Embed Size (px)
Citation preview

Article No. jmbi.1999.2899 available online at http://www.idealibrary.com on J. Mol. Biol. (1999) 290, 717±730
Molecular and Cellular Analysis of Grb2 SH3 DomainMutants: Interaction with Sos and Dynamin
Michel Vidal, Nathalie Goudreau, Fabrice Cornille, Didier Cussac,Edith Gincel and Christiane Garbay*
DeÂpartement dePharmacochimie MoleÂculaire etStructurale, U266 INSERM-UMR 8600 CNRS, UniversiteÂRene Descartes-UFR desSciences Pharmaceutiques etBiologiques, 4, Avenue del'Observatoire, 75270 ParisCedex 06, France
E-mail address of the [email protected]
Abbreviations used: Grb2, growthbound protein 2; SH2, Src homologhomology 3; EGF, epidermal growtsevenless; GST, glutathione S-transfepidermal growth factor receptor o22; NOE, nuclear Overhauser effectspectroscopy; DQF-COSY, double qspectroscopy ®ltered correlated spetotal correlated spectroscopy.
0022-2836/99/280717±14 $30.00/0
Quantitative analysis of Grb2/dynamin interaction through plasmon res-onance analysis (BIAcore) using Grb2 mutants showed that the high af®-nity measured between Grb2 and dynamin is essentially mediated by theN-SH3 domain of Grb2. In order to study the interactions between Grb2and either dynamin or Sos in more detail, Grb2 N-SH3 domains contain-ing different mutations have been analysed. Two mutations were locatedon the hydrophobic platform binding proline-rich peptides (Y7V andP49L) and one (E40T) located in a region that we had previously shownto be essential for Grb2/dynamin interactions. Through NMR analysis,we have clearly demonstrated that the structure of the P49L mutant isnot folded, while the other E40T and Y7V mutants adopt folded struc-tures that are quite similar to that described for the reference domain.Nevertheless, these point mutations were shown to alter the overallstability of these domains by inducing an equilibrium between a foldedand an unfolded form. The complex formed between the peptideVPPPVPPRRR, derived from Sos, and the E40T mutant was shown tohave the same 3D structure as that described for the wild-type SH3domain. However, the VPPPVPPRRR peptide adopts a slightly differentorientation when it is complexed with the Y7V mutant. Finally, the af®-nity of the proline-rich peptide GPPPQVPSRPNR, derived from dynamin,for the Grb2 N-SH3 domain was too low to be analyzed by NMR. Thus,the interaction between either Sos or dynamin and the SH3 mutants weretested on a cellular homogenate by means of a far-Western blot analysis.In these conditions, the P49L mutant was shown to be devoid of af®nityfor Sos as well as for dynamin. The Y7V SH3 mutant displayed adecrease of af®nity for both Sos and dynamin, while the E40T mutantexhibited a decrease of af®nity only for dynamin. These results supportthe existence of two binding sites between dynamin and the Grb2 N-SH3domain.
# 1999 Academic Press
Keywords: Grb2; SH3 domains; mutants; Sos; dynamin
*Corresponding authoring author:
factor receptor-y 2; SH3, Srch factor; Sos, Son oferase; ER 22,verexpressing clone; NOESY, NOEuantumctroscopy; TOCSY,
Introduction
Grb2 (growth factor receptor bound protein 2) isthe most extensively studied adaptor proteininvolved in growth factor-stimulated signalingpathways (Lowenstein et al., 1992; Chardin et al.,1995). Grb2 is composed of one SH2 (Src homology2) domain surrounded by two SH3 domains(Maignan et al., 1995). It binds tyrosine-phosphory-lated proteins by means of its SH2 domain, andinteracts with several proteins containing proline-rich motifs through its SH3 domains (Simon &Schreiber, 1995). Sos, the nucleotide exchangefactor for Ras (Buday & Downward, 1993; Rozakis-
# 1999 Academic Press

718 Analysis of Grb2 SH3 Domain Mutants
Adcock et al., 1993), and dynamin, a protein essen-tial for receptor endocytosis (Herskovits et al., 1993;Miki et al., 1994), are two important targets of theGrb2 SH3 domains.
The interactions involving SH3 domains and iso-lated proline-rich peptides derived from variousprotein targets are, in general, characterized byaf®nities that are several orders of magnitudelower than those determined using the whole pro-teins, suggesting the existence of additional stabi-lizing interactions between the two patners. Forinstance, in the case of the Grb2/Sos interaction,both the N and C-terminal SH3 domains wereshown to be important for binding, since speci®camino acid mutations in each of the SH3 domainswere reported to be detrimental to the Grb2/Sosbinding af®nity. However, it was shown that theN-terminal SH3 domain contributes more to Sosbinding than its C-terminal counterpart (Rozakis-Adcock et al., 1993; Li et al., 1993). Likewise, otherstudies have reported that both SH3 domains ofGrb2 are important for its interaction with dyna-min (Scaife et al., 1994; Miki et al., 1994). Moreover,we have recently proposed the existence of asecond binding site involved in the interactionbetween Grb2 N-SH3 domain and dynamin, whichis different from the proline-rich binding site andwhich is located at the surface of the SH3 domain.This second binding site is constituted by a stretchof ten amino acid residues, the 36-45 region(WYKAELNGKD), and was shown to be essentialfor Grb2/dynamin interaction but not involved inGrb2/Sos recognition (Vidal et al., 1998).
In the ®rst part of this study, to evaluate theimportance of each of the Grb2 SH3 domains inbinding to dynamin, we have used the surfaceplasmon resonance technique to determine the af®-nity between these two partners, and betweendynamin and two Grb2 analogues bearing theP49L and the P206L mutations, respectively. Thesepoint mutations are located within the N or C-terminal SH3 domains and have been shown to bedetrimental to polyproline peptide binding(Rozakis-Adcock et al., 1993). Our current resultsshow that only the N-SH3 domain is essential formediating the high af®nity of binding betweenGrb2 and dynamin. Therefore in the second part ofthis study, we have undertaken a more detailedcharacterization of the interaction between differ-ent Grb2 N-SH3 mutants and proline-rich peptidesderived from both Sos and dynamin. Speci®cally,three mutants of Grb2 N-SH3 domain have beensynthesized and analyzed at the molecular level.The ®rst two mutants, Y7V and P49L, had pointmutation located at the level of the proline-richpeptide recognition platform (Goudreau et al.,1994; Terasawa et al., 1994; Wittekind et al., 1994,1997; Guruprasad et al., 1995), while the last one,E40T, had a point mutation located within thesecond binding site for dynamin (Vidal et al., 1998).As a ®rst step, the af®nity of these three mutantsfor the proline-rich peptides derived from both Sosand dynamin was measured by ¯uorescence spec-
troscopy. All three mutants were then studied byNMR spectroscopy, both alone and in the presenceof a proline-rich peptide, in order to investigate ifthese domains and their complexes were foldedproperly. Finally, to investigate the effects of thesemutations on the interaction between the Grb2 N-SH3 domain and the whole Sos and dynamin pro-teins, competition experiments on ER 22 cellextracts (®broblast over-expressing EGF-receptor)have been performed through far-Western blotanalysis.
Results
Only the Grb2 N-SH3 domain is essential formediating Grb2/dynamin high-affinity binding
The af®nity and the rates of interaction betweendynamin and different Grb2 mutants weremeasured with a BIAcore biosensor through sur-face plasmon resonance (SPR) in order to detectbinding in real time (Fagerstam et al., 1992;Johnsson et al., 1991). The different Grb2 proteinswere immobilized on the sensor chip and the bind-ing of dynamin was monitored by the increase inresponse unit (RU) following a dynamin ¯ow overthe immobilized Grb2 samples. The early bindingphase (<240 seconds) was used to determine theassociation constant (kass) between dynamin andthe different Grb2 proteins. The dissociation phase(kdiss) of these interactions was then measuredusing an over¯ow of the Grb2 species at the end ofthe dynamin injection. The af®nity constant ofthese complexes was determined as the ratio ofkdiss/kass (see Materials and Methods).
As can be seen in Figure 1, Grb2 and its C-term-inal P206L mutant exhibited similar associationand dissociation pro®les, while the N-terminalP49L mutant did not elicit a measurable associ-ation. Therefore, from the data analysis, similarassociation (105 Mÿ1 sÿ1) and dissociation kinetics(3 � 10ÿ3 sÿ1) were derived for the interactionbetween dynamin and either Grb2 or its C-terminalSH3 (P206L) mutant, corresponding in both casesto an equilibrium dissociation constant value (Kd)of 25(�10) nM. This result clearly shows that theN-SH3 domain is essential for mediating the highaf®nity of binding between Grb2 and dynamin,since no association was measured when the N-terminal SH3 domain was mutated.
Structural characteristics of the two bindingsites of Grb2 N-SH3 using selected mutants
Selection of the mutants
Since the Grb2 N-SH3 domain appeared to bethe most important recognition parameter involvedin the interaction with dynamin or with Sos(Rozakis-Adcock et al., 1993; this work), we havedecided to characterize, at the molecular level, theinteractions between different Grb2-N-SH3mutants and proline-rich peptides derived from
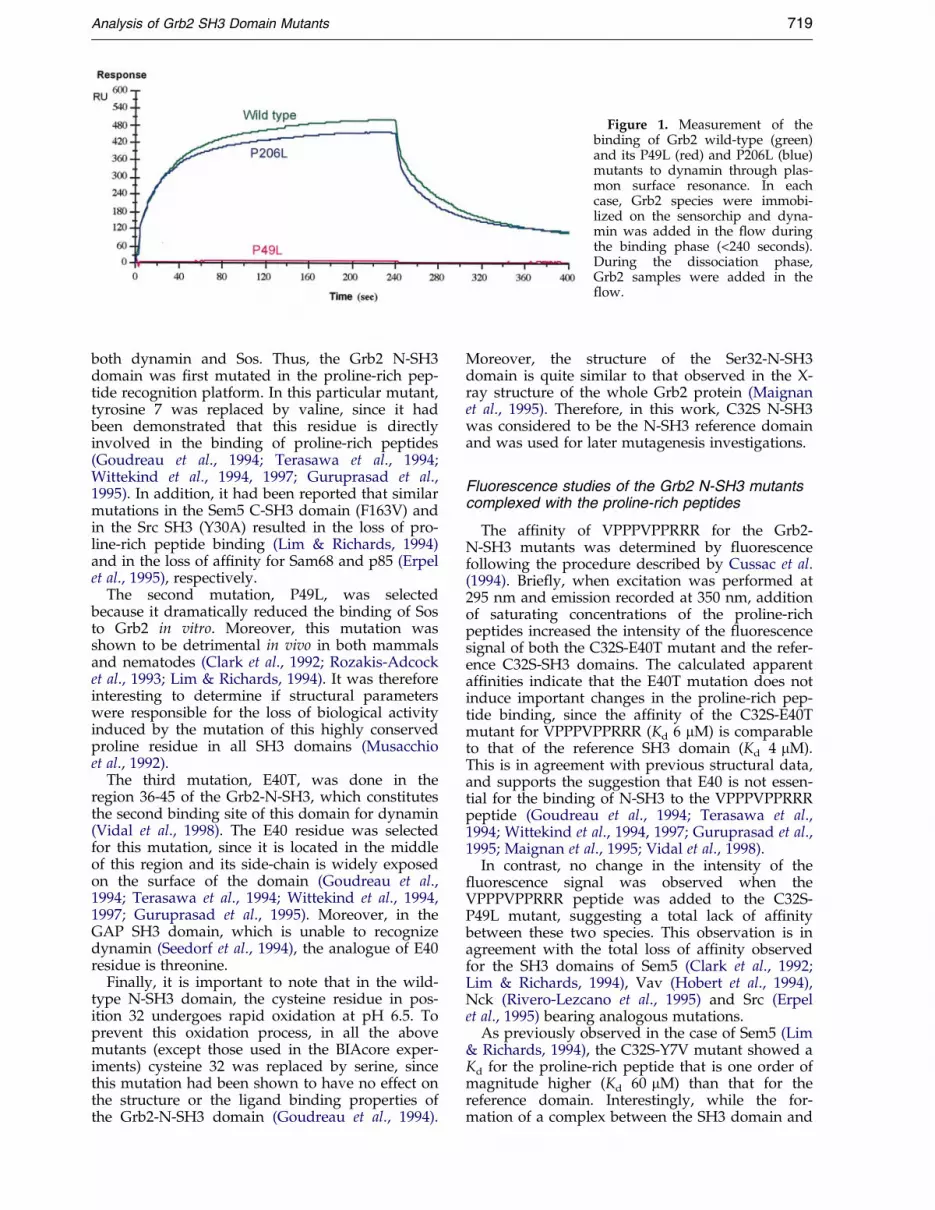
Figure 1. Measurement of thebinding of Grb2 wild-type (green)and its P49L (red) and P206L (blue)mutants to dynamin through plas-mon surface resonance. In eachcase, Grb2 species were immobi-lized on the sensorchip and dyna-min was added in the ¯ow duringthe binding phase (<240 seconds).During the dissociation phase,Grb2 samples were added in the¯ow.
Analysis of Grb2 SH3 Domain Mutants 719
both dynamin and Sos. Thus, the Grb2 N-SH3domain was ®rst mutated in the proline-rich pep-tide recognition platform. In this particular mutant,tyrosine 7 was replaced by valine, since it hadbeen demonstrated that this residue is directlyinvolved in the binding of proline-rich peptides(Goudreau et al., 1994; Terasawa et al., 1994;Wittekind et al., 1994, 1997; Guruprasad et al.,1995). In addition, it had been reported that similarmutations in the Sem5 C-SH3 domain (F163V) andin the Src SH3 (Y30A) resulted in the loss of pro-line-rich peptide binding (Lim & Richards, 1994)and in the loss of af®nity for Sam68 and p85 (Erpelet al., 1995), respectively.
The second mutation, P49L, was selectedbecause it dramatically reduced the binding of Sosto Grb2 in vitro. Moreover, this mutation wasshown to be detrimental in vivo in both mammalsand nematodes (Clark et al., 1992; Rozakis-Adcocket al., 1993; Lim & Richards, 1994). It was thereforeinteresting to determine if structural parameterswere responsible for the loss of biological activityinduced by the mutation of this highly conservedproline residue in all SH3 domains (Musacchioet al., 1992).
The third mutation, E40T, was done in theregion 36-45 of the Grb2-N-SH3, which constitutesthe second binding site of this domain for dynamin(Vidal et al., 1998). The E40 residue was selectedfor this mutation, since it is located in the middleof this region and its side-chain is widely exposedon the surface of the domain (Goudreau et al.,1994; Terasawa et al., 1994; Wittekind et al., 1994,1997; Guruprasad et al., 1995). Moreover, in theGAP SH3 domain, which is unable to recognizedynamin (Seedorf et al., 1994), the analogue of E40residue is threonine.
Finally, it is important to note that in the wild-type N-SH3 domain, the cysteine residue in pos-ition 32 undergoes rapid oxidation at pH 6.5. Toprevent this oxidation process, in all the abovemutants (except those used in the BIAcore exper-iments) cysteine 32 was replaced by serine, sincethis mutation had been shown to have no effect onthe structure or the ligand binding properties ofthe Grb2-N-SH3 domain (Goudreau et al., 1994).
Moreover, the structure of the Ser32-N-SH3domain is quite similar to that observed in the X-ray structure of the whole Grb2 protein (Maignanet al., 1995). Therefore, in this work, C32S N-SH3was considered to be the N-SH3 reference domainand was used for later mutagenesis investigations.
Fluorescence studies of the Grb2 N-SH3 mutantscomplexed with the proline-rich peptides
The af®nity of VPPPVPPRRR for the Grb2-N-SH3 mutants was determined by ¯uorescencefollowing the procedure described by Cussac et al.(1994). Brie¯y, when excitation was performed at295 nm and emission recorded at 350 nm, additionof saturating concentrations of the proline-richpeptides increased the intensity of the ¯uorescencesignal of both the C32S-E40T mutant and the refer-ence C32S-SH3 domains. The calculated apparentaf®nities indicate that the E40T mutation does notinduce important changes in the proline-rich pep-tide binding, since the af®nity of the C32S-E40Tmutant for VPPPVPPRRR (Kd 6 mM) is comparableto that of the reference SH3 domain (Kd 4 mM).This is in agreement with previous structural data,and supports the suggestion that E40 is not essen-tial for the binding of N-SH3 to the VPPPVPPRRRpeptide (Goudreau et al., 1994; Terasawa et al.,1994; Wittekind et al., 1994, 1997; Guruprasad et al.,1995; Maignan et al., 1995; Vidal et al., 1998).
In contrast, no change in the intensity of the¯uorescence signal was observed when theVPPPVPPRRR peptide was added to the C32S-P49L mutant, suggesting a total lack of af®nitybetween these two species. This observation is inagreement with the total loss of af®nity observedfor the SH3 domains of Sem5 (Clark et al., 1992;Lim & Richards, 1994), Vav (Hobert et al., 1994),Nck (Rivero-Lezcano et al., 1995) and Src (Erpelet al., 1995) bearing analogous mutations.
As previously observed in the case of Sem5 (Lim& Richards, 1994), the C32S-Y7V mutant showed aKd for the proline-rich peptide that is one order ofmagnitude higher (Kd 60 mM) than that for thereference domain. Interestingly, while the for-mation of a complex between the SH3 domain and

720 Analysis of Grb2 SH3 Domain Mutants
the proline-rich peptide is characterized by anincrease in ¯uorescence in most cases, the inter-action with the proline-rich peptide resulted in astrong decrease in ¯uorescence in the case of theY7V mutant. This result may re¯ect changes in thestructure of the complex in relation to the absenceof Y7.
It had been reported that the proline-rich peptideGPPPQVPSRPNR, derived from dynamin, wasalso able to inhibit the Grb2/dynamin interaction(Miki et al., 1994; Scaife et al., 1994; Vidal et al.,1998). This peptide was synthesized and its af®nityfor the Grb2 N-SH3 mutants measured. The ¯uor-escence signal of the reference N-SH3 domain wasvery slightly modi®ed by the addition of the dyna-min proline-rich peptide and therefore its af®nitycould only be estimated to be around 300 mM (asmeasured by the displacement of VPPPVPPRRRcomplexed to the N-SH3 domain). As expected,since the E40 residue is not involved in the proline-rich recognition platform, the af®nity of the pep-tide GPPPQVPSRPNR for C32S-E40T mutant wasalmost the same as that measured for the referencedomain.
Figure 2. The 1D 1H NMR spectra of Grb2 N-SH3 domaand NH regions of the C32S Grb2 N-SH3 domain and it(c) C32S-E40T mutant; (d) C32S-Y7V mutant. The reference das an unfolded form (U) and the two other mutants are in eq
NMR analysis of the Grb2 N-SH3 mutants aloneand in complex with VPPPVPPRRR
SH3 domains alone. The N-SH3 mutants were ®rstanalyzed alone (Figure 2). As it had been reportedby several groups (Goudreau et al., 1994; Terasawaet al., 1994; Wittekind et al., 1994, 1997; Guruprasadet al., 1995), the wild-type Grb2 N-SH3 domain iswell structured in solution (phosphate buffer, pH 6,100 mM NaCl) and adopts a compact b-barrel-likestructure. Moreover, in a previous study wedemonstrated that the more stable mutant C32S-N-SH3 adopts a folded structure that is almost identi-cal with that of the wild-type domain (Goudreauet al., 1994). The folding of the domain is character-ized by, among other things, a large spreading ofthe NMR resonances in the 1H NMR spectra, ascan be seen in Figure 2(a). In contrast to this, theNMR resonances of C32S-P49L are not dispersedat all (see Figure 2(b) for the NH region) and allfall within the same regions of the spectra, whichis characteristic of an unstructured and/orunfolded protein (Zhang et al., 1994, 1997; Farrowet al., 1995; Zhang & Forman-Kay, 1995, 1997). Evi-
ins in phosphate buffer (pH 6, 100 mM NaCl): aromatics three mutants. (a) Reference; (b) C32S-P49L mutant;omain exists as a folded form (F), the C32S-P49L mutantuilibrium between the two forms.

Analysis of Grb2 SH3 Domain Mutants 721
dence for such an unfolding was further con®rmedby 2D analysis, where no NOE consistent with thefolded structure was observed (data not shown).
The spectra of C32S-E40T and C32S-Y7Vmutants (Figure 2(c) and (d), respectively) show alarge number of resonances compared to the refer-ence spectra (Figure 2(a)). In fact, the spectrashown in Figure 2(c) and (d) are more or lessequivalent to the sum of the spectra shown inFigure 2(a) and (b) for the folded reference N-SH3domain and the unfolded C32S-P49L mutant,respectively. This observation seems to indicatethat both the Y7V and the E40T mutants exist atpH 6.0 as a 1:1 mixture of folded and unfoldedforms. Evidence for such equilibrium betweenfolded and unfolded forms was further providedby the presence of exchange peaks in the NOESYand ROESY spectra (data not shown). Moreover,similar equilibrium behaviors as a function of pH,ionic strength and temperature have been reportedfor several other SH3 domains, such as those ofDrk (Zhang et al., 1994, 1997; Farrow et al., 1995;Zhang & Forman-Kay, 1995, 1997), a-spectrin(Viguera et al., 1994), and Grb2 (Goudreau et al.,1994; Wittekind et al., 1994).
Analysis of the 1D and 2D NMR spectra for theC32S-E40T SH3 mutant revealed that the resonancechemical shifts for the folded form of this domainwere very similar to those of the reference (C32S)SH3 domain (Goudreau et al., 1994). Only someresidues that are located nearby the mutationpoint, in the three-dimensional space, such as L17,K26, V27, L28, K38, A39, L41, G43, K44, D45 andG46 showed some slight differences in their chemi-cal shifts. Moreover, the distribution of NOEs forthis mutant was almost identical with that of thereference (C32S) N-SH3 domain (Goudreau et al.,1994), with the exception of the two inter-strandNOEs, NH(E40)-NH(K26) and NH(E40)-Ha(V27),which were missing in the case of the C32S-E40TSH3 mutant. In a similar way, the chemical shiftsand the distribution of NOEs for the folded formof the C32S-Y7V SH3 mutant were very similar tothose of the reference SH3 domain (data notshown), with only slight variations in chemicalshifts observed for residues K6, D8, F9, T12, N51and Y52. Altogether, these results seem to indicatethat the E40T and Y7V mutations did not alter thethree-dimensional structures of these N-SH3domains but instead affected only their overallfolding stability.
SH3 domains complexed with VPPPVPPRRR.Due to its low af®nity, the proline-rich peptideGPPPQVPSRPNR derived from dynamin did notform a suf®ciently stable complex with Grb2 SH3mutants to be studied by NMR. Therefore, only thecomplexes formed between the N-SH3 mutantsand the proline-rich peptide VPPPVPPRRR derivedfrom Sos were studied.
First, the addition of the VPPPVPPRRR peptideto the C32S-P49L SH3 mutant did not induce anychange in the NMR spectra of the domain. This is
in agreement with our ¯uorescence data anddemonstrates once again that the proline-rich pep-tide is unable to interact with the P49L mutant.This result is not surprising in the light of ourNMR data, where we have shown that this mutantexists as unfolded form in solution.
In contrast, and in agreement with its 60mMbinding af®nity, the addition of VPPPVPPRRR tothe C32S-E40T SH3 mutant resulted in a complexformation that was also accompagnied by a displa-cement of the equilibrium between the folded andunfolded forms towards entirely the folded form(data not shown). The VPPPVPPRRR/C32S-E40Tcomplex is quite similar to the one we describedfor the reference domain (Goudreau et al., 1994), asdemonstrated by the observation of the same 15intermolecular NOEs, such as NOEs involving theVal5 methyl groups of the proline-rich peptidewith the aromatic protons H2,6 and H3,5 of F9, H6
and H7 of W36 and H3,5 of Y52 of the SH3 domain,NOEs involving b and g protons of Pro6 with H2
and indolic NH of W36, NOEs involving Ha ofArg8 with H2 and indolic NH of W36 and, ®nally,NOEs involving b0 and g protons of Pro2 with thearomatic protons H2,6 and H3,5 of Y7, and H3,5 ofY52 (data not shown). In all cases, the resonanceassignments were obtained by the combined use ofTOCSY, NOESY and COSY experiments throughthe sequence-speci®c method. The assignment ofthe proline-rich peptides was achieved asdescribed (Goudreau et al., 1994).
In the case of the C32S-Y7V mutant, the additionof the VPPPVPPRRR peptide resulted in chemicalshift changes that were slightly different fromthose observed in the reference complex(Goudreau et al., 1994). Speci®cally, changes in 1Hchemical shifts upon proline-rich peptide additionwere observed for the following residues: V7, F9,K10, T12, A13, D15, E16, L17, E30, D33, Q34, N35,W36, K38, F47, P49, K50, N51 and Y52, whereasthe others remained unaffected. In addition, theintermolecular NOEs observed between theVPPPVPPRRR peptide and the C32S-Y7V mutantwere, in part, similar to those observed in the refer-ence complex and, in part, different. For instance,the NOEs involving the methyl groups of Val5 andthe aromatic protons H2,6 and H3,5 of F9, H6 andH7 of W36, and H3,5 of Y52, those involving the band g protons of Pro6 and the H2 and indolic NHof W36 and those involving the Ha of Arg8 andthe H2 and indolic NH of W36 were all conserved.In contrast, instead of observing intermolecularNOEs between the b and g protons of Pro2 and thearomatic protons H2,6 and H3,5 of Y7 and H3,5 ofY52 (as in the reference complex), NOEs betweenthe b and g protons of Pro3 and the aromatic pro-tons H2,6, H3,5 of Y52 and the methyl group of V7were observed (Figure 3). In the light of theseresults, the 3D structure of the complex wasexpected to be somewhat different from that of thewild-type N-SH3 domain and was thereforesolved.

Figure 3. Portion of the 1H 2D NOESY spectrum of the side-chain region of C32S-Y7V Grb2 N-SH3 domain com-plexed with VPPPVPPRRR. Some NOEs have been labeled: crosspeaks involving Y52 aromatic protons and b and gprotons of P3, crosspeaks involving V5 methyl groups of VPPPVPPRRR and F9 and Y52 aromatic protons, and cross-peaks involving V5 methyl groups and H6 and H7 of W36. Amino acids from VPPPVPPRRR are underlined.
722 Analysis of Grb2 SH3 Domain Mutants
Structure of the VPPPVPPRRR/C32S-Y7VN-SH3 complex
The three-dimensional structure of the complexbetween the C32S-Y7V N-SH3 mutant andVPPPVPPRRR was calculated using a simulatedannealing protocol with the Discover softwarepackage and AMBER force®eld as described(Goudreau et al., 1994; and see Materials andMethods). The RMSD of the eight selected struc-tures (see Materials and Methods) against theirmean coordinate are 0.95(�0.1) AÊ for the back-bone and 1.66(�0.14) AÊ for the heavy atoms,when residues 1, 13 to 16 and 27 to 35 from theSH3 domain and the last two residues, 9 and10, from the proline-rich peptide are excluded.The local RMSD values (Berndt et al., 1993),which show the local convergence of the struc-ture, are higher in ¯exible regions. In accord-ance, residues 13 to 16, and residues 27 to 35,which both have high RMSD values, correspondto a part of the long hairpin loop linking strands1 and 2, and to the poorly de®ned loop joining
strands 3 and 4. The structural statistics for theeight individual simulating structures of thecomplex are presented in Tables 1 and 2. TheRamachandran plot for the ordered regions (resi-dues 2 to 9, 16 to 29, 36 to 56 of C32S-Y7V N-SH3 and residues 1 to 8 of VPPPVPPRRR) ofthe ensemble of minimized complex structures isshown in Figure 4. The peptide residues fall intothe (ÿ70 �,140 �) area, which is typical of a poly-proline type II helix. For the C32-Y7V SH3domain, most of the residues fall into theextended area (ÿ90 �,140 �). The structure of theY7V N-SH3 mutant in this complex is very simi-lar to that of C32S reference domain (Goudreauet al., 1994) as can be seen in Figure 5. TheRMSD between the reference C32S N-SH3 andthe C32S-Y7V N-SH3 mutant is 1.4 AÊ for thebackbone atoms (N,Ca,C and O). The interactionsbetween the domain and the proline-rich peptideproceed essentially through hydrophobic con-tacts, as shown in Table 3. In previous analysesof the wild-type complex (Goudreau et al., 1994;

Table 1. Root-mean-square deviations (AÊ ) for the eightselected structures of the C32S-Y7V/VPPPVPPRRR com-plex
Backbone All atoms
All residues 1.25 � 0.14 2.1 � 0.192-12, 17-26, 36-56 (protein)1-8 (peptide)
0.95 � 0.251.66 � 0.2
2-12, 17-26, 36-56 (protein) 0.71 � 0.07 1.45 � 0.11
Table 2. Structural statistics for the eight selected struc-tures of the C32S-Y7V/VPPPVPPRRR complex
A. RMSD from experimentalDistance restraints (AÊ ) 0.0648 � 0.001Dihedral restraints (deg) 0.16 � 0.017
B. RMSD from idealized geometryBonds (AÊ ) 0.04 � 0.001Angles (deg.) 1.63 � 0.03Dihedral o (deg.) 0.15 � 0.05
C. Discover potential energyEnoe (kcal molÿ1) 2.4 � 0.5ELJ (kcal molÿ1) ÿ248 � 5Etot (kcal molÿ1) ÿ341 � 16
Analysis of Grb2 SH3 Domain Mutants 723
Wittekind et al., 1994), the proline-rich peptiderecognition platform of the SH3 domain hadbeen divided into three subsites (S1, S2 and S3),which were shown to interact respectively withPro2, Val5 and Arg8 of the proline-rich peptide.This platform still interacts with the proline-richpeptide in the C32S-Y7V mutant complex, butthe residues involved in binding are slightlydifferent from the peptide point of view. Indeed,Pro3 interacts with the platform instead of Pro2,while Val5 and Arg8 are still involved in theinteraction. All of these interactions are describedin Table 3. This complex is shown in Figure 5(a).Thus, the proline-rich peptide adopts a slightlydifferent orientation on the surface of the SH3domain, as is shown in Figure 5(b).
Figure 4. Ramachandran plot, generated by PROCHECK (29, 36-56) of the N-SH3 domain C32S-Y7V mutant and resistructures; 81.2 % of residues are in most favored regions, 1are in the disallowed region.
Biological interactions of Grb2 N-SH3 mutantdomains on ER 22 cellular extracts
After the physicochemical analysis of the inter-actions involving Grb2 N-SH3 mutants, wedecided to perform their biochemical studies. Thus,the effects of the mutations in the Grb2 N-SH3domain for both the recognition of Sos and dyna-min were studied by competition experimentsusing cellular extracts of ER 22 cells. Far-Westernblot analyses were carried out using GST-Grb2-N-SH3-SH2 as a probe to ensure that no interaction
Laskowski et al., 1993, 1996) for the ordered part (2-9, 16-dues 1-8 of the proline-rich peptide for the ensemble of8.7 % are in the additionally allowed regions and 0.3 %

Figure 5. (a) The 3D structures of the N-SH3 domains of Grb2 complexed with the proline-rich peptideVPPPVPPRRR. Left, C32S-Y7V Grb2 N-SH3 mutant; right, reference domain. The interacting side-chains from theproline-rich peptide are in red, the peptide backbone is in yellow, the interacting side-chains from the SH3 domainare in orange, and the backbone of the SH3 is in blue. The essential differences appear at the level of subsite S1,which comprises amino acid residues Y7 and Y52 in the reference domain and interacts with Pro2 of the peptide.When Y7 is mutated as V7, the proline-rich peptide interacts with V7 and Y52 side-chains by means of Pro3 ringinstead of Pro2. (b) Superposition of the 3D structures of the N-SH3 domains of Grb2 complexed with the proline-rich peptide: in yellow, the reference domain and in red, the mutant domain. Only tyrosine 7 or valine 7 are shown.It appears that the difference is mainly in the way the peptide binds the protein.
724 Analysis of Grb2 SH3 Domain Mutants
with the C-SH3 domain of Grb2 could occur. Inorder to avoid potential interactions involving theSH2 domain, by means of phosphorylation pro-cesses, the cells were blocked in the G0 phase byserum starving for 24 hours before lysis. In theseconditions, only the interactions involving the N-SH3 domain were expected to be detected.
After staining with an anti-GST antibody andrevelation by the ECL method, the amounts of Sos(170 kDa) and dynamin (100 kDa) were quanti®edand the results are presented in Figure 6. The refer-ence C32S N-SH3 domain used as a competitorwas able to displace the complexes formed
between GST-Grb2-N-SH3-SH2 and Sos andbetween GST-Grb2-N-SH3-SH2 and dynamin incellular extracts (Figure 6, column ref). In agree-ment with the fact that it retains a high af®nity forSos, the E40T-C32S mutant was able to displacethe N-SH3-Sos complex (Figure 6, column E40T) asshown by the decrease of the retained 170 kDaprotein. However, this mutant was not able todisplace the 100 kDa protein, corresponding todynamin, in agreement with its lower af®nity forthis protein. This result shows the importance ofthe E40 amino acid residue, located in the 36-45region, in binding to dynamin.

Table 3. Interactions observed between the peptidefrom Sos (VPPPVPPRRR) and the Y7V SH3 domain(hydrogen bonds, hydrophobic and van der Waals con-tacts)
SH3domain Peptide
A. Hydrogen bondsAsp15 Arg8Glu16 Arg8Asp33 Arg9Trp36 Pro6Asn51 Pro4Tyr52 Pro3
B. Hydrophobic and van der Waals contactsVal7 Pro3Phe9 Val5
Asp15 Arg8Glu16 Arg8Asp33 Arg9Asn35 Pro6Trp36 Pro6Phe47 Arg8Pro49 Val5
Pro6Lys50 Pro6Asn51 Pro3
Pro4Pro6
Tyr52 Pro3Val5
Analysis of Grb2 SH3 Domain Mutants 725
The C32S-Y7V mutant at a similar level slightlyinhibited the interaction between Grb2 N-SH3-SH2and either Sos or dynamin (Figure 6, column Y7V).The Y7V mutation, located in the proline-rich rec-ognition platform, re¯ects the importance of the Y7
Figure 6. Quanti®cation of the displacement by N-terminal SH3 domain mutants of Sos and dynaminbound to GST-Grb2-N-SH3-SH2 fusion protein on ER 22cell homogenate. When a mutant possesses af®nity foreither Sos or dynamin, the corresponding band intensitydecreases.
aromatic residue in the recognition of both pro-teins. Finally, comparison of these results suggeststhat the Grb2/dynamin interaction proceedsthrough at least two regions of the Grb2 N-SH3domain (the hydrophobic platform and the 36-45region), whereas the Grb2/Sos interaction involvesonly the hydrophobic recognition platform of theproline-rich peptides.
Finally, as expected from NMR and ¯uorescencedata and in agreement with previous biologicalresults (Rozakis-Adcock et al., 1993; Lim &Richards, 1994), the C32S-P49L mutant was unableto compete with either Sos or dynamin when com-plexed with GST-Grb2-N-SH3-SH2.
Our current results support our previous study,where we demonstrated the importance of the N-SH3 36-45 sequence in mediating the Grb2/dyna-min interaction in addition to the proline-rich pep-tide interactions (Vidal et al., 1998). In contrast, inthe case of Sos, our current data suggest that itsinteraction with the Grb2 N-SH3 domain involvesonly the proline-rich hydrophobic platform. There-fore, the high af®nity of binding between Grb2 andSos is most probably achieved through similaradditional interactions involving the C-SH3domain as previously proposed (Rozakis-Adcocket al., 1993; Li et al., 1993).
Discussion
The NMR analyses of Grb2 N-SH3 domainmutants showed that E40T and Y7V mutations didnot modify the overall structures of these SH3domains but instead altered their overall stabilityby inducing an equilibrium between a folded andan unfolded form. In contrast, the P49L mutationtotally disrupted the three-dimensional structure ofthe domain. This observation provides an expla-nation for the total loss of biological activityobserved for the P49L mutation in Grb2 analoguesand for the lethality in the nematode Caenorhabditiselegans observed for the analogous mutation inSem-5 (Clark et al., 1992; Rozakis-Adcock et al.,1993; Lim & Richards, 1994). However, althoughthis proline residue plays a key role in the foldingof the SH3 domain, its presence is not essential toensure the proper folding of SH3 domains, sincethe GAP SH3 domain, which is devoid of a prolineresidue at this position, adopts a perfectly foldedstructure in solution (Yang et al., 1994). However,SH3 domains lacking this proline residue seem tohave particular binding properties, since GAP SH3and Vav N-SH3 do not possess the classical hydro-phobic platform and do not seem to bind the con-ventional proline-rich peptides (Yang et al., 1994;Ramos-Morales et al., 1995).
The existence of an equilibrium between a foldedand an unfolded form, as observed when Y7 orE40 are mutated, has been reported for severalSH3 domains. It was shown that the equilibriumcould be modi®ed through changes in pH, tem-perature or ionic strengh, or even by the addition

726 Analysis of Grb2 SH3 Domain Mutants
of a proline-rich ligand (Viguera et al., 1994; Zhanget al., 1994, 1997; Farrow et al., 1995; Zhang &Forman-Kay, 1995, 1997). Therefore, it would bevery interesting to determine if such an equili-brium exists in the entire Grb2 protein. Indeed, ifthe folding-unfolding of Grb2 N-SH3 domain existsat the cellular level, it could play an essential rolein the regulation of the Ras signaling pathway. Forinstance, the unfolding of the Grb2 N-SH3 domain,as related to local ionic strength or pH modi®-cations, could promote Grb2/Sos dissociation andtherefore be complementary to the effect of Sosphosphorylation by MAPKinases, which wasdescribed as a factor for Ras desactivation(Rozakis-Adcock et al., 1995; Kharlund et al., 1995;Cherniack et al., 1995). Moreover, such an equili-brium may suggest a possible physiological rolefor deletion mutants of Grb2, particularly the Ash-s protein (Watanabe et al., 1995). Indeed, thismutant, corresponding to the N-SH3 domain ofGrb2, could adopt a folded or unfolded structurein function of ionic strength in the cell and couldparticipate in the Ras signaling pathway regu-lation.
Another interesting observation of our results, isthe formation of slightly different complexesbetween VPPPVPPRRR and either C32S or C32S-Y7V SH3 domains. In the latter complex, the pro-line-rich peptide still interacts with the hydro-phobic platform but it adopts a differentconformation at the surface of the Y7V SH3domain mutant. Moreover, the af®nity of the pep-tide for this SH3 domain is one order of magnitudelower. This might provide a low binding betweenSos and the C32S-Y7V SH3 domain. Indeed, in theentire protein, the VPPPVPPRRR sequence prob-ably cannot easily adopt the conformation necess-ary to interact with this mutant, which mightproduce an important af®nity decrease betweenthe proteins in vivo.
Here, we showed through surface plasmon res-onance studies that only the N-terminal SH3domain is really important for mediating the highaf®nity of binding of Grb2 for dynamin. Indeedthe P49L mutant, in which the N-SH3 domain isnot folded, is unable to bind dynamin, whereas theP206L mutant retains its af®nity. While it has notbeen shown to be the case, one could expect thatthe P206L C-SH3 mutation acts similarly to P49LN-SH3, based on structure alignment (Musacchioet al., 1992). Moreover, this mutation was shown tohave effects on ligand binding similar to that of itsanalogue in the N-SH3 domain (Fukazawa et al.,1995). However the dynamin proline-rich peptideGPPPQVPSRPNR, which was proposed to interactwith Grb2, did not give a complex with the Grb2N-SH3 domain suf®ciently stable to be analyzedthrough NMR. These results support the assump-tion that additional interactions might existbetween dynamin and this SH3 domain to accountfor the high af®nity measured between the twoproteins.
Moreover, on cellular homogenates, with respectto the fact that SH3 domains bind proline-richregions, we show that the Y7V mutation, locatedin the hydrophobic platform, affects the binding ofthe N-SH3 domain of Grb2 to both Sos and dyna-min. In contrast, the E40T mutation, located in the36-45 sequence, modi®es only the binding to dyna-min. These results support the suggestion that Sosbinds the N-SH3 domain of Grb2 at the proline-rich recognition platform, while dynamin bindsthis domain through the hydrophobic platformand through the 36-45 sequence, by means ofanother unknown mechanism (Vidal et al., 1998).
Dynamin is a protein that has been shown toplay a crucial role in the endocytosis pathway (fora review, see Liu & Robinson, 1995). The GTPaseactivity of dynamin was shown to be related to thebinding of protein containing SH3 domains (Goutet al., 1993) such as Grb2, PI3-Kinase and evenamphiphysin (Wigge et al., 1997). The GTPaseactivity regulation is a very complex process, sincedifferent events take place, such as dynamin oligo-merization (Tuma & Collins, 1995), phosphoryl-ation (Robinson et al., 1994) and binding tomicrotubules (Shoetner & Vallee, 1992), phospholi-pids (Zheng et al., 1996) or calcium (Liu &Robinson, 1995). It was even proposed that theSH2 domain of Grb2 could play a role, since aphosphopeptide derived from the insulin-receptorwas described to activate dynamin GTPase activity(Ando et al., 1994). Recent data have shown that,while the N-SH3 domain of Grb2 is essential for itsbinding to dynamin, it failed to exert any effect onreceptor-mediated endocytosis (Owen et al., 1998).Moreover, transfection experiments of amphiphy-sin exert a negative effect on endocytosis in ®bro-blasts (Wigge et al 1997), showing that dynamin/amphiphysin binding is the essential parameter forreceptor endocytosis. Therefore, what could be therole of Grb2/dynamin complex in cells? In a pre-vious paper, we have shown that EGF stimulationon ER 22 cells produced an increase of Grb2/Sospool, concomitant with a decrease of Grb2/dyna-min complex. These results suggested that Grb2/Sos increase might happen, at least in part, at theexpense of the Grb2/dynamin complex, whichcould consequently liberate free dynamin (Vidalet al., 1998). Thus, Grb2/dynamin preformed com-plexes might constitute an ``inactive storage form''of dynamin, in terms of endocytosis, which couldbe liberated near the membrane following receptoractivation. This pool of dynamin could self-assem-ble (Takei et al., 1995) at the clathrin lattice, bindGTP and then could be trapped by another part-ner, such as amphiphysin, which has a high af®-nity for dynamin and is able to bind the AP2complex (Damke et al., 1994; Shupliakov et al.,1997). By preventing dynamin oligomerization, asrecently described, amphiphysin could redistributedynamin along the neck of the budding vesicle atthe clathrin-coated pit. Thus, the potential role ofthe Grb2/dynamin complex in endocytosis, whichhas been largely controversial, could therefore

Analysis of Grb2 SH3 Domain Mutants 727
consist only of the recruitment and liberation ofdynamin near the membrane, following growthfactor stimulation.
Materials and Methods
Surface plasmon resonance (BIAcore)
Surface plasmon resonance (SPR) experiments werecarried out on a Pharmacia Biosensor BIAcore 2000apparatus. Grb2 or its mutants were covalently bound tothe dextran matrix of a CM5 sensorchip (Pharmacia Bio-sensor), via their free amino groups, by the N-ethyl-N0-(3-dimethylaminopropyl)-carbodiimide-N-hydroxysucci-nimide activation method followed by a treatment with1 M ethanolamine. Immobilizations were performed in10 mM potassium acetate buffer (pH 7). The amountof protein bound to the sensorchip was monitored by thechange in refractive index. In order to allow themeasurement of kinetic parameters, the matrix wascontrolled to make sure it was not saturated. Bindingexperiments were performed at 25 �C in HBS runningbuffer (20 mM Hepes (pH 7.4), 1 mM DTT, 150 mMNaCl, 0.005 % (v/v) Tween 20) with a ¯ow-rate of20 ml/minute.
After each binding experiment, the sensorchip wasregenerated by a three minute pulse of 10 ml of 3 M gua-nidinium chloride in HBS buffer, a treatment that didnot produce signi®cant changes in baseline and responseafter 40 runs. Thus, estimation of kinetic parameters wasrealized by repetitive injections of a whole range of con-centration of dynamin over different densities ofimmobilized Grb2 entities. Analysis of experimental datawas performed with the interactive software BIA evalu-ation v2.0 (Pharmacia Biosensor). The signal observedwhen an interaction takes place is measured in resonanceunits, Ru , which corresponds to the amount of boundprotein. The signal change rate is described by the sim-pli®ed equation dRu/dt � constant ÿ C ks Ru where C isthe concentration of the injected protein andks � kass � C � kdiss. The dissociation rate constant wasalso measured in free buffer ¯ow, after the end of thedynamin injection. The af®nity constant was de®ned asthe ratio kdiss/kass (Johnsson et al., 1991; Fagerstam et al.,1992).
Peptide synthesis and purifications
Dynamin was puri®ed from rat brain as described(Montiel et al., 1997). The proline-rich peptides and SH3domains were prepared by solid-phase synthesis. Theprotected peptide chains were assembled according tothe stepwise method of Merri®eld on an Applied Biosys-tems 431A peptide synthesizer with HOBt/DCC ascoupling reagents. Synthetic peptides corresponding toGPPPQVPSRPNR and VPPPVPPRRR were synthesizedusing ten equivalents of amino acids, while SH3 domainswere synthesized using 20 equivalents, as described(Yang et al., 1994). The peptides were puri®ed byHPLC using a C4 (for SH3 domains) or a C8 (fordeca- and dodecapeptides) Vydac 5 mm column(220 mm � 10 mm) and a linear gradient of 10-90 % B in90 min (where A is 0.1 % TFA; B is 70 % CH3CN, 0.09 %TFA) at a ¯ow-rate of 1.5 ml/minute with detection at214 nm. The identity of the peptides was checked byelectrospray mass spectroscopy.
Cells and preparation of cell lysates
Adherent ER 22 cells (hamster ®broblasts overexpres-sing the human epidermal growth factor receptor, a kindgift from Dr J. Pouyssegur, So®a Antipolis, Nice, France)were routinely grown in Dulbecco's modi®ed Eagle'smedium supplemented with 10 % (v/v) fetal calf serumcontaining the antibiotic G418 (200 mg/ml) and 2 mML-glutamine (GIBCO-BRL) and were incubated at 37 �Cin 5 % CO2. Cells were grown to con¯uence and wereserum-starved for 24 hours.
Cells were rapidly washed with ice-cold PBS (137 mMNaCl, 2.7 mM KCl, 9.6 mM Na2HPO4, 1.47 mMKH2PO4) and solubilized for 30 minutes at 4 �C in 1 mlof HNTG lysis buffer (50 mM Hepes (pH 7.5), 150 mMNaCl, 10 % glycerol, 1 % Triton X-100, 1 mM EGTA,1 mM MgCl2, 1 mM Na3VO4, 10 mM Na4P2O7, 10 mMNaF, 1 mg/ml leupeptin, 1 mg/ml pepstatin A, 1 mg/mltrypsin inhibitor, 1 mg/ml chymostatin, 1 mg/ml anti-pain, 2 mg/ml aprotinin, 10 mg/ml benzamidine, 1 mMphenylmethylsulfonyl ¯uoride). Lysates were clearedby centrifugation at 15,000 rpm for ten minutes, andthe protein concentration was determined (Bio-Radmicroassay).
GST fusion proteins
DNA sequences corresponding to Grb2 (residues 1 to210) and Grb2 N-SH3-SH2 (residues 1 to 160) wereampli®ed by PCR and cloned into the expression vectorpGEX2T at BamHI and EcoRI restriction sites. Bacteriatransformed with the recombinant plasmids weregrown, induced with IPTG, and disrupted by sonication(Fath et al., 1994). The GST-SH3 fusion proteins werepuri®ed by af®nity chromatography on glutathione-agarose beads (Sigma) followed by elution with 10 mMreduced glutathione.
Antibodies
The anti-glutathione S-transferase (GST) monoclonalantibody was purchased from Hybridolab Pasteur Insti-tute (Paris, France). The peroxydase anti-mouse antibodywas purchased from Sigma .
Far-Western blot analysis
Proteins from a total cell lysate were fractionated bySDS-PAGE (7.5 % (w/v) polyacrylamide gel) and trans-ferred onto PVDF membrane as described (Parker et al.,1996). Non-speci®c binding to the ®lters was blocked byadding 2 % (w/v) skimmed milk in PBS containing0.05 % Tween 20 for two hours at room temperature. The®lters were incubated with (50 mg/ml) or without SH3domains overnight and then GST-Grb2 N-SH3-SH2 pro-tein (1.75 mg/ml) in blocking buffer was added beforeincubation for three hours at room temperature. Afterwashes in PBS containing 0.05 % Tween 20, bound pro-teins were detected by successive incubations with theanti-GST Mab (0.25 mg/ml), peroxidase conjugated anti-mouse antibody and revealed by the ECL method(Amersham). Quantitative evaluation of band intensitywas performed on a Bio-pro®l V6.0 Vilber-Lourmat(Marne-La-ValleÂe, France). Data are mean values (�SEM)of three independent assays.

728 Analysis of Grb2 SH3 Domain Mutants
Fluorescence studies
Fluorescence measurements were carried out with aPerkin-Elmer LS 50B luminescence spectrophotometer at20 �C. The excitation wavelength used in most of theexperiments was 280 nm (6 nm slit), and the emissionwavelength was 350 nm (10 nm slit). In a typical assay,aliquots of peptide solution were added to 800 ml of1 mM SH3 solution in 20 mM phosphate buffered saline(100 mM NaCl, pH 6.0), and the mixture was stirred in acuvette three minutes prior to analysis with the spectro-photometer. The peptide solution was added until nosigni®cant SH3-dependent changes in ¯uorescence inten-sity was observed. After background subtraction andvolume correction, the dissociation constant (Kd) was cal-culated as described by Cussac et al. (1994).
NMR experiments
The C32S Grb2-N-SH3 reference domain as well as themutants C32S-E40T and C32S-Y7V were dissolved at1 mM concentration, in 2H2O or H2O/2H2O (9:1) 20 mMsodium phosphate buffer (pH 6.0), containing 100 mMNaCl.
The NMR samples of the N-SH3 mutants/proline-richpeptide complexes were about 1 mM in a 1:1 ratio of theGrb2-N-SH3 domain and VPPPVPPRRR in 2H2O orH2O/2H2O (9:1) 20 mM sodium phosphate buffer(pH 6.0) containing 100 mM NaCl. However, for theC32S-Y7V mutant complex, the buffer contained500 mM NaCl.
The 2D 1H-NMR spectra were recorded at 20 �C andat 600 MHz on a BruÈ ker AMX600 spectrometer. All the2D experiments were acquired in the phase-sensitivemode using the TPPI method (Marion & WuÈ trich, 1993).HOHAHA spectra were recorded with mixing times of50 and 80 ms, and with a clean TOCSY pulse sequence(Bax & Davis, 1985). NOESY spectra were recorded withmixing times of 70, 100 and 200 ms (Macura et al., 1981).
The conversion of NOEs into distance constraints wasdone by classi®cation of weak, medium and strong NOEintensities into distance intervals of 1.8-2.5 AÊ , 1.8-3.5 AÊ
and 1.8-5.0 AÊ , respectively. 3JNH-Ha and 3JHaHb couplingconstants were measured with DQF-COSY (Rance et al.,1983) and converted into � and angles when appro-priate.
Structure calculations
The structure calculations for the C32S-Y7V SH3/VPPPVPPRRR complex were done using a simulatedannealing protocol with the Discover (version 95.0) soft-ware/package and AMBER force®eld (Clore et al., 1986;MSI, San Diego CA) as described (Goudreau et al., 1994).In all, 786 NOE constraints were used: 158 were sequen-tial (i ÿ j � 1), 24 medium (1 < i ÿ j < 4), 184 long dis-tance (i ÿ j > 4), 420 intraresidual; 16 contraintscorresponding to hydrogen bonds, as found by exchangeNMR experiments were included as distance constraints(two constraints per hydrogen bond); 17 intermolecularrestraints were used in the calculation. Peptidic angleswere maintained as trans for both the C32S-Y7V SH3domain and the VPPPVPPRRR peptide with a force con-stant of 200 kcalÿ1 molÿ2. Twenty � torsion angles wererestrained within the range ÿ150 � < � < ÿ90 �. Thesetorsion angles correspond to protein residues with 3JNH-
Ha > 8 Hz. For the proline-rich peptide, seven � torsionangles within the range ÿ105 � < � < ÿ45 � and six tor-
sion angles within the range 115 � < < 175 � were intro-duced as constraints to maintain the proline-rich peptidein the polyproline II helix conformation. The referenceC32S-N-SH3/VPPPVPPRRR complex was used as astarting point for the calculations. Fifty structures weregenerated and eight were selected on the basis of energyand NOE violations.
Brookhaven Protein Data Bank accession number
The coordinates have been deposited in the Brook-haven Protein Data Bank under reference 1AZE.
Acknowledgements
The authors gratefully acknowledge Drs B. Tocque, M.Duchesne and F. Parker from ``RhoÃne Poulenc Rorer'' fortheir help in biological experiments, and for providingGST-Grb2-N-SH3-SH2 protein. Dr P. Chardin isacknowledged for providing Grb2 mutants.
The authors thank C. LENOIR for her assistance inpeptide synthesis, K. Wecker and Drs S. Bouaziz and G.Tiraboschi for their help in iconographic preparation ofthe manuscript. We acknowledge C. Dupuis for herinvaluable help in drafting this manuscript.
The authors particularly thank Professor B. P. Roquesfor his scienti®c help to develop this project.
This work was supported by the ``Ligue Nationalecontre le Cancer'' (Comite de Paris) and M.V. thanks``RhoÃne Poulenc Rorer'' for his ®nancial support.
References
Ando, A., Yonezawa, K., Gout, I., Nakata, T., Ueda, H.,Hara, K., Kitamura, Y., Noda, Y., Takenawa, T.,Hirokawa, N., Water®eld, M. D. & Kasuga, M.(1994). A complex of Grb2-dynamin binds to tyro-sine-phosphorylated insulin receptor substrate-1after insulin treatment. EMBO J. 13, 3033-3038.
Bax, A. & Davis, D. G. (1985). MLEV-17 based two-dimensional homonuclear magnetization transferspectroscopy. J. Magn. Reson. 65, 355-360.
Berndt, K. D., GuÈ ntert, P. & WuÈ thrich, K. (1993). NMRsolution structure of dendrotoxin K from venom ofDendroapis polypepis polypepis. J. Mol. Biol. 234, 735-750.
Buday, L. & Downward, J. (1993). Epidermal growthfactor regulates p21Ras through the formation of acomplex of receptor, Grb2 adapter protein, and Sosnucleotide exchange factor. Cell, 73, 611-620.
Chardin, P., Cussac, D., Maignan, S. & Ducruix, A.(1995). The Grb2 adaptor. FEBS Letters, 369, 47-51.
Cherniack, A. D., Klarlund, J. K., Conway, B. R. &Czech, P. (1995). Disassembly of Son-of-sevenlessproteins from Grb2 during p21ras desensitizationby insulin. J. Biol. Chem. 270, 1485-1488.
Clark, S. G., Stern, M. J. & Horvitz, H. R. (1992). C.elegans cell-signalling gene Sem-5 encodes a proteinwith SH2 and SH3 domains. Nature, 356, 340-344.
Clore, G. M., BruÈ nger, A. T., Karplus, M. & Gronenborn,A. M. (1986). Application of molecular dynamicswith interproton distance restraints to three-dimen-sional protein structure determination. J. Mol. Biol.191, 523-551.

Analysis of Grb2 SH3 Domain Mutants 729
Cussac, D., Frech, M. & Chardin, P. (1994). Binding ofthe Grb2 SH2 domain to phosphotyrosine motifsdoes not change the af®nity of its SH3 domains forSos proline-rich motifs. EMBO J. 13, 4011-4021.
Damke, H., Bada, T., Warnock, D. E. & Schmid, S. L.(1994). Induction of mutant dynamin speci®callyblocks endocytic coated vesicle formation. J. CellBiol. 127, 915-934.
Erpel, T., Superti-Furga, G. & Courtneige, S. A. (1995).Mutational analysis of the Src SH3 domain: thesame residues of the ligand binding surface areimportant for intra- and intermolecular interactions.EMBO J. 14, 963-975.
Fagerstam, L. G., Frostell-Karlsson, A., Karlsson, R.,Persson, B. & Ronnberg, I. (1992). Biospeci®c inter-action analysis using surface plasmon resonancedetection applied to kinetic, binding site and con-centration analysis. J. Chromatog. 597, 397-410.
Farrow, N. A., Zhang, O., Forman-Kay, J. D. & Kay,L. E. (1995). Comparison of the backbone dynamicsof a folded and an unfolded SH3 domain existingin equilibrium in aqueous buffer. Biochemistry, 34,868-878.
Fath, I., Schweighoffer, F., Rey, I., Multon, M. C.,Boiziau, J., Duchesne, M. & TocqueÂ, B. (1994). Clon-ing of a Grb2 isoform with apoptotic properties.Science, 264, 971-974.
Fukazawa, T., Reedquist, K. A., Trub, T., Soltoff, S.,Panchamoorthy, G., Druker, B., Cantley, L.,Shoelson, S. E. & Band, H. (1995). The SH3 domain-binding T cell tyrosyl phosphoprotein p120.Demonstration of its identity with the c-cbl proto-oncogene product and in vivo complexes with Fyn,Grb2, and phosphatidylinositol 3-kinase. J. Biol.Chem. 270, 19141-19150.
Goudreau, N., Cornille, F., Duchesne, M., Parker, F.,TocqueÂ, B., Garbay, C. & Roques, B. P. (1994). NMRstructure of the N-terminal SH3 domain of Grb2and its complex with a proline-rich peptide fromSos. Nature Struct. Biol. 1, 898-907.
Gout, I., Dhand, R., Hiles, I. D., Fry, M. J., Panayotou,G., Das, P., Truong, O., Totty, N. F., Hsuan, J.,Booker, G. W., Campbell, I. D. & Water®eld, M. D.(1993). The GTPase dynamin binds to and is acti-vated by a subset of SH3 domains. Cell, 75, 25-36.
Guruprasad, L., Dhanaraj, V., Timm, D., Blundell, T. L.,Gout, I. & Water®eld, M. D. (1995). The crystalstructure of the N-terminal SH3domain of Grb2.J. Mol. Biol. 248, 856-866.
Herskovits, J. S., Burgess, C. C., Obar, R. A. & Vallee,R. B. (1993). Effects of mutant rat dynamin on endo-cytosis. J. Cell Biol. 122, 565-578.
Hobert, O., Jallal, B., Schlessinger, J. & Ullrich, A. (1994).Novel signaling pathway suggested by SH3domain-mediated p95vav/heterogeneous ribonu-cleoprotein K interaction. J. Biol. Chem. 269, 20225-20228.
Johnsson, U., Fagerstam, L., Ivarsson, B., Johnsson, B.,Karlsson, R., Lundh, K., Lofas, S., Persson, B., Roos,H. & Ronnberg, I., et al. (1991). Real-time biospeci®cinteraction analysis using surface plasmon reson-ance and a sensorchip technology. Biotechniques, 11,620-627.
Kharlund, J. K., Cherniack, A. D. & Czech, M. P. (1995).Divergent mechanisms for homologous desensitiza-tion of p21ras by insulin and growth factors. J. Biol.Chem. 270, 23421-23428.
Laskowski, R. A., MacArthur, M. W., Moss, D. S. &Thornton, J. M. (1993). PROCHECK: a program to
check the stereochemical quality of protein struc-tures. J. Appl. Crystallog. 26, 283-291.
Laskowski, R. A., Rullmann, J. A. C., MacArthur, M. W.,Kaptein, R. & Thornton, J. M. (1996). AQUA andPROCHECK-NMR: programs for checking thequality of protein structures solved by NMR. J. Biol.Mol. NMR, 8, 477-486.
Li, N., Batzer, A., Daly, R., Yajnik, V., Skolnik, E.,Chardin, P., Bar-Sagi, D., Margolis, B. &Schlessinger, J. (1993). Guanine-nucleotide-releasingfactor hSos1 binds to Grb2 and links receptor tyro-sine kinases to Ras signalling. Nature, 363, 85-88.
Lim, W. A. & Richards, F. M. (1994). Critical residues inan SH3 domain from Sem-5 suggest a mechanismfor proline-rich peptide recognition. Nature Struct.Biol. 1, 221-225.
Liu, J.-P. & Robinson, P. J. (1995). Dynamin and endocy-tosis. Endocrinol. Rev. 16, 590-607.
Lowenstein, E. J., Daly, R. J., Batzer, A. G., Li, W.,Margolis, B., Lammers, R., Ullrich, A., Skolnik, E. Y.,Bar-Sagi, D. & Schlessinger, J. (1992). The SH2 andSH3 domain-containing protein Grb2 links receptortyrosine kinases to ras signaling. Cell, 70, 431-442.
Macura, S., Huang, Y., Suter, D. & Ernst, R. R. (1981).Two-dimensional chemical exchange and cross-relaxation spectroscopy of coupled nuclear spins.J. Magn. Reson. 43, 259-281.
Maignan, S., Guilloteau, J. P., Fromage, N., Arnoux, B.,Becquart, J. & Ducruix, A. (1995). Crystal structureof the mammalian Grb2 adaptor. Science, 268, 291-293.
Marion, D. & WuÈ thrich, K. (1993). Application of phasesensitive two-dimensional correlated spectroscopy(COSY) for measurements of 1H-1H spin-spin coup-ling constants in proteins. Biochem. Biophys. Res.Commun. 113, 967-974.
Miki, H., Miura, K., Matuoka, K., Nakata, T., Hirokawa,N., Orita, S., Kaibuchi, K., Takai, Y. & Takenawa, T.(1994). Association of Ash/Grb2 with dynaminthrough the Src homology 3 domain. J. Biol. Chem.269, 5489-5492.
Montiel, J. L., Cussac, D., Cornille, F., Vidal, M., Garbay,C. & Roques, B. P. (1997). Rapid and ef®cient puri®-cation of rat brain dynamin using an af®nity col-umn of the carboxy-terminal SH3 domain of Grb2.Protein Pept. Letters, 4, 195-202.
Musacchio, A., Gibson, T., Lehto, V. P. & Saraste, M.(1992). SH3-an abundant protein domain in searchof a function. FEBS Letters, 307, 55-61.
Owen, D. J., Wigge, P., Vallis, Y., Moore, J. D., Evans,P. R. & McMahon, H. T. (1998). Crystal structure ofthe amphiphysin-2 SH3 domain and its role in theprevention of dynamin ring formation. EMBO J. 15,5273-5285.
Parker, F., Maurier, F., Delumeau, I., Duchesne, M.,Faucher, D., Debusche, L., Dugue, A.,Schweighoffer, F. & TocqueÂ, B. (1996). A Ras-GTPase-activating protein SH3-domain-binding pro-tein. Mol. Cell Biol. 16, 2561-2569.
Ramos-Morales, F., Romero, F., Schweighoffer, F.,Bismuth, G., Camonis, J., Tortolero, M. & Fischer, S.(1995). The proline-rich region of Vav binds to Grb2and Grb3-3. Oncogene, 11, 1665-1669.
Rance, M., Sorensen, O. W., Bodenhausen, G., Wagner,G., Ernst, R. R. & Wuthrich, K. (1983). Improvedspectral resolution in COSY 1H NMR spectra ofproteins via double quantum ®ltering. Biochem. Bio-phys. Res. Commun. 117, 479-485.

730 Analysis of Grb2 SH3 Domain Mutants
Rivero-Lezcano, O. M., Marcilla, A., Sameshima, J. H. &Robbins, K. C. (1995). Wiskott-Aldrich syndromeprotein physically associates with Nck through Srchomology 3 domains. Mol. Cell. Biol. 15, 5725-5731.
Robinson, P. J., Liu, J. P., Powell, K. A., Fykse, E. M. &SuÈ dhof, T. C. (1994). Phosphorylation of dynamin Iand synaptic-vesicle recycling. Trends Neurosci. 17,348-353.
Rozakis-Adcock, M., Fernley, R., Wade, J., Pawson, T. &Bowtell, D. (1993). The SH2 and SH3 domains ofmammalian Grb2 couple the EGF receptor to theRas activator mSos1. Nature, 363, 83-85.
Rozakis-Adcock, M., Van Der, Geer P., Mbamalu, G. &Pawson, T. (1995). MAP kinase phosphorylation ofmSos1 promotes dissociation of mSos1-Shc andmSos1-EGF receptor complexes. Oncogene, 11, 1417-1426.
Scaife, R., Gout, I., Water®eld, M. D. & Margolis, R. L.(1994). Growth factor-induced binding of dynaminto signal transduction proteins involves sorting todistinct ans separate proline-rich dynaminsequences. EMBO J. 13, 2574-2582.
Seedorf, K., Kostka, G., Lammers, R., Bashkin, P., Daly,R., Burgess, W. H., Van der Bliek, A. M.,Schlessinger, J. & Ullrich, A. (1994). Dynamin bindsto SH3 domains of phospholipase Cg and Grb2.J. Biol. Chem. 269, 16009-16014.
Shpetner, H. S. & Vallee, R. B. (1992). Dynamin is aGTPase stimulated to high levels of activity bymicrotubules. Nature, 355, 733-735.
Shupliakov, O., LoÈw, P., Grabs, D., Gad, H., Chen, H.,David, C., Takei, K., De Camilli, P. & Brodin, L.(1997). Synaptic vesicle endocytosis impaired bydisruption of dynamin-SH3 domain interactions.Science, 276, 259-263.
Simon, J. A. & Schreiber, S. L. (1995). Grb2 SH3 bindingto peptides from Sos: evaluation of a general modelfor SH3-ligand interactions. Curr. Biol. 2, 53-60.
Takei, K., McPherson, P. S., Schmid, S. L. & De Camilli,P. (1995). Tubular membrane invaginations coatedby dynamin rings are induced by GTP-gS in nerveterminals. Nature, 374, 186-189.
Terasawa, H., Kohda, D., Hatanaka, H., Tsuchiya, S.,Ogura, K., Nagata, K., Ishii, S., Mandiyan, V.,Ullrich, A., Schlessinger, J. & Inagaki, F. (1994).Structure of the N-terminal SH3 domain of Grb2complexed with a peptide from the guanine nucleo-tide releasing factor Sos. Nature Struct. Biol. 1, 891-897.
Tuma, P. L. & Collins, C. A. (1995). Dynamin formspolymeric complexes in the presence of lipid ves-icles. Characterization of chemically cross-linkeddynamin molecules. J. Biol. Chem. 270, 26707-26714.
Vidal, M., Montiel, J. L., Cussac, D., Cornille, F.,Duchesne, M., Parker, F., TocqueÂ, B., Roques, B. P.& Garbay, C. (1998). Differential interactions of thegrowth factor receptor-bound protein 2 N-SH3
domain with Son of Sevenless and dynamin. J. Biol.Chem. 273, 5343-5348.
Viguera, A. R., Martinez, J. C., Filimonov, V. V., Mateo,P. L. & Serrano, L. (1994). Thermodynamic and kin-etic analysis of the SH3 domain of spectrin shows atwo-state folding transition. Biochemistry, 33, 2142-2150.
Watanabe, K., Fukuchi, T., Hosoya, H., Shirasawa, T.,Matuoka, K., Miki, H. & Takenawa, T. (1995). Spli-cing isoforms of rat Ash/Grb2. J. Biol. Chem. 270,13733-13739.
Wigge, P., Vallis, Y. & McMahon, H. T. (1997). Inhi-bition of receptor-mediated endocytosis by theamphiphysin SH3 domain. Curr. Biol. 7, 554-560.
Wittekind, M., Mapelli, C., Farmer, B. T., II, Suen, K. L.,Goldfarb, V., Tsao, J., Lavoie, T., Barbacid, M.,Meyers, C. A. & Mueller, L. (1994). Orientation ofpeptide fragments from Sos proteins bound to theN-terminal SH3 domain of Grb2 determined byNMR spectroscopy. Biochemistry, 33, 13531-13539.
Wittekind, M., Mapelli, C., Lee, V., Goldfarb, V.,Friedrichs, M. S., Meyers, C. A. & Mueller, L.(1997). Solution structure of the Grb2 N-terminalSH3 domain complexed with a ten-residue peptidederived from Sos: direct re®nement against NOEs,J-couplings and 1H and 13C chemical shifts. J. Mol.Biol. 267, 933-952.
Yang, Y. S., Garbay, C., Duchesne, M., Fromage, N.,TocqueÂ, B. & Roques, B. P. (1994). Solution structureof GAP SH3 domain by 1H NMR and spatialarrangement of essential Ras signaling-involvedsequence. EMBO J. 13, 1270-1279.
Zhang, O., Kay, L. E., Olivier, J. P. & Forman-Kay, J. D.(1994). Backbone 1H and 15N resonance assign-ments of the N-terminal SH3 domain of drk infolded and unfolded states using enhanced-sensi-tivity pulsed ®eld gradient NMR techniques.J. Biomol. NMR, 4, 845-858.
Zhang, O. & Forman-Kay, J. D. (1995). Structural charac-terization of folded and unfolded states of an SH3domain in equilibrium in aqueous buffer. Biochemis-try, 34, 6784-6794.
Zhang, O., Forman-Kay, J. D., Shortle, D. & Kay, L. E.(1997). Triple-resonance NOESY-based experimentswith improved spectral resolution:applications tostructural characterization of unfolded, partiallyfolded and folded proteins. J. Biomol. NMR, 9, 181-200.
Zhang, O. & Forman-Kay, J. D. (1997). NMR studies ofunfolded states of an SH3 domain in aqueous sol-ution and denaturating conditions. Biochemistry, 36,3959-3970.
Zheng, J., Cahill, S. M., Lemmon, M. A., Fushman, D.,Schlessinger, J. & Cowburn, D. (1996). Identi®cationof the binding site for acidic phospholipids on thePH domain of dynamin: implications for stimu-lation of GTPase activity. J. Mol. Biol. 255, 14-21.
Edited by A. R. Fersht
(Received 18 February 1999; received in revised form 24 May 1999; accepted 24 May 1999)





























