Embed Size (px)
Citation preview

Properties of Mouse Spinal Lamina I GABAergic Interneurons.
Kimberly J. Dougherty, Michael A. Sawchuk, and Shawn HochmanDepartment of Physiology, Emory University School of Medicine, Atlanta, Georgia
AbstractLamina I is a sensory relay region containing projection cells and local interneurons involved inthermal and nociceptive signaling. These neurons differ in morphology, sensory response modality,and firing characteristics. We examined intrinsic properties of mouse lamina I GABAergic neuronsexpressing enhanced green fluorescent protein (EGFP). GABAergic neuron identity was confirmedby a high correspondence between GABA immunolabeling and EGFP fluorescence. Morphologiesof these EGFP+/GABA+ cells were multipolar (65%), fusiform (31%), and pyramidal (4%). In wholecell recordings, cells fired a single spike (44%), tonically (35%), or an initial burst (21%) in responseto current steps, representing a subset of reported lamina I firing properties. Membrane properties oftonic and initial burst cells were indistinguishable and these neurons may represent one functionalpopulation because, in individual neurons, their firing patterns could interconvert. Single spike cellswere less excitable with lower membrane resistivity and higher rheobase. Most fusiform cells (64%)fired tonically while most multipolar cells (56%) fired single spikes. In summary, lamina I inhibitoryinterneurons are functionally divisible into at least two major groups both of which presumablyfunction to limit excitatory transmission.
INTRODUCTIONGABA is the major inhibitory transmitter in the CNS. In the spinal cord, GABAergicinterneurons are concentrated in the superficial laminae (I-III) where they reduce excitabilityby both pre- and postsynaptic inhibition. Axo-axonic GABAergic synapses onto primaryafferent terminals produce presynaptic inhibition (Alvarez et al. 1992; Rudomin and Schmidt1999; Schmidt et al. 1998) while postsynaptically, GABAergic neurons reduce the excitabilityof both projection neurons (Alvarez et al. 1992) and interneurons (Jankowska 1992).
Glutamic acid decarboxylase (GAD) is the rate-limiting enzyme in GABA synthesis. Thereare two isoforms of GAD expressed in the mature CNS—GAD65 and GAD67. A neuron isGABAergic if it contains either or both enzymes (Soghomonian and Martin 1998). Bothenzymes are produced in cell bodies in all regions of the spinal cord except lamina IX (Barberet al. 1982; Ma et al. 1994). Almost all GAD+ boutons in the spinal gray matter are labeledwith both 65 and 67 isoforms (Mackie et al. 2003). GABAergic neurons have been identifiedin transgenic mice expressing enhanced green fluorescent protein (EGFP) under the control ofa GAD67 regulatory element (Heinke et al. 2004; Oliva et al. 2000). In hippocampus andneocortex, while EGFP expression was observed only in a subpopulation of somatostatin-containing interneurons, 99% of EGFP+ neurons were GAD67+ and hence GABAergic (Olivaet al. 2000). Recently, GABAergic neurons have been targeted in lamina II in these mice andfound to comprise a larger fraction (35%; 54% in colchicine-treated) of the total identifiedGABAergic neurons and were a diverse population both neurochemically and physiologically(Heinke et al. 2004).
Address for reprint requests and other correspondence: S. Hochman, Whitehead Biomedical Research Bldg., Rm. 644, Emory UniversitySchool of Medicine, 615 Michael St., Atlanta GA 30322 (E-mail: E-mail: [email protected])..
NIH Public AccessAuthor ManuscriptJ Neurophysiol. Author manuscript; available in PMC 2009 May 8.
Published in final edited form as:J Neurophysiol. 2005 November ; 94(5): 3221–3227. doi:10.1152/jn.00184.2005.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Lamina I is the most superficial lamina of the sensory dorsal horn (laminae I-VI) and receivesinput mainly from nociceptive mechanoreceptors and thermal afferents (Christensen and Perl1970). Lamina I is made up of ascending tract neurons and local excitatory and inhibitoryinterneurons. Unlike lamina II, which contains mostly propriospinal and local interneurons(see Willis and Coggeshall 1991), lamina I, containing a relatively large number of ascendingtract neurons, is one of the main sensory channels that transmits thermal, nociceptive, and otherinteroceptive stimuli to higher structures (Craig et al. 2001; Gauriau and Bernard 2004; Spikeet al. 2003). Local inhibitory GABAergic interneurons make up ~25% of the neuronalpopulation (Polgar et al. 2003; Todd and McKenzie 1989) and serve to limit afferentinformation by both pre- and postsynaptic inhibitory mechanisms (Alvarez et al. 1992).
Cells in this region have been divided into several classes based on morphological andphysiological properties. Morphologically, cells were divided into three main classes—fusiform (with longitudinal and ventral dendritic arbors or exclusively longitudinal arbors),pyramidal, multipolar (with many or few dendritic branches), and flattened (Han et al. 1998;Lima and Coimbra 1986; Prescott and De Koninck 2002). Dendritic extension of all three celltypes is greatest in the rostrocaudal axis (Chery et al. 2000; Lima and Coimbra 1986). LaminaI neurons in rat can also be divided into categories based on firing properties. Five differentsubtypes were distinguished: tonic, phasic, initial burst, delayed onset, and single spike(Prescott and De Koninck 2002; Ruscheweyh and Sandkuhler 2002). Additionally, lamina Iprojection neurons were classified as either gap- or bursting-firing neurons (Ruscheweyh et al.2004). Prescott and DeKoninck (2002) correlated firing with cell morphology. For example,most fusiform cells fire tonically, whereas multipolar cells fire with either delayed onset or asingle spike after intracellular current injection. In cat, lamina I neurons have also beenclassified based on the types of sensory stimuli that they respond to (i.e., modality). Theseproperties also correlate with morphology—fusiform cells are nociceptive specific (noxious),pyramidal cells are thermoreceptive specific (warm and cool), and multipolar cells are eitherpolymodal nociceptive (noxious and thermal) or nociceptive specific (Han et al. 1998).However, this relationship may not hold true in rodent (Todd et al. 2002, 2005).
While lamina I ascending tract neurons have been studied extensively (i.e., Andrew and Craig2002; Craig and Andrew 2002; Ruscheweyh et al. 2004), currently no studies havecharacterized lamina I neurons based on transmitter phenotype (but see Torsney andMacDermott 2004). We used transgenic mice containing EGFP under the control of a GAD67regulatory element (Oliva et al. 2000) to identify spinal GABAergic neurons (cf. Heinke et al.2004). The aims were twofold. The first was to characterize the morphology of GAD67-EGFPneurons and their overlap with GABA immunolabeling in lamina I. The second goal was torecord from lamina I EGFP+ cells and characterize their properties in relation to priorobservations in similarly located unidentified cell types (Prescott and De Koninck 2002;Ruscheweyh and Sandkuhler 2002). We show that EGFP is a reliable marker of GABAergicphenotype in lamina I and that these GABAergic neurons are divisible into at least two groupsrepresenting a subset of the firing properties observed for neurons in this lamina.
Some of these data have been presented in abstract form (Dougherty et al. 2003, 2004).
METHODSAll experimental procedures complied with the National Institutes of Health guidelines foranimal care and the Emory Institutional Animal Care and Use Committee. HomozygoticGAD67-EGFP mice obtained from Jackson Laboratory (Bar Harbor, ME) were used in allexperiments. Lamina I was identified between the dorsal white matter and the relativelytranslucent substantia gelatinosa. No cell >20 μm from the edge of the white matter was
Dougherty et al. Page 2
J Neurophysiol. Author manuscript; available in PMC 2009 May 8.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

considered (Chery et al. 2000). Some lamina I neurons may have been excluded fromconsideration because lamina I is thicker in the central part of the cord.
ImmunohistochemistryMice at postnatal day (P) 14 were anesthetized with urethan (2 mg/kg ip), perfused with 1:3vol/body wt ice-cold 0.9% NaCl, 0.1% NaNO2, 1 unit/ml heparin, followed by equal volume/body weight of 4% paraformaldehyde or modified Lana's fixative (4% paraformaldehyde, 0.2%picric acid, 0.16 M PO3); pH 6.9. All spinal cords were isolated and postfixed 1 h, cryoprotectedin 10% sucrose, 0.1 M PO3, pH 7.4 until sectioned in 10 μm thick slices on a cryostat (Leitz1720).
Ten nonconsecutive sections (100 μm apart) from lumbar segments 1-3 of two mice were usedfor EGFP/GABA and EGFP/NeuN comparison. All incubations and washes forimmunohistological processing were performed in 0.1M PO3-buffered saline containing 0.3%triton X-100 (PBS-T). Tissue was washed overnight in PBS-T at 4°C followed by incubationin primary antibody for 48-72 h (either rabbit anti-GABA, 1:500 [Chemicon, Temecula, CA]or mouse anti-NeuN 1:500 [Sternberger Monoclonals, Lutherville, MD]). Slides were thenwashed three times for 30 min and incubated in Cy3 anti-rabbit (GABA) or anti-mouse (NeuN)conjugated secondary antibody at 1:250 (Jackson ImmunoResearch, West Grove, PA). LaminaI was defined as the first layer of cells in the dorsal spinal gray. Lamina I cells were countedin nonconsecutive transverse and parasagittal sections (100 μm apart) for the GABA/EGFPand in transverse sections for NeuN/EGFP using the Neurolucida image analysis system(MicroBrightField, Williston, VT). All cell counts can only be regarded as estimates sincestereological techniques were not used. Images of transverse sections (Fig. 1, A-C) wereoverlaid in Corel-Draw 12.0 (Ottawa, Ontario).
For cell morphology, cells in lamina I were counted in L1-6 parasagittal sections in two miceat P14. A total of 1,496 cells were classified by morphology. Criteria for discerning fusiform,pyramidal, and multipolar cell types were soma shape and number of primary processes(Prescott and De Koninck 2002). Because cells were examined in two dimensions, it is possiblethat primary process were missed. However, morphology is most evident in parasagittal andhorizontal sections because most primary processes extend in the rostral-caudal plane. Theflattened morphology can only be identified in the horizontal plane; however, flattened cellsare not GABAergic (Lima et al. 1993).
ElectrophysiologyMice (P4-19, mean = 12) were anesthetized with urethan (2 mg/kg ip) and decapitated, and thespinal cord was carefully dissected out of the body cavity and placed in a cooled (<4°C)artificial cerebrospinal fluid (ACSF) containing (in mM): 250 sucrose, 2.5 KCl, 2 CaCl2,1MgCl2, 25 glucose, 1.25 NaH2PO4, and 26 NaHCO3 at a pH of 7.4. The ACSF wascontinuously oxygenated with 95% O2-5% CO2. Parasagittal (150 μm) or transverse (250 μm)spinal slices were cut from lumbar cord. Slices were left to recover at room temperature for ≥1h prior to the onset of experimentation.
Patch electrodes were prepared from 1.5 mm OD capillary tubes (World Precision Instruments,Sarasota, FL) using a two-stage puller (Narishige PP83) to produce resistance values rangingfrom 5 to 8 MΩ. The intracellular recording solution contained (in mM): 140 K-gluconate(KF), 0.2 EGTA, and 10 HEPES, pH 7.3. In some electrodes, a support solution of 4 mM ATPand 1 mM GTP was also included.
The recording chamber was continuously perfused with oxygenated ACSF [that contained (inmM): 125 NaCl, 2.5 KCl, 2 CaCl2, 1 MgCl2, 25 glucose, 1.25 NaH2PO4, and 26 NaHCO3 at
Dougherty et al. Page 3
J Neurophysiol. Author manuscript; available in PMC 2009 May 8.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

a pH of 7.4] at a rate of ~2 ml/min. Whole cell patch-clamp recordings were undertaken atroom temperature using the Multiclamp amplifier (Axon Instruments; Union City, CA) filteredat 5 kHz (4-pole low-pass Bessel). EGFP+ lamina I interneurons were identified usingepifluorescent illumination. Position of the cell in lamina I was verified using differential-interference contrast optics (DIC) at ×40 to show that the cell was in or adjacent to the whitematter in that focal plane (Chen and Gu 2005). Then the electrode was lowered into the slice,and the cell was targeted for whole cell patch-clamp recordings using DIC. Both voltage- andcurrent-clamp data were acquired on computer with the pCLAMP acquisition softwareClampex (v 9.0; Axon Instruments). Immediately after rupture of the cell membrane (in voltageclamp at -80 mV), the current-clamp recording configuration was used to determine restingmembrane potential. Series resistance was subtracted. Most experiments were conducted inthe current-clamp mode, although in a few cases, voltage-clamp recordings were made. Cellswere brought to -80 mV by injecting bias current through the headstage. Then, a series of 1shyperpolarizing and depolarizing current steps were undertaken. Liquid junction potentialswere not corrected for. Firing type was determined by the response to current steps at and abovethreshold (Prescott and De Koninck 2002; Ruscheweyh and Sandkuhler 2002). Resistance wascalculated from the average of steady-state responses to 1s long hyperpolarizing voltage steps.Exponential curve fit of responses to hyperpolarizing current steps was used to calculatemembrane time constant (τm). Rheobase was the minimum current step magnitude required torecruit an action potential. Voltage threshold was determined at rheobase by measuring themembrane potential at the inflection point at which the action potential was initiated (e.g.,Fedirchuk and Dai 2004). The membrane potential value at threshold was largely insensitiveto the holding potential in the range of holding potential values tested (-60 to -90 mV; r = 0.12).Maximal firing frequency was calculated as the reciprocal of the interspike interval betweenthe first two spikes at the highest current step. Capacitance was calculated by dividing τm byresistance (τm = RC).
A total of 244 lamina I EGFP+ cells were recorded from. For analysis of firing properties, onlycells with resting membrane potentials more negative than -50 (n = 133) were included. Foranalysis of cellular properties based on morphology, cells with membrane potentials morenegative than -40 (n = 57) were considered because the additional cells had similar propertiesto those in the former group.
Statistical comparisons between firing properties were made using one-way ANOVAs (InStat,GraphPad Software, San Diego, CA). Membrane properties of the two morphological cell typeswere statistically compared using a Student's t-test and reported as means ± SE.
RESULTSGAD67-EGFP cells represent the majority of lamina I GABAergic neurons
The number of cells expressing endogenous EGFP was compared with the number of cellsimmunolabeled for GABA in the same section in lamina I of lumbar cord at P14 (Fig. 1). BothEGFP fluorescence and GABA immunolabeling were observed predominantly in the dorsalhorn. An overlay of EGFP and GABA labeling shows that most but not all EGFP+ cells werealso GABA+ in the dorsal horn (Fig. 1C). In lamina I, most of the EGFP+ cells are GABA+
(73%), whereas the percentage of GABA+ cells expressing EGFP is 68%. EGFP-expressingneurons made up 19% of the total population of lamina I neurons. Thus, although most laminaI cells are EGFP+ and GABA+, some cells appear to express only EGFP or GABA.
Lamina I EGFP+ cell morphology was determined by the shape of the soma and number ofprimary dendrites seen in parasagittal sections as described by Prescott and DeKoninck(2002). This was also examined at P14. The percentages reported here are averages from twomice. Multipolar cells have round somas and several primary dendrites. These constituted 65%
Dougherty et al. Page 4
J Neurophysiol. Author manuscript; available in PMC 2009 May 8.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

of the EGFP expressing neurons (Fig. 2A) and resembled type IIa cells described by Lima andCoimbra (1986). Fusiform cells have a flattened soma and two primary dendrites and accountedfor 31% of EGFP+ cells (Fig. 2B). EGFP+ pyramidal shaped cells, with three primary dendrites,were rare (4%). Dendrites could frequently be followed for long distances by endogenousEGFP fluorescence (Fig. 2C).
Membrane properties of EGFP+ neuronsWhole cell recordings were made from visually identified EGFP+ neurons (n = 244) in laminaI. Due to stringent inclusion criteria (see METHODS), 133 cells were further analyzed. In responseto depolarizing current steps from a holding potential of -80 mV, these neurons displayed oneof three different firing patterns (Fig. 3A). The highest incidence of cells responded with asingle spike to current steps, comprising 44% of sampled neurons. Cells that fired tonicallythroughout the current step represented 35% of the population, whereas 21% of cells fired withseveral spikes at current step onset (termed initial burst). Only one EGFP+ neuron respondedwith delayed firing during current step application and will not be considered further in thisstudy.
Passive and active membrane properties were measured for the populations of cells with restingmembrane potentials more negative than -50 mV and divided into three groups based on theaforementioned differences in firing properties (Table 1). Resting membrane potentials did notdiffer between the three groups. Tonic and initial burst cells were statistically indistinguishablefor all passive membrane and threshold properties measured. It is possible that these cellsrepresent one group with modifiable firing properties. First, some cells that fired tonically alsoinstead fired an initial burst with larger amplitude current steps (n = 19; Fig. 3B). Second, othercells classified as tonic based on initial recordings converted into initial burst cells later in therecording (n = 10; Fig. 3C1), and the reverse conversion was also seen (n = 4; Fig. 3C2). Incontrast, this never occurred in cells firing single spikes.
Cells firing single spikes differed significantly from the other two groups in several membraneproperties (Table 1). Rheobase was higher in single spike cells (135 pA) than tonic cells (65pA) and initial burst cells (71 pA), suggesting that single spike neurons are less excitable.Single spiking cells also had lower resistance and shorter τm values than that of both tonic andinitial burst (424 compared with 721 and 618 MΩ and 16 compared with 30 and 28 ms,respectively). Interestingly, there was a strong inverse correlation between τm and rheobase(Fig. 4A; r = -0.68) indicating that τm contributed significantly to the value of rheobase. Longerτm values for tonic and initial burst cells support a greater capacity for synaptic integration.Capacitance, maximum firing frequency, and spike height were not significantly differentbetween groups. Membrane time constant (τm) was well correlated with membrane resistance(Fig. 4A, r = 0.39) and cell capacitance values (r = 0.53), and there was an inverse correlationbetween cell capacitance and resistance (r = -0.42). Threshold voltage was similar betweencell populations suggesting that there are no differences in the voltage-dependent activation ofNa+ spikes in these cell populations.
Firing properties were compared with cell morphology in 25 identified fusiform cells and 32identified multipolar cells. Most fusiform cells fired tonically (64%) with a minority displayinginitial burst (24%) and single spike activity (12%). In contrast, the majority of multipolar cellsfired a single spike (56%) with a minority displaying tonic (31%) and initial burst firing (13%).Multipolar and fusiform cells also had differences in cellular membrane properties (Table 2).As compared with multipolar cells, fusiform cells had a longer τm (30 vs. 19 ms). Fusiformcells also had larger cell capacitance (50 vs. 34 pF) and lower rheobase (74 vs. 110 pA),suggesting that fusiform cells are larger and more excitable than multipolar cells. We thencompared the distribution of membrane properties in each morphological cell type separatedas single-spike, tonic and initial burst firing populations (Fig. 4B). Single-spike cells in both
Dougherty et al. Page 5
J Neurophysiol. Author manuscript; available in PMC 2009 May 8.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

morphological types had higher rheobase, lower resistances, and shorter τm. Thus thedifferences in membrane properties between morphological types are at least partly due to thehigher incidence of single spike neurons in multipolar cells and a higher incidence of tonic andinitial burst neurons in fusiform cells.
DISCUSSIONWe used transgenic mice expressing EGFP under the control of a GAD67 regulatory element(Oliva et al. 2000) to characterize morphological and physiological properties of lamina IGABAergic interneurons. The view taken was that an understanding of the response propertiesof local inhibitory interneurons would provide insight into mechanisms controlling excitabilityat this nodal point in sensory processing. The primary finding of the present work is that laminaI inhibitory interneurons are functionally divisible into at least 2 major groups.
Morphology of GAD67-EGFP neurons and overlap with GABA immunolabeling in lamina IThe present study demonstrates that transgenic mice expressing EGFP fluorescence under thecontrol of a GAD67 regulatory element can be used to identify some GABAergic neurons inlamina I. We noted that 73% of EGFP+ cells were also GABA+. Additionally, EGFP+ cellsmake up 19% of the population of lamina I neurons. Because ~25% of lamina I is GABAergic(Polgar et al. 2003), approximately three out of four GABAergic neurons can be visualized bythe endogenous EGFP. This further confirms the reliability of EGFP as a reporter of theGABAergic phenotype (Heinke et al. 2004; Oliva et al. 2000). However, because ~25% ofGABA+ cells are not EGFP+, it is possible that EGFP is not reporting a subpopulation ofGABAergic inhibitory interneurons and that these unlabeled interneurons represent anotherfunctional neuron subpopulation. This is unlikely for the following reasons. First, thepercentage of double-labeled cells increases after blocking axonal transport (Dougherty et al.2004; Heinke et al. 2004), suggesting that, in many neurons, GABA and/or EGFP is traffickingout of somata. Second, neuron identity relies on fluorescence and several neurons may be belowdetection threshold. Third, we have observed a considerable heterogeneity of peptide contentin these neurons (Dougherty et al. 2004), indicating that EGFP+ expression is found inneurochemically diverse populations of GABAergic neurons. Finally, EGFP+ neurons weremostly fusiform (31%) and multipolar (65%) and an earlier study suggested that populationsof multipolar and fusiform cells in lamina I are GABAergic, whereas flattened and pyramidalcells are not GABAergic (Lima et al. 1993).
It is possible that the number of fusiform cells was overestimated in morphological countsbecause cells were counted in parasagittal sections. Morphology is best determined in theparasagittal plane in the lateral half of the cord and in the horizontal plane in the medial half.Because lamina I is difficult to delineate in the horizontal plane, the parasagittal plane wasused (as in Prescott and De Koninck 2002). In thin sections (10 μm), it is possible that cells ofother morphologies appear fusiform, particularly those in the medial part of the cord. However,only 31% of cells were fusiform in this plane which seems reasonable when compared withthe Lima et al. (1993) study where 43% of GABAergic neurons were shown to be fusiform.Morphological classifications and comparisons are complicated due to inconsistencies betweengroups. Lima and Coimbra (1986) have classified lamina I neurons into four main groups—fusiform, multipolar, flattened, and pyramidal. Studies which correlated morphology witheither modality or firing (Han et al. 1998; Prescott and De Koninck 2002), did not include adistinct flattened morphology but included flattened cells in the multipolar category.Additionally, Han et al. (1998) targeted larger cells, presumably mostly projection neurons, socells called multipolar were likely flattened because multipolar cells are probably localinterneurons.
Dougherty et al. Page 6
J Neurophysiol. Author manuscript; available in PMC 2009 May 8.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Electrophysiological characterization of lamina I GAD67-EGFP+neuronsMeasured passive and threshold properties of EGFP+ neurons are comparable to previousestimates from unidentified lamina I neurons (Graham et al. 2004; Prescott and De Koninck2002; Ruscheweyh and Sandkuhler 2002). Because recordings were undertaken at roomtemperature, results cannot be directly compared with in vivo studies. With respect to firingproperties,Prescott and DeKoninck (2002) identified four firing properties in unidentified adultrat lamina I neurons—tonic, phasic, delayed, and single spike. Ruscheweyh and Sandkuhler(2002) found all four of these properties along with initial burst in unidentified neurons inlamina I of younger rats (P18-28). Presently, three of these firing properties were found inlamina I GABAergic interneurons in the young mouse—tonic, single spike, and initial burst.Interestingly, Ruscheweyh et al. (2004) observed two firing patterns in identified lamina Iprojection neurons (called gap and bursting firing) that were never seen in our EGFP+ neurons,demonstrating that firing properties divide, at least partly, into functional classes.
While we treated the population of neurons that fired tonically or with an initial burst as separatepopulations based on previous work (Ruscheweyh and Sandkuhler 2002), several observationssuggest that these neurons may constitute one functional class with modifiable firing properties.First, passive membrane and threshold properties were indistinguishable between thesepopulations. Second, for a range of current steps both firing types could be observed in 28%of these neurons (Fig. 3D). Third, initial burst and tonic firing properties were oftenbidirectionally interconvertible in individual neurons during the course of a recording (Fig. 3,E and F). Last, modulatory transmitters can reversibly convert firing properties between thesetwo types (Garraway and Hochman 2001a). Because there is evidence of continuedmonoaminergic modulation of dorsal horn neurons in acute slice preparations (Garraway andHochman 2001b), such shifts are possible.
In comparison, the passive and threshold properties of single spike cells differed considerablyfrom the aforementioned cells. They were less excitable (higher rheobase) and had lowerresistance and membrane time constant (τm) values. Because these values were well correlatedin individual neurons, it is clear that a lower membrane resistivity contributes to reduced cellexcitability.
Correlation of morphology and firing propertiesFusiform and multipolar morphologies were the predominant cell types of lamina I EGFP+
neurons. Because cells of all three firing properties were found in both morphological celltypes, morphology and intrinsic membrane properties are shaped at least partly by independentmechanisms. Nonetheless, the majority of fusiform cells were tonic or initial burst (88%), andthe majority of multipolar cells were single spike (56%). These differences in incidence andtheir associated differences in membrane properties led multipolar cells to have acomparatively higher rheobase and shorter τm values. These properties, and the smallercapacitance of multipolar cells, are consistent with the results obtained by Prescott andDeKoninck (2002), indicating that fusiform cells are larger and more excitable than multipolarcells.
The cells termed phasic by Prescott and DeKoninck (2002) are analogous to the initial burstcells in our study and in the work of Ruschweyh and Sandkuhler (2002). With this in mind,the incidence of the three major firing types in fusiform neurons here is very similar to thatreported by Prescott and DeKoninck (2002). Because not all fusiform neurons are GABAergic(Lima et al. 1993), the implication is that firing properties for this cell morphology are notdistinguished based on transmitter phenotype and excitatory interneurons with the samemorphology express a similar range in firing properties. Comparing our work to Prescott andDeKoninck (2002), multipolar cells in both GABAergic neurons and lamina I cells in general
Dougherty et al. Page 7
J Neurophysiol. Author manuscript; available in PMC 2009 May 8.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

are mostly single spike (56 and 58%, respectively) with initial burst/phasic cells making up asmall percentage of the multipolar population (13 and 5%, respectively). However, we neverobserved delayed firing, whereas Prescott and DeKo-ninck (2002) never observed tonic firingin multipolar neurons. We cannot explain these striking differences but offer several possibleexplanations. First, it is possible that the cells expressing delayed firing were flattened cellsthat were not differentiated from multipolar cells. Flattened cells are not GABAergic (Lima etal. 1993) and so would not have been targeted based on EGFP expression in the current study.Second, Prescott and DeKoninck (2002) studied adult rat, and we studied the young mouse,so it is possible that firing properties differ between species or developmentally (but seeHochman et al. 1997). Last, it is possible that delayed, tonic, and initial burst cells are oneneuron population whose firing properties are sculpted by neuromodulation, as observed inother studies (e.g., Garraway and Hochman 2001a) and that unknown factors bias differentstudies toward different neuromodulatory “states”.
We conclude that the firing properties of the lamina I inhibitory apparatus are functionallydivisible into at least two major groups representing a subset of the firing properties observedfor neurons in this lamina.
ACKNOWLEDGMENTSWe are indebted to M. Hatcher and M. Daugherty for expert technical assistance and to Dr. Stefan Clemens and D.Machacek for comments on the manuscript.
GRANTS This work was supported by the National Institute of Neurological Disorders and Stroke Grants NS-45248and NS-40893 to S. Hochman and fellowship award NS-49784 to K. J. Dougherty.
REFERENCESAlvarez FJ, Kavookjian AM, Light AR. Synaptic interactions between GABA-immunoreactive profiles
and the terminals of functionally defined myelinated nociceptors in the monkey and cat spinal cord. JNeurosci 1992;12:2901–2917. [PubMed: 1494939]
Andrew D, Craig AD. Responses of spinothalamic lamina I neurons to maintained noxious mechanicalstimulation in the cat. J Neurophysiol 2002;87:1889–1901. [PubMed: 11929909]
Barber RP, Vaughn JE, Roberts E. The cytoarchitecture of GABAergic neurons in rat spinal cord. BrainRes 1982;238:305–328. [PubMed: 7046873]
Chen M, Gu JG. A P2X receptor-mediated nociceptive afferent pathway to lamina I of the spinal cord.Mol Pain 2005;1:4. [PubMed: 15813988]
Chery N, Yu XH, De Koninck Y. Visualization of lamina I of the dorsal horn in live adult rat spinal cordslices. J Neurosci Methods 2000;96:133–142. [PubMed: 10720677]
Christensen BN, Perl ER. Spinal neurons specifically excited by noxious or thermal stimuli: marginalzone of the dorsal horn. J Neurophysiol 1970;33:293–307. [PubMed: 5415075]
Craig AD, Andrew D. Responses of spinothalamic lamina I neurons to repeated brief contact heatstimulation in the cat. J Neurophysiol 2002;87:1902–1914. [PubMed: 11929910]
Craig AD, Krout K, Andrew D. Quantitative response characteristics of thermoreceptive and nociceptivelamina I spinothalamic neurons in the cat. J Neurophysiol 2001;86:1459–1480. [PubMed: 11535691]
Dougherty KJ, Sawchuk M, Hochman S. Properties of GABAergic neurons in spinal cord lamina I ofGAD67-EGFP expressing transgenic mice. Soc Neurosci Abstr 2003;260:17.
Dougherty KJ, Sawchuk M, Hochman S. Continuing characterization of GABAergic neurons in spinalcord lamina I of GAD67-EGFP expressing transgenic mice. Soc Neurosci Abstr 2004;292:14.
Fedirchuk B, Dai Y. Monoamines increase the excitability of spinal neurones in the neonatal rat byhyperpolarizing the threshold for action potential production. J Physiol 2004;557:355–361. [PubMed:15090607]
Dougherty et al. Page 8
J Neurophysiol. Author manuscript; available in PMC 2009 May 8.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Garraway SM, Hochman S. Modulatory actions of serotonin, norepinephrine, dopamine, andacetylcholine in spinal cord deep dorsal horn neurons. J Neurophysiol 2001a;86:2183–2194.[PubMed: 11698510]
Garraway SM, Hochman S. Pharmacological characterization of serotonin receptor subtypes modulatingprimary afferent input to deep dorsal horn neurons in the neonatal rat. Br J Pharmacol 2001b;132:1789–1798. [PubMed: 11309251]
Gauriau C, Bernard JF. A comparative reappraisal of projections from the superficial laminae of the dorsalhorn in the rat: the forebrain. J Comp Neurol 2004;468:24–56. [PubMed: 14648689]
Graham BA, Brichta AM, Callister RJ. In vivo responses of mouse superficial dorsal horn neurons toboth current injection and peripheral cutaneous stimulation. J Physiol 2004;561:749–763. [PubMed:15604230]
Han ZS, Zhang ET, Craig AD. Nociceptive and thermoreceptive lamina I neurons are anatomicallydistinct. Nat Neurosci 1998;1:218–225. [PubMed: 10195146]
Heinke B, Ruscheweyh R, Forsthuber L, Wunderbaldinger G, Sandkuhler J. Physiological,neurochemical and morphological properties of a subgroup of GABAergic spinal lamina II neuronsidentified by expression of green fluorescent protein in mice. J Physiol 2004;560:249–266. [PubMed:15284347]
Hochman S, Garraway SM, Pockett S. Membrane properties of deep dorsal horn neurons from neonatalrat spinal cord in vitro. Brain Res 1997;767:214–219. [PubMed: 9367250]
Jankowska E. Interneuronal relay in spinal pathways from proprioceptors. Prog Neurobiol 1992;38:335–378. [PubMed: 1315446]
Lima D, Avelino A, Coimbra A. Morphological characterization of marginal (lamina I) neuronsimmunoreactive for substance P, enkephalin, dynorphin and gamma-aminobutyric acid in the ratspinal cord. J Chem Neuroanat 1993;6:43–52. [PubMed: 7679910]
Lima D, Coimbra A. A Golgi study of the neuronal population of the marginal zone (lamina I) of the ratspinal cord. J Comp Neurol 1986;244:53–71. [PubMed: 3950090]
Ma W, Behar T, Chang L, Barker JL. Transient increase in expression of GAD65 and GAD67 mRNAsduring postnatal development of rat spinal cord. J Comp Neurol 1994;346:151–160. [PubMed:7962709]
Mackie M, Hughes DI, Maxwell DJ, Tillakaratne NJ, Todd AJ. Distribution and colocalization ofglutamate decarboxylase isoforms in the rat spinal cord. Neuroscience 2003;119:461–472. [PubMed:12770560]
Oliva AA Jr, Jiang M, Lam T, Smith KL, Swann JW. Novel hippocampal interneuronal subtypesidentified using transgenic mice that express green fluorescent protein in GABAergic interneurons.J Neurosci 2000;20:3354–3368. [PubMed: 10777798]
Polgar E, Hughes DI, Riddell JS, Maxwell DJ, Puskar Z, Todd AJ. Selective loss of spinal GABAergicor glycinergic neurons is not necessary for development of thermal hyperalgesia in the chronicconstriction injury model of neuropathic pain. Pain 2003;104:229–239. [PubMed: 12855333]
Prescott SA, De Koninck Y. Four cell types with distinctive membrane properties and morphologies inlamina I of the spinal dorsal horn of the adult rat. J Physiol 2002;539:817–836. [PubMed: 11897852]
Rudomin P, Schmidt RF. Presynaptic inhibition in the vertebrate spinal cord revisited. Exp Brain Res1999;129:1–37. [PubMed: 10550500]
Ruscheweyh R, Ikeda H, Heinke B, Sandkuhler J. Distinctive membrane and discharge properties of ratspinal lamina I projection neurones in vitro. J Physiol 2004;555:527–543. [PubMed: 14694142]
Ruscheweyh R, Sandkuhler J. Lamina-specific membrane and discharge properties of rat spinal dorsalhorn neurons in vitro. J Physiol 2002;541:231–244. [PubMed: 12015432]
Schmidt BJ, Hochman S, MacLean JN. NMDA receptor-mediated oscillatory properties: potential rolein rhythm generation in the mammalian spinal cord. Ann NY Acad Sci 1998;860:189–202. [PubMed:9928312]
Soghomonian JJ, Martin DL. Two isoforms of glutamate decarboxylase: why? Trends Pharmacol Sci1998;19:500–505. [PubMed: 9871412]
Spike RC, Puskar Z, Andrew D, Todd AJ. A quantitative and morphological study of projection neuronsin lamina I of the rat lumbar spinal cord. Eur J Neurosci 2003;18:2433–2448. [PubMed: 14622144]
Dougherty et al. Page 9
J Neurophysiol. Author manuscript; available in PMC 2009 May 8.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Todd AJ, McKenzie J. GABA-immunoreactive neurons in the dorsal horn of the rat spinal cord.Neuroscience 1989;31:799–806. [PubMed: 2594201]
Todd AJ, Puskar Z, Spike RC, Hughes C, Watt C, Forrest L. Projection neurons in lamina I of rat spinalcord with the neurokinin 1 receptor are selectively innervated by substance p-containing afferentsand respond to noxious stimulation. J Neurosci 2002;22:4103–4113. [PubMed: 12019329]
Todd AJ, Spike RC, Young S, Puskar Z. Fos induction in lamina I projection neurons in response tonoxious thermal stimuli. Neuroscience 2005;131:209–217. [PubMed: 15680704]
Torsney C, MacDermott AB. NK1 receptor expressing rat spinal cord dorsal horn cells: primary afferentsynaptic inputs and alterations in the absence of GABAergic/glycinergic inhibition. Soc NeurosciAbst 2004;292:9.
Willis, WD.; Coggeshall, RE. Sensory Mechanisms of the Spinal Cord. Vol. 2nd ed.. Plenum; New York:1991.
Dougherty et al. Page 10
J Neurophysiol. Author manuscript; available in PMC 2009 May 8.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig. 1.Comparison of enhanced green fluorescent protein (EGFP) fluorescence to GABAimmunolabeling in lamina I neurons in P14 glutamic acid decarboxylase 67 (GAD67)-EGFPmice. A: EGFP+ cells are evident throughout the dorsal horn. B: somatic GABA labeling isrestricted to the superficial laminae; however, GABA+ processes are seen throughout the dorsalhorn. C: overlay of A and B shows that most of the EGFP+ cells in lamina I are also GABA+
(colored yellow; filled arrows), but there are EGFP+ cells that are not GABA+ (stick arrow)and GABA+ cells that are not EGFP+ (open arrow). Transverse spinal slice (10 μm) is fromlumbar cord. Scale bar is 50 μm.
Dougherty et al. Page 11
J Neurophysiol. Author manuscript; available in PMC 2009 May 8.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig. 2.Morphological cell types in lamina I at P14. There are four main morphological cell types oflamina I EGFP+ cells. Mean values for multipolar (A), fusiform (B), and pyramidal (not shown)are 65, 31, and 4%, respectively. Dendritic extent could be easily followed with endogenousEGFP fluorescence (C). Cells are shown in live slices cut in the parasagittal plane. Scale barsare 10 μm. →, the primary process used to identify cell morphology in A and B and the somain C.
Dougherty et al. Page 12
J Neurophysiol. Author manuscript; available in PMC 2009 May 8.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig. 3.Firing properties displayed by lamina I EGFP+ cells. In response to 1 s long current steps,EGFP+ lamina I neurons fired tonically (A1, n = 46) with a single spike (A2, n = 59) with aninitial burst (A3, n = 28) or after a delay (not shown, n = 1). B: several cells classified as tonicfired with an initial burst in response to larger-amplitude current steps (n = 19). C: during thecourse of recordings, some cells that initially fired tonically subsequently switched to firingan initial burst (C1, n = 10) in response to current steps of the same magnitude. Conversely,some cells firing with an initial burst switched to tonic firing (C2, n = 4) later during therecordings. All scale bars are 20 mV and 100 ms (as shown in A).
Dougherty et al. Page 13
J Neurophysiol. Author manuscript; available in PMC 2009 May 8.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
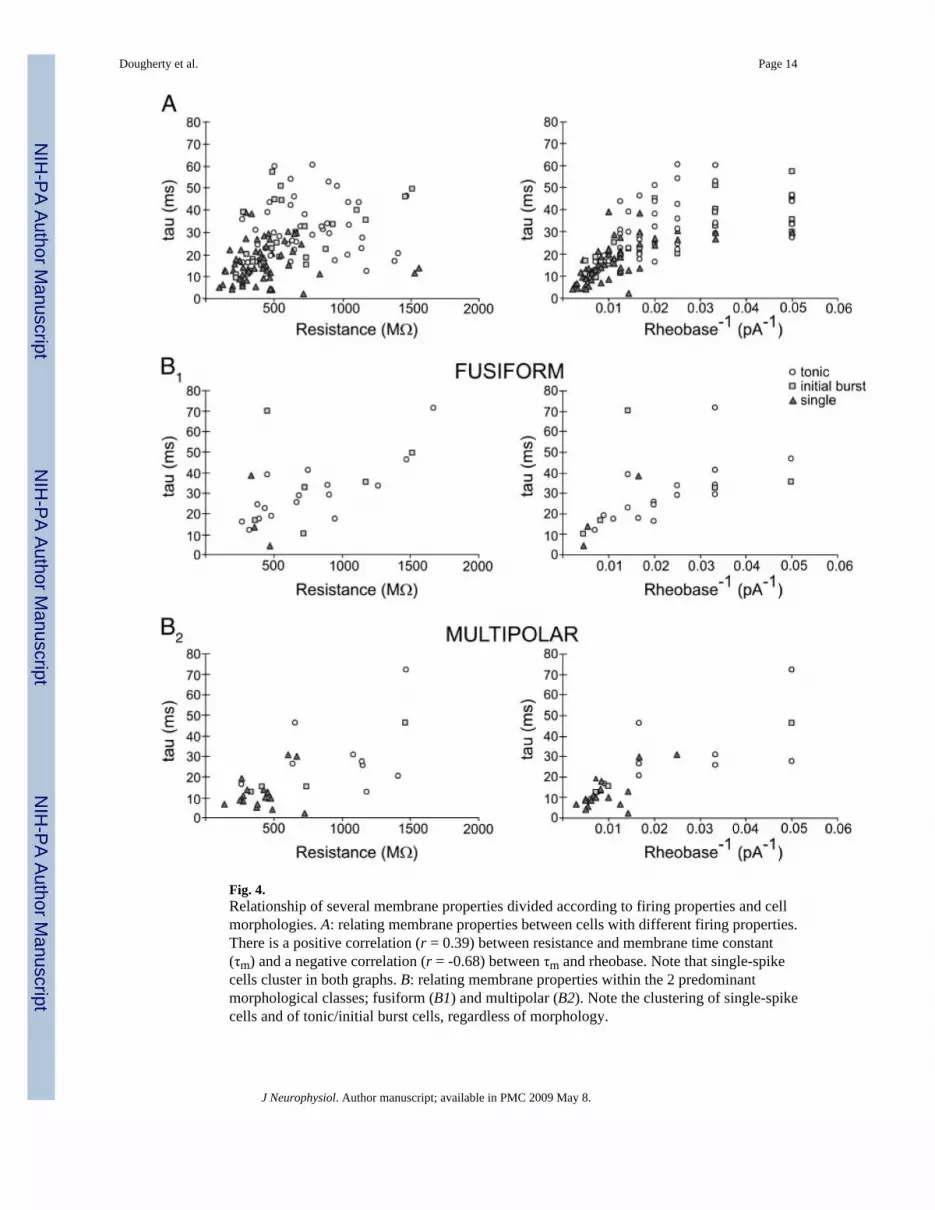
Fig. 4.Relationship of several membrane properties divided according to firing properties and cellmorphologies. A: relating membrane properties between cells with different firing properties.There is a positive correlation (r = 0.39) between resistance and membrane time constant(τm) and a negative correlation (r = -0.68) between τm and rheobase. Note that single-spikecells cluster in both graphs. B: relating membrane properties within the 2 predominantmorphological classes; fusiform (B1) and multipolar (B2). Note the clustering of single-spikecells and of tonic/initial burst cells, regardless of morphology.
Dougherty et al. Page 14
J Neurophysiol. Author manuscript; available in PMC 2009 May 8.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Dougherty et al. Page 15
Table 1Comparison of intrinsic membrane properties between cells with different firing properties
Single (59) Tonic (46) Initial Burst (28) ANOVA (P)
Membrane potential, mV -62.8 ± 1.1 -63.5 ± 1.0 -61.5 ± 1.1 0.52
Resistance, MΩ 424 ± 35* 721 ± 48 618 ± 69 <0.0001
τm, ms 16.3 ± 1.2* 30.3 ± 2.0 28.1 ± 2.8 <0.0001
Capacitance, pF 46.2 ± 3.8 48.9 ± 3.9 51.6 ± 5.5 0.69
Rheobase, pA 135 ± 12* 65 ± 5 72 ± 11 <0.0001
Maximum firing frequency, s-1 N/A 38.6 ± 2.1 33.1 ± 2.6 0.34
Spike height, mV 45.3 ± 1.6 53.5 ± 2.4 47.2 ± 3.5 <0.05
Voltage threshold, mV -43.8 ± 1.2 -44.5 ± 1.2 -47.6 ± 1.6 0.17
*Significantly different from both tonic and initial burst.
J Neurophysiol. Author manuscript; available in PMC 2009 May 8.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Dougherty et al. Page 16
Table 2Comparison of intrinsic membrane properties within the two predominant morphological classes
Fusiform (25) Multipolar (32)
Membrane potential, mV -58.8 ± 2.2 -59.9 ± 1.9
Resistance, MΩ 723 ± 82 607 ± 69
τm, ms 30.0 ± 3.4 18.7 ± 2.6*
Capacitance, pF 47.8 ± 6.3 33.9 ± 3.1*
Rheobase, pA 74 ± 12 110 ± 12*
Max firing frequency, s-1 41.2 ± 3.5 39.6 ± 4.6
Spike height, mV 56.1 ± 2.9 50.0 ± 2.2
Voltage threshold, mV -48.5 ± 1.7 -43.5 ± 1.5*
*P < 0.05 via t-test.
J Neurophysiol. Author manuscript; available in PMC 2009 May 8.





























