Embed Size (px)
Citation preview

Microbiology 1:
Revision Notes
2008/2009
Version 1.3
Richard Smith
January 6, 2009
Abstract
This document is meant to help with revision for the UWE Microbi-ology 1 examination. It is designed to be used with the Anki �ashcardswhich are available at http://blahah.wordpress.com. At that address youwill also �nd links and textbooks which should supplement this material.I think I've covered everything in the syllabus and in the sample/pastpapers - please let me know if you think I've missed anything. There maybe updates as I hone these notes - keep checking the site.
You can click red entries in the Table of Contents to go directly to thesection described.
-Rik
Contents
I An Overview 10
1 History 10
1.1 Spontaneous generation (aka Aristotelian Abiogenesis) . . . . . . 10
1.2 Antoni van Leeuwenhoek . . . . . . . . . . . . . . . . . . . . . . 10
1.3 Francisco Redi . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
1.4 Louis Pasteur . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
1.5 Robert Koch . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
1.5.1 Limitations of Koch's postulates . . . . . . . . . . . . . . 12
1.6 and so on, up to date... . . . . . . . . . . . . . . . . . . . . . . . 12
1

2 Classi�cation 13
2.1 Linnaeus and the binomial nomenclature . . . . . . . . . . . . . . 13
2.2 Modern taxonomy . . . . . . . . . . . . . . . . . . . . . . . . . . 13
2.3 Phylogenetics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
2.3.1 Ernst Haeckel's recapitulation theory . . . . . . . . . . . . 14
2.3.2 Gene transfer . . . . . . . . . . . . . . . . . . . . . . . . . 14
2.3.3 Ribosomal (16s) RNA . . . . . . . . . . . . . . . . . . . . 15
3 Nutrition 16
3.1 Nutritional categories . . . . . . . . . . . . . . . . . . . . . . . . 16
3.1.1 Heterotrophy . . . . . . . . . . . . . . . . . . . . . . . . . 16
3.1.2 Chemo- & Photoheterotrophy . . . . . . . . . . . . . . . . 16
3.1.3 Autotrophy . . . . . . . . . . . . . . . . . . . . . . . . . . 16
3.1.4 Chemo- & Photoautotrphy . . . . . . . . . . . . . . . . . 17
3.2 Litho- and Organotrophy . . . . . . . . . . . . . . . . . . . . . . 17
4 Archaea 17
4.1 Euryarchaeota . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
4.2 Crenarchaeota . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
4.3 Korarchaeota . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
5 Bacteria 19
5.1 The most important phyla . . . . . . . . . . . . . . . . . . . . . . 19
5.1.1 Proteobacteria . . . . . . . . . . . . . . . . . . . . . . . . 19
5.1.2 Other Gram-negative phyla . . . . . . . . . . . . . . . . . 22
5.1.3 Gram Positive Bacteria . . . . . . . . . . . . . . . . . . . 23
5.2 Biochemistry (some important terms to learn) . . . . . . . . . . . 24
5.2.1 Peptidoglycan . . . . . . . . . . . . . . . . . . . . . . . . . 24
5.2.2 Exotoxins . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
5.2.3 Endotoxins . . . . . . . . . . . . . . . . . . . . . . . . . . 25
5.2.4 Enterotoxins . . . . . . . . . . . . . . . . . . . . . . . . . 25
5.2.5 Bacteriocins . . . . . . . . . . . . . . . . . . . . . . . . . . 25
2

5.2.6 Siderophores . . . . . . . . . . . . . . . . . . . . . . . . . 26
5.3 Reproduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26
5.3.1 Binary Fission . . . . . . . . . . . . . . . . . . . . . . . . 26
5.4 Endospores . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
5.5 Morphology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28
5.6 Some important example Bacteria . . . . . . . . . . . . . . . . . 30
5.6.1 Escherichia coli . . . . . . . . . . . . . . . . . . . . . . . . 31
5.6.2 Staphylococcus aureus . . . . . . . . . . . . . . . . . . . . 31
5.6.3 Pseudomonas aeruginosa . . . . . . . . . . . . . . . . . . 32
5.6.4 Enterococcus faecalis . . . . . . . . . . . . . . . . . . . . . 33
5.6.5 Bacillus subtilis . . . . . . . . . . . . . . . . . . . . . . . . 34
6 Fungi 35
6.1 Importance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35
6.2 Morphology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35
6.2.1 Yeasts . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35
6.2.2 Multicellular fungi . . . . . . . . . . . . . . . . . . . . . . 36
6.2.3 Dimorphism . . . . . . . . . . . . . . . . . . . . . . . . . . 36
6.3 Structure . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37
6.4 Nutrition . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37
6.5 Reproduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37
6.6 Classi�cation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37
6.6.1 Zygomycota . . . . . . . . . . . . . . . . . . . . . . . . . . 37
6.6.2 Chytridiomycota . . . . . . . . . . . . . . . . . . . . . . . 38
6.6.3 Ascomycota . . . . . . . . . . . . . . . . . . . . . . . . . . 38
6.6.4 Basidiomycota . . . . . . . . . . . . . . . . . . . . . . . . 39
7 Virii 41
7.1 Structure . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
7.2 Viral genome . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
7.3 Bacteriophages . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
7.3.1 Viroids and Prions . . . . . . . . . . . . . . . . . . . . . . 42
3

8 Protista 44
8.1 The 'Algae' . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44
8.1.1 Structural characteristics of algal protists . . . . . . . . . 44
8.1.2 Euglenophyta . . . . . . . . . . . . . . . . . . . . . . . . . 45
8.2 Dino�agellata . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46
8.3 Diatoms . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47
8.4 Chlorophyta . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
8.5 Rhodophyta . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
8.6 `The Protozoa' . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
8.7 Amoebas with external shells (Foramanifera & Radiolaria) . . . . 50
8.8 Sporozoans (Apixomplexa) . . . . . . . . . . . . . . . . . . . . . 50
8.9 Slime moulds and water moulds . . . . . . . . . . . . . . . . . . . 51
8.9.1 Oomycota (water moulds) . . . . . . . . . . . . . . . . . . 51
8.9.2 Myxomycota (plasmodial slime moulds) . . . . . . . . . . 51
8.9.3 Dictyostelida (cellular slime moulds) . . . . . . . . . . . . 52
9 Growth & Death 53
9.1 Factors a�ecting microbial growth . . . . . . . . . . . . . . . . . 53
9.1.1 Temperature . . . . . . . . . . . . . . . . . . . . . . . . . 53
9.1.2 pH . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 53
9.1.3 Oxygen . . . . . . . . . . . . . . . . . . . . . . . . . . . . 53
9.1.4 Carbon Dioxide . . . . . . . . . . . . . . . . . . . . . . . . 54
9.1.5 Osmotic Pressure . . . . . . . . . . . . . . . . . . . . . . . 54
9.1.6 Light . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 55
9.2 The kinetics of microbial growth . . . . . . . . . . . . . . . . . . 55
9.2.1 Lag phase . . . . . . . . . . . . . . . . . . . . . . . . . . . 55
9.2.2 Log phase (also exponential or logarithmic phase) . . . . 56
9.2.3 Stationary phase . . . . . . . . . . . . . . . . . . . . . . . 56
9.2.4 Death phase (also exponential or logarithmic decline) . . 57
9.3 Batch culture and continuous culture . . . . . . . . . . . . . . . . 57
9.4 Growth in multicellular organisms . . . . . . . . . . . . . . . . . 58
9.5 Culture media . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58
4

9.6 Chemically de�ned media . . . . . . . . . . . . . . . . . . . . . . 59
9.7 Selective and di�erential media . . . . . . . . . . . . . . . . . . . 59
9.8 Microbial death kinetics . . . . . . . . . . . . . . . . . . . . . . . 60
9.8.1 The D value, z value, F value . . . . . . . . . . . . . . . . 60
9.8.2 D and z values in the food industry (possible long answerexam question, learn this!) . . . . . . . . . . . . . . . . . 61
II Microbial control 63
10 The control of microorganisms 63
10.1 Classi�cation of pathogens . . . . . . . . . . . . . . . . . . . . . . 64
10.2 Sterilisation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 65
10.2.1 Sterilisation by heat . . . . . . . . . . . . . . . . . . . . . 65
10.2.2 The Autoclave . . . . . . . . . . . . . . . . . . . . . . . . 65
10.2.3 Tyndallisation / Intermittent steaming . . . . . . . . . . . 66
10.2.4 Pasteurisation . . . . . . . . . . . . . . . . . . . . . . . . . 66
10.3 Sterilisation by irradiation . . . . . . . . . . . . . . . . . . . . . . 67
10.3.1 UV Radiation . . . . . . . . . . . . . . . . . . . . . . . . . 67
10.3.2 Ionising radiation . . . . . . . . . . . . . . . . . . . . . . . 68
10.3.3 Gamma radiation . . . . . . . . . . . . . . . . . . . . . . . 68
10.4 Filtration . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 68
10.5 Phenols and phenolics . . . . . . . . . . . . . . . . . . . . . . . . 69
10.6 Alcohols . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 69
10.7 Halogens . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 69
10.8 Heavy metals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70
10.9 Aldehydes: Glutaraldehyde and Formaldehyde . . . . . . . . . . 70
10.10Surfactants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 71
11 Antimicrobial agents 72
11.1 Antibiotics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 72
11.1.1 Fleming and Penicillin . . . . . . . . . . . . . . . . . . . . 73
11.1.2 Types of antibiotic . . . . . . . . . . . . . . . . . . . . . . 74
11.2 Anti fungal agents . . . . . . . . . . . . . . . . . . . . . . . . . . 75
11.3 Anti viral agents . . . . . . . . . . . . . . . . . . . . . . . . . . . 75
5

III The importance of microorganisms 76
12 Disease and microorganisms 76
12.1 Classi�cation of diseases . . . . . . . . . . . . . . . . . . . . . . . 76
12.2 Some bacterial diseases of humans . . . . . . . . . . . . . . . . . 77
12.2.1 Waterborne transmission: cholera . . . . . . . . . . . . . . 77
12.2.2 Airborne transmission: `strep' throat . . . . . . . . . . . . 78
12.2.3 Contact transmission: syphilis . . . . . . . . . . . . . . . 79
12.2.4 Vector-borne transmission: plague . . . . . . . . . . . . . 79
12.2.5 Food poisoning: botulism . . . . . . . . . . . . . . . . . . 79
12.2.6 Food infection: salmonellosis . . . . . . . . . . . . . . . . 80
12.3 Fungi and disease . . . . . . . . . . . . . . . . . . . . . . . . . . . 80
12.3.1 Ergot . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81
12.3.2 Mycotoxins . . . . . . . . . . . . . . . . . . . . . . . . . . 82
12.3.3 Parasitism . . . . . . . . . . . . . . . . . . . . . . . . . . . 82
12.3.4 Fungal diseases of plants . . . . . . . . . . . . . . . . . . . 82
12.4 Protists and human disease . . . . . . . . . . . . . . . . . . . . . 82
12.4.1 Malaria . . . . . . . . . . . . . . . . . . . . . . . . . . . . 82
12.4.2 African Sleeping Sickness (or Trypanosomiasis) . . . . . . 83
12.5 Viral disease . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83
12.5.1 Airborne transmission: in�uenza . . . . . . . . . . . . . . 83
12.5.2 Transmission by water or food: viral gastroenteritis . . . 84
12.5.3 Latent and slow (persistent) viral infections . . . . . . . . 84
12.5.4 Viruses and cancer . . . . . . . . . . . . . . . . . . . . . . 84
12.5.5 Virus vaccine . . . . . . . . . . . . . . . . . . . . . . . . . 85
12.5.6 Interferons (IFNs) . . . . . . . . . . . . . . . . . . . . . . 85
12.6 Immunity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 85
12.6.1 Non-speci�c immunity . . . . . . . . . . . . . . . . . . . . 86
12.6.2 Speci�c immunity . . . . . . . . . . . . . . . . . . . . . . 87
12.7 Vaccines . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 88
12.7.1 Types of vaccines . . . . . . . . . . . . . . . . . . . . . . . 88
12.7.2 Developing a vaccine . . . . . . . . . . . . . . . . . . . . . 90
6

13 Importance in natural world: associations 91
13.1 Types of microbial associations . . . . . . . . . . . . . . . . . . . 91
13.1.1 Parasitism: . . . . . . . . . . . . . . . . . . . . . . . . . . 91
13.1.2 Mutualism: . . . . . . . . . . . . . . . . . . . . . . . . . . 91
13.1.3 Commensalism: . . . . . . . . . . . . . . . . . . . . . . . . 91
13.2 Microbial association with animals . . . . . . . . . . . . . . . . . 91
13.2.1 Termites . . . . . . . . . . . . . . . . . . . . . . . . . . . . 91
13.2.2 The honey gude bird . . . . . . . . . . . . . . . . . . . . . 92
13.2.3 Giant tube worms . . . . . . . . . . . . . . . . . . . . . . 92
13.2.4 Humans . . . . . . . . . . . . . . . . . . . . . . . . . . . . 92
13.3 Microbial associations with plants . . . . . . . . . . . . . . . . . 93
13.3.1 Ectomycorrhizae . . . . . . . . . . . . . . . . . . . . . . . 93
13.3.2 Endomycorrhizae . . . . . . . . . . . . . . . . . . . . . . . 93
13.3.3 Nitrogen-�xing bacteria . . . . . . . . . . . . . . . . . . . 93
13.4 Microbial associations with other microorganisms . . . . . . . . . 94
13.4.1 Lichens . . . . . . . . . . . . . . . . . . . . . . . . . . . . 94
13.4.2 E.coli and Enterococcus faecalis . . . . . . . . . . . . . . . 94
13.5 A note on secondary metabolites, because they feature in an examquestion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 95
14 Importance in natural world: the environment 97
14.1 The carbon cycle . . . . . . . . . . . . . . . . . . . . . . . . . . . 97
14.2 The nitrogen cycle . . . . . . . . . . . . . . . . . . . . . . . . . . 98
14.3 Bene�cial e�ects of microorganisms in the environment . . . . . . 98
14.3.1 Solid waste treatment: composting and land�ll . . . . . . 98
14.3.2 Waste water treatment . . . . . . . . . . . . . . . . . . . . 99
14.3.3 Bioremediation . . . . . . . . . . . . . . . . . . . . . . . . 100
14.4 Harmful e�ects of microorganisms in the environment . . . . . . 101
7

15 Human indigenous microbiota (normal �ora) 103
15.1 Associations Between Humans and the Normal Flora . . . . . . . 103
15.2 Tissue speci�city . . . . . . . . . . . . . . . . . . . . . . . . . . . 104
15.3 The composition of the normal �ora . . . . . . . . . . . . . . . . 104
15.3.1 Normal �ora of the skin . . . . . . . . . . . . . . . . . . . 105
15.3.2 Normal �ora of the conjunctiva . . . . . . . . . . . . . . . 105
15.3.3 Normal �ora of the respiratory tract . . . . . . . . . . . . 106
15.3.4 Normal Flora of the Urogenital Tract . . . . . . . . . . . 106
15.3.5 Normal Flora of the Oral Cavity . . . . . . . . . . . . . . 107
15.3.6 Normal Flora of the Gastrointestinal Tract . . . . . . . . 108
15.4 Bene�cial e�ects of the normal �ora . . . . . . . . . . . . . . . . 109
15.5 Harmful e�ects of the normal �ora . . . . . . . . . . . . . . . . . 110
16 Industrial applications 112
16.1 Food and beverage industries . . . . . . . . . . . . . . . . . . . . 112
16.1.1 Alcohol (wines & spirits) . . . . . . . . . . . . . . . . . . 112
16.1.2 Beer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 112
16.1.3 Dairy products . . . . . . . . . . . . . . . . . . . . . . . . 113
16.1.4 Bread . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 114
16.1.5 Microorganisms as food . . . . . . . . . . . . . . . . . . . 114
16.1.6 Microbial spoilage of food . . . . . . . . . . . . . . . . . . 114
16.2 Microbial production of biochemicals . . . . . . . . . . . . . . . . 115
16.2.1 Acetone . . . . . . . . . . . . . . . . . . . . . . . . . . . . 115
16.2.2 Amino acids (and MSG) . . . . . . . . . . . . . . . . . . . 116
16.2.3 Citric acid . . . . . . . . . . . . . . . . . . . . . . . . . . . 116
16.2.4 Vitamins . . . . . . . . . . . . . . . . . . . . . . . . . . . 116
16.2.5 Industrial enzymes . . . . . . . . . . . . . . . . . . . . . . 116
16.2.6 Syrups . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 117
16.3 Genetically modi�ed organisms . . . . . . . . . . . . . . . . . . . 117
16.4 Microbial mining . . . . . . . . . . . . . . . . . . . . . . . . . . . 117
8

17 Lab techniques 119
17.1 Units . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 119
17.2 Safety . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 119
17.2.1 Aseptic technique . . . . . . . . . . . . . . . . . . . . . . . 119
17.2.2 Microbiological safety cabinets . . . . . . . . . . . . . . . 120
17.3 Pure culture . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 121
17.3.1 Streak plate . . . . . . . . . . . . . . . . . . . . . . . . . . 121
17.3.2 Pour plate . . . . . . . . . . . . . . . . . . . . . . . . . . . 121
17.4 Preparing specimens . . . . . . . . . . . . . . . . . . . . . . . . . 122
17.4.1 Smear . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 122
17.4.2 Wet Mount . . . . . . . . . . . . . . . . . . . . . . . . . . 123
17.5 Counting Methods . . . . . . . . . . . . . . . . . . . . . . . . . . 123
17.5.1 Total cell count - haemocytometer . . . . . . . . . . . . . 123
17.5.2 Colony forming units . . . . . . . . . . . . . . . . . . . . . 123
17.5.3 Serial dilution - estimating viable cell numbers . . . . . . 124
17.5.4 Spread plate (Miles & Misra) . . . . . . . . . . . . . . . . 125
17.5.5 Pour plate . . . . . . . . . . . . . . . . . . . . . . . . . . . 125
17.5.6 Turbidimetric . . . . . . . . . . . . . . . . . . . . . . . . . 126
17.6 Staining methods . . . . . . . . . . . . . . . . . . . . . . . . . . . 126
17.6.1 Types of stains . . . . . . . . . . . . . . . . . . . . . . . . 126
17.6.2 Here's how to Gram-stain a specimen: . . . . . . . . . . . 127
17.6.3 Here's how to apply the Ziehl-Nielsen acid-fast stain to aspecimen: . . . . . . . . . . . . . . . . . . . . . . . . . . . 128
17.6.4 Special stains . . . . . . . . . . . . . . . . . . . . . . . . . 128
17.6.5 The endospore stain . . . . . . . . . . . . . . . . . . . . . 128
9

Part I
An Overview
1 History
1.1 Spontaneous generation (aka Aristotelian Abiogene-sis)
Until 1859 the prevalent (non-religious) theory to explain growth of new or-ganisms from inanimate matter was 'spontaneous generation'. The theory wassuggested by Aristotle, among others, hence the name 'Aristotelian Abiogene-sis' (abiogenesis = [greek] �birth without life�). The theory held that life arosespontaneously and frequently without reliance on former life. An example ofwhere this belief was applied is to the arrival of maggots on meat. The theorywas eventually disproven by Pasteur in 1859, although several other earlier ex-perimentors made signi�cant contributions (Francisco Redi, Schwann, Cagniardde la Tour etc.).
1.2 Antoni van Leeuwenhoek
Antoni van Leeuwenhoek (1632-1723) a.k.a �the father of microbiology�. Hewas the �rst person to describe and accurately draw bacteria, protists, rotifers,and spermatazoa - he called them all 'animalcules'. He also made accuratemicroscopic observations of muscle �bres and the blood �ow in capillaries.
During his lifetime, Leeuwenhoek ground over 500 optical lenses, putting themto use in more than 400 di�erent types of microscope. Only 9 of these still exist.He communicated extensively with the British Royal Society and provided themwith many slides which have been preserved to this day.
1.3 Francisco Redi
Francesco Redi (1626-97) challenged the idea that maggots arose spontaneouslyfrom rotting meat. In the �rst major experiment to challenge spontaneousgeneration, he placed meat in a variety of open, covered and partially coveredcontainers. After a �ew days he observed maggots on the open and partiallycovered meat, but not on the meat covered with a gauze (which had prevented�ies from landing). He also captured maggots and observed them metamorphoseinto �ies. Whilst this was evidence supporting the fact that maggots do notarise spontaneously, many still held that spontaneous generation was true formicroscopic organisms.
10

Figure 1: Leeuwenhoek's microscope. The lens (a) was held between twobrass plates and used to view the specimen, which was placed on the mountingpin (b). Focusing was achieved by means of two screws (c) and (d). Some ofLeeuwenhoek's microscopes could magnify up to 300 times. Original source:Antony van Leeuwenhoek and his little animals by C. E. Dobell (1932).
1.4 Louis Pasteur
Pasteur �nally disproved spontaneous generation in 1859 with his swan-necked�ask experiments. These �asks were �lled with meat broth, and had a long, slen-der neck which followed several bends, preventing dust and other material fromsettling into the �ask. The fact that no bacterial growth occured inthe �asks,despite air still being able to pass into the �asks, showed that it is germinationrather than spontaneous generation which gives rise to organisms.
1.5 Robert Koch
The �rst proof of the involvement of bacteria in disease and the de�nitive proofof the germ theory of disease came from the German Robert Koch. In 1876Koch showed the relationship between the cattle disease anthrax and a bacilluswhich we now know as Bacillus anthracis.
11

Koch's Postulates
1. The microorganism must be present in every instance of the disease andabsent from healthy individuals.
2. The microorganism must be capable of being isolated and grown in pureculture.
3. When the microorganism is inoculated into a healthy host, the same dis-ease condition must result.
4. The same microorganism must be re-isolated from the experimentally in-fected host.
1.5.1 Limitations of Koch's postulates
Despite their value, it is now realised that Koch's postulates do have certainlimitations. It is known for example that certain agents responsible for causingdisease (e.g. viruses and prions) can't be grown in vitro, but only in host cells.
Also, the healthy animal in Postulate 3 is seldom human, so a degree of extrap-olation is necessary � if agent X does not cause disease in a laboratory animal,can we be sure it won't in humans?
Furthermore, some diseases are caused by more than one organism, and someorganisms are responsible for more than one disease.
1.6 and so on, up to date...
� After the discovery of viruses in the late 1800s, it soon became known thatthey were the cause of many diseases.
� A major development of the 20th Century was the development of antibi-tiocs and other antimicrobial agents.
� The second half of the 20th Century is characterised (in microbiologi-cal terms) by the great leaps made in molecular genetics. Sequencingof genomes, robosomal RNA sequencing etc have led to unprecedentedinsight into phylogeny and biochemistry.
n.b. other history is discussed in the relevant section.
12

2 Classi�cation
2.1 Linnaeus and the binomial nomenclature
Linnaeus (real name Carl von Linné), a swedish botanist, created the binomialclassi�cation system which is used universally in modern taxonomy (viruses sitoutside of the biological taxonomic system). In addition, he also discovered andclassi�ed many of the species recognised today.
It is important to know the rules for using this binomial (two-name) system:
� the generic (genus) name is always given a capital letter
� the speci�c (species) name is given a small letter
� the generic and speci�c name are italicised, or, if this isn't possible,underlined
� the generic name may be shortened in practise, in which case the abbre-viation is dotted.
Some examples:
� if typed: Escherichia coli
� if handwritten: Escherichia coli
� abbreviated: E. coli or E. coli
2.2 Modern taxonomy
There are 8 main taxonomic ranks in use today: domain, kingdom, phylum,class, order, family, genus, species.
Whereas Linnaeus classi�ed for ease of identi�cation, it is now generally ac-cepted that classi�cation should re�ect the Darwinian principle of common de-scent.
Since the 1960s a trend called cladistic taxonomy (or cladistics or cladism) hasemerged, arranging taxa in an evolutionary tree. If a taxon includes all thedescendants of some ancestral form, it is called monophyletic, as opposed toparaphyletic. Other groups are called polyphyletic.
A new formal code of nomenclature, the PhyloCode, to be renamed "Inter-national Code of Phylogenetic Nomenclature" (ICPN), is currently under de-velopment, intended to deal with clades, which do not have set ranks, unlikeconventional Linnaean taxonomy. It is unclear, should this be implemented,how the di�erent codes will coexist.
13

Domains are a relatively new grouping, suggested by Carl Woese et. al. Thethree-domain system was �rst invented in 1990, but not generally accepted untillater. Now, the majority of biologists accept the domain system, but a largeminority use the �ve-kingdom method. One main characteristic of the three-domain method is the separation of Archaea and Bacteria, previously groupedinto the single kingdom Bacteria (a kingdom also sometimes called Monera).Consequently, the three domains of life are conceptualized as Archaea, Bacteria,and Eukaryota (comprising the nuclei-bearing eukaryotes). A small minorityof scientists add Archaea as a sixth kingdom, but do not accept the domainmethod.
Thomas Cavalier-Smith, who has published extensively on the classi�cation ofprotists, has recently proposed that the Neomura, the clade which groups to-gether the Archaea and Eukarya, would have evolved from Bacteria, more pre-cisely from Actinobacteria.
2.3 Phylogenetics
Phylogenetics is the study of evolutionary relatedness among various groups oforganisms (e.g., species, populations), which is discovered through molecularsequencing data.
2.3.1 Ernst Haeckel's recapitulation theory
During the late 19th century, Ernst Haeckel's recapitulation theory, or biogeneticlaw, was widely accepted. This theory was often expressed as "ontogeny recapit-ulates phylogeny", i.e. the development of an organism exactly mirrors the evo-lutionary development of the species. Haeckel's early version of this hypothesis[that the embryo mirrors adult evolutionary ancestors] has since been rejected,and the hypothesis amended as the embryo's development mirroring embryos ofits evolutionary ancestors. Most modern biologists recognize numerous connec-tions between ontogeny and phylogeny, explain them using evolutionary theory,or view them as supporting evidence for that theory.
2.3.2 Gene transfer
Organisms can generally inherit genes in two ways: vertical gene transfer andhorizontal gene transfer. Vertical gene transfer is the passage of genes fromparent to o�spring, and horizontal gene transfer or lateral gene transfer oc-curs when genes jump between unrelated organisms, a common phenomenon inprokaryotes.
Lateral gene transfer has complicated the determination of phylogenies of or-ganisms since inconsistencies have been reported depending on the gene chosen.
14

2.3.3 Ribosomal (16s) RNA
Carl Woese came up with the three-domain theory of life (eubacteria, archaeaand eukaryotes) based on his discovery that the genes encoding ribosomal RNAare ancient and distributed over all lineages of life with little or no lateral genetransfer. Therefore rRNA are commonly recommended as molecular clocks forreconstructing phylogenies.
This has been particularly useful for the phylogeny of microorganisms, to whichthe species concept does not apply and which are too morphologically simple tobe classi�ed based on phenotypic traits.
15

3 Nutrition
3.1 Nutritional categories
Microorganisms can be categorised according to how they obtain their carbonand energy. Carbon is the most abundant component of the microbial cell,and most microorganisms obtain their carbon in the form of organic molecules,derived directly or indirectly from other organisms. This mode of nutrition isthe one that is familiar to us as humans (and all other animals); all the food weeat is derived as complex organic molecules from plants and other animals (andeven some representatives of the microbial world such as mushrooms!).
3.1.1 Heterotrophy
Microorganisms which obtain their carbon in this way are described as het-erotrophs, and include all the fungi and protozoans as well as most types ofbacteria.
Microorganisms as a group are able to incorporate the carbon from an incred-ibly wide range of organic compounds into cellular material. In fact there ishardly any such compound occurring in nature that cannot be metabolised bysome microorganism or other, explaining in part why microbial life is to befound thriving in the most unlikely habitats. Many synthetic materials can alsoserve as carbon sources for some microorganisms, which can have considerableeconomic signi�cance.
3.1.2 Chemo- & Photoheterotrophy
� The great majority of heterotrophs obtain energy as well as carbon fromthe same organic source. Such organisms release energy by the chemicaloxidation of organic nutrient molecules, and are therefore termed chemo-heterotrophs.
� Those few heterotrophs which do not follow this mode of nutrition includethe green and purple non-sulphur bacteria. These are able to carry outphotosynthesis and are known as photoheterotrophs.
3.1.3 Autotrophy
A signi�cant number of bacteria and all of the algae do not, however, take uptheir carbon preformed as organic molecules in this way, but derive it insteadfrom carbon dioxide. These organisms are called autotrophs, and again we candraw a parallel with higher organisms, where all members of the plant kingdomobtain their carbon in a similar fashion.
16

3.1.4 Chemo- & Photoautotrphy
We can also categorise microorganisms nutritionally by the way they derive theenergy they require to carry out essential cellular reactions. Autotrophs thusfall into two categories.
� Chemoautotrophs obtain their energy as well as their carbon from in-organic sources; they do this by the oxidation of inorganic molecules suchas sulphur or nitrite.
� Photoautotrophs have photosynthetic pigments enabling them to con-vert light energy into chemical energy.
3.2 Litho- and Organotrophy
There is one �nal subdivision of nutritional categories in microorganisms! Whetherorganisms are chemotrophs or phototrophs, they need a molecule to act as asource of electrons (reducing power) to drive their energy generating systems.
� Those able to use an inorganic electron donor such as H2O, H2S or am-monia are called lithotrophs
� Those requiring an organic molecule to ful�l the role are organotrophs.
Most (but not all) microorganisms are either photolithotrophic autotrophs (al-gae, blue-greens) or chemo-organotrophic heterotrophs (most bacteria). For thelatter category, a single organic compound can often act as the provider of car-bon, energy and reducing power. The substance used by chemotrophs as an en-ergy source may be organic (chemoorganotrophs) or inorganic (chemolithotrophs).
4 Archaea
� An archaeaon is a single-celled prokaryote. It has no cell nucleus and nocell organelles.
� Initially, archaea were seen as extremophiles that lived in harsh environ-ments, such as hot springs and salt lakes, but they have since been foundin a broad range of habitats, such as soils, oceans, and marshlands.
� Archaea are particularly numerous in the oceans, and the archaea inplankton may be one of the most abundant groups of organisms on theplanet.
� These prokaryotes are now recognized as a major part of life on Earth andmay play an important role in both the carbon cycle and nitrogen cycle.
17

� No clear examples of archaeal pathogens or parasites are known, but theyare often mutualists or commensals.
� Archaea reproduce asexually and divide by binary �ssion, fragmen-tation, or budding.
� In contrast to bacteria and eukaryotes, no species of archaea areknown that form spores.
� Archaean cell membranes di�er greatly from those found in other formsof life - they do not contain muramic acid.
� Most possess a cell wall and one or more �agella.
4.1 Euryarchaeota
The largest phylum of archaea, including the halophiles (salt-loving, Halobac-teria) and methanogenic forms. Halobacteria appear in large reddish bloomsin salt evaporations plains. The red colouration is caused by the bacterial pig-ment bacteriorhodopsin. Methanogens such asMethanococcus andMethanobac-
terium are unique among all life forms in their ability to generate methane fromsimple carbon compounds. Some methanogens are found in the gut of humansand, signi�cantly, ruminants. Methanogens are also used in sewage treatment.
4.2 Crenarchaeota
Most abundant archaea in the marine environment. They stain gram negativeand have diverse morphology. Examples include Sulfolobus solfataricus. Thisorganism was originally isolated from geothermally-heated sulfuric springs inItaly, and grows at 80 °C and pH of 2-4.
4.3 Korarchaeota
These organisms are known only from 16s ribosomal fragments found in envi-ronmental samples - whole individuals have never been recorded. Thus, nothingis currently known about their metabolic processes.
18

5 Bacteria
Bacteria are grouped into 23 phyla. They are ubiquitous in every habitat onEarth, forming the majority of the world's biomass.
5.1 The most important phyla
5.1.1 Proteobacteria
This is by far the biggest single phylum of Bacteria. The size of the group ismatched by its diversity, both morphological and physiological; most forms ofmetabolism are represented, and the wide range of morphological forms givesrise to the group's name. (Proteus was a mythological Greek god who was ableto assume many di�erent forms.) The reason such a diverse range of organismshave been assigned to a single taxonomic grouping is that their 16S rRNA indi-cates a common ancestor (thought to be photosynthetic, though few membersnow retain this ability). At the time of writing more than 460 genera and 1600species had been identi�ed, all of them Gram-negative and representing almosthalf of all accepted bacterial genera. These include many of the best knownGram-negative bacteria of medical, industrial and agricultural importance. Fortaxonomic purposes, the Proteobacteria have been divided into �ve classes re-�ecting their presumed lines of descent and termed the Alphaproteobacteria,Betaproteobacteria, Gammaproteobacteria, Deltaproteobacteria and Epsilon-proteobacteria.
Photosynthetic Proteobacteria:
The purple sulphur and purple non-sulphur bacteria are the only members ofthe Proteobacteria to have retained the photosynthetic ability of their presumedancestor. The type of photosynthesis they carry out, however, is quite distinctfrom that carried out by plants, algae and cyanobacteria, di�ering in two im-portant respects:
� it is anoxygenic � no oxygen is produced by the process
� it utilizes bacteriochlorophyll a and/or b, which have di�erent ab-sorbance properties to chlorophylls a and b.
Nitrifying Proteobacteria:
This group comprises aerobic Gram-negative chemolithoautotrophs that derivetheir energy from the oxidation of inorganic nitrogen compounds (either am-monia or nitrite), and their carbon from CO2 . The nitrifying bacteria arerepresented in both the α- and β-Proteobacteria. The oxidation of ammonia
19

through to nitrate is a two-stage process, with speci�c bacteria carrying outeach stage (ammonia to nitrite and nitrite to nitrate). This is re�ected in thegeneric names of the bacteria, bearing the pre�x Nitroso- or Nitro- accordingto whether they carry out the �rst or second reaction. Nitrifying bacteria playan essential role in the cycling of nitrogen in terrestrial, marine and freshwaterhabitats. Nitrite, which is toxic to many forms of life, rarely accumulates inthe environment, due to the activity of the Nitrobacteria. As was the case withthe purple photosynthetic bacteria, several cell forms are represented among thenitri�ers.
Iron- and sulphur-oxidising Proteobacteria
Two further groups of environmentally signi�cant chemolithoautotrophs derivetheir energy through the oxidation of reduced iron and sulphur respectively.
Hydrogen-oxidising Proteobacteria
This diverse group of bacteria are united by their ability to derive energy byusing hydrogen gas as a donor of electrons, and oxygen as an acceptor.
Nitrogen-�xing Proteobacteria
Nitrogen-�xing bacteria may be free-living in the soil (e.g. Azotobacter), orform a symbiotic relationship with cells on the root hairs of leguminous plantssuch as peas, beans and clover (e.g. Rhizobium).
Closely related to Rhizobium, but unable to �x nitrogen, are members of thegenus Agrobacterium. Like Rhizobium, these enter the tissues of plants, butinstead of forming a mutually bene�cial association, cause cell proliferation andtumour formation. A. tumefaciens has proved to be a valuable tool in the ge-netic engineering of plants.
Methanotrophic Proteobacteria
Methanotrophs are strict aerobes, requiring oxygen for the oxidation of methane.The methane-generating bacteria, however, are anaerobes; methanotrophs areconsequently to be found at aerobic/anaerobic interfaces such as topsoil, whereexist the both the oxygen and the methane they require. The methane is �rstlyoxidised to methanol, then to formaldehyde, by means of separate enzyme sys-tems. Some of the carbon in formaldehyde is assimilated into organic cellularmaterial, while some is further oxidised to carbon dioxide.
Enteric Proteobacteria
This is a large group of rod-shaped bacteria, mostly motile by means of per-itrichous �agella. They are facultative aerobes, characterised by their ability in
20

anaerobic conditions to carry out fermentation of glucose and other sugars togive a variety of products.
The most thoroughly studied of all bacteria, Escherichia coli is a member of thisgroup, as are a number of important pathogens of humans such as Salmonella,Shigella and Yersinia (the causative agent of plague).
Vibrio and related genera
A few other genera, including Vibrio and Aeromonas, are also facultative anaer-obes able to carry out the fermentative reactions described above, but are dif-ferentiated from the enteric bacteria by being oxidase-positive.
Vibrio and Photobacterium both include examples of marine bioluminescentspecies; these are widely found both in seawater and associated with �sh andother marine life. The luminescence, which requires the presence of oxygen,is due to an oxidation reaction carried out by the enzyme luciferase. Vibrio
cholerae is the causative agent of cholera, a debilitating and often fatal formof acute diarrhoea transmitted in faecally contaminated water. It remains amajor killer in many third world countries. Several species of Vibrio, includingV. cholerae, have been shown to possess two circular chromosomes instead ofthe usual one.
The Pseudomonads
Members of this group of proteobacteria, the most important genus of which isPseudomonas, are straight or curved rods with polar �agella. They are chemo-heterotrophs. A characteristic of many pseudomonads is the ability to utilise anextremely wide range of organic compounds (maybe over 100!) for carbon andenergy, something that makes them very important in the recycling of carbonin the environment.
Several species are signi�cant pathogens of animals and plants; Pseudomonasaeruginosa is an e�ective coloniser of wounds and burns in humans, while P.
syringae causes chlorosis (yellowing of leaves) in a range of plants. Because oftheir ability to grow at low temperatures, a number of pseudomonads are im-portant in the spoilage of food.
Predatory proteobacteria
Bdellovibrio is a unique genus. It is a very small comma-shaped bacterium,which actually attacks and lives inside other Gram-negative bacteria. Poweredby its �agellum, it collides with its prey at high speed and penetrates even thickcell walls by a combination of enzyme secretion and mechanical boring. It takesup residence in the periplasmic space, between the plasma membrane and cellwall. The host's nucleic acid and protein synthesis cease, and its macromoleculesare degraded, providing nutrients for the invader, which grows into a long helical
21

cell. This eventually divides into several motile progeny cells, which are thenreleased.
Another group of bacteria that may be regarded as predatory are theMyxobac-teria. These are rod-shaped bacteria lacking �agella, which yet are motile bygliding along a solid surface, aided by the excretion of extracellular polysaccha-rides. For this reason they are sometimes referred to as the gliding bacteria.They are heterotrophs, typically requiring complex organic nutrients, whichthey obtain by the lysis of other types of bacteria. Thus, unlike Bdellovibrio,they digest their prey before they ingest it. When a rich supply of nutrients isnot available, many thousands of cells may aggregate to form fruiting bodies,inside which myxospores develop. These are able to resist drought and lack ofnutrients for many years. Myxobacteria exhibit the most complex life cycles ofany procaryote so far studied.
Spirilla
Collected together under this heading are several genera of aerobic (mostlymicroaerophilic) spiral-shaped bacteria with polar �agella. These include free-living, symbiotic and parasitic types.
Spirilla such as Aquaspirillum and Magnetospirillum contain magnetosomes,intracellular particles of iron oxide (magnetite, Fe3O4). Such magnetotacticbacteria have the remarkable ability to orientate themselves with respect to theearth's magnetic �eld (magnetotaxis). Two important pathogens of humansare included in the spirilla; Campylobacter jejuni is responsible for foodbornegastroenteritis, while Helicobacter pylori has in recent times been identi�ed asthe cause of many cases of peptic ulcers.
Rickettsia
This group comprises arthropod-borne intracellular parasites of vertebrates, andincludes the causative agents of human diseases such as typhus and RockyMountain spotted fever. The bacteria are taken up by host phagocytic cells,where they multiply and eventually cause lysis.
Neisseria and related Proteobacteria
All members of this loose collection of bacteria are aerobic non-motile cocci,typically seen as pairs, with �attened sides where they join. Some however onlyassume this morphology during stationary growth phase. Many are found inwarm-blooded animals, and some species are pathogenic. The genus Neisseriaincludes species responsible for gonorrhoea and meningitis in humans.
5.1.2 Other Gram-negative phyla
1. Cyanobacteria - photosynthetic, probably the origin of plant chloro-
plasts
22

2. Chlamidiae - important in diseases including blindness and STIs
3. Spirochaetes - helical shape, corkscrew movement. One sp. causes
syphilis
5.1.3 Gram Positive Bacteria
Gram-positive bacteria are divided into two large phyla, the Firmicutes and theActinobacteria. Some 2500 species are known, but a substantial proportion ofthese belong to just a handful of genera. Gram-positive bacteria mostly have achemoheterotrophic mode of nutrition and include among their number severalimportant human pathogens, as well as industrially signi�cant forms.
The base composition of an organism's DNA can be expressed as the percentageof cytosine and guanine residues (per cent GC content); the technique is usedwidely in microbial taxonomy, and the Gram-positive bacteria are divided intothose whose GC content is signi�cantly over or under 50 per cent.
Phylum Firmicutes: The low GC Gram-positive bacteria
The spore-forming Gram-positive bacteria include two large genera, Clostridiumand Bacillus. Although not particularly close in phylogenetic terms, they areboth capable of propagation by endospores.
Several species of Clostridium are serious human pathogens including C. bo-
tulinum (botulism) and C. tetani (tetani). These conditions are due to theproduction of bacterial exotoxins.
Bacillus species are chemoautotrophs and are usually motile. Only a few speciesare pathogenic in humans, notably B. anthraxis (anthrax)
The non-spore-forming low GC Gram-positive bacteria include a number ofmedically and industrially signi�cant genera, a few of which are discussed below.The lactic acid bacteria are a taxonomically diverse group containing both rods(Lactobacillus) and cocci (Streptococcus, Lactococcus), all characterised by theirfermentative metabolism with lactic acid as end-product. Although they areable to tolerate oxygen, these bacteria do not use it in respiration. They aresaid to be aerotolerant.
The genus Streptococcus remains a large one, although some members havebeen assigned to new genera in recent years, e.g. Enterococcus, Lactococcus.
Streptococci are classi�ed in a number of ways on the basis of phenotypic char-acteristics. Many species produce haemolysis when grown on blood agar.
Phylum Actinobacteria: The high GC Gram-positive bacteria
The actinomycetes are aerobic, �lamentous bacteria that form branching myceliasuper�cially similar to those of the Fungi. Remember, however, that the acti-nomycetes are procaryotes and the fungi are eucaryotes, so the mycelia formed
23

by the former are appreciably smaller. The best known genus is Streptomyces.A high proportion of therapeutically useful antibiotics derive from Streptomyces
species.
5.2 Biochemistry (some important terms to learn)
5.2.1 Peptidoglycan
Peptidoglycan, also known as murein, is a polymer consisting of sugars andamino acids that forms a mesh-like layer outside the plasma membrane of bac-teria, forming the cell wall. The sugar component consists of alternating residuesof β-(1,4) linked N-acetylglucosamine and N-acetylmuramic acid residues.
Attached to the N-acetylmuramic acid is a peptide chain of three to �ve aminoacids. The peptide chain can be cross-linked to the peptide chain of anotherstrand forming the 3D mesh-like layer.
Some Archaea have a similar layer of pseudopeptidoglycan.
Peptidoglycan serves a structural role in the bacterial cell wall, giving structuralstrength, as well as counteracting the osmotic pressure of the cytoplasm. Acommon misconception is that peptidoglycan gives the cell its shape; however,whereas peptidoglycan helps maintain the structure of the cell, it is actually theMreB protein that facilitates cell shape.
Peptidoglycan is also involved in binary �ssion during bacterial cell reproduc-tion.
The peptidoglycan layer is substantially thicker in Gram-positive bacteria (20to 80 nanometers) than in Gram-negative bacteria (7 to 8 nanometers), with theattachment of the S-layer. Peptidoglycan forms around 90% of the dry weightof Gram-positive bacteria but only 10% of Gram-negative strains. In Gram-positive strains, it is important in attachment roles and stereotyping purposes.For both Gram-positive and Gram-negative bacteria, particles of approximately2 nm can pass through the peptidoglycan.
5.2.2 Exotoxins
An exotoxin is a toxin excreted by a microrganism, including bacteria, fungi,algae, and protozoa. An exotoxin can cause damage to the host by destroyingcells or disrupting normal cellular metabolism.
Both Gram negative and Gram positive bacteria produce exotoxins. They arehighly potent and can cause major damage to the host. Exotoxins may besecreted, or, similar to endotoxins, may be released during lysis of the cell.
Most exotoxins can be destroyed by heating. They may exert their e�ect locallyor produce systemic e�ects. Well known exotoxins include the botulinum toxin
24

produced by Clostridium botulinum, the Corynebacterium diphtheriae exotoxinwhich is produced during life threatening symptoms of diphtheria.
Exotoxins are susceptible to antibodies produced by the immune system, butmany exotoxins are so toxic that they may be fatal to the host before the immunesystem has a chance to mount defenses against it.
5.2.3 Endotoxins
Endotoxins (not to be confused with enterotoxin) are toxins associated with cer-tain bacteria. Classically, an "endotoxin" is a toxin which, unlike an "exotoxin",is not secreted in soluble form by live bacteria, but is a structural componentin the bacteria which is released mainly when bacteria are lysed.
The de�nitive examples of endotoxin are lipopolysaccharide (LPS) or lipo-oligo-saccharide (LOS) found in the outer membrane of variousGram-negativebacteria and is an important cause of their ability to cause disease. The termLPS is often used exchangeably with endotoxin, owing to its historical discovery.In the 1800s it became understood that bacteria could secrete toxins into theirenvironment, which became broadly known as "exotoxin". The term endotoxincame from the discovery that portions of Gram-negative bacteria itself can causetoxicity, hence the name endotoxin. Studies of endotoxin over the next 50 yearsrevealed that the e�ects of "endotoxin" were, in fact, due to lipopolysaccharide.
5.2.4 Enterotoxins
An enterotoxin is a protein toxin released by a microorganism in the intestine.
Enterotoxins are frequently cytotoxic and kill cells by altering the permeabilityof the epithelial cells of the intestinal wall. They are mostly pore forming toxins,secreted by bacteria, that assemble to form pores in cell membranes. This causesthe cells to die.
5.2.5 Bacteriocins
Bacteriocins are proteinaceous toxins produced by bacteria to inhibit the growthof similar or closely related bacterial strain(s). They are typically considered tobe narrow spectrum antibiotics, though this has been debated. They are struc-turally, functionally, and ecologically diverse. Bacteriocins were �rst discoveredby A. Gratia in 1925.
Bacteriocins are of interest in medicine because they are made by non-pathogenicbacteria that normally colonize the human body. Loss of these harmless bacteriafollowing antibiotic use may allow opportunistic pathogenic bacteria to invadethe human body.
25

5.2.6 Siderophores
A Siderophore (Greek for iron carrier) is an iron chelating (a type of binding)compound secreted by microorganisms such as bacteria and fungi. In responseto iron limitation in their environment, microbe siderophore production is dere-pressed. This is followed by excretion of the siderophore into the extracellularenvironment. Once outside the cell, the siderophore acts to sequester and solu-bilize the iron.
Siderophores e�ectively bind with iron by forming an octahedral siderophore-iron complex. Siderophores are then recognized by cell speci�c receptors onthe outer membrane of the cell. Following binding to these receptors they aretransported across the cell membrane by a number of processes including butnot limited to gating mechanisms and speci�c protein channels.
5.3 Reproduction
5.3.1 Binary Fission
Fig: Binary �ssion in prokaryotes
Bacteria grow to a �xed size and then reproduce through binary �ssion, a form ofasexual reproduction. Under optimal conditions, bacteria can grow and divideextremely rapidly, and bacterial populations can double as quickly as every9.8 minutes. Bacteria such as Pseudomonas and Clostridia spp. can have ageneration time of 10 mins in optimal conditions. The average bacterial gener-ation time (under ideal conditions) is around 20 minutes. In cell division, two
26

identical clone daughter cells are produced. Some bacteria, while still repro-ducing asexually, form more complex reproductive structures that help dispersethe newly-formed daughter cells. Examples include fruiting body formationby Myxobacteria and aerial hyphae formation by Streptomyces, or budding.Budding involves a cell forming a protrusion that breaks away and produces adaughter cell.
5.4 Endospores
Bacterial endospores are highly resistant to hostile physical and chemical con-ditions. Only a few genera of bacteria such as Bacillus and Clostridium arecapable of forming endospores. These are a dormant form of the bacteriumthat allows it to survive sub-optimal environmental conditions. Because thesespores are resistant to heat, radiation, disinfectants, and desiccation (dryingout), they are di�cult to eliminate from medical and pharmaceutical materialsand are a frequent cause of contamination.
Spores have a tough outer covering made of keratin and are highly resistant toheat and chemicals. The keratin also resists staining, so specialized proceduresare necessary to stain endospores:
� Malachite green stain is forced into the spore by heating the cells.
� Vegetative cells are then decolourised with water and stained pink withsafranin counterstain.
Endospores may be located in the middle of the cells (central), at the end(terminal), or between the end and the middle of the cells (subterminal). Theendospores themselves may be round or oval.
27
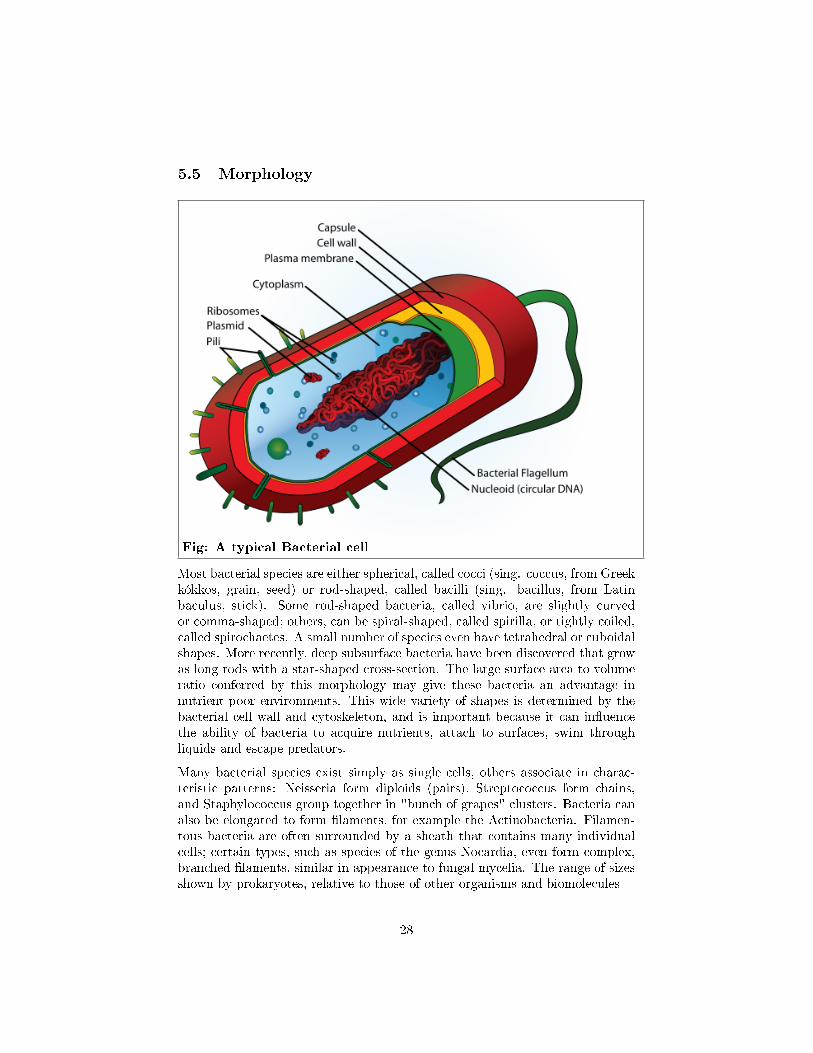
5.5 Morphology
Fig: A typical Bacterial cell
Most bacterial species are either spherical, called cocci (sing. coccus, from Greekkókkos, grain, seed) or rod-shaped, called bacilli (sing. bacillus, from Latinbaculus, stick). Some rod-shaped bacteria, called vibrio, are slightly curvedor comma-shaped; others, can be spiral-shaped, called spirilla, or tightly coiled,called spirochaetes. A small number of species even have tetrahedral or cuboidalshapes. More recently, deep subsurface bacteria have been discovered that growas long rods with a star-shaped cross-section. The large surface area to volumeratio conferred by this morphology may give these bacteria an advantage innutrient-poor environments. This wide variety of shapes is determined by thebacterial cell wall and cytoskeleton, and is important because it can in�uencethe ability of bacteria to acquire nutrients, attach to surfaces, swim throughliquids and escape predators.
Many bacterial species exist simply as single cells, others associate in charac-teristic patterns: Neisseria form diploids (pairs), Streptococcus form chains,and Staphylococcus group together in "bunch of grapes" clusters. Bacteria canalso be elongated to form �laments, for example the Actinobacteria. Filamen-tous bacteria are often surrounded by a sheath that contains many individualcells; certain types, such as species of the genus Nocardia, even form complex,branched �laments, similar in appearance to fungal mycelia. The range of sizesshown by prokaryotes, relative to those of other organisms and biomolecules
28

Bacteria often attach to surfaces and form dense aggregations called bio�lms orbacterial mats. These �lms can range from a few micrometers in thickness toup to half a meter in depth, and may contain multiple species of bacteria, pro-tists and archaea. Bacteria living in bio�lms display a complex arrangement ofcells and extracellular components, forming secondary structures such as micro-colonies, through which there are networks of channels to enable better di�usionof nutrients. In natural environments, such as soil or the surfaces of plants, themajority of bacteria are bound to surfaces in bio�lms. Bio�lms are also impor-tant in medicine, as these structures are often present during chronic bacterialinfections or in infections of implanted medical devices, and bacteria protectedwithin bio�lms are much harder to kill than individual isolated bacteria.
Even more complex morphological changes are sometimes possible. For exam-ple, when starved of amino acids, Myxobacteria detect surrounding cells in aprocess known as quorum sensing, migrate towards each other, and aggregate toform fruiting bodies up to 500 micrometres long and containing approximately100,000 bacterial cells. In these fruiting bodies, the bacteria perform separatetasks; this type of cooperation is a simple type of multicellular organisation.For example, about one in 10 cells migrate to the top of these fruiting bodiesand di�erentiate into a specialised dormant state called myxospores, which aremore resistant to drying and other adverse environmental conditions than areordinary cells.
29

Fig: Bacterial morphology. MAKE SURE YOU LEARN THIS, POS-SIBLE EASY MARKS!
5.6 Some important example Bacteria
Learning a bit about each of these bacteria should help guarantee you somemarks. Take special note of their main features, i.e.:
� are they gram+ve or -ve?
� what is their shape and size?
� are they �agellated?
� where are they most often encountered and why are they important?
I've tried to include the most important stu�, but reading around the linksprovided will help you gain a good understanding.
30

5.6.1 Escherichia coli
E. coli is Gram-negative, facultative anaerobic and non-sporulating. The bestunderstood bacterium, it is an inhabitant of the human gut, and an occa-sional pathogen. It is also used for the manipulation of genes from all andsundry other organisms. It is rod-shaped, �agellated, and approx. 3µm (mi-crons/micrometres) in length. When binary �ssion occurs, the cells dividelength-ways (see �g).
5.6.2 Staphylococcus aureus
S. aureus is a facultatively anaerobic, Gram-positive coccus, which appearsas grape-like clusters when viewed through a microscope and has large, round,golden-yellow colonies, often with haemolysis, when grown on blood agar plates.The bacterial cells are approximately 1 micron in diameter.
A spherical bacterium, Staphylococcus aureus is frequently found in the noseand skin of a person. About 20% of the population are long-term carriers ofS. aureus. S. aureus can cause a range of illnesses from minor skin infections,such as pimples, impetigo (may also be caused by Streptococcus pyogenes), boils,cellulitis folliculitis, furuncles, carbuncles, scalded skin syndrome and abscesses,to life-threatening diseases such as pneumonia, meningitis, osteomyelitis endo-carditis, Toxic shock syndrome (TSS), and septicemia.
S. aureus is catalase positive (meaning that it can produce the enzyme "cata-lase") and able to convert hydrogen peroxide (H2O2) to water and oxygen, whichmakes the catalase test useful to distinguish staphylococci from enterococci andstreptococci.
31

5.6.3 Pseudomonas aeruginosa
Like other members of the genus, Pseudomonas aeruginosa is a free-living bac-terium, commonly found in soil and water. However, it occurs regularly onthe surfaces of plants and occasionally on the surfaces of animals. Members ofthe genus are well known to plant microbiologists because they are one of thefew groups of bacteria that are true pathogens of plants. In fact, Pseudomonasaeruginosa is occasionally a pathogen of plants. However, Pseudomonas aerugi-nosa has become increasingly recognized as an emerging opportunistic pathogenof clinical relevance. Several di�erent epidemiological studies track its occur-rence as a nosocomial pathogen and indicate that antibiotic resistance is in-creasing in clinical isolates.
Pseudomonas aeruginosa is an opportunistic pathogen, meaning that it exploitssome break in the host defenses to initiate an infection. In fact, Pseudomonasaeruginosa is the epitome of an opportunistic pathogen of humans. The bac-terium almost never infects uncompromised tissues, yet there is hardly any tissuethat it cannot infect if the tissue defenses are compromised in some manner. Itcauses urinary tract infections, respiratory system infections, dermatitis, softtissue infections, bacteremia, bone and joint infections, gastrointestinal infec-tions and a variety of systemic infections, particularly in patients with severeburns and in cancer and AIDS patients who are immunosuppressed.
Pseudomonas aeruginosa is a Gram-negative rod measuring 0.5 to 0.8 µm by1.5 to 3.0 µm. Almost all strains are motile by means of a single polar �agellum.
The bacterium is very common in soil and water, and on surfaces in contactwith soil or water. Its metabolism is respiratory and never fermentative, butit will grow in the absence of O2 if NO3 is available as a respiratory electronacceptor.
The typical Pseudomonas bacterium in nature might be found in a bio�lm,attached to some surface or substrate, or in a planktonic form, as a unicellularorganism, actively swimming by means of its �agellum. Pseudomonas is one ofthe most vigorous, fast-swimming bacteria seen in hay infusions and pond watersamples.
32

In its natural habitat Pseudomonas aeruginosa is not particularly distinctive asa pseudomonad, but it does have a combination of physiological traits that arenoteworthy and may relate to its pathogenesis.
� Pseudomonas aeruginosa has very simple nutritional requirements. It is oftenobserved "growing in distilled water", which is evidence of its minimal nutri-tional needs. In the laboratory, the simplest medium for growth of Pseudomonasaeruginosa consists of acetate as a source of carbon and ammonium sulfate asa source of nitrogen.
� P. aeruginosa possesses the metabolic versatility for which pseudomonads areso renowned. Organic growth factors are not required, and it can use more thanseventy-�ve organic compounds for growth.
� Its optimum temperature for growth is 37 degrees, and it is able to grow attemperatures as high as 42 degrees.
� It is tolerant to a wide variety of physical conditions, including temperature.It is resistant to high concentrations of salts and dyes, weak antiseptics, andmany commonly used antibiotics.
� Pseudomonas aeruginosa has a predilection for growth in moist environments,which is probably a re�ection of its natural existence in soil and water.
5.6.4 Enterococcus faecalis
Enterococcus faecalis is a nonmotile, gram-positive, (sort of) spherical bac-terium. It can be observed singly, in pairs, or in short chains, and is mostoften found in the large intestine of humans. It is a facultative anaerobe with afermentative metabolism. It can often be confused with S. pneumonia, but E.faecalis contains many identi�cation features that can be veri�ed with testing.
33

E. faecalis is listed as the �rst to the third leading cause of nosocomial infections.Most of these infections occur after surgery of the abdomen or a puncturingtrauma, but can also be linked to the increased use of IV's and catheters, whichare considered compromising devises. It is also responsible for urinary tractinfections, bacterimia, endocarditis, meningitis, and can be found in woundinfections along with many other bacteria.
E. faecalis was �rst identi�ed as a Strep D bacterium (Streptococcus faecalis)because of its characteristic Strep D speci�c cell wall carbohydrate. It wasn'tuntil 1984, that it was classi�ed as an Enterococcus. E. faecalis is among themost antibiotic resistant bacteria known. It contains many natural antibioticresistances, along with several acquired immunities transferred in R-plasmidsamong promiscuous bacteria. More than 25% of the E. faecalis genome is ex-ogenously acquired, leading to its resistance to the strongest antibiotics and insome cases all antibiotics. It is also considered to be a carrier of vancomycinresistance for other genera of bacteria.
5.6.5 Bacillus subtilis
Bacillus subtilis is a Gram-positive, rod-shaped and endospore-forming aerobicbacterium. It is found in soil and rotting plant material and is non-pathogenic.It is one of the most studied gram-positive bacteria. One feature that hasattracted a lot of interest in B. subtilis is its ability to di�erentiate and formendospores.
34

6 Fungi
We may de�ne true fungi as primarily terrestrial, spore-bearing organisms, lack-ing chlorophyll and having a heterotrophic, absorptive mode of nutrition. Some80,000 species are known and it is thought possible that at least a million moreremain to be described! True fungi are a monophyletic group; that is, theyare all thought to descend from a common ancestor, some 550 million years ago.
6.1 Importance
Fungi are of great importance economically and socially, and may have ben-e�cial or detrimental e�ects. Many fungi, particularly yeasts, are involved inindustrial fermentation processes. These include, for example, the production ofbread and alcohol, while other fungi are essential to the cheese-making process.Many antibiotics, including penicillin, derive from fungi, as does the immuno-suppressive drug cyclosporin.
Along with bacteria, fungi are responsible for the decomposition and reprocess-ing of vast amounts of complex organic matter; some of this is recycled to theatmosphere as CO2, while much is rendered into a form that can be utilised byother organisms. The other side of this coin is seen in the activity of fungi thatdegrade and destroy materials of economic importance such as wood, paper andleather, employing essentially the same biochemical processes.
Additionally, some fungi may cause disease; huge damage is caused to cropsand other commercially valuable plants, while a number of human diseases,particularly of the skin and scalp, are also caused by fungi.
6.2 Morphology
Fungi range in size and shape from unicellular yeast forms to large multicellularmushrooms.
6.2.1 Yeasts
Yeasts are unicellular. They are non-motile, lacking �agella. They reproduceeither:
� asexually, by budding or transverse �ssions
� sexually, by spore formation
35
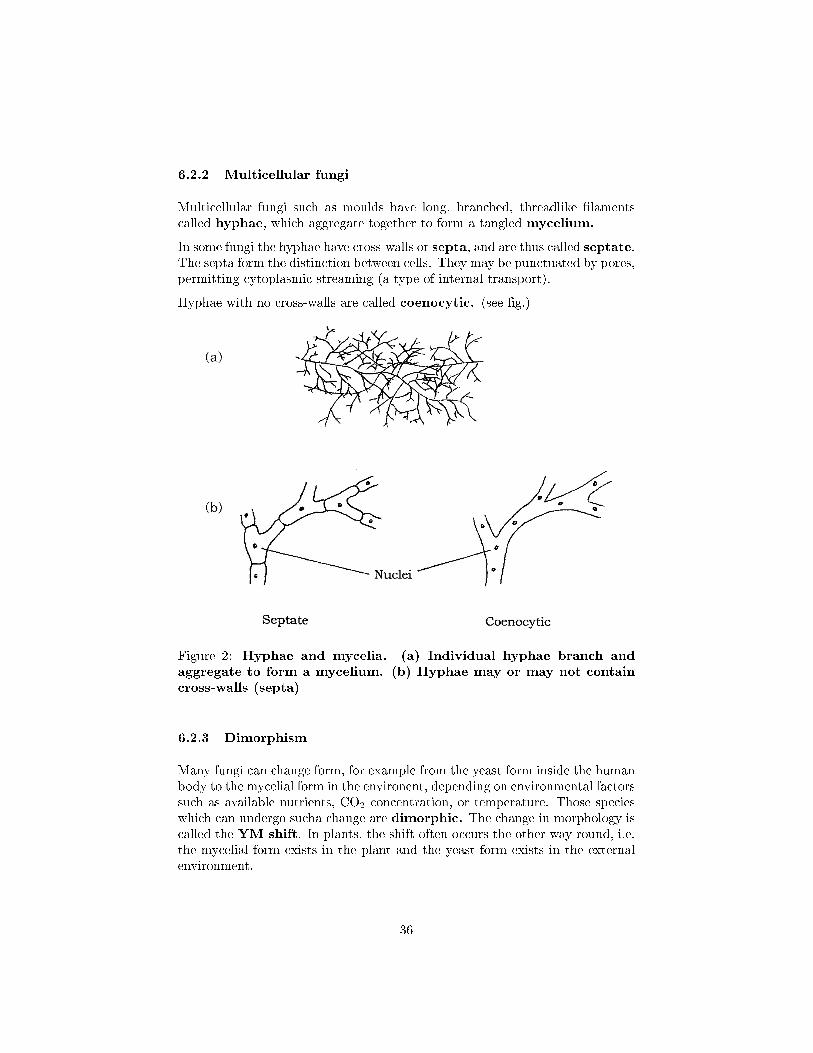
6.2.2 Multicellular fungi
Multicellular fungi such as moulds have long, branched, threadlike �lamentscalled hyphae, which aggregate together to form a tangled mycelium.
In some fungi the hyphae have cross-walls or septa, and are thus called septate.The septa form the distinction between cells. They may be punctuated by pores,permitting cytoplasmic streaming (a type of internal transport).
Hyphae with no cross-walls are called coenocytic. (see �g.)
Figure 2: Hyphae and mycelia. (a) Individual hyphae branch andaggregate to form a mycelium. (b) Hyphae may or may not containcross-walls (septa)
6.2.3 Dimorphism
Many fungi can change form, for example from the yeast form inside the humanbody to the mycelial form in the environent, depending on environmental factorssuch as available nutrients, CO2 concentration, or temperature. Those specieswhich can undergo sucha change are dimorphic. The change in morphology iscalled the YM shift. In plants, the shift often occurs the other way round, i.e.the mycelial form exists in the plant and the yeast form exists in the externalenvironment.
36

6.3 Structure
Whereas plant and algal cell walls are made up of cellulose, fungal cell wallsare made of chitin (the same stu� as arthopod exoskeletons!). It is a polymerwhose repeating subunit is N-acetylglucosamine.
6.4 Nutrition
Most fungi are saprobic, that is they obtain their nutrition from decaying mat-ter, which they grow over and through, frequently secreting enzymes extracel-lularly to break down complex molecules to simpler forms that can be absorbedby the hyphae. Some species can be symbiotic, eg. in lichens, and some areparasitic on a host organism. Most fungi are able to synthesize their own aminoacids and proteins from carbohydrates and simple nitrogenous compounds. Al-though fungi are unable to move, they can swiftly colonise new territory as aresult of the rapid rate at which their hyphae grow. All energy is concentratedon adding length rather than thickness; this growth pattern leads to an increasein surface area and is an adaptation to an absorbtive way of life. Carbohydratesare stored mainly in the form of glycogen. Metabolism is mainly aerobic, butsome yeasts can function as facultative anaerobes.
6.5 Reproduction
Although there is great variety among the patterns of reproduction found inthe fungi, all share in common the pattern of reproducing by spores. Theseare non-motile reproductive cells that rely on being carried by animals or onthe wind for their dispersal. The hyphae that bear the spores usually projectup into the air, aiding dispersal. One of the main reasons we have to practiceaseptic technique in the lab is that fungal spores are pretty well ubiquitous(found everywhere) - they will germinate and grow anywhere they can �nd asuitable medium. Spores of the common black bread mould Rhizopus have beenfound in the air over the North Pole, and hundreds of miles out to sea.
In some fungi the aerial spore-bearing hyphae are developed into large complexstructures called fruiting bodies. The most familiar example of a fruiting bodyis the mushroom. Many people think that the mushroom itself is the wholefungus but it only represents a part of it; most is buried away out of sight belowthe surface of the soil or rotting material, a network of nearly invisible hyphae.
6.6 Classi�cation
6.6.1 Zygomycota
The Zygomycota is a relatively small phylum, comprising less than a thousandspecies. Its members are typically found in soil, or on decaying organic matter,
37

including animal droppings. Some members of the group are of great impor-tance in the formation of a mutualistic association with plant roots known as amycorrhiza. Members of the Zygomycota are characterised by the formation ofa dormant form, the zygospore, which is resistant to unfavourable environmen-tal conditions. Hyphae are coenocytic, with numerous haploid nuclei, but fewdividing walls or septa.
Familiar examples of this group areMucor and the black bread mould Rhizopus.
6.6.2 Chytridiomycota
The chytrids are believed to have been the �rst of the fungal groups to divergefrom a common ancestor many millions of years ago. They di�er from all otherfungal groups by possessing �agellated zoospores. At one time, the Fungiwere de�ned by their lack of �agella, so the chytrids were assigned to the Pro-tista. However, molecular evidence, including the possession of a chitinous cellwall, suggests that it would be more appropriate to place them among the Fungi.
Some members of the chytrids may live saprobically on decaying plant andanimal matter, while others are parasites of plants and algae. Another group liveanaerobically in the rumen of animals such as sheep and cattle. In recent yearsthere has been evidence that a parasitic species of chytrid is at least partiallyresponsible for the dramatic decline in frog populations in certain parts of theworld. Some chytrids are unicellular, while others form mycelia of coenocytichyphae. Reproduction may be asexual by means of motile zoospores, or sexual.The latter may involve fusion of gametes to produce a diploid zygote, but thereis no dicaryotic stage in the life cycle.
6.6.3 Ascomycota
The Ascomycota are characterised by the production of haploid ascospores
through the meiosis of a diploid nucleus in a small sac called an ascus. Forthis reason they are sometimes called the sac fungi or cap fungi. Many of thefungi that cause serious plant diseases such as Dutch elm disease and powderymildew belong to this group. They include some 30,000 species, among themyeasts, food spoilage moulds, brown fruit rotting fungi and tru�es.
Around half of ascomycote species exist in association with algae to form lichens.Most ascomycetes produce mycelia that super�cially resemly those of zygomycetes,but di�er in that they have distinct, albeit perforated cross walls (septa) sepa-rating each cell.
Asexual reproduction in most ascomycetes involves the production of airbornespores called conidia. These are carried on the ends of specialised hyphaecalled conidiophores, where they may be pinched o� as chains or clusters.Note that the conidia are not contained within sporangia; they may be naked orprotected by a �ask-like structure called the pycnidium. Asexual reproduction
38

by conidia formation is a means of rapid propagation for the fungus in favourableconditions. the characteristic green, pink or brown colour of many moulds isdue to the pigmentation of the conidia, which are produced in huge numbersand dispersed by air currents. The conidia germinate to form another mycelium(haploid).
In the case of unicellular yeasts, asexual reproduction occurs as the results ofbudding, a pinching o� of a protuberance from the cell, which eventually growsto full size.
Sexual reproduction in some ascomycetes involves separate + and − matingstrains similar to those seen in sygomycetes, whilst in other cases an individualwill be self-fertile, and thus able to mate with itself.
6.6.4 Basidiomycota
This large group of some 25,000 species contains the true mushrooms and toad-stools as well as other familiar fungi such as pu�balls and bracket fungi. In factthe great majority of the fungi that we see in �elds and woodlands belong tothe Basidiomycota. They are of great economic importance in the breakdownof wood and other plant material. The group derives its common name of theclub fungi from the way that the spore-bearing hyphae involved in reproductionare swollen at the tips, resembling clubs.
Asexual reproduction occurs much less frequently in basidiomycetes than in theother types of fungi. When it does occur, it is generally by means of conidia, al-though some types are capable of fragmenting their hyphae into individual cells,each of which then acts like a spore and germinates to form a new mycelium.
Sexual reproduction in a typical mushroom involves the fusion of haploid hyphaebelonging to two compatible mating types to produce a dicaryotic myceliumin which each cell has two haploid nuclei. The most striking feature of thissecondary mycelium is the clamp connection; this is unique to the Basidiomy-cota and is a device for ensuring that as growth continues, each new cell hasone nucleus from each of the parent mating strains. This dicaryotic secondarymycelium continues to grow, overwhelming any remaining haploid hyphae fromthe parent fungi.
When the secondary mycelium has been developing for some time, it formsa dense compact ball or button, which pushes up just above the surface andexpands into a basidiocarp; this is the mushroom itself. Stalk formation andupward growth is extremely rapid; a stalk or stipe of 10 cm can be formed in onlyabout 6�9 hours. The growth is initially towards light (positive phototropism)and then upward (negative geotropism).
As the cap expands, �eshy �aps radiating from the centre of its underside openup. These are the gills, made up of compacted hyphae with numerous basidiaarranged at right angles. As each basidium matures, its two nuclei �nally fuse,and then undergo meiosis to produce four haploid basidiospores.
39

A single large mushroom can produce millions of basidiospores in the space ofa few days. They are discharged from the end of the basidia and then fall bygravity from the gills. Air currents then carry them away for dispersal. Upon�nding a suitable substratum, the spores germinate into a haploid myceliumjust below the surface of the soil, thus completing the life cycle.
40

7 Virii
All viruses are obligate intracellular parasites; they inhabit a no-man's-landbetween the living and the non-living worlds, and possess characteristics ofboth. They are now known to di�er radically from the simplest true organisms,bacteria, in a number of respects:
� they cannot be observed using a light microscope
� they have no internal cellular structure
� they contain either DNA or RNA, but not both
� they are incapable of replication unless occupying an appropriate livinghost cell
� they are incapable of metabolism
� individuals show no increase in size.
When inside a host cell, viruses show some of the features of a living organism,such as the ability to replicate themselves, but outside the cell they are justinert chemical tructures, thus fuelling the debate as to whether they can beconsidered to be life forms. A particular virus has a limited host range, that is,it is only able to infect certain cell types. Nobody is sure how viruses evolved.
7.1 Structure
The demonstration by Wendel Stanley in 1935 that a preparation of tobaccomosaic virus could be crystallised was an indication of the relative chemicalhomogeneity of viruses, and meant that they could not be thought of in thesame terms as other living things. Compared to even the most primitive cellularorganism, viruses have a very simple structure. An intact viral particle, orvirion, has in essence just two components: a core of nucleic acid, surroundedand protected by a protein coat or capsid, the combination of the two beingknown as the nucleocapsid. In certain virus types, the nucleocapsid is furthersurrounded by a membranous envelope, partly derived from host cell material.Most viruses are smaller than even the smallest bacterial cells.
7.2 Viral genome
The genetic material of a virus may be either RNA or DNA, and either of thesemay be single-stranded or double-stranded. The genome may furthermore becircular or linear. An additional variation in the viral genome is seen in certainRNA viruses, such as the in�uenza virus; here, instead of existing as a single
41

molecule, it is segmented, existing as several pieces, each of which may encodea separate protein. In some plant viruses, the segments may be present inseparate particles, so in order for replication to occur, a number of virions needto co-infect a cell, thereby complementing each other (multipartite genomes)!Double-stranded RNA is always present in the segmented form.
The size of the genome varies greatly; it may contain as few as four genes or asmany as over 200. These genes may code for both structural and non-structuralproteins; the latter include enzymes such as RNA/DNA polymerases requiredfor viral replication.
7.3 Bacteriophages
Viruses that infect bacterial cells are called bacteriophages (phages for short),which means, literally, `bacteria eaters'. Perhaps the best understood of allviral replication cycles are those of a class of bacteriophages which infect E.
coli, known as the T-even phages. These are large, complex viruses, with acharacteristic head and tail structure. The double-stranded, linear DNA genomecontains over 100 genes, and is contained within the icosahedral head. Thegrowth cycle is said to be lytic, because it culminates in the lysis (=bursting)of the host cell.
7.3.1 Viroids and Prions
Although some viruses are exceedingly small and simple, even simpler infectiondiseases exist. Over 16 di�erent plant diseases - for example, potato spindle-tuber disease - are cuased by a group of inectious agents called viroids. Theseare circular, single-stranded RNAs, about 250 to 370 nucleotides long, that canbe transmitted between plants by mechanical means or by way of pollen andovules are are replicated in their hosts. The single-stranded RNs normally existsas a closed circle collapsed into a rodlike shape by intrastrand base pairing.Viroids are found principally in the nucleolus of infected cells; between 200and 10,000 copies may be present. A plant may be growing without showingsymptoms - that is, it may have a latent infection. The same viroid, when inother species, might cause a disease.
There is evidence than an infectious agent di�erent from both viruses and vi-roids can cause disease in livestock and humans. The agent has been calleda prion (for proteinaceous infectious particle). The best studied of these pri-ons causes a degenerative disorder of the central nervous system in sheeps andgoats; this disorder is named scrapie. A�icted animals lose coordination oftheir movements, tend to scrape or rub their skin, and eventually cannot walk.No nucleic acid has yet been detected in the agent. It seems to be a membraneprotein, often called PrP (for prion protein). The PrP gene is present in manynormal vertebrates and invertebrates, and the prion protein is bound to the
42

surface of neurons. Presumably an altered PrP is at least partly responsible forthe disease.
Prions have also been called virinos andNemavirus in the scienti�c literature,and in some works and textbooks they are used interchangably.
Some slow virus diseases may be due to prions. This is particularly true ofcertain neurological diseases of humans and animals. Bovine spongiformencephalopathy (BSE or �mad cow disease�), kuru, fatal familial insom-nia, the Creutzfeldt-Jakob disease (CJD), and Gerstmann-Sträussler-Scheinker syndrome (GSS) appear to be prion diseases. They result inprogressive degenration of the brain and eventual death. It has been shownthat eating meat from cattle with BSE can cause a variant of Creutzfeldt-Jacobdisease in humans.
43

8 Protista
The Protista represents a very diverse group of organisms, united by their pos-session of eucaryotic characteristics, and failure to �t satisfactorily into theanimal, plant or fungal kingdoms. Some scientists limit use of the name to uni-cellular organisms, while others also include organisms such as the macroscopicalgae, which are not accommodated conveniently elsewhere.
It has been found helpful in the past to think of Protists as being dividedinto those with characteristics that are plant�like (the Algae), animal-like (theProtozoa) and fungus-like (the water moulds and slime moulds). It should beborne in mind, however, that molecular evidence suggests such a division to bearti�cial; on the basis of molecular and cytological comparison, the `animal-like'protozoan Trypanosoma, for example, is closely related to the photosynthetic(and therefore `plant-like') Euglena.
8.1 The 'Algae'
The Algae is a collective name traditionally given to several phyla of primitive,and mostly aquatic plants, making up a highly diverse group of over 30,000species. They display a wide variety of structure, habitat and life-cycle, rangingfrom single-celled forms to massive seaweeds tens of metres in length. Mostalgae share a number of common features which caused them to be groupedtogether. Among these are:
� possession of the pigment chlorophyll
� deriving energy from the sun by means of oxygenic photosynthesis
� �xing carbon from CO2 or dissolved bicarbonate
In this discussion, multicellular forms, including the Phaeophyta (brown algae)and Rhodophyta (red algae), are not discussed at great length and are includedfor the sake of completeness.
8.1.1 Structural characteristics of algal protists
All algal types are eucaryotic, and therefore contain the internal organelles stan-dard to most eukaryotes: nuclei, mitochondria, endoplasmic reticulum, ribo-somes, Golgi body, and in most instances, chloroplasts. With the exceptionof one group (the Euglenophyta), all have a cellulose cell wall, which is fre-quently modi�ed with other polysaccharides, including pectin and alginic acids.In some cases, the cell wall may be forti�ed with deposits of calcium carbonateor silica. This is permeable to small molecules and ions, but impermeable tomacromolecules.
44

8.1.2 Euglenophyta
The is a group of unicellular �agellated organisms, which probably represent themost ancient group of algal protists. Individuals range in size from 10-500µm.Euglenophytes are commonly found in fresh water, particularly that with a highorganic content, and to a lesser extent, in soil, brackish water and salt water.Members of this group have a well-de�ned nucleus, and chloroplasts containingchlorophylls a and b.
The storage product of photosynthesis is a β-1,3-linked glucan called paramy-lon, found almost exclusively in this group. Euglenophytes lack a cellulose cellwall but have instead, situated within the plasma membrane, a �exible pelliclemade up of interlocking protein strips, a characteristic which links them to cer-tain protozoan species. A further similarity is the way in which locomotion isachieved by the undulation of a terminal �agellum. Movement towards a lightsource is facilitated in many euglenids by two structures situated near the baseof the �agellum; these are the para�agellar body and the stigma or eyespot.The latter is particularly conspicuous, as it is typically an orange-red colour,and relatively large.
Figure 3: Euglena. Euglena has a number of features in common with thezoo�agellates, but its possession of chloroplasts has meant that it has tradition-ally been classi�ed among the algae
Reproduction is by binary �ssion (i.e. by asexual means only). Division starts atthe anterior end, and proceeds longitudinally down the length of the cell, givingthe cell a characteristic `two-headed' appearance. During mitosis, the chromo-somes within the nucleus replicate, forming pairs that split longitudinally. Sincethe euglenophyte is usually haploid, it thus becomes diploid for a short period.As �ssion proceeds, one daughter cell retains the old �agellum, while the otherone generates a new one later. As in the binary �ssion of bacteria, the progenyare genetically identical, i.e. clones.
45

When conditions are unfavourable for survival due to failing nutrient supplies,the cells round up to form cysts surrounded by a gelatinous covering; these havean increased complement of paramylon granules, but no �agella. An importantrespect in which euglenids may be at variance with the notion of `plant-likeprotists' is their ability to exist as heterotrophs under certain conditions. Whenthis happens, they lose their photosynthetic pigments and feed saprobically ondead organic material in the water.
8.2 Dino�agellata
The dino�agellates (also known variously as Pyrrophyta, or `�re algae') arechie�y marine planktonic types, comprising some 2000 species. This is anotherunicellular group, but one whose cells are often covered with armoured platesknown as thecae (sing: theca). They are generally bi�agellate, with the twodissimilar �agella lying in part within the longitudinal and lateral grooves thatrun around the cell. The beating of the �agella causes the cell to spin like a topas it moves through the water (the group takes its name from the Greek word`to whirl').
Although many non-photosynthetic (chemoheterotrophic) types exist, most di-no�agellates are photosynthetic, containing chlorophyll a and c plus certaincarotenoids and xanthophylls, which give them a red/golden appearance. As agroup, they are second only to the diatoms (see below) as the primary photo-synthetic producers in the marine environment. Some dino�agellates form en-dosymbiotic relationships with marine animals such as corals and sea anemones;these are termed zooxanthellae.
An unusual feature of dino�agellate ultra-structure is that the chromosomescontain little, if any, histone protein, and exist almost permanently in the con-densed form. Some tropical species of dino�agellate emit light, the only algaeto do so. Due to an enzyme�substrate (luciferin�luciferase) interaction, this cancause a spectacular glow in the water at night, especially when the water isdisturbed, for example by a ship. Bioluminescence of this kind has proved tobe a useful `tagging' system for cells in biological research.
Other marine dino�agellates can produce metabolites that act as nerve toxinsto higher animals. Shell�sh such as mussels and oysters can concentrate thesewith no harm to themselves, but they can be fatal to humans who consumethem.
Sometimes, when conditions are highly favourable, an explosion of growth re-sults in the development of huge `red tides' of dino�agellates in coastal waters.This produces a build-up of toxins, and may lead to the death of massive num-bers of �sh and other marine life. The greatly increased incidence of theseblooms in recent decades is probably due to pollution by fertilisers containingnitrates and phosphates.
46

Reproduction by asexual means involves binary �ssion. In armoured forms, thetheca may be shed before cell division, or split along suture lines; in eithercase, daughter cells must regenerate the missing sections. Sexual reproductionis known to occur in some dino�agellates, and is probably more widespread.Gametes produced by mitosis fuse to produce a diploid zygote; this undergoesmeiosis to reinstate the haploid condition in the o�spring. In some species wesee isogamy, the fusion of identical, motile gametes, while in others, anisogamyoccurs, in which gametes of dissimilar size fuse. Fusion may occur betweengenetically identical gametes, or only when the gametes come from geneticallydistinct populations.
8.3 Diatoms
The diatoms, which belong to the division Chrysophyta (the golden-brown al-gae), make up the majority of phytoplankton in marine food chains, and assuch are the most important group of algal protists in terms of photosyntheticproduction. Over 10,000 species of diatom are recognised, but some experts feelthat the real number is many times greater than this.
As with the dino�agellates, chlorophylls a and c are present, but not chlorophyllb. Their colour is due to carotenoids and xanthophylls (particularly fucoxanthin)masking the chlorophyll. Diatoms have their cells surrounded by a silica-basedshell known as a frustule, composed of two overlapping halves (the epithecaand the hypotheca). Microbiologists are rarely able to resist the temptation toliken this structure to that of a petri dish, and with good reason. With theelectron microscope it can be seen that the frustule is perforated with numeroustiny pores that connect the protoplast of the cell with the outside environment.
Diatom classi�cation is based almost entirely on the shape and pattern of theseshells, which are uniform for a particular species, and often have a very strikingappearance. When diatoms die, their shells fall to the bottom of the sea, andcan accumulate in thick layers where they represent a valuable mineral resource.This �ne, light material (diatomaceous earth) has a number of applications, forexample in �ltration systems, and also as a light abrasive in products such assilver polish or toothpaste.
Reproduction is usually asexual by binary �ssion, but a sexual phase with theproduction of haploid gametes can occur. Chrysophytes are unusual among thethree primitive groups of algae in that they are diploid. In diatoms, asexualreproduction involves mitotic cell division, with each daughter cell receivingone half of the parental frustule, and synthesizing a new one to complementit. The newly formed half, however, always acts as the hypotheca (lower half)of the new cell; consequently, one in two daughter cells will be slightly smallerthan the parent, an e�ect which is heightened over a number of generations.This process continues until a critical size is reached, and the diatoms undergoa phase of sexual reproduction, which re-establishes the normal frustule size. In
47

species whose frustules have a degree of elasticity, the daughter cells are able toexpand, and the problem of cell diminution does not arise.
In bilaterally symmetrical (long, thin) diatoms, meiosis in parental cells pro-duces identical, non-motile gametes, which fuse to form a zygote. The radiallysymmetrical (round) forms provide an example of the third pattern of gametefusion found in the algae: oogamy. Here, there is a clear distinction between thesmall, motile sperm cell and the larger, immobile egg cell. Both are produced bymeiosis in the parental cell, followed, in the case of the male, by several roundsof mitosis, to give a large number of sperm cells.
8.4 Chlorophyta
The green algae have always attracted a lot of interest because, as a group, theyshare a good deal in common with the higher plants in terms of ultrastructure,metabolism and photosynthetic pigments, pointing to the likelihood of a com-mon ancestor. They possess both chlorophyll a and b and certain carotenoids,store carbohydrate in the form of starch, and generally have a rigid cell wallcontaining cellulose. The starch is stored in structures called pyrenoids, whichare found within chloroplasts. There are two phylogenetically distinct lines ofgreen algae, the Charophyta and the Chlorophyta; the latter are much the biggergroup, but the charophytes seem to be more closely related to green plants.
Chlorophytes demonstrate a wide variety of body forms, ranging from unicel-lular types to colonial, �lamentous, membranous and tubular forms. The vastmajority of species are freshwater aquatic, but a few marine and pseudoterres-trial representatives exist.
The genus usually chosen to illustrate the unicellular condition in chlorophytesis Chlamydomonas. This has a single chloroplast, similar in structure and shapeto that of a higher plant, and containing a pyrenoid. Situated together at theanterior end is a pair of smooth or whiplash �agella, whose regular, orderedcontractions propel it through the water. A further structural feature foundin Chlamydomonas and other motile forms of green algae is the stigma or eye-spot; this is made up of granules of a carotenoid pigment and is at least partiallyresponsible for orienting the cell with respect to light.
Reproduction in Chlamydomonas and other unicellular types under favourableconditions of light, temperature and nutrients, occurs asexually by the produc-tion of zoospores. A single haploid adult loses its �agella and undergoes mitosisto produce several daughter cells, which then secrete cell walls and �agella andtake up an independent existence of their own. This can result in a tremendousincrease in numbers; a single cell can divide as many as eight times in one day.Sexual reproduction in Chlamydomonas, which occurs when conditions are lessfavourable, di�ers in detail according to the species.
Any one of the three variants of gamete production seen in the algae may be seen:isogamy, anisogamy and oogamy. In all cases, two haploid gametes undergo a
48

fusion of both cytoplasm and nuclei to give a diploid zygote. The gametes maysimply be unmodi�ed haploid adult cells, or they may arise through mitoticcleavage of the adult, depending on the species.
8.5 Rhodophyta
The red coloration of the rhodophytes is due to the pigments phycoerythrinand phycocyanin, which mask the chlorophylls present, in this case a and d.The biggest single di�erence between the red algae and the other groups wehave looked at so far is that they lack �agella at any stage of their life cycle.Thus they are completely lacking in any motile forms, even in the reproductivestages; the gametes rely on being passively dispersed. Almost all the red algaeare multicellular marine species, inhabiting habitats ranging from shallow rockpools to the ocean's deeps.
Red algae are the source of several complex polysaccharides of commercial value.Agar and agarose are used in the laboratory in microbial growth media andelectrophoresis gels respectively, whilst carrageenan is an important thickeningagent in the food industry. In addition, Porphyra species are cultivated in Japanfor use in sushi dishes.
8.6 `The Protozoa'
The name Protozoa comes from the Greek, meaning `�rst animal', and was orig-inally applied to single-celled organisms regarded as having animal-like charac-teristics (multicellular animals were termed Metazoa). Protozoans as a grouphave evolved an amazing range of variations on the single-celled form, partic-ularly with respect to the di�erent means of achieving movement. they are amorphologically diverse group of well over 50,000 species; although the majorityare free-living, the group also includes commensal forms and some extremelyimportant parasites of animals and humans.
Most protozoans are found in freshwater or marine habitats, where they forma signi�cant component of plankton, and represent an important link in thefood chain. Although water is essential for the survival of protozoans, many areterrestrial, living saprobically in moist soil.
A protozoan needs to pack all the functions of an entire eucaryotic organisminto a single cell; consequently a protozoan cell may be much more complex thana single animal cell, which is dedicated to a single function. Thus, protozoansdisplay most of the typical features of a eucaryotic cell, but they may also haveevolved certain specialised features. The single cell is bounded by the typicalbilayer membrane discussed earlier, but depending on the type in question, thismay in turn be covered by a variety of organic or inorganic substances to forman envelope or shell.
49

One of the most characteristic structural features of protozoans is the contrac-tile vacuole, whose role is to pump out excess amounts of water that enterthe cell by osmosis. The activity of the contractile vacuole is directly related tothe osmotic potential di�erential between the cell and its surroundings. This isvitally important for freshwater protozoans, since the hypotonic nature of theirenvironment means that water is continually entering the cell. The contractilevacuole often has a star-shaped appearance, the radiating arms being canalsthat drain water from the cytoplasm into the vacuole.
Most protozoans have a heterotrophic mode of nutrition, typically ingestingparticulate food such as bacteria, and digesting them in phagocytic vacuoles.Since they actively `hunt' their food rather than simply absorbing it across thecell surface, it is not surprising that the majority of protozoans are capableof movement. The structural features used to achieve locomotion (e.g. cilia,�agella) are among the characteristics used to classify the protozoans.
Types of protozoan which are important (but probably do not need to be coveredin any depth for this module) include:
1. zoo�agellates (Mastigophora) - distinguished by their long, whip-like�agellum. Includes choano�agellates.
2. ciliates (Ciliophora) - largest, most complex group of protozoans. In-cludes well-known individuals such as Paramecium. Characterised by pos-session of cilia. Uniquely, they have two types of nuclei, macronuclei(similar to nucleus in other eukaryotes) and micronuclei (up to 80 percell, used only in conjugation.
3. amoebas (Sarcodina) - characterised by pseudopodia (�false feet�)
8.7 Amoebas with external shells (Foramanifera & Radi-olaria)
Some types of amoeba have an external shell covering the cell. The Foraminiferasecrete a shell of protein coated with calcium carbonate; their shells are coveredwith pores through which their long, �lamentous pseudopodia project. Someforaminiferans are zooplankton, microscopic organisms living at the surface ofthe sea, while others are bottom-dwellers. It is the discarded shells of countlesslong-dead creatures such as these that make up deposits of limestone manyhundreds of metres in depth. Many thousands of shells are needed to form just1 g of such a deposit! The White Cli�s of Dover are an example of a limestonedeposit made up largely of foraminiferan shells.
8.8 Sporozoans (Apixomplexa)
Members of this group are all parasitic, infecting a range of vertebrates andinvertebrates. They have complex life cycles involving both haploid and diploid
50

phases and infecting more than one host. Probably the best known is Plasmod-ium, the causative agent of malaria, which spends part of its life in a species ofAnopheles mosquito.
8.9 Slime moulds and water moulds
The �nal group to consider in this chapter are the so-called `fungus-like' protists.Its members are phylogenetically diverse, and its two principal groupings, theslime moulds and the water moulds, are placed far apart from each other inmodern classi�cation systems.
8.9.1 Oomycota (water moulds)
Water moulds resemble true fungi in their gross structure, comprising a mass ofbranched hyphae. At the cellular and molecular level however, they bear verylittle resemblance, and are not at all closely related. The Oomycota derive theirname from the single large egg cell that is fertilised to produce a diploid zygoteas part of the sexual reproduction cycle.
Many water moulds play an important role in the decomposition of dead plantsand animals in freshwater ecosystems, while others are parasitic on the gills of�sh.
Terrestrial members of the Oomycota include a number of important plantpathogens, such as rusts and mildews, which can have a devastating e�ect oncrops such as tobacco and potatoes.
8.9.2 Myxomycota (plasmodial slime moulds)
At one stage in their life cycle, the plasmodial or acellular slime moulds existas a single-celled amoeboid form. Two of these haploid amoebas fuse to givea diploid cell, which then undergoes repeated divisions of the nucleus, withoutany accompanying cell division; the result is a plasmodium, a mass of cytoplasmthat contains numerous nuclei surrounded by a single membrane.
This retains the amoeboid property of cytoplasmic streaming, so the wholemultinucleate structure is able to move in a creeping fashion. This 'feedingplasmodium' (mass of protoplasm containing several nuclei and bound by acytoplasmic membrane), which may be several centimetres in length and oftenbrightly coloured, feeds phagocytically (by engul�ng solid particles in the cellmembrane to be internally digested) on rotting vegetation.
Fruiting bodies develop from the plasmodium when it is mature or when con-ditions are unfavourable, and a cycle of sexual reproduction is entered. Whenfavourable conditions return, meiosis gives rise to haploid spores, which germi-nate to produce the amoeboid form once more.
51

8.9.3 Dictyostelida (cellular slime moulds)
A unicellular amoeboid form also �gures in the life cycle of the other group ofslime moulds, the Dictyostelida. This haploid amoeba is the main vegetativeform, but when food supplies are scarce, large numbers aggregate to form aslug-like blob, super�cially not unlike the plasmodium described above. Unlikethe plasmodium, however, this aggregate is fully cellular, so each componentcell retains its plasma membrane.
52

9 Growth & Death
9.1 Factors a�ecting microbial growth
9.1.1 Temperature
Microorganisms as a group are able to grow over a wide range of temperatures,from around freezing to above boiling point. For any organism, the minimumand maximum growth temperatures de�ne the range over which growth is pos-sible; this range is typically about 25�30 � C. Growth is slower at low tem-peratures because enzymes work less e�ciently and also because lipids tend toharden and there is a loss of membrane �uidity. Growth rates increase with tem-perature until the optimum temperature is reached, then the rate falls again.The optimum and limiting temperatures for an organism are a re�ection of thetemperature range of its enzyme systems, which in turn are determined by theirthree-dimensional protein structures. The optimum temperature is generallycloser to the maximum growth temperature than the minimum. Once the op-timum value is passed, the loss of activity caused by denaturation of enzymescauses the rate of growth to fall away sharply.
9.1.2 pH
Microorganisms are strongly in�uenced by the prevailing pH of their surround-ings. As with temperature, we can de�ne minimum, optimum and maximumvalues for growth of a particular type. The pH range (between minimum andmaximum values) is greater in fungi than it is in bacteria. Most microorgan-isms grow best around neutrality (pH 7). Many bacteria prefer slightly alkalineconditions but relatively few are tolerant of acid conditions, and fewer still areacidophilic.
9.1.3 Oxygen
Oxygen is present as a major constituent (20 per cent) of our atmosphere, andmost life forms are dependent upon it for survival and growth. Such organismsare termed aerobes. Not all organisms are aerobes however; some anaerobesare able to survive in the absence of oxygen, and for some this is actually anecessity.
Aerobic organisms require oxygen to act as a terminal electron acceptor in theirrespiratory chains. Such organisms, when grown in laboratory culture, musttherefore be provided with enough oxygen to satisfy their requirements. Fora shallow layer of medium such as that in a petri dish, su�cient oxygen isavailable dissolved in surface moisture. In a deeper culture such as a �ask ofbroth however, aerobes will only grow in the surface layers unless additional
53

oxygen is provided (oxygen is poorly soluble in water). This is usually done byshaking or mechanical stirring.
Obligate anaerobes cannot tolerate oxygen at all. They are cultured in spe-cial anaerobic chambers, and oxygen excluded from all liquid and solid media.Facultative anaerobes are able to act like aerobes in the presence of oxygen,but have the added facility of being able to survive when conditions becomeanaerobic. Aerotolerant anaerobes are organisms that are basically anaerobic;although they are not inhibited by atmospheric oxygen, they do not utilise it.Microaerophiles require oxygen, but are only able to tolerate low concentra-tions of it (2�10 per cent), �nding higher concentrations harmful. Organismsinoculated into a static culture medium will grow at positions that re�ect theiroxygen preferences.
9.1.4 Carbon Dioxide
Autotrophic organisms are able to use carbon dioxide as a carbon source; whengrown in culture, these are provided with bicarbonate in their growth mediumor incubated in a CO2-enriched atmosphere. However, heterotrophic bacteriaalso require small amounts of carbon dioxide, which is incorporated into variousmetabolic intermediates. This dependency can be demonstrated by the failureof these organisms to grow if carbon dioxide is deliberately removed from theatmosphere.
9.1.5 Osmotic Pressure
Osmosis is the di�usion of water across a semipermeable membrane from a lessconcentrated solution to a more concentrated one, equalising concentrations.The pressure required to make this happen is called the osmotic pressure. Ifa cell were placed in a hypertonic solution (one whose solute concentration ishigher), osmosis would lead to a loss of water from the cell (plasmolysis). Thisis the basis of using high concentrations of salt or other solutes in preservingfoods against microbial attack. In the opposite situation, water would pass froma dilute (hypotonic) solution into the cell, causing it to swell and burst. Therigid cell walls of bacteria prevent them from bursting; this, together with theirminute size, makes them less sensitive to variations in osmotic pressure thanother types of cell. They are generally able to tolerate NaCl concentrations ofbetween 0.5 and 3.0 per cent.
Haloduric (`salt-tolerant') bacteria are able to tolerate concentrations ten timesas high, but prefer lower concentrations, whereas halophilic (`salt-loving')forms are adapted to grow best in conditions of high salinity such as thosethat prevail in the Dead Sea in the Middle East. In order to do this withoutplasmolysis occurring, they must build up a higher internal solute concentration,which they do by actively concentrating potassium ions inside the cell.
54

9.1.6 Light
Phototrophic organisms require light in order to carry out photosynthesis. Inthe laboratory, care must be taken that light of the correct wavelength is used,and that the source used does not also act as a heat source. Fluorescent lightproduces little heat, but does not provide the wavelengths in excess of 750 nmneeded by purple and green photosynthetic bacteria.
9.2 The kinetics of microbial growth
Unicellular organisms divide by binary �ssion; each cell grows to full size, repli-cates its genetic material then divides into two identical daughter cells. Byidentical means, two cells divide into four, four into eight and so on, leading toan exponential increase in cell numbers:
1 99K 2 99K 4 99K 8 99K 2n
If we were to plot the number of cells in a population against time, we wouldget an exponential curve. It is more convenient when plotting a growth curveto plot the logarithm of cell numbers of against time, giving us a straight line.
Such exponential growth cannot continue inde�nitely, however, and growth usu-ally slows down due to either the supply of nutrients becoming exhausted, orbecause metabolism leads to an accumulation of harmful waste substances. Uni-cellular growth usually occurs in a series of di�erent phases:
1. lag phase
2. exponential (log, logarithmic) phase
3. stationary phase
4. death phase (exponential or logarithmic decline)
In standard bacterial growth curves one keeps track of cell growth by somemeasure or estimation of cell number.
9.2.1 Lag phase
� Upon a change in environment (especially from a rich environment to apoor environment), or when going from stationary phase to exponentialphase, there is a lag before division resumes.
55

� For example, stationary phase Escherichia coli placed in an excess of ster-ile broth will go through a lag phase during which they increase in cell sizebut do not divide. They will divide only once they have reached the sizeof a cell which is about to divide during exponential growth under thoseconditions.
� During this time a culture is said to be in lag phase.
� During lag phase cells increase in mass but do not divide.
� In other words, there is no change in number, but an increase in mass.
� "The length of the lag phase is determined in part by characteristics ofthe bacterial species and in part by conditions in the media�both themedium from which the organisms are taken and the one to which theyare transferred. Some species adapt to the new medium in an hour or two;others take several days. Organisms from old cultures, adapted to limitednutrients and large accumulated wastes, take longer to adjust to a newmedium than do those transferred from a relatively fresh, nutrient-richmedium."
9.2.2 Log phase (also exponential or logarithmic phase)
� Exponential growth is a physiological state marked by back-to-back divi-sion cycles such that the population doubles in number every generationtime.
� Note that during exponential growth there is no change in average cellmass, though individuals cells are constantly changing in mass as theyincrease in mass, then divide thus rapidly decreasing in mass (while in-creasing in number).
9.2.3 Stationary phase
� Stationary phase is classically de�ned as a physiological point where therate of cell division equals the rate of cell death, hence viable cell numberremains constant.
� Note that when cell division = 0 and cell death = 0, then the rate of celldivision = rate of cell death.
� In other words, when cells stop dividing but have not yet started dyingthey are in stationary phase.
� A way to distinguish these possibilities is to compare viable count withtotal count.
56

� If both total counts and viable counts don't change then you know thatthere is both no cell division and no cell death.
� If total count increases while viable counts remain constant, then youknow that you are observing a true balance between ongoing cell divisionand cell death.
� Stationary phase usually occurs when cell concentration is so great andthat some aspect of the environment is no longer able to serve the require-ments of exponential growth.
� Stationary phase is a time of signi�cant physiological change and partic-ularly involves the physiological adaptation of cells to survival throughperiods of little growth.
9.2.4 Death phase (also exponential or logarithmic decline)
� In single celled microorganisms cell death is the point at which reinitiationof division is no longer possible.
� Note that the concept of cell death is actually dependent on how oneattempts to reinitiate growth.
� Particularly, there are ways to gently revive some microbes from phys-iological states that would result in permanent lack of growth in othergrowth environments.
� An analogous situation would be a person with an injury that is inevitablyfatal in a third-world hospital, but readily treated in a �rst-world hospital.
� Death phase is a physiological point at which cell deaths exceed cell births.
� More speci�cally, viable count declines.
� "During the decline phase, many cells undergo involution�that is, theyassume a variety of unusual shapes, which makes them di�cult to iden-tify." (p. 140, Black, 1996)
9.3 Batch culture and continuous culture
The phases of growth described above apply to a batch culture. In this formof culture, appropriate nutrients and other conditions are provided for growth,then an inoculum is added and the culture incubated. No further nutrients areadded and no waste products are removed, thus conditions in the culture arecontinually changing. This results in active growth being of limited durationfor the reasons outlined above.
57

Sometimes it is desirable to keep the culture in the logarithmic phase, for ex-ample if the cells are being used to produce alcohol or antibiotics.
In a continuous culture, nutrient concentrations and other conditions are heldconstant, and the cells are held in a state of exponential growth. This is achievedby continuously adding fresh culture medium and removing equal volumes of theold. Parameters such as pH can also be monitored and adjusted. The equipmentused to do this is called a chemostat. It produces a steady-state culture whosepopulation size is kept constant by careful control of �ow rates and nutrientconcentrations.
9.4 Growth in multicellular organisms
If uninterrupted, growth in fungi proceeds radially outwards from the initiatingspore, allowing the fungal colony to colonise new regions potentially rich innutrients. Actual growth occurs solely at the hyphal tip; as this happens, theterminal cell grows longer, until eventually a new cross wall or septum is formed.Cells away from the tip do not become any longer during hyphal extension,however hyphae in this region may develop into aerial reproductive structures.Older hyphae at some distance from the tip may become completely empty ofcytoplasm. Cell counts and turbidometric measurements are not appropriateto estimate growth of fungi; however total mycelial mass can be measured andits change plotted against time. A fungal growth cycle shows roughly the samephases of growth as described above for bacteria.
9.5 Culture media
A culture medium is nutrient material prepared in the laboratory for thegrowth of microorganisms. Microorganisms that grow in size and number on aculture medium are referred to as a culture. In order to use a culture mediummust be sterile, meaning that it contains no living organisms. This is importantbecause we only want microorganisms that we add to grow and reproduce, notothers. We must have the proper nutrients, pH, moisture, and oxygen levels(or no oxygen) for a speci�c microorganism to grow. Many culture media areavailable for microbial growth. Media are constantly being developed for theuse of identi�cation and isolation of bacteria in the research of food, water, andmicrobiology studies.
The most popular and widely used medium in microbiology laboratories is thesolidifying agent agar. Agar is a complex polysaccharide derived from red algae.Very few microorganisms can degrade agar, so it usually remains in a solid form.Agar media are usually contained in test tubes or Petri dishes. The test tubesare held at a slant and are allowed to solidify on an angle, called a slant. A slantincreases the surface area for organism growth. When they solidify in a verticaltube it is called a deep. The shallow dishes with lids to prevent contamination
58

are called Petri dishes. Petri dishes are named after their inventor, Julius Petri,who in 1887 �rst poured agar into glass dishes.
9.6 Chemically de�ned media
For a medium to support microbial growth, it must provide an energy source,as well as carbon, nitrogen, sulfur, phosphorous, and any other organic growthfactors that the organism cannot make itself, source for the microorganisms toutilize.
A chemically de�ned medium is one whose exact chemical composition isknown. Chemically de�ned media must contain growth factors that serve as asource of energy and carbon. Chemically de�ned media are used for the growthof autotrophic bacteria.
Heterotrophic bacteria and fungi are normally grown on complex media, whichare made up of nutrients, such as yeasts, meat, plants, or proteins (the exactcomposition is not quite known and can vary with each mixture). In complexmedia, the energy, carbon, nitrogen, and sulfur needed for microbial growth areprovided by protein. Proteins are large molecules that some microorganisms canuse directly. Partial digestion by acids and enzymes break down proteins intosmaller amino acids called peptones. Peptones are soluble products of proteinhydrolysis. These small peptones can be digested by bacteria. Di�erent vitaminsand organic growth factors can be provided by meat and yeast extracts.
If a complex medium is in a liquid form it is called a nutrient broth.
If agar is added, it is called a nutrient agar.
9.7 Selective and di�erential media
In health clinics and hospitals, it is necessary to detect microorganisms that areassociated with disease. Selective and di�erential media are therefore used.
Selective media are made to encourage the growth of some bacteria whileinhibiting others. An example of this is bismuth sul�te agar. Bismuth sul�teagar is used to isolate Salmonella typhi from fecal matter. Salmonella typhi isa gram-negative bacterium that causes salmonella.
Di�erential media make it easy to distinguish colonies of desired organismsfrom nondesirable colonies growing on the same plate. Pure cultures of microor-ganisms have identi�able reactions with di�erent media. An example is bloodagar.
Blood agar is a dark red/brown di�erential medium that contains red bloodcells used to identify bacterial species that destroy red blood cells. An exampleof this type of bacterium is Streptococcus pyogenes, the agent that causes strepthroat. Blood agar is not selective, and will grow a wide range of organisms.
59

MacConkey agar is both selective and di�erential. MacConkey agar containsbile salts and crystal violet, which inhibit the growth of gram-positive bacteria,and lactose, in which gram-negative bacteria can grow.
Rose bengal chloramphenicol is a selective medium for the enumeration ofyeasts and moulds. It is not di�erential.
Mannitol salt agar is a selective and di�erential medium. Mannitol is thedi�erential part, it indicates organisms that ferment mannitol. If mannitol fer-mentation is occuring, lactic acid will be produced and the pH will dop causingthe MSA plate to turn yellow. The salt portion is selective for halophiles; or-ganisms that cannot withstand a high salt concentration will be unable to growwell.
Enrichment cultures are usually liquids and provide nutrients and environ-mental conditions that provide for the growth of certain microorganisms, butnot others.
9.8 Microbial death kinetics
When microorganisms are exposed to any of the treatments designed to killthem, they are not all killed instantaneously. During a given time period, onlya certain proportion of them will die.
Suppose we had 1000 cells (an unrealistically small number, but it keeps thearithmetic simple) and that 10 per cent were killed each minute. After oneminute, 900 cells would remain, and after the second minute 10 per cent ofthese would die, leaving us with 900 = 90 = 810 survivors.
After a further minute, another 10 per cent of the survivors would be killed,so 810 = 81 = 729 would be left. A plot of the surviving cells against time ofexposure gives a graph such as shown below.
The curve is exponential; theoretically, there will never be zero survivors, butafter a while we are going to have less than one cell, let us say one tenth of a cell,which clearly can not happen. What this really means is that in a given unitvolume, there will be a one in 10 chance of there being a cell present. Sterilityis generally assumed when this �gure falls as low as one in a million.
Since only a proportion of the surviving population is killed per unit time, itfollows that the more cells you have initially, the longer it will take to eliminatethem. The steepness of the slope of the graph is an indication of the e�ectivenessof heat sterilisation.
9.8.1 The D value, z value, F value
The decimal reduction time or D value is the time needed to reduce thepopulation by one log cycle, equal to a factor of ten (i.e. to kill 90 per cent of
60

Figure 4: The kinetics of cell death. (a) During heat sterilisation, the number ofliving cells decreases by the same proportion per unit time, giving an exponentialcurve. (b)When plotted on a logarithmic scale, the decrease in numbers is seenas a straight line, whose slope is a re�ection of the rate of killing. The time periodbetween A and A is the decimal reduction time (D): the time taken to reducethe population to one-tenth of its size. The total period until the point SA isthe sterility assurance value, when there is only a one in a million probabilityof any cells having survived.
the population) using a particular heat treatment. The D value applies to aparticular temperature; at higher temperatures, the rate of killing is enhanced,and so the D value is reduced). The D value is usually written with a subscript,indicating the temperature for which it applies, e.g. D121 represents the D valueat 121°C.
The increase in temperature required to reduce D by a factor of 10 is the Zvalue.
Another way to describe heating e�ectiveness is with the F value. The Fvalue is the time in minutes at a speci�c temperature (usually 250°F or 121.1°C)needed to kill a population of cells or spores.
Since in real life, the microbial population is certain to be a mixed one, thenthe critical factor is the death rate of the most resistant species, that is, theone with the highest percentage of survivors per minute. Sterilisation protocolsshould therefore be based on the rate of destruction of endospores.
9.8.2 D and z values in the food industry (possible long answerexam question, learn this!)
The food processing industry makes extensive use of D and z values. After afood has been canned, it must be heated to eliminate the risk of botulism arisingfrom Clostridium botulinum spores. See the relevant section (food poisoning)for details of botulism. Heat treatment is carried out long enough to reduce apopulation of 1012 C. botulinum spores to 100 (one spore); thus there is a verysmall chance of any can having a viable spore.
61

The D value for these spores at 121°C is 0.204 minutes. Therefore it would take12D or 2.5 minutes to reduce 1012 spores to one spore by heating at 121°C.
Note: If you're having trouble following this, join the club. Here's an attemptat an explanation....
The D value represents the time taken to kill 90% of the population. If D is0.204 minutes and you start with 1012 spores, then after 0.204 minutes you have1011 spores left. After another 0.204 (D) you have 1010 spores left, and so on.
In other words, each time D time passes, the population is reduced by onepower
The z value for C. botulinum spores is 10°C�that is, it takes a 10°C changein temperature to alter the D value tenfold. If the cans were to be processedat 111°C rather than at 121°C, the D value would increase by tenfold to 2.04minutes and the 12D value to 24.5 minutes.
62

Part II
Microbial control
10 The control of microorganisms
It is very important to control microbial growth in surgical and hospital settings,as well as in industrial and food preparation facilities. There are many termsused to describe the �ght to control microorganisms. Here are the importantones to learn:
Sterilization is the destruction of all microorganisms and viruses, as well asendospores. Sterilization is used in preparing cultured media and cannedfoods. It is usually performed by steam under pressure, incineration, or asterilizing gas such as ethylene oxide.
Antisepsis is the reduction of pathogenic microorganisms and viruses on livingtissue. Treatment is by chemical antimicrobials, like iodine and alcohol.Antisepsis is used to disinfect living tissues without harming them.
Commercial sterilization is the treatment to kill endospores in commer-cially canned products. An example is the bacteria Clostridium botulinum,which causes botulism.
Aseptic means to be free of pathogenic contaminants. Examples include properhand washing, �ame sterilization of equipment, and preparing surgical en-vironments and instruments.
Any word with the su�x -cide or �cidal indicates the death or destruction ofan organism. For example, a bactercide kills bacteria. Other examples arefungcides, germicides and virucides. Germicides include ethylene oxide,propylene oxide, and aldehydes. For the same reason, these germicidesare also used in preserving specimens in laboratories.
Disinfection is the destruction or killing of microorganisms and viruses onnonliving tissue by the use of chemical or physical agents. Examples ofthese chemical agents are phenols, alcohols, aldehydes, and surfac-tants. Pasteurisation is an example of a physical method.
Degerming is the removal of microorganisms by mechanical means, such ascleaning the site of an injection. This area of the skin is degermed byusing an alcohol wipe or a piece of cotton swab soaked with alcohol. Handwashing also removes microorganisms by chemical means.
Pasteurisation uses heat to kill pathogens and reduce the number of foodspoilage microorganisms in foods and beverages. Examples are pasteurizedmilk and juice.
63

Sanitation is the treatment to remove or lower microbial counts on objectssuch as eating and drinking utensils to meet public health standards. Thisis usually accomplished by washing the utensils in high temperatures orscalding water and disinfectant baths.
Bacterostatic, fungistatic, and virustastic agents � or any word with thesu�x -static or -stasis � indicate the inhibition of a particular type ofmicroorganism. These are unlike bactericides or fungicides that kill or de-stroy the organism. Germistatic agents include refrigeration, freezing,and some chemicals.
10.1 Classi�cation of pathogens
There are four categorie of pathogens:
1. Hazard Group 1: unlikely to cause human disease;
2. Hazard Group 2: can cause human disease and may be a hazard toemployees; it is unlikely to spread to the community and there is usuallye�ective prophylaxis or treatment available;
3. Hazard Group 3: can cause severe human disease and may be a serioushazard to employees; it may spread to the community, but there is usuallye�ective prophylaxis or treatment available
4. Hazard Group 4: causes severe human disease and is a serious hazardto employees; it is likely to spread to the community and there is usuallyno e�ective prophylaxis or treatment available.
Laboratories wishing to work with organisms at hazard group 3 and 4 must takethese extra precautions:
� Negative air pressure required
� Outgoing air to be �ltered (double �lter at cat 4 level)
� Cat 4 labs accessed through air-lock
� Observation window
� Sealable to allow disinfection
� At cat 4 there must be an incinerator on-site and all equipment mustremain in the room, also all surfaces must be non-porous and easy toclean
64

Table 1: Temperature reached at given steam pressuresPressure
(kPa) (psi) Temperature
0 0 10069 10 115
103.5 15 121138 20 126172.5 25 130
10.2 Sterilisation
One of the oldest forms of anitmicrobial treatment is that of heating, and inmost cases this remains the preferred means of sterilisation, provided that itdoes not cause damage to the material in question.
10.2.1 Sterilisation by heat
Boiling at 100 � C for 10 minutes is usually enough to achieve sterility, providedthat organisms are not present in high concentrations; in fact most bacteriaare killed at about 70 � C. If, however, endospores of certain bacteria (notablyBacillus and Clostridium) are present, they can resist boiling, sometimes forseveral hours. The causative agents of some particularly nasty conditions, suchas botulism and tetanus, are members of this group. In order to destroy theheat resistant endospores, heating beyond 100 � C is required, and this can beachieved by heating under pressure in a closed vessel (see Table).
10.2.2 The Autoclave
A typical laboratory treatment is 15 minutes at a pressure of 103 kpa (15 psi),raising the temperature of steam to 121 � C. This is carried out in an autoclave,which is, to all intents and purposes, a large-scale pressure cooker. Air is drivenout of the system so that the atmosphere is made up entirely of steam; thedesired temperature will not be reached if this is not achieved. Large loads, orlarge volumes of liquids, may need a longer treatment time in order for the heatto penetrate throughout. Modern autoclaves include probes designed to assessthe temperature within a load rather than that of the atmosphere.
Autoclaves work due to the latent heat of condensation. Latent heat isthe heat transfered from one medium to another. In an autoclave, pressurisedsteam condenses on any surface, transfering large amounts of heat energy to thematerial it lands on, which kills microorganisms.
Some other methods commonly used to test autoclave e�ectiveness include:
65

� Spores of Bacillus stearothermophilus are introduced into a system alongwith the material to be sterilised; if subsequent testing shows that thespores have all been destroyed, it is reasonable to assume that the sys-tem has also destroyed any other biological entity present. This test issometimes called the spore strip test. The problem with this method isthat it is retrospective. If a problem has arisen, then this will be discov-ered only when the spores have germinated. This is probably too late totake e�ective action other than to call in an engineer to prevent furtherproblems.
� Special tape calledAutoclave Tape which changes colour if the necessarytemperature is reached can act as a more convenient but less reliableindicator.
� Browne's Tubes are glass tubes that contain heat sensitive dyes. Thesechange colour after su�cient time at the desired temperature. Before heatexposure, the contents of the tube appear red. As heating progresses, thecolour changes to green. Only when the tube is green can sterilisationconditions be considered adequate.
� Bowie Dick tape is applied to articles being autoclaved. If the processhas been satisfactory, dark brown stripes will appear across the tape. Palebrown stripes are suggestive of poor heat penetration, and an unsatisfac-tory sterilisation process.
These methods give an immediate indication of the success or otherwise of anautoclave run but they are only suggestive of a successful sterilisation.
10.2.3 Tyndallisation / Intermittent steaming
An e�ect similar to that achieved by autoclaving can be obtained by a methodcalled intermittent steaming or tyndallisation (after the Irish physicist JohnTyndall, who was one of the �rst to demonstrate the existence of heat-resistantmicrobial forms). This is used for those substances or materials that might bedamaged by the high temperatures used in autoclaving. The material is heatedto between 90 and 100 � C for about 30 minutes on each of three successivedays, and left at 37 � C in the intervening periods. Vegetative cells are killed o�during the heating period, and during the 37 � C incubation, any endosporesthat have survived will germinate. Once these have grown into more vegetativecells, they too are killed in the next round of steam treatment. Clearly this isquite a long-winded procedure, and it is therefore reserved for those materialswhich might be harmed by steam sterilisation.
10.2.4 Pasteurisation
Pasteurisation is a process which slows microbial growth in foods. The pro-cess was named after its creator, French chemist and microbiologist Louis Pas-
66

teur. The �rst pasteurisation test was completed by Louis Pasteur and ClaudeBernard on April 20, 1862.
Unlike sterilization, pasteurisation is not intended to kill all pathogenic micro-organisms in the food or liquid. Instead, pasteurisation aims to reduce thenumber of viable pathogens so they are unlikely to cause disease (assuming thepasteurisation product is refrigerated and consumed before its expiration date).Commercial-scale sterilization of food is not common because it adversely a�ectsthe taste and quality of the product.
Pasteurisation is an example of disinfection.
Pasteurisation typically uses temperatures below boiling since at temperaturesabove the boiling point for milk, casein micelles will irreversibly aggregate (or"curdle"). There are two main types of pasteurisation used today: High Tem-perature/Short Time (HTST) and Extended Shelf Life (ESL) treatment. Ultra-high temperature (UHT or ultra-heat treated) is also used for milk treatment.In the HTST process, milk is forced between metal plates or through pipesheated on the outside by hot water, and is heated to 71.7 °C (161 °F) for 15-20seconds. UHT processing holds the milk at a temperature of 138 °C (250 °F)for a fraction of a second. ESL milk has a microbial �ltration step and lowertemperatures than HTST.[1] Milk simply labeled "pasteurisation " is usuallytreated with the HTST method, whereas milk labeled "ultra-pasteurisation "or simply "UHT" has been treated with the UHT method.
10.3 Sterilisation by irradiation
Certain types of irradiation are used to control the growth of microorganisms.These include both ionising and non-ionising radiation.
10.3.1 UV Radiation
The most widely used form of non-ionising radiation is ultraviolet (UV) light.Wavelengths around 260 nm are used because these are absorbed by the purineand pyrimidine components of nucleic acids, as well as certain aromatic aminoacids in proteins. The absorbed energy causes a rupture of the chemical bonds,so that normal cellular function is impaired. UV light causes the formationof thymine dimers, where adjacent thymine nucleotides on the same strandare linked together, inhibiting DNA replication. Although many bacteria arecapable of repairing this damage by enzyme-mediated photoreactivation, virusesare much more susceptible. UV lamps are commonly found in food preparationareas, operating theatres and specialist areas such as tissue culture facilities,where it is important to prevent contimination. Because they are also harmfulto humans (particularly the skin and eyes), UV lamps can only be operated insuch areas when people are not present. UV radiation has very poor penetratingpowers; a thin layer of glass, paper of fabric is able to impede the passage of
67

the rays. The chief application is therefore in the sterilisation of work surfacesand the surrounding air, although it is increasingly �nding and application inthe treatment of water supplies.
10.3.2 Ionising radiation
Ionising radiations have a shorter wavelength and much higher energy, givingthem greater penetrating powers. The e�ect of ionising radiations is due tothe production of highly reactive free radicals, which disrupt the structure ofmacromolecules such as DNA and proteins. Surgical supplies such as syringes,catheters and rubber gloves are commonly sterilised employing gamma (γ) raysfrom the isotope cobalt 60 (60Co).
10.3.3 Gamma radiation
Gamma radiation has been approved for use in over 40 countries for the preser-vation of food, which it does not only by killing pathogens and spoilage organ-isms but also by inhibiting processes that lead to sprouting and ripening. Thepractice has aroused a lot of controversy, largely due to concerns about healthand safety, although the �rst patent applications for its use date back nearly ahundred years! Although the irradiated product does not become radioactive,there is a general suspicion on the part of the public about anything to do withradiation, which has led to its use on food being only very gradually accepted byconsumers. In this respect Europe lags behind the USA, where during the 1990sa positive attitude towards irradiation of food both by professional bodies andthe media has led to a more widespread acceptance of the technology. Gammaradiation is used in situations where heat sterilisation would be inappropriate,because of undesirable e�ects on the texture, taste or appearance of the product.This mainly relates to fresh produce such as meat, poultry, fruit and vegeta-bles. Irradiation is not suitable for some foodstu�s, such as those with a highfat content, where unpleasant tastes and odours result. Ionising radiations havethe great advantage over other methods of sterilisation that they can penetratepackaging.
10.4 Filtration
Many liquids such as solutions of antibiotics or certain components of culturemedia become chemically altered at high temperatures (such substances arecalled heat labile ie. damaged or altered by heat), so the use of any of theheat regimes described above is not appropriate. Rather than killing the mi-croorganisms, an alternative approach is simply to isolate them.
This can be done for liquids and gases by passing them through �lters of an ap-propriate pore size. Filters used to be made from materials such as asbestos and
68

sintered glass, but have been largely replaced by membrane �lters, commonlymade of nitrocellulose or polycarbonate. These can be purchased ready-sterilisedand the liquid passed through by means of pressure or suction. Supplies of airor other gases can also be �lter-sterilised in this way. A pore size of 0.22 µmis commonly used; this will remove bacteria plus, of course, anything bigger,such as yeasts; however, mycoplasma and viruses are able to pass through poresof this size. With a pore size 10 times smaller than this, only the smallest ofviruses can pass through, so it is important that an appropriate pore size ischosen for any given task.
A drawback with all �lters, but especially those of a small pore size, is thatthey can become clogged easily. Filters in general are relatively expensive,and are not the preferred choice if alternative methods are available. Highe�ciency particulate air (HEPA) �lters create clean atmospheres in areas suchas operating theatres and laboratory laminar-�ow hoods.
10.5 Phenols and phenolics
Phenols are compounds derived from pheno (carbolic acid) molecules. Phenolicsdisrupt the plasma by denaturing proteins; they also disrupt the plasma mem-brane of the cell. Joseph Lister used phenol in the late 1800s to reduce infectionduring surgery. Phenolic disinfectants are useful for disinfecting laboratory worksurfaces.
Examples include Dettol and TCP
10.6 Alcohols
Alcohols are e�ective against bacterial fungi and viruses. However, they, arenot e�ective against fungal spores or bacterial endospores. Alcohols that arecommonly used are isopropanol (rubbing alcohol) and ethanol (the alcoholwe drink). Alcohols denature proteins and disrupt cytoplasmic membranes.Pure alcohol is not as e�ective as 70 percent because the denaturing of proteinsrequires water. Alcohols are good to use because they evaporate rapidly. Adisadvantage is that they may not contact the microorganisms long enough tobe e�ective. Alcohol is commonly used in swabbing the skin prior to an injection.
10.7 Halogens
Halogens are nonmetallic, highly resistive chemical elements. Halogens are ef-fective against vegetative bacterial cells, fungal cells, fungal spores, protozoancysts, and many viruses. Halogen-containing antimicrobial agents include io-dine, which inhibits protein function. Iodine is used in surgery and by campersto disinfect water. An iodophur is an iodine-containing compound that is
69

longer-lasting than iodine and does not stain the skin. Other halogen agentsinclude:
� Chlorine (Cl2). Used to treat drinking water, swimming pools, and insewage plants to treat waste water. Chlorine products such as sodiumhypochlorite (household bleach) are e�ective disinfectants.
� Chlorine dioxide (ClO2). A gas that can disinfect large areas.
� Chloroamines. Chemicals containing chlorine and ammonia. They areused as skin antiseptics and in water supplies.
� Bromine. Used to disinfect hot tubes because it does not evaporate asquickly as chlorine in high temperatures.
� Oxidizing agents. Kill microorganisms by oxidizing their enzymes, thuspreventing metabolism. Hydrogen peroxide, for example, disinfects andsterilizes inanimate objects, such as food processing and medical equip-ment, and is also used in water puri�cation.
Examples include Bleach, Domestos, Parazone etc.
10.8 Heavy metals
Arsenic, zinc, mercury, silver, nickel, and copper are called heavy met-als due to their high molecular weights. They inhibit microbial growth becausethey denature enzymes and alter the three-dimensional shapes of proteins thatinhibit or eliminate the protein's function. Heavy metals are bacteriostatic andfungistatic agents. An example is silver nitrate. At one time, hospitals requirednewborn babies to receive a one percent cream of silver nitrate to their eyes toprevent blindness caused by Neisseria gonorhoeae, which could enter the baby'seyes while passing through the birth canal of a mother who was infected. To-day, antibiotic ointments that are less irritating are used. Another example isthe use of copper in swimming pools, �sh tanks, and in reservoirs to control al-gae growth. Copper interferes with chlorophyll, thus a�ecting metabolism andenergy.
10.9 Aldehydes: Glutaraldehyde and Formaldehyde
Aldehydes function in microbial growth by denaturing proteins and inactivatingnucleic acids. Two types are glutaraldehyde that is a liquid and formaldehydethat is a gas.
� Glutaraldehyde is used in a two percent solution to kill bacteria, fungi,and viruses on medical and dental equipment.
70

� Healthcare workers and morticians dissolve gaseous formaldehyde inwater, making a 37 percent solution of formalin. Formalin is used indisinfecting dialysis machines, surgical equipment, and embalming bodiesafter death.
� Gaseous agents, such as ethylene oxide, propylene oxide, andbeta-propiolactone, are used on equipment that cannot be sterilizedeasily with heat, chemicals, or radiation. Certain items, like pillows,mattresses, dried or powered food, plastic-ware, sutures, and heart-lungmachines, are placed in a closed chamber, then �lled with these gases.Gaseous agents denature proteins.
10.10 Surfactants
Surfactants are chemicals that act on surfaces by decreasing the tension of waterand disrupting cell membranes. Examples are household soaps and detergents.
71

11 Antimicrobial agents
� The term chemotherapy is most closely associated in the minds of mostpeople with the treatment of cancer. In fact the term was �rst used byPaul Ehrlich to describe any use of a drug or other chemical substancefor the treatment of disease; thus, it has much wider terms of reference.In our present discussion, we shall con�ne ourselves to chemotherapy asit relates to the treatment of infectious diseases. It was Ehrlich who, 100years ago, observed how certain dyes would stain bacteria but not thesurrounding tissues, leading him to formulate the idea of selective toxic-ity, whereby a substance would selectively target harmful microorganismsbut leave human tissues undamaged. He tested hundreds of syntheticcompounds in the search for his `magic bullet' before �nding, in 1910,an arsenic-containing drug, Salvarsan, which was e�ective against Tre-ponema pallidum, the causative agent of syphilis.
� It was another 20 years before another signi�cant antimicrobial drug wasdeveloped, when the German chemist Gerhard Domagk showed a syn-thetic dye, Prontosil, to be active against a range of Gram-positive bac-teria. The active component of prontosil was shown soon afterwards tobe sulphanilamide. In the following decade, numerous derivatives of sul-phanilamide were synthesised, many of which were more potent antimicro-bial agents than the parent molecule. This class of compounds is knowncollectively as the sulphonamides, or sulfa drugs. In the years leadingup to the Second World War, sulphonamides dramatically improved themortality rates due to pneumonia and puerperal fever.
� Nowadays, sulphonamides have largely been replaced by antibiotics be-cause of their side-e�ects, and because, due to wholesale and indiscrim-inate use in the early years, bacterial resistance to sulphonamides hasbecome widespread. Some synthetic comounds are still useful as antimi-crobial agents, however. Isoniazid is one of the principal agents usedin the treatment of tuberculosis. It is nearly always given in associationwith another antimicrobial agent because of the high incidence of resistantforms of the mycobacteria that cause the disease.
11.1 Antibiotics
The other major breakthrough in the treatment of infectious diseases was ofcourse the discovery of naturally occurring antimicrobial agents, or antibiotics.These are metabolites produced by certain microorganisms, which inhibit thegrowth of certain other microorganisms. The de�nition has been extended toinclude semisynthetic derivatives of these naturally occuring molecules.
� Antibiotic theapy has been used in some form for many centuries. Manyhundreds of years ago, the Chinese used mouldy soybean curd in the treat-
72

ment of boils and South American Indians controlled foot infections bywearing sandals which had become furry with mould!
� In the late 19th century, Tyndall made the observation that a culturemedium cloudy with bacterial growth would clear when mould grew onthe surface.
� Around the same time Pasteur and Joubert demonstrated that culturedanthrax bacilli could be inactivated in the presence of certain other mi-croorganisms from the environment.
� By the early 1920s the search was on for the isolation of a microbiallyproduced antibacterial agent, and Gratia and Dath isolated a substancefrom a soil actinomycete which came to be known as actinomycin. How-ever, although potent against a number of pathogens, actinomycin is tootoxic to be useful therapeutically.
� One of the best known of all stories of scienti�c discovery is that of howSir Alexander Fleming discovered penicillin in 1928...
11.1.1 Fleming and Penicillin
Fleming was also looking for a naturally occurring antimicrobial agent. On oneoccasion, he noticed that a plate culture of Staphylococcus aureus had becomecontaminated by the growth of a mould; around it were clear areas, where theS. aureus did not grow. The mould was subsequently identi�ed as Penicil-lium notatum . and the substance that had di�used through the agar fromit, preventing bacterial growth, became known as penicillin. Further investiga-tion revealed that broth from a culture of the Penicillium mould was inhibitorytowards the growth of a number of other Gram-positive pathogens, and re-mained so even when diluted several hundred times. Critically, when tested onmice, it was, for the most part, harmless.
When it came to purifying the active ingredient and using it in vivo however,a number of problems were encountered. The penicillin proved to be impure,only produced in minute amounts, and unstable in the acid conditions of thestomach, thereby limiting its therapeutic potential.
After publishing a few papers on the subject, Fleming ceased work on penicillinand it was left to Howard Florey and Ernst Chain in 1939 to take up thechallenge of producing it in su�cient quantities and in a pure enough form fortherapeutic use. Early work in Oxford had to continue in the United States be-cause of the German air raids in Britain. The American entry into the SecondWorld War in late 1941 meant that the development of penicillin was awardedwar project status, giving it greatly added impetus. As a result of their endeav-ours, the yield of penicillin rose hugely, and in 1945, Fleming, Chain and Floreyshared the Nobel Prize for their work.
73

� Other antibiotics were also isolated during this period, most notably strep-tomycin, isolated by SelmanWaksman from Streptomyces griseus, whichwas to prove so e�ective against tuberculosis.
In 1942 there was only enough penicillin in the world to treat a few hundredindividuals, but by the end of the Second World War production had grown tosuch an extent that 7 million people a year could be treated. By the mid-1950s,such well-known antibiotics as tetracycline, chloramphenicol and neomycinhad been isolated. The discovery of a few naturally occurring compounds hadrevolutionised the treatment of infectious diseases.
New antibiotics are still being sought today. Of the thousands isolated so far,only a small proportion have proved to be of any real therapeutic or commercialvalue. This is because, like the actinomycin mentioned above, most of the sub-stances isolated harm not only bacteria but humans too. A key prerequisite forany chemotherapeutic agent is selective toxicity. An obvious way of achievingthis is for a compound to direct its e�ect against a metabolic or physiologicalfunction found in microbial cells but not in the host. We shall look at someexamples of this later in this chapter. Those chemotherapeutic agents whichinhibit the same process in host as in pathogen, or which cause harm to thehost in some other way, are said to have side-e�ects. These may include directlytoxic e�ects, hypersensitivity (allergic) reactions or adverse e�ects on the host'snormal resident micro�ora. One of the reasons why penicillin was, and contin-ues to be, so successful, was that the target of its action is unique to bacteria,so it its degree of selective toxicity is high.
Penicillin is usually most e�ective against Gram-positive bacteria.
11.1.2 Types of antibiotic
All antibiotics have the common property of interfering in some way with anormal, critical function of the target bacterial cell. The most commonly usedantibiotics exert their e�ect by one of the following methods:
1. Inhibition of cell wall synthesis (group I). The main group which workin this way are the β-lactam antibiotics, so-called because they containa β-lactam ring in their structure. Included among this group are thepenicillins and the cephalosporins.
2. Disruption of cell membranes (group II)
3. Interference with protein synthesis (group III)
4. Interference with nucleic acid synthesis (group IV)
74

11.2 Anti fungal agents
Fungi are eucaryotes, and therefore are una�ected by those agents which se-lectively target uniquely procaryotic features such as peptidoglycans and 70Sribosomes. Therein lies the problem: anything that damages fungal cells islikely to damage human cells too.
Polyene antibiotics such as amphotericin and nystatin (both produced by speciesof Streptomyces) act on the sterol components of membranes; their use is limited,because human cells can also be a�ected by their action (to use a term welearnt earlier in this chapter, they have a low therapeutic index). Nystatinis used topically against Candida infections, while amphotericin B is generallyused against systemic infections of fungal origin. The latter substance can havea wide range of serious side-e�ects, but in some cases infections are so severethat the physician is faced with no alternative. Synthetic compounds such asthe imidazoles have a similar mode of action to the polyenes; they are e�ectiveagainst super�cial mycoses (fungal infections of the skin, mouth and urinogenitaltract). Griseofulvin, a natural antibiotic produced by a species of Penicillium,is another antifungal agent whose use is restricted; it works by interfering withmitosis and not surprisingly has a range of side-e�ects. Although used to treatsuper�cial infections, it is taken orally.
11.3 Anti viral agents
In spite of the looming threat of resistant strains, there is no doubt that an-tibiotics have been hugely successful in the control of bacterial diseases. Wehave, however, been a lot less successful when it comes to �nding a treatmentfor diseases caused by viruses; a quick revision of their modus operandi shouldmake it clear why this is so. Viruses survive by entering a host cell and hijackingits replicative machinery, thus a substance interfering with the virus is likely toharm the host as well. A number of compounds have been developed however,which are able to act selectively on a viral target. All antiviral agents act byinterfering with some aspect of the virus's replication cycle. A number of suchcompounds have been found, but only a few have been approved for use in hu-mans. One of the �rst antiviral agents to be approved for use was amantidine,which inhibits uncoating of the in�uenza A virus by preventing the formationof acid conditions in the host cell's endocytotic vesicles. Its speci�city for thevirus is due to selective binding to M2, a matrix protein. Amantidine's e�cacyis dependent on administration within the early stages of an infection. It canbe administered prophylactically, but may have side-e�ects.
75

Part III
The importance of microorganisms
12 Disease and microorganisms
12.1 Classi�cation of diseases
Epidemiologists measure the frequency of diseases within a given population inregards to the geographical size of the area and the amount of damage the dis-ease in�icts on the population. Diseases can be classi�ed as endemic, sporadic,epidemic, or pandemic.
� An endemic disease is the average or normal number of cases of a diseasein a certain population. The number of people contracting the disease andthe severity of the disease is so low that it raises little concern and doesnot constitute a health problem. An example is the varicella-zoster virus(the virus that causes chickenpox). Chickenpox is an endemic disease thatusually a�ects children and is seasonal. An endemic disease can give riseto epidemics.
� A sporadic disease occurs when there are small numbers of isolated casesreported. Sporadic diseases do not threaten the population.
� An epidemic disease arises when the level of disease in a certain popu-lation exceeds the endemic level. This disease will cause an increase inmortality rate and the rate of morbidity. The level of destruction will beso large that it will cause a signi�cant public health concern.
� A disease becomes pandemic when it is distributed throughout the world.For example, in 1918 the swine �u (in�uenza) reached pandemic propor-tions. Some experts consider the HIV virus to be pandemic.
� In a common source epidemic, large numbers of the population aresuddenly infected from the same source. These epidemics usually areattributed to a contaminated supply of water or improperly prepared orhandled food. An example is people who eat contaminated chicken salad ata college cafeteria. Everyone who eats the chicken salad on this particularday will become infected and feel ill. The epidemic will subside very fast,though, as the source of infection is eradicated.
� A propagated epidemic occurs from person-to-person contact. Thedisease causing agent moves from a person who is infected to a personwho is not infected. In a propagated epidemic, the number of new casesrises and falls much slower than in common source epidemics, making the
76

pathogen much harder to isolate and thus eliminate. An example is a ��u�virus.
� Pathognomonic is a word that refers to the speci�c characteristics ofdisease.
� Immunity is the speci�c resistance to disease.
� Virulence is the degree of pathogenicity or the capacity of an organismto produce disease.
� Pathology is the study of disease. It is derived from Pathos (�su�ering�)and logos which means (�science�). Pathology is that branch of discourseconcerned with the structural and functional changes that occur due to adisease-causing agent or pathogen. A pathologist is a scientist or physicianwho studies the cause of diseases, or etiology, and pathogenesis, themanner in which a disease develops
12.2 Some bacterial diseases of humans
12.2.1 Waterborne transmission: cholera
The causative agentVibrio cholerae is ingested in faecally contaminated water orfood. The bacteria attach by means of adhesins to the intestinal mucosa, where,without actually penetrating the cells, they release the cholera exotoxin. Thiscomprises an `A' and several `B' subunits; the former is the active ingredient,while the latter attach to epithelial cells by binding to a speci�c glycolipid inthe membrane. This allows the passage of the `A' subunit into the cell, whereit causes the activation of an enzyme called adenylate cyclase. This results inuncontrolled production of cyclic AMP, causing active secretion of chloride andwater into the intestinal lumen. The outcome of this is huge �uid loss (10 l ormore per day) through profuse and debilitating diarrhoea. In the young, oldand sick, death through dehydration and salt depletion can follow within a veryshort time. If proper liquid and electrolyte replacement therapy is available,recovery rates can be very high. Although it is now very rare in the developedworld (in 2002, only two cases were reported in the USA), cholera is a majorkiller in the third world. It is easily preventable by means of clean water suppliesand improved sanitation, however when these services break down, for exampleduring war or after an earthquake, cholera outbreaks quickly follow. This is thesituation currently developing in Zimbabwe.
77
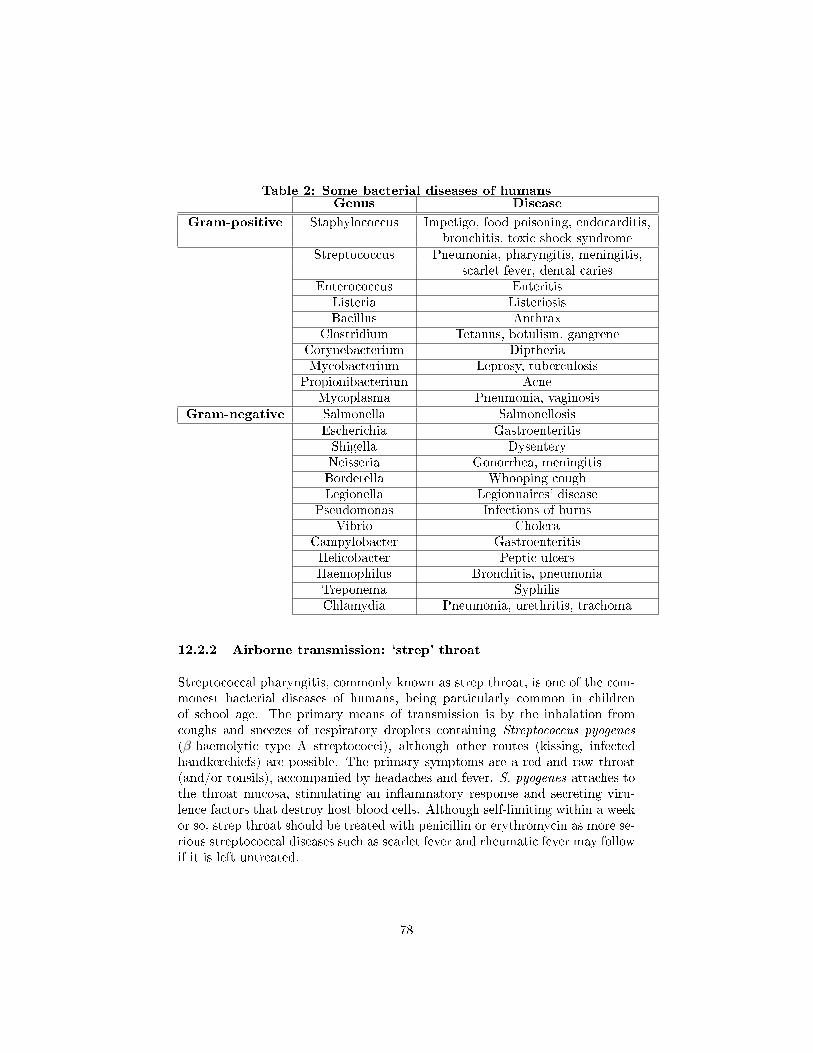
Table 2: Some bacterial diseases of humansGenus Disease
Gram-positive Staphylococcus Impetigo, food poisoning, endocarditis,bronchitis, toxic shock syndrome
Streptococcus Pneumonia, pharyngitis, meningitis,scarlet fever, dental caries
Enterococcus EnteritisListeria ListeriosisBacillus Anthrax
Clostridium Tetanus, botulism, gangreneCorynebacterium DiptheriaMycobacterium Leprosy, tuberculosis
Propionibacterium AcneMycoplasma Pneumonia, vaginosis
Gram-negative Salmonella SalmonellosisEscherichia GastroenteritisShigella DysenteryNeisseria Gonorrhea, meningitisBordetella Whooping coughLegionella Legionnaires' disease
Pseudomonas Infections of burnsVibrio Cholera
Campylobacter GastroenteritisHelicobacter Peptic ulcersHaemophilus Bronchitis, pneumoniaTreponema SyphilisChlamydia Pneumonia, urethritis, trachoma
12.2.2 Airborne transmission: `strep' throat
Streptococcal pharyngitis, commonly known as strep throat, is one of the com-monest bacterial diseases of humans, being particularly common in childrenof school age. The primary means of transmission is by the inhalation fromcoughs and sneezes of respiratory droplets containing Streptococcus pyogenes
(β-haemolytic type A streptococci), although other routes (kissing, infectedhandkerchiefs) are possible. The primary symptoms are a red and raw throat(and/or tonsils), accompanied by headaches and fever. S. pyogenes attaches tothe throat mucosa, stimulating an in�ammatory response and secreting viru-lence factors that destroy host blood cells. Although self-limiting within a weekor so, strep throat should be treated with penicillin or erythromycin as more se-rious streptococcal diseases such as scarlet fever and rheumatic fever may followif it is left untreated.
78

12.2.3 Contact transmission: syphilis
Causative organisms of sexually transmitted diseases such as Neisseria gonor-
rhoeae (gonorrhoea) and Treponema pallidum (syphilis) are extremely sensitiveto the e�ects of environmental factors such as UV light and desiccation. Theyare therefore unable to live outside of their human host, and rely for transmis-sion on intimate human contact. The spirochaete T. pallidum enters the bodythrough minor abrasions, generally on the genitalia or mouth, where a charac-teristic lesion called a chancre develops. The disease may proceed no furtherthan this, but if T. pallidum enters the bloodstream and passes around thebody, the more serious secondary stage develops, lasting some weeks. Followinga latent period of several years, around half of secondary syphilis cases go onto develop into the tertiary stage of the disease, whose symptoms may includemental retardation, paralysis and blindness. Congenital syphilis is caused byT. pallidum being passed from a mother to her unborn child. The primaryand secondary stages of syphilis are readily treated by penicillin; however, thetertiary stage is much less responsive to such therapy.
12.2.4 Vector-borne transmission: plague
A limited number of bacterial diseases reach their human hosts via an insectintermediary from their main host, usually another species of mammal. Plague(bubonic plague, the Black Death) has been responsible for the deaths of untoldmillions of people in terrible epidemics such as the ones that wiped out as muchas one-third of the population of Europe in the Middle Ages. It is caused bythe Gram-negative bacterium Yersinia pestis, whose normal host is a rat, butcan be spread to humans by �eas. The bacteria pass to the lymph nodes, wherethey multiply, causing the swellings known as bubos. Y. pestis produces anexotoxin, which prevents it from being destroyed by the host's macrophages;instead, it is able to multiply inside them.
From the lymph nodes, the bacteria spread via the bloodstream to other tissuessuch as the liver and lungs. Once established in the lungs (pneumonic plague),plague can spread from human to human by airborne transmission in respira-tory droplets. Untreated, plague has a high rate of fatality, particularly for thepneumonic form of the disease. Early treatment with streptomycin or tetracy-cline, however, is largely successful. Improved public health measures and theawareness of the dangers of rats and other rodents have meant that con�rmedcases of plague are now relatively few.
12.2.5 Food poisoning: botulism
Clostridium botulinum is a Gram-positive, rod shaped bacterium that producesthe neurotoxin botulin, which causes the �accid muscular paralysis seen in bo-tulism. It is also the main paralytic agent in botox. It is an anaerobic spore-
79

former, which produces oval, subterminal endospores and is commonly found insoil.
Normal symptoms usually include dry mouth, double and/or blurred vision,di�culty swallowing, muscle weakness, drooping eyelids, di�cult breathing,slurred speech, vomiting, urinary incontinence and sometimes diarrhea. Thesesymptoms will eventually lead to body paralysis. The respiratory muscles area�ected as well, which may cause death due to respiratory failure. These are allsymptoms of the muscle paralysis caused by the bacterial toxin.
In all cases illness is caused by the toxin made by C. botulinum, not by thebacterium itself, thus it is a food poisoning organism, rather than a foodinfection organism. The pattern of damage occurs because the toxin a�ectsnerves that are �ring more often.
12.2.6 Food infection: salmonellosis
Over 1200 types of Salmonella exist. All are potentially dangerous to people.Salmonella live in the intestinal tracts of humans and animals and are contin-ually passed from person to animal, animal to person, and person to person ina continuous cycle. The prime sources of Salmonella contaminants of our foodsupply come from the intestines of animals. Vermin such as rodents, roachesand �ies also carry Salmonella.
Salmonellosis has occurred from the consumption of contaminated foods suchas cheese, milk, eggs, meat, poultry, pastries, and cakes. Salmonellosis is causedwhen we eat foods which contain the organism. This is a food infection. Theseorganisms continue to grow and multiply in the small intestines. The result issickness eight to 24 hours after we eat the contaminated food.
Salmonellosis is characterized by an abrupt onset of diarrhea, nausea, abdominalpain, prostration, chills, fever and vomiting. These symptoms vary in intensityfrom slight to severe. The symptoms rarely cause death except to infants or theelderly who may rapidly dehydrate
Salmonellosis is the most widespread of all foodborne illness. More than 20,000cases are reported to the Center for Disease Control annually. This is probablyonly a small percent of the cases that occur each year.
12.3 Fungi and disease
A limited number of fungi are pathogenic to humans (see table above). Mycoses(sing: mycosis) in humans may be cutaneous, or systemic; in the latter, sporesgenerally enter the body by inhalation, but subsequently spread to other organsystems via the blood, causing serious, even fatal disease.
Cutaneous mycoses are the most common fungal infections found in humans,and are caused by fungi known as dermatophytes, which are able to utilise
80

Table 3: Some fungal diseases of humansDisease Fungus
Histoplasmosis Histoplasma capsulatumBlastoplasmosis Blastomyces dermatitidisCryptococcosis Cryptococcus neoformans
Cutaneous mycoses Trichophyton spp.Pneumocystis pneumonia Pneumocystis cariniiCandidiasis (`thrush') Candida albicans
Aspergillosis Aspergillus fumigatus
the keratin of skin, hair or nails by secreting the enzyme keratinase. Popularnames for such infections include ringworm and athletes' foot. They are highlycontagious, but not usually serious conditions.
Systemic mycoses can be much more serious, and include conditions such ashistoplasmosis and blastomycosis. The former is caused by Histoplasma capsu-
latum, and is associated with areas where there is contamination by bat or birdexcrement. It is thought that the number of people displaying clinical symp-toms of histoplasmosis represents only a small proportion of the total numberinfected. If con�ned to the lungs, the condition is generally self-limiting, butif disseminated to other parts of the body such as the heart or central nervoussystem, it can be fatal. The causative agents of both diseases exhibit dimor-phism; they exist in the environment as mycelia but convert to yeast at thehigher temperature of their human host.
Aspergillus fumigatus is an example of an opportunistic pathogen, that is, anorganism which, although usually harmless, can act as a pathogen in individ-uals whose resistance to infection has been lowered. Other opportunistic my-coses include candidiasis (`thrush') and Pneumocystis pneumonia. The latteris found in a high percentage of acquired immune de�ciency syndrome (AIDS)patients, whose immune defences have been compromised. The causative or-ganism, Pneumocystis carinii, was previously considered to be a protozoan, andhas only been classed as a fungus in the last decade, as a result of DNA/RNAsequence evidence. It lives as a commensal in a variety of mammals, and isprobably transmitted to humans through contact with dogs. The incidence ofopportunistic mycoses has increased greatly since the introduction of antibi-otics, immunosuppressants and cytotoxic drugs. Each of these either suppressesthe individual's natural defences, or eliminates harmless microbial competitors,allowing the fungal species to �ourish.
12.3.1 Ergot
Members of the genus Claviceps may infect a variety of grains, particularly rye,when they come into �ower, giving rise to the condition called ergot. No great
81

damage is caused to the crop, but as the fungus develops in the maturing grain,powerful hallucinatory compounds are produced, which cause ergotism in thosewho consume bread made from the a�ected grain. This was relatively commonin the Middle Ages, when it was known as St Anthony's Fire. The hallu-cinatory e�ects of ergotism have been put forward by some as an explanationfor outbreaks of mass hysteria such as witch hunts and also for the cause ofthe abandonment of the ship, the Mary Celeste. The e�ects can go beyond thepsychological causing convulsions and even death. In small controlled amounts,the drugs derived from ergot can be medically useful in certain situations suchas the induction of childbirth and the relief of migraine headaches.
12.3.2 Mycotoxins
Many fungi produce natural mycotoxins; these are secondary metabolites,which, if consumed by humans, can cause food poisoning that can sometimes befatal. Certain species of mushroom ('toadstools') including the genus Amanitacontain syubstances that are highly poisonous to humans. Other examples ofmycotoxin illnesses include ergotism and a�atoxin poisoning. A�atoxins are car-cinogenic toxins produced by Aspergillus �avus that grows on stored peanuts.In the early 1960s, the turkey industry in the UK was almost crippled by 'TurleyX disease', caused by the consumption of feed contaminated by A. �avus.
12.3.3 Parasitism
It is thought likely that all animals are parasitised by one fungus or another.extraordinary though it may seem, there are even fungi that act as predatorson small soil animals such as nematode worms, producing constrictive hyphalloops that tighten, immobilising the prey.
12.3.4 Fungal diseases of plants
Fungi also cause disease in plants, and can have a devastating e�ect on cropsof economic importance, either on the living plant or in storage subsequent toharvesting. Rusts, smuts and mildews are all examples of common plant diseasescaused by fungi.
12.4 Protists and human disease
12.4.1 Malaria
Protist: Plasmodium
Vector: Anopholes Mosquito
82

� According to the World Health Organization, 300-500 million cases ofmalaria occur each year
� Malaria results in 1.5-2.7 million deaths per year (much more than AIDS)
� Most cases occur in Africa and South America
� Symptoms include fever, headache, vomitting and other �u-like symptoms
� The protist lives inside the bloodstrea, eventually clogging capillaries anddestroying blood cells, which will lead to death if not treated
12.4.2 African Sleeping Sickness (or Trypanosomiasis)
Protist: Trypanosoma
Vector: Tse Tse Fly
� Occurs mostly in sub-saharan africa
� Symptoms include fever, headaches, pain in joints followed by a phasewhen the parasite infects the central nervous system, causing confusion,lack of coordination, and uncontrolled sleepiness. Without treatment, thehost will die
12.5 Viral disease
Viruses are responsible for causing some of the most serious infectious diseasesto a�ect humans.
12.5.1 Airborne transmission: in�uenza
In�uenza is a disease of the respiratory tract caused by members of the Or-
thomyxoviridae. Transmission occurs as a result of inhaling airborne respiratorydroplets from an infected individual. Infection by the in�uenza virus resultsin the destruction of epithelial cells of the respiratory tract, leaving the hostopen to secondary infections from bacteria such as Haemophilus in�uenzae andStaphylococcus aureus. It is these secondary infections that are responsible forthe great majority of fatalities caused by in�uenza. Generally, su�erers fromin�uenza recover completely within 10�14 days, but some people, notably theelderly and those with chronic health problems, may develop complications suchas pneumonia.
83

12.5.2 Transmission by water or food: viral gastroenteritis
The cause of gastroentiritis may be bacterial or viral. The major cause of theviral form is the human rotavirus , which, together with the Norwalk virus, isresponsible for the majority of reported cases. The rotavirus has a segmented,dsRNA genome, and is a non-enveloped virus.
The virus damages the villi in the upper part of the intestinal tract, a�ectingnormal ion transport, and resulting in the characteristic water loss. Transmis-sion of gastroenteritis is via the faecal�oral route, that is, by the ingestion offaecally contaminated food or water. Poor hygiene practice or contaminatedwater supplies are usually to blame for the perpetuation of the cycle. Normally,the condition is self-limiting, lasting only a couple of days; the normal treat-ment is �uid replacement therapy. In areas where clean water supplies are notavailable, however, the outcome can be much more serious. In the Third World,the condition is a major killer; it is the principal cause of infant mortality, andthe cause of some �ve to ten million deaths per year.
12.5.3 Latent and slow (persistent) viral infections
After an infection has passed, a virus may sometimes remain in the body forlong periods, causing no harm. It may be reactivated, however, by stress orsome change in the individual's health, and initiate a disease state. Well knownexamples of latent viral infections are cold sores and shingles, both caused bymembers of the herpesvirus family. A virus of this sort will remain with anindividual throughout their lifetime.
Whereas latent virus infections are characterised by a sudden increase in virusproduction, in persistent (slow) infections the increase is more gradual, buildingup over several years. Such infections have a serious e�ect on the target cells, andare generally fatal. An example is the measles virus, which can re-manifest itselfafter many years in a rare condition called subacute sclerosing panencephalitis.
12.5.4 Viruses and cancer
A number of chemical and physical agents are known to trigger the uncontrolledproliferation of cells that characterise cancers, but in the last two decades it hasbecome clear that at least six types of human cancer can be virally induced.How do cells lose control of their division, and how are viruses able to bring thisabout? It is now known that cells contain genes called protooncogenes, involvedin normal cell replication. They are normally under the control of other, tumour-suppressor genes, but these can be blocked by proteins encoded by certain DNAviruses. When this happens, the protooncogene functions as an oncogene, andcell division is allowed to proceed uncontrolled. Retroviruses have a di�erentmechanism; they carry their own, altered, version of the cellular oncogene, whichbecomes integrated into the host's genome and leads to uncontrolled cell growth.
84

12.5.5 Virus vaccine
Smallpox, once the scourge of millions, was in 1979 the �rst infectious diseaseto be declared successfully eradicated. This followed a worldwide campaign ofvaccination by the World Health Organisation over the previous decade, andwas made feasible by the fact that humans are the only reservoir for the virus.Vaccination is a preventative strategy that aims to stimulate the host immunesystem, by exposing it to the infectious agent in question in an inactivated orincomplete form.
12.5.6 Interferons (IFNs)
Interferons are natural proteins produced by the cells of the immune systemof most vertebrates in response to challenges by foreign agents such as viruses,parasites and tumor cells. Interferons belong to the large class of glycopro-teins known as cytokines. Interferons are produced by a wide variety of cellsin response to the presence of double-stranded RNA, a key indicator of viralinfection. Interferons assist the immune response by inhibiting viral replicationwithin host cells, activating natural killer cells and macrophages, increasingantigen presentation to lymphocytes, and inducing the resistance of host cellsto viral infection.
Along with their natural function, interferons have antiviral, and antisepticproperties when administered as drugs.
Interferon therapy is used (in combination with chemotherapy and radiation)as a treatment for many cancers.
More than half of hepatitis C patients treated with interferon respond with viralelimination (sustained virological response), better blood tests and better liverhistology (detected on biopsy). There is some evidence that giving interferonimmediately following infection can prevent chronic hepatitis C. However, peopleinfected by HCV often do not display symptoms of HCV infection until monthsor years later making early treatment di�cult.
12.6 Immunity
An immune system is an organism's protection from invading organisms and for-eign substances, such as bacteria, viruses, fungi, protozoa, pollen, transplantedtissues, and insect venom. Parts of these substances are called antigens, or im-munogens. Antigens can be polysaccharides or proteins and provoke a speci�cimmune response in organisms.
Immunity is a defensive response of a host when a foreign substance or organ-ism invades it.
85

12.6.1 Non-speci�c immunity
Non-speci�c immunity is the �rst line of defense against invading organisms.
� Mast cells - Mast cells are a type of innate immune cell that resides inthe connective tissue and in the mucous membranes, and are intimatelyassociated with defense against pathogens, wound healing, but are alsooften associated with allergy and anaphylaxis. When activated, mastcells rapidly release characteristic granules, rich in histamine and heparin,along with various hormonal mediators, and chemokines, or chemotacticcytokines into the environment. Histamine dilates blood vessels, caus-ing the characteristic signs of in�ammation, and recruits neutrophils andmacrophages.
� Phagocytosis - The word 'phagocyte' literally means 'eating cell'. Theseare immune cells that engulf, i.e. phagocytose, pathogens or particles. Toengulf a particle or pathogen, a phagocyte extends portions of its plasmamembrane, wrapping the membrane around the particle until it is en-veloped (i.e. the particle is now inside the cell). Once inside the cell, theinvading pathogen is contained inside an endosome which merges with alysosome. The lysosome contains enzymes and acids that kill and digestthe particle or organism. Phagocytes generally patrol the body searchingfor pathogens, but are also able to react to a group of highly specializedmolecular signals produced by other cells, called cytokines. The phago-cytic cells of the immune system include macrophages, neutrophils, anddendritic cells.
Phagocytosis of the hosts' own cells is common as part of regular tis-sue development and maintenance. When host cells die, either internallyinduced by processes involving programmed cell death (also called apopto-sis), or caused by cell injury due to a bacterial or viral infection, phagocyticcells are responsible for their removal from the a�ected site. By helpingto remove dead cells preceding growth and development of new healthycells, phagocytosis is an important part of the healing process followingtissue injury.
� Neutrophils - Neutrophils, along with two other cell types; eosinophilsand basophils (see below), are known as granulocytes due to the presenceof granules in their cytoplasm, or as polymorphonuclear cells (PMNs) dueto their distinctive lobed nuclei. Neutrophil granules contain a variety oftoxic substances that kill or inhibit growth of bacteria and fungi. Similarto macrophages, neutrophils attack pathogens by activating a respiratoryburst. The main products of the neutrophil respiratory burst are strongoxidizing agents including hydrogen peroxide, free oxygen radicals andhypochlorite. Neutrophils are the most abundant type of phagocyte, nor-mally representing 50 to 60% of the total circulating leukocytes, and are
86

usually the �rst cells to arrive at the site of an infection. The bone marrowof a normal healthy adult produces more than 100 billion neutrophils perday, and more than 10 times that many per day during acute in�amma-tion.
� Macrophages - Macrophages, from the Greek, meaning "large eatingcell", are large phagocytic leukocytes, which are able to move outside ofthe vascular system by moving across the cell membrane of capillary vesselsand entering the areas between cells in pursuit of invading pathogens. Intissues, organ-speci�c macrophages are di�erentiated from phagocyticcells present in the blood called monocytes. Macrophages are the moste�cient phagocytes, and can phagocytose substantial numbers of bacteriaor other cells or microbes. The binding of bacterial molecules to receptorson the surface of a macrophage triggers it to engulf and destroy the bac-teria through the generation of a �respiratory burst�, causing the releaseof reactive oxygen species. Pathogens also stimulate the macrophage toproduce chemokines, which summons other cells to the site of infection.
� Basophils and Eosinophils - Basophils and eosinophils are cells relatedto the neutrophil (see above). When activated by a pathogen encounter,basophils releasing histamine are important in defense against parasites,and play a role in allergic reactions (such as asthma). Upon activation,eosinophils secrete a range of highly toxic proteins and free radicals thatare highly e�ective in killing bacteria and parasites, but are also responsi-ble for tissue damage occurring during allergic reactions. Activation andtoxin release by eosinophils is therefore tightly regulated to prevent anyinappropriate tissue destruction.
� Natural killer (NK) and lymphokine activated killer (LAK) cells- NK cells attack host cells that have been infected by microbes, but donot directly attack invading microbes. For example, NK cells attack anddestroy tumor cells, and virus-infected cells, through a process known as"missing-self." This term describes cells with low levels of a cell-surfacemarker called MHC I (major histocompatibility complex) - a situationthat can arise in viral infections of host cells.
12.6.2 Speci�c immunity
Antibodies are proteins that are made by the body in response to an antigenand can combine speci�cally with that antigen. The immune system recognizesa body or substance within the organism as self or nonself. Self is any bodyor substance that belongs to the organism. Nonself is any body or organismthat doesn't belong to the organism. An antigen is recognized as nonself by theimmune system.
An antigen causes the organism to form antibodies and specialized lympho-cytes that target the speci�c antigen. Lymphocytes can be eitherT-lymphocytes
87

or B-lymphocytes. Once the infection is defeated, some of the T and B cellsbecomy memory cells. If the antigen invades again, these lymphocytes and an-tibodies attack the antigen, making the antigen inactive or destroyed. This iscalled a speci�c immune response against an antigen.
12.7 Vaccines
A vaccine is a suspension that contains a part of a pathogen that induces theimmune system to produce antibodies that combat the antigen. The conceptof a vaccine stems from the variolation process that was used in eighteenth-century England to protect people from smallpox.
The variolation process requires that a needle tip of smallpox be placed in thevein of a patient. Nearly all the patients contracted a mild case of smallpox,which left them with antibodies that protected them from contracting the dis-ease. Half of the patients who contracted smallpox died. By contrast, only onepercent who received the variolation process died.
Edward Jenner noticed that dairymaids who contracted cowpox, which isrelated chemically to smallpox, were immune to smallpox. Jenner discoveredthat injecting cowpox into the skin of a healthy person prevented them fromdeveloping smallpox. Jenner's discovery enabled Louis Pasteur to develop thetechnique of creating vaccines.
The injection of an antigen induces the primary immune and secondary immuneresponses in the patient. The primary immune response produces antibodiesand the secondary immune response produces memory cells that attack afuture invasion of the antigen.
Vaccines play an important role in controlling the spread of viruses. A viruscannot be treated with antibiotics. However, you can minimize catching the �uby getting a �u shot, which is a vaccine against a particular strain of �u virus.
Vaccines also prevent bacterial infections such as typhoid, but are not as e�ec-tive on bacteria as they are on viruses. However, bacteria infections are treatablewith antibiotics, which is a common method of combating bacterial diseases.
Scientists use vaccines to provide herd immunity to a population. Herd im-munity requires that most�not all�of the population be immunized in orderto prevent an epidemic of a disease. An outbreak of a disease would be isolatedto a small percentage of the population and therefore have a minimum e�ect.
12.7.1 Types of vaccines
Passive Immunisation. Protection is conferred by giving the person anti-bodies. Mainly given as pre- or immediately post exposure prophylaxis.Protection is usually short-lived as the recipients immune system is not
88

involved. Tetanus, botulism, hepatitis B, and rabies are amongst the dis-ease for which passive immunisation (post-exposure) can be important.Also used for snake and spider bites.
Active Immunisation. These vaccines initiate the recipient's own defencesystem to mount a defence. The majority of commonly administered vac-cines are active immunisations. Immunity can be life-long, however inmany cases boosters are given periodically
Attenuated whole agent. a.k.a live-attenuated. These vaccines are de-signed for people who have a normal immune system. The attenuatedwhole agent vaccine uses weakened living microbes to mimic the real in-fection to produce 95 percent immunity over a long term without theneed of a supplemental vaccination called a booster. Common attenu-ated whole agent vaccines include those for Tuberculosis bacillus, measles,rubella, Sabin polio, and mumps. There is a risk that live microorganismsor virus can regain their strength resulting in the patient contracting thedisease.
Inactivated whole agent. These vaccines are not designed for people whohave an abnormal immune system. The inactivated whole agent vaccineuses dead microbes that were killed by phenol or formalin. Commoninactivated whole agent vaccines include those for pneumonia, Salk polio,rabies, in�uenza, typhoid, and pertussis (commonly known as whoopingcough).
Toxoids. The toxoid vaccine is made of toxins produced by a virus or bacte-ria that has been inactive. They are then used against toxins that areproduced by a disease-causing microorganism. Patients require a boostervaccination every 10 years because the toxoid vaccine does not providelifelong immunity. Common toxoid vaccines include those for diphtheriaand tetanus.
Subunit. These vaccines have few side e�ects. The subunit vaccine uses frag-ments of a microorganism to create an immune response. Subunit vaccinesare produced by using genetic engineering techniques to insert the genes ofan antigen into another organism are called recombinant vaccines. Com-mon subunit vaccines include those for hepatitis B.
Surrogate vaccines. There are couple of vaccines that do not contain thebacterium they are used to protect against. BCG contains Mycobacterium
bovis but is used to protect against Mycobacterium tuberculosis and crossprotects against Mycobacterium leprae. The vaccine used to eradicatesmallpox contained the related cowpox virus.
Conjugated. These are fairly new in development and are designed for chil-dren under 24 months whose immune system normally does not respond
89

well to vaccines based on capsular polysaccharides. These polysaccha-rides are T-independent antigens. The vaccine is produced by combingthe polysaccharides with a protein.
Nucleic acid. These vaccines are in the animal testing stage. The nucleicacid vaccine, which is also called the DNA vaccine, contains plasmids ofnaked DNA and is designed to produce protein that stimulates an immuneresponse. The nucleic acid vaccine has a strong e�ect on large parasitesand viruses.
12.7.2 Developing a vaccine
Vaccines are developed by cultivating a large quantity of pathogen, which isa disease causing organism. Some pathogens, such as the rabies virus, can becultivated in animals. For example, a chick embryo is commonly used to growviruses and is the method used to develop the in�uenza vaccine.
When vaccines were �rst introduced against measles and polio they would onlygrow in humans. However, with the development of cell culture techniques, cellsfrom humans and primates enable large-scale viral growth.
Scientists use recombinant vaccines because they do not need an animal host togrow the microorganism. An example is hepatitis B.
90

13 Importance in natural world: associations
13.1 Types of microbial associations
Microorganisms in nature do not exist as pure cultures but alongside numerousother organisms, microbial or otherwise, with which they may have to competein the never-ending struggle for survival. In a number of cases, this coexistencemay extend beyond merely sharing the same environmental niche; some mi-croorganisms form a close physical association with another type of organism,from which special bene�ts may accrue for one or both parties. Such associa-tions are termed collectively symbiosis ([greek] `living together'). Three generalforms of symbiotic relationship may be de�ned:
13.1.1 Parasitism:
An association from which one partner derives some or all of its nutritionalrequirements by living either in or on the other (the host), which usually su�ersharm as a result.
13.1.2 Mutualism:
An association from which both participants derive bene�t. The relationship isfrequently obligatory, that is, both are dependent upon the other for survival.Non-obligatory mutualism is sometimes called protocooperation.
13.1.3 Commensalism:
An association from which one participant (the commensal) derives bene�t,and the other is neither bene�ted nor harmed. The relationship is not usuallyobligatory.
13.2 Microbial association with animals
13.2.1 Termites
Termites are insects belonging to the order Isoptera that are found particularlyin tropical regions. Their famous ability to destroy trees and wooden structuressuch as buildings and furniture is due to a resident population of �agellatedprotozoans in their hindgut, which are able to break down cellulose. Termiteslack the enzymes necessary to do this, and would thus starve to death if theprotozoans were not present. In return, they are able to provide the anaerobicconditions required by the protozoans to ferment the cellulose to acetate, carbon
91

dioxide and hydrogen. The acetate is then utilised as a carbon source by thetermites themselves.
In addition to the protozoans, anaerobic bacteria resident in the hindgut alsoplay an important role in the metabolism of the termites. Acetogenic andmethanogenic species compete for the carbon dioxide and hydrogen producedby the protozoans. The former contribute more acetate for the termite to use,whilst the latter produce signi�cant amounts of methane. Some methanogensexist as endosymbionts within the protozoans.
In other types of termite, no resident population of cellulose digesters is present.Instead, the termite ingests a fungus, which proivdes the necessary cellulolyticenzymes.
13.2.2 The honey gude bird
Another example of a hosts's staple diet being indigestible without the assistanceof resident microorganisms is provided by the brightly coloured African bird thehoney guide. The honey guide eats beeswax, and relies on a two-stage digestionprocess by bacteria (Micrococcus cerolyticus) and yeast (Candida albicans) torender it in a usable form.
13.2.3 Giant tube worms
At the bottom of the deepest oceans, around geothermal vents, live enormous(two metres or more) tube worms belonging to the genus Riftia. These lackany sort of digestive system, but instead contain in their body cavity a tissueknown as the trophosome. This comprises vascular tissue plus cells packedwith endosymbiotic bacteria. These are able to generate ATP and NADPHby the oxidation of hydrogen sulphide generated by volcanic activity and �xcarbon dioxide via the Calvin cycle, providing the worm with a supply of organicnutrients. Hydrogen sulphide is transported to the trophosome from the worm'sgill plume by a form of haemoglobin present in its blood.
13.2.4 Humans
Warm-blooded animals such as humans play host in their lower intestinal tractto vast populations of bacteria. Although some of these are capable of producinguseful metabolites such as vitamin K, most live as commensals, neither bene�t-ing nor harming their host. It could be argued, however, that the very presenceof the resident intestinal micro�ora acts as an important defence against coloni-sation by pathogens, thus making the association more one of mutualism. Seesection on human microbiology.
92

13.3 Microbial associations with plants
The roots of almost all plants form mutualistic associations with fungi, knownas mycorrhizae, which serve to enhance the uptake of water and mineral nutri-ents, especially phosphate, by the plants. The bene�cial e�ect of a mycorrhizalassociation is particularly noticeable in soils with a poor phosphorus content.In return, the plant supplies reduced carbon in the form of carbohydrates to thefungi.
Unlike other plant-microorganism interactions that occur in the rhizosphere (theregion around the surface of a plant's root system), mycorrhizal associationsinvolve the formation of a distinct, integrated structure comprising root cellsand fungal hyphae.
13.3.1 Ectomycorrhizae
In ectomycorrhizae the plant partner is always a tree; the fungus surrounds theroot tip, and hyphae spread between (but do not enter) root cells.
13.3.2 Endomycorrhizae
In the case of the more common endomycorrhizae, the fungal hyphae actuallypenetrate the cells by releasing cellulolytic enzymes. Arbuscular mycorrhizaeare found in practically all plant types, including `lower' plants (mosses, ferns).They form highly branched arbuscules within the root cells that gradually lyse,releasing nutrients into the plant cells. In contrast to pathogenic fungi, mycor-rhizal fungi are often rather non-speci�c in their choice of `partner' plant.
13.3.3 Nitrogen-�xing bacteria
The ability of crop plants to thrive is frequently limited by the supply of availablenitrogen; although there is a lot of it in the atmosphere, plants are unable toutilise it, and instead must rely on an inorganic supply (both naturally-occurringand in the form of fertilisers). However, certain bacterial species are able to `�x'atmospheric nitrogen into a usable form.
Some of these, notably Rhizobium spp. form a mutualistic relationship withleguminous plants such as peas, beans and clover, converting nitrogen to ammo-nia, which the legume can incorporate into amino acids. In return, the bacteriareceive a supply of organic carbon, which they can use as an energy source forthe �xation of nitrogen.
The free-living Rhizobium enters the plant via its root hairs, forming an infectionthread and infecting more and more cells. Normally rod-shaped, they proliferateas irregularly-shaped bacteroids, densely packing the cells and causing them to
93

swell, forming root nodules. Rhizobium requires oxygen as a terminal electronacceptor in oxidative phosphorylation, but the nitrogenase enzyme, which �xesthe nitrogen, is sensitive to oxygen. The right microaerophilic conditions aremaintained by means of a unique oxygen-binding pigment, leghaemoglobin.This is only synthesised by means of a collaboration between both partners.Nitrogen �xation requires a considerable input of energy in the form of ATP(16 molecules for every molecule of nitrogen), so when ammonia is in plentifulsupply the synthesis of the nitrogenase enzyme is repressed.
13.4 Microbial associations with other microorganisms
13.4.1 Lichens
The most familiar example of mutualism between microorganisms is that oflichens, which comprise a close association between the cells of a fungus (usuallybelonging to the Ascomycota) and a photosynthetic alga or cyanobacterium.
Although many di�erent fungal species may take part in lichens, only a limitednumber of algae or cyanobacteria do so.
Lichens are typically found on exposed hard surfaces such as rocks, tree barkand roofs, and grow very slowly at a rate of a milimetre or two per year. Theyoften occupy particularly harsh environments, from the polar regions to thehottest deserts.
The photosynthetic partner usually exists as a layer of cells scattered amongfungal hyphae. Often unicellular, it �xes carbon dioxide as organic matter,which the heterotrophic fungus absorbs and utilises.
The fungal member provides anchorage and supplies inorganic nutrients andwater, as well as protecting the alga from excessive exposure to sunlight.
Although lichens are tolerant of extremes of temperature and water loss, theyhave a well-known sensitivity to atmospheric pollutants such as the oxides ofnitrogen and sulphur. Their presence in an urban setting is therefore a usefulindicator of air quality. Lichens were used for many years as a source of brightlycoloured dyes for the textile industry; they are also used in the perfume industry.The dye is litmus paper is derived from a lichen, belonging to the genus Roccella.
It should be stressed that lichens are not just a mixture of fungal and algalcells. They are distinctive structures with properties not possessed by either oftheir component species. Indeed, the relationship between the two partners of alichen is so intimate that the composite organisms are given taxonomic status.Many thousands of species of lichen have been identi�ed.
13.4.2 E.coli and Enterococcus faecalis
The gut bacteria E. coli and Enterococcus faecalis cooperate to utilise arginine.As shown by the �gure below, neither can usefully metabolise this amino acid
94

Figure 5: Protocooperation can make available an otherwise unutilisable sub-strate. Individually, neither Enterococcus faecalis nor E. coli is able to utilisearginine; however, working together they can convert it into putrescine, whichcan then be metabolised further by either organism to produce energy
on its own; however, neither is dependent on the reaction.
13.5 A note on secondary metabolites, because they fea-ture in an exam question
There is a question in the example paper which is very confusingly phrased. Itrefers to 'secondary products', which is not a standard or de�ned term.
If you come across this term, it refers to secondary metabolites.
Secondary metabolites are organic compounds that are not directly involved inthe normal growth, development or reproduction of organisms. Unlike primarymetabolites, absence of secondary metabolities results not in immediate death,but in long-term impairment of the organism's survivability, fecundity (ability toundergo healthy development) or aesthetics, or perhaps in no signi�cant changeat all.
All biologically produced antibiotics are secondary products e.g strep-tomycin, penicillin etc.
In addition, other important pharmaceuticals are produced as secondary metabo-lites - things such as:
95

� Insulin
� Factor VIII
� Human growth factor
� Interferons
96

14 Importance in natural world: the environment
All organisms may be placed into one of three categories with respect to theirpart in the global �ow of energy:
1. (Primary) Producers: autotrophs that obtain energy from the sun orchemical sources (e.g. green plants, photosynthetic bacteria, chemolithotrophicbacteria). They use the energy to synthesise organic material from carbondioxide and water.
2. Consumers: heterotrophs that derive energy through the consumptionof other organisms (producers or other consumers). They may serve as alink between the primary producers and the decomposers.
3. Decomposers: organisms that break down the remains and waste prod-ucts of producers and consumers, obtaining energy and releasing nutrients,including CO2, that can be reused by the producers.
Natural systems exist in a balance; carbon and all the other elements that makeup living things are subject to repeated recycling, so that they are available todi�erent organisms in di�erent forms. Algae, green plants and certain bacteriacapture light energy, then use it to synthesise organic carbon compounds fromcarbon dioxide and water. What happens to all this organic carbon? It does notjust accumulate, but is recycled by other living things, which convert it back tocarbon dioxide by respiration. Many other elements such as sulphur, nitrogen,and iron are similarly changed from one form to another in this way, by a cyclicseries of reactions. Microorganisms are responsible for most of these reactions,oxidising and reducing the elements according to their metabolic needs. Thecontinuation of life on Earth is dependent on the cycling of �nite resources inthis way.
14.1 The carbon cycle
Atmospheric CO2 is �xed into organic compounds by plants, together with pho-totrophic and chemoautotrophic microorganisms. The organic compounds thussynthesised undergo cellular respiration and CO2 is returned to the atmosphere.The carbon may have been passed along a food chain to consumers before thisoccurs. Carbon dioxide is also produced by the decomposition of dead plant, an-imal and microbial material by heterotrophic bacteria and fungi. Methanogenicbacteria produce methane from organic carbon or CO2. This in turn is oxidisedby methanotrophic bacteria; carbon may be incorporated into organic materialor lost as CO2.
97

14.2 The nitrogen cycle
Nitrogen is essential to all living things as a component of proteins and nucleicacids. Although elemental nitrogen makes up three quarters of the Earth'satmosphere, only a handful of life forms are able to utilise it for metabolicpurposes. These are termed nitrogen-�xing bacteria, and incorporate thenitrogen into ammonia.
The nitrogenase enzyme complex responsible for the reaction is very sensitiveto oxygen, and is thought to have evolved early in the Earth's history, whenthe atmosphere was still largely oxygen-free. Many nitrogen-�xing bacteria areanaerobes; those that are not have devised ways of keeping the cell interioranoxic. Azotobacter species, for example, utilise oxygen at a high rate, so thatit never accumulates in the cell, inactivating the nitrogenase. Many cyanophytes(blue-greens) carry out nitrogen �xation in thick-walled heterocysts which helpmaintain anoxic conditions.
Some nitrogen-�xing bacteria such as Rhizobium infect the roots of leguminousplants such as peas, beans and clover, where they form nodules and form amutually bene�cial association.
Ammonia produced by nitrogen �xation is assimilated as amino acids, whichcan then form proteins and feed into pathways of nucleotide synthesis. Organicnitrogen in the form of dead plant and animal material plus excrement re-enters the environment, where it undergoes mineralisation at the hands of arange of microorganisms, involving the deamination of amino acids to theircorresponding organic acid. This process of mineralisation may occur aerobicallyor anaerobically, in a wide range of microorganisms.
The process of nitri�cation, by which ammonia is oxidised stepwise �rstly tonitrite and then to nitrate, involves two di�erent groups of bacteria.
Denitri�cation is the reduction, under anaerobic conditions, of nitrite andnitrate to nitrogen gas. The nitrate thus formed may su�er a number offates. It may act as an electron acceptor in anaerobic respiration, becomingreduced to nitrogen via a series of intermediates including nitrite. This processof denitri�cation occurs in anaerobic conditions such as waterlogged soils. Al-ternatively, it can be reduced once again to ammonia and thence converted toorganic nitrogen.
14.3 Bene�cial e�ects of microorganisms in the environ-ment
14.3.1 Solid waste treatment: composting and land�ll
We in the modern Western world are often described as living in a `throwawaysociety'. On average, each of us generates around 2 tonnes of solid waste ma-terial per year, and all of this must be disposed of in some way! Most of it
98

ends up in land�ll sites, huge holes in the ground where refuse is deposited toprevent it being a hazard. The non-biodegradable components (metals, plas-tics, rubble, etc.) remain there more or less inde�nitely; however, over a periodof time biodegradable material (food waste, textiles, paper, etc.) undergoes adecomposition process. The rate at which this happens is dependent on thenature of the waste and the conditions of the land�ll, but could take severaldecades.
Aerobic processes give way to anaerobic ones and a signi�cant result of thelatter is the generation of methane. Modern land�ll sites incorporate systemsthat remove this to prevent it being a �re/explosion hazard, and may put it togood use as a fuel source. Many householders separate organic waste items suchas vegetable peelings and grass cuttings and use them to make compost.
This practice, apart from providing a useful gardening supplement, also sub-stantially reduces the volume of material that has to be disposed of by othermeans (see above). We have already mentioned the role of microorganisms inthe recycling of carbon in the biosphere; these same processes serve to degradethe organic waste, especially the cellulose, resulting in a considerable reductionof the bulk.
Fungi and bacteria, particularly actinomycetes, break down the organic matterto produce CO2, water and humus, a relatively stable organic end product.Compost is not really a fertiliser, since its nitrogen content is not high, butit nevertheless provides nutrients to a soil and generally helps to improve itscondition. Composting is carried out on a large scale by local authorities usingthe waste generated in municipal parks and gardens.
14.3.2 Waste water treatment
The aim of wastewater treatment is the removal of undesirable substances andhazardous microorganisms in order that the water may safely enter a water-course such as a river or stream. Further puri�cation procedures are requiredbefore it can be used as drinking water. Wastewater treatment is fundamen-tal to any developed society, and greatly reduces the incidence of waterbornediseases such as cholera. Wastewater may come from domestic or commer-cial sources; highly toxic industrial e�uents may require pre-treatment beforeentering a water treatment system.
Sewage is the term used to describe liquid wastes that contain faecal matter(human or animal). The e�ectiveness of the treatment process is judged chie�yby the reduction of the wastewater's biochemical oxygen demand (BOD).This is a measure of the amount of oxygen needed by microorganisms to oxidiseits organic content. A high BOD leads to the removal of oxygen from water, acertain indicator of pollution. Wastewater treatment usually occurs in stages,the �rst of which (primary treatment) is purely physical, and involves theremoval of �oating objects followed by sedimentation, a process that removes
99

up to a third of the BOD value. Secondary treatment involves microbialoxidation, leading to a substantial further reduction in BOD.
This may take one of two forms, both of which are aerobic, the traditionaltrickling �lter and the more recent activated sludge process. In the former,the wastewater is passed slowly over beds of stones or pieces of moulded plastic.These develop a bio�lm comprising bacteria, protozoans, fungi and algae, andthe resulting treated water has its BOD reduced by some 80�85 per cent.
Activated sludge plants achieve an even higher degree of BOD reduction. Herethe wastewater is aerated in tanks that have been seeded with a mixed microbialsludge. The main component of this is the bacterium Zoogloea, which secretesslime, forming aggregates called �ocs, around which other microorganisms suchas protozoans attach. Some of the water's organic content is not immediatelyoxidised, but becomes incorporated into the �ocs. After a few hours' residencein the tank, the sludge is allowed to settle out, and the treated water passesout of the system. Before being discharged to a watercourse, it is treated withchlorine to remove any pathogenic microorganisms that may remain. The prin-cipal operating problem encountered with activated sludge is that of bulking.This is caused by �lamentous bacteria such as Sphaerotilus natans; it results inthe sludge not settling properly and consequently passing out with the treatedwater. Both secondary treatment processes result in some surplus sludge, whichundergoes anaerobic digestion, resulting in the production of methane and CO2.The methane can be used as a fuel to power the plant, and any remaining sludgeis dewatered and used as a soil conditioner. Care must be taken in this context,however, that the sludge does not have a high heavy metal content.
14.3.3 Bioremediation
Perhaps the biggest problem facing the developed world at the start of thetwenty �rst century is that of pollution of the environment. Our dependence onthe products of the chemical industries has resulted in the production of vastamounts of toxic waste material. One way of dealing with such (mostly organic)waste is to encourage the growth of bacteria and fungi that are able to oxidisethe pollutants, a process known as bioremediation.
We have seen how microorganisms are able to utilise an enormous range of or-ganic compounds as carbon sources; the Gram-negative bacterium Burkholde-
ria cepacia can use over 100 such compounds. Many organisms can metabolisenot only naturally occurring substances, but also synthetic ones, making themvaluable allies in the process of bioremediation. Often the most e�ective mi-croorganisms to use are those found living naturally at the contaminated site,since they have demonstrated the ability to survive the toxic e�ects of the pollu-tant, although in other cases specially adapted or genetically modi�ed bacteriamay be introduced (bioaugmentation). Examples of the use of microorgan-isms include the treatment of toxic waste sites, chemical spills, pesticides ingroundwater and oil spills.
100

One of the �rst successful applications of bioremediation came in the aftermathof the Exxon Valdez disaster in 1989 when thousands of tons of crude oil werereleased o� the coast of Alaska. Depending on the circumstances, bioremedi-ation procedures may occur in situ, or the contaminated soil or water may beremoved to a specialist facility for treatment.
14.4 Harmful e�ects of microorganisms in the environ-ment
The natural processes of bioconversion that are so important in the global re-cycling of elements may have unwanted consequences for humans. Prominentamong these is acid mine drainage, a frequently encountered problem in miningregions. Bacterial oxidation of mineral sulphides, particularly the ubiquitousiron pyrite, leads to the release of a highly acidic leachate into streams andrivers. This also contains dissolved metals, including ferric iron. When it mixeswith stream water, the pH is raised su�ciently for the iron to precipitate as un-sightly orange ferric hydroxides, blanketing the stream bed and wiping out plantand animal life. The main culprits in the formation of acid mine drainage aresulphur-oxidising bacteria, notably Acidithiobacillus ferrooxidans; as we shallsee in the �nal chapter, under controlled conditions this same organism can alsoprovide economic bene�ts to the mining industry by extracting valuable metalsfrom low grade ores.
Another area in which environmental microorganisms can have detrimental ef-fects is that of biodeterioration, whereby economically important materialssuch as wood, paper, textiles, petroleum and even metals and concrete may besubject to damage by a range of microorganisms, mainly fungi and bacteria.
The most important microorganisms in the biodeterioration of wood are mem-bers of the Basidiomycota. Wood is only susceptible to fungal attack whenits moisture level reaches around 30 per cent. The major component of woodthat is subject to microbial attack is cellulose, although some forms can alsodegrade lignin. There are two main forms of rot; white rot, which involves thedegradation of lignin as well as cellulose, and brown rot, in which the ligninis unaltered. The dry rot fungus Serpula lacrymans produces thick strands ofhyphae called rhizomorphs, which it uses to conduct water and nutrients fromdamper areas. These are very strong, and able to travel over brickwork andmasonry barriers. S. lacrymans is able to generate water as a metabolic endproduct and thus, once established, is able to grow even on dry wood. Dryrot �ourishes in areas of static dampness such as badly ventilated, uninhabitedproperties.
Because cellulose is also an important component of paper and textiles, itsbreakdown is clearly of great economic importance. Degradation by fungi, and,to a lesser extent, bacteria, results in a loss of strength of the material in ques-tion. The paper-making process provides warm, wet conditions rich in nutrients,
101

ideal for microbial growth, which can clog up machinery and discolour the �n-ished product. A variety of biocides are used in an e�ort to minimise microbialcontamination.
The discoloration referred to above raises the point that biodeterioration of amaterial need not necessarily a�ect its physical or chemical make-up; aestheticdamage can lessen the economic value of a material by altering its appearance.The blackening of shower curtains by moulds growing on surface detritus, fa-miliar to generations of students, is another example of this!
102

15 Human indigenous microbiota (normal �ora)
In a healthy animal, the internal tissues, e.g. blood, brain, muscle, etc., arenormally free of microorganisms. However, the surface tissues, i.e., skin andmucous membranes, are constantly in contact with environmental organisms andbecome readily colonized by various microbial species. The mixture of organismsregularly found at any anatomical site is referred to as the normal �ora, exceptby researchers in the �eld who prefer the term "indigenous microbiota".The normal �ora of humans consists of a few eucaryotic fungi and protists,but bacteria are the most numerous and obvious microbial components of thenormal �ora.
15.1 Associations Between Humans and the Normal Flora
E. coli is the best known bacterium that regularly associates itself with humans,being an invariable component of the human intestinal tract. Even though E.
coli is the most studied of all bacteria, and we know the exact location andsequence of 4,288 genes on its chromosome, we do not fully understand itsecological relationship with humans.
In fact, not much is known about the nature of the associations between humansand their normal �ora, but they are thought to be dynamic interactions ratherthan associations of mutual indi�erence. Both host and bacteria are thought toderive bene�t from each other, and the associations are, for the most part, mu-tualistic. The normal �ora derive from their host a steady supply of nutrients,a stable environment, and protection and transport. The host obtains from thenormal �ora certain nutritional and digestive bene�ts, stimulation of the devel-opment and activity of immune system, and protection against colonization andinfection by pathogenic microbes.
While most of the activities of the normal �ora bene�t their host, some ofthe normal �ora are parasitic (live at the expense of their host), and someare pathogenic (capable of producing disease). Diseases that are produced bythe normal �ora in their host may be called endogenous diseases. Mostendogenous bacterial diseases are opportunistic infections, meaning that thethe organism must be given a special opportunity of weakness or let-down inthe host defenses in order to infect. An example of an opportunistic infection ischronic bronchitis in smokers wherein normal �ora bacteria are able to invadethe weakened lung.
Sometimes the relationship between a member of the normal �ora and its hostcannot be deciphered. Many of the normal �ora that are not predominant intheir habitat, even though always present in low numbers, are thought of ascommensal bacteria. However, if a presumed commensal relationship is studiedin detail, parasitic or mutualistic characteristics often emerge.
103

15.2 Tissue speci�city
Most members of the normal bacterial �ora prefer to colonize certain tissuesand not others. This "tissue speci�city" is usually due to properties of both thehost and the bacterium. Usually, speci�c bacteria colonize speci�c tissues byone or another of these mechanisms.
1. Tissue tropism is the bacterial preference or predilection for certain tis-sues for growth. One explanation for tissue tropism is that the host pro-vides essential nutrients and growth factors for the bacterium, in additionto suitable oxygen, pH, and temperature for growth.
2. Speci�c adherence. Most bacteria can colonize a speci�c tissue or sitebecause they can adhere to that tissue or site in a speci�c manner thatinvolves complementary chemical interactions between the two surfaces.Speci�c adherence involves biochemical interactions between bacterial sur-face components (ligands or adhesins) and host cell molecular recep-tors. The bacterial components that provide adhesins are molecular partsof their capsules, �mbriae, or cell walls. The receptors on human cells ortissues are usually glycoprotein molecules located on the host cell or tissuesurface.
3. Bio�lm formation. Some of the indigenous bacteria are able to con-struct bio�lms on a tissue surface, or they are able to colonize a bio�lmbuilt by another bacterial species. Many bio�lms are a mixture of mi-crobes, although one member is responsible for maintaining the bio�lmand may predominate. The classic bio�lm that involves components ofthe normal �ora of the oral cavity is the formation of dental plaque on theteeth. Plaque is a naturally-constructed bio�lm, in which the consortiaof bacteria may reach a thickness of 300-500 cells on the surfaces of theteeth. These accumulations subject the teeth and gingival tissues to highconcentrations of bacterial metabolites, which result in dental disease.
15.3 The composition of the normal �ora
The normal �ora of humans are exceedingly complex and consist of more than200 species of bacteria. The makeup of the normal �ora may be in�uenced byvarious factors, including genetics, age, sex, stress, nutrition and diet of theindividual.
Three developmental changes in humans, weaning, the eruption of the teeth,and the onset and cessation of ovarian functions, invariably a�ect the composi-tion of the normal �ora in the intestinal tract, the oral cavity, and the vagina,respectively. However, within the limits of these �uctuations, the bacterial �oraof humans is su�ciently constant to a give general description of the situation.
104

A human �rst becomes colonized by a normal �ora at the moment of birthand passage through the birth canal. In utero, the fetus is sterile, but whenthe mother's water breaks and the birth process begins, so does colonizationof the body surfaces. Handling and feeding of the infant after birth leads toestablishment of a stable normal �ora on the skin, oral cavity and intestinaltract in about 48 hours. It has been calculated that a human adult houses about1012 bacteria on the skin, 1010 in the mouth, and 1014 in the gastrointestinaltract. The latter number is far in excess of the number of eucaryotic cells in allthe tissues and organs which comprise a human.
15.3.1 Normal �ora of the skin
The adult human is covered with approximately 2 square meters of skin. Thedensity and composition of the normal �ora of the skin varies with anatomicallocale. The high moisture content of the axilla, groin, and areas between the toessupports the activity and growth of relatively high densities of bacterial cells,but the density of bacterial populations at most other sites is fairly low, generallyin 100s or 1000s per square cm. Most bacteria on the skin are sequestered insweat glands.
The skin microbes found in the most super�cial layers of the epidermis and theupper parts of the hair follicles are Gram-positive cocci (Staphylococcus epider-midis and Micrococcus sp.) and corynebacteria such as Propionibacterium sp.
These are generally nonpathogenic and considered to be commensal, althoughmutualistic and parasitic roles have been assigned to them. For example, staphy-lococci and propionibacteria produce fatty acids that inhibit the growth of fungiand yeast on the skin. But, if Propionibacterium acnes, a normal inhabitantof the skin, becomes trapped in hair follicle, it may grow rapidly and cause in-�ammation and acne. Sometimes potentially pathogenic Staphylococcus aureusis found on the face and hands in individuals who are nasal carriers. This isbecause the face and hands are likely to become inoculated with the bacteriaon the nasal membranes. Such individuals may autoinoculate themselves withthe pathogen or spread it to other individuals or foods.
15.3.2 Normal �ora of the conjunctiva
A variety of bacteria may be cultivated from the normal conjunctiva, but thenumber of organisms is usually small. Staphylococcus epidermidis and certaincoryneforms (Propionibacterium acnes) are dominant. Staphylococcus aureus,some streptococci, Haemophilus sp. and Neisseria sp. are occasionally found.The conjunctiva is kept moist and healthy by the continuous secretions from thelachrymal glands. Blinking wipes the conjunctiva every few seconds mechan-ically washing away foreign objects including bacteria. Lachrymal secretions(tears) also contain bactericidal substances including lysozyme. There is littleor no opportunity for microorganisms to colonize the conjunctiva without special
105

mechanisms to attach to the epithelial surfaces and some ability to withstandattack by lysozyme.
Pathogens which do infect the conjunctiva (e.g. Neisseria gonorrhoeae andChlamydia trachomatis) are thought to be able to speci�cally attach to theconjunctival epithelium. Newborn infants may be especially prone to bacterialattachment. Since Chlamydia and Neisseria might be present on the cervicaland vaginal epithelium of an infected mother, silver nitrate or an antibiotic maybe put into the newborn's eyes to avoid infection after passage through the birthcanal.
15.3.3 Normal �ora of the respiratory tract
A large number of bacterial species colonize the upper respiratory tract (na-sopharynx). The nares (nostrils) are always heavily colonized, predominantlywith Staphylococcus epidermidis and corynebacteria, and often (in about 20% ofthe general population) with Staphylococcus aureus, this being the main carriersite of this important pathogen. The healthy sinuses, in contrast are sterile.The pharynx (throat) is normally colonized by streptococci and various Gram-negative cocci. Sometimes pathogens such as Streptococcus pneumoniae, Strep-tococcus pyogenes, Haemophilus in�uenzae and Neisseria meningitidis colonizethe pharynx.
The lower respiratory tract (trachea, bronchi, and pulmonary tissues) is virtu-ally free of microorganisms, mainly because of the e�cient cleansing action ofthe ciliated epithelium which lines the tract. Any bacteria reaching the lowerrespiratory tract are swept upward by the action of the mucociliary blanket thatlines the bronchi, to be removed subsequently by coughing, sneezing, swallowing,etc. If the respiratory tract epithelium becomes damaged, as in bronchitis or vi-ral pneumonia, the individual may become susceptible to infection by pathogenssuch as H. in�uenzae or S. pneumoniae descending from the nasopharynx.
15.3.4 Normal Flora of the Urogenital Tract
Urine is normally sterile, and since the urinary tract is �ushed with urine everyfew hours, microorganisms have problems gaining access and becoming estab-lished. The �ora of the anterior urethra, as indicated principally by urine cul-tures, suggests that the area my be inhabited by a relatively consistent normal�ora consisting of Staphylococcus epidermidis, Enterococcus faecalis and somealpha-hemolytic streptococci. Their numbers are not plentiful, however. In ad-dition, some enteric bacteria (e.g. E. coli, Proteus) and corynebacteria, whichare probably contaminants from the skin, vulva or rectum, may occasionally befound at the anterior urethra.
The vagina becomes colonized soon after birth with corynebacteria, staphylo-cocci, streptococci, E. coli, and a lactic acid bacterium historically named "Do-derlein's bacillus" (Lactobacillus acidophilus). During reproductive life, from
106

puberty to menopause, the vaginal epithelium contains glycogen due to the ac-tions of circulating estrogens. Doderlein's bacillus predominates, being able tometabolize the glycogen to lactic acid. The lactic acid and other products ofmetabolism inhibit colonization by all except this lactobacillus and a select num-ber of lactic acid bacteria. The resulting low pH of the vaginal epithelium pre-vents establishment by most other bacteria as well as the potentially-pathogenicyeast, Candida albicans. This is a striking example of the protective e�ect ofthe normal bacterial �ora for their human host.
15.3.5 Normal Flora of the Oral Cavity
The presence of nutrients, epithelial debris, and secretions makes the mouth afavorable habitat for a great variety of bacteria. Oral bacteria include strep-tococci, lactobacilli, staphylococci and corynebacteria, with a great number ofanaerobes, especially bacteroides.
The mouth presents a succession of di�erent ecological situations with age, andthis corresponds with changes in the composition of the normal �ora. At birth,the oral cavity is composed solely of the soft tissues of the lips, cheeks, tongueand palate, which are kept moist by the secretions of the salivary glands. Atbirth the oral cavity is sterile but rapidly becomes colonized from the environ-ment, particularly from the mother in the �rst feeding. Streptococcus salivariusis dominant and may make up 98% of the total oral �ora until the appearanceof the teeth (6 - 9 months in humans). The eruption of the teeth during the�rst year leads to colonization by S. mutans and S. sanguis. These bacteria re-quire a nondesquamating (nonepithelial) surface in order to colonize. They willpersist as long as teeth remain. Other strains of streptococci adhere strongly tothe gums and cheeks but not to the teeth. The creation of the gingival crevicearea (supporting structures of the teeth) increases the habitat for the variety ofanaerobic species found. The complexity of the oral �ora continues to increasewith time, and bacteroides and spirochetes colonize around puberty.
The normal bacterial �ora of the oral cavity clearly bene�t from their host whoprovides nutrients and habitat. There may be bene�ts, as well, to the host. Thenormal �ora occupy available colonization sites which makes it more di�cult forother microorganisms (nonindigenous species) to become established. Also, theoral �ora contribute to host nutrition through the synthesis of vitamins, andthey contribute to immunity by inducing low levels of circulating and secretoryantibodies that may cross react with pathogens. Finally, the oral bacteria exertmicrobial antagonism against nonindigenous species by production of inhibitorysubstances such as fatty acids, peroxides and bacteriocins.
On the other hand, the oral �ora are the usual cause of various oral diseases inhumans, including abscesses, dental caries, gingivitis, and periodontal disease.If oral bacteria can gain entrance into deeper tissues, they may cause abscessesof alveolar bone, lung, brain, or the extremities. Such infections usually containmixtures of bacteria with Bacteroides melaninogenicus often playing a dominant
107

role. If oral streptococci are introduced into wounds created by dental manip-ulation or treatment, they may adhere to heart valves and initiate subacutebacterial endocarditis.
15.3.6 Normal Flora of the Gastrointestinal Tract
The bacterial �ora of the gastrointestinal (GI) tract of animals has been studiedmore extensively than that of any other site. The composition di�ers betweenvarious animal species, and within an animal species. In humans, there aredi�erences in the composition of the �ora which are in�uenced by age, diet,cultural conditions, and the use of antibiotics. The latter greatly perturbs thecomposition of the intestinal �ora.
In the upper GI tract of adult humans, the esophagus contains only the bac-teria swallowed with saliva and food. Because of the high acidity of thegastric juice, very few bacteria (mainly acid-tolerant lactobacilli) canbe cultured from the normal stomach. However, at least half the popula-tion in the United States is colonized by a pathogenic bacterium, Helicobacterpylori. Since the 1980s, this bacterium has been known to be the cause ofgastric ulcers, and it is probably a cause of gastric and duodenal cancer aswell. The Australian microbiologist, Barry Marshall, received the Nobel Prizein Physiology and Medicine in 2005, for demonstrating the relationship betweenHelicobacter and gastric ulcers.
The proximal small intestine has a relatively sparse Gram-positive �ora, con-sisting mainly of lactobacilli and Enterococcus faecalis. This region has about105 - 107 bacteria per ml of �uid. The distal part of the small intestine con-tains greater numbers of bacteria (108/ml) and additional species, includingcoliforms (E. coli and relatives) and Bacteroides, in addition to lactobacilli andenterococci.
The �ora of the large intestine (colon) is qualitatively similar to that found in fe-ces. Populations of bacteria in the colon reach levels of 1011/ml feces. Coliformsbecome more prominent, and enterococci, clostridia and lactobacilli can be regu-larly found, but the predominant species are anaerobic Bacteroides and anaero-bic lactic acid bacteria in the genus Bi�dobacterium (Bi�dobacterium bi�dum).These organisms may outnumber E. coli by 1,000:1 to 10,000:1. Sometimes,signi�cant numbers of anaerobic methanogens (up to 1010/gm) may reside inthe colon of humans. This is our only direct association with archaea as normal�ora.
At birth the entire intestinal tract is sterile, but bacteria enter with the �rstfeed. The initial colonizing bacteria vary with the food source of the infant.In breast-fed infants, bi�dobacteria account for more than 90% of the total in-testinal bacteria. Enterobacteriaceae and enterococci are regularly present, butin low proportions, while bacteroides, staphylococci, lactobacilli and clostridiaare practically absent. In bottle-fed infants, bi�dobacteria are not predomi-nant. When breast-fed infants are switched to a diet of cow's milk or solid food,
108

bi�dobacteria are progressively joined by enterics, bacteroides, enterococci lac-tobacilli and clostridia. Apparently, human milk contains a growth factor thatenriches for growth of bi�dobacteria, and these bacteria play an important rolein preventing colonization of the infant intestinal tract by non indigenous orpathogenic species.
The composition of the �ora of the gastrointestinal tract varies along the tract(at longitudinal levels) and across the tract (at horizontal levels) where cer-tain bacteria attach to the gastrointestinal epithelium and others occur in thelumen. There is frequently a very close association between speci�c bacteriain the intestinal ecosystem and speci�c gut tissues or cells (evidence of tissuetropism and speci�c adherence). Gram-positive bacteria, such as the strepto-cocci and lactobacilli, are thought to adhere to the gastrointestinal epitheliumusing polysaccharide capsules or cell wall teichoic acids to attach to speci�creceptors on the epithelial cells. Gram-negative bacteria such as the entericsmay attach by means of speci�c �mbriae which bind to glycoproteins on theepithelial cell surface.
It is in the intestinal tract that we see the greatest e�ect of the bacterial �ora ontheir host. This is due to their large mass and numbers. Bacteria in the humanGI tract have been shown to produce vitamins and may otherwise contributeto nutrition and digestion. But their most important e�ects are in their abilityto protect their host from establishment and infection by alien microbes andtheir ability to stimulate the development and the activity of the immunologicaltissues.
On the other hand, some of the bacteria in the colon (e.g. Bacteroides) havebeen shown to produce metabolites that are carcinogenic, and there may be anincreased incidence of colon cancer associated with these bacteria. Alterationsin the GI �ora brought on by poor nutrition or perturbance with antibioticscan cause shifts in populations and colonization by nonresidents that leads togastrointestinal disease.
15.4 Bene�cial e�ects of the normal �ora
The e�ects of the normal �ora are inferred by microbiologists from experimentalcomparisons between "germ-free" animals (which are not colonized by anymicrobes) and conventional animals (which are colonized with a typical normal�ora). Brie�y, some of the characteristics of a germ-free animals that are thoughtto be due to lack of exposure to a normal �ora are:
1. vitamin de�ciencies, especially vitamin K and vitamin B12
2. increased susceptibility to infectious disease
3. poorly developed immune system, especially in the gastrointestinal tract
4. lack of "natural antibody" or natural immunity to bacterial infection
109

Because these conditions in germ-free mice and hamsters do not occur in con-ventional animals, or are alleviated by introduction of a bacterial �ora (at theappropriate time of development), it is tempting to conclude that the humannormal �ora make similar contributions to human nutrition, health and devel-opment. The overall bene�cial e�ects of microbes are summarized below.
1. The normal �ora synthesize and excrete vitamins in excess of theirown needs, which can be absorbed as nutrients by their host. Forexample, in humans, enteric bacteria secrete Vitamin K and Vitamin B12, andlactic acid bacteria produce certain B-vitamins. Germ-free animals may bede�cient in Vitamin K to the extent that it is necessary to supplement theirdiets.
2. The normal �ora prevent colonization by pathogens by competingfor attachment sites or for essential nutrients. This is thought to betheir most important bene�cial e�ect, which has been demonstrated in the oralcavity, the intestine, the skin, and the vaginal epithelium. In some experiments,germ-free animals can be infected by 10 Salmonella bacteria, while the infectiousdose for conventional animals is near 106 cells.
3. The normal �ora may antagonize other bacteria through the pro-duction of substances which inhibit or kill nonindigenous species. Theintestinal bacteria produce a variety of substances ranging from relatively non-speci�c fatty acids and peroxides to highly speci�c bacteriocins, which inhibitor kill other bacteria.
4. The normal �ora stimulate the development of certain tissues,i.e., the caecum and certain lymphatic tissues (Peyer's patches) inthe GI tract. The caecum of germ-free animals is enlarged, thin-walled, and�uid-�lled, compared to that organ in conventional animals. Also, based on theability to undergo immunological stimulation, the intestinal lymphatic tissuesof germ-free animals are poorly-developed compared to conventional animals.
5. The normal �ora stimulate the production of natural antibodies.Since the normal �ora behave as antigens in an animal, they induce an immuno-logical response, in particular, an antibody-mediated immune (AMI) response.Low levels of antibodies produced against components of the normal �ora areknown to cross react with certain related pathogens, and thereby prevent in-fection or invasion. Antibodies produced against antigenic components of thenormal �ora are sometimes referred to as "natural" antibodies, and such anti-bodies are lacking in germ-free animals.
15.5 Harmful e�ects of the normal �ora
Harmful e�ects of the normal �ora, some of which are observed in studies withgerm-free animals, can be put in the following categories. All but the last twoare fairly insigni�cant.
110

1. Bacterial synergism between a member of the normal �ora anda potential pathogen. This means that one organism is helping another togrow or survive. There are examples of a member of the normal �ora supplyinga vitamin or some other growth factor that a pathogen needs in order to grow.This is called cross-feeding between microbes. Another example of synergism oc-curs during treatment of "staph-protected infections" when a penicillin-resistantstaphylococcus that is a component of the normal �ora shares its drug resistancewith pathogens that are otherwise susceptible to the drug.
2. Competition for nutrients. Bacteria in the gastrointestinal tract mustabsorb some of the host's nutrients for their own needs. However, in general,they transform them into other metabolisable compounds, but some nutrient(s)may be lost to the host. Germ-free animals are known to grow more rapidly ande�ciently than conventional animals. One explanation for incorporating antibi-otics into the food of swine, cows and poultry is that the animal grows fasterand can therefore be marketed earlier. Unfortunately, this practice contributesto the development and spread of bacterial antibiotic resistance within the farmanimals, as well as humans.
3. Induction of a low grade toxemia. Minute amounts of bacterial toxins(e.g. endotoxin) may be found in the circulation. Of course, it is these smallamounts of bacterial antigen that stimulate the formation of natural antibodies.
4. The normal �ora may be agents of disease. Members of the normal�ora may cause endogenous disease if they reach a site or tissue where theycannot be restricted or tolerated by the host defenses. Many of the normal �oraare potential pathogens, and if they gain access to a compromised tissue fromwhich they can invade, disease may result.
5. Transfer to susceptible hosts. Some pathogens of humans that aremembers of the normal �ora may also rely on their host for transfer to otherindividuals where they can produce disease. This includes the pathogens thatcolonize the upper respiratory tract such as Neisseria meningitidis, Streptococcuspneumoniae, Haemophilus in�uenzae and Staphylococcus aureus, and potentialpathogens such as E. coli, Salmonella or Clostridium in the gastrointestinaltract.
111

16 Industrial applications
16.1 Food and beverage industries
16.1.1 Alcohol (wines & spirits)
Ethanol results from the fermentation process, because the conversion of sugarto carbon dioxide and water is incomplete. Although, in principle, wine can bemade from almost any fruit juice with a high sugar content, the vast majority ofcommercially produced wines derive from the fermentation of the sugar presentin grape. Such fermentation reactions may be initiated by yeasts naturally foundon the grape skin; however the results of such fermentations are erratic and maybe unpalatable.
In commercial winemaking the must (juice) resulting from the crushed grapesis treated with sulphur dioxide to kill o� the natural micro�ora, and then in-oculated with the yeast Saccharomyces cerevisiae, variety ellipsoideus. Spe-cially developed strains are used, which produce a higher percentage of alcohol(ethanol) than naturally occurring yeasts. Fermentation proceeds for a few daysat a temperature of 22�27 �C for red wines (lower for whites), after which thewine is separated from the skins by pressing. This is followed by ageing in oakbarrels, a process that may last several months, and during which the �avourdevelops.
Malolactic fermentation is a secondary fermentation carried out on certain typesof wine. Malic acid, which has a sharp taste, is converted to the milder lacticacid, imparting smoothness to the wine. A secondary product of malolacticfermentation is diacetyl, which imparts a `buttery' �avour to the wine. Spiritssuch as brandy and rum result from the products of a fermentation processbeing concentrated by distillation. This gives a much higher alcohol contentthan that of wines.
16.1.2 Beer
Beer is produced by the fermentation of barley grain. The procedure varies ac-cording to the type of beer, but follows a series of clearly de�ned steps. Grain,unlike grapes, contains no sugar to serve as a substrate for the yeast, so beforefermentation can begin, it is soaked in water and allowed to germinate. Thisstimulates the production of the enzymes necessary for the conversion of starchto maltose (`malting'). An additional source of starch may be introduced dur-ing the next stage, mashing, in which the grains are ground up in warm water,and further digestion takes place. The liquid phase or wort is drained o� andhops are added. They impart �avour and colour to the �nished product and alsopossess antimicrobial properties, thereby helping to prevent contamination. Themixture is boiled, inactivating the enzymes, precipitating proteins and killing
112

o� any microorganisms. In the next stage, the wort is �ltered and transferredto the fermentation vessel where yeast is introduced.
Two species of yeast are commonly used in the brewing process, both belongingto the genus Saccharomyces. S. cerevisiae is mainly used in the production ofdarker beers such as traditional English ales and stouts, whereas S. carlsbergen-sis (no prizes for guessing where this one was developed!) gives lighter coloured,less cloudy, lager-type beers. Cells of S. cerevisiae are carried to the surface ofthe fermentation by carbon dioxide bubbles (top fermenters), while S. carlsber-gensis cells form a sediment at the bottom (bottom fermenters). `Spent' yeastmay be dried, and used as an animal food supplement.
Fermentation takes about a week to complete, at a temperature appropriate foreach type of yeast (S. carlsbergensis prefers somewhat lower temperatures thanS. cerevisiae). Following fermentation, the beer is allowed to age or `rest' forsome months in the cold. Beers destined for canning or bottling are �ltered toremove remaining microorganisms.
Beers typically have an alcohol content of around 4 per cent. Small amounts ofother secondary products such as amyl alcohol and acetic acid are also produced,and contribute to the beer's �avour. `Light' or low-carbohydrate beers areproduced by reducing the levels of complex carbohydrates. The yeast do notpossess the enzymes necessary to cope with these branched molecules, so asupplement of debranching enzymes may be added to aid their breakdown.
16.1.3 Dairy products
Milk can be fermented to produce a variety of products, including butter, yo-ghurt and cheese. In each case, acid produced by the fermentation processcauses coagulation or curdling of the milk proteins.
In cheese-making, this coagulation is e�ected by the addition of the proteaserennin, or by the action of lactic acid bacteria (especially Streptococcus lactis
and S. cremoris). Coagulation allows the separation of the semisolid curdfrom the liquid whey. The subsequent steps in the cheese-making processdepend on the speci�c type of cheese. Following separation, the curd of mostcheeses is pressed and shaped, removing excess liquid and �rming the texture.During the ripening process, salt is often added, and �avour develops due tocontinuing microbial action on the protein and fat components of the cheese.
In some cases, a fresh inoculation of microorganisms is made at this point, suchas the addition of Penicillium spores to Camembert and Brie. The length ofthe ripening period varies from a month to more than a year according to type,with the harder cheeses requiring the longer periods.
Yoghurt is another milk derivative. Thickened milk is exposed to the action oftwo bacteria, Streptococcus thermophilus and Lactobacillus bulgaricus, both ofwhich ferment lactose present in milk into lactic acid. In addition, L. bulgaricus
113

contributes aromatics responsible for imparting �avour to the yoghurt. Otherdairy products, such as soured cream and buttermilk, are also produced bymeans of the fermentative properties of species of streptococci and lactobacilli.
16.1.4 Bread
The biological agent responsible for bread production is yeast. In fact baker'syeast and brewer's yeast are just di�erent strains of the same species, Saccha-romyces cerevisiae. In breadmaking, aerobic, rather than anaerobic conditionsare favoured, so sugar present in the dough is converted all the way to carbondioxide rather than to alcohol. It is this that causes the bread to rise. Anysmall amount of ethanol that may be produced is evaporated during the bakingprocess.
Many other popular foodstu�s are the result of microbial fermentation processes.These include vinegar, soy sauce and sauerkraut.
Silage is animal fodder made from the fermentation of grass and other plantmaterial by the action of lactic acid bacteria.
16.1.5 Microorganisms as food
Some microorganisms are used as foodstu�s.
Perhaps the most obvious of these are mushrooms, the stalked fruiting bodiesof certain species of basidiomycete, notably Agaricus bisporus. These are grownin the dark at favourable temperatures, in order to stimulate the production offruiting bodies.
Another fungus, Fusarium forms the basis of QuornTM , a processed myco-protein that has been used as a meat substitute for some years in the UK.Whereas mushrooms are grown as agricultural products, QuornTM must beproduced under highly regulated sterile conditions.
Other microbial food sources include certain algae (seaweed), which form animportant part of the diet in some parts of the world, and bacteria and yeastgrown in bulk as single-cell protein (SCP, sold as Pruteen) for use as a protein-rich animal food supplement.
The cyanobacterium Spirulina has been collected from dried-up ponds in partsof central Africa for use as a food supplement since time immemorial and is nowavailable at health stores in the West.
16.1.6 Microbial spoilage of food
Unfortunately for us, fresh foods such as meats, fruit and vegetables providea rich source of nutrients, which a wide range of heterotrophic microorganisms
114

�nd just as attractive as we do. Certain microbial types are associated withparticular foodstu�s, depending on their chemical composition and physicalfactors such as pH and water content. Acidic foods such as fruits, for example,tend to favour the growth of fungi rather than bacteria.
Often, spoilage organisms come from the same source as the food, for examplesoil on vegetables, or meat exposed to intestinal contents following slaughter.Others are introduced as contaminants during transport, storage or preparation.Among the most commonly found spoilage organisms are a number of humanpathogens, including Pseudomonas, Salmonella, Campylobacter and Listeria.Thus, although microbial spoilage may merely lead to foodstu�s being renderedunpalatable, it can also result in serious and even fatal illness (`food poison-ing'). Whilst observable changes to foodstu�s are only likely after the microbialpopulation has reached a considerable size, food poisoning can result from thepresence of much smaller numbers of contaminants.
Some foodstu�s are more susceptible to spoilage than others: fresh items suchas meat, �sh, dairy produce and fruit and vegetables are all highly perishable.Foods such as rice and �our, on the other hand, are much more resistant, becausehaving no water content they do not provide suitable conditions for microbialgrowth. Drying is one of a number of methods of food preservation, all designedto prevent growth of microorganisms by making conditions unfavourable. Othermethods include heating/canning, drying, pickling, smoking and, in many coun-tries, irradiation.
16.2 Microbial production of biochemicals
Many products of microbial metabolism �nd an application in the food andother industries. These include amino acids, steroids, enzymes and antibiotics.Microbial growth conditions are adjusted so that production of the metabolitein question takes place at an optimal rate. Often an unnaturally high rateof production is achieved by the use of a mutated or genetically engineeredstrain of microorganism, or by manipulating culture conditions to favour excessmetabolite production.
16.2.1 Acetone
The development of a microbial means of producing acetone was vital to the al-lied e�ort in the First World War. Acetone was a crucial precursor in explosivesmanufacture and the demands of war soon outstripped supply by traditionalmethods. The problem was solved when Chaim Weismann isolated a strainof Clostridium acetobutylicum that could ferment molasses to acetone and bu-tanol (another industrially useful product). Nowadays, acetone is made morecheaply from petrochemicals.
115

16.2.2 Amino acids (and MSG)
Microbially produced amino acids are used in the food industry, in medicine andas raw materials in the chemical industry. The one produced in the greatestquantities by far is glutamic acid (in excess of half a billion tonnes per year),with most of it ending up as the �avour enhancer monosodium glutamate. Theamino acids aspartic acid and phenylalanine are components of the arti�cialsweetener aspartame and are also synthesised on a large scale.
16.2.3 Citric acid
A number of organic acids are produced industrially by microbial means, mostnotably citric acid, which has a wide range of applications in the food andpharmaceutical industries. This is mostly produced as a secondary metaboliteby the large-scale culture of the mould Aspergillus niger.
16.2.4 Vitamins
Certain microorganisms serve as a ready source of vitamins. In many cases thesecan be synthesised less expensively by chemical means; however, ribo�avin (bythe mould Ashbya gossypii) and vitamin B12 (by the bacteria Propionibac-
terium shermanii and Pseudomonas denitri�cans) are produced by large-scalemicrobial fermentation. Microorganisms play a partial role in the productionof ascorbic acid (vitamin C). Initially, glucose is reduced chemically to sor-bitol, which is then oxidised by a strain of Acetobacter suboxydans to the hexosesorbose. Chemical modi�cations convert this to ascorbic acid.
16.2.5 Industrial enzymes
Enzymes of fungal and bacterial origin have been utilised for many centuries in avariety of processes. It is now possible to isolate and purify the enzymes neededfor a speci�c process and the worldwide market is currently worth around abillion pounds. The most useful industrial enzymes include proteases, amylases,lipases and pectinase. Enzymes have been added to cleaning products such aswashing powders, carpet shampoos and stain removers since the 1960s, and thisremains one of the principal industrial applications of enzymes.
Proteases are the most widely used enzymes in this context; working in com-bination with a surfactant, they hydrolyse protein-based stains such as blood,sweat and various foods. Greasy and oily stains present a di�erent challenge,made all the more di�cult by the move towards lower washing temperatures.The inclusion of lipases aids the removal of stains such as butter, salad dress-ing and lipstick, while amylases deal with starch-based stains such as cereal orcustard.
116

The food and detergent industries between them account for around 80 per centof all enzyme usage.
16.2.6 Syrups
Syrups and modi�ed starches are used in a wide range of foodstu�s, includingsoft drinks, confectionery and ice cream, as well as having a wealth of other ap-plications. Di�erent enzymes or combinations of enzymes are used to producethe desired consistencies and physical properties. High fructose corn syrup(HFCS) is a sweetener used in a multitude of food products. It is some 75 percent sweeter than sucrose and has several other advantages. HFCS is a mixtureof fructose, dextrose (a form of glucose) and disaccharides, and is produced bythe action of a series of three enzymes on the starch (amylose and amylopectin)of corn (maize). Alpha amylase hydrolyses the internal α-1, 4-glycosidic bondsof starch, but is not able to degrade ends of the chain. The resulting di- andoligosaccharides are broken down to the monomer glucose by the action of glu-coamylase, then �nally glucose isomerase converts some of the glucose to itsisomer, fructose.
16.3 Genetically modi�ed organisms
Recombinant DNA technology can be used to genetically modify microorgan-isms so that they produce commercially important proteins such as humaninsulin. This is done by incorporating the gene for the desired protein into anappropriate cloning vector, and inserting it into a host cell such as E. coli orSaccharomyces cerevisiae.
The initial application of this technology was in the microbial production ofmedically important proteins such as insulin and epidermal growth factor,however other proteins may also be produced by these means. These includeenzymes used in diagnostic and analytical applications, where a higher purityof preparation is required than, for example, the enzymes used in detergents.These are often derived originally from other microorganisms; for example thethermostable DNA polymerase from Thermus aquaticus used in PCR (poly-merase chain reaction) is now commonly made by recombinant E. coli cells thathave been transformed with the T. aquaticus gene. Many of the more recentrecombinant human proteins to be developed for therapeutic use have been toocomplex for expression in a microbial system (e.g. Factor VIII), so it has beennecessary to employ cultured mammalian cells.
16.4 Microbial mining
An unexpected application for microorganisms is to be found in the mining in-dustry. Acidophilic bacteria including Acidithiobacillus ferrooxidans are increas-ingly being used to extract valuable metals, notably copper, from low-grade ores
117

that would not be worth working by conventional technologies. A. ferrooxidansis the organism largely responsible for the phenomenon of acid mine drainage;by carrying out the same reactions in a di�erent context, however, it can beput to a bene�cial use. Tailings (mineral waste with a low metal content) aregathered in huge tips and acidi�ed water sprinkled over them, stimulating thegrowth of indigenous bacteria.
Bacterial oxidation results in soluble copper sulphate leaching from the tip andbeing collected for copper extraction. This bacterial action is known as directbioleaching, but if you follow the process in, you will see that A. ferrooxidanshas not �nished yet! This remarkable organism can also oxidise iron from itsferrous to ferric form; the resulting ferric sulphate is a potent oxidising agent,which, when recycled to the tip, carries out indirect (chemical) bioleaching,and so the cycle continues. A. ferrooxidans has a number of other unusualfeatures, which enable it to survive in this hostile environment; it thrives inacidic conditions (pH < 2.0), and has an unusually high tolerance of metal ionssuch as copper.
Bacteria are also involved in the extraction of other metals such as uraniumand gold; the methodologies di�er slightly, but still involve the conversion of aninsoluble compound to a soluble one. It is only in the last 20 years or so that theeconomic possibilities of biohydrometallurgy have been realised, and now asigni�cant proportion of the world's copper and other metals is produced in thisway. The method is inexpensive but rather slow; it may take years to extractthe copper from a large tip. However as high-grade copperbearing ores becomeincreasingly scarce, bioleaching seems likely to play an increasingly importantrole.
Sulphur-oxidising bacteria also have a role to play in the coal mining indus-try. Increased environmental awareness in many countries means that it is nolonger acceptable to burn o� the sulphur content of coal as sulphur dioxide,so an alternative must be found. One possibility is the biodesulphurisation ofcoal, using sulphur-oxidising bacteria to remove the sulphur before combustion.Whilst technically feasible, economic considerations mean that this has not yetbeen widely adopted.
118

17 Lab techniques
17.1 Units
In science, the metric system is standard. The metric system is part of thethe Système International d'Unités (SI system).
The trick to learning the metric system is to memorize the meaning of six pre-�xes. In the metric system, each pre�x means that a meter is either multipliedor divided by a multiple of 10.
Pre�x Value in metres
Kilo (km) (kilo = 1,000) 1000m or 1x10−3mDeci (dm) (deci = 1/10) 0.1m or 1x10−1m
Centi (cm) (centi = 1/100) 0.01m or 1x10−2mMilli (mm) (milli = 1/1000) 0.001m or 1x10−3m
Nano (nm) (nano = 1/1,000,000,000) 0.000000001m or 1x10−9mPico (pm) (pico = 1/1,000,000,000,000) 0.000000000001m or 1x10−12m
You should also see section 15.4.2 about Colony Forming Units (CFU).
17.2 Safety
17.2.1 Aseptic technique
Aseptic technique is the name given to the procedures used by microbiologiststo prevent microbial contamination of themselves, which may result in infection,contamination of the environment they are working in, and contamination of thespecimen they are working on, which is especially important when a pure cultureis desired. It is used whenever specimens are to be transferred between media,for example, when subculturing. Such a procedure, using a �ame sterilizationmethod, might occur as follows
1. A person would assemble the closed tube or �ask from which�and theclosed tube or �ask to which�the specimen is to be transferred, an in-oculating loop, and a �re source, all on a clean, preferably microbe-freesurface with some overhead protection from airborne microbes.
2. The person would start the �re, and move the end of the inoculating loop,in a slow back-and-forth motion, through the top of the blue part of the�ame. The person would not allow the loop to touch anything except thespecimen itself, until the entire procedure is �nished.
3. Preparing to execute the specimen transfer, the person would hold both ofthe tubes or �asks in one hand, typically the opposite of the writing hand.The person would then open the tube or �ask containing the specimensource and brie�y hold the top of it in the �ame, to kill unwanted microbes.
119

4. Quickly, so as to minimize the possible time for contamination of thespecimen in the source tube or �ask, the person would use the inoculatingloop with their writing hand to retrieve the specimen, and then sterilizethe top of the tube or �ask again before immediately closing it.
5. Keeping in mind that the specimen on the inoculating loop could be con-taminated during every moment it is exposed, the person would repeatthe previous step identically with the tube or �ask in which the specimenis to be deposited; however, the person would be depositing the sampleinto the tube or �ask.
17.2.2 Microbiological safety cabinets
The microbiological safety cabinet (MSC) is a basic tool in the laboratory forthose who work with biological agents, including pathogenic micro-organisms(including genetically modi�ed micro-organisms), cell cultures and human andanimal tissues and �uids.
� Class I cabinet. An open fronted cabinet designed to protect the oper-ator by continuously drawing air into the front of the cabinet.
This is the traditional cabinet of microbiology and clinical diagnostic lab-oratories. Its construction is simpler than the other types and is thereforecheaper to produce and buy. It works by drawing air into the cabinetaway from the worker and then through a High E�ciency Particulate Air[HEPA] �lter. It is the cabinet of choice when the key requirement is theprotection of the worker.
� Class II cabinet. An open fronted cabinet designed to protect the oper-ator from exposure and the work from external contamination. Inward airis directed downwards into a plenum below the work surface and is �lteredbefore being redirected into the work area as a laminar down �ow of cleanair. The balance of this laminar down �ow with the incoming air providesan air curtain at the open front which provides the operator protection.The split between exhausted and recycled air on each cycle is normally inthe range 20/80 to 30/70.
These cabinets are designed so that the work area is kept clean by adown-�ow of HEPA-�ltered air across the work. With good working tech-nique it is possible to achieve both a high degree of product protectionand a operator protection of the same order as that provided by Class Icabinets. However, their performance is more a�ected by other factors,such as operator movement and air movements outside the cabinet. Theyare typically used for cell and tissue culture applications and for otherwork where product protection from contamination is important.
120

� Class III cabinet. A totally enclosed cabinet in which operations areconducted through gloves attached to glove ports. Air enters the cabinetthrough a HEPA �lter at the side or rear of the cabinet and is exhaustedin a similar way to a class I cabinet.
Class III cabinets are designed to completely contain the hazardous agentand are used mainly for work with hazard group 4 organisms or work withhazard group 3 organisms deemed to be at high risk, for example wherehighly concentrated samples are being handled. They o�er the greatestprotection to the worker and work, but movements are more restrictedand this may a�ect dexterity.
Laminar �ow hoods should not be confused with MSCs. These hoods provide a�ltered air-�ow which is intended for product protection only. The cabinetsmust not be confused with Class II cabinets as they have no in�ow, indeedthe air passes over the product being worked on [e.g. cell culture] and so theyo�er no operator protection. Therefore, laminar �ow hoods must not be usedwith any biological material of human origin. Other animal-derived materialor tissue may only be used after a full risk assessment that con�rms the totalnon-pathogenic and non-allergenic status of the material. They also must notbe used when handling biological agents [pathogenic organisms].
17.3 Pure culture
17.3.1 Streak plate
The standard method of obtaining a pure bacterial culture is the creation of astreak plate. A wire inoculating loop is used to spread out a drop of bacterialsuspension on an agar plate in such a way that it becomes progressively moredilute; eventually, individual cells will be deposited on the agar surface.
Following incubation at an appropriate temperature, a succession of cell divi-sions occurs, resulting in the formation of a bacterial colony, visible to the nakedeye. Colonies arise because movement is not possible on the solid surface andall the progeny stay in the same place.
A colony represents, in theory at least, the o�spring of a single cell and itsmembers are therefore genetically identical. (In reality, a clump of cells may bedeposited together and give rise to a colony; this problem can be overcome byrepeated isolation and restreaking of single colonies.)
17.3.2 Pour plate
An alternative method for the isolation of pure cultures is the pour plate. Inthis method, a dilute suspension of bacteria is mixed with warm molten agar,
121

Figure 6: The streak plate. Streaking the sample across the agar surface even-tually results in individual cells being deposited. Repeated cycles of cell divisionlead to the production of visible, isolated colonies.
and poured into an empty petri plate.
As the agar sets, cells are immobilised, and once again their progeny are allkept together, often within, as well as on, the agar. This method is especiallyuseful for the isolation of bacteria that are unable to tolerate atmospheric levelsof oxygen.
17.4 Preparing specimens
There are two ways to prepare a specimen to be observed under a light compoundmicroscope. These are a smear and a wet mount.
17.4.1 Smear
A smear is a preparation process where a specimen is spread on a slide. Youprepare a smear using the heat �xation process:
1. Use a clean glass slide.
2. Take a loop of the culture.
3. Place the live microorganism on the glass slide.
122

4. The slide is air dried then passed over a Bunsen burner about three times.
5. The heat causes the microorganism to adhere to the glass slide. This isknown as �xing the microorganism to the glass slide.
6. Stain the microorganism with an appropriate stain.
17.4.2 Wet Mount
A wet mount is a preparation process where a live specimen in culture �uid isplaced on a concave glass side or a plain glass slide. The concave portion of theglass slide forms a cup-like shape that is �lled with a thick, syrupy substance,such as carboxymethyl cellulose. The microorganism is free to move aboutwithin the �uid, although the viscosity of the substance slows its movement.This makes it easier for you to observe the microorganism. The specimen andthe substance are protected from spillage and outside contaminates by a glasscover that is placed over the concave portion of the slide.
17.5 Counting Methods
Several methods exist for the measurement of bacterial numbers, most of whichare also applicable to the enumeration of other unicellular forms such as yeasts.Such methods fall into two main categories: those that count total cell num-bers, and those that count viable cells only.
17.5.1 Total cell count - haemocytometer
Total cell counts are generally done by direct microscopic examination. A spe-cialised glass slide called a haemocytometer is employed, which carries anetched grid of known area. The depth of the liquid sample is also known, so bycounting the number of cells visible in the �eld of view, the number of cells perunit volume can be determined. The method may be made more accurate bythe use of a �uorescent dye such as acridine orange, which binds to DNA, andhence avoids confusion with non-cellular debris. However, such methods cannotdi�erentiate between living and non-living cells. Their usefulness is further lim-ited by the fact that the smallest bacteria are di�cult to resolve as individualcells by light microscopy.
17.5.2 Colony forming units
Colony-forming unit (CFU) is a measure of viable bacterial or fungal numbers.Unlike in direct microscopic counts where all cells, dead and living, are counted,CFU measures viable cells.
123

Figure 7: Serial dilution
The theory behind the technique of CFU establish that a single bacterium cangrow and become a colony, via binary �ssion. These colonies are clearly dif-ferent between each other, both microscopical and macroscopical. However,some bacteria do not separate completely during the sample preparation pro-cess (Staphylococcus, Streptococcus) and the results of the count will be belowthe number of individual cells using direct methods.
17.5.3 Serial dilution - estimating viable cell numbers
In order that the sample to be plated out contains an appropriate number ofcells, the original sample is subjected to serial dilution. In the example below,it is diluted by a factor of ten at each stage to give a �nal dilution of 10-5 (onein a hundred thousand). Samples of 0.1 ml of each dilution are plated out on asuitable solid medium and colonies allowed to develop.
TNTC = too numerous to count. In the earlier tubes, the suspension of cellsis too concentrated, resulting in too many colonies to count. In the �nal tube,the suspension is so dilute that there are no cells in the sample taken.
The 10=3 dilution is used to calculate the concentration of cells in the origi-
124

nal culture, as it falls within the range of 30=300 colonies regarded as beingstatistically reliable.
Colony count (10=3 dilution) = 65
∴ Number of cfu (colony forming units) per ml diluted suspension = 65 Ö 10 =650
∴ Number of cfu per ml original susp'n = 650 Ö 103= 6.5 Ö 105
17.5.4 Spread plate (Miles & Misra)
The Miles & Misra technique is a viable cell enumeration technique whichfollows the preparation of serial dilutions.
� Materials required:
1. A calibrated dropping pipette, delivering drops of about 0.02ml
2. 6 plates of clear nutrient agar
� The plates are divided into numbered sectors.
� The inoculum / suspension is deposited as drops of 0.02ml from a heightof 2.5cm on to the medium where it spreads over an area of 1.5 � 2.0cmdiameter.
� Each of the 6 plates receives one drop of each dilution in separate num-bered sectors.
� The plates are incubated for 18 � 24 hours and observed for growth.
� Sectors where more than 20 colonies are present without any con�uenceare utilized to make the viable counts.
� Viable count per 0.02ml for a dilution is obtained by taking the averageof counts for that dilution in all the six plates.
17.5.5 Pour plate
A pour plate is another alternative for enumerating viable cells, and follows aserial dilution.
0.1ml of each dilution is placed in a separate Petri dish. Liquid agar is thenadded to cover the base of the dish, and the mixture swirled gently to mix.The plates are allowed to set under a laminar �ow hood, then inverted andincubated. After a few days, colonies may be counted to give concentration incfu/mL.
125

17.5.6 Turbidimetric
Sometimes it is desirable to have an estimate of bacterial numbers immediately.A useful method for doing this is based on how cloudy or turbid the liquid growthmedium becomes due to bacterial growth. Turbidimetric methods measure thechange in optical density or absorbance of the medium, that is, how much abeam of light is scattered by the suspended particulate matter. They can becarried out very quickly by placing a sample in a spectrophotometer.
Values of optical density can be directly related to bacterial numbers or mass byreference to a standard calibration curve. Thus, an estimate of bacterial num-bers, albeit a fairly approximate one, can be obtained almost instantaneouslyduring an experimental procedure. Other indirect methods of measuring celldensity include wet and dry weight estimations, and the measurement of cellcomponents such as total nitrogen, protein or nucleic acid.
17.6 Staining methods
Not all specimens can be clearly seen under a microscope. Sometimes the spec-imen blends with other objects in the background because they absorb andre�ect approximately the same light waves. You can enhance the appearance ofa specimen by using a stain. A stain is used to contrast the specimen from thebackground.
A stain is a chemical that adheres to structures of the microorganism and ine�ect dyes the microorganism so the microorganism can be easily seen under amicroscope.
Stains used in microbiology are either basic or acidic.
Basic stains are cationic and have positive charge. Common basic stains aremethylene blue, crystal violet, safranin, and malachite green.These are ideal for staining chromosomes and the cell membranes of manybacteria.
Acid stains are anionic and have a negative charge. Common acidic stainsare eosin and picric acid. Acidic stains are used to stain cytoplasmicmaterial and organelles or inclusions.
17.6.1 Types of stains
There are two types of Stains: simple and di�erential.
Simple stain: A simple stain has a single basic dye that is used to show shapesof cells and the structures within a cell. Methylene blue, safranin, car-bolfuchsin and crystal violet are common simple stains that are found inmost microbiology laboratories.
126

Di�erential stain: A di�erential stain consists of two or more dyes and is usedin the procedure to identify bacteria. Two of the most commonly useddi�erential stains are the Gram stain and the Ziehl-Nielsen acid-faststain.
In 1884 Hans Christian Gram, a Danish physician, developed the Gram stain.Gram-stain is a method for the di�erential staining of bacteria. Gram-positivemicroorganisms stain purple. Gram-negative microorganisms stainpink. Staphylococcus aureus, a common bacterium that causes food poisoning,is Gram-positive. Escherichia coli is gram-negative.
The Ziehl-Nielsen acid-fast stain, developed by Franz Ziehl and Friedrick Nielsen,is a red dye that attaches to the waxy material in the cell walls of bacteria suchas Mycobacterium tuberculosis, which is the bacterium that causes tuberculosis,andMycobacterium leprae, which is the bacterium that causes leprosy. Microor-ganisms that retain the Ziehl-Nielsen acid-fast stain are called acid-fast. Thosethat do not retain it turn blue because the microorganism doesn't absorb theZiehl-Nielsen acid-fast stain.
17.6.2 Here's how to Gram-stain a specimen:
1. Prepare the specimen using the heat �xation process (see �Smear� earlierin this chapter).
2. Place a drop of crystal violet stain on the specimen.
3. Apply iodine on the specimen using an eyedropper. The iodine helps thecrystal violet stain adhere to the specimen. Iodine is a mordant, which isa chemical that �xes the stain to the specimen.
4. Wash the specimen with ethanol or an alcohol-acetone solution, then washwith water.
5. Wash the specimen to remove excess iodine. The specimen appears purplein color.
6. Wash the specimen with an ethanol or alcohol-acetone decolourising solu-tion.
7. Wash the specimen with water to remove the dye.
8. Apply the safranin stain to the specimen using an eyedropper.
9. Wash the specimen.
10. Use a paper towel and blot the specimen until the specimen is dry.
11. The specimen is ready to be viewed under the microscope. Gram-positivebacteria appear purple and gram-negative bacteria appear pink.
127

17.6.3 Here's how to apply the Ziehl-Nielsen acid-fast stain to aspecimen:
1. Prepared the specimen (see �Smear� earlier in this chapter).
2. Apply the red dye carbol-fuchsin stain generously using an eyedropper.
3. Let the specimen sit for a few minutes.
4. Warm the specimen over steaming water. The heat will cause the stain topenetrate the cell wall.
5. Wash the specimen with an alcohol-acetone decolorizing solution consist-ing of 3 percent hydrochloric acid and 95 percent ethanol. The hydrochlo-ric acid will remove the colour from non�acid-fast cells and the back-ground. Acid-fast cells will stay red because the acid cannot penetratethe cell wall.
6. Apply methylene blue stain on the specimen using an eyedropper.
17.6.4 Special stains
Special stains are paired to dye speci�c structures of microorganisms such asendospores, �agella, and gelatinous capsules. One stain in the pair is usedas a negative stain. A negative stain is used to stain the background of themicroorganism, causing the microorganism to appear clear. A second stain isused to colorize speci�c structures within the microorganism. For example,nigrosin and India ink are used as a negative stain and methylene blue is usedas a positive stain.
17.6.5 The endospore stain
The Schae�er-Fulton endospore stain is a special �agellar stain that is used tocolourise the endospore.
1. Prepare the specimen (see �Smear� earlier in this chapter).
2. Heat the malachite green stain over a Bunsen burner until it becomes�uid.
3. Apply the malachite green to the specimen using an eyedropper.
4. Wash the specimen for 30 seconds.
5. Apply the safranin stain using an eyedropper to the specimen to stainparts of the cell other than the endospore.
6. Observe the specimen under the microscope. Endospores will appeargreen.
128





























