Embed Size (px)
Citation preview

1999, 19(12):8033. Mol. Cell. Biol.
Steven Bharadwaj, Adnan Ali and Nick Ovsenek of Heat Shock Factor 1 In Vivo
RegulationChaperone Complex Function in Multiple Components of the HSP90
http://mcb.asm.org/content/19/12/8033Updated information and services can be found at:
These include:
REFERENCEShttp://mcb.asm.org/content/19/12/8033#ref-list-1at:
This article cites 45 articles, 29 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on Novem
ber 9, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
on Novem
ber 9, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

MOLECULAR AND CELLULAR BIOLOGY,0270-7306/99/$04.0010
Dec. 1999, p. 8033–8041 Vol. 19, No. 12
Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Multiple Components of the HSP90 Chaperone ComplexFunction in Regulation of Heat Shock Factor 1 In Vivo
STEVEN BHARADWAJ, ADNAN ALI, AND NICK OVSENEK*
Department of Anatomy and Cell Biology, College of Medicine, University of Saskatchewan,Saskatoon, Saskatchewan, Canada S7N 5E5
Received 22 April 1999/Returned for modification 14 May 1999/Accepted 30 August 1999
Rapid and transient activation of heat shock genes in response to stress is mediated in eukaryotes by the heatshock transcription factor HSF1. It is well established that cells maintain a dynamic equilibrium betweeninactive HSF1 monomers and transcriptionally active trimers, but little is known about the mechanism linkingHSF1 to reception of various stress stimuli or the factors controlling oligomerization. Recent reports haverevealed that HSP90 regulates key steps in the HSF1 activation-deactivation process. Here, we tested thehypothesis that components of the HSP90 chaperone machine, known to function in the folding and maturationof steroid receptors, might also participate in HSF1 regulation. Mobility supershift assays using antibodiesagainst chaperone components demonstrate that active HSF1 trimers exist in a heterocomplex with HSP90,p23, and FKBP52. Functional in vivo experiments in Xenopus oocytes indicate that components of the HSF1heterocomplex, as well as other components of the HSP90 cochaperone machine, are involved in regulatingoligomeric transitions. Elevation of the cellular levels of cochaperones affected the time of HSF1 deactivationduring recovery: attenuation was delayed by immunophilins, and accelerated by HSP90, Hsp/c70, Hip, or Hop.In immunotargeting experiments with microinjected antibodies, disruption of HSP90, Hip, Hop, p23, FKBP51,and FKBP52 delayed attenuation. In addition, HSF1 was activated under nonstress conditions after immuno-targeting of HSP90 and p23, evidence that these proteins remain associated with HSF1 monomers and functionin their repression in vivo. The remarkable similarity of HSF1 complex chaperones identified here (HSP90,p23, and FKBP52) and components in mature steroid receptor complexes suggests that HSF1 oligomerizationis regulated by a foldosome-type mechanism similar to steroid receptor pathways. The current evidence leadsus to propose a model in which HSF1, HSP90 and p23 comprise a core heterocomplex required for rapidconformational switching through interaction with a dynamic series of HSP90 subcomplexes.
The heat shock response is characterized by increased syn-thesis of heat shock proteins (Hsps) which prevent denatur-ation of cellular proteins under a variety of stress conditions. Inmulticellular organisms, this is directed at the transcriptionallevel by transient activation of the heat shock transcriptionfactor HSF1 (reviewed in references 26 and 44). The stressactivation and attenuation pathways involve a number of mod-ifications to the HSF1 molecule. Under nonstress conditions,HSF1 exists as transcriptionally inactive non-DNA-bindingmonomers that, in response to various stress stimuli, assem-ble into active homotrimers capable of binding to heat shockelements (HSEs) in hsp gene promoters. These oligomericchanges involve dynamic inter- and intramolecular interactionsbetween conserved hydrophobic heptad repeats. Additionalsteps include hyperphosphorylation, the functional impact ofwhich is unclear, as well as activation of the transcriptionalactivation domain. Numerous studies have shown that trimerformation is tightly associated with increased HSE binding andthat transcription is regulated independently of trimerization.Active trimers are eventually converted back to inactive mono-mers after an appropriate amount of Hsp expression or uponreturn to nonstress conditions.
Although the peptide sequence requirements and oligomer-ization properties have essentially been determined, the sig-naling pathways through which various stress inducers activateHSF1, as well as the folding mechanisms controlling monomer-
to-trimer transitions, have not been elucidated. Since inductionof protein unfolding and accumulation of damaged proteins isa common feature of various stress conditions, it has beensuggested that Hsps, as molecular chaperones sensitive to thelevel of denatured proteins, provide a basis for HSF1 regula-tion. An autoregulatory role for Hsp/c70 is supported by nu-merous observations in various model systems (1, 3, 4, 7, 11, 13,25, 31). However, a mechanism by which Hsp/c70 influencesdifferent steps of the HSF1 activation-deactivation pathwayhas not been clearly defined. Hsp/c70 may function to maintainHSF1 in its inactive monomeric state or to disassemble trimersfollowing stress (5, 27, 37) and also, along with Hdj1, to repressthe transcriptional activation domain during attenuation (42).However, since Hsp/c70 has not been linked to trimer forma-tion or transcriptional upregulation in the initial phases of theactivation pathway, it appears that other chaperones or cellularfactors must be involved in modulating HSF1.
HSP90 has emerged as a key factor in the regulation ofHSF1. The initial evidence came from affinity chromatographyexperiments demonstrating HSF1 interaction with HSP90 invitro (28) and later from in vitro reconstitution experimentsshowing HSF1 interaction with HSP90 and several associatedcochaperones (29). Two recent reports provided more directclues. Using an in vitro HeLa cell extract system, Zou et al.demonstrated enhanced threshold activation of HSF1 with theHSP90-binding agent geldanamycin, or with antibody immu-nodepletion of HSP90, and concluded that HSP90 acts as anHSF1 repressor (45). Using the Xenopus oocyte model system,our laboratory has shown that HSP90 is present in immunecomplexes with both active and inactive HSF1 and further thatdisruption of HSP90 in vivo with geldanamycin or microin-
* Corresponding author. Mailing address: Department of Anatomyand Cell Biology, College of Medicine, University of Saskatchewan,107 Wiggins Rd., Saskatoon, SK, Canada S7N 5E5. Phone: (306)966-4069. Fax: (306) 966-4298. E-mail: [email protected].
8033
on Novem
ber 9, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

jected antibodies activates HSF1, delays recovery, and inhibitsHSF1-mediated transcription after heat shock (2). This evi-dence led us to suggest a model for HSF1 regulation in whichHSP90 forms a complex with HSF1 and participates in mod-ulating both monomer-trimer transitions and transcriptionalactivity (2).
HSP90 functions as part of a large and dynamic heterocom-plex along with a number of cochaperones including Hip (p48),Hop (p60), p23, Hsp/c70, and the immunophilins Cyp-40,FKBP51, and FKBP52. This multifunctional chaperone com-plex prevents aggregation of unfolded proteins under stressfulconditions and also regulates the folding and maturation of anumber of signal transduction molecules and receptors (forreviews, see references 12 and 36). Receptors for progesterone(PR) and glucocorticoid (GR) are well-characterized cellulartargets of the HSP90 chaperone complex (reviewed in refer-ence 35). HSP90-receptor interactions establish and maintainhigh-affinity hormone binding conformations through a step-wise assembly process involving sequential formation of anintermediate complex containing HSP90, Hsp/c70, Hip, andHop and a mature complex containing HSP90, p23, and animmunophilin. The molecular details of HSP90 function andthe specific roles of cochaperones in the steroid folding path-ways are currently being investigated by several groups usingthe cell-free reticulocyte lysate model system.
It had previously been hypothesized that assembly mecha-nisms similar to the steroid receptor pathways may also beinvolved in the folding of various HSP90 target substrates,including HSF1 (29). Here we use Xenopus oocytes as an invivo model to investigate the functional roles of HSP90 co-chaperones in HSF1 regulation under relevant physiologicalconditions. Immunotargeting experiments involving direct nu-clear microinjection of antibodies against individual HSP90chaperone complex proteins resulted in either activation ofHSF1 under nonstress conditions or significant delay of atten-uation. In addition, mobility supershifts of HSF1-HSE com-plexes were observed with antibodies against HSP90, p23, andFKBP52, suggesting these proteins assemble into a heterocom-plex with active HSF1 trimers. The data provide evidence thatmultiple components of the HSP90 chaperone machinery par-ticipate in regulating HSF1 in vivo.
MATERIALS AND METHODS
Oocyte manipulations and stress treatments. Xenopus laevis frogs were pur-chased from Xenopus I (Ann Arbor, Mich.). Ovary portions were surgicallyremoved from adult female frogs, and follicular cells were removed from oocytesby treatment in calcium-free OR2 buffer (82.5 mM NaCl, 2.5 mM KCl, 1 mMMgCl2, 1 mM NaH2PO4, 5 mM HEPES [pH 7.8], 10 mg of streptomycin sulfateper liter, 10 mg of benzylpenicillin per liter) containing 2 mg of collagenase (typeII; Sigma) per ml for 3 h at 18°C. Oocytes were washed extensively, allowed torecover for 4 h in OR2 (as above, with 1 mM CaCl2) at 18°C, and maintained inOR2 during experimental treatments. Only stage VI oocytes were selected forexperiments. Nuclei and cytoplasm were obtained under OR2 by scoring animalhemispheres with a needle and gently squeezing the equatorial region withwatchmaker’s forceps. The nonshock temperature was 18°C; unless otherwiseindicated, heat shock was performed at 33°C for 1 h. In all experiments, aminimum of 20 oocytes was used for each sample.
Immunotargeting and protein overexpression. For immunotargeting experi-ments, antibodies were diluted (1:1) in sterile H2O immediately prior to micro-injection. Antibody solutions (15 nl) were injected directly into the nucleus orcytoplasm as indicated. Injected oocytes were incubated for 30 min at 18°C andthen further incubated at the nonshock temperature (18°C), heat shocked at33°C, or heat shocked and then placed at control temperature during recoveryfor the indicated times. Protein extracts were prepared for gel mobility shiftanalysis and immunoblotting immediately after treatments. HSP90 monoclonalantibody (MAb) SPA 830 was from StressGen, Victoria, British Columbia, Can-ada; anti-HSF1 antiserum was provided by R. Morimoto, Northwestern Univer-sity, Evanston, Ill.; anti-YY1 polyclonal antibody (PAb), anti-human IkB-a PAb,and NFkB (p65) PAb sc-372-G were from Santa Cruz Biotechnology, SantaCruz, Calif.; anti-Hsp70 MAb clone 3a3 and Cyp-40 PAb PA3-023 were fromAffinity BioReagents, Golden, Colo.; Hip (clone 2G6), Hop (clone f5), FKBP51
(clone Hi51), and FKBP52 (clone Hi52c) MAbs were gifts from D. Smith,University of Nebraska, Omaha; p23 MAb clone JJ3 was a gift from D. Toft,Mayo Graduate School, Rochester, Minn. Anti-human PTP-1B antibody clone15 was from Transduction Laboratories, Lexington, Ky.; anti-human CREB PAbwas from New England Biolabs, Beverly, Mass.
For overexpression experiments, capped mRNAs encoding human chaperoneswere synthesized in vitro, using T7 RNA polymerase (Pharmacia Biotech); 50 nlof mRNA solution (2 mg/ml) was injected directly into oocyte cytoplasm. Ex-pression plasmids encoding wild-type Hip (33), Hop (9), Cyp-40 (22), FKBP51(30), and FKBP52 (30) were gifts from D. Smith. Expression plasmid for humanp23 (20) was a gift from D. Toft. Purified bovine HSP90 was a gift from R. Matts,Oklahoma State University, Stillwater; purified Hsp/c70 was from StressGen. Forelevation of Hsps, 20 ng of each protein was injected directly into oocyte nuclei.Identical concentrations of bovine serum albumin (BSA) were used for controls.
Protein extracts and gel mobility shift assays. Protein extracts were preparedby homogenizing oocytes in buffer C (50 mM Tris-Cl [pH 7.9], 20% glycerol, 50mM KCl, 0.1 mM EDTA, 2 mM dithiothreitol, 10 mg of aprotinin per ml, 10 mgof leupeptin per ml) (14) in a Dounce homogenizer. Homogenates were trans-ferred to Eppendorf tubes and spun for 5 min at 15,000 3 g (4°C). The resultantsupernatants were immediately frozen in liquid nitrogen and stored at 280°C.Oocytes were homogenized in a volume of 10 ml of buffer C per oocyte.
DNA mobility shift assays were performed with HSE oligonucleotide probesas previously described (17). DNA-binding reaction mixtures contained 10 ml ofextract (1 oocyte equivalent by volume, or 20 mg of soluble protein). Bindingreactions were performed with 1 mg of poly(dI-dC), 10 mM Tris (pH 7.8), 50 mMNaCl, 1 mM EDTA, 0.5 mM dithiothreitol, and 5% glycerol in a final volume of20 ml. Reaction mixtures were incubated on ice for 20 min and immediatelyloaded onto 5% nondenaturing polyacrylamide gels containing 6.7 mM Tris-Cl(pH 7.5), 1 mM EDTA, and 3.3 mM sodium acetate. Gels were electrophoresedfor 2.5 h at 150 V, dried, and exposed to autoradiography with X-ray film (XAR;Kodak). Antibody recognition experiments were performed by adding antibodiesdirectly into DNA-binding reactions or by mixing antibodies with oocyte extractsfor 20 min on ice prior to DNA-binding reactions.
Immunoblotting. Protein extracts were fractionated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE; 10% acrylamide gel) and elec-troblotted onto polyvinylidene difluoride or nitrocellulose membranes, and theblots were blocked (2 h at room temperature) in TBST (20 mM Tris-Cl [pH 7.6],137 mM NaCl, 0.1% [vol/vol] Tween 20) containing 5% milk powder. Antibodieswere diluted in TBST with 2.5% milk (1:5,000), and blots were incubated inprimary antibody for 2 h at room temperature. Blots were washed in TBST andincubated with secondary antibody (horseradish peroxidase-conjugated goat an-ti-rabbit or anti-mouse immunoglobulin G) (Bio-Rad) diluted 1:10,000 in TBST–2.5% milk for 2 h at room temperature. Blots were washed, and proteins werevisualized by chemiluminescence (Renaissance system; Dupont NEN) and au-toradiography with X-ray film (XAR; Kodak).
CAT assays. Chloramphenicol acetyltransferase (CAT) assays were performedwith 1 oocyte equivalent of whole-cell extract from uninjected or microinjectedoocytes as previously described (2, 32). Antibody injections or heat shock treat-ments were performed 4 h after plasmid injections, and then oocytes wereincubated at the nonshock temperature for 12 h to allow for CAT expression. Apool of at least 20 microinjected oocytes was used for each experimental treat-ment. The acetylated products were separated by thin-layer chromatography andvisualized by autoradiography.
RESULTS
Antibodies against HSP90, p23, and FKBP52 recognizeheat-activated HSF1. In previous work, we demonstrated thatHSP90 associates in a complex with active HSF1 and showedevidence that HSP90 could regulate some of the dynamicchanges that occur during heat shock and attenuation (2).HSP90 is known to interact with a number of protein cofactorsin the assembly and maturation process of cellular substratessuch as PR and GR. Therefore, we were interested in testingthe hypothesis that these additional constituents of the HSP90chaperone machine participate in controlling the major struc-tural and oligomeric transitions that control the HSE-bindingactivity of HSF1.
Gel mobility supershift experiments were used to examine ifantibodies against known components of the HSP90 chaper-one complex could recognize heat-induced HSF1-HSE com-plexes. Antibodies against each of the chaperones (HSP90,Hip, Hop, Hsp/c70, p23, Cyp-40, FKBP51, and FKBP52) weremixed (individually) with aliquots of a crude extract of heat-shocked oocytes in DNA-binding reactions with labeled HSE(Fig. 1A). Clear mobility supershifts of HSF1 were observed
8034 BHARADWAJ ET AL. MOL. CELL. BIOL.
on Novem
ber 9, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
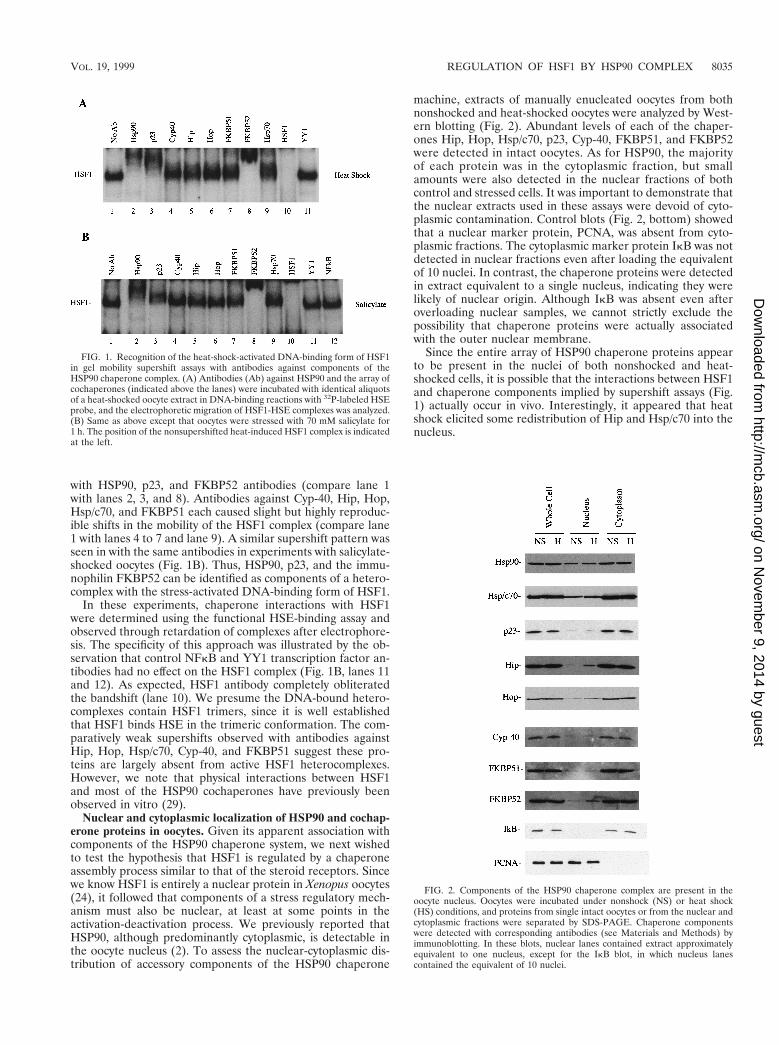
with HSP90, p23, and FKBP52 antibodies (compare lane 1with lanes 2, 3, and 8). Antibodies against Cyp-40, Hip, Hop,Hsp/c70, and FKBP51 each caused slight but highly reproduc-ible shifts in the mobility of the HSF1 complex (compare lane1 with lanes 4 to 7 and lane 9). A similar supershift pattern wasseen in with the same antibodies in experiments with salicylate-shocked oocytes (Fig. 1B). Thus, HSP90, p23, and the immu-nophilin FKBP52 can be identified as components of a hetero-complex with the stress-activated DNA-binding form of HSF1.
In these experiments, chaperone interactions with HSF1were determined using the functional HSE-binding assay andobserved through retardation of complexes after electrophore-sis. The specificity of this approach was illustrated by the ob-servation that control NFkB and YY1 transcription factor an-tibodies had no effect on the HSF1 complex (Fig. 1B, lanes 11and 12). As expected, HSF1 antibody completely obliteratedthe bandshift (lane 10). We presume the DNA-bound hetero-complexes contain HSF1 trimers, since it is well establishedthat HSF1 binds HSE in the trimeric conformation. The com-paratively weak supershifts observed with antibodies againstHip, Hop, Hsp/c70, Cyp-40, and FKBP51 suggest these pro-teins are largely absent from active HSF1 heterocomplexes.However, we note that physical interactions between HSF1and most of the HSP90 cochaperones have previously beenobserved in vitro (29).
Nuclear and cytoplasmic localization of HSP90 and cochap-erone proteins in oocytes. Given its apparent association withcomponents of the HSP90 chaperone system, we next wishedto test the hypothesis that HSF1 is regulated by a chaperoneassembly process similar to that of the steroid receptors. Sincewe know HSF1 is entirely a nuclear protein in Xenopus oocytes(24), it followed that components of a stress regulatory mech-anism must also be nuclear, at least at some points in theactivation-deactivation process. We previously reported thatHSP90, although predominantly cytoplasmic, is detectable inthe oocyte nucleus (2). To assess the nuclear-cytoplasmic dis-tribution of accessory components of the HSP90 chaperone
machine, extracts of manually enucleated oocytes from bothnonshocked and heat-shocked oocytes were analyzed by West-ern blotting (Fig. 2). Abundant levels of each of the chaper-ones Hip, Hop, Hsp/c70, p23, Cyp-40, FKBP51, and FKBP52were detected in intact oocytes. As for HSP90, the majorityof each protein was in the cytoplasmic fraction, but smallamounts were also detected in the nuclear fractions of bothcontrol and stressed cells. It was important to demonstrate thatthe nuclear extracts used in these assays were devoid of cyto-plasmic contamination. Control blots (Fig. 2, bottom) showedthat a nuclear marker protein, PCNA, was absent from cyto-plasmic fractions. The cytoplasmic marker protein IkB was notdetected in nuclear fractions even after loading the equivalentof 10 nuclei. In contrast, the chaperone proteins were detectedin extract equivalent to a single nucleus, indicating they werelikely of nuclear origin. Although IkB was absent even afteroverloading nuclear samples, we cannot strictly exclude thepossibility that chaperone proteins were actually associatedwith the outer nuclear membrane.
Since the entire array of HSP90 chaperone proteins appearto be present in the nuclei of both nonshocked and heat-shocked cells, it is possible that the interactions between HSF1and chaperone components implied by supershift assays (Fig.1) actually occur in vivo. Interestingly, it appeared that heatshock elicited some redistribution of Hip and Hsp/c70 into thenucleus.
FIG. 1. Recognition of the heat-shock-activated DNA-binding form of HSF1in gel mobility supershift assays with antibodies against components of theHSP90 chaperone complex. (A) Antibodies (Ab) against HSP90 and the array ofcochaperones (indicated above the lanes) were incubated with identical aliquotsof a heat-shocked oocyte extract in DNA-binding reactions with 32P-labeled HSEprobe, and the electrophoretic migration of HSF1-HSE complexes was analyzed.(B) Same as above except that oocytes were stressed with 70 mM salicylate for1 h. The position of the nonsupershifted heat-induced HSF1 complex is indicatedat the left.
FIG. 2. Components of the HSP90 chaperone complex are present in theoocyte nucleus. Oocytes were incubated under nonshock (NS) or heat shock(HS) conditions, and proteins from single intact oocytes or from the nuclear andcytoplasmic fractions were separated by SDS-PAGE. Chaperone componentswere detected with corresponding antibodies (see Materials and Methods) byimmunoblotting. In these blots, nuclear lanes contained extract approximatelyequivalent to one nucleus, except for the IkB blot, in which nucleus lanescontained the equivalent of 10 nuclei.
VOL. 19, 1999 REGULATION OF HSF1 BY HSP90 COMPLEX 8035
on Novem
ber 9, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

Effect of chaperone overexpression on HSF1 in vivo. In thenext experiments, we were interested in determining, underrelevant physiological conditions, the potential for each com-ponent of the HSP90 chaperone machine to function as mod-ulators of HSF1. The first strategy was to overexpress individ-ual proteins by microinjection of in vitro-synthesized mRNAsinto oocytes and then assess any subsequent effects on theHSE-binding function of HSF1 in vivo. It was possible to attainapproximately twofold increases in the amount of cochaper-ones with cytoplasmic injection of mRNA (see Fig. 4, right),but this approach failed to elevate Hsp/c70 and HSP90 levels(data not shown), probably because these are already veryabundant proteins. We solved this problem by microinjectingpurified Hsp/c70 and HSP90 directly into oocyte nuclei, afterwhich we could detect substantial elevation of their respectivelevels (Fig. 3, right). HSF1 activity was then examined in oo-cytes in which each component of the HSP90 chaperone com-plex was individually elevated, either in nonshocked cells, im-mediately following heat shock, or at several time pointsduring recovery. In each experiment, uninjected oocytes fromthe same ovary were used as parallel controls, since we haveoften observed that the recovery profile of HSF1 can varybetween cell batches from different females. HSF1 remainedinactive under nonstress conditions after twofold elevation ofeach chaperone, and the relative amounts of induced HSF1-HSE complexes were similar to those in uninjected controlsimmediately following heat shock (Fig. 3 and 4). Elevation ofHSP90 and Hsp/c70 resulted in a slightly accelerated rate ofrecovery relative to uninjected controls (Fig. 3). Elevating Hipand Hop resulted in marked acceleration in the rate of recov-ery, but no significant effect on recovery was observed afteroverexpression of p23 (Fig. 4B). Interestingly, each of theimmunophilins Cyp-40, FKBP51, and FKBP52 delayed recov-ery relative to uninjected controls (Fig. 4B). With each chap-erone, the same pattern was consistently observed in replicateexperiments, regardless of the time frame of recovery in con-trols (data not shown). Despite contrasting results with immu-nophilins and other chaperones, the simplest interpretation ofthese experiments is that multiple components of the HSP90machine are involved in the process of trimer disassembly
during attenuation. Nascent Hip, Hop, and microinjected Hspscould have increased the efficiency of trimer disassembly eitherdirectly via their independent chaperone activities or by pro-moting the formation of additional active chaperone com-plexes. Alternatively, it is possible that nascent immunophilinshad disrupted endogenous chaperone complexes, normally re-quired for efficient trimer disassembly, through competitiveinteractions with the tetratricopeptide-binding site of HSP90,as has been reported to occur in vitro (8, 10). Note that incontrols with injected BSA, HSF1 activity was not affectedunder nonshock conditions, immediately after heat shock, orduring recovery (Fig. 3). Thus, the effects on HSF1 observed inthese experiments appeared to be a specific result of increasedchaperone concentration or activity rather than simply thepresence of additional proteins.
HSF1 activity is affected by immunotargeting chaperones invivo. An alternative strategy to overexpression of HSP90 chap-erone components is to disrupt or deplete their activities invivo. Direct microinjection of specific antibodies into Xenopusoocytes provides a unique opportunity to examine the functionof components of HSP90 chaperone complexes in vivo. This isparticularly useful because specific chaperone activities can besuppressed in a relatively short time frame, avoiding cellgrowth and viability problems that would be inevitable conse-quences of genetic or antisense knockouts of key chaperones.We have previously shown that microinjection of specific an-tibodies is an effective means to target specific protein activi-ties in oocytes. For example, injected antibodies against HSP90induced HSF1 activity under nonshock conditions, demon-
FIG. 3. Effects of elevated levels of HSP90 and Hsp/c70 on the HSE-bindingactivity of HSF1 under nonshock conditions, during heat shock, and duringrecovery. The levels of Hsp/c70 (top) and HSP90 (middle) were elevated bydirect microinjection of purified proteins into nuclei. Similar amounts of BSAwere injected into nuclei (bottom), and then oocytes were incubated at nonshock(NS) temperatures, heat shocked (time zero), or heat shocked and allowed torecover at the nonshock temperature (for times indicated at the top). Left, gelmobility shift assays comparing HSF1 activity in oocytes with elevated Hsps andidentically treated uninjected controls; right, immunoblots showing relative lev-els of HSP90 or Hsp/c70 in uninjected (U) and injected (Inj.) oocyte nuclei.
FIG. 4. Effects of elevated levels of components of the HSP90 chaperonemachine on HSF1. The levels of Hip and Hop (A) and the immunophilins Cyp-40, FKBP51, and FKBP52 or p23 (B) were increased by microinjection of cor-responding mRNAs, and HSF1 was compared with uninjected controls by gelshift assay (left). Immunoblots showing relative levels of cochaperones in mRNA-injected oocytes (Inj.) and uninjected (U) controls are shown on the right.
8036 BHARADWAJ ET AL. MOL. CELL. BIOL.
on Novem
ber 9, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
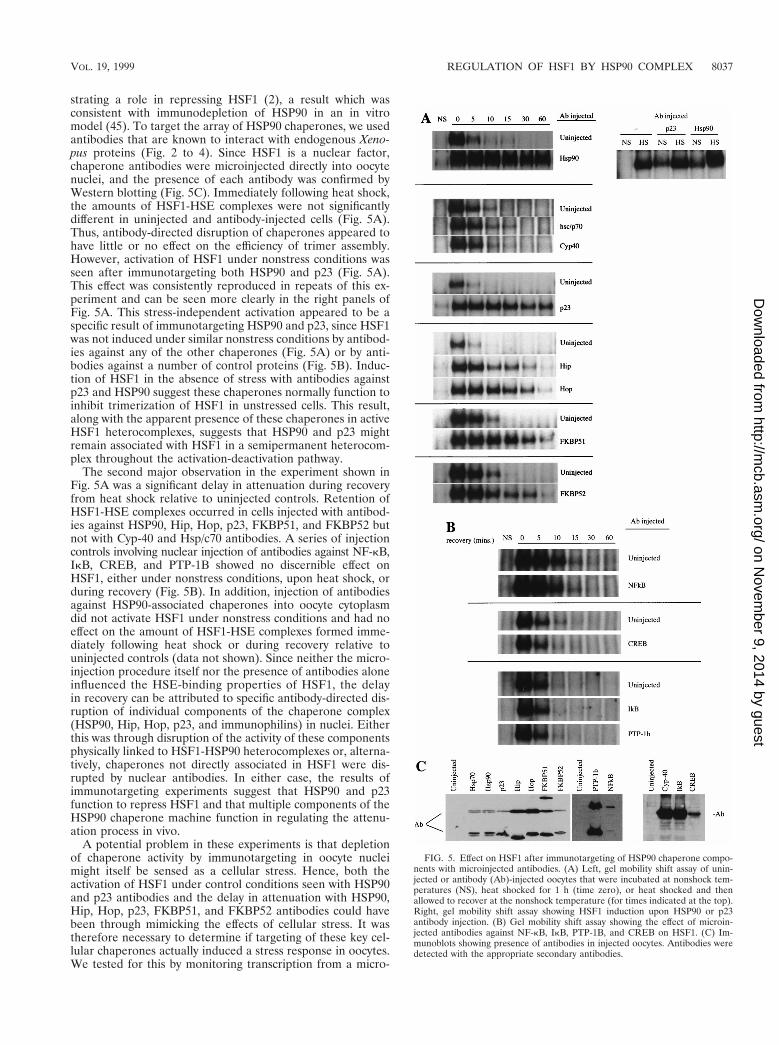
strating a role in repressing HSF1 (2), a result which wasconsistent with immunodepletion of HSP90 in an in vitromodel (45). To target the array of HSP90 chaperones, we usedantibodies that are known to interact with endogenous Xeno-pus proteins (Fig. 2 to 4). Since HSF1 is a nuclear factor,chaperone antibodies were microinjected directly into oocytenuclei, and the presence of each antibody was confirmed byWestern blotting (Fig. 5C). Immediately following heat shock,the amounts of HSF1-HSE complexes were not significantlydifferent in uninjected and antibody-injected cells (Fig. 5A).Thus, antibody-directed disruption of chaperones appeared tohave little or no effect on the efficiency of trimer assembly.However, activation of HSF1 under nonstress conditions wasseen after immunotargeting both HSP90 and p23 (Fig. 5A).This effect was consistently reproduced in repeats of this ex-periment and can be seen more clearly in the right panels ofFig. 5A. This stress-independent activation appeared to be aspecific result of immunotargeting HSP90 and p23, since HSF1was not induced under similar nonstress conditions by antibod-ies against any of the other chaperones (Fig. 5A) or by anti-bodies against a number of control proteins (Fig. 5B). Induc-tion of HSF1 in the absence of stress with antibodies againstp23 and HSP90 suggest these chaperones normally function toinhibit trimerization of HSF1 in unstressed cells. This result,along with the apparent presence of these chaperones in activeHSF1 heterocomplexes, suggests that HSP90 and p23 mightremain associated with HSF1 in a semipermanent heterocom-plex throughout the activation-deactivation pathway.
The second major observation in the experiment shown inFig. 5A was a significant delay in attenuation during recoveryfrom heat shock relative to uninjected controls. Retention ofHSF1-HSE complexes occurred in cells injected with antibod-ies against HSP90, Hip, Hop, p23, FKBP51, and FKBP52 butnot with Cyp-40 and Hsp/c70 antibodies. A series of injectioncontrols involving nuclear injection of antibodies against NF-kB,IkB, CREB, and PTP-1B showed no discernible effect onHSF1, either under nonstress conditions, upon heat shock, orduring recovery (Fig. 5B). In addition, injection of antibodiesagainst HSP90-associated chaperones into oocyte cytoplasmdid not activate HSF1 under nonstress conditions and had noeffect on the amount of HSF1-HSE complexes formed imme-diately following heat shock or during recovery relative touninjected controls (data not shown). Since neither the micro-injection procedure itself nor the presence of antibodies aloneinfluenced the HSE-binding properties of HSF1, the delayin recovery can be attributed to specific antibody-directed dis-ruption of individual components of the chaperone complex(HSP90, Hip, Hop, p23, and immunophilins) in nuclei. Eitherthis was through disruption of the activity of these componentsphysically linked to HSF1-HSP90 heterocomplexes or, alterna-tively, chaperones not directly associated in HSF1 were dis-rupted by nuclear antibodies. In either case, the results ofimmunotargeting experiments suggest that HSP90 and p23function to repress HSF1 and that multiple components of theHSP90 chaperone machine function in regulating the attenu-ation process in vivo.
A potential problem in these experiments is that depletionof chaperone activity by immunotargeting in oocyte nucleimight itself be sensed as a cellular stress. Hence, both theactivation of HSF1 under control conditions seen with HSP90and p23 antibodies and the delay in attenuation with HSP90,Hip, Hop, p23, FKBP51, and FKBP52 antibodies could havebeen through mimicking the effects of cellular stress. It wastherefore necessary to determine if targeting of these key cel-lular chaperones actually induced a stress response in oocytes.We tested for this by monitoring transcription from a micro-
FIG. 5. Effect on HSF1 after immunotargeting of HSP90 chaperone compo-nents with microinjected antibodies. (A) Left, gel mobility shift assay of unin-jected or antibody (Ab)-injected oocytes that were incubated at nonshock tem-peratures (NS), heat shocked for 1 h (time zero), or heat shocked and thenallowed to recover at the nonshock temperature (for times indicated at the top).Right, gel mobility shift assay showing HSF1 induction upon HSP90 or p23antibody injection. (B) Gel mobility shift assay showing the effect of microin-jected antibodies against NF-kB, IkB, PTP-1B, and CREB on HSF1. (C) Im-munoblots showing presence of antibodies in injected oocytes. Antibodies weredetected with the appropriate secondary antibodies.
VOL. 19, 1999 REGULATION OF HSF1 BY HSP90 COMPLEX 8037
on Novem
ber 9, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

injected reporter construct (hsp70-CAT) under control of thehsp70 promoter, which is known to be switched on by heatshock and various other stresses (2, 6, 23) (Fig. 6). If immu-nodepletion of chaperones elicited a stress response, we ex-pected to observe induction of HSF1-mediated transcriptionfrom the hsp70 promoter. In controls, CAT activity was in-duced by heat shock relative to unstressed oocytes, but noactivation was observed in antibody-injected oocytes (Fig. 6).This experiment was repeated several times with differentbatches of oocytes yielding similar results (data not shown). Inaddition, Hsp levels were unaffected by nuclear injection of anyof the chaperone antibodies (data not shown). It thereforeappears that immunodepletion of each chaperone was notperceived as a cellular stress condition. Thus, the effects ofdelayed attenuation and activation of HSF1 observed in im-munotargeting experiments were probably through direct dis-ruption of HSP90 chaperone complexes that modulate HSF1.
DISCUSSION
Here we tested the hypothesis that constituents of theHSP90 chaperone machine participate in the regulation ofHSF1 under both nonstress and heat shock conditions. Previ-ous work identifying HSP90 as a suppressor of HSF1 in HeLacell extracts (45) and a modulator of HSF1 activities in Xeno-pus oocytes (2) prompted us to examine the possibility thatHSP90 cochaperones also function in HSF1 regulatory path-way. Key predictions were that cochaperones, if present innuclei, could be detected in the HSF1-HSP90 heterocomplexand that disruption of their activities in vivo would have animpact on the DNA-binding activity of HSF1. Our results weregenerally consistent with these predictions. All of the HSP90chaperone complex proteins, although predominantly cyto-plasmic, were present at low levels in oocyte nuclei before andafter heat shock (Fig. 2). Antibodies against HSP90, p23, andFKBP52 significantly supershifted HSF1 complexes in func-tional HSE-binding assays (Fig. 1). Therefore, we identifyHSP90, p23, and the immunophilin FKBP52 as components ofa mature heterocomplex with the stress-activated form ofHSF1. Knockout or overexpression of each of the cochaper-ones had profound impacts on the HSE-binding activity ofHSF1, either during attenuation or prior to heat shock. Takentogether, our results provide strong evidence that dynamicchanges in oligomerization states of HSF1 are chaperoned bycomponents of the HSP90 chaperone machine. We concludethat HSP90 and associated cochaperones are required for fold-ing of HSF1.
In the present analysis, cochaperone interactions with HSF1were detected with the functional HSE-binding assay andretardation of DNA-binding complexes. HSP90, p23, andFKBP52 appeared to interact stably with HSF1 under theseassay conditions. In contrast, antibodies against Hip, Hop,
Hsp/c70, Cyp-40, and FKBP51 caused only slight shifts of theHSF1-HSE complex. This finding suggests that these lattercomponents of the HSP90 chaperone machine are absent fromactive HSF1 heterocomplexes. However, previous in vitro re-constitution experiments have demonstrated HSF1 interactionwith HSP90, Hsp/c70, Hop, p23, and each of the immuno-philins (29), and so the existence of larger heterocomplexesshould not yet be excluded. Putative interactions betweenHSF1 trimers and Hip, Hop, Hsp/c70, or the other immuno-philins could be more transient and hence difficult to detect, ortheir antibody interactions could be sterically hindered in alarger heterocomplex. Although the current data suggest thatHSF1 associates only with HSP90, p23, and FKBP52, it isimportant to consider that the electrophoretic mobility shiftassay/supershift technique selects for cochaperones present inactive DNA-binding complexes and excludes different HSF1-chaperone complexes that may exist at different points of theactivation-deactivation profile.
In overexpression experiments (Fig. 3 and 4), we found thatelevation of HSP90, Hsp/c70, Hip, and Hop decreased the timeof deactivation to the non-DNA-binding state during recovery.Our findings with Hsp/c70 are consistent with previous resultsobtained with mammalian and Drosophila cells after elevationof Hsp/c70 (27, 37), although this effect has not been reportedbefore with elevated levels of HSP90, Hop, and Hip. A simpleinterpretation is that these chaperones increased the efficiencyof trimer disassembly directly through their independent chap-erone activities. But it was confusing that the immunophilins(Cyp-40, FKBP51, and FKBP52) had the contrary effect ofdelaying recovery relative to uninjected controls. Since HSP90chaperone complexes are highly dynamic, it is conceivablethat nascent chaperones actually exerted their effects throughremodeling of existing chaperone complexes in vivo. Conse-quently, Hip, Hop, and the Hsps may have accelerated recov-ery by promoting formation of additional active chaperonecomplexes. This could explain the opposite effects seen withimmunophilins, which could have disrupted preexistingchaperone complexes through competitive interactions, thusdelaying recovery. An intriguing possibility is that nascent im-munophilins competed with Hop in vivo for the tetratricopep-tide-binding site of HSP90, as has been demonstrated in vitro(8). Thus the delay of attenuation with immunophilins mightbe akin to the delay seen with injected Hop antibodies (Fig.5A). It is difficult to fully explain the results of overexpressionexperiments because little is known about the behavior andfunction of HSP90 cochaperones in vivo, and we do not knowhow nascent cochaperones may have shifted the equilibrium ofHSP90-containing complexes. Also, we cannot exclude thepossibility that overexpressed cochaperones indirectly affectedHSF1 complexes either by aiding or blocking normal cellularsignaling and recovery. However, these results, combined withdetection of cochaperones in HSF1 heterocomplexes, imply amore direct functional link between the HSP90 chaperonemachine and regulation of HSF1 trimer disassembly.
Antibody-directed knockouts of HSP90 cochaperones invivo also showed profound effects on HSF1 activity. In consid-ering the results of immunotargeting experiments, it is impor-tant to note that microinjected chaperone antibodies did notactivate expression from the hsp70 promoter (Fig. 6). Thisimplies that effects on HSF1 were manifested by specific tar-geting of chaperone complexes that function to regulate HSF1rather than by mimicking a stress response. It is difficult, how-ever, to rule out the possibility that general signaling pathwaysrequired for HSF1 activity were disrupted by immunodepletionof key chaperones in vivo.
It was interesting that the HSE-binding activity of HSF1 was
FIG. 6. Effect of microinjected chaperone antibodies on HSF1-mediatedtranscription in unstressed cells. Oocytes were injected with hsp70-CAT andantibodies (Ab.) against each component of the HSP90 complex as indicated.CAT expression in antibody-injected oocytes was compared to that in similarlytreated nonshocked (NS) or heat-shocked (33°C, 1 h; HS) samples withoutinjected antibody (2). CAT assays were performed as described in Materials andMethods.
8038 BHARADWAJ ET AL. MOL. CELL. BIOL.
on Novem
ber 9, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

activated under nonshock conditions after targeting HSP90and p23 (Fig. 5). This reveals that both chaperones may func-tion to maintain HSF1 in the repressed monomeric state. Toour knowledge, our results are the first indication that p23 mayfunction as a repressor of HSF1. This was likely a specific effectof diminishing specific target protein activity in our experi-ments because HSF1 was not activated by antibodies againstany of the other chaperones or by different control antibodies.It is also interesting that HSF1 remained transcriptionally in-active (Fig. 6), and so disruption of HSP90 and p23 appearedto elicit only a partially activated conformation. This stress-independent partial activation is consistent with the idea thatadditional independent modifications of the HSF1 moleculeare necessary for transcriptional activity. Our findings withHSP90 targeting in vivo are also in agreement with the resultsof HSP90-immunodepletion in the in vitro extract system de-scribed by Zou et al. (45) and have hitherto been observed inthis laboratory (2). We note that activation of HSF1 seen herewas not observed after p23 immunodepletion in HeLa cellextracts (45) and assume that this contradiction is attributableto differences in the respective model systems.
A second major effect of immunotargeting was a significantdelay in the rate of decline of HSE complexes during recovery,which was seen with HSP90, Hip, Hop, p23, FKBP51, andFKBP52 antibodies (Fig. 5). The retention of HSE binding isinterpreted as evidence that these proteins function in theprocess of HSF1 trimer disassembly. Given the presence ofHSP90, p23, and FKBP52 in active heterocomplexes, we cansuggest that these cochaperones may function directly to un-fold trimers, or at least to potentiate disassembly. It also ap-pears that Hip, Hop, and other cochaperones influence thisprocess, despite the apparent absence of these proteins fromthe active HSF1 heterocomplex.
The immunotargeting and overexpression experiments de-scribed here were performed in vivo under relevant physiolog-ical conditions, but some inconsistencies were apparent incomparing the results of these assays with individual cochap-erones. For example, elevation of Hsp/c70 and Cyp-40 eitheraccelerated or delayed attenuation, but corresponding resultswere not obtained with injected antibodies. This could reflectdifferences in the ability of particular antibodies to interactwith target proteins or diminish chaperone activities in vivo.This was likely the case with Hsp70, for which a role in HSF1regulation has already been established (5, 27, 37, 42). Sincethe Cyp-40 homologue Cpr7 was shown to suppress the heatshock response in yeast (15), a similar role for Cyp-40 in higherorganisms should not be discounted. Also of note is that mi-croinjected antibodies against p23 significantly delayed recov-ery, but overexpression had no apparent effect on attenuation.This might suggest that exogenous p23 either did not acquirean active conformation in oocytes or failed to disrupt preex-isting chaperone complexes. It is also possible that endogenousp23 associates stably in HSF1 complexes, precluding any effectof exogenous proteins. Regardless of these discrepancies, thecombined data provide evidence that many of the differentHSP90 cochaperones play a role in HSF1 regulation.
The results reported here support the previous hypothesis ofNair et al. (29) that HSF1 is regulated similarly to steroidreceptors. Numerous in vitro studies using the rabbit reticulo-cyte lysate system have revealed that PR and GR pathwaysinvolve at least eight different protein constituents of theHSP90 complex and that assembly of the ligand-binding con-formation requires a complex and well ordered series ofdynamic interactions (9, 33, 35). In these pathways, p23 isassociated with HSP90, and both proteins are required formaintenance of the high-affinity hormone binding state (19,
20). One possible model for HSF1 regulation is that HSP90and p23 comprise a semipermanent core heterocomplex re-quired for rapid interconversion between monomers and tri-mers and that oligomerization is controlled by a foldosome-typemechanism involving a dynamic series of HSP90 subcomplexescontaining different cochaperones. This possibility is supportedby our observations of both HSP90 and p23 in the active HSF1heterocomplex, activation of HSF1 and delay of attenuationafter antibody disruption of both molecules, and the apparentinfluence of multiple cochaperones on the DNA-binding activ-ity of HSF1. The putative HSF1 core complex could be com-pared to HSP90 and p23 in telomerase complexes, in whichboth chaperones are required for assembly and remain asso-ciated with active enzyme through its cycling (18). Assembly-disassembly of HSF1 involves a dynamic equilibrium underdifferent cellular conditions, and as with steroid receptors (43),this occurs constantly. This observation leads us to suggest thatHSP90 and p23 act coordinately to maintain HSF1 monomersin an inactive state that is competent for rapid assembly intotrimers. Both molecules could remain associated with HSF1 asit is assembled into trimers, and the data suggest that bothcould be directly involved in the process of trimer disassembly.
But what are the other cochaperones doing in what is prob-ably a dynamic milieu? Studies of steroid complex formationhave shown it is a multistep process involving transient asso-ciation with Hsp/c70, Hip, and Hop. It is remarkable that thecomponents we have identified in active HSF1 heterocom-plexes, HSP90, p23, and FKBP52 are precisely the same as inlate stage or mature steroid receptor complexes (35). Thisfinding supports the hypothesis that a similar foldosome-typechaperone process occurs with HSF1. Our model predicts thatthe putative HSF1 core complex interacts with various cochap-erones in a stepwise manner, with HSP90 and p23 acting asorganizers of intermediate complexes. This could involve ad-dition and release of individual components or interaction withHSP90 subcomplexes containing different combinations of co-chaperones. As in steroid folding, we suggest that Hsp/c70 andHSP90 participate in a highly coordinated manner, along withtheir respective interacting or organizing proteins, Hop andHip, to modulate HSF1 folding. Hip, Hop, and Hsp/c70 prob-ably interact transiently with the putative core heterocom-plex as HSF1 molecules are chaperoned through oligomericchanges. This view is consistent with the known autoregulatoryroles of both HSP90 and Hsp/c70 and could explain the veryweak but reproducible supershifts of HSF1-HSE complexesthat we observe with their respective antibodies (Fig. 1).
The apparent interaction of FKBP52 with the stress-acti-vated heterocomplex is, to our knowledge, the first indicationthat immunophilins could be involved in HSF1 regulation. Itsputative role in this regard is consistent with known immu-nophilin function. Immunophilins are peptidyl-prolyl isomer-ase family proteins that mediate protein folding and are iden-tified as targets of immunosuppressive drugs cyclosporin A(Cyp-40) and FK506 (FKBPs) (16, 41). These are tetratrico-peptide repeat-containing proteins that interact with HSP90complexes (30, 38, 39), but their precise roles in receptorcomplexes have not yet been elucidated. Our observation thatantibody disruption of FKBP52 delayed attenuation suggestsits main function is to potentiate trimer disassembly. Sinceimmunophilins are also thought to be involved in targetedprotein trafficking (34), we raise the intriguing prospect thatFKBP52 mediates rapid relocalization of HSF1 to and fromstress granules within the nucleus, a phenomenon that hasrecently been observed in HeLa cells (21). We have not deter-mined whether FKBP52 remains associated with HSF1 mono-
VOL. 19, 1999 REGULATION OF HSF1 BY HSP90 COMPLEX 8039
on Novem
ber 9, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

mers or if stress granules are present in heat-shocked oocytenuclei.
It is tempting to speculate that the sequential order of in-teractions or mechanistic function of chaperones in the HSF1folding pathway is similar to those in steroid receptor folding.However, given the disparate physical characteristics and bio-logical functions of HSF1, it is likely that its folding pathway issignificantly different. At the very least, we have identifiedpotential roles for HSP90 and p23 in HSF1 repression and forHSP90, Hsp/c70, Hip, Hop, p23, and immunophilins in trimerdisassembly. This suggests a new paradigm for stress generegulation in which neither HSP90 nor Hsp70, nor any of thecochaperones, functions independently to regulate activity.Rather, it is more likely that HSF1 regulation in vivo involvesa series of coordinated interactions with multiple componentsof the large HSP90 chaperone complex. Our current modeldoes not exclude potential regulatory roles of additional fac-tors such as a recently identified human HSF1-binding protein,HSBP1 (40).
A number of important questions remain to be addressedregarding HSP90-HSF1 heterocomplexes. For example, whatare the stoichiometric relationships between different chaper-one components and HSF1, and what are the dynamics ofthese interactions at each phase of the response? In addition,what are the specific functional roles of each chaperone inoligomerization, phosphorylation, and transcription? Anotherquestion is whether the HSF1-HSP90 chaperone complex com-prises or contributes to the cellular stress-sensing mechanism.One idea is that an HSF1 heterocomplex containing key cel-lular chaperones could be directly responsive to the intracel-lular environment and transmit signals from stress-inducedchanges in the levels of denatured or misfolded proteins di-rectly to the HSF1 molecule. Alternatively, HSP90-cochaper-one complexes could act downstream of the stress-sensingmechanism to chaperone HSF1 oligomeric transitions. It isimportant to emphasize that the present work points to studiesof the steroid folding pathways for clues to help unravel fun-damental mechanisms of the stress response. Further studiesusing the Xenopus oocyte and other models can now be aimedat elucidating the dynamics and functional details of the HSF1-HSP90 chaperone complex and its potential relationship to thecellular stress-sensing mechanism.
ACKNOWLEDGMENTS
We are grateful to D. Smith for antibodies and expression vectorsfor Hip, Hop, and immunophilins, R. Morimoto for HSF1 antibody,D. Toft for p23 reagents, A. Wolffe for hsp70-CAT, and B. Roeslerfor providing various antibodies. We thank R. O’Carroll, J. Xavier,P. Mercier, and G. Davies for advice and technical assistance.
S.B. was supported by graduate scholarships from the Natural Sci-ences and Engineering and Medical Research Councils of Canada.A.A. was supported by a postdoctoral fellowship from the HealthServices Utilization Research Council of Saskatchewan. This researchwas supported by a Medical Research Council operating grant to N.O.
REFERENCES
1. Abravaya, K. A., M. Myers, S. P. Murphy, and R. I. Morimoto. 1992. Thehuman heat shock protein hsp70 interacts with HSF, the transcription factorthat regulates heat shock gene expression. Genes Dev. 6:1153–1164.
2. Ali, A., S. Bharadwaj, R. O’Carroll, and N. Ovsenek. 1998. HSP90 interactswith and regulates the activity of heat shock factor 1 in Xenopus oocytes.Mol. Cell. Biol. 18:4949–4960.
3. Ananthan, J., A. L. Goldberg, and R. Voellmy. 1986. Abnormal proteinsserve as eukaryotic stress signals and trigger the activation of heat shockgenes. Science 232:522–525.
4. Baler, R., W. J. Welch, and R. Voellmy. 1992. Heat shock gene regulation bynascent polypeptides and denatured proteins: hsp70 as a potential autoreg-ulatory factor. J. Cell Biol. 117:1151–1159.
5. Baler, R., J. Zou, and R. Voellmy. 1996. Evidence for a role of hsp70 in the
regulation of the heat shock response of mammalian cells. Cell Stress Chap-erones 1:33–39.
6. Bharadwaj, S., A. Hnatov, A. Ali, and N. Ovsenek. 1998. Regulation of theDNA-binding and transcriptional activities of heat shock factor 1 is uncou-pled in Xenopus oocytes pineapplea. Biochim. Biophys. Acta 1402:79–85.
7. Boorstein, W. R., and E. A. Craig. 1990. Transcriptional regulation of SSA3,an hsp70 gene from Saccharomyces cerevisiae. Mol. Cell. Biol. 10:3262–3267.
8. Carrello, A., E. Ingley, R. F. Minchin, S. Tsai, and T. Ratajczak. 1999. Thecommon tetratricopeptide repeat acceptor site for steroid receptor-associ-ated immunophilins and hop is located in the dimerization domain of Hsp90.J. Biol. Chem. 274:2682–2689.
9. Chen, S., and D. F. Smith. 1998. Hop as an adaptor in the heat shock protein70 (Hsp70) and Hsp90 chaperone machinery. J. Biol. Chem. 273:35194–35200.
10. Chen, S., W. P. Sullivan, D. O. Toft, and D. F. Smith. 1998. Differentialinteractions of p23 and the TPR-containing proteins Hop, Cyp40, FKBP52and FKBP51 with Hsp90 mutants. Cell Stress Chaperones 3:118–129.
11. Craig, E. A., and C. A. Gross. 1991. Is Hsp70 the cellular thermometer?Trends Biochem. Sci. 16:1472–1477.
12. Csermely, P., T. Schnaider, C. Soti, Z. Prohaszka, and G. Nardai. 1998. The90-kDa molecular chaperone family: structure, function, and clinical appli-cations. A comprehensive review. Pharmacol. Ther. 79:129–168.
13. DiDomenico, B. J., G. E. Bugaisky, and S. Lindquist. 1982. The heat shockresponse is self-regulated at both the transcriptional and posttranscriptionallevels. Cell 31:593–603.
14. Dignam, J. D., R. M. Lebovitz, and R. G. Roeder. 1983. Accurate transcrip-tion initiation by RNA polymerase II in a soluble extract from isolatedmammalian nuclei. Nucleic Acids Res. 11:1475–1489.
15. Duina, A. A., H. M. Kalton, and R. F. Gaber. 1998. Requirement for Hsp90and a CyP-40-type cyclophilin in negative regulation of the heat shock re-sponse. J. Biol. Chem. 273:18974–18978.
16. Fruman, D. A., S. J. Burakoff, and B. E. Bierer. 1994. Immunophilins inprotein folding and immunosuppression. FASEB J. 8:391–400.
17. Gordon, S., S. Bharadwaj, A. Hnatov, A. Ali, and N. Ovsenek. 1997. Distinctstress-inducible and developmentally regulated heat shock transcription fac-tors in Xenopus oocytes. Dev. Biol. 181:47–63.
18. Holt, S. E., D. L. Aisner, J. Baur, V. Tesmer, M. Dy, M. Oullette, J. Trager,G. Morin, D. O. Toft, J. W. Shay, W. E. Wright, and M. A. White. 1999.Functional requirement of p23 and Hsp90 in telomerase complexes. GenesDev. 13:817–826.
19. Hutchison, K. A., L. F. Stancato, J. K. Owens-Grillo, J. L. Johnson, P.Krishna, D. O. Toft, and W. B. Pratt. 1995. The 23-kDa acidic protein inreticulocyte lysate is the weakly bound component of the hsp foldosome thatis required for assembly of the glucocorticoid receptor into a functionalheterocomplex with hsp90. J. Biol. Chem. 270:18841–18847.
20. Johnson, J. L., T. G. Beito, C. J. Krco, and D. O. Toft. 1994. Characterizationof a novel 23-kilodalton protein of unactive progesterone receptor com-plexes. Mol. Cell. Biol. 14:1956–1963.
21. Jolly, C., Y. Usson, and R. I. Morimoto. 1999. Rapid and reversible relocal-ization of heat shock factor 1 within seconds to nuclear stress granules. Proc.Natl. Acad. Sci. USA 96:6769–6774.
22. Kieffer, L. J., T. W. Seng, W. Li, D. G. Osterman, R. E. Handschumacher,and R. M. Bayney. 1993. Cyclophilin-40, a protein with homology to the P59component of the steroid receptor complex. Cloning of the cDNA andfurther characterization. J. Biol. Chem. 15:12303–12310.
23. Landsberger, N., M. Ranjan, G. Almouzni, D. Stump, and A. P. Wolffe. 1995.The heat shock response in Xenopus oocytes, embryos, and somatic cells: aregulatory role for chromatin. Dev. Biol. 170:62–74.
24. Mercier, P. A., J. Foksa, N. Ovsenek, and J. T. Westwood. 1997. Xenopusheat shock factor 1 is a nuclear protein before heat stress. J. Biol. Chem.272:14147–14151.
25. Mifflin, L. C., and R. E. Cohen. 1994. hsc70 moderates the heat shock (stress)response in Xenopus laevis oocytes and binds to denatured protein inducers.J. Biol. Chem. 269:15718–15723.
26. Morimoto, R. I. 1998. Regulation of the heat shock transcriptional response:cross talk between a family of heat shock factors, molecular chaperones, andnegative regulators. Genes Dev. 12:3788–3796.
27. Mosser, D. D., J. Duchaine, and B. Massie. 1993. The DNA-binding activityof the human heat shock transcription factor is regulated in vivo by hsp70.Mol. Cell. Biol. 13:5427–5438.
28. Nadeau, K., A. Das, and C. T. Walsh. 1993. Hsp90 chaperonins possessATPase activity and bind heat shock transcription factors and peptidyl prolylisomerases. J. Biol. Chem. 268:1479–1487.
29. Nair, S. C., E. J. Toran, R. A. Rimerman, S. Hjermstad, T. E. Smithgall, andD. F. Smith. 1996. A pathway of multi-chaperone interactions common todiverse regulatory proteins: estrogen receptor, Fes tyrosine kinase, heatshock transcription factor HSF1, and the aryl hydrocarbon receptor. CellStress Chaperones 1:237–250.
30. Nair, S. C., R. A. Rimerman, E. J. Toran, S. Chen, V. Prapapanich, R. N.Butts, and D. F. Smith. 1997. Molecular cloning of human FKBP51 andcomparisons of immunophilin interactions with Hsp90 and progesteronereceptor. Mol. Cell. Biol. 17:594–603.
8040 BHARADWAJ ET AL. MOL. CELL. BIOL.
on Novem
ber 9, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

31. Nunes, S. L., and S. K. Calderwood. 1995. Heat shock factor 1 and the heatshock cognate 70 protein associate in high molecular weight complexes in thecytoplasm of NIH-3T3 cells. Biochem. Biophys. Res. Commun. 213:1–6.
32. Ovsenek, N., and J. J. Heikkila. 1990. DNA sequence specific binding activityof the Xenopus heat shock transcription factor is heat inducible before themidblastula transition. Development 110:427–433.
33. Prapapanich, V., S. Chen, and D. F. Smith. 1998. Mutation of Hip’s carboxy-terminal region inhibits a transitional stage of progesterone receptor assem-bly. Mol. Cell. Biol. 18:944–952.
34. Pratt, W. B., M. Czar, L. F. Stancato, and J. K. Owens. 1993. The hsp56immunophilin component of steroid receptor heterocomplexes: could this bethe elusive nuclear localization signal-binding protein? J. Steroid Biochem.Mol. Biol. 46:269–279.
35. Pratt, W. B., and D. O. Toft. 1997. Steroid receptor interactions with heatshock protein and immunophilin chaperones. Endocrine Rev. 18:306–360.
36. Pratt, W. B. 1998. The hsp90-based chaperone system: involvement in signaltransduction from a variety of hormone and growth factor receptors. Proc.Soc. Exp. Biol. Med. 217:420–434.
37. Rabindran, S. K., J. Wisniewski, L. Li, G. C. Li, and C. Wu. 1994. Interactionbetween heat shock factor and hsp70 is insufficient to suppress induction ofDNA-binding activity in vivo. Mol. Cell. Biol. 14:6552–6560.
38. Ratajczak, T., A. Carrello, P. J. Mark, B. J. Warner, R. J. Simpson, R. L.
Moritz, and A. K. House. 1993. The cyclophilin component of the unacti-vated estrogen receptor contains a tetratricopeptide repeat domain andshares identity with p59 (FKBP59). J. Biol. Chem. 268:13187–13192.
39. Renoir, J. M., C. Radanyi, L. E. Faber, and E. E. Baulieu. 1990. Thenon-DNA-binding heterooligomeric form of mammalian steroid hormonereceptors contains a hsp90-bound 59-kilodalton protein. J. Biol. Chem. 265:10740–10745.
40. Satyal, S. H., D. Chen, S. G. Fox, J. M. Kramer, and R. I. Morimoto. 1998.Negative regulation of the heat shock transcriptional response by HSBP1.Genes Dev. 12:1962–1974.
41. Schreiber, S. L. 1991. Chemistry and biology of the immunophilins and theirimmunosuppressive ligands. Science 251:283–287.
42. Shi, Y., D. D. Mosser, and R. I. Morimoto. 1998. Molecular chaperones asHSF1-specific transcriptional repressors. Genes Dev. 12:654–666.
43. Smith, D. F. 1993. Dynamics of Hsp90-progesterone receptor binding andthe disactivation model for steroid receptor complexes. Mol. Endocrinol.7:1418–1429.
44. Wu, C. 1995. Heat shock transcription factors: structure and regulation.Annu. Rev. Cell Dev. Biol. 11:441–469.
45. Zou, Z., Y. Guo, T. Guettouche, D. Smith, and R. Voellmy. 1998. Repressionof heat shock transcription factor HSF1 activation by hsp90 (HSP90 com-plex) that forms a stress-sensitive complex with HSF1. Cell 94:471–480.
VOL. 19, 1999 REGULATION OF HSF1 BY HSP90 COMPLEX 8041
on Novem
ber 9, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from





























