Embed Size (px)
Citation preview

119
第十四届中国古脊椎动物学学术年会论文集. 董为主编. 北京:海洋出版社, 2014. 119-134
Proceedings of the Fourteenth Annual Meeting of the Chinese Society of Vertebrate Paleontology
DONG Wei (ed). Beijing: China Ocean Press, 2014. 119-134
中国南方中-晚更新世的马来熊化石
江左其杲 1 丛浩亮 1
马 睿 1 冯 贺 1
刘金毅 2
(1 中国地质大学地学院,北京 100083;2 中国科学院古脊椎动物与古人了类研究所,北京 100044)
摘 要 重新研究了中国南方更新世的熊属 Ursus 化石,利用传统的形态比较和多变量统
计的分析方法,在广西中-晚更新世诸洞穴和云南富民河上洞的熊类化石中辨认出马来熊标
本,首次明确肯定了马来熊在这些地区的存在。马来熊早在中更新世就已出现于我国的南
方地区,如广西和云南。中-晚更新世的化石马来熊稍大于现生种,M1 的中附尖常与后小
尖紧密相连,M2 的跟座稍长于现生种,m1 的前下后尖较高,与现生种有一些区别,但这
些仍然构不成种间的差别,可以作为马来熊的一个化石亚种,即 Ursus malayanus
praemalayanus。
关键词 广西;云南;中-晚更新世;马来熊
1 前言
马来熊 Ursus (Helarctos) malayanus 为现生最小的熊,其形态也十分特殊。对于它
在熊科中的系统发育位置也有多种不同的观点 [1]。作为化石,在世界上它的发现并不
多见。在中国,马来熊化石尤其少见,有关的发现研究存在不少问题:要么是无确切
的产地信息,要么是标本的分类鉴定存有疑问。
在我国以往发现的熊类化石中,与马来熊有关的记述仅有以下几处:Zdansky 曾
经将山西晚上新世的一具带 m1 的残破下颌鉴定为 Ursus cf. boeckhi [2]。而 Thenius 则
认为它代表了一类原始的马来熊,命名为 Ursus sinomalayanus。Erdbrink [3]及邱占祥
[4]
对此也表达了相同的观点。Von Koenigswald 曾经在中药铺购得若干马来熊的牙齿,据
此研究命名为 Ursus praemalayanus,不过 Erdbrink 则认为它们就是现生的马来熊
(Ursus malayanus),可惜这批标本没有确切的产地信息 [4]。裴文中
[5]在研究广西中-
晚更新世诸洞穴的熊类标本时,发现若干跟座较短的 M2,个体较小,暂订为 Ursus sp.,
并且认为可能属于马来熊。同时他也发现若干形态较为奇特的 m1,与一般黑熊(Ursus
thibetanus)有所不同,但没有做进一步探讨。
武竹仙等描述了湖北郧西地点晚更新世的熊科化石,将一些尺寸较小的牙齿归入
马来熊 [6],但事实上这些牙齿在形态上更接近于黑熊。
基金项目:国家自然科学基金项目(批准号:40972013)和中国地质大学(北京)大学生科技立项及创新型实验项目
(批准号:201311415016).
刘金毅:男,49 岁,研究员,从事第四纪哺乳动物学的研究. [email protected]

120
由此看来,到目前为止在我国境内尚无确定的有明确产地记录的马来熊化石。在
地质历史时期,我国境内马来熊是否的确存在过?它又是最早何时在我国何地出现
的?它的地史地理分布究竟如何?这些问题显然值得做深入的研究和厘清。本研究通
过对我国南方更新世熊属化石的再研究,在广西中-晚更新世诸洞穴以及云南中更新
世富民河上洞的熊类标本中辨认出马来熊化石,确认了马来熊在我国更新世时期的存
在与可能的地理分布范围。拟从牙齿的形态和尺寸大小两个方面来进行讨论分析。
2 材料与方法
2.1 材料
本项研究的标本收藏于中国科学院古脊椎动物与古人类研究所标本馆,主要为更
新世中国南方的熊属 Ursus 牙齿化石(详见表 1,黑熊类的标本主要用于对比),其中
可资鉴定为马来熊的牙齿数目很少,只有 8 个 M1(RV87029.1~8),5 个 M2(V5071,
V5076~5079),3 个 m1(RV87029.9~11),8 个 m2(RV87029.12~19)。上述这些标本
产自中-晚更新世的广西诸洞穴,它们早先由裴文中记述和研究过,或鉴定为 Ursus
thibetanus,或为 Ursus sp. [5]。另外还有两件标本来自云南富民河上洞,即 M1
(RV38058.1~2,原先曾被鉴定为 Ursus angustidens[7]),。马来熊标本稀少,可能和马
来熊不冬眠,很少进入洞穴有关[8];另一方面也可能和收集化石有关:由于马来熊牙
齿较小,且它的 P4 原尖常常不发育,形态与其他熊科差异较大,极有可能未将这些
标本归入到熊类中。
为了对比研究,作者还详细测量和研究了现收藏于中国科学院动物研究所标本馆
的现生马来熊头骨标本 6 件(H721,H1039,H1136,H1138,H1143 和未标号的标本
1 件)。
表 1 本研究用的熊类标本及产地
Table 1 Ursid specimens for study and comparison
产地
Locality
时代
Age
原定分类
previous taxa
本文修改分类
emended taxa
标本数目
Number of the specimens
广西柳城
巨猿洞
早更新世
Ursus aff. thibetanus Ursus thibetanus primitinus M1:28; M2:29;
m1:12; m2:22
云南富民河
上洞
中更新世 Ursus angustidens Ursus thibetanus kokeni
Ursus malayanus
praemalayanus
M1:18; M2:18;
m1:9; m2: 11
广西其他洞穴 中晚更新世
Ursus thibetanus
Ursus sp.
Ursus thibetanus kokeni
Ursus malayanus
praemalayanus
M1:52; M2:59;
m1:16; m2:35
2.2 方法
本研究拟采用传统的形态比较和多变量统计的方法对广西其他山洞和云南富民
河上洞的疑似马来熊标本进行深入研究和分析,以确定它们的分类属性。对拟研究的
121
牙齿做了多项测量(见下文),并对所测量的数据做主成分分析(Principal Component
Analysis,PCA)。
主成分分析(PCA)是古生物学上常用的数理统计方法,其旨在通过对原始数据
进行分析,提取出主成分(Principal Component,PC),使主成分的方差达到最大,不
同主成分之间互相垂直,每个主成分都是原先几个初始变量的线性组合,第一主成分
(PC1)值的方差最大,代表最大的差异,故而代表最重要的几何特征(即数据分离
最明显的项),第二主成分(PC2)代表第二重要的特征,而且其与第一主成分没有重
复,以此类推。据此制作第一(PC1)和第二主成分(PC2)的散点图,分析和比较它
们在图区的相对位置,分析研究不同群体间的相对差异。本研究使用 SPSS 20.0
(IBM SPSS Statistics)软件进行统计分析。
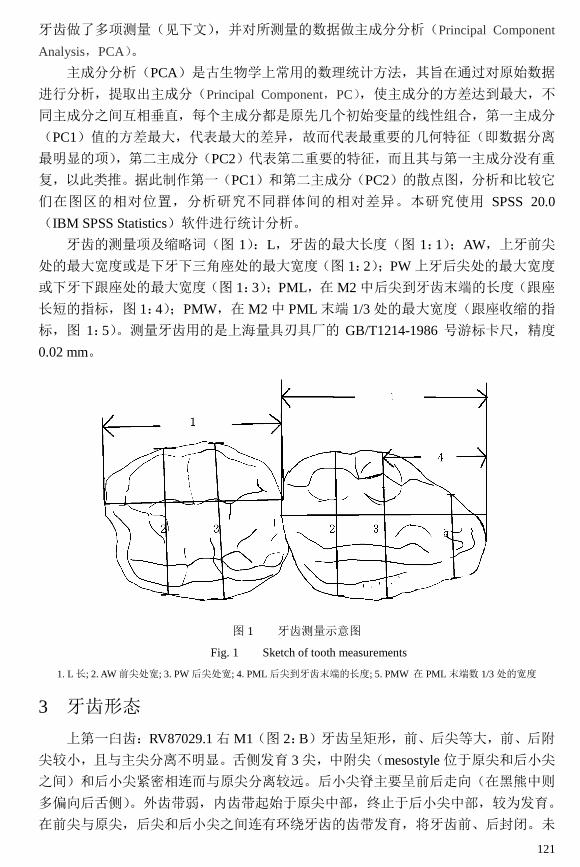
牙齿的测量项及缩略词(图 1):L,牙齿的最大长度(图 1:1);AW,上牙前尖
处的最大宽度或是下牙下三角座处的最大宽度(图 1:2);PW 上牙后尖处的最大宽度
或下牙下跟座处的最大宽度(图 1:3);PML,在 M2 中后尖到牙齿末端的长度(跟座
长短的指标,图 1:4);PMW,在 M2 中 PML 末端 1/3 处的最大宽度(跟座收缩的指
标,图 1:5)。测量牙齿用的是上海量具刃具厂的 GB/T1214-1986 号游标卡尺,精度
0.02 mm。
图 1 牙齿测量示意图
Fig. 1 Sketch of tooth measurements
1. L 长; 2. AW 前尖处宽; 3. PW 后尖处宽; 4. PML 后尖到牙齿末端的长度; 5. PMW 在 PML 末端数 1/3 处的宽度
3 牙齿形态
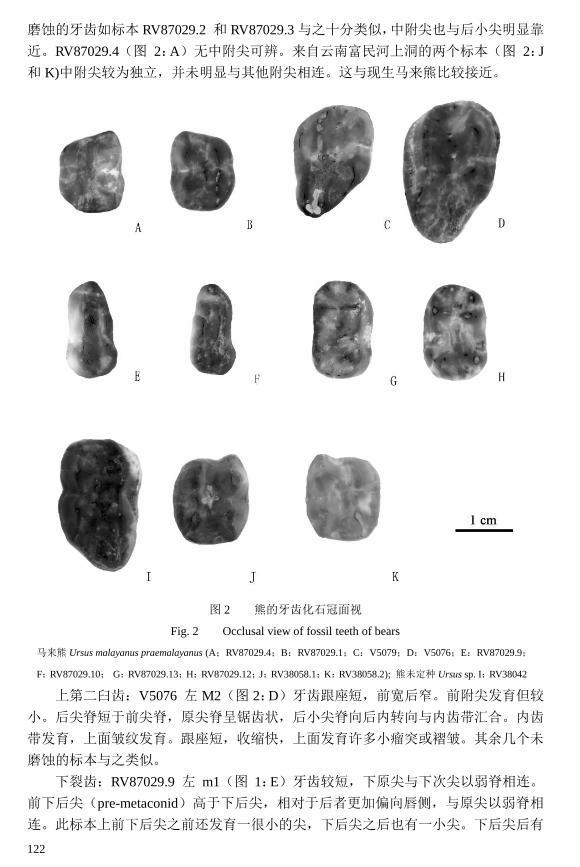
上第一臼齿:RV87029.1 右 M1(图 2:B)牙齿呈矩形,前、后尖等大,前、后附
尖较小,且与主尖分离不明显。舌侧发育 3 尖,中附尖(mesostyle 位于原尖和后小尖
之间)和后小尖紧密相连而与原尖分离较远。后小尖脊主要呈前后走向(在黑熊中则
多偏向后舌侧)。外齿带弱,内齿带起始于原尖中部,终止于后小尖中部,较为发育。
在前尖与原尖,后尖和后小尖之间连有环绕牙齿的齿带发育,将牙齿前、后封闭。未
122
磨蚀的牙齿如标本 RV87029.2 和 RV87029.3 与之十分类似,中附尖也与后小尖明显靠
近。RV87029.4(图 2:A)无中附尖可辨。来自云南富民河上洞的两个标本(图 2:J
和 K)中附尖较为独立,并未明显与其他附尖相连。这与现生马来熊比较接近。
图 2 熊的牙齿化石冠面视
Fig. 2 Occlusal view of fossil teeth of bears
马来熊 Ursus malayanus praemalayanus (A;RV87029.4;B:RV87029.1;C:V5079;D:V5076;E:RV87029.9;
F:RV87029.10; G:RV87029.13;H:RV87029.12;J:RV38058.1;K:RV38058.2); 熊未定种 Ursus sp. I:RV38042
上第二臼齿:V5076 左 M2(图 2:D)牙齿跟座短,前宽后窄。前附尖发育但较
小。后尖脊短于前尖脊,原尖脊呈锯齿状,后小尖脊向后内转向与内齿带汇合。内齿
带发育,上面皱纹发育。跟座短,收缩快,上面发育许多小瘤突或褶皱。其余几个未
磨蚀的标本与之类似。
下裂齿:RV87029.9 左 m1(图 1:E)牙齿较短,下原尖与下次尖以弱脊相连。
前下后尖(pre-metaconid)高于下后尖,相对于后者更加偏向唇侧,与原尖以弱脊相
连。此标本上前下后尖之前还发育一很小的尖,下后尖之后也有一小尖。下后尖后有

123
一弱脊平行矢状面向后延伸,止于下三角座与下跟座之间的凹谷。下内尖 1 个。下次
尖以脊和下原尖相连,其前内侧有一 V 字型凹槽。其余两个标本磨蚀严重,但可见前
下后尖与 RV87029.9(图 2:F) 一样,高于下后尖,更加靠近唇侧。
下第二臼齿:RV87029.12 左 m2(图 2:H)牙齿短宽,下前尖较发育,位于牙齿
前内侧。下原尖前侧也有 1 个小尖。下后尖与下原尖以横脊相连(下后尖相对于下原
尖靠后)。下跟座的齿尖有一定磨损。其余未磨蚀的标本大多与此标本相类似。
RV87029.13(图 2:G)与之类似,但下后尖与下原尖间的脊是垂直于长轴的。
小结: 上述广西其他山洞和云南富民河上洞的牙齿标本,在形态上显然与现生的
马来熊十分接近:牙齿短而宽,齿尖发育但并不复杂。其中 m1 和部分 m2 具有最明
显的马来熊牙齿特征。M1,M2,m2 也是相对更加接近马来熊而非同区分布的柯氏黑
熊(Ursus thibetanus kokeni)。
4 牙齿测量与分析
上第一臼齿:对于 M1,本研究共计测量了 98 件标本三组测量数据(表 2),并对
此做了主成分分析,得到第一、第二主成分的散点图(图 3)。从统计分析的结果(表 2)
可以看出,第一主成分解释全部变量的 73%,和各项测量项都成正相关(见表 2,PC1
的值为正),且和宽度(AW 和 PW)的关系更大些。第二主成分则解释了变量的 21%,
与长度(L)呈正相关(见表 2,PC2 的值为正),而和牙齿宽度呈负相关(见表 2,
AW 和 PW 的 PC2 值为负),其中和牙齿前尖处的宽(AW)呈更加明显的负相关(见
表 2,AW 的 PC2 值= - 0.504,小于 PW 对应的值)。这样一来比较长而窄的牙齿就会
在 PC2 上的分值会较大。
表 2 M1 主成分分析主成分得分系数及比重。
Table 2 Component loadings of Principal Component Analysis of M1.
主成分
项目 1 2 3
成分得分系数矩阵
长 L 0.335 1.101 0.141
前尖处宽 AW 0.412 -0.504 1.540
后尖处宽 PW 0.420 -0.382 -1.624
单一主成分解释的比重/% 72.705 20.670 6.625
累计主成分解释的比重/% 72.705 93.375 100.000
从图 3 可以看出,现生马来熊位于图区的左下方,体现了其个体较小而宽度较大
的特点。中-晚更新世广西诸洞穴的马来熊在第二主成分上的分值与现生马来熊一致,
但第一主成分较大,代表其个体较大。富民河上洞的疑似马来熊(Ursus malayanus
praemalayanus ?)的标本完全落入马来熊的变异范围内,而和富民河上洞的黑熊标本(U.
t. kokeni Fumin)差距较大(注意图 3 中╳和△的比较),这确认了它们就是马来熊。我
国中-晚更新世的柯氏黑熊(U. t. kokeni)明显处于图区的上部。云南富民河上洞的黑
熊标本(U. t. kokeni Fumin)落入广西中-晚更新世诸洞穴的黑熊标本标本(U. t. kokeni
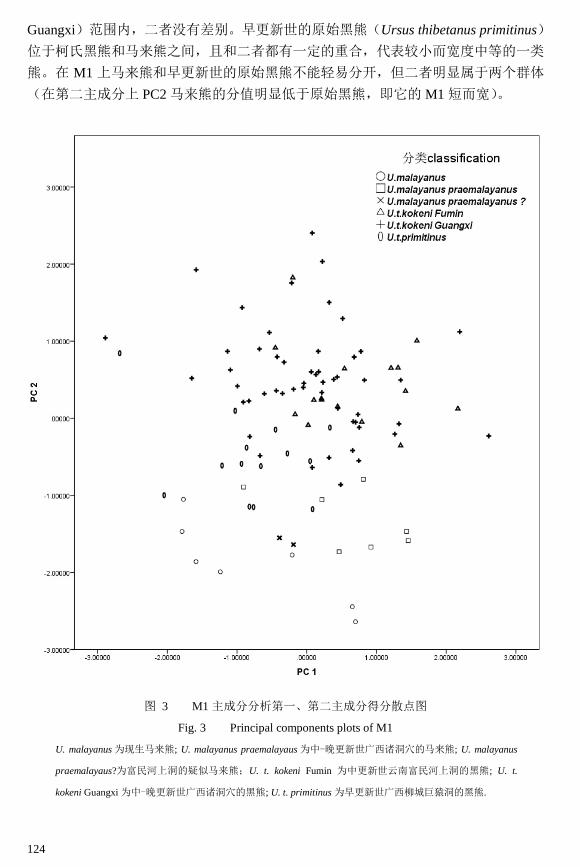
124
Guangxi)范围内,二者没有差别。早更新世的原始黑熊(Ursus thibetanus primitinus)
位于柯氏黑熊和马来熊之间,且和二者都有一定的重合,代表较小而宽度中等的一类
熊。在 M1 上马来熊和早更新世的原始黑熊不能轻易分开,但二者明显属于两个群体
(在第二主成分上 PC2 马来熊的分值明显低于原始黑熊,即它的 M1 短而宽)。
图 3 M1 主成分分析第一、第二主成分得分散点图
Fig. 3 Principal components plots of M1
U. malayanus 为现生马来熊; U. malayanus praemalayaus 为中-晚更新世广西诸洞穴的马来熊; U. malayanus
praemalayaus?为富民河上洞的疑似马来熊;U. t. kokeni Fumin 为中更新世云南富民河上洞的黑熊; U. t.
kokeni Guangxi 为中-晚更新世广西诸洞穴的黑熊; U. t. primitinus 为早更新世广西柳城巨猿洞的黑熊.
125
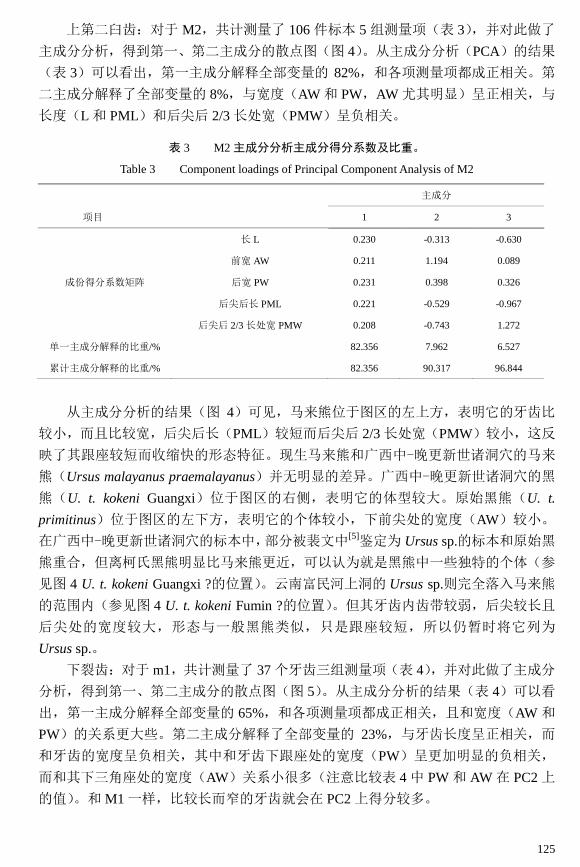
上第二臼齿:对于 M2,共计测量了 106 件标本 5 组测量项(表 3),并对此做了
主成分分析,得到第一、第二主成分的散点图(图 4)。从主成分分析(PCA)的结果
(表 3)可以看出,第一主成分解释全部变量的 82%,和各项测量项都成正相关。第
二主成分解释了全部变量的 8%,与宽度(AW 和 PW,AW 尤其明显)呈正相关,与
长度(L 和 PML)和后尖后 2/3 长处宽(PMW)呈负相关。
表 3 M2 主成分分析主成分得分系数及比重。
Table 3 Component loadings of Principal Component Analysis of M2
从主成分分析的结果(图 4)可见,马来熊位于图区的左上方,表明它的牙齿比
较小,而且比较宽,后尖后长(PML)较短而后尖后 2/3 长处宽(PMW)较小,这反
映了其跟座较短而收缩快的形态特征。现生马来熊和广西中-晚更新世诸洞穴的马来
熊(Ursus malayanus praemalayanus)并无明显的差异。广西中-晚更新世诸洞穴的黑
熊(U. t. kokeni Guangxi)位于图区的右侧,表明它的体型较大。原始黑熊(U. t.
primitinus)位于图区的左下方,表明它的个体较小,下前尖处的宽度(AW)较小。
在广西中-晚更新世诸洞穴的标本中,部分被裴文中[5]鉴定为 Ursus sp.的标本和原始黑
熊重合,但离柯氏黑熊明显比马来熊更近,可以认为就是黑熊中一些独特的个体(参
见图 4 U. t. kokeni Guangxi ?的位置)。云南富民河上洞的 Ursus sp.则完全落入马来熊
的范围内(参见图 4 U. t. kokeni Fumin ?的位置)。但其牙齿内齿带较弱,后尖较长且
后尖处的宽度较大,形态与一般黑熊类似,只是跟座较短,所以仍暂时将它列为
Ursus sp.。
下裂齿:对于 m1,共计测量了 37 个牙齿三组测量项(表 4),并对此做了主成分
分析,得到第一、第二主成分的散点图(图 5)。从主成分分析的结果(表 4)可以看
出,第一主成分解释全部变量的 65%,和各项测量项都成正相关,且和宽度(AW 和
PW)的关系更大些。第二主成分解释了全部变量的 23%,与牙齿长度呈正相关,而
和牙齿的宽度呈负相关,其中和牙齿下跟座处的宽度(PW)呈更加明显的负相关,
而和其下三角座处的宽度(AW)关系小很多(注意比较表 4 中 PW 和 AW 在 PC2 上
的值)。和 M1 一样,比较长而窄的牙齿就会在 PC2 上得分较多。
主成分
项目 1 2 3
长 L 0.230 -0.313 -0.630
前宽 AW 0.211 1.194 0.089
成份得分系数矩阵 后宽 PW 0.231 0.398 0.326
后尖后长 PML 0.221 -0.529 -0.967
后尖后 2/3 长处宽 PMW 0.208 -0.743 1.272
单一主成分解释的比重/% 82.356 7.962 6.527
累计主成分解释的比重/% 82.356 90.317 96.844
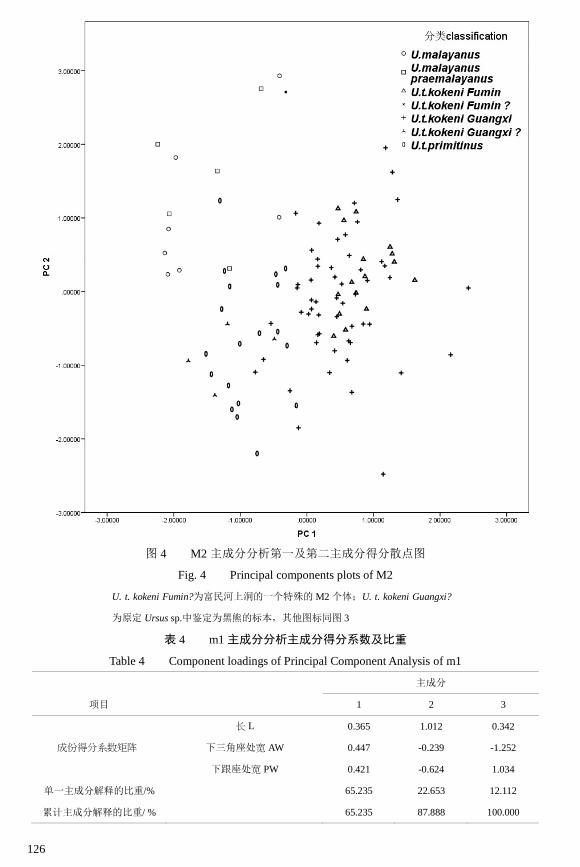
126
图 4 M2 主成分分析第一及第二主成分得分散点图
Fig. 4 Principal components plots of M2
U. t. kokeni Fumin?为富民河上洞的一个特殊的 M2 个体;U. t. kokeni Guangxi?
为原定 Ursus sp.中鉴定为黑熊的标本,其他图标同图 3
表 4 m1 主成分分析主成分得分系数及比重
Table 4 Component loadings of Principal Component Analysis of m1
主成分
项目 1 2 3
长 L 0.365 1.012 0.342
成份得分系数矩阵 下三角座处宽 AW 0.447 -0.239 -1.252
下跟座处宽 PW 0.421 -0.624 1.034
单一主成分解释的比重/% 65.235 22.653 12.112
累计主成分解释的比重/ % 65.235 87.888 100.000
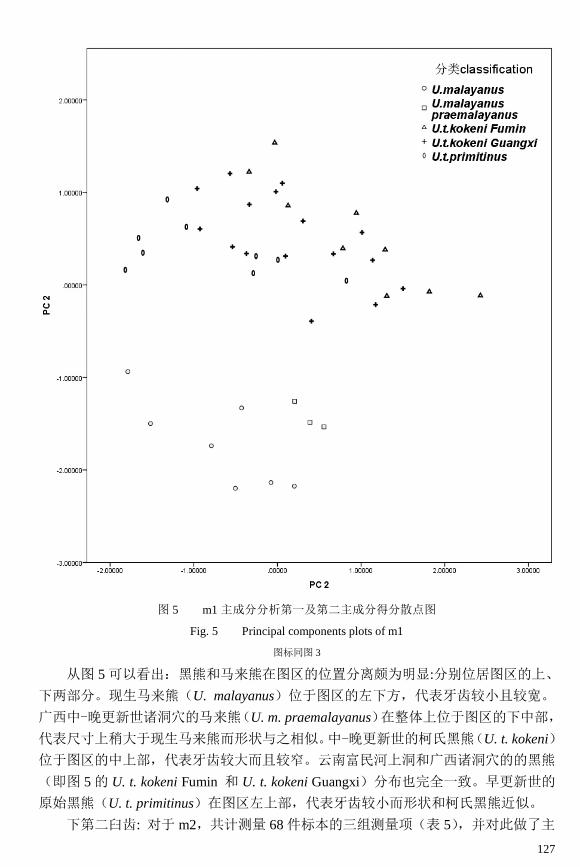
127
图 5 m1 主成分分析第一及第二主成分得分散点图
Fig. 5 Principal components plots of m1
图标同图 3
从图 5 可以看出:黑熊和马来熊在图区的位置分离颇为明显:分别位居图区的上、
下两部分。现生马来熊(U. malayanus)位于图区的左下方,代表牙齿较小且较宽。
广西中-晚更新世诸洞穴的马来熊(U. m. praemalayanus)在整体上位于图区的下中部,
代表尺寸上稍大于现生马来熊而形状与之相似。中-晚更新世的柯氏黑熊(U. t. kokeni)
位于图区的中上部,代表牙齿较大而且较窄。云南富民河上洞和广西诸洞穴的的黑熊
(即图 5 的 U. t. kokeni Fumin 和 U. t. kokeni Guangxi)分布也完全一致。早更新世的
原始黑熊(U. t. primitinus)在图区左上部,代表牙齿较小而形状和柯氏黑熊近似。
下第二臼齿: 对于 m2,共计测量 68 件标本的三组测量项(表 5),并对此做了主

128
成分分析,得到第一、第二主成分的散点图(图 6)。从主成分分析的结果(表 5)可
以看出,第一主成分解释全部变量的 81%,和各项测量项都成正相关,各个测量项的
相关程度几乎相等。第二主成分解释了全部变量的 14%,与牙齿长度(L)呈正相关,
而和下三角座处宽度(AW)呈负相关,与下跟座处的宽度(PW)则几乎没有关系。
表 5 m2 主成分分析主成分得分系数及比重
Table 5 Component loadings of Principal Component Analysis of m2
从图 6 可以看出,马来熊主要分布于图区的左下方,现生马来熊标本(U.
malayanus)较少而无法和广西中-晚更新世诸洞穴的马来熊(U. m. praemalayanus)区
分。柯氏黑熊(U. t. kokeni)位于图区中央,偏于马来熊的右上方,代表牙齿较大而
下前尖处的宽度较小,更加偏向于前窄后宽的形状。原始黑熊(U. t. primitinus)位于
马来熊的左上方,代表牙齿很小,而形态上则和柯氏黑熊近似。部分原始黑熊与马来
熊重合,而马来熊和柯氏黑熊也无法截然区分。
小结:总体来讲,主成分分析的结果突出说明了这批标本的尺寸(第一主成分 PC1)
和形状特点(第二主成分 PC2)。从图区的分布来看,广西其他诸洞穴和云南富民河上
洞的熊类标本中确有部分标本明显与现生马来熊更加接近,反之与黑熊比较远,显示
出短宽的特点。由此看来在中-晚更新世的广西诸洞穴和云南富民河上洞中确实存在
马来熊。在 M1 与 m1 中(图 3 和图 5),化石和现生马来熊又稍有不同,表现在 PC1
分值稍大(尺寸较大),而 PC2 分值相当(形状一致)。化石马来熊大于现生种其实是
完全可以理解的:这种现象并非个例,因为许多物种(如黑熊和大熊猫等)在地质历
史时期均呈现了类似的演化规律[5]。在 M2 与 m2 中(图 4 和图 6),化石和现生马来
熊混在一块,无法彼此分离。此外值得注意的是:马来熊的牙齿在大小尺寸上与早更
新世的原始黑熊颇为接近,除了 m1 的区分十分明显以外,其他很难与原始黑熊截然
区分。
5 相关问题讨论
马来熊为东洋界分子,现在在中国境内仅仅分布在最南方[9],相应的化石发现也
极少。马来熊为一种杂食性的熊,牙齿与其他熊类有较为明显的不同:犬齿巨大,犬
齿后脊发育并带有明显的锯齿;前臼齿 P1 ~ P3、p1 ~ p3 排列紧密,常常失去几个前
臼齿 [10];P4 的原尖通常不发育,但对应处仍有一膨突。由于发育程度不一,马来熊
的 P4 长宽比值变化很大,类似北极熊(Ursus maritimus);臼齿短宽,附尖发育弱;
主成分
项目 1 2 3
长 L 0.360 1.207 0.823
成份得分系数矩阵 下三角座处宽 AW 0.366 -1.069 1.121
下跟座处宽 PW 0.386 -0.112 -1.828
单一主成分解释的比重/% 80.923 12.760 6.317
累计主成分解释的比重/% 80.923 93.683 100.000
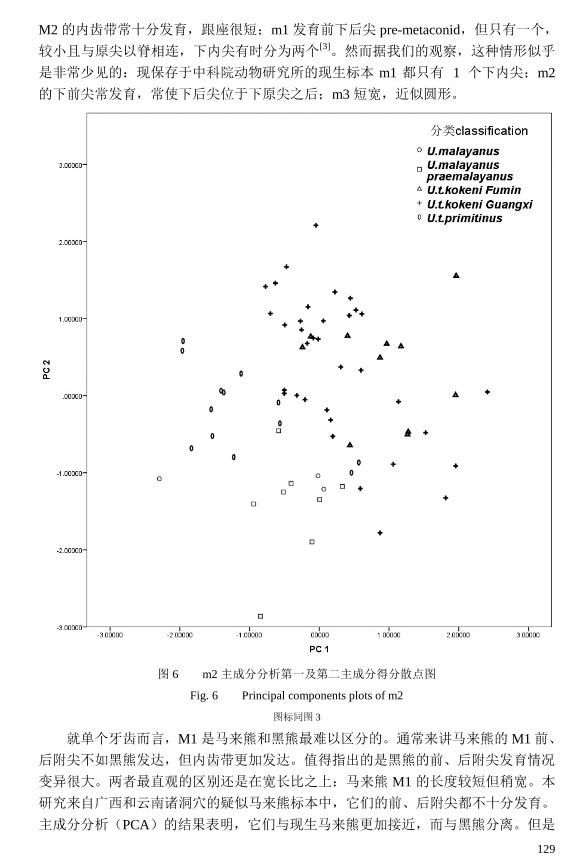
129
M2 的内齿带常十分发育,跟座很短;m1 发育前下后尖 pre-metaconid,但只有一个,
较小且与原尖以脊相连,下内尖有时分为两个[3]。然而据我们的观察,这种情形似乎
是非常少见的:现保存于中科院动物研究所的现生标本 m1 都只有 1 个下内尖;m2
的下前尖常发育,常使下后尖位于下原尖之后;m3 短宽,近似圆形。
图 6 m2 主成分分析第一及第二主成分得分散点图
Fig. 6 Principal components plots of m2
图标同图 3
就单个牙齿而言,M1 是马来熊和黑熊最难以区分的。通常来讲马来熊的 M1 前、
后附尖不如黑熊发达,但内齿带更加发达。值得指出的是黑熊的前、后附尖发育情况
变异很大。两者最直观的区别还是在宽长比之上:马来熊 M1 的长度较短但稍宽。本
研究来自广西和云南诸洞穴的疑似马来熊标本中,它们的前、后附尖都不十分发育。
主成分分析(PCA)的结果表明,它们与现生马来熊更加接近,而与黑熊分离。但是

130
化石标本中的中附尖常与后小尖相连紧密,与现生马来熊稍有不同,后者的中附尖常
呈孤立状。
现生马来熊的 M2 形态比较有特点:内齿带通常十分发育,跟座较短(在现生熊
属 Ursus 中是最短的,一些北极熊个体除外)。在广西诸洞穴的标本中,M2 的内齿带
都比较发育,很好地体现了马来熊的特征。从主成分分析(PCA)结果来看,它们与
现生马来熊近乎一致(图 4),彼此间难以区分。从统计数据来看,广西标本的跟座稍
长(跟座长指数 PML/L 为 0.51 ~ 0.67),而现生种较短(PML/L 仅为 0.38 ~ 0.57),体
现了化石标本与现生种的细微不同。这或许是化石标本体型增大的缘故?是否如此?
尚有待今后的深入调查与研究。
下裂齿 m1 是熊类最有鉴定意义的牙齿。产自广西诸洞穴的下裂齿标本呈现出短
宽的形态特点;在未磨蚀的标本中,下后尖为单个且与下原尖以脊相连;下后尖后没
有延伸向下次尖的脊。在这些方面,它们显然与现生的马来熊一致,区别于黑熊。其
余的两个标本虽然磨蚀严重,但前下后尖的形态及主成分分析(PCA)的结果都倾向
支持它们隶属马来熊。在化石标本中,m1 的前下后尖常高于下后尖,但现生标本的
两尖高度相近。
在广西诸洞穴的标本中,m2 的下前尖较发育,下后尖在下原尖之后,与马来熊
的特征相符,而与黑熊稍有不同。虽然某些标本的下前尖并不发育,下后尖与下原尖
并行排列,但它们的宽长比明显更接近于马来熊。值得进一步指出的是现生马来熊标
本中也存在类似的形态(下前尖不发育,下后尖与下原尖并行),客观地讲,马来熊
m2 的形态变化较大。
Erdbrink[3]列出了 Von Koenigswald 在中药铺购买的马来熊牙齿化石,并给出了一
些描述。除了 M2 以外,广西诸洞穴的这批标本与 Von Koenigswald 购买的马来熊化
石标本在尺寸大小上彼此吻合(表 6)。事实上,在广西的标本中除了一个长度为
16.32 mm 的 M2 外,其余的牙齿范围为 18.10 ~ 22.36 mm,与 Erdbrink 所列差距其实
也不大。爪哇岛 Punung 和越南 Duoi U’Oi 两地点的马来熊体型的大小与我国中-晚更
新世的同类标本相似(表 6)。由此不难发现:地质历史时期的马来熊较现生种要稍大
些,与黑熊和大熊猫一样,马来熊从中更新世到现在也经历了体型逐步变小的过程。
总体来讲,广西和云南中-晚更新世诸洞穴的这批熊类牙齿,无论是在形态上还
是大小尺寸上都与现生的马来熊十分接近,区别于现生和化石黑熊。虽说它们尺寸上
稍大于现生的马来熊,但与 Von Koenigswald 订为 Ursus praemalayanus 的标本比较吻
合,考虑到 Von Koenigswald 的标本可能同样来自中-晚更新世的中国南方[3],因此有
理由认为广西和云南的这批标本与 Von Koenigswald 购买的标本应为同一物种,据此
鉴定为 U. praemalayanus。然而本研究的形态比较和数理统计分析的结果表明:虽说
化石马来熊稍大于现生种,且形态上也有若干不同,不过这种差异很细微,通常情况
下两者相互很难区分(如 M2 和 m2),这些不同更像是个体间或是居群间,而非种间
的形态差异。Erdbrink 认定 Von Koenigswald 购买的那些标本就是现生马来熊一样,本
研究倾向支持 Erdbrink 的这一结论。因此,广西和云南中-晚更新世诸洞穴的这批熊
标本可以作为现生马来熊的一个史前亚种,即 Ursus malayanus praemalayanus。

131
表 6 马来熊牙齿长度统计
Table 6 Length of teeth of sun bears mm
牙齿 广西
诸洞穴
云南富民
河上洞
Von Koenigswald
购买[10]
Punung
爪哇岛[14]
Duoi U’Oi
越南)[15]
现生种
M1 15.90~17.86
(n=7)
15.84~15.96
(n=2)
15.50~18.00
(n=17)
14.22~16.04
(n=7)
M2 16.32~22.36
(n=5)
19.50~25.00
(n=19)
20.00~24.00
(n=4)
17.78~23.14
(n=7)
m1 18.00~18.42
(n=3)
17.00~19.00
(n=18)
18.00
(n=2)
16.90~18.20
(n=2)
15.00~17.62
(n=7)
m2 15.82~19.32
(n=8)
16.00~18.50
(n=12)
17.00
(n=2)
15.18~18.94
(n=3)
由此看来,在中-晚更新世时期我国的南方(主要是广西和云南地区)就已出现
马来熊,其地理分布范围较现今的要稍大些(譬如广西地区)。另外湖北郧西也曾经
报道有马来熊化石的发现[6],化石马来熊在我国的分布似乎一度非常广泛。然而郧西
的标本鉴定值得商榷,郧西的标本包括若干犬齿、一个 M1、一个 M2 和两个 m2 [6]。
从图版和测量来看,郧西的 M1 标本虽然长度上很小、接近马来熊,但明显较马来熊
狭窄,其宽长比落入黑熊的变异范围。郧西的 M2 虽然小,但仍然处在黑熊的变异范
围之内,且其前尖处宽(AW)明显小于后尖处宽(PW),这与黑熊相似,而在马来
熊中前尖处宽(AW)通常大于后尖处宽(PW),这可能与其内齿带发育有关。虽然
这个标本的跟座也较短,但整体形态还是更加接近黑熊。两个 m2 都磨损较为严重,
但从轮廓上来看下后尖和下原尖并行排列,下前尖似乎未发育,另外它们的大小和宽
长比也完全落入黑熊的变异范围之内。这样看来,这一批郧西的标本,除了较小的
M1 之外,其实和黑熊标本没什么区别,而和马来熊相差很远。因此,这些郧西标本
实际上属于黑熊,可能代表了一些极端小的个体。
Zdansky 描述了在山西沁县发现的上新世晚期的熊类化石,归入 Ursus cf.
boeckhi [2],Thenius、邱占祥(都认为归入中华马来熊 Ursus sinomalayanus。它的前臼
齿列较短,联合部高,m1 的前下后尖和下原尖以脊相连,下内尖双分,符合马来熊
的一般特征[4],但其尺寸相对较大,大小为 20.7 mm × 9.7 mm,长度明显大于中-晚
更新世的化石和现生马来熊。其宽长比为 0.47,处于马来熊的下限之外(0.53 ~ 0.60),
但比大多数的黑熊稍宽(0.42 ~ 0.48)。
纳玛熊 Ursus namadicus 是 Falconer 和 Cautley 在描述纳玛动物群时建立的,至
今也只有当时建立种时的材料。Lydekker [10]和 Pilgrim
[11]都认为该种最接近于马来熊。
化石出产于中更新世的地层中 [11-12]。Pilgrim
[10]认为它的形态比马来熊更进步,而不可
能马来熊的直系祖先。纳玛熊体型较大,P4 约为 16.51 mm × 13.46 mm,M1 为
19.00 mm × 18.03 mm,M2 长 26.92 mm[10]。每个牙齿都大于中-晚更新世的化石和现
生马来熊的上限。P4 的原尖大而靠后,与后尖分离明显,与现生马来熊明显不同。根

132
据图版分析,M2 的前尖处宽(AW)似乎大于后尖处宽(PW),和马来熊类似,但它
的跟座较长,与马来熊则不同[10]。其实,马来熊的主要特征是其牙齿较宽,纳玛熊在
此方面表现尤为突出:它 M1 的宽长比为 0.95,高于现生种(0.92)和中-晚更新世的
马来熊(0.86),远大于熊属其他种类。在这一特征上它有些接近南美短面熊
Arctotherium(M1 的宽长比通常大于 1),但在 Arctotherium 中(A. angustidens 除外),
P4 的原尖通常较小而退化,接近牙齿的内侧中央;M2 的跟座较短,类似现生马来熊,
也与纳玛熊不同,而且南美短面熊从未在南美以外被发现过 [12],因此纳玛熊不太可能
属于 Arctotherium。从目前掌握的资料来看,Pilgrim [11]的观点似乎更加可取,即它们
可能在形态上接近于马来熊,但不是后者的直接祖先,可能代表了马来熊演化史上的
一个绝灭旁支。
致谢 中国科学院古脊椎动物与古人类研究所的邱占祥院士在研究中提供了相关文
献和热心指导。在研究测量标本的过程中,笔者得到中国科学院古脊椎动物与古人类
研究所标本馆的陈津女士及中国科学院动物研究所曹小平女士的大力帮助。此外,中
国科学院古脊椎动物与古人类研究所的史勤勤博士和中国地质大学(北京)欧强副教
授在论文修改和图版制作方面给予帮助。在此一并表示衷心感谢。
参 考 文 献
1 Yu L, Li Q, Ryder O A, et al. Phylogeny of the bears (Ursidae) based on nuclear and mitochondrial genes. Molecular
Phylogenetics and Evolution, 2004, 32(2): 480-494.
2 Zdansky O. Weitere Bemerkungen über fossile Carnivoren aus China. Palaeont Sin, 1927, Ser C, 4(4): 1-28.
3 Erdbrink D P. A review of fossil and recent bears of the Old World, with remarks on their phylogeny based upon their
dentition. Deventer Drukkerij Jan De Lange, 1953 (1-2): 1-597.
4 邱占祥,邓涛,王伴月.甘肃东乡首次发现熊亚科化石——龙担哺乳动物群补充报道之二(英文).古脊椎动物学
报, 2009, 47(4): 245-264.
5 裴文中. 广西柳城巨猿洞及其它山洞之食肉类、长鼻目和啮齿类化石. 中国科学院古脊椎动物与古人类研究所集
刊,1987, 18: 1-134.
6 武竹仙. 郧西人:黄龙洞遗址发掘报告. 北京: 科学出版社, 2006. 163-170.
7 Augeri D M. On the biogeographic ecology of the Malayan sun bear. Ph.D. Thesis, University of Cambridge, 2005.
1-349.
8 卞美年,贾兰坡. 云南之洞穴及岩洞沉积. 中国地质学会志, 1938, 18(3-4): 325-349.
9 髙耀亭等.中国动物志,兽纲,第八卷:食肉目. 北京:科学出版社,1987. 76-102.
10 Lydekker R. Indian Tertiary and post-Tertiary Vertebrata. Part 6. Siwalik and Narbada Carnivora. Palaeont Indica, 1884,
10(2): 178-363.
11 Pilgrim G E. The fossil Carnivora of India. Pal Indica, NS, 1932, 18: 49-51.
12 Nanda A C. Comments on the Pinjor Mammalian Fauna of the Siwalik Group in relation to the post-Siwalik faunas of
Peninsular India and Indo-Gangetic Plain. Quaternary International, 2008, 192(1): 6-13.

133
13 Soibelzon L H, Rincon A D. The fossil record of the short-faced bears (Ursidae, Tremarctinae) from Venezuela:
Systematic, biogeographic, and paleoecological implications. Neues Jahrbuch für Geologie und Paläontologie-
Abhandlungen, 2007, 244(3): 287-298.
14 Badoux D M. Fossil Mammals from Two Fissure Deposits at Punung (Java): With Some Remarks on Migration and
Evolution in South East Asia. Doctoral dissertation. Hague, Drukkerij en Uitgeversmij. v/h Kemink en Zoon NV, 1959:
81-87.
15 Bacon A M, Demeter F, Duringer P, et al. The Late Pleistocene Duoi U’Oi cave in northern Vietnam: palaeontology,
sedimentology, taphonomy and palaeoenvironments. Quaternary Science Reviews, 2008, 27(15): 1627-1654.
MIDDLE AND LATE PLEISTOCENE SUN BEARS
(URSIDAE, CARNIVORA) FROM SOUTHERN CHINA
JIANGZUO Qi-gao1 CONG Hao-liang
1 MA Rui
1 FENG He
1 LIU Jin-yi
2
(1 China University of Geosciences, Beijing 100083;
2 Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044)
ABSTRACT
The Pleistocene fossil bears teeth from Southern China are restudied and revised in
present paper. Some specimens from Guangxi Zhuang Autonomous Region (eight M1, five
M2, three m1, eight m2) and Yunnan Province (two M1, a possible M2) could be referred to
sun bears Ursus (Helarctos) malayanus.
These specimens are basically similar to the modern sun bears in both size and
morphology, but they also display some minor difference. The fossils are somewhat large
and narrow in teeth. The mesocone of M1, apart from protocone, is often connected tightly
with metaconule. Some specimens even develop no mesocone (possibly fused with
metaconule). In modern sun bears, however, the mesocone is more distinct and not
connected with any cones. As regards M2, there are no siginificant differences between
fossil and modern sun bears, except the talon in which fossil bears is slightly long ( PML,
measured as length between apex of metacone to the tip of the talon).The lower carnassial is
the most diagnostic teeth for Ursus. The pre-metaconid is connected with protoconid with a
ridge weaker than modern bear. In all three fossil teeth the pre-metaconid is higher than
metaconid, while the two cones are of the same height in the modern specimens. The only
specimen with an unworn talonid displays only one entoconid. In the teeth of m2, the

134
paraconid is often developed, and the protoconid is generally situated anterior to the
metaconid due to the development of paraconid, except some specimens in which the ridge
between metaconid and protoconid is almost perpendicular to the long axis of tooth. On the
basis of the similarities in both size and morphology, the present authors would refer these
fossil bears from Guangxi and Yunnan to Ursus malayanus, and consider then as a fossil
subspecies of modern sun bears, namely Ursus malayanus praemalayanus. Moreover, Sun
bears from Java and Vietnam from middle and late Pleistocene could be referred to the same
subspecies. Teeth of small bears from Yunxi, Hubei Province is, however, much narrower
than those of sun bears and resemble to black bears morphologically, thus the fossil sun bear
is only limited to the Southernmost part of China in Middle and Late Pleistocene.
Key words Guangxi, Yunnan, Middle-Late Pleistocene, sun bears




























