Embed Size (px)
Citation preview

Mutations of Tubulin Glycylation Sites Reveal Cross-talk betweenthe C Termini of �- and �-Tubulin and Affect the Ciliary Matrixin Tetrahymena*
Received for publication, July 22, 2004, and in revised form, October 6, 2004Published, JBC Papers in Press, October 18, 2004, DOI 10.1074/jbc.M408324200
Virginie Redeker,a,b,c Nicolette Levilliers,c,d Emilie Vinolo,a,e Jean Rossier,a Danielle Jaillard,f
Dylan Burnette,g,h Jacek Gaertig,g and Marie-Helene Bred,i
From the aEcole Superieure de Physique et de Chimie Industrielles de la Ville de Paris, Laboratoire de Neurobiologie,UMR 7637 CNRS, 10 rue Vauquelin, 75005 Paris, France, the dLaboratoire de Biologie Cellulaire 4, UMR 8080 CNRS,Universite Paris-Sud, 91405 Orsay cedex, France, the fCentre Commun de Microscopie Electronique, Universite Paris-Sud,91405 Orsay cedex, France, and the gDepartment of Cellular Biology, University of Georgia, Athens, Georgia 30602
Two types of polymeric post-translational modifica-tions of �/�-tubulin, glycylation and glutamylation, oc-cur widely in cilia and flagella. Their respective cellularfunctions are poorly understood. Mass spectrometryand immunoblotting showed that two closely relatedspecies, the ciliates Tetrahymena and Paramecium,have dramatically different compositions of tubulinpost-translational modifications in structurally identi-cal axonemes. Whereas the axonemal tubulin of Parame-cium is highly glycylated and has a very low glutamyla-tion content, the axonemal tubulin of Tetrahymena isglycylated and extensively glutamylated. In addition,only the �-tubulin of Tetrahymena undergoes detyrosi-nation. Mutations of the known glycylation sites in Tet-rahymena tubulin affected the level of each polymericmodification type in both the mutated and nonmutatedsubunits, revealing cross-talk between �- and �-tubulin.Ultrastructural analyses of glycylation site mutants un-covered defects in the doublet B-subfiber of axonemesand revealed an accumulation of dense material in theciliary matrix, reminiscent of intraflagellar transportparticles seen by others in Chlamydomonas. We proposethat polyglycylation and/or polyglutamylation stabilizethe B-subfiber of outer doublets and regulate the in-traflagellar transport.
Microtubules are subject to a set of post-translational modi-fications (PTMs)1 whose significance has emerged only recently(1–4). Among PTMs, two polymeric modifications, glutamyla-
tion and glycylation, substantially increase the heterogeneityof the �/�-tubulin heterodimer. These tubulin modifications,referred to as polyglutamylation and polyglycylation, corre-spond to the addition of a peptide polymer consisting of severalglutamates (5) or glycines (6) onto the �-carboxyl group of aglutamate of the primary sequence of tubulin. These two PTMswill be referred to as “polymodifications” throughout this re-port. Polymodifications generate peptide branches of variablelengths distributed on several glutamate acceptor sites in theC-terminal tails of �- and �-tubulin (7–9). Both polymodifica-tion types are enriched in flagella and cilia of protists andmetazoan cells (4) and were implicated in axoneme motility (10,11). Whereas polyglycylation is restricted to axonemes in theciliated and flagellated metazoan cells, polyglutamylation oc-curs in both axonemes and basal bodies (12–15). In ciliates,both polymodification types are not only present in cilia andbasal bodies (15–18), but also occur on the more dynamic in-tracytoplasmic microtubules (17, 19, 20). Ciliates assemble upto 17 types of distinct microtubular arrays in a single cell (19,21–23). In these highly differentiated cells, the microtubularnetworks are involved in nuclear divisions, intracellular trans-port, organelle positioning, and are associated with specializedorganelles that function in osmotic regulation, feeding, excre-tion, and locomotion. The �- and �-tubulins of ciliates arebiochemically heterogeneous (17, 20, 24), suggesting that struc-tural differences among tubulin isoforms are important in gen-erating functionally distinct types of microtubules in a singlecell. However, genetic and biochemical studies showed that inciliates only one or two gene isotypes of �- and �-tubulin formthe bulk of microtubules (6, 25–28). Therefore, the large num-ber of tubulin isoforms present in ciliates is mainly the result ofPTMs (see refs. 6 and 29). Thus, ciliates provide a favorablemodel for assessing the function of tubulin PTMs.
Recently, by means of site-directed mutagenesis, three andfive glycylation sites were identified in �- and �-tubulin ofTetrahymena, respectively (30). Whereas all three sites of �-tu-bulin are dispensable, similar sites in �-tubulin are essentialfor cell survival. Specifically, among the five sites on �-tubulin,single and double site mutations did not affect cells, but triplesite mutations resulted in either hypomorphic or lethal pheno-types, in a site-specific manner. Viable triple site �-tubulinmutants grow and move slowly and have occasional defects incytokinesis (30). The lethal mutants have disorganized andparalyzed axonemes and are completely blocked in cytokinesis,
* This work was supported by CNRS, the Association pour la Recherchecontre le Cancer, and the Universite Paris-Sud. Activities at the Univer-sity of Georgia were supported by the National Institutes of Health GrantGM54017 and National Science Foundation Grant 235826 (to J. G.). Thecosts of publication of this article were defrayed in part by the payment ofpage charges. This article must therefore be hereby marked “advertise-ment” in accordance with 18 U.S.C. Section 1734 solely to indicate thisfact.
b Present address: Laboratoire d’Enzymologie et de Biochimie Struc-turales, UPR 9063 CNRS, 91198 Gif-sur-Yvette, France.
c The authors contributed equally to the work.e Present address: Unite de Regulation Enzymatique des Activites
Cellulaires, URA 2185 CNRS, Institut Pasteur, 75724 Paris Cedex15, France.
h Present address: Dept. of Molecular, Cellular and DevelopmentalBiology, Yale University, New Haven, CT 06520.
i To whom correspondence should be addressed: Tel.: 33-(0)169-156-480; Fax: 33-(0)169-156-803; E-mail: [email protected].
1 The abbreviations used are: PTM, post-translational modification;OD, outer doublet; IFT, intraflagellar transport; WT, wild type; mAb,monoclonal antibody; HPLC, high-performance liquid chromatography;
MALDI-TOF, matrix-assisted laser desorption ionization time-of-flight;MS, mass spectrometry; MAP, microtubule-associated protein.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 280, No. 1, Issue of January 7, pp. 596–606, 2005© 2005 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org596
by guest on February 8, 2016http://w
ww
.jbc.org/D
ownloaded from

before their death. The axonemes in the lethal mutants lackthe central pair and contain peripheral singlets instead ofdoublets (31).
The five glycylatable glutamates in �-tubulin of Tetrahy-mena, deduced from site-directed mutagenesis and immuno-blotting (Glu437, Glu438, Glu439, Glu440, and Glu442), are homol-ogous to the glycylation sites identified directly by massspectrometry in Paramecium (9). Less is known about the sitelocation and function of glutamylation in both ciliate species.Immunological data showed the presence of glutamylated tu-bulin in ciliates (17). Because glycylation and glutamylationsites are located in proximity within the C-terminal tails of �-or �-tubulin (32–34), it is possible that the mutations of glycyl-ation sites of Tetrahymena (30, 31) also affect glutamylation.
Here, a structural analysis of WT and mutated Tetrahymenatubulins was undertaken to systematically characterize PTMson each tubulin subunit and to determine how mutations of theknown glycylation sites affect the level and the extent of eachpolymodification type.
By mass spectrometry and immunoblotting, we show that, incontrast to axonemal tubulin of Paramecium, which exhibits aquantitatively minor polyglutamylation (6, 17), the Tetrahy-mena �/�-tubulin is extensively polyglutamylated. Noticeably,the mutations of the known glycylation sites not only affectedglycylation, but also glutamylation on the same tubulin sub-unit. Unexpectedly, mutations of glycylation sites on one tubu-lin subunit (� or �) affected the levels of polymodifications onthe nonmutated subunits of the �/�-tubulin dimers. Thus,cross-talk between �- and �-tubulin exists, which affects thetotal levels of polymodifications on microtubules. Our ultra-structural analyses of cilia of the viable polymodification mu-tants showed that the B-subfiber of the outer doublets (ODs) isstrongly affected. Furthermore, the glycylation site mutantsaccumulate electron-dense aggregates, similar to the in-traflagellar transport (IFT) particles of Chlamydomonas, sug-gesting that tubulin polymodifications are involved in IFT.
MATERIALS AND METHODS
Cell Culture—Paramecium tetraurelia and Tetrahymena ther-mophila strains were grown as described previously (20, 29).
Tetrahymena Mutants—Site-directed mutagenesis of the major�-tubulin of Tetrahymena encoded by the ATU1 gene (Atu1p) and themajor �-tubulin encoded by the BTU1 gene (Btu1p) was performedas described before (30). In T. thermophila, the wild type (WT)sequence of the �-tubulin C-terminal tail domain is 427DATAEEEGE-FEEEEGEN443, with the known polyglycylation sites underlined. In thepresent work, viable �-tubulin mutants with a triple substitution ofadjacent glutamates (E) by aspartate (D) (30) or alanine (A) residueswere used. The tail sequences of these mutants are 427DATAEEEG-EFEDDDGEN443 (the mutant named �EDDD440) and 427DATAEEEG-EFEAAAGEN443 (�EAAA440). A quadruple glycylation site �-tubulinmutant with the tail sequence 427DATAEEEGEFEDDDGDN443 (�ED-DDGD442) was also used. For �-tubulin, the WT C-terminal tail se-quence is 437IETAEGEGEEEGY449, with the known polyglycylationsites underlined, whereas the tail sequence of the viable mutant usedhere is 437IETAEGEGAAAGY449 (�AAA447) (30).
Preparation of Axonemal Tubulin—The axonemal tubulin of Para-mecium was extracted from cilia as described previously (35). WT andmutant Tetrahymena cilia were prepared as described (30), and axon-emal tubulin was extracted exactly as done for Paramecium.
Protein Electrophoresis and Immunoblotting—Axonemal proteinswere separated by SDS-PAGE on 10% polyacrylamide mini-gels (36),containing 0.1% (wt/wt) SDS (99% pure, BDH, Poole, UK) at pH 8.3,according to Suprenant et al. (24). Under these conditions, the Parame-cium or Tetrahymena �-tubulin migrates faster than �-tubulin. Pro-teins were transferred onto nitrocellulose by the method of Kyhse-Andersen (37). The blots were stained with Ponceau red, washed inantibody buffer, and then incubated overnight with one of the followingprimary antibodies: TAP 952 or AXO 49 monoclonal antibodies (mAbs),raised against Paramecium axonemal tubulin (38) and directed againstmono- and polyglycylated tubulins, respectively (20); GT335 mAb di-rected against glutamylated tubulin (39); TAP 9311 (38) or DM1A (40;
purchased from Amersham Biosciences) mAbs, directed against N- andC-terminal sequences of �-tubulin, respectively; C140 (41) polyclonalantibodies directed against an N-terminal sequence of �-tubulin. Afterextensive washing, blots were incubated with peroxidase-labeled sheepanti-mouse and donkey anti-rabbit IgG antibodies and processed forenhanced chemiluminescence (ECL) (Amersham Biosciences). TheGT335 and C140 antibodies were kindly provided by Dr. P. Denoulet(University of Paris VI, France) and Dr. J. M. Andreu (Centro deInvestigaciones Biologicas, Madrid, Spain), respectively.
Proteolytic Digestion of Axonemal Tubulins—Axonemal tubulin, typ-ically 0.2–0.5 mg, was digested with the endoproteinase Asp-N (se-quencing grade protease, Roche Applied Science) at an enzyme to tu-bulin protein ratio of 1:400 (w/w) in 50 mM Tris-HCl, pH 8.5, at 37 °C for6 h. After digestion, the peptide mixture was frozen at �20 °C. Prior tochromatography, the peptide sample was briefly sonicated and clarifiedby centrifugation at 12,000 � g for 2 min.
Purification of the C-terminal Tubulin Peptides—The endoproteinaseAsp-N digestion of Tetrahymena tubulin produces the C-terminal pep-tides of �- and �-tubulin beginning with an aspartate at positions 431and 427, respectively, when the digestion is complete. Following incom-plete digestion, longer �-tubulin C-terminal peptides were generated,beginning with an aspartate at position 424. The C-terminal peptideswere separated on an arginine-Sepharose column and either desaltedon a C18 Sep-Pak cartridge prior to direct analysis of the total peptidepools, as previously described (7), or further separated by reverse-phaseHPLC. In the latter case, about 100 �g of digested tubulin was injectedinto a C18 reverse-phase HPLC column (5 �m, 250 � 1 mm, HypersilBDS-C18, flow rate of 200 �l/min). Peptides were eluted with a gradientof solvent A (0.1% trifluoroacetic acid) and solvent B (80% acetonitrile,0.1% trifluoroacetic acid) consisting of 1% solvent B for 10 min, 1% to5% solvent B in 1 min, 5% to 50% solvent B in 60 min, and 50% to 100%solvent B in 10 min. Peptide elution was monitored at 214 nm.
MALDI-TOF Mass Spectrometry—Mass spectra were acquired in thelinear mode on a MALDI-TOF mass spectrometer (Voyager-STR, Per-septive Biosystems, Inc., Framingham, MA) equipped with a delayedextraction device. Desorption was produced by a nitrogen laser beam(� � 337 nm). Delayed extraction time was set at 200 ns. The samplewas mixed 1:1 (v:v) with a saturated solution of either sinapinic acid(3,5-dimethoxy-4-hydroxicinnamic acid, Aldrich) in 30% acetonitrile,0.1% aqueous trifluoroacetic acid or 2,5-dihydroxybenzoic acid (Aldrich)in 0.1% aqueous trifluoroacetic acid, and analyzed in the negative orpositive ion mode, respectively. Except when indicated, the mass spec-tra shown were recorded in linear and positive ion modes, with 2,5-dihydroxybenzoic acid as the matrix. External calibration was per-formed using a mixture of neurotensin, adrenocorticotropic hormone(residues 18–39) and adrenocorticotropic hormone (residues 7–38) withaverage m/z of 1671.95, 2464.71, and 3658.17, respectively, for thedeprotonated ions in the negative ion mode, and of 1673.95, 2466.71,and 3660.17 for the protonated ions.
Transmission Electron Microscopy—Cells were washed with 10 mM
Tris, pH 7.4, and fixed in 1% glutaraldehyde in 50 mM cacodylate buffer,pH 7.4, at room temperature for 1 h, washed three times in cacodylatebuffer, and postfixed in 1% osmium tetroxide in cacodylate buffer for 1 hat room temperature. After three washes in cacodylate buffer, the cellswere embedded in 2.5% agarose and dehydrated before inclusion inEpon. Thin sections were obtained with a Leica ultramicrotome andcontrasted with uranyl acetate and lead citrate. Sections were observedwith a Philips 208 electron microscope.
FIG. 1. Western blot analysis of axonemal proteins of P. tetrau-relia (P) and T. thermophila (T) probed with TAP 952 and AXO49 mAbs (1: 20,000), specific for mono- and polyglycylated tubu-lin, respectively, and with an anti �-tubulin mAb, TAP 9311(1: 500).
Tubulin Post-translational Modifications and Cilia Integrity 597
by guest on February 8, 2016http://w
ww
.jbc.org/D
ownloaded from

RESULTS
Post-translational Modifications of Axonemal Tubulins ofParamecium and Tetrahymena—We examined the status ofpost-translational modifications of the C-terminal peptidesof axonemal tubulin extracted from WT Tetrahymena and com-pared the Tetrahymena data with those of the already exten-sively characterized axonemal tubulin of Paramecium.
Western blots with TAP 952 and AXO 49 mAbs, which rec-ognize mono- and polyglycylated tubulins, respectively, showedthat the axonemal tubulins of both ciliates contain mono- andpolyglycylated sites in both subunits (Fig. 1). Note that, underconditions described under “Materials and Methods,” ciliate�-tubulins migrate faster than �-tubulins (24). Noticeably, a
striking difference in reactivity with the anti-glutamylated tu-bulin mAb, GT335, indicated the presence of a higher level ofglutamylation in Tetrahymena than in Paramecium (Fig. 2).
We purified the C-terminal tubulin peptides from cilia of bothspecies by arginine-Sepharose chromatography (7), following theendoproteinase Asp-N digestion. An analysis of the resultingtotal peptide pools by MALDI-TOF mass spectrometry gave abroad picture of the modifications of both �- and �-tubulin C-terminal peptides (Fig. 3). This approach shows the diversity andreveals the most abundant peptide variants. Fig. 3 compares theC-terminal peptide pools of Paramecium (panels a and b) andTetrahymena (panels c and d) axonemal tubulins. Using Asp-N,�-tubulins from both Paramecium and Tetrahymena werecleaved at the N-terminal side of Asp427, whereas �-tubulin wascleaved mainly at Asp424, but also less often at Asp431, generating“long” and “short” � peptides, respectively.
In the Paramecium spectrum (Fig. 3a), the predominant �
peptide (experimental average m/z � 2139.42) is a hexaglycy-lated variant (calculated average m/z � 2140.03). The otherions observed in this mass spectrum form a series with differ-ences of 57 Da, the mass of a single glycine residue. The rangeof masses of these isoforms indicates addition of 4–18 glycineresidues. In the spectrum shown in Fig. 3b, the long peptides of�-tubulin were observed. The two predominant peptides ofisotypes �1 and �2 are triglycylated, and additions of 1–19glycines were detected. A similar composition of the glycylated
FIG. 2. Western blot analysis of axonemal proteins of Parame-cium (P), Tetrahymena (T), WT, and �EDDD440 mutant (EDDD),probed with GT335 mAb (1:50,000), specific for glutamylatedtubulin, and with anti �-tubulin mAb, DM1A (1:4,000), and anti�-tubulin antibodies, C140 (1:4,000).
FIG. 3. Mass spectra of the C-terminal peptides of axonemal �- and �-tubulins from Paramecium and WT Tetrahymena. a and b, �-and �-tubulin peptides, respectively, from Paramecium. In panel a, the �-tubulin peptide series is observed together with short �-tubulin peptidesstarting at residue Asp431. In the latter �-tubulin peptides series, the first glycylated peptides are completely annotated with the isotypeidentification, �1 or �2, and the number of additional glycine residues. The other glycylated peptides of this series are marked by isotype pairs withan asterisk. A clearer mass spectrum of the �-tubulin peptides is presented in panel b, where only long Paramecium �-tubulin peptides startingat residue Asp424, in the mass range from 2800 to 3900, are observed. The two predominant peptides of isotypes �1 and �2 are triglycylated(experimental average m/z of 2909.31 for �1 and 2923.17 for �2; calculated average m/z of 2909.98 and 2924.00, respectively). c and d, �- and�-tubulin peptides, respectively, from WT Tetrahymena. In d, only long �-tubulin peptides were observed. Curved arrows indicate increments ofglycine units added to the nonmodified isoform (�0G) and to each glutamylated peptide series (1E, 2E, and so on). In these experiments, theC-terminal peptides of �- and �-tubulin were analyzed in positive and negative ion modes, respectively.
Tubulin Post-translational Modifications and Cilia Integrity598
by guest on February 8, 2016http://w
ww
.jbc.org/D
ownloaded from

�-tubulin peptides of axonemal tubulin of Paramecium wasobserved previously using a different purification scheme (9),whereas an HPLC separation allowed the detection of lessabundant peptides representing highly modified isoforms with19–32 additional glycine residues (6). The consistency of Para-mecium results obtained with two different methods validatesour experimental approach for a comparative study with theTetrahymena tubulins (see below).
In the mass spectrum obtained for Tetrahymena �-tubulin(Fig. 3c), the predominant molecular ion (experimental averagem/z � 2142.61) corresponds to the C-terminal peptide of thetetraglycylated �-tubulin (calculated average m/z � 2143.0).Other ions corresponding to addition of up to at least 12 glycineresidues were observed. Fig. 3d presents the molecular ions of
the Tetrahymena �-tubulin. This mass spectrum showed anunexpected complexity and revealed two types of ion series, onewith increments of 57 Da, the mass of a glycine residue, andone with increments of 129 Da, the mass of a glutamate resi-due. Thus, Tetrahymena �-tubulin is glycylated but also exten-sively glutamylated. A further dissection of the mass spectrumindicates that the first molecular ion (experimental averagem/z � 2682.06) corresponds to a long �-tubulin C-terminalpeptide starting at Asp424, which is also detyrosinated at its Cterminus and lacks additional modifications (calculated m/z �2681.75). The most abundant ions form a series correspondingto additions of 1 to at least 11 glutamates, with a predominanttriglutamylated peptide. Other ion series that appear to be lessabundant correspond to the addition of 1 to 5 or 6 glycineresidues to either the nonmodified peptide, or to peptides si-multaneously glutamylated at different levels. In these series,the nonglycylated form is predominant. We conclude that Tet-rahymena �-tubulin is glycylated, extensively glutamylated,and detyrosinated, and often two or three modification typesexist on the same tubulin molecule.
In addition to the peptide pool analyses, some apparentlyless abundant but highly modified isoforms were detected byMALDI-TOF MS after HPLC separation of the peptide pool.High levels of polymodifications were revealed by the massspectra of two HPLC fractions corresponding to �- and �-tubu-lin C-terminal peptides (Fig. 4, a and b). In Fig. 4a, minor formsof �-tubulin peptides were found to be glutamylated by additionof one to three glutamates, and each glutamylated peptide wasglycylated by four to six glycines. In this mass spectrum, as inthe spectrum of the peptide pool (see above), the tetraglycyl-ated �-tubulin isoform is the predominant ion. In anotherHPLC fraction, �-tubulin was found to be glutamylated withthe addition of up to 17 glutamate residues (Fig. 4b).
Table I compares the types of polymodifications detected inWT Paramecium and Tetrahymena in peptide pools and HPLCfractions. It appears that, in Paramecium, both �- and �-tubu-lin subunits are glycylated to a larger extent than in Tetrahy-mena. The most striking difference involves polyglutamylation.Whereas this modification was not detected in Paramecium byMS, even in minor HPLC fractions, glutamylation is the mostabundant modification of the Tetrahymena �-tubulin and canbe detected in HPLC fractions of the �-tubulin subunit as well.
Composition of Post-translational Modifications in the Axon-emal Tubulin from Glycylation Site Mutants of Tetrahymena—The structural approach described above was used to analyzethe post-translational modifications, and particularly the poly-modifications, in the tubulin C-terminal peptides of Tetrahy-
FIG. 4. Mass spectra of selected HPLC fractions of axonemal �-and �-tubulin peptides from WT Tetrahymena (a and b, respec-tively). In a, curved arrows indicate increments of glycine units in each� peptide series (nonglutamylated and glutamylated). b shows the massspectrum of long �-tubulin peptides that are glutamylated. One pep-tide, marked with an open triangle, has not been identified with cer-tainty. Its molecular mass could match with an internal �-tubulinpeptide (sequence 165–202), but also with a long C-terminal �-tubulinpeptide starting at residue Asp424 with 10 additional glutamates and 3additional glycines. This latter possibility seems less probable, becauseno other ions are associated to this peptide in a separate series. HPLC-purified peptides were analyzed in positive ion mode.
TABLE IPost-translational modifications identified in the arginine-Sepharose-purified C-terminal peptide pools and HPLC-purified fractions from
Paramecium and from WT and mutant Tetrahymena axonemal tubulinsNote that the �-tubulin C-terminal peptides of Tetrahymena were found to be detyrosinated.
Species Strains Sequences of the identified peptidesPeptide pools HPLC-purified fractions
Additionalglycines
Additionalglutamates
Additionalglycines
Additionalglutamates
Paramecium WT �:427DATAEEEGEFEEEGEQ442 4–18 (6maj) NDa 4–32b ND�1:431DYEEVGIETAEGEGEEGEG449 1–19 (3maj) ND 3–34b ND�2:431DYEEVGIETAEGEGEEGEA449 1–19 (3maj) ND 3–34b ND
Tetrahymena WT �:427DATAEEEGEFEEEEGEN443 4–12 (4maj) ND 0–12 0–7�:431DYEEVGIETAEGEGEEEG448 0–6 (0maj) 0–11 (3maj) 0–10 0–19
�EDDD440 �:427DATAEEEGEFEDDDGEN443 c 0–7 (1maj) 0–2 (0maj) 0–42 0–2�:431DYEEVGIETAEGEGEEEG448 0–26 (0maj) ND 0–37 ND
�EAAA440 �:427DATAEEEGEFEAAAGEN443 0–3 (0,1maj) ND�:431DYEEVGIETAEGEGEEEG448 0–8 (0maj) ND
�AAA447 �:427DATAEEEGEFEEEEGEN443 2–26 (4maj) ND�:431DYEEVGIETAEGEGAAAG448 No peptide No peptide
a ND, not detected.b See Ref. 6.c Underlined bold characters represent the mutated sites.
Tubulin Post-translational Modifications and Cilia Integrity 599
by guest on February 8, 2016http://w
ww
.jbc.org/D
ownloaded from

mena glycylation site mutants. Depending on the mutated sitelocation, triple mutants of the five known glycylatable sites in�-tubulin are either viable (hypomorphic) or nonviable (30).Purification of mutant tubulins from the nonviable mutants isnot feasible. Therefore, to examine how glycylation site muta-tions affect the composition of polymodifications, we used hy-pomorphic triple mutants having substitutions of glutamatesby aspartates or alanines in �-tubulin (�EDDD440 and�EAAA440). Like the �EDDD440 mutant (30), �EAAA440 grewand swam more slowly than the WT and underwent infrequentarrests in cytokinesis. We also analyzed a triple mutant of�-tubulin (�AAA447), which previously was found to have anormal phenotype (30).
Western blot analyses of WT axonemal tubulin showed thatthe AXO 49 mAb reacts more strongly with the polyglycylatedepitopes on �-tubulin than on �-tubulin (Fig. 5). In �EDDD440,the reactivity with the �-tubulin region was much weakercompared with WT. This is consistent with the previous quan-titative immunoblotting analysis, which showed that the levelof AXO 49 epitopes in �EDDD440 �-tubulin was only 24% ofWT, and supported the conclusion that sites of glycylation wereeliminated in this mutant (30). However, under the presentimproved gel electrophoresis conditions (see “Materials andMethods”), the AXO 49 mAb revealed a smear of slowly-migrat-ing isoforms in the region of mutant �-tubulin, contrasting withthe WT �-tubulin which appeared as a sharp band (Fig. 5).Note that in previous studies a correlation has been establishedbetween the presence of a smear of AXO 49 reactive isoformsand the presence of hyperglycylated variants (e.g. refs 10, 20).Thus, despite an overall decrease in the extent of glycylation of�-tubulin in the �EDDD440 mutant, a subset of isoforms may
contain abnormally long chains of polyglycine. This conclusionwas subsequently confirmed using MS (see below). In�EAAA440, both tubulin subunits were only faintly detected(Fig. 5), indicating the presence of a lower extent of polyglycy-lation compared with the WT and �EDDD440. In �AAA447,previous results showed an AXO 49 reactivity with the �-tubu-lin band only, leading to the conclusion that in this mutant allglycylation sites on �-tubulin were eliminated (30). Westernblots with GT335 showed that axonemal tubulins of �EDDD440
(Fig. 2), �EAAA440 and �AAA447 (not shown) were less reactivecompared with WT, indicating that all these mutations affectnot only glycylation but also glutamylation.
Post-translational modifications of axonemal tubulin of the�EDDD440 mutant were further characterized by mass spec-trometry. In the first step, the pools of �- and �-tubulin C-terminal peptides were analyzed by MALDI-TOF MS (Fig. 6).The mass spectrum of the �-tubulin peptides from the�EDDD440 mutant (Fig. 6a) was complex. The first molecularion (experimental average m/z � 1871.16) was identified as anonmodified �-tubulin peptide starting at residue Asp427 (cal-culated average m/z � 1871.02). The prominent series of ionscorresponds to glycylated forms with one to seven glycine res-idues added. The monoglycylated form represents the predom-inant peptide of this series and of the entire spectrum. A secondseries of ions corresponding to monoglutamylated �-tubulinpeptides was observed. The latter ions were detected in amonoglycylated form, which represents the most abundant ionof this series, and in bi-, tri-, and tetraglycylated form. Anadditional less abundant C-terminal peptide of �EDDD440
�-tubulin was both biglutamylated and monoglycylated. Thus,the MS analysis of peptide pools of �EDDD440 �-tubulin showsthat: (i) the predominant isoform in the mutant is monoglycy-lated, whereas it is tetraglycylated in WT; (ii) glutamylated�-tubulin is present in the mutant, whereas it was not detectedin similar pools of WT. This lack of detection in WT could beexplained by the difficulty of some peptides to desorb from thematrix when the overall heterogeneity of the peptide mixture isvery high. This ion suppression observed for some less abun-dant peptides present in a complex mixture has been reportedas spectral suppression (42). The sensitivity of detection isusually increased for less heterogeneous peptide mixtures. Thisexplains the detection of low abundance of peptides in HPLC-purified fractions, as is in fact the case for glutamylated WT�-tubulin peptides comprising one to seven additional gluta-mates (Table I).
In the mass spectrum of �-tubulin peptides from the
FIG. 5. Western blot analysis of axonemal proteins of Tetrahy-mena WT, �EDDD440 (EDDD), and �EAAA440 (EAAA) mutantsprobed with AXO 49 (1:10,000), specific for polyglycylated tubu-lin, and with an anti-�-tubulin mAb, DM1A (1:500).
FIG. 6. Mass spectra of the C-terminal peptides of axonemal �- and �-tubulins from �EDDD440 Tetrahymena mutant (a and b,respectively). In a, the mass spectrum clearly shows the �-tubulin C-terminal peptides series. Curved arrows indicate increments of glycine unitsin the monoglutamylated peptide series (1E). In this mass range, other peptides can be observed: the �s peptide corresponding to a shortnonmodified C-terminal �-tubulin peptide starting at residue Asp431; the �(327–344) peptide corresponds to an internal peptide of �-tubulin fromresidue 327 to residue 344. b shows the only long C-terminal peptide series of �-tubulin starting at residue Asp424. The peptides marked with anopen triangle were unidentified peptide ions. Peptides were analyzed in positive ion mode.
Tubulin Post-translational Modifications and Cilia Integrity600
by guest on February 8, 2016http://w
ww
.jbc.org/D
ownloaded from

�EDDD440 mutant (Fig. 6b), a unique series of ions was ob-served. It begins with the predominant ion (experimental av-erage m/z � 2682.13) corresponding to a detyrosinated, non-glycylated, and nonglutamylated �-tubulin peptide starting atresidue Asp424 (calculated average m/z � 2681.77). The follow-ing ions in the same series correspond to the addition of 1–26glycine residues. Even though the number of added glycineresidues is considerably higher compared with WT, the non-modified peptide is still predominant. The most striking differ-ence with the WT is the absence of glutamylated �-tubulinpeptides, whereas this modification is predominant in the WT.
The �EDDD440 tubulin peptides were also subjected toHPLC separation prior to MALDI-TOF MS. As shown in Fig. 7,for two HPLC fractions analyzed, high levels of glycylationwere observed for both �- (Fig. 7a) and �-tubulin (Fig. 7b)peptides with the detection of up to 42 and 37 additional gly-cine residues, respectively (Table I). These results are in agree-ment with the detection of a smear of AXO 49 immunoreactivetubulin isoforms in �EDDD440 (see Fig. 5), indicating the pres-ence of highly modified isoforms. Such hyperglycylated iso-forms were not observed in HPLC fractions of WT tubulinpeptides (Fig. 4). Thus, in addition to an overall decrease inglycylation on �-tubulin, hyperglycylation affects subsets of �-and �-tubulin isoforms in the �EDDD440 mutant.
In conclusion, for the �EDDD440 mutant, the most intriguingresult lies in the composition of the nonmutated �-tubulin.Whereas the predominant modification in WT �-tubulin is glu-tamylation, this modification was not detected in the�EDDD440 �-tubulin, even after HPLC purification. Further-more, the maximal number of added glycine residues increasesconsiderably in �EDDD440 �-tubulin. These data strongly sug-gest that mutations of polymodification sites in the C-terminaltail of �-tubulin induce extensive changes in the composition ofPTMs in the nonmutated �-tubulin subunit.
To examine further the phenomenon of cross-talk betweenthe �- and �-tubulin subunits, we analyzed additional Tetra-hymena mutants. First, another �-tubulin mutant, �EAAA440,was examined, in which the same three modification sites(mutated to aspartates in the �EDDD440 mutant) were substi-tuted by alanines.
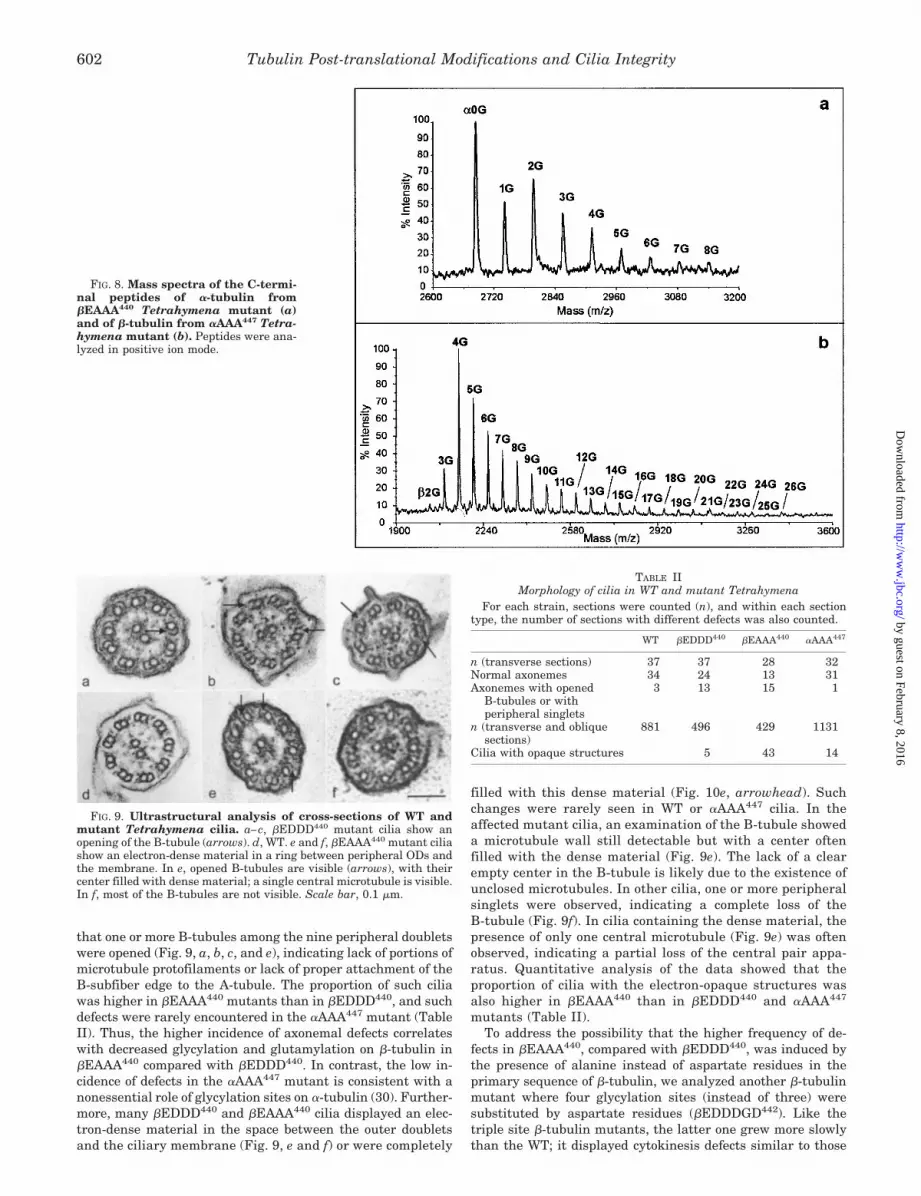
The mass spectrum of the �-tubulin C-terminal peptides ofthe �EAAA440 mutant (Fig. 8a) showed a series of ions withmass increments of 57 Da, the mass of one glycine residue. Thefirst ion of the series, which is also the predominant ion of themass spectrum (experimental average m/z � 2684.68) corre-sponds to a C-terminal detyrosinated and nonpolymodified�-tubulin peptide starting at residue Asp424 (calculated aver-age m/z � 2683.78). The additional ions correspond to thepresence of one to eight glycine residues. Thus, the maximalnumber of glycine residues is considerably lower comparedwith �EDDD440 �-tubulin. As in the case of the �EDDD440
mutant, we did not detect any glutamylation on the �-tubulinsubunit, confirming the general influence of the �-tubulin mu-tations on the composition of polymodifications in the nonmu-tated �-tubulin.
To determine whether the influence of the polymodificationsite status on the nonmutated subunit is reciprocal, we examined�-tubulin of a mutant of �-tubulin glycylation sites, �AAA447.Previous study showed that the glycylation sites on �-tubulin arenot essential and their absence does not change the gross phe-notype (30). The mass spectrum of �-tubulin C-terminal peptidesof the �AAA447 tubulin mutant (Fig. 8b) also contained a series ofions with mass increments of 57 Da. The predominant �-tubulinC-terminal peptide (experimental average m/z � 2142.62) was atetraglycylated form (calculated average m/z � 2143.02), and thewhole series of glycylated isoforms reflects additions of 2–26
glycine residues. The maximal number of additional glycine res-idues in the �-tubulin subunit is therefore higher in the �AAA447
mutant than in WT. Glutamylation was not detected in thepeptide pool on �-tubulin of the �AAA447 mutant. Because glu-tamylation appears to be quantitatively minor also on WT �-tu-bulin, an effect of the �AAA447 mutation on glutamylation of thenonmutated � subunit could be below the limit of detection.Therefore, the only noticeable effect observed following the mu-tation of polymodification sites in the C-terminal tail of �-tubulinis an increase in the maximal glycylation level of the nonmutated�-tubulin subunit.
Ultrastructural Analysis of Cilia in Glycylation Site MutantCells—Previous studies showed severe defects in the structureof axonemes in the nonviable �-tubulin mutant (�DDDE440),including lack of central pair and conversions of peripheraldoublets into singlets (31). The viable hypomorphic mutantsin which we analyzed the composition of modifications by MS(above) display slow motility and growth (30), but the ultra-structure of their cilia has not been studied. Because weobserved that the �EAAA440 mutant displays a more signif-icant loss of polymodifications compared with �EDDD440, wefurther tested whether any structural changes could be de-tected in cilia of viable mutants and whether the extent ofthese changes correlates with the extent of loss ofpolymodifications.
A transmission electron microscopy examination of thin sec-tioned cells revealed a class of cilia with profound defects in theaxonemal doublets. In a subpopulation of cilia from �EDDD440
and �EAAA440 mutants, cross-sections of the axonemes showed
FIG. 7. Mass spectra of selected HPLC fractions of axonemal �-and �- tubulin peptides from �EDDD440 Tetrahymena mutant (aand b, respectively). a, the mass spectrum shows an incredible het-erogeneity of the �-tubulin C-terminal peptides. The series identified as�s peptides corresponds to short �-tubulin peptides starting at residueAsp427 and ending at residue Glu437. The presence of the three gly-cylatable sites that were mutated from glutamate to aspartate explainsthis type of cleavage with the endoproteinase Asp-N. These short �-tu-bulin peptides are shown in an extensively glycylated form. The iden-tification of these peptides was confirmed by tandem mass spectrometryusing a nanoelectron spray quadrupole time-of-flight mass spectrome-ter. The ion series marked with an asterisk corresponds to short �-tu-bulin C-terminal peptides with one additional glutamate residue, andadditions of 28–42 glycine residues. b, highly glycylated forms of thelong C-terminal �-tubulin peptide starting at residue Asp424 are ob-served. Peptides were analyzed in positive ion mode.
Tubulin Post-translational Modifications and Cilia Integrity 601
by guest on February 8, 2016http://w
ww
.jbc.org/D
ownloaded from
that one or more B-tubules among the nine peripheral doubletswere opened (Fig. 9, a, b, c, and e), indicating lack of portions ofmicrotubule protofilaments or lack of proper attachment of theB-subfiber edge to the A-tubule. The proportion of such ciliawas higher in �EAAA440 mutants than in �EDDD440, and suchdefects were rarely encountered in the �AAA447 mutant (TableII). Thus, the higher incidence of axonemal defects correlateswith decreased glycylation and glutamylation on �-tubulin in�EAAA440 compared with �EDDD440. In contrast, the low in-cidence of defects in the �AAA447 mutant is consistent with anonessential role of glycylation sites on �-tubulin (30). Further-more, many �EDDD440 and �EAAA440 cilia displayed an elec-tron-dense material in the space between the outer doubletsand the ciliary membrane (Fig. 9, e and f) or were completely
filled with this dense material (Fig. 10e, arrowhead). Suchchanges were rarely seen in WT or �AAA447 cilia. In theaffected mutant cilia, an examination of the B-tubule showeda microtubule wall still detectable but with a center oftenfilled with the dense material (Fig. 9e). The lack of a clearempty center in the B-tubule is likely due to the existence ofunclosed microtubules. In other cilia, one or more peripheralsinglets were observed, indicating a complete loss of theB-tubule (Fig. 9f). In cilia containing the dense material, thepresence of only one central microtubule (Fig. 9e) was oftenobserved, indicating a partial loss of the central pair appa-ratus. Quantitative analysis of the data showed that theproportion of cilia with the electron-opaque structures wasalso higher in �EAAA440 than in �EDDD440 and �AAA447
mutants (Table II).To address the possibility that the higher frequency of de-
fects in �EAAA440, compared with �EDDD440, was induced bythe presence of alanine instead of aspartate residues in theprimary sequence of �-tubulin, we analyzed another �-tubulinmutant where four glycylation sites (instead of three) weresubstituted by aspartate residues (�EDDDGD442). Like thetriple site �-tubulin mutants, the latter one grew more slowlythan the WT; it displayed cytokinesis defects similar to those
FIG. 8. Mass spectra of the C-termi-nal peptides of �-tubulin from�EAAA440 Tetrahymena mutant (a)and of �-tubulin from �AAA447 Tetra-hymena mutant (b). Peptides were ana-lyzed in positive ion mode.
FIG. 9. Ultrastructural analysis of cross-sections of WT andmutant Tetrahymena cilia. a–c, �EDDD440 mutant cilia show anopening of the B-tubule (arrows). d, WT. e and f, �EAAA440 mutant ciliashow an electron-dense material in a ring between peripheral ODs andthe membrane. In e, opened B-tubules are visible (arrows), with theircenter filled with dense material; a single central microtubule is visible.In f, most of the B-tubules are not visible. Scale bar, 0.1 �m.
TABLE IIMorphology of cilia in WT and mutant Tetrahymena
For each strain, sections were counted (n), and within each sectiontype, the number of sections with different defects was also counted.
WT �EDDD440 �EAAA440 �AAA447
n (transverse sections) 37 37 28 32Normal axonemes 34 24 13 31Axonemes with opened
B-tubules or withperipheral singlets
3 13 15 1
n (transverse and obliquesections)
881 496 429 1131
Cilia with opaque structures 5 43 14
Tubulin Post-translational Modifications and Cilia Integrity602
by guest on February 8, 2016http://w
ww
.jbc.org/D
ownloaded from

previously described (30) and a great cell size heterogeneity. Inthis mutant, even a higher proportion (20%) of cilia was foundto contain aggregates of electron-dense material (see Fig. 10).Stacked structures were observed between one peripheral dou-blet and the ciliary membrane (Fig. 10a, arrow) and looked likeIFT structures described in Chlamydomonas (43, 44). Theywere frequently located between all ODs and the membrane(Fig. 10d) and sometimes were found as a massive dense struc-ture (arrow at the bottom of Fig. 10d and upper cilium in Fig.10a). Similarly as in �EAAA440, some cilia contained one cen-tral microtubule (Fig. 10d, arrowhead). In Fig. 10c, bridgeswere visible that link the B-tubule with the ciliary membrane.Fig. 10b represents one cilium displaying several peripheralsinglets. The fact that not all axonemal cross-sections showabnormalities could be due to their localized distributionalong the axoneme. Even if axoneme defects were less fre-quent in the �AAA447 mutant, Fig. 10e is informative regard-ing the gradual change of Tetrahymena mutant cilia. In thosecilia belonging to an oral apparatus, different degrees ofdefects were observed. Whereas in some cilia, intact periph-eral doublets were observed with a dense material at theirproximities, in other cilia, outer singlets were visible (ar-rows), and in one cilium, only a massive dense material waspresent, without any visible microtubule (arrowhead). Be-cause the opaque structures were observed in different mu-tants, in which tubulin polyglycylatable site mutations eitherinvolved charge conservation (�EDDDGD442), or not(�EAAA440), the most straightforward explanation is that theultrastructural defects in the mutants are induced bychanges in the levels of tubulin polymodifications.
DISCUSSION
Different Compositions of Tubulin Post-translational Modi-fications in Tetrahymena and Paramecium Contribute to As-sembly of Structurally Identical 9�2 Axonemes—We have pre-viously shown that, in Paramecium, axonemal tubulin is highlyglycylated, whereas glutamylation represents a quantitativelyminor PTM, given that it was only detected by immunoblottingand not by mass spectrometry (6, 17). The present study re-vealed that the Tetrahymena axonemal tubulin is glutamylatedat much higher level compared with Paramecium. The �-tubu-lin subunit in Tetrahymena appears to be more glutamylatedthan the � subunit. In contrast to glutamylation, the level ofglycylation was found to be higher in �- than in �-tubulin. Thisindicates that in Tetrahymena the dominant substrate forpolyglycylation is the � subunit, whereas the � subunit ispreferred for polyglutamylation. This is in contrast to Parame-cium where polyglycylation affects similarly both tubulin sub-units. In addition, the entire axonemal �/�-tubulin of Parame-cium is glycylated, whereas nonmodified �-tubulin was readilydetected in Tetrahymena.
In Tetrahymena, a substantial portion of tubulin isoforms,especially in the � subunit, is both glutamylated and glycylatedon the same molecule, and each level of glutamylation coexistswith one or more levels of glycylation. The coexistence of bothpolymodifications in a single molecule has also been found inGiardia lamblia and in the axonemes of sea urchin and bullsperm (32–34), but in those species such isoforms appeared tobe only minor variants. Given the presence of numerous gluta-mates within the primary sequence of the C-terminal tail of �-and �-tubulin (six and nine, respectively), the array of poly-
FIG. 10. Ultrastructural analysis ofcross-sections of �EDDDGD442 (a–d)and � AAA447 (e) mutant cilia. a,stacked particles (arrow) are visible be-tween one outer doublet and the mem-brane. The upper cilium contains a mas-sive dense material. In b, singletperipheral microtubules are visible. In c,the arrow shows bridges connecting theB-tubule to the membrane. In d, numer-ous oral cilia contain the opaque struc-tures between several ODs and the mem-brane (arrows). In one of these cilia, onecentral microtubule is missing (arrow-head). In e, arrows show oral cilia withsinglet peripheral microtubules. A ciliumis filled with an electron-dense materialwith no microtubule visible (arrowhead).Scale bar, 0.1 �m.
Tubulin Post-translational Modifications and Cilia Integrity 603
by guest on February 8, 2016http://w
ww
.jbc.org/D
ownloaded from

modified variants could be generated by the presence of closelyjuxtaposed or even shared polyglutamylation and polyglycylationsites. Alternatively, a single site could bear lateral chains combin-ing glutamate and glycine residues, as previously suggested (32).
Another striking difference between the two ciliate species isthe unique presence of detyrosination of �-tubulin in Tetrahy-mena. In Paramecium, the �-tubulin genes all lack the C-terminal tyrosine codon entirely (28),2 and the MS analyses didnot reveal any proteolytic event at the C terminus (6). Incontrast, the majority of �-tubulin peptides in Tetrahymenawas found to be detyrosinated. A novel type of �-tubulin car-boxypeptidase is likely involved in Tetrahymena, because thepenultimate amino acid in the �-tubulin is glycine and notglutamate as in most other species.
The differences between Tetrahymena and Parameciumwere unexpected, given that both ciliates belong to the samephylogenetic class known as Oligohymenophorea (45) and havea similar cytoskeleton design (46). The axonemes of these cili-ates appear to be identical based on electron microscopy. Thus,different combinations of PTMs in various organisms may con-tribute to assembly of essentially identical structures. Further-more, some species appear to lack entirely one of the polymodi-fications types; for example, in Trypanosoma, which assemblesboth axonemes and complex pellicular microtubules, tubulin isglutamylated but lacks glycylation (47). In contrast, in Para-mecium, glutamylation is quantitatively minor. It appearstherefore that the two polymodification types, despite theirdistinct chemical structure, are involved in similar functionsand could cooperate (as suggested for mammalian sperm mo-tility, see Ref. 13) or even substitute for each other. Growingevidence indicates that PTMs act on microtubules by modulat-ing their affinities for interactors such as microtubule-associ-ated proteins (MAPs) and molecular motors (see below). Indistinct species, PTMs and interactors that are dependent onPTMs could co-evolve while maintaining a similar structuraldesign of microtubular organelles.
Cross-talk between Polymodification Sites in �- and �-Tubu-lin Subunits—We found that mutations of three of the fiveglycylation sites in Tetrahymena �-tubulin change glycylationand glutamylation levels of axonemal tubulin on both the mu-tated and nonmutated tubulin subunits.
The �EDDD440 tubulin displayed a decrease of reactivitywith GT335 (Fig. 2), whereas a decrease of the number ofadditional glycines was observed in the most abundant �-tu-bulin isoform (monoglycylated in �EDDD440 instead of tetra-glycylated in the WT, Table I). This decrease of glycylation inthe predominant isoform is in accordance with the fact thatonly two of the five glycylation sites remain in the �EDDD440
mutant. However, we also detected quantitatively minor hy-perglycylated �-tubulin peptides in the same mutant. Twomechanistic explanations for this complex effect can be pro-posed. First, the elimination of sites could allow for addition ofmore glycines to the remaining sites, due to decreased compe-tition for the glycylase enzyme. Second, the elimination ofadjacent sites may increase the processivity of the glycylaseenzyme, in accordance with the detection of hyperglycylatedshort � peptides, 427DATAEEEGEFE437, containing Glu437 as asole remaining known glycylation site. Regardless of the mech-anism of hyperglycylation, our data suggest that the multiplesites of glycylation on �-tubulin influence each other.
Unexpectedly, mutations of �-tubulin glycylation sites af-fected dramatically both polymodifications in �-tubulin. Glu-tamylation, which is the most abundant PTM in WT �-tubulin,could not be detected in the �EDDD440 �-tubulin by mass
spectrometry. In contrast, the maximal number of added gly-cines, which reaches 10 in WT �-tubulin, is considerably in-creased (up to 37) in the �EDDD440 �-tubulin.
The strong effect of mutations on the nonmutated subunitsuggests the existence of cross-talk between the two subunits ofthe tubulin dimers. This cross-talk could either take placewithin the same dimer or between two subunits of adjacenttubulin dimers within a microtubule, given that both tubulinand microtubules could be substrates of both polymodificationenzymes, as shown for polyglutamylation (48). A simple modelcould assign to the �-tubulin subunit a role in the initial bind-ing of enzymes responsible for deposition of polymodificationson �-tubulin. Specifically, the tubulin glutamylase could have�-tubulin as a binding site for glutamylating the � subunit, andthe mutation of the tail domain of �-tubulin could affect theenzyme binding either directly, or through a conformationalchange in �-tubulin. There could also be a requirement of acertain level of glycylation on the �-tubulin tail for the bindingof the glutamylase. It is highly relevant that the best charac-terized tubulin modification enzyme, the tubulin-tyrosine li-gase, forms a tight complex with �/�-tubulin, involving a bind-ing site on �-tubulin in addition to its catalytic target, theC-terminal end of �-tubulin (49). It is therefore possible that alltubulin-amino acid ligases (tubulin-tyrosine ligase and the stillunknown glycylases and glutamylases) bind to �-tubulin whilethey modify the tails of �- or �/�-tubulin.
A comparison of �EDDD440 and �EAAA440 mutantsstrengthens the idea that the � subunit is critical for thepolymodifications on both �- and �-tubulin. The overall poly-glycylation extent on �- and �-tubulin of �EAAA440 was muchlower than that of �EDDD440 (Fig. 5 and Table I). In the�EAAA440 mutant, the presence of alanine instead of gluta-mate or aspartate residues in �-tubulin could decrease theaffinity of the modifying enzyme(s). This could be due to the netcharge decrease or to a conformational change in �-tubulin.
It is important to note that we also observed a reciprocalinfluence of the � subunit on �-tubulin, but it was more subtle.A mutation of the �-tubulin glycylation sites (�AAA447) in-creased the maximal number of added glycines to the tail of thenonmutated � subunit but did not affect the level of glycylationof the most abundant isoform (four glycines). In addition, glu-tamylation was not detected in the � subunit of this mutant. Itis therefore possible that the binding and/or activity of theglycylase(s) and glutamylase(s) involve a cooperative interac-tion of tails of both tubulin subunits. Various studies suggestthat the C-terminal tails are highly flexible domains at thesurface of microtubules, and are able to participate in numer-ous interactions with tubulins and other proteins such asMAPs and motors (50–52).
Regardless of the nature of the cross-talk mechanism, ourresults show the existence of interdependence of polymodifica-tion sites between the two subunits of the tubulin dimers.Because the mutations of the known glycylation sites on �-tu-bulin also affect glutamylation, the observed phenotypicchanges (Refs. 30 and 31 and this report) could represent theloss of both glycylation and glutamylation. Furthermore, thephenotype of the �-tubulin mutants may result from changesencompassing both subunits. Finally, it can not be ruled outthat the mutant phenotypes resulting from mutations in onesubunit are suppressed to some extent by potential compensatorychanges in the mutated and nonmutated subunits (such as in-creased enzyme processivity leading to hyperglycylation of minorisoforms). The dissection of respective functional contributions ofglutamylation and glycylation will require inactivation of genesencoding the glutamylation and glycylation enzymes.2 P. Dupuis-Williams, personal communication.
Tubulin Post-translational Modifications and Cilia Integrity604
by guest on February 8, 2016http://w
ww
.jbc.org/D
ownloaded from

Mutations in the Tubulin Polymodification Sites Affect theB-subfiber of Axonemal Outer Doublets and the Ciliary Ma-trix—Thazhath et al. (31) showed that a lethal deficiency intubulin polyglycylation affects the structure of the axoneme,manifested by the lack of the central pair and replacement ofperipheral doublets by singlets. We show here that the B-tubule of doublets is primarily affected by a mild deficiency inpolymodifications. In most cases, the viable mutant axonemeshave opened B-tubules. Such a defect may result from eitherinsufficient number of protofilaments or lack of proper bindingof the most lateral protofilament of the B-tubule to the A-tu-bule. Our analyses of the viable mutants revealed additionalchanges that were not obvious in the lethal mutants, includingaccumulation of the dense material between the doublets andthe ciliary membrane, suggestive of a defect in IFT (see below).
There appears to be a correlation between the strong effect ofthe polymodification site mutations on the B-tubule and thedistribution of polymodifications within the axoneme. Mul-tigner et al. (53) showed that, in sea urchin sperm flagella, theA-tubules contain unmodified tyrosinated tubulin, whereas theB-tubules contain detyrosinated tubulin that is extensivelypolyglycylated. A predominant occurrence of polyglutamylatedtubulin in the B-tubule was shown in Spermatozopsis similis(54). Thus, it appears that polymodifications have a functioneither in the assembly or in the attachment of the B-tubule tothe A-tubule in axonemes.
Previous data showed a potential role for polymodificationsin the maturation of organelles such as cilia and basal bodies(18, 54). Polyglycylation follows a sequential mechanism,monoglycylation occurring in newly assembled microtubularstructures and lengthening of polyglycine chains in matureones (18). Accordingly, recent epitope tagging experimentsshowed that a lethal glycylation site mutation affects newlyformed cilia as well as mature ones (55). Therefore, in Tetra-hymena, polymodifications could mediate the B-tubule closureduring the course of axonemal assembly or stabilize the B-tubule after the assembly. In both cases, glycylation or glu-tamylation could act directly or indirectly. Surprisingly,whereas long-lived microtubules in animal tissue cultured cellshave been shown to be extensively post-translationally modi-fied (56–58), the A-tubule of sea urchin sperm flagella, which isa stable microtubule, contains at least 95% of unmodified tu-bulin (53). Thus, a direct involvement of polymodifications inaxonemal microtubule stability seems unlikely. The axonemestabilization process could result from the doublet microtubuleconformation per se (as opposed to a singlet conformation)and/or from interactions between axonemal microtubules andaccessory proteins, in particular those which may be requiredfor the doublet formation such as tektins (59, 60). The poly-modifications could specifically recruit MAPs for assembly andstabilization of the B-tubule. In vitro experiments showed thatpolyglutamylation is involved in the interaction between tubu-lin and MAPs (61, 62), and a correlation between glycylationand microtubule cold stability has been established in gerbilcochlea (63), presumably involving cold MAPs (64).
Our results also suggest that polymodifications affect in-traflagellar transport. IFT involves the movement of large pro-tein complexes (IFT particles) along the doublet microtubules(65–67). This process is required for flagellar and ciliary as-sembly as well as maintenance, due to the turnover of flagellarcomponents (68, 69). IFT particles and flagellar or ciliary pre-cursors are moved in the flagella or cilia by kinesin-II (44, 70,71) and are recycled by the cytoplasmic dynein DHC1b (72).The IFT particles were shown to be preferentially associatedwith the B-subfiber of the outer doublet microtubules; theyappear as electron-dense material located between the flagellar
membrane and the peripheral doublets (43, 44, 73). Our elec-tron microscopy observations of hypomorphic �-tubulin mu-tants showed cilia with an unusual electron-dense material, inthe space between the ODs and the flagellar membrane, thatresembles IFT particles. This material may represent gatheredIFT particles, which were already shown to accumulate inmutants affected in IFT motors (44, 72–74). From MS data,�EDDD440 and �EAAA440 showed a decrease in the overallglutamylation levels of �/�-tubulin and in the glycylation levelof the predominant isoform of �-tubulin, in comparison withthe WT. Importantly, the maximal number of additional gly-cines analyzed in the peptide pool was lower in �EAAA440 (upto eight glycines) than in �EDDD440 �-tubulin (up to 26 gly-cines), and glutamylation was not detected in the �-tubulinpeptides of the former mutant (Table I), showing that�EAAA440 is more deficient in both polymodifications. Thisdeficiency correlates with a higher proportion of cilia contain-ing an unusual dense material adjacent to the ODs of�EAAA440.
The polyglycine and polyglutamate chains are known to bedistributed in a polarized fashion along the axoneme (10, 13,54, 75, 76) and therefore could potentially act as regulators ofIFT along the length of the axoneme. In addition, as observedin mammalian sperm flagella, individual doublets react differ-ently with the anti-tubulin polymodification mAbs (12, 13, 77).A differential modification of specific ODs could provide localenvironments more or less favorable for the anterograde andretrograde IFT. In glycylation site mutants, the decrease in thelevel of polymodifications could alter the kinetics of IFT and/orcomposition of IFT cargoes and induce destabilization of theB-tubule and microtubules of the central pair. We also need toconsider an alternative explanation, namely that the polymodi-fications are primarily required at the level of assembly ofmicrotubules. The potential effect on IFT could therefore beindirect and result from the structural defects in the B-tubules,which are tracks for IFT. Nevertheless, the observation ofnumerous cross-sections containing the opaque structures nearintact B-tubules favors the hypothesis that IFT defects precedethe structural defects of the B-tubules.
The effect on IFT could be mediated by changes in the motilityparameters of molecular motors. The processivity of kinesins wasshown to be regulated by the interactions between conservedbasic residues of the motor proteins and the acidic C termini oftubulin (78–80). The C-terminal sequence of tubulin, which in-teracts with kinesin (81), includes the polyglycylation and poly-glutamylation sites. Accordingly, microinjection of anti-polyglu-tamylated tubulin mAb (GT335) into melanophores selectivelyimpaired kinesin-dependent pigment granule dispersion (82).The affinity of kinesin to tubulin was shown to depend on thelength of the polyglutamyl chain (83), and flagellar dynein alsoappeared to depend on polyglutamylation and polyglycylation forinteractions with microtubules (10, 11). Polyglutamylation is al-ready known to affect the binding of MAPs and motors to tubu-lins in vitro (61, 62, 83). It is conceivable that, in the Tetrahymenamutants, lack of proper levels of polymodifications disrupts thebinding of these interactors. Polymodifications could specificallyaffect the interactions between IFT motors (kinesin-II and IFTdynein) and the axonemal microtubules and thus regulate theintraflagellar transport.
Acknowledgments—We are grateful to J. Degrouard and A.Fleury-Aubusson for help in figure editing. We are indebted to thecolleagues who generously provided us with antibodies. We thank G.Fryd-Versavel for help in ciliate stock maintenance, F. Iftode for adviceon electron microscopy, N. Lambin for help in manuscript editing, andM. Pradels for technical assistance.
Tubulin Post-translational Modifications and Cilia Integrity 605
by guest on February 8, 2016http://w
ww
.jbc.org/D
ownloaded from

REFERENCES
1. MacRae, T. H. (1997) Eur. J. Biochem. 244, 265–2782. Luduena, R. F. (1998) Int. Rev. Cytol. 178, 207–2753. Rosenbaum, J. (2000) Curr. Biol. 10, R801–R8034. Westermann, S., and Weber, K. (2003) Nat. Rev. Mol. Cell. Biol. 4, 938–9475. Edde, B., Rossier, J., Le Caer, J. P., Desbruyeres, E., Gros, F., and Denoulet,
P. (1990) Science 247, 83–856. Redeker, V., Levilliers, N., Schmitter, J. M., Le Caer, J. P., Rossier, J., Ad-
outte, A., and Bre, M. H. (1994) Science 266, 1688–16917. Redeker, V., Rossier, J., and Frankfurter, A. (1998) Biochemistry 37,
14838–148448. Schneider, A., Plessmann, U., Felleisen, R., and Weber, K. (1998) FEBS Lett.
429, 399–4029. Vinh, J., Langridge, J. I., Bre, M. H., Levilliers, N., Redeker, V., Loyaux, D.,
and Rossier, J. (1999) Biochemistry 38, 3133–313910. Bre, M. H., Redeker, V., Quibell, M., Darmanaden-Delorme, J., Bressac, C.,
Cosson, J., Huitorel, P., Schmitter, J. M., Rossier, J., Johnson, T., Adoutte,A., and Levilliers, N. (1996) J. Cell Sci. 109, 727–738
11. Gagnon, C., White, D., Cosson, J., Huitorel, P., Edde, B., Desbruyeres, E.,Paturle-Lafanechere, L., Multigner, L., Job, D., and Cibert, C. (1996) J. CellSci. 109, 1545–1553
12. Fouquet, J. P., Kann, M. L., Pechart, I., and Prigent, Y. (1997) Tissue Cell 29,573–583
13. Kann, M. L., Prigent, Y., Levilliers, N., Bre, M. H., and Fouquet, J. P. (1998)Cell Motil. Cytoskeleton 41, 341–352
14. Million, K., Larcher, J. C., Laoukili, J., Bourguignon, D., Marano, F., andTournier, F. (1999) J. Cell Sci. 112, 4357–4366
15. Pechart, I., Kann, M. L., Levilliers, N., Bre, M. H., and Fouquet, J. P. (1999)Biol. Cell 91, 685–697
16. Adoutte, A., Delgado, P., Fleury, A., Levilliers, N., Laine, M. C., Marty, M. C.,Boisvieux-Ulrich, E., and Sandoz, D. (1991) Biol. Cell 71, 227–245
17. Bre, M. H., de Nechaud, B., Wolff, A., and Fleury, A. (1994) Cell Motil.Cytoskeleton 27, 337–349
18. Iftode, F., Clerot, J. C., Levilliers, N., and Bre, M. H. (2000) Biol. Cell 92,615–628
19. Fleury, A., Callen, A. M., Bre, M. H., Iftode, F., Jeanmaire-Wolf, R., Levilliers,N., and Clerot, J. C. (1995) Protoplasma 189, 37–60
20. Bre, M. H., Redeker, V., Vinh, J., Rossier, J., and Levilliers, N. (1998) Mol.Biol. Cell 9, 2655–2665
21. Cohen, J., and Beisson, J. (1988) in Paramecium (Gortz, H.-D., ed) pp.363–392, Springer-Verlag, Berlin
22. Gaertig, J., and Fleury, A. (1992) Protoplasma 167, 74–8723. Gaertig, J. (2000) J. Eukaryot. Microbiol. 47, 185–19024. Suprenant, K. A., Hays, E., LeCluyse, E., and Dentler, W. L. (1985) Proc. Natl.
Acad. Sci. U. S. A. 82, 6908–691225. Dupuis, P. (1992) EMBO J. 11, 3713–371926. Gaertig, J., Thatcher, T. H., McGrath, K. E., Callahan, R. C., and Gorovsky,
M. A. (1993) Cell Motil. Cytoskeleton 25, 243–25327. McGrath, K. E., Yu, S. M., Heruth, D. P., Kelly, A. A., and Gorovsky, M. A.
(1994) Cell Motil. Cytoskeleton 27, 272–28328. Dupuis-Williams, P., Klotz, C., Mazarguil, H., and Beisson, J. (1996) Biol. Cell
87, 83–9329. Gaertig, J., Cruz, M. A., Bowen, J., Gu, L., Pennock, D. G., and Gorovsky, M. A.
(1995) J. Cell Biol. 129, 1301–131030. Xia, L., Hai, B., Gao, Y., Burnette, D., Thazhath, R., Duan, J., Bre, M. H.,
Levilliers, N., Gorovsky, M. A., and Gaertig, J. (2000) J. Cell Biol. 149,1097–1106
31. Thazhath, R., Liu, C., and Gaertig, J. (2002) Nat. Cell Biol. 4, 256–25932. Mary, J., Redeker, V., Le Caer, J. P., Rossier, J., and Schmitter, J. M. (1996)
J. Biol. Chem. 271, 9928–993333. Plessmann, U., and Weber, K. (1997) J. Protein Chem. 16, 385–39034. Weber, K., Schneider, A., Westermann, S., Muller, N., and Plessmann, U.
(1997) FEBS Lett. 419, 87–9135. Geuens, G., Hill, A. M., Levilliers, N., Adoutte, A., and DeBrabander, M. (1989)
J. Cell Biol. 108, 939–95336. Laemmli, U. K. (1970) Nature 227, 680–68537. Kyhse-Andersen, J. (1984) J. Biochem. Biophys. Methods 10, 203–20938. Callen, A. M., Adoutte, A., Andreu, J. M., Baroin-Tourancheau, A., Bre, M. H.,
Calvo Ruiz, P., Clerot, J. C., Delgado, P., Fleury, A., Jeanmaire-Wolf, R.,Viklicky, V., Villlalobo, E., and Levilliers, N. (1994) Biol. Cell 81, 95–119
39. Wolff, A., de Nechaud, B., Chillet, D., Mazarguil, H., Desbruyeres, E., Aude-bert, S., Edde, B., Gros, F., and Denoulet, P. (1992) Eur. J. Cell Biol. 59,425–432
40. Blose, S. H., Meltzer, D. I., and Feramisco, J. R. (1984) J. Cell Biol. 98,847–858
41. Arevalo, M. A., Nieto, J. M., Andreu, D., and Andreu, J. M. (1990) J. Mol. Biol.214, 105–120
42. Mock, K., Sutton, C. W., and Cottrell, J. S. (1992) Rapid Commun. MassSpectrom. 6, 233–238
43. Kozminski, K. G., Johnson, K. A., Forscher, P., and Rosenbaum, J. L. (1993)Proc. Natl. Acad. Sci. U. S. A. 90, 5519–5523
44. Kozminski, K. G., Beech, P. L., and Rosenbaum, J. L. (1995) J. Cell Biol. 131,1517–1527
45. Baroin-Tourancheau, A., Delgado, P., Perasso, R., and Adoutte, A. (1992) Proc.Natl. Acad. Sci. U. S. A. 89, 9764–9768
46. Fleury, A., Delgado, P., Iftode, F., and Adoutte, A. (1992) Dev. Genet. 13,247–254
47. Schneider, A., Plessmann, U., and Weber, K. (1997) J. Cell Sci. 110, 431–43748. Regnard, C., Audebert S., Desbruyeres E., Denoulet P., and Edde B. (1998)
Biochemistry 37, 8395–840449. Wehland, J., and Weber, K. (1987) J. Cell Biol. 104, 1059–106750. Sackett, D. L. (1995) in Subcellular Biochemistry, Vol. 24, Proteins: Structure,
Function, and Engineering (Biswas, B. B., and Roy, S., eds) pp. 255–302,Plenum Press, New York
51. Nogales, E. (2000) Annu. Rev. Biochem. 69, 277–30252. Lowe, J., Li, H., Downing, K. H., and Nogales, E. (2001) J. Mol. Biol. 313,
1045–105753. Multigner, L., Pignot-Paintrand, I., Saoudi, Y., Job, D., Plessmann, U., Rudi-
ger, M., and Weber, K. (1996) Biochemistry 35, 10862–1087154. Lechtreck, K. F., and Geimer, S. (2000) Cell. Motil. Cytoskeleton 47, 219–23555. Thazhath, R., Jerka-Dziadosz, M., Duan, J., Wloga, D., Gorovsky, M. A.,
Frankel, J., and Gaertig, J. (2004) Mol. Biol. Cell 15, 4136–414756. Bre, M. H., Kreis, T. E., and Karsenti, E. (1987) J. Cell Biol. 105, 1283–129657. Wehland, J., and Weber, K. (1987) J. Cell Sci. 88, 185–20358. Schulze, E., Asai, D. J., Bulinski, J. C., and Kirschner, M. (1987) J. Cell Biol.
105, 2167–217759. Norrander, J. M., and Linck, W. (1994) in Microtubules (Hyams, J. S., and
Lloyd, C. W., eds) pp. 201–220, Wiley-Liss, New York60. Nojima, D., Linck, R. W., and Egelman, E. H. (1995) Curr. Biol. 5, 158–16761. Boucher, D., Larcher, J. C., Gros, F., and Denoulet, P. (1994) Biochemistry 33,
12471–1247762. Bonnet, C., Boucher, D., Lazereg, S., Pedrotti, B., Islam, K., Denoulet, P., and
Larcher, J. C. (2001) J. Biol. Chem. 276, 12839–1284863. Bane, B. C., MacRae, T. H., Xiang, H., Bateman, J., and Slepecky, N. B. (2002)
Cell Tissue Res. 307, 57–6764. Bosc, C., Andrieux, A., and Job, D. (2003) Biochemistry 42, 12125–1213265. Rosenbaum, J. L., Cole, D. G., and Diener, D. R. (1999) J. Cell Biol. 144,
385–38866. Rosenbaum, J. L., and Witman, G. B. (2002) Nat. Rev. Mol. Cell. Biol. 3,
813–82567. Snell, W. J., Pan, J., and Wang, Q. (2004) Cell 117, 693–69768. Song, L., and Dentler, W. L. (2001) J. Biol. Chem. 276, 29754–2976369. Marshall, W. F., and Rosenbaum, J. L. (2001) J. Cell Biol. 155, 405–41470. Piperno, G., Mead, K., and Henderson, S. (1996) J. Cell Biol. 133, 371–37971. Brown, J. M., Marsala, C., Kosoy, R., and Gaertig, J. (1999) Mol. Biol. Cell 10,
3081–309672. Pazour, G. J., Dickert, B. L., and Witman, G. B. (1999) J. Cell Biol. 144,
473–48173. Pazour, G. J., Wilkerson, C. G., and Witman, G. B. (1998) J. Cell Biol. 141,
979–99274. Porter, M. E., Bower, R., Knott, J. A., Byrd, P., and Dentler, W. (1999) Mol.
Biol. Cell 10, 693–71275. Kann, M. L., Prigent, Y., and Fouquet, J. P. (1995) Tissue Cell 27, 323–32976. Huitorel, P., White, D., Fouquet, J. P., Kann, M. L., Cosson, J., and Gagnon, C.
(2002) Mol. Reprod. Dev. 62, 139–14877. Fouquet, J. P., Prigent, Y., and Kann, M. L. (1996) Mol. Reprod. Dev. 43,
358–36578. Okada, Y., and Hirokawa, N. (2000) Proc. Natl. Acad. Sci. U. S. A. 97, 640–64579. Thorn, K. S., Ubersax, J. A., and Vale, R. D. (2000) J. Cell Biol. 151, 1093–110080. Skiniotis, G., Cochran, J. C., Muller, J., Mandelkow, E., Gilbert, S. P., and
Hoenger, A. (2004) EMBO J. 23, 989–99981. Tucker, C., and Goldstein, L. S. B. (1997) J. Biol. Chem. 272, 9481–948882. Klotz, A., Rutberg, M., Denoulet, P., and Wallin, M. (1999) Cell Motil. Cytoskel-
eton 44, 263–27383. Larcher, J. C., Boucher, D., Lazereg, S., Gros, F., and Denoulet, P. (1996)
J. Biol. Chem. 271, 22117–22124
Tubulin Post-translational Modifications and Cilia Integrity606
by guest on February 8, 2016http://w
ww
.jbc.org/D
ownloaded from

Dylan Burnette, Jacek Gaertig and Marie-Hélène BréVirginie Redeker, Nicolette Levilliers, Emilie Vinolo, Jean Rossier, Danielle Jaillard,
Tetrahymena-Tubulin and Affect the Ciliary Matrix in β- and αMutations of Tubulin Glycylation Sites Reveal Cross-talk between the C Termini of
doi: 10.1074/jbc.M408324200 originally published online October 18, 20042005, 280:596-606.J. Biol. Chem.
10.1074/jbc.M408324200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/280/1/596.full.html#ref-list-1
This article cites 80 references, 34 of which can be accessed free at
by guest on February 8, 2016http://w
ww
.jbc.org/D
ownloaded from





























