Embed Size (px)
Citation preview

Mitochondrial Induction of Adult T Cell Leukemia DerivedFactor (ADF/hTx) After Oxidative Stresses in RetinalPigment Epithelial Cells
Chiaki D. Gauntt,* Akihiro Ohira,* Osamu Honda,* Kazuteru Kigasawa,fToyoshi Fujimoto,X Hiroshi Masutani,§ Junji Yodoi,§ and Yoshihito Honda*
Purpose. Adult T cell leukemia derived factor (ADF) is a human homologue of thioredoxin(hTx), which exhibits scavenging activity with reactive oxygen intermediates. In their previousstudy, the authors found that after transient retinal ischemia, the expression of thioredoxin inrat retinal pigment epithelium (RPE) layer increased markedly. The present investigation is todetermine intracellular ADF localization in RPE after transient ischemia and in cultured hu-man RPE cells after oxidative insult by H2Oa.
Methods. The authors employed immunoelectron microscopy to examine ADF localization inRPE. Labeling density analysis was performed to supplement the main observation in theexperiment of transient retinal ischemia. 3-(4,5-dimethylthiazol-2yl)-2-5-diphenyltetrazoliumbromide (MIT) assay was performed to verify the protective role of recombinant ADF (rADF)against HaOjj.
Results. In immunogold electron microscopy, sparse ADF-positive labeling was seen in thecytosol and mitochondria in normal rat RPE and in untreated cultured RPE cells. After oxida-tive stress, it was concentrated in mitochondria in both groups. MTT assay proved that rADFprotected cultured RPE from the toxicity*of H2O2.
Conclusions. This study shows the induction of ADF/hTx in mitochondria of RPE after oxida-tive stresses and its protective effect on cultured RPE exposed to Y\tO2- The data indicate thepossibly important role of ADF/hTx in the protection of retinal cells from the oxidativestresses associated with retinal ischemic disease and probably with regular visual activity. InvestOphthalmol Vis Sci. 1994;35:2916-2923.
Adult T cell leukemia derived factor (ADF) was firstdiscovered in 1985 as an inducible protein that hasseveral co-cytokine functions.1""' Soon it was proventhat ADF is the human homologue of thioredoxin(hTx), which is found in most species and keeps mark-edly high homology among species.6"0 Thioredoxinpossesses dithiol-disulfide oxidoreductase activity,which catalyzes the refolding of disulfide-containingproteins.10"13 It also has a scavenging function against
From the *Departments of Ophthalmology and ^Anatomy, Faculty of Medicine,Kyoto University; the ^Department of Ophthalmology, Tohai University School ofMedicine; and §The Institute for Virus Research, Kyoto University, Japan.Supported by a Grant-in-Aid for Scientific Research from the Ministry of Education,Science and Culture of Japan (#04671072, AO).Submitted for publication August 6, 1993; revised December 14, 1993; acceptedFebruary 10, 1994.Proprietary interest category: N.Reprint requests: Chiaki D. Gauntt, Ml), Department of Ophthalmology, Faculty ofMedicine, Kyoto University, Sakyo-ku, Kyoto, 606, Japan.
reactive oxygen intermediates, especially hydroperox-ide.14 Thioredoxin is expressed in many animal celltypes, including human cells.15 It is strongly expressedin malignant or activated tissue and is induced by ultra-violet or x-ray irradiation and viral infection.51G~20 It iswell known that when the blood supply recovers aftertissue ischemia, superoxide is produced and by super-oxide dismutase (SOD) it is converted to H2O2. Thesereactive oxygen intermediates are reported to be re-sponsible for various tissue injuries21"24 and also clini-cal retinal ischemic disease.25"27 In rat retina, we foundthat ADF localized mainly in the ganglion cell layerand inner plexiform layer under normal conditionsthrough light microscopic immunochemical tech-niques.28 Recently, through the same techniques, weobserved strong ADF induction in retinal pigment epi-thelial cells (RPE) in the rat retinal ischemia/reperfu-sion model.28
2916Investigative Ophthalmology & Visual Science, June 1994, Vol. 35, No. 7Copyright © Association for Research in Vision and Ophthalmology

Mitochondrial ADF Induction 2917
The present study examined the electron micro-scopic intracellular localization of ADF under normalconditions and after ischemia-reperfusion in rat RPEand in cultured human RPE cells using immunocyto-chemical methodology to determine the ADF induc-tion and distribution in detail. The immunogold parti-cle density of the electron micrographs was alsoanalyzed in vivo to evaluate the result with semiquanti-tative data. Our hypothesis that the phenomenon isfor cellular protection is supported by the result of the3-(4,5-dimethylthiazol-2yl)-2-5-diphenyltetrazoliumbromide (MTT) assay, proving the protective effect ofrADF against the toxicity of H2O2.
MATERIALS AND METHODS
Rat Model of Retinal Transient Ischemia
Four adult male albino Sprague-Dawley rats (S-D rats),each weighing 200 to 250 g, were used in this study.Because mammalian retinas are controlled by circa-dian rhythms, the rats used in this experiment weremaintained in 12-hour light-dark cycles and werekilled at identical hours during the day. The animalswere treated according to the ARVO Statement forthe Use of Animals in Ophthalmic and Vision Re-search during the study. We created photodynamictransient retinal venous thrombosis according to a pre-viously described method.29"30 In brief, we injectedphotosensitizing dye rose bengal (Sigma, St. Louis,MO) in dosages of 80 mg/kg intravenously into therats and, under direct observation, exposed a retinalvein to light using a Haag-Streit photo slit-lamp(Haag-Streit, Bern, Switzerland) at a 3 X 0.5 mm slitlight size and 127 W/m2 until the vessel was completelyoccluded. When albino rats are used, the choroidalvascular bed also collapses in the area of light expo-sure so that the entire neural retina and RPE becomesischemic. This thrombosis dissolves spontaneously in ashort time (at most a few days). As a control, two male,nontreated, albino S-D rats of the same weight and agegroup were chosen. We also created a control retinathat was exposed to the same amount of light withoutrose bengal administration and an unexposed controlthat was injected with rose bengal. By light micro-scopic immunohistochemistry, we observed no signifi-cant change in RPE of these controls.28
Immunogold Electron Microscopy of ADF/hTx inRPE of Retinal Transient Ischemic Model. Four modelrats 72 hours after thrombosis by rose bengal and twocontrol rats without the treatment were anesthetizedby intraperitoneal injection of sodium pentobarbital.They were perfusion-fixed through the left heart ven-tricle with freshly prepared 4% paraformaldehyde(PFA) containing 0.2% glutaraldehyde, pH 7.3. Thenthe eyes were enucleated and dissected. The eyecups
were further fixed by immersion in the same fixative at4°C for an additional 30 minutes. The retina was cutinto single cubic pieces and immersed in 2.3 M su-crose-0.1 M phosphate buffer solution for 60 min-utes. The pieces were placed on specimen holders andfrozen in liquid nitrogen. Ultrathin cryosections (50 to100 nm in thickness) were prepared with an ultrami-crotome (Reichert FC4, Leica, Germany) and placedon carbon formvar-coated grids. The sections werefirst incubated with polyclonal affinity-purified anti-ADF antibody 4 /tg/ml in 10 mM glycine-PBS, or non-immune rabbit IgG as a control, for 30 minutes atroom temperature, then with colloidal gold conju-gated anti-rabbit IgG antibody in dilution of 1:40(Amersham, Tokyo, Japan) for another 30 minutes atroom temperature. Subsequently, these sections werepoststained for 5 minutes on ice with 2% uranyl ace-tate, then embedded in 2% methylcellulose containing0.4% uranyl acetate.33 The grids were examined withan electron microscope (1200 EX, JEOL, Tokyo, Ja-pan). From each animal, 20 ultrathin sections wereexamined.
The antibody was raised by immunization with the10 mer synthetic peptide corresponding to the C-ter-minal sequence of ADF protein conjugated with bo-vine serum albumin (BSA) in the presence of completeFreund's adjuvant. The specific anti-ADF peptide wasaffinity purified using Sepharose CL-4B coupled withADF C-terminal 10 mer peptide conjugated with BSA,after absorption by BSA-sepharose.12 Cold inhibitionwas used to test antibody specificity. rADF was elec-trophoresed. After blotting, the membrane was incu-bated with 3 Mg/m' of anti-ADF antibody that hadbeen preincubated with 0 to 200 ng of rADF. Singlebands at 13 kD were detected in all the lanes except forthe lane incubated with 200 ng of rADF added to anti-ADF antibody. The bands were more intensive on adose-dependent basis when the rADF that was addedto the antibody decreased.13
Immunogold Particle Density Analysis. Two groupseach of 25 ultrathin sections were prepared frommodel and control RPE. Then 100 random electronmicrographs were taken from each group. We ob-served in a pilot study that immunolabeling in the mi-tochondria of model RPE markedly increased while inother parts of the cell it stayed the same after transientischemia. We paid attention to the mitochondrial im-munolabeling density and compared it to that of otherparts. The surface area in /nm2 of mitochondria and thecytoplasm were first measured using a Digigrammermodel G-2 (Mutoh, Tokyo, Japan), and the gold parti-cles were directly counted in the measured area. Thedata were processed using Lotus 1-2-3 on a Macintosh(Apple Computer, Cupertino, CA) model Ilsi. The to-tal measured areas of mitochondria of model RPE andcontrol RPE and cytoplasm of the model and the con-

2918 Investigative Ophthalmology & Visual Science, June 1994, Vol. 35, No. 7
trol were 139.4 nm2, 150.1 nm2, 387.3 jan2, and 365.3/im2, respectively. Only well-defined mitochondriawere measured, and particles on the mitochondrialmembrane were included in the calculation. For themeasurement of cytoplasm, the areas were carefullychosen from the zones that were clearly visualized.The particles seen around the endoplasmic reticulumwere counted as existing in the cytoplasm. BecauseADF/hTx is a soluble protein, we considered that theywere actually in the cytoplasm. The particles in thenucleus, cell membrane, and other organellae weresparse and did not show significant differences fromthose of control micrographs. The mean and standarddeviation were calculated and shown on a histogram.
Human RPE Cell Line
RPE cell line (K-1034) was established from a 26-year-old human eye that had to be enucleated because of anorbital angioma.81 Tenets of the Declaration of Hel-sinki were followed, and informed consent was ob-tained. Institutional human experimentation commit-tee approval was granted. The cells usually exhibit aflat epitheloid shape or a thin spindle shape. They arestrongly positive for human keratin and partially posi-tive for melanin pigment when stained with Masson'smethod. They proliferate rapidly on a collagen type-1coated plastic culture dish surface and actively phago-cytize latex beads 1 fim in diameter. These cells werefirst cultured in Ham's F-12 medium containing 10%fetal bovine serum until just before they achieved con-fluency. Cells at the 70th to 80th passage levels wereused in this experiment. Cells were subsequently incu-bated for 6 hours in serum-free Ham's F-12 mediumsupplemented with 100 fxM H2O2 in the same incuba-tor as a culture dish of untreated cells used as a con-trol.
Light Micrographic Immunohistochemistry of ADF/hTX in Cultured Human RPE. The cultured RPE cellsexposed to H2O2 and normally maintained cells (con-trol) were rinsed with PBS and then fixed with 2% PFAcontaining 25% Bouin solution (2,4,6-trinitrophenolsaturated in water:37% formalin solution: 100% aceticacid = 15:5:1) for 1 hour on ice, submerged in 100%ethanol containing 0.3% H2O2 for 6 minutes to eraseendogenous peroxidase, and then incubated with thesame polyclonal affinity-purified anti-ADF antibody 2/ig/ml overnight at 4°C, horseradish peroxidase(HRP)-conjugated anti-rabbit IgG 10 Mg/ml 90 min-utes at 37°C, and 3-amino-9-ethylcarbazole (AEC)(Biomeda, Foster City, CA) 10 minutes at room tem-perature, respectively. For the control staining, thefirst antibody was replaced with 2 Mg/ml °f normalrabbit IgG. The samples were observed through lightmicroscopy. The experiment was repeated threetimes.
Immunogold Electron Microscopy of ADF/hTx inCultured Human RPE. The cultured RPE cells incu-bated with H2O2 and nontreated control cells werefixed for 1 hour on ice in 2% PFA containing 0.2%glutaraldehyde (pH 7.4) and then further fixed in 2%PFA at 4°C overnight. The cells were gently scrapedoff and centrifuged at 1200 rpm for 5 minutes, form-ing loose pellets. The pellets were washed in PBS, em-bedded in 10% gelatin, cut into small blocks, and thenimmersed in 2.3 M sucrose containing 4% polyvinylal-cohol, standing overnight at 4°C. They were mountedon specimen folders and frozen in liquid nitrogen.
Using the previously described methods, ultrathincryosections were prepared, incubated with a series ofantibodies, embedded, and examined with electron mi-croscope. Thirty ultrathin sections from each groupwere examined. The experiment was repeated threetimes.
Toxicity Assay of H2O2. Human cultured RPE cellswere preincubated with and without rADF (1 to 100Mg/ml) for 1 hour in 100 /ul of serum-free Ham's F-12medium in 96-well culture plates (Corning, NY). Sub-sequently, various concentrations of H2O2 were ap-plied to each well. After 24 hours of incubation, MTT(Sigma, St. Louis, MO) solution (5 fig/ml MTT in PBS)was added to all wells (10 fil to each well), followed byincubation at 37°C for 4 hours. An aliquot of 100 fil of0.04 N HCl-isopropanol was added to all wells andmixed thoroughly to dissolve the blue MTT formazan.The toxicity of H2O2 was estimated by reduced absor-bance as measured by an ELISA reader (MicroplateReader, model 3550, BioRad, Richmond, CA), with atest wave length of 570 nm and a reference wavelength of 630 nm.34 Each assay was carried out in tripli-cate.
For precise comparison, all tissues, cell sampleprocessing, and immunologic staining procedureswere carried out simultaneously under identical con-ditions.
RESULTS
Rat Model of Transient Retinal Ischemia
Immunogold Electron Microscopy of ADF/hTx inRPE of Retinal Transient Ischemic Model. In the non-treated S-D rat RPE, immunogold particles were ob-served along the endoplasmic reticulum and in thesurrounding cytoplasm. The mitochondria weresparsely labeled (Fig. 1A). The cell membrane and thenucleus were faintly labeled (data not shown). Thecontrol showed no background labeling (Fig. IB).
In the RPE of the transient retinal ischemicmodel, many gold particles were observed in mito-chondria, whereas immunolabeling in the cytoplasmand the endoplasmic reticulum were similar to that of

Mitochondrial ADF Induction 2919
ER
ER
B
FIGURE l. Immunoelectron micrographs of retinal pigmentepithelium of normal rat. Bar = 0.2 ^m. (A) ADF labelingassociates mainly with endoplasmic reticulum and surround-ing cytoplasm. Mitochondria are only sparsely labeled by thegold particles. (B) Control micrograph shows little back-ground labeling. M = mitochondria; N = nucleus; ER =endoplasmic reticulum.
treated cells were stained faintly (Fig. 4B). The controlshowed no background staining (data not shown).
Immunogold Electron Microscopy of ADF/hTx inCultured Human RPE. Through immunocytochemicalinvestigation, the RPE cells exposed to H2O2 indicatedabundant ADF-positive immunolabeling intensively inmitochondria and moderately in cytoplasm aroundmitochondria (Fig. 5A). The normally maintained cellsshowed moderately sparse labeling of ADF/hTx in cy-toplasm and in mitochondria {Fig. 5B). The controlmicrographs suggested no background staining (Fig.5C).
The viability of the RPE treated with 100 nMH2O2 for 6 hours was between 86% and 94%, and thatof control without the treatment was between 92% and96%. This excludes the possibility of dead cell samples.
Toxicity Assay of H2O2. The analysis in MTT assaydemonstrated that the sensitivity of the RPE cellsagainst H2O2 was effectively reduced by the preincu-bation with rADF (Fig. 6). After preincubation with 1
ER
M
nontreated RPE (Fig. 2A). The background immuno-labeling was absent in the control electron micro-graphs (Fig. 2B).
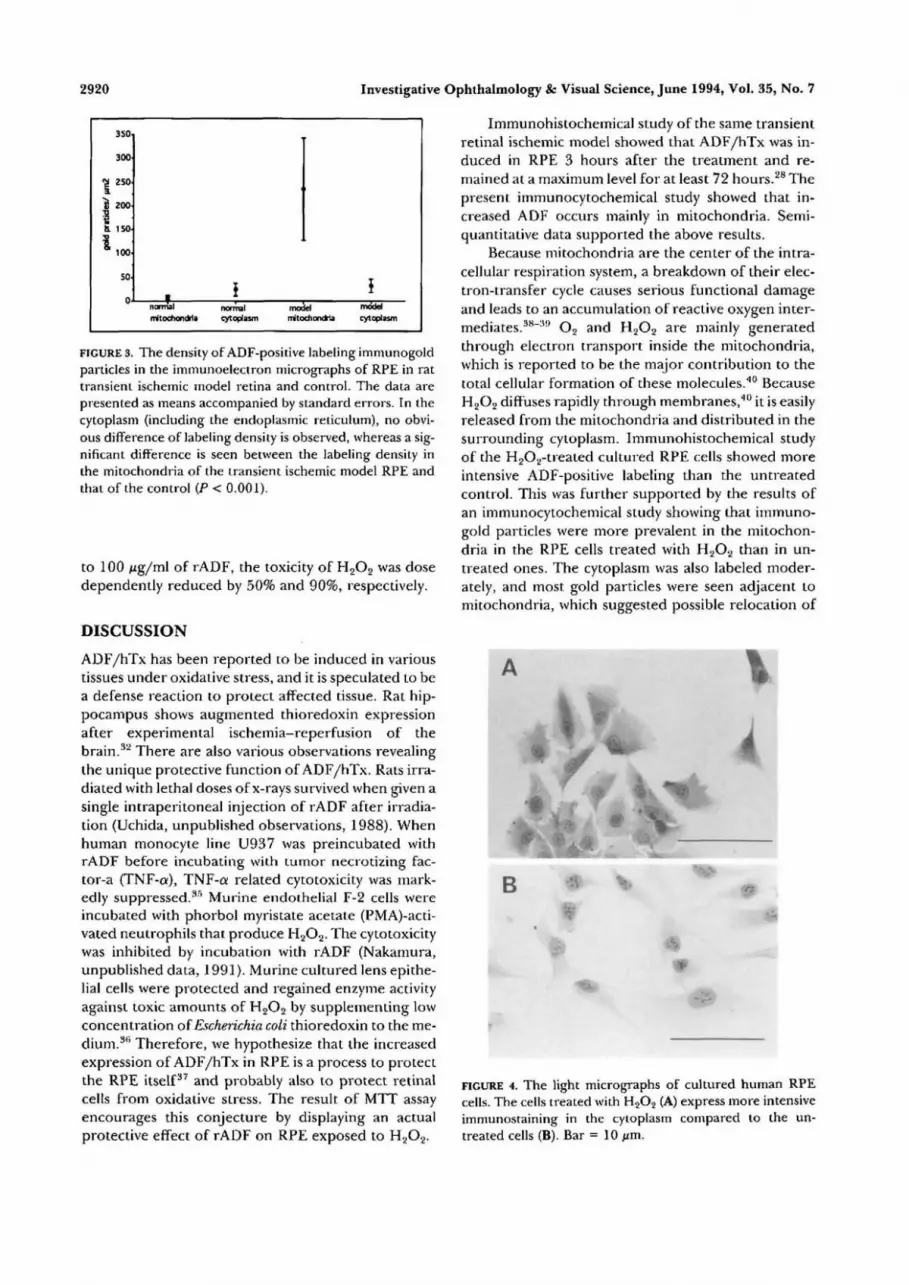
The histogram showed that the density of goldparticles in mitochondria of RPE was significantlygreater in the ischemic model retina than in the con-trol (P < 0.001 by Student's i-test), whereas the densityin the cytoplasm and on the endoplasmic reticulumdid not show any difference between the two groups(Fig. 3).
Human RPE Cell Line
Light Microscopic Immunohistochemistry of ADF/hTx in Cultured Human RPE. The light micrographs ofcultured RPE cells showed that the cells exposed toH2O2 (Fig. 4A) expressed ADF intensively, and un-
B
FIGURE 2. Immunoelectron micrographs of RPE of transientretinal ischemic rat model. Bar = 0.2 fim. (A) Mitochondriaare heavily labeled for ADF. Density of gold particles in thecytoplasm and endoplasmic reticulum are equivalent to thatof control RPE. (B) Control micrograph shows little back-ground labeling. M = mitochondria; ER = endoplasmic retic-ulum.
2920 Investigative Ophthalmology & Visual Science, June 1994, Vol. 35, No. 7
111
350,
300
250
200
ISO
10a
50'
0- fnOFTTUl
mitochondria
inormalcytoplasm
j
Inrtodd moddmitochoncfcia cytoplasm
FIGURE 3. The density of ADF-positive labeling immunogoldparticles in the immunoelectron micrographs of RPE in rattransient ischemic model retina and control. The data arepresented as means accompanied by standard errors. In thecytoplasm (including the endoplasmic reticulum), no obvi-ous difference of labeling density is observed, whereas a sig-nificant difference is seen between the labeling density inthe mitochondria of the transient ischemic model RPE andthat of the control (P < 0.001).
to 100 Mg/m' of rADF, the toxicity of H2O2 was dosedependently reduced by 50% and 90%, respectively.
DISCUSSION
ADF/hTx has been reported to be induced in varioustissues under oxidative stress, and it is speculated to bea defense reaction to protect affected tissue. Rat hip-pocampus shows augmented thioredoxin expressionafter experimental ischemia-reperfusion of thebrain.32 There are also various observations revealingthe unique protective function of ADF/hTx. Rats irra-diated with lethal doses of x-rays survived when given asingle intraperitoneal injection of rADF after irradia-tion (Uchida, unpublished observations, 1988). Whenhuman monocyte line U937 was preincubated withrADF before incubating with tumor necrotizing fac-tor-a (TNF-a), TNF-a related cytotoxicity was mark-edly suppressed.35 Murine endothelial F-2 cells wereincubated with phorbol myristate acetate (PMA)-acti-vated neutrophils that produce H2O2. The cytotoxicitywas inhibited by incubation with rADF (Nakamura,unpublished data, 1991). Murine cultured lens epithe-lial cells were protected and regained enzyme activityagainst toxic amounts of H2O2 by supplementing lowconcentration of Escherichia coli thioredoxin to the me-dium.3'1 Therefore, we hypothesize that the increasedexpression of ADF/hTx in RPE is a process to protectthe RPE itself37 and probably also to protect retinalcells from oxidative stress. The result of MTT assayencourages this conjecture by displaying an actualprotective effect of rADF on RPE exposed to H2O2.
Immunohistochemical study of the same transientretinal ischemic model showed that ADF/hTx was in-duced in RPE 3 hours after the treatment and re-mained at a maximum level for at least 72 hours.28 Thepresent immunocytochemical study showed that in-creased ADF occurs mainly in mitochondria. Semi-quantitative data supported the above results.
Because mitochondria are the center of the intra-cellular respiration system, a breakdown of their elec-tron-transfer cycle causes serious functional damageand leads to an accumulation of reactive oxygen inter-mediates.38"40 O2 and H2O2 are mainly generatedthrough electron transport inside the mitochondria,which is reported to be the major contribution to thetotal cellular formation of these molecules.'10 BecauseH2O2 diffuses rapidly through membranes,40 it is easilyreleased from the mitochondria and distributed in thesurrounding cytoplasm. Immunohistochemical studyof the H2O2-treated cultured RPE cells showed moreintensive ADF-positive labeling than the untreatedcontrol. This was further supported by the results ofan immunocytochemical study showing that immuno-gold particles were more prevalent in the mitochon-dria in the RPE cells treated with H2O2 than in un-treated ones. The cytoplasm was also labeled moder-ately, and most gold particles were seen adjacent tomitochondria, which suggested possible relocation of
FIGURE 4. The light micrographs of cultured human RPEcells. The cells treated with H2O2 (A) express more intensiveimmunostaining in the cytoplasm compared to the un-treated cells (B). Bar = 1 0 ^m.

Mitochondrial ADF Induction 2921
M
ER 1 ER
FIGURE 5. The immunoelectron micrographs of cultured human RPE cells. Bar = 0.2 jtm. (A)The cells treated with HyOa exhibit intensive ADF distribution in mitochondria. Moderateimmunolabeling is seen also in the surrounding cytoplasm. (B) The untreated cells show onlySlight labeling both in mitochondria and in cytoplasm. (C) The control micrograph shows nobackground labeling. M = mitochondria; N - nucleus; ER = endoplasmic reticulum.
the antigen. It is likely that, during both transient reti-nal ischemia and H2O2 exposure, the reactive oxygenintermediates were detoxified by ADF/hTx induced inmitochondria.
There are reports indicating other proteins
H2O2conc.(nM)
0 1 10 100rADF cone. (nQ/m\)
FIGURE 6. Effect of rADF on the toxicity of H2O2. RPE cellswere preincubated with rADF at 37°C for 48 hours, and thetoxicity of H2O2 on the treated and nontreated RPE wasexamined. The toxicity was estimated as follows: % OD re-duction = (T - S) X100/(T - B). S, absorbance in the pres-ence of H2O2; T, absorbance in the absence of H2OS; B,absorbance of medium alone. Each bar represents the mean± SD of four separate experiments. The significance of dif-ference of % OD reductions between the tested wells and thecontrol was determined by Student's Mest (P < 0.01) in allthree different concentrations of H2O2.
change their localization under various stresses. Mito-chondrial specific manganese-superoxide dismutase(Mn-SOD) is induced under various stresses, includingoxidative insult,41"47 whereas copper-zinc SOD usuallydoes not increase under these stresses,43 It is hypothe-sized that Mn-SOD induction is critical in the cellularantioxidant protection system, and mitochondrialO2-synthesis is the main factor of toxic intracellularoxidation.42'15 Genetic analysis has revealed that thepromotor sequences of ADF9-48 and human Mn-SOD4'1 genes show significant resemblance. Both lackTATA boxes and CAT boxes, are rich in GC, and pos-sess Spl binding sites. This may suggest that thiore-doxin, which detoxifies H2O2 produced from super-oxide by the function of SOD, and Mn-SOD behave ina similar manner under certain stressful conditions.
The transient retinal ischemic model in this studyis considered to be partially analogous to clinical isch-emic retinopathy. It is particularly important to notethat the retina consumes the largest amount of oxygenin the body and that outer segments of the photore-ceptor cells contain polyunsaturated fatty acids in highconcentrations that are particularly vulnerable to radi-cals and reactive oxygen.50"52 The retina is incessantlyexposed to light, including ultraviolet that is known toproduce oxidative stresses and degrade DNA. In der-matology, the natural defense of skin against light iswidely known. It is reasonable to assume that the ret-ina is also carefully armored by self-protecting systems

2922 Investigative Ophthalmology & Visual Science, June 1994, Vol. 35, No. 7
against oxidative stresses. Wilson observed in this reti-nal ischemic model using albino S-D rat that choroidalvascular bed also collapsed in the area of retinal vascu-lar thrombosis.30 We propose two distinct specula-tions. The highly differentiated photoreceptor cellsmay not possess enough of a self-protecting system,and RPE cells take over the role in response to certainsignals from photoreceptors. Another speculation isthat ADF induction may be part of a self-protectiveresponse of RPE cells.
Outer segments of retinal photoreceptor cellscontain retinol, which was recently reported to inducethioredoxin.53 Through the autophagocytosis of pho-toreceptor outer segments by RPE, retinoids are recy-cled in the physiological visual cycle of reduction andoxidation. Thioredoxin may participate in this processthrough redox regulation. On the other hand, thiore-doxin has been reported to reactivate some enzymesthat were inactivated through oxidation,36 suggestingthat ADF/hTX may form a general cycle in most tis-sues that reverses oxidative insults. The function andnatural protection of ADF/hTx have not been studiedextensively yet. More work is needed to comprehendthese subjects.
Key Words
ADF/hTx, retinal ischemia, retinal pigment epithelial cells,mitochondria, oxidative stress
References
1. Wakasugi N, Tagaya Y, Wakasugi H, et al. Adult T-cellleukemia-derived factor/thioredoxin, produced byboth human T-lymphotropic virus type 1- and Ep-stein-Barr virus-transformed lymphocytes, acts as anautocrine growth factor and synergizes with interleu-kin 1 and interleukin 2. Proc Natl Acad Sci USA.1990;87:8282-8286.
2. Wakasugi H, Rimsky L, Mahe Y, et al. Epstein-Barrvirus-containing B-cell line produces an interleukin 1that it uses as a growth factor. Proc Natl Acad Sci USA.1987;84:804-808.
3. Teshigawara K, Maeda M, Nishino K, et al. Adult Tleukemia cells produce a lymphokine that augumentsinterleukin 2 receptor expression. JMol Cell Immunol.1985;2:l7-26.
4. Yodoi J, Uchiyama T. IL-2 receptor dysfunction andadult T-cell leukemia. Immunol Rev. 1986;92:135-156.
5. Tagaya Y, Okada M, Sugie K, et al. IL-2 receptor/Tac-(p55) inducing factor: Purification and characteriza-tion of adult T cell leukemia-derived factor. J Im-munol. 1988;140:2614-2620.
6. Yodoi J, Tursz T. ADF, a growth-promoting factor de-rived from adult T cell leukemia and homologous tothioredoxin: Involvement in lymphocyte immortaliza-tion by HTLV-I and EBV. Adv Cancer Res.1991;57:381-411.
7. Tagaya Y, Maeda Y, Mitsui A, et al. ATL-derived fac-tor (ADF), An IL-2 receptor/Tac inducer homologousto thioredoxin: Possible involvement of dithiol-reduc-tion in the IL-2 receptor induction. EMBO J.1989;8:757-764.
8. Holmgren A. Thioredoxin: The amino acid sequencesof the protein from escherichia coli B. EurJ Biochem.1968;6:475-484.
9. Wollman E, d'Auriol E, Rimsky L, et al. Cloning andexpression of a cDNA for human thioredoxin. J BiolChem 1988;263:15506-15512.
10. Yodoi J, Uchiyama T. Diseases associated with HTLV-1 virus: IL-2 receptor dysregulation and redox regula-tion. Immunol Today. 1992;13:405-411.
11. Holmgren A. Thioredoxin catalyzes the reduction ofinsulin disulfides by dithiothreitol and dihydrolipo-amide./ Biol Chem. 1979;254:9627-9632.
12. Tagaya Y, Masutani H, Nakamura H, et al. Role ofATL-derived factor (ADF) in the normal and abnor-mal cellular activation: Involvement of dithiol relatedreduction. Mol Immunol. 1990;27:1279-1289.
13. Yamauchi A, Masutani H, Tagaya Y, et al. Lymphocytetransformation and thiol compounds: The role ofADF/thioredoxin as an endogenous reducing agent.Mol Immunol. 1992;29:263-270.
14. Mitsui A, Hirakawa T, Yodoi J. Reactive oxygen-re-ducing and protein-refolding activities of adult T cellleukemia derived factor/human thioredoxin. BiochemBiophysRes Commun. 1992;1220-1226.
15. Rozell B, Hansson H. Immunohistochemical localiza-tion of thioredoxin and thioredoxin reductase in adultrats. EurJ Cell Biol. 1985;38:79-86.
16. Kusama K, Saitoh T, Masutani H, Nakamura H, YodoiJ, Moro I. Adult T cell leukemia derived factor (ADF)in oral epithelial lesions. J Oral Pathol Med.1991;20:421-424.
17. Iwai T, Fujii S, Nanbu Y, et al. Expression of adultT-cell leukaemia-derived factor, a human thioredoxinhomologue, in the human ovary throughout the men-strual cycle. Virchows Arch A Pathol Anat Histopathol.1992;420:213-217.
18. Fujii S, Nambu Y, Nonogaki H, et al. Coexpression ofadult T-cell leukemia-derived factor, a human thiore-doxin homologue, and human papillomavirus DNA inneoplastic cervical squamous epithelium. Cancer.1991;68:1583-1591.
19. Nakamura H, Masutani H, Tagaya Y, et al. Expressionand growth-promoting effect of adult T-cell leukemia-derived factor: A human thioredoxin homologue inhepatocellular carcinoma. Cancer. 1992;69:2091-2097.
20. Wakita H, Yodoi J, Masutani H, Toda K, Takigawa M.Immunohistochemical distribution of adult T-cell leu-kemia-derived factor/thioredoxin in epithelial compo-nents of normal and pathologic human skin condi-tions. J Invest Dermatol. 1992;99:101-107.
21. Garcia J, Anderson M. Physiopathology of cerebralischemia. Crit Rev Neurobiol. 1989;4:303-324.
22. McCord J. Oxygen-derived free radicals in post-isch-emic tissue injury. N EnglJ Med. 1985;312:159-163.
23. Fukuchi T, Kobayashi A, Kaneko M, Ichiyama A, Ya-

Mitochondrial ADF Induction 2923
mazaki N. Possible involvement of free radicals andantioxidants in the early stages of the development ofcardiomyopathy in BIO 14.6 Syrian Hamster. JpnHeart J. 1991;32:655-666.
24. Rozga J. The role of reactive oxygen metabolites inthe patogenesis of ulcerative colitis: A speculative syn-thesis. Mater Med Pol. 1989;21:263-268.
25. Feberowski N, Stefansson E, Davidson R. Local hypo-thermia protects the retina from ischemia. Invest Oph-thalviol VisSci. 1989;30:2309-2313.
26. Szabo M, Droy-Lefaix M, Doly M, Carre C, BraquestP. Ischemia and reperfusion-induced histologicchanges in rat retina: Demonstration of a free radical-mediated mechanism. Invest Ophthalmol Vis Sci.1991;32:1471-1478.
27. Kagan V, Shvedova A, Novikov K, Kozliv Y. Light-in-duced free radical oxidation of membrane lipids inphotoreceptors of frog retina. Biochem Biophy Acta.1973;330:76-79.
28. Ohira A, Honda O, Gauntt CD, et al. Oxidative stressinduces adult T cell leukemia derived factor/thiore-doxin in the rat retina. Lab Invest. 1994;70:279-285.
29. Ohira A, de Juan EJ, Stefansson E, Faberowski N, Da-vidson R. Retinal ischemia and cell proliferation in therat: The role of soluble mitogens. Graefes Arch ClinExp Ophthalmol. 1990;228:195-199.
30. Wilson C, Hatchell D. Photodynamic retinal vascularthrombosis. Invest Ophthalmol Vis Sci. 1991;32:2357-2365.
31. Kigasawa K, Soushi S, Tanaka Y, Obazawa H. Mor-phorogic and chromosomal study of a human retinalpigment epithelial cell line. Jap j Ophthalmol.1994;38:10-15.
32. Tomimoto H, Akiguchi I, Wakita H, Kimura J, HoriK, Yodoi J. Astroglial expression of ATL-derived fac-tor, a human thioredoxin homologue, in the gerbilbrain after transient global ischemia. Brain Res.1993;625:l-8.
33. Tokuyasu K. Immunocytochemistry on ultrathin fro-zen section. HistochemJ. 1980;12:381-403.
34. Mosmann T. Rapid colorimetric assay for cellulargrowth and survival: Application to proliferation andcytotoxisity assays. J Immunol Methods. 1983;65:55-63.
35. Matsuda M, Masutani H, Nakamura H, et al. Protec-tive activity of adult T cell leukemia-derived factor(ADF) against tumor necrotic factor-dependent cyto-toxicity on U937 cells. J Immunol. 1991;147:3837-3841.
36. Spector A, Yan G, Huang R, McDermott M, GascoyneP, Pigiet V. The effect of H2O2 upon thioredoxin-enriched lens epithelial cells. J Biol Chem. 1988;263:4984-4990.
37. Dorey C, Khouri G, Syniuta L, Curran S, Weiter J.Superoxide production by porcine retinal pigment epi-thelium in vitro. Invest Ophthalmol Vis Sci. 1989;30:1047-1054.
38. Shlafer M, Myers CL, Adkins S. Mitochondrial hydro-
gen peroxide generation and activities of glutathioneperoxidase and superoxide dismutase following globalischemia. J Mol Cell Cardiol. 1987; 19:1195-1206.
39. Corbisier P, Raes M, Michiels C, et al. Respiratory ac-tivity of isolated rat liver mitochondria following invitro exposure to oxygen species: A threshold study.Mech Aging Dev. 1990;51:249-263.
40. Chance B, Sies H, Boveris A. Hydroperoxide metabo-lism in mammalian organs. Physiol Rev. 1979;59:527-605.
41. Shiki Y, Meyrick BO, Brigham KL, Burr IM. Endo-toxin increases superoxide dismutase in cultured bo-vine pulmonary endothelial cells. Am J Physiol.1987;252:C436-C440.
42. White JA, Scandalios JG. In vitro synthesis, importa-tion and processing of Mn-superoxide dismutase(SOD-3) into maize mitochondria. Biochem BiophysActa. 1987;926:16-25.
43. HolleyJA, Janssen YM, Mossman BT, Taatjes DJ. In-creased manganese superoxide dismutase protein intype II epithelial cells of rat lungs after inhalation ofcrocidolite asbestos or cristobalite silica. AmJ Pathol.1992:141:475-485.
44. Del VP, Shaffer JB. Regulation of antioxidant enzymeexpression in LPS-treated bovine retinal pigment epi-thelial and corneal endothelial cells. Curr Eye Res.1991;10:919-925.
45. Asoh K, Watanabe Y, Mizoguchi H, et al. Induction ofmanganese superoxide dismutase by tumor necrosisfactor in human breast cancer MCF-7 cell line and itsTNF-resistant variant. Biochem Biophys Res Commun.1989;162:794-801.
46. Summers RW, Maves BV, Reeves RD, Arjes LJ, Ober-ley LW. Irradiation increases superoxide dismutase inrat intestinal smooth muscle. Free Radic Biol Med.1989;6:261-270.
47. Warner BB, Burhans MS, Clark JC, Wispe JR. Tumornecrosis factor-alpha increases Mn-SOD expression:Protection against oxidant injury. Am J Physiol.1991;260:L296-L301.
48. Tonissen KF, Wells JR. Isolation and characterizationof human thioredoxin-encoding genes. Gene.1991;102:221-228.
49. Hartman JT. European Patent Application. NewYork: Bio-Technology Corp; 1988: No. 88104880.5.
50. Organisciak D, Wang H, Li Z, Tso M. The protectiveeffect of ascorbate in retinal light damage of rats. In-vest Ophthalmol Vis Sci. 1985;26:1580-1588.
51. Anderson R, Rapp L, Wiegand R. Lipid peroxidationand retinal degeneration. Curr Eye Res. 1984;3:223-227.
52. Fliesler S, Anderson R. Chemistry and metabolism oflipids in the vertebrate retina. Prog Lipid Res.1983:22:79-131.
53. An G, Wu R. Thioredoxin gene expression is tran-scriptionally up-regulated by retinol in monkey con-ducting airway epithelial cells. Biochem Biophys ResCommun. 1992; 183:170-175.





























