Embed Size (px)
Citation preview

GENERAL AND COMPARATIVE ENDOCRINOLOGY 45, 189-203 (1981)
Monoaminergic Innervation of the Caudal Neurosecretory System of the Brook Trout Salvelinus fontinalis in Relation to
Osmotic Stimulation1
CBLINE AUDET AND GASTON CHEVALIERS
Dkpartement des Sciences Biologiques, Universitk du Qkbec d MontrPal, Que’bec, H3C 3P8, Canada
Montrt?al, C.P. 8888,
Accepted February 2, 1981
The distribution of monoamine fluorescence reaction (Falck and Hillarp technique) was investigated in the posterior spinal cord of the brook trout, Salvelinus fontinalis, in relation with the caudal neurosecretory system. A semiquantitative analysis of fluorescence was made following variations of external salinity, injections of salt solutions, and drugs aimed at modifying hydromineral balance. Under all experimental conditions, an extensive network of beaded fibers innervate a number of caudal neurosecretory cells whereas some others do not appear to be contacted by fluorescent varicosities. Three types of aminergic cells differ- ent from nonfluorescent caudal neurosecretory neurons were observed in the posterior spinal cord: (i) green fluorescent CSF-contacting neurons interspersed between ordinary ependymal cells; (ii) small perikarya with discrete spots of intense yellow fluorescence; (iii) small somata with diffuse green fluorescence. No fluorescence was observed in neurohemal region or urophysis. The action of nialamide and reserpine indicated that histofluorescent reaction was due to monoamines. In comparison to freshwater exposure to demineralized water for 3 days, but not 7 days, increased markedly the fluorescence of beaded fibers and varicosities contiguous to caudal neurosecretory cells while transfer to seawater for I and 7 days reduced them significantly. Injections of isotonic NaCl, hypertonic Na,SO,, and hypertonic choline chloride in freshwater fish, were less effective in producing fluorescence changes; however, these treatments showed a common tendency to enhance frequency of caudal neurosecretory cells with abutting varicosities in the most anterior part of the popu- lation. Acetazolamide and aldactone treatments also induced significant variations of aminergic innervations. These findings indicate a monoaminergic participation in the control of the function of caudal neurosecretory cells in relation to iono- and osmoregulation.
Various lines of evidence have contrib- uted to the demonstration of the existence of an afferent innervation of the teleost caudal neurosecretory system. Electro- physiological studies in several species had indicated neural input to caudal neuro- secretory cells (Ishibashi, 1962; Bennett and Fox, 1962; Yagi and Bern, 1965); sub- sequently, electron microscopic identifica- tion of synaptic connections with the caudal neurosecretory neurons was then reported in many species (Fridberg, 1963; Fridberg et al., 1966; Bern et al., 1965; Sano et al.,
l Supported by National Science and Engineering Research Council of Canada.
2 To whom reprint requests should be addressed.
1966; Baumgarten et al., 1970; Kobayashi et al., 1979). As well, monoaminergic in- nervation of caudal neurosecretory neurons was demonstrated with the use of the Falck- Hillarp histofluorescent technique (Falck et al., 1962) in Esox lucius (Baum- garten et al., 1970), Gillichthys mirabilis and Tilapia mossarnbica (Lederis et al., 1974; Swanson et al., 1975) and Cyprinus carpio (Kobayashi, 1979, 1980), and the presence of monoamine oxidase activity was histochemically shown in nerve termi- nals in close contact with caudal neurose- cretory perikarya in the carp (Luppa et al., 1968; Kobayashi et al., 1979). Investigation of aminergic innervation in the urophysis revealed a positive histofluorescent reac-
189 0016-6480/81/100189-15$01.00/O Copyright @ 1981 by Academic Press, Inc. All rights of reproduction in any form reserved.

190 AUDET AND CHEVALIER
tion in the pike (Baumgarten et al., 1970) (albeit inconsistent) and in carp (Kobayashi et al., 1980) but not in Tilapia (Lederis et al., 1974) and Gillichthys (Swanson, 1975). Furthermore, noradrenaline, but not adren- aline or dopamine, was chemically detected both in the caudal spinal cord (Kobayashi et al., 1980) and urophysis of the carp, and noradrenergic, but not adrenergic or dopaminergic pharmacological stimulation was found to stimulate in vitro secretion of urotensin II in the isolated posterior spinal cord of the carp (Lederis et al., 1978; Ichikawa, 1979).
Cholinergic innervation of the caudal neurosecretory system has also been pro- posed in some species on the basis of bio- chemical assays of acetylcholine (Kobay- ashi, 1963; Ichikawa, 1978), histochemical demonstrations of acetylcholinesterase ac- tivity (Uemura, 1965; Luppa et al., 1968; Kobayashi, 1979)) ultrastructural findings (Baumgarten et al., 1970; Kobayashi et al., 1979)) and pharmacological stimulation of isolated spinal cord preparations by cholin- ergic (muscarinic) drugs (Lederis et al., 1978; Ichikawa, 1979).
Unfortunately, the physiological signifi- cance of both aminergic and cholinergic control systems remains to be established. Yagi and Bern (1965) have, however, re- ported that transection of the spinal cord anterior to the caudal neurosecretory cells of Tilapia resulted in a loss of spontaneous activity of the neurosecretory neurons and in abolition of the responses of the neurose- cretory cells to osmotic stimuli. Variations in the monoamine histofluorescence around caudal neurosecretory cells was also noted in Gillichthys after transfer from sea water to fresh water (Swanson et al., 1975).
In order to investigate further the role of the nervous control in the osmoregulatory function of the caudal neurosecretory sys- tem, the formaldehyde-induced fluores- cence technique (Falck et al., 1962) was used to study the distribution of mono- amines in the posterior spinal cord of the
brook trout Salvelinus fontinalis. Inasmuch as caudal neurosecretory cells of S. fon- tinalis, a euryhaline species, were previ- ously reported to respond to changes of external salinity (Chevalier, 1976, 1978), qualitative and semiquantitative analyses of histofluorescence were made in order to compare the distribution and intensity of fluorescence in fish submitted to various treatments to determine if altering internal as well as external ionic and osmotic condi- tions could induce differences in histoflu- orescent reactions. First, aminergic flu- orescence was investigated in trout ex- posed for various times to either fresh water, deionized water, or sea water, con- ditions to which caudal neurosecretory cells of S. fontinalis respond by varying their synthetic activity of neurosecretory material (Chevalier, 1976, 1978). Histoflu- orescence was also analyzed in the caudal spinal cord of trout injected with isotonic NaCl, hypertonic Na,SO,, and choline chloride. These treatments were aimed at increasing extracellular body fluid and in- creasing and decreasing selectively plasma level of sodium and chloride (Bourguet et al., 1964; Mayer and Nibelle, 1970; Mayer-Gostan and Kirsch, 1979).
Moreover, a similar study was carried out in S. fontinalis treated with drugs which alter hydromineral metabolism. Acetazol- amide, an inhibitor of carbonic anhydrase, which has been reported to block sodium and chloride absorption mechanisms in gills of freshwater species (Maetz and Garcia- Romeu, 1964; Kerstetter et al., 1970) and aldactone, an antialdosterone agent, which induced hypertrophy of interrenal cells (Oliverau, 1964) and also reduced sodium influx in eel gills (Chan et al., 1967) were used to modify plasma electrolyte concen- tration.
MATERIALS AND METHODS
Fish
Hatchery-reared brook trouts, Salvelinus fontinalis, were kept in recirculated aerated tap water aquaria,

INNERVATION OF NEUROSECRETORY CELLS 191
maintained at lo”, with a 12: 12 hr photoperiod. A total of 115 fish of either sex, of an average length of 15.7 + 2.9 cm and of average weight of 35.9 + 18 g, were used throughout the year.
Histological Techniques
All trouts were anesthetized with tricaine methane- sulfonate (MS 222) (100 mgliter) and were then sac- rificed by decapitation. The posterior part of the spinal cord, from the 12th preterminal verterbra to thefilum terminale, was dissected out, frozen in liquid propane, cooled in liquid nitrogen, and then freeze-dried for 5 days as described by Parent et al. (1978). The Falck and Hillarp reaction (Falck et al., 1962) was carried out with paraformaldehyde (70% relative humidity) for 1 hr at 80” and the freeze-dried material was embedded in paraffin in a vacuum oven. Series of four &pm-thick sagittal sections were dry-mounted on one slide, em- bedded in Entellan, and examined under a Leitz fluo- rescence microscope equipped with a Leitz Ploemopak-2 system for reflected light microscopy, an excitation filter BG12 and a barrier filter K530. After examination in the fluorescence microscope, one slide out of four was routinely stained with hematox- ylene-eosin for topographic histological study.
The specificity of the observed fluorescence for monoamines was checked by the following methods:
(1) Two freeze-dried specimens were not exposed to formaldehyde vapors.
(2) Nialamide (Sigma) (100 mgkg), an inhibitor of monoamine oxidase, was injected intraperitoneally in five trouts, 3 hr prior to sacrifice (Fremberg and Merl- ing, 1975).
(3) Reserpine (Sigma) (30 mg/kg), a monoamine de- pleting agent, was given intraperitoneally and 24 hr before sacrifice.
to five fish, 3
In order to minimize the effects of circadian varia- tion in tissue amine contents, all fish were sacrificed between 11:00 AM and 14:00 PM.
Osmotic Manipulations Variations of external salinity. One group of 12 fish
were fasted for 5 days in fresh water and subsequently transferred to individual aquaria containing 10 liters of deionized water (< 1 mosm/kg) changed twice daily. All fish were unfed during these treatments; 6 were maintained in this medium for 3 days and the remain- ing fish for 7 days.
Another group of six fish were directly transferred from the freshwater stock aquarium to aquaria con- taining reconstituted 25% full-strength seawater (Marinemix, Marine Enterprises, N.D.) and sacrificed after 1 day in this medium, while a last group of six trouts were progressively adapted to full-strength sea- water according to the following schedule: 25% sea- water after Day 1, 50% seawater after Day 3, 75% seawater after Day 5, and 100% seawater after Day 6.
The latter experimental animals were killed at Day 7 from start of adaptation to seawater. To ensure that differences in fluorescence between various experi- mental groups would not be due to inherent variations in the Falck-Hillarp reaction (Swanson et al., 1975), fish of different experimental groups were processed simultaneously.
Injections of salt solutions. Groups of five freshwater-adapted Salvelinus fontinalis were injected intraperitoneally with one of the following salt solu- tions, 3 hr before sacrifice: isotonic (0.85%) NaCl, hypertonic (0.3 M) Na,SO,, or hypertonic (0.3 M) choline chloride. The volume of the injections was 1 ml/kg. A second group of five fish was similarly given two injections 3 and 24 hr prior to sacrifice.
Treatments with aldactone and acetazolamide. Al- dactone (SC 9420) (Searle) was injected intraperitone- ally in four freshwater-adapted fish (50 mgkg in 0.85% saline, 48, 24, and 3 hr prior to sacrifice). As controls, two fish received three injections of 0.85% saline (0.2 ml). A single injection of 30 mg/kg of acetazolamide (Sigma) was administered to a group of four fish 3 hr before sacrifice. For this last treatment, controls were the same as for nialamide since the nature and volume of the vehicle were the same.
Analysis offluorescence. One section out of four in each slide was studied in fluorescent microscopy, which yielded from 15 to 55 observations of sagittal sections per fish for the visual analysis of the nature and distribution of fluorescence. In addition, for com- parison purposes, a semiquantitative analysis of fluo- rescence was carried out through the use of an arbi- trary scale (Swanson et al., 1975): fluorescent indices ( 1- 10) were assigned to each section depending on the visible intensity and number of fluorescent fibers and varicosities: 10 indicated large amounts of fluorescent fibers and varicosities and 0 indicated no fluorescence. All measurements were carried out by the same ob- server .on coded slides to avoid bias. Fluorescence could then be compared between different regions of the caudal spinal cord. For convenience, the regions considered were defined as follows: (i)filum terminale and first terminal vertebral region; (ii) the second and the third preterminal vertebral region; (iii) the fourth and fifth preterminal vertebral region.
Histograms of the number of those neurosecretory cell bodies surrounded by varicosities were also plot- ted for each of the three regions of the posterior spinal cord for each of the various experimental groups (Audet, 1980). A detailed report of these data will not be presented here for sake of brevity but the most salient results are outlined.
Statistical analysis. Semiquantitative data of fluorescent index were evaluated with the sign test, a nonparametric statistical test (Mendenhall, 1971). The latter is based on the comparison of paired observa- tions. Although parameters such as means and SE could not be tested statistically, they were represented

192 AUDET AND CHEVALIER
in histograms for their informative value. (Figs. 7, 8, minale. Only green and yellow fluorescence and 13).
RESULTS
A. Variation of External Salinity
was observedin the posterior spinal cord of S. fontinalis. Fluorescent material was present as beaded fibers, and isolated var- icosities, and small nonneurosecretory cell perikarya (Fig. 1). No fluorescence was ob- a. Fresh water. In Salvelinus fontinalis,
the caudal neurosecretory cells are distrib- served in the perikarya of any caudal neu- uted ventrolaterally with regard to the rosecretory cells (Fig. 2). ependymal canal and are restricted to a re- Fluorescent beaded fibers were observed gion of the spinal cord posterior to the running mostly parallel to the spinal cord - fourth preterminal vertebra; they are con- axis. All beaded fibers and isolated var- tinuously present to the tip of thefilum ter- icosities showed green and yellow fluores-
FIG. 1. Longitudinal section of posterior spinal cord of freshwater trout Salvelinusfontinali.s treated with nialamide (100 mg/kg). The formaldehyde-induced fluorescence appears as beaded fibers primar- ily in the region of the caudal neurosecretory cells (c). The urophysis (u) displayed no fluorescence. Some CSF-contacting neurons (s) interspersed between cence. (ec) ependymal canal. (m) meninges (x 1500).
ependymal cells showing
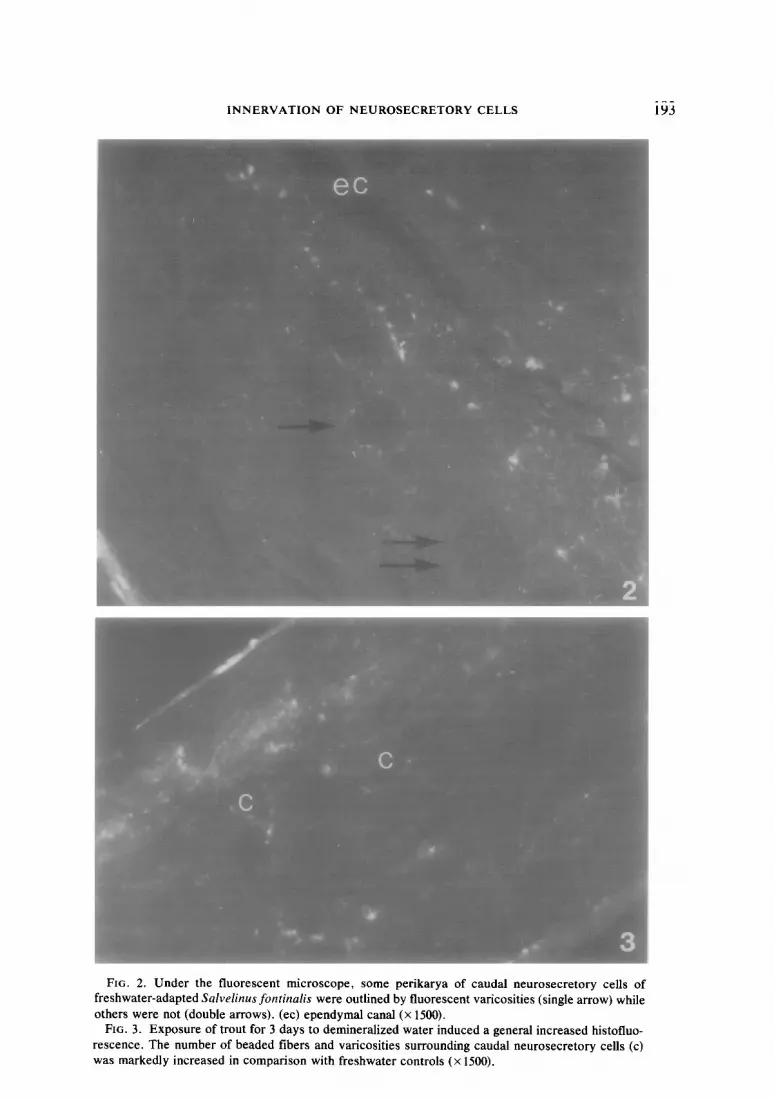
INNERVATION OF NEUROSECRETORY CELLS 193

194 AUDET AND CHEVALIER
cence, the latter color being predominant in the distal region of the caudal neurosecre- tory cells, while in more anterior zones of spinal cord, only green fluorescence was present.
Both the intensity of fluorescence and the number of varicosities in close contact with caudal neurosecretory perikarya exhibited an increasing cephalocaudal gradient from the fourth to fifth preterminal vertebrae to the j%un terminale (Figs. 8, 9). However, in all fish studied, there were always a number of caudal neurosecretory cell bodies which were not outlined by fluores- cent varicosities (Fig. 2). This group of cells was distributed throughout the entire population of the caudal neurosecretory neurons, but they were encountered most frequently in the most cranial part of this population. Motor neurons are not present in those areas of the spinal cord posterior to the 4th preterminal vertebra; when they are encountered more craniad, most of them are not accompanied by varicosities.
Cell types which showed a positive fluorescent reaction in the posterior spinal cord can be classified into three categories. First, bipolar, CSF-contacting neurons, with bright green fluorescence were en- countered between epithelial ependymal cells (Fig. 4); their beaded axons run per- pendicularly to the central canal. Second, small cell bodies with short cell processes showed discrete spots of intensely yellow fluorescence (Fig. 5). Third, another cate- gory of less numerous small cell bodies ex- hibited green diffuse fluorescence (Fig. 6). The latter two cell categories were found in isolated groups of two or three and were most abundant in the distal regions of the 1st vertebra andfilum terminale.
Nialamide treatment induced a general enhancement of the intensity of fluores- cence (Z-’ < 0.01) in all three regions of the posterior spinal cord in freshwater-adapted S. fontinalis. In particular, the number of varicosities increased in the area of the spi- nal cord where caudal neurosecretory cells
were concentrated, and the proportion of yellow varicosities became greatly en- hanced. The entire urophysis, however, appeared almost devoid of fluorescence (Fig. 1). Specimens showed no induction of fluorescence when exposure to formal- dehyde was omitted. Control fish injected with only the vehicle 0.85% saline showed no differences in fluorescence when com- pared to uninjected freshwater animals. Re- serpine treatment reduced the intensity of the fluorescence (P < 0.01) but never elimi- nated it completely, possibly due to large differences in resistance to the drug among fish.
b. Deionized water. Fish exposed to de- mineralized water for 3 days exhibited a higher index of fluorescence compared to freshwater-adapted fish (P < 0.01) (Figs. 3, 7): the intensity and number of both beaded fibers and isolated varicosities were in- creased in all regions of the posterior spinal cord studied (Fig. 8). Also, neurosecretory cell bodies in contact with varicosities were significantly more numerous at the level of second to third and fourth to fifth vertebral regions when compared to their respective freshwater controls (P < 0.01). A more prolonged exposure to demineralized water (7 days) did not induce any significant dif- ference in the amount of fluorescence or distribution of varicosities compared to the freshwater controls.
c. Seawater. Transfer of freshwater- adapted Salvelinus fontinalis to 25% sea- water for 1 day is accompanied by a marked reduction in all spinal cord regions of the fluorescence level (P < 0.01) (Figs. 7, 8) due to reduction in fluorescence intensity, number of beaded fibers, and number of varicosities in contact with neurosecretory cell bodies. Likewise, progressive exposure to full strength seawater for 7 days brought about a significant lowering of the fluores- cence index of the same order as a l-day exposure in all posterior spinal cord. Var- icosities in contact with caudal neurose- cretory cells were seen less frequently at

INNERVATION OF NEUROSECRETORY CELLS
FIG. 4. Longitudinal section of posterior spinal cord of freshwater-adapted Salvelinus fontinalis showing bright green, fluorescent CSF-contacting neurons located in a plane perpendicular to the ependymal canal. Beaded axons can be followed for a certain distance toward more peripheral zones of the spinal cord. (ec) ependymal canal (X 2500). Small perikarya with yellow granular fluorescence (Fig. 5) and others with diffuse yellow fluorescence (Fig. 6) are two other aminergic cell types found in the posterior spinal cord of freshwater Salvelinus fontinalis (X 6000).
the level of the most posterior region and 2nd- 3rd preterminal vertebral level.
B. Injection of Salt Solution
a. Isotonic 0.85% NaCl. A single ip in- jection of 1 ml/kg of isotonic NaC13 hr prior to sacrifice induced a slight decrease (P < 0.01) of fluorescence in the second and third preterminal vertebral region only (Fig. 13). Also, more varicosities were as-
sociated with neurosecretory perikarya only in the region of the fourth to fifth pre- terminal vertebrae compared to fresh water controls. A few large beaded fibers of green fluorescence were uniquely observed in the latter region. In addition, administration of two injections, 24 and 3 hr before sacrifice, induced the appearance of more var- icosities near cell bodies in the entire caudal spinal cord; but nevertheless the overall

196 AUDET AND CHEVALIER
I I
Fresh waler Deiomzed water beo water
FIG. 7. Histogram of formaldehyde-induced fluo- rescence index (determined by intensity and frequency of beaded fibers and isolated varicosities based on the use of an arbitrary scale) in sections of posterior spinal cord of the brook trout Salvelinus jkztinalis ex- posed to media of variuos external salinities. N = 6 fish in each experiment. (a) different from freshwater control (P < 0.01). (Sign test of nonparametric statics.) (cf. Materials and Methods).
level of fluorescence was not different from fresh water controls (N.S.).
b. 0.3 M Na2S0,. When hypertonic (0.3 M) Na,SO, is injected 3 hr before sacrifice, a slight reduction of fluorescence is ob- served in the second to third preterminal
a ra
DW SW
FIG. 8. Histogram of formaldehyde-induced fluo- rescence index for three subregions of posterior spinal cord of S. fontinalis exposed to media of various salinities. (FW) freshwater. (DW) demineralized water. (SW) seawater, Empty columns: filum ter- minale and first terminal vertebral region, stippled columns: second to third preterminal vertebral region and striped columns: third to fourth preterminal ver- tebral region. Difference from freshwater control. (a) (P < 0.01) and (b) (P < 0.05) (Sign test).
100 -
80,
FIG. 9. Histogram of the relative frequency of caudal neurosecretory cells in contact with fluorescent varicosities in the posterior spinal cord of S. fontinalis adapted to fresh water. The presence of two popula- tions of neurosecretory neurons is evident. Index 0 indicates the frequency with which cells with no fluorescent innervation were noted. Indexes 1, 2, 3, 4 stand for observations in which, respectively, 1- 25%, 26- 50%, 51- 75%. 76- 100% of cells observed had contacting varicosities, thus suggesting a second neu- rosecretory cell type. Empty columns:&m terminale and 1st terminal vertebral region, stippled columns: second to third preterminal vertebral region and striped columns: third to fourth preterminal vertebral region.
region (P < 0.01) but not in the other areas. The proportion of caudal neurosecretory somata in contact with green varicosities was also higher than in the controls over the entire caudal spinal cord (Fig. 11). There was no difference in the fluorescence index between control and treated animals after two injections of 0.3 M Na,SO, 24 and 3 hr prior to sacrifice, but this treatment did re- sult in an increased number of caudal neu- rosecretory neurons with contacting var- icosities in the fourth to fifth preterminal vertebral region; however, this number was lowered in the most posterior spinal region.
c. 0.3 M Choline chloride. No variation of the fluorescence index in fish injected once with 0.3 A4 choline chloride was ob- served with regards to controls. However,
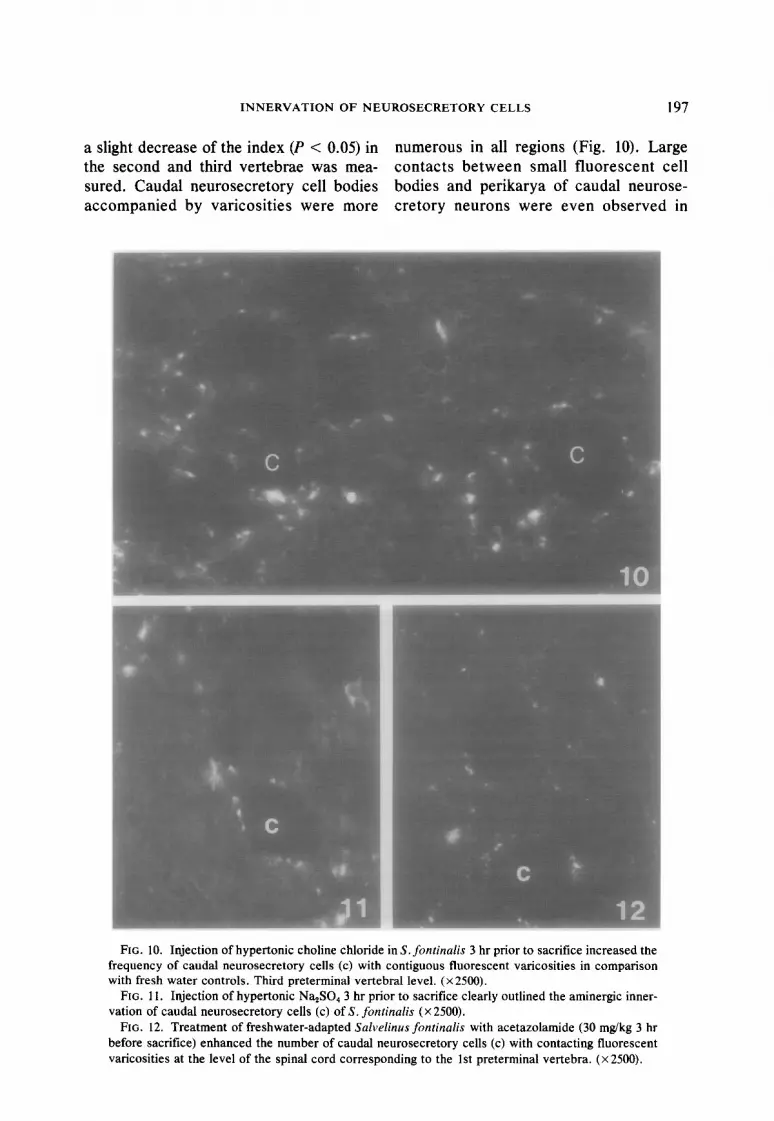
INNERVATION OF NEUROSECRETORY CELLS 197
a sl light decrease of the index (P < 0.05) in numerous in all regions (Fig. 10). L; arge the second and third vertebrae was mea- contacts between small fluorescent cell sur ed. Caudal neurosecretory cell bodies bodies and perikarya of caudal neur ose- act :ompanied by varicosities were more cretory neurons were even observec ;1 in
FIG. 10. Injection of hypertonic choline chloride in S. fontinalis 3 hr prior to sacrifice increased the
frequency of caudal neurosecretory cells (c) with contiguous fluorescent varicosities in comparison with fresh water controls. Third preterminal vertebral level. (x2500).
FIG. 11. Injection of hypertonic Na,SO, 3 hr prior to sacrifice clearly outlined the aminergic inner- vation of caudal neurosecretory cells (c) of S. fontinalis (X 2500).
FIG. 12. Treatment of freshwater-adapted Salvelinus fontinalis with acetazolamide (30 mgkg 3 hr before sacrifice) enhanced the number of caudal neurosecretory cells (c) with contacting fluorescent varicosities at the level of the spinal cord corresponding to the 1 st preterminal vertebra. (x 2500).

198 AUDET AND CHEVALIER
some but not all fish. In the 3 out of 11 fish that survived two injections 24 and 3 hr prior to sacrifice, only a small increase of varicosities in contact with caudal neurose- cretory cell bodies in the fourth to fifth vertebral area was observed.
C. Aldactone and Acetazolamide Treatments
Aldactone, 50 mg/kg in a volume of 0.2 ml 0.85% saline, administered intraperito- neally to freshwater trout 48, 24, and 3 hr prior to sacrifice caused a slight but signifi- cant (P < 0.01) diminution of fluorescence at the level of the second and third verte- brae exclusively; however, the incidence of caudal neurosecretory cells with abutting varicosities was sharply augmented in all regions of posterior spinal cord. Acetazol- amide-treated fish (one dose of 30 mg/kg 3 hr prior to sacrifice) showed a general reduction of the fluorescence index (P < O.Ol), due primarily to a reduction in 2nd- 3rd preterminal vertebral zone (P < 0.01). However, the number of caudal neu-
IO
t
: =* : : t
ii 4
5
3hr 3-24hr 3hr FW 0.85X NaCl 0.3M 1 Cholin.
chloride
FIG. 13. Histogram of fluorescence index for the three subregions of posterior spinal cord of freshwater S. fontinalis injected with either one or two salt solu- tions (0.85% NaCl, 0.3 M Na,SO, and 0.3 M choline chloride), respectively, at 3 hr and 24 and 3 hr prior to sacrifice. Empty columns: fihm terminale and 1st terminal vertebral region, stippled columns: second to third preterminal vertebral region and striped col- umns: third to fourth preterminal vertebral region. (a) Different from uninjected freshwater fish (P < 0.01) and (b) (P < 0.05). (Sign test.) (cf. Materials and Methods).
rosecretory somata with contacting var- icosities was clearly increased in the re- gions of the j&m terminale- 1st vertebral and the third and fourth vertebral zones but not in second and third vertebral zones (Fig. 12). Vehicle-injected fish (0.2 ml 0.85% NaCl) showed no variation when compared to freshwater-adapted control fish.
DISCUSSION
In Salvelinus fontinalis adapted to fresh- water, an extensive innervation of caudal neurosecretory cells was observed. The presence of green and yellow varicosities, and the increased level of associated fluo- rescence induced by nialamide as well as its decrease following reserpine treatments, indicated specificity for biogenic amines. No attempt was made in this histoflu- orescent study to distinguish between nor- adrenaline and dopamine, those primary catecholamines yielding a similar yellow- green fluorescence after paraformaldehyde treatment. That the observed variations in the color of the fluorescence are due to dif- ferences in concentration as opposed to the presence of different species of catechol- amines, is supported by the increase in the number of yellow varicosities with a yellow color observed following nialamide treat- merit. Biochemical investigations of cate- cholamines in the carp have demonstrated the presence of noradrenaline only (Ko- bayashi, 1980); noradrenaline has also been reported to stimulate release of urotensin I in the isolated urophysis of carp (Ichikawa, 1979). The presence of mono- aminergic terminals in contact with caudal neurosecretory cells have previously been unequivocally demonstrated at the electron microscopic level with the aid of the selec- tive marker 5hydroxydopamine (Audet et al., 1980).
On the basis of afferent aminergic inner- vation, two populations of caudal neurose- cretory cells appear to be present in the trout S. fontinalis: under all the experi- mental conditions tested in the present

INNERVATION OF NEUROSECRETORY CELLS 199
study, only part of the cells appeared to ex- hibit aminergic connections, and this por- tion is concentrated topographically in the most posterior part of the whole popula- tion. This pattern of nonuniform aminergic innervation is similar to that observed in the pike (Baumgarten et al., 1970) and in the carp (Kobayashi et al., 1980), but different from that found in Tilapia (Lederis et al., 1974) and Gillichthys (Swanson et al., 1975) in which the entire population was inner- vated. The varicosities observed in each of the above-mentioned species all appeared yellow-green in color. Parenthetically, in contrast to the findings of Baumgarten (1970) and Kobayashi et al. (1979), no au- tofluorescent orange granules within caudal neurosecretory cells were evidenced in S. fontinalis.
S. fontinalis resembles Esox lucius, Tilapia mossambica, and Gillich thys mirabilis in the relative scarcity of aminer- gic innervation in the urophysis proper but in this, it differs from the carp (Kobayashi, 1980); this is not entirely unexpected in that the method used in the latter case was the glyoxylate paraformaldehyde reaction, a reaction which is known to intensify monoamine fluorescence.
While no fluorescent caudal neurosecre- tory cell somata were observed, three other cell types with aminergic perikarya were identified. Green fluorescent CSF-con- tatting neurons send their axonal projec- tions to more peripheral areas where caudal neurosecretory cells appear contacted by green varicosities; this suggests a possible functional link between both types of neurons. Such CSF-contacting cells have also been described in the posterior spinal cord of Esox lucius (Baumgarten et al., 1970) and Leucisius rutilus (Wilem and Fridberg, 1972); they are reminescent of similar paraventricular cells found in the diencephalum of cyclostomes, elasmo- branchs, and many teleost fishes and for which a role in osmoregulation has been re- peatedly proposed (cf. Santer, 1977). An- other cell type presented intense spots of
yellow fluorescence and has already been reported in the pike (Baumgarten et al., 1970) and in the carp (Kobayashi ef al., 1979, 1980). Large zones of contact be- tween some perikarya of these yellow cells and cell bodies of caudal neurosecretory cells were conspicuous in S. fontinalis after injection of hypertonic solution of choline chloride.
Coincident with this population was one which differed by virtue of its green color and the diffuse intracellular distribution of this fluorescence. This is the first report of such perikarya in fish. However, the ques- tion of axonal pathways of small aminergic perikarya and the origin of the innervation of caudal neurosecretory cells remains un- answered. The high frequency with which these aminergic neurons are found in the region of the spinal cord where caudal neu- rosecretory cells are concentrated leads one to posit a functional relationship be- tween small aminergic neurons and caudal neurosecretory cells.
In spite of its well-known limitations (Fuxe and Jonsson, 1973; Kopin et al., 1974), semiquantitative studies with the Falc k - Hillarp tee hnique are useful for comparison purposes, since in other sys- tems gross differences in fluorescence have been shown to reflect functional changes (Soest et al., 1973; Swanson et al., 1975; Fremberg and Merling, 1975; Schipper et al., 1978).
Variations observed in the present study, in the fluorescence level after changing ex- ternal salinities is in striking correspon- dence with variations in neurosecretory synthetic activity of caudal neurosecretory cells evidenced in S. fontinalis by previous ultrastructural and radioautographic studies (Chevalier, 1976, 1978). Exposure to de- ionized water for 3 days, which was found to induce stimulation of synthesis of pro- teinaceous material, is accompanied by a substantial increase of the fluorescence index, while 7-day exposure to deionized water (which brought synthetic activity back to the freshwater level) corresponds to

200 AUDET AND CHEVALIER
a reduction of fluorescence to the control level. The 3-day exposure is notable for in- creasing the number of caudal neurosecre- tory perikarya with contacting varicosities in the anterior region. In comparison with freshwater fish, this suggests an activation of this particular group of cells through af- ferent aminergic stimulation in response to exposure to demineralized water. How- ever, this activation is no longer observable in S. fontinalis after 7 days in deionized water. On the other hand, transfer of the trout from freshwater to 25% seawater for 1 day, as well as progressive adaptation to 100% seawater for 7 days, both of which accompanied by a significant reduction in aminergic index and number of beaded fi- bers and varicosities in contact with cell bodies, has been accompanied by di- minished synthetic activity of secretory material in most caudal neurosecretory cells in fish submitted to identical condi- tions (Chevalier, 1976).
Therefore, opposing osmotic stimuli equivalent on the one hand to water loading with reduction of plasma electrolyte levels and on the other hand to salt loading re- sulted in opposite variations in the amount of fluorescence in nerve terminals contact- ing caudal neurosecretory cells. However, the nature of the technique used did not allow to establish that the cells were the same in both cases. Similarly, electrophy- siological studies (Yagi and Bern, 1965) have demonstrated in Tilapia mossambica that some caudal neurosecretory neurons responded to low-sodium infusion while other neurons were stimulated by high- sodium infusion. This indirect evidence actually indicates that aminergic innerva- tion plays a role in the regulation of the osmo/ionoregulatory function of some neu- rosecretory cells in S. fontinalis and that this role is important. Since no aminergic innervation was observed in the neuro- hemal region, aminergic regulation would seem to be in relation to cell synthetic ac- tivity rather than to release.
The changes measured in both the inten- sity of fluorescence in the posterior spinal cord and in the frequency of aminergically innervated caudal neurosecretory cells in Salvelinus fontinalis after various experi- mental conditions designed to modify its hydromineral balance, are summarized in Table 1. All intraperitoneal injections of salt solutions were less efficient than external variations in salinity, in evoking differences in overall levels of fluorescence in the pos- terior spinal cord; injections of isotonic NaCl, hypertonic Na,SO,, and hypertonic choline chloride, designed to increase vol- ume of extracellular fluid and to alter sodium and chloride plasma levels, respec- tively , fail to induce specific differences.
As judged by the variation of fluores- cence observed after injections of salt solu- tions, at least under the present conditions, one is led to ask to what extent short stimuli such as injections of ionic solutions, could induce the same modifications of internal milieu as long-acting external salinity changes, which are capable of stimulating the caudal neurosecretory system, presum- ably by disrupting the hydromineral bal- ance. Every solution injected resulted in similar decreases, but this occurs exclu- sively in the region of the 2nd and 3rd pre- terminal vertebrae. In addition, the inci- dence of cells with aminergic innervation was increased in most cases in all regions of the posterior spinal cord. The common re- sponse that ionic injections induced points to a non-specific effect, possibly due to an increase of extracellular body fluid volume (2-4% body wt) as osmotic triggering fac- tor. It should be noted that injections of smaller volumes (0.2 ml/kg) of isotonic saline (used as vehicle in control experi- ments for drug treatments) failed to elicit any of the changes in fluorescence ob- served after injection of larger volumes. However, two sequential injections of hypertonic Na,S04, and choline chloride, 24 and 3 hr before sacrifice, unexpectedly resulted in no changes of fluorescence indi-

INNERVATION OF NEUROSECRETORY CELLS 201
TABLE 1 VARIATIONS OF FLUORESCENCE IN POSTERIOR SPINAL CORD
Frequency of varicosities in contact with caudal neurosecretory cells
Experimental conditions
Fluorescence j&m terminale- intensity in 1 st terminal
posterior spinal cord vertebra 2nd-3rd vertebrae
4th-5th vertebrae
External medium Demineralized water
3 days 7 days
Sea water (25%) 1 day
(100%) 7 days
Injection NaCl, 0.85%
Single Two
Na‘$304, 0.3 M Single Two
Choline chloride, 0.3 M Single Two
Drugs Aldactone Acetazolamide Vehicle
tt* -(N.S.)a
J (2nd-3rd p.v.)* -(N.S.)
;(2nd-3rd p.v.)** -(N.S.)
:(2nd-3rd p.v.)** -(N.S.)
&(2nd-3rd p.v.)* J (2nd-3rd p.v.)*
-(N.S.)
t
t t
t t t
t t t
t t t
t t t
Note. Summary of variations of fluorescence in caudal spinal cord of Salvelinus fontinalis in comparison to freshwater controls following manipulations designed to alter hydromineral metabolism. Double arrows, varia- tions of fluorescence index in the three compartments considered together. Diagonal arrows, slight variations in one compartment; p.v., preterminal vertebra.
*P < 0.01. ** P < 0.05. n Nonsignificant, according to nonparametric statistics (sign test).
ces similar to those produced by a single 1 ml/kg injection. This hypothesis presup- poses that differences observed in fluores- cence after various treatments are all cor- related with innervation of the same cell population.
Alternatively, one cannot reject the pos- sible interpretation that changes in fluores- cent varicosities surrounding caudal neuro- secretory cells might be due, to some ex- tent, to a generalized change in aminergic varicosities in the central nervous system due, for example, to shock; since the most
anterior segment of the spinal cord studied here (containing mainly motoneuron cell bodies) showed parallel variations in the number of varicosities accompanying cell bodies in every injection experiment.
Intraperitoneal administration of both acetazolamide and aldactone in order to lower plasma sodium level, produced mod- ifications of fluorescence similar to those observed after injection of salt solutions. This suggests that the action of these inhib- itor of Na influx in gills of freshwater- adapted fish has a functional correlate in

202 AUDET AND CHEVALIER
the aminergic regulation of caudal neurose- cretory cells.
Thus, it can be concluded that the vari- ous changes observed in the fluorescence concentration of monoaminergic innerva- tion of caudal neurosecretory cells of Sal- velinus fontinafis after modification of ex- ternal salinity, injection of ionic solutions, and administration of drugs affecting hy- dromineral metabolism strongly suggest aminergic participation in the control of caudal neurosecretory cells although no specific stimulating signal for this nervous control could be definitely identified.
ACKNOWLEDGMENTS
The authors wish to thank Dr. Andre Parent from Universite Laval, Quebec, for technical advices on Falck-Hillarp technique, and Dr. Serge Boileau and Mrs. Andree Beauregard-Lacroix for technical assis- tance. We are also grateful to Searle Pharmaceuticals for providing Aldactone, and to Mr. S. Sacks for cor- recting English manuscript.
REFERENCES
Audet, C. (1980). “Innervation du systeme caudal neurosecreteur de la truite mouchetee, Salvelinus fontinalis en relation avec l’osmoregulation.” M. SC. Thesis. Universite du Quebec P Montreal, Canada.
Audet, C., Gauthier, L., and Chevalier, G. (1980). Monoaminergic control of the caudal neurose- cretory system of the trout Salvelinusfontinalis in relation to osmoregulation. In “Eight Interna- tional Symposium on Neurosecretion” (D. Farner and K. Lederis, eds.), Plenum, New York, in press.
Baumgarten, H. G., Falck, B., and Wartenberg, H. (1970). Adrenergic neurons in the spinal cords of the pike (Esox lucius) and their relation to the caudal neurosecretory system. 2. Zellforsch. 107, 479- 498.
Bennet, M. V. L., and Fox, S. (1962). Electrophysiol- ogy of caudal neurosecretory cells in the skate and fluke. Gen. Comp. Endocrinol. 2, 77-95.
Bern, H. A., Yagi, K., and Nishioka, R. S. (1965). Structure and function of the caudal neurosecre- tory system of fishes. Arch. Anut. Microscop. Morphol. Exp. 54, 217-238.
Bourguet, J., Lahlouh, B., and Maetz, J. (1964). Mod- ifications experimentales de l’equilibre hydromin- era1 et osmoregulation chez Curussius uurutus. Gen. Comp. Endocrinol. 4, 563-576.
Chart, D. K., Chesterston, I., Henderson, I. W., and
Rankin, J. C. (1967). Studies on the experimental alteration of water and electrolyte composition in the eel (Anguillu unguillu L.). J. Endocrinol. 37, 297- 3 17.
Chevalier, G. (1976). Ultrastructural studies of caudal neurosecretory cells of Salvelinus fontinulis in relation to external salinity. Gen. Comp. Endo- crinol. 29, 44 1 - 454.
Chevalier, G. (1978). In vivo incorporation of (3H) leucine and (3H) tyrosine by caudal neurosecre- tory cells of the trout Sulvelinusfontinulis in rela- tion to osmotic manipulations. A radioautographic study. Gen. Comp. Endocrinol. 36, 223-228.
Falck, B., Hillarp, N. A., Thieme, G., and Torp, A. (1962). Fluorescence of catecholamines and re- lated compounds condensed with formaldehyde. J. Histochem. Cytochem. 10, 348-354.
Fremberg, M., and Meurling, P. (1975). Catechol- amine fluorescence in the pituitary of the eel, Anguillu unguilla, with special reference to its variation during background adaptation. Cell. Tiss. Res. 157, 53-72.
Fridberg, G. (1963). Electron microscopy of the caudal neurosecretory system in Leuciscus rutilus and Phoxinus phoxinus. Acta Zool. (Stockh.) 44, 245- 267.
Fridberg, G., Bern, H. A., and Nishioka, R. S. (1966). The caudal neurosecretory system of the isospon- dilus teleost, Albulu vulpes from different habitats. Gen. Comp. Endocrinol. 6, 195-212.
Fuxe, K., and Jonsson, G. (1973). The histochemical fluorescence method for the demonstration of catecholamine. J. Histochem. Cytochem. 21 (4) 293-311.
Ichikawa, T. (1978). Acetylcholine in the urophysis of several species of teleosts. Gen. Comp. Endo- crinol. 35, 226-233.
Ichikawa, T. (1979). Release of urotensin I by neuro- transmitters and ultrastructural changes in the carp urophysis in vitro. Gunmu Sympo. Endo- crinol. 16, 87-96.
Ishibashi, T. (1962). Electrical activity of the caudal neurosecretory cells in the eel Anguillu juponicu with special reference to synaptic transmission. Gen. Comp. Endocrinol. 2, 415-424.
Kerstetter, T. H., Kirschner, L. B., and Rafuse, D. (1970). On the mechanisms of sodium ion trans- port by the irrigated gills of rainbow trout (Sulma guirdneri). J. Gen. Physiol. 56, 342-359.
Kobayashi, H., Uemura, H., Oota, Y., and Ishii, S. (1963). Cholinergic substance in the caudal neuro- secretory storage organ of fish. Science 141, 714-716.
Kobayashi, Y., Ichikawa, T., and Kobayashi, H. (1979). Innervation of the caudal neurosecretory system of the teleost. Gunmu Sympo. Endocrinol. 16, 81-86.
Kobayashi, Y., Kobayashi, H., Ohshiro, S., Osumi,

INNERVATION OF NEUROSECRETORY CELLS 203
Y., and Fujiwara, M. (1980). Monoaminergic in- nervation of the caudal neurosecretory system of the carp, Cyprinus carpio. Zbl. Vet. Med. C. Anat. Histol. Embryol. 9, 65-72.
Kopin, I. J., Palkovitz, M., Kobayashi, R. M., and Jacobowitz, D. M. (1974). Quantitative relation- ship of catecholamine content and histofluores- cence in brain of rats. Brain Res. 80, 229-235.
Lederis, K., Bern, H. A., Medakovic, M., Chan, D. K. O., Nishioka, R. S., Letter, A., Swanson, D., Gunther, R., Tesanovic, M., and Horne, B. (1974). Recent functional studies on the caudal neurosecretory system of teleost fishes. In “Neurosecretion-The Final Neuroendocrine Pathway” (F. Knowles and L. Vollrath, eds.), pp. 94- 103. Springer-Verlag, New York.
Lederis, K., Gill, V., and Ichikawa, T. (1978). Affer- ent stimulation of cerebral and urophysial neuro- secretory neurons in fishes, in “VIIIth Interna- tional Symposium on Comparative Endocrinol- ogy” (P. J. Gaillard and H. H. Boer, eds.), pp. 305- 308. Elsevier/North-Holland, Amsterdam.
Luppa, H., Weiss, J., und Feustel, G. (1968). His- tochemische Untersuchungen zur Lokalisation von Acetylcholinesterase, Monoaminooxidase und Monoaminen im kaudalen neurosekretoris- then System von Cyprinus carpio. Z. Zellforsch. 89, 499-- 508.
Maetz, J., and Garcia-Romeu, F. (1964). The mecha- nism of sodium and chloride uptake by the gills of a freshwater, Carassius auratus. II. Evidence for NH,+/Na+ and HCO,-/Cl- exchanges. J. Gen. Physiol. 47, 1209- 1227.
Mayer, N., and Nibelle, J. (1970). Kinetics of the min- eral balance in the eel Anguilla anguilla in relation to external salinity changes and intravascular saline infusions. Comp. Biochem. Physiol. 35, 553- 566.
Mayer-Gostan, N., and Kirsch, R. (1979). Kinetic study of Na+ and Cl- exchanges in freshwater eels following intravascular infusion of sodium chloride, sodium sulfate and choline chloride. Comp. Biochem. Physiol. 63A, 415-422.
Mendenhall, W. (1971). “Introduction to Probability and Statistics,” 3rd Ed. Duxbury Press, Bel- mount, Calif.
Olivereau, M. (1964). Action de l’aldactone sur l’inter- renal anterieur de I’Anguille. J. Physiol. 56, 625- 626.
Parent, A., Dube, L., Bradford, Jr., M. R., and Northcutt, R. G. (1978). The organization of monoamine-containing neurons in the brain of the sunfish (Lepomis gibossus) as revealed by fluo- rescence microscopy. J. Comp. Neurol. 182, 495- 516.
Sano, Y., Iida, Y., und Takemoto, S. (1966). Weitere elektronenmikroskopische Untersuchungen am Kaudalen neurosekretorischen System von Fis- then. Z. Zellforsch. 75, 328- 338.
Santer, R. M. (1977). Monoaminergic nerves in the central and peripheral nervous systems of fishes. Gen. Pharmacol. 8, 157- 172.
Soest, S. W., Farner, D. S., and Oksche, A. (1973). Fluorescence microscopy of neurons containing primary catecholamines in the ventral hypothala- mus of the white-crowned sparrow, Zonotrichia leucophrys gambelii. Z. Zellforsch. 141, l- 18.
Swanson, D. D., Nishioka, R. S., and Bern, H. A. (1975). Aminergic innervation of the cranial and caudal neurosecretory systems in the teleost Gil- lichthys mirabilis. Acta Zool. (Stockh.) 56, 225- 237.
Uemura, H. (1965). Histochemical studies of the dis- tribution of cholinesterase and alkaline phos- phatase in the vertebrate neurosecretory system. Annot. Zool. Japan 38, 79-96.
Wilen, P., and Fridberg, G. (1973). Ultrastructural studies on the ontogenesis of the caudal neuro- secretory system in the roach Leuciscus rutilus. Z. Anat. Entwickl.-gesch. 139, 207-216.
Yagi, K., and Bern, H. A. (1965). Electrophysiologic analysis of the response of the caudal neurose- cretory system of Tilapia mossambica to osmotic manipulations. Gen. Comp. Endocrinol. 5, 509- 526.





























