjustments of photosynthetic capacity and internal mesophyll conductance to current and previous light availabilities and leaf age in Mediterranean evergreen species Quercus ilex, Plant Cell Environ., 29 (2006) 1159-1178. [112] P. Montpied, A. Granier, E. Dreyer, Seasonal time-course of gradients of photosynthetic capacity and mesophyll conductance to CO2 across a beech (Fagus sylvatica L.) canopy, J. Exp. Bot., 60 (2009) 2407-2418. [113] L.P. Mullin, S.C. Sillett, G.W. Koch, K.P. Tu, M.E. Antoine, Physiological consequences of height-related morphological variation in Sequoia sempervirens foliage, Tree Physiol., 29 (2009) 999-1010. [114] Ü. Niinemets, Stomatal conductance alone does not explain the decline in foliar photosynthetic rates with increasing tree age and size in Picea abies and Pinus sylvestris, Tree Physiol., 22 (2002) 515-535. [115] M.G. Ryan, B.J. Yoder, Hydraulic limits to tree height and tree growth. What keeps trees from growing beyond a certain height?, BioScience, 47 (1997) 235-242. [116] K. Steppe, Ü. Niinemets, R.O. Teskey, Tree size- and age-related changes in leaf physiology and their influence on carbon gain, in: F.C. Meinzer, T. Dawson, B. Lachenbruch (Eds.) Size- and age-related changes in tree structure and function, Springer, Berlin, 2011, pp. 235-253. [117] B.J. Yoder, M.G. Ryan, R.H. Waring, A.W. Schoettle, M.R. Kaufmann, Evidence of reduced photosynthetic rates in old trees, Forest Sci., 40 (1994) 513-527. [118] O. Kull, A. Koppel, Net photosynthetic response to light intensity of shoots from different crown positions and age in Picea abies (L.) Karst, Scand. J. Forest Res., 2 (1987) 157-166. [119] Ü. Niinemets, Distribution patterns of foliar carbon and nitrogen as affected by tree dimensions and relative light conditions in the canopy of Picea abies, Trees: Struct. Funct., 11 (1997) 144-154. [120] D.R. Woodruff, F.C. Meinzer, B. Lachenbruch, Height-related trends in leaf xylem anatomy and shoot hydraulic characteristics in a tall conifer: Safety versus efficiency in water transport, New Phytol., 180 (2008) 90-99. [121] M. Williams, E.B. Rastetter, D.N. Fernandes, M.L. Goulden, S.C. Wofsy, G.R. Shaver, J.M. Melillo, J.W. Munger, S.M. Fan, K.J. Nadelhoffer, Modelling the soil-plant-atmosphere continuum in a Quercus–Acer stand at Harvard Forest: the regulation of stomatal conductance by light, nitrogen and soil/plant hydraulic properties, Plant Cell Environ., 19 (1996) 911-927.

37
[122] D. Epron, D. Godard, G. Cornic, B. Genty, Limitation of net CO2 assimilation rate by internal resistances to CO2 transfer in the leaves of two tree species (Fagus sylvatica L. and Castanea sativa Mill.), Plant Cell Environ., 18 (1995) 43-51. [123] A. Ohsumi, T. Kanemura, K. Homma, T. Horie, T. Shiraiwa, Genotypic variation of stomatal conductance in relation to stomatal density and length in rice (Oryza sativa L.), Plant Prod. Sci., 10 (2007) 322-328. [124] T. Cai, L.B. Flanagan, R.S. Jassal, T.A. Black, Modelling environmental controls on ecosystem photosynthesis and the carbon isotope composition of ecosystem-respired CO2 in a coastal Douglas-fir forest, Plant Cell Environ., 31 (2008) 435-453. [125] T. Keenan, S. Sabate, C. Gracia, The importance of mesophyll conductance in regulating forest ecosystem productivity during drought periods, Glob. Change Biol., 16 (2010) 1019-1034. [126] C.P. Bickford, D.T. Hanson, N.G. McDowell, Influence of diurnal variation in mesophyll conductance on modelled 13C discrimination: Results from a field study, J. Exp. Bot., 61 (2010) 3223-3233. [127] G.D. Farquhar, L.A. Cernusak, Ternary effects on the gas exchange of isotopologues of carbon dioxide., Plant Cell Environ., (2012) doi: 10.1111/j.1365-3040.2012.02484.x [128] R.J. Oliver, G. Taylor, J.W. Finch, Assessing the impact of internal conductance to CO2 in a land-surface scheme: Measurement and modeling of photosynthesis in Populus nigra. , Agr. Forest Meteorol., 152 (2012) 240-251. [129] N. Ubierna, J.D. Marshall, Estimation of canopy average mesophyll conductance using �13C of phloem contents. , Plant Cell Environ., 34 (2011) 1521-1535. [130] L. Sack, C.M. Streeter, N.M. Holbrook, Hydraulic analysis of water flow through leaves of sugar maple and red oak, Plant Physiol., 134 (2004) 1824-1833. [131] E. Steudle, J. Frensch, Water transport in plants: Role of the apoplast, Plant Soil, 187 (1996) 67-79. [132] N. Kodama, A. Cousins, K.P. Tu, M.M. Barbour, Spatial variation in photosynthetic CO2 carbon and oxygen isotope discrimination along leaves of the monocot triticale (Triticum×Secale) relates to mesophyll conductance and the Peclet effect, Plant Cell Environ., 34 (2011) 1548-1562. [133] B. Duan, Y. Li, X. Zhang, H. Korpelainen, C. Li, Water deficit affects mesophyll limitation of leaves more strongly in sun than in shade in two contrasting Picea asperata populations, Tree Physiol., 29 (2009) 1551-1561. [134] R.Y. Soolanayakanahally, R.D. Guy, S.N. Silim, E.C. Drewes, W.R. Schroeder, Enhanced assimilation rate and water use efficiency with latitude through increased photosynthetic capacity and internal conductance in balsam poplar (Populus balsamifera L.), Plant Cell Environ., 32 (2009) 1821-1832. [135] J. Flexas, J. Galmes, A. Galle, J. Gulias, A. Pou, M. Ribas-Carbo, M. Tomas, H. Medrano, Improving water use efficiency in grapevines: Potential physiological targets for biotechnological improvement, Aust. J. Grape Wine R., 16 (2010) 106-121. [136] J. Galmes, M.A.N. Conesa, J.M. Ochogavia, J.A. Perdomo, D.M. Francis, M. Ribas-Carbo, R. Save, J. Flexas, H. Medrano, J. Cifre, Physiological and morphological adaptations in relation to water use efficiency in Mediterranean accessions of Solanum lycopersicum, Plant, Cell & Environment, 34 (2011) 245-260. [137] L. Sack, N.M. Holbrook, Leaf hydraulics, in: Annu. Rev. Plant Biol., 2006, pp. 361-381. [138] K. Aasamaa, A. Sober, Hydraulic conductance and stomatal sensitivity to changes of leaf water status in six deciduous tree species, Biol. Plantarum., 44 (2001) 65-73. [139] T.J. Brodribb, T.S. Feild, G.J. Jordan, Leaf maximum photosynthetic rate and venation are linked by hydraulics, Plant Physiol., 144 (2007) 1890-1898.

38
[140] Y.T. Hanba, S.I. Miyazawa, I. Terashima, The influence of leaf thickness on the CO2 transfer conductance and leaf stable carbon isotope ratio for some evergreen tree species in Japanese warm-temperate forests, Funct. Ecol., 13 (1999) 632-639. [141] L. Sack, P.D. Cowan, N. Jaikumar, N.M. Holbrook, The 'hydrology' of leaves: Co-ordination of structure and function in temperate woody species, Plant Cell Environ., 26 (2003) 1343-1356. [142] L. Sack, K. Frole, Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees., Ecology, 87 (2006) 483-491. [143] S. von Caemmerer, J.R. Evans, Determination of the Average Partial Pressure of CO2 in Chloroplasts From Leaves of Several C3 Plants, Funct. Plant Biol., 18 (1991) 287-305. [144] J.A. Bunce, How do leaf hydraulics limit stomatal conductance at high water vapour pressure deficits?, Plant Cell Environ., 29 (2006) 1644-1650. [145] Y.A.N. Li, H.A.O. Xu, S. Cohen, Long-term hydraulic acclimation to soil texture and radiation load in cotton, Plant Cell Environ., 28 (2005) 492-499. [146] H. Cochard, A. Nardini, L. Coll, Hydraulic architecture of leaf blades: Where is the main resistance?, Plant Cell Environ., 27 (2004) 1257-1267. [147] K.A. Mott, Leaf hydraulic conductivity and stomatal responses to humidity in amphistomatous leaves, Plant Cell Environ., 30 (2007) 1444-1449. [148] J.P. Ferrio, M. Cuntz, C. Offermann, R. Siegwolf, M. Saurer, A. Gessler, Effect of water availability on leaf water isotopic enrichment in beech seedlings shows limitations of current fractionation models, Plant Cell Environ., 32 (2009) 1285-1296. [149] J.P. Ferrio, A. Pou, I. Florez-Sarasa, A. Gessler, N. Kodama, J. Flexas, M. Ribas-Carbo, The Peclet effect on leaf water enrichment correlates with leaf hydraulic conductance and mesophyll conductance for CO2, Plant Cell Environ., 35 (2012) 611-625. [150] A. Kahmen, K. Simonin, K. Tu, G.R. Goldsmith, T.E. Dawson, The influence of species and growing conditions on the 18O enrichment of leaf water and its impact on 'effective path length', New Phytol., 184 (2009) 619-630. [151] Y. Zhou, K. Grice, Y. Chikaraishi, H. Stuart-Williams, G.D. Farquhar, N. Ohkouchi, Temperature effect on leaf water deuterium enrichment and isotopic fractionation during leaf lipid biosynthesis: Results from controlled growth of C3 and C4 land plants, Phytochemistry, 72 (2011) 207-213. [152] H. Craig, L.I. Gordon, Deuterium and oxygen 18 variations in the ocean and marine atmosphere, Marine Geochemistry, Narragansett Marine Laboratory, University of Rhode Island,, 3 (1965) 277-374. [153] G. Dongmann, H.W. Nurnberg, H. Forstel, K. Wagener, On the enrichment of H2
18O in the leaves of transpiring plants, Radiation and Environmental Biophysics, 11 (1974) 41-52. [154] G.D. Farquhar, J. Lloyd, Carbon and oxygen isotope effects in the exchange of carbon dioxide between terrestrial plants and the atmosphere., In Stable Isotopes and Plant Carbon-Water Relations (eds Ehleringer J.R., Hall A.E. & Farquhar G.D.), Academic Press, San Diego., (1993) pp. 47 70. [155] M.M. Barbour, R.A. Fischer, K.D. Sayre, G.D. Farquhar, Oxygen isotope ratio of leaf and grain material correlates with stomatal conductance and grain yield in irrigated wheat, Funct. Plant. Biol., 27 (2000) 625-637. [156] M. Cuntz, J. Ogee, G.D. Farquhar, P. Peylin, L.A. Cernusak, Modelling advection and diffusion of water isotopologues in leaves, Plant Cell Environ., 30 (2007) 892-909. [157] M. Centritto, F. Loreto, K. Chartzoulakis, The use of low [CO2] to estimate diffusional and non-diffusional limitations of photosynthetic capacity of salt-stressed olive saplings, Plant Cell Environ., 26 (2003) 585-594. [158] R. Schaufele, J. Santrucek, H. Schnyder, Dynamic changes of canopy-scale mesophyll conductance to CO2 diffusion of sunflower as affected by CO2 concentration and abscisic acid, Plant Cell Environ., 34 (2011) 127-136.

39
[159] F.P. Gomes, M.A. Oliva, M.S. Mielke, A.A.F. de Almeida, H.G. Leite, L.A. Aquino, Photosynthetic limitations in leaves of young Brazilian Green Dwarf coconut (Cocos nucifera L. 'nana') palm under well-watered conditions or recovering from drought stress, Environ. Exp. Bot., 62 (2008) 195-204. [160] G. Grassi, F. Ripullone, M. Borghetti, S. Raddi, F. Magnani, Contribution of diffusional and non-diffusional limitations to midday depression of photosynthesis in arbutus unedo L, Trees - Struct. Funct., 23 (2009) 1149-1161.

40
Figure legends Figure 1. Mesophyll conductance is greatest the more a species is phylogenetically evolved. Average ± S.E. values for gs and gm in different pooled groups of plants. Data from liverworts and hornworts from [17], data for all other groups from [41]. Figure 2. Mesophyll conductance reflects the CO2 diffusion pathway, which is composed of air, water and lipid barriers. Comparison of CO2 diffusion pathway within the fully expanded needle of evergreen conifer Abies alba (a-c) and within the leaf of broad-leaved deciduous Populus tremula (d-f) according to light (a, b, d), scanning (c) and transmission electron (e, f) micrographs. Leaf cross-sections (a, d) illustrate the CO2 gas phase diffusion pathway from ambient air (Ca) to sub-stomatal cavities (Ci) and from sub-stomatal cavities to outer surface of cell walls, Ci,w. The CO2 concentration drawdown, Ca-Ci, depends on stomatal conductance, while the drawdown Ci-Ci,w is characterized by internal gas-phase diffusion conductance (gias) that is determined by effective mesophyll thickness and porosity. The micrographs of palisade tissue (b, e) demonstrate parallel diffusion pathways within cells (indicated with arrows for representative cells) that is determined by the exposure of chloroplasts to intercellular airspace. The representative micrographs of palisade cells (c, f) illustrate CO2 diffusion pathways in liquid phase from mesophyll cell outer surface to chloroplasts. The CO2 drawdown from outer surface of cell walls to chloroplasts (Cc, Ci,w-Cc) is determined by liquid-phase diffusion conductance gliq that consists of cell wall (cw), plasma membrane (pm, not visible in the micrographs), cytoplasm (cyt), chloroplast envelope (env, not visible in the micrographs), and chloroplast stroma (chl). Micrographs from Peguero-Pina et al. [62] (a-c) and Tosens et al. [15] (d-f), with permission. Figure 3. Altered expression of aquaporins in relation to mesophyll conductance limitations of photosynthesis. Average (error bars show +SE) net assimilation rate, stomatal conductance to CO2 and mesophyll diffusion conductance (upper panels) and relative photosynthetic limitation due to limited biochemical capacity, stomatal conductance and mesophyll diffusion conductance (lower panels) in transformed plants with reduced (AS, RNA-interference) and over-expressed (OE) aquaporins and corresponding controls (either wild type or plants transformed with the same construct used for AS and OE, but lacking the modified aquaporin expression phenotype). As the data represent one-to-one correspondence between data pairs (control vs. AS and control vs. OE and AS vs. OE within the given study), the averages between the treatments and corresponding controls and between AS and OE were compared by paired t-tests that is a more powerful statistical test than standard ANOVA [141]. Statistical significance as: ns – not significant, * - P<0.05, ** - P < 0.01, *** - P < 0.001. The data are for Arabidopsis thaliana aquaporins atpip1;2-1 and atpip2;3-1 [32], Nicotiana tabacum aquaporin NtAQP1 [33-35] and Hordeum vulgare aquaporin HvPIP2;1 [36]. The data correspond to saturating light, leaf temperature of 25 °C and chamber CO2 concentration of 280-400 µmol mol-1. The relative limitations of photosynthesis were calculated according to [43] and the three imitations sum up to 1. Maximum carboxylase activity of Rubisco, Vcmax, needed for these calculations was determined by inverse modeling as in [44]. Means between treatment and corresponding controls and between AS and OE were compared by paired t-tests (ns – not significant, * - P<0.05, ** - P < 0.01, *** - P < 0.001). Figure 4. Co-regulation of stomatal conductance, mesophyll conductance and photosynthesis in wild type and mutants with altered expression of aquaporins. Net assimilation rate (An) in relation to (a) stomatal conductance to CO2 (gs) and (b) mesophyll diffusion conductance (gm), and correlations between (c) the CO2 drawdown due to limited gs (difference of CO2 concentrations in ambient air and sub-stomatal cavities, Ca-Ci=An/gs) and (d) the CO2 drawdown due to limited gm

41
(difference in CO2 concentrations in sub-stomatal cavities and chloroplasts, Ci-Cc = An/gm). Every data point corresponds to an average value of either control, aquaporin overexpressed, aquaporin antisense or aquaporin-inhibited plants reported in original studies. Data sources for transformed plants as in Fig. 3. Data for HgCl2 treated (0.3-1.2 mM) and non-treated control plants were from [39] (Vicia faba) and [37, 38] (Nicotiana tabacum). Environmental conditions during measurements as in Fig. 3. The number of data points in different panels differs because not all studies reported the whole suite of characteristics An, gs, gm, Ci and Cc. Figure 5. Increasing CO2 concentration reduces mesophyll conductance. gm correlates with Ci over a broad range of ambient CO2 concentrations and across a number of different species and growth forms. The isotopic method (δ13C) and the combined chl fluorescence and infrared gas exchange analysis method are shown in closed and open circles, respectively. Data are taken or calculated from [86], [88], [157], [89], [99], [96], [90], [91], [97], [158], [87], [92], [93], [94], [11]. Figure 6. Mesophyll conductance limitations increase as photosynthesis declines (total limitation increase) in response to stress. Empty circles represent stomatal limitation (SCL), filled circles mesophyll conductance limitation (MCL) and triangles biochemical limitation (BL). Data have been compiled from the following references: [83] , [124], [102], [108], [66, 90], [67, 71], [101], [159], [160], [104], [107], [105], [84], [81] and [109].
Figure 7. Mesophyll conductance co-regulated with hydraulic conductance. A) Strong relationship between average leaf hydraulic conductance (Kleaf) and mesophyll diffusion conductance (gm) for different plant species, based on literature data (see Supplemental Information for the list of references). B) Comparative response of different species to changes in growing conditions affecting both gm and Kleaf: high/low water stress for Quercus, Fagus and Pinus; canopy height for Pseudotsuga; a combination of drought and vein severing for Vitis.
Figure 8. Mesophyll conductance and hydraulic conductance co-regulate with the effective pathlength of mesophyll water movement. Strong relationship between treatment averages for leaf lamina hydraulic conductance (Klamina), scaled effective pathlength (Leff) and mesophyll conductance for CO2 (gm) in Grenache grape. Treatment averages for gm were calculated excluding values above 200 mmol CO2 m
-2 s-1, which were beyond the linear range for the relationship between gm and Leff (see inset with individual leaf values). Circles: control plants, triangles: drought plants. Open symbols: intact leaves. Closed symbols: vein-severed leaves. Data from [149]. Figure 9. Altered expression of PIP 1 aquaporins results in co-variations of the mesophyll conductance and the effective pathlength of mesophyll water transfer. Upper panel: the relationship between gm and the path length of mesophyll water transfer (L) in wild-type (circles), anti-sense (downwards triangles) and over-expressing (up-wards triangles) tobacco plants with altered levels of NtAQP1. Empty dots are individual plants within each genotype, and filled dots average values per genotype. Lower panel: potential explanation for the opposite relationships found between gm and L within and between genotypes, based on the findings by Otto et al. [51]: negative correlations between gm and the path length of mesophyll water transfer occur between genotypes, likely reflecting that altered expression of one aquaporin class results in different proportions of aquaporin classes within tetramers; positive correlations between gm and the path length of mesophyll water transfer occur between individuals within each genotype, likely reflecting different concentrations of aquaporins rather than different proportions of aquaporin classes within tetramers.

42
FIGURE 1

43
FIGURE 2

44
FIGURE 3

45
FIGURE 4

46
FIGURE 5

47
relative TL
0,0 0,2 0,4 0,6 0,8 1,0
rela
tive
SC
L, M
CL,
BL
0,0
0,2
0,4
0,6
0,8
1,0
TL vs MCL TL vs SL TL vs BL
FIGURE 6

48
g m (
mm
ol C
O2
m-2
s-1
)
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Herbs monocotsHerbs dicots Deciduous angiosp. Ev. angiosp. Ev. gymnosp.
gm=0.0268+0.0156*Kleaf
r2=0.659 N=20 P<0.001
Kleaf (mmol H2O s-1 MPa-1 m-2)0 5 10 15 20 25
gm (
mm
ol C
O2
m-2
s-1)
0.0
0.5
1.0
1.5
Kleaf (mmol H2O s-1 MPa-1 m-2)0 5 10 15 20 25
g m (
mm
ol C
O2
m-2
s-1
)
0.0
0.1
0.2
0.3
0.4
QuercusFagusPinusPseudotsugaVitis
gm=-0.0036+0.0206*Kleaf
r2=0.594 N=14 P<0.001
A)
B)
FIGURE 7
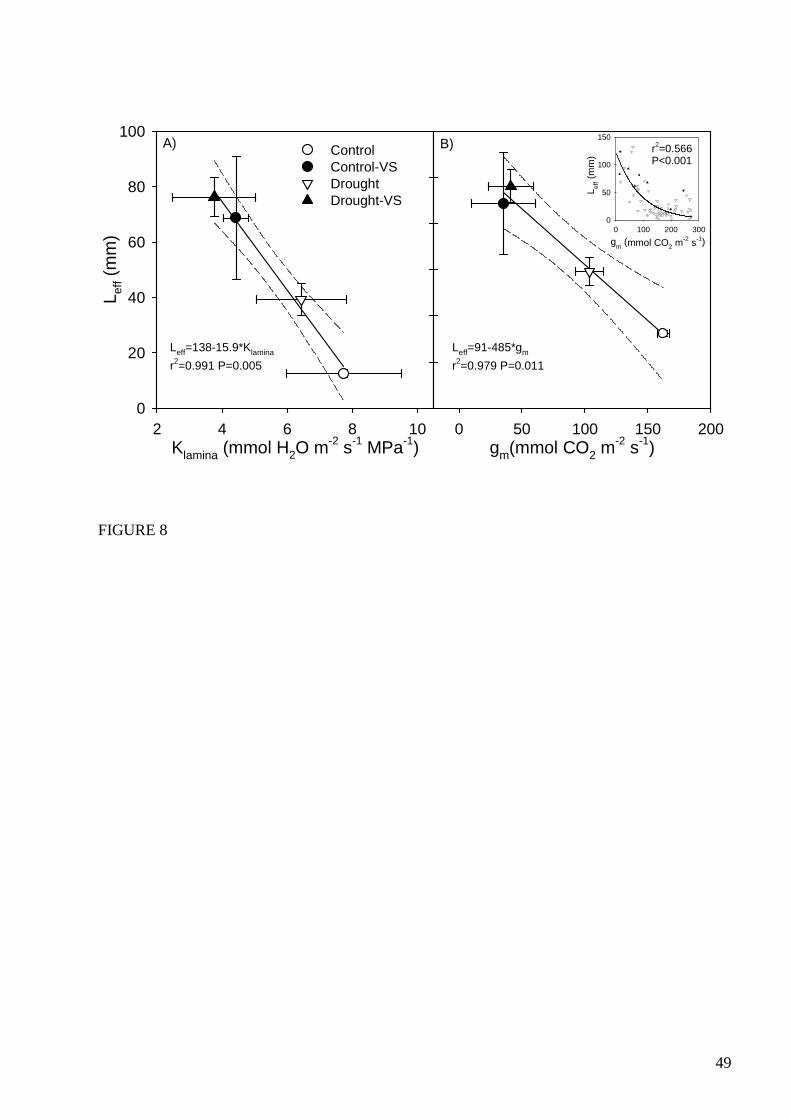
49
Klamina (mmol H2O m-2 s-1 MPa-1)2 4 6 8 10
L eff (
mm
)
0
20
40
60
80
100
gm(mmol CO2 m-2 s-1)
0 50 100 150 200
ControlControl-VSDroughtDrought-VS
Leff=91-485*gm
r2=0.979 P=0.011
A) B)
Leff=138-15.9*Klamina
r2=0.991 P=0.005
gm (mmol CO2 m
-2 s-1)0 100 200 300
L eff (
mm
)
0
50
100
150
r2=0.566P<0.001
FIGURE 8

50
AS WT OE
Plant 1 Plant 1 Plant 1
Plant 2 Plant 2 Plant 2
Increasing CO2 transport and declining water transport (high L)
Increasingboth
CO
2and
water
transport
FIGURE 9

Appendix A: Supplementary data Table S1. Average and standard deviation of leaf hydraulic conductance (Kleaf) and mesophyll diffusion conductance (gm) across the different genera included in Fig. 8. The “OUT” column indicates the values not included in the linear regression plotted in Fig. 8a.
Genus Kleaf
(mmol H2O s-1 MPa-1 m-2)
gm
(mmol CO2 m-2 s-1)
OUT References Kleaf References gm
Herbaceous monocots
Triticum 3.1 ±0.00 0.44 ±0.072 X [1],[2],[3] [4],[7],[5],[6],[8] Oryza 7.5 ±3.89 0.56 ±0.410 X [9] [10],[11],[8] Herbaceous dicots
Phaseolus 19.5 ±4.72 0.28 ±0.115 [12],[13] [14],[15],[16],[17],[18],[8],[19] Helianthus 13.2 ±3.31 0.79 ±0.545 X [12],[20],[21] [22],[23],[17],[24],[25] Vicia 15.7 ±0.00 0.40 ±0.060 [26] [6],[27] Glycine 11.2 ±0.00 0.49 ±0.165 X [28] [29],[30],[31] Arabidopsis 11.5 ±1.90 0.20 ±0.000 [32] [33],[34] Nicotiana 26.0 ±4.00 0.30 ±0.205 X [35] [36],[37],[33],[30],[38],[39],[31],[40],[41],[42],[8]Gossypium 12.2 ±1.10 1.06 ±0.745 X [28],[43] [22],[44] Woody deciduous angiosperms
Quercus 7.5 ±1.71 0.14 ±0.032 [45],[46],[48],[20],[47] [31],[49],[6],[50],[51],[52],[53],[54] Fagus 7.1 ±0.46 0.16 ±0.094 [46] [55],[19] _Acer 7.4 ±1.35 0.05 ±0.011 [56],[20] [58],[17],[57],[50] alnus 7.9 ±0.00 0.12 ±0.017 [59] [17],[57],[50] populus 10.6 ±2.20 0.22 ±0.059 [45] [60],[17],[57],[50],[51],[61],[52],[18] betula 11.0 ±3.00 0.77 ±0.335 X [47],[62] [63],[64],[65],[66] Tilia 7.0 ±0.10 0.92 ±0.415 X [45] [65],[24] Juglans 8.0 ±3.00 0.15 ±0.035 [67] Corylus 7.8 ±0.56 0.18 ±0.000 [20] [50] Castanea 4.4 ±0.83 0.11 ±0.000 [20] [22],[55],[68] Vitis 11.0 ±2.18 0.16 ±0.044 [1],[47] [69],[1],[70],[71],[15],[33],[72] Woody evergreen angiosperms
Quercus 3.0 ±1.44 0.08 ±0.013 [20] [73],[17],[74],[6],[75],[76],[52] Olea 8.9 ±0.28 0.22 ±0.135 [20] [77],[78],[79],[33],[80],[75] Camellia 6.0 ±0.00 0.10 ±0.025 [21] [17],[74] Hedera 10.4 ±0.26 0.15 ±0.000 [47] [6] eucalyptus 9.9 ±0.42 0.15 ±0.014 [81] [82],[83] [49],[6],[8],[84],[85],[86] Laurus 2.2 ±0.56 0.08 ±0.000 [20] [75] Woody evergreen gymnosperms
Pinus 5.3 ±1.12 0.13 ±0.020 [87],[48],[88] [14],[50],[89] Pseudotsuga 5.1 ±0.67 0.11 ±0.087 [48],[90] [14],[91],[50],[92],[90]

References Appendix A
1. J. P. Ferrio, A. Pou, I. Florez-Sarasa, A. Gessler, N. Kodama, J. Flexas, M. Ribas-Carbo, The Péclet effect on leaf water enrichment correlates with leaf hydraulic conductance and mesophyll conductance for CO2, Plant Cell Environ. 35 (2012) 611-625.
2. M. Gallardo, J. Eastham, P. J. Gregory, N. C. Turner, A comparison of plant hydraulic conductances in wheat and lupins, J. Exp. Bot. 47 (1996) 233-239.
3. P. J. Gregory, J. Eastham, Growth of shoots and roots, and interception of radiation by wheat and lupin crops on a shallow, duplex soil in response to time of sowing, Aust. J. Agric. Res. 47 (1996) 427-447.
4. J. R. Evans, L. Vellen, Wheat cultivars differ in transpiration efficiency and CO2 diffusion inside their leaves, Crop Research in Asia: Achievements and Perspective1996) 326-329.
5. F. Loreto, G. Di Marco, D. Tricoli, T. D. Sharkey, Measurements of mesophyll conductance, photosynthetic electron transport and alternative electron sinks of field grown wheat leaves, Photosynthesis Research 41 (1994) 397-403.
6. F. Loreto, P. C. Harley, G. d. Marco, T. D. Sharkey, G. Di-Marco, Estimation of mesophyll conductance to CO2 flux by three different methods, Plant Physiol. 98 (1992) 1437-1443.
7. N. Kodama, A. Cousins, K. P. Tu, M. M. Barbour, Spatial variation in photosynthetic CO2 carbon and oxygen isotope discrimination along leaves of the monocot triticale (Triticum ¥ Secale) relates to mesophyll, Plant Cell Environ 34, 1548â€"1562 (2011) 1548-1562.
8. S. Von Caemmerer, J. R. Evans, Determination of the average partial pressure of CO2 in chloroplasts from leaves of several C3 plants, Aust. J. Plant Physiol. 18 (1991) 287-305.
9. V. Stiller, H. R. Lafitte, J. S. Sperry, Embolized conduits of rice (Oryza sativa, Poaceae) refill despite negative xylem pressure. American Journal of Botany 92: 1970-1974., American Journal of Botany 92 (2005) 1970-1974.
10. Y. T. Hanba, M. Shibasaka, Y. Hayashi, T. Hayakawa, K. Kasamo, I. Terashima, M. Katsuhara, Overexpression of the Barley Aquaporin HvPIP2;1 Increases Internal CO2 Conductance and CO2 Assimilation in the Leaves of Transgenic Rice Plants, Plant Cell Physiol. 45 (2004) 521-529.
11. A. Scartazza, M. Lauteri, M. C. Guido, E. Brugnoli, Carbon isotope discrimination in leaf and stem sugars, water- use efficiency and mesophyll conductance during different developmental stages in rice subjected to drought, Aust. J. Plant Physiol. 25 (1998) 489-498.
12. A. Kahmen, K. Simonin, K. Tu, G. R. Goldsmith, T. E. Dawson, The influence of species and growing conditions on the 18-O enrichment of leaf water and its impact on 'effective path length', New Phytol. 184 (2009) 619-630.
13. A. Söber, Hydraulic conductance, stomatal conductance, and maximal photosynthetic rate in bean leaves, Photosynthetica 34 (1998) 599-603.
14. E. H. De Lucia, D. Whitehead, M. J. Clearwater, The relative limitation of photosynthesis by mesophyll conductance in co-occurring species in a temperate rainforest dominated by the conifer Dacrydium cupressinum, Functional Plant Biol. 30 (2003) 1197-1204.
15. J. Flexas, J. Bota, F. Loreto, G. Cornic, T. D. Sharkey, Diffusive and metabolic limitations to photosynthesis under drought and salinity in C-3 plants, Plant. Biol. 6 (2004) 269-279.
16. M. D. Flowers, E. L. Fiscus, K. O. Burkey, F. L. Booker, J. J. B. Dubois, Photosynthesis, chlorophyll fluorescence, and yield of snap bean (Phaseolus vulgaris L.) genotypes differing in sensitivity to ozone, Environmental and Experimental Botany 61 (2007) 190-198.
17. Y. T. Hanba, H. Kogami, I. Terashima, The effect of internal CO2 conductance on leaf carbon isotope ratio, Isotopes in Environmental and Health Studies 39 (2003) 5-13.
18. E. L. Singsaas, D. R. Ort, E. H. DeLucia, Elevated CO2 effects on mesophyll conductance and its consequences for interpreting photosynthetic physiology, Plant Cell Environ. 27 (2004) 41-50.
19. C. R. Warren, M. Löw, R. Matyssek, M. Tausz, Internal conductance to CO2 transfer of adult Fagus sylvatica: Variation between sun and shade leaves and due to free-air ozone fumigation, Environmental and Experimental Botany 59 (2007) 130-138.

20. A. Nardini, Are Sclerophylls and Malacophylls Hydraulically Different?, Biologia Plantarum 44 (2001) 239-245.
21. C. Scoffoni, A. D. McKown, M. Rawls, L. Sack, Dynamics of leaf hydraulic conductance with water status: quantification and analysis of species differences under steady state, J. Exp. Bot. 2011)
22. E. Brugnoli, A. Scartazza, M. Lauteri, M. C. Monteverdi, C. Maguas, Carbon isotope discrimination in structural and non-structural carbohydrates in relation to productivity and adaptation to unfavourable conditions, Stable Isotopes: Integration of Biological, Ecological and Geochemical Processes1998) 133-146.
23. H. Eichelmann, A. Laisk, Ribulose-1,5-bisphosphate carboxylase/oxygenase content, assimilatory charge, and mesophyll conductance in leaves, Plant Physiol. 119 (1999) 179-189.
24. A. Laisk, F. Loreto, Determining photosynthetic parameters from leaf CO2 exchange and chlorophyll fluorescence1: Ribulose-1,5-bisphosphate carboxylase/oxygenase specificity factor, dark respiration in the light, excitation distribution between photosystems, alternative electron transport rate, and mesophyll diffusion resistance, Plant Physiol. 110 (1996) 903-912.
25. A. Laisk, A. Sumberg, Partitioning of the leaf CO2 exchange into components using CO2 exchange and fluorescence measurements, Plant Physiol. 106 (1994) 689-695.
26. K. A. Mott, Leaf hydraulic conductivity and stomatal responses to humidity in amphistomatous leaves, Plant Cell Environ 30 (2007) 1444-1449.
27. I. Terashima, K. Ono, Effects of HgCl2 on CO2 Dependence of Leaf Photosynthesis: Evidence Indicating Involvement of Aquaporins in CO2 Diffusion across the Plasma Membrane, Plant Cell Physiol. 43 (2002) 70-78.
28. J. A. Bunce, How do leaf hydraulics limit stomatal conductance at high water vapour pressure deficits?, Plant Cell Environ 29 (2006) 1644-1650.
29. C. J. Bernacchi, P. B. Morgan, D. R. Ort, S. P. Long, The growth of soybean under free air [CO <sub>2</sub>] enrichment (FACE) stimulates photosynthesis while decreasing in vivo Rubisco capacity, Planta 220 (2005) 434-446.
30. J. Flexas, M. Ribas-Carbo, J. Bota, J. Galmes, M. Henkle, S. Martinez-Canellas, H. Medrano, Decreased Rubisco activity during water stress is not induced by decreased relative water content but related to conditions of low stomatal conductance and chloroplast CO2 concentration, New Phytol. 172 (2006) 73-82.
31. J. S. Gillon, D. Yakir, Internal conductance to CO2 diffusion and C18OO discrimination in C3 leaves, Plant Physiol. 123 (2000) 201-213.
32. P. Martre, R. Morillon, F. Barrieu, G. B. North, P. S. Nobel, M. J. Chrispeels, Plasma Membrane Aquaporins Play a Significant Role during Recovery from Water Deficit, Plant Physiol. 130 (2002) 2101-2110.
33. J. Flexas, A. Diaz-Espejo, J. Galmes, R. Kaldenhoff, H. Medrano, M. Ribas-Carbo, Rapid variations of mesophyll conductance in response to changes in CO2 concentration around leaves, Plant. Cell. Environ. 30 (2007) 1284-1298.
34. J. Flexas, M. F. Ortuño, M. Ribas-Carbo, A. Diaz-Espejo, I. D. Florez-Sarasa, H. Medrano, Mesophyll conductance to CO2 in Arabidopsis thaliana, New Phytol. 175 (2007) 501-511.
35. F. Siefritz, M. T. Tyree, C. Lovisolo, A. Schubert, R. Kaldenhoff, PIP1 plasma membrane aquaporins in tobacco: from cellular effects to function in plants, Plant Cell 14 (2002) 869-876.
36. C. J. Bernacchi, A. R. Portis, H. Nakano, S. Von-Caemmerer, S. P. Long, Temperature response of mesophyll conductance. Implications for the determination of Rubisco enzyme kinetics and for limitations to photosynthesis in vivo, Plant Physiol. 130 (2002) 1992-1998.
37. J. R. Evans, S. Von Caemmerer, B. A. Setchell, G. S. Hudson, The relationship between CO2 transfer conductance and leaf anatomy in transgenic tobacco with a reduced content of Rubisco, Aust. J. Plant Physiol. 21 (1994) 475-495.
38. J. Flexas, M. Ribas-Carbo, D. T. Hanson, J. Bota, B. Otto, J. Cifre, N. McDowell, H. Medrano, R. Kaldenhoff, Tobacco aquaporin NtAQP1 is involved in mesophyll conductance to CO2 in vivo, The Plant Journal 48 (2006) 427-439.
39. J. Galmes, H. Medrano, J. Flexas, Acclimation of Rubisco specificity factor to drought in tobacco: Discrepancies between in vitro and in vivo estimations, J. Exp. Bot. 57 (2006) 3659-3667.

40. M. Peisker, H. Apel, Inhibition by light of CO2 evolution from dark respiration: Comparison of two gas exchange methods, Photosynthesis Research 70 (2001) 291-298.
41. G. D. Price, S. Von Caemmerer, J. R. Evans, J. W. Yu, J. Lloyd, V. Oja, P. Kell, K. Harrison, A. Gallagher, M. R. Badger, Specific reduction of chloroplast carbonic anhydrase activity by antisense RNA in transgenic tobacco plants has a minor effect on photosynthetic CO2 assimilation, Planta 193 (1994) 331-340.
42. T. D. Sharkey, T. L. Vassey, P. J. Vanderveer, R. D. Vierstra, Carbon metabolism enzymes and photosynthesis in transgenic tobacco (Nicotiana tabacum L.) having excess phytochrome, Planta 185 (1991) 287-296.
43. Y. Li, H. Xu, S. Cohen, Long-term hydraulic acclimation to soil texture and radiation load in cotton, Plant Cell Environ. 28 (2005) 492-499.
44. S. Ennahli, H. J. Earl, Physiological limitations to photosynthetic carbon assimilation in cotton under water stress, Crop Sci. 45 (2005) 2374-2382.
45. K. Aasamaa, A. Söber, Sensitivity of stem and petiole hydraulic conductance of deciduous trees to xylem sap ion concentration, Biologia Plantarum 54 (2010) 299-307.
46. I. Aranda, L. Gil, J. A. Pardos, Seasonal changes in apparent hydraulic conductance and their implications for water use of European beech (Fagus sylvatica L.) and sessile oak [Quercus petraea (Matt.) Liebl] in South Europe, Plant Ecology 179 (2005) 155-167.
47. L. Sack, P. J. Melcher, M. A. Zwieniecki, N. M. Holbrook, The hydraulic conductance of the angiosperm leaf lamina: a comparison of three measurement methods, J. Exp. Bot. 53 (2002) 2177-2184.
48. D. M. Johnson, D. R. Woodruff, K. A. McCulloh, F. C. Meinzer, Leaf hydraulic conductance, measured in situ, declines and recovers daily: leaf hydraulics, water potential and stomatal conductance in four temperate and three tropical tree species, Tree Physiol. 29 (2009) 879-887.
49. P. C. Harley, F. Loreto, G. Di-Marco, T. D. Sharkey, Theoretical Considerations when Estimating the Mesophyll Conductance to CO2 Flux by Analysis of the Response of Photosynthesis to CO2, Plant Physiol. 98 (1992) 1429-1436.
50. D. K. Manter, J. Kerrigan, A/Ci curve analysis across a range of woody plant species: Influence of regression analysis parameters and mesophyll conductance, J. Exp. Bot. 55 (2004) 2581-2588.
51. T. L. Pons, M. H. M. Westbeek, Analysis of differences in photosynthetic nitrogen-use efficiency between four contrasting species, Physiologia Plantarum 122 (2004) 68-78.
52. O. Roupsard, P. Gross, E. Dreyer, Limitation of photosynthetic activity by CO2 availability in the chloroplasts of oak leaves from different species and during drought, Ann. Sci. For. 53 (1996) 243-254.
53. V. Velikova, T. Tsonev, P. Pinelli, G. A. Alessio, F. Loreto, Localized ozone fumigation system for studying ozone effects on photosynthesis, respiration, electron transport rate and isoprene emission in field-grown Mediterranean oak species, Tree Physiol. 25 (2005) 1523-1532.
54. C. R. Warren, E. Dreyer, Temperature response of photosynthesis and internal conductance to CO2: results from two independent approaches, J. Exp. Bot. 57 (2006) 3057-3067.
55. D. Epron, D. Godard, G. Cornic, B. Genty, Limitation of net CO2 assimilation rate by internal resistances to CO2 transfer in the leaves of two tree species (Fagus sylvatica L. and Castanea sativa Mill.), Plant Cell Environ. 18 (1995) 43-51.
56. H. Cochard, J. S. Venisse, T. S. Barigah, N. Brunel, S. Herbette, A. Guilliot, M. T. Tyree, S. Sakr, Putative Role of Aquaporins in Variable Hydraulic Conductance of Leaves in Response to Light, Plant Physiol. 143 (2007) 122-133.
57. Y. T. Hanba, S. I. Miyazawa, H. Kogami, I. Terashima, Effects of leaf age on internal CO2 transfer conductance and photosynthesis in tree species having different types of shoot phenology, Functional Plant Biol. 28 (2001) 1075-1084.
58. Y. T. Hanba, H. Kogami, I. Terashima, The effect of growth irradiance on leaf anatomy and photosynthesis in Acer species differing in light demand, Plant Cell Environ. 25 (2002) 1021-1030.
59. B. J. Bond, K. L. Kavanagh, Stomatal behavior of four woody species in relation to leaf-specific hydraulic conductance and threshold water potential, Tree Physiol. 19 (1999) 503-510.

60. B. Genty, S. Meyer, C. Piel, F. Badeck, R. Liozon, CO diffusion inside leaf mesophyll of ligneous plants2, Photosynthesis: Mechanisms and Effects1998) 3961-3966.
61. M. Ridolfi, E. Dreyer, Responses to water stress in an ABA-unresponsive hybrid poplar (Populus koreana x trichocarpa cv. Peace). III. Consequences for photosynthetic carbon assimilation, New Phytol. 135 (1997) 31-40.
62. A. Sellin, E. O. Unapuu, P. Kupper, Effects of light intensity and duration on leaf hydraulic conductance and distribution of resistance in shoots of silver birch (Betula pendula), Physiologia Plantarum 134 (2008) 412-420.
63. H. Eichelmann, V. Oja, B. Rasulov, E. Padu, I. Bichele, H. Pettai, T. M+Âls, I. Kasparova, E. Vapaavuori, A. Laisk, Photosynthetic parameters of birch (Betula pendula Roth) leaves growing in normal and in CO2- and O3-enriched atmospheres, Plant Cell Environ. 27 (2004) 479-495.
64. H. Eichelmann, V. Oja, B. Rasulov, E. Padu, I. Bichele, H. Pettai, U. Niinemets, A. Laisk, Development of leaf photosynthetic parameters in Betula pendula Roth leaves: Correlations with photosystem I density, Plant. Biol. 6 (2004) 307-318.
65. A. Laisk, H. Eichelmann, V. Oja, B. Rasulov, E. Padu, I. Bichele, H. Pettai, O. Kull, Adjustment of leaf photosynthesis to shade in a natural canopy: rate parameters, Plant Cell Environ 28 (2005) 375-388.
66. A. Laisk, V. Oja, B. Rasulov, H. R+ñmma, H. Eichelmann, I. Kasparova, H. Pettai, E. Padu, E. Vapaavuori, A computer-operated routine of gas exchange and optical measurements to diagnose photosynthetic apparatus in leaves, Plant Cell Environ. 25 (2002) 923-943.
67. C. Piel, E. Frak, X. Le Roux, B. Genty, Effect of local irradiance on CO2 transfer conductance of mesophyll in walnut, J. Exp. Bot. 53 (2002) 2423-2430.
68. M. Lauteri, A. Scartazza, M. C. Guido, E. Brugnoli, Genetic variation in photosynthetic capacity, carbon isotope discrimination and mesophyll conductance in provenances of Castanea sativa adapted to different environments, Funct. Ecol. 11 (1997) 675-683.
69. H. Düring, Stomatal and mesophyll conductances control CO2 transfer to chloroplasts in leaves of grapevine (Vitis vinifera L.), Vitis 42 (2003) 65-68.
70. G. Fila, F. W. Badeck, S. Meyer, Z. Cerovic, J. Ghashghaie, Relationships between leaf conductance to CO2 diffusion and photosynthesis in micropropagated grapevine plants, before and after ex vitro acclimatization, J. Exp. Bot. 57 (2006) 2687-2695.
71. J. Flexas, J. Bota, J. M. Escalona, B. Sampol, H. Medrano, Effects of drought on photosynthesis in grapevines under field conditions: an evaluation of stomatal and mesophyll limitations, Functional Plant Biol. 29 (2002) 461-471.
72. B. Sampol, J. Bota, D. Riera, H. Medrano, J. Flexas, Analysis of the virus-induced inhibition of photosynthesis in malmsey grapevines, New Phytol. 160 (2003) 403-412.
73. I. Fleck, K. Peña-Rojas, X. Aranda, Mesophyll conductance to CO2 and leaf morphological characteristics under drought stress during Quercus ilex L. resprouting, Ann. For. Sci. 67 (2010)
74. Y. T. Hanba, S. I. Miyazawa, I. Terashima, The influence of leaf thickness on the CO2 transfer conductance and leaf stable carbon isotope ratio for some evergreen tree species in Japanese warm-temperate forests, Funct. Ecol. 13 (1999) 632-639.
75. U. Niinemets, A. Cescatti, M. Rodeghiero, T. Tosens, Leaf internal diffusion conductance limits photosynthesis more strongly in older leaves of Mediterranean evergreen broad-leaved species, Plant Cell Environ. 28 (2005) 1552-1566.
76. U. Niinemets, A. Cescatti, M. Rodeghiero, T. Tosens, Complex adjustments of photosynthetic potentials and internal diffusion conductance to current and previous light availabilities and leaf age in Mediterranean evergreen species Quercus ilex, Plant Cell Environ 28 (2006) 1552-1566.
77. G. Bongi, F. Loreto, Gas-Exchange Properties of Salt-Stressed Olive (Olea europea L.) Leaves, Plant Physiol. 90 (1989) 1408-1416.
78. M. Centritto, F. Loreto, K. Chartzoulakis, The use of low [CO2] to estimate diffusional and non-diffusional limitations of photosynthetic capacity of salt-stressed olive saplings, Plant Cell Environ. 26 (2003) 585-594.
79. A. Diaz-Espejo, E. Nicolás, J. E. Fern+índez, Seasonal evolution of diffusional limitations and photosynthetic capacity in olive under drought, Plant Cell Environ. 30 (2007) 922-933.

80. F. Loreto, M. Centritto, K. Chartzoulakis, Photosynthetic limitations in olive cultivars with different sensitivity to salt stress, Plant Cell Environ. 26 (2003) 595-601.
81. C. J. Blackman, T. J. Brodribb, G. J. Jordan, Leaf hydraulic vulnerability is related to conduit dimensions and drought resistance across a diverse range of woody angiosperms, New Phytol. 188 (2010) 1113-1123.
82. F. Costa e Silva, A. Shvaleva, P. Maroco, M. H. Almeida, M. M. Chaves, J. S. Pereira, Responses to water stress in two Eucalyptus globulus clones differing in drought tolerance, Tree Physiol. 24 (2004) 1165-1172.
83. C. Scoffoni, A. Pou, K. Aasamaa, L. Sack, The rapid light response of leaf hydraulic conductance: new evidence from two experimental methods, Plant Cell Environ 31 (2008) 1803-1812.
84. C. R. Warren, The photosynthetic limitation posed by internal conductance to CO 2 movement is increased by nutrient supply, J. Exp. Bot. 55 (2004) 2313-2321.
85. C. R. Warren, Does growth temperature affect the temperature responses of photosynthesis and internal conductance to CO2? A test with Eucalyptus regnans, Tree Physiol. 28 (2008) 11-19.
86. C. R. Warren, Soil water deficits decrease the internal conductance to CO2 transfer but atmospheric water deficits do not, J. Exp. Bot. 59 (2008) 327-334.
87. J. C. Domec, A. Noormets, J. S. king, G. Sun, S. G. McNulty, M. J. Gavazzi, J. L. Boggs, E. A. Treasure, Decoupling the influence of leaf and root hydraulic conductances on stomatal conductance and its sensitivity to vapour pressure deficit as soil dries in a drained loblolly pine plantation, Plant Cell Environ. 32 (2009) 980-991.
88. D. M. Johnson, F. C. Meinzer, D. R. Woodruff, K. A. McCulloh, Leaf xylem embolism, detected acoustically and by cryo-SEM, corresponds to decreases in leaf hydraulic conductance in four evergreen species, Plant Cell Environ 32 (2009) 828-836.
89. C. R. Warren, Why does photosynthesis decrease with needle age in Pinus pinaster?, Trees - Structure and Function 20 (2006) 157-164.
90. D. R. Woodruff, F. C. Meinzer, B. Lachenbruch, D. M. Johnson, Coordination of leaf structure and gas exchange along a height gradient in a tall conifer, Tree Physiol. 29 (2009) 261-272.
91. G. J. Ethier, N. J. Livingston, D. L. Harrison, T. A. Black, J. A. Moran, Low stomatal and internal conductance to CO2 versus Rubisco deactivation as determinants of the photosynthetic decline of ageing evergreen leaves, Plant Cell Environ. 29 (2006) 2168-2184.
92. C. R. Warren, N. J. Livingston, D. H. Turpin, Water stress decreases the transfer conductance of Douglas-fir (Pseudotsuga menziesii) seedlings, Tree Physiol. 24 (2004) 971-979.

Appendix B Leaf water enrichment models and scaled effective path length
Steady-state isotopic enrichment over source water at the site of evaporation (e) has been described by the Craig & Gordon model [1-2]:
e k v k ea
ei
(Eq. S1)
where + is the equilibrium fractionation between liquid water and vapour [3]; k is the kinetic fractionation as vapour diffuses from leaf intercellular spaces to the atmosphere [4-5], v is the isotopic enrichment of atmospheric water vapour relative to plant source water, and ea/ei is the ratio of ambient to intercellular vapour pressures.
The steady-state isotopic enrichment of mean lamina mesophyll water (LsP) can be described by the above steady-state Craig & Gordon model corrected for the gradient from xylem source water to enriched water at the evaporating sites, the so-called Péclet effect [6]:
LsP e
1 e
with
D
lv (Eq. S2)
where (Péclet number) is the ratio of convection to diffusion, v is the linear velocity of water movement (m s-1), l is the length of the water pathway along which diffusion occurs (i.e. the distance between the xylem and the evaporation surface), and D the tracer-diffusivity (m2 s-1) of heavy water isotopologues (either H2
18O or 2H1HO) in ‘normal’ water. Linear velocity through the water pathway (v) can be related to transpiration rate (E, mol H2O m-2 s-
1) as follows [7]:
C
Ekv (Eq. S3)
where C is the molar concentration of water (55.56 103 mol m-3) and k is a scaling factor to convert the velocity of water moving through the leaf as a slab (E/C) to the actual velocity in a porous medium, and usually ranges from 102 to 103. Accordingly, can be also expressed as:
DC
LE
eff (Eq. S4)
where Leff stands for the scaled effective pathlength, i.e. the product of l and k. Thus, Leff is expected to be much larger than the actual distance between the xylem and the evaporation sites, and would vary not only with changes in l but also with changes in the tortuosity of the water pathway (e.g. varying the proportion of apoplastic and cell-to-cell water pathways; [8]).

References Appendix B
1. H. Craig, L.I. Gordon, Deuterium and oxygen-18 variations in the ocean and the marine atmosphere. In: Tongiorgi T (ed) Proceeding of the conference on stable isotopes in oceanographic studies and palaeotemperatures, Spoleto, Italy, V. Lishi e F., Pisa, 1965, pp 9-130.
2. G. Dongmann, H.W. Nurnberg, H. Forstel, K. Wagener (1974) On the enrichment of H2
18O in the leaves of transpiring plants. Radiation and Environmental Biophysics 11 (1974) 41-52.
3. M. Majoube Fractionnement en oxygene-18 et en deuterium entre l'eau et sa vapeur. Journal de Chimie et Physique 58 (1971) 1423-1436.
4. G.D. Farquhar, K.T. Hubick, A.G. Condon, R.A. Richards, Carbon Isotope Fractionation and Plant Water-Use Efficiency. In: Rundel PW, Ehleringer JR, Nagy KA (eds) Stable Isotopes in Ecological Research, vol 68. Springer cop., New York, 1989, pp 21-40.
5. C.D. Cappa, M.B. Hendricks, D.J. DePaolo, R.C. Cohen, Isotopic fractionation of water during evaporation. Journal of Geophysical Research-Atmospheres 108 (2003) 4525.
6. G.D. Farquhar, J. Lloyd, Carbon and oxygen isotope effects in the exchange of carbon dioxide between terrestrial plants and the atmosphere, in: J.R. Ehleringer, A.E. Hall, G.D. Farquhar (Eds.), Stable Isotopes and Plant Carbon–Water Relations, Academic Press, San Diego, 1993, pp. 47–70.
7. M.M. Barbour, G.D. Farquhar Do pathways of water movement and leaf anatomical dimensions allow development of gradients in H2
18O between veins and the sites of evaporation within leaves. Plant, Cell and Environment 27 (2004) 107-121.
8. E. Steudle, J. Frensch, Water transport in plants: Role of the apoplast. Plant Soil 187 (1996) 67-79.





























