Embed Size (px)
Citation preview

Biochemical and Molecular Rules of Nutrients
Moderate Food Restriction Affects Skeletal Muscle and Liver GrowthHormone Receptors Differently in Pigs1–4
Sylvie Combes, Isabelle Louveau5 and Michel Bonneau
Institut National de la Recherche Agronomique, Station de Recherches Porcines, 35590 Saint Gilles, France
ABSTRACT The present study was conducted to determine the influence of food restriction on growth hormonereceptor (GHR) in porcine skeletal muscle (longissimus dorsi and trapezius) and liver in relationship to plasmagrowth hormone binding protein (GHBP). At 76 d of age, pigs were allotted to one of three groups. In GroupR100kg-196d, pigs were fed 70% of control food intake and slaughtered at 100 kg. Control pigs had free accessto food and were slaughtered either at the same weight (Group C100kg-161d) or at the same age (Group C130kg-196d) as R100kg-196d pigs. Plasma insulin-like growth factor-I concentrations tended to be lower in food-restrictedpigs than in control pigs at 40 kg (P õ 0.1) and 70 kg (Põ 0.05). At slaughter, there were no significant differencesamong the three groups. In liver, 125I-labeled bovine GH specific binding was higher (P õ 0.05) in food-restrictedpigs than in control pigs, and GHR mRNA level was higher (P õ 0.05) in food-restricted pigs than in C100kg-161d pigs. In trapezius, 125I-labeled bovine GH specific binding was lower (P õ 0.05) in food-restricted pigs thanin C130kg-196d pigs, and the level of GHR mRNA was higher (P õ 0.01) in food-restricted pigs than in controlpigs. The levels of GHR in longissimus dorsi were not affected by food restriction. The level of plasma GHBP waslower (P õ 0.05) in food-restricted pigs than in C130kg-196d pigs. These data indicate that nutritional statusregulates GHR in a tissue-specific manner and that there is no simple relationship between plasma GHBP andhepatic GHR in pigs. J. Nutr. 127: 1944–1949, 1997.
KEY WORDS: • growth • growth hormone receptor • food restriction • muscle • pigs
Growth hormone (GH)6 and insulin-like growth factor-I et al. 1993 and 1994). A 50–66% restriction of food intake(IGF-I) play a major role in growth regulation and direct sub- decreased hepatic GH binding in steers (Breier et al. 1988)strate availability toward lean body mass. In pigs, GH adminis- and GHR mRNA levels in young pigs (Dauncey et al. 1994).tration markedly increases muscle growth (Campbell et al. Similarly 48–72 h of food deprivation decreased both GH1989, Chung et al. 1985). The mechanisms involved in GH binding (Baxter et al. 1981, Maes et al. 1983) and GHRaction remain unclear. Because the first step in GH action is mRNA levels (Bornfeldt et al. 1989, Straus and Takemotobinding to a specific cell surface receptor, changes in GH 1990) in rat liver. The regulation of GHR has been studiedreceptor (GHR) levels may provide some insight into the im- more in liver than in other tissues and especially in skeletal
muscle, which is a potentially important target tissue for GH.portance of GH actions. Numerous studies indicate that nutri-Indeed, the presence of GH binding sites has been demon-tional status influences the GH/IGF-I axis by affecting hor-strated from the last third of gestation to the adult stage inmonal and/or receptor level. Food restriction and deprivationpig skeletal muscle (Schnoebelen-Combes et al. 1996). How-are known to decrease plasma IGF-I concentrations in severalever, it is not known whether the change in growth inducedspecies, including pigs (Buonomo and Baile 1991, Daunceyby food restriction is associated with change in GH bindinglevel in that tissue. To our knowledge, there is only a recent
1 Presented in part in the annual meeting of the American Society of Animal study that demonstrates that a 50% restriction of food intakeScience, July 24–26, 1996, Rapid City, SD [Combes, S., Louveau, I. & Bonneau increases the level of GHR mRNA in skeletal muscleM. (1996) Effect of feed restriction of GH receptor (GHR) in porcine skeletal muscle (Dauncey et al. 1994).and liver. J. Anim. Sci. 74 (suppl. 1) : 142 (abs.)].
2 S. Combes was supported by a grant from the Institut Technique du Porc, In addition to GHR in tissues, a plasma GH binding proteinParis, France. (GHBP) has been identified in several species, including pigs
3 This study was supported by a grant from the Institut National de la Recher- (Davis et al. 1992). It is believed that GHBP results from ache Agronomique (AIP ‘‘Controle de la Differenciation Tissulaire’’).proteolytic cleavage of GHR in pigs, as in humans and rabbit.4 The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked ‘‘advertisement’’ Serum GHBP measurement may be useful as a simple indicatorin accordance with 18 USC section 1734 solely to indicate this fact. of hepatic GHR status because liver is the major source of5 To whom correspondence should be addressed.
GHBP mRNA in rats (Tiong and Herington 1991). To our6 Abbreviations used: bGH, bovine growth hormone; C100kg-161d, controlpigs slaughtered at the same weight as food-restricted pigs; C130kg-196d, con- knowledge, only one study reports a positive correlation be-trol pigs slaughtered at the same age as food-restricted pigs; GH, growth hor- tween these two variables in food-restricted rats (Villares etmone; GHBP, growth hormone binding protein; GHR, growth hormone receptor, al. 1994). It remains to be determined whether plasma GHBPhGH; human growth hormone; IGF-I, insulin-like growth factor-I; LD, longissimusdorsi; R100kg-196d, food-restricted pigs; TR, trapezius. and hepatic GHR levels also vary in a parallel manner in pigs.
0022-3166/97 $3.00 q 1997 American Society for Nutritional Sciences.Manuscript received 7 April 1997. Initial review completed 16 May 1997. Revision accepted 30 June 1997.
1944
/ 4w2a$$0004 09-11-97 09:32:34 nutra LP: J Nut November
by guest on June 12, 2013jn.nutrition.org
Dow
nloaded from

1945GROWTH HORMONE RECEPTOR IN FOOD-RESTRICTED PIGS
TABLE 1
Animal characteristics and effect of a 30% restriction of food intake on growth performance of pigs1,2
SignificanceGroup
R100kg-196d vs. R100kg-196d vs.C130kg-196d C100kg-161d R100kg-196d C130kg-196d C100kg-161d
Age at start of the study, d 76.4 { 1.4 76.4 { 1.4 76.4 { 1.4 NS NSWeight at start of the study, kg 30.6 { 0.4 31.0 { 0.7 30.1 { 0.3 NS NSAge at slaughter, d 196.0 { 3.6 161.0 { 1.6 196.0 { 3.6 NS 0.001Weight at slaughter, kg 129.9 { 1.8 100.9 { 0.6 100.4 { 0.7 0.001 NSGrowth rate, kg/d 0.832 { 0.016 0.820 { 0.013 0.589 { 0.010 0.001 0.001Food intake, kg/d 2.46 { 0.05 2.21 { 0.05 1.68 { 0.02 0.001 0.001
1 Values are means { SEM, n Å 9. NS Å not significant (P ú 0.05).2 C100kg-161d and C130kg-196d, control pigs slaughtered at the same age as food-restricted pigs; R100kg-196d, food-restricted pigs.
frozen tissues were cut into small pieces and homogenized (1:5, wt/v)In all studies performed to date, extreme changes in nutri-in ice-cold 50 mmol/L Tris-HCl buffer (pH 7.4) containing 250tional status for a short time were investigated. The presentmmol/L sucrose, 1 mmol/L EDTA and 1 mmol/L phenylmethylsulfo-study was undertaken to examine the influence of 30% foodnyl fluoride, using a Polytron homogenizer. After centrifugation, theintake restriction on GHR levels in two skeletal muscles (onefinal pellet was resuspended in 50 mmol/L Tris-HCl buffer (pH 7.4),white and one red) compared with liver, considering GHR and the microsomal membrane preparations were stored at 0207CmRNA and also GH-binding sites. Plasma IGF-I and GHBP until binding studies were performed. Protein concentration was esti-
levels were also determined. A moderate intensity but long mated using the bicinchoninic acid assay (Pierce, Rockford, IL) withfood restriction was chosen because the growth rates obtained bovine serum albumin as a standard.are close to those observed in pig production. Growth hormone binding assays. All GH binding assays were
performed in triplicate as previously described (Schnoebelen-Combeset al. 1996). Briefly, microsomal membrane proteins (400 mg for liverMATERIALS AND METHODS and 500 mg/tube for muscle) were incubated with 125I-labeled bovineGH (bGH, 0.6 kBq/tube or 0.6 mg/L) in 25 mmol/L Tris-HCl (pHAnimals and sample collection. Pigs were reared in compliance7.4) containing 10 mmol/L CaCl2 , 5 g/L bovine serum albumin, 0.2with national regulations for the humane care and use of animals ing/L NaN3 (binding buffer). Nonspecific binding was determined byresearch (certificate of authorization to experiment on living animalsaddition of an excess of unlabeled bGH (250 ng/tube). After a 48-hno. 04793 delivered by the French Department of Agriculture to M.incubation at room temperature, 2 mL of ice-cold binding buffer wasBonneau). Nine litters of Large White1 Pietrain pigs from the INRAadded to stop the reaction. Bound hormone and free hormone wereherd were used (Table 1). At 76 d of age, three female pigs fromseparated by centrifugation at 4000 1 g for 20 min at 47C.each litter were allotted to one of three groups. All pigs had free
Growth hormone binding protein determination. The GHBPaccess to water and received the same commercial diet based onlevel was determined using HPLC gel filtration as previously describedwheat, barley, corn and soybean meal. The diet contained 12.4 MJ(Tar et al. 1990). After filtration through a 0.45-mm Millipore mini-digestible energy/kg, 17.5% crude protein, 0.95% lysine, 38.8% starchfilter, plasma (100 mL) was incubated overnight at 47C with 100 mLand 4.1% lipids. In Group R100kg-196d, pigs were fed 70% of controlof buffer [0.1 mol/L KH2PO4 (pH 7.0), 1 g/L bovine serum albumin]food intake and slaughtered at 100 kg. Control pigs had free accesscontaining 125I-labeled human GH (hGH Serono Laboratories, Ge-to food and were slaughtered either at the same weight (Groupnova, Switzerland) with a specific radioactivity of 3.7–5.6 MBq/mg.C100kg-161d) or at the same age (Group C130kg-196d) as R100kg-A parallel incubation was performed in the presence of an excess of196d pigs. Pigs were fed once a day. Blood samples were collected byunlabeled hGH (2 mg) to evaluate nonspecific binding. The entirejugular puncture at 40 and 70 kg. Pigs were killed by exsanguinationincubation mixture was placed onto a HPLC Protein Pak 300swafter electrical stunning. Blood and tissue sampling were performedcolumn (Waters, Milford, MA; 0.751 30 cm). Elution was performed20–24 h after the last meal. Blood, liver, longissimus dorsi (LD, ausing a degassed buffer (0.1 mol/L Na2SO4, 0.1 mol/L KH2PO4, pHwhite muscle) and trapezius (TR, a red muscle) samples were collected7.0). The binding of GH is expressed as the radioactivity in thewithin 15 min after death. Plasma and tissues for binding studies wereGHBP peak divided by the total radioactivity (GH and the GHBPstored at 0207C; tissues for mRNA analysis were frozen in liquidpeaks). The somatogenic specificity of hGH binding to plasma GHBPnitrogen and stored at 0707C until assayed.in pigs was previously demonstrated (Schnoebelen-Combes et al.Plasma insulin-like growth factor-I concentrations. Plasma IGF-1996).I concentrations were determined using a double antibody radioim-
RNA isolation and RNAse protection assays. Total RNA wasmunoassay after acid-ethanol extraction (Daughaday et al. 1980).isolated from tissues using the guanidium thiocyanate methodThe assay was performed using recombinant IGF-I (human recombi-(Chomczynski and Sacchi 1987). Quantity and quality of isolatednant IGF-I, receptor quality, Mallinckrodt, St Louis, MO) as a tracertotal RNA were evaluated spectrophotometrically and confirmed withwith a specific radio-activity of 5.3 MBq/mg and as a standard. Ahorizontal gel electrophoresis. The abundance of GHR mRNA waspolyclonal antibody raised in rabbit (Claus et al. 1992) was used atquantified using a sensitive solution hybridization-RNAse protectiona final dilution of 1:40,000. All the samples were analyzed within aassay. Porcine GHR cDNA in pGEM 3Zf- (Promega, Madison, WI)single assay. The intraassay CV for a plasma sample containing 5.4was kindly provided by Terry D. Etherton (Penn State University,and 63.6 nmol/L of IGF-I were 8.8 and 12.9%, respectively. As recom-University Park, PA). To generate antisense riboprobe, the plasmidmended by Bang et al. (1995), a validation of the assay was performedwas linearized by digestion with Hind II and transcribed with Promega(Louveau and Bonneau 1996). The data indicated that althoughkit using T7 polymerase (Promega) in the presence of [a32P]CTP (Duacid-ethanol extraction lead to IGF-I levels that differed from thosePont de Nemours, Les Ulis, France). The RNA probe consisted ofobtained for glycyl-glycine/G-50 extracted plasma, there was a close375 nucleotides, of which 368 were the complementary sequencerelationship (r Å 0.97, P õ 0.001) between the values obtained by/654 to /1021 of the GHR. To check for possible differences inthese two methods.quantification and/or loading, samples were also assayed for 18S RNAMembrane preparation. Microsomal membranes were prepared
as previously described (Meserole and Etherton 1984). In brief, using human 18S cDNA (pT7 RNA 18S) obtained from Ambion
/ 4w2a$$0004 09-11-97 09:32:34 nutra LP: J Nut November
by guest on June 12, 2013jn.nutrition.org
Dow
nloaded from

1946 COMBES ET AL.
(Austin, TX). To obtain a low specific activity probe, the linearizedplasmid was transcribed with RiboMax large scale system kit usingT7 polymerase (Promega). The 18S RNA probe consisted of 116nucleotides and protected an RNA doublet of 80 and 70 bp.
Solution hybridization-RNAse protection assays were performedin triplicate on total RNA. Samples from the three pigs within eachlitter were always analyzed on the same gel. Assays were conductedas follows: 20 mg of total RNA was incubated with 32P-GHR RNAprobe (7 kBq) and 32P-18S RNA probe (0.4 kBq, 1 mg) overnight at457C in 29 mL of hybridization buffer [20 mmol/L Tris (pH 7.5), 1mmol/L EDTA, 0.4 mmol/L NaCl, 0.1% SDS, 75% formamide]. Theunhybridized strands were digested with ribonucleases A and T1(RNAse Cocktail, Ambion) for 45 min at 377C. Digestion by RNAseswas followed by an incubation with Proteinase K (Eurobio, Les Ulis,France), phenol-chloroform extraction, and ethanol precipitation ofhybridized RNA. Samples and 32P-DNA size markers (pGEM DNAMarker, Promega) were then resuspended in gel dye and separatedby size on 6% polyacrylamide/6 mol/L urea gel. Radioactive bandswere visualized by autoradiography of dried gels using Kodak X-OMAT AR film and double intensifying screens for 1–3 d at 0707C.Unhybridized probe digested or undigested by RNAses was used ascontrol. The relative intensities of the protected bands were quanti-fied using Densylab software (Microvision Instruments, Evry, France).The data were not normalized to the abundance of 18S RNA. How-ever, when the 18S RNA was barely detectable because of loss ofthe sample, the value of GHR was not considered.
Statistical analysis. Data were subjected to ANOVA using theGeneral Linear Model procedure of SAS (1989). The model includedthe main effects of restriction and litter. A further analysis was per-formed, using the contrast statement of the General Linear Modelprocedure, to compare means for the C130kg-196d or the C100kg-161d group to those for the R100kg-196d group.
RESULTS
FIGURE 1 Growth curves of control (C130kg-196d, C100kg-Growth rates, age and weight at slaughter. Growth curves161d) and food-restricted (R100kg-196d) pigs during the experimentaland growth performance of control (C100kg-161d andperiod. Values are means, n Å 9. Bars for SEM were smaller than theC130kg-196d) and food-restricted (R100kg-196d) pigs are pre-symbols.sented in Figure 1 and Table 1, respectively. The achieved
overall levels of restriction were 24 and 32% comparativelyto the C100kg-161d and C130kg-196d groups, respectively. 161d and C130kg-196d) (Fig. 6). In LD, there was a wideGrowth rates were 28 and 29% lower (P õ 0.001) in food- individual variation, and GHR mRNA level did not differrestricted (Group R100kg-196d) pigs than in Groups C100kg- significantly among the three groups (Fig. 6).161d and C130kg-196d, respectively.
Plasma insulin-like growth factor-I concentrations. Plasma DISCUSSIONIGF-I concentrations (Fig. 2) were lower in food-restrictedpigs (R100kg-196d) than in control pigs (C100kg-161d and The present study is, to our knowledge, the first to assess
the influence of a moderate food restriction on GHR, consider-C130kg-196d) at 40 kg (P õ 0.1) and 70 kg (P õ 0.05). Atslaughter, there were no significant differences among the ing both GHR mRNA and GH binding levels in skeletal
muscle as compared with liver. This study also investigatedthree groups.Growth factor binding in liver, skeletal muscle and plas- the possibility of a co-regulation of GHBP and GHR. In the
present work, pigs were subjected to a 24–32% restriction ofma. In liver, 125I-labeled bGH specific binding was higher (Põ 0.05) in restricted pigs (R100kg-196d) than in control pigs food intake, which resulted in a 28–29% decrease in growth
rate. The finding of a lower plasma IGF-I concentration in(C100kg-161d and C130kg-196d) (Fig. 3). In TR, 125I-labeledbGH specific binding was lower (P õ 0.05) in food-restricted food-restricted pigs compared with controls is in accordance
with previous studies conducted in pigs and other species (Buo-pigs (R100kg-196d) than in C130kg-196d pigs (Fig. 3). Theeffect of food restriction on 125I-labeled bGH specific binding nomo and Baile 1991, Dauncey et al. 1993 and 1994). How-
ever, differences in plasma IGF-I concentrations between food-was not significant in LD (Fig. 3). 125I-labeled hGH bindingto plasma GHBP was higher (P õ 0.05) in C130kg-196d pigs restricted and control pigs were low. At slaughter, the lack of
a significant difference among the groups may be related tothan in R100kg-196d or C100kg-161d pigs (Fig. 4). No differ-ence was observed between C100kg-161d and R100kg-196d a greater variability among pigs. The observation that food
restriction induced an elevation in both GH binding and GHRpigs.Expression of growth hormone receptor mRNA in liver mRNA levels in liver is not consistent with data reported
previously. A reduction of GH binding in liver has been ob-and skeletal muscle. Growth hormone receptor mRNA ex-pression was assessed by RNAse protection assay (Fig. 5). In served in rats (Villares et al. 1994) and cattle (Breier et al.
1988). It has also been reported that food restriction decreasedliver, the level of GHR mRNA was higher (P õ 0.05) inrestricted pigs (R100kg-196d) than in C100kg-161d pigs (Fig. GHR mRNA levels in the liver of pigs (Dauncey et al. 1994)
but had no effect on GHR mRNA levels in the liver of rats6). In TR, GHR mRNA level was higher (P õ 0.01) in re-stricted pigs (R100kg-196d) than in control pigs (C100kg- (Villares et al. 1994). In the present study, food restriction
/ 4w2a$$0004 09-11-97 09:32:34 nutra LP: J Nut November
by guest on June 12, 2013jn.nutrition.org
Dow
nloaded from

1947GROWTH HORMONE RECEPTOR IN FOOD-RESTRICTED PIGS
volving development (Breier et al. 1989), GH administration(Chung and Etherton 1986), 50% restriction of food intake(Dauncey et al. 1994) in pigs or food deprivation in rats (Maeset al. 1983), hepatic GHR level and plasma IGF-I concentra-tion did not vary in a parallel manner in the current study.This lack of parallelism is consistent with the hypothesis of apost-receptor defect as suggested during protein restriction inrats (Fliesen et al. 1989, Maiter et al. 1989, Thissen et al.1990).
The parallel study of GHR levels in two skeletal musclescomparatively to liver indicates that food restriction affectedGHR differently in liver and skeletal muscle. This is in accor-dance with a previous study in pigs (Dauncey et al. 1994).Our results also demonstrate that the tissue specificity is ex-pressed at the GH binding level. Indeed, although a clearincrease in GH binding level was observed in liver, there waseither no change or a slight reduction in GH binding in skele-tal muscle of food-restricted pigs. Our results also suggest thatLD and TR muscles may not respond similarly to food restric-tion. The differences are too small to draw any conclusions.Therefore, further studies are needed to determine whetherthe effect of food restriction on GHR is muscle specific.
There is little information regarding the influence of foodrestriction on GHBP levels. In the present study, plasmaGHBP levels were slightly lower in food-restricted than incontrol pigs of the same age, but there was no significantdifference between animals of the same weight. The small but
FIGURE 2 Plasma insulin-like growth factor-I (IGF-I) concentra-tion in control (C130kg-196d, C100kg-161d) and food-restricted(R100kg-196d) pigs. Values are means { SEM, n Å 9 or n Å 18 forcontrol pigs at 40 and 70 kg. Means are significantly different, *P õ0.05.
induced a slight reduction in GH binding associated with aslight increase in GHR mRNA level in TR muscle. Foodrestriction had no significant effect on GH binding and GHRmRNA levels in LD muscle, whereas an increase in the GHRmRNA content in that tissue was observed previously(Dauncey et al. 1994). To our knowledge, there are no otheravailable data in skeletal muscle.
The discrepancies between our study and previous findingscould be related to the extent of food restriction and/or theage of the pigs. First, compared with the other studies, the useof a less severe (24–32% vs. 43–66%) but longer (119 vs.10–40 d) food restriction might have allowed a progressivemetabolic adaptation to food restriction, which could eventu-ally have resulted in an elevation in hepatic GHR level. Sec-ond, in the present study, pigs were older (161 and 196 d ofage) than the 49-d-old pigs used by Dauncey et al. (1994).The importance of age in IGF-I and GHR regulation has beensuggested in other situations. It has been observed that GH-induced increase in GH binding to liver is much higher in30-d-old pigs (81%) than in 120-d-old pigs (15%) (Ambler etal. 1992). Similarly, it has been shown that the effect of adietary protein restriction on plasma IGF-I and hepatic GHRis age-dependent in rats (Fliesen et al. 1989). Protein restric-tion in 3-wk-old rats induced a decrease in plasma IGF-I (66%)and in hepatic GH binding. These effects were progressively
FIGURE 3 Growth hormone (GH) specific binding to liver, longissi-attenuated with increasing age. At 12 wk of age, there was amus dorsi (LD) and trapezius (TR) in control (C130kg-196d, C100kg-slight reduction in plasma IGF-I (17%) and a slight increase 161d) and food-restricted (R100kg-196d) pigs. Microsomal membranes
in hepatic GH binding. Our findings are in accordance with were incubated with 125I-bGH in the absence or the presence of anthese latter observations and support the hypothesis that the excess of unlabeled bGH (250 ng) for 48 h at room temperature asinfluence of food restriction on the GH/IGF-I axis is also age described in Materials and Methods. Values are means { SEM, n Å 9.
Means are significantly different, *P õ 0.05 and **P õ 0.01.dependent. In contrast to results for several other studies in-
/ 4w2a$$0004 09-11-97 09:32:34 nutra LP: J Nut November
by guest on June 12, 2013jn.nutrition.org
Dow
nloaded from
1948 COMBES ET AL.
ACKNOWLEDGMENTS
We wish to thank F. Giovanni, J. Portanguen and L. Sofer fortheir technical assistance. We are grateful to M. Douaire for letting ususe Densylab software (Laboratoire de Genetique Animale, ENSAR,Rennes, France). Bovine GH was a gift from USDA Animal HormoneProgram (Belstville, MD). The antiserum against IGF-I was kindlyprovided by U. Weiler (University of Hohenheim, Stuttgart, Ger-many). Special appreciation is extended to M-C Postel-Vinay (IN-SERM U344, Hopital Necker, Paris, France) for her helpful com-ments on the manuscript.
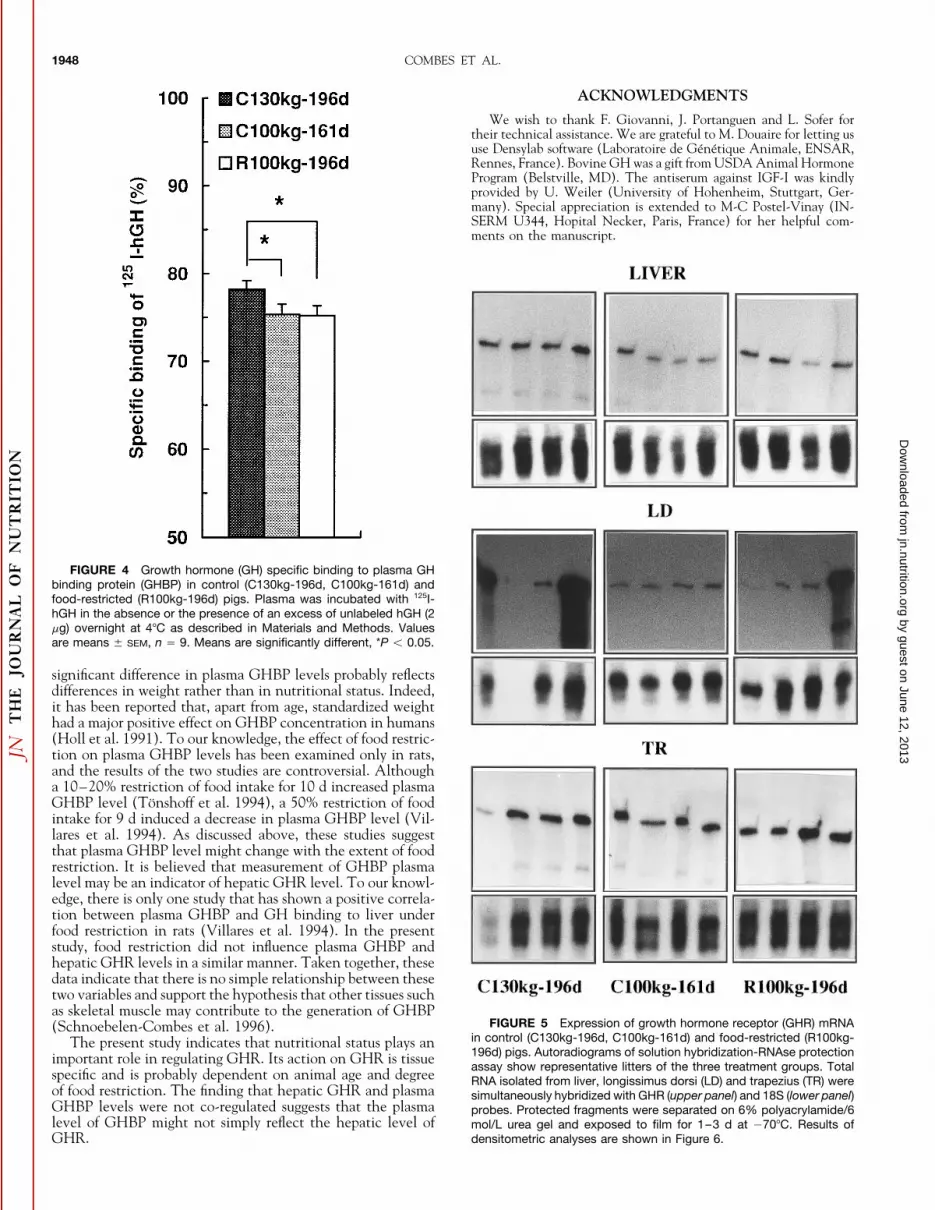
FIGURE 4 Growth hormone (GH) specific binding to plasma GHbinding protein (GHBP) in control (C130kg-196d, C100kg-161d) andfood-restricted (R100kg-196d) pigs. Plasma was incubated with 125I-hGH in the absence or the presence of an excess of unlabeled hGH (2mg) overnight at 47C as described in Materials and Methods. Valuesare means { SEM, n Å 9. Means are significantly different, *P õ 0.05.
significant difference in plasma GHBP levels probably reflectsdifferences in weight rather than in nutritional status. Indeed,it has been reported that, apart from age, standardized weighthad a major positive effect on GHBP concentration in humans(Holl et al. 1991). To our knowledge, the effect of food restric-tion on plasma GHBP levels has been examined only in rats,and the results of the two studies are controversial. Althougha 10–20% restriction of food intake for 10 d increased plasmaGHBP level (Tonshoff et al. 1994), a 50% restriction of foodintake for 9 d induced a decrease in plasma GHBP level (Vil-lares et al. 1994). As discussed above, these studies suggestthat plasma GHBP level might change with the extent of foodrestriction. It is believed that measurement of GHBP plasmalevel may be an indicator of hepatic GHR level. To our knowl-edge, there is only one study that has shown a positive correla-tion between plasma GHBP and GH binding to liver underfood restriction in rats (Villares et al. 1994). In the presentstudy, food restriction did not influence plasma GHBP andhepatic GHR levels in a similar manner. Taken together, thesedata indicate that there is no simple relationship between thesetwo variables and support the hypothesis that other tissues suchas skeletal muscle may contribute to the generation of GHBP
FIGURE 5 Expression of growth hormone receptor (GHR) mRNA(Schnoebelen-Combes et al. 1996).in control (C130kg-196d, C100kg-161d) and food-restricted (R100kg-The present study indicates that nutritional status plays an196d) pigs. Autoradiograms of solution hybridization-RNAse protectionimportant role in regulating GHR. Its action on GHR is tissueassay show representative litters of the three treatment groups. Totalspecific and is probably dependent on animal age and degree RNA isolated from liver, longissimus dorsi (LD) and trapezius (TR) were
of food restriction. The finding that hepatic GHR and plasma simultaneously hybridized with GHR (upper panel) and 18S (lower panel)GHBP levels were not co-regulated suggests that the plasma probes. Protected fragments were separated on 6% polyacrylamide/6level of GHBP might not simply reflect the hepatic level of mol/L urea gel and exposed to film for 1–3 d at 0707C. Results of
densitometric analyses are shown in Figure 6.GHR.
/ 4w2a$$0004 09-11-97 09:32:34 nutra LP: J Nut November
by guest on June 12, 2013jn.nutrition.org
Dow
nloaded from

1949GROWTH HORMONE RECEPTOR IN FOOD-RESTRICTED PIGS
insulin-like growth factor I, somatotropin, and metabolic hormone in swine.J. Anim. Sci. 69: 755–760.
Campbell, R. G., Steele, N. C., Caperna, T. J., McMurtry, J. P., Solomon, M. B. &Mitchell, A. D. (1989) Interrelationships between sex and exogenousgrowth hormone administration on performance, body composition and pro-tein accretion of growing pigs. J. Anim. Sci. 67: 177–186.
Chomczynski, P. & Sacchi, N. (1987) Single step method of RNA isolationby acid guanidium thiocyanate-phenol-chloroform extraction. Anal. Biochem.162: 156–159.
Chung, C. S., Etherton, T. D. & Wiggins, J. P. (1985) Stimulation of swinegrowth by porcine growth hormone. J. Anim. Sci. 60: 118–130.
Chung, S. C. & Etherton, T. D. (1986) Characterization of porcine growth hor-mone (pGH) binding to porcine liver microsomes: chronic administration ofpGH induces pGH binding. Endocrinology 119: 780–786.
Claus, R., Weiler, U., Hofacker, S. & Meng, H. (1992) Cycle dependent changesof growth hormone (GH), insulin-like growth factor (IGF-I) and insulin in bloodplasma of sows and their relation to progesterone and oestradiol. GrowthRegul. 2: 116–121.
Daughaday, W. H., Mariz, I. K. & Blethen, S. L. (1980) Inhibition of access ofbound somatomedin to membrane receptor and immunobinding sites: a com-parison of radioreceptor and radioimmunoassay of somatomedin in nativeand acid-ethanol–extracted serum. J. Clin. Endocrinol. Metab. 51: 781–788.
Dauncey, M. J., Burton, K. A., White, P., Harrison, A. P., Gilmour, R. S., Duchamp,C. & Cattaneo, D. (1994) Nutritional regulation of growth hormone receptorgene expression. FASEB J. 8: 81–88.
Dauncey, M. J., Rudd, B. T., White, D. A. & Shakespear, R. A. (1993) Regula-tion of insulin-like growth factor binding proteins in young growing animalsby alteration of energy status. Growth Regul. 3: 198–207.
Davis, S. L., Graf, M., Morisson, C. A., Hall, T. R. & Swift, P. J. (1992) Identifica-tion and partial purification of serum growth hormone binding protein in do-mestic animal species. J. Anim. Sci. 70: 773–780.
Fliesen, T., Maiter, D., Gerard, G., Underwood, L. E., Maes, M. & Ketelsleger, J. M.(1989) Reduction of serum insulin-like growth factor-I by dietary restriction isage dependent. Pediatr. Res. 26: 415–419.
Holl, R. W., Snehotta, R., Siegler, B., Scherbaum, W. & Heinze, E. (1991) Bind-ing protein for human growth hormone: effects of age and weight. Horm. Res.35: 190–197.
Louveau, I. & Bonneau, M. (1996) Effect of a growth hormone infusion onplasma insulin-like growth factor-I in Meishan and Large White pigs. Reprod.Nutr. Dev. 36: 301–310.
Maes, M., Underwood, L. E. & Ketelslegers, J. M. (1983) Plasma soma-tomedin-C in fasted and refed rats: close relationship with changes in liverFIGURE 6 Abundance of growth hormone receptor (GHR) mRNAsomatogenic but not lactogenic binding sites. J. Endocrinol. 97: 243–252.per 20 mg total RNA isolated from liver, longissimus dorsi (LD) and
Maiter, D., Fliesen, T., Underwood, L. E., Maes, M., Gerard, G., Davenport, M. L. &trapezius (TR) in control (C130kg-196d, C100kg-161d) and food-re-Ketelslegers, J. M. (1989) Dietary protein restriction decreases insulin-likestricted (R100kg-196d) pigs. GHR mRNA was quantified using solution growth factor-I independent of insulin and liver growth hormone binding.
hybridization-RNAse protection assay as described in Materials and Endocrinology 124: 2604–2611.Methods. Representative autoradiograms are shown in Figure 5. Values Meserole, V. K. & Etherton, T. D. (1984) Insulin binding to liver microsomes
from lean and obese swine during growth to market weight. J. Anim. Sci. 59:are means { SEM, n Å 9. Means are significantly different, *P õ 0.05650–657.and **P õ 0.01.
SAS Institute Inc. (1989) SAS/STATt User’s Guide, version 6, 4th ed., vol. 2. SASInstitute, Cary, NC.
Schnoebelen-Combes, S., Louveau, I., Postel-Vinay, M.-C. & Bonneau, M.LITERATURE CITED (1996) Ontogeny of GH receptor and GH-binding protein in the pig. J. Endo-crinol. 148: 249–255.Ambler, G. R., Breier, B. H., Blair, A.S.H., McCutcheon, S. N., Silbergeld, A. &
Straus, D. S. & Takemoto, C. D. (1990) Effect of fasting on insulin-like growthGluckman, P. D. (1992) The interrelationship between and the regulationfactor I (IGF-I) and growth hormone receptor mRNA levels and IGF-I geneof hepatic growth hormone receptors and circulating GH binding protein intranscription in rat liver. Mol. Endocrinol. 4: 91–100.the pig. Acta Endocrinol. 126: 155–161.
Tar, A., Hocquette, J.-F., Souberbielle, J. C., Clot, J. P., Brauner, R. & Postel-Bang, P., Baxter, R. C., Blum, W. F., Breier, B. H., Clemmons, D. R., Hall, K.,Vinay, M.-C. (1990) Evaluation of growth hormone binding proteins in hu-Hintz, R. L., Holly, J.M.P., Rosenfeld, R. G. & Zapf, J. (1995) Valid mea-man plasma using high pressure liquid chromatography gel filtration. J. Clin.surements of total IGF concentrations in biological fluids. RecommendationsEndocrinol. Metab. 71: 1202–1207.from the 3rd international symposium on insulin-like growth factors. Endocri-
Thissen, J. P., Triest, S., Maes, M., Underwood, L. E. & Ketelslegers, J. M. (1990)nology 136: 816–817.The decreased plasma concentration of insulin-like growth factor-I in protein-Baxter, R. C., Bryson, J. M. & Turtle, J. R (1981) The effect of fasting on liverrestricted rats is not due to decreased numbers of growth hormone receptorsreceptor for prolactin and growth hormone. Metabolism 30: 1086–1090.on isolated hepatocytes. J. Endocrinol. 124: 159–165.Bornfeldt, K. E., Arnqvist, H. J., Enberg, B., Mathews, L. S. & Norsted, G. (1989)
Tiong, T. S. & Herington, A. C. (1991) Tissue distribution, characterization, andRegulation of insulin-like growth factor-I (IGF-I) and growth hormone receptorregulation of messenger ribonucleic acid for growth hormone receptor andgene expression by diabetes and nutritional state in rat tissues. J. Endocrinol.serum binding protein in the rat. Endocrinology 129: 1628–1634.122: 651–656.
Tonshoff, B., Eden, S., Weiser, E., Carlsson, B., Robinson, I.C.A.F., Blum, W. F. &Breier, B. H., Gluckman, P. D. & Bass, J. J. (1988) The somatotrophic axis inMehls, O. (1994) Reduced hepatic growth hormone (GH) receptor geneyoung steers: influence of nutritional status and oestradiol-17b on hepaticexpression and increased plasma GH binding protein in experimental uremia.high and low-affinity somatotrophic binding sites. J. Endocrinol. 116: 169–Kidney Int. 45: 1085–1092.177.
Villares, S.M.F., Goujon, L., Maniar, S., Delehaye-Zervas, M. C., Martini, J.-F.,Breier,B. H.,Gluckman,P. D.,Blair,H. T.&MacCutcheon,S. N. (1989) Somato-Kleincknecht, C. & Postel-Vinay, M.-C. (1994) Reduced food intake is thetrophic receptors in hepatic tissue of the developing male pig. J. Endocrinol.main cause of low growth hormone receptor in uremic rats. Mol. Cell. Endocri-122: 25–31.
Buonomo, F. C. & Baile, C. A. (1991) Influence of nutritional deprivation on nol. 106: 51–56.
/ 4w2a$$0004 09-11-97 09:32:34 nutra LP: J Nut November
by guest on June 12, 2013jn.nutrition.org
Dow
nloaded from





























