Embed Size (px)
Citation preview

Journal of Electromyography and Kinesiology 17 (2007) 57–73
www.elsevier.com/locate/jelekin
Muscles within muscles: Coordination of 19 muscle segmentswithin three shoulder muscles during isometric motor tasks
J.M.M. Brown a, J.B. Wickham b,*, D.J. McAndrew a, X.-F. Huang a
a Department of Biomedical Science, University of Wollongong, Wollongong 2500, Australiab Department of Anatomy and Physiology, School of Human Biosciences, La Trobe University, Vic. 3086, Australia
Received 10 April 2005; received in revised form 24 September 2005; accepted 26 October 2005
Abstract
The aim of the present study was to determine how the intra-muscular segments of three shoulder muscles were coordinated to pro-duce isometric force impulses around the shoulder joint and how muscle segment coordination was influenced by changes in movementdirection, mechanical line of action and moment arm (ma).
Twenty male subjects (mean age 22 years; range 18–30 years) with no known history of shoulder pathologies, volunteered to partic-ipate in this experiment. Utilising an electromyographic technique, the timing and intensity of contraction within 19 muscle segments ofthree superficial shoulder muscles (Pectoralis Major, Deltoid and Latissimus Dorsi) were studied and compared during the production ofrapid (e.g. approximately 400 ms time to peak) isometric force impulses in four different movement directions of the shoulder joint (flex-ion, extension, abduction and adduction).
The results of this investigation have suggested that the timing and intensity of each muscle segment’s activation was coordinatedacross muscles and influenced by the muscle segment’s moment arm and its mechanical line of action in relation to the intended directionof shoulder movement (e.g. flexion, extension, abduction or adduction). There was also evidence that motor unit task groups wereformed for individual motor tasks which comprise motor units from both adjacent and distant muscles.
It was also confirmed that for any particular motor task, individual muscle segments can be functionally classified as prime mover,synergist or antagonist – classifications which are flexible from one movement to the next.� 2005 Elsevier Ltd. All rights reserved.
Keywords: Electromyography; Functional differentiation; Muscle segment; Shoulder muscles; Motor control; Motor unit task group
1. Introduction
It is now well accepted that motor units, within individ-ual muscle segments of single skeletal muscles, can be inde-pendently controlled by the central nervous system (CNS)to produce particular motor outcomes [3,21]. This phe-nomenon, which may be termed ‘functional differentiation’[18], has been described within a number of individual skel-etal muscles, including the Tensor Fascia Latae [18], theGluteus Maximus and Medius [16], the Triceps Surae [3],the Biceps Brachii [2] and the Pectoralis Major [19]. Ettema
1050-6411/$ - see front matter � 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jelekin.2005.10.007
* Corresponding author. Tel.: +61 3 94795725; fax: +61 3 94795784.E-mail address: [email protected] (J.B. Wickham).
et al. [5] has suggested that large absolute differences inmoment arms between different segments of a single musclepartially explain this phenomenon.
More recent studies on the Deltoid [24] have shown thatskeletal muscle has a greater potential for functional differ-entiation than hitherto accepted. Wickham and Brown [24]have determined that the Deltoid muscle, commonlydescribed as having only three (anterior, middle and poster-ior heads) functional muscle segments [10], is composed ofat least seven muscle segments which all have the potentialto be independently coordinated by the CNS. Functionallyindependent muscle segments provided the CNS greaterflexibility to ‘‘fine tune’’ the activity of its Deltoid motorunits when controlling movements of the shoulder joint.

58 J.M.M. Brown et al. / Journal of Electromyography and Kinesiology 17 (2007) 57–73
Furthermore, Wickham and Brown [24] have suggestedthat the seven individual muscle segments of the Deltoidmay be functionally classified as either prime mover, syner-
gist or antagonist muscle segments based upon theirmechanical lines of action (joint movement most likelyfrom approximation of a segment’s origin and insertion)and periods of activation. Attempts to assign functionalclassifications to individual muscle segments, classificationshitherto used to describe whole muscle function [1] are use-ful in characterising the activity of each individual musclesegment. For example, the prime mover segments of theDeltoid were found to have the most efficient mechanicallines of action, the earliest and longest durations of activa-tion and the highest amplitudes of myoelectric activity.Deltoid muscle segments with increasingly divergentmechanical lines of action (synergist segments) were foundto have increasingly later onsets, shorter periods of activa-tion and lower intensities of myoelectric activity. Finally,antagonist segments, with mechanical lines of action oppos-ing the movement, were activated last, had the shortestperiods of activation and variable amplitudes of myoelec-tric activity [24].
Applying functional classifications to muscle segmentswithin a single muscle is useful to provide a clearer under-standing of how the CNS controls motor unit activitywithin a muscle during a range of motor tasks. However,there have been few studies into how the CNS controlsthe muscle segments of individual muscles constituting amuscle group around a particular joint. We do not as yetunderstand how the muscle segments of individual muscles,within a muscle group, are coordinated together to producemotor tasks or how that coordination is affected by factorssuch as movement direction and moment arm.
Therefore, the aim of the present study was to determinehow the muscle segments of individual shoulder muscleswere coordinated together to produce isometric forceimpulses around the shoulder joint and how that coordina-
Fig. 1. The experimental setup. Depicted is one of the subjects seated in the eplane. In this position subjects performed the static tasks of shoulder joint fle
tion was influenced by movement direction, mechanicalline of action and moment arm. This investigation com-pared the activation of 19 muscle segments within threesuperficial shoulder muscles (Pectoralis Major, Deltoidand Latissimus Dorsi) during the execution of rapid (e.g.400 ms time to peak) isometric force impulses in four differ-ent movement directions (shoulder-flexion, -extension,-abduction and -adduction). Our intention was to under-stand how individual muscle segments were coordinatednot only within a single muscle, but across a group of mus-cles that controlled a single joint.
2. Methods
2.1. Procedures
Twenty male subjects (mean age 22 years; range 18–30years) with no known history of shoulder pathologies, vol-unteered to participate in this experiment.
The subjects sat in an adjustable dental chair within awire cage with their extended right upper limb positionedsnugly within an arm cast (Fig. 1). The arm cast was immo-bilised by attachment to the wire cage to permit the sub-jects to perform a series of isometric shoulder joint tasks.The arm cast, so fitted, ensured that the right upper limbwas fixed securely in space and did not move, other thandue to unavoidable compression of its soft tissues, duringany of the isometric motor tasks investigated.
Each subject performed four isometric tasks; shoulder-abduction, -adduction, -flexion and -extension, at 75% ofMVC, a movement time (MT) approximating 400 ms andwith an inter-trial period of at least 30 s. Each task was per-formed with the right upper limb secured in 20� of abduc-tion in the coronal plane with subject feedback onmovement accuracy being provided by an oscilloscope.
Ten trials were recorded from each isometric movementtask. Following an auditory trigger signal, each subject was
xperimental chair with the arm cast set at 20� of abduction in the frontalxion, extension, abduction and adduction.
J.M.M. Brown et al. / Journal of Electromyography and Kinesiology 17 (2007) 57–73 59
asked to match an electronically generated force–timecurve on an oscilloscope which provided the subject withfeedback on movement direction, movement speed andthe force of contraction. The order in which the trials wereperformed was randomised to minimise muscular fatigueand order effects. To permit normalisation of the intensitydata, three MVC contractions were recorded for eachmovement direction at the beginning of the experiment.The MVC contractions were recorded with an inter-trailinterval of 3 min.
2.2. Force recordings
Two load cells, connecting the arm cast to the wire cage,recorded the isometric force produced by the subjects in the
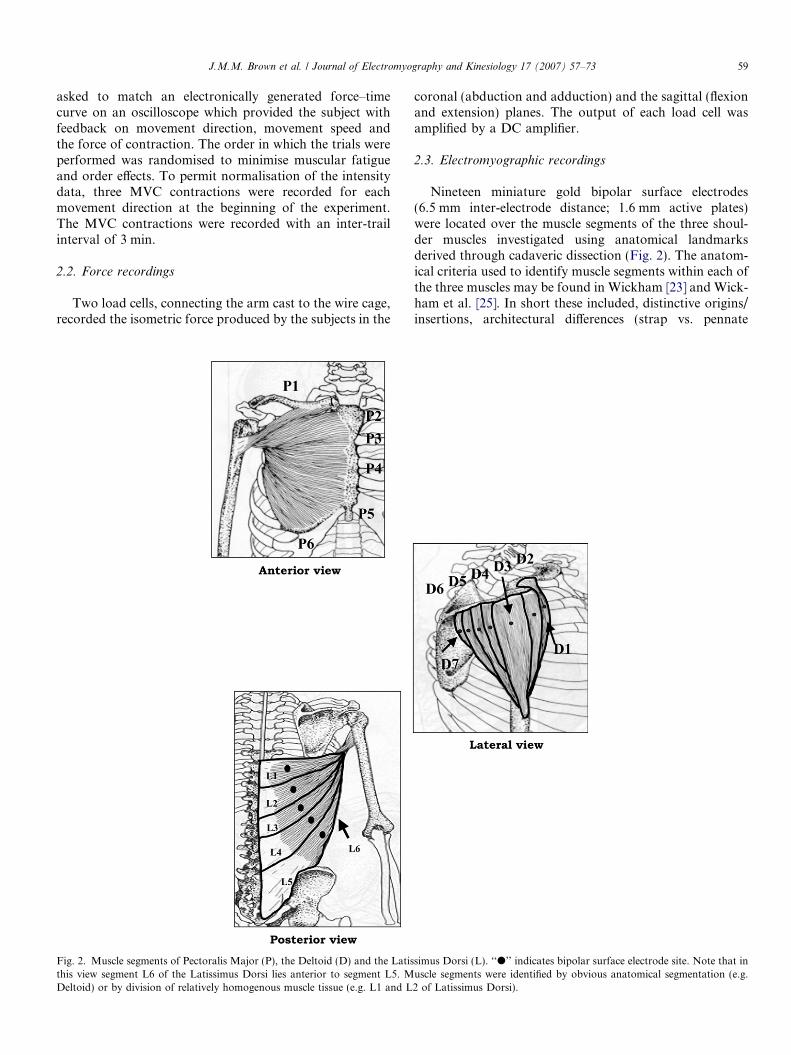
Fig. 2. Muscle segments of Pectoralis Major (P), the Deltoid (D) and the Latisthis view segment L6 of the Latissimus Dorsi lies anterior to segment L5. MDeltoid) or by division of relatively homogenous muscle tissue (e.g. L1 and L
coronal (abduction and adduction) and the sagittal (flexionand extension) planes. The output of each load cell wasamplified by a DC amplifier.
2.3. Electromyographic recordings
Nineteen miniature gold bipolar surface electrodes(6.5 mm inter-electrode distance; 1.6 mm active plates)were located over the muscle segments of the three shoul-der muscles investigated using anatomical landmarksderived through cadaveric dissection (Fig. 2). The anatom-ical criteria used to identify muscle segments within each ofthe three muscles may be found in Wickham [23] and Wick-ham et al. [25]. In short these included, distinctive origins/insertions, architectural differences (strap vs. pennate
simus Dorsi (L). ‘‘d’’ indicates bipolar surface electrode site. Note that inuscle segments were identified by obvious anatomical segmentation (e.g.2 of Latissimus Dorsi).

60 J.M.M. Brown et al. / Journal of Electromyography and Kinesiology 17 (2007) 57–73
fibered), intramuscular fascial thickenings and >10� differ-ence in lines of action between adjacent segments. One ormore of these criteria needed to be present. In all 19 bipolarsurface electrodes were applied to each subject; seven overthe Deltoid and six each over the Pectoralis Major and theLatissimus Dorsi (Fig. 1).
Electrode gel was injected into the electrode wells byutilising a syringe coupled to a 1 mm cannula. The surfaceelectrodes were secured to the subject by double-sided tapeafter the skin had been shaved, abraded and washed withalcohol to reduce skin resistance. Reference surface elec-trodes (3M paediatric electrodes) were positioned overthe acromion, clavicle and the anterior superior iliac spine.The 19 bipolar surface electrodes and their reference elec-trodes were connected to differential preamplifiers andHUMTEC 100 EMG amplifiers (input impedance of1 · 1013 X, CMRR of 110 db, signal to noise ratio of1000:1, 1000k gain). The raw EMG signals were amplifiedand filtered (10 Hz high pass and 1 kHz low pass; Butter-worth filters) prior to storage.
The isometric force impulse and the raw EMG wave-forms were stored on an IBM compatible microcomputerafter A–D conversion at 1 kHz. A 2000 ms recording per-iod, commencing at least 200 ms before the beginning ofthe each motor task, was recorded for each trial.
2.4. Data analysis
Data analysis of the EMG waveforms utilised a digitalsignal processing (DSP) package for determinations ofmuscle segment contraction intensity and timing (Fig. 3).
Fig. 3. The analysis screen window that was used to locate the temporal and cshows a representative raw EMG signal with its 20 Hz linear envelope displaydetector set at 10% of peak segmental intensity (horizontal line) that was usedsignal was used when there was a low signal to noise ratio and the threshold detfrom the 100 kg prime load cell is displayed in the bottom window. The durataverage V/ms (total Vms divided by duration). Representative data from one
This program permitted the raw EMG signals to be pro-cessed (rectified, low passed, high passed, etc.) prior toanalysis.
2.5. Timing analysis
Firstly, the time of force onset and the timing of forcepeak were determined with the temporal features of musclesegment activation determined in relation to these values(Fig. 3).
To determine the timing of each muscle segment, theraw EMG waveforms were rectified and smoothed with a20 Hz low pass filter (Fig. 3). Threshold detectors (10%peak amplitude), combined with visual analysis, were usedto locate the onset (ON), peak (PEAK) and end (OFF) ofactivity within each muscle segment (Fig. 3). From thisdata the duration of segmental activation could also bedetermined. ON and OFF were determined in relation tothe beginning of the isometric force onset while PEAKwas measured in relation to the timing of peak isometricforce (Fig. 3).
2.6. Intensity analysis
Each EMG waveform, representing the activity of a seg-ment within a particular trial was integrated to determinethe intensity of muscle segment contraction. The EMGwaveforms were full wave rectified and then integrated(10 ms bins) to determine the muscle segment’s intensityof activation or discharge rate (DR: mVms/ms). The dis-charge rate was the average intensity of motor unit con-
onsequently the intensity parameters of the EMG signals. The top windowed in the middle window. The middle window also displays the thresholdto locate the on/off positions of the EMG signals. Visual detection of theector could not determine an appropriate on/off location. The force recordion of each burst was integrated (time reset; 1 ms bins) to give a value ofsubject (segment D1) during a 75% MVC shoulder joint flexion task.

J.M.M. Brown et al. / Journal of Electromyography and Kinesiology 17 (2007) 57–73 61
traction throughout the muscle segments period of activa-tion. This value was then normalised against the dischargerate found during the MVC contractions associated witheach particular movement direction.
2.7. Biomechanical analysis
Three-dimensional cartesian coordinates of each of the19 muscle segments were determined through use of a ver-tical milling machine. This sort of machine, which is com-monly used in engineering workshops for shaping metalvery accurately (to within 0.01 mm), was considered morethan adequate for our biomechanical analysis. By securinga cadaver (bisected at the waist) to a specially designedframe in front of the milling machine, 3D coordinates ofeach muscle segment and the axis of rotation of the shoul-der joint were determined (Fig. 4). This was made possibleby securing custom made pointers to the mill vice and thenmanipulating the milling machine X, Y and Z mill slides toapproximate the pointers at each segmental attachmentpoint. A detailed description of the utilisation of the verti-cal milling machine may be found in Wickham et al. [25].AutoCAD was utilised to graphically represent the biome-chanical relationships (e.g. mechanical lines of action)between each of the muscle segments and the axis of shoul-der joint rotation. In addition, moment arms (ma) weredetermined for each individual segment for both shoulderabduction/adduction and flexion/extension.
2.8. Functional classifications of muscle segments
The functional classifications of each muscle segmentwere simplified from those previously proposed by Wick-ham and Brown [24]. In the present study, functional clas-sifications of muscle segments were applied as followed:
Fig. 4. A composite representation (AutoCad) of the lines of action to thehumerus for each segment of the Pectoralis Major (P), the Deltoid (D) andthe Latissimus Dorsi (L). Coronal view with the upper limb abducted to20� in the coronal plane. Representative data from one cadaver.
(a) Prime Mover segment: A muscle segment which wasactivated first and had an agonist moment arm forthe intended motor task. Prime mover segments hadmuscle fibres with mechanical lines of action similar,or identical to, the direction of the intended jointmotion.
(b) Synergist segment: A muscle segment, with an agonistmuscle arm, which was activated significantly(p < 0.05) later than the prime mover segment/s.Often, the synergist segments had mechanical linesof action divergent away from the prime mover seg-ments and the intended movement direction.
(c) Antagonist segment: A muscle segment with an antag-onist moment arm which was activated with, or sig-nificantly (p < 0.05) after, the prime mover segment/s. In all cases, antagonist segments had mechanicallines of action and moment arms that opposed thedirection of intended joint movement.
The functional classification of a muscle segment wasdetermined after review of all muscles segments’ contribu-tions, from the three muscles investigated, to the motortask. Therefore particular functional classifications (e.g.prime mover segment) for a given motor task may beapplied to the muscle segments of one or more muscles.
2.9. Statistical evaluations
Repeated measures analysis of variance and StudentNeumann Keuls techniques were used to evaluate the data.Pearson’s correlation coefficients were used to determinethe relationship between the timing of activity in musclesegments of adjacent muscles. Statistical evaluations weredesigned to determine whether the timing and contractionintensity of muscle segment activity varied significantly:(a) within each individual muscle and (b) across all threemuscles investigated. The level of significance was set at(p < 0.05).
3. Results
3.1. Biomechanical data
Table 1 provides a biomechanical analysis of each of the19 muscle segments derived from one representative cada-ver. Note that the largest muscle segment cross-sectionalareas were found in segments P1 and P2 (clavicular head)of the Pectoralis Major, segment D3 (middle head) of theDeltoid and segment L2 (upper fibres) of Latissimus Dorsi.Within both the Deltoid and the Latissimus Dorsi, the‘‘stronger’’ muscle segments were found towards the mid-dle of the muscle. In contrast, the upper segments (clavicu-lar head) of the Pectoralis Major had the greatest forcegeneration potential.
Table 2 illustrates the moment arms for each muscle seg-ment derived for both flexion/extension and abduction/adduction of the shoulder joint. All muscle segments of
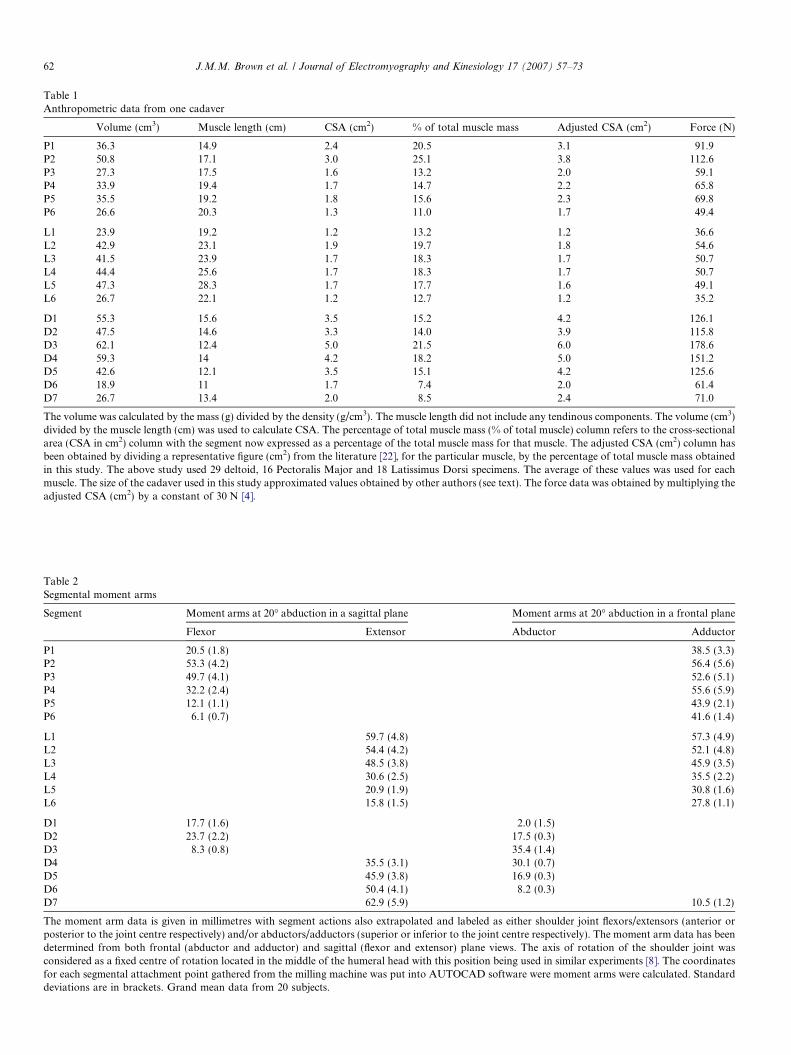
Table 1Anthropometric data from one cadaver
Volume (cm3) Muscle length (cm) CSA (cm2) % of total muscle mass Adjusted CSA (cm2) Force (N)
P1 36.3 14.9 2.4 20.5 3.1 91.9P2 50.8 17.1 3.0 25.1 3.8 112.6P3 27.3 17.5 1.6 13.2 2.0 59.1P4 33.9 19.4 1.7 14.7 2.2 65.8P5 35.5 19.2 1.8 15.6 2.3 69.8P6 26.6 20.3 1.3 11.0 1.7 49.4
L1 23.9 19.2 1.2 13.2 1.2 36.6L2 42.9 23.1 1.9 19.7 1.8 54.6L3 41.5 23.9 1.7 18.3 1.7 50.7L4 44.4 25.6 1.7 18.3 1.7 50.7L5 47.3 28.3 1.7 17.7 1.6 49.1L6 26.7 22.1 1.2 12.7 1.2 35.2
D1 55.3 15.6 3.5 15.2 4.2 126.1D2 47.5 14.6 3.3 14.0 3.9 115.8D3 62.1 12.4 5.0 21.5 6.0 178.6D4 59.3 14 4.2 18.2 5.0 151.2D5 42.6 12.1 3.5 15.1 4.2 125.6D6 18.9 11 1.7 7.4 2.0 61.4D7 26.7 13.4 2.0 8.5 2.4 71.0
The volume was calculated by the mass (g) divided by the density (g/cm3). The muscle length did not include any tendinous components. The volume (cm3)divided by the muscle length (cm) was used to calculate CSA. The percentage of total muscle mass (% of total muscle) column refers to the cross-sectionalarea (CSA in cm2) column with the segment now expressed as a percentage of the total muscle mass for that muscle. The adjusted CSA (cm2) column hasbeen obtained by dividing a representative figure (cm2) from the literature [22], for the particular muscle, by the percentage of total muscle mass obtainedin this study. The above study used 29 deltoid, 16 Pectoralis Major and 18 Latissimus Dorsi specimens. The average of these values was used for eachmuscle. The size of the cadaver used in this study approximated values obtained by other authors (see text). The force data was obtained by multiplying theadjusted CSA (cm2) by a constant of 30 N [4].
Table 2Segmental moment arms
Segment Moment arms at 20� abduction in a sagittal plane Moment arms at 20� abduction in a frontal plane
Flexor Extensor Abductor Adductor
P1 20.5 (1.8) 38.5 (3.3)P2 53.3 (4.2) 56.4 (5.6)P3 49.7 (4.1) 52.6 (5.1)P4 32.2 (2.4) 55.6 (5.9)P5 12.1 (1.1) 43.9 (2.1)P6 6.1 (0.7) 41.6 (1.4)
L1 59.7 (4.8) 57.3 (4.9)L2 54.4 (4.2) 52.1 (4.8)L3 48.5 (3.8) 45.9 (3.5)L4 30.6 (2.5) 35.5 (2.2)L5 20.9 (1.9) 30.8 (1.6)L6 15.8 (1.5) 27.8 (1.1)
D1 17.7 (1.6) 2.0 (1.5)D2 23.7 (2.2) 17.5 (0.3)D3 8.3 (0.8) 35.4 (1.4)D4 35.5 (3.1) 30.1 (0.7)D5 45.9 (3.8) 16.9 (0.3)D6 50.4 (4.1) 8.2 (0.3)D7 62.9 (5.9) 10.5 (1.2)
The moment arm data is given in millimetres with segment actions also extrapolated and labeled as either shoulder joint flexors/extensors (anterior orposterior to the joint centre respectively) and/or abductors/adductors (superior or inferior to the joint centre respectively). The moment arm data has beendetermined from both frontal (abductor and adductor) and sagittal (flexor and extensor) plane views. The axis of rotation of the shoulder joint wasconsidered as a fixed centre of rotation located in the middle of the humeral head with this position being used in similar experiments [8]. The coordinatesfor each segmental attachment point gathered from the milling machine was put into AUTOCAD software were moment arms were calculated. Standarddeviations are in brackets. Grand mean data from 20 subjects.
62 J.M.M. Brown et al. / Journal of Electromyography and Kinesiology 17 (2007) 57–73
D2
D6D5D4D3
P4
P1P2P3
P6P5
D1
Segment
J.M.M. Brown et al. / Journal of Electromyography and Kinesiology 17 (2007) 57–73 63
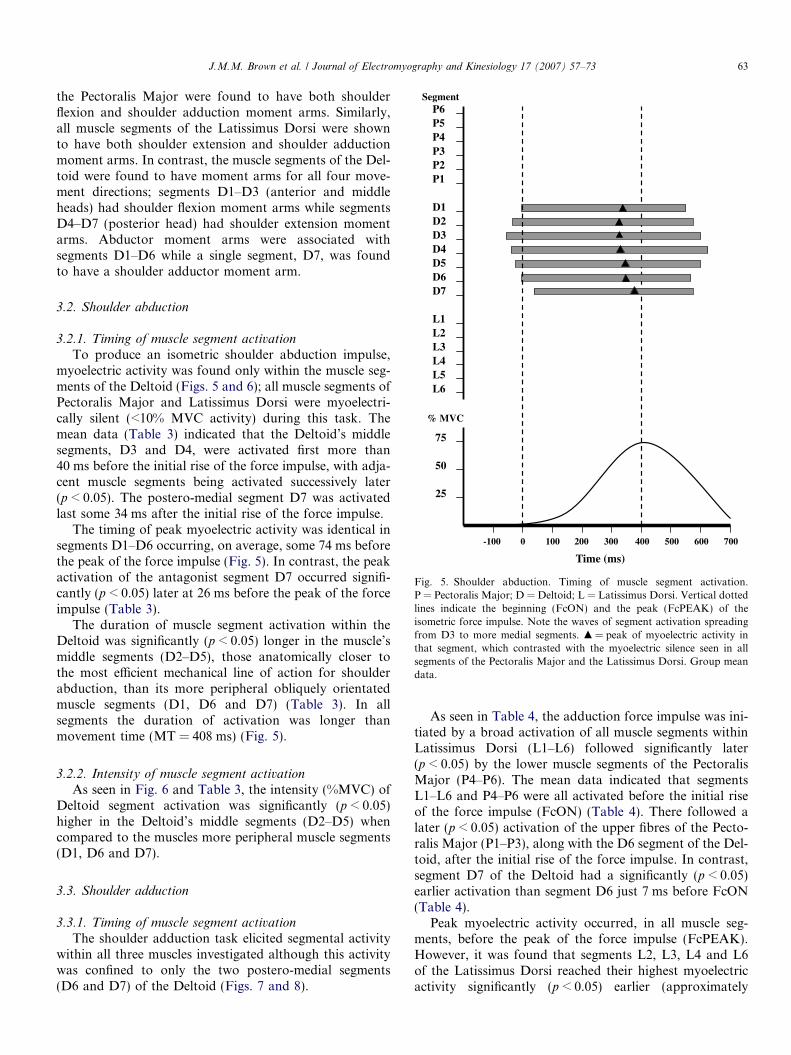
the Pectoralis Major were found to have both shoulderflexion and shoulder adduction moment arms. Similarly,all muscle segments of the Latissimus Dorsi were shownto have both shoulder extension and shoulder adductionmoment arms. In contrast, the muscle segments of the Del-toid were found to have moment arms for all four move-ment directions; segments D1–D3 (anterior and middleheads) had shoulder flexion moment arms while segmentsD4–D7 (posterior head) had shoulder extension momentarms. Abductor moment arms were associated withsegments D1–D6 while a single segment, D7, was foundto have a shoulder adductor moment arm.
L1
L3L2
L5L4
L6
D7
% MVC
50
25
75
-100 0 100 200 300 400 500 600 700
Time (ms)
Fig. 5. Shoulder abduction. Timing of muscle segment activation.P = Pectoralis Major; D = Deltoid; L = Latissimus Dorsi. Vertical dottedlines indicate the beginning (FcON) and the peak (FcPEAK) of theisometric force impulse. Note the waves of segment activation spreadingfrom D3 to more medial segments. m = peak of myoelectric activity inthat segment, which contrasted with the myoelectric silence seen in allsegments of the Pectoralis Major and the Latissimus Dorsi. Group meandata.
3.2. Shoulder abduction
3.2.1. Timing of muscle segment activation
To produce an isometric shoulder abduction impulse,myoelectric activity was found only within the muscle seg-ments of the Deltoid (Figs. 5 and 6); all muscle segments ofPectoralis Major and Latissimus Dorsi were myoelectri-cally silent (<10% MVC activity) during this task. Themean data (Table 3) indicated that the Deltoid’s middlesegments, D3 and D4, were activated first more than40 ms before the initial rise of the force impulse, with adja-cent muscle segments being activated successively later(p < 0.05). The postero-medial segment D7 was activatedlast some 34 ms after the initial rise of the force impulse.
The timing of peak myoelectric activity was identical insegments D1–D6 occurring, on average, some 74 ms beforethe peak of the force impulse (Fig. 5). In contrast, the peakactivation of the antagonist segment D7 occurred signifi-cantly (p < 0.05) later at 26 ms before the peak of the forceimpulse (Table 3).
The duration of muscle segment activation within theDeltoid was significantly (p < 0.05) longer in the muscle’smiddle segments (D2–D5), those anatomically closer tothe most efficient mechanical line of action for shoulderabduction, than its more peripheral obliquely orientatedmuscle segments (D1, D6 and D7) (Table 3). In allsegments the duration of activation was longer thanmovement time (MT = 408 ms) (Fig. 5).
3.2.2. Intensity of muscle segment activation
As seen in Fig. 6 and Table 3, the intensity (%MVC) ofDeltoid segment activation was significantly (p < 0.05)higher in the Deltoid’s middle segments (D2–D5) whencompared to the muscles more peripheral muscle segments(D1, D6 and D7).
3.3. Shoulder adduction
3.3.1. Timing of muscle segment activation
The shoulder adduction task elicited segmental activitywithin all three muscles investigated although this activitywas confined to only the two postero-medial segments(D6 and D7) of the Deltoid (Figs. 7 and 8).
As seen in Table 4, the adduction force impulse was ini-tiated by a broad activation of all muscle segments withinLatissimus Dorsi (L1–L6) followed significantly later(p < 0.05) by the lower muscle segments of the PectoralisMajor (P4–P6). The mean data indicated that segmentsL1–L6 and P4–P6 were all activated before the initial riseof the force impulse (FcON) (Table 4). There followed alater (p < 0.05) activation of the upper fibres of the Pecto-ralis Major (P1–P3), along with the D6 segment of the Del-toid, after the initial rise of the force impulse. In contrast,segment D7 of the Deltoid had a significantly (p < 0.05)earlier activation than segment D6 just 7 ms before FcON(Table 4).
Peak myoelectric activity occurred, in all muscle seg-ments, before the peak of the force impulse (FcPEAK).However, it was found that segments L2, L3, L4 and L6of the Latissimus Dorsi reached their highest myoelectricactivity significantly (p < 0.05) earlier (approximately

Fig. 6. Shoulder abduction. Intensity of muscle segment contraction in Latissimus Dorsi (L1–L6), Deltoid (D1–D7) and Pectoralis Major (P1–P7).Diagram based on group mean data. Note that muscle segment activity is limited to the Deltoid muscle only and more intense in the middle (adductor)segments of the muscle.
Table 3Shoulder abduction; timing (ms) and intensity (%MVC)
On (ms) Peak (ms) Duration(ms)
Intensity(%MVC)
x s.d. x s.d. x s.d. x s.d.
D1 �04* 26 �68 43 554* 96 38* 24D2 �35* 19 �85 58 617 87 83 33D3 �53 19 �82 37 636 76 87 17D4 �42 23 �81 31 657 96 88 20D5 �24* 24 �59 31 624 81 73 23D6 �05* 21 �66 41 560* 84 58* 30D7 34* 26 �26* 66 528* 70 51* 30
Mean and standard deviations. Significant (p < 0.05) difference: *to seg-ment D3. Movement time (FcOn to FcPeak) of 408 ms.
64 J.M.M. Brown et al. / Journal of Electromyography and Kinesiology 17 (2007) 57–73
�117 ms) than all other muscle segments (approximately�65 ms) investigated (Table 4).
The duration of muscle segment activation was found tobe longest in segments L1–L6, D7 and P3–P6 (approxi-mately 584 ms). Segments P1, P2 and D6 were all foundto have significantly (p < 0.05) shorter periods of activation(approximately 495 ms) during this task (Table 4). Allactive muscle segments had periods of activity longer thanadduction movement time of 390 ms (Table 4).
3.3.2. Intensity of muscle segment activation
The intensities of muscle segment activation (%MVC)were similar throughout the muscle segments of LatissimusDorsi and the lower segments of the Pectoralis Major(Table 4). All muscle segments of Latissimus Dorsi (L1–L6), and the lower segments of Pectoralis Major (P4–P6)had significantly (p < 0.05) higher intensities of activation(approximately 64% MVC) than the other activated musclesegments (approximately 34% MVC) (Table 4 and Fig. 8).
3.3.3. The sequence of muscle segment activation
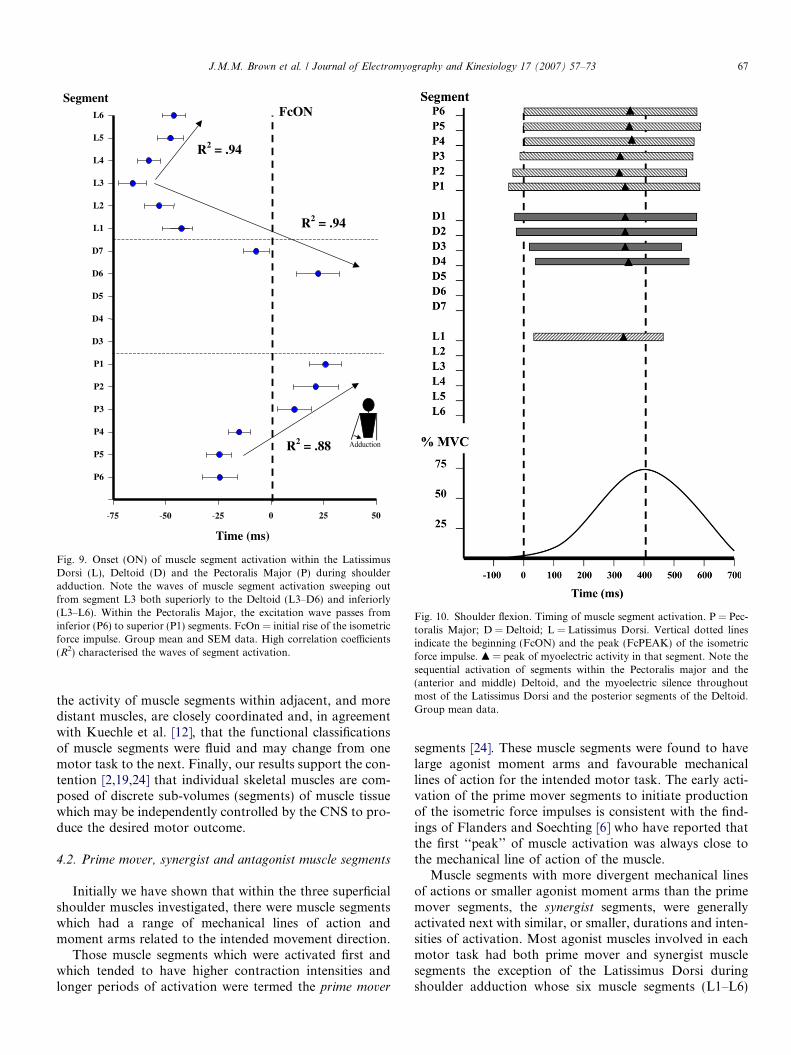
Correlation of muscle segment activation times duringthe adduction motor task (Fig. 9) indicated that each mus-cle segment, within the three muscles investigated, was acti-vated in a sequence that ignored anatomical boundariesbetween adjacent muscles. As seen in Fig. 9, the CNS pro-duced three waves of muscle segment activation (sequentialactivation of muscle segments) during the adduction motortask. Two muscle segment activation waves, centred onsegment L3, progressively activated the muscle segmentsof Latissimus Dorsi as well as the postero-medial segmentsof the Deltoid. The other wave of muscle segment activa-tion began within segments P5 and P6 (lower fibres) andthen progressively activated higher muscle segments ofthe Pectoralis Major. All three muscle segment activationwaves were associated with high correlation coefficients(R2) of between 0.88 and 0.94 (Fig. 9).
3.4. Shoulder flexion
3.4.1. Timing of muscle segment activation
The shoulder flexion task was accomplished by wide-spread activation of the Pectoralis Major (P1–P6), the ante-rior four segments of the Deltoid (D1–D4) and only onesegment (L1) of the antagonist Latissimus Dorsi(Fig. 10). The task was initiated by simultaneous activationof four adjacent muscle segments on the anterior aspect ofthe shoulder; P1 and P2 (clavicular head) with D1 and D2(anterior head) (Fig. 10). A significantly (p < 0.05) lateractivation occurred in segments P3–P6, D3, D4 and L1(Table 5). The remaining segments (D5–D7; L2–L6) ofthe Deltoid and the Latissimus Dorsi respectively remainedinactive during this task (Fig. 11).
All active muscle segments across the three muscle inves-tigated, except segment P4, reached their maximum inten-
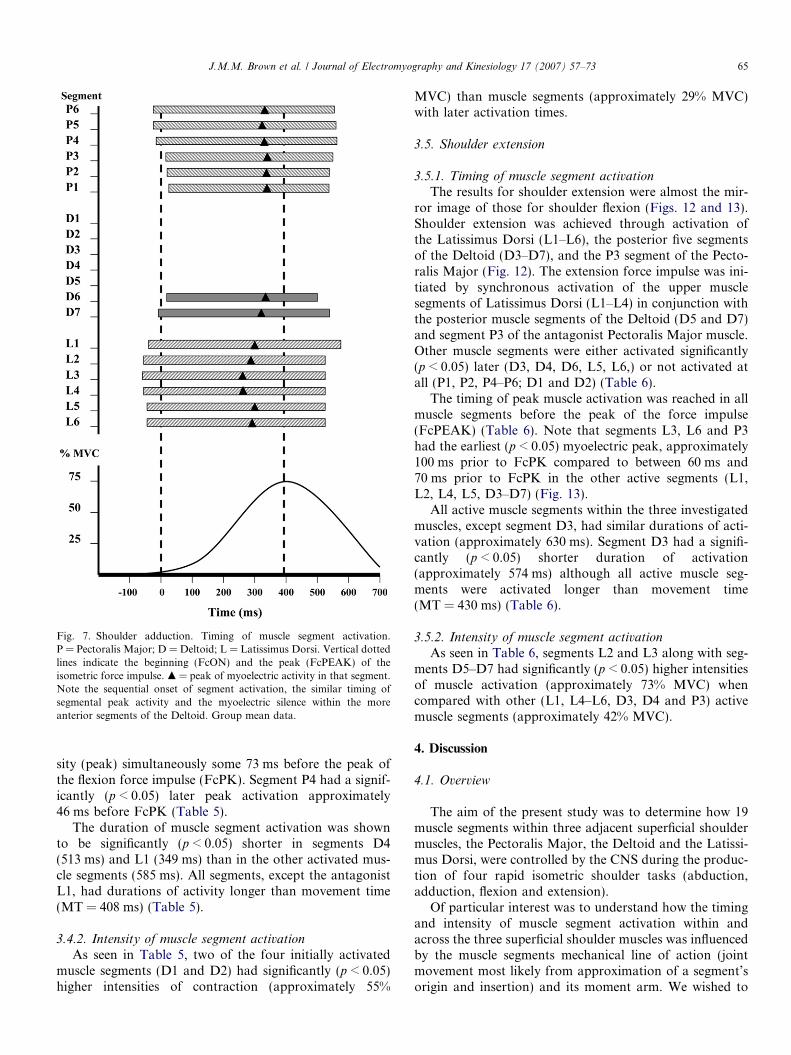
Fig. 7. Shoulder adduction. Timing of muscle segment activation.P = Pectoralis Major; D = Deltoid; L = Latissimus Dorsi. Vertical dottedlines indicate the beginning (FcON) and the peak (FcPEAK) of theisometric force impulse. m = peak of myoelectric activity in that segment.Note the sequential onset of segment activation, the similar timing ofsegmental peak activity and the myoelectric silence within the moreanterior segments of the Deltoid. Group mean data.
J.M.M. Brown et al. / Journal of Electromyography and Kinesiology 17 (2007) 57–73 65
sity (peak) simultaneously some 73 ms before the peak ofthe flexion force impulse (FcPK). Segment P4 had a signif-icantly (p < 0.05) later peak activation approximately46 ms before FcPK (Table 5).
The duration of muscle segment activation was shownto be significantly (p < 0.05) shorter in segments D4(513 ms) and L1 (349 ms) than in the other activated mus-cle segments (585 ms). All segments, except the antagonistL1, had durations of activity longer than movement time(MT = 408 ms) (Table 5).
3.4.2. Intensity of muscle segment activation
As seen in Table 5, two of the four initially activatedmuscle segments (D1 and D2) had significantly (p < 0.05)higher intensities of contraction (approximately 55%
MVC) than muscle segments (approximately 29% MVC)with later activation times.
3.5. Shoulder extension
3.5.1. Timing of muscle segment activationThe results for shoulder extension were almost the mir-
ror image of those for shoulder flexion (Figs. 12 and 13).Shoulder extension was achieved through activation ofthe Latissimus Dorsi (L1–L6), the posterior five segmentsof the Deltoid (D3–D7), and the P3 segment of the Pecto-ralis Major (Fig. 12). The extension force impulse was ini-tiated by synchronous activation of the upper musclesegments of Latissimus Dorsi (L1–L4) in conjunction withthe posterior muscle segments of the Deltoid (D5 and D7)and segment P3 of the antagonist Pectoralis Major muscle.Other muscle segments were either activated significantly(p < 0.05) later (D3, D4, D6, L5, L6,) or not activated atall (P1, P2, P4–P6; D1 and D2) (Table 6).
The timing of peak muscle activation was reached in allmuscle segments before the peak of the force impulse(FcPEAK) (Table 6). Note that segments L3, L6 and P3had the earliest (p < 0.05) myoelectric peak, approximately100 ms prior to FcPK compared to between 60 ms and70 ms prior to FcPK in the other active segments (L1,L2, L4, L5, D3–D7) (Fig. 13).
All active muscle segments within the three investigatedmuscles, except segment D3, had similar durations of acti-vation (approximately 630 ms). Segment D3 had a signifi-cantly (p < 0.05) shorter duration of activation(approximately 574 ms) although all active muscle seg-ments were activated longer than movement time(MT = 430 ms) (Table 6).
3.5.2. Intensity of muscle segment activation
As seen in Table 6, segments L2 and L3 along with seg-ments D5–D7 had significantly (p < 0.05) higher intensitiesof muscle activation (approximately 73% MVC) whencompared with other (L1, L4–L6, D3, D4 and P3) activemuscle segments (approximately 42% MVC).
4. Discussion
4.1. Overview
The aim of the present study was to determine how 19muscle segments within three adjacent superficial shouldermuscles, the Pectoralis Major, the Deltoid and the Latissi-mus Dorsi, were controlled by the CNS during the produc-tion of four rapid isometric shoulder tasks (abduction,adduction, flexion and extension).
Of particular interest was to understand how the timingand intensity of muscle segment activation within andacross the three superficial shoulder muscles was influencedby the muscle segments mechanical line of action (jointmovement most likely from approximation of a segment’sorigin and insertion) and its moment arm. We wished to
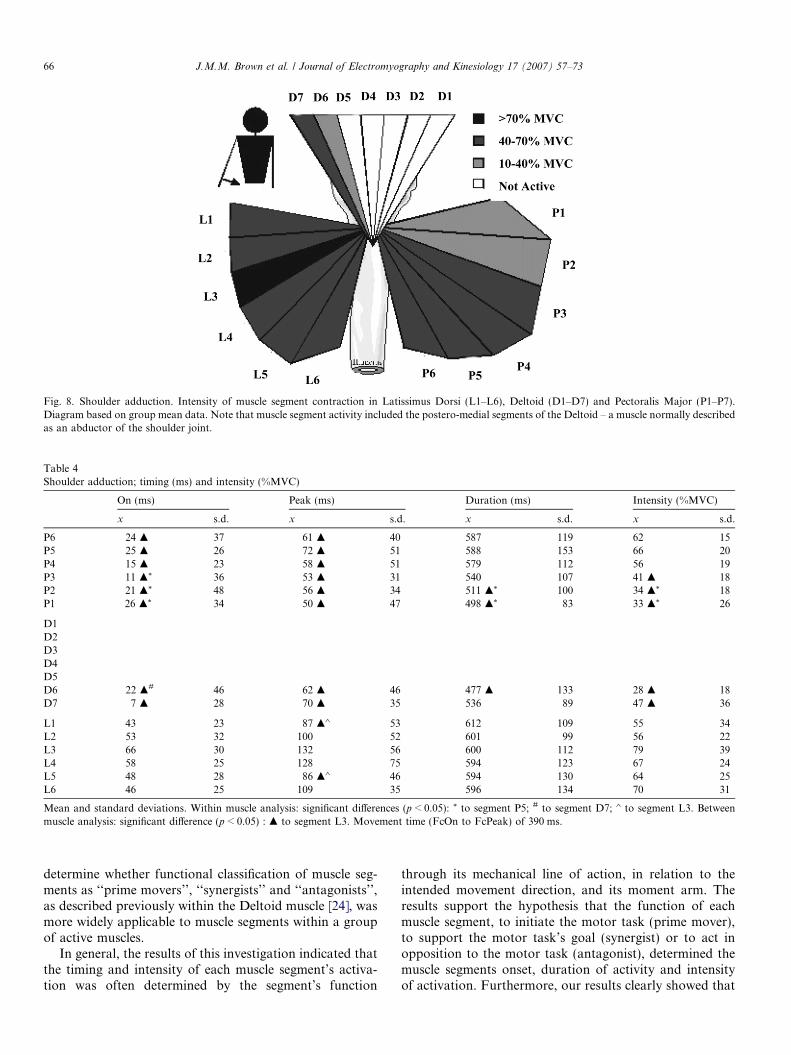
Fig. 8. Shoulder adduction. Intensity of muscle segment contraction in Latissimus Dorsi (L1–L6), Deltoid (D1–D7) and Pectoralis Major (P1–P7).Diagram based on group mean data. Note that muscle segment activity included the postero-medial segments of the Deltoid – a muscle normally describedas an abductor of the shoulder joint.
Table 4Shoulder adduction; timing (ms) and intensity (%MVC)
On (ms) Peak (ms) Duration (ms) Intensity (%MVC)
x s.d. x s.d. x s.d. x s.d.
P6 �24 m 37 �61 m 40 587 119 62 15P5 �25 m 26 �72 m 51 588 153 66 20P4 �15 m 23 �58 m 51 579 112 56 19P3 11 m* 36 �53 m 31 540 107 41 m 18P2 21 m* 48 �56 m 34 511 m* 100 34 m* 18P1 26 m* 34 �50 m 47 498 m* 83 33 m* 26
D1D2D3D4D5D6 22 m# 46 �62 m 46 477 m 133 28 m 18D7 �7 m 28 �70 m 35 536 89 47 m 36
L1 �43 23 �87 m� 53 612 109 55 34L2 �53 32 �100 52 601 99 56 22L3 �66 30 �132 56 600 112 79 39L4 �58 25 �128 75 594 123 67 24L5 �48 28 �86 m� 46 594 130 64 25L6 �46 25 �109 35 596 134 70 31
Mean and standard deviations. Within muscle analysis: significant differences (p < 0.05): * to segment P5; # to segment D7; � to segment L3. Betweenmuscle analysis: significant difference (p < 0.05) : m to segment L3. Movement time (FcOn to FcPeak) of 390 ms.
66 J.M.M. Brown et al. / Journal of Electromyography and Kinesiology 17 (2007) 57–73
determine whether functional classification of muscle seg-ments as ‘‘prime movers’’, ‘‘synergists’’ and ‘‘antagonists’’,as described previously within the Deltoid muscle [24], wasmore widely applicable to muscle segments within a groupof active muscles.
In general, the results of this investigation indicated thatthe timing and intensity of each muscle segment’s activa-tion was often determined by the segment’s function
through its mechanical line of action, in relation to theintended movement direction, and its moment arm. Theresults support the hypothesis that the function of eachmuscle segment, to initiate the motor task (prime mover),to support the motor task’s goal (synergist) or to act inopposition to the motor task (antagonist), determined themuscle segments onset, duration of activity and intensityof activation. Furthermore, our results clearly showed that
FcON
R2 = .94
R2 = .88
R2 = .94
L6
L5
L4
L3
L2
L1
D7
D6
D5
D4
D3
P1
P2
P3
P4
P5
P6
Segment
25 500-25-50-75
Time (ms)
Fig. 9. Onset (ON) of muscle segment activation within the LatissimusDorsi (L), Deltoid (D) and the Pectoralis Major (P) during shoulderadduction. Note the waves of muscle segment activation sweeping outfrom segment L3 both superiorly to the Deltoid (L3–D6) and inferiorly(L3–L6). Within the Pectoralis Major, the excitation wave passes frominferior (P6) to superior (P1) segments. FcOn = initial rise of the isometricforce impulse. Group mean and SEM data. High correlation coefficients(R2) characterised the waves of segment activation.
Fig. 10. Shoulder flexion. Timing of muscle segment activation. P = Pec-toralis Major; D = Deltoid; L = Latissimus Dorsi. Vertical dotted linesindicate the beginning (FcON) and the peak (FcPEAK) of the isometricforce impulse. m = peak of myoelectric activity in that segment. Note thesequential activation of segments within the Pectoralis major and the(anterior and middle) Deltoid, and the myoelectric silence throughoutmost of the Latissimus Dorsi and the posterior segments of the Deltoid.Group mean data.
J.M.M. Brown et al. / Journal of Electromyography and Kinesiology 17 (2007) 57–73 67
the activity of muscle segments within adjacent, and moredistant muscles, are closely coordinated and, in agreementwith Kuechle et al. [12], that the functional classificationsof muscle segments were fluid and may change from onemotor task to the next. Finally, our results support the con-tention [2,19,24] that individual skeletal muscles are com-posed of discrete sub-volumes (segments) of muscle tissuewhich may be independently controlled by the CNS to pro-duce the desired motor outcome.
4.2. Prime mover, synergist and antagonist muscle segments
Initially we have shown that within the three superficialshoulder muscles investigated, there were muscle segmentswhich had a range of mechanical lines of action andmoment arms related to the intended movement direction.
Those muscle segments which were activated first andwhich tended to have higher contraction intensities andlonger periods of activation were termed the prime mover
segments [24]. These muscle segments were found to havelarge agonist moment arms and favourable mechanicallines of action for the intended motor task. The early acti-vation of the prime mover segments to initiate productionof the isometric force impulses is consistent with the find-ings of Flanders and Soechting [6] who have reported thatthe first ‘‘peak’’ of muscle activation was always close tothe mechanical line of action of the muscle.
Muscle segments with more divergent mechanical linesof actions or smaller agonist moment arms than the primemover segments, the synergist segments, were generallyactivated next with similar, or smaller, durations and inten-sities of activation. Most agonist muscles involved in eachmotor task had both prime mover and synergist musclesegments the exception of the Latissimus Dorsi duringshoulder adduction whose six muscle segments (L1–L6)

Table 5Shoulder flexion; timing (ms) and intensity (%MVC)
On (ms) Peak (ms) Duration(ms)
Intensity(%MVC)
x s.d. x s.d. x s.d. x s.d.
P6 3 m* 44 �53 29 596 88 27 .* 16P5 �2 m* 36 �59 49 602 86 31 .* 19P4 4 m* 32 �46 c 41 576 82 27 .* 16P3 �11 m* 37 �80 52 567 99 24 .* 13P2 �38 32 �88 49 576 136 33 .* 17P1 �47 26 �75 34 625 123 42 . 22
D1 �27 23 �70 29 586 126 55 24D2 �29 23 �75 33 594 125 55 24D3 11 m# 28 �82 38 548 130 44 .# 16D4 28 m# 23 �65 25 513 m 96 19 .# 15D5D6D7
L1 35 m 20 �85 20 349 m 43 11 . 24L2L3L4L5L6
Mean and standard deviations. Within muscle analysis: significant differ-ences (p < 0.05): * to segment P1; # to segment D2. Between muscleanalysis: significant differences (p < 0.05) : m to segment P1; c to segmentP2; . to segment D1. Movement time (FcOn to FcPeak) of 408 ms.
68 J.M.M. Brown et al. / Journal of Electromyography and Kinesiology 17 (2007) 57–73
all appeared to have a prime mover function (Fig. 6 andTable 4). This notable exception will be discussed furtherbelow.
We have also noted in the results that muscle segmentswith antagonist moment arms, whether located within anantagonist muscle (e.g. D1–D6 Deltoid during shoulderadduction) or as a single segment within an agonist muscle(e.g. D7 during shoulder abduction), had characteristicperiods of activation (Figs. 4, 6, 9 and 11).
Fig. 11. Shoulder Flexion. Intensity of muscle segment contraction in LatiDiagram based on group mean data. Note that the highest intensities of musclePectoralis Major. Also note the unique antagonist role played by muscle segm
In general, antagonist muscle segments tended to havedelayed onsets, shorter periods of activity and lower con-traction intensities than the prime mover segments (e.g.D7 in shoulder abduction; D6 during shoulder adductionand L1 during shoulder flexion) (Tables 3–5). An exceptionwas segment P3 (Pectoralis Major) during shoulder exten-sion which was co-activated with the prime mover seg-ments of the Latissimus Dorsi (Table 6). This was anunexpected result given that numerous reports [11,15] of‘‘triphasic’’ muscle activation patterns during rapid isomet-ric motor tasks have suggested that the antagonist (Ant)muscle burst is always activated after the initial agonist(Ag1) period of activation.
However, the involvement of antagonist muscle seg-ments in these rapid isometric motor tasks is consistentwith the findings of Flanders and Soechting [6] who statethat antagonist muscle activity, in part, is important tocounteract internal joint forces which may cause tissuedamage.
In general these results, obtained from the superficialshoulder muscle group, broadly support the previous find-ings of Wickham and Brown [24] for the Deltoid muscle.These findings, with notable exceptions to these generalisa-tions, will now be discussed in more detail with reference toeach movement direction.
4.3. Influence of movement direction
The most simple motor control strategies were employedby the CNS during the shoulder abduction task (Table 3).Here the middle muscle segments of the Deltoid (D3 andD4) were found to have the most favourable mechanicallines of action and the greatest moment arms to produceshoulder abduction in the coronal plane. Shoulder abduc-tion was initiated by activation of the prime mover seg-
ssimus Dorsi (L1–L6), Deltoid (D1–D7) and Pectoralis Major (P1–P7).segment contraction were seen in adjacent segments of the Deltoid and theent L1.
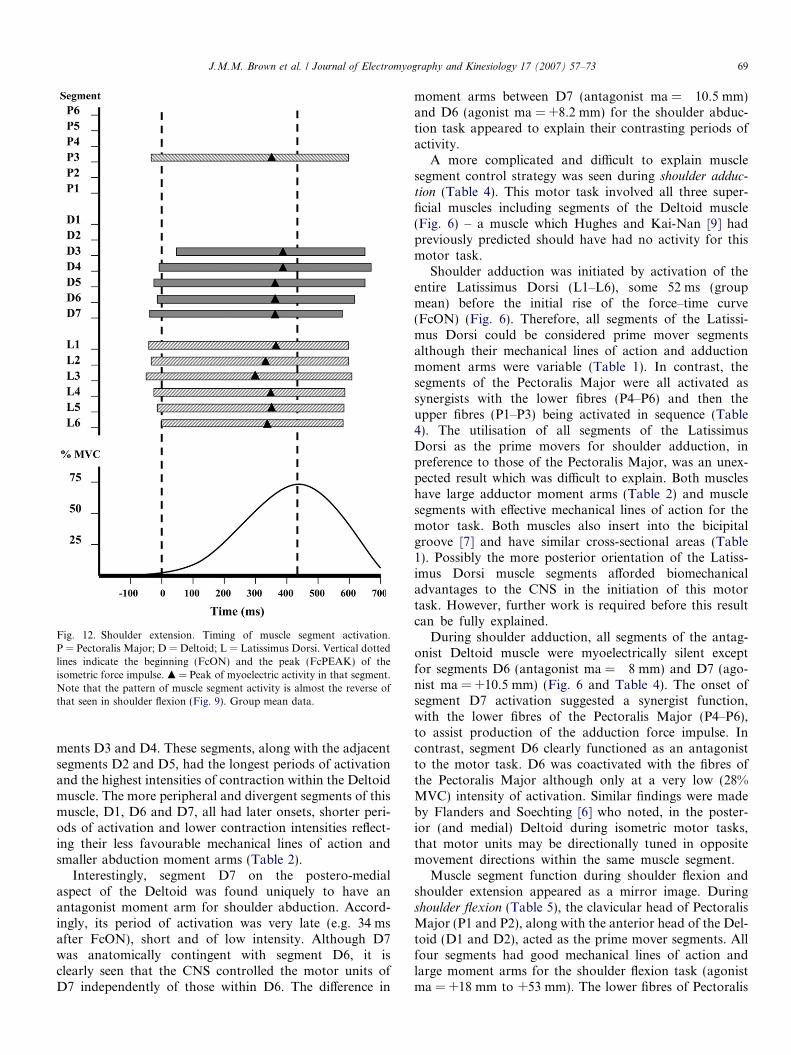
Fig. 12. Shoulder extension. Timing of muscle segment activation.P = Pectoralis Major; D = Deltoid; L = Latissimus Dorsi. Vertical dottedlines indicate the beginning (FcON) and the peak (FcPEAK) of theisometric force impulse. m = Peak of myoelectric activity in that segment.Note that the pattern of muscle segment activity is almost the reverse ofthat seen in shoulder flexion (Fig. 9). Group mean data.
J.M.M. Brown et al. / Journal of Electromyography and Kinesiology 17 (2007) 57–73 69
ments D3 and D4. These segments, along with the adjacentsegments D2 and D5, had the longest periods of activationand the highest intensities of contraction within the Deltoidmuscle. The more peripheral and divergent segments of thismuscle, D1, D6 and D7, all had later onsets, shorter peri-ods of activation and lower contraction intensities reflect-ing their less favourable mechanical lines of action andsmaller abduction moment arms (Table 2).
Interestingly, segment D7 on the postero-medialaspect of the Deltoid was found uniquely to have anantagonist moment arm for shoulder abduction. Accord-ingly, its period of activation was very late (e.g. 34 msafter FcON), short and of low intensity. Although D7was anatomically contingent with segment D6, it isclearly seen that the CNS controlled the motor units ofD7 independently of those within D6. The difference in
moment arms between D7 (antagonist ma = �10.5 mm)and D6 (agonist ma = +8.2 mm) for the shoulder abduc-tion task appeared to explain their contrasting periods ofactivity.
A more complicated and difficult to explain musclesegment control strategy was seen during shoulder adduc-tion (Table 4). This motor task involved all three super-ficial muscles including segments of the Deltoid muscle(Fig. 6) – a muscle which Hughes and Kai-Nan [9] hadpreviously predicted should have had no activity for thismotor task.
Shoulder adduction was initiated by activation of theentire Latissimus Dorsi (L1–L6), some 52 ms (groupmean) before the initial rise of the force–time curve(FcON) (Fig. 6). Therefore, all segments of the Latissi-mus Dorsi could be considered prime mover segmentsalthough their mechanical lines of action and adductionmoment arms were variable (Table 1). In contrast, thesegments of the Pectoralis Major were all activated assynergists with the lower fibres (P4–P6) and then theupper fibres (P1–P3) being activated in sequence (Table4). The utilisation of all segments of the LatissimusDorsi as the prime movers for shoulder adduction, inpreference to those of the Pectoralis Major, was an unex-pected result which was difficult to explain. Both muscleshave large adductor moment arms (Table 2) and musclesegments with effective mechanical lines of action for themotor task. Both muscles also insert into the bicipitalgroove [7] and have similar cross-sectional areas (Table1). Possibly the more posterior orientation of the Latiss-imus Dorsi muscle segments afforded biomechanicaladvantages to the CNS in the initiation of this motortask. However, further work is required before this resultcan be fully explained.
During shoulder adduction, all segments of the antag-onist Deltoid muscle were myoelectrically silent exceptfor segments D6 (antagonist ma = �8 mm) and D7 (ago-nist ma = +10.5 mm) (Fig. 6 and Table 4). The onset ofsegment D7 activation suggested a synergist function,with the lower fibres of the Pectoralis Major (P4–P6),to assist production of the adduction force impulse. Incontrast, segment D6 clearly functioned as an antagonistto the motor task. D6 was coactivated with the fibres ofthe Pectoralis Major although only at a very low (28%MVC) intensity of activation. Similar findings were madeby Flanders and Soechting [6] who noted, in the poster-ior (and medial) Deltoid during isometric motor tasks,that motor units may be directionally tuned in oppositemovement directions within the same muscle segment.
Muscle segment function during shoulder flexion andshoulder extension appeared as a mirror image. Duringshoulder flexion (Table 5), the clavicular head of PectoralisMajor (P1 and P2), along with the anterior head of the Del-toid (D1 and D2), acted as the prime mover segments. Allfour segments had good mechanical lines of action andlarge moment arms for the shoulder flexion task (agonistma = +18 mm to +53 mm). The lower fibres of Pectoralis
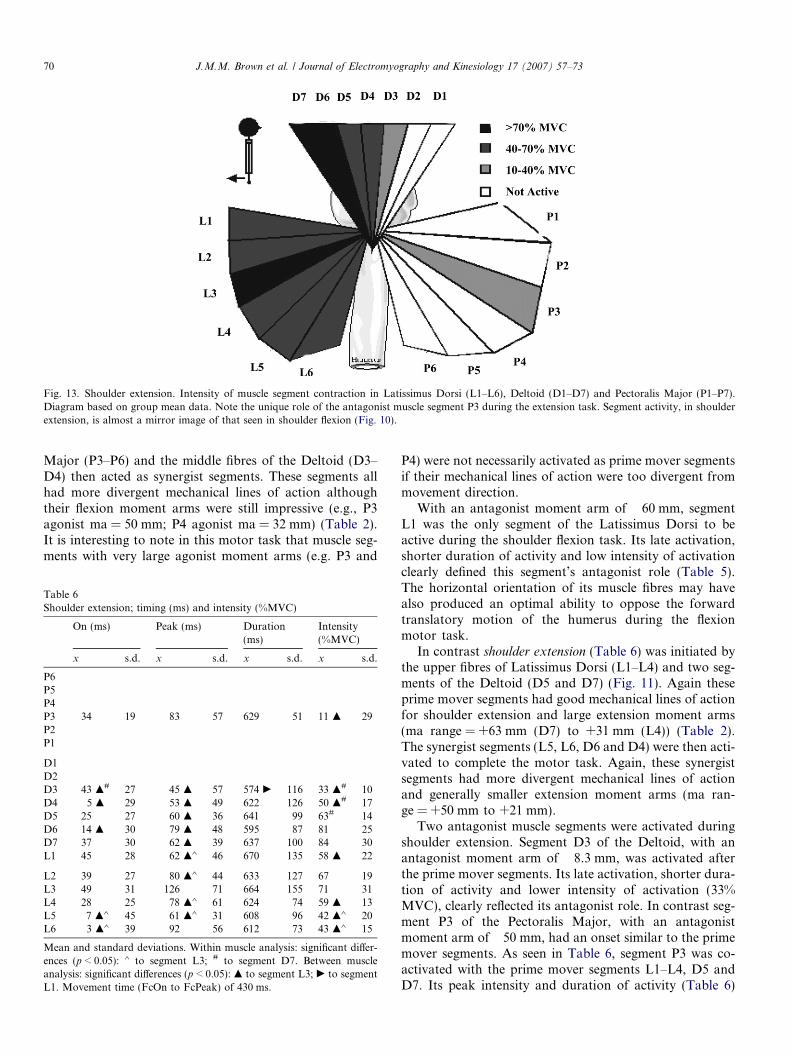
Fig. 13. Shoulder extension. Intensity of muscle segment contraction in Latissimus Dorsi (L1–L6), Deltoid (D1–D7) and Pectoralis Major (P1–P7).Diagram based on group mean data. Note the unique role of the antagonist muscle segment P3 during the extension task. Segment activity, in shoulderextension, is almost a mirror image of that seen in shoulder flexion (Fig. 10).
70 J.M.M. Brown et al. / Journal of Electromyography and Kinesiology 17 (2007) 57–73
Major (P3–P6) and the middle fibres of the Deltoid (D3–D4) then acted as synergist segments. These segments allhad more divergent mechanical lines of action althoughtheir flexion moment arms were still impressive (e.g., P3agonist ma = 50 mm; P4 agonist ma = 32 mm) (Table 2).It is interesting to note in this motor task that muscle seg-ments with very large agonist moment arms (e.g. P3 and
Table 6Shoulder extension; timing (ms) and intensity (%MVC)
On (ms) Peak (ms) Duration(ms)
Intensity(%MVC)
x s.d. x s.d. x s.d. x s.d.
P6P5P4P3 �34 19 �83 57 629 51 11 m 29P2P1
D1D2D3 43 m# 27 �45 m 57 574 c 116 33 m# 10D4 �5 m 29 �53 m 49 622 126 50 m# 17D5 �25 27 �60 m 36 641 99 63# 14D6 �14 m 30 �79 m 48 595 87 81 25D7 �37 30 �62 m 39 637 100 84 30L1 �45 28 �62 m� 46 670 135 58 m 22
L2 �39 27 �80 m� 44 633 127 67 19L3 �49 31 �126 71 664 155 71 31L4 �28 25 �78 m� 61 624 74 59 m 13L5 �7 m� 45 �61 m� 31 608 96 42 m� 20L6 3 m� 39 �92 56 612 73 43 m� 15
Mean and standard deviations. Within muscle analysis: significant differ-ences (p < 0.05): � to segment L3; # to segment D7. Between muscleanalysis: significant differences (p < 0.05): m to segment L3; c to segmentL1. Movement time (FcOn to FcPeak) of 430 ms.
P4) were not necessarily activated as prime mover segmentsif their mechanical lines of action were too divergent frommovement direction.
With an antagonist moment arm of �60 mm, segmentL1 was the only segment of the Latissimus Dorsi to beactive during the shoulder flexion task. Its late activation,shorter duration of activity and low intensity of activationclearly defined this segment’s antagonist role (Table 5).The horizontal orientation of its muscle fibres may havealso produced an optimal ability to oppose the forwardtranslatory motion of the humerus during the flexionmotor task.
In contrast shoulder extension (Table 6) was initiated bythe upper fibres of Latissimus Dorsi (L1–L4) and two seg-ments of the Deltoid (D5 and D7) (Fig. 11). Again theseprime mover segments had good mechanical lines of actionfor shoulder extension and large extension moment arms(ma range = +63 mm (D7) to +31 mm (L4)) (Table 2).The synergist segments (L5, L6, D6 and D4) were then acti-vated to complete the motor task. Again, these synergistsegments had more divergent mechanical lines of actionand generally smaller extension moment arms (ma ran-ge = +50 mm to +21 mm).
Two antagonist muscle segments were activated duringshoulder extension. Segment D3 of the Deltoid, with anantagonist moment arm of �8.3 mm, was activated afterthe prime mover segments. Its late activation, shorter dura-tion of activity and lower intensity of activation (33%MVC), clearly reflected its antagonist role. In contrast seg-ment P3 of the Pectoralis Major, with an antagonistmoment arm of �50 mm, had an onset similar to the primemover segments. As seen in Table 6, segment P3 was co-activated with the prime mover segments L1–L4, D5 andD7. Its peak intensity and duration of activity (Table 6)

J.M.M. Brown et al. / Journal of Electromyography and Kinesiology 17 (2007) 57–73 71
were also similar to the prime mover segments although itsintensity of activation was significantly (p < 0.05) lower at11% of MVC. It is difficult to explain why this particularantagonist muscle segment, the only segment of the Pecto-ralis Major to be activated during shoulder extension,should have such an early activation although its horizon-tal muscle fibre orientation may be implicated. Activationof the P3 segment may have been required early to protectthe anterior shoulder from injury during the vigorous acti-vation (approximately 73% of MVC group mean) of theprime mover segments; a role commonly given to antago-nist muscle action [6]. If this hypothesis is true, it is evidentthat the CNS has considerable flexibility regarding the tim-ing of antagonist muscle activity during the production ofmotor tasks.
With regard to the peak of muscle segment activation, itis clear that for all muscle segments involved in the fourmotor tasks investigated, maximal motor unit activationoccurred before the peak of the isometric force–time curve.In general, prime mover segments had earlier peak intensi-ties when compared to synergist segments although thiswas not always the case. There was, however, some limitedevidence to suggest that the peak activation of some antag-onist muscle segments was significantly (p < 0.05) laterthan the agonist muscle segments (Table 3).
4.4. Motor unit task groups
It was evident that to produce each of the four move-ment tasks, motor unit task groups were formed bothwithin, and where appropriate, across the three shouldermuscles, and then activated (timing and intensity) sequen-tially to produce the movement task. Motor unit taskgroups may be defined as a group of motor units, fromwithin a muscle or across several muscles, which are activesimultaneously for a particular motor task [14]. It isthought that differences in moment arms, or physiologicalfactors such as fibre type, may explain the existence ofmotor unit task groups, and therefore functionally inde-pendent muscle segments within muscle [5].
In the present study, each motor task was accomplishedthrough the formation of at least two, and up to four, dif-ferent motor unit task groups which often comprisedmotor units from adjacent, and more distant, muscles.For example, during shoulder abduction two motor unittask groups were formed within the Deltoid to achievethe motor task. The first motor unit task group formedincluded the prime mover segments (D3, D4), a commonfeature in all movement tasks in this study. A second motorunit task group was then formed by the remaining synergistand antagonist segments (D1, D2, D5, D6 and D7) whichall had smaller abduction moment arms, or in the case ofD7, an adduction moment arm. Contrary to the hypothesisof Flanders and Soechting [6], there was no evidence tosuggest that the motor units of the middle (D3) and poster-ior (D4–D7) Deltoid were tightly coupled during thismotor task.
Shoulder adduction was accomplished with the forma-tion of three motor unit task groups. The first containedall motor units within the Latissimus Dorsi muscle whichacted together as the prime movers for the motor task. Asecond group was formed by motor units within the lowerfibres of Pectoralis Major along with the agonist segmentD7 of the Deltoid. A third group was formed by motorunits within the upper fibres of the Pectoralis Major alongwith the antagonist D6 segment of the Deltoid. Fig. 8clearly shows the three motor unit task groups and con-firms that the activation of segments (motor units) withineach task group was sequential away from muscle segmentswith higher agonist moment arms.
Two motor unit task groups were formed during shoul-der flexion while three were formed during shoulder exten-sion. In both movement tasks, the first group contained theprime mover segments while the second comprised syner-gist segments. All motor unit task groups contained motorunits from a number of muscles. The antagonist segmentseither formed their own motor unit task group (shoulderextension) or were distributed either to the first (shoulderextension) or the second (shoulder flexion) motor unit taskgroup.
4.5. Muscle segment function and the CNS
The production of muscle force is a product ofmoment arm, muscle size, the muscle’s length tension rela-tionship as well as the timing and intensity of motor unitactivation; the latter controlled by the CNS [12]. Ourresults support the suggestion that the CNS, through itsupper and lower motoneurons, has the flexibility to ‘‘finetune’’ and coordinate the timing and intensity of motorunit activity both within a single muscle and betweenadjacent and more distant muscles, to meet the intendedgoals of the motor task.
It has been previously demonstrated, in monkey cortex,that pyramidal tract neurons from layer 5 of M1 (the pri-mary motor cortex) directly innervate lower motor neuronsin layer IX of the spinal cord [20]. These are the motoneu-rons which have direct control over the activation of amuscle’s constituent fibres.
Given that there is a proven relationship between theactivity of upper motoneurons within a cortical column,and the subsequent EMG activity within an active muscle[17], the results of this study may be used to gain furtherinsight into the strategies used by the CNS to control vol-untary movement.
Specifically, our study supports the contention that spe-cific motor columns within the human M1 (primary motorcortex), have the ability to independently control discretesubpopulations of motor units within a single skeletal mus-cle. This has previously only been confirmed in animalstudies [13,17]. Therefore, our results give further insightinto the complexity of the task faced by the CNS to controleach and every muscle involved in even the most simple ofmotor tasks.

72 J.M.M. Brown et al. / Journal of Electromyography and Kinesiology 17 (2007) 57–73
4.6. Limitations
Several limitations in the experimental design may haveinfluenced the results.
Firstly, it should not be concluded that the 19 musclesegments identified in this study represented the ‘‘ultimate’’design imposed by the CNS. In some instances (e.g. Latiss-imus Dorsi), the anatomical borders of each muscle seg-ment were found by simple division of relativelyhomogenous muscle tissue. Even when the muscle segmentswere clearly defined anatomically (e.g. Deltoid), it may bepossible that each segment could be further subdivided intoever smaller functional units. It is not beyond possibilitythat the ‘‘ultimate muscle segment’’ would be representedby a single muscle spindle and its associated muscle fibers!However, it remains probable that similar findings, tothose found here, would have been found if fewer, or more,muscle segments had been investigate. The exciting possi-bility now exists for the application of a multi-electrodefine-wire EMG technique, to an in vitro analysis of animalmuscle, whereby more muscle segments may be investi-gated and greater clarity provided to their neuromuscularcontrol.
A second limitation was the problem of crosstalkbetween adjacent bipolar electrode pairs. If detectable lev-els of crosstalk were present, the ability to differentiatebetween the activity of adjacent muscle segments wouldbe compromised. Considerable effort was made to mini-mize the effects of electrode crosstalk. The bipolar elec-trodes were specifically designed to have small activeplates (1.6 mm in diameter) and inter-electrode distances(6.5 mm). This electrode design, using the crosstalk ‘‘ruleof thumb’’ reported by Basmajian and DeLuca [1], wouldsuggest meaningful pickup of muscle fiber activity onlywithin approximately 6.5 mm of each bipolar electrodepair. Adjacent bipolar electrode pairs were generallyspaced more than 2 cm from each other so as to minimize,as much as practicable, electrode crosstalk. In addition,regardless of possible electrode crosstalk, the results pre-sented here provide strong evidence of the sequential acti-vation of adjacent muscle segments to control theseisometric tasks (Fig. 8). It would have been expected thathigh levels of electrode crosstalk would have negated thepossibility of this result.
5. Conclusion
The aim of this study was to determine how 19 musclesegments within the Latissimus Dorsi, Deltoid and Pecto-ralis Major, were controlled by the CNS to produce fourisometric shoulder motor tasks.
The results of this investigation have suggested that thetiming and intensity of each muscle segment’s activationwere coordinated across muscles and influenced by themuscle segment’s moment arm and its mechanical line ofaction in relation to the intended shoulder movement direc-tion (e.g. flexion, extension, abduction or adduction).
There was also evidence that motor unit task groups wereformed for individual motor tasks which comprised motorunits from both adjacent and distant muscles. In addition,the results supported the hypothesis that individual musclessegments may be functionally classified as prime mover,synergist or antagonist; functional classifications whichwere generally determined by the muscles segmentsmoment arm and/or mechanical line of action.
Acknowledgements
The authors wish to thank the University of Wollon-gong for technical and financial support to Dr. Wickhamduring his doctoral studies and to Dr. Brown during hissabbatical leave in Germany. Thank you also to Mr. LukeBones and Mr. Daniel Wickham for assistance with prepa-ration of the illustrations and to the reviewers for theexcellent comments and suggestions. The work describedin this study was approved by the Human Ethics Commit-tee of the University of Wollongong.
References
[1] Basmajian JV, DeLuca C. Muscles alive; their functions revealed byelectromyography. 5th ed. Baltimore: Williams & Wilkins; 1985.
[2] Brown JMM, Solomon C, Paton ME. Further evidence of functionaldifferentiation within biceps brachii. Electromyograph Clin Neuro-physiol 1993;33:301–9.
[3] Campbell KM, Norman BA, Biggs L, Blanton PL, Lehr RP.Electromyographic investigation of the relative activity amongfour components of the triceps surae. Am J Phys Med1973;52(1):30–41.
[4] Enoka RM. The neuromechanical basis of kinesiology, 2nd ed. In:Human kinetics, 1994.
[5] Ettema GJC, Styles G, Kippers V. The moment arms of 23 musclesegments of the upper limb with varying elbow and forearm positions:Implications for motor control. Human Movement Sci1998;17(2):201–20.
[6] Flanders M, Soechting JF. Arm muscle activation for static forces inthree-dimensional space. J Neurophysiol 1990;64(6):1818–37.
[7] Gray’s Anatomy. The anatomical basis of medicine and surgery.Sydney: Churchill Livingstone; 1990.
[8] Hogfors C, Sigholm G, Herberts P. Biomechanical model of thehuman shoulder – I. Elements. J Biomech 1987;20(2):157–66.
[9] Hughes RE, Kai-Nan A. Force analysis of rotator cuff muscles. ClinOrthopaed Related Res 1996;1(330):75–83.
[10] Jenkins DB. In: Hollinshead’s functional anatomy of the limbs andback. Sydney: W.B. Saunders; 1998.
[11] Karst GM, Hassan Z. Antagonist muscle activity during forearmmovements under varying kinematic and loading conditions. ExpBrain Res 1987;67:391–401.
[12] Kuechle DK, Newman SR, Itoi E, Niebur GL, Morrey BF, An K-N.The relevance of the moment arm of shoulder muscles with respect toaxial rotation of the glenohumeral joint in four positions. ClinBiomech 2000;15(5):322–9.
[13] Lemon R. The output map of the primate motor cortex. TrendsNeurosci 1988;11:501–6, 1989, erratum appears in Trends NeurosciApr;12(4):158..
[14] Loeb GE. Motorneuron task groups: Coping with kinematic heter-ogeneity. J Exp Biol 1985;115:137–46.
[15] MacKinnon CD, Rothwell JC. Time-varying changes in corticospinalexcitability accompanying the triphasic EMG pattern. J. Physiol.1998;506P:114P.

J.M.M. Brown et al. / Journal of Electromyo
[16] Manueddu Y, Blanc W, Taillard W. Study of the functioning of theGluteus Medius and Maximus: An electromyographic analysis. AnnKinesither 1989;16:193–201.
[17] Muir RB, Lemon RN. Corticospinal neurons with a special role inprecision grip. Brain Res 1983;61:312–6.
[18] Pare EB, Stern JR, Schwartz JM. Functional differentiation withinthe tensor fascia latae. J Bone Joint Surg A 1981;63(9):239–46.
[19] Paton ME, Brown JMM. An electromyographic analysis of func-tional differentiation in human pectoralis major muscle. J Electro-myograph Kinesiol 1994;4(3):161–9.
[20] Shinoda Y, Yokota J, Futami T. Divergent projection of individualcorticospinal axons to motoneurons of multiple muscles in themonkey. Neurosci Lett 1981;23:7–12.
[21] Soderberg GL, Dostal WF. Electromyographic study of three parts ofthe gluteus medius muscle during functional activities. Phys Therapy1978;58(6):691–6.
[22] Veeger HEV, VanDerHelm FCT, VanDerWoude LHV, Pronk GM,Rozendal RH. Inertia and muscle contraction parameters formusculoskeletal modelling of the shoulder mechanism. J Biomech1991;24(7):615–29.
[23] Wickham JB. Muscles within Muscles: The neuromotor activationpatterns of intramuscular segments. PhD thesis, Department ofBiomedical Science, University of Wollongong, 2002.
[24] Wickham JB, Brown JMM. Muscles within muscles: the neuromotorcontrol of intra-muscular segments. Eur J Appl Physiol 1998;78:219–25.
[25] Wickham JB, Brown JMM, McAndrew DJ. Muscles within muscles:Anatomical and functional segmentation of selected shoulder jointmusculature. J Musculoskeletal Res 2004;8(1):57–73.
Mark Brown has a Doctoral qualification fromthe University of Queensland and is currently aSenior Lecturer and Assistant Dean at theUniversity of Wollongong. He was Head of theDepartment of Biomedical Science between1997 and 2001 and is the immediate past VicePresident (exercise science) of Australia’s pro-fessional Exercise Science association (AAESS).His research interests include functional seg-mentation of skeletal muscle and sustainabletransport.
James Wickham received a Bachelor of SportsScience (exercise science) from the Universityof New South Wales in 1992 and a Diplomaof Education (physical education) in 1993from the same institution. In 1995 he receivedan Honours Degree from the University ofWollongong and a PhD in 2002. Since 1999James has been employed as an AnatomyLecturer at La Trobe University in Mel-bourne. His research interests include usingelectromyography to quantify shoulder muscleactivation patterns and functional differentia-
tion within skeletal muscles.
graphy and Kinesiology 17 (2007) 57–73 73
Darryl McAndrew received a Bachelor of Sci-ence (Human Movement Science) from theUniversity of Wollongong in 1993 and is cur-rently an Associate Lecturer within theDepartment of Biomedical Science, UoW. He isconcurrently completing a PhD focusing on thefibre type characteristics of segmental muscleand a M.Sc. in OH & S, specialising in Occu-pational Hygeine. His research interests includeidentifying muscle fibre type characteristics viamechanomyography and occupational exposureto industrial noise.
Xu-Feng Huang received his PhD degree from
the University of New South Wales in 1992. He is currently an Associate Professor in theDepartment of Biomedical Science and theDirector of Neurobiology Research Centre forMetabolic and Psychiatric Disorders in theUniversity of Wollongong. His research interestincludes the central regulation of energybalance.




























