Embed Size (px)
Citation preview
Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/26802432
MultiplematernaloriginsofnativemodernandancienthorsepopulationsinChina
ArticleinAnimalGenetics·October2009
DOI:10.1111/j.1365-2052.2009.01950.x·Source:PubMed
CITATIONS
26
READS
92
13authors,including:
CeiridwenJEdwards
UniversityofHuddersfield
179PUBLICATIONS1,485CITATIONS
SEEPROFILE
SamWeining
NorthwestA&FUniversity
43PUBLICATIONS738CITATIONS
SEEPROFILE
LiLiu
StanfordUniversity
74PUBLICATIONS1,141CITATIONS
SEEPROFILE
FuqiuLi
thesecondhospitalJilinUniversity
491PUBLICATIONS7,969CITATIONS
SEEPROFILE
AllcontentfollowingthispagewasuploadedbySamWeiningon10June2014.
Theuserhasrequestedenhancementofthedownloadedfile.Allin-textreferencesunderlinedinbluearelinkedtopublicationsonResearchGate,lettingyouaccessandreadthemimmediately.

doi:10.1111/j.1365-2052.2009.01950.x
Multiple maternal origins of native modern and ancient horsepopulations in China
C. Z. Lei*, R. Su†, M. A. Bower‡, C. J. Edwards§,1, X. B. Wang*, S. Weining¶, L. Liu**, W. M. Xie*,
F. Li*, R. Y. Liu††, Y. S. Zhang*, C. M. Zhang* and H. Chen*,‡‡
*Shaanxi Key Laboratory of Molecular Biology for Agriculture, College of Animal Science and Technology, Northwest A & F University,
Yangling, Shaanxi 712100, China. †Henan Veterinary Medicine Engineering and Technology Center, Luoyang, Henan 471000, China.‡Glyn Daniel Laboratory for Archaeogenetics, McDonald Institute for Archaeological Research, Downing Street, Cambridge CB2 3ER, UK.§Smurfit Institute of Genetics, Trinity College, Dublin 2, Ireland. ¶College of Agronomy, Northwest A & F University, Yangling, Shaanxi
712100, China. **Archaeology Program, La Trobe University, Melbourne, Vic. 3086, Australia. ††College of Animal Science and
Technology, Guizhou University, Guiyang, Guizhou 550025, China. ‡‡Institute of Cellular and Molecular Biology, Xuzhou Normal
University, Xuzhou, Jiangsu 221116, China
Summary To obtain more knowledge of the origin and genetic diversity of domestic horses in China, this
study provides a comprehensive analysis of mitochondrial DNA (mtDNA) D-loop sequence
diversity from nine horse breeds in China in conjunction with ancient DNA data and evi-
dence from archaeological and historical records. A 247-bp mitochondrial D-loop sequence
from 182 modern samples revealed a total of 70 haplotypes with a high level of genetic
diversity. Seven major mtDNA haplogroups (A–G) and 16 clusters were identified for the 182
Chinese modern horses. In the present study, nine 247-bp mitochondrial D-loop sequences of
ancient remains of Bronze Age horse from the Chifeng region of Inner Mongolia in China (c.
4000–2000a BP) were used to explore the origin and diversity of Chinese modern horses and
the phylogenetic relationship between ancient and modern horses. The nine ancient horses
carried seven haplotypes with rich genetic diversity, which were clustered together with
modern individuals among haplogroups A, E and F. Modern domestic horse and ancient
horse data support the multiple origins of domestic horses in China. This study supports the
argument that multiple successful events of horse domestication, including separate intro-
ductions of wild mares into the domestic herds, may have occurred in antiquity, and that
China cannot be excluded from these events. Indeed, the association of Far Eastern mtDNA
types to haplogroup F was highly significant using Fisher�s exact test of independence
(P = 0.00002), lending support for Chinese domestication of this haplogroup. High diversity
and all seven mtDNA haplogroups (A–G) with 16 clusters also suggest that further work is
necessary to shed more light on horse domestication in China.
Keywords archaeology, Chinese horse, haplogroup, mitochondrial DNA D-loop, origin.
Introduction
The origin of the domestic horse has been a focus of
multi-disciplinary research for decades, but conclusions are
controversial. While osteoarchaeological data indicate a
restricted area in Central Asia for the earliest horse
domestication, genetic studies show widespread origins.
Horses have played a significant role in China�s political,
economic and military history, but the origin of domesti-
cated horses in China remains unclear. Literary sources
describe the importation of horses into China from different
parts of the world throughout history (Xie 1987), a situa-
tion that could have led to a complex mosaic pattern of
horse breeds. To explore this complexity, this study provides
a comprehensive analysis of mitochondrial DNA (mtDNA)
D-loop sequence diversity from nine native horse breeds in
China. The results of this genetic analysis are put into
Address for correspondence
H. Chen, Shaanxi Key Laboratory of Molecular Biology for Agriculture,
College of Animal Science and Technology, Northwest A & F University,
Yangling, Shaanxi 712100, China.
E-mail: [email protected]
1Present address: RLAHA, University of Oxford, Dyson Perrins Building,
South Parks Road, Oxford OX1 3QY, UK
Accepted for publication 10 June 2009
� 2009 The Authors, Journal compilation � 2009 Stichting International Foundation for Animal Genetics, Animal Genetics, 40, 933–944 933

context within a framework of ancient DNA, archaeological
and historical records. As the evidence for horse domesti-
cation is complex and ephemeral, multiple lines of evidence
from interdisciplinary research need to be integrated to
achieve a better understanding of the origins of the domestic
horse in China.
Archaeological and historical background
Wild horse (Equus ferus Boddaert, 1785) remains occur
widely in Late Palaeolithic sites in China, particularly in the
north. Small numbers of horse bones have also been found
at some Neolithic sites (Olsen 1988; Yuan & Flad 2006),
although it is not possible, based on morphology alone, to
assign them as domestic or wild. Based on our current
understanding of the available archaeological evidence,
horses were almost certainly first domesticated in Central
Asia some 5000 years ago and later introduced to China
(Levine et al.2003; Mashkour 2006). However, it is essen-
tial to note that the archaeological record for these regions
is sparse and limited, and archaeozoological data collection
and analysis are rare.
Within China, horse remains began to be more common
throughout the Bronze Age (2nd and 1st millennia BC). Horse
remains, incorporated in human burials, have been reported
from almost all the Bronze Age Qijia culture sites in the Upper
Yellow River region, northwest China, dating around
2000 BC. Such mortuary practice has been interpreted as
evidence of horse domestication; however, this is inconclu-
sive. If this claim is plausible, domestic horses were used only
for limited purposes, primarily for food and burial offerings
(Linduff 2003; Yuan & Flad 2003; Flad et al. 2007).
In the Middle and Lower Yellow River regions, domesti-
cated horses did not appear in the archaeological record
until the Late Shang period (c. 1300–1046 BC) (Linduff
2003; Yuan & Flad 2005, 2006). If these domestic horses
were imported into China, it is not clear through which
routes they came at this time, nor is it certain if the Qijia
horses found in the Upper Yellow River were related to those
from the Late Shang period. Horses may have been intro-
duced together with chariots to the Central Plains, and were
primarily symbols of status. For example, large numbers of
horses appeared as prestige sacrificial items in the Shang
royal ritual ceremonies at the capital city in Anyang,
Henan, whereas similar horse sacrifices at small scales were
also practised in other areas, such as Laoniupo in Shaanxi
and Qianzhangda in Shandong (Linduff 2003; Yuan & Flad
2005, 2006). It seems that horses had special relationships
with the Shang royal elite at this time, as oracle-bone
inscriptions recorded that even the Shang king fed horses in
the stables (Linduff 2003; Yuan & Flad 2005, 2006).
The practice of using horses and chariots in elite burials
continued during the next millennium, but diminished as
the millennium drew to a close. At the same time, however,
against the background of the development of equine
pastoral nomadism, horses gained an increasingly impor-
tant military role throughout the dynastic eras in Chinese
history, as demonstrated in the archaeological record and
ancient documents (Yuan & Flad 2005, 2006). In the
Western Zhou period (1045–771 BC), raising good breeding
horses was an important task, which was sometimes
directly overseen by the kings. It is believed that the
ancestors of the Qin dynasty specialized in raising horses for
the Western Zhou kings in the northwestern region of
China (Hsu & Linduff 1988). During the Spring and
Autumn period (770–476 BC), the military power of a state
was often measured by the number of horse-chariot units it
possessed. Early evidence for horseback riding in East Asia,
as depicted on a buckle-like bronze fitting, is from the Upper
Xiajiadian culture (1000–300 BC) in Inner Mongolia (Inner
Mongolia Team 1975). Numerous life-sized terracotta
horses along with chariots and soldiers arranged in military
formations were entombed in the Qin (221–207 BC) Em-
peror�s mausoleum near Xi�an, to show the political, mili-
tary and symbolic power of the Emperor. The demand for
the horses increased dramatically during the Han dynasty
(206 BC–AD 220) as the Chinese heartland was constantly
facing military threats from mounted nomads in the
northern steppes. The Eurasian steppes were seen as the
region where the finest horses were produced. Throughout
the Chinese history, dynastic rulers adopted various
strategies, such as marriage, warfare, tribute and trade, to
ensure a plentiful supply of horses from this region. In the
late Tang dynasty (around the 9th century AD), for example,
China paid the exorbitant price of a million bolts of silk per
year for 100 000 steppe horses (Mair 2003).
In south China, the earliest evidence for domesticated
horses is from bronze figures from the Dian culture (4th
century BC–AD 1st century) in Yunnan (Zhang 1998). Horses
were depicted as being of short built and as being used
primarily as a riding animal by the elite. Based on their
stature and morphology, these horses may have been related
to Mongolian ponies from the Central Asiatic steppes
(Chiou-Peng 2004). Starting in the Tang dynasty, horses
became an indispensable commodity in the ancient trade
system known as chama gudao, the Ancient Tea-Horse Road,
in south-west China, mainly involving the bilateral trade
between Tibet (producing horses) and the Yunnan-Sichuan
region (producing tea). The horses from Yunnan, which
were originated in other areas, became highly desirable for
the imperial court in the Central Plains (Zhang 1998, 2006).
From an economic perspective, the horse has been an
essential component of Chinese society because of its
importance for riding, draught and transportation. It was
ranked at the top of the six traditional domestic animals
(horse, pig, cattle, sheep and goat, dog and chicken) (Xie
1987). However, the demand for horses for transport and
agriculture has rapidly decreased in recent decades and as a
result, some horse breeds are today on the brink of extinc-
tion. Nevertheless, the horse is still an important source of
� 2009 The Authors, Journal compilation � 2009 Stichting International Foundation for Animal Genetics, Animal Genetics, 40, 933–944
Lei et al.934

animal power in mountainous and pastoral regions in
northern and south-western parts of China such as Xinji-
ang, Tibet, Inner Mongolia, Gansu, Qinghai, Yunnan and
Guizhou.
As a result of the limited findings of early horse remains
and because of the long history of transporting and trading
horses across the vast area of China, the origins and early
history of domesticated horses in China remain a mystery.
Population genetics can, therefore, offer an important line of
evidence to help us address this question.
Genetic research
Mitochondrial DNA D-loop sequence diversity has been an
important tool for understanding the origin of domesticated
horses (Kim et al. 1999; Lister et al. 2001; Vila et al. 2001;
Hill et al. 2002; Jansen et al. 2002; Cozzi et al. 2004; Royo
et al. 2005; Luıs et al. 2006; McGahern et al. 2006a;
Glazewska et al. 2007). Vila et al. (2001) suggested that the
mtDNA data from domesticated horses fell into six different
maternal haplogroups (A–F). Jansen et al. (2002) indicated
that seven maternal haplogroups (A–G) could additionally
be grouped into 17 distinct phylogenetic clusters. McGahern
et al. (2006a) also defined two new clusters A7 and F3, so
that the seven haplogroups (A–G) were further grouped into
19 distinct phylogenetic clusters. These results are generally
regarded as evidence for multiple occurrences of horse
domestication. However, the mtDNA results can also be
explained as mares from multiple separate lineages
contributing to the modern genetic pool. Levine (2006)
has suggested that as the original domesticated population
expanded, horses from wild populations were introduced
into the domestic herds over time.
China has rich horse genetic resources; the number of
Chinese domestic horses was 7.639 million in 2004 (Statistic
report in China, 2004). Horses are widespread throughout
14 provinces of North-west, South-west and North-east
China, in addition to Hubei province of Central China. Fifteen
native domestic horse breeds remaining as several separate
populations are recorded in China today (Hendricks 1995).
Based on the origin, habitats and body size, the 15 Chinese
breeds can be grouped into five horse types: Mongolian,
South-western, Hequ, Kazahk and Tibetan (Xie 1987). In
addition, China has pony or small horse populations dis-
tributed mainly in the Shaanxi, Sichuan, Guangxi, Guizhou
and Yunnan provinces. In China, an adult Equus caballus
individual with a height at the withers less than or equal to
106 cm is classified as a pony, while the one with a height
greater than 106 cm is called a common horse (Xie 1987).
Despite the presence of diverse native horse breeds and
populations, research on equine genetics is not well devel-
oped in China and only a limited number of reports on the
mtDNA diversity of Chinese domestic horses are available.
Previously, mtDNA of three short horses and three common
horses in Yunnan was digested by 20 restriction endonuc-
leases and results indicated high genetic diversity and
multiple maternal origins (Wang et al. 1994). mtDNA
D-loop sequences have also been analysed for four Mongo-
lian individuals, seven Tibetan individuals and 10 Guan-
shan individuals (Mang et al. 2005; McGahern et al. 2006a;
Xu et al. 2007). Furthermore, a 262-bp mtDNA D-loop
sequence from nine Chinese ancient horse remains indicates
multiple maternal origins (Cai et al. 2007a). Modern and
ancient mtDNA sequence analysis has provided critical
evidence on the origins and dispersal routes of horse, pig,
cattle, goat and sheep (Jung et al. 2002; Anderung et al.
2005; Keyser-Tracqui et al. 2005; Larson et al. 2005,
2007a,b; Beja-Pereira et al. 2006; Fernandez et al. 2006;
McGahern et al. 2006b; Cai et al. 2007b). In this study,
mtDNA D-loop sequences of 182 animals (48 from Gen-
Bank) from nine modern Chinese domestic horse breeds and
of nine previously published ancient horse specimens were
analysed to cast new light on the origin and genetic diver-
sity of domestic horses in China.
Materials and methods
Specimen collection and DNA extraction
All 134 blood samples representing five local domestic horse
breeds were collected in China. The genomic DNA was
extracted from blood by a standard phenol–chloroform
method. In addition, 48 mtDNA D-loop sequences from
five Chinese local domestic horse breeds were available from
GenBank (AF014413–AF014417, AF056071, DQ297622–
DQ297633, DQ297634–DQ297635, DQ297637–DQ297638,
DQ327838–327847, DQ986464–DQ986479) and nine mtDNA
D-loop sequences from Bronze Age Chinese horse remains were
also obtained from GenBank (DQ900922–DQ900930). The
distributions of all nine Chinese domestic horse breeds and the
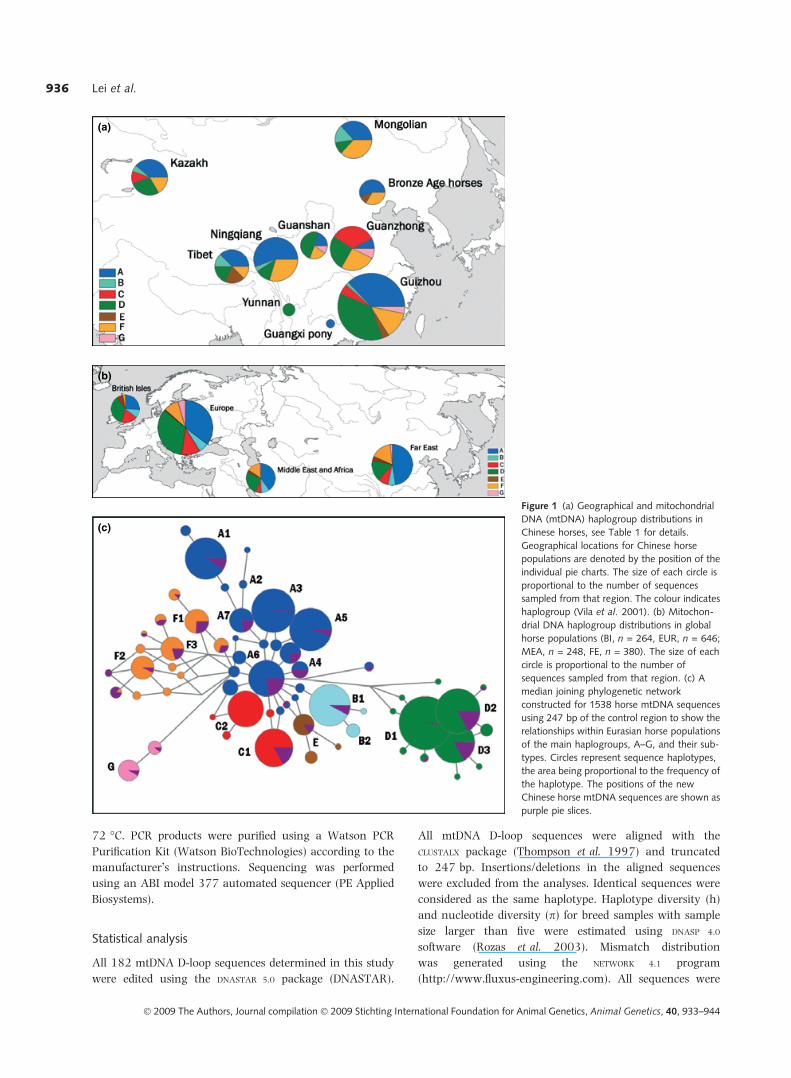
sites of the ancient horse remains are shown in Fig. 1a.
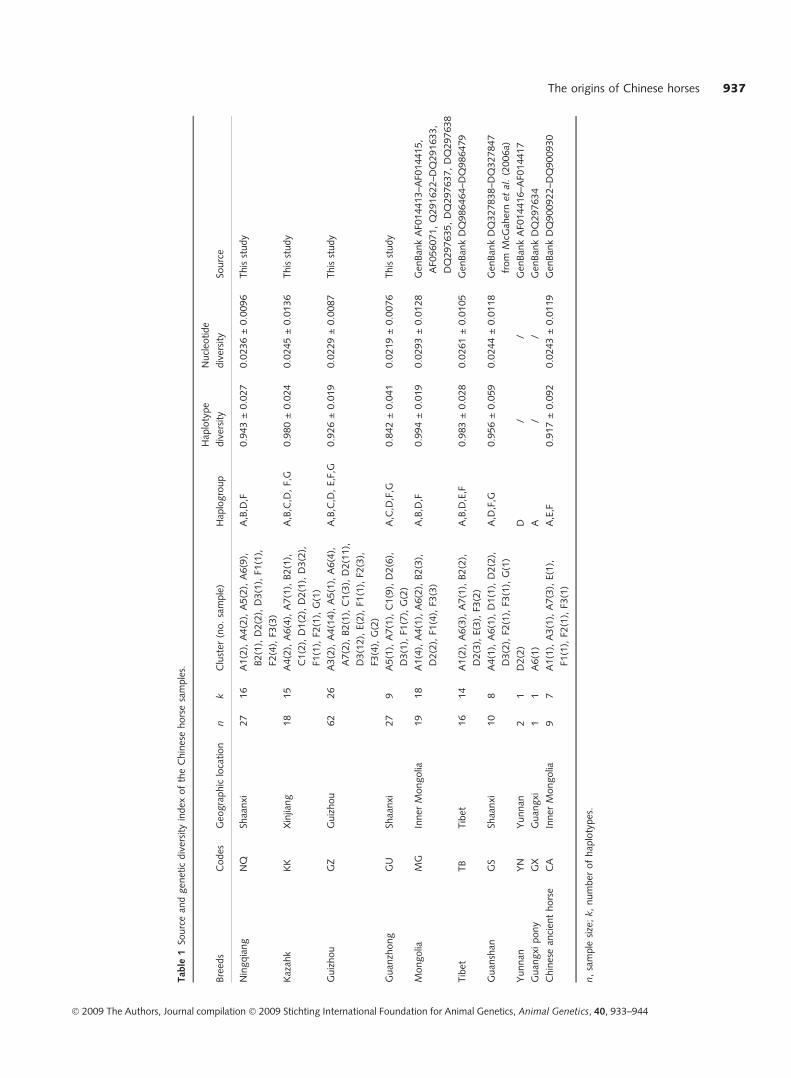
Geographical locations and number of horse samples are shown
in Table 1.
PCR amplification and sequencing
To amplify the 247-bp hypervariable region I sequence of
the equine mtDNA D-loop region (McGahern et al. 2006a)
between positions 15 494 and 15 740 of reference
sequence X79547 (Xu & Arnason 1994), a pair of primers
were used according to the published sequences, HA:
5¢-AGTCTCACCATCAACACCCAAAGC-3¢ and HB: 5¢-CCT
GAAGTAGGAACCAGATG-3¢ (Ivankovic et al. 2002). PCR
amplifications were conducted in a 50-ll volume containing
5 ll of 10· buffer, 1.5 mM MgCl2, 0.25 mM dNTPs, 0.2 lM
each primer, 1.5 U Taq DNA polymerase (TaKaRa Biosys-
tems) and approximately 50 ng genomic DNA. The PCR
was carried out using a standard program with 4 min
denaturation at 95 �C, 35 cycles for 30 s at 94 �C, 60 s at
55 �C, and 90 s at 72 �C, and final extension for 10 min at
� 2009 The Authors, Journal compilation � 2009 Stichting International Foundation for Animal Genetics, Animal Genetics, 40, 933–944
The origins of Chinese horses 935
72 �C. PCR products were purified using a Watson PCR
Purification Kit (Watson BioTechnologies) according to the
manufacturer�s instructions. Sequencing was performed
using an ABI model 377 automated sequencer (PE Applied
Biosystems).
Statistical analysis
All 182 mtDNA D-loop sequences determined in this study
were edited using the DNASTAR 5.0 package (DNASTAR).
All mtDNA D-loop sequences were aligned with the
CLUSTALX package (Thompson et al. 1997) and truncated
to 247 bp. Insertions/deletions in the aligned sequences
were excluded from the analyses. Identical sequences were
considered as the same haplotype. Haplotype diversity (h)
and nucleotide diversity (p) for breed samples with sample
size larger than five were estimated using DNASP 4.0
software (Rozas et al. 2003). Mismatch distribution
was generated using the NETWORK 4.1 program
(http://www.fluxus-engineering.com). All sequences were
(a)
(b)
(c)
Figure 1 (a) Geographical and mitochondrial
DNA (mtDNA) haplogroup distributions in
Chinese horses, see Table 1 for details.
Geographical locations for Chinese horse
populations are denoted by the position of the
individual pie charts. The size of each circle is
proportional to the number of sequences
sampled from that region. The colour indicates
haplogroup (Vila et al. 2001). (b) Mitochon-
drial DNA haplogroup distributions in global
horse populations (BI, n = 264, EUR, n = 646;
MEA, n = 248, FE, n = 380). The size of each
circle is proportional to the number of
sequences sampled from that region. (c) A
median joining phylogenetic network
constructed for 1538 horse mtDNA sequences
using 247 bp of the control region to show the
relationships within Eurasian horse populations
of the main haplogroups, A–G, and their sub-
types. Circles represent sequence haplotypes,
the area being proportional to the frequency of
the haplotype. The positions of the new
Chinese horse mtDNA sequences are shown as
purple pie slices.
� 2009 The Authors, Journal compilation � 2009 Stichting International Foundation for Animal Genetics, Animal Genetics, 40, 933–944
Lei et al.936
Tab
le1
Sourc
ean
dgen
etic
div
ersi
tyin
dex
of
the
Chin
ese
hors
esa
mple
s.
Bre
eds
Codes
Geo
gra
phic
loca
tion
nk
Clu
ster
(no.
sam
ple
)H
aplo
gro
up
Hap
loty
pe
div
ersi
ty
Nucl
eotide
div
ersi
tySo
urc
e
Nin
gqia
ng
NQ
Shaa
nxi
27
16
A1(2
),A
4(2
),A
5(2
),A
6(9
),
B2(1
),D
2(2
),D
3(1
),F1
(1),
F2(4
),F3
(3)
A,B
,D,F
0.9
43
±0.0
27
0.0
236
±0.0
096
This
study
Kaz
ahk
KK
Xin
jiang
18
15
A4(2
),A
6(4
),A
7(1
),B2(1
),
C1(2
),D
1(2
),D
2(1
),D
3(2
),
F1(1
),F2
(1),
G(1
)
A,B
,C,D
,F,
G0.9
80
±0.0
24
0.0
245
±0.0
136
This
study
Guiz
hou
GZ
Guiz
hou
62
26
A3(2
),A
4(1
4),
A5(1
),A
6(4
),
A7(2
),B2(1
),C
1(3
),D
2(1
1),
D3(1
2),
E(2),
F1(1
),F2
(3),
F3(4
),G
(2)
A,B
,C,D
,E,
F,G
0.9
26
±0.0
19
0.0
229
±0.0
087
This
study
Guan
zhong
GU
Shaa
nxi
27
9A
5(1
),A
7(1
),C
1(9
),D
2(6
),
D3(1
),F1
(7),
G(2
)
A,C
,D,F
,G0.8
42
±0.0
41
0.0
219
±0.0
076
This
study
Mongolia
MG
Inner
Mongolia
19
18
A1(4
),A
4(1
),A
6(2
),B2(3
),
D2(2
),F1
(4),
F3(3
)
A,B
,D,F
0.9
94
±0.0
19
0.0
293
±0.0
128
Gen
Ban
kA
F014413–A
F014415,
AF0
56071,
Q291622–D
Q291633,
DQ
297635,
DQ
297637,
DQ
297638
Tib
etTB
Tib
et16
14
A1(2
),A
6(3
),A
7(1
),B2(2
),
D2(3
),E(
3),
F3(2
)
A,B
,D,E
,F0.9
83
±0.0
28
0.0
261
±0.0
105
Gen
Ban
kD
Q986464–D
Q986479
Guan
shan
GS
Shaa
nxi
10
8A
4(1
),A
6(1
),D
1(1
),D
2(2
),
D3(2
),F2
(1),
F3(1
),G
(1)
A,D
,F,G
0.9
56
±0.0
59
0.0
244
±0.0
118
Gen
Ban
kD
Q327838–D
Q327847
from
McG
aher
net
al.
(2006a)
Yunnan
YN
Yunnan
21
D2(2
)D
//
Gen
Ban
kA
F014416–A
F014417
Guan
gxi
pony
GX
Guan
gxi
11
A6(1
)A
//
Gen
Ban
kD
Q297634
Chin
ese
anci
ent
hors
eC
AIn
ner
Mongolia
97
A1(1
),A
3(1
),A
7(3
),E(
1),
F1(1
),F2
(1),
F3(1
)
A,E
,F0.9
17
±0.0
92
0.0
243
±0.0
119
Gen
Ban
kD
Q900922–D
Q900930
n,
sam
ple
size
;k,
num
ber
of
hap
loty
pes
.
� 2009 The Authors, Journal compilation � 2009 Stichting International Foundation for Animal Genetics, Animal Genetics, 40, 933–944
The origins of Chinese horses 937

deposited in Genbank (accession nos AY136785–
AY136786, EU203727–EU203858).
In addition, the 247-bp region was examined in a total
dataset of 1538 horses (the 134 new sequences recorded
here and 1404 GenBank accessions) sampled from 119
populations from three broad geographical regions: Europe
(EUR), Middle East and Africa (MEA), and the Far East (FE).
Diversity values were estimated using the ARLEQUIN program
(Schneider et al. 2000) and an analysis of molecular vari-
ance (AMOVA) was performed to investigate the partitioning
of mtDNA molecular variance among these three geo-
graphical groups. To test the concordance of haplogroup
F with Far Eastern populations, Fisher�s exact test of inde-
pendence (Sokal & Rohlf 1995) was performed in a 2 · 2
contingency table featuring EUR/MEA populations vs. FE
populations, with counts of haplogroup F vs. non-haplo-
group F sequences. Median joining networks (Bandelt
et al.1999) of the total dataset and subsets of the data
according to the geographical regions defined above were
drawn in NETWORK 4.1. Star contraction pre-processing was
performed (Forster et al.2001). Parameters and down
weighting of hypervariable sites were according to Jansen
et al. (2002). Maximum parsimony calculations were per-
formed on the output networks (Polzin & Daneschmand
2003). Haplotype data were plotted geographically using
ARCGIS 9 (Esri).
Results
Sequence variation
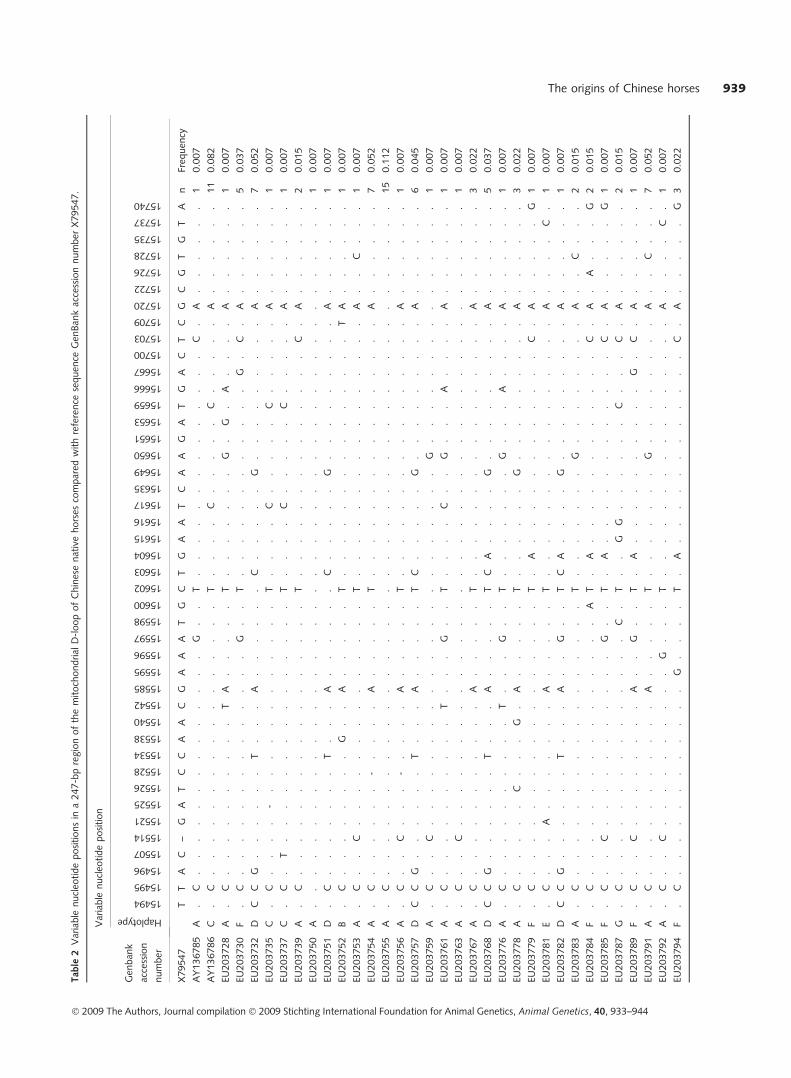
A 247-bp equine mtDNA D-loop region sequence was
examined in 182 Chinese domestic horses. Seventy mtDNA
haplotypes were identified and defined by 47 polymorphic
nucleotide sites: 43 transitions, two transversions and two
coexistence sites of transition and transversion, suggesting
high polymorphisms and a strong bias towards transitions
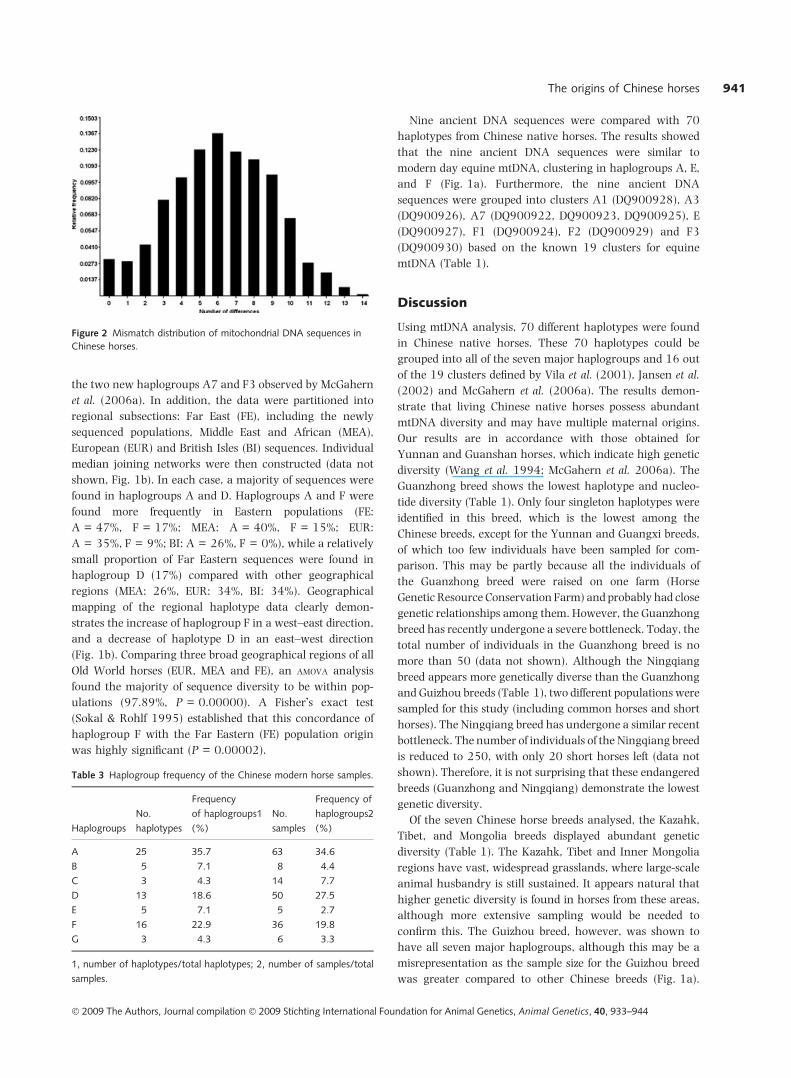
(Table 2). The mismatch analysis performed for Chinese
horse breeds (Fig. 2) shows a monomodal distribution with
a major peak at six mutational differences. Sequence dif-
ferences ranged from one to 14 base pairs. Among the 70
haplotypes, 27 haplotypes (38.6%) were detected more than
once and were thus shared by individuals among breeds or
within breeds. The largest haplotype consisted of 18 indi-
viduals, with another two haplotypes containing 15 and 12
individuals respectively, and 23 haplotypes possessing two
to nine individuals each. Forty-three haplotypes (61.4%)
were singletons: 10 (23.3%) from the Guizhou breed, nine
(20.9%) from the Mongolia breed, eight (18.6%) from the
Kazahk breed, seven (16.3%) from the Tibetan breed, four
(9.3%) from the Guanzhong breed, three (7.0%) from the
Ningqiang breed and two (4.6%) from the Guanshan breed.
The number of haplotypes identified in each breed (except
the Yunnan and Guangxi breeds) ranged from eight to 26,
and haplotype diversity values ranged from 0.842 ± 0.041
in the Guanzhong breed to 0.994 ± 0.019 in the Mongolia
breed. Similarly, the Mongolia breed showed the highest
nucleotide diversity value (0.02926 ± 0.01282), whereas
the Guanzhong breed displayed the lowest nucleotide
diversity value (0.02194 ± 0.00763), indicating abundant
genetic diversity in Chinese domestic horse breeds (Table 1).
The Chinese breed nucleotide diversity values compare
favourably with those estimated from the broad geo-
graphical regions (EUR = 0.029123 ± 0.01511; MEA =
0.02576 ± 0.01356; FE = 0.02594 ± 0.01363).
Nine ancient DNA sequences of 247 bp of the D-loop
region cited from GenBank carried seven haplotypes. Three
(DQ900922, DQ900923, DQ900925) out of these nine
ancient sequences shared the same haplotype. Interestingly,
one modern haplotype was identical to the ancient sequence
DQ900929, one to DQ900924, and another to DQ900927.
The nine ancient sequences were differentiated by a total of
17 polymorphic nucleotide sites, all of which were transi-
tions. These nine ancient DNA sequences were shown to
display high level of haplotype diversity (0.917 ± 0.092)
and nucleotide diversity (0.0243 ± 0.0119) (Table 1).
Phylogenetic analysis
Previous research grouped equine mtDNA haplotypes into
seven major haplogroups (A–G) (Vila et al. 2001; Jansen
et al. 2002) and 19 clusters (Jansen et al. 2002; McGahern
et al. 2006a). mtDNA haplotypes in Chinese domestic
horses demonstrated the presence of these seven major
haplogroups (A–G) that included 16 out of the 19 clusters,
except clusters A2, B1 and C2 (Fig. 1a,b and Table 1). A, D
and F were the predominant haplogroups, comprising
35.7%, 18.6% and 22.9% of the 70 haplotypes respectively,
whereas haplogroups B, C, E and G comprised 7.1%, 4.3%,
7.1% and 4.3% respectively. Furthermore, among the 182
individuals, haplogroups A–G showed similar distribution
patterns (Table 3). The results indicated that the majority
of sequences in Chinese domestic horse breeds belong to
haplogroups A, D and F. Different horse breeds displayed
distinctive profiles of the previously reported seven major
haplogroups A–G and only 16 of the 17 clusters (Vila et al.
2001; Jansen et al. 2002) (Table 1). At least four major
haplogroups and seven clusters were observed in each of the
seven Chinese domestic horse breeds. All of the seven major
haplogroups in addition to 14 clusters were observed in the
Guizhou breed (Table 1). These data are consistent with
multiple maternal origins in living native Chinese domestic
horses.
Mitochondrial data from the living Chinese horse popu-
lations were grouped with 1404 previously published
sequences. A median-joining network of the complete
dataset (1538 horses) was drawn to determine the rela-
tionships of the seven main haplogroups, A–G (Fig. 1c,
purple pie slices represent the relative positions of the new
Chinese sequences). The network confirms the position of
� 2009 The Authors, Journal compilation � 2009 Stichting International Foundation for Animal Genetics, Animal Genetics, 40, 933–944
Lei et al.938
Tab
le2
Var
iable
nucl
eotide
posi
tions
ina
247-b
pre
gio
nof
the
mitoch
ondrial
D-l
oop
of
Chin
ese
nat
ive
hors
esco
mpar
edw
ith
refe
rence
sequen
ceG
enBan
kac
cess
ion
num
ber
X79547.
Gen
ban
k
acce
ssio
n
num
ber
Var
iable
nucl
eotide
posi
tion
Haplotype
15494
15495
15496
15507
15514
15521
15525
15526
15528
15534
15538
15540
15542
15585
15595
15596
15597
15598
15600
15602
15603
15604
15615
15616
15617
15635
15649
15650
15651
15653
15659
15666
15667
15700
15703
15709
15720
15722
15726
15728
15735
15737
15740
X79547
TT
AC
–G
AT
CC
AA
CG
AA
AT
GC
TG
AA
TC
AA
GA
TG
AC
TC
GC
GT
GT
An
Freq
uen
cy
AY
136785
A.
C.
..
..
..
..
..
..
.G
..
T.
..
..
..
..
..
..
.C
.A
..
..
..
10.0
07
AY
136786
C.
C.
..
..
..
..
..
..
..
..
T.
..
.C
..
..
.C
..
..
.A
..
..
..
11
0.0
82
EU203728
A.
C.
..
..
..
..
.T
A.
..
..
T.
..
..
..
G.
G.
A.
..
.A
..
..
..
10.0
07
EU203730
F.
C.
..
..
..
..
..
..
.G
..
T.
..
..
..
..
..
.G
.C
.A
..
..
..
50.0
37
EU203732
DC
CG
..
..
..
T.
..
A.
..
..
.C
..
..
.G
..
..
..
..
.A
..
..
..
70.0
52
EU203735
C.
C.
..
.-
..
..
..
..
..
..
T.
..
.C
..
..
.C
..
..
.A
..
..
..
10.0
07
EU203737
C.
C.
T.
..
..
..
..
..
..
..
T.
..
.C
..
..
.C
..
..
.A
..
..
..
10.0
07
EU203739
A.
C.
..
..
..
..
..
..
..
..
T.
..
..
..
..
..
..
.C
.A
..
..
..
20.0
15
EU203750
A.
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
10.0
07
EU203751
D.
C.
..
..
..
T.
..
A.
..
..
.C
..
..
.G
..
..
..
..
.A
..
..
..
10.0
07
EU203752
B.
C.
..
..
..
.G
..
A.
..
..
T.
..
..
..
..
..
..
..
TA
..
..
..
10.0
07
EU203753
A.
C.
.C
..
..
..
..
..
..
..
T.
..
..
..
..
..
..
..
.A
..
C.
..
10.0
07
EU203754
A.
C.
..
..
.-
..
..
A.
..
..
T.
..
..
..
..
..
..
..
.A
..
..
..
70.0
52
EU203755
A.
C.
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
15
0.1
12
EU203756
A.
C.
.C
..
.-
..
..
A.
..
..
T.
..
..
..
..
..
..
..
.A
..
..
..
10.0
07
EU203757
DC
CG
..
..
..
T.
..
A.
..
..
TC
..
..
.G
..
..
..
..
.A
..
..
..
60.0
45
EU203759
A.
C.
.C
..
..
..
..
..
..
..
..
..
..
..
G.
..
..
..
..
..
..
..
10.0
07
EU203761
A.
C.
..
..
..
..
.T
..
.G
..
T.
..
.C
..
G.
..
A.
..
.A
..
..
..
10.0
07
EU203763
A.
C.
.C
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
10.0
07
EU203767
A.
C.
..
..
..
..
..
A.
..
..
T.
..
..
..
..
..
..
..
.A
..
..
..
30.0
22
EU203768
DC
CG
..
..
..
T.
..
A.
..
..
TC
A.
..
.G
..
..
..
..
.A
..
..
..
50.0
37
EU203776
A.
C.
..
..
..
..
.T
..
.G
..
T.
..
..
..
G.
..
A.
..
.A
..
..
..
10.0
07
EU203778
A.
C.
..
..
C.
..
G.
A.
..
..
T.
..
..
.G
..
..
..
..
.A
..
..
..
30.0
22
EU203779
F.
C.
..
..
..
..
..
..
..
..
T.
A.
..
..
..
..
..
.C
.A
..
..
.G
10.0
07
EU203781
E.
C.
..
A.
..
..
..
A.
..
..
T.
..
..
..
..
..
..
..
.A
..
..
C.
10.0
07
EU203782
DC
CG
..
..
..
T.
..
A.
.G
..
TC
A.
..
.G
..
..
..
..
.A
..
..
..
10.0
07
EU203783
A.
C.
..
..
..
..
..
..
..
..
T.
..
..
..
G.
..
..
..
.A
..
C.
..
20.0
15
EU203784
F.
C.
..
..
..
..
..
..
..
.A
T.
A.
..
..
..
..
..
.C
.A
.A
..
.G
20.0
15
EU203785
F.
C.
.C
..
..
..
..
..
.G
..
T.
A.
..
..
..
..
..
.C
.A
..
..
.G
10.0
07
EU203787
G.
C.
..
..
..
..
..
..
..
C.
T.
.G
G.
..
..
.C
..
.C
.A
..
..
..
20.0
15
EU203789
F.
C.
.C
..
..
..
..
A.
.G
..
T.
A.
..
..
..
..
.G
.C
.A
..
..
..
10.0
07
EU203791
A.
C.
..
..
..
..
..
A.
..
..
T.
..
..
..
G.
..
..
..
.A
..
C.
..
70.0
52
EU203792
A.
C.
.C
..
..
..
..
..
G.
..
T.
..
..
..
..
..
..
..
.A
..
..
C.
10.0
07
EU203794
F.
C.
..
..
..
..
..
.G
..
..
T.
A.
..
..
..
..
..
.C
.A
..
..
.G
30.0
22
� 2009 The Authors, Journal compilation � 2009 Stichting International Foundation for Animal Genetics, Animal Genetics, 40, 933–944
The origins of Chinese horses 939

Tab
le2
Continued
.
Gen
ban
k
acce
ssio
n
num
ber
Var
iable
nucl
eotide
posi
tion
Haplotype
15494
15495
15496
15507
15514
15521
15525
15526
15528
15534
15538
15540
15542
15585
15595
15596
15597
15598
15600
15602
15603
15604
15615
15616
15617
15635
15649
15650
15651
15653
15659
15666
15667
15700
15703
15709
15720
15722
15726
15728
15735
15737
15740
EU203795
AA
C.
..
..
..
..
..
A.
..
..
..
..
..
..
G.
..
..
..
.A
..
..
..
10.0
07
EU203797
C.
C.
.C
..
..
..
..
..
..
..
T.
..
.C
..
..
.C
..
..
.A
..
..
..
10.0
07
EU203798
A.
C.
..
..
..
..
..
..
..
.A
..
..
..
..
..
..
..
..
..
..
..
..
30.0
22
EU203804
A.
C.
..
..
..
..
..
A.
..
..
..
..
..
..
G.
..
A.
..
.A
..
..
..
10.0
07
EU203805
DC
CG
..
..
..
T.
..
..
..
..
TC
A.
..
.G
..
..
..
.C
.A
..
..
..
10.0
07
EU203813
F.
C.
.C
..
..
..
..
..
..
.A
T.
A.
..
..
..
..
..
.C
.A
.A
..
.G
10.0
07
EU203814
G.
C.
..
..
..
..
..
..
..
C.
T.
..
G.
..
..
..
..
.C
.A
..
..
..
10.0
07
EU203817
A.
C.
..
..
..
..
.T
A.
..
..
T.
..
..
..
G.
..
A.
..
.A
..
..
..
10.0
07
EU203818
A.
C.
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
.C
..
..
..
..
10.0
07
EU203820
DC
CG
..
..
.T
T.
..
..
..
..
TC
..
..
.G
..
..
..
..
.A
..
..
..
10.0
07
EU203821
F.
C.
..
..
..
..
..
..
..
..
T.
A.
..
..
..
.C
..
.C
.A
.A
..
.G
50.0
37
EU203822
B.
C.
..
..
..
.G
..
A.
..
.A
T.
..
..
..
..
..
..
..
TA
..
..
..
10.0
07
EU203823
A.
C.
..
..
..
..
..
..
..
..
T.
..
..
..
GA
..
..
..
.A
..
..
..
10.0
07
EU203824
F.
C.
..
..
..
..
..
..
.G
..
T.
A.
..
T.
..
..
.G
TC
.A
..
.T
..
10.0
07
EU203825
DC
CG
..
A.
..
T.
..
A.
..
..
TC
A.
..
.G
..
..
..
..
.A
..
..
..
10.0
07
EU203826
A.
C.
..
..
..
..
..
..
..
..
T.
..
..
..
G.
..
..
..
.A
..
..
..
20.0
15
EU203827
DC
CG
..
..
..
T.
..
..
..
..
TC
..
..
.G
..
..
..
..
.A
..
..
..
10.0
07
EU203829
E.
C.
..
A.
..
..
..
..
.G
..
T.
..
..
..
G.
..
..
..
.A
..
..
..
20.0
15
EU203841
A.
C.
..
..
..
..
..
..
..
..
T.
..
..
..
G.
..
..
..
AA
A.
..
..
10.0
07
EU203842
A.
C.
..
..
..
..
.T
..
.G
..
T.
..
..
T.
G.
..
A.
.C
.A
..
..
..
20.0
15
EU203846
A.
C.
..
..
..
..
..
..
..
..
T.
..
..
..
..
..
..
..
.A
..
C.
..
20.0
15
EU203848
E.
C.
..
A.
..
..
..
..
.G
..
T.
A.
..
..
..
..
..
.C
.A
..
..
.G
10.0
07
EU203851
F.
C.
..
..
..
..
..
..
.G
..
T.
A.
..
..
..
..
.G
.C
.A
..
..
..
10.0
07
EU203852
A.
C.
..
..
..
..
..
..
.G
..
T.
..
..
..
G.
..
..
..
.A
..
..
..
10.0
07
EU203856
F.
C.
..
..
..
..
..
..
.G
..
T.
A.
..
..
..
..
..
.C
.A
..
..
.G
10.0
07
� 2009 The Authors, Journal compilation � 2009 Stichting International Foundation for Animal Genetics, Animal Genetics, 40, 933–944
Lei et al.940
the two new haplogroups A7 and F3 observed by McGahern
et al. (2006a). In addition, the data were partitioned into
regional subsections: Far East (FE), including the newly
sequenced populations, Middle East and African (MEA),
European (EUR) and British Isles (BI) sequences. Individual
median joining networks were then constructed (data not
shown, Fig. 1b). In each case, a majority of sequences were
found in haplogroups A and D. Haplogroups A and F were
found more frequently in Eastern populations (FE:
A = 47%, F = 17%; MEA: A = 40%, F = 15%; EUR:
A = 35%, F = 9%; BI: A = 26%, F = 0%), while a relatively
small proportion of Far Eastern sequences were found in
haplogroup D (17%) compared with other geographical
regions (MEA: 26%, EUR: 34%, BI: 34%). Geographical
mapping of the regional haplotype data clearly demon-
strates the increase of haplogroup F in a west–east direction,
and a decrease of haplotype D in an east–west direction
(Fig. 1b). Comparing three broad geographical regions of all
Old World horses (EUR, MEA and FE), an AMOVA analysis
found the majority of sequence diversity to be within pop-
ulations (97.89%, P = 0.00000). A Fisher�s exact test
(Sokal & Rohlf 1995) established that this concordance of
haplogroup F with the Far Eastern (FE) population origin
was highly significant (P = 0.00002).
Nine ancient DNA sequences were compared with 70
haplotypes from Chinese native horses. The results showed
that the nine ancient DNA sequences were similar to
modern day equine mtDNA, clustering in haplogroups A, E,
and F (Fig. 1a). Furthermore, the nine ancient DNA
sequences were grouped into clusters A1 (DQ900928), A3
(DQ900926), A7 (DQ900922, DQ900923, DQ900925), E
(DQ900927), F1 (DQ900924), F2 (DQ900929) and F3
(DQ900930) based on the known 19 clusters for equine
mtDNA (Table 1).
Discussion
Using mtDNA analysis, 70 different haplotypes were found
in Chinese native horses. These 70 haplotypes could be
grouped into all of the seven major haplogroups and 16 out
of the 19 clusters defined by Vila et al. (2001), Jansen et al.
(2002) and McGahern et al. (2006a). The results demon-
strate that living Chinese native horses possess abundant
mtDNA diversity and may have multiple maternal origins.
Our results are in accordance with those obtained for
Yunnan and Guanshan horses, which indicate high genetic
diversity (Wang et al. 1994; McGahern et al. 2006a). The
Guanzhong breed shows the lowest haplotype and nucleo-
tide diversity (Table 1). Only four singleton haplotypes were
identified in this breed, which is the lowest among the
Chinese breeds, except for the Yunnan and Guangxi breeds,
of which too few individuals have been sampled for com-
parison. This may be partly because all the individuals of
the Guanzhong breed were raised on one farm (Horse
Genetic Resource Conservation Farm) and probably had close
genetic relationships among them. However, the Guanzhong
breed has recently undergone a severe bottleneck. Today, the
total number of individuals in the Guanzhong breed is no
more than 50 (data not shown). Although the Ningqiang
breed appears more genetically diverse than the Guanzhong
and Guizhou breeds (Table 1), two different populations were
sampled for this study (including common horses and short
horses). The Ningqiang breed has undergone a similar recent
bottleneck. The number of individuals of the Ningqiang breed
is reduced to 250, with only 20 short horses left (data not
shown). Therefore, it is not surprising that these endangered
breeds (Guanzhong and Ningqiang) demonstrate the lowest
genetic diversity.
Of the seven Chinese horse breeds analysed, the Kazahk,
Tibet, and Mongolia breeds displayed abundant genetic
diversity (Table 1). The Kazahk, Tibet and Inner Mongolia
regions have vast, widespread grasslands, where large-scale
animal husbandry is still sustained. It appears natural that
higher genetic diversity is found in horses from these areas,
although more extensive sampling would be needed to
confirm this. The Guizhou breed, however, was shown to
have all seven major haplogroups, although this may be a
misrepresentation as the sample size for the Guizhou breed
was greater compared to other Chinese breeds (Fig. 1a).
Table 3 Haplogroup frequency of the Chinese modern horse samples.
Haplogroups
No.
haplotypes
Frequency
of haplogroups1
(%)
No.
samples
Frequency of
haplogroups2
(%)
A 25 35.7 63 34.6
B 5 7.1 8 4.4
C 3 4.3 14 7.7
D 13 18.6 50 27.5
E 5 7.1 5 2.7
F 16 22.9 36 19.8
G 3 4.3 6 3.3
1, number of haplotypes/total haplotypes; 2, number of samples/total
samples.
Figure 2 Mismatch distribution of mitochondrial DNA sequences in
Chinese horses.
� 2009 The Authors, Journal compilation � 2009 Stichting International Foundation for Animal Genetics, Animal Genetics, 40, 933–944
The origins of Chinese horses 941

Other native horse breeds in China might have similar
genetic diversity levels if sufficient samples were sequenced.
McGahern et al. (2006a), in their study of 962 horses
from 72 populations, revealed a significant biogeographical
association of eastern equine mtDNA types with haplogroup
F and a significant association of European equine mtDNA
types with haplogroup D. Using an increased dataset of
1538 horses from 119 populations, we confirmed that
haplotype F occurred more frequently in Eastern popula-
tions (17%) when compared with other Eurasian popula-
tions (MEA 15%, EUR 9%, BI 0%), whereas a relatively
small proportion of Far Eastern sequences were found in
haplogroup D (17%) compared with other geographical
regions (MEA 26%, EUR 34%, BI 34%). This biogeograph-
ical association of Far Eastern equine mtDNA types with
haplogroup F was highly significant (P = 0.00002), sug-
gesting that there has been a degree of genetic isolation and
differentiation of eastern and western horse populations and
lending support to the idea that haplogroup F may have
been taken into the domestic gene pool in the Far East.
Based on our data, we cannot exclude the possibility that
an early horse domestication event may have taken place
independently in China. Neither can we conclusively dem-
onstrate that the post-domestication inclusion of native
Chinese wild mares into the domestic horse breeding pop-
ulation is responsible for the patterns we observed in our
data. It is probable that multiple processes, which we may
never disentangle, have resulted in the observed modern
haplotype descriptions. For example, it is likely that the
constant procurement of horses from many regions by the
imperial court and the high mobility of horses and their
military utilization in past millennia, as described above,
resulted in considerable mixing of horse populations (Xie
1987), including input from native Chinese breeds. This
could result in the abundant genetic diversity we observed
in our data.
We compared our living native horse genetic data with
sequences from ancient horses dating to the Bronze Age
(c. 4000–2000a BP) from the Chifeng archaeological sites in
Inner Mongolia, northeast China (Cai et al. 2007a).
The majority of ancient samples (five) were shown to
possess haplogroup A, whereas haplogroup E was found in
one specimen (Table 1). Three ancient samples were char-
acterized by mtDNA haplogroup F, which was strongly
associated with Eastern breeds (McGahern et al. 2006a).
This demonstrates that haplotypes A, E and F were present
in horse populations in China between 4000 and 2000 BP.
However, without additional analyses (e.g. stable isotope
analysis), it is not possible to state whether the Bronze Age
horses were Chinese native horses, or whether they had
been imported from a distance.
Unlike the situation for many other large domesticated
animal species that demonstrate remarkably clear geo-
graphical patterning (MacHugh & Bradley 2001), caution
must be used when inferring population structure from
equine mtDNA data. Discrete horse populations share
mtDNA sequence with other populations despite geo-
graphical distance (McGahern et al. 2006b). For example, in
the Chifeng Bronze Age horses, the most common sequence
haplogroup (five out of nine individuals) was also the most
common globally, with 38% out of 1538 sequences repre-
sented in haplogroup A. The other two haplotypes repre-
sented in the Bronze Age data are of interest, as they are
more rare, with haplotype F occurring at 11% of the total
global population sequenced to date, and more significantly
with haplotype E, occurring at only 3% in living horse
populations. The likelihood of randomly sampling haplotype
E from a wild population is extremely low; therefore, it is
probable that haplotype E was more commonly occurring in
past populations.
Conclusions
Archaeological and osteological evidence has, thus far,
pointed to Central Asia as the region where the earliest
horse domestication took place, but genetic data, including
those in this study, show that the domestication of horses
was not a single discrete event. Multiple horse domestica-
tion events (i.e. sampling and re-sampling of wild horse
populations) may have occurred later in other regions
including China. Far Eastern domestic horses, like those in
Eurasia, originated from a diverse founder population,
sampled during multiple domestication events and/or via
trade and exchange over short and/or long geographical
distances, or else resulting from repeated post-domestication
resampling of native wild mare populations. Our data
demonstrate that much of this genetic diversity has been
preserved in domestic native horse populations across Chi-
na, particularly in the Inner Mongolian and south-western
regions. This finding is consistent with the scenario attested
in the archaeological and historical records that horses were
continuously brought into dynastic centres from the
northern steppes and south-western China. It is hoped that
further archaeogenetic study of ancient horse remains
within a multidisciplinary approach (including archaeozo-
ology and bioarchaeology) will provide more insights into
the origins and history of domestic horses in China.
Acknowledgements
This work is supported by Programme for New Century
Excellent Talents in University of 2007 (NCET-07-0699),
Research Program of Northwest A&F University (07ZR004).
MAB is supported by the Leverhulme Trust and the Isaac
Newton Trust; CJE is supported by the Irish Research
Council for Science Engineering and Technology (Basic
Research Grant Scheme project number SC/202/510).
Special thanks to Dr E.W. Hill (University College Dublin,
Ireland), Dr Richard Meadow (Harvard University, USA)
and Dr W.-S. Liu (The Pennsylvania State University, USA)
� 2009 The Authors, Journal compilation � 2009 Stichting International Foundation for Animal Genetics, Animal Genetics, 40, 933–944
Lei et al.942

for their valuable suggestions and English editing, which
improved the manuscript.
References
Anderung C., Bouwman A., Persson P., Carretero J.M., Ortega A.I.,
Elburg R., Smith C., Arsuaga J.L., Ellegren H. & Gotherstrom A.
(2005) Prehistoric contacts over the Straits of Gibraltar indicated
by genetic analysis of Iberian Bronze Age cattle. Proceedings of the
National Academy of Sciences of the United States of America 102,
8431–5.
Bandelt H.J., Forster P. & Rohl A. (1999) Median-joining networks
for inferring intraspecific phylogenies. Molecular Biology and
Evolution 16, 37–48.
Beja-Pereira A., Caramelli D., Lalueza-Fox C. et al. (2006) The
origin of European cattle: evidence from modern and ancient
DNA. Proceedings of the National Academy of Sciences of the United
States of America 103, 8113–8.
Cai D.W., Han L., Xie C.Z., Li S.N., Zhou H. & Zhu H. (2007a)
Mitochondrial DNA analysis of Bronge age horses recovered from
Chifeng region, Inner Mongolia, China. Progress in Natural Science
17, 544–50.
Cai D.W., Han L., Zhang X.L., Zhou H. & Zhu H. (2007b) DNA
analysis of archaeological sheep remains from China. Journal of
Archaeoloical Science 34, 1347–55.
Chiou-Peng T.H. (2004) Horsemen in the Dian Culture of Yunnan.
In: Gender and Chinese Archaeology (Ed. by K.M. Linduff & Y. Sun),
pp. 289–314. Altamira Press, Walnut Creek, CA.
Cozzi M.C., Strillacci M.G., Valiati P., Bighignoli B., Cancedda M. &
Zanotti M. (2004) Mitochondrial D-loop sequence variation
among Italian horse breeds. Genetics Selection and Evolution 36,
663–72.
Fernandez H., Hughes S., Vigne J.D., Helmer D., Hodgins G., Miquel
C., Hanni C., Luikart G. & Taberlet P. (2006) Divergent mtDNA
lineages of goats in an Early Neolithic site, far from the initial
domestication areas. Proceedings of the National Academy of
Sciences of the United States of America 103, 15375–9.
Flad R., Yuan J. & Li S. (2007) Zooarcheological evidence of animal
domestication in northwest China. In: Late Quaternary Climate
Change and Human Adaptation in Arid China (Ed. by D.B. Madsen,
F.H. Chen & X. Gao), pp. 167–203. Elsevier, Amsterdam,
Holland.
Forster P., Torroni A., Renfrew C. & Rohl A. (2001) Phylogenetic
star contraction applied to Asian and Papuan mtDNA evolution.
Molecular Biology and Evolution 18, 1864–81.
Glazewska I., Wysocka A., Gralak B., Prus R. & Sell J. (2007) A new
view on dam lines in Polish Arabian horses based on mtDNA
analysis. Genetics Selection and Evolution 39, 609–19.
Hendricks B.L. (1995) International Encyclopedia of Horse Breeds.
University of Oklahoma Press, London.
Hill E.W., Bradley D.G., Al-Barody M., Ertugrul O., Splan R.K.,
Zakharov I. & Cunningham E.P. (2002) History and integrity of
thoroughbred dam lines revealed in equine mtDNA variation.
Animal Genetics 33, 287–94.
Hsu C.Y. & Linduff K. (1988) Western Chou Civilization. Yale
University Press, New Haven, CT.
Inner Mongolia Team, Institute of Archaeology CASS (1975) Ning-
cheng Nanshangen yizhi fajue baogao (Report of excavation at
Nanshangen in Ningcheng). Kaogu Xuebao, 117-40 (in Chinese).
Ivankovic A., Kavar T., Caput P., Mioc B., Pavic V. & Dovc P.
(2002) Genetic diversity of three donkey populations in the
Croatial region. Animal Genetics 33, 169–77.
Jansen T., Forster P., Levine M.A., Oelke H., Hurles M., Renfrew C.,
Weber J. & Olek K. (2002) Mitochondrial DNA and the origins of
the domestic horse. Proceedings of the National Academy of Sciences
of the United States of America 99, 10905–10.
Jung Y.H., Han S.H., Shin T. & Oh M.Y. (2002) Genetic charac-
terization of horse bone excavated from the Kwakji archaeologi-
cal site, Jeju, Korea. Molecule Cells 14, 224–30.
Keyser-Tracqui C., Blandin-Frappin P., Francfort H.P., Ricaut F.X.,
Lepetz S., Crubezy E., Samashev Z. & Ludes B. (2005) Mito-
chondrial DNA analysis of horses recovered from a frozen tomb
(Berel site, Kazahkhstan, 3rd Century BC). Animal Genetics 36,
203–9.
Kim K.I., Yang Y.H., Lee S.S., Park C., Ma R., Bouzat J.L. & Lewin
H.A. (1999) Phylogenetic relationships of Cheju horses to other
horse breeds as determined by mtDNA D-loop sequence poly-
morphism. Animal Genetics 30, 102–8.
Larson G., Dobney K., Albarella U. et al. (2005) Worldwide phy-
logeography of wild boar reveals multiple centers of pig domes-
tication. Science 307, 1618–21.
Larson G., Albarella U., Dobney K. et al. (2007a) Ancient DNA,
pig domestication, and the spread of the Neolithic into Europe.
Proceedings of the National Academy of Sciences of the United States
of America 104, 15276–81.
Larson G., Cucchi T., Fujita M. et al. (2007b) Phylogeny and an-
cient DNA of Sus provides insights into neolithic expansion in
Island Southeast Asia and Oceania. Proceedings of the National
Academy of Sciences of the United States of America 104, 4834–9.
Levine M. (2006) DNA and horse domestication: the archaeologist�scut. In: Equids in Time and Space (Ed. by M. Mashkour), pp. 192–
201. Oxbow Books, Oxford, UK.
Levine M., Renfrew C. & Boyle K. (2003) Prehistoric Steppe Adapta-
tion and the Horse. McDonald Institute for Archaeological
Research, Cabridge, UK.
Linduff K.M. (2003) A walk on the wild side: late Shang appro-
priation of horses in China. In: Prehistoric Steppe Adaptation and
the Horse (Ed. by M. Levine, C. Renfrew & K. Boyle), pp. 139–62.
McDonald Institute for Archaeological Research, Cambridge, UK.
Lister A.M., Vila C., Ellegren H., Gotherstrom A., Leonard J.A. &
Wayne R.K. (2001) Tales from the DNA of domestic horses.
Science 292, 218–9.
Luıs C., Bastos-Silveira C., Cothran E.G. & do Mar Oom M. (2006)
Iberian origins of New World horse breeds. Journal of Heredity 97,
107–13.
MacHugh D.E. & Bradley D.G. (2001) Livestock genetic origins:
Goats buck the trend. Proceedings of the National Academy of
Sciences of the United States of America 98, 5382–4.
Mair V. (2003) The horse in late Prehistoric China: wresting culture
and control from the �Barbarians�. In: Prehistoric Steppe Adaptation
and the Horse (Ed. by M. Levine, C. Renfrew & K. Boyle), pp. 163–
87. McDonald Institute for Archaeological Research, Cambridge,
UK.
Mang L., Li J.L. & Shi Y.F. (2005) Sequence comparing of mtDNA
D-loop varied region in Chinese Mongolian horse and external
thoroughbred horse. Hereditas (Beijing) 27, 91–94 (in Chinese).
Mashkour M. (2006) Equids in Time and Space: Papers in Honour of
Vera Eisenmann. Oxbow, Oxford, UK.
� 2009 The Authors, Journal compilation � 2009 Stichting International Foundation for Animal Genetics, Animal Genetics, 40, 933–944
The origins of Chinese horses 943

McGahern A., Bower M.A.M., Edwards C.J. et al. (2006a) Evidence
for biogeographic patterning of mitochondrial DNA sequences in
Eastern horse populations. Animal Genetics 37, 494–7.
McGahern A.M., Edwards C.J., Bower M.A., Heffernan A., Park
S.D.E., Brophy P.O., Bradley D.G., MacHugh D.E. & Hill E.W.
(2006b) Mitochondrial DNA sequence diversity in extant Irish
horse populations and in ancient horses. Animal Genetics 37,
498–502.
Olsen S.J. (1988) The horse in ancient China and its cultural
influence in some other areas. Proceedings of the Academy of
Natural Sciences of Philadelphia 140, 151–89.
Polzin T. & Daneschmand S.V. (2003) On Steiner trees and mini-
mum spanning trees in hypergraphs. Operations Research Letters
31, 12–20.
Royo L.J., Alvarez I., Beja-Pereira A., Molina A., Fernandez I.,
Jordana J., Gomez E., Gutierrez J.P. & Goyache F. (2005) The
origins of Iberian horses assessed via mitochondrial DNA. Journal
of Heredity 96, 663–9.
Rozas J., Sanchez-DelBarrio J.C., Messegyer X. & Rozas R. (2003)
DnaSP, DNA polymorphism analyses by the coalescent and other
methods. Bioinformatics 19, 2496–7.
Schneider S., Roessli D. & Excoffier L. (2000) ARLEQUIN: A Software
for Population Genetic Data Analysis (Version 2.0). Genetics
and Biometry Laboratory, University of Geneva, Geneva,
Switzerland.
Sokal R.R. & Rohlf F.J. (1995) Biometry: The Principles and Practice of
Statistics in Biological Research. Freeman, New York, NY.
Statistic Report in China Editing Commission (2004) Statistic Report
in China. Statistic Report in China Editing Commission, Beijing,
China.
Thompson J.D., Gibson T.J., Plewniak F., Jeanmougin F. & Higgins
D.G. (1997) The CLUSTAL-X windows interface: flexible strategies
for multiple sequences alignment aided by quality analysis tools.
Nucleic Acids Research 25, 4876–82.
Vila C., Leonard J.A., Gotherstrom A., Marklund S., Sandberg K.,
Liden K., Wayne R.K. & Ellegren H. (2001) Widespread origins of
domestic horse lineages. Science 291, 474–7.
Wang W., Liu A.H., Lin S.Y., Lan H., Su B., Xie D.W. & Shi L.M.
(1994) Multiple genotypes of mitochondrial DNA within a horse
population from a small region in Yunnan province of China.
Biochemical Genetics 32, 371–8.
Xie C.X. (1987) Horse and Ass Breeds in China. Shanghai Scientific
and Technical Publishing House, Shanghai, China.
Xu X. & Arnason U. (1994) The complete mitochondrial DNA
sequence of the horse, Equus caballus: extensive heteroplasmy of
the control region. Gene 148, 357–62.
Xu S., Luosang J., Hua S., He J., Ciren A., Wang W., Tong X., Liang
Y., Wang J. & Zheng X. (2007) High altitude adaptation and
phylogenetic analysis of Tibetan horse based on the mitochon-
drial genome. Journal of Genetics and Genomics 34, 720–9.
Yuan J. & Flad R. (2003) Two issues concerning ancient domesti-
cated horses in China. Bulletin of the Museum of Far Eastern
Antiquities 75, 110–26.
Yuan J. & Flad R. (2005) New zooarchaeological evidence for
changes in Shang dynasty animal sacrifice. Journal of Anthropo-
logical Archaeology 24, 252–70.
Yuan J. & Flad R. (2006) Research on early horse domestication in
China. In: Equids in Time and Space (Ed. by M. Mashkour), pp.
124–31. Oxbow Books, Oxford, UK.
Zhang Z. (1998) Jinning Shizhaishan. Yunnan Meishu Press, Kun-
ming, China.
Zhang Y. (2006) Chama gudao yu chama maoyi de lishi yu jiazhi
(The history and significance of Tea-horse Road and Tea-horse
trade). Xizang Daxue Xuebao 11, 34–40.
� 2009 The Authors, Journal compilation � 2009 Stichting International Foundation for Animal Genetics, Animal Genetics, 40, 933–944
Lei et al.944





























