Embed Size (px)
Citation preview

J Comp Physiol A (1988) 163:489-506 Joumal of
Sensory, Comparative .~u~,, and
Physiology A ~ a . ~ . . , Physiology
�9 Springer-Verlag 1988
Morphological and electrophysiological properties of a novel in vitro preparation: the electrosensory lateral line lobe brain slice
William B. Mathieson* and Leonard Maler Department of Anatomy, Faculty of Health Sciences, University of Ottawa, Ottawa, K1H 8M5, Canada
Accepted March 4, 1988
Summary. An in vitro brain slice preparation of the electrosensory lateral line lobe (ELL) of weakly electric fish was developed. The morphology of this slice was studied and revealed that most ELL neu- rons and synapses retained their normal appear- ance for at least 10 h in vitro. The electrophysio- logical characteristics of the main ELL output neu- rons, the pyramidal cells, were measured. Extracel- lular electrode recordings demonstrated that pyra- midal cells are capable of spontaneous, rhythmic spike activity. Intracellular recordings showed that intrinsic oscillations in membrane potential under- lie the bursting behavior. The majority of pyrami- dal cells respond to depolarizing current pulses with an initial lag in spike firing followed by a non-accommodating, higher frequency spike train.
Time and voltage-dependent properties of pyr- amidal cell responsiveness, as well as the effects of pharmacological blocking agents indicated that rhythmic activity and repetitive firing are domi- nated by a persistent, subthreshold sodium con- ductance (gNa) which activates at depolarized lev- els and is the driving force behind the membrane potential oscillations and the sustained (non-ac- commodating) spike firing. In addition, a tran- sient, outward potassium conductance (gA) is re-
Abbreviations: A CSF artificial cerebrospinal fluid; 4-AP 4-ami- no-pyridine; BP basilar pyramidal cell; D M L dorsal molecular layer; EGTA ethyleneglycol-bis(B-aminoethylether)-N,N'te- traacetic acid; ELL electrosensory lateral line lobe; EOD elec- tric organ discharge; EPSP excitatory postsynaptic potential; FPP fast prepotential; IPSP inhibitory postsynaptic potential; L Y Lucifer Yellow; NBP non-basilar pyramidal cell; Rin input resistance; SP slow potential; TEA tetraethyl ammonium; tsf tractus stratum fibrosum; T T X tetrodotoxin; Vrn membrane potential; V M L ventral molecular layer
* Present address : Department of Medical Physiology, Faculty of Medicine, University of Calgary, 3330 Hospital Dr., N.W., Calgary, Alberta, Canada, T2N 4N1
sponsible for the lag in spike firing by acting as a ' brake' during the initial 50-200 ms of a depolar- izing stimulus.
Calcium currents and calcium-dependent po- tassium conductance add to the interval between spontaneous bursts but appear insufficient for spike frequency accommodation.
The in vitro behaviour of pyramidal cells differs substantially from the behaviour of the same cell type in vivo. These observations raise possibilities that intrinsic membrane properties together with local synaptic interactions may regulate pyramidal cell responsiveness.
Introduction
Certain weakly electric fish of the order gymnoti- form (apteronotids) emit a nearly sinusoidal, con- stant frequency electric organ discharge (EOD). Electroreceptor organs distributed about the body surface are highly sensitive to electric field distur- bances caused by nearby objects or to mixing of their own EOD with those of neighboring fish. Two physiologically distinct types of tuberous re- ceptors encode changes in EOD amplitude (P-type afferents) or timing of the zero-crossing of the EOD waveform (T-type afferents) (Scheich et al. 1973). Both types of electroreceptive afferents pro- ject centrally to form 3 complete somatotopic maps of the body surface within the electrosensory later- al line lobe (ELL) of the medulla (Carr et al. 1982; Heiligenberg and Dye 1982).
The ELL is a highly laminated structure, con- taining at least 11 distinct cell types (Maler 1979). The main components of the ELL circuitry are illustrated schematically in Fig. 1. This study fo-

490 W.B. Mathieson and L. Maler: ELL pyramidal cell electrophysiology in vitro
pf ::!---- - - - - ~ !- --=!~--: 1~1~11111 VM[_
I ' / ~ j / Dh k
P-afferentl T-afferent �9 Fig. l. Simplified schematic diagram of the ELL circuitry. Pri- mary afferent input from tuberous and ampullary receptors at the fish's body surface form the deep fiber layer (DFL) on the ventral surface of the ELL. The pyramidal cell layer (PYR) contains cell bodies of 3 different classes of neurons: basilar pyramidals (BP), non-basilar pyramidals (NBP) and poly- morphic cells (POL). Basilar pyramidals possess a long basilar dendrite which extends ventrally into the deep neuropil (DNL) where it receives excitatory synaptic input (open circles) from P-type afferent terminals. Non-basilar pyramidals lack a basilar dendrite and therefore do not receive direct primary afferent input. Pyramidal cell axons leave the ELL via the plexiform layer (not shown). Two classes of interneurons make inhibitory synaptic contact (closed circles) with basilar and non-basilar pyramidals: granule cells, which occupy the granule cell layer (Gr), and polymorphic cells. BPs, NBPs, polymorphic cells and a subset of granule cells have large apical dendrites which ex- tend dorsally into the molecular layers. Here they receive de- scending, excitatory contact: patallel fibers (p.f.) from neurons in the nucleus praeeminentialis enter the ELL via the tractus stratum fibrosum (TSF) and ramify in the ventral molecular layer (VML). PFs originating in the lobus caudalis of the cerbel- lum ramify in the dorsal molecular layer (DML)
cuses on the electrical behavior of 2 main ELL output neurons : basilar and non-basilar pyramidal cells, also referred to as E and I cells according to physiological criteria (Saunders and Bastian 1984). The cytology of the ELL and the synaptic connectivity of pyramidal neurons led to the pre- diction that basilar pyramidal cells (BPs) would respond to increased electrosensory input with a central excitatory receptive field, flanked by a larger annulus of surround inhibition. Conversely, non-basilar pyramidals (NBPs) were predicted to respond with an opposite receptive field pattern (e.g. off-center, on-surround) (Maler et al. 1981 b). These predictions were subsequently verified by physiological recordings in vivo (Bastian 1981b, 1986a, b; Saunders and Bastian 1984).
Pyramidal cells also receive a large descending input to their apical dendrites in the molecular layer (Maler 1979). The dorsal molecular layer (DML) contains parallel fibers originating from
granule cells of the posterior eminentia granularis, whereas parallel fibers in the ventral molecular layer (VML) originate in the nucleus praeeminen- tialis. Both descending inputs form a recurrent pathway for electrosensory signal analysis by ELL pyramidal neurons (Sas and Maler 1987).
Electrophysiological studies employing several species of gymnotiform fish have examined the responsiveness of pyramidal cells to a variety of electrical stimuli. Enger and Szabo (1965) first re- ported that ELL pyramidal neurons are sensitive to motion and direction of objects placed in their receptive field. Many of the units responded in a phasic manner, leading the authors to speculate on their possible role as edge detectors for moving objects. More recently, Bastian (1981 b) measured the sensitivity of E and I units to object movement; both cell types responded abruptly to amplitude modulations, and rapidly adapted their spike fre- quencies to near-baseline levels.
It is important to note that this phasic firing pattern can only partially be accounted for by re- ceptor adaptation. Recordings of receptor activity in response to amplitude modulations showed that P-type afferents are only partially and slowly adapting (Bastian 1986a; Hopkins 1976; Shum- way and Maler, submitted). However, convergence of afferent input onto pyramidal cells results in a dramatic increase in the degree of phasic firing (Bastian 1981b, 1986a). The disparity in temporal coding at receptor vs. pyramidal cell level suggests that pyramidal cells have a spike adaptation mech- anism that may be mediated in part by (1) interac- tions of pyramidal cells with local inhibitory inter- neurons (Maler et al. 1981 b) or (2) by intrinsic bio- physical properties of the pyramidal cells them- selves; it is also possible that fatigue or autoinhi- bition of primary afferent terminals may contrib- ute to adaptation. Since intracellular recording from pyramidal cells is extremely difficult in vivo, very little is known about possible intrinsic mecha- nisms that may govern their response characteris- tics.
Further studies in vivo demonstrate that ELL output is regulated by the activity of descending input to the DML. This feedback pathway acts as a gain control mechanism capable of altering E and I cell sensitivity to electrosensory input, spike frequency adaptation and their receptive fields as mapped with moving objects (Bastian 1986 a). Moreover, the same descending input me- diates adjustments in pyramidal cell responsiveness to compensate for fluctuations in EOD intensity (Bastian 1986b). The synaptic connectivity in- volved in parallel fiber input to the ELL suggests

W.B. Mathieson and L. Maler: ELL pyramidal cell electrophysiology in vitro 491
that such a gain control is mediated indirectly via local inhibitory interneurons (Maler et al. 1981 b). Again, however, an unexplored possibility remains that descending control may also act directly on pyramidal cells to change their intrinsic firing properties.
In order to characterize pyramidal cell electro- physiology at the intracellular level, we have devel- oped an ELL slice preparation. Measurements of resting behavior and stimulus-evoked spike firing, together with the ionic conductances which under- lie pyramidal cell activity in vitro indicate that pyr- amidal cells undergo dramatic changes in their fir- ing characteristics when dissociated from the nor- mal peripheral input and descending control. These results identify several conductances intrin- sic to pyramidal cell membranes which may serve as targets for synaptic modulation of electrosen- sory processing during appropriate behavioral or environmental conditions.
Materials and methods
Slice preparation. ELL brain slices were prepared from the gym- notiform species, Apteronotus leptorhynchus, maintained at 25 ~ in aquarium water adjusted for conductivity to 5-10 kf2. cm. Adult fish (> 15 g) were pretreated by injecting (i.m.) 25 gl of solution containing 0.25 mg 2-amino-5-phosphovaleric acid (2-APV) and 1.2 mg L-glutamic acid diethylester (GDEE), two acidic amino acid antagonists which interfere with N-methyl-D- aspartic acid (NMDA) and quisqualate-type receptor binding (McLennan 1983). The injected fish were kept for 20 min in oxygenated tank water and anesthetized with diethylether prior to surgery (approx. 5 ml ether stirred into 1 L tank water). The rationale for this protocol was that the ELL contains signif- icant quantities of glutamate and aspartate (Nadi and Maler 1987), two excitatory neurotransmitters. In vivo studies indicate that blockade of their receptor binding can protect hyperexcit- able tissue from convulsive activity (Meldrum et al. 1983) and reduce the damaging effects of brain anoxia (Gerhardt et al. 1986). Additional studies implicate anoxia in the development of spreading depression (Milan et al. 1985), a phenomenon which is suppressed by ether anesthesia (van Herreveld and Stature 1953). Without these pretreatments, the viability of the ELL slice was greatly reduced; intraceltular electrode recording indicated that a majority of neurons had undergone depolariza- tion block and had been rendered electrically unresponsive, a condition that was prevented by the above treatments.
The anesthetized fish were then submerged in a small dish of ice-cold artificial cerebrospinal fluid (ACSF) (see below) oxy- genated with a 95% O2/COz mixture. After dissecting away the cranium, a razor cut ranseeted the brain transversely to the longitudinal axis of the ELL. This placed the slice angle parallel to the orientation of the pyramidal cells' apical den- drites and thus minimized their amputation. A transverse orien- tation had an additional benefit of sparing the majority of par- allel fiber afferents originating in the caudal lobe, allowing orthodromic activation of the ELL circuitry via descending in- put.
All cranial nerves, the ganglia of the anterior lateral line nerve (ALLN) and the spinal cord were quickly severed and
the tissue block containing the ELL was removed from the skull. The tissue was then fixed to a small aluminium block with a drop of cyanoacrylate glue, submerged in chilled ACSF and sectioned with a vibrating microtome to an optimal thick- ness of 500-700 gin. The slices were transferred to a nylon mesh in a bath and incubated either submerged or at the interface between ACSF and the humidified, oxygenated atmosphere in the bath chamber. The total time elapsed between excision of the ELL and the placement of the slice onto the bath was 7-9 min.
Since the composition of gymnotiform CSF had not been analyzed, ACSF was developed based on the composition of mammalian ACSF (Schwartzkroin 1977; Schubert and Mitz- doff 1979) and modified according to our empirical observa- tions. Standard ACSF solution contained (in mM): NaC1 131; NaHCOa 20; KCt 2; KH2PO 4 1.25; MgSO4 2; CaCI2 2.5; dextrose 10; pH set to 7.4 before bubbling with 95% 02/5% COg. Test solutions containing TEA + or 4 - A P were osmoti- cally balanced by appropriate reductions in NaC1. For ACSF containing Mn z+ (2-10 mM), Ca + + was reduced to 0.2 mM, MgSO 4 was omitted to prevent precipitation and pH was buf- fered to 7.4 with 20 m M HEPES, 11.4 m M TRIS.
Recording apparatus. Glass microelectrodes (WPI capillary tub- ing) were filled with either 2 M NaC1 for extracetlular recording (10-20 Mr2 resistance) or 3 M KC1 for intracellular recording (50-100 M~? resistance). In some experiments cells were im- paled with electrodes filled with 2 M potassium methyl sulfate or 3 M potassium acetate. To improve the stability of intracellu- lar recordings, electrode tips were coated with a silicon solution ( 'Aqua-Sil ' , 0.4%; Pierce Chemical Co.). This promoted seal- ing of the hydrophobic plasma membrane to the charged glass tubing following impalement of the cell.
The recording electrode was positioned over the pyramidal cell layer under visual guidance and lowered into the tissue with a Midgard Microstep driver. For intracellular recording, pyramidal cells were stimulated directly via a high impedance bridge balance circuit on the preamplifier. Following the initial impalement of the cells, hyperpolarizing current (~0.4 nA) was frequently passed through the recording electrode for ~ 3 min to help stabilize the membrane potential (Vm). Bipolar, Ag- AgC1 electrodes were used to activate pyramidal cells ortho- dromically, or antidromically by stimulating granule cells in the lobus caudalis or axons in the plexiform layer, respectively.
Passive electrical properties of the membrane, such as rest- ing potential (Vm), input resistance (Rin), and time constant (rm) were routinely measured during intracellular recording. VR was measured by recording the voltage drop after penetration of the electrode. Rin was measured by delivering 30 ms hyperpo- larizing pulses to the cell and calculating the slope of the cur- rent/voltage regression line. The membrane time constant was measured by delivering a square pulse of intracellular hyperpo- larizing current (e.g. 0.3 hA, 80 ms duration) and plotting time vs. (1-Vo/Vm~x) (semilog plot; Connors and Kriegstein 1986).
All intracellular recording data presented here were taken from cells which met the following criteria: the electrode was situated within the pyramidal cell layer of the lateral, centro- lateral or centro-medial segments of the ELL. All cells used in the study had a Vm < -- 60 mV and action potential amplitude > 60 mV. Each unit was held at least long enough for the Vm to stabilize and measure the passive membrane properties as well as spontaneous firing behaviour.
EGTA injection. The effect of removing free intracellular C a 2 +
on pyramidal cell firing was tested by injecting the specific calci- um chelator, EGTA (ethyleneglycol-bis(-aminoethyl ether)- N,N'-tetra-acetic acid) directly into the cell soma. Hyperpolar-

492 W.B. Mathieson and L. Maler: ELL pyramidal cell electrophysiology in vitro
izing current (0.3-0.7 nA DC for up to 1 h) was passed through micropipettes filled with a solution of 1.8 M K acetate plus 0.2 M K EGTA.
Intracellular labelling. Identification of cell types recorded phys- iologically was accomplished by intracellular injection of the fluorescent dye, Lucifer Yellow (LY). Electrodes were filled with a 4% solution of LY inl M lithium chloride (R=100- 150 M~2). The dye was injected by passing i nA hyperpolarizing pulses through the recording electrode (1 Hz; 200 ms duration) for as long as possible before losing the unit (e.g. 45 min).
The electrode was pulled up and the tissue emersed in 4% formaldehyde in 0.1 M Na phosphate buffer, pH 7.4 overnight. The fixed slice was glued to a vibratome block and cut into 100 gm thick sections and washed in buffer. The sections were dehydrated through a graded series of alcohol, cleared and cov- erslipped in dimethyl sulfoxide (DMSO) (Grace and Llinfis 1985). The resulting slides were viewed with epifluorescence microscopy through an FITC filter and photographed.
Cytology. After the recording session, representative slices were prepared for histological examination by immersion directly into a 2% paraformaldehyde/2% glutaraldehyde solution in 0.12 M Na-cacodylate buffer (pH 7.4) and stored overnight at 4 ~ The slices were then osmicated, dehydrated and embedded in Araldite. The tissue blocks were trimmed down to the depth and location that included pyramidal cells recorded physiologi- cally, and sectioned for light and electron microscopy.
Results
Viability of slices
The tissue was considered viable when, during each pass of the electrode through the slice, several cells could be punctured and recorded intracellularly. The majority of these units had resting potentials of 60 mV or greater. Following a 1-2 h recovery period, it was typical of the healthiest slices to re- main active for up to 10 h.
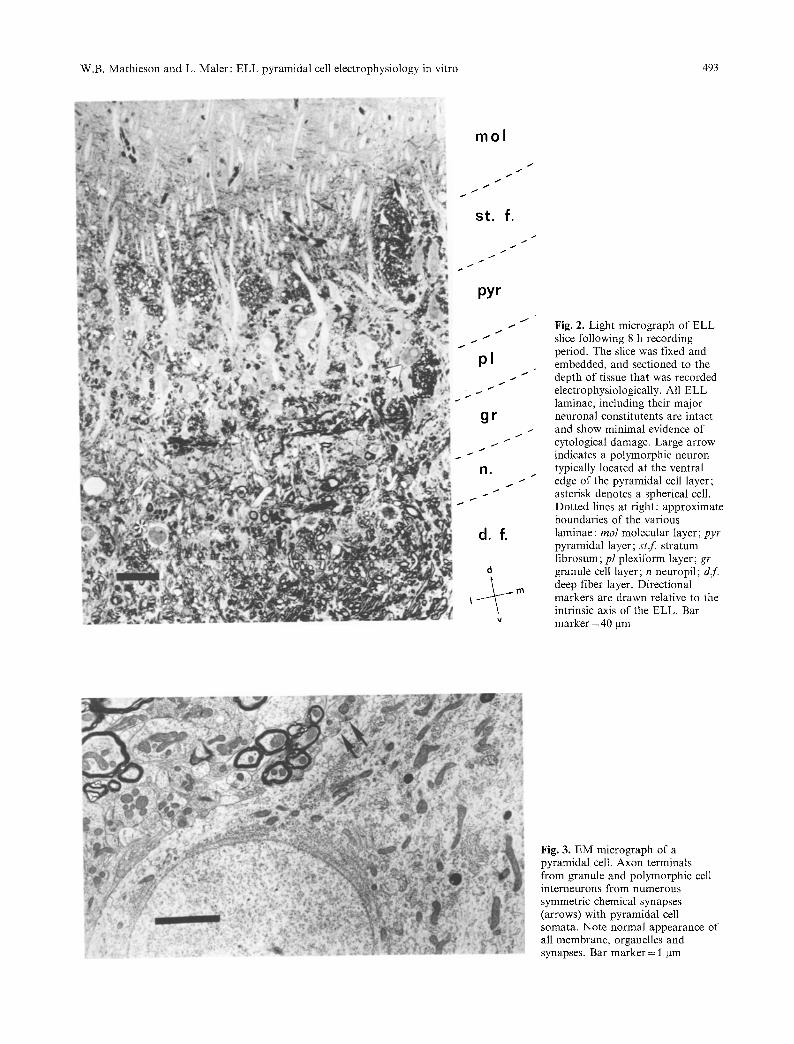
Areas of tissue which contained electrode re- cording sites were examined with light and electron microscopy so that a correlation could be made between the cytological appearance and the viabi- lity of the tissue. Recordings were made in all three ELL segments which receive tuberous electrorecep- tive input (Heiligenberg and Dye 1982; Mathieson et al. 1986). The viability and appearance of the slices were identical in each of these segments. Fig- ure 2 illustrates the appearance of an area of an ELL slice which had been recorded in vitro for 8 h. All of the ELL laminae and major cell types described in vivo were easily recognizable and dis- played minimal evidence of disruption (Figs. 2, 3). As expected, the greatest damage occurred within the laminae containing fiber tracts oriented per- pendicular to the plane of section. As a conse- quence, the deep fiber layer (dr), tractus stratum fibrosum (tst) and to a lesser extent the plexiform
layer (pl) contained varying amounts of extracellu- lar edema and vacuolated axonal processes. Unra- velling of the myelin sheath frequently occurred, particularly with the larger fibers. In spite of the disruption of these fiber tracts, the majority of ax- ons displayed normal internal ultrastructure; mito- chondrial swelling was not apparent and neurofila- ments were evenly spaced throughout the axo- plasm.
In the neuropil layer, electrosensory afferent terminals synapsed with dendritic processes from basilar pyramidal cells, granule cells, polymorphic cells and spherical cell somata. Although occasion- al evidence of darkened terminals and separation of synapticjunctions were found, most electrotonic and chemical synapses were qualitatively similar to those reported in vivo (Maler etal. 1981b; Mathieson et al. 1986).
Granule cells, polymorphic cells, basilar and non-basilar pyramidal cells appeared normal, al- though occasional pycnotic pyramidal cell bodies were present in even the most healthy slices. It was also possible to identify axon terminals from polymorphic and granule cell interneurons synaps- ing with pyramidal cell somata. Apical dendrites from these cell types remained intact in the ventral and dorsal molecular layers where they were often observed receiving asymmetric chemical synapses from descending parallel fibers.
Other local interneurons, such as neurons of the ventral molecular layer and stellate cells, were identified in the ELL slice and appeared normal at the light microscopic level (Fig. 2). These cells are relatively scarce and were not identified in EM sections.
Intracellular labelling with Lucifer Yellow
Since all of the recording data presented in this study were taken from the pyramidal cell layer, representative samples of intracellular units were injected with Lucifer Yellow (LY) in an attempt to correlate electrophysiological behaviour with the morphological identity of the cell. Of ten cells filled with LY, 6 were identified as non-basilar pyr- amids and 4 were basilar pyramids. Figure 4 illus- trates the resulting fluorescence micrographs. Basi- lar and non-basilar pyramidal cell somata are ap- proximately of equal diameter and are alternately distributed across the ELL (Maler 1979), a pattern which reflects the nearly equal numbers of each cell type injected with LY. Although it was possible that the electrode data presented in the following sections included occasional representation from polymorphic cells, it is reasonable to assume that
W.B. Mathieson and L. Maler: ELL pyramidal cell electrophysiology in vitro 493
mol
/
I
/
t
st. f. I
pyr
/
t /
I /
t
pl
gr /
/
I1. s
t
s
d , f.
"4
Fig. 2. Light micrograph of ELL slice following 8 h recording period. The slice was fixed and embedded, and sectioned to the depth of tissue that was recorded electrophysiologically. All ELL laminae, including their major neuronal constitutents are intact and show minimal evidence of cytological damage. Large arrow indicates a polymorphic neuron typically located at the ventral edge of the pyramidal cell layer; asterisk denotes a spherical cell. Dotted lines at right: approximate boundaries of the various laminae : tool molecular layer; pyr pyramidal layer; st .f stratum fibrosum; pl plexiform layer; gr granule cell layer; n neuropil; dr. deep fiber layer. Directional markers are drawn relative to the intrinsic axis of the ELL. Bar marker = 40 gm
Fig. 3. EM micrograph of a pyramidal cell. Axon terminals from granule and polymorphic cell interneurons from numerous symmetric chemical synapses (arrows) with pyramidal cell somata. Note normal appearance of all membrane, organelles and synapses. Bar marker = 1 ~tm

494 W.B. Mathieson and L. Maler: ELL pyramidal cell electrophysiology in vitro
Fig. 4A, B. Lucifer Yellow-injected basilar and non-basilar pyr- amidal cells. A This cell was identified as a non-basilar pyrami- dal cell by its morphology and position in the ELL. A large apical dendrite extends dorsally into the ventral and dorsal molecular layers where it bifurcates and divides into an exten- sive apical dendritic tree. Numerous somatic dendrites are visi- ble extending a short distance into the surrounding pyramidal cell layer. Conspicuously absent is a basilar dendrite, present only in basilar pyramidal cells. B This cell was identified as a basilar pyramidal cell due to the presence of its long, basilar dendrite, which extends ventrally into the deep neuropil layer where it receives direct primary afferent input. The basilar den- drite is smooth and unbranched as it descends through the plexiform and granule cell layers, then divides into an extensive basilar bush within the neuropil. Note that the BP has fewer somatic dendrites than the NBP, but has a long apical dendrite
the large majority of recordings were from basilar and non-basilar pyramidal cells.
Non-basilar pyramidals had cell bodies which measured approximately 20 x 60 gm, a single api- cal dendrite which bifurcated into 15-20 branches ramifying into the full extent of the ventral and dorsal molecular layers (Fig. 4A). Due to the diffi- culty in reconstructing the apical dendritic tree in
LY-filled cells, the full dimension of the apical den- drites was not measured. Partial reconstruction measurements indicated that they extended a mini- mum of 340 txm dorsoventrally by 120 gm trans- versely. From the soma emerged numerous tiny somatic dendrites which arborized within the pyra- midal layer. The initial segment and proximal por- tion of the single axon was usually labelled, al- though the LY did not diffuse far enough along the axon to describe its course. None of the la- belled pyramidal cells showed any evidence of dye- coupling with adjacent neuronal processes. Allow- ing for species size differences and histological shrinkage, the somatic and dendritic measurements were compatible with those previously reported for NBPs (Maler 1979; Saunders and Bastian 1984).
Figure 4B illustrates the elongated shape of a LY-filled basilar pyramidal cell. The soma mea- sured approximately 20 gm across and usually pre- sented fewer somatic dendrites than the NBP, con- sistent with Maler's (1979) Golgi data. Instead, a single highly elongated basilar dendrite descended approximately 200-400 gm, arborizing within the deep neuropil. This basilar trunk has a diameter of approximately 10 gm, substantially greater than the 2-3 gm reported by Maler (1979). The basilar dendrite terminated in a compact bush, 30-50 Ixm in diameter. The apical dendrite resembled that of an NBP.
The 200-400 lain length of the basilar dendrite of LY-filled BPs was considerably greater than that reported by either Maler (1979) (40-70 gin) in Eigenmania or Saunders and Bastian (ave. 137 ~tm) in Apteronotus; presumably this was due to less shrinkage in slices dehydrated and cleared in DMSO (Grace and Llinfis 1985). From the pro- file of the BP in Fig. 4, it is possible that primary afferent input, which contacts the basilar dendrite at its distal bush in the neuropil (Maler et al. 1981 b), might be electrotonically distant from the soma and the spike initiating initial segment. Possi- ble functional significance of these measurements will be elaborated upon in the Discussion.
Synaptic connectivity
In transverse slices, synaptic activation of pyrami- dal cells was achieved by stimulation of descending parallel fibers from the lobus caudalis. Figure 5 A illustrates synaptic potentials recorded from pyra- midal cells following stimulation of the descending input. Activation of primary afferent fibers was less successful at evoking synaptic potentials. Pri- mary afferents reach the ELL by coursing longitu- dinally in the caudal direction. Therefore, in the

W.B. Mathieson and L. Maler: ELL pyramidal cell electrophysiology in vitro 495
2 m s
B
2 sec
Fig. 5. A Excitatory postsynaptic potentials (EPSPs). Stimula- tion of granule cells in the lobus caudalis activates descending input from parallel fibers which, in turn, contact pyramidal cell apical dendrites. Following a 3-6 ms delay, suprathreshold stimulation evoked all-or-nothing spikes or bursts of spikes. Hyperpolarizing the cell with 0.2-0.4 nA steady-state current injection prevented the cell from reaching threshold for spike initiation, revealing an EPSP. B Spontaneous, hyperpolarizing inhibitory postsynaptic potentials (IPSPs) were occasionally en- countered. In this recording, a depolarizing slow potential with a burst of spikes is visible at the start of the trace, followed by rhythmic, hyperpolarizing IPSPs
1.000 ~ . ~
> [
u~
0 .100
0.070 I 0 5 10 15 20 25 30
TIME (MSEC)
Fig. 6. Insert: affect of a hyperpolarizing intracellular current pulse (0.3 nA, 80 ms) on a pyramidal cell. Time axis. 10 ms; vertical axis 5 mV, i nA. Semilog plot of (1-V/Vmax) vs time. This cell has a time constant of about 9 ms. Note that the first 3-4 points (about 1 ms) have a steeper decline which is not well approximated by the exponential. This time period is contaminated by electrode artefact and so curve peeling was not used. The deviations from exponential decay which occur at later times (15 ms) may represent slowly activating nonlinear conductances, but these do not greatly affect the estimates of the time constant
transverse plane, many of their fibers are perpen- dicular to the orientation of the slice and are sec- tioned as they enter the ELL, rendering them unre- sponsive. If it had been desirable to study the syn- aptic interactions of primary afferent fibers with pyramidal cell properties, orientation of the ELL slice in the longitudinal plane would preserve the majority of afferents in the deep fiber layer.
Synaptic activity generated from the large numbers of interneurons was occasionally evident by the presence of spontaneous, hyperpolarizing IPSPs, recorded from pyramidal cells using acetate electrodes (Fig. 5B). IPSPs were rarely encoun- tered when using chloride electrodes, presumably due to reversal of their direction by intracellular chloride injection (Eccles 1964).
Passive membrane characteristics
Intracellular electrode recordings were obtained from more than 140 pyramidal neurons; passive membrane parameters such as input resistance for the membrane (R~n), membrane time constant (rm), and resting membrane potential (Vm) were mea- sured. In addition, spike amplitude was routinely measured during brief depolarizing current pulses as a measure of the stability of the recording and overall health of the tissue. The membrane time constant was measured by applying a hyperpolar- izing current step to the cell and plotting the log of the slope of the voltage response as a function of time. The assumptions underlying this method
have been thoroughly reviewed and discussed (see Jack et al. 1975). Two assumptions are needed: (1) the hyperpolarizing current step does not activate large nonlinear conductances; as the insert to Fig. 6 shows, this assumption appears to be valid, (2) the cut dendrites of the cell seal themselves and are not a short-circuit; it is not possible to prove this assumption valid, but the apparent health and long-term viability of the impaled neurons suggest that sealing has occurred. The voltage response to the current step consists of a very brief period (1 ms) of rapid voltage change followed by a sec- ond phase of slower voltage change. The early ra- pid shift is presumably due to charging of the elec- trode capacitance, charging the interior of the neu- ron and its processes and charging the membrane of the soma. Since we cannot precisely measure the large contribution made by electrode capaci- tance, we used only the later part of the voltage response to estimate the time constant (Fig. 6). As discussed by Jack et al. (1975, pp. 168-171) this can be approximated by a single exponential since the interior of the cell and its dendrites is approxi- mately isopotential even for extensive dendritic trees. The data in Table 1 summarizes the calcu- lated membrane parameters.
Spontaneous activity
Extracellular, single unit recordings tu the pyra- midal cell layer of healthy slices revealed spontane- ous activity which followed 2 characteristic pat-

496 W.B. Mathieson and L. Maler: ELL pyramidal cell electrophysiology in vitro
Table 1. Electrophysiological properties of ELL pyramidal cells in vitro
Input Resting Spike Time resistance potential amplitude constant, Mf~ mV mV Tm
m s
# Sampled 132 96 141 8
AVE 27.1 73.9 68.2 6.75 _+SE _+1.16 _+0.96 +0.79 _+1.44
b
C 1
0 n A " ' ~ ' ' " ~ ' 0 n A 0 . 3 n A t
Fig. 7a-e. Spontaneous slow and fast pre-potentials, a lntracel- lular recording of strongly rhythmic cell with large, slow prepo- tentials (SPs). The oscillation in Vm typically consisted of a slow rising phase followed by a steeper repolarization. Note that these oscillations can occur entirely below spike threshold. Fast prepotentials (FPPs, arrows) frequently were superim- posed on the depolarized portion of the SPs. b FPPs recorded spontaneously in superimposed oscilloscope sweeps from the cell in a at three different points along the SP trajectory. As Vm depolarized, the FPPs increased in frequency and amplitude. Manually clamping the Vm to different values with DC injec- tions similarly affected FPP occurrence, c A strongly rhythmic cell became nearly quiescent when hyperpolarized by intracellu- lar current injection. Calibration markers in: a I s, 5 mV; b 5 ms, 2 mV; e 0.5 s, 3 mV
terns: (1) low frequency tonic firing of action po- tentials; (2) short bursts of action potentials of 2-20 spikes per burst, and interburst intervals of 1 to 2 s. The following sections employed intraso- matic electrode recording techniques to show that rhythmic activity results from intrinsic membrane properties which are characteristic of healthy pyra- midal cells in vitro.
Oscillations in V,,
The values reported in Table I indicated that for cells used in this study, the resting membrane po- tential averaged about 74 mV, with a range of 60-90 inV. However, many units displayed an un- stable Vm which oscillated above and below the resting potential in a stereotyped fashion. The tra- jectories of several examples of these voltage excur- sions are illustrated in Fig. 7. More robust oscilla- tions consisted of a slow depolarizing ramp, fol-
lowed by a steep repolarization. Rhythmicity did not depend on the electrolyte used in the recording electrode; identical patterns of slow potentials (SP) were observed while recording with potassium chloride, acetate or methyl sulfate electrodes. In a stable recording, the frequency and amplitude of SPs was remarkably constant, although it varied from cell to cell within a range of 0.5-1 Hz and 2-15 inV. The oscillations can remain entirely be- low spike threshold. In more depolarized neurons, however, the SPs traversed the subthreshold range for spike firing, and evoked a rhythmic burst of action potentials which resembled the extracellu- larly recorded activity. From a total of 131 cells, pyramidal cells were placed into 3 classes on the basis of their spontaneous behaviour; (a) approxi- mately 1/3 of those sampled were non-rhythmic at any potential, (b) 1/3 were 'weakly' rhythmic, e.g. they displayed oscillations in Vm during sub- threshold, depolarizing current injection but not at rest, (c) 1/3 were considered 'strongly' rhythmic since Vm oscillated even at the resting membrane potential. It is obviously possible that the 'strongly' rhythmic class are merely cells more de- polarized due to injury.
SPs were strongly voltage-dependent; hyperpo- larizing current injection reduced their amplitude and frequency, subthreshold depolarization in- creased both amplitude and frequency. In many cells, step-changes in Vm by as little as 5 mV above and below resting Vm were sufficient to switch the cells' firing behaviour from tonic to bursting (Fig. 7c).
While recording from rhythmic cells, measure- ments of input resistance were made over the course of several consecutive membrane oscilla- tions. Surprisingly, even during the more robust SPs, no detectable changes in Rin were measured at different points in the rhythm. This indicates that either the number of channels involved in gen- erating the different phases of the SPs is quite small, or that activation-inactivation of inward currents are offset by inactivation-activation of outward currents to produce no net change in Rin. Another possibility is that the channel openings generating SPs occur far enough out on the den- drites that fluctuations in Rin are too small to mea- sure at the soma.
Fast prepo ten tials
Superimposed upon the SP were small, depolariz- ing spikelets which superficially resembled so- called fast prepotentials (FPPs) described in hippo- campal neurons (Spencer and Kandel 1961). Their

W.B. Mathieson and L. Maler: ELL pyramidal cell electrophysiology in vitro 497
amplitude varied from 2-5 mV in a voltage-depen- dent manner; as the Vm oscillated across a slow potential, FPPs increased in frequency and ampli- tude reaching a maximum at more depolarized vol- tages (Fig. 7 a, b). FPPs can be completely elimi- nated by a 5 mV hyperpolarization, and reach maximal frequencies and amplitudes with less than 10 mV depolarization. The degree of voltage-sensi- tivity with respect to Vm suggests that FPPs origi- nate electrotonically near the cell soma.
Correlation of ceil type with rhythmic potentials
An interesting possibility emerged that slow oscil- lations in membrane potential might exist in either basilar or non-basilar pyramidal cells, but not both. In order to test this, cells were characterized according to their rhythmic membrane properties, then injected with Lucifer Yellow. In a sample of ten cells, no correlation was found between cell type and auto-rhythmicity; nearly equal numbers of NBP and BP neurons exhibited membrane oscil- lations. Conversely, non-rhythmic cells were equally likely to be BP or NBP cells.
Repetitive firing properties
F/I data
Input-output relationships for pyramidal neurons were investigated by delivering suprathreshold, de- polarizing pulses and calculating the firing fre- quencies from the 1st, 2nd and 3rd interspike inter- vals of the resultant spike trains. The calculated spike frequencies were plotted against total current injected to give a series of F/I curves for each neu- ron ( N = 30). A graph illustrating the F/I curves from an exemplary neuron is shown in Fig. 8. Most cells responded to increasingly intense stimuli by raising their spike frequencies in a roughly-sigmoi- dal fashion. The slope of the curve was shallow just above rheobase, and steepest at intermediate stimulus intensities. The spike frequency usually plateaued at higher current levels. In addition, the spike frequencies calculated from the 3rd inter- spike intervals were consistently higher than those calculated from the 1 st interval.
Spike accommodation
Longer duration depolarizing stimuli (0.4-1 s) evoked 2 characteristic patterns of spike potentials. First, approximately 2/3 of pyramidal cells (35/50) responded with a slowly developing train of spikes which increased in frequency to the point of tonic
200 .~ . . . . . . . . . . . .
--160' ~ ~ ~ , ~ +
ZI'~ 120 3 r d
�9 = ,P 8 0
~" 2rid
40 1 st
.;r ] 1 l I I 0.4 0.5 0.6 0.7 0.8 TOTAL IHJECTEO CURRENT (nA)
Fig. 8. F/I curves from a neuron calculated from the 1st, 2nd, and 3rd interspike intervals, The sigmoidal shape of spike fre- quency increases is obvious in all 3 curves, The 1st interval, representing ' instantaneous' frequency values was consistently lower than both 2nd (intermediate) or 3rd (quasi-steady-state) interspike interval values
firing, or rapid bursts of action potentials (Fig. 9 a - c). These neurons showed no indication of spike accommodation. In contrast to the non-accommo- dating neurons, roughly 1/3 of the cells tested (15/50) responded in a more phasic manner (Fig. 9d), by exhibiting partial to near total spike accommodation. It should be noted that the occur- rence of these two response types was complicated by one or more voltage-dependent processes. An accommodative cell could often be made to fire tonically by increasing the stimulus intensity or by current clamping Vm to a more depolarized level for several seconds and then applying an addition- al depolarizing pulse; the ionic basis for this com- plex behaviour was not investigated.
Stimulus-response lag
One of the more conspicuous features of pyramidal cell firing in response to direct stimulation was a lag period at the start of a depolarizing pulse which lasted up to 250 ms. As illustrated by the spike trains in Fig. 9, a pronounced lag preceded both tonic or bursting-type repetitive firing. In some cells, (e.g., Fig. 9c) the lag period so dominated the initial 200 ms that the spike frequency was held below 25 Hz, then abruptly increased to nearly 100 Hz.
The response characteristics of in vitro pyrami- dal cells was strikingly different from their in vivo counterparts. Bastian's laboratory has recorded from pyramidal cells (E and I cells) in situ while the fish received behaviourally relevant, excitatory stimuli. As shown by the response of an E cell in Fig. 9e (compliments of Dr. Bastian, personal communication), pyramidal cells in vivo respond to excitatory input with a fast, high frequency

498 W.B. Mathieson and L. Maler: ELL pyramidal cell electrophysiology in vitro
b
a I J
I L_ 2 n A
, 1 2 o m v 5 0 m s
e ,I~
I I ,~%~&'N'~g~t~WWW##,~
I T ,I,I,,,, ll]tT!~r:'
4 m y
2 0 r n s 5 m s
Fig. 9a--e. Repetitive firing characteristics of pyramidal cells in vitro and in vivo. a-e Pyramidal cells recorded in vitro stimulated directly by intracellular current injection. The range of stimulus responses are shown from 3 different cells. Each responded to the stimulation with a slowly developing train of spike potentials (250 Hz maximum) which displayed no evidence of spike frequency accommodation. The 3 responses also illustrate the range in magnitude of the ' lag' in spike frequency, d Accommodating cells' response. Approximately 1/3 of all cells tested responded with varying degrees of spike frequency accommodation, e Cells recorded in vivo (compliments of J. Bastian). While a fish was presented with a simulated EOD modulation (lower trace), pyramidal cell response was measured extracellularly (upper trace). Left panel: at the onset of increased electrical stimulation, the cell responded with an abrupt burst of spikes (600 Hz maximum, arrow) which rapidly adapted. Right panel: expanded view of initial burst response. Marker = 20 ms (left panel); 5 ms (right panel). Note different time scales in a-d versus e
burst of action potentials, followed by rapid spike adaptation. Spike frequencies of 450-600 Hz are not uncommon during the initial 50 ms of the burst (compared to 25-100 Hz in vitro). Moreover, the response is extremely abrupt, following a brief la- tency (10-15ms) for conduction of the afferent volley and a single synaptic delay (Bastian 1981 a). The functional significance of these discrepancies will be addressed in the Discussion.
Ionic conductances
Fast sodium conductance (gNa)
From the many examples of spontaneous and stim- ulus-evoked spike-potentials presented in these re- sults, it is evident that ELL pyramidal neurons display a fast sodium current typical of most inver- tebrate and vertebrate neurons. The rapid rise of the depolarizing phase and short duration of the spike (1 ms at 1/2 maximum amplitude), indicate rapid activation and inactivation kinetics of sodi- um channels, and the threshold for spike genera- tion was in the range of 10-15 mV depolarized from rest. In addition, 0.2 gM TTX was sufficient to block gNa, and thereby eliminate fast somatic spikes.
Subthreshold sodium conductance
In addition to classic fast-Na spikes, several lines of evidence suggested that, in the voltage range subthreshold for spike potentials, the excitability of pyramidal cells is to a large extent determined by the presence of a persistent Na conductance (gNaP).
TTX sensitivity
At resting membrane potentials, fast Na channels are for the most part closed. However, in cells which display slow oscillations, another, persistent gNa provided the driving force for the SPs. Addi- tion of 0.2 gM TTX to the bath ACSF completely and reversibly eliminated pyramidal cell rhythmic behaviour (N--10). The same effect was obtained by removal of Na § from the ACSF by substitution with TRIS (N= 3). The strip chart recording in Fig. 10 illustrates at a slow speed the time course of TTX-blockade of membrane oscillations. Dur- ing TTX-block, the interval between SPs increased, and the amplitude of the depolarizing phase of the voltage shift diminished until the Vm was essen- tially flat. Although the hyperpolarizing phase ap- peared unaffected by TTX, an increase in SP inter-

W.B. Mathieson and L. Maler: ELL pyramidal cell electrophysiology in vitro 499
l . l , , ' l ' l ~ ! i . . . . , ' ; . . , ' ! ' . . . .
1 8 ~ 2 0 a 2 3 ~
m v
2 6 s e c
Fig. 10. TTX block of slow potentials, The strip chart record shows at slow speed a strongly rhythmic cell with regular, 0.4 Hz SPs. TTX (0.2 IxM) added to the bath abolished the voltage oscillations, demonstrating a Na-dependent component of the rhythm. During the early stages of the TTX block (e.g. at T= 20'), the slope of the depolarizing ramp of the SP gradu- ally diminished until the SP was eliminated. This slice was main- tained in the interface condition, which was reflected by the long delay before the drug had an effect. Also note throughout the records the upwardly directed pen-strokes which correspond to intermittent, depolarizing current injections that monitored TTX block of spike potentials; fast and persistent Na channels were equally sensitive to TTX
val resulted from a reduction in the slope of the depolarizing ramp. Not shown in Fig. 10 is the observation that fast-prepotentials were also Na- dependent. Both TTX application and Na substi- tution abolished the brief depolarizing spikelets which frequently were superimposed upon the crest of SPs.
Activation of a subthreshold inward current was particularly obvious during depolarizing cur- rent pulses. Following an initial lag, a conspicuous depolarizing ramp developed in the voltage range preceding spike firing (Fig. 11). The slope of the ramp varied from cell to cell, so that cells which displayed a strong oscillation in Vm also displayed a large depolarizing ramp during injection of cur- rent pulses. In 4 separate experiments, addition of 0.2 gM TTX abolished the ramp providing addi- tional evidence for the activation of a subthreshold sodium current (gNaP).
Calcium conductance (gCa)
gCa blockade. Cells which displayed regular oscil- lations in Vm (N= 3), were incubated in low calci- um ACSF containing 2 m M Mn 2 § to block calci- um currents (Bagust and Kerkut 1980). As shown in Fig. 12, calcium blockade altered the trajectories of the SPs by progressively increasing their fre- quency and at the same time diminishing their am- plitude. With time the rhythm had degraded into very irregular and rapid fluctuations in Vm, 2-3 mV in amplitude, which, since calcium influx was blocked, probably was a reflection of the per- sistent Na-current and IA (see below). Washout
a b
5m= lOre=
Fig. 11 a, b. TTX sensitivity of gNa(P). When pyramidal cells were given brief, depolarizing current pulses, they responded with a subthreshold depolarizing ramp in Vm (trace a, arrow). As the ramp traversed the voltage threshold for spike initiation, a single spike or burst of spikes resulted. The depolarizing ramp was abolished by TTX-treatment (b), providing additional evi- dence for the presence of a persistent, subthreshold sodium conductance
1 2 0 ' ACSF w a s h
Fig. 12. Effect of manganese block of calcium influx on sponta- neous slow potentials. A strip chart recording of a strongly rhythmic cell before and after addition of 2 mM Mn + +/0 mM Ca + + to the bath solution. During the block of Ca-channels, the interburst interval gradually decreased (e.g. at 12' and 23) until the rhythm had degraded into very rapid, small amplitude oscillations in membrane potential (e.g. 120'). Washout with normal ACSF completely restored the pre-manganese rhythm
with normal ACSF completely recovered the pre- test rhythm. The shortening of the interval between successive oscillations caused by Mn +§ block strongly suggested that a Ca-dependent outward current, possibly gK(Ca), adds to the hyperpolariz- ing phase of the SPs, thereby regulating the fre- quency of rhythmic firing. The decrease in ampli- tude of the voltage shifts also suggested that calci- um influx either contributes directly to the depolar- izing phase of the SP, or indirectly by activating a calcium-dependent inward (Na) current. In either case, since Mn2+-block also interrupts synaptic transmission (Bagust and Kerkut 1980), the residu- al, irregular potentials following calcium channel blockade must be intrinsic to the pyramidal cell.
Calcium spikes. The presence of an active calcium current was demonstrated directly by evoking cal- cium spike-potentials during depolarizing current injections. The magnitude of this current appeared extremely small, however, and required TTX blockade of sodium channels (gNa and gNaP), as well as TEA and 4-AP block of potassium channels

500 W.B. Mathieson and L. Maler: ELL pyramidal cell electrophysiology in vitro
i,o m v
l O m s Fig. 13. Ca + +-spikes. Slices were perfused with 0.2 g M TTX to block Na-channels, 2 m M 4-AP to block IA, and 10-15 m M TEA to block remaining K-channels. Low threshold Ca-spikes (several spikes superimposed) displayed a characteristic broad shape. Stimulus current = 0.7 nA
(gK and gA) in order to sufficiently unmask gCa to evoke clean calcium spikes (Fig. 13). Under these conditions, low threshold (e.g., 0.5nA) spikes with amplitudes within the range of 5-20 mV were recorded in 10 separate experiments.
Calcium-dependent potassium conductance, gK(Ca)
The fact that a majority of ELLL pyramidal cells in vitro fire repetitively with little or no accommo- dation, in addition to the observed difficulty in evoking calcium spike potentials, leads to the spec- ulation that gK(Ca) is not an important compo- nent of their membrane conductance. To assess the contribution of gK(Ca) to spike firing, slices were incubated with ACSF containing the bee venom toxin, apamin, reported to specifically block gK(Ca) in several vertebrate preparations (Hugues et al. 1982). Alternatively, pyramidal cells were injected intracellularly with potassium EGTA, a specific calcium chelator, to block the activity of calcium-dependent potassium currents. Both treatments had little or no effect on the firing
patterns of non-accommodating cells (data not shown), but dramatically altered the repetitive fir- ing of more strongly accommodating cells, making them fire tonically (Fig. 14). These results suggest that the majority of pyramidal cells have little gK(Ca), whereas the repetitive firing of accommo- dating cells is strongly influenced by this outward conductance.
IA
Several lines of evidence suggested that ELL pyra- midal cells have a large A-current which regulates the input-output relationships as recorded intracel- lularly. First, repetitive firing in response to depo- larizing pulses was typically preceded by a variable delay and subsequent low spike-frequency period; the duration of this lag roughly corresponds to the inactivation times for Ia (cf. Fig. 9). Since IA inactivates with time at positive membrane poten- tials (Connor and Stevens 1971 ; Neher 1971), cur- rent-clamping Vm at different values should affect the lag in spike reactivity, if indeed it is mediated by IA. Figure 15A illustrates the voltage-depen- dence of the lag period. At each membrane poten- tial tested, the magnitude of the depolarizing pulse was adjusted so that the same final stimulus volt- age was reached. At depolarized V~,'s, the response lag was reduced, whereas hyperpolarization in- creased lag time indicating de-inactivation of an outward conductance.
The presence of an/A-like current during repet- itive firing was also demonstrated pharmacologi- cally, with the addition of 1 m M 4-AP to the bath solution. At this dose, 4-AP has been shown to selectively block A-channels (Thompson 1977;
A
my I lOOms
Fig. 14A-C. Effect of apamin or EGTA on spike frequency accommodation. A An accommodating cell was perfused with 400 n M apamin. 10 min after bath application of the drug, accommodation was eliminated. Washout with ACSF partially restored the accommodating response. B, C Spike frequency accommodation is illustrated from 2 experiments before (0') and after (10') intracellular injection of K + EGTA. Vm was manually clamped to a constant value and stimulus amplitudes were kept constant. While EGTA injection eliminated accommodation within 10 min, it had no effect on the ' l ag ' in spike frequency seen in the cell in B

W.B. Mathieson and L. Maler: ELL pyramidal cell electrophysiology in vitro 501
A
i!7 ~ r If Ill [IFl!tl!!
a
B take into account the cellular properties of pyrami- dal cells themselves. The results presented in this study are the first attempt to describe pyramidal cell physiology at the intracellular level, and to identify the cellular properties which have func- tional consequences for the output of the ELL.
:!:: ' : ' , ~i ~L i I i"
[ t i
20 --- m v
5 0 m s Fig. 15 A, B. Effect of/A-like current on stimulus-response ' lag ' . A The cell was stimulated with depolarizing current pulses (400 ms duration); this was done while the cell was (1) depolar- ized with a long constant current pulse, so as to inactivate IA, (2) at resting Vm, and (3) while the cell was hyperpolarized with long constant current pulse so as to remove IA inactivation. In the lower trace, Vm was hyperpolarized by a steady-state current of - 0 . 3 hA; a prominent lag in spike firing was evident. At resting Vm (middle trace) the lag was substantially reduced and at depolarized Vm (top trace, + 0.4 nA D.C.) the stimulus evoked a much smaller lag. In each trace stimulus intensity of the depolarizing pulse was adjusted to give the same total current injected. B During - 0 . 3 nA D.C. hyperpolarization, the stimulus evoked a spike train with a moderate lag. Middle trace: At the same Vm and stimulus pulse, 58 rain after addition of 1 m M 4-AP; the lag was greatly reduced and spike frequency increased. Top trace: 80 rain after 4-AP addition; lag was elimi- nated
Zbicz and Weight 1985). As indicated in Fig. 15 B, 4-AP abolished the response lag to direct stimula- tion, and allowed the cell to fire an immediate, high-frequency burst of spikes.
Discussion
To date, the electrophysiology of the ELL has been explored primarily with extracellular electrode re- cordings of pyramidal neurons in response to var- ious behavioural stimuli (Enger and Szabo 1965; Scheich 1977; Bastian 1981b, 1986a, b; Partridge et al. 1981 ; Matsubara 1981). Although pyramidal cell activity under these conditions correlates well with predictions made from neuroanatomical dia- grams (Bastian 1986 a), none of the interpretations
Pyramidal cell electrophysiology in vitro
The laminar organization of the ELL, which segre- gates the cellular and fiber tracts into discrete layers facilitated electrode recording of a pure pop- ulation of basilar and non-basilar pyramidal cells. Intracellular labelling with Lucifer Yellow con- firmed this important assumption; most or all of the physiological data reported here are represen- tative of electrical activity of BP and NBP neurons.
The results of these data demonstrate that pyr- amidal cells in vitro are capable of exhibiting volt- age-dependent, rhythmic oscillations in membrane potential; the presence or absence of these slow potentials appear to correlate with spontaneous bursting or tonic spike activity recorded extracellu- larly. When stimulated directly with depolarizing current injection, they respond with an initial ' lag' in spike firing lasting up to 250 ms in which the maximum rate of firing is restricted to 100 Hz or less. Following the lag period, the majority of cells enter a period of higher frequency spike firing which persists throughout the duration of the stim- ulus pulse; they display little or no spike frequency accommodation and as such are tonic neurons (Fig. 9).
Ionic conductances
Characterization of the membrane conductance of pyramidal cells indicates that spontaneous rhythmic activity and the repetitive firing proper- ties are mediated by the actions of at least 6 differ- ent ionic currents. Two currents, in particular, dominate the physiological behaviour of these cells: a persistent, subthreshold sodium current (/Nap) and a transient outward current similar to the A-current (IA).
Several lines of evidence support the conclusion that gNaP contributes substantially to pyramidal cell activity. First, the spontaneous, slow potentials are driven by voltage-dependent processes which activate in the voltage range subthreshold to spike initiation. Altering Vm by as little as 5 mV can transform a non-rhythmic cell into one that dis- plays robust SPs. Furthermore, SPs are sodium dependent; blockade of Na influx eliminates rhythmic activity. The progressive flattening of the

502 W.B. Mathieson and L. Maler: ELL pyramidal cell electrophysiology in vitro
SP rising phase which accompanies TTX block suggests that gNaP drives the depolarizing compo- nent of membrane oscillations.
Additional evidence for gNaP activation is seen in the response to positive current injection. A subthreshold current pulse elicits a slowly growing depolarizing ramp in the voltage trace, often trig- gering the appearance of SPs. This inward ramp is sensitive to TTX block. It is reasonable to as- sume that the persistent, depolarizing action of gNaP is largely responsible for the lack of spike frequency accommodation when challenged with long duration stimuli. Recent studies in various mammalian neurons (e.g. cortical pyramidal cells) have indicated that gNaP may typically be an im- portant conductance which helps to regulate a neu- ron's behaviour at threshold (Stafstrom et al. 1984).
Ia is an important current in many vertebrate and invertebrate neurons and appears to be a ma- jor determinant of interspike interval (Rogawski 1985). Due to the time and voltage properties of IA, the action of this current is especially obvious during the early response to stimulation. The kinet- ics of Ia are such that activation occurs rapidly (within ms) and inactivation proceeds with a time constant of 200-300 ms, depending on the mem- brane potential (Neher 1971). This outward cur- rent counteracts the depolarizing effect of positive current injection, resulting in the prominent lag period in spike train formation clearly visible in the stimulus-response profiles (cf. Fig. 9). The im- portance of IA to electroresponsiveness is most ob- vious when the current is blocked with 4-AP or inactivated by steady-state depolarization; under these conditions pyramidal cells become highly ex- citable and react rapidly to depolarizing stimuli by firing a high frequency burst of spikes (Fig. 15).
Ia may also influence spontaneous oscillations in membrane potential. Current clamping Vm to depolarized levels results in SPs of increased fre- quency and amplitude. Depolarizing the cell not only activates gNaP, but in addition inactivates (with time) Ia, which would further unmask the voltage excursion driven by gNaP. Thus, some of the variability in the strength of pyramidal cells' rhythmic behaviour may be due to differences in the strength of IA.
In addition to the classic sodium and delayed rectifier currents, calcium and calcium-dependent potassium currents influence pyramidal cell activi- ty, although the contribution of these last two cur- rents is far less demonstrable than that of either Ia or gNaP. By virtue of the inability to demon- strate calcium spikes in at least 1/2 of the cells tested, /Ca is probably not a major contributor to
the total membrane conductance. Even in the re- maining fraction of cells that generated Ca-spikes, a massive blockade of Na and K channels was required in order to sufficiently unmask the pres- ence of/Ca- In light of the relatively weak calcium influx, it is possible that calcium entry via voltage activated channels does not play a major direct role in pyramidal cell excitability.
Calcium currents, however small, indirectly regulate rhythmic activity and repetitive firing by activating gK(Ca). This is evident from the effects of eliminating calcium from the bath or blocking Ca channels with manganese. In spontaneously rhythmic cells these treatments substantially re- duced the interval between SPs, indicating that a calcium-dependent outward current (gK(Ca)) reg- ulates the frequency of membrane oscillations. Spike frequency accommodation, as well, depends on gK(Ca). In those cells which displayed accom- modation, blockade of gK(Ca) with apamin or chelating calcium with EGTA injection eliminated the accommodation whereas non-accommodating cells were unaffected by these drugs. The observa- tion that the majority of cells were non-accommo- dating provides further support to the notion that calcium and calcium-dependent potassium chan- nels play minor roles in pyramidal cell physiology in vitro. The fraction of cells which displayed ac- commodation appear to belong to a different func- tional class; it will be important for future studies to determine whether this functional distinction corresponds to a morphologically unique popula- tion of pyramidal cells.
Fast pre-potentials (FPPs)
An intriguing aspect of pyramidal cell electrophysi- ology is the presence of spontaneous spikelets, re- ferred to as fast pre-potentials. Their appearance does not seem to correspond to injury potentials, since many neurons deemed healthy by physiologi- cal criteria displayed stable patterns of FPPs throughout long duration recording sessions. In general, they were sensitive to voltage manipula- tion and TTX treatment, similar to that of the larger amplitude slow potentials. The FPPs in- creased in frequency and amplitude when the cell was depolarized, even though this moved the cell closer to the sodium reversal potential. However, the small voltage shifts (< 5 mV) would have a negligible effect on the large driving force pro- duced by the sodium gradient. Increasing depolar- izing current presumably activates greater numbers of the sodium channel patches which produce the

W.B. Mathieson and L. Maler : ELL pyramidal cell electrophysiology in vitro 503
FPPs, thus increasing both their frequency and am- plitude.
The duration and amplitude of FPPs observed in ELL pyramidal cells resemble similar potentials recorded in hippocampal pyramidal neurons in vivo (Spencer and Kandel 1961) and in vitro (Swartzkroin 1977). The precise nature of hippo- campal FPPs has yet to be defined, but has been proposed to originate from localized patches of dendritic membrane referred to as 'hot spots' (Spencer and Kandel 1961) or from electrotonic coupling potentials transmitted via gap junctions (MacVicar and Dudek 1980, 1981).
Although ultrastructural studies of the ELL have identified the existence of gap junctions be- tween NBPs and ascending processes from granule cell dendrites (Maler et al. 1981 b), the intracellular recordings presented here argue against FPPs be- ing electrotonic coupling potentials; FPP sensitivi- ty to current injection or oscillation in Vm suggests that they arise on or near the cell soma. Further- more, the granule cell dendritic processes are long and thin (e.g. 50-100 Ixm from the pyramidal cells, 2-5 gm diameter, Maler 1979). It seems likely that the granule cells are electrotonically too remote from the NBPs to be influenced by such small am- plitude fluctuations in Vm-
In light of the size measurements taken from LY-filled basilar pyramidal cells, it seems feasible that FPPs might function to amplify electrosensory input from the periphery, in a manner similar to that proposed for hippocampal neurons (Kandel and Spencer 1961). Since primary afferents synapse onto the distal bush of the basilar dendrite, EPSPs must be conveyed up to 400 gm before reaching the soma. Localized patches of membrane with ac- tive sodium conductance, situated part way along the basilar dendritic trunk could act as FPP gener- ating sites and provide a mechanism to enhance the EPSPs. Localizing sodium 'hot spots' on the dendritic trunk would also render them voltage- and TTX-sensitive, consistent with the recording data. This hypothesis predicts that only basilar pyramids should have FPPs. Further combined morphological and physiological studies will be re- quired to resolve this issue.
E L L pyramidal cells in vitro vs in vivo
Accommodation
Although pyramidal cell physiology has been in- vestigated in intact fish almost exclusively at the extracellular level, it is interesting to draw compar- isons between their response properties in vivo vs
in vitro. Bastian recorded E and I cell activities from immobilized fish while manipulating the in- tensity of their electroreceptive input (Bastian 1981b, 1986b). By examining the contrasting pat- terns of spike responses evoked in vitro vs in vivo, it is clear that removal of the ELL from its normal ascending and descending connections dramati- cally alters pyramidal cell excitability and elec- troresponsiveness.
Several possible explanations could account for the discrepancies in firing behaviour under the two conditions. As shown by Bastian's extracellular re- cordings (and illustrated in Fig. 9), pyramidal cells in situ (E cells) respond to excitatory electrosen- sory input with two distinct phases: an early, very brief high frequency burst of spikes (up to 600 Hz for a period of 20-50 ms), followed by a second phase of low frequency firing (spike adaptation). In vivo, therefore, pyramidal cells are both highly excitable and phasic in their response to peripheral input.
Both of these properties are, to a large extent, lost after slice preparation. Pyramidal cells in vitro have an early phase of low responsiveness (100-200 Hz), lasting for about 100-250 ms; this phase appears to result from a strong/A-like con- ductance. The second phase of response is usually non-accommodative, due to the combined influ- ences of gNaP activation and a weak gK(Ca).
At least some of these characteristics can be explained strictly in terms of neurocircuitry. In the intact brain, basilar pyramidal cells receive mono- synaptic excitation from P-type primary afferents as well as a disynaptic, inhibitory pathway via granule cell interneurons (Maler etal. 1981). Therefore, during an increase in afferent stimula- tion, BPs experience a rapid excitation (monosyn- aptic pathway) followed by a delayed inhibition (disynaptic pathway). In the slice preparation, however, pyramidal cells were stimulated by direct current injection, which bypassed the synaptically mediated excitation-inhibition sequence probably present in in vivo processing. Thus the adaptation of ELL basilar pyramids to sustained electrorecep- tive input is probably due to disynaptic inhibition rather than to accommodation based on outward K currents such as Ix(c,). In support of this hypoth- esis recent studies have shown that (a) a subset of ELL granule cell interneurons is GABAergic (Maler and Mugnaini 1986); and (b) bicuculine, a GABA antagonist, causes E cells (basilar pyra- mids) to become more tonic (Shumway and Maler, in press). Furthermore, Bastian (1986a) has re- ported that lesions of the descending input to pyra- midal cells reversibly converts their response from

504 W.B. Mathieson and L. Maler: ELL pyramidal cell electrophysiology in vitro
phasic to tonic. Again, this strongly suggests that the intrinsic membrane properties of pyramidal cells are such that they respond tonically to a sus- tained depolarizing input. This conclusion is made intuitively plausible by the temporal response char- acteristics of the E-cells (basilar pyramids). Bastian (1981b) has shown that E-cells respond optimally to amplitude modulations (AMs) of about 60 Hz. A long lasting outward current such as IK(ca~, would presumably act as a low pass filter and block the response to such high frequency modulations; IN,(p), however, would add to the primary afferent evoked depolarization, thus permitting the E cell to fire at higher rates. Thus, it may be that the presence of a strong, persistent INa(p) and a weak IK(c,) is a cellular specialization which optimizes E-cells for responding to rapidly modulated prima- ry afferent input. In nature these modulations might be produced by rapid movement of objects with respect to the electroreceptors.
A certain number of cells (approx. 1/3) did show spike frequency accommodation which was primarily due to IK~c,). It will be important to de- termine if these accommodating cells are a differ- ent morphological cell type, or whether pyramidal cells can, under certain circumstances, display a more powerful IK(c,) or a reduced IN,(p).
I-cells (non-basilar pyramids) also adapt in vivo to sustained electroreceptive input (Bastian 1981 b). The input to non-basilar pyramids is quite complex (Maler et al. 1981 a) and it is not evident how the circuitry could account for the adaptive properties of these cells. Bastian (1981 b) reported that I-cells respond best to AMs with lower fre- quency (about 16 Hz) and it will be interesting to determine whether this is a circuitry property or due to a greater IK~c,) in these cells.
Spike-frequency lag
It is not yet possible to understand why pyramidal cells in vivo can produce a rapid, high frequency response to primary afferent input while in vitro the same cell type will respond to intense depolar- ization with only low frequency firing (lag phase). One possible explanation is in the inactivation of the A-current. The normal input to pyramidal cells in vivo (from primary afferents and descending in- put) may keep these neurons in a state of tonic depolarization. Since IA inactivates when the cell is depolarized (Connor and Stevens 1971; Neher 1971; and see Fig. 15) this would mean that in vivo, 1A would not normally counteract the depo- larizing effect of primary afferent input; with all extrinsic input severed in vitro, pyramidal cells
would be hyperpolarized and IA inactivation would be removed. Subsequent depolarization would therefore be strongly counteracted by the activa- tion of IA.
This possibility is weakened, however, since Bastian (1986a, b) demonstrated that even after partial surgical removal of descending input, E- units could still respond even more vigorously to increased primary afferent input. This removal of descending input appeared to reduce tonic depolar- ization of pyramidal cells (Bastian 1986b) and IA inactivation would be expected to be removed, ren- dering the cell less, not more, responsive.
An alternative hypothesis is that synaptic in- put, present in vivo but lost in vitro, is capable of suppressing Ia. Of particular interest is the re- cent report that in cultured hippocampal neurons, IA was blocked by acetylcholine, even in conditions where an M-current was not detectable (Nakajima et al. 1986). This finding may be important for ELL functioning, since pyramidal neurons receive a descending, cholinergic input probably emanat- ing from eurydendroid cells of the caudal lobe (Maler et al. 1981 a; Phan and Maler 1983). This input could produce the intense early response of pyramidal cells in vivo by tonically suppressing A- channel activity. Although this hypothesis is ele- gantly simple, it lacks experimental support. The ELL slice preparation is, however, well suited to testing its validity since it is possible to stimulate descending fiber-systems in the slice, or to apply putative neurotransmitters or modulators. It will be especially important to test whether the de- scending cholinergic input can, in fact, modify IA.
gK(Ca) modulation. Similarly, gK(Ca) is a poten- tial target for synaptic modulation in pyramidal neurons. In hippocampal CA1 cells, for example, blockade of gK(Ca) with acetylcholine (Cole and Nicoll 1983) or noradrenaline (Madison and Nicoll 1982) is accompanied by a reduction in spike fre- quency accommodation during repetitive firing, similar to the apamin and EGTA blockade of ac- commodation recorded in ELL pyramidal cells in vitro.
The potential for neuromodulation of IA and/ or gK(Ca) may be an important feature of electro- sensory processing in nature. Modifying these channel properties may extend the range of re- sponses of ELL neurons as required during a vari- ety of behavioural situations and seasonal fluctua- tions in the life of the fish. For example, the fre- quencies of EOD amplitude modulations (AMs) detected by a fish vary over a wide spectrum, from the high frequency AMs encountered during elec-

W.B. Mathieson and L. Maler : ELL pyramidal cell electrophysiology in vitro 505
trolocation of rapidly moving objects, to low fre- quency AMs (e.g. beat frequencies) produced by mixing of the EODs of 2 or more conspecifics dur- ing social interactions. The temporal responsive- ness of pyramidal cells may need to be matched to the variable frequencies of these signals.
Furthermore, the sensitivity of electroreceptors varies with seasonal fluctuations in water resistivity (Knudsen 1974). It is probable that pyramidal cells must somehow adjust their own excitability to compensate for the changing electroreceptive in- put.
Two recent reports by Bastian (1986a, b) de- scribe one mechanism which could be used for this purpose in ELL processing of electrosensory sig- nals. Descending fibers originating in the nucleus praeeminentialis of the midbrain are a part of a feedback loop which acts as a gain control for pyr- amidal cell responsiveness. When Bastian inter- rupted these descending fiber tracts, pyramidal cell excitability and surround inhibition increased.
Descending input could also impose long-last- ing control on pyramidal cell excitability by modu- lating one or more of its ionic conductances. For example, modifying IA would significantly change the early response for pyramidal cells, while modu- lating gK(Ca) or gNaP would alter the late re- sponse.
General implications for in vitro model systems
In spite of the many advantages to the in vitro approach to neurophysiology, it is clear from the data presented in this study that the interpretation of such work must be made with great care. Many reports on the in vitro properties of central neurons are taken as an accurate representation of their in vivo counterparts, in spite of not having a solid basis for comparison. As with ELL pyramidal cells, interruption of the normal synaptic drive may be sufficient to radically alter such basic physiolog- ical parameters as the input-output relationships of a cell, or the functional state of the ion channels that make up the composite membrane conduc- tance. Technical advances in tissue maintenance will continue to extend the life of neural prepara- tions, and these concerns will become even greater as long-term changes in membrane properties ex- press themselves.
The data to emerge from this and future studies of ELL neurons are unique in these respects. Un- like similar data reported in more complex model systems, the cellular properties of ELL neurons examined in vitro can be interpreted within a grow- ing framework of neurocircuitry and sensory phys-
iology. In this way, the physiological consequences of tissue preparation and maintenance can be re- cognized, and even utilized, to formulate and test predictions about the mechanisms that govern elec- trosensory processing.
Acknowledgements. We wish to thank Dr. J. Bastian for provid- ing the photographs of pyramidal cell firing in vivo. and B. Ellis for assistance in preparing the figures. This was supported by MRC grant MT6027 to L.M.
References
Bagust J, Kerkut GA (1980) The use of the transition elements manganese, cobalt, and nickel as synaptic blocking agents on isolated, hemisected, mouse spinal cord. Brain Res 182:474477
Bastian J (1981a) Electrolocation. I. How the electroreceptors of Apteronotus albifrons code for moving objects and other electrical stimuli. J Comp Physiol 144:456M79
Bastian J (1981b) Electrolocation. II. The effects of moving objects and other electrical stimuli on the activities of two categories of posterior lateral line lobe cells in Apteronotus albifrons. J Comp Physiol 144:481M94
Bastian J (1986a) Gain control in the electrosensory system mediated by descending inputs to the electrosensory lateral line lobe. J Neurosci 6:533-562
Bastian J (1986b) Gain control in the electrosensory system: a role for the descending projections to the electrosensory lateral line lobe. J Comp Physiol A 158:505 516
Carr CE, Maler L, Sas E (1982) Peripheral organization and central projections of the electrosensory nerves in gymnoti- form fish. J Comp Neurol 211:139 153
Cole AE, Nicoll RA (1983) Acetylcholine mediates a stow syn- aptic potential in hippocampal pyramidal cells. Science 221 : 1299 1301
Connor JA, Stevens CF (1971) Voltage clamp studies of a tran- sient outward membrane current in gastropod neural soma. J Physiol 213:21-30
Connors BW, Kriegstein AR (1986) Cellular physiology of the turtle visual cortex : Distinctive properties of pyramidal and stellate neurons. J Neurosci 6:164~177
Eccles JC (1964) The physiology of synapses. Springer, Berlin Heidelberg New York, pp 173-188
Enger PS, Szabo T (1965) Activity of central neurons involved in electrolocation in some weakly electric fish. J Neurophy- siol 28 : 800-818
Gerhardt SC, Bernard P, Pastor G, Boast CA (1986) Effects of systemic administration of the NMDA antagonist, CPP, on ischemic brain damage in gerbils. Neurosci Abstr 12:59
Grace AA, LlinAs R (1985) Morphological artifacts induced in intracellularly stained neurons by dehydration: circum- vention using rapid dimethyl sulfoxide clearing. Neurosci 16:461-476
Heiligenberg W, Dye J (1982) Labelling of electroreceptive af- ferents in a gymnotid fish by intracellular injection of HRP: the mystery of multiple maps. J Comp Physiol 148:287-296
Hopkins CD (1976) Stimulus filtering and eleetroreception: tu- berous electroreceptors in three species of gymnotoid fish. J Comp Physiol 111:171-207
Hugues M, Romey G, Duval D, Vincent JP, Lazsunski M (1982) Apamin as a selective blocker of the calcium-depen- dent potassium channel in neuroblastoma cells: voltage- clamp and biochemical characterization of the toxin recep- tor. Proc Natl Acad Sci USA 79 : 1308-1312

506 W.B. Mathieson and k Maler: ELL pyramidal cell electrophysiology in vitro
Jack JJB, Noble D, Tsien RW (1975) Electric current flow in excitable cells. Clarendon Press, Oxford
Knudsen E (1974) Spatial aspects of the electric fields generated by weakly electric fish. J Comp Physiol 99:103-118
MacVicar BA, Dudek FE (1980) Dye-coupling between CA3 pyramidal cells in slices of rat hippocampus. Brain Res 196: 494497
MacVicar BA, Dudek FE (1981) Electronic coupling between pyramidal cells: a direct demonstration in rat hippocampal slices. Science 213 : 782-784
Madison DV, Nicoll RA (1982) Noradrenaline blocks accom- modation of pyramidal cell discharge in the hippocampus. Nature 299 : 636-638
Maler L (1979) The posterior lateral line lobe of certain gymno- toid fish: quantitative light microscopy. J Comp Neurol 183 : 323-363
Maler L, Mugnaini E (1986) Immunohistochemical identifica- tion of GABAergic synapses in the electroseusory lateral line lobe of a weakly electric fish (Apteronotus leptorhyn- chus). Soc Neurosci Abstr 84: 312
Maler L, Collins M, Mathieson WB (1981a) The distribution of acetylcholinesterase and choline acetyl transferase in the cerebellum and posterior lateral line lobe of weakly electric fish (Gymnotidae). Brain Res 226:320-325
Maler L, Sas E, Rogers J (1981 b) The cytology of the posterior lateral line lobe of high-frequency weakly electric fish (Gym- notidae): dendritic differentiation and synaptic specificity in a simple cortex. J Comp Neurol 195:87-139
Mathieson WB, Heiligenberg W, Maler L (1986) Ultrastructur- al studies of synaptic targets of physiologically identified electrosensory afferent synapses in the gymnotiform fish, Eigenmannia. J Comp Neurol 255:526-537
McLennan H (1983) Receptors for the excitatory amino acids in the central nervous system. Prog Neurobiol 20:251~71
Matsubara J (1981) Neural correlates of a nonjammable elec- troloeation system. Science 211:722-1725
Meldrum MJ, Chroucher G, Badman G, Collins JF (1983) Antiepileptic action of excitatory amino acid antagonists in the photosensitive baboon, Papio papio. Neurosci Lett 39:101-104
Milan F, Janigro D, DiGregorio D, Gorio A (1985) GM1 treat- ment in vivo protects membrane ATPase activities and pre- vents mitochondrial damage in hippocampal slices. Neu- rosci Abstr 11 : 376
Nadi S, Maler L (1987) The laminar distributions of amino acids in the caudal cerebellum and electrosensory lateral line lobe of weakly electric fish (Gymnotidae). Brain Res 425: 218-224
Neher E (1971) Two fast transient current components during voltage clamp on snail neurons. J Gen Physiol 58 : 36-53
Nakajima Y, Nakajima S, Leonard RS, Yamagucbi K (1986) Acetylcholine raises excitability by inhibiting the fast tran- sient potassium current in cultured hippocampal neurons. Proc Natl Acad Sci USA 83 : 3022-3026
Partridge BL, Heiligenberg W, Matsubara J (1981) The neural basis of a sensory filter in the jamming avoidance response: no grandmother cells in sight. J Comp Physiol 145:153- 163
Phan M, Maler L (1983) Distribution of muscarinic receptors in the caudal cerebellum and electrosensory lateral line lobe of gymnotiform fish. Neurosci Lett 42:137-143
Rogawski MA (1985) The A current: how ubiquitous a feature �9 of excitable cells is it? Trends Neurosci 8:214-219 Sas E, Maler L (1987) The organization of afferents to the
caudal lobe of the cerebellum of the gymnotid fish Apterono- tus leptorhynchus. Anat Embryol 177:55-79
Saunders J, Bastian J (1984) The physiology and morphology of two types of electrosensory neurons in the weakly electric fish Apteronotus leptorhynchus. J Comp Physiol A 154:199-209
Scheich H (1977) Neural basis of communication in the high frequency electric fish, Eigenmannia virescens (jamming avoidance response). J Comp Physiol 113 : 181-255
Scheich H, Bullock TH, Hamstra RH (1973) Coding properties of two classes of afferent nerve fibers : high frequency elec- troreceptors in the electric fish, Eigenmannia. J Neurophy- siol 36 : 3%60
Schubert P, Mitzdorf U (1979) Analysis and quantitative evalu- ation of the depressive effect of adenosine on evoked poten- tials in hippocampal slices. Brain Res 172:186-190
Schwartzkroin PA (1977) Further characteristics of hippocam- pal CA1 cells in vitro. Brain Res 128:53-68
Shumway CA, Maler L (in press) Bicucculine affects both tem- poral and receptive field properties of pyramidal cells in the electrosensory lateral line lobe. Soc Neurosci Abstr
Spencer WA, Kandel ER (1961) Electrophysiology of hippo- campal neurons. IV. Fast prepotentials. J Neurophysiol 24:272-285
Stafstrom CE, Schwindt PC, Flatman JA, Crill WE (1984) Properties of subthreshold response and action potential recorded in layer V neurons from cat sensorimotor cortex in vitro. J Neurophysiol 52:224263
Thompson SH (1977) Three pharmacologically distinct potassi- um channels in molluscan neurons. J Physiol 265:465-488
van Herreveld A, Stamm JS (1953) Effect of pentobarbitol and ether on the spreading cortical depression. Am J Physiol 173:164-170
Zbicz KL, Weight FF (1985) Transient voltage and calcium- dependent outward currents in hippocampal CA3 pyrami- dal neurons. J Neurophysiol 53:1038 1057





























