Embed Size (px)
Citation preview

C
Modeling auditory transducer dy
namicsBjorn Nadrowski and Martin C. GopfertDepartment of Cellular Neurobiology, BlumenbachInstitute, University of Gottingen, Max-Planck Institutefor Experimental Medicine, Gottingen, Germany
Correspondence to Bjorn Nadrowski, Department ofCellular Neurobiology, Blumenbach Institute, Universityof Gottingen, Max-Planck Institute for ExperimentalMedicine, Hermann-Rein-Str. 3, 37075 Gottingen,GermanyTel: +49 551 3899 401; fax: +49 551 3899 439;e-mail: [email protected]
Current Opinion in Otolaryngology & Head and
Neck Surgery 2009, 17:400–406
Purpose of review
This article reviews the literature on the modeling of auditory transducer dynamics.
Theoretical descriptions and computational models of transducer dynamics are
presented and discussed.
Recent findings
Since the introduction of the gating-spring model of hair cell mechanotransduction in
1983, theories of auditory transducer dynamics have been developed along with the
accumulation of electrophysiological and mechanical data. Recent findings suggest that
the auditory transduction apparatus might be very similar across vertebrates and
invertebrates, and that auditory transducer dynamics can shape the performance of
entire hearing organs.
Summary
The sense of hearing relies on a small number of transduction modules that convert
minute mechanical stimuli into electrical signals. Models have been proposed that
describe how this transduction works. These models may help to understand the
biophysics of mechanoelectrical signal transduction, the contribution of transducer
dynamics to auditory signal processing, and to link transducer function and genes.
Keywords
hearing, mechanotransduction, modeling
Curr Opin Otolaryngol Head Neck Surg 17:400–406� 2009 Wolters Kluwer Health | Lippincott Williams & Wilkins1068-9508
Introduction
In the last decade, our understanding of the molecular
mechanisms underlying the impressive performance of
the hearing system has made significant progress. The
introduction of the gating-spring model in 1983 [1] has
triggered the development of a number of theoretical and
computational models that describe transducer dynamics.
In this article, we will review these models and discuss
how transducer dynamics influences auditory system
performance.
The gating-spring model ofmechanotransductionIn the inner ear, specialized hair cells transduce minute
mechanical stimuli into electrical signals (for reviews, see
[2,3]). Deflection of the hair bundle, sitting on top of each
hair cell, opens ion channels that allow for the influx of
potassium and calcium into the cell body [4]. This causes
a depolarization of the cell membrane [5] that triggers
synaptic vesicle release [6], and finally action potentials
[7].
The gating-spring model of mechanotransduction posits
that the ion channels that mediate this transduction are
mechanically gated [1]: a hypothesized elastic element
opyright © Lippincott Williams & Wilkins. Unautho
1068-9508 � 2009 Wolters Kluwer Health | Lippincott Williams & Wilkins
(the gating spring) is directly connected to each channel,
so that the channel is more likely to be open if the tension
in the gating spring is increased. In the simplest case, the
ion channel can assume either an open or a closed
configuration (two-state model) [1,8], and the gating
spring is modeled as a Hookean spring. The transition
from the closed to the open configuration shortens the
gating spring by a certain distance (the gating swing)
[1,9].
In hair cells, the transducers can be found at the tips of
the stereocilia that make up the hair bundle [1,10]. Forces
applied to the tip of the bundle lead to the entire bundle
rotating about its base, creating a shearing motion
between adjacent stereocilia that stretches the gating
springs [9]. The tips of adjacent stereocilia are linked
together by fine filaments [11]. These tip links seem to be
required for funneling forces to the transduction channels
[9,11–14,15��], although they are most probably too stiff
for being the gating springs proper [16,17�].
Experimental evidence suggests that hair bundles move
as a unit, with the stereocilia never splaying apart [7,18–
20]. In accordance, forces applied to the tip of the hair
bundle are thought to be simultaneously distributed
among all gating springs (parallel force transmission)
[9]. In many models, the hair bundle has therefore been
rized reproduction of this article is prohibited.
DOI:10.1097/MOO.0b013e3283303443

C
Modeling auditory transducer dynamics Nadrowski and Gopfert 401
idealized as a two-stereocilia structure (Fig. 1 [21]) that
can be projected onto a one-dimensional system (Fig. 2).
In this picture, the relaxation of the gating springs upon
channel opening reduces the stiffness of the entire hair
bundle. The minimal stiffness occurs when the channels’
open probability is one-half. This nonlinear gating com-
pliance has been observed in the hair bundle’s mechanics
[9], providing strong support for the gating-spring model
(reviewed in [10,22,23]).
As many sensory systems, hair cell transducers display
adaptation to sustained stimuli [1,24]. This adaptation is
thought to be brought about by a set of adaptation motors,
most probably myosin-1c [25], that are connected to the
transduction channels [26]. The motors are supposed to
promote adaptation by regulating the tension in the gating
springs [26]. Early experimental evidence suggested that
these motors could be described by a linear force–velocity
relationship [27]. The force that acts on the motors is given
by the gating-spring tension. If the gating-spring tension is
opyright © Lippincott Williams & Wilkins. Unauth
Figure 1 The gating-spring model of hair cell transduction
The hair bundle’s response to step stimuli illustrates adaptation in the gati200 ms). (b) Evolution of the transducer current of the channels. (c) Sketcprobability curves. Deflecting the bundle by a certain step amplitude [e.g., 60 nthe step [point (ii) in (b)]. The relation between deflection and open probabilitcurve [left curve with the points (i) and (ii) indicating the open probabilities andand (b)]. When repeating this procedure at the end of a sustained deflection,by a certain distance (black arrow). Curves are calculated using the hair buoperate in monostable state. Incomplete adaption manifests itself in two wayarrows) is less than the bundle’s deflection; and the open probability in statiopermission from [10].
zero, the motors will crawl up the stereocilia with a certain
velocity. This velocity decreases linearly as the gating-
spring tension is increased. If the gating-spring tension is
larger than the stall force of the motors, the motors slip
down the stereocilia [26,27]. Transducer adaptation in
hair cells is incomplete [28], a phenomenon that can be
accounted for by complementing the gating-spring model
with an extent spring (Fig. 2) [28].
Many of the models for hair cell mechanotransduction
utilize the framework presented in Figs 1 and 2; a projec-
tion of the entire hair bundle geometry to one-dimensional
space, with linear elastic elements. The differences
between models are mainly restricted to the characteriz-
ation of the dynamics of the adaptation motors and the
characterization of the effects of calcium on the individual
components of the transduction apparatus (see below).
The adaptation mediated by the mechanical movements
of the adaptation motors is termed slow adaptation, as it
orized reproduction of this article is prohibited.
ng-spring model. (a) Deflection applied to the hair bundle’s tip (60 nm,h of an idealized hair bundle at different times. (d) Displacement-openm as in (a)] elicits a certain open probability immediately after the onset ofy for different step amplitudes defines the displacement-open probabilitydeflections immediately before and after the step of 60 nm depicted in (a)adaptation has shifted the resulting displacement-open probability curvendle model proposed in [21], with motor parameters altered in order tos: the horizontal shift of the displacement-open probability curves (blacknary state (iii) is larger than in the unperturbed bundle (i). Adapted with

C
402 Hearing science
Figure 2 Basic components of the gating spring model of
mechanotransduction
ESGSS
B
D
AMF
C
AF
The transduction module proper consists of a serial arrangement of a setof adaptation motors (AM), a mechanosensitive ion channel (C), and agating spring (GS). The adaptation motors can exert forces on the actinfilaments (AF) of the stereocilia, enabling them to actively crawl up thesefilaments (to the right), thereby increasing the tension in the gatingsprings. High tension in the gating springs increases the probability ofchannel opening, which relaxes the gating spring by a distance, thegating swing. Because the gating springs cannot transmit compressionforces [12], a slackening element (S) is needed. Incomplete adaptationis mediated by the extent spring (ES): large forces (F) lead to highertension in the extent spring, thereby increasing the open probability ofthe channels. If the tension in the gating springs is high, the adaptationmotors will slip (movement to the left). A given external force (F) isbalanced by the tension in the spring element corresponding to the bulkstiffness of the hair bundle, the tension in the gating spring and thetension in the extent spring. AF, actin filament; AM, adaptation motor; B,bulk stiffness of hair bundle; C, mechanosensitive ion channel; D,distance; ES, extent spring; F, force; GS, gating spring; S, slackeningelement.
occurs on timescales of dozens of milliseconds [26]. The
hair bundle also exhibits a phenomenon called fast adap-
tation, a fast calcium-mediated reclosure of the channels
following immediately upon opening of the channels that
operates on sub-millisecond timescales [29]. Recent evi-
dence also suggests that this fast adaptation requires
myosin-1c [30], but also see [31�].
Generic descriptions: Hopf bifurcation andstochastic resonanceIt has long been speculated that the cochlea is more than
just a passive frequency analyzer. Gold [32] concluded in
1948 that no passive liquid-bathed resonator could
achieve the high-frequency resolution that he had
measured in humans. He, therefore, proposed that some
kind of active element would act as a regenerating device,
effectively reducing the high-damping coefficients of the
liquid environment of the ear.
Hopf bifurcation
A number of experiments have shown that the cochlea is
endowed with an active amplifier (reviewed in [33]).
Features of this ‘cochlear amplifier’ [34] include active
amplification of weak stimuli [35], nonlinear compression
(the range of input amplitudes is compressed in a smaller
range of output amplitudes) [36,37], sharp frequency
selectivity [37,38], and spontaneous otoacoustic emis-
sions (ears do emit sound; discovered in 1977, see
opyright © Lippincott Williams & Wilkins. Unautho
[39]). One of the candidate sources of the cochlear
amplifier is the mechanical activity of hair bundles
[3,40,41], which display all of the above features [42–45].
All these four hallmarks of the cochlear amplifier are
generic properties of a dynamical system that is poised
close to a dynamic instability, called Hopf bifurcation [46].
A Hopf bifurcation occurs when a dynamical system passes
from a quiescent state to an oscillatory state due to a
variation of a control parameter. Close to this critical point,
any system undergoing this transition can be described by
using a generic equation (Hopf oscillator) and accordingly
displays generic properties that include the aforemen-
tioned four key characteristics [47,48]. It has, therefore,
been proposed that both the cochlea and the hair bundle
operate close to a Hopf bifurcation [21,47,49,50].
The basic gating-spring model does not allow for Hopf
bifurcations [21,50]; if, however, calcium feedback is
included, the gating-spring model can display such a
bifurcation and oscillatory states [21,49–51].
Stochastic resonance
Biological systems are noisy. In the case of transducer
dynamics, fluctuations arise from Brownian motion,
stochastic channel opening/closing (channel clatter), and
stochastic binding/unbinding of molecular motors. Add-
ing noise to a Hopf bifurcation results in a smeared-out
transition between quiescent and oscillatory state,
reducing the sensitivity of the system [52]. In certain
conditions, however, noise can also be beneficial for the
detection of small signals. The mechanism underlying
this beneficial effect is called stochastic resonance [53].
This mechanism is quite simple. Imagine a sensor that
can essentially be described by a potential energy land-
scape consisting of two wells separated by an energy
barrier. A weak stimulus might not suffice to allow for
the passage from one well to the other; adding noise of an
appropriate intensity will allow this passage, resulting in
an enhancement of the system’s response.
The stochastic transition between the open and closed
states of the transducer channel provides such a double-
well situation. Experiments on single hair cell have
provided evidence for stochastic resonance; an optimal
signal-to-noise ratio is obtained for physiological noise
intensities [54,55]. These experimental data could be
reproduced via stochastic simulations using the frame-
work of the gating-spring model [56]. In this analysis,
adding fluctuations stemming from both Brownian
motion and channel clatter was essential in order to
obtain a quantitative agreement between the simulations
and the experiments.
Like Hopf oscillators, systems that make use of stochastic
resonance can display phase locking to external stimuli
rized reproduction of this article is prohibited.

C
Modeling auditory transducer dynamics Nadrowski and Gopfert 403
[57] and small signal amplification [58]. Methods to
distinguish between amplification based on stochastic
resonance and Hopf oscillations have been proposed
[59]. According to theory, stochastic resonance can prin-
cipally occur if a system operates in a bistable regime [58].
Modeling suggests that sensory hair bundles can display
both bistable regimes and Hopf bifurcations [21,50],
suggesting that hair cells may utilize both mechanisms
for signal amplification.
Auditory transducer dynamics as the basis foractive hair bundle movementsThe simple gating-spring model described in the first
section does not allow for active oscillations [21,50,60��],
which are key characteristics of hair bundle dynamics
[45,51]. By supplementing the gating-spring model with
negative calcium feedback, spontaneous oscillations as
well as many other features of active hair bundle
dynamics can be explained.
Negative calcium feedback
Different mechanisms of calcium feedback have been
proposed. All these mechanisms assume that this feed-
back is negative, with calcium entering through open
channels initiating some process that recloses the
channels.
Three mechanisms of calcium feedback have been pro-
posed: kinetic reclosure, in which high calcium concen-
tration leads to a conformational change of the channel
that favors the closed configuration by altering the
kinetic transition rates between closed and open states
[31�,49,61–63,64�]; release mechanism, in which calcium
leads to conformational changes of one or more of the
elements in the force transmission line, ultimately
reducing gating-spring tension, thus leading to channel
reclosure [29,30,31�,51,65]; and motor feedback, in
which calcium changes the characteristics of the adapta-
tion motors, which then leads to altered rates of motor
crawling and slipping [10,21,31�,47,50,51,66,67�]. These
mechanisms are not mutually exclusive, and combi-
nations of them have been used [31�,51].
The first model that was proven to display a Hopf
bifurcation was based on the kinetic reclosure mechanism
[49]. This model necessitated two calcium binding sites
and six different channel states in order to allow for Hopf
bifurcations with physiological parameters; later models
based on kinetic reclosure used four [62,63] and even 10
states [31�,64�]. To explain fast adaptation, both the
kinetic reclosure mechanism [9,61] and the release mech-
anism [29,30] have been proposed. The two mechanisms
have different mechanical correlates; following a positive
deflection, the release mechanism predicts that the bun-
dle moves even further in the positive direction due to
opyright © Lippincott Williams & Wilkins. Unauth
the relaxation of the gating springs, before the channels
reclosure. By contrast, the kinetic reclosure mechanism
predicts a movement in the opposite direction because
of channel reclosure. Whether hair bundles use one of
these mechanisms, or even both of them, is unresolved
[30,61,66].
Adaptation motors
Many recent models still use adaptation motors with
linear force–velocity characteristics, but assume that
these characteristics (stall force and slope) are effected
by the calcium concentration at the motors’ location
[21,27,31�,50,51,66,67�]; increasing calcium thereby
always weakens the adaptation motors.
A more complex motor model that allows for self-sus-
tained oscillations of an ensemble of molecular motors
when coupling them to an elastic element [68] has been
proposed [47]. In this model, the internal calcium con-
centration corresponds to the control parameter of the
Hopf bifurcation. The result is a system that is tuned
automatically to the critical point, a self-tuned critical
oscillator [47].
Fluctuations and noise
Three main fluctuation sources can be distinguished in
hair bundle dynamics [21,47,50]: stochastic binding/
unbinding of molecular motors (motor noise); stochastic
opening/closing of the ion channels (channel clatter); and
Brownian motion, inevitably present in all systems at
finite temperature.
Whereas the first descriptions of hair bundle dynamics did
not explicitly include noise in their models [27–29,49], an
early study [8] made use of a property of channel clatter
(the variance of receptor current as a function of the
channels’ open probability) in order to estimate the num-
ber of channels per hair bundle. Measurements of the
variance of bundle movement, in turn, have been used to
characterize properties of the adaptation motors, indicating
that the motors’ stepping frequency declines with gating-
spring tension [69]. Experimental evidence for the ear’s
ability to detect subthreshold stimulus amplitudes (stimuli
that do not elicit increased firing rates) by phase locking
has been presented for auditory nerve fibers [70,71] and for
hair bundle movements [44]. In a model that includes
motor noise [47], this phase locking has been reproduced.
Because noise necessarily limits sensitivity, phase locking
might be an essential mechanism for low-level stimulus
detection.
From transducer dynamics to auditory systemperformanceActive transducer dynamics explains the hair bundle’s
ability to undergo a Hopf bifurcation. By coupling
orized reproduction of this article is prohibited.
C
404 Hearing science
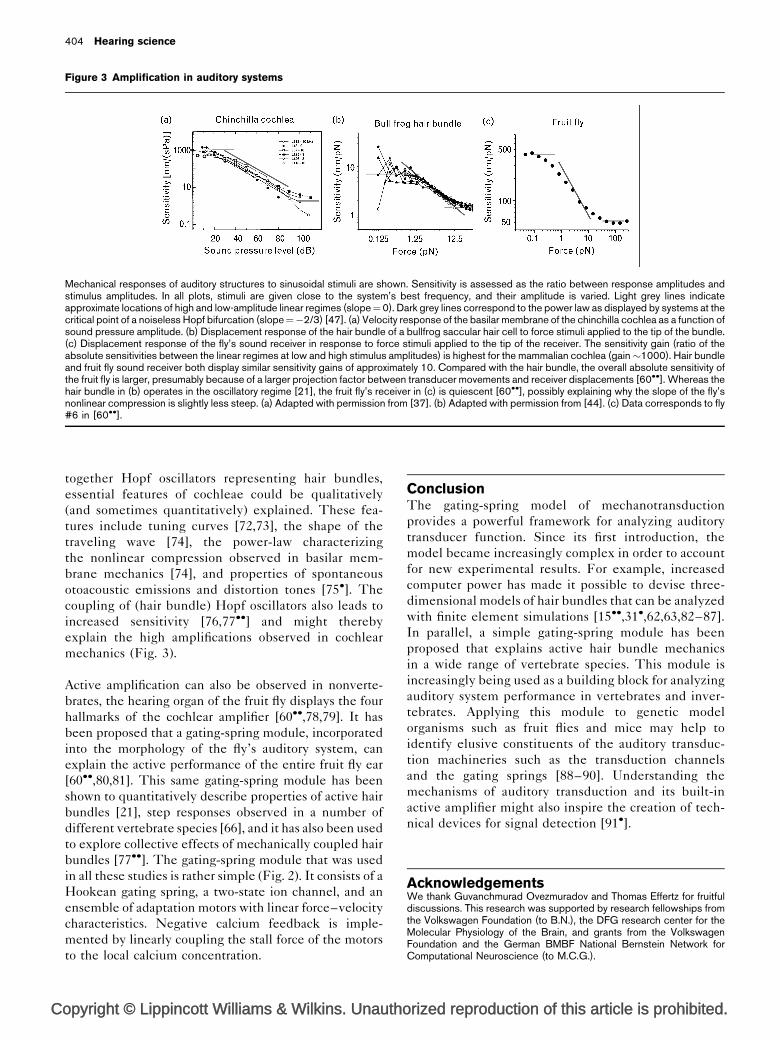
Figure 3 Amplification in auditory systems
Mechanical responses of auditory structures to sinusoidal stimuli are shown. Sensitivity is assessed as the ratio between response amplitudes andstimulus amplitudes. In all plots, stimuli are given close to the system’s best frequency, and their amplitude is varied. Light grey lines indicateapproximate locations of high and low-amplitude linear regimes (slope¼0). Dark grey lines correspond to the power law as displayed by systems at thecritical point of a noiseless Hopf bifurcation (slope¼�2/3) [47]. (a) Velocity response of the basilar membrane of the chinchilla cochlea as a function ofsound pressure amplitude. (b) Displacement response of the hair bundle of a bullfrog saccular hair cell to force stimuli applied to the tip of the bundle.(c) Displacement response of the fly’s sound receiver in response to force stimuli applied to the tip of the receiver. The sensitivity gain (ratio of theabsolute sensitivities between the linear regimes at low and high stimulus amplitudes) is highest for the mammalian cochlea (gain�1000). Hair bundleand fruit fly sound receiver both display similar sensitivity gains of approximately 10. Compared with the hair bundle, the overall absolute sensitivity ofthe fruit fly is larger, presumably because of a larger projection factor between transducer movements and receiver displacements [60��]. Whereas thehair bundle in (b) operates in the oscillatory regime [21], the fruit fly’s receiver in (c) is quiescent [60��], possibly explaining why the slope of the fly’snonlinear compression is slightly less steep. (a) Adapted with permission from [37]. (b) Adapted with permission from [44]. (c) Data corresponds to fly#6 in [60��].
together Hopf oscillators representing hair bundles,
essential features of cochleae could be qualitatively
(and sometimes quantitatively) explained. These fea-
tures include tuning curves [72,73], the shape of the
traveling wave [74], the power-law characterizing
the nonlinear compression observed in basilar mem-
brane mechanics [74], and properties of spontaneous
otoacoustic emissions and distortion tones [75�]. The
coupling of (hair bundle) Hopf oscillators also leads to
increased sensitivity [76,77��] and might thereby
explain the high amplifications observed in cochlear
mechanics (Fig. 3).
Active amplification can also be observed in nonverte-
brates, the hearing organ of the fruit fly displays the four
hallmarks of the cochlear amplifier [60��,78,79]. It has
been proposed that a gating-spring module, incorporated
into the morphology of the fly’s auditory system, can
explain the active performance of the entire fruit fly ear
[60��,80,81]. This same gating-spring module has been
shown to quantitatively describe properties of active hair
bundles [21], step responses observed in a number of
different vertebrate species [66], and it has also been used
to explore collective effects of mechanically coupled hair
bundles [77��]. The gating-spring module that was used
in all these studies is rather simple (Fig. 2). It consists of a
Hookean gating spring, a two-state ion channel, and an
ensemble of adaptation motors with linear force–velocity
characteristics. Negative calcium feedback is imple-
mented by linearly coupling the stall force of the motors
to the local calcium concentration.
opyright © Lippincott Williams & Wilkins. Unautho
ConclusionThe gating-spring model of mechanotransduction
provides a powerful framework for analyzing auditory
transducer function. Since its first introduction, the
model became increasingly complex in order to account
for new experimental results. For example, increased
computer power has made it possible to devise three-
dimensional models of hair bundles that can be analyzed
with finite element simulations [15��,31�,62,63,82–87].
In parallel, a simple gating-spring module has been
proposed that explains active hair bundle mechanics
in a wide range of vertebrate species. This module is
increasingly being used as a building block for analyzing
auditory system performance in vertebrates and inver-
tebrates. Applying this module to genetic model
organisms such as fruit flies and mice may help to
identify elusive constituents of the auditory transduc-
tion machineries such as the transduction channels
and the gating springs [88–90]. Understanding the
mechanisms of auditory transduction and its built-in
active amplifier might also inspire the creation of tech-
nical devices for signal detection [91�].
AcknowledgementsWe thank Guvanchmurad Ovezmuradov and Thomas Effertz for fruitfuldiscussions. This research was supported by research fellowships fromthe Volkswagen Foundation (to B.N.), the DFG research center for theMolecular Physiology of the Brain, and grants from the VolkswagenFoundation and the German BMBF National Bernstein Network forComputational Neuroscience (to M.C.G.).
rized reproduction of this article is prohibited.

C
Modeling auditory transducer dynamics Nadrowski and Gopfert 405
References and recommended readingPapers of particular interest, published within the annual period of review, havebeen highlighted as:� of special interest�� of outstanding interest
Additional references related to this topic can also be found in the CurrentWorld Literature section in this issue (p. 416).
1 Corey DP, Hudspeth AJ. Kinetics of the receptor current in bullfrog saccularhair-cells. J Neurosci 1983; 3:962–976.
2 Hudspeth AJ. How the ear’s works work. Nature 1989; 341:397–404.
3 Hudspeth AJ. Making an effort to listen: mechanical amplification in the ear.Neuron 2008; 59:530–545.
4 Lumpkin EA, Marquis RE, Hudspeth AJ. The selectivity of the hair cell’smechanoelectrical-transduction channel promotes Ca2þ flux at low Ca2þ
concentrations. Proc Natl Acad Sci U S A 1997; 94:10997–11002.
5 Corey DP, Hudspeth AJ. Ionic basis of the receptor potential in a vertebratehair cell. Nature 1979; 281:675–677.
6 Keen EC, Hudspeth AJ. Transfer characteristics of the hair cell’s afferentsynapse. Proc Natl Acad Sci U S A 2006; 103:5537–5542.
7 Hudspeth AJ, Corey DP. Sensitivity, polarity, and conductance change inresponse of vertebrate hair cells to controlled mechanical stimuli. Proc NatlAcad Sci U S A 1977; 74:2407–2411.
8 Holton T, Hudspeth AJ. The transduction channel of hair-cells from the bullfrogcharacterized by noise-analysis. J Physiol (Lond) 1986; 375:195–227.
9 Howard J, Hudspeth AJ. Compliance of the hair bundle associated with gatingof mechanoelectrical transduction channels in the bullfrog’s saccular hair cell.Neuron 1988; 1:189–199.
10 Hudspeth AJ, Gillespie PG. Pulling springs to tune transduction: adaptationby hair-cells. Neuron 1994; 12:1–9.
11 Pickles JO, Comis SD, Osborne MP. Cross-links between stereocilia in theguinea-pig organ of corti, and their possible relation to sensory transduction.Hear Res 1984; 15:103–112.
12 Kachar B, Parakkal M, Kurc M, et al. High-resolution structure of hair-cell tiplinks. Proc Natl Acad Sci U S A 2000; 97:13336–13341.
13 Assad JA, Shepherd GM, Corey DP. Tip-link integrity and mechanical trans-duction in vertebrate hair cells. Neuron 1991; 7:985–994.
14 Lumpkin EA, Hudspeth AJ. Detection of Ca2þ entry through mechanosensi-tive channels localizes the site of mechanoelectrical transduction in hair-cells.Proc Natl Acad Sci U S A 1995; 92:10297–10301.
15
��Beurg M, Fettiplace R, Nam J-H, et al. Localization of inner hair cell mechan-otransducer channels using high-speed calcium imaging. Nat Neurosci 2009;12:553–558.
This experimental and computational study observes calcium-induced fluores-cence changes in the stereocilia of rat cochlear inner hair cells following stepdeflections using fast confocal microscopy. Calcium signals are larger in rows withshorter stereocilia than in the row containing the tallest stereocilia. In contrast toprevious studies, this indicates that the mechanoelectrical transduction channelsare located at the lower ends (instead of at the upper ends or both ends) of the tiplinks.
16 Sotomayor M, Corey DP, Schulten K. In search of the hair-cell gating springelastic properties of ankyrin and cadherin repeats. Structure (Camb) 2005;13:669–682.
17
�Sotomayor M, Schulten K. The allosteric role of the Ca2þ switch in adhesionand elasticity of c-cadherin. Biophys J 2008; 94:4621–4633.
This study performs a molecular dynamics simulation of mechanical properties ofthe tip-link protein c-cadherin and their dependence on calcium. C-cadherinbehaves consistent with a viscoelastic element, but with very high stiffness(minimum 25 mN/m). This study supports a role for tip links in force transmission,but not as the elastic element forming the gating spring.
18 Crawford AC, Evans MG, Fettiplace R. Activation and adaptation of trans-ducer currents in turtle hair cells. J Physiol (Lond) 1989; 419:405–434.
19 Karavitaki KD, Corey DP. Hair bundle mechanics at high frequencies: a test ofseries or parallel transduction. In: Nuttall AL, Ren T, Gillespie P, et al., editors.Auditory mechanisms: processes and models. Singapore: World ScientificPublishing Co. Pte. Ltd; 2006. pp. 286–292.
20 Kozlov AS, Risler T, Hudspeth AJ. Coherent motion of stereocilia assures theconcerted gating of hair-cell transduction channels. Nat Neurosci 2007;10:87–92.
21 Nadrowski B, Martin P, Julicher F. Active hair-bundle motility harnesses noiseto operate near an optimum of mechanosensitivity. Proc Natl Acad Sci U S A2004; 101:12195–12200.
opyright © Lippincott Williams & Wilkins. Unauth
22 Roberts WM, Howard J, Hudspeth AJ. Hair-cells: transduction, tuning, andtransmission in the inner-ear. Annu Rev Cell Biol 1988; 4:63–92.
23 Phillips KR, Biswas A, Cyr JL. How hair cells hear: the molecular basis of hair-cell mechanotransduction. Curr Opin Otolaryngol Head Neck Surg 2008;16:445–451.
24 Eatock RA, Corey DP, Hudspeth AJ. Adaptation of mechanoelectrical trans-duction in hair cells of the bullfrog’s sacculus. J Neurosci 1987; 7:2821–2836.
25 Holt JR, Gillespie SKH, Provance DW, et al. A chemical-genetic strategyimplicates myosin-1c in adaptation by hair cells. Cell 2002; 108:371–381.
26 Howard J, Hudspeth AJ. Mechanical relaxation of the hair bundle mediatesadaptation in mechanoelectrical transduction by the bullfrogs saccular haircell. Proc Natl Acad Sci U S A 1987; 84:3064–3068.
27 Assad JA, Corey DP. An active motor model for adaptation by vertebrate hair-cells. J Neurosci 1992; 12:3291–3309.
28 Shepherd GMG, Corey DP. The extent of adaptation in bullfrog saccular hair-cells. J Neurosci 1994; 14:6217–6229.
29 Wu YC, Ricci AJ, Fettiplace R. Two components of transducer adaptation inauditory hair cells. J Neurophysiol 1999; 82:2171–2181.
30 Stauffer EA, Scarborough JD, Hirono M, et al. Fast adaptation in vestibular haircells requires myosin-1c activity. Neuron 2005; 47:541–553.
31
�Beurg M, Nam J-H, Crawford A, et al. The actions of calcium on hair bundlemechanics in mammalian cochlear hair cells. Biophys J 2008; 94:2639–2653.
An experimental and computational study reporting that the mechanical non-linearities observed in rat outer hair cells are strongly reduced in the presenceof constant low calcium concentrations. The article uses a finite element simulationof a three-dimensional hair bundle model with a kinetic reclosure scheme andproposes that myosins are not directly involved in force generation and nonlinearmechanics in the outer hair cell.
32 Gold T. Hearing 0.2. The physical basis of the action of the cochlea. Proc RSoc Lond Ser B Biol Sci 1948; 135:492–498.
33 Robles L, Ruggero MA. Mechanics of the mammalian cochlea. Physiol Rev2001; 81:1305–1352.
34 Davis H. An active process in cochlear mechanics. Hear Res 1983; 9:79–90.
35 Lukashkin AN, Walling MN, Russell IJ. Power amplification in the mammaliancochlea. Curr Biol 2007; 17:1340–1344.
36 Rhode WS. Observations of the vibration of the basilar membrane in squirrelmonkeys using the mossbauer technique. J Acoust Soc Am 1971; 49:1218–1231.
37 Ruggero MA, Rich NC, Recio A, et al. Basilar-membrane responses to tonesat the base of the chinchilla cochlea. J Acoust Soc Am 1997; 101:2151–2163.
38 Robles L, Ruggero MA, Rich NC. Basilar-membrane mechanics at the baseof the chinchilla cochlea 0.1. Input-output functions, tuning curves, andresponse phases. J Acoust Soc Am 1986; 80:1364–1374.
39 Kemp DT. Otoacoustic emissions: concepts and origins. In: Manley GA, FayRR, Popper AN, editors. Springer Handbook of auditory research. Activeprocesses and otoacoustic emissions in hearing. New York, USA: Springer;2008. pp. 1–38.
40 Manley GA, Gallo L. Otoacoustic emissions, hair cells, and myosin motors.J Acoust Soc Am 1997; 102:1049–1055.
41 Kennedy HJ, Crawford AC, Fettiplace R. Force generation by mammalian hairbundles supports a role in cochlear amplification. Nature 2005; 433:880–883.
42 Martin P, Hudspeth AJ. Active hair-bundle movements can amplify a hair cell’sresponse to oscillatory mechanical stimuli. Proc Natl Acad Sci U S A 1999;96:14306–14311.
43 Martin P, Hudspeth AJ, Julicher F. Comparison of a hair bundle’s spontaneousoscillations with its response to mechanical stimulation reveals the underlyingactive process. Proc Natl Acad Sci U S A 2001; 98:14380–14385.
44 Martin P, Hudspeth AJ. Compressive nonlinearity in the hair bundle’s activeresponse to mechanical stimulation. Proc Natl Acad Sci U S A 2001;98:14386–14391.
45 Crawford AC, Fettiplace R. The mechanical properties of ciliary bundles ofturtle cochlear hair cells. J Physiol (Lond) 1985; 364:359–379.
46 Strogatz SH. Nonlinear dynamics and chaos: Westview Press; 2001. pp.248–254.
47 Camalet S, Duke T, Julicher F, et al. Auditory sensitivity provided by self-tunedcritical oscillations of hair cells. Proc Natl Acad Sci U S A 2000; 97:3183–3188.
orized reproduction of this article is prohibited.

C
406 Hearing science
48 Eguiluz VM, Ospeck M, Choe Y, et al. Essential nonlinearities in hearing. PhysRev Lett 2000; 84:5232–5235.
49 Choe Y, Magnasco MO, Hudspeth AJ. A model for amplification of hair-bundlemotion by cyclical binding of Ca2þ to mechanoelectrical-transduction chan-nels. Proc Natl Acad Sci U S A 1998; 95:15321–15326.
50 Vilfan A, Duke T. Two adaptation processes in auditory hair cells together canprovide an active amplifier. Biophys J 2003; 85:191–203.
51 Martin P, Bozovic D, Choe Y, et al. Spontaneous oscillation by hair bundles ofthe bullfrog’s sacculus. J Neurosci 2003; 23:4533–4548.
52 Hoffmann KH. The Hopf-bifurcation of two-dimensional systems under theinfluence of one external noise source. Zeitschrift Fur Physik B-CondensedMatter 1982; 49:245–252.
53 Wiesenfeld K, Moss F. Stochastic resonance and the benefits of noise: fromice ages to crayfish and squids. Nature 1995; 373:33–36.
54 Jaramillo F, Wiesenfeld K. Mechanoelectrical transduction assisted byBrownian motion: a role for noise in the auditory system. Nat Neurosci1998; 1:384–388.
55 Jaramillo F, Wiesenfeld K. Physiological noise level enhances mechanoelec-trical transduction in hair cells. Chaos Solitons Fractals 2000; 11:1869–1874.
56 Lindner JF, Bennett M, Wiesenfeld K. Stochastic resonance in the mechan-oelectrical transduction of hair cells. Phys Rev E Stat Nonlin Soft Matter Phys2005; 72 (5 Pt 1):1–4.
57 Neiman A, Silchenko A, Anishchenko V, et al. Stochastic resonance: noise-enhanced phase coherence. Phys Rev E 1998; 58:7118–7125.
58 Jung P, Hanggi P. Amplification of small signals via stochastic resonance.Phys Rev A 1991; 44:8032–8042.
59 Balakrishnan J. Self-tuning to the hopf bifurcation in fluctuating systems.J Phys A Math Gen 2005; 38:1627–1652.
60
��Nadrowski B, Albert JT, Gopfert MC. Transducer-based force generationexplains active process in Drosophila hearing. Curr Biol 2008; 18:1365–1372.
This experimental, theoretical, and computational study presents a simple math-ematical model that explains the performance of the Drosophila ear. Adapting asimple gating-spring model to the morphology of the fly ear, a large number ofexperimental results are quantitatively reproduced.
61 Cheung ELM, Corey DP. Ca2þ changes the force sensitivity of the hair-celltransduction channel. Biophys J 2006; 90:124–139.
62 Nam J-H, Cotton JR, Grant W. A virtual hair cell, I: addition of gating springtheory into a 3-d bundle mechanical model. Biophys J 2007; 92:1918–1928.
63 Nam J-H, Cotton JR, Grant W. A virtual hair cell, Ii: evaluation of mechano-electric transduction parameters. Biophys J 2007; 92:1929–1937.
64
�Nam J-H, Fettiplace R. Theoretical conditions for high-frequency hair bundleoscillations in auditory hair cells. Biophys J 2008; 95:4948–4962.
A study performing finite-element simulations of a three-dimensional model of a hairbundle, using the kinetic reclosure scheme of negative feedback to generateoscillations. Oscillation frequencies mainly depend on the speed of calciumkinetics. Loading the bundle with an elastic element representing the tectorialmembrane sharpens the response.
65 Bozovic D, Hudspeth AJ. Hair-bundle movements elicited by transepithelialelectrical stimulation of hair cells in the sacculus of the bullfrog. Proc NatlAcad Sci U S A 2003; 100:958–963.
66 Tinevez J-Y, Julicher F, Martin P. Unifying the various incarnations of activehair-bundle motility by the vertebrate hair cell. Biophys J 2007; 93:4053–4067.
67
�Clausznitzer D, Lindner B, Julicher F, et al. Two-state approach to stochastichair bundle dynamics. Phys Rev E 2008; 77:41901–41913.
A theoretical and computational study that maps the dynamics of a simple gating-spring model to the Fitzhugh–Nagumo model of neural excitation. It is shown thatanalytic results for the power spectral density and response function can bederived when the system displays relaxation oscillations and therefore allows for agood separation of timescales, a situation that has been observed in oscillating hairbundles of the bullfrog.
68 Julicher F, Prost J. Spontaneous oscillations of collective molecular motors.Phys Rev Lett 1997; 78:4510–4513.
opyright © Lippincott Williams & Wilkins. Unautho
69 Frank JE, Markin V, Jaramillo F. Characterization of adaptation motors insaccular hair cells by fluctuation analysis. Biophys J 2002; 83:3188–3201.
70 Hillery CM, Narins PM. Neurophysiological evidence for a traveling wave in theamphibian inner-ear. Science 1984; 225:1037–1039.
71 Koppl C. Phase locking to high frequencies in the auditory nerve and cochlearnucleus magnocellularis of the barn owl, Tyto alba. J Neurosci 1997; 17:3312–3321.
72 Magnasco MO. A wave traveling over a hopf instability shapes the cochleartuning curve. Phys Rev Lett 2003; 90:058101.1–058101.4.
73 Stoop R, Vyver J, Kern A. Limit cycles, noise, and chaos in hearing. MicroscRes Tech 2004; 63:400–412.
74 Duke T, Julicher F. Active traveling wave in the cochlea. Phys Rev Lett 2003;90:158101.1–158101.4.
75
�Vilfan A, Duke T. Frequency clustering in spontaneous otoacoustic emissionsfrom a lizard’s ear. Biophys J 2008; 95:4622–4630.
This theoretical and computational study considers an ensemble of coupled Hopfoscillators that describe the bobtail lizard cochlea. It finds frequency and amplitudecharacteristics that are consistent with experimental results.
76 Risler T, Prost J, Julicher F. Universal critical behavior of noisy coupledoscillators: a renormalization group study. Phys Rev E Stat Nonlin Soft MatterPhys 2005; 72 (1 Pt 2):016130.
77
��Dierkes K, Lindner B, Julicher F. Enhancement of sensitivity gain and fre-quency tuning by coupling of active hair bundles. Proc Natl Acad Sci U S A2008; 105:18669–18674.
A theoretical and computational study that investigates the collective effects ofelastic coupling between hair bundles. It is found that the amplificatory gainincreases approximately 40-fold when 9�9 bundles are coupled together, sup-porting the idea that collective effects of coupled hair bundles might be animportant source of amplification in hearing.
78 Gopfert MC, Humphris ADL, Albert JT, et al. Power gain exhibited by motilemechanosensory neurons in Drosophila ears. Proc Natl Acad Sci U S A 2005;102:325–330.
79 Gopfert MC, Albert JT, Nadrowski B, et al. Specification of auditory sensitivityby Drosophila trp channels. Nat Neurosci 2006; 9:999–1000.
80 Albert JT, Nadrowski B, Gopfert MC. Mechanical signatures of transducergating in the Drosophila ear. Curr Biol 2007; 17:1000–1006.
81 Nadrowski B, Gopfert MC. Level-dependent auditory tuning: transducer-based active processes in hearing and best-frequency shifts. CommunicativeIntegr Biol 2009; 2:7–10.
82 Nam J-H, Cotton JR, Peterson EH, et al. Mechanical properties and con-sequences of stereocilia and extracellular links in vestibular hair bundles.Biophys J 2006; 90:2786–2795.
83 Silber J, Cotton J, Nam J-H, et al. Computational models of hair cell bundlemechanics: III. 3-d utricular bundles. Hear Res 2004; 197:112–130.
84 Cotton J, Grant W. Computational models of hair cell bundle mechanics: I.Single stereocilium. Hear Res 2004; 197:96–104.
85 Cotton J, Grant W. Computational models of hair cell bundle mechanics: II.Simplified bundle models. Hear Res 2004; 197:105–111.
86 Cotton JR, Grant JW. A finite element method for mechanical response of haircell ciliary bundles. J Biomech Eng 2000; 122:44–50.
87 Nam JH, Cotton JR, Grant JW. Effect of fluid forcing on vestibular hair bundles.J Vestib Res 2005; 15:263–278.
88 Gillespie PG, Dumont RA, Kachar B. Have we found the tip link, transductionchannel, and gating spring of the hair cell? Curr Opin Neurobiol 2005;15:389–396.
89 Albert JT, Nadrowski B, Gopfert MC. Drosophila mechanotransduction:linking proteins and functions. Fly 2007; 1:238–241.
90 Corey DP. What is the hair cell transduction channel? J Physiol 2006; 576(Pt 1):23–28.
91
�Ahn KH. A bio-inspired electromechanical system: artificial hair cell. ElectronTransport In Nanosystems, NATO Science for Peace and Security Series BPhysics and Biophysics; 2008. pp. 351–359.
A theoretical and computational study of a proposed electrical circuit mimickingthe hair bundle’s oscillatory mechanism in order to detect small amplitude forces ina noisy environment.
rized reproduction of this article is prohibited.





























