Embed Size (px)
Citation preview

This article was downloaded by: [Institut Pasteur]On: 15 August 2012, At: 23:29Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Avian PathologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/cavp20
Molecular study of the quasispecies evolution ofa typical pigeon paramyxovirus type 1 after serialpassages in pigeons by contactC. Barbezange a b & V. Jestin aa AFSSA (French Agency for Food Safety), Avian and Rabbit Virology Immunology andParasitology Unit, BP53, 22440 Ploufragan, Ploufragan, Franceb University of Cambridge, Centre for Veterinary Sciences, Department of VeterinaryMedicine, Madingley Road, Cambridge, CB3 0ES, UK
Version of record first published: 19 Oct 2010
To cite this article: C. Barbezange & V. Jestin (2005): Molecular study of the quasispecies evolution of a typical pigeonparamyxovirus type 1 after serial passages in pigeons by contact, Avian Pathology, 34:2, 111-122
To link to this article: http://dx.doi.org/10.1080/03079450500059321
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss, actions,claims, proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly orindirectly in connection with or arising out of the use of this material.

Molecular study of the quasispecies evolution of a typicalpigeon paramyxovirus type 1 after serial passages inpigeons by contact
C. Barbezange1,2 and V. Jestin1*
1AFSSA (French Agency for Food Safety), Avian and Rabbit Virology Immunology and Parasitology Unit, BP53, 22440Ploufragan, France, and 2Present address: University of Cambridge, Centre for Veterinary Sciences, Department ofVeterinary Medicine, Madingley Road, Cambridge CB3 0ES, UK
The quasispecies nature of a typical pigeon paramyxovirus type 1 (pPMV-1) was, for the first time, studiedunder conditions close to the natural infectious environment. The virus was serially passaged in pigeons bysuccessive contacts. Viral heterogeneity was analysed in the kidneys and brain of five pigeons from the lastcontact, by reverse transcriptase-polymerase chain reactions performed on RNA directly extracted from theorgan and targeting the P and HN genes of the virus. The viral diversity following in vivo passage was foundto be different from that in the inoculum, but demonstrated the reality of the quasispecies concept forpPMV-1 strains. Moreover, some aberrant genomic RNAs comprising insertions in the P gene editing site ordeletions in the HN gene were also detected, with possible consequences for the pathogenicity and infectivityof the virus.
Introduction
Pathogenic strains of avian paramyxovirus type 1(APMV-1 or NDV) are responsible for Newcastle disease(ND), a worldwide sanitary and economically importantdisease of poultry. Pigeon paramyxoviruses type 1(pPMV-1s) are antigenic variants of APMV-1 (Alexan-der et al. , 1984a, 1985; Collins et al. , 1989; Jestin et al. ,1989), which first appeared in the Middle East in the late1970s (Kaleta et al. , 1985) and are responsible for aparticular form of ND in the pigeon species. The pPMV-1 strains reached Europe in 1981 and were responsiblefor a panzootic in 1985. Despite various controlmeasures including vaccination, pPMV-1 infection re-mains enzootic in pigeons in many countries (Alexander,2001).
Moreover, pPMV-1 strains represent a current threatfor poultry species because they can infect and causepathology in chicken broilers, breeders and layers, asdemonstrated by the natural outbreaks described inGreat Britain in 1984 (Alexander et al. , 1984b) and in1996 in pheasants (Alexander et al. , 1997). However, thepathogenicity of pPMV-1 strains is not yet completelyunderstood.
The pPMV-1s, as all APMV-1s, belong to the newlycreated genus Avulavirus within the viral family Para-myxoviridae (see the website of the InternationalCommittee on Taxonomy for Viruses: http://www.dan-forthcenter.org/iltab/ICTVnet). The 15-kb non-segmen-ted single-stranded RNA genome of negative polarityencodes the six structural proteins (Millar & Emmerson,1988). The nucleoprotein (NP), phosphoprotein (P) andRNA-directed RNA polymerase (L) form the transcrip-
tional and replicative complex with the genomic RNA(Curran & Kolakofsky, 1999). The matrix protein (M) isinvolved in the budding process of the newly formedvirus particles (Peeples et al. , 1992), while the haemag-glutinin-neuraminidase (HN) and fusion protein (F),two glycoproteins anchored in the virion envelope, areinvolved in the binding and/or fusion to the host cellmembrane, and, for HN, in the release of the newlyformed virus particles (Yusoff & Tan, 2001). The P genealso encodes two non-structural proteins, called V andW, which seem to be involved in the pathogenesis of thevirus (Steward et al. , 1995; Mebatsion et al. , 2001; Parket al. , 2003).
The poor fidelity of the RNA-dependent RNA poly-merase of RNA viruses results in the coexistence ofclosely related but distinct variants called quasispecies(Domingo et al. , 1996; Domingo, 2002). RNA virusesare therefore a complex and dynamic population ofmutants. This structure is likely to be involved in thesurvival, evolution and pathogenesis of RNA viruses(Domingo & Holland, 1997; Domingo et al. , 2003). Theheterogeneity of the APMV-1s has been known since thelate 1950s (Granoff, 1964) and was shown in vitro usingthe plaque assay test performed on different cell culturesystems (Schloer & Hanson, 1968; Lomniczi, 1974).Kaleta et al. (1980) showed the presence of virulentsubpopulations within lentogenic strains of APMV-1,and a change of virulence was observed for strainsserially passaged in chickens by parental routes ofinoculation (intracerebral or intramuscular) (Alexander& Parsons, 1984, 1986; Islam et al. , 1994; Kommers
*To whom correspondence should be addressed. Tel: �/33 2 96 01 62 22. Fax: �/33 2 96 01 62 63. E-mail: [email protected]
Received 7 September 2004. Provisionally accepted 3 November 2004. Accepted 21 December 2004
Avian Pathology (April 2005) 34(2), 111�/122
ISSN 0307-9457 (print)/ISSN 1465-3338 (online) # 2005 Houghton Trust LtdDOI: 10.1080/03079450500059321
Dow
nloa
ded
by [
Inst
itut P
aste
ur]
at 2
3:29
15
Aug
ust 2
012

et al. , 2003). Kissi (1988) and Kissi & Lomniczi (1988)proposed a hypothesis for the influence of the quasis-pecies nature of pPMV-1 on pathogenicity based on theuse of Hungarian pPMV-1 strains passaged in embryo-nated chicken eggs. The authors suggested that pPMV-1was a heterogeneous population in the pigeon species,but following serial passages in chickens it could becomemore homogeneous and lead to the selection of morevirulent subpopulations. However, this hypothesis for theheterogeneity of pPMV-1 has never been proven at themolecular level, nor confirmed in in vivo experiments.
In order to gain a better understanding of pPMV-1pathogenicity in pigeons and chickens, we studied thequasispecies nature of a typical pPMV-1 strain beforeand after serial passages in pigeons by contact. First, weanalysed the quasispecies of the 99299 isolate, previouslyclassically characterized at the molecular level (Barbe-zange & Jestin, 2003a), in the P, F and HN genes becauseof their potential role in the pathogenicity. This allowedus to determine the initial heterogeneity status of theinoculum, since the strain was obtained as an infectiousallantoic fluid by the reference method of isolation(see Methods). Virus heterogeneity was then studiedmolecularly after six passages in pigeons. The kidneysand the brain were analysed as these two organs havebeen shown to be among the principal targets for thevirus. In fact, a previous study successfully reproducedthe disease with the 99299 isolate in conventional(i.e. non-specific pathogen free) pigeons placed in con-tact with infected ones (Barbezange & Jestin, 2003b).
Methods
Virus strain. pPMV-1 strain 99299 was isolated according to reference
methods (Anonymous, 1992; Picault, 1993a, 1993b) in 1999 in France
from an infected pigeon flock exhibiting green diarrhoea, torticollis and
mortality. This strain displayed an Intra Cerebral Pathogenicity Index
(ICPI) of 1.4, and was grouped in the P group of APMV-1 as defined by
Collins et al. (1989) using monoclonal antibodies. Its fusion cleavage
motif was 112RRQKRFIG119 (Barbezange & Jestin, 2003a). Therefore
this strain was classified as a typical pPMV-1 and was representative of
recent European isolates (Meulemans et al. , 2002). Putative specific
motifs of the pigeon variant strains were also described in the editing
site of the P gene and the intracellular domain of the HN gene
(Barbezange & Jestin, 2003a), which were included in the fragments
focused on in the present study.
Animals. Weaned, conventional hybrid 4-week-old meat pigeons were
obtained from a commercial breeding flock in Brittany, France, with no
significant disease history recorded. Parents in the loft were vaccinated
against pigeon paramyxovirosis once a year with a commercial oil-
adjuvated inactivated vaccine. The pigeons were maintained in quar-
antine for 4 weeks in an experimental biosafety BL3 containment
facility (‘‘quarantine BL3’’), where each pigeon tested negative for
APMV-1 antibodies using the haemagglutination inhibition test.
Experimental design. At the end of the quarantine period, 10 pigeons
were transferred into another biosafety BL3 containment facility
(‘‘infected BL3’’) and were inoculated oculo-nasally with 106 EID50
(mean egg infectious dose) per animal of 99299 isolate (Inoculated
group). Six days after inoculation, cloacal swabs were taken from all
animals and virus excretion was detected by a reverse transcriptase
(RT)-nested polymerase chain reaction (PCR) targeting the NP gene
(Barbezange & Jestin, 2002). The following day, 10 pigeons were
transferred from the quarantine BL3 and mixed with those inoculated
to form the Contact 1 group. Six days later, cloacal swabs were taken
from all the pigeons in the Contact 1 group, and examined for the
presence of viral RNA using the same RT-nested PCR. The next day,
the pigeons in the Contact 1 group were transferred to a clean cage in
the same infected BL3 and mixed with 10 susceptible pigeons (Contact
2) from the quarantine BL3. The remaining inoculated pigeons were
then killed. Five successive contacts (Contacts 1 to 5) were performed
following the same procedure (detection of infection and excretion by
the pigeons of the previous contact, change of cage, mixing birds
together, then killing the earlier group). At each passage, the clinical
signs were recorded.
At the end of the experiment (i.e. 6 days after the last contact), the
body condition of the Contact 5 pigeons was also evaluated. Their
kidneys and central nervous system (brain) were aseptically collected,
and 9 mm3 organ aliquots were stored at �/708C in 1.5 ml RNase-free
microtubes. The samples were first tested by the RT-nested PCR
targeting the NP gene to select five positive pigeons from the Contact
5 group. Then samples from these five selected pigeons were further
processed to analyse the quasispecies.
Characterization of the initial quasispecies nature of the 99299 isolate.
The infectious allantoic fluid used for inoculation was also used to
characterize the initial quasispecies status of the 99299 isolate. RNA
was extracted from 400 ml with the RNeasy† Mini kit (QIAGEN),
according to the manufacturer’s instructions. Reverse transcription was
carried out with MMLV reverse transcriptase and hexamers on 1 mg
RNA, and a part of the P, F and HN genes, amplified by PCR, were
used to analyse the heterogeneity (Figure 1). The three PCRs were
performed with Platinum† Taq DNA polymerase High Fidelity
(Invitrogen). The reaction mix contained 0.1 U/ml polymerase, 0.05
mM each dNTP, 2 mM MgSO4 and 0.5 mM each primer and a 10�1
dilution of the cDNA. For the P gene, a 700-base pair (bp) fragment
was amplified with primers Upa 5?-TGGCCACCTTTACAGATG-3?and Lpg 5?-AACGCCTCCATCATAGACAT-3?, using the following
conditions: 948C for 2 min, one cycle; 948C for 30 sec, 538C for 30 sec,
688C for 1 min 20 sec, 40 cycles. For the F gene, a 663-bp fragment was
amplified with primers Ufa 5?-TTAGAAAAAACACGGGTAGAA-3?and Lfc 5?-AATTTTTATACAGTCCAATTC-3?, using the same con-
ditions except 528C as annealing temperature and 1 min for the
polymerization step at 688C. The HN gene had a 642-bp fragment
amplified with primers Uhnc 5?-TTGCGCTAGAGAATGATGAA-3?and Lhnb 5?-AAAAGAATACCCTCCCTGTT-3?, using the same
conditions as for the F gene fragment.
The PCR products were purified using the QIAquick† Gel Extrac-
tion kit (QIAGEN) and cloned using the ZeroBlunt† TOPO† PCR
cloning Kit for sequencing (Invitrogen) according to the manufacturers’
instructions. Plasmid purifications were performed using the QIAprep†
Spin Miniprep kit (QIAGEN) according to the manufacturers’ instruc-
tions. Several clones with inserts (17 or 18) were sequenced in both
directions by the dye terminator method (ABI PRISM DyeTerminator
Cycle Sequencing Ready Reaction Kit; Applied Biosystems) on an
automatic DNA sequencer ABI 373XL (Applied Biosystems) with M13
forward and reverse universal primers and/or specific inner primers. The
sequence of each clone was assembled with the CAP program and
compared with the consensus sequences previously published
AJ415882, AJ306304, AJ415886 (Barbezange & Jestin, 2003a).
Characterization of the 99299 quasispecies nature following in vivo serial
passages. Frozen tissue samples were thawed and homogenized in 300 ml
TRIzol† reagent with a disposable pestle in a 1.5 ml microtube (Piston
Pellet; Merck Eurolab), before an additional 700 ml TRIzol† reagent
was added. After 20 sec vortexing and 10 min room-temperature
incubation, the samples were processed according to the manufacturer’s
instructions. The RNA pellet from each sample was dissolved in sterile
RNase-free water, spectrophotometrically quantified and stored at �/
708C.
Using total RNA extracted from organ aliquots, two direct RT-PCRs
were developed to target a shorter part of the P and HN genes (Figure
1). Reverse transcriptions were performed on 1 mg RNA according to
the manufacturer’s instructions, in a 20-ml reaction mix with MMLV
(Invitrogen) and specific primers targeting the viral genomic negative
strand: Upa 5?-TGGCCACCTTTACAGATG-3? for the P gene or
Uhn1 5?-ATCCGTTCTACCGCATCAC-3? for the HN gene. Both
PCRs were performed with Platinum† Taq DNA polymerase High
Fidelity (Invitrogen) with the mix containing 0.1 U/ml polymerase, 0.05
mM each dNTP, 2 mM MgSO4 and 0.4 mM each primer and a 10�1 or
112 C. Barbezange and V. Jestin
Dow
nloa
ded
by [
Inst
itut P
aste
ur]
at 2
3:29
15
Aug
ust 2
012

10�2 dilution of the cDNA. For the P gene, a 390-bp fragment was
amplified with the primers Upe 5?-TGGCGACACACAGCTCAAGA-
3? and Lpg 5?-AACGCCTCCATCATAGACAT-3?, and the following
programme was used: 948C for 2 min, one cycle; 948C for 30 sec, 558Cfor 30 sec, 688C for 40 sec, 50 cycles. For the HN gene, a 265-bp
fragment was amplified with the primers Uhnc 5?-TTGCGCTAGA-
GAATGATGAA-3? and Lhnh 5?-AGTGCCAGCGGAGATTCAAG-
3?, and the following programme was used: 948C for 2 min, one cycle;
948C for 30 sec, 53.58C for 30 sec, 688C for 30 sec, 50 cycles.
To confirm that these shorter PCRs are able to detect the initial
quasispecies of the inoculum, these differences in RT protocol (with
specific primers) and PCR technique (10�2 diluted cDNA) were also
applied to RNA extracted from the infectious allantoic fluid used as
inoculum.
The PCR products were purified, cloned, sequenced and compared
with the consensus sequences as already described.
Biological properties of the viral populations. The virus present in the
brain and kidneys from the selected pigeons in the Contact 5 group was
directly titrated by weighing and homogenizing an organ aliquot in 1 ml
phosphate-buffered saline. After clarification, the supernatant was 10-
fold serially diluted and used to inoculate chicken embryo kidney cells
plated in 96-well plates. The cytopathic effect was recorded daily, and
the titre was determined 4 days after cell infection using the method of
Reed & Muench (1938).
The virulence of the virus present in each organ was assessed by
sequencing PCR products targeting the cleavage motif of the F protein.
For this purpose, PCR products were obtained with Ufj 5?-CTCCAAACCCTACACCAGGA-3? and NDV425 5?-TACAAGC-
CAAACAAAATGC-3? primers on the RNA previously extracted for
the P and HN quasispecies analysis (RT performed using Ufa 5?-TTAGAAAAAACACGGGTAGAA-3? primer).
The virulence of each virus isolated from each organ was also
assessed. Once the virus isolation was performed (in embryonated
chicken eggs according to the reference method using a similar organ
aliquot), the ICPI was determined according to the international
reference method (Office International des Epizooties, 2000) using
1-day-old chickens. In addition, the F cleavage site of each isolate was
also determined using the RT-PCR already mentioned.
Finally, the pPMV-1 antigenic profile of each isolate (first adjusted to
4 haemagglutination units) was also checked by the haemagglutination
inhibition test (Picault, 1993a), using the specific monoclonal antibody
161/617 (Collins et al. , 1989).
Results
Characterization of the initial quasispecies nature of the99299 isolate. A PCR product of the F, HN and P geneswas cloned, and between 10 548 and 11 254 bases weresequenced for each PCR product (Table 1). Thus, 17 (forthe P gene) and 18 (for the HN and F genes) clones perPCR product were analysed and compared with theconsensus sequence previously published.
Less than one-third of mutant clones were obtainedfor the F gene, and 13 out of the 18 clones were identicalrepresenting the master sequence, which was found to beexactly the same as the consensus sequence. The viralpopulation was considered as homogeneous in the Fgene even though five mutants differed only by a single,but different, nucleotide from the consensus sequence.
The HN and P genes were each characterized by alarge proportion of mutant clones (83% and 88%,respectively). For HN (Figure 2), even if three clonesout of the 18 (ino.1.a) were identical to the consensus, nomaster sequence (i.e. identical sequence for at leastseveral, if not a majority of clones) was found. Nineclones (ino.1.b to j) showed one to three differencescompared with the consensus. The six remaining clonesdiffered from the consensus by a common mutation(C�/T at position 182 of the HN ORF), which resultedin a Thr to Ile substitution in the predictive proteinsequence. Two of those clones (ino.1.k) had only thismutation, while the four others (ino.1.l to o) exhibitedone to three additional nucleotide mutations. For P(Figure 3), two clones out of 17 (ino.1.a) were identicalto the consensus. Three clones (ino.1.b to d) were foundto be mutants from the consensus by one nucleotide.However, the master sequence, represented by fiveidentical clones (ino.1.e), differed from the consensusby two non-synonymous variations: G�/A at position281 (Gly to Glu substitution) and C�/T at position 413(Thr to Ile substitution) of the P ORF. The sevenremaining clones (ino.1.f to l) shared those two specificdifferences with the master sequence, and exhibited oneto four additional mutations.
Figure 1. Amplification, cloning and sequencing strategy used to analyse the quasispecies of the 99299 pPMV-1 isolate before and after
passages in pigeons. The top drawing represents the virus genome with the schematization of the six genes. The in vitro section shows the
portions of the P, F and HN genes targeted to analyse the initial quasispecies status of the 99299 isolate used as inoculum. The functional
sites are also marked. The in vivo section shows the parts of the P and HN genes focused on to analyse the evolution of the viral
quasispecies after passage in pigeons (arrows indicate the positions used to describe subpopulations in the inoculum).
In vivo quasispecies nature of pigeon PMV-1 113
Dow
nloa
ded
by [
Inst
itut P
aste
ur]
at 2
3:29
15
Aug
ust 2
012

Therefore, it was possible to consider two mainsubpopulations in both the HN and P genes of the99299 inoculum strain, defined by the positions 182 inthe HN gene and 281 and 413 in the P gene, respectively.
Serial passages in pigeons. The six serial passages(one inoculation followed by five contacts) were success-ful since it was possible to maintain the infection(as monitored by the NP-based RT-nested PCR)through the six groups of 10 pigeons each. Throughoutthe experiment, the morbidity remained under 50% andthe mortality approximately 20% to 30%. Although itwas possible to record typical signs of paramyxovirosisfor all the inoculated pigeons (tremors, prostration,diarrhoea and torticollis) and most of the typical signsfor the pigeons from the first three contacts (tremors,prostration and diarrhoea), no typical clinical signs werenoticed for the last two contacts. However, as shown inTable 2, pigeons 1 to 3 of the last contact displayedcachexia.
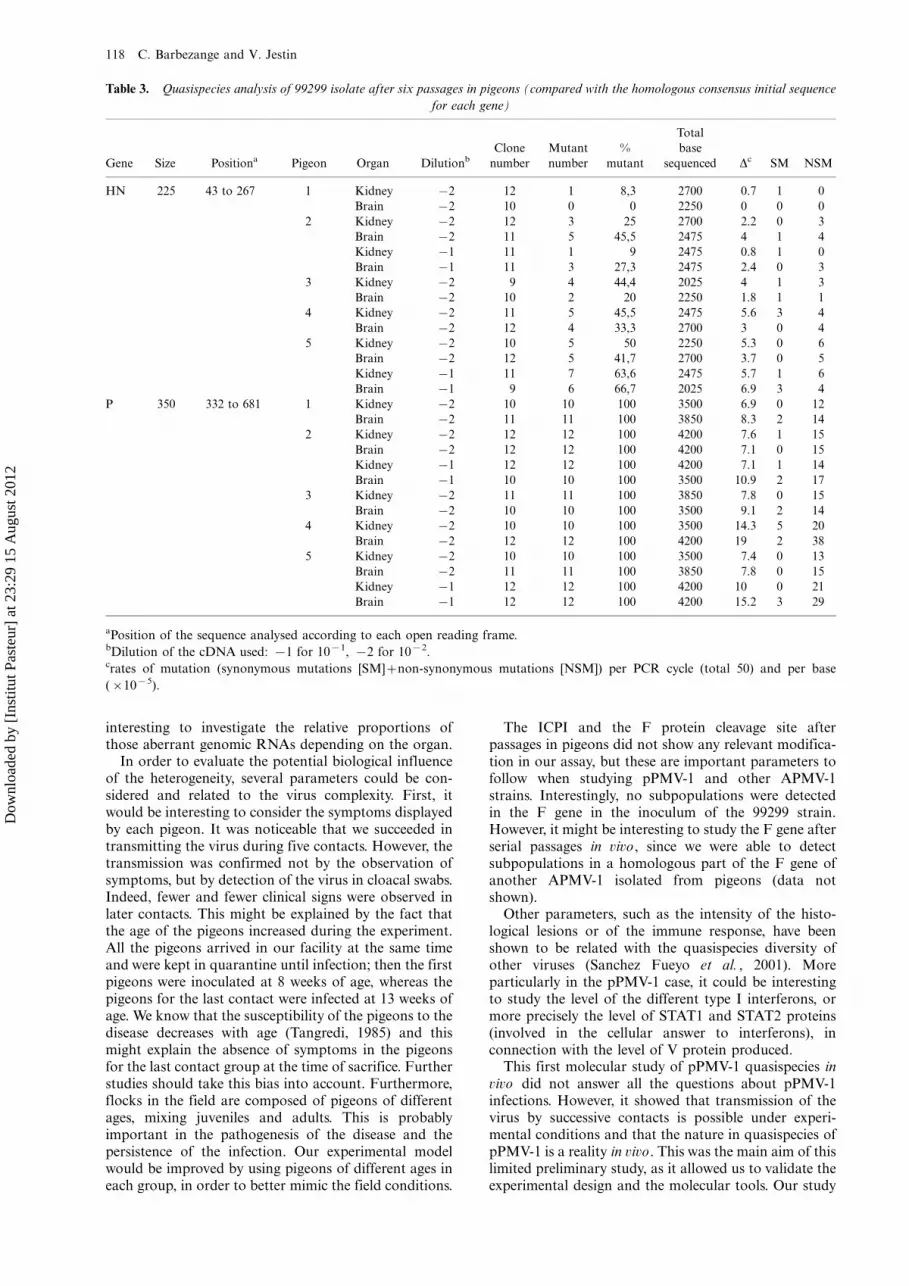
Characterization of the 99299 quasispecies nature after invivo serial passages. Two RT-PCRs were designed toamplify a shorter part of the HN and P genes (Figure 1),which would include positions 182 of the HN gene and413 of the P gene, respectively. The subpopulationsdescribed in the inoculum were also detected by theseshorter RT-PCRs, which used specific primers for the RTand 10�2 diluted cDNA for the PCR (Figures 2 and 3).The virus genome was then directly targeted by thosetwo specific RT-PCRs in brain and kidney samples offive pigeons from the last contact group (contact 5, i.e.sixth passage). Between nine and 12 clones for eachcloned PCR product were sequenced in both directionsand the sequence was determined without any ambiguity.Thus, between 2025 and 2700 bases for HN gene andbetween 3500 and 4200 bases for the P gene wereanalysed per organ sample (Table 3).
Overall, the mutation rate observed varied from 0 to6.9�/10�5 mutations per nucleotide per PCR cycle forthe HN clones, and from 6.9�/10�5 to 15.2�/10�5
mutations per nucleotide per PCR cycle for the P clones.Out of the two subpopulations defined in the inocu-
lum by position 413 in the P gene, only the dominantsubpopulation was recovered after passages in pigeons.Thus, the virus population appeared homogeneous atthis position with a thymidine residue for the fivepigeons analysed (Figure 4). Similarly in the HN gene,most of the clones analysed were characterized by acytosine residue at position 182. However, it was possibleto recover both subpopulations described at this position
in the inoculum for one sample, the kidney of pigeon 5(Figure 5).
Considering only the mutations shared by severalclones to define a subpopulation, it was then possible todefine new subpopulations compared with the inoculum,mainly in the P gene and for pigeons 4 and 5 (Figure 4).Thus, position 674 could be used to define subpopula-tions for both pigeons, whereas positions 385, 510 and626 defined subpopulations in the P gene mainly forpigeon 4. Moreover, some clones with insertions in theediting site of the P gene at position 402 (Figure 4) weredetected in four pigeons (1, 2, 3 and 5), mainly in brainsamples (10 clones for the brain samples versus four forthe kidney samples). Interestingly, eight clones (B.2.f,B.4.d and B.5.i) with deletions at position 148 in the HNgene (Figure 5) were detected for three pigeons (2, 4 and5) but only in brain samples.
Thus, to summarize, pigeons 4 and 5 displayed ahigher genetic heterogeneity for the viral populationspresent in their brain and kidneys (Table 2).
Biological properties of the viral populations. Neither avariation in the sequence of the cleavage site of the Fprotein of the virus nor an obvious modification in itsICPI could account for the difference in body conditiondisplayed by pigeons 1 to 3 compared with pigeons 4 and5 (Table 2).
Despite four passages in embryonated chicken eggs,no virus could be isolated from the brain of pigeons 3 to5. On the contrary, pPMV-1s (reacting with the specificmonoclonal antibody 161/617) were easily isolated fromthe brain of pigeons 1 and 2, and from the kidneys of allfive pigeons. However, the virus present in brain samplescould not be titrated directly using chicken embryokidney cells as no cytopathic effect being recorded.Considering the kidney samples only, pigeons 1 and 5displayed the highest and lowest virus titres, respectively,whereas intermediate virus titres were obtained forpigeons 2 to 4 (Table 2).
Discussion
The concept of the viral quasispecies offers interestingprospects to explain the pathogenesis and the evolutionof RNA viruses. However, the study of the subpopula-tions is subject to debate. One of the main points ofdiscussion is the way subpopulations should be de-scribed. Whereas Domingo and colleagues (Domingoet al. , 1996, 2003; Domingo & Holland, 1997; Ruiz-Jarabo et al. , 2000, 2002) consider each mutation todefine subpopulations, Smith et al. (1997) recommendlimiting the definition of the subpopulations to the
Table 1. Initial quasispecies analysis of the F, HN and P genes of the 99299 inoculum
Gene
Clone
number Sizea (bp) Positionb
Mutant
number
%
mutants
Total base
sequenced Dntc
Number
of SM
Number
of NSM
F 18 586 1 to 585 5 28 10548 1.2 1 4
HN 18 602 43 to 644 15 83 10836 6.2 3 24
P 17 662 20 to 681 15 88 11254 8.7 4 35
aLength of the sequence compared with the consensus.bPosition of the sequence analysed according to each open reading frame.cRates of mutations (synonymous mutations [SM]�/non-synonymous mutations [NSM]) per PCR cycle (total 40) and per base
(�/10�5).
114 C. Barbezange and V. Jestin
Dow
nloa
ded
by [
Inst
itut P
aste
ur]
at 2
3:29
15
Aug
ust 2
012

50 60 70 80 90 100 110 120 130 140 150 160 170. . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . .
99299 AGAGAAGCGAAGAGCACATGGCGCTTGGTTTTCCGGATCGCAGTTTTACTTTTAATAGTGGTGACCCTAGCTATCTCTGTAGCTGCCCTGGTATACAGCATGGGGGCTAGTACGCCCCGCGACCTCGTAGino.1.a (3) ..................................................................................................................................ino.1.b (1) ..................................................................................................................................ino.1.c (1) ..................................................................................................................................ino.1.d (1) ..................................................................................................................................ino.1.e (1) ..................................................................................................................................ino.1.f (1) ..................................................................................................................................ino.1.g (1) .............A....................................................................................................................ino.1.h (1) ..........................................................................................................G.......................ino.1.i (1) ..................................................................................................................................ino.1.j (1) ..................................................................................................................................ino.1.k (2) ..................................................................................................................................ino.1.l (1) ..................................................................................................................................ino.1.m (1) ..................................................................................................................................ino.1.n (1) ..................................................................................................................................ino.1.o (1) ..................................................................................................................................
ino.2.a (6) ..................................................................................................................................ino.2.b (3) ..................................................................................................................................ino.2.c (1) ..................................................................................................................................
180 190 200 210 220 230 240 250 260 270 280 290 300. . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . .
99299 GCATATCGACTATGGTTTCTAAGGCGGAGGACAGGATTACATCTTTACTCAGTTCAAACCAAGATGTAGTAGACAGAATATACAAGCAAGTGGCCCTTGAATCTCCGCTGGCACTACTAAATACTGAATCino.1.a (3) ..................................................................................................................................ino.1.b (1) ..................................................................................................................................ino.1.c (1) .........................................................G........................................................................ino.1.d (1) ................................................................................................................................C.ino.1.e (1) ..................................................................................................................................ino.1.f (1) ..................................................................................................................................ino.1.g (1) ..................................................................................................................................ino.1.h (1) ..................................................................................................................................ino.1.i (1) ............C.....................................................................................................................ino.1.j (1) ..................................................................................................................................ino.1.k (2) .........T........................................................................................................................ino.1.l (1) .........T........................................................................................................................ino.1.m (1) .........T........................................................................................................................ino.1.n (1) .........T.........................................................................T..............................................ino.1.o (1) .........T........................................................................................................................
ino.2.a (6) ............................................................................................... ino.2.b (3) .........T..................................................................................... ino.2.c (1) .......A.T.....................................................................................
310 320 330 340 350 360 370 380 390 400 410 420 430. . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . .
99299 TATAATCATGAATGCAATAACGTCTCTCTCTCATCAAATTAGCGGGGCTGCAAACAATAGCAGGTGTGGGGCGCCTGTTCATGACCCAGATTATATCGGGGGGATAGGTAAAGAACTCATAGTAGACGACino.1.a (3) ..................................................................................................................................ino.1.b (1) ..................................................................................................................................ino.1.c (1) ..................................................................................................................................ino.1.d (1) ..................................................................................................................................ino.1.e (1) ..................................................................................................................................ino.1.f (1) ..................................................................................................................................ino.1.g (1) ..............................................................................................T...................................ino.1.h (1) .............................................................................................................G....................ino.1.i (1) ..................................................................................................................................ino.1.j (1) .............................................................................G.............................T......................ino.1.k (2) ..................................................................................................................................ino.1.l (1) ....................................................................................................................C.............ino.1.m (1) ..................................................................................................................................ino.1.n (1) ..................................................................................................................................ino.1.o (1) ..................................................................................................................................
ino.2.a (6) ino.2.b (3) ino.2.c (1)
440 450 460 470 480 490 500 510 520 530 540 550 560. . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . .
99299 ATCAGTGGTGTCACATCATTCTACCCATCTGCATACCAAGAACACCTGAATTTTATCCCAGCGCCCACTACAGGATCAGGCTGCACTCGGATACCCTCATTTGATATGAGTGCTACCCACTATTGTTATAino.1.a (3) ..................................................................................................................................ino.1.b (1) ............G.....................................................................................................................ino.1.c (1) ..................................................................................................................................ino.1.d (1) ..................................................................................................................................ino.1.e (1) ..................................................................................................................................ino.1.f (1) ........................................G.........................................................................................ino.1.g (1) ..................................................................................................................................ino.1.h (1) ..................................................................................................................................ino.1.i (1) ..................................................................................................................................ino.1.j (1) ..........................................................................................................C.......................ino.1.k (2) ..................................................................................................................................ino.1.l (1) ..................................................................................................................................ino.1.m (1) ..................................................................................................................................ino.1.n (1) ..................................................................................................................................ino.1.o (1) ...............................................................................A..................................................
ino.2.a (6) ino.2.b (3) ino.2.c (1)
570 580 590 600 610 620 630 640. . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . .
99299 CTCATAACGTGATATTGTCTGGCTGTATAGATCACTCACATTCACATCAATACTTAGCACTTGGTGTGCTTCGGACATCTGCino.1.a (3) ..................................................................................ino.1.b (1) ..................................................................................ino.1.c (1) ..................................................................................ino.1.d (1) ..................................................................................ino.1.e (1) ...G..............................................................................ino.1.f (1) ..................................................................................ino.1.g (1) ..................................................................................ino.1.h (1) ..................................................................................ino.1.i (1) ............................G.....................................................ino.1.j (1) ..................................................................................ino.1.k (2) ..................................................................................ino.1.l (1) ..................................................................................ino.1.m (1) .C................................................................................ino.1.n (1) ..................................................................................ino.1.o (1) ............C...........................C.....C...................................
ino.2.a (6) ino.2.b (3) ino.2.c (1)
182
Figure 2. Alignment of clone sequences obtained for the HN gene of the 99299 strain inoculum. The published consensus sequence
(AJ415886) of 99299 is used as a reference, and only differences from it are shown for the clones. Each unique clone sequence was given a
codification including ino (for inoculum), 1 or 2 (for the dilution of the cDNA used for the PCR, 1 for 10�1 and 2 for 10�2) and a
lowercase letter. Then, the number of clones found for each unique sequence is indicated in brackets. Position 182, used to define
subpopulations, is highlighted by an arrow.
In vivo quasispecies nature of pigeon PMV-1 115
Dow
nloa
ded
by [
Inst
itut P
aste
ur]
at 2
3:29
15
Aug
ust 2
012

20 30 40 50 60 70 80 90 100 110 120 130| . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . .
99299 CTGAGATAGAGGATATATTTGAGACCAGTGGAACTGTCATTGACAGTATAATTACGGCCCAGGGCAGATCAGCAGAGACTGTCGGGAAGAGCGCAATCCCGCAAGGCAAGACAAAGGCCCino.1.a (2) ........................................................................................................................ino.1.b (1) ........................................................................................................................ino.1.c (1) ........................................................................................................................ino.1.d (1) ........................................................................................................................ino.1.e (5) ........................................................................................................................ino.1.f (1) ........................................................................................................................ino.1.g (1) ........................................................................................................................ino.1.h (1) ........................................................................................................................ino.1.i (1) ........................................................................................................................ino.1.j (1) ........................................................................................................................ino.1.k (1) ...............................................................................................G........................ino.1.l (1) .................C......................................................................................................
ino.2.a (4) ------------------------------------------------------------------------------------------------------------------------ino.2.b (6) ------------------------------------------------------------------------------------------------------------------------ino.2.c (1) ------------------------------------------------------------------------------------------------------------------------
140 150 160 170 180 190 200 210 220 230 240 250| . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . .
99299 TGAGCATAGCACGGGAGAAGCACGGGAGCATCCAGCCATCCGCCGGCCAGGACACCTCTAACCAACAAGACAAACCAGACAAACAGCCACCCGCATCCGAGCAGGTGACTCCACACAACGino.1.a (2) ........................................................................................................................ino.1.b (1) ........................................................................................................................ino.1.c (1) ........................................................................................................................ino.1.d (1) ........................................................................................................................ino.1.e (5) ........................................................................................................................ino.1.f (1) ........................................................................................................................ino.1.g (1) ........................................................................................................................ino.1.h (1) ........................................................................................................................ino.1.i (1) ................................................................................G.......................................ino.1.j (1) ........................................................................................................................ino.1.k (1) ........................................................................................................................ino.1.l (1) .............................................................................C..........................................
ino.2.a (4) ------------------------------------------------------------------------------------------------------------------------ino.2.b (6) ------------------------------------------------------------------------------------------------------------------------ino.2.c (1) ------------------------------------------------------------------------------------------------------------------------
260 270 280 290 300 310 320 330 340 350 360 370| . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . .
99299 ATCCTCTGACCGCACCCAAAGGACCGCTCCCCACTCAGGCCGCAGGTGATGCTGGCGACACACAGCTCAAGACCGGAGCAAGCAACTCTCTTTTGTCAATGCTTGACAAGCTGAGCAATAino.1.a (2) ........................................................................................................................ino.1.b (1) ......................................................................................C.................................ino.1.c (1) ........................................................................................................................ino.1.d (1) .....................A..................................................................................................ino.1.e (5) .....................A..................................................................................................ino.1.f (1) .....................A..................................................................................................ino.1.g (1) .....................A......................................................G...........................................ino.1.h (1) .....................A..................................................................................................ino.1.i (1) .....................A..................................................................................................ino.1.j (1) .....................A....................................................................C.............................ino.1.k (1) .....................A..................................................................................................ino.1.l (1) .....................A.......................................T.T........................................................
ino.2.a (4) ------------------------------------------------------------------------................................................ino.2.b (6) ------------------------------------------------------------------------................................................ino.2.c (1) ------------------------------------------------------------------------................................................
380 390 400 410 420 430 440 450 460 470 480 490| . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . .
99299 AATCGTCTAATGCTAAAAAGGGGCCAAGGTCGACCCCCCAGGAGGGCCACCACCAACCTCCGGCCCAACAACACGGGAATCAGTCGAGCCGTGGAAACAACCAGGAAGGACCGCAGCATCino.1.a (2) ........................................................................................................................ino.1.b (1) ........................................................................................................................ino.1.c (1) ........................................................................................................................ino.1.d (1) ........................................................................................................................ino.1.e (5) .................................T......................................................................................ino.1.f (1) .................................T.............................T........................................................ino.1.g (1) .................................T......................................................................................ino.1.h (1) .................................T......................................................................................ino.1.i (1) .................................T......................................................................................ino.1.j (1) .................................T......................................................................................ino.1.k (1) .................................T.............................................C........................................ino.1.l (1) .................................T......................................................................................
ino.2.a (4) ........................................................................................................................ino.2.b (6) .................................T......................................................................................ino.2.c (1) .................................T..........A...........................................................................
500 510 520 530 540 550 560 570 580 590 600 610| . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . .
99299 AGGCCAAAGCTGCCCCTGGAAGCCGGGGCACAGACGCGAGCACAGCATATCATGGACAATGGAAGGAGTCACAACTATCAGCTGGTGCAACCCCTCATGTGCTCCAGTCAGGGCGGAGCCino.1.a (2) ........................................................................................................................ino.1.b (1) ........................................................................................................................ino.1.c (1) .............G..........................................................................................................ino.1.d (1) ........................................................................................................................ino.1.e (5) ........................................................................................................................ino.1.f (1) ........................................................................................................................ino.1.g (1) ........................................................................................................................ino.1.h (1) ........................................................................................................................ino.1.i (1) ........................................................................................................................ino.1.j (1) ........................................................................................................................ino.1.k (1) ........................................................................................................................ino.1.l (1) ........................................................................................................................
ino.2.a (4) ........................................................................................................................ino.2.b (6) ........................................................................................................................ino.2.c (1) ........................................................................................................................
620 630 640 650 660 670 680| . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | .
99299 AAGACAGTACTCCTGTACCTGTGGAGCATGTCCAGCTACCTGTCGACTTTGTGCAGGCGATGino.1.a (2) ..............................................................ino.1.b (1) ..............................................................ino.1.c (1) ..............................................................ino.1.d (1) ..............................................................ino.1.e (5) ..............................................................ino.1.f (1) ..............................................................ino.1.g (1) ..............................................................ino.1.h (1) ..........................................................C...ino.1.i (1) ..............................................................ino.1.j (1) ..............................................................ino.1.k (1) ..............................................................ino.1.l (1) .....................................................T........
ino.2.a (4) ..............................................................ino.2.b (6) ..............................................................ino.2.c (1) ..............................................................
281
413
Figure 3. Alignment of clone sequences obtained for the P gene of the 99299 strain inoculum. The published consensus sequence
(AJ415882) of 99299 is used as a reference, and only differences from it are shown for the clones. Each unique clone sequence was given a
codification including ino (for inoculum), 1 or 2 (for the dilution of the cDNA used for the PCR, 1 for 10�1 and 2 for 10�2) and a
lowercase letter. Then, the number of clones found for each unique sequence is indicated in brackets. Arrows used to highlight the positions
used to define subpopulations.
116 C. Barbezange and V. Jestin
Dow
nloa
ded
by [
Inst
itut P
aste
ur]
at 2
3:29
15
Aug
ust 2
012

obvious mutations found in several clones. Since ourstudy is the first to analyse the pPMV-1 quasispecies atthe molecular level, we were cautious in our definition ofthe subpopulations and heterogeneity by taking intoaccount only the mutations found in several clones.However, the position and nature of all the mutationsfound are displayed on Figure 2, 3, 4 and 5.
Most published studies on viral quasispecies usedclassical Taq polymerase and nested PCRs to amplifythe targeted virus genome, and between 10 and 20 clonesfor each sample. The use of a proof-reading polymeraseand of direct PCRs in our study limited error incorpora-tion during the amplification, and the mutation rates forthe heterogeneous samples were found to be 10-foldabove the error rate of the high fidelity polymerases(Mullan et al. , 2001; Malet et al. , 2003).
Our work provides original observations furthersupporting the nature in quasispecies of pPMV-1 invivo. First, it is noticable that only one of the two initialsubpopulations described in the P gene was recoveredafter serial passages in pigeons, and that the two initialsubpopulations in the HN gene were only detected inone sample from one pigeon. This could not only be dueto the conditions of the PCR, as it was possible to detectthe P and HN subpopulations in the inoculum even byusing cDNA diluted at 10�2 (Figures 2 and 3). Themissing subpopulations might then represent a memoryfor the quasispecies (Ruiz-Jarabo et al. , 2000) and beundetectable by the technique used, or simply havedisappeared during the contact passages (Kissi et al. ,1999). Each contact would need to be studied to betterunderstand this phenomenon. The fragments of the Pand HN genes targeted for the quasispecies study afterpassages in pigeons were smaller than those used beforepassage in vivo. The aim was to limit the number ofsequencing reactions to two per clone (one forward andone reverse), but of course this might lead to loss ofinformation. However, the description of other novelsubpopulations showed that the quasispecies equilibriumwas modified in vivo. Nevertheless, a great variabilitywas noticed between pigeons in the viral heterogeneity
level and in the subpopulations found. Such an indivi-
dual host variability has already been described for other
virus quasispecies (Gretch & Polyak, 1997; Hotzel et al. ,2002) and has to be taken into account in further studies.
Interestingly, insertions in the P gene and deletions inthe HN gene were observed more frequently in the brain
samples. These insertions and deletions were not sequen-
cing errors, as the electrophoregrams showed clearsignals (data not shown). Differences in the quasispecies
complexity according to the organ were observed for
other viruses (Laskus et al. , 2000; Ruster et al. , 2001).Depending on the cell type, some mechanisms might
occur more or less rapidly after infection, modifying the
concentration of ions or other oligo-elements and so,disrupting the working of the viral polymerase (Shap-
shak et al. , 1999). The insertions in the P gene editing
site on the viral genomic strand might be due to thestuttering of the polymerase by a similar mechanism to
that responsible for the appearance of mRNA coding for
the V and W proteins (Steward et al. , 1993). Theseinsertions might have an influence on the level of V and
W proteins produced in the infected cells with conse-
quences on pathogenesis, since the V protein is believedto play a role in inhibiting the host interferon response
(Kato et al. , 1997; Delenda et al. , 1998; Komatsu et al. ,
2004). The deletions in the HN gene introduce apremature stop codon after the transmembrane domain
of the protein. Therefore, truncated HN proteins might
be expressed at the surface of the cells and incorporatedinto newly formed virus particles, possibly with con-
sequences for the binding abilities and the infectivity of
the virus (Huang et al. , 2004). However, the questionarises whether these aberrant and defective genomic
RNAs, detected inside the infected cells, are actually
packaged into the newly formed virus particles, excretedand transmitted to susceptible hosts, as shown in other
models (Cabot et al. , 2000; Kissi et al. , 1999; Wang et
al. , 2003). This question could be assessed by studyingthe virus heterogeneity after virus purification from both
organs and cloacal and tracheal swabs. It would also be
Table 2. Biological properties of the viral populations present in the kidneys and brain of the pigeons of the sixth passage, and
relationship with their body condition
Isolation (embryonated chicken eggs)
Pigeon
Body
condition Organ Heterogeneitya Virus titreb
F cleavage
site (organ)
Passage ICPI F cleavage site
(allantoic fluid)
1 Cachexia Kidney �/ 104.5 RRQKR/FIG 1 1.3 RRQKR/FIG
Brain �/ No CPE RRQKR/FIG 1 1.3 RRQKR/FIG
2 Cachexia Kidney �/ 103.3 RRQKR/FIG 1 1.3 RRQKR/FIG
Brain �/�/ No CPE RRQKR/FIG 2 1.2 RRQKR/FIG
3 Cachexia Kidney �/ 102.7 RRQKR/FIG 1 1.4 RRQKR/FIG
Brain �/ No CPE RRQKR/FIG Negc �/d �/
4 Good Kidney �/�/ 102.45 RRQKR/FIG 1 1.2 RRQKR/FIG
Brain �/�/ No CPE RRQKR/FIG Neg �/ �/
5 Good Kidney �/�/ 101.6 RRQKR/FIG 1 1.2 RRQKR/FIG
Brain �/�/�/ No CPE RRQKR/FIG Neg �/ �/
aCodification of the virus heterogeneity considering both genes: �/, few or no mutation; �/, few mutations and deletion in HN or
insertion in P but no clear subpopulations; �/�/, more mutations and deletion in HN or insertion in P and clear subpopulation; �/�/�/
, more mutations and deletion in HN and insertion in P and clear subpopulation.bVirus titre in the organ, expressed as TCID50/mg (mean infectious dose for tissue culture, using chicken embryo kidney cells, per
milligram of organ). No CPE, no cytopathic effect observed.cNo haemagglutination activity detected after four passages in embryonated chicken eggs.dNot done.
In vivo quasispecies nature of pigeon PMV-1 117
Dow
nloa
ded
by [
Inst
itut P
aste
ur]
at 2
3:29
15
Aug
ust 2
012
interesting to investigate the relative proportions ofthose aberrant genomic RNAs depending on the organ.
In order to evaluate the potential biological influenceof the heterogeneity, several parameters could be con-sidered and related to the virus complexity. First, itwould be interesting to consider the symptoms displayedby each pigeon. It was noticeable that we succeeded intransmitting the virus during five contacts. However, thetransmission was confirmed not by the observation ofsymptoms, but by detection of the virus in cloacal swabs.Indeed, fewer and fewer clinical signs were observed inlater contacts. This might be explained by the fact thatthe age of the pigeons increased during the experiment.All the pigeons arrived in our facility at the same timeand were kept in quarantine until infection; then the firstpigeons were inoculated at 8 weeks of age, whereas thepigeons for the last contact were infected at 13 weeks ofage. We know that the susceptibility of the pigeons to thedisease decreases with age (Tangredi, 1985) and thismight explain the absence of symptoms in the pigeonsfor the last contact group at the time of sacrifice. Furtherstudies should take this bias into account. Furthermore,flocks in the field are composed of pigeons of differentages, mixing juveniles and adults. This is probablyimportant in the pathogenesis of the disease and thepersistence of the infection. Our experimental modelwould be improved by using pigeons of different ages ineach group, in order to better mimic the field conditions.
The ICPI and the F protein cleavage site afterpassages in pigeons did not show any relevant modifica-tion in our assay, but these are important parameters tofollow when studying pPMV-1 and other APMV-1strains. Interestingly, no subpopulations were detectedin the F gene in the inoculum of the 99299 strain.However, it might be interesting to study the F gene afterserial passages in vivo, since we were able to detectsubpopulations in a homologous part of the F gene ofanother APMV-1 isolated from pigeons (data notshown).
Other parameters, such as the intensity of the histo-logical lesions or of the immune response, have beenshown to be related with the quasispecies diversity ofother viruses (Sanchez Fueyo et al. , 2001). Moreparticularly in the pPMV-1 case, it could be interestingto study the level of the different type I interferons, ormore precisely the level of STAT1 and STAT2 proteins(involved in the cellular answer to interferons), inconnection with the level of V protein produced.
This first molecular study of pPMV-1 quasispecies invivo did not answer all the questions about pPMV-1infections. However, it showed that transmission of thevirus by successive contacts is possible under experi-mental conditions and that the nature in quasispecies ofpPMV-1 is a reality in vivo. This was the main aim of thislimited preliminary study, as it allowed us to validate theexperimental design and the molecular tools. Our study
Table 3. Quasispecies analysis of 99299 isolate after six passages in pigeons (compared with the homologous consensus initial sequence
for each gene)
Gene Size Positiona Pigeon Organ Dilutionb
Clone
number
Mutant
number
%
mutant
Total
base
sequenced Dc SM NSM
HN 225 43 to 267 1 Kidney �/2 12 1 8,3 2700 0.7 1 0
Brain �/2 10 0 0 2250 0 0 0
2 Kidney �/2 12 3 25 2700 2.2 0 3
Brain �/2 11 5 45,5 2475 4 1 4
Kidney �/1 11 1 9 2475 0.8 1 0
Brain �/1 11 3 27,3 2475 2.4 0 3
3 Kidney �/2 9 4 44,4 2025 4 1 3
Brain �/2 10 2 20 2250 1.8 1 1
4 Kidney �/2 11 5 45,5 2475 5.6 3 4
Brain �/2 12 4 33,3 2700 3 0 4
5 Kidney �/2 10 5 50 2250 5.3 0 6
Brain �/2 12 5 41,7 2700 3.7 0 5
Kidney �/1 11 7 63,6 2475 5.7 1 6
Brain �/1 9 6 66,7 2025 6.9 3 4
P 350 332 to 681 1 Kidney �/2 10 10 100 3500 6.9 0 12
Brain �/2 11 11 100 3850 8.3 2 14
2 Kidney �/2 12 12 100 4200 7.6 1 15
Brain �/2 12 12 100 4200 7.1 0 15
Kidney �/1 12 12 100 4200 7.1 1 14
Brain �/1 10 10 100 3500 10.9 2 17
3 Kidney �/2 11 11 100 3850 7.8 0 15
Brain �/2 10 10 100 3500 9.1 2 14
4 Kidney �/2 10 10 100 3500 14.3 5 20
Brain �/2 12 12 100 4200 19 2 38
5 Kidney �/2 10 10 100 3500 7.4 0 13
Brain �/2 11 11 100 3850 7.8 0 15
Kidney �/1 12 12 100 4200 10 0 21
Brain �/1 12 12 100 4200 15.2 3 29
aPosition of the sequence analysed according to each open reading frame.bDilution of the cDNA used: �/1 for 10�1, �/2 for 10�2.crates of mutation (synonymous mutations [SM]�/non-synonymous mutations [NSM]) per PCR cycle (total 50) and per base
(�/10�5).
118 C. Barbezange and V. Jestin
Dow
nloa
ded
by [
Inst
itut P
aste
ur]
at 2
3:29
15
Aug
ust 2
012

340 350 360 370 380 390 400 410 420 430 440 450 460 470 480 490 500 510. . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | .
99299 CCGGAGCAAGCAACTCTCTTTTGTCAATGCTTGACAAGCTGAGCAATAAATCGTCTAATGCTAAAAAGGGG---CCAAGGTCGACCCCCCAGGAGGGCCACCACCAACCTCCGGCCCAACAACACGGGAATCAGTCGAGCCGTGGAAACAACCAGGAAGGACCGCAGCATCAGGCCAAAG
K.1.a (8) .......................................................................---..........T...............................................................................................K.1.b (1) .......................................................................---..........T....................G..........................................................................K.1.c (1) .......................................................................---..........T...............................................................................................B.1.a (6) .......................................................................---..........T...............................................................................................B.1.b (1) ..................................................................G....---..........T...............................................................................................B.1.c (1) .......................................................................---..........T....................................................................G..........................B.1.d (2) .......................................................................---..........T...........................................................................G...................B.1.e (1) .......................................................................G--..........T...............................................................................................
K.2.a (17) .......................................................................---..........T...............................................................................................K.2.b (1) ...............................C.......................................---..........T...............................................................................................K.2.c (1) .......................................................................---..........T...............................................G...............................................K.2.d (1) .......................................................................---..........T...............................................................................................K.2.e (1) ..................................................C....................---..........T...............................................................................................K.2.f (1) .......................................................................---..........T...............................................................................................K.2.g (1) .......................................................................GGG..........T...............................................................................................B.2.a (13) .......................................................................---..........T...............................................................................................B.2.b (1) ........................................................G..............---..........T...............................................................................................B.2.c (1) .......................................................................---..........T...............................................................................................B.2.d (1) .......................................................................---..........T...............................................................................................B.2.e (1) .......................................................................---..........T...............................................................................................B.2.f (3) .......................................................................G--..........T...............................................................................................B.2.g (2) .......................................................................GG-..........T...............................................................................................
K.3.a (9) .......................................................................---..........T...............................................................................................K.3.b (1) .......................................................................G--..........T...............................................................................................K.3.c (1) .......................................................................GG-..........T...............................................................................................B.3.a (5) .......................................................................---..........T...............................................................................................B.3.b (1) ..............C........................................................---..........T...............................................................................................B.3.c (1) ..............................G........................................---..........T...............................................................................................B.3.d (1) .......................................................................---..........T...............................................................................................B.3.e (1) .......................................................................---..........T..........................................A....................................................B.3.f (1) .......................................................................G--..........T...............................................................................................
K.4.a (3) .......................................................................---..........T...............................................................................................K.4.b (1) .................................G.....................................---..........T...............................................................................................K.4.c (1) .......................................................................---..........T...............................................................................................K.4.d (1) ..............................C........................................---..........T............................................G..................................................K.4.e (2) .......................................................................---..........T...............................................................................................K.4.f (1) .......................................................................---..........T...............................................................................................K.4.g (1) .......................................................................---..........T...............................................................................................B.4.a (9) .....................................................C.................---..........T...............................................................................................B.4.b (1) .......................C.............................C.................---..........T...............................................................................................B.4.c (1) .....................................................C.................---..........T...............................................................................................B.4.d (1) .....................................................C.................---..........T...............................................................................................
K.5.a (12) .......................................................................---..........T...............................................................................................K.5.b (2) .......................................................................---..........T...............................................................................................K.5.c (1) ..............................G........................................---..........T...............................................................................................K.5.d (1) .......................................................................---..........T...............................................................................................K.5.e (1) .......................................................................---..........T...............................................................................................K.5.f (1) .......................................................................---..........T...............................................................................................K.5.g (2) .......................................................................---..........T...............................................................................................K.5.h (1) .....................................................C.................---..........T...............................................................................................K.5.i (1) .......................................................................G--..........T...............................................................................................B.5.a (8) .......................................................................---..........T...............................................................................................B.5.b (1) .......................................................................---..........T..............G................................................................................B.5.c (1) .......................................................................---..........T...............................................................................................B.5.d (1) .......................................................................---..........T...............................................................................................B.5.e (5) .......................................................................---..........T...............................................................................................B.5.f (1) .......................................................................---..........T......................................................................................T........B.5.g (1) .......................................................................---..........T......................................G........................................................B.5.h (1) .......................................................................---..........T...............................................................................................B.5.i (1) .......................................................................---..........T...................T........................................G..................................B.5.j (2) .......................................................................G--..........T...............................................................................................B.5.k (1) .......................................................................G--..........T...............................................................................................
520 530 540 550 560 570 580 590 600 610 620 630 640 650 660 670 680. . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . .
99299 CTGCCCCTGGAAGCCGGGGCACAGACGCGAGCACAGCATATCATGGACAATGGAAGGAGTCACAACTATCAGCTGGTGCAACCCCTCATGTGCTCCAGTCAGGGCGGAGCCAAGACAGTACTCCTGTACCTGTGGAGCATGTCCAGCTACCTGTCGACTTTGTGCAGGCGATG
K.1.a (8) .............................................................................................................................................................................K.1.b (1) .............................................................................................................................................................................K.1.c (1) ..................................................C..........................................................................................................................B.1.a (6) .............................................................................................................................................................................B.1.b (1) .............................................................................................................................................................................B.1.c (1) .............................................................................................................................................................................B.1.d (2) .............................................................................................................................................................................B.1.e (1) .............................................................................................................................................................................
K.2.a (17) .............................................................................................................................................................................K.2.b (1) .............................................................................................................................................................................K.2.c (1) .............................................................................................................................................................................K.2.d (1) .....................................................................................................................................................................G.......K.2.e (1) ..................................................................................................................G..........................................................K.2.f (1) ..................A......................................................................................................C...................................................K.2.g (1) .............................................................................................................................................................................B.2.a (13) .............................................................................................................................................................................B.2.b (1) .............................................................................................................................................................................B.2.c (1) ................................................................................................................G............................................................B.2.d (1) ......................................C.......................................................................................C............C.......C.........................B.2.e (1) ....................................................................---......................................................................................................B.2.f (3) .............................................................................................................................................................................B.2.g (2) .............................................................................................................................................................................
K.3.a (9) .............................................................................................................................................................................K.3.b (1) ................................................................................................................G............................................................K.3.c (1) .............................................................................................................................................................................B.3.a (5) .............................................................................................................................................................................B.3.b (1) .............................................................................................................................................................................B.3.c (1) .............................................................................................................................................................................B.3.d (1) ....................................................................................................G........................................................................B.3.e (1) ........................G....................................................................................................................................................B.3.f (1) .............................................................................................................................................................................
K.4.a (3) .............................................................................................................................................................................K.4.b (1) .............................................................................................................................................................................K.4.c (1) .......................................................................................................................................................C.....................K.4.d (1) .............................................................................................................................................................................K.4.e (2) .C...................................................................................................................A.......................................................K.4.f (1) .C...................................................................................................................A........................................C..............K.4.g (1) .C.........................................G.........................................................................A................................................C......B.4.a (9) .....................................................................................................................................................................G.......B.4.b (1) .....................................................................................................................................................................G.......B.4.c (1) .........................................................................................................................C...........................................G.......B.4.d (1) ........................................................A...........................................................................................G................G.......
K.5.a (12) .............................................................................................................................................................................K.5.b (2) ..........................................G..................................................................................................................................K.5.c (1) .............................................................................................................................................................................K.5.d (1) ......................................................G......................................................................................................................K.5.e (1) ...........................................................C..........................................................................................................C......K.5.f (1) ............................................................................................T................................................................................K.5.g (2) .....................................................................................................................................................................G.......K.5.h (1) .....................................................................................................................................................................G.......K.5.i (1) .............................................................................................................................................................................B.5.a (8) .............................................................................................................................................................................B.5.b (1) .............................................................................................................................................................................B.5.c (1) ...........................................................................................................................................................A.................B.5.d (1) .C...........................................................................................................................................................................B.5.e (5) .....................................................................................................................................................................G.......B.5.f (1) .....................................................................................................................................................................G.......B.5.g (1) .............................G.......................................................................................................................................G.......B.5.h (1) .....................................................................................A............C..................................................................G.......B.5.i (1) ....................G................................................................................................................................................G.......B.5.j (2) .............................................................................................................................................................................B.5.k (1) .....................................................................................C.......................................................................................
510 626
413402385
674
Figure 4. Alignment of clone sequences obtained for the P gene of the 99299 strain after serial passages in pigeons. As in Figure 3, each
sequence is compared with the published consensus sequence (AJ415882) of 99299, and only differences from it are shown for the clones.
Due to the presence of one to three nucleotide insertions for some clones, the numbering is shifted forward by 3 from position 402, therefore
position 406 on the figure is in fact position 403 on the gene. Each unique clone sequence was given a codification including the organ
(K, kidney; B, brain), the pigeons (numbered 1 to 5) and a lowercase letter. Then, the number of clones found for each unique sequence is
indicated in brackets. Arrows used to point out the remarkable positions mentioned in the text.
In vivo quasispecies nature of pigeon PMV-1 119
Dow
nloa
ded
by [
Inst
itut P
aste
ur]
at 2
3:29
15
Aug
ust 2
012

50 60 70 80 90 100 110 120 130 140 150 160 170. . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . .
99299 AGAGAAGCGAAGAGCACATGGCGCTTGGTTTTCCGGATCGCAGTTTTACTTTTAATAGTGGTGACCCTAGCTATCTCTGTAGCTGCCCTGGTATACAGCATGGGGGCTAGTACGCCCCGCGACCTCGTAG
K.1.a (11) ..................................................................................................................................K.1.b (1) ..................................................................................................................................B.1.a (10) ..................................................................................................................................
K.2.a (19) ..................................................................................................................................K.2.b (1) ...............G..................................................................................................................K.2.c (1) ............................................................................................................................C.....K.2.d (1) ..................................................................................................................................K.2.e (1) ..............................................................................................................C...................B.2.a (14) ..................................................................................................................................B.2.b (2) ..................................................................................................................................B.2.c (1) ..................................................................................................................................B.2.d (1) ............................C.....................................................................................................B.2.e (1) ..................................................................................................................................B.2.f (3) .........................................................................................................-........................
K.3.a (5) ..................................................................................................................................K.3.b (1) ...............................................................G..................................................................K.3.c (1) ..................................................................................................................................K.3.d (1) ..................................................................................................................................K.3.e (1) ..................................................................................................................................B.3.a (8) ..................................................................................................................................B.3.b (1) .................................................................................................A................................B.3.c (1) ..................................................................................................................................
K.4.a (6) ..................................................................................................................................K.4.b (1) .................................................................................................................A................K.4.c (1) .................................................................................................................A................K.4.d (1) ...............................................G..................................................................................K.4.e (1) ........................................................G.........................................................................K.4.f (1) ....G.............................................................................................................................B.4.a (9) ..................................................................................................................................B.4.b (1) ...............................................................G..................................................................B.4.c (1) ..................................................................................................................................B.4.d (1) .........................................................................................................-........................
K.5.a (9) ..................................................................................................................................K.5.b (10) ..................................................................................................................................K.5.c (1) ............................................................................................................................C.....K.5.d (1) .....G............................................................................................................................B.5.a (10) ..................................................................................................................................B.5.b (1) ..................................................................................................................................B.5.c (1) ..................................................................................................................................B.5.d (1) ..................................................................................................T...............................B.5.e (1) ..................................................................................................................................B.5.f (1) ..................................................................................................................................B.5.g (1) ..................................................................................................................................B.5.h (1) ..................................................................................................................................B.5.i (4) .........................................................................................................-........................
180 190 200 210 220 230 240 250 260. . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . .
99299 GCATATCGACTATGGTTTCTAAGGCGGAGGACAGGATTACATCTTTACTCAGTTCAAACCAAGATGTAGTAGACAGAATATACAAGCAAGTGGCC
K.1.a (11) ...............................................................................................K.1.b (1) ........................................................................................G......B.1.a (10) ...............................................................................................
K.2.a (19) ...............................................................................................K.2.b (1) ...............................................................................................K.2.c (1) ...............................................................................................K.2.d (1) .........................................................................................C.....K.2.e (1) ...............................................................................................B.2.a (14) ...............................................................................................B.2.b (2) ................................................C..............................................B.2.c (1) .......................................................G.......................................B.2.d (1) ...............................................................................................B.2.e (1) ..............................G................................................................B.2.f (3) ...............................................................................................
K.3.a (5) ...............................................................................................K.3.b (1) ...............................................................................................K.3.c (1) ..G............................................................................................K.3.d (1) ........................................................G......................................K.3.e (1) ...................................................................C...........................B.3.a (8) ...............................................................................................B.3.b (1) ...............................................................................................B.3.c (1) ...................................................................G...........................
K.4.a (6) ...............................................................................................K.4.b (1) ...............................................................................................K.4.c (1) ....................................C..........................................................K.4.d (1) ...............................................................................................K.4.e (1) ...........G...................................................................................K.4.f (1) ...............................................................................................B.4.a (9) ...............................................................................................B.4.b (1) ...............................................................................................B.4.c (1) ..G............................................................................................B.4.d (1) ...............................................................................................
K.5.a (9) ...............................................................................................K.5.b (10) .........T.....................................................................................K.5.c (1) ................................G..............................................................K.5.d (1) ...............................................................................................B.5.a (10) ...............................................................................................B.5.b (1) ....................................C..........................................................B.5.c (1) .....................................................................G.........................B.5.d (1) ...............................................................................................B.5.e (1) ...................G...........................................................................B.5.f (1) ............................................................................G..................B.5.g (1) ..........................................................................................C....B.5.h (1) ....................G.............................G............................................B.5.i (4) ...............................................................................................
182
148
Figure 5. Alignment of clone sequences obtained for the HN gene of the 99299 strain after serial passages in pigeons. As in Figure 2,
each sequence is compared with the published consensus sequence (AJ415886) of 99299 and only differences from it are shown for the
clones. Each unique clone sequence was given a codification including the organ (K, kidney; B, brain), the pigeons (numbered 1 to 5) and
a lowercase letter. Then, the number of clones found for each unique sequence is indicated in brackets. Arrows used to point out the
remarkable positions mentioned in the text.
120 C. Barbezange and V. Jestin
Dow
nloa
ded
by [
Inst
itut P
aste
ur]
at 2
3:29
15
Aug
ust 2
012

also highlighted that, due to the great individualvariability between pigeons, the number and nature ofsamples and biological parameters to study should beincreased to draw objective correlations between viralheterogeneity and pathogenicity. Indeed, it would beinteresting to study other organs. We have collected andstored the spleen, lungs and liver from the pigeons usedin our present study and these are available for futureuse. It would also be useful to further characterize theviral subpopulations excreted in the digestive andrespiratory tracts, paying particular attention to thedifferent contact groups. For such large studies, techni-ques using differential gel electrophoresis mobility, suchas single-strand conformation polymorphism (Mc Kech-nie & Mc Cruden, 2001), would be useful to obtain anoverview on viral heterogeneity.
Several studies using only pPMV-1 are needed to fullyunderstand its pathogenesis. However, the influence ofvaccination and/or co-infection, for example with im-munosuppressive agents such as circovirus (Todd, 2000),on the nature of pPMV-1 quasispecies is also worthstudying. Indeed, such an influence has been shown forother viruses (Collins et al. , 2000; Essajee et al. , 2002),and such a hypothesis relating to an interaction betweenthe quasispecies nature of pPMV-1, vaccination and co-infections, together with the fact that suspect cases aremore systematically reported, might explain why moreand more cases of pigeon paramyxovirosis are recordedin Europe (Commission of the European Community,2000; Terregino et al. , 2003) despite the strengthening ofcontrol measures.
Acknowledgements
C.B. was supported by a grant from the DepartmentCouncil of Cotes d’Armor (France) as a Ph.D. student.The authors are grateful to C. de Boisseson and V. Beven(AFSSA-Ploufragan, Biosecurity and Viral Geneticsunit) for teaching and performing part of the sequen-cing; Y. Morin and his team from the SEEPA service(AFSSA-Ploufragan, experimental facilities) for takingpart in the experiments; M.-O. Le Bras and K. Loubou-tin (AFSSA-Ploufragan, Avian and Rabbit Virology,Immunology and Parasitology unit) for performing theICPI tests, the virus isolations and the F gene PCRs; andDr A. Thackray (University of Cambridge, Departmentof Veterinary Medicine) for correcting the Englishlanguage.
References
Alexander, D.J. (2001). Newcastle disease. British Poultry Science , 42 ,
5�/22.
Alexander, D.J. & Parsons, G. (1984). Avian paramyxovirus type 1
infections of racing pigeons. 2. Pathogenicity experiments in pigeons
and chickens. Veterinary Record , 114 , 466�/468.
Alexander, D.J. & Parsons, G. (1986). Pathogenicity for chickens of
avian paramyxovirus type 1 isolates obtained from pigeons in Great
Britain during 1983�/85. Avian Pathology, 15 , 487�/493.
Alexander, D.J., Russell, P.H. & Collins, M.S. (1984a). Paramyxovirus
type 1 infections of racing pigeons. 1. Characterization of isolated
viruses. Veterinary Record , 114 , 444�/446.
Alexander, D.J., Wilson, G.W., Thain, J.A. & Lister, S.A. (1984b). Avian
paramyxovirus type 1 infection of racing pigeons. 3. Epizootiological
considerations. Veterinary Record , 115 , 213�/216.
Alexander, D.J., Russell, P.H., Parsons, G., Abu Elzein, E.M.E.,
Ballouh, A., Cernik, K., Engstrom, B., Fevereido, M., Fleury,
H.J.A., Guittet, M., Kaleta, E.F., Kihm, U., Kosters, J., Lomniczi,
B., Meister, J., Meulemans, G., Nerome, K., Petek, M., Pokomunski,
S., Polten, B., Prip, M., Richter, R., Saghy, E., Samberg, Y.,
Spanoghe, L. & Tumova, B. (1985). Antigenic and biological
characterisation of avian paramyxovirus type 1 isolates from
pigeons*/an international collaborative study. Avian Pathology, 14 ,
365�/376.
Alexander, D.J., Manvell, R.J., Frost, K.M., Pollitt, W.J., Welchman, D.
& Perry, K. (1997). An outbreak of Newcastle disease in pheasants in
Great Britain in May 1996. Veterinary Record , 140 , 20�/22.
Anonymous (1992). Council Directive 92/66/EEC of 14 July 1992
introducing Community measures for the control of Newcastle
disease. Official Journal of the European Communities , L260 , 1�/20.
Barbezange, C. & Jestin, V. (2002). Development of a RT-nested PCR
test detecting pigeon Paramyxovirus-1 directly from organs of
infected animals. Journal of Virological Methods , 106 , 197�/207.
Barbezange, C. & Jestin, V. (2003a). Molecular characterisation of three
avian paramyxovirus type 1 isolated from pigeons in France. Virus
Genes , 26 , 175�/183.
Barbezange, C. & Jestin, V. (2003b). Monitoring of pigeon paramyx-
ovirus type-1 in organs of pigeons naturally infected with Salmonella
Typhimurium. Avian Pathology, 32 , 277�/283.
Cabot, B., Martell, M., Esteban, J.I., Sauleda, S., Otero, T., Esteban, R.,
Guardia, J. & Gomez, J. (2000). Nucleotide and amino acid
complexity of hepatitis C virus quasispecies in serum and liver.
Journal of Virology, 74 , 805�/811.
Collins, K.R., Mayanjakizza, H., Sullivan, B.A., Quinonesmateu, M.E.,
Toossi, Z. & Arts, E.J. (2000). Greater diversity of HIV-1 quasispecies
in HIV-infected individuals with active tuberculosis. Journal of
Acquired Immune Deficiency Syndromes , 24 , 408�/417.
Collins, M.S., Alexander, D.J., Brockman, S., Kemp, P.A. & Manvell,
R.J. (1989). Evaluation of mouse monoclonal antibodies raised
against an isolate of the variant avian paramyxovirus type 1
responsible for the current panzootic in pigeons. Archives of Virology,
104 , 53�/61.
Commission of the European Community. (2000). In D.J. Alexander
(Ed.), Proceedings of the Joint Sixth Annual Meetings of the
National Newcastle Disease and Avian Influenza Laboratories of
Countries of the European Union, Brussels, 29�/30 November 1999.
Curran, J. & Kolakofsky, D. (1999). Replication of paramyxoviruses.
Advances in Virus Research , 54 , 403�/422.
Delenda, C., Taylor, G., Hausmann, S., Garcin, D. & Kolakofsky, D.
(1998). Sendai viruses with altered P, V, and W protein expression.
Virology, 242 , 327�/337.
Domingo, E. (2002). Quasispecies theory in virology. Journal of
Virology, 76 , 463�/465.
Domingo, E. & Holland, J.J. (1997). RNA virus mutations and fitness
for survival. Annual Reviews in Microbiology, 51 , 151�/178.
Domingo, E., Escarmis, C., Sevilla, N., Moya, A., Elena, S.F., Quer, J.,
Novella, I.S. & Holland, J.J. (1996). Basic concepts in RNA virus
evolution. FASEB Journal , 10 , 859�/864.
Domingo, E., Escarmis, C., Baranowski, E., Ruiz-Jarabo, C.M.,
Carrillo, E., Nunez, J.I. & Sobrino, F. (2003). Evolution of foot-
and-mouth disease virus. Virus Research , 91 , 47�/63.
Essajee, S.M., Yogev, R., Pollack, H., Greenhouse, B., Krasinski, K. &
Borkowsky, W. (2002). Recombinant glycoprotein vaccines for hu-
man immunodeficiency virus-infected children and their effects on
viral quasispecies. Clinical and Diagnostic Laboratory Immunology, 9 ,
79�/82.
Granoff, A. (1964). Nature of the Newcastle disease virus population.
In R.P. Hanson (Ed.), Newcastle Disease Virus: An Evolving
Pathogen . Madison, USA: University of Wisconsin Press.
Gretch, D.R. & Polyak, S.J. (1997). The quasispecies nature of hepatitis
C virus: research methods and biological implications. In Groupe
Francais d’Etudes Moleculaires des Hepatites (GEMHEP) (Eds.),
Hepatitis C Virus: Genetic Heterogeneity and Viral Load (pp. 57�/69).
Paris: John Libbey Eurotext.
Hotzel, I., Kumpula-Mc Whirter, N. & Cheevers, W.P. (2002). Rapid
evolution of two discrete regions of the caprine arthritis-encephalitis
virus envelope surface glycoprotein during persistent infection. Virus
Research , 84 , 17�/25.
In vivo quasispecies nature of pigeon PMV-1 121
Dow
nloa
ded
by [
Inst
itut P
aste
ur]
at 2
3:29
15
Aug
ust 2
012

Huang, Z., Panda, A., Elankumaran, S., Govindarajan, D., Rock-
emann, D.D. & Samal, S.K. (2004). The hemagglutinin-neuramini-
dase protein of Newcastle disease virus determines tropism and
virulence. Journal of Virology, 78 , 4176�/4184.
Islam, M.A., Ito, T., Takakuwa, H., Takada, A., Itakura, C. & Kida, H.
(1994). Acquisition of pathogenicity of a newcastle disease virus
isolated from a japanese quail by intracerebral passage in chickens.
Japanese Journal of Veterinary Research , 42 , 147�/156.
Jestin, V., Cherbonnel, M., Morin, M., Guittet, M. & Bennejean, G.
(1989). Characterization of French avian paramyxovirus type 1
(PMV1) isolates with a panel of monoclonal antibodies to the
Ploufragan strain of Newcastle disease virus. Archives of Virology,
105 , 189�/198.
Kaleta, E.F., Siegmann, O., Jank-Ladwig, R. & Glunder, G. (1980).
Isolation and biological properties of virulent subpopulations from
lentogenic Newcastle disease virus strains. Comparative Immunology,
Microbiology and Infectious Diseases , 2 , 485�/496.
Kaleta, E.F., Alexander, D.J. & Russell, P.H. (1985). The first isolation
of the avian PMV-1 virus responsible for the current panzootic in
pigeons? Avian Pathology, 14 , 553�/557.
Kato, A., Kiyotani, K., Sakai, Y., Yoshida, T. & Nagai, Y. (1997). The
paramyxovirus, Sendai virus, V protein encodes a luxury function
required for viral pathogenesis. EMBO Journal , 16 , 578�/587.
Kissi, B. (1988). Studies on the virulence of pigeon paramyxovirus-1
(PMV-1). I. Changes in the virulence of pigeon PMV-1 strains
isolated in Hungary upon passage in chickens, embryonated hen’s
eggs and pigeons. Acta Veterinaria Hungarica , 36 , 283�/292.
Kissi, B. & Lomniczi, B. (1988). Studies on the virulence of pigeon
paramyxovirus-1 (PMV-1). II. Mechanism of the virulence increase
of pigeon PMV-1. Acta Veterinaria Hungarica , 36 , 293�/298.
Kissi, B., Badrane, H., Audry, L., Lavenu, A., Tordo, N., Brahimi, M. &
Bourhy, H. (1999). Dynamics of rabies virus quasispecies during
serial passages in heterologous hosts. Journal of General Virology, 80 ,
2041�/2050.
Komatsu, T., Takeuchi, K., Yokoo, J. & Gotoh, B. (2004). C and V
proteins of Sendai virus target signaling pathways leading to IRF-3
activation for the negative regulation of interferon-beta production.
Virology, 325 , 137�/148.
Kommers, G.D., King, D.J., Seal, B.S. & Brown, C.C. (2003). Virulence
of six heterogeneous-origin Newcastle disease virus isolates before
and after sequential passages in domestic chickens. Avian Pathology,
32 , 81�/93.
Laskus, T., Radkowski, M., Wang, L.F., Nowicki, M. & Rakela, J.
(2000). Uneven distribution of hepatitis C virus quasispecies in
tissues from subjects with end-stage liver disease: confounding effect
of viral adsoption and mounting evidence for the presence of low-
level extrahepatic replication. Journal of Virology, 74 , 1014�/1017.
Lomniczi, B. (1974). Plaque assay for avirulent (lentogenic) strains of
Newcastle disease virus. Applied Microbiology, 27 , 1162�/1163.
Malet, I., Belnard, M., Agut, H. & Cahour, A. (2003). From RNA to
quasispecies: a DNA polymerase with proofreading activity is highly
recommended for accurate assessment of viral diversity. Journal of
Virological Methods , 109 , 161�/170.
Mc Kechnie, V.M. & Mc Cruden, E.A.B. (2001). Optimisation and
sensitivity of single-stranded conformation polymorphism for the
detection of hepatitis C virus quasi-species. Journal of Virological
Methods , 92 , 131�/139.
Mebatsion, T., Verstegen, S., Devaan, L.T.C., Romer-Oberdorfer, A. &
Schrier, C.C. (2001). A recombinant Newcastle disease virus with
low-level V protein expression is immunogenic and lacks pathogeni-
city for chicken embryos. Journal of Virology, 75 , 420�/428.
Meulemans, G., Van Den Berg, T.P., Decaesstecker, M. & Boschmans,
M. (2002). Evolution of pigeon Newcastle disease virus strains. Avian
Pathology, 31 , 515�/519.
Millar, N.S. & Emmerson, P.T. (1988). Molecular cloning and
nucleotide sequencing of Newcastle disease virus. In D.J. Alexander
(Ed.), Newcastle Disease (pp. 79�/97). Boston: Kluwer Academic
Publishers.
Mullan, B., Kennywalsh, E., Collins, J.K., Shanahan, F. & Fanning, L.J.
(2001). Inferred hepatitis C virus quasispecies diversity is influenced
by choice of DNA polymerase in reverse transcriptase-polymerase
chain reactions. Analytical Biochemistry, 289 , 137�/146.
Office International des Epizooties. (2000). Newcastle Disease. In OIE
Standards Commission (Ed.), OIE Manual of Standards for Diag-
nostic Tests and Vaccines (pp. 221�/226). Paris: OIE.
Park, M.S., Garcia-Sastre, A., Cros, J.F., Basler, C.F. & Palese, P. (2003).
Newcastle disease virus V protein is a determinant of host range
restriction. Journal of Virology, 77 , 9522�/9532.
Peeples, M.E., Wang, C., Gupta, K.C. & Coleman, N. (1992). Nuclear
entry and nucleolar localization of the Newcastle disease virus
(NDV) matrix protein occur early in infection and do not require
other NDV proteins. Journal of Virology, 66 , 3263�/3269.
Picault, J.P. (1993a). Maladie de Newcastle: identification du virus par
inhibition de l’hemagglutination. In COFRAC (Ed.), Texte Francais
de Reference d’une technique d’analyse Pr112/00/VA 30/00 . Paris:
COFRAC.
Picault, J.P. (1993b). Myxoviroses aviaires a virus hemagglutinant:
isolement par ovoculture et recherche de l’activite hemagglutinante.
In COFRAC (Ed.), Texte Francais de Reference d’une technique
d’analyse Pr112/00/VA 10/00 . Paris: COFRAC.
Reed, L.J. & Muench, H. (1938). A simple method of estimating fifty
percent endpoints. American Journal of Hygiene , 27 , 493�/497.
Ruiz-Jarabo, C.M., Arias, A., Baranowski, E., Escarmis, C. &
Domingo, E. (2000). Memory in viral quasispecies. Journal of
Virology, 74 , 3543�/3547.
Ruiz-Jarabo, C.M., Arias, A., MolinaParis, C., Briones, C., Bara-
nowski, E., Escarmis, C. & Domingo, E. (2002). Duration and fitness
dependence of quasispecies memory. Journal of Molecular Biology,
315 , 285�/296.
Ruster, B., Zeuzem, S., Krumpkonvalinkova, V., Berg, T., Jonas, S.,
Severin, K. & Roth, W.K. (2001). Comparative sequence analysis of
the core- and NS5-region of hepatitis C virus from tumor and
adjacent non-tumor tissue. Journal of Medical Virology, 63 , 128�/
134.
Sanchez Fueyo, A., Gimenez Barcons, M., Puig Basagoiti, F., Rimola,
A., Sanchez Tapias, J.M., Saiz, J.C. & Rodes, J. (2001). Influence of
the dynamics of the hypervariable region 1 of hepatitis C virus (HCV)
on the histological severity of HCV recurrence after liver transplan-
tation. Journal of Medical Virology, 65 , 266�/275.
Schloer, G. & Hanson, R.P. (1968). Plaque morphology of Newcastle
disease virus as influenced by cell type and environmental factors.
American Journal of Veterinary Research , 29 , 883�/895.
Shapshak, P., Segal, D.M., Crandall, K.A., Fujimura, R.K., Zhang,
B.T., Xin, K.Q., Okuda, K., Petito, C.K., Eisdorfer, C. & Goodkin,
K. (1999). Independent evolution of HIV type 1 in different brain
regions. AIDS Research and Human Retroviruses , 15 , 811�/820.
Smith, D.B., Mc Allister, J., Casino, C. & Simmonds, P. (1997). Virus
‘‘quasispecies’’: making a mountain out of a mohehill? Journal of
General Virology, 78 , 1511�/1519.
Steward, M., Vipond, B., Millar, N.S. & Emmerson, P.T. (1993). RNA
editing in Newcastle disease virus. Journal of General Virology, 74 ,
2539�/2547.
Steward, M., Samson, A.C.R., Errington, W. & Emmerson, P.T. (1995).
The Newcastle disease virus V protein binds zinc. Archives of
Virology, 140 , 1321�/1328.
Tangredi, B.P. (1985). Avian paramyxovirus type 1 infection in pigeons:
clinical observations. Avian Diseases , 29 , 1252�/1255.
Terregino, C., Cattoli, G., Grossele, B., Bertoli, E., Tisato, E. & Capua,
I. (2003). Characterization of Newcastle disease virus isolates
obtained from Eurasian collared doves (Streptopelia decaocto) in
Italy. Avian Pathology, 32 , 63�/68.
Todd, D. (2000). Circoviruses: immunosuppressive threats to avian
species: a review. Avian Pathology, 29 , 373�/394.
Wang, W.-K., Sung, T.-L., Lee, C.-N., Lin, T.-Y. & King, C.-C. (2003).
Sequence diversity of the capsid gene and the nonstructural gene
NS2B of Dengue-3 virus in vivo. Virology, 303 , 181�/191.
Yusoff, K. & Tan, W.S. (2001). Newcastle disease virus: macromolecules
and opportunities. Avian Pathology, 30 , 439�/455.
Translations of the abstract in French, German and Spanish are available on the Avian Pathology website.
122 C. Barbezange and V. Jestin
Dow
nloa
ded
by [
Inst
itut P
aste
ur]
at 2
3:29
15
Aug
ust 2
012

Non-English Abstracts
Molecular study of the quasispecies evolution of a typicalpigeon paramyxovirus type 1 after serial passages inpigeons by contact
C. Barbezange1,2 and V. Jestin1*
1AFSSA (French Agency for Food Safety), Avian and Rabbit Virology Immunology and Parasitology Unit, BP53, 22440Ploufragan, France, and 2Present address: University of Cambridge, Centre for Veterinary Sciences, Department ofVeterinary Medicine, Madingley Road, Cambridge CB3 0ES, UK
La nature en quasi-especes d’un paramyxovirus de type 1-variant pigeon typique (pPMV-1) a ete pour lapremiere fois etudiee dans des conditions proches de l’infection naturelle. Le virus a subi plusieurs passageschez le pigeon par contacts successifs. L’heterogeneite virale a ete analysee dans les reins et le cerveau de 5pigeons preleves lors du dernier contact, par retro-transcription suivie d’amplification genique (RT-PCR)directement a partir des ARNs extraits de ces organes et en ciblant les genes viraux P et HN. Bien que ladiversite virale consecutive aux passages in vivo se soit revelee differente de celle presente dans l’inoculum, larealite du concept de quasi-especes pour les souches pPMV-1 est demontree. De plus, ont ete detectes desARNs genomiques aberrants presentant des insertions au niveau du site d’edition du gene P ou des deletionsdans le gene HN, avec des consequences possibles sur la pathogenicite et l’infectivite virales.
Zum erstenmal wurde die Natur der Quasispezies eines typischen Paramyxovirus Typ 1 (pPMV-1) unterBedingungen, die der naturlichen Infektionsumgebung sehr nahe kamen, untersucht. Das Virus wurde durchaufeinanderfolgenden Kontakt in Tauben serienpassagiert. Die Virusheterogenitat wurde in den Nieren undGehirnen von funf Tauben aus dem letzten Kontakt analysiert. Dabei richtete sich die RT-PCR mit derdirekt aus den Organen extrahierten RNS auf die P- und HN-Gene des Virus. Die Virusvielfalt nach den invivo-Passagen unterschied sich von der des Inokulums, aber spiegelte die Realitat des Quasispezieskonzeptsfur pPMV-1-Stamme wieder. Daruber hinaus wurden einige abweichende Genom-RNS, die Insertionen ander Bearbeitungsstelle des P-Gens oder Deletionen im HN-Gen beinhalteten, entdeckt, was Konsequenzenfur die Pathogenitat und Infektiositat des Virus haben kann.
La naturaleza de cuasiespecies de un tıpico paramixovirus tipo 1 de la palomas (pPMV-1) fue estudiada porprimera vez bajo condiciones similares a las del entorno donde se da la infeccion natural. El virus fue pasadoseriadamente en palomas por contacto. La heterogeneidad viral fue analizada en los rinones y cerebro decinco palomas del ultimo contacto, mediante RT-PCRs de ARN directamente extraıdo de los organos yamplificando los genes P y HN del virus. La diversidad viral tras pases in vivo fue diferente respecto a la delinoculo, pero demostro la realidad del concepto de cuasiespecies para las cepas pPMV-1. Ademas, algunosARNs genomicos aberrantes con inserciones en el punto de edicion del gen P o delecciones en el gen HNfueron tambien detectados, con posibles consecuencias para la patogenicidad e infectividad de los virus.
*To whom correspondence should be addressed. Tel: �/33 2 96 01 62 22. Fax: �/33 2 96 01 62 63. E-mail: [email protected]
Received 7 September 2004. Provisionally accepted 3 November 2004. Accepted 21 December 2004
Avian Pathology (April 2005) 34(2), 1�/2
ISSN 0307-9457 (print)/ISSN 1465-3338 (online) # 2005 Houghton Trust LtdDOI: 10.1080/03079450500059321
Dow
nloa
ded
by [
Inst
itut P
aste
ur]
at 2
3:29
15
Aug
ust 2
012





























