Embed Size (px)
Citation preview

N-acetylcysteineasapotentialtreatmentforpersistentbreeding-inducedendometritisinthemare
by
MichelleDianeCaissie
AThesispresentedto
TheUniversityofGuelph
Inpartialfulfillmentofrequirementsforthedegreeof
DoctorofVeterinarySciencein
PopulationMedicine
Guelph,Ontario,Canada
©MichelleDianeCaissie,August,2019

ii
ABSTRACT
N-acetylcysteineasapotentialtreatmentforpersistentbreeding-inducedendometritisinthemare
MichelleCaissieAdvisor:UniversityofGuelph,2019Dr.TraceyChenier
Persistentbreeding-inducedendometritis(PBIE)isamajorcauseofinfertilityinmares.
PBIEsusceptiblemaresdonotclearinflammationpost-breedinginatimelyfashion,ultimately
resultinginearlyembryonicdeath.N-acetylcysteine(NAC),amucolytic,hasanti-inflammatory
properties, affects inflammatory cytokines and is a mild inhibitor of nitric oxide synthase.
Increased nitric oxide (NO) levels and alterations in inflammatory cytokines have been
documented inPBIEmares.Theobjectiveofthisstudywastodetermine ifNACtreatmentof
PBIE-susceptiblemareswouldlowerNOandinflammatorycytokinelevels,therebyresultingin
the resolution of clinical signs associatedwith PBIE. The effects of NAC histologically on the
endometriumwerealsodetermined.Arandomized,blinded,cross-overdesignclinicaltrialwas
performedutilizingPBIEsusceptiblemares(n=9). Intrauterineinfusionof180mlsof3.3%NAC
wasperformed12hourspriortoinsemination.Maresweresampledforendometrialcytology
and intra-uterine fluid to determine interleukin-6 concentration (ELISA) and nitric oxide
(Colorimetric assay) levels at 12 and 60 hours post-insemination. Endometrial biopsies were
takenatthesametimepointspost-inseminationforscoringbasedonthedegreeofinflammation

iii
presentandtodeterminegeneexpressionofinflammatorycytokinesbyqPCR.Clinicalsignsof
endometrialedemaandintra-uterinefluidvolumeswereassessedat12andthenevery24hours
postbreeding.StatisticalanalysiswasperformedusingarepeatedmeasuresANOVAandaMann
WhitneyWilcoxonTest.Pre-breedingintrauterinetreatmentwithNACdidnotimproveclinical
signs in PBIE-affectedmares, nor did it affectNO levels, IL-6 concentration or cytokine gene
expression.Differencesacrosstimepointspost-AIwerenotedforIL-6geneexpression,which
was highest at 12 hours post-AI (p=0.003). Expression of other cytokines evaluated did not
changeovertime.TreatmentwithNACdidhaveasignificanteffectonthedegreeofendometrial
inflammation noted histologically, with treated cycles displaying more diffuse and severe
neutrophil infiltrationcomparedtocontrolcycles.TheeffectsofNAContheendometriumof
PBIE-susceptiblemaresseeninthisstudyisconcerning.Furtherresearchisrequiredtoevaluate
thesafetyofNACinthetreatmentofPBIE-susceptiblemares.

iv
ACKNOWLEDGEMENTS
Firstly, I would like to thankmy supervisor, Dr. Tracey Chenier for her guidance and
endless support throughout theDVScprogram.Researchwasanewerconcept tomewhen I
began,butDr.ChenierprovidedmewitheverythingIneededtocompletethisproject.Starting
withthebasics,progressingthroughtothemoredifficultelements,shehelpedmetogrowasa
researcher.Shewasconsistentlyavailableforfeedbackandassistance,evenlateatnightoron
weekends.Shehadauniqueabilitytocreateopenandconstructivebrainstormingsessionswhile
makingmefeelcomfortabletoaskquestions,howeversimpletheymayhavebeen.Whenthe
programbecameoverwhelming,sheknewexactlywhattosaytopushmeoutsideofmycomfort
zoneandtomovemyprogressforward.Dr.Chenierwasnotonlyagreatmentorforresearch,
shealsowasagreatmentorasaclinicianandasateacher.Duringmyfirstseasonasaresident
inTheriogenology,Dr.ChenierallowedmetogrowasaclinicianasIlearnedtomanagemyown
researchmares,whileprovidingmewith theguidanceand support I needed. Having sucha
knowledgeableandskilledcliniciantolearnfromwasaninvaluableopportunityforme,andone
thatIwillbeforevergratefulfor.Ithoroughlyenjoyedworkingwithher,bothintheclinic,and
onthisresearchproject.
Thepastthreeyearscouldalsonothavebeenpossiblewithoutmyothermentor,Dr.
CathyGartley.Dr.Gartleyhasbeenthereforme,providingvaluableinsightoneverythingfrom
tohowtodealwiththemostdifficultofcases,tofeedbackregardingmyresearchand,even
adviceonlifeoutsidetheclinic.Shehasbeenencouraging,approachable,andextremely
knowledgeable.Iwasalwaysexcitedtoseecaseswithher,notonlyforthelearning

v
opportunitiesbutalsoforthebyproductsofconversationandlaughterthatwouldemergefrom
spendingtimetogetherintheclinic.Dr.Gartleyalsotookthetimetodiscussvariousconditions
withmeoutsidetheclassroomandagreedtocoordinateaspecialtopicscoursetohelpprepare
meforboards.Icannotexpressmygratitudeforherdedicationtoteaching.
InadditiontothementorshipthatbothDr.ChenierandDr.Gartleyhaveprovidedme,I
willforevercherishtheirfriendship.
Totheothermembersofmythesiscommittee,Dr.ElizabethScholtz,Dr.JoanneHewson
andDr.RonJohnson,Ioweagreatdealofthanks.Fromtheverybeginningofthisprogram,they
wereencouraging,enthusiastic,insightfulandprovidedmewithinvaluableguidancethroughthe
completionofthisresearchproject.Iamespeciallygratefultothemforhelpingmeredesignmy
researchprojectandforalltheirhardworkduringtheeditingphaseofthisthesis.
ManythanksgotoDr.MonicaAntenosforteachingmeeverythingthereistoknowabout
pipettingandrunningELISAs.Iwouldnothavebeenabletocompletethissectionofmyresearch
projectwithout her. I thoroughly valued her teaching, knowledge, opinion, and patience. I
enjoyedmyexperienceworkingwithherandamforeverthankfulforherhelp.Aspecialthank
youtoDr.AllisonMacKayandDr.LizSt.Johnfortheirencouragementandhelpwhileworkingin
thelabaswell.
IowemygratitudetoDr.DorotheeBienzleandDr.MaryEllenClarkforperformingthe
qPCR analysis for this project. Special thanks goes to Dr. Rob Foster for his insight on the
endometrialbiopsyanalysis,forbeingonmyqualifyinganddefenseexaminationcommittee,and
forallowingmeattendhisgraduatereproductivepathologycourse.Also,thankyoutoDr.Paula

vi
MenziesandDr.TerriO’Sullivanforteachingmeaboutsmallruminantsandswinereproduction.
MygratitudetoWilliamSearsforrunningandbeingpatientduringthestatisticalanalysis.
IamgratefultoKarenDaPonteasafriend,ateacher,andatherapist.Shehasbeena
wealthofknowledgethroughoutmyDVSc.Karenhasbeenapillarofsupportthroughdifficult
cases,challengesoftheprogram,andinlife.Shewillalwaysholdaspecialplaceinmyheart,and
willforeverbemyfriend.ToSarahDyck,athankyouforbeingafantasticresearchassistantand
friendduringtheclinicalphaseofmyresearchproject.Iwouldhavenotbeenabletogetover
thelearningcurvewithouther,andforthatIamgrateful.ThankyoualsotothestaffattheArkell
Research Facility, most notably Danielle Watson, for their help with the mares during this
researchproject.AspecialthankstoStephanieBourgonforhelpingmewithmyELISAs,andto
PaigeVeyseyforhelpingmewithmyresearchandforbeingagoodresidentmate.
IwouldalsoliketothankDr.FirdousKhanforallhishelpatthebeginningofmyprogram,
andforprovidingmewithinvaluableinsightonmyresearchproject.Iappreciatehis,Dr.Mariana
DieldeAmorimandDr.RasaLevstein’sencouragementandguidancethroughoutmypreparation
fortheboardexam.
This project would not have been possible without the generous contributions from
EquineGuelphandtheOntarioMinistryofAgriculture,FoodandRuralAffairs(OMAFRA)andto
themIowemanythanks.
Finally,Iwouldliketothankmyfamilyfortheirendlesssupport.Therearenotenough
words to express how much they mean to me. To Blaire, my partner, who provided
encouragementwhenthingsgotdifficultandwhocelebratedsuccesseswithme.Thepastthree
yearshavenotbeeneasyforhimeither,buthislove,patienceandsupportneverwavered.Not

vii
onlydidhelivethroughthedaytodaywithme,healsohelpedmemasterMicrosoftWordand
Excelandhelpedmewithmywork,sowecouldspendtimetogether.Tomyparents,Daveand
Diane,theyhavebeentherefromthebeginning:fromencouragingmetofollowmydreams,to
talkingmethroughpanickingaboutanundergraduatephysics100course,totalkingmethrough
thefinalweeksofmyDVScandtellingmetonevergiveup.Iamespeciallyappreciativetomy
dad,whoremindedmetotakethingsdaybyday,andtomymomforalwayslookingoutformy
well-being,andtellingmeit’sokaytotakeabreak.Tomysiblings,DanielleandMatt,mybrother-
in-lawRick,andmynieceandnephews,Spencer,HaileyandKoen,thankyouforunderstanding
thatIcouldnotcomehometospendasmuchtimewithyouasIwanted.Tomypartner’sfamily,
theMacKenzies,Bill,Heather,Laura,Peter,Candace,EmmaandGrace,thankyouforbeinga
greatsupportsystemandforgivingmeahomeawayfromhome.Lastly,Iamthankfulformy
furryfriends,CooperandOreowhohavebeenmyconstantstudycompanions.

viii
DECLARATIONOFWORKPERFORMED
IdeclarethatIperformedalltheworkdescribedinthisthesiswiththeexceptionofthefollowing:
BacterialcultureofendometrialswabswasperformedattheAnimalHealthLaboratoryatthe
UniversityofGuelph.
Real-time quantitative PCR on endometrial tissue was performed by Dr. Bienzle’s research
laboratoryattheUniversityofGuelphintheDepartmentofPathobiology.
The preparation and staining with Hematoxylin and Eosin (H&E) of endometrial tissues for
histopathologicalevaluationwasperformedbytheAnimalHealthLaboratoryattheUniversityof
Guelph.
Histological evaluation and scoring of inflammation for these sampleswas performed byDr.
TraceyChenierintheDepartmentofPopulationMedicine,withtheinputofDr.RobFosterinthe
DepartmentofPathobiologyattheUniversityofGuelph.

ix
TABLEOFCONTENTSABSTRACT......................................................................................................................................ii
ACKNOWLEDGEMENTS.................................................................................................................iv
DECLARATIONOFWORKPERFORMED......................................................................................viii
LISTOFFIGURES...........................................................................................................................xii
LISTOFTABLES............................................................................................................................xiv
LISTOFAPPENDICES....................................................................................................................xv
LISTOFABBREVIATIONS.............................................................................................................xvii
CHAPTERONE:LITERATUREREVIEW............................................................................................1
1.1INTRODUCTIONANDSIGNIFICANCEOFPERSISTENTBREEDINGINDUCEDENDOMETRITITS(PBIE)..............................................................................................................1
1.2THEIMMUNESYSTEM&THENORMALINFLAMMATORYRESPONSE...............................2
1.3RISKFACTORSFORPBIE......................................................................................................7
1.4PATHOPHYSIOLOGYOFPBIE.............................................................................................11
1.4.1HumoralandInnateImmunity...................................................................................11
1.4.2UterineClearance.......................................................................................................15
1.4.3MucociliaryApparatusandLymphatics.....................................................................19
1.4.4CytokineProfilesofSusceptibleandResistantMares..............................................21
1.5DIAGNOSISOFPBIE...........................................................................................................23
1.6TREATMENTOFPBIE..........................................................................................................25
1.6.1EcbolicTherapy...........................................................................................................25
1.6.2UterineLavage............................................................................................................27
1.6.3Immunomodulation....................................................................................................28
1.6.4N-AcetylCysteine........................................................................................................32
1.7RESEARCHOBJECTIVES......................................................................................................35
1.8REFERENCES.......................................................................................................................36
CHAPTERTWO:THEEFFECTSOFTREATMENTWITHN-ACETYLCYSTEINEONCLINICALSIGNSINPBIESUSCEPTIBLEMARES...........................................................................................................57
2.1ABSTRACT..........................................................................................................................57
2.2INTRODUCTION..................................................................................................................58
2.3MATERIALSANDMETHODS..............................................................................................61
2.3.1SelectionofPBIESusceptibleMares..........................................................................62

x
2.3.2ClinicalTrial.................................................................................................................62
2.3.3InflammationasAssessedbyEndometrialBiopsies..................................................64
2.3.4EndometrialCytology(%Neutrophils).......................................................................65
2.3.5StatisticalAnalyses.....................................................................................................65
2.4RESULTS.............................................................................................................................66
2.4.1BiopsyInflammationScore........................................................................................66
2.4.2EstimatedIntra-UterineFluidVolumes(IUF).............................................................67
2.4.3EndometrialEdema....................................................................................................68
2.4.4EndometrialCytology(%Neutrophils).......................................................................68
2.4.5InteractionsBetweenTreatmentCyclesandHoursPost-AI(Appendix1)...............68
2.5DISCUSSION.......................................................................................................................68
2.6REFERENCES.......................................................................................................................76
CHAPTERTHREE:THEEFFECTOFTREATMENTOFPBIESUSCEPTIBLEMARESWITHN-ACETYLCYSTEINEONNITRICOXIDELEVELSANDINFLAMMATORYCYTOKINEPROFILES.....................93
3.1ABSTRACT..........................................................................................................................93
3.2INTRODUCTION..................................................................................................................94
3.3MATERIALSANDMETHODS..............................................................................................97
3.3.1MareSelection,StudyDesignandSampling.............................................................97
3.3.2Real-TimeQuantitativePCR.......................................................................................99
3.3.3Intra-UterineFluidCollection.....................................................................................99
3.3.4ELISADeterminationofInterleukin-6ConcentrationinUterineFluid....................100
3.3.5NitricOxideConcentration.......................................................................................100
3.3.6StatisticalAnalyses...................................................................................................101
3.4 RESULTS......................................................................................................................102
3.4.1EffectofIntra-UterineTreatmentof3.3%NACinPBIESusceptibleMares............102
3.4.2ChangesinEndometrialInflammationinResponsetoAIOverTime.....................102
3.4.3IL-6ConcentrationasMeasuredbyELISA................................................................103
3.4.4Nitricoxide................................................................................................................103
3.5DISCUSSION.....................................................................................................................103
3.6REFERENCES.....................................................................................................................110
CHAPTERFOUR:GENERALDISCUSSION....................................................................................122
4.1POSSIBLELINKSBETWEENMRNAEXPRESSIONANDPBIECLINICALSIGNS...................122
4.2LIMITATIONS....................................................................................................................124

xi
4.3OVERALLCONCLUSIONSANDFUTUREDIRECTIONS.......................................................127
4.4REFERENCES.....................................................................................................................132
APPENDICES...............................................................................................................................134

xii
LISTOFFIGURESCHAPTERONE
Figure1:Simplifiedflowchartdisplayingcytokineinteractions
CHAPTERTWO
Figure1:Graphicalrepresentationoftheclinicaltrialtimeline.
Figure2(a-d):PhotomicrographsofendometrialbiopsiesofPBIE-susceptiblemaresassignedan
inflammatorybiopsyscorerangingfrom1-4.
Figure3:DifferencesinmeanbiopsyinflammationscoreofPBIE-susceptiblemaresintreatedand
controlcycles.
Figure4:Photomicrographofendometrialbiopsydisplayingerosionoftheendometrialluminal
epitheliumfollowingintra-uterinetreatmentwith3.3%NAC.
Figure5:Photomicrographofendometrialbiopsydisplayingneutrophilinfiltrationofthestratum
spongiosumandglandularlumenfollowingintra-uterinetreatmentwith3.3%NAC.
Figure6:MeanbiopsyinflammationscoreofPBIE-susceptiblemaresat12and60hourspost-AI.
Figure7:MedianIUFofPBIE-susceptiblemaresintreatedandcontrolcycles.
Figure8:MedianIUFofPBIE-susceptiblemaresat12,36,60and84hourspost-AI.
Figure9:MeanendometrialedemascoreofPBIE-susceptiblemaresintreatedandcontrolcycles.
Figure10:MeanendometrialedemascoreofPBIE-susceptiblemaresat12,36,60and84hours
post-AI.
Figure11:Mean%neutrophilsofPBIE-susceptiblemaresintreatedandcontrolcycles.
Figure12:Mean%neutrophilsofPBIE-susceptiblemaresat12and60hourspost-AI.

xiii
CHAPTERTHREE
Figure1:MedianIL-6concentrationofPBIE-susceptiblemaresintreatedandcontrolcycles.
Figure2:MediannitricoxideofPBIE-susceptiblemaresintreatedandcontrolcyclespost-AI.
Figure3:MedianIL-6mRNAexpressionat12and60hourspost-AI.
Figure4:Median IL-6mRNAexpression ina subsetofPBIE-susceptiblemaresatbaseline,12
hoursand60hoursintreatedandcontrolcycles.
Figure5:MedianIL-1βmRNAexpressioninasubsetofPBIE-susceptiblemaresatbaseline,12
hoursand60hoursintreatedandcontrolcycles.
Figure6:MedianIL-10mRNAexpressioninasubsetofPBIE-susceptiblemaresatbaseline,12
hoursand60hoursintreatedandcontrolcycles.
Figure7:MedianTNF-αmRNAexpressioninasubsetofPBIE-susceptiblemaresatbaseline,12
hoursand60hoursintreatedandcontrolcycles.
Figure8:MedianIL-6concentrationofPBIE-susceptiblemaresat12and60hourspost-AI.
Figure9:MediannitricoxideofPBIE-susceptiblemaresat12and60hourspost-AI.

xiv
LISTOFTABLESCHAPTERONE
Table1:mRNAexpressionof select cytokines inequineendometritis literature,organizedby
study.
Table2:mRNAexpressionof select cytokines inequineendometritis literature,organizedby
cytokine.
CHAPTERTWO
Table1:InflammationbiopsyscoreassignmentforeachPBIE-susceptiblemareintreatedand
controlcyclesat12and60hourspost-AI.
CHAPTERTHREE
Table1:PairsofreferencegenesusedinclinicaltrialforeachPBIE-susceptiblemare.
Table2:Primersequencesforamplificationofreferencegenesandselectedcytokines.
Table3:MedianmRNAexpressionofIL-6,IL-10,TNF-αandIL-1βfortreatedandcontrolcycles
inPBIE-susceptiblemares.
Table4:MedianmRNAexpressionofIL-6,IL-10,TNF-αandIL-1βat12and60hourspost-AIin
PBIE-susceptiblemares.
Table5:MedianmRNAexpressionofIL-6,IL-10,TNF-αandIL-1βatbaseline,12and60hours
post-AIforasubsetofPBIE-susceptiblemares.

xv
LISTOFAPPENDICESCHAPTERTWO
Appendix1:Interactionsbetweentreatmentcyclesandhourspost-AIforoutcomesmeasured
inchapter2
Figure1:Meanbiopsyinflammationscoreintreatedandcontrolcyclesat12and60hours
postAI.
Figure2:MedianIUFintreatedandcontrolcyclesat12,36,60and84hourspostAI.
Figure3:Meanendometrialedemascoreintreatedandcontrolcyclesat12,36,60and84
hourspostAI.
Figure4:Mean%neutrophilsintreatedandcontrolcyclesat12and60hourspostAI.
CHAPTERTHREE
Appendix2:IL-6validationprocess
Table1:CalculatedvaluesofpercentrecoveryandlinearityforfinalIL-6validation.
Appendix3:Maineffectsoftreatmentandhourspost-AIoncytokinesevaluated
Figure1:MedianIL-6mRNAexpressioninPBIE-susceptiblemaresintreatedandcontrol
cycles.
Figure2:MedianIL-10mRNAexpressioninPBIE-susceptiblemaresintreatedandcontrol
cycles.
Figure3:MedianTNF-αmRNAexpressioninPBIE-susceptiblemaresintreatedandcontrol
cycles.
Figure4:MedianIL-1βmRNAexpressioninPBIE-susceptiblemaresintreatedandcontrol
cycles.

xvi
Figure5:MedianIL-6mRNAexpressioninPBIE-susceptiblemaresintreatedandcontrol
cyclesat12and60hourspost-AI.
Figure6:MedianIL-10mRNAexpressioninPBIE-susceptiblemaresat12and60hours
post-AI.
Figure7:MedianTNF-αmRNAexpressioninPBIE-susceptiblemaresat12and60hours
post-AI.
Figure8:MedianIL-1βmRNAexpressioninPBIE-susceptiblemaresat12and60hours
post-AI.
CHAPTERFOUR
Appendix2:IL-6validationprocess
Table1:CalculatedvaluesofpercentrecoveryandlinearityforfinalIL-6validation.
Appendix4:ProcessesforvalidationattemptsforIL-10,TNF-αandIL-1βELISAkits
xvii
LISTOFABBREVIATIONSAHL AnimalHealthLaboratory IgG ImmunoglobulinGANOVA AnalysisofVariance IgM ImmunoglobulinMAIC AkaikeInformationCriterion Ig(T) ImmunoglobulinTAI ArtificialInsemination iNOS InducibleNitricOxideSynthaseBSA BovineSerumAlbumin IFN-γ Interferon-γCDE ChronicDegenerative
EndometritisIL-1α Interleukin-1α
COPD ChronicObstructivePulmonaryDisease
IL-1RA Interleukin1ReceptorAgonist
CUI ChronicUterineInfection IL-1RN Interleukin1ReceptorAgonistGene
CD4+ ClusterofDifferentiation4 IL-6 Interleukin-6CD8+ ClusterofDifferentiation8 IL-8 Interleukin-8
C3 ComplementComponent3 IL-10 Interleukin-10C3b ComplementComponent3b IUF Intra-UterineFluidVolumeC5 ComplementComponent5 KCI PotassiumchlorideC5a ComplementComponent5a LRS LactatedRinger’sSolution
C5b ComplementComponent5b LPS LipopolysaccharidescDNA ComplementaryDNA LL LowerConfidenceLimitCT CycleThreshold MHC MajorHistocompatibilityComplexCOX-2 Cyclooxygenase-2 mRNA MessengerRNACRISP-3 CysteineRichSecretory
Protein3mOsm Milliosmole
DNA Deoxyribonucleicacid MS MultipleSclerosis
DUC DelayedUterineClearance MCWE MycobacterialCellWallExtractEMG Electromyography NAC N-acetylCysteineeNOS EndothelialNitricOxide
SynthaseNET NeutrophilExtracellularTrap
ELISA Enzyme-LinkedImmunosorbentAssay
nNOS NeuronalNitricOxideSynthase
EeF2 EukaryoticTranslationElongationFactor2
NO NitricOxide
FBS FetalBovineSerum NOS NitricOxideSynthase
GAPDH Glyceraldehyde3-phosphatedehydrogenase
NF-κB NuclearFactor-κB
H&E HematoxylinandEosin NOD NucleotideoligomerizationdomainhCG HumanChorionic
GonadotropinPBIE PersistentBreedingInduced
EndometritisHpf HighPowerField(400x) PBS PhosphateBufferedSaline
xviii
PRP PlateletRichPlasma TEM TransmissionElectronMicroscopyPMN Polymorphonuclear
neutrophilsTX TreatmentCycle
PGF2α ProstaglandinF2αTNF-α TumorNecrosisFactor-α
PGFM ProstaglandinF2αmetabolite SEM ScanningElectronMicroscopyRQ RelativeQuantification SE StandardErrorqPCR(qRT-PCR)
ReverseTranscriptaseQuantitativePCR
SDHA SuccinateDehydrogenaseComplexFlavoproteinSubunitA
RNA RibonucleicAcid UL UpperConfidenceLimitRPS23 RibosomalProteinS23

1
CHAPTERONE:LITERATUREREVIEW
1.1INTRODUCTIONANDSIGNIFICANCEOFPERSISTENTBREEDINGINDUCED
ENDOMETRITITS(PBIE)
In2010,horsescontributed19.6billiondollarstotheCanadianeconomy(Evans2012).At
thattime,thehorsepopulationinCanadawas963500,ofwhich41%werepartofthebreeding
sector.Twenty-twopercentofthenationalequinepopulationresidesinOntario,representing
thesecondlargesthorsepopulationwithinCanada,followingAlberta.InOntario,14%ofhorses
arewithin thebreedingsector (Evans2012).Thehealthof thebreeding industry is therefore
criticaltotheoverallsuccessofthehorseindustry.Despitethis,reproductiveefficiencyinhorses
is generally low, for a variety of reasons, including infertility caused by uterine disease.
Endometritis is the third most common medical condition encountered overall in equine
veterinary practice (Traub-Dargatzet al. 1991), and approximately 60%ofmares affectedby
endometritisarebarren(Overbecketal.2011).Causesofendometritisincludeacutebacterial
infections,chronic infections,andpersistentbreeding-induced inflammation.Estimatesof the
prevalenceofPBIEvarydependinguponthestudyandthemethodusedtodefinethecondition.
Whendefinedas“anyfluidaccumulationwithintheuterusonedayfollowingbreeding”,PBIE
was present in 13.7% of intensively managed Thoroughbred broodmares (Zent et al. 1998).
AnotherstudyidentifiedPBIEin43%ofamixedpopulationofmares(Newcombe1997).Given
thefairlylargenumberofhorsesinthebreedingsectorbothinOntarioandCanadaasawhole,
endometritis and more specifically PBIE, causes a significant economic impact to both the
CanadianandOntariobreedingindustries.

2
Maresaffectedbyendometritishaveimpairedfertility,leadingtodecreasedpregnancy
rates and the requirement for repeat breedings. Endometritis impairs fertility by two
mechanisms:prematureluteolysiscausedbyinflammatory-drivenprostaglandinrelease,andby
thedirect toxiceffectsofuterine inflammationon theembryo (Neelyetal. 1979;Troedsson
1997;Troedssonetal.2001b).Ashort-livedinflammatoryreactionisanormalresponsefollowing
breedinginallmares(Kotilainenetal.1994;Katilaetal.1995)andisnecessarysincenon-viable
spermatozoa,inflammatorydebrisandothercomponentsoftheejaculatemustbeclearedfrom
thefemale’sreproductivetractpriortoembryodescentonday5or6post-breeding(Betteridge
et al. 1982; Freeman et al. 1992). A subset of mares fail to clear the normal inflammatory
responseinatimelyfashion,leadingtopersistentinflammationandearlyembryonicdeathat
the time of embryo descent.Mares inwhich this occurs are considered susceptible to PBIE.
Understanding this post-breeding immune response and susceptibility to PBIE is key to
understandingmethodsthatmayreducetheimpactofPBIEonmarereproductiveperformance.
1.2THEIMMUNESYSTEM&THENORMALINFLAMMATORYRESPONSE
Theimmunesystemisdividedintotwobroadcategories:theinnateimmunesystemand
theadaptiveimmunesystem.Responsebytheinnateimmunesystemdoesnotrequireprevious
exposuretoanantigen;thesameresponsewilloccurregardlessoftheincitingcause.Theinnate
immune system consists of physical barriers, phagocytic cells and plasma proteins. By
comparison,theadaptiveimmunesystemhasamemorycomponent,whichprimesitsresponse
tospecificagentssuchasacertainspeciesofbacteriaorvirus,followinginitialexposure(Tizard
2013).Adaptiveimmunityisfurtherdividedintohumoralandcell-mediatedimmunity,wherethe

3
formeristargetedagainstextracellularpathogensandthelatteristargetedagainstintracellular
pathogens.Intheuterus,theinnateimmunesystemplaysthemostimportantroleinthedefense
againstequineendometritis (Snyder2017). This canbeexplainedby the fact that the innate
immuneresponseallowsforamorerapidresponsethantheadaptiveimmuneresponse.While
the adaptive immune response does play a small role in uterine defense, it requires diligent
regulationsincetheformationofantibodiestowardsantigenicmaterial,inthisinstancesemen,
wouldbedisastrousforfuturefertility(Troedsson2006;Lyle2011;Foster2017).
Researchersfirstbelievedthatendometritiswasduetobacterialcontaminationatthe
timeofbreeding(PycockandAllen1990;Asburyetal.1980;Mitchelletal.1982;Liuetal.1985;
Watsonetal.1987a;Watsonetal.1988;Williamsonetal.1987;Troedsson1991).Pycocketal.
(1990)characterized the inflammatory response inmares followingbacterial inoculationwith
Streptococcuszooepidemicus.Amassiveinfluxofneutrophilsintotheuterinelumenwithin30
minutes of inoculation occurred in all mares. Since that study, the inflammatory pathway
following bacterial inoculation has been further elucidated. An increase in arachidonic acid
metabolitesleadstoincreasedvascularpermeability,causingneutrophilinfiltration(Watsonet
al.1987b;LeBlancandCausey2009).ProstaglandinE2, leukotrieneB4aswellasother serum
proteins arepresentwithin theuterine lumenat this time andmay act as chemoattractants
(Watsonetal.1987a;Watsonetal.1987c;Watsonetal.1988;Pycocketal.1990).Antibodies,
suchasIgGorcomplement(C3)thenattachtothesurfaceofthebacteria,providingbindingsites
for neutrophil attachment, which allows opsonisation to occur. This leads to bacterial
phagocytosisbytheneutrophils.Complement (C3)and IgG levelsdecreasewithinthefirst24
hours, which is likely a result of increased consumption during the inflammatory response

4
(Troedsson1991).Thisinitialresponseisfollowedbyasecondarypeakinneutrophilnumbers
twelve hours following inoculation. This secondary peak is without a concurrent increase in
serumproteins,butinsteadisaccompaniedbyanincreaseinlocalimmunoglobulinsecretions
(IgA) (Williamson et al. 1987). Immunoglobulins identified in the uterus of mares following
bacterialinoculationconsistofIgA,IgG,IgG(T)andIgM(KenneyandKahleel1975;Asburyetal.
1980),where IgA isproducedbytheendometriumandIgGpassivelydiffuses intotheuterine
lumenfromtheperipheralcirculation(Mitchelletal.1982;Liuetal.1985).Inflammatoryexudate
is removed within 48 hours following inoculation via uterine contractions caused by
prostaglandinF2αrelease,asevidencedbylackofintra-luminalfluidvisibleonultrasound.Excess
fluidisalsoremovedvialymphaticsfollowingclosureofthecervix.
The inflammatory response to deposition of semen in the uterus is believed to be
essentiallyidenticaltothatdescribedabove,andhasbeenbettercharacterized(Kotilainenetal.
1994; Katila et al. 1995). Spermatozoa cause post-breeding inflammation via neutrophil
chemotaxisandcomplementactivation(Troedsson1995).Morespecifically,oncespermenter
theuterus,theyactivatecomplementpresentwithinuterinesecretions.Thisleadstocleavage
ofC5factorintoC5aandC5b,whichcreatesachemotacticsignaltoneutrophils.Asinbacterial
endometritis, prostaglandins, leukotriene B4 and other serum proteins may also act as
chemoattractants(Watsonetal.1987a;Watsonetal.1987c;Watsonetal.1988;Pycocketal.
1990). Neutrophils enter the uterine lumen, become activated and bind to sperm via
complementC3bfactororcomplement-independentfactors,allowingforthe initiationofthe
inflammatoryresponse.Furthermore,neutrophilextracellulartraps(NETs)andtraditionalligand
binding may enhance this neutrophil attachment, ultimately leading to phagocytosis of the

5
spermatozoa by neutrophils (Dahms and Troedsson 2002; Alghamdi and Foster 2005).
Concurrently, prostaglandin F2α, a metabolite of arachidonic acid via the cyclooxygenase
pathway, is released within the uterine lumen, initiating uterine contractions to assist with
physical evacuation of fluid and debris (Troedsson et al. 1995). Activated neutrophils are
responsibleforprostaglandinrelease,andthisispartofthenormalresponsetobreedinginan
attempt to clear the uterus of physiologic inflammation prior to embryo descent (Troedsson
1991).
Theimmediatecontractileresponsefollowingbreedingismediatedviaadrenergicnerves
andlocallyviaprostaglandinsinseminalplasma(Baeetal.2001).Concurrentoxytocinrelease
furtherheightenstheresponseofthemyometrium.Alexanderetal.(1995)demonstratedthat
oxytocinreleaseoccursfollowinginteractionwithastallion,includingstallionvocalizationand
physicalcontact,aswellasfollowingartificialinsemination.Madilletal.(2000)showedthatsuch
breedingstimulialsocausesuterinecontractionsinteasedmares.Asmeasuredbymyometrial
electrodes and a Grass polygraph, these contractions were found to be almost immediate
following the stimuli, andalmost always coincidedwithoxytocin release (Madilletal. 2000).
Oxytocin is released from the pituitary gland and binds to endometrial receptors,which are
upregulatedwhileestrogenlevelsriseandprogesteronelevelsdecrease,ondays14-17ofthe
estruscycle(StullandEvans1986;Jonesetal.1991).Therefore,theresponsetooxytocinrelease
isgreatestduringestrus.Thisisalsotruewhenexogenousoxytocinisadministeredduringestrus
anddiestrus.Myometrialcontractionsaremorefrequentbutshortlivedinestruscomparedto
diestrus,likelyduetoanincreasednumberofreceptors(Troedssonetal.1995).Oxytocinbinding
toup-regulatedreceptorsleadstoprostaglandinsynthesisandcontributestoincreaseduterine

6
contractions(Sharpetal.1997).Inaddition,prostaglandinF2αisknowntoup-regulateoxytocin
receptors, creating a positive feedback loop. Interestingly, oxytocin can lead tomyometrial
contractionsviaactivationofdirectandindirectpathways.Whenactivationtakesplaceviathe
direct pathway, myometrial contractions occur regardless of prostaglandin levels; however,
prostaglandinlevelsmustbeelevatedformyometrialcontractionstotakeplaceviatheindirect
pathway. This was demonstrated through the administration of phenylbutazone to block
prostaglandin synthesis. Normal mares treated with oxytocin and phenylbutazone did have
myometrial contractions but these were without concurrent prostaglandin F2α metabolite
(PGFM)plasmaelevations.Additionally,whenthesemaresweretreatedwithphenylbutazone
only,theyshoweddecreaseduterineclearance(Cadarioetal.1995).Therefore,phenylbutazone
blocksprostaglandin-mediateduterinecontractions,butdoesnothaveaneffectonoxytocin-
mediatedclearance.Conflictingresultswerenotedbythesameresearchersin1999,whenPGFM
levelswereonlyelevatedinmareswithintra-uterinefluidfollowingtreatmentwithoxytocin,but
notinnormalmares.Theseworkershypothesizedthatuterineinflammationprimedtheuterus
forPGF2αrelease(Cadarioetal.1999).
Regardless of themechanisms involved,myometrial contractions lead to clearance of
inflammatoryproductsfromtheuterus.Uterinecontractions,incombinationwithuterinefolds,
contributetoclearanceviathecervix.Inaddition,clearancevialymphaticsisdependentupon
myometrialcontractions(LeBlancetal.1995).Ifthepressurewithintheuterusisnotsufficient,
lymphaticdrainagecannotoccur,leadingtofluidcollectioninthelumeninsteadofitsremoval
viathelymphatics(Causey2006).Amucociliaryapparatusalsoexistswithintheuterus,where

7
mucusisproducedbyendometrialcellsandthenpropelledbycilia.This,combinedwithuterine
contractions,minimizesbacterialattachmentandmaximizesremovalofinflammatorycellsand
debris from the uterus (Causey 2007). The combination of innate immune response and
mechanicalclearanceresultintheinflammatoryresponsebeingshort-livedfollowingbreeding
innormalmares(Troedsson1999).
1.3RISKFACTORSFORPBIE
Factors associated with susceptibility to PBIE in mares include poor perineal
conformation,age,anatomiclocationoftheuterus,andfluidretentionduringdiestrus(Ricketts
andAlonso1991;Troedssonetal.1993a;LeBlancetal.1994a;LeBlancetal.1998;Brinksoetal.
2003; Woodward et al. 2012; Canisso et al. 2016). Reproductive anatomy is an important
componentofthemare’suterinedefensesystem.Thethreeanatomicalbarriersthatprotectthe
uterus fromexternal contamination are: apposition and vertical angulation of the vulvar lips
(perineal conformation), the vestibulo-vaginal sphincter and the cervix (Pascoe 2007; Pycock
2009;Bergfelt2009).Mareswithnormalperinealconformationhaveatleasttwo-thirdsofthe
vulvarlengthbelowthepelvicbrim,lessthan10-degreeanteriortilttothevulva,andvulvarlips
thatarewellapposedtopreventfecalcontaminationofthevagina(Pascoe2007;Pycock2009
McCue2008;Canissoetal.2016).Perinealconformationmaybealteredfromnormalinsome
instances.Forexample,leanathleticmareswithverylittleperinealfatmayhavearecessedanus
leadingtocranialanglingofthevulva,thereforepullingmorethanone-thirdofthevulvaabove
thepelvicbrim (Hurtgen2006;Pascoe2007). Similar changesmayoccur inoldermareswith
increasedparity as seenby lengtheningof the vulvaand increase in theangleofdeclination

8
(Pascoe1979).Thevestibulo-vaginalsphincterisformedbymucosalfoldsatthedemarcationof
thevestibulefromthevaginaattheleveloftheexternalurethralorifice(Bergfelt2009).Mares
with an incompetent vestibulo-vaginal sphincter aspirate air into the vagina (pneumovagina)
during exercise, or following urination, a term commonly referred to as “wind-sucking”.
Pneumovagina leads to irritation and inflammation of the vagina, vestibule and cervix,
predisposingthemaretosecondarybacterialinfectionandendometritis(Pascoe2007;McCue
2008;Canissoetal.2016).
Abnormalitiesaffectingthecervixincludefibrosis,tortuosityandincompetence.Cervical
malfunctioncanoccurinoldermaidenmares,mareswithrepeatedembryorecoveryattempts
or following abortion or dystocia-induced damage (Pycock 2007; McCue 2008; LeBlanc and
Causey 2009). An inability to dilate the cervix following breeding, as seenwith fibrosis post
dystociaorinoldermaidenmares,canleadtodecreaseduterineclearance(Pycock2007).Reilas
etal. (2016) showed thatocclusionof thecervixbywayofacatheter for25hours following
artificialinsemination(AI)leadstoincreasedintra-uterinefluidvolumesandincreasedneutrophil
numberinnormalmares.Inaddition,theysawaworseningoffibrosisonendometrialbiopsies
instudymareswherefluidwasretainedforeither25hoursor24hourswithsomefluidremoved
at6hours,andspeculatedthatretentionof fluid intheuterusmay leadtoalterations inthe
inflammatoryresponse.
Other anatomical abnormalities include stretching of the broad ligamentwith cranial
displacementoftheuterusoverthepelvicbrim,whichleadstodecreaseduterineclearancedue
to simple gravitational effects (LeBlanc and Causey 2009). In fact, uterine clearance of a
radiocolloidasassessedbyscintigraphydemonstratedthatmareswithamoreventraluterine

9
cervicalanglecleared<30%of theradiocolloid intwohours,comparedtonormalmaresthat
cleared >50%. Mares with delayed clearance also tended to be over the age of 12, were
pluriparousandhadahistoryofendometritis(LeBlancetal.1998).Therefore,manyoftheserisk
factorsareaffectedbothbyageofthemareandparity.
Carnevale andGinther (1992) first showed thatolderponymares indiestrushad less
uterine contractility as assessed by trans-rectal ultrasound prior to and following breeding,
compared to youngermares under the same conditions. The oldermares also had a higher
incidenceofendometritis,asassessedbyendometrialbiopsy,andincreasedintra-uterinefluid.
Woodwardetal.(2012)examinedtheassociationbetweenmareage,uterinefluidretentionand
endometrial biopsy scores. In their study, mares over 9 years of age were more likely to
accumulatefluidthanyoungermaresandevenmoresowhenmareswereolderthan17years.
AhigherpercentageofoldermaresalsohadKenneybiopsyscoresofIIborIII.Ageistherefore
likelyassociatedwithamoresevereendometrialbiopsyscore.Ageandbiopsyscoreappearto
leadtoincreasedfluidretentionandPBIEsusceptibility.However,thisstudydidnotassessthe
effectsofparityasaconfoundingvariable,asthereproductivehistoryofthemaresinthisstudy
wasunknown.SimilarresultswerenotedformareageinastudybyBarbacinietal.(2003),where
theincidenceofuterinefluidaccumulationpost-breedingwashigherinmaresaged10-16years
oldcomparedtoyoungermares.Thisstudyalsorevealedthatbarrenmaresweremorelikelyto
demonstrateuterinefluidretentioncomparedtomaidenandpost-parturientmares.Increased
levels of uterine fluid (>2cm) post-AI were associatedwith lower pregnancy rates; per cycle
pregnancyrateswere41.9%inmareswithfluidretentioncomparedto56.2%inmareswithout
fluid. Clearly, pluriparous mares tend have an increased likelihood of abnormal perineal

10
conformation,anincompetentvestibulo-vaginalsphincterandafibroticcervix,allofwhichcan
contributetosusceptibilitytoPBIE(Hurtgen2006;LeBlancandCausey2009;Christoffersenand
Troedsson2017).
The association between uterine biopsy grade and susceptibility to chronic uterine
infections(CUI)wasassessedbyTroedssonetal.(1993a).Inthisstudy,maresweredividedinto
four categories based on severity of histologic findings (1=normal to 4=severe) and were
challengedwith a bacterial inoculation of Streptococcus zooepidemicus during estrus.Mares
were considered susceptible to CUI if endometrial cytology revealed >1 neutrophil/5hpf and
endometrialculturehad>20bacterialcoloniesat96hourspostchallenge.MareswithgradeI
endometrialbiopsiesweremorelikelytoberesistanttotheinoculation,whereasmareswitha
biopsy score of grade IIIweremore likely to be susceptible to CUI, indicating an association
betweenseverityofendometrialbiopsyscoreandsusceptibilitytoCUI.However,therewasno
associationbetweenbiopsygradesIIaorIIbandsusceptibility,limitingtheapplicationofbiopsy
gradetopredictsusceptibility.InasimilarstudybyWilliamsonetal.(1989),mareswereclassified
assusceptibleorresistanttoCUIbasedonreproductivehistoryandendometrialbiopsyscores.
Followinginoculation,mareswerere-classifiedbasedontheirabilitytoclearaninoculationof
Streptococcus zooepidemicus after 10 days. Susceptibility was more highly correlated with
reproductivehistorythanendometrialbiopsy.However,astudydesignlimitationexistedinthat
mares were only classified as susceptible ten days following bacterial inoculation, allowing
considerable time for less susceptiblemares toclear the infectionprior to thesampling time
point.Inaddition,biopsyscoreswerenotdefinedusingthecurrentendometrialbiopsygrading

11
system described by Kenney and Doig (1986), limiting the capability to compare with other
studies.
1.4PATHOPHYSIOLOGYOFPBIE
1.4.1HumoralandInnateImmunity
Muchresearchhas focusedonmaresusceptibilityversus resistance toacutebacterial
endometritisandPBIE.Possibledifferencesbetweenmareswithrespecttotheirabilitytoclear
intra-uterineinfectionwerefirstdescribedbyHughesandLoy(1969)andPetersonetal.(1969),
where some mares appeared to be resistant to infections, while others did not. The initial
inflammatoryreactionappearedtobethesameamongstthesemares,butdifferedwithtimein
thatsomemares(susceptible)maintainedthe infection,andothers (resistant)cleared it.This
suggestedthatanalteredinflammatorydefensemechanismmaybeinvolved.Subsequentwork
focusedonpossibledifferencesinthehumoralimmuneresponse,specificallydifferinglevelsof
immunoglobulinsbetweensusceptibleandresistantmares.Asburyetal.(1980)foundincreased
intra-uterinelevelsofIgA,IgGandIgTinmaressusceptibletoendometritisfollowingbacterial
inoculationwithStreptococcuszooepidemicus,comparedtonormalmares.Inaddition,Mitchell
etal.(1982)notedhighertotalimmunoglobulin,IgAandIgGinsusceptiblemares,asdefinedby
endometrialbiopsies.Similarly,Williamsonetal. (1983)foundelevated levelsof IgA inmares
withactiveinfections,comparedtoresistantmares.Conversely,Troedssonetal.(1993c)found
nosignificantdifferencesbetweensusceptibleandresistantmares.ThisstudyexaminedIgA,IgG
andcomplementcleavagefactorC3expressioninmaresthatwereeitherresistantorsusceptible
toabacterialinoculationofStreptococcuszooepidemicus.Inbothgroups,levelsofIgA,IgGand

12
C3decreasedinthefirst24hoursfollowinginoculation.LevelsofIgGandC3roseinresistant
mares by 36 hours, however they remained low in susceptiblemares, suggesting continued
bacterial opsonisation. No differences were detected in IgA levels between groups. The
susceptibilityofendometritiswasthusnotattributedtodeficienciesinimmunoglobulins.
Watson and Dixon (1993) showed upregulation of MHC Class II expression in the
endometrialepitheliumofsusceptiblemares,comparedtonormalmaresinestrus.Themajority
ofcellswithMHCClassIIexpressionconsistedofTlymphocytes,butalsoincludedsomeepithelial
cells. When assessed histologically, endometrial T lymphocytes were increased in non-bred
mares susceptible to endometritis, compared to resistantmares, both in estrus and diestrus
(Fumosoetal.2007).Followingartificialinsemination,PBIE-susceptiblemareshadhigherlevels
of T lymphocytes in diestrus only, compared to resistantmares (Fumosoet al. 2007). This is
interesting to the understanding of PBIE, since no differences in T lymphocyte numberwere
observedinnormalmaresregardlessofstageofcycle(WatsonandThomson1996).Tunonetal.
(1999)characterizedthetypeofTlymphocyteexpressioninnormalmares.TheyfoundthatCD4+
Tcellswereelevatedat6hoursfollowinginsemination,butnotat48hours.Nodifferencesin
CD8+Tcellswerenotedateithertimepoint.SusceptiblemaresalsohadmoreCD4+expression
thanCD8+expressionandmoreCD8+cellscomparedtoresistantmares(WatsonandThomson
1996),likelyduetopersistentantigenicstimulationinaffectedmares.
Alterations in uterineneutrophil chemotaxis, phagocytosis, andbactericidal activity in
susceptiblemareshavebeenotherareasofresearchinunderstandingPBIE.Decreasedopsonins
fromuterinesecretionsofmaressusceptibletobacterialendometritiswasnotedbyAsburyet
al.(1982).In1985,Liuetal.demonstratedthatuterinesecretionsofmaresinfusedwithbacterial

13
inoculum,withgradeIIIuterinebiopsiesandahistoryofuterineinfections,hadlessneutrophil
elasticity and chemotaxis compared tomareswith grade I biopsies and normal reproductive
histories.Theythenshowedthatneutrophilmigrationwasdelayedfollowing inoculationwith
Streptococcuszooepidemicusinsusceptiblemarescomparedtoresistantmares.Inafollow-up
study, Lui et al. (1986) found that neutrophil levels in susceptiblemareswere elevated and
remained as such from 15 to 25 hours post inoculation compared to resistant mares.
Interestingly,neutrophilmigrationwasdecreasedinsusceptiblemaresat20hours;however,by
25 hours, migration had resumed in these mares. Watson et al. (1987c) showed further
neutrophil dysfunction, in that uterine-derived neutrophils displayed decreased phagocytic
capabilities in susceptible mares compared to resistant mares pre-inoculation. Following
induction of inflammation with 1% oyster glycogen, susceptible mare neutrophils displayed
decreasedbactericidalactivitywhenincubatedfor24hourswithagenitalstrainofStreptococcus
zooepidemicus,asassessedbypercentsurvivalofbacteria.Themechanismsbywhichneutrophil
functioncouldbeaffectedinsusceptiblemareswereinvestigatedbyTroedssonetal.(1993d),
who compared phagocytic ability of neutrophils in susceptible and resistantmares following
bacterialinoculationinestruswithStreptococcuszooepidemicus.Theauthorsconcludedthatthe
decreasedphagocyticabilityofneutrophilsseeninsusceptiblemareswascausedbydifferences
inuterinesecretionsandnotbydysfunctionalityoftheneutrophilsthemselves.Thiswasmade
evident by the demonstration of adequate phagocytosis when neutrophils from susceptible
mares were placed in an optimal environment. In addition, susceptible mares in this study
actuallyhadimprovedchemotaxiscomparedtoresistantmares.ThisisincontrasttoLiuetal.
(1985) who did not see chemotaxis improvement when uterine lavage fluid containing

14
neutrophils taken from mares with grade III biopsies were placed in the presence of
chemoattractants. Troedson (1993d) suggested that differences in components of uterine
secretionscouldresultinvariationsinopsonisationbycomplementandantibodies(Troedsson
etal.1993d).Thiswassupportedbythedecreasedbactericidalactivityofneutrophilsseenin
vitrowhenfactorsrequiredfortheclassicalandalternatecomplementpathwayswereremoved
(Watsonetal.1988).Troedsson(1999)suggestedthatstationaryageduterinefluid,asisoften
the case with PBIE-susceptible mares, can lead to inactivation of complement proteins,
decreasingneutrophilphagocytic success. Furthermore,neutrophilsmusthavea surface (the
endometrium)onwhichtoperformphagocytosis,andthissurfacemaynotbeavailablewhen
there is intra-luminal fluid present (Troedsson 1999). In summary, susceptible mares have
functional neutrophils and the same rate of phagocytosis as resistant mares; however, the
functionalandphagocyticabilitiesofthoseneutrophilsappeartobereducedwhenexposedto
the altered uterine fluid of PBIE-susceptible mares. Susceptible mares also have increased
numbers of neutrophils for prolonged periods of time following induction of inflammation,
comparedtoresistantmares(Liuetal.1986;Watsonetal.1987c;NikolakopoulosandWatson
1997;WoodwardandTroedsson2013).Therefore,theuterinefluidofsusceptiblemarescauses
alteredneutrophilfunctionality,whichcouldcontributetodifferencesinboththetimingoftheir
presencewithintheuterinelumenandtheirlevels.
Increased intra-uterine neutrophil number and altered uterine secretions can have a
directeffectonspermatozoa,furthercontributingtodecreasedfertility.Spermatozoaincubated
intheuterinesecretionsofinflameduteriinvitrodemonstratedalteredvelocityandprogressive
motility, which was attributed to the presence of neutrophils binding to the spermatozoa

15
(Alghamdietal.2001).Someoftheseeffectscouldbemediatedbythepresenceorabsenceof
seminal plasma,whose effects on fertility and the post-breeding inflammatory response are
controversial.Inlargeconcentrations,seminalplasmahasbeenshowntobedetrimentaltothe
qualityofthawedsemen(Morrelletal.2014);however,italsomayshortenthedurationofpost-
breedinginflammationwhenpresentinsmallervolumes(Troedssonetal.2000;Troedssonetal.
2001). Mares inseminated with sperm, milk extender and seminal plasma demonstrated
decreaseduterinecontractionsandincreasedintrauterineneutrophilconcentrations,compared
toAIwithspermandextenderalone.Thissuggestsbothanincreasedintensityanddurationof
inflammation post-breeding when seminal plasma is used (Portus et al. 2005). However,
Troedssonetal.(2000)showedthatseminalplasmadecreasesneutrophilphagocytosisofsperm
and Alghamdi et al. (2004) demonstrated that seminal plasma suppresses neutrophil sperm
bindinginadose-dependentmanner,invitro.Fifteen-daypregnancyrateswerealsohigherin
maresinseminatedwithspermatozoaresuspendedinseminalplasmasuspensionwithoutsemen
extender compared to spermatozoa resuspended in semen extender alone (Alghamdi et al.
2004). It has thus been suggested that seminal plasma or some of its components could be
beneficialinthemanagementofPBIEinsusceptiblemares(Troedssonetal.2000;Alghamdiet
al.2004).
1.4.2UterineClearance
Failure to clearuterine fluidpost-breedinghasbecome themost important andmost
commonlyuseddefinitionforPBIE inmares(Brinskoetal.2003). This inabilitytoclear intra-
uterinefluidischaracteristicofthecondition,andoccursduetodefectsinthenormaluterine
clearancemechanisms,whichvarybystageoftheestrouscycle.Evansetal.(1986)demonstrated

16
the influence of reproductive steroid environment on uterine clearance in anestrus maiden
maresusinguterineinfusionofStreptococcuszooepidemicus,chromium-labeledmicrospheres,
andcharcoal. Maresinthestudyweretreatedwitheitherestradiolorprogesteroneandwere
infused with Streptococcus zooepidemicus, chromium-labeled microspheres, and charcoal.
Estrogen-treatedmareshadrapiduterineclearanceofmicrospheresandbacteriaby3dayspost-
inoculation. By comparison, uterine clearance of progesterone-treatedmareswas poor and
produced persistent bacterial infections following inoculation, demonstrating the inhibitory
effectsofprogesteroneonuterineclearance.Subsequently,in1987hisgroupdemonstrateda
correlation between decreased uterine clearance and age, where youngmares cleared non-
antigenicmarkersmoreefficientlythanoldmareseveninanestrogen-dominatedenvironment
(Evansetal. 1987). Troedssonand Liu (1991)demonstrated thatmares resistant tobacterial
infections cleared a significantly greater amount of intra-uterine non-antigenic chromium-
labelled microspheres (51Cr) than susceptible mares. Mares were classified as potentially
susceptibleorresistanttouterineinfectionbasedonreproductivehistory,biopsyscoreandper
rectum examination. They were challenged with an intra-uterine infusion of Streptococcus
zooepidemicusandinoculatedwithmicrospheres.Resistantmareshadanegligiblenumberof
microspheresrecoveredbyuterinelavage24hourspostinoculationwhereasalargenumberof
microsphereswererecoveredinsusceptiblemaresat96hourspostinoculation.
Uterine contractility also appears to be different between susceptible and resistant
mares.Troedssonetal.(1993b)usedelectromyography(EMG)followingsurgicalimplantationof
myometrialelectrodestoassessmyometrialactivity.Resistantmaresshowedgreaterintensity,
durationandfrequencyofmyometrialcontractionscomparedtosusceptiblemares, following

17
intrauterine bacterial challenge. Furthermore, LeBlanc et al. (1994a) infused the uterus of
resistantandsusceptiblemareswitha radiocolloidandmonitoredclearancevia scintigraphy.
Resistantmaresclearedtheproductwithintwohoursof infusion,whereassusceptiblemares
displayeddelayedclearance.Thesamegroup (Neuwirthetal.1994) showed that susceptible
maresonlyclearedlessthan15%oftheradiocolloidwithintwohoursofinfusion,comparedto
over50%oftheproductclearedbythistimepointinresistantmares.Thesefindingscouldinpart
beexplainedbysusceptiblemarespossiblyhavinginherentlyabnormalpatternsofpropagation
of myometrial contractions, leading to retention of the fluid. Mares with delayed uterine
clearance (DUC) weremore likely to have an opposing pattern of propagation compared to
normalmares.Morespecifically,insteadoftheuterinecontractionscommencingintheuterine
hornsfollowedbytheuterinebody,contractionsinaffectedmareswereinitiatedintheuterine
bodyandthentheuterinehorns (vonReitzensteinetal.2001;LeBlancandCausey2009). In
addition, Rigby et al. in 2001 showed that susceptible mares may have decreased uterine
clearanceduetoan intrinsiccontractiledefectof themyometrium.Reproductive tractswere
removedfromyoungnulliparousmaresaswellasfromolderresistantandsusceptiblemares,
and both longitudinal and circular strips of myometrium were utilized in isometric tension
studies.StripswereexposedtoKCl,oxytocinandPGF2αandresponseswerecomparedbetween
thethreegroups.Resultsofthisin-vitrostudyshowedthatthemyometriumofsusceptiblemares
generatedlesstensionthanthatofbothyoungnulliparousandolderresistantmares;however,
theywerenotabletodeterminetheexactmechanismbywhichthisoccurred.
Differing levels of nitric oxide (NO) may further contribute to decreased myometrial
contractilityand thereforedecreaseduterine clearance (Alghamdietal. 2005).Nitricoxide is

18
formedfromL-arginineviathenitricoxidesynthase(NOS)enzyme.ThreeisoformsofNOShave
been identified and includeneuronalNOS (nNOS), endothelialNOS (eNOS) and inducibleNO
(iNOS).BothiNOSandeNOShavebeenidentifiedintheuterusandtheirpatternsofexpression
vary; eNOS is constitutively produced whereas iNOS is induced by inflammatory conditions
(Moncada and Higgs 1993). It is likely that the persistence of inflammation seen in PBIE
susceptiblemares leads to increased levels ofNO (Woodwardet al. 2013b).Nitric oxide is a
smooth muscle relaxant, and high levels could account for the diminished myometrial
contractility seen inPBIE-susceptiblemares. In supportof this theory, susceptiblemareshad
higherlevelsofNOintheiruterinesecretions13hoursfollowingartificialinsemination,aswell
ashigherNOSmRNAexpressionintheirbiopsies,comparedwithresistantmares(Alghamdiet
al.2005).Differencesinnitricoxidelevelsbetweensusceptibleandresistantmareswerefurther
classifiedbyWoodwardetal. (2013b).Specifically, total intrauterineNO insusceptiblemares
increasedatsixand12hourspost-AI,whereaslevelsremainedstableinresistantmaresover24
hours. Uterine iNOSmRNA expression was not different between susceptible and resistant
maresatanytimepoint.Khanetal.(2018)performedinvitroexperimentswherebymyometrium
obtainedfromsusceptiblemareswastreatedwithanitricoxidesynthaseinhibitortodetermine
itseffectsoncontractility.Resultsshowedthatmyometrialcontractilityimprovedwhentreated
withtheinducibleNOsynthaseinhibitor1,400Wdihydrochloride.
Additional mechanisms for the delayed uterine clearance in susceptible mares may
include abnormalities in the electrical pattern for contractility, smooth muscle fatigue, and
unsuitablehormonesignaling(LeBlancandCausey2009).MareswithGradeIIIbiopsyscoreshave
upto40%lessuterineperfusioncomparedtonormalmares,duetoendometrialvesselelastosis,

19
leading to decreased amounts of hormones, neurotransmitter and inflammatory mediators
deliveredtotheuterus(Esteller-Vicoetal.2007).In1999,NikolakopoulosandWatsonshowed
that susceptible and resistant mares have similar patterns of systemic oxytocin secretion
followingAI;however,susceptiblemareshavelowerlevelsofPGFMcomparedtoresistantmares
followingbreeding.Prostaglandinmetabolite(PGFM)isusedasamarkerforlevelsofPGF2α,and
decreasedlevelsareattributedtoapossibledefectinitsproductionorreleaseattheoxytocin
receptors.Asnotedabove,whenadministeredexogenousoxytocin,susceptiblemaresproduce
minimallevelsofPGFMcomparedtoresistantmares.Itisthereforepossiblethatdifferencesin
prostaglandinplayaroleintheintrinsicabnormalitiesinuterinecontractilityandthatthesemay
becausedbydecreaseduterineperfusion.
1.4.3MucociliaryApparatusandLymphatics
Somemares susceptible toPBIEmayhaveanalteredmucociliary apparatusandpoor
lymphaticdrainage.Freemanetal. (1990)showedthatmareswithurinepooling,asourceof
inflammation,havegreaterhistologicalstainingforcarboxylatedacidmucins intheuterus,as
comparedtonormalanestrusandtransitionalmares.Furthermore,Causeyetal.(2000)showed
thatmareswithDUChaveincreasedmucusproductioncomparedtonormalmares,asassessed
by endometrial biopsy staining. Increasedmucusproduction is believed tobe in response to
uterineinflammationorirritation(Freemanetal.1990).Thismucusalsotendstobelocalizedin
fibroticnestsandinareasof increasedinflammation.This is important,asmucusflowcanbe
affectedbyincreasesinvolumeandduetoanincreaseinviscosity.Dilutionofmucuswithintra-
luminalfluidcansimilarlyimpacttheeffectivenesswithwhichmucusisabletobindandmove
particles(Causey2007).Therefore,whileexcessmucuscanbedetrimental, insufficientmucus

20
allowsadhesionofsubstratestotheendometrialepithelium,leadingtodecreasedclearance.The
issueisfurthercompoundedasincreasedfluidvolumewithintheuterusleadstoseparationof
theendometrialfolds,whichnotonlyleadstoapoorenvironmentforneutrophilphagocytosis,
butalsoaffectsmucociliaryclearance.Causey(2006,2007)speculatedthatmucociliaryflowis
bestachievedwhentheendometrialwallsapposeeachotherclosely,andthattheciliabecome
ineffectivewhentheyarefurtherapartduetofluidaccumulations.Somemaressusceptibleto
endometritisdemonstrate lossofcilia fromtheendometrialepithelium, leadingtodecreased
movement of particles and fluid (Ferreira-Dias et al. 1994; Ferreira-Dias et al. 1999). More
specifically,mareswithgradeIIuterinebiopsiesdemonstratedpatchyareasofmicrovilliloss,and
mareswithgradeIIIuterinebiopsieshadareastotallydevoidofmicrovilluscellsandciliatedcells
onscanningelectronmicroscopy(SEM)andtransmissionelectronmicroscopy(TEM)(Ferreira-
Diasetal.1994;Ferreira-Diasetal.1999).Mareswithimpairedlymphatics,apendulousuterus,
or with impaired cervical drainage show increased fluid volumes within the uterus, further
contributingtotheproblem(LeBlancetal.1995).Whiletheexactmechanismsbywhichuterine
mucociliaryclearanceaffectsPBIEhavenotbeenelucidated,itisevidentthatitlikelycontributes
significantly.
Themovementofspermatozoatotheoviductfollowingbreedingiscausednotonlyby
uterine contractions, but also by endometrial epithelial ciliary flow (Troedsson et al. 2005).
Failureofciliarymovementcouldalsoaffectspermtransport,furthercontributingtodecreased
fertility.Infact,Causey(2006)statedthatspermtransportisalteredintheoviductsofsusceptible
mares,leadingtolowernumbersofspermatozoainthecaudalisthmus,anothermechanismby
whichfertilitymaybediminishedinsusceptiblemares.

21
1.4.4CytokineProfilesofSusceptibleandResistantMares
Cytokines are cell-derived polypeptides involved in inflammatory mediation, which
recently have been found to be different between PBIE susceptible and resistant mares.
Cytokinesareproducedatsitesofinflammationbystromal,endothelialandimmunecells,and
oncepresent,managetheinflammatoryresponse(FeghaliandWright1997;Tizard2013).They
can be categorized into pro-inflammatory cytokines, which as their name implies, promote
inflammation, and anti-inflammatory cytokines, which control inflammation and prevent its
persistence.Cytokinesmediatethetypeofinflammationthattakesplace,howlongitlastsand
whatother cellsmaybecome involved,byhavingbothagonisticandantagonist functionson
differenttypesofcells(FeghaliandWright1997).Someofthemostcommonlyidentifiedpro-
inflammatorycytokinesintheuterusincludethoseoftheinterleukin-1family(IL-1αandIL-1β),
interferon-γ(IFN-γ),tumornecrosisfactor-α(TNF-α)andinterleukin8(IL-8).IL-1αandIL-1βare
involved in therecruitmentofotherpro-inflammatorycytokines,suchasTNF-α. IFN-γaids in
neutrophilmigration from thevasculature into the siteof inflammation.TNF-α is involved in
prostaglandinproduction,apoptosisandaidsneutrophils,whichhavebeenrecruitedbyIL-8,in
their ability to neutralize pathogens (Tizard 2013). Anti-inflammatory cytokines such as
interleukin-10(IL-10)andinterleukin-1RA(IL-1RA),where interleukin-1RN(IL-1RN) isthegene
whichencodes for IL-1RA,are important for theregulationof inflammation. In fact, IL-10has
beenshowntoinhibittheproductionofIL-1αandIL-1β,IL-6,IL-8andTNF-αinhumans(Tizard
2013). Finally, interleukin-6 (IL-6) can act as both a pro-inflammatory and anti-inflammatory
cytokine (Tizard 2013) and canbe present in the uterus ofmares following an inflammatory

22
stimulus(Woodwardetal.2013a).Asimplifiedschematicofinteractionsbetweencytokinescan
befoundinFigure1.
DifferinglevelsofendometrialmRNAexpressionofthesecytokinescanbenotedbetween
resistant and susceptible mares at different time points following insemination or bacterial
inoculation (summarized in Tables 1 and 2). Briefly, PBIE-susceptible mares appear to have
elevatedbaselinemRNAexpressionofpro-inflammatorycytokines(IL-1β,IL-6,IL-8andTNF-α)
anddecreasedmRNAexpressionofanti-inflammatorycytokines(IL-10) inestruscomparedto
normalmares(Fumosoetal.2003;Fumosoetal. 2007).FollowingAI,PBIEsusceptiblemares
havepersistentelevationsofpro-inflammatorycytokineexpression(IL-1β,IL-8andTNF-α)atday
7post-AI(Fumosoetal.2003;Fumosoetal.2007).IncreasedmRNAexpressionofIL-8at3hours
post-inoculationwithE.coliwasfoundinnormalmarescomparedtoPBIEmares(Christoffersen
etal.2012),andmRNAexpressionoftheanti-inflammatorycytokine,IL-10waselevatedbyday
7post-AIinsusceptiblemares(Fumosoetal.2007).Bycomparison,however,Woodwardetal.
(2013a)foundthatsusceptiblemareshadlowerlevelsofanti-inflammatoryIL-1RNandIL-10and
pro-inflammatory IL-6 at six hours post-AI, and found no differences at 24 hours post-AI
compared to resistantmares. In contrast to Fumosoet al. (2007), other studies did not find
differences in IL-8 expression between susceptible and resistant mares. Disparities amongst
studiescouldbeattributedtodifferencesinthestudydesignincludingmareinclusioncriteria,
samplesizeandtimepoints.IncreasedmRNAexpressionofpro-inflammatorycytokinesoccurs
innormalandsusceptiblemares followingAIaswellaspost infusionofextender,phosphate
bufferedsaline(PBS)andseminalplasmainnormalmares(Palmetal.2008).

23
1.5DIAGNOSISOFPBIE
AsunderstandingofPBIEpathophysiologyevolvedoverthepastfourdecadesofscientific
investigation,sohasthedefinitionanddiagnosisofthiscondition.Earlystudies(HughesandLoy
1969;Petersen1969)identifiedthatprolongedandexcessivefluidretentionoccurredfollowing
bacterial inoculation of somemares. These studies highlighted the impact of acute bacterial
endometritisonthefertilityofbroodmares.However,mareswithPBIEmayormaynothavea
bacterialcomponenttothedisease(Kotilainenetal.1994;Troedsson1995).Overtime,failure
ofuterineclearancebecame the focusof study,and the termDUCwasused todescribe the
condition(Evansetal.1986;LeBlancetal.1998;Rigbyetal.2001).Methodsto identifyDUC
mares included failure to clear radiocolloid, India ink or chromium-labelled microspheres.
However,thesemethodswereexpensiveandimpracticalforidentifyingmaresinclinicalpractice.
Brinsko(2003)recommendedthatmaresbedefinedashavingDUC/PBIEiftheyretainedgreater
than2cmofintrauterinefluidasassessedbytrans-rectalultrasoundexaminationat72hours
post-AI.Forthefirsttime,practitionershadapracticalmethodavailabletoidentifyPBIEmares
for treatment. However, this method only identified a lack of clearance and not persistent
inflammationwithintheuterus,alsotypicalofPBIE,limitingitsclinicalapplication.Troedssonet
al. (1993a)demonstrated correlationsbetweenhistological endometrial lesions andamare’s
clinical response to challenge with Streptococcus zooepidemicus, suggesting mares could be
identifiedas“potentiallysusceptible”onthebasisofanendometrialbiopsyscoreofIII.However,
correlationofsusceptibilitytoPBIEwithgradesofIIaorIIbwerepoor.Woodwardetal.(2013a,
2013b)andAlghamdietal. (2005)demonstratedasimilarapproachusing inseminationwith
killedorlivesperm,respectively.Mareswerefirstscreenedas“potentiallysusceptible”onthe

24
basisofbiopsyscore(Woodwardetal.2013a;Woodwardetal.2013b)aswellasaclinicalhistory
of low fertility rates, poor perineal conformation, and endometritis (Alghamdi et al. 2005).
Selected mares were inseminated and then monitored by trans-rectal ultrasound for fluid
retention up to 96 hours later. In addition, endometrial cytology at 96 hours was used to
demonstratepersistenceof the inflammatory response.Onlymareswithbothpersistenceof
fluidandcytologicalevidenceof inflammationat96hourspost-inseminationweredefinedas
beingsusceptibletoPBIE.Thisapproachhassincebeenusedinseveralresearchstudiestoselect
andclassifymaressusceptibletoPBIE,althoughthetimepointusedfordeterminationhasvaried
between 48, 72 and 96 hours across studies (Troedsson et al. 1991; Alghamdi et al. 2005;
Woodwardetal.2013a;Woodwardetal.2013b;Khanetal.2018).Fromaclinicalstandpoint,
thisstrictcriterionmightexcludemareswithlessseverediseasethatwouldbesubfertileand
mayprogresstomoreseverediseaseovertime.Inaddition,inseminationwithadoseofkilled
semen prior to breeding to diagnose PBIEmay define the condition clearly for experimental
studies,howeverit isnotpracticalforpractitionersinthefield. Inclusionofadditionalcriteria
such as perineal conformation, persistent endometrial edema (Pycock 2009), and breeding
history(subfertility,repeatedendometritisandhistoryoffluidaccumulationinpreviouscycles)
would likely further improve this definition and classification methodology. In other words,
mares could be classified as likely to be susceptible based on their reproductive history and
perineal conformation. Thiswould allow practitioners to plan for increasedmonitoring post-
breedingandtohaveapossibletreatmentplanprepared intheeventthatthemareshowed
signs of PBIE such as any accumulation of fluid, persistence of edema near ovulation or the
developmentofedemapost-ovulation.

25
1.6TREATMENTOFPBIE
TreatmentofPBIEcanbedividedintothreebroadcategories:improvingmechanicaluterine
clearance,dilutingandremovingdebris,andmodifyingandimprovingtheimmuneresponse.
1.6.1EcbolicTherapy
Ecbolic therapies including oxytocin and prostaglandin are shown to increase uterine
clearanceby increasingmyometrial activity (Troedsson 1997; LeBlanc andCausey 2009). The
effectsofoxytocinonuterineclearancewereshownwhenbothsusceptibleandresistantmares
cleared >90% of radiocolloid following its administration (20U IV). This is in comparison to
susceptible mares only clearing negligible amounts and normal mares clearing 50-80% of
radiocolloidwhennot treatedwithoxytocin (Leblancetal.1994b).Oxytocin isawidelyused
treatmentforPBIE(LeBlancandCausey2009),withrecommendeddosagesrangingfrom10-25U
giveninthefirst48hourspost-breeding.Cadarioetal.(1999)showedthatbothsusceptibleand
resistantmareshavedose-dependentresponsestooxytocinadministration,withdosesof10U
resultinginthestrongestandmostprolongeduterinecontractionsascomparedto5Uand2.5U.
Administration of oxytocin results in uterine pressure changes within 30-65 seconds of
administration, and high amplitude uterine contractions for 30 minutes, resulting in almost
immediatevoidanceoftheuterus(Cadarioetal.1999;LeBlancetal.2009).Propulsionofuterine
fluidwasdescribedbyAllen(1991)inonemarefollowingtreatmentinestruswithoxytocin(5U
IV).Intra-uterinefluidmovedfromthecranialuterus,tothecaudaluterusandintothecranial
vagina by 1-hour post administration, as assessed by trans-rectal ultrasound. Furthermore,
administrationofoxytocin(10USC,5UIV;10UIV)intheperi-ovulatoryperioddoesnotresultin
areductionoffertility(Allen1991).However,duetoitsshortdurationofaction,oxytocinmust

26
begiveneveryfewhours.Unfortunately,somemaresdonotrespondtooxytocinadministration,
andcontinuetoaccumulateuterinefluiddespitetreatment(Hurtgen2006;LeBlancandCausey
2009).Factorssuspectedtoaffecttheresponsetooxytocintreatmentincludedecreasedornon-
functional oxytocin receptors in the endometrium, a pendulous uterus, or a closed cervix
(LeBlanc and Causey 2009).Moreover, inappropriately high dosages of oxytocin result in an
abnormalpropagationofuterinecontractions,therebyrenderingthetreatmentunsuccessful.In
fact,CampbellandEngland(2002)determinedthatdosesabove30Ucauseddecreaseduterine
contractions compared to lower doses. In addition, Rasch et al. (1996) determined that
pregnancyrateswereimprovedwhen15UofoxytocinwasadministeredIVcomparedto25UIV
seven to eight hours following AI (78% versus 52%) in mares with any intra-uterine fluid
accumulationpriortobreeding.
Oxytocin treatment causes prostaglandin release from the endometrium, potentially
prolonging itsdurationofaction.Administrationofprostaglandinssuchascloprostenolalone
alsopromotesuterineclearance,withweakerbutmoreprolongedcontractionsnotedfollowing
its administration compared to oxytocin (LeBlanc and Causey 2009). Low amplitude uterine
contractionslastuptofourorfivehours,whichnotonlyleadstophysicalclearanceofthefluid
butalsoaidswithlymphaticflow(Combsetal.1996;Troedsson1997;LeBlancandCausey2009).
Prostaglandins may also cause cervical relaxation, further aiding mechanical clearance.
ProstaglandinE2(misoprostol)appliedtothecervixhasalsobeenadministeredinthetreatment
ofPBIE,asittooleadstodilationofthecervixduetoitsdirectabilitytobreakdowncollagenand
decreasetotalcollagencontentinthecervix(Tangetal.2007;LeBlancandCausey2009).Care
mustbetakenwhenadministeringprostaglandinsforthetreatmentofPBIE.Ifgiveninthefirst

27
48hours followingovulation,mares couldhavedecreasedpregnancy rates, due to luteolytic
effects and decreased progesterone production from the early developing corpus luteum
(Troedssonetal.2001a;Nieetal.2002;Brendemuehl2002;Nieetal.2003).However,when
prostaglandinwasgivennolessthan12hourspriortoovulation,noeffectsonluteal lifespan
wereseen.Therefore,ifPBIEmaresaretobetreatedwithprostaglandins,treatmentshouldbe
limitedtowithinthistimeframe(Troedssonetal.2001a;Nieetal.2003).
1.6.2UterineLavage
The use of ecbolic drugs is often combinedwith therapeutic uterine lavage to dilute
irritants, as well as to clear inflammatory debris and excessive fluid from the uterus. Post-
breeding lavage induces uterine contractions, attracts newneutrophils and stimulates serum
opsoninstoaidintheinflammatoryresponse,duetothemildlyirritatingnatureoftheinfusion
(Brinskoetal.1990).Performinguterinelavageat>4hourspost-breedingisnotdetrimentalto
fertility(Brinskoetal.1990;Brinskoetal.1991)andhasbeenshowntoincreasepregnancyrates
inPBIEsusceptiblemares,whencombinedwithecbolictherapy4-8hourspost-breeding(Knutti
etal.2000).Furthermore,pregnancyratesdoubledinthegrouplavagedat4-8hourscompared
to those lavaged at 18-20 hours post-breeding; although this effect was not statistically
significant.
Theeffectonfertilityofpre-breedinguterinelavagewithLactatedRinger’sSolution(LRS)
immediately prior to inseminationwith fresh-chilled semenwas explored byVanderwall and
Woods(2003).Inthisstudy,sixoutoftenmareshadresidualfluidasdetectedbytrans-rectal
ultrasound immediately following the procedure and five out of six of thesemares became
pregnant. Three of the four mares that did not accumulate fluid following lavage became

28
pregnant.Noneofthepregnantmareshadresidualfluidanygreaterthan39mmindepth.This
study thereforeprovidesanotheroption for the treatmentofPBIE,especially in caseswhere
morethanonebreedingisperformedinanattempttoclearanyinflammationcausedbythefirst
breeding.
1.6.3Immunomodulation
AdditionaltreatmentsforPBIEincludeapproachestocorrectthealteredinflammatory
responseofsusceptiblemares.Theseincludeadministrationofanti-inflammatoryagentssuchas
glucocorticoids, and intrauterine infusion of agents such as Mycobacterial cell wall extract
(MCWE), platelet rich plasma (PRP) and chelating agents such as tris-EDTA (LeBlanc 2010;
WoodwardandTroedsson2015;Canissoetal.2016).
Twomainstudiesevaluatedtheefficacyofglucocorticoidtreatmentontheclinicalsigns
ofPBIE (Dell’Aqua Jretal. 2006;Buccaetal. 2008). Fivedosesof0.1mg/kgofprednisolone
acetate every 12 hours, beginning in estrus,with the last treatment given at the timeof AI,
resulted in reductionofuterine fluid volumeand improved clarity inmareswithahistoryof
uterinefluidaccumulation.However,areductionofuterineneutrophilfunctioninallmareswas
also seen (Dell’ Aqua Jr et al. 2006). Similarly, treatment with a single dose of 50mg of
dexamethasoneIVatthetimeofbreedingdecreaseduterineedema,uterinefluidvolumeand
uterinefluidturbidityinsusceptiblemares(Buccaetal.2008).Inbothstudies,pregnancyrates
wereimprovedintreatedmareshavingahistoryofintra-uterinefluidaccumulation(Dell’Aqua
Jretal.2006)orthosemareswithmorethanthreePBIEriskfactors(Buccaetal.2008),butnot
innormalmares.Interestingly,nodifferencesinneutrophilfunctionwerenotedbetweencontrol
andtreatedcycles(Buccaetal.2008).Reductionofneutrophilfunctionmayoccurbyblocking

29
thecyclo-oxygenaseand5-lipoxygenasepathwaysofinflammation(WoodwardandTroedsson
2015).Glucocorticoidsmaybebeneficial due to their effectsonNOproductionand cytokine
expression,astheyreducemRNAexpressionofIL-1,whichplaysaroleintheactivationofiNOS
(Barnes1998);however,whenmeasuringthedirecteffectsofdexamethasone inPBIEonNO
production, no effect was noted in both iNOS mRNA expression and total intrauterine NO
(Woodwardetal.2013b).Woodwardetal.(2015)showedadecreaseinIL-1βmRNAexpression
following treatment with dexamethasone in susceptible mares, but no differences in other
cytokineswerenoted.Overall,itappearsthatthepost-breedinginflammatoryresponseisless
severeandresolvesmorerapidlywhenglucocorticoidsareused.
Other treatments aimed at altering the inflammatory response include MCWE or
Propionibacteriumacnes,whichenhancestheimmuneresponsebycausingcytokinereleaseand
byactivatingimmunecells(PhillipsandFilion2001;Fumosoetal.2003;Woodwardetal.2013b;
Woodwardetal.2015).TreatmentofPBIEsusceptiblemareswithMCWEreturnedintra-uterine
pro-inflammatorycytokinelevelstothosemoretypicalofresistantmares,morespecificallywith
significantdown-regulationofIL-1β(Fumosoetal.2003;Woodwardetal.2015).Treatmentof
PBIE-susceptiblemareswithMCWE24hourspriortobreedingalsodecreasedintrauterineNO
levelscomparedtonon-treatedcontrolmares(Woodwardetal.2013b).Thiscanbeexplained
by the fact that reducing the up-regulation of pro-inflammatory cytokines leads to less
inflammationandthereforefewerinflammatoryproducts,suchasNO.
Morerecently,intrauterineinfusionofPRPhasbeeninvestigatedasapotentialtherapy
forPBIE.Plasmaisthoughttoenhancephagocytosisbyincreasingtheamountofcomplement
presentinuterinefluid(LiuandTroedsson2008).Metcalfetal.(2012)examinedtheeffectof

30
PRP treatment prior to AI on endometrialmRNA expression of cytokines in PBIE susceptible
mares.Interleukin-1β,IL-6andIL-8weredown-regulatedandiNOSexpressionreduced,inPRP-
treatedcycles,comparedtountreatedcycles. Theseresultswereattributedto therenewing
effectsofPRPbytheformationofgrowthfactors,cytokinesandchemokines.TheeffectofPRP
onpregnancyrateswasassessed inPBIE-susceptiblebarrenmares(Metcalf2014).Treatment
with PRP resulted in decreased retention of intra-luminal fluidwhen compared to untreated
cycles,whichmayhaveledinparttosignificantlyincreasedpregnancyratesduringtreatment
cycles(67%pregnancyrate inthetreatedgroupcomparedto19%inthenon-treatedgroup).
Reghinietal. (2016) examined theeffect of PRP treatment fourhourspost-AI inmareswith
chronicdegenerativeendometritis (CDE).Percentageofneutrophilsaswellasdepthof intra-
uterinefluidaccumulationswerebothreduced24hourspost-AIinCDE-affectedmarestreated
withPRPwhencomparedtocontrolcycles.However,nosignificanteffectsofPRPtreatmenton
NO levels were noted in this study. Effects of treatment with PRP prior to AI compared to
treatmentfollowingAIinPBIE-susceptiblemareswasassessedinastudybySegabinazzietal.
(2017). Both treatmentprotocols resulted in decreasedneutrophils on endometrial cytology,
higherconceptionratesandlowerCOX-2immunostainingwhencomparedtountreatedcycles.
However,noeffectofPRPtreatmentonintra-uterinefluidlevelswasobserved.Althoughthere
aresomeinconsistenciesinresultsbetweenstudies,theoveralleffectthatPRPdecreasesuterine
inflammationisclear.Morestudiesarerequiredinordertodeterminetheexactmechanismsof
actionandoptimaltreatmentprotocol.
Two proteins identified in seminal plasma also appear to have beneficial effects on
breeding-inducedinflammation:lactoferrinandCRISP-3(Dotyetal.2011;Troedssonetal.2014),

31
andresearchhasfocusedonwhethertheseproteinscouldbeofbenefitinthetreatmentofPBIE.
Morespecifically,lactoferrinhasbeenshowntoenhanceneutrophilphagocytosisofnon-viable
spermatozoa, assisting with clearance of sperm from the uterus prior to embryo descent
(Troedssonetal.2014).Incontrast,CRISP-3decreasesneutrophilbindingtolivespermatozoa,
therebyimprovingchancesoffertilization(Dotyetal.2011).Fedorkaetal.(2016)examinedthe
effects of infusionof onebillion spermatozoa alone, or in combinationwith seminal plasma,
CRISP-3orlactoferrinonmRNAexpressionofselectcytokinesinPBIEmares.Asimilarstudywas
repeated in PBIE-resistantmares (Fedorka et al. 2017). Results showed that only lactoferrin
decreasedIL-1RNmRNAexpressionwhencomparedtoothertreatmentsandnoothereffectsof
infusionwithlactoferrinorCRISP-3werenotedinPBIE-susceptiblemares(Fedorkaetal.2016).
However, infusion of PBIE-resistantmareswith seminal plasma and spermatozoa resulted in
increasedmRNAexpressionof thepro-inflammatory cytokines IL-8 and IL-1β, anddecreased
levelsofTNF-α(Fedorkaetal.2017).InfusionoflactoferrinalonesuppressedTNF-αexpression,
butnoeffectsofCRISP-3werenoted.TheseresultswereincomparisontothemRNAexpression
notedintheothertreatmentcycles.Duetothepromisingeffectsoflactoferrinoninflammatory
cytokineexpression, itsuseasapost-breedingtreatmentwasexplored(Fedorkaetal.2018).
Intra-uterine infusion of three different doses of human recombinant lactoferrin (50µg/ml,
250µg/mland500µg/mldilutedin10mlofLRS)inPBIEmares6hoursfollowingAIresultedina
decreasedneutropil:epithelial cell ratioat18hourspost treatment,butdidnotchangepost-
breeding intra-uterine fluid accumulation compared to the control cycle. Increased mRNA
expressionofIL-1RNanddecreasedmRNAexpressionofIFNγwerealsoseenintreatedcycles.

32
Giventheseresults,treatmentwithlactoferrinseemspromising,yetmoreresearchisneeded
priortoitsclinicalusage.
1.6.4N-AcetylCysteine
N-acetyl cysteine (NAC) is a mucolytic agent used in the treatment of bacterial
endometritiswhenthepresenceofabiofilmissuspected,orincaseswhereexcessuterinemucus
is present inhorses (Ferris et al. 2016).N-acetyl cysteinedisruptsdisulphidebondsbetween
mucinpolymers, reducingmucusviscosity (LeBlanc2010). Inadditionto itsmucolyticeffects,
NAC has demonstrated antioxidant and anti-inflammatory properties in multiple models of
diseaseinbothhumanandveterinarymedicine.Thismaybeinpartduetoitsmetabolism,asit
isde-acetylatedtocysteine.Cysteineisaprecursorofglutathione,whichdecreasesfreeradical
formation (Bisselinget al. 2004). Increased levels of glutathionewere detected in circulating
neutrophilsinahumanclinicaltrialevaluatingNACforthetreatmentofcysticfibrosis.Airway
neutrophilcountssignificantlydecreasedfollowinghigh-doseNACtreatmentinthesepatients
(Tirouvanziametal.2006). Inahumanchronicobstructivepulmonarydisease(COPD) invitro
model,NACwasevaluatedforbothitsmucolyticpropertiesandroleasananti-oxidant(Cazzola
etal.2017).Thestudyfoundthathigherconcentrationsofthedrugreducedpro-oxidativefactors
andenhancedanti-oxidativefactors,inadditiontoinhibitingpro-inflammatorycytokines(IL-1β,
Il-8,TNF-α),suggestingadose-dependenteffectofNAC.
N-acetyl cysteine affects cytokine and LPS-inducedproductionofNO in rat peritoneal
macrophages, C6 glial cells and primary astrocytes (Pahan et al. 1997). This in vitro study
demonstratedthatwhenthesecellswereincubatedwithNACtwohourspriortotheadditionof
anLPSstimulus,NOproductionwas inhibitedat the levelof iNOS.Thiswasattributedtothe

33
inhibitionofnuclearfactorkappa-light-chainenhancerofactivatedBcells(NF-κB).Similareffects
were noted in a murine LPS-induced preterm labor model, where multiple aspects of the
inflammatory response were reduced by NAC pre-treatment (Paintlia et al. 2008). The anti-
inflammatoryeffectsofNACwerefurtherinvestigatedinaratmultiplesclerosismodel.Here,
treatmentwithNACblockedtheinductionofmonocyte/macrophageproducedTNF-α,IFN-γ,IL-
1βandiNOSaswellassplenocyteproductionofIFN-γ.Inaddition,splenocyteproductionofthe
anti-inflammatorycytokineIL-10wasincreasedintheNAC-treatedgroup,demonstratingaclear
impactofNAConcytokineproduction(Stanislausetal.2005).Finally,studieshaveexaminedthe
effectsofNAConNOproductionspecifically(Bergaminietal.2001;Majanoetal.2004).Inboth
studies, inflammation was induced by LPS in rat models. Treatment with NAC resulted in
inhibitionofiNOS,andMajanoetal.(2004)wereabletodemonstratethatthisinhibitionmay
haveoccurredpartiallyatthelevelofNF-κB.
N-acetyl cysteine has been effective for treatment of inflammation in horses in
experimentalstudies,specificallyincasesofmucosalinjuryintherightdorsalcolon,andcorneal
ulceration(Haffneretal.2003;Rottingetal.2003).ThesestudiesshowthatNAChasmorethan
justmucolyticeffects.Itsabilitytoaltercytokineexpression,iNOSinductionandneutrophillevels
providesevidencethatitmaybeusefulincasesofuterineinflammation,suchasPBIE.
MultiplerecentstudieshaveexaminedtheutilityofNACfortreatmentofendometritis
(Witteetal.2012;Gores-Lindholmetal.2013;Melkusetal.2013).Potentialadverseeffectsof
NAC on the uterus have been assessed (Witte et al. 2012; Melkus et al. 2013). First, oral
administration of NAC was evaluated in normal mares during estrus. The effects on mucus
viscosityweredeterminedone-andfive-daysfollowingtreatment,inadditiontoevaluationof

34
endometrial epithelial integrity, neutrophil infiltration and COX-2 staining on endometrial
biopsiestakenondayfive.OralNACtreatmenthadnoeffectonendometrialepithelialintegrity
ormucus viscosity, but did result in decreased infiltration of neutrophils and COX-2 staining
comparedtountreatedmares (Witteetal.2012).Furtherstudiesconsistedofhealthymares
treatedwith intra-uterine infusionof5%NACduringestrus (Melkusetal.2013).Endometrial
biopsiesweretakenat24-and72-hours followingtreatment inorder todetermineepithelial
integrity, neutrophil infiltration and COX-2 staining. Results showed that no damage to the
epitheliumwassustained,andthatat72hourspost-treatment,fewerneutrophilsandlessCOX-
2stainingwaspresentintreatedmarescomparedtocontrols.ThisdemonstratedthatNACwas
safe,andthatitmayalsohaveananti-inflammatoryeffectintheuterus(Melkusetal.2013).In
amoreclinicalstudy,Gores-Lindholmetal. (2013)assessedendometrialstructure,neutrophil
activityandreproductiveperformanceinbothfertileandbarrenmarestreatedwithintra-uterine
NAC.Theyshowedthatperipheralneutrophilreleaseofoxygenradicalswasreducedby3.0%
NACsolution,therebypossiblydiminishingexcessiveintra-uterineneutrophil-induceddamage,
andthatendometrialepithelialarchitecturewasmaintained.Decreasedmucuswasnotedatthe
histological level in fertile mares but not in barren mares treated with NAC. These results
conflicted with those by Witte et al. (2012) and could be explained by differing routes of
administrationofNAC.Thesestudiessuggestthatintrauterineinfusionororaltreatmentwith
NACisnotdetrimentalontheendometrium,mayormaynotaffectmucusproductioninfertile,
healthy mares, and likely reduces inflammation in the uterus, as evidenced by decreased
neutrophilinfiltrationandCOX-2staining.

35
Basedonitsanti-inflammatory,anti-oxidantandNOmodulatingeffects,theutilityofNAC
as anadjunct for the treatmentofPBIE in susceptiblemares isworthyof further study. This
researchwasthereforeundertakenwiththegoaloffurtheringourunderstandingoftheeffects
ofusingNACinthemanagementPBIEinmares.
1.7RESEARCHOBJECTIVES
Thespecificobjectivesofthisresearchwere:
1. TodetermineifmaressusceptibletoPBIEtreatedwithN-acetylcysteinewillresolvetheir
clinicalsignsmorerapidlyintreatedcyclescomparedtocontrolcycles,and
2. TodetermineifmaressusceptibletoPBIEwillhavelowerconcentrationsofnitricoxideand
a modified inflammatory cytokine profile following treatment with N-acetyl cysteine
comparedtocontrolcycles.

36
1.8REFERENCES
1. Alexander, S.L., Irvine, C.H.G., Shand,N. and Evans,M.J. (1995). Is luteinizing hormone
secretionmodulatedbyendogenousoxytocininthemare?Studiesontheroleofoxytocin
andfactorsaffectingitssecretioninestrousmares.Biol.Reprod.Mono.1,361-371.
2. Alghamdi, A.S. and Foster, D.N. (2005). Seminal DNase frees spermatozoa entangled in
neutrophilextracellulartraps.Biol.Reprod.73,1174-1181.
3. Alghamdi,A.S.,Foster,D.N.,Carlson,C.S.andTroedsson,M.H.T.(2005).Nitricoxidelevels
and nitric oxide synthase expression in uterine samples from mares susceptible and
resistanttopersistentbreeding-inducedendometritis.Am.J.Reprod.Immunol.53,230–237.
4. Alghamdi,A.S.,Foster,D.N.andTroedsson,M.H.T.(2004).Equineseminalplasmareduces
spermbindingtopolymorphonuclearneutrophils(PMNs)andimprovesthefertilityoffresh
semeninseminatedintoinflameduteri.Reproduction.127,593-600.
5. Alghamdi,A.S.,Troedsson,M.H.T.,Laschkewitsch,T.andXue,J.L.(2001).Uterinesecretion
frommareswithpost-breedingendometritisaltersspermmotioncharacteristicsinvitro.
Theriogenology.55,1019-1028.
6. Allen,W.E.(1991).Investigationsintotheuseofexogenousoxytocinforpromotinguterine
drainageinmaressusceptibletoendometritis.Vet.Rec.128,593-594.
7. Asbury,A.C.,Halliwell,R.E.W.,Foster,G.W.andLongino,S.J.(1980).Immunoglobulinsin
uterinesecretionsofmareswithdifferingresistancetoendometritis.Theriogenology.14,299-308.
8. Asbury,A.C.,Schultz,K.T.,Klesius,P.H.,Foster,G.W.andWashburn,S.M.(1982).Factors
affectingphagocytosis of bacteriabyneutrophils in themare’suterus. J. Reprod. Fertil.Suppl01,151-159.
9. Bae, S.-E., Corcoran,B.M. andWatson, E.D., (2001). Immunohistochemical studyof the
distribution of adrenergic and peptidergic innervation in the equine uterus and cervix.
Reproduction.122,275-282.

37
10. Barbacini,S.,Necchi,D.,Zavaglia,G.andSquires,E.L.(2003).Retrospectivestudyonthe
incidence of postinsemination uterine fluid in mares inseminated with frozen/thawed
semen.J.EquineVet.Sci.23,493-496.
11. Barnes,P.J.(1998).Anti-inflammatoryactionsofglucocorticoids:molecularmechanisms.
Clin.Sci.94,557-572.
12. Bergamini,S.,Rota,C.,Canali,R.,Staffieri,M.,Daneri,F.,Bini,A.,Giovannini,F,Tomasi,A.
andIannone,A.(2001).N-acetylcysteineinhibitsinvivonitricoxideproductionbyinduciblenitricoxidesynthase.NitricOxide.5,349-360.
13. Bergfelt, D.R. (2009). Anatomy and physiology of the mare. In: Equine Breeding
ManagementandArtificialInsemination,2nded,Ed:J.C.Samper,Elsevier,Missouri.pp113-
131.
14. Betteridge, K.J., Eaglesome, M.D., Mitchell, D., Floods, P.F. and Beriault, R. (1982).
Developmentofhorseembryosup to twenty-twodaysafterovulation:observationson
freshspecimens.J.Anat.135,191-209.
15. Bisseling,T.M.,Roes,E.M.,Raijmakers,M.T.M.,Steegers,E.A.P.,Peters,W.H.M.andSmits,
P. (2004).N-acetylcysteine restores nitric oxide –mediated effects in the fetoplacental
circulationofpreeclampticpatients.Am.J.Obstet.Gynecol.191,328-333.
16. Brendemuehl,J.P.(2002).Effectofoxytocinandcloprostenolonlutealformation,function
andpregnancyratesinmares.Theriogenology.58,623-626.
17. Brinsko,S.P.,Rigby,S.L.,Varner,D.D.andBlanchard,T.L. (2003).Apracticalmethodfor
recognizing mares susceptible to post-breeding endometritis. Proc. Am. Assoc. Equine.Pract.49,363–365.
18. Brinsko, S.P., Varner, D.D., Blanchard, T.L. and Meyers, S.A. (1990). The effect of
postbreedinguterinelavageonpregnancyratesinmares.Theriogenology.33,465-475.
19. Brinsko, S.P., Varner, D.D. and Blanchard, T.L. (1991). The effect of uterine lavage
performedfourhourspost-inseminationonpregnancyratesinmares.Theriogenology.35,1111-1119.

38
20. Bucca,S.,Carli,A.,Buckley,T,Dolci,G.andFogarty,U.(2008).Theuseofdexamethasone
administeredtomaresatbreedingtimeinthemodulationofpersistentmating induced
endometritis.Theriogenology.70,1093-1100.
21. Cadario,M.E.,Thatcher,W.W.,Klapstein,E.,Merrit,A.M.,Archbald,L.F.,Thatcher,M.J.and
LeBlanc,M.M.(1999).Dynamicsofprostaglandinsecretion,intrauterinefluidanduterine
clearance in reproductively normal mares and mares with delayed uterine clearance.
Theriogenology.52,1181-1192.
22. Cadario, M.E., Thatcher, M.-J. D. and LeBlanc, M.M. (1995). Relationship between
prostaglandinanduterineclearanceofradiocolloidinthemare.Biol.Reprod.Mono.1,495-500.
23. Campbell, M.L.H. and England, G.C.W. (2002). A comparison of the ecbolic efficacy of
intravenousandintrauterineoxytocintreatments.Theriogenology.58,473-477.
24. Canisso, I.F., Stewart, J. and Coutinho da Silva, M.A. (2016). Endometritis - managing
persistentpost-breedingendometritis.Vet.Clin.Equine.32,465-480.
25. Carnevale, E.M. and Ginther, O.J. (1992). Relationships of age to uterine function and
reproductiveefficiencyinmares.Theriogenology.37,1101-1115.
26. Causey,R.C.(2006).Makingsenseofequineuterineinfections:Themanyfacesofphysical
clearance.Vet.J.172,405-421.
27. Causey,R.C.(2007).Mucusandthemare:Howlittleweknow.Theriogenology.68,386-394.
28. Causey,R.C.,Ginn,P.,Katz,B.P.,Hall,B.J.,Anderson,K.J.andLeBlanc,M.M.(2000).Mucus
production by endometrium of reproductively healthy mares and mares with delayed
uterineclearance.J.Reprod.Fertil.Suppl56,333-339.
29. Cazzola,M.,Calzetta,L.,Facciolo,F.,Rogliani,P.andMatera,M.G.(2017).Pharmacological
investigationontheanti-oxidantandanti-inflammatoryactivityofN-acetylcysteineinan
exvivomodelofCOPDexacerbation.Resp.Res.18.

39
30. Christoffersen,M.andTroedsson,M.H.T.(2017). Inflammationandfertility inthemare.
Reprod.Dom.Anim.Suppl52,14-20.
31. Christoffersen,M.,Woodward, E.M., Bojesen,A.M., Petersen,M.R., Squires, E.L., Lehn-
Jensen, H. and Troedsson,M.H.T. (2012). Effect of immunomodulatory therapy on the
endometrial inflammatory response to induced infectious endometritis in susceptible
mares.Theriogenology.78,991-1004.
32. Combs,G.B.,LeBlanc,M.M.,Neuwirth,L.andTran,T.Q.(1995).EffectsofprostaglandinF2α,
cloprostenol and fenprostalene on uterine clearance of radiocolloid in the mare.
Theriogenology.45,1449-1455.
33. Dahms,B.J.andTroedsson,M.H.T.(2002).Theeffectofseminalplasmacomponentson
opsonisationandPMN-phagocytosisofequinespermatozoa.Theriogenology.58,457-460.
34. Dell’AquaJr.,J.A.,Papa,F.O.,Lopes,M.D.,Alvarenga,M.A.,Macedo,L.P.andMelo,C.M.
(2006).Modulationof acuteuterine inflammatory response after artificial insemination
withequinefrozensemen.Anim.Reprod.Sci.94,270-273.
35. Doty,A.,Buhi,W.C.,Benson,S.,Scoggin,K.E.,Pozor,M.,Macpherson,M.,Mutz,M.and
Troedsson,M.H.T.(2011).EquineCRISP3modulatesinteractionbetweenspermatozoaand
polymorphonuclearneutrophils.BiolReprod.85,157-164.
36. Esteller-Vico, A., Liu, I.K.M., Steffey, E.P., Vaghan, M.E., Brosnan, R.J. (2007). Effect of
vascular degeneration on utero-ovarian blood flow and perfusion in the cyclic mare.
Theriogenology.68,506.
37. Evans,V.(2012)HorseRacinginCanada-TheEconomicsofHorseRacingin2010.Equine
Canada,p.1–4.
38. Evans,M.J.,Hamer,J.M.,Gason,L.M.,Graham,C.S.,Asbury,A.C.andIrvine,C.H.(1986).
Clearanceofbacteriaandnon-antigenicmarkers following intra-uterine inoculation into
maidenmares:Effectofsteroidhormoneenvironment.Theriogenology.26,37–50.
39. Evans,M.J., Hamer, J.M., Gason, L.M. and Irvine, C.H. (1987). Factors affecting uterine
clearanceofinoculatedmaterialsinmares.J.Reprod.Fertil.Suppl.35,327–334.

40
40. Fedorka, C.E., Scoggin, K.E., Boakari, Y.L., Hoppe, N.E., Squires, E.L., Ball, B.A. and
Troedsson, M.H.T. (2018). The anti-inflammatory effect of exogenous lactoferrin on
breeding-induced endometritis when administered post-breeding in susceptible mares.
Theriogenology.114,63-69.
41. Fedorka,C.E.,Scoggin,K.E.,Woodward,E.M.,Squires,E.L.,Ball,B.A.,andTroedsson,M.H.T.
(2016).TheeffectofseminalplasmaproteinsonendometrialmRNAcytokineexpressionin
maressusceptibletopersistentmating-inducedendometritis.Reprod.Dom.Anim.52,89-96.
42. Fedorka,C.E.,Woodward,E.M.,Scoggin,K.E.,Esteller-Vico,A.,Squires,E.L.,Ball,B.A.and
Troedsson,M.H.T.(2017).Theeffectofcysteine-richsecretoryprotein-3andlactoferrinon
endometrialcytokinemRNAexpressionafterbreedinginthehorse.J.EquineVet.Sci.48,136-142.
43. Feghali,C.A.andWright,T.M.(1997).Cytokinesinacuteandchronicinflammation.Front.Biosci.2,12-26.
44. Ferreira-Dias,G.,Nequin,L.G.andKing,S.S.(1994).Morphologiccharacteristicsofequine
endometriumclassifiedasKenneycategoriesI,II,andIII,usinglightandscanningelectron
microscopy.Am.J.Vet.Res.55,1060-1065.
45. Ferreira-Dias, G., Nequin, L.G. and King, S.S. (1999). Morphologic comparisons among
equine endometrium categories I, II, and III, using light and transmission electron
microscopy.Am.J.Vet.Res.60,49-55.
46. Ferris,R.A.,McCue,P.M.,Borlee,G.I.,Loncar,K.D.,Hennet,M.L.andBorlee,B.R.(2016).Invitroefficacyofnonantibiotictreatmentsonbiofilmdisruptionofgram-negativepathogens
andaninvivomodelofinfectiousendometritisutilizingisolatesfromtheequineuterus.
Clin.Microbiol.54,631-639.
47. Foster, R.A. (2017) Female reproductive system and mammae. In: Pathologic Basis ofVeterinaryDisease.6thed.,J.F.Zachary,Elsevier,Missouri.pp1147-1193.
48. Freeman,D.A.,Roszel,J.F.,Slusher,S.H.andCastro,M.(1990).Variationinglycogenand
mucins in the equine uterus related to physiologic and pathologic conditions.
Theriogenology.33,799-808.

41
49. Freeman, D.A.,Woods, D.L., Vanderwall, D.K. andWeber, J.A. (1992). Embryo-initiated
oviductaltransportinmares.J.Reprod.Fertil.95,535-538.
50. Fumoso,E.A.,Aguilar,J.,Giguere,S.,Rivulgo,M.,Wade,J.andRogan,D.(2007).Immune
parameters inmaresresistantandsusceptibletopersistentpost-breedingendometritis:
effectsofimmunomodulation.Vet.Immunol.Immunopathol.118,30-39.
51. Fumoso, E., Giguere, S.,Wade, J., Rogan,D., Videla-Dorna, I. andBowden, R.A. (2003).
EndometrialIL-1β,IL-6andTNF-α,mRNAexpressioninmaresresistantorsusceptibleto
post-breeding endometritis. Effects of estrous cycle, artificial insemination and
immunomodulation.Vet.Immunol.Immunopathol.96,31-41.
52. Gores-Lindholm, A.R., LeBlanc, M.M., Causey, R., Hitchborn, A., Fayrer-Hosken, R.A.,
Kruger,M.,Vandenplas,M.L.,Flores,P.andAhlschwede,S.(2013).Relationshipsbetween
intrauterine infusion of N-acetylcysteine, equine endometrial pathology, neutrophil
function,post-breedingtherapy,andreproductiveperformance.Theriogenology.80,218-227.
53. Haffner, J.C., Fecteau, K.A. and Eiler, H. (2003). Inhibition of collagenase breakdown of
equinecorneasbytetanusantitoxin,equineserumandacetylcysteine.Vet.Ophthalmol.6,67-72.
54. Hughes,J.P.andLoy,R.G.(1969).Investigationsoftheeffectofintrauterineinoculationof
Streptococcuszooepidemicusinthemare.Proc.15th.Ann.Conv.Am.Assoc.EquinePract.,285-289.
55. Hurtgen,J.P.(2006).Pathogenesisandtreatmentofendometritisinthemare:Areview.
Theriogenology.66,560-566.
56. Jones,D.M.,Fielden,E.D.andCarr,D.H.(1991).Somephysiologicalandpharmacological
factorsaffectinguterinemotilityasmeasuredbyelectromyographyinthemare.J.Reprod.Fertil.Suppl44,357-368.
57. Katila, T. (1995). Onset and duration of uterine inflammatory response of mares after
inseminationwithfreshsemen.Biol.Reprod.Mono.1,515-517.
58. Kenney,R.M.(1978).Cyclicandpathologicchangesofthemareendometriumasdetected
bybiopsy,withanoteonearlyembryonicdeath.J.Am.Vet.Med.Assoc.172,241–62.

42
59. Kenney, R.M. and Doig, P.A. (1986). Equine Endometrial Biopsy. In:Current Therapy inTheriogenology.2nded.,D.A.Morrow,Saunders,Philadelphia.pp723-729.
60. Kenney,R.M.andKhaleel,S.A.(1975).Bacteriostaticactivityofthemareuterus:aprogress
reportonimmunoglobulins.J.Reprod.Fertil.Suppl23,357-358.
61. Khan,F.A.,Chenier,T.S.,Foster,R.A.,Hewson,J.andScholtz,E.L.(2018).Endometrialnitric
oxide synthase activity inmares susceptibleor resistant topersistent breeding-induced
endometritisandtheeffectofaspecific iNOS inhibitor invitro.Reprod.Dom.Anim.53,718-724.
62. Knutti,B.,Pycock,J.F.,vanderWeijden,G.C.andKupfer,U.(2000).Theinfluenceofearly
post-breeding uterine lavage on pregnancy rate in mares with intrauterine fluid
accumulationsafterbreeding.EquineVet.J.12,267-270.
63. Kotilainen,T.,Huhtinen,M.andKatila,T.(1994).Sperm-inducedleukocytosisintheequine
uterus.Theriogenology.41,629-636.
64. LeBlanc,M.M.(2010).Advancesinthediagnosisandtreatmentofchronicinfectiousand
post-mating-inducedendometritisinthemare.Reprod.Dom.Anim.Suppl45,21-27.
65. LeBlanc,M.M.andCausey,R.C.(2009).Clinicalandsubclinicalendometritisinthemare:
boththreatstofertility.Reprod.Dom.Anim.Suppl3,10-22.
66. LeBlanc, M.M., Johnson, R.D., Calderwood Mays, M.B. and Valderrama, C. (1995).
LymphaticclearanceofIndiainkinreproductivelynormalmaresandmaressusceptibleto
endometritis.Biol.Reprod.Mono.1,501–506.
67. LeBlanc,M.M.,Neuwirth,L.,Asbury,A.C.,Tran,T.,Mauragis,D.andKlapstein,E.(1994a).
Scintigraphicmeasurementofuterineclearanceinnormalmaresandmareswithrecurrent
endometritis.EquineVet.J.26,109–113.
68. LeBlanc,M.M.,Neuwirth, L., Jones, L., Cage, C. andMauragis, D. (1998).Differences in
uterinepositionofreproductivelynormalmaresandthosewithdelayeduterineclearance
detectedbyscintigraphy.Theriogenology.50,49–54.

43
69. LeBlanc,M.M., Neuwirth, L.,Mauragis, D., Klapstein, E. and Tran, T. (1994b). Oxytocin
enhancesclearanceofradiocolloidfromtheuterinelumenofreproductivelynormalmares
andmaressusceptibletoendometritis.EquineVet.J.26,279-282.
70. Liu,I.K.M.,Cheung,A.T.W.,Walsh,E.M.andAyin,S.(1986).Thefunctionalcompetenceof
uterine-derived polymorphonuclear neutrophils (PMN) from mares resistant and
susceptibletochronicuterineinfection:Asequentialmigrationanalysis.Biol.Reprod.35,1168-1174.
71. Liu, I.K.M., Cheung, A.T.W., Walsh, E.M., Miller, M.E. and Lindenberg, P.M. (1985).
Comparisonofperipheralbloodanduterine-derivedpolymorphonuclearleukocytesfrom
maresresistantandsusceptibletochronicendometritis:Chemotacticandcellelastimetry
analysis.Am.J.Vet.Res.46,917-920.
72. Liu,I.K.M.andTroedsson,M.H.T.(2008).Thediagnosisandtreatmentofendometritisin
themare:Yesterdayandtoday.Theriogenology.70,415-420.
73. Lyle, S. (2011). Immunological considerations. In: Equine Reproduction. 2nd ed. Ed:
McKinnon,A.O.,Squires,E.L.,Vaala,W.E.,Warner,D.D.,Wiley-Blackwell,NewJersey.pp.
2587-2596.
74. Madill,S.,Troedsson,M.H.T.,SantschiE.M.andMalone,E.D.(2002).Dose-responseeffect
ofintramuscularoxytocintreatmentonmyometrialcontractionofreproductivelynormal
maresduringestrus.Theriogenology.58,479-481.
75. Majano, P.L., Medina, J., Zubia, I., Sunyer, L., Lara-Pezzi, E., Maldonado-Rodriguez, A.,
Lopez-Cabrera,M. andMoreno-Otero, R. (2004). N-acetyl-cysteinemodulates inducible
nitricoxidesynthasegeneexpressioninhumanhepatocytes.J.Hepatol.40.
76. McCue,P.M.(2008).Theproblemmare:Managementphilosophy,diagnosticprocedures,
andtherapeuticoptions.J.EquineVet.Sci.28,619-626.
77. Melkus,E.,Witte,T.,Walter,I.,Heuwieser,W.andAurich,C.(2013).Investigationsonthe
endometrialresponsetointrauterineadministrationofN-acetylcysteineinoestrousmares.
Reprod.Dom.Anim.48,591-597.

44
78. Metcalf, E.S. (2014). The effect of platelet-rich plasma (PRP) on intraluminal fluid and
pregnancyrates inmaressusceptible topersistentmating-inducedendometritis (PMIE).
EquineVet.Sci.34,128.
79. Metcalf,E.S.,Scoggin,K.andTroedsson,M.H.T.(2012).Theeffectofplatelet-richplasma
on endometrial pro-inflammatory cytokines in susceptible mares following semen
deposition.J.EquineVet.Sci.32,498.
80. Mitchell, G., Liu, I.K.M., Perryman, L.E., Stabenfeldt, G.H. and Hughes, J.P. (1982).
Preferentialproductionandsecretionofimmunoglobulinsbytheequineendometrium–a
mucosalimmunesystem.J.Reprod.Fert.Suppl.32,161-168.
81. Moncada,S.andHiggs,A.(1993).TheL-arginine-nitricoxidepathway.N.Engl.J.Med.329,2002-2012.
82. Morrell, J.M., Georgakas, A., Lundeheim, N., Nash, D., Davies Morel, M.C.G. and
Johannisson,A.(2014).Effectofheterologousandhomologousseminalplasmaonstallion
spermquality.Theriogenology.82,176-183.
83. Neely, D.P., Kindahl, H., Stabenfeldt, G.H., Edqvist, L.E. and Hughes, J.P. (1979).
Prostaglandin release patterns in the mare: physiological, pathophysiological and
therapeuticresponses.JReprod.Fert.Suppl27,181-189.
84. Neuwirth,L.,LeBlanc,M.M.,Mauragis,D.,Klapstein,E.andTran,T.(1994).Scintigraphic
measurementofuterineclearanceinmares.Vet.Radiol.Ultrasound.36,64-68.
85. Nikolakopoulos, E. and Watson, E.D. (1997). Does artificial insemination with chilled,
extended semen reduce the antigenic challenge to the mare’s uterus compared with
naturalservice?Theriogenology.47,583-590.
86. Nikolakopoulos, E. and Watson, E.D. (1999). Uterine contractility is necessary for the
clearance of intrauterine fluid but not bacteria after bacterial infusion in the mare.
Theriogenology.52,413-423.
87. NewcombeJ.R.(1997).Theeffectoftheincidenceanddepthofintra-uterinefluidinearly
dioestrusonpregnancyrateinmares.Pferdeheilkunde.13,545.

45
88. Nie,G.J., Johnson, K.E.,Wenzel, J.G.W. andBraden, T.D. (2002). Effect of periovulatory
ecbolicsonlutealfunctionandfertility.Theriogenology.58,461-463.
89. Nie,G.J.,Johnson,K.E.,Wenzel,J.G.W.andBraden,T.D.(2003).Lutealfunctioninmares
followingadministrationofoxytocin,cloprostenolorsalineonday0,1or2post-ovulation.
Theriogenology.60,1119-1125.
90. Overbeck, W., Witte, T.S. and Heuwieser, W. (2011). Comparison of three diagnostic
methodstoidentifysubclinicalendometritisinmares.Theriogenology.75,1311-1318.
91. Pahan,K.,Sheikh,F.G.,Namboodiri,A.M.S.andSingh,I.(1997).N-acetylcysteineinhibits
induction of NO production by endotoxin or cytokine stimulated rat peritoneal
macrophages,C6glialcellsandastrocytes.FreeRadic.Biol.Med.24,39-48.
92. Paintlia, M.K., Paintlia, A.S., Singh, A.K. and Singh, I. (2008). Attenuation of
lipopolysaccharideinducedinflammatoryresponseandphospholipidsmetabolismatthe
feto-maternalinterfacebyN-acetylcysteine.Pediatr.Res.64,334-339.
93. Palm,F.,Walter,I.,Budik,S.,Kolodziejek,J.,Nowotny,N.andAurich,C.(2008).Influence
ofdifferentsemenextendersandseminalplasmaonPMNmigrationandonexpressionof
IL-1β,IL-6,TNF-αandCOX-2mRNAinequineendometrium.Theriogenology.70,843-851.
94. Pascoe,R.R.(1979).Observationsonthelengthandangleofdeclinationofthevulvaand
itsrelationtofertilityinthemare.J.Reprod.Fertil.Suppl27,299-305.
95. Pascoe,R.R.(2007).Vulvarconformation.In:CurrentTherapyinEquineReproduction.1st
ed.,Ed:J.C.Samper,J.F.Pycock,A.O.McKinnon,SaundersElsevier,Missouri.pp140-145.
96. Peterson, F.B., McFeely, R.A. and David, J.S.E. (1969). Studies on the pathogenesis of
endometritisinthemare.Proc.Am.Assoc.Equine.,279-282.
97. Phillips,N.C.andFilion,M.C.(2001).Therapeuticpotentialofmycobacterialcell-wallDNA
complexes.ExpertOpin.Investig.Drugs10,2157-2165.
98. Portus, B.J., Reilas, T. and Katila, T. (2005). Effect of seminal plasma on uterine
inflammation,contractilityandpregnancyratesinmares.EquineVet.J.37,515-519.

46
99. Pycock,J.F.(2007).Therapyformareswithuterinefluid,1stedn,Ed:CurrentTherapyin
Equine Reproduction. 1st ed., Ed: J. C. Samper, J.F. Pycock, A.O. McKinnon, Saunders
Elsevier,Missouri.pp93-104.
100. Pycock, J.F. (2009). Breeding management of the problem mare. In: Equine Breeding
ManagementandArtificialInsemination,2nded,Ed:J.C.Samper,Elsevier,Missouri.pp139-
164.
101. Pycock,J.F.andAllenW.E.(1990).Inflammatorycomponentsinuterinefluidfrommares
withexperimentallyinducedbacterialendometritis.EquineVet.J.22,422-425.
102. Rasch, K., Schoon, H.A., Sieme, H. and Klug, E. (1996). Histomorphological endometrial
status and influence of oxytocin on the uterine drainage and pregnancy rate inmares.
EquineVet.J.28,455-460.
103. Reghini, M.F.S., Neto, C.R., Segabinazzi, L.G., Castro Chaves, M.M.B., Dell’Aqua, C.P.F.,
Bussiere,M.C.C.,Dell’AquaJr.,J.A.,Papa,F.O.andAlvarenga,M.A.(2016).Inflammatory
response in chronicdegenerativeendometritismares treatedwithplatelet-richplasma.
Theriogenology.86,516-522.
104. Reilas,T.,RiveraDelAlamo,M.M.,Liepina,E.,Yeste,M.andKatila,T.(2016).Effectsonthe
equine endometriumof cervical occlusion after insemination.Theriogenology.85, 617–624.
105. Ricketts,S.W.andAlonso,S.(1991).Theeffectofageandparityonthedevelopmentof
equinechronicendometrialdisease.EquineVet.J.23,189–192.
106. Rigby, S.L., Barhoumi, R., Burghardt, R.C., Colleran, P., Thompson, J.A., Varner, D.D.,
Blanchard,T.L.,Brinsko,S.P.,Taylor,T.,Wilkerson,M.K.andDelp,M.D.(2001).Mareswith
delayeduterine clearancehavean intrinsicdefect inmyometrial function.Biol.Reprod.65,740–747.
107. Rotting,A.K.,Freeman,D.E.,Eurell,J.A.C.,Constable,P.D.andWallig,M.(2003).Effectsof
acetylcysteineandmigrationofresidenteosinophilsinaninvitromodelofmucosalinjury
andrestitutioninequinerightdorsalcolon.Am.J.Vet.Res.10,1205-1212.

47
108. Segabinazzi,L.G.,Friso,A.M.,Correal,S.B.,Crespilho,A.M.,Dell’AquaJr,J.A.,Miro,J.,Papa,
F.O.,Alvarenga,M.A. (2017).Uterineclinical findings, fertility rate, leucocytemigration,
and COX-2 protein levels in the endometrial tissue of susceptible mares treated with
platelet-richplasmabeforeandafterAI.Theriogenology.104,120-126.
109. Sharp,D.C., Thatcher,M.-J., Salute,M.E. and Fuchs,A.-R. (1997). Relationshipbetween
endometrialoxytocinreceptorsandoxytocin-inducedprostaglandinF2αreleaseduringtheoestrouscycleandearlypregnancyinponymares.JReprod.Fert.109,137-144.
110. Stanislaus,R.,Gilg,A.G.,Singh,A.K.,Singh, I. (2005).N-acetyl-L-cysteineamelioratesthe
inflammatory disease process in experimental autoimmune encephalomyelitis in Lewis
rats.J.Autoimmun.2.
111. Stull, C.L. andEvans, J.W. (1986).Oxytocinbinding in theuterusof the cyclingmare. J.EquineVet.Sci.6,114-119.
112. Snyder,P.W.(2017).Diseasesofimmunity.In:PathologicBasisofVeterinaryDisease.6thedn.,J.F.Zachary,Elsevier,Missouri.pp242-285.e5.
113. Tang, O.S., Gemzell-Danielsson, K. and Ho, P.C. (2007). Misoprostol: pharmacokinetic
profiles,effectsontheuterusandside-effects.Int.J.Gynaecol.Obstet.99,S160-S167.
114. Tirouvanziam,R.,Conrad,C.K.,Bottiglieri,T.,Herzenberg,L.A.,Moss,R.B.andHerzenberg,
L.A. (2006). High dose oral N-acetylcysteine, a glutathione prodrug, modulates
inflammationincysticfibrosis.Proc.Natl.Acad.Sci.103,4628-4633.
115. TizardI.R.(2013).Thedefenseofthebody.In:VeterinaryImmunology.9thed.,I.R.Tizard,ElsevierSaunders,Missouri.pp1-10.
116. Tizard I.R. (2013). Innate immunity: proinflammatory and antimicrobial mediators. In:
VeterinaryImmunology.9thed.,I.R.Tizard,ElsevierSaunders,Missouri.pp21-29.
117. TizardI.R.(2013).Regulationofadaptiveimmunity.In:VeterinaryImmunology.9thed.,I.R.Tizard,ElsevierSaunders,Missouri.pp209-224.
118. Traub-Dargatz,J.L.,Salman,M.D.andVoss,J.L.(1991).Medicalproblemsoradulthorses,
asrankedbyequinepractioners.J.Am.Vet.Med.Assoc.198,1745-1747.

48
119. Troedsson,M.H.T.(1991).Uterinedefensemechanismsinthemare.(thesis).
120. Troedsson,M.H.T.(1995).Uterineresponsetosemendepositioninthemare.Proc.Soc.Theriogenol.130-135.
121. Troedsson,M.H.T. (1997). Therapeutic considerations formating-induced endometritis.
Pferdeheilkunde.13,516-520.
122. Troedsson,M.H.T.(1999).Uterineclearanceandresistancetopersistentendometritisin
themare.Theriogenology.52,461–471.
123. Troedsson,M.H.T. (2006).Breeding-inducedendometritis inmares.Vet.Clin.Equine22,705-712.
124. Troedsson,M.H.T.,Ababneh,M.M.,Ohlgreen,A.F.,Madill,S.,Vetscher,N.andGregas,M.
(2001a).EffectofprostaglandinF2αonpregnancyratesandlutealfunctioninthemare.
Theriogenology.55,1891-1899.
125. Troedsson, M.H.T., de Morales, M.J. and Liu, I.K.M. (1993a). Correlations between
histologicendometriallesionsinmaresandclinicalresponsetointrauterineexposurewith
Streptococcuszooepidemicus.Am.J.Vet.Res.54,570-572.
126. Troedsson,M.H.T.,Desvousges,A.,Alghamdi,A.S.,Dahms,B.,Dow,C.A.,Hayna,J.,Valesco,
R., Collahan, P.T.,Macpherson,M.L., Pozor,M. and Buhi,W.C. (2005). Components in
seminalplasmaregulatingspermtransportandelimination.Anim.Reprod.Sci.89,171–186.
127. Troedsson,M.H.T.,Esteller-Vico,A.,Scoggin,K.E.,Woodward,E.M.,Squires,E.L.,Ball,B.A.
andMaxwell,H. (2014). Equine seminal plasmaderived lactoferrin regulatesbindingof
polymorphonuclearneutrophils(PMNs)tospermatozoa.J.EquineVet.Sci.34,49.
128. Troedsson,M.H.T.,Lee,C.-S.,Franklin,R.andCrabo,B.G.(2000).Theroleofseminalplasma
inpost-breedinguterineinflammation.J.Reprod.Fertil.Suppl56,341-349.
129. Troedsson,M.H.T.,Loset,K.,Alghamdi,A.M.,Dahms,B.andCrabo,B.G.(2001b)Interaction
betweenequinesemenandtheendometrium:theinflammatoryresponsetosemen.Anim.Reprod.Sci.68,273-278.

49
130. Troedsson,M.H.T.andLiu,I.K.M.(1991).Uterineclearanceofnon-antigenicmarkers(51Cr)
inresponsetoabacterialchallengeinmarespotentiallysusceptibleandresistanttochronic
uterineinfections.J.Reprod.Fertil.Suppl44,283–8.
131. Troedsson,M.H.T., Liu, I.K.M., Ing,M. and Pascoe, J. (1995). Smoothmuscle electrical
activityintheoviduct,andtheeffectofoxytocin,prostaglandinF2α,andprostaglandinE2
onthemyometriumandtheoviductofthecyclingmare.Biol.Reprod.Mono.1,475-488.
132. Troedsson,M.H.T.,Liu,I.K.M.,Ing,M.,Pascoe,J.andThurmond,M.(1993b).Multiplesite
electromyography recordings of uterine activity following an intrauterine bacterial
challengeinmaressusceptibleandresistanttochronicuterineinfection.J.Reprod.Fertil.99,307–313.
133. Troedsson,M.H.T.,Liu, I.K.M.andThurmond,M.(1993c). Immunoglobulin(IgGandIgA)
and complement (C3) concentrations in uterine secretion following an intrauterine
challenge of Streptococcus zooepidemicus in mares susceptible to versus resistant to
chronicuterineinfection.Biol.Reprod.49,502-506.
134. Troedsson,M.H.T.,Liu,I.K.M.andThurmond,M.(1993d).Functionofuterineandblood-
derived polymorphonuclear neutrophils in mares susceptible and resistant to chronic
uterineinfection:phagocytosisandchemotaxis.Biol.Reprod.49,507-514.
135. Tunon,A.-M.,Nummijarvi,A.,Katila,T.,Magnusson,U.andRodriguez-Martinez,H.(1999).
Distribution of T cells in the endometrium of themare 6 and 48 h after insemination.
Reprod.Dom.Anim.34,443-444.
136. Vanderwall,D.K.andWoods,G.L. (2003).Effecton fertilityofuterine lavageperformed
immediatelypriortoinseminationinmares.J.Am.Vet.Med.Assoc.222,1108-1110.
137. vonReitzenstein,M.,Callahan,M.A.,Hansen,P.J.andLeBlanc,M.M.(2002).Abberations
inuterinecontractilepatternsinmareswithdelayeduterineclearanceafteradministration
ofdetomidineandoxytocin.Theriogenology.58,887-898.
138. Watson, E.D. and Dixon, C.E. (1993). An immunohistological study of MHC class II
expressionandTlymphocytesintheendometriumofthemare.EquineVet.J.25,120-124.

50
139. Watson, E.D. and Thomson, S.R.M. (1996). Lymphocyte subsets in the endometriumof
genitallynormalmaresandmaressusceptibletoendometritis.EquineVet.J.28,106-110.
140. Watson,E.D.,Stokes,C.R.andBourne,F.J.(1987a).Cellularandhumoralmechanismsin
maressusceptibleandresistanttopersistentendometritis.Vet.Immunol.Immunopathol.16,107–21.
141. Watson, E.D., Stokes, C.R. and Bourne, F.J. (1987b). Influence of arachidonic acid
metabolites in vitro and in uterinewashings onmigration of equine neutrophils under
agarose.Res.Vet.Sci.43,203-207
142. Watson E.D., Stokes, C.R. and Bourne, F.J. (1988). Concentration of immunoreactive
leukotrieneB4inuterinelavagefluidfrommareswithexperimentallyinducedandnaturally
occurringendometritis.J.Vet.Pharmacol.Therap.11,130-134.
143. Watson,E.D.,Stokes,C.R.,David,J.S.E.,andBourne,F.J.(1987c).Concentrationsofuterine
luminalprostaglandinsinmareswithacuteandpersistentendometritis.EquineVet.J.19,31-37.
144. Williamson,P.,Dunning,A.,O’Connor,J.andPenhale,W.J.(1983).Immunoglobulinlevels,
proteinconcentrationsandalkalinephosphataseactivityinuterineflushingsfrommares
withendometritis.Theriogenology.19,441-448.
145. Williamson, P.,Munyua, S.Martin, R. and Penhale,W.J. (1987). Dynamics of the acute
uterine response to infection, endotoxin infusion and physical manipulation of the
reproductivetractinthemare.J.Reprod.Fert.Suppl35,317-325.
146. Williamson, P., Munyua, S.J.M. and Penhale, J. (1989). Endometritis in the mare: A
comparisonbetweenreproductivehistoryanduterinebiopsyasatechniqueforpredicting
susceptibilityofmarestouterineinfection.Theriogenology.32,351-357.
147. Witte, T.S.,Melkus, E.,Walter, I., Senge, B., Schwab, S., Aurich, C. and Heuwieser,W.
(2012). Effects of oral treatment with N-acetylcysteine on the viscosity of intrauterine
mucusandendometrialfunctioninestrousmares.Theriogenology.78,1199-1208.

51
148. Woodward,E.M.,Christoffersen,M.,Campos,J.,Betancourt,A.,Horohov,D.,Scoggin,K.E.,
Squires,E.L.andTroedsson,M.H.T.(2013a).Endometrialinflammatorymarkersoftheearly
response inmares susceptibleor resistant topersistentbreeding-inducedendometritis.
Reproduction.145,289-296.
149. Woodward,E.M.,Christoffersen,M.,Campos,J.,Horohov,D.W.,Scoggin,K.E.,Squires,E.
andTroedsson,M.H.T.(2013b).Aninvestigationofuterinenitricoxideproductioninmares
susceptibleandresistanttopersistentbreeding-inducedendometritisandtheeffectsof
immunomodulation.Reprod.Dom.Anim.48,554–561.
150. Woodward, E.M., Christoffersen, M., Campos, J., Squires, E.L. and Troedsson, M.H.T.
(2012). Susceptibility to persistent breeding-induced endometritis in the mare:
Relationship to endometrial biopsy score and age, and variations between seasons.
Theriogenology.78,495-501.
151. Woodward, E.M., Christoffersen,M., Horohow, D., Squires, E.L. and Troedsson,M.H.T.
(2015). The effect of treatment with immune modulators on endometrial cytokine
expressioninmaressusceptibletopersistentbreeding-inducedendometritis.EquineVet.J.47,235-239.
152. Woodward,E.M.andTroedsson,M.H.T.(2013)Equinebreeding-inducedendometritis.J.EquineVet.Sci.33,673–82.
153. WoodwardE.M.andTroedsson,M.H.T.(2015).Inflammatorymechanismsofendometritis.
EquineVet.J.47,384-389.
154. Zent, W.W., Troedsson, M.H.T. and Xue, J.-L. (1998). Postbreeding uterine fluid
accumulationinanormalpopulationofThoroughbredmares:Afieldstudy.Proc.AnnualConventionAAEP.44,64-65.
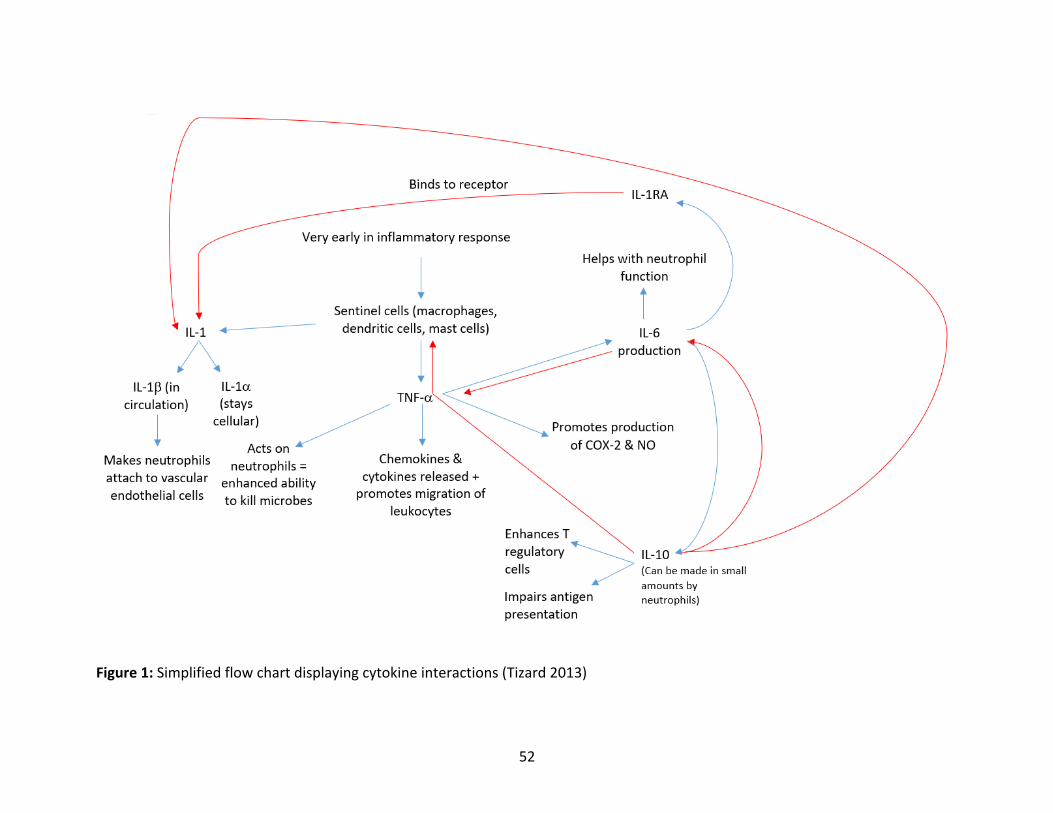
52
Figure1:Simplifiedflowchartdisplayingcytokineinteractions(Tizard2013)
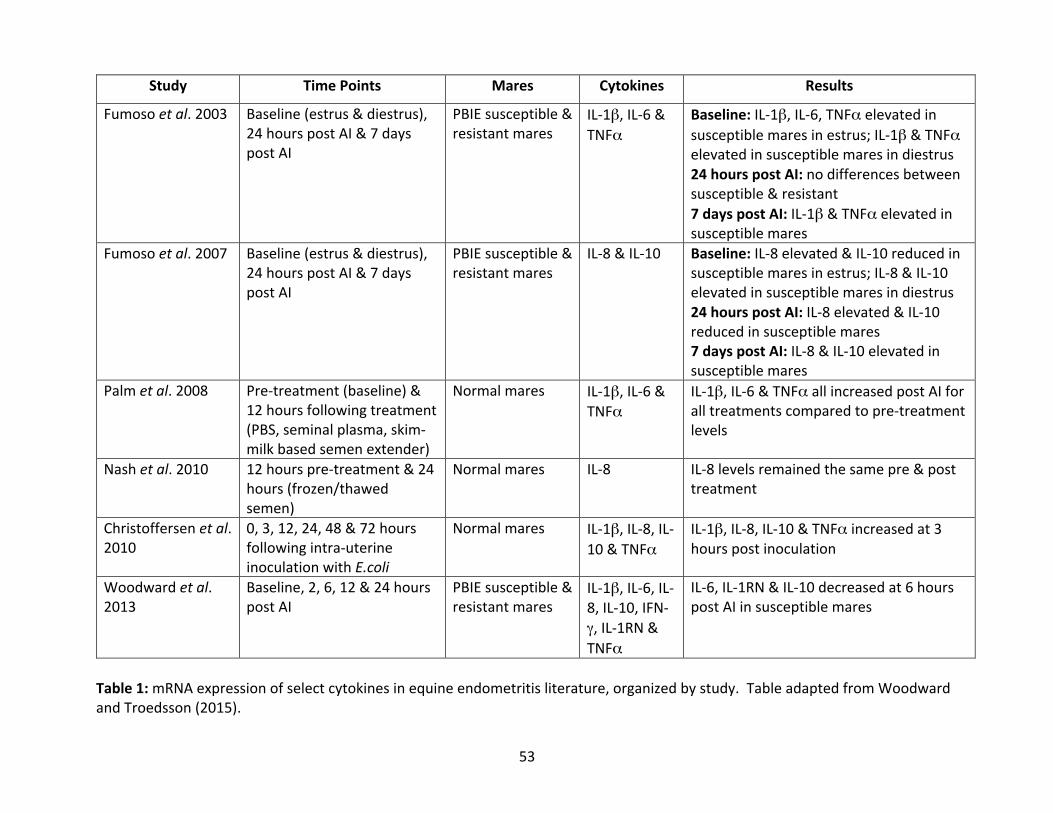
53
Study TimePoints Mares Cytokines Results
Fumosoetal.2003 Baseline(estrus&diestrus),24hourspostAI&7dayspostAI
PBIEsusceptible&resistantmares
IL-1β,IL-6&TNFα
Baseline:IL-1β,IL-6,TNFαelevatedinsusceptiblemaresinestrus;IL-1β&TNFαelevatedinsusceptiblemaresindiestrus24hourspostAI:nodifferencesbetweensusceptible&resistant7dayspostAI:IL-1β&TNFαelevatedinsusceptiblemares
Fumosoetal.2007 Baseline(estrus&diestrus),24hourspostAI&7dayspostAI
PBIEsusceptible&resistantmares
IL-8&IL-10 Baseline:IL-8elevated&IL-10reducedinsusceptiblemaresinestrus;IL-8&IL-10elevatedinsusceptiblemaresindiestrus24hourspostAI:IL-8elevated&IL-10reducedinsusceptiblemares7dayspostAI:IL-8&IL-10elevatedinsusceptiblemares
Palmetal.2008 Pre-treatment(baseline)&12hoursfollowingtreatment(PBS,seminalplasma,skim-milkbasedsemenextender)
Normalmares IL-1β,IL-6&TNFα
IL-1β,IL-6&TNFαallincreasedpostAIforalltreatmentscomparedtopre-treatmentlevels
Nashetal.2010 12hourspre-treatment&24hours(frozen/thawedsemen)
Normalmares IL-8 IL-8levelsremainedthesamepre&posttreatment
Christoffersenetal.2010
0,3,12,24,48&72hoursfollowingintra-uterineinoculationwithE.coli
Normalmares IL-1β,IL-8,IL-10&TNFα
IL-1β,IL-8,IL-10&TNFαincreasedat3hourspostinoculation
Woodwardetal.2013
Baseline,2,6,12&24hourspostAI
PBIEsusceptible&resistantmares
IL-1β,IL-6,IL-8,IL-10,IFN-γ,IL-1RN&TNFα
IL-6,IL-1RN&IL-10decreasedat6hourspostAIinsusceptiblemares
Table1:mRNAexpressionofselectcytokinesinequineendometritisliterature,organizedbystudy.TableadaptedfromWoodwardandTroedsson(2015).
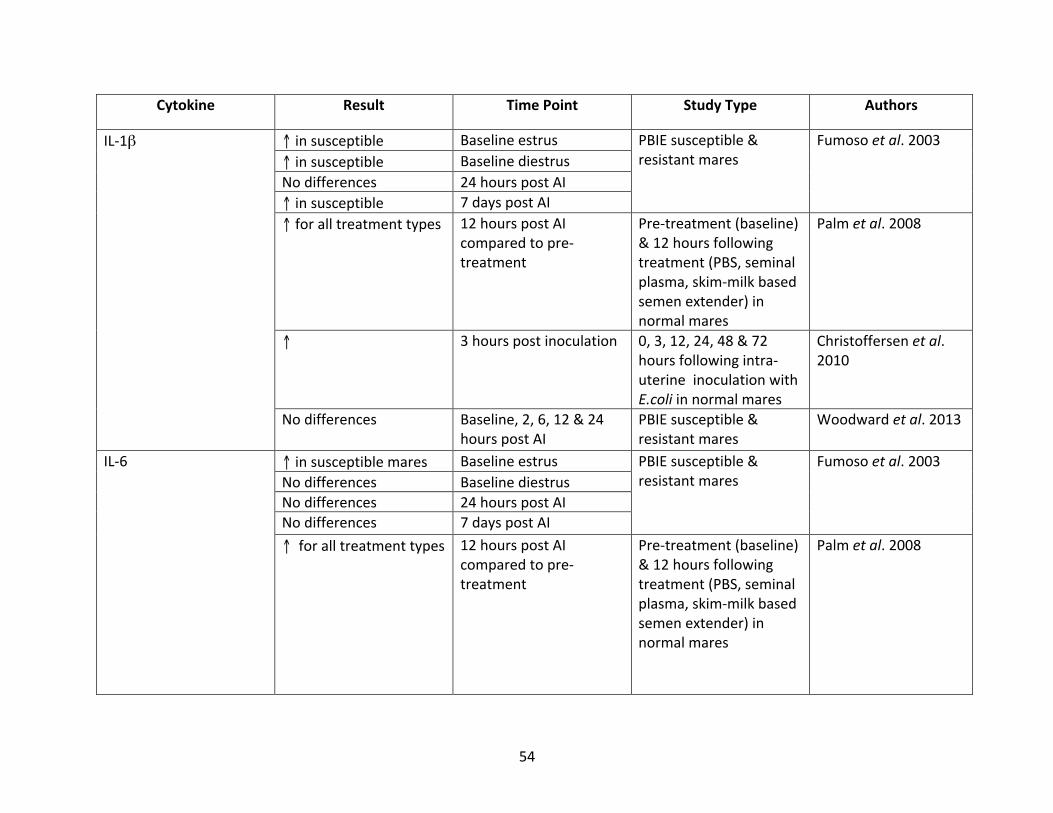
54
Cytokine Result TimePoint StudyType Authors
IL-1β ↑insusceptible Baselineestrus PBIEsusceptible&resistantmares
Fumosoetal.2003↑insusceptible BaselinediestrusNodifferences 24hourspostAI↑insusceptible 7dayspostAI↑foralltreatmenttypes 12hourspostAI
comparedtopre-treatment
Pre-treatment(baseline)&12hoursfollowingtreatment(PBS,seminalplasma,skim-milkbasedsemenextender)innormalmares
Palmetal.2008
↑ 3hourspostinoculation 0,3,12,24,48&72hoursfollowingintra-uterineinoculationwithE.coliinnormalmares
Christoffersenetal.2010
Nodifferences Baseline,2,6,12&24hourspostAI
PBIEsusceptible&resistantmares
Woodwardetal.2013
IL-6 ↑insusceptiblemares Baselineestrus PBIEsusceptible&resistantmares
Fumosoetal.2003Nodifferences BaselinediestrusNodifferences 24hourspostAINodifferences 7dayspostAI↑foralltreatmenttypes 12hourspostAI
comparedtopre-treatment
Pre-treatment(baseline)&12hoursfollowingtreatment(PBS,seminalplasma,skim-milkbasedsemenextender)innormalmares
Palmetal.2008
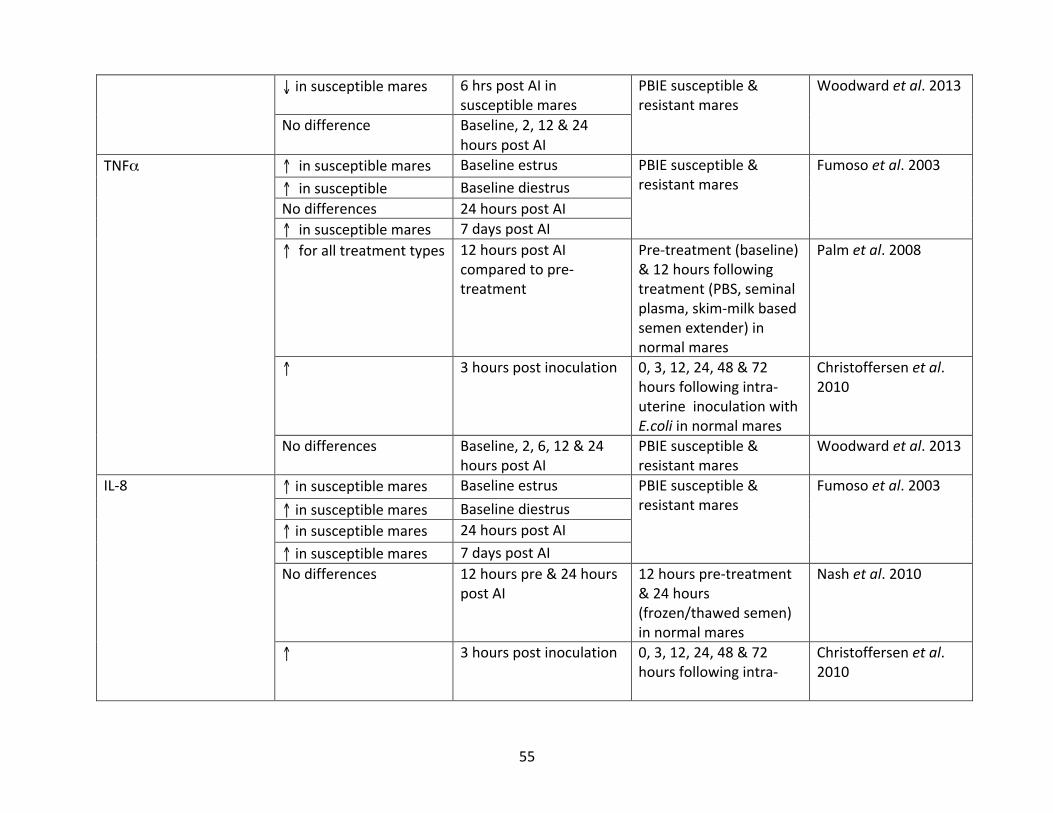
55
↓insusceptiblemares 6hrspostAIinsusceptiblemares
PBIEsusceptible&resistantmares
Woodwardetal.2013
Nodifference Baseline,2,12&24hourspostAI
TNFα ↑insusceptiblemares Baselineestrus PBIEsusceptible&resistantmares
Fumosoetal.2003↑insusceptible BaselinediestrusNodifferences 24hourspostAI↑insusceptiblemares 7dayspostAI↑foralltreatmenttypes 12hourspostAI
comparedtopre-treatment
Pre-treatment(baseline)&12hoursfollowingtreatment(PBS,seminalplasma,skim-milkbasedsemenextender)innormalmares
Palmetal.2008
↑ 3hourspostinoculation 0,3,12,24,48&72hoursfollowingintra-uterineinoculationwithE.coliinnormalmares
Christoffersenetal.2010
Nodifferences Baseline,2,6,12&24hourspostAI
PBIEsusceptible&resistantmares
Woodwardetal.2013
IL-8 ↑insusceptiblemares Baselineestrus PBIEsusceptible&resistantmares
Fumosoetal.2003↑insusceptiblemares Baselinediestrus↑insusceptiblemares 24hourspostAI↑insusceptiblemares 7dayspostAINodifferences 12hourspre&24hours
postAI12hourspre-treatment&24hours(frozen/thawedsemen)innormalmares
Nashetal.2010
↑ 3hourspostinoculation 0,3,12,24,48&72hoursfollowingintra-
Christoffersenetal.2010
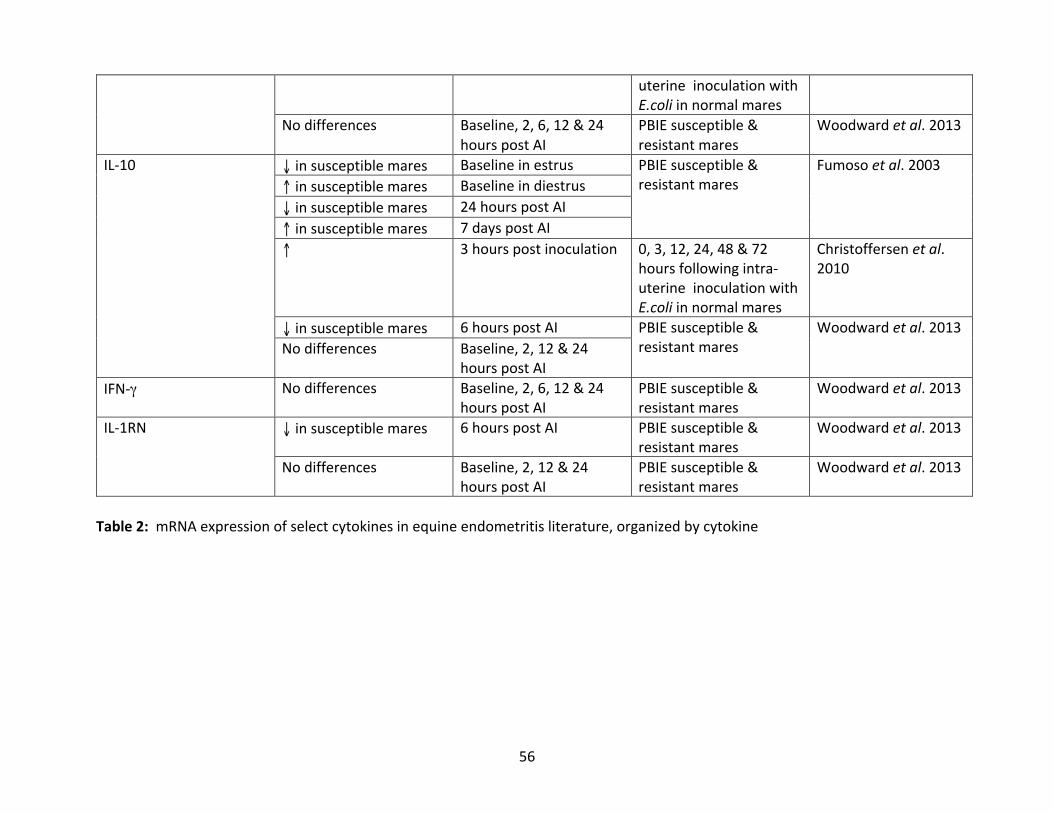
56
uterineinoculationwithE.coliinnormalmares
Nodifferences Baseline,2,6,12&24hourspostAI
PBIEsusceptible&resistantmares
Woodwardetal.2013
IL-10 ↓insusceptiblemares Baselineinestrus PBIEsusceptible&resistantmares
Fumosoetal.2003↑insusceptiblemares Baselineindiestrus↓insusceptiblemares 24hourspostAI↑insusceptiblemares 7dayspostAI↑ 3hourspostinoculation 0,3,12,24,48&72
hoursfollowingintra-uterineinoculationwithE.coliinnormalmares
Christoffersenetal.2010
↓insusceptiblemares 6hourspostAI PBIEsusceptible&resistantmares
Woodwardetal.2013Nodifferences Baseline,2,12&24
hourspostAIIFN-γ Nodifferences Baseline,2,6,12&24
hourspostAIPBIEsusceptible&resistantmares
Woodwardetal.2013
IL-1RN ↓insusceptiblemares 6hourspostAI PBIEsusceptible&resistantmares
Woodwardetal.2013
Nodifferences Baseline,2,12&24hourspostAI
PBIEsusceptible&resistantmares
Woodwardetal.2013
Table2:mRNAexpressionofselectcytokinesinequineendometritisliterature,organizedbycytokine

57
CHAPTERTWO:THEEFFECTSOFTREATMENTWITHN-ACETYLCYSTEINE
ONCLINICALSIGNSINPBIESUSCEPTIBLEMARES
2.1ABSTRACT
Persistentbreedinginducedendometritis(PBIE)isamajorcauseofinfertilityinmares.
Uterine inflammationat the timeof embryonicdescentultimately results inearlyembryonic
death.Asevereendometrialbiopsygrade(IIborIII)hasbeenidentifiedasariskfactorforPBIE.
Intra-uterinefluidaccumulation(>2cmindepth),pathologicendometrialedemaandelevated
intra-uterine neutrophil levels are all clinical features of PBIE. N-acetyl cysteine (NAC), a
mucolyticusedtotreatbacterialendometritisinmares,hasanti-inflammatoryproperties,and
was investigated to determine if intra-uterine infusion before breeding would result in the
improvement of clinical signs associated with PBIE. In addition, the effects of NAC at the
histological level on the endometrium were determined. A randomized, blinded, cross-over
designclinicaltrialwasperformedutilizingPBIEsusceptiblemares(n=9).Intrauterineinfusionof
3.3% NAC was performed 12 hours prior to insemination, and endometrial cytology and
endometrialbiopsysampleswereobtainedat12and60hourspost-insemination.Endometrial
biopsieswerescoredbasedonthedegreeofinflammation(neutrophils)present.Clinicalsignsof
endometrialedemaandintra-uterinefluidvolumeswereassessedbytrans-rectalultrasoundat
12andthenevery24hourspostbreeding.StatisticalanalysisutilizedrepeatedmeasuresANOVA
andaMannWhitneyWilcoxonTest.TreatmentwithNACdidnotimproveclinicalsignsinPBIE-
affected mares. However, treatment with NAC had a significant effect on the degree of
endometrialinflammationnotedhistologically,withtreatedcyclesdisplayingmorediffuseand

58
severeneutrophilinfiltrationcomparedtocontrolcycles.TheeffectsofNAContheendometrium
of PBIE-susceptible mares seen in this study is concerning. Further research is required to
evaluatethesafetyofNACinthetreatmentofPBIE-susceptiblemares.
2.2INTRODUCTION
Uterineinflammationisanormalresponsetobreedingandisbelievedtocleartheuterus
ofdebris,spermatozoaandinflammatoryproductspriortoembryodescent.Insomemares,the
inflammatoryresponseisprolonged,leadingtoearlyembryonicdeathandinfertility.Affected
maresareconsideredsusceptibletopersistentbreeding-inducedendometritis(PBIE)(Troedsson
2006;WoodwardandTroedsson2015;ChristoffersenandTroedsson2017).
ConsiderableresearchhasfocusedonthereasonssomemaresaresusceptibletoPBIE,
withthemajorityofstudiesfocusingonthe immunesystemandmechanicalclearanceofthe
uterus(NikolakopoulosandWatson,1999;Watson2000;LeBlancandCausey2009;Troedsson
2006;WoodwardandTroedsson2015;ChristoffersenandTroedsson2017).Multipleaspectsof
immune dysfunction have been investigated as potential contributors to PBIE. An altered
humoralimmuneresponse,specificallydifferinglevelsofimmunoglobulinsbetweensusceptible
andresistantmares,hasbeenidentified(Asburyetal.1980;Mitchelletal.1982;Williamsonet
al.1983;Troedsson1991).Incontrast,Troedssonetal(1993b)foundthatPBIE-susceptiblemares
didnothaveimmunoglobulindeficiencyat24hoursfollowingAI,disprovingthepreviousbelief
that immunoglobulin alterationswere the cause of PBIE. Troedsson et al. (1993c) and other
researchersdemonstratedthatPBIE-susceptiblemareshadelevatednumbersof intra-uterine
neutrophilswithreducedfunctionalityandphagocyticproperties(Luietal.1986;Watsonetal.

59
1987;Watsonetal.1988;NikolakopoulosandWatson1997;Troedssonetal.1993c;Woodward
andTroedsson2013).Additionalstudieshaverecentlyfurtheredourunderstandingoftherole
of inflammation inPBIE. Differences inmRNAexpressionofbothpro-andanti-inflammatory
cytokineshavebeenidentifiedeitheratbaselinelevels,oratmultipletimepointsfollowingAI,
betweennormalandaffectedmares.Persistentexpressionofpro-inflammatorycytokines for
prolonged periods of time, as well as reduced expression of anti-inflammatory cytokines
followingbreeding,maycontributetothemanifestationofPBIE(Fumosoetal.2003;Fumosoet
al. 2007; Christoffersenet al. 2012;Woodwardet al. 2013a; Christoffersen and Troedsson,
2017).PBIE-susceptiblemaresarealsoknowntohaveanunderlyingmyometrialdefect,causing
decreaseduterine contractility (Troedssonetal. 1993a; LeBlancetal. 1994a;Neuwirthetal.
1994;Rigbyetal.2001;vonReitzenseinetal.2001;LeBlancandCausey2009).Inaddition,intra-
uterinenitricoxidelevelsareelevatedinPBIE-susceptiblemares,furthercontributingtoreduced
uterine contractions (Alghamdi et al. 2005; Woodward et al. 2013b; Khan et al. 2018). A
reductioninuterinecontractilityleadstodecreaseduterineclearance,afeatureofPBIE.From
these studies, it is clear that themechanisms involved in uterine health following breeding,
includingtheimmuneresponse,persistentinflammationanduterineclearance,arecomplexand
interdependent.DysregulationofanyofthesemechanismshavethepotentialtoleadtoPBIEin
mares.
ClinicalsignsofPBIEincludeabnormalendometrialedema(Samper2007),persistenceof
a>2cmdepthofintra-uterinefluidbeyond48hourspost-AIdetectedontrans-rectalultrasound
(Brinsko 2003; Pycock 2007), presence of inflammatory cells on endometrial cytology and

60
endometrial histopathology consistent with uterine inflammation (LeBlanc 2010; Troedsson
2011;Canissoetal.2016).
Treatmentmodalities for PBIE do exist, yet these are notwithout limitations. Ecbolic
therapies, including oxytocin and prostaglandin, increase uterine clearance by increasing
myometrialactivity,andcanbehighlyeffectiveat improvinguterineclearance(LeBlancetal.
1994b, Combs et al. 1996; Troedsson 1997; Cadario et al. 1999; LeBlanc and Causey 2009).
However, some mares are refractory to oxytocin and continue to accumulate fluid despite
therapy(Hurtgen2006;LeBlancandCausey2009).Additionally,prostaglandinsadministeredas
ecbolic therapy post-ovulationmay interfere with progesterone production from the corpus
luteum, potentially resulting in early embryonic death. Prostaglandins are therefore only
recommendedinthepre-ovulatoryperiod,limitingtheiruseintreatmentofPBIE(Troedssonet
al. 2001; Nie et al. 2002; Brendemuehl 2002; Nie et al. 2003). Immunomodulators, such as
glucocorticoids,areuseful inmareswithsterile inflammationbutshouldbeavoided inmares
with concurrent bacterial involvement or othermedical conditions such as laminitis (LeBlanc
2010).Therefore,alternativetreatmentmodalitiesforPBIEareneeded.
N-acetylcysteine(NAC)isamucolyticagentthatdisruptsdisulphidebondslinkingmucin
polymers(LeBlanc2010)andhasanti-inflammatoryandanti-oxidantproperties(Bergaminietal.
2001;Majanoetal.2004;Cazzolaetal.2017).N-acetylcysteineinhibitedtheenzymeresponsible
forNOsynthesisandblockedpro-inflammatorycytokineproductioninamultiplesclerosis(MS)
model using rats, and enhanced anti-oxidative factors in in vitro human chronic obstructive
pulmonarydisease(COPD)research(Stanislausetal.2005).N-acetylcysteineiscurrentlyused
as an intra-uterine infusion inmareswith excessive uterinemucus production or in cases of

61
bacterial endometritis complicated by a biofilm (LeBlanc and Causey, 2009; LeBlanc 2010;
Troedsson2012).InfusionofNACimprovedfertilityratesofbarrenmareswhenusedduringthe
priorcycleor48hourspre-breeding(Gores-Lindholmetal.2009).Oral(Witteetal.2012)and
intra-uterine(Melkusetal.2013)NACadministrationdecreasedneutrophilnumberandCOX-2
staininginuterinehistologicalsamplesinnormalmares.N-acetylcysteineappearstobesafefor
use in horses: it was reported to cause no adverse effects on normal mare endometrium
following intra-uterine infusion of either a 3.3% or 5% solution (Gores-Lindholmet al. 2013;
Melkusetal.2013)orfollowingoraladministration(10mg/kg)(Witteetal.2012).Nostudiesto
datehaveexaminedtheeffectsofNAContheresolutionofclinicalsignsofPBIEinsusceptible
mares. Based on its anti-inflammatory and anti-oxidant effects, we investigated NAC as a
potentialtreatmentforPBIEinsusceptiblemares.
Theobjectiveofthisstudywastodeterminetheefficacyofintra-uterineinfusionofNAC
asatreatmentforPBIEinsusceptiblemares.Wehypothesizedthatintra-uterinetreatmentof
PBIE-susceptiblemareswithNACwouldresultinmorerapidresolutionofexcessiveendometrial
edema, percent neutrophils on endometrial cytology, intra-uterine fluid volume and
inflammationasassessedbyendometrialbiopsy,comparedtountreatedcycles.
2.3MATERIALSANDMETHODS
AlltheproceduresperformedonanimalsinthisstudywereapprovedbytheUniversity
ofGuelphAnimalCareCommittee(AnimalUtilizationProtocolnumber3213),inaccordance
withtheCanadianCouncilonAnimalCareguidelines.

62
2.3.1SelectionofPBIESusceptibleMares
MarespreviouslyidentifiedassusceptibletoPBIE(Khanetal.2018)wereutilizedinthis
study.MareswereselectedinamannersimilartothatofAlghamdietal.(2005),andWoodward
etal.(2013a,2013b;2015).PotentiallyPBIE-susceptiblemareswereidentifiedonthebasisofa
KenneybiopsyscoreofIIborIII,negativebacterialcultureand<2neutrophilsperhighpower
field(hpf)onendometrialcytology.StudymareswerethenconfirmedtobePBIE-susceptibleby
failuretoclearintra-uterinefluidfollowingintra-uterinechallengewithkilledspermatozoa(i.e.
persistence of greater than 2cm of intrauterine fluid 96 hours following insemination).
Immediatelypriortothecommencementoftheclinicaltrial,studymareswereagainscreened
toensuretheywerenegativeforbacterialendometritis,andpre-trialendometrialbiopsieswere
obtained.BacterialculturesweresubmittedtotheAnimalHealthLaboratory(AHL,Guelph,ON,
Canada).Onlymareswithoutbacterialendometritiswereenrolledinthetrial.
2.3.2ClinicalTrial
Ablinded,randomized,crossoverdesignedclinicaltrialwasperformedusingtenPBIE-
susceptiblemares(Figure1).MareswerehousedattheArkellEquineResearchStationatthe
UniversityofGuelphandwereofvariousbreeds(Standardbredandmixedbreed)andages(7-19
years). The order of Treatment (N-acetyl cysteine intra-uterine infusion) and Control (saline)
cycles for each mare were assigned via random number allocation, with a washout period
between experimental cycles that consisted of at least one estrous cycle. The primary
investigatorconductingclinicalexaminationsandallpersonnelinvolvedinthedailycareofthe
mareswereblindedtotreatmentandcontrolassignmentofthemaresduringdatacollection.
OnlycycleswhichfitthedefinitionofPBIE,(>2cmofintra-uterinefluidforatleast48hourspost

63
AI)wereincludedintheanalysis. Forthisreason,onemarewasremovedfromthestatistical
analysisforbothtreatmentandcontrolcycles.
Maresweremonitoredbytrans-rectalpalpationandultrasoundandestruswasinduced
with an intra-muscular dose (125mcg) of cloprostenol (Estrumate®, Intervet Canada Corp.,
KirklandQC,CAN)asneeded.Onceinestruswitha35mmorlargerfollicleidentified,2,500IUof
humanchorionicgonadotropin(hCG,Chorulon®,IntervetCanadaCorp.,Kirkland,QC,Canada)
wasadministeredintravenouslytoinduceovulation.Duringtreatmentcycles(TX),180mlofa
3.3%NACsolutionconsistingof30mlofN-acetylcysteine(AcetylcysteineInjection20%solution,
AlvedaPharmaceuticals Inc., Toronto,ON,Canada)diluted in150mlof0.9% sodiumchloride
(0.9%SodiumChlorideInjectionUSP,BaxterCorporation,Mississauga,ON,Canada)wasinfused
into the uterus using a 21-inch insemination pipette (Partnar Animal Health, Ilderton, ON,
Canada). During control cycles (CONTROL), 180ml of sterile saline (0.9% Sodium Chloride
InjectionUSP)was infused into theuterus. Uterine infusions forboth treatmentandcontrol
cycleswereperformedatthetimeasthehCGinjection.Twelvehoursfollowinginfusion,mares
wereartificiallyinseminatedwith1x109killedspermatozoain30mlofsemenextenderprepared
aspreviouslydescribed(Woodwardetal.2013a;Woodwardetal.2013b;Woodwardetal.2015;
Khanetal.2018).Maresweremonitoredviatrans-rectalultrasoundbeginning12hourspost-AI,
andthiswasrepeatedat24hourintervalsthroughtoandincluding84hourspost-AI(12,36,60
and84hourspost-AI),todetermineendometrialedemascore(0=none;1=minimal;2=moderate;
3=normalmaximaland4=excessiveorpathological;DieldeAmorimetal.2016)andintra-uterine
fluid volume (IUF). The IUF volume was calculated using the formula IUF
(ml)=length*width*height,eachmeasured incentimeters,wherethe IUFwasthemeanvalue

64
derivedfrom2-5calculationsoffluidasassessedintheuterinebodyaswellasfromwithinthe
leftandrightuterinehorns.
Endometrialcytology,endometrialfluid(thissamplewasusedforadifferentexperiment)
and endometrial biopsy samples were taken, in that order, at 12 and 60 hours post-AI.
Endometrial cytology samples were aseptically obtained using a guarded swab (Kalayjian
Industries Inc., SignalHill, CA,USA) (Khanetal. 2018).Briefly, theguarded swabwaspassed
throughthecervix,theswabwasadvancedbeyondthesheathandthenrotatedseveraltimes
beforebeingleftinplaceforaminimumof30seconds,andthentheswabwasretractedintothe
sheath.Theoutersheathwasthenrotatedtocollectendometrialcellsinthecap,andtheentire
unitwas then removed. Endometrial biopsieswere taken aseptically at thebaseof the right
uterinehorn,bytrans-rectalguidanceofaPilling-Weckbiopsypunch(JorgensenLaboratories,
Loveland, CO, USA), through the cervix, as previously described (Kenney 1978). Endometrial
biopsy samples were placed in 10% formalin solution (Surgipath® 10% Neutral Buffered
Formalin, Leica Biosystems Richmond, Inc., Richmond, IL, USA) and submitted to the Animal
HeathLaboratory(UniversityofGuelph)forembeddinginparaffinwax,sectioningandroutine
stainingwithHematoxylinandEosin(H&E).
2.3.3InflammationasAssessedbyEndometrialBiopsies
Endometrial biopsies were examined for the degree and distribution of neutrophils,
lymphocytesandplasmacells(‘inflammatorycells’)byaBoardCertifiedTheriogenologist(TC),
blindedtomaretreatmentassignments.Asemi-quantitative inflammationscorebetween1-4
(1=mild; 2=moderate; 3=marked and superficial; 4= severe and diffuse) was assigned to
endometrialbiopsies,andwasbasedonthecompilationofthreeseparateobservationsmade

65
acrossaminimumof10highpowerfields(Figure2;Table1).Thefirstobservationconsistedof
thenumberofneutrophilsperhighpowerfieldasadaptedfromAlvarengaetal.(2016)(1=0-3
neutrophils/hpf);2=(4-10neutrophils/hpf);3=10-30neutrophils/hpf);4=>30neutrophils/hpf).
Thesecondobservationconsistedofthedepthofinflammatorycellinfiltration,withscoresof1-
3 having inflammatory cells present in themost superficial areas only (stratum compactum)
whereasthosewithdiffuseinfiltrationofinflammatorycellsdeepintotheendometrium(stratum
spongiosum) were given a score of 4. The third observation consisted of the presence of
endometrial luminalepitheliumerosion,asdefinedbylossofepitheliumwithevidenceofcell
proliferationonboththeedgesandbelowtheareaoferosion(cellsbecomingthinandstretched
ontheedges;presenceofhyperchromaticcellsbelow).Thepresenceoferosioncontributedto
amoreseverescore(3or4).
2.3.4EndometrialCytology(%Neutrophils)
Endometrial cytology slideswere prepared and stainedwithmodifiedWright Giemsa
stain(PROTOCOLTM,Hema3TMStatPack,FisherBrand®,Pittsburg,PA),aspreviouslydescribed
(Khanetal.2018).Thepercentageofneutrophilstototalcellswasdetermined(endometrialcells
and neutrophils combined) across multiple fields of view, at 400x. This accounted for any
variability of cell distribution. The mean percentage of neutrophils was calculated after a
minimumof500totalcellswerecounted.
2.3.5StatisticalAnalyses
StatisticalanalyseswereperformedusingtheSASsoftwareprogram(SASInstitute,Inc.,
Version9.3,Cary,NC,USA).Differences in IUF,%neutrophilsanduterineedemascoreswere
comparedbetweenNAC-treated(TX)andsaline(CONTROL)cycles,aswellasacrosstimepoints

66
using repeatedmeasures ANOVA.Mare identification (ID) was included as a variable in the
statistical model, but was removed, as no significance was detected. Differences in biopsy
inflammationscorebetweenNAC-treated(TX)andsaline(CONTROL)aswellasacrosstimepoints
wereanalyzedusingtheMannWhitneyWilcoxontest.TheAkaikeinformationcriterion(AIC)was
utilizedtodeterminethebestfitmodel.NormalityofthedatawastestedusingtheShapiro-Wilk
test. Data for estimated intrauterine fluid volume (IUF) required logarithmic transformation,
however all other outcome parameters were normally distributed. Results are presented as
medians+/-lowerlimit(LL)andupperlimit(UL)whenlogtransformationwasrequiredandas
means+/-standarderror(SE)whennodatatransformationswereperformed.Wheretherewas
noeffectoftreatment,dataforNAC-treated(TX)andsaline(CONTROL)cycleswerecombinedto
thenexaminetheeffectofnumberofhourspost-AI.Similarly,wheretherewasnoeffectofhours
post AI, data for 12 and 60 hours post-AI were combined to then examine the effects of
treatment.Correlationbetween%neutrophilsandbiopsyinflammationscorewasanalyzedusing
the Spearman correlation coefficient, to account for the significance in the model and
distributionofthedata.CorrelationbetweenIUFanduterineedemascorewasassessedusing
the Pearson correlation coefficient, since the model had no effects and data was normally
distributed.Thepvaluewassetat<0.05forstatisticalsignificanceforallstatisticalanalyses.
2.4RESULTS
2.4.1BiopsyInflammationScore
Pre-trialbiopsieswereavailablefor7ofthe9PBIE-susceptiblemaresusedinthisstudy.
Themeanpre-trial(baseline)biopsyinflammationscorewas1.14+/-0.65.Biopsyinflammation

67
scorewassignificantlygreaterintheNACtreatedcycle(TXmean=3.0+/-0.5)comparedtothe
salinetreatedcycle(CONTROLmean=2.2+/-0.5),whenbiopsyinflammationscoresat12and
60hourspost-AIwerecombinedforeachtreatmentcycle(p=0.01)(Figure3).Theendometrial
effectsofNACincludederosionsoftheluminalepitheliumandmarkedinfiltrationofneutrophils
inthestratumcompactum(Figures2d,4and5).Oneofthedifferences incriteriabetweena
biopsyinflammationscoreof1-3comparedtoabiopsyinflammationscoreof4wasthepresence
ofdeep-seatedneutrophil infiltration.The incidenceofan inflammationscoreof4wasmuch
greaterinNACtreatedcycleswithtimepointscombined(8/18)incomparisontosalinetreated
cycles(1/18)withtimepointscombined(Table1).Therewasnosignificanttimeeffectonbiopsy
inflammationscorefrom12to60hourspost-AIwhenbothtreatmentcycleswerecombined;
mean(12hourspostAI)was2.8+/-0.5whereasmean(60hourspost-AI)was2.3+/-0.5(p=0.12)
(Figure6).
2.4.2EstimatedIntra-UterineFluidVolumes(IUF)
TreatmentwithNACdidnotaffectcalculatedIUFwhentimepointswerecombinedwithin
eachtreatmentcycle(p=0.78;TXmedian=3.9cm3;range=1.1-14.7cm3;CONTROLmedian=4.5
cm3;range=1.2-16.8cm3)(Figure7).Incontrast,theeffectoftimewassignificant(p=0.005),
withmoreintra-uterinefluidpresentat36hourspost-AI(median=9.8cm3;range=2.6-36.2cm3)
comparedto12hourspost-AI(median=3.4cm3;range=1-11.6cm3),60hourspost-AI(median
= 3.9cm3; range = 0.9-17.5cm3) and 84 hours post-AI (median = 2.5cm3; range = 0.6-9.5cm3)
(Figure8).

68
2.4.3EndometrialEdema
TreatmentwithNACdidnotaffectendometrialedemascores(p=0.90;TXmean=1.7+/-
0.6;CONTROLmean=1.7+/-0.6)(Figure9).Endometrialedemascorewassignificantlyhigher
(p=0.04)at12hours(mean=2.1+/-0.6)and36hours(mean=2.0+/-0.6)comparedto60hours
post-AI(mean=1.3+/-0.6)and84hours(mean=1.3+/-0.6)post-AI(Figure10).Theestimated
IUFanduterineedemascoreswerenotsignificantlycorrelated(r=-0.14;p=0.45).
2.4.4EndometrialCytology(%Neutrophils)
TreatmentwithNACdidnotaffect%neutrophilsonendometrialcytology (p=0.39;TX
mean = 48.4% +/- 12.5%; CONTROL mean = 40.9% +/- 12.5%) (Figure 11). Evidence of
inflammationdecreasedovertimepost-AI(p=0.005).The%neutrophilswassignificantlyhigher
at12hourspost-AI(mean=57.7%+/-12.5%)comparedto60hourspost-AI(mean=31.6%+/-
12.5%) (Figure12).Additionally,%neutrophilsandbiopsy inflammationscorewerepositively
correlated(r=0.43;p=0.006).
2.4.5InteractionsBetweenTreatmentCyclesandHoursPost-AI(Appendix1)
There was no significant interaction between treatment cycle and hours-post-AI for
biopsy inflammation score (p=0.12;Appendix1: figure1), IUF (p=0.84;Appendix1: figure2),
endometrialedema(p=0.65;Appendix1:figure3),nor%neutrophils(p=0.98;Appendix1:figure
4).
2.5DISCUSSION
Intra-uterineinfusionofPBIE-susceptiblemareswitha3.3%NACsolution12hourspre-
AIresultedinmoreinflammationonendometrialbiopsycomparedtotreatmentwithsaline12

69
hourspre-AI.ThisisincontrasttopreviousstudieswheretreatmentwithNACdidnotappearto
bedetrimental to theendometrium,andtheendometrial luminalepitheliumremained intact
(Witteetal.2012;Gores-Lindholmetal.2013;Melkusetal.2013).Melkusetal.(2013)reported
thatintra-uterineinfusionof5%NACinnormalmaresresultedinlessendometrialinflammation
onbiopsyat72hourspost-treatmentcomparedtocontrolmarestreatedwithsaline.However,
NACtreatedandcontrolnormalmaresbothhadhighernumbersofneutrophils/hpf24hours
followingtreatment(10.9neutrophils/hpfinNACgroup;8.3neutrophils/hpfinsalinegroup).This
resolved by 72 hours, with treatedmares having significantly less neutrophils than controls.
Normal mares treated orally with NAC for four days had fewer neutrophils present in the
endometriumonedayfollowingtreatment,comparedtountreatedcontrols(Witteetal.2012).
ItisinterestingthatintheWitteetal.(2012)andMelkusetal.(2013)studies,normalmaresin
estrus hadmore neutrophilswithout provocation of inflammation, such asAI. In the studies
mentionedabove,maresweresimplytreatedinestrusandbiopsieswereobtained.Untreated
maresorthosetreatedwithsalinehadgreaterthan2neutrophils/hpf,whichisconsideredtobe
indicativeofendometritis(Woodwardetal.2013a)(4.8neutrophils/hpfwithnotreatment;3.9
neutrophils/hpfwithsaline infused intra-uterineat72hoursposttreatment).Thereasonsfor
thisincreaseinsupposedlynormalmareswithoutaninflammatorychallengearenotknown,but
could include differences in the original classification as normalmares, posing as a possible
limitationtotheirstudy,ortotheirritatingnatureoftheinfusionofNACitself.Gores-Lindholm
etal. (2013)reportedthatthedegreeofacute inflammation inendometrialbiopsieswasnot
differentthreedaysfollowingintra-uterineinfusionofa3.3%NACsolutionascomparedtopre-
treatment within a mixed group of 20 mares (10 barren treated; 6 fertile treated; 6 fertile

70
control).However,maresinthatstudyweretreatedwithuterinelavagewithlactatedRinger’s
solution(LRS)dailyfortwodaysbetweentheinitialinfusionandtheprocurementofthebiopsy,
possiblyaidingresolutionofanyinflammationresultingfromtheNACinfusionbythetimethe
biopsy was taken. In contrast to the above studies, which utilized either normal or mixed
populationsofmares,ourtrialinvolvedonlyPBIE-affectedmares,selectedpartiallyonthebasis
ofpoorendometrialbiopsyscores(GradeIIbandIII).Normalmaresarelikelymorecapableof
clearing any inflammatory reaction caused by infusion of NAC or other irritating substances.
However, this inflammatory reactionmaynotbeclearedas rapidly inmares thatarealready
susceptibletouterineinflammation,suchasthoseusedinourstudy.
Inflammatoryscoresof4werealmostexclusivelyassignedtomaresintheNACtreated
cycles (Table 1). The inflammation noted in these biopsies consisted almost entirely of
neutrophils and was diffusely deep-seated, extending into the stratum spongiosum and
occasionallyintotheglandularlumen(Figures2dand5).Additionally,severelyaffectedmares,
withascoreof3or4,oftendisplayedsignsofareactive luminalepitheliumindicativeofcell
injury. The biopsies in our study also had some areas of erosion or complete loss of the
endometrialluminalepithelium(Figure4)andthesechangesweretakenintoconsiderationfor
thebiopsyinflammationscore(scoreof3or4).Thesechanges,aswellasthosementionedabove,
areallsuggestiveofamarkedendometrialresponsetotheirritatingnatureofNAC.Witteetal.
(2012) reported that treatment with NAC caused increased epithelial cell proliferation and
suggestedittobeevidenceofendometrialregeneration.Gores-Lindholmetal.(2013)mentioned
anincreaseinepithelialcellheightinsalinetreatedfertilemarescomparedtoNACtreatedfertile
mares. The changes in these studies could suggest endometrial reactivity; however, the

71
epitheliumremainedintact.Thebiopsieswithareasoferosionwereverysimilarinappearance
tothoseseeninmarestreatedwithintra-uterinekerosene,whichwaspreviouslyreportedto
causesevereinflammation(R.Foster,Personalcommunication;Bracheretal.1991).Mareswith
GradeIIIbiopsyscoresthatreceivedanintra-uterineinfusionof50mlofcommerciallyavailable
kerosene demonstrated a more prolonged and severe inflammatory response histologically,
compared to similarly treated mares with Grade I or II biopsy scores (Bracher et al. 1991).
However,inthatstudy,mononuclearcellswerethepredominantinflammatorycelltypepresent,
comparedtothepredominanceofneutrophilsseeninourstudyfollowingtheNACinfusion.One
previousstudyusingintra-uterineNAC(Gores-Lindholmetal.2013)demonstratedanincrease
in lymphocytesandplasmacellsonlyonendometrialbiopsy followingNAC infusion inbarren
mares,buttheauthorsnotedthatthesechangesweretoochronicinnaturetobeduetoNAC
infusion.
Theosmolalityof3.3%NACsolutionusedinthepresentstudywasconsiderablyhigher
thanthe0.9%salineusedinthecontrolcycles(saline=280mOsm;NAC=668mOsm).Whileno
studieshaveexaminedtheeffectsofhyperosmoticsolutionsontheendometriumspecifically,
multiple studies have shown that hyperosmotic stressors are damaging to cells and are pro-
inflammatoryinnature(Nemethetal.2002;Luoetal.2005;Schwartzetal.2008;Brockeretal.
2012). Exposure of cells to hyperosmotic solutions leads to an efflux of fluid out of the cell,
shrinkageofthecell,alterationinproteinsynthesis,anddamagetoboththecytoskeletonand
nucleus. This can lead to cellular apoptosis (Brocker et al. 2012). It is possible that the
inflammatory reaction seen in our study was further exacerbated by insemination 12 hours
followingNACinfusion,limitingtheabilityofthealreadycompromisedendometriumtoclearthe

72
inflammation. In previous studies that administered intra-uterine NAC, treatment was often
given 24-36 hours prior to breeding (LeBlanc and Causey 2009; Gores-Lindholm et al. 2009;
LeBlanc2010;Lyle2012;Troedsson2012;Gores-Lindholmetal.2013;Buczkowskaetal.2015),
perhapsallowingforsomerecoverypriortoanotherinsult.Giventhebiopsyfindingsinourstudy,
onecouldspeculatethatintra-uterineinfusionofNACmaynotbeasinertaspreviouslybelieved.
TreatmentwithNACdidnot improveclinical signsofPBIE, including IUF. Intrauterine
fluid retention occurs in PBIE mares for several reasons, including lack of cervical dilation,
pendulousuterusandpooruterinecontractility.Cervicalpatencyisknowntoimproveuterine
drainage (Pycock 2007, LeBlanc andCausey2009, Reliaset al. 2016). Some studymareshad
fibrotic and tortuous cervices that resulted fromuseas repeatedembryodonors inprevious
research studies. Therefore, somemares retained considerable fluid, leading to a significant
amountofvariabilityinIUFlevelsbetweenmares.Thismayhavelimitedtheapparentefficacyof
intra-uterineNACtreatmentinourstudy.
The PBIE-susceptible mares in our study demonstrated the expected inflammatory
responseovertime,includingchangesinIUF,uterineedemascoresand%neutrophils.IUFwas
mostelevatedat36hoursanddiminishedthereafter.Myometrialcontractionsareoneofthe
mainmechanismsofuterineclearance(Troedsson2006;LeBlancandCausey2009;Troedsson
andWoodward2016;ChristoffersenandTroedsson2017).Normalmaresclearintra-uterinefluid
within24hoursfollowingAI(Katila1995);itisconsideredabnormaltofindanyintra-uterinefluid
greater than 24 hours post-ovulation and into diestrus (Newcombe 1998). However, PBIE-
susceptiblemaresdonotdemonstratenormaluterineclearance.LeBlancetal.(1994a;1995)
found that susceptible mares cleared negligible amounts of radiocolloid 4 hours following

73
infusion;furthermore,Indiainkwasmaintainedforupto72hourspost-infusion.Furthermore,
Troedsson et al. (1993a) showed that susceptible mares inoculated with Streptococcus
zooepidemicus had a marked decrease in myometrial activity between 7-17 hours post-
inoculationcomparedtoresistantmares.Inourstudy,IUFwasgreatestat36hoursbutnotat12
hourspost-AI.Troedssonetal.(1993a)demonstratedthatthepatternofuterinecontractilityof
PBIE-susceptible and normal mares was similar initially following a challenge, however the
durationand intensityofmyometrial contractions rapidlydeclined inPBIE-susceptiblemares,
whereasinresistantmaresitdidnot.ThismayexplaintheincreaseinIUFseenfrom12to36
hoursinourstudymares.
EndometrialedemaandestimatedIUFwerenotsignificantlycorrelatedinthisstudy.This
iscontradictorytoKotilainenetal.(1994)wherepre-treatment(AI,extender,seminalplasma,
PBS)uterinefluidlevelwaspositivelycorrelatedwithuterineedema.Inaddition,Samper(2007),
foundthatpresenceoffluidatthetimeofovulationwasmorecommonlynotedinmareswith
gradethree,fourandfiveedemascores.Thesedifferencescouldbeaccountedforbythefact
thatpre-treatmentfluidlevelscouldvaryfrompost-treatmentfluidlevelsandthatthegroupof
maresinthosestudieswerenormalandnotscreenedasPBIE-susceptible.
Percent neutrophils on cytologywere highest at 12 hours post-AI in our studywhen
treatmentcycleswerecombined.Aninfluxofneutrophilsintotheendometriumoccursfollowing
infusionwithspermatozoainnormalmares,andwasfirstdocumentedbyKotilainenetal.(1994).
Innormalmares,neutrophilsappearat4hours,peakat8hoursandareabsentby48hourspost-
breeding(Katila1995).Thisresponseisverysimilartothatnotedinnormalmaressubjectedto
intra-uterineinfusionofStreptococcuszooepidemicusfourhoursfollowingbreeding(Troedsson

74
1995).AsimilarpatternofneutrophilinfiltrationoccursinPBIE-susceptiblemares,butismuch
moresevere.Neutrophilsweresignificantlyelevatedinsusceptiblemaresasearlyas2hourspost
AI, compared to 6 hours post AI in resistant mares (Woodward et al. 2013a). Endometrial
neutrophil infiltration peaked in both groups at 6 hours post AI, and decreased thereafter,
howeverneutrophilswereconsistentlyelevatedinsusceptiblemaresthroughout(Woodwardet
al.2013a).Overall,persistentinflammationwasafeatureofourPBIE-susceptiblestudymares,
consistentwithpreviousfindings.
DoseandtimingofadministrationofNACmayhaveaffectedefficacyinthepresentstudy.
A3.3%solutionwasselectedbasedonpreviousstudiesusingintra-uterineNACinmaresasa
mucolytic(LeBlancandCausey2009;LeBlanc2010;Lyle2012;Troedsson2012;Gores-Lindholm
etal.2013;Buczkowskaetal.2015;Scoggin2016).Sub-fertilemareswithahistoryofmucus
hyper-secretion that received an intrauterine infusion of 3.3% NAC pre-breeding, attained a
pregnancyrateof77%(Gores-Lindholmetal.2013).However,researchinotherfields,suchas
incysticfibrosis(humans)androdentpulmonarystudies,suggeststhatthedoseofNACneeds
tobesubstantialenoughtocreateananti-inflammatoryeffect(Pahanetal.1997;Tirouvanziam
etal.2006).ItisthereforepossiblethattheselecteddoseofNACinourstudywasadequatefor
itspreviouslyknownmucolyticeffects,butnothighenoughtocausedirectanti-inflammatory
effects.NACwasadministeredpre-breedinginourclinicaltrial,inthesamefashionasprevious
studies(Gores-Lindholmetal.2013).Ratintra-peritoneal,cellcultureandtissuemodelstudies
havedemonstratedtheanti-inflammatoryeffectsofNACwhenadministeredasapre-treatment
priortoinductionofinflammation(Pahanetal.1997;Majanoetal.2004;Cazzolaetal.2017).
Together,thisevidenceprovidedthebasisforthetimingofNACinfusioninourstudy.However,

75
since no other studies have examined NAC treatment specifically in PBIE mares, treatment
followingtheinflammatoryresponseinducedbybreedingmightprovemorebeneficial.Infact,
oralNACtreatmentofpatientsclinicallyillwithcysticfibrosisledtomodulationofthepulmonary
inflammationseenwiththiscondition(Tirouvanziametal.2006).
Inconclusion,intra-uterinetreatmentwith3.3%NACdidnotimproveclinicalsignsinPBIE
mares.NACinfusionwasassociatedwithasignificantandprolongedacuteinflammatoryreaction
oftheendometrium,whichwassuspectedtobeduetothehyperosmoticnatureofNAC.Further
studiesareneededtoexploretheeffectsofhypertonicsolutionsontheendometriuminboth
normal and PBIE-susceptible mares. Additional studies evaluating alternative non-irritating
immunomodulatory treatments such as platelet-rich plasma, which might normalize the
inflammatoryresponseofPBIEmares,wouldbeofvalue.

76
2.6REFERENCES
1. Alghamdi,A.S.,Foster,D.N.,Carlson,C.S.andTroedsson,M.H.T.(2005).Nitricoxidelevelsand nitric oxide synthase expression in uterine samples from mares susceptible andresistanttopersistentbreeding-inducedendometritis.Am.J.Reprod.Immunol.53,230–237.
2. Asbury,A.C.,Halliwell,R.E.W.,Foster,G.W.andLongino,S.J.(1980).Immunoglobulinsin
uterinesecretionsofmareswithdifferingresistancetoendometritis.Theriogenology.14,299-308.
3. Bergamini,S.,Rota,C.,Canali,R.,Staffieri,M.,Daneri,F.,Bini,A.,Giovannini,F,Tomasi,A.
andIannone,A.(2001).N-acetylcysteineinhibitsinvivonitricoxideproductionbyinduciblenitricoxidesynthase.NitricOxide.5,349-360.
4. Bracher,V.,Neuschaefer,A.andAllen,R.W.(1991).Theeffectofintra-uterineinfusionof
keroseneontheendometriumofmares.J.Reprod.Fert.Suppl44,706-707.5. Brendemuehl,J.P.(2002).Effectofoxytocinandcloprostenolonlutealformation,function
andpregnancyratesinmares.Theriogenology.58,623-626.6. Brinsko,S.P.,Rigby,S.L.,Varner,D.D.andBlanchard,T.L. (2003).Apracticalmethodfor
recognizing mares susceptible to post-breeding endometritis. Proc. Am. Assoc. Equine.Pract.49,363–365.
7. Brocker, C., Thompson,D.C. andVasiliou, V. (2012). The role of hyperosmotic stress in
inflammationanddisease.Biomol.Concepts.3,345-364.8. Cadario,M.E.,Thatcher,W.W.,Klapstein,E.,Merrit,A.M.,Archbald,L.F.,Thatcher,M.J.and
LeBlanc,M.M.(1999).Dynamicsofprostaglandinsecretion,intrauterinefluidanduterineclearance in reproductively normal mares and mares with delayed uterine clearance.Theriogenology.52,1181-1192.
9. Canisso, I.F., Stewart, J. and Coutinho da Silva, M.A. (2016). Endometritis - managing
persistentpost-breedingendometritis.Vet.Clin.Equine.32,465-480.10. Cazzola,M.,Calzetta,L.,Facciolo,F.,Rogliani,P.andMatera,M.G.(2017).Pharmacological
investigationontheanti-oxidantandanti-inflammatoryactivityofN-acetylcysteineinanexvivomodelofCOPDexacerbation.Resp.Res.18.
11. Christoffersen,M.andTroedsson,M.H.T.(2017). Inflammationandfertility inthemare.
Reprod.Dom.Anim.Suppl52,14-20.

77
12. Christoffersen,M.,Woodward, E.M., Bojesen,A.M., Petersen,M.R., Squires, E.L., Lehn-Jensen, H. and Troedsson,M.H.T. (2012). Effect of immunomodulatory therapy on theendometrial inflammatory response to induced infectious endometritis in susceptiblemares.Theriogenology.78,991-1004.
13. Combs,G.B.,LeBlanc,M.M.,Neuwirth,L.andTran,T.Q.(1995).EffectsofprostaglandinF2α,
cloprostenol and fenprostalene on uterine clearance of radiocolloid in the mare.Theriogenology.45,1449-1455.
14. Fumoso,E.A.,Aguilar,J.,Giguere,S.,Rivulgo,M.,Wade,J.andRogan,D.(2007).Immune
parameters inmaresresistantandsusceptibletopersistentpost-breedingendometritis:effectsofimmunomodulation.Vet.Immunol.Immunopathol.118,30-39.
15. Fumoso, E., Giguere, S.,Wade, J., Rogan,D., Videla-Dorna, I. andBowden, R.A. (2003).
EndometrialIL-1β,IL-6andTNF-α,mRNAexpressioninmaresresistantorsusceptibletopost-breeding endometritis. Effects of estrous cycle, artificial insemination andimmunomodulation.Vet.Immunol.Immunopathol.96,31-41.
16. Gores-Lindholm, A.R., LeBlanc, M.M., Causey, R., Hitchborn, A., Fayrer-Hosken, R.A.,
Kruger,M.,Vandenplas,M.L.,Flores,P.andAhlschwede,S.(2013).Relationshipsbetweenintrauterine infusion of N-acetylcysteine, equine endometrial pathology, neutrophilfunction,post-breedingtherapy,andreproductiveperformance.Theriogenology.80,218-227.
17. Hurtgen,J.P.(2006).Pathogenesisandtreatmentofendometritisinthemare:Areview.
Theriogenology.66,560-566.18. Katila, T. (1995). Onset and duration of uterine inflammatory response of mares after
inseminationwithfreshsemen.Biol.Reprod.Mono.1,515-517.19. Kenney,R.M.(1978).Cyclicandpathologicchangesofthemareendometriumasdetected
bybiopsy,withanoteonearlyembryonicdeath.J.Am.Vet.Med.Assoc.172,241–62.20. Khan,F.A.,Chenier,T.S.,Foster,R.A.,Hewson,J.andScholtz,E.L.(2018).Endometrialnitric
oxide synthase activity inmares susceptibleor resistant topersistent breeding-inducedendometritisandtheeffectofaspecific iNOS inhibitor invitro.Reprod.Dom.Anim.53,718-724.
21. Kotilainen,T.,Huhtinen,M.andKatila,T.(1994).Sperm-inducedleukocytosisintheequine
uterus.Theriogenology.41,629-636.22. LeBlanc,M.M.(2010).Advancesinthediagnosisandtreatmentofchronicinfectiousand
post-mating-inducedendometritisinthemare.Reprod.Dom.Anim.Suppl45,21-27.

78
23. LeBlanc,M.M.andCausey,R.C.(2009).Clinicalandsubclinicalendometritisinthemare:
boththreatstofertility.Reprod.Dom.Anim.Suppl3,10-22.
24. LeBlanc, M.M., Johnson, R.D., Calderwood Mays, M.B. and Valderrama, C. (1995).Lymphaticclearanceofindiainkinreproductivelynormalmaresandmaressusceptibletoendometritis.Biol.Reprod.Mono.1,501–506.
25. LeBlanc,M.M.,Neuwirth,L.,Asbury,A.C.,Tran,T.,Mauragis,D.andKlapstein,E.(1994a).Scintigraphicmeasurementofuterineclearanceinnormalmaresandmareswithrecurrentendometritis.EquineVet.J.26,109–113.
26. LeBlanc,M.M., Neuwirth, L.,Mauragis, D., Klapstein, E. and Tran, T. (1994b). Oxytocin
enhancesclearanceofradiocolloidfromtheuterinelumenofreproductivelynormalmaresandmaressusceptibletoendometritis.EquineVet.J.26,279-282.
27. Liu,I.K.M.,Cheung,A.T.W.,Walsh,E.M.andAyin,S.(1986).Thefunctionalcompetenceof
uterine-derived polymorphonuclear neutrophils (PMN) from mares resistant andsusceptibletochronicuterineinfection:Asequentialmigrationanalysis.Biol.Reprod.35,1168-1174.
28. Luo, L., De-Quan, L., Corrales, R. and Pflugfelder, S. (2005). Hyperosmolar saline is a
proinflammatorystressonthemouseocularsurface.EyeContactLens31,186-193.29. Majano, P.L., Medina, J., Zubia, I., Sunyer, L., Lara-Pezzi, E., Maldonado-Rodriguez, A.,
Lopez-Cabrera,M. andMoreno-Otero, R. (2004). N-acetyl-cysteinemodulates induciblenitricoxidesynthasegeneexpressioninhumanhepatocytes.J.Hepatol.40.
30. Melkus,E.,Witte,T.,Walter,I.,Heuwieser,W.andAurich,C.(2013).Investigationsonthe
endometrialresponsetointrauterineadministrationofN-acetylcysteineinoestrousmares.Reprod.Dom.Anim.48,591-597.
31. Mitchell, G., Liu, I.K.M., Perryman, L.E., Stabenfeldt, G.H. and Hughes, J.P. (1982).
Preferentialproductionandsecretionofimmunoglobulinsbytheequineendometrium–amucosalimmunesystem.J.Reprod.Fert.Suppl.32,161-168.
32. Nemeth,Z.H.,Deitch,E.A.,Szabo,C.andHasko,G. (2002).Hyperosmoticstress induces
nuclear factor-κB activation and interleukin-8 production in human intestinal epithelialcells.Am.J.Pathol.161,987-996.
33. Nie,G.J., Johnson, K.E.,Wenzel, J.G.W. andBraden, T.D. (2002). Effect of periovulatory
ecbolicsonlutealfunctionandfertility.Theriogenology.58,461-463.

79
34. Nie,G.J.,Johnson,K.E.,Wenzel,J.G.W.andBraden,T.D.(2003).Lutealfunctioninmaresfollowingadministrationofoxytocin,cloprostenolorsalineonday0,1or2post-ovulation.Theriogenology.60,1119-1125.
35. Neuwirth,L.,LeBlanc,M.M.,Mauragis,D.,Klapstein,E.andTran,T.(1994).Scintigraphic
measurementofuterineclearanceinmares.Vet.Radiol.Ultrasound.36,64-68.36. NewcombeJ.R.(1997).Theeffectoftheincidenceanddepthofintra-uterinefluidinearly
dioestrusonpregnancyrateinmares.Pferdeheilkunde.13,545.37. Nikolakopoulos, E. and Watson, E.D. (1997). Does artificial insemination with chilled,
extended semen reduce the antigenic challenge to the mare’s uterus compared withnaturalservice?Theriogenology.47,583-590.
38. Nikolakopoulos, E. and Watson, E.D. (1999). Uterine contractility is necessary for the
clearance of intrauterine fluid but not bacteria after bacterial infusion in the mare.Theriogenology.52,413-423.
39. Pahan,K.,Sheikh,F.G.,Namboodiri,A.M.S.andSingh,I.(1997).N-acetylcysteineinhibits
induction of NO production by endotoxin or cytokine stimulated rat peritonealmacrophages,C6glialcellsandastrocytes.FreeRadic.Biol.Med.24,39-48.
40. Paintlia, M.K., Paintlia, A.S., Singh, A.K. and Singh, I. (2008). Attenuation of
lipopolysaccharideinducedinflammatoryresponseandphospholipidsmetabolismatthefeto-maternalinterfacebyN-acetylcysteine.Pediatr.Res.64,334-339.
41. Pycock,J.F.(2007).Therapyformareswithuterinefluid,1stedn,Ed:CurrentTherapyin
Equine Reproduction. 1st edn., Ed: J. C. Samper, J.F. Pycock, A.O. McKinnon, SaundersElsevier,Missouri.pp93-104.
42. Rigby, S.L., Barhoumi, R., Burghardt, R.C., Colleran, P., Thompson, J.A., Varner, D.D.,
Blanchard,T.L.,Brinsko,S.P.,Taylor,T.,Wilkerson,M.K.andDelp,M.D.(2001).Mareswithdelayeduterine clearancehavean intrinsicdefect inmyometrial function.Biol.Reprod.65,740–747.
43. Samper,J.C.(1997).Ultrasonographicappearanceandpatternofuterineedematotimeof
ovulationinmares.Proc.Annul.ConventionAAEP.43,189-191.44. Samper, J.C. (2007).How to interpret endometrial edema in broodmares.Proc.Annul.
ConventionAAEP.53,571-572.

80
45. Schwartz,L.,Abolhassani,M.,Pooya,M.,Steyaert,J.-M.,Wertz,X.,Isreal,M.,Guais,A.andChaumet-Riffaud, P. (2008). Hyperosmotic stress contributes to mouse colonicinflammation through the methylation of protein phosphatase A2. Am. J. Physiol.Gastrointest.LiverPhysiol.295,G934-G941.
46. Stanislaus,R.,Gilg,A.G.,Singh,A.K.,Singh, I. (2005).N-acetyl-L-cysteineamelioratesthe
inflammatory disease process in experimental autoimmune encephalomyelitis in Lewisrats.J.Autoimmun.2.
47. Tirouvanziam,R.,Conrad,C.K.,Bottiglieri,T.,Herzenberg,L.A.,Moss,R.B.andHerzenberg,
L.A. (2006). High dose oral N-acetylcysteine, a glutathione prodrug, modulatesinflammationincysticfibrosis.Proc.Natl.Acad.Sci.103,4628-4633.
48. Troedsson,M.H.T.(1991).Uterinedefensemechanismsinthemare.(thesis).49. Troedsson,M.H.T.(1995).Uterineresponsetosemendepositioninthemare.Proc.Soc.
Theriogenol.130-135.50. Troedsson,M.H.T. (1997). Therapeutic considerations formating-induced endometritis.
Pferdeheilkunde.13,516-520.51. Troedsson,M.H.T. (2006).Breeding inducedendometritis inmares.Vet.Clin.Equine22,
705-712.52. Troedsson,M.H.T.(2011).Endometritis.In:EquineReproduction.2ndedn,Ed:McKinnon,
A.O.,Squires,E.L.,Vaala,W.E.,Warner,D.D.,Wiley-Blackwell,NewJersey.pp.2608–2619.53. Troedsson, M.H.T and Woodward, E.M. (2016). Our current understanding of the
pathophysiology of equine endometritis with an emphasis on breeding-inducedendometritis.Biol.Reprod.16,8-12.
54. Troedsson,M.H.T.,Ababneh,M.M.,Ohlgreen,A.F.,Madill,S.,Vetscher,N.andGregas,M.
(2001). EffectofprostaglandinF2α onpregnancy ratesand luteal function in themare.Theriogenology.55,1891-1899.
55. Troedsson,M.H.T.,Liu,I.K.M.,Ing,M.,Pascoe,J.andThurmond,M.(1993a).Multiplesiteelectromyography recordings of uterine activity following an intrauterine bacterialchallengeinmaressusceptibleandresistanttochronicuterineinfection.J.Reprod.Fertil.99,307–313.
56. Troedsson,M.H.T.,Liu,I.K.M.andThurmond,M.(1993b).Immunoglobulin(IgGandIgA)and complement (C3) concentrations in uterine secretion following an intrauterinechallenge of Streptococcus zooepidemicus in mares susceptible to versus resistant tochronicuterineinfection.Biol.Reprod.49,502-506.

81
57. Troedsson,M.H.T.,Liu,I.K.M.andThurmond,M.(1993c).Functionofuterineandblood-
derived polymorphonuclear neutrophils in mares susceptible and resistant to chronicuterineinfection:phagocytosisandchemotaxis.Biol.Reprod.49,507-514.
58. vonReitzenstein,M.,Callahan,M.A.,Hansen,P.J.andLeBlanc,M.M.(2002).Abberations
inuterinecontractilepatternsinmareswithdelayeduterineclearanceafteradministrationofdetomidineandoxytocin.Theriogenology.58,887-898.
59. Watson,E.D.(2000).Post-breedingendometritisinthemare.Anim.Reprod.Sci.60-61,221-
232.60. Watson,E.D., Stokes,C.R. andBourne, F.J. (1987).Cellular andhumoralmechanisms in
maressusceptibleandresistanttopersistentendometritis.Vet.Immunol.Immunopathol.16,107–21.
61. Watson E.D., Stokes, C.R. and Bourne, F.J. (1988). Concentration of immunoreactive
leukotrieneB4inuterinelavagefluidfrommareswithexperimentallyinducedandnaturallyoccurringendometritis.J.Vet.Pharmacol.Therap.11,130-134.
62. Williamson,P.,Dunning,A.,O’Connor,J.andPenhale,W.J.(1983).Immunoglobulinlevels,
proteinconcentrationsandalkalinephosphataseactivityinuterineflushingsfrommareswithendometritis.Theriogenology.19,441-448.
63. Witte, T.S.,Melkus, E.,Walter, I., Senge, B., Schwab, S., Aurich, C. and Heuwieser,W.
(2012). Effects of oral treatment with N-acetylcysteine on the viscosity of intrauterinemucusandendometrialfunctioninestrousmares.Theriogenology.78,1199-1208.
64. Woodward,E.M.andTroedsson,M.H.T.(2013)Equinebreeding-inducedendometritis.J.
EquineVet.Sci.33,673–82.65. WoodwardE.M.andTroedsson,M.H.T.(2015).Inflammatorymechanismsofendometritis.
EquineVet.J.47,384-389.66. Woodward,E.M.,Christoffersen,M.,Campos,J.,Betancourt,A.,Horohov,D.,Scoggin,K.E.,
Squires,E.L.andTroedsson,M.H.T.(2013a).Endometrialinflammatorymarkersoftheearlyresponse inmares susceptibleor resistant topersistentbreeding-inducedendometritis.Reproduction.145,289-296.
67. Woodward,E.M.,Christoffersen,M.,Campos,J.,Horohov,D.W.,Scoggin,K.E.,Squires,E.
andTroedsson,M.H.T.(2013b).Aninvestigationofuterinenitricoxideproductioninmaressusceptibleandresistanttopersistentbreeding-inducedendometritisandtheeffectsofimmunomodulation.Reprod.Dom.Anim.48,554–561.

82
68. Woodward, E.M., Christoffersen, M., Horohow, D., Squires, E.L. and Troedsson, M.H.T.
(2015). The effect of treatment with immune modulators on endometrial cytokineexpressioninmaressusceptibletopersistentbreeding-inducedendometritis.EquineVet.J.47,235-239.
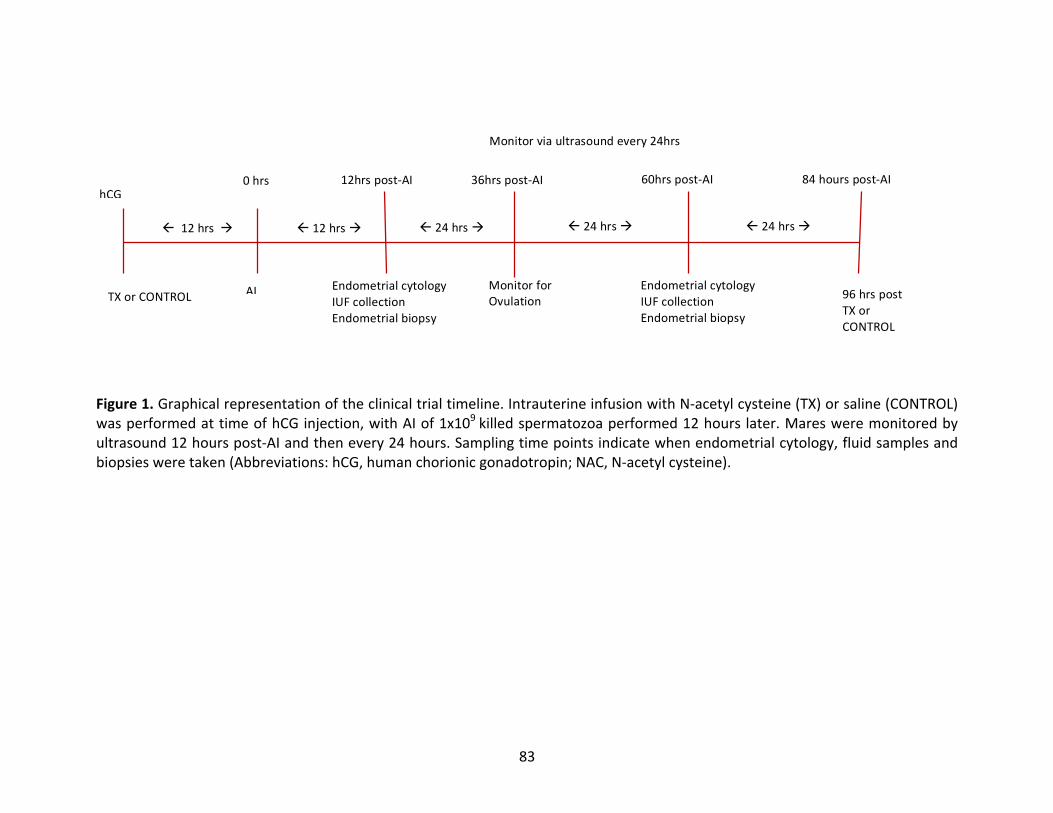
83
Figure1.Graphicalrepresentationoftheclinicaltrialtimeline.IntrauterineinfusionwithN-acetylcysteine(TX)orsaline(CONTROL)wasperformedattimeofhCGinjection,withAIof1x109killedspermatozoaperformed12hours later.Maresweremonitoredbyultrasound12hourspost-AIandthenevery24hours.Samplingtimepointsindicatewhenendometrialcytology,fluidsamplesandbiopsiesweretaken(Abbreviations:hCG,humanchorionicgonadotropin;NAC,N-acetylcysteine).
ß24hrsà
hCG
TXorCONTROL
ß12hrsà
AI
ß12hrsà
96hrspostTXorCONTROL
Monitorviaultrasoundevery24hrs
12hrspost-AI 36hrspost-AI 60hrspost-AI 84hourspost-AI
ß24hrsà
EndometrialcytologyIUFcollectionEndometrialbiopsy
EndometrialcytologyIUFcollectionEndometrialbiopsy
MonitorforOvulation
0hrs
ß24hrsà
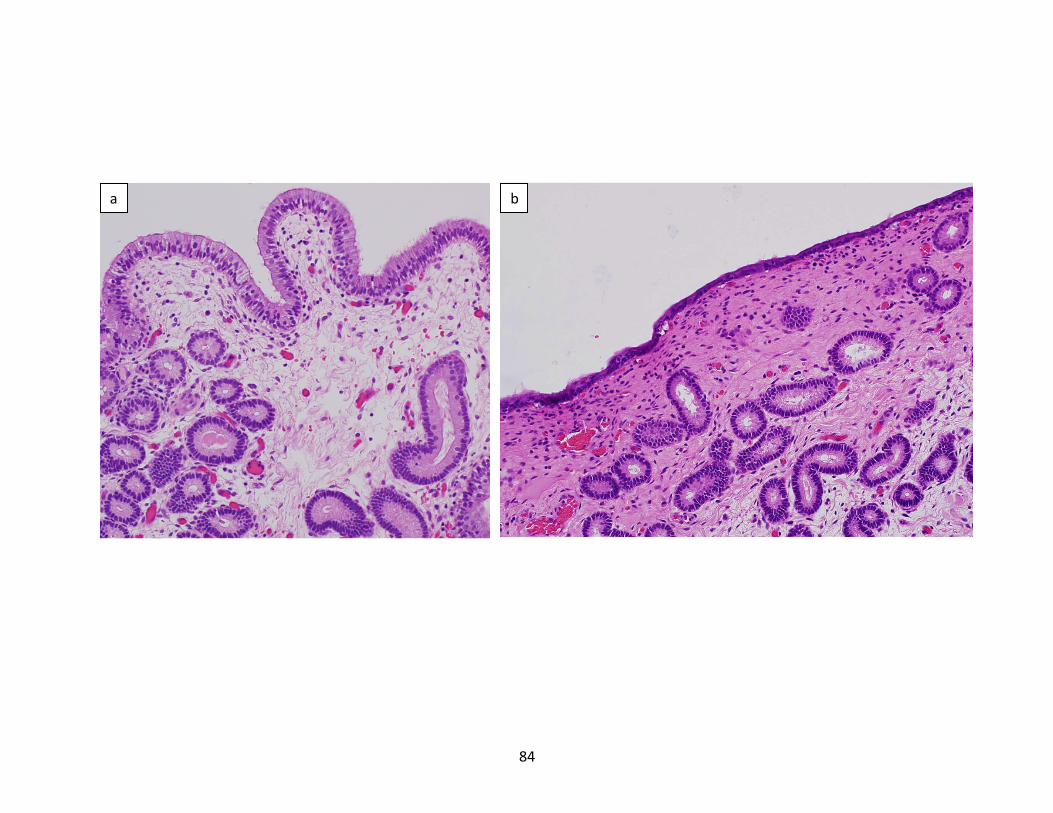
84
a b
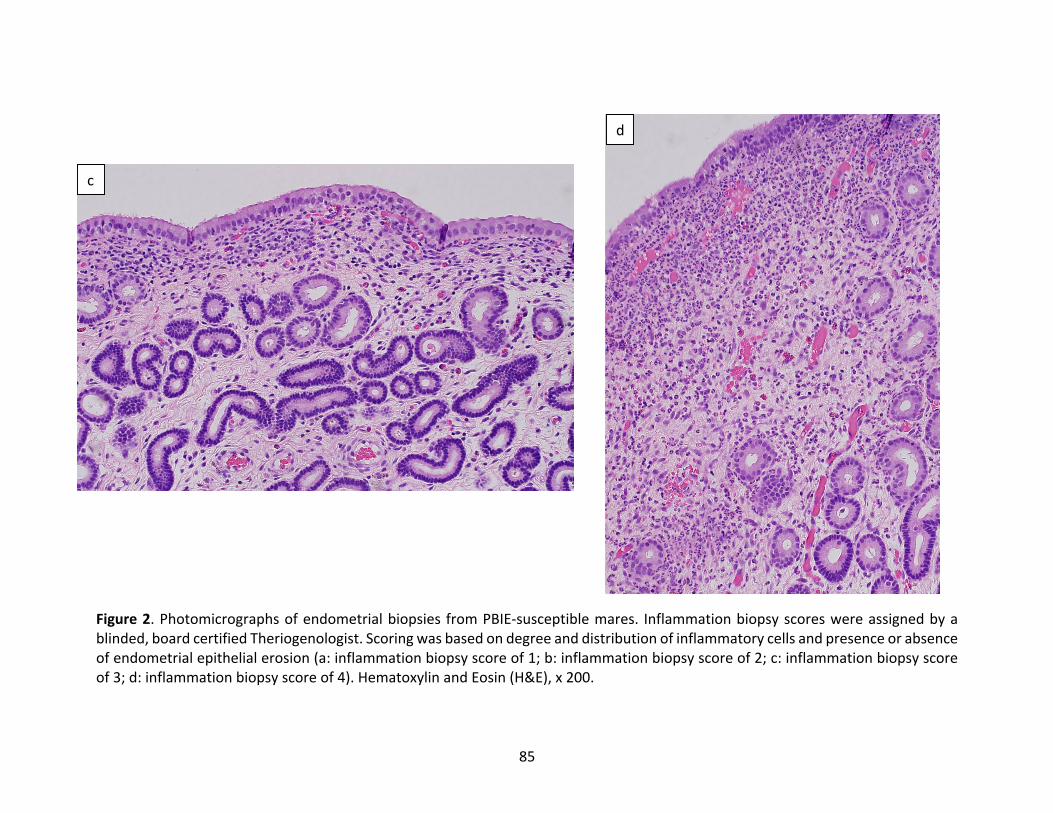
85
Figure2.Photomicrographsofendometrialbiopsies fromPBIE-susceptiblemares. Inflammationbiopsyscoreswereassignedbyablinded,boardcertifiedTheriogenologist.Scoringwasbasedondegreeanddistributionofinflammatorycellsandpresenceorabsenceofendometrialepithelialerosion(a:inflammationbiopsyscoreof1;b:inflammationbiopsyscoreof2;c:inflammationbiopsyscoreof3;d:inflammationbiopsyscoreof4).HematoxylinandEosin(H&E),x200.
c
d
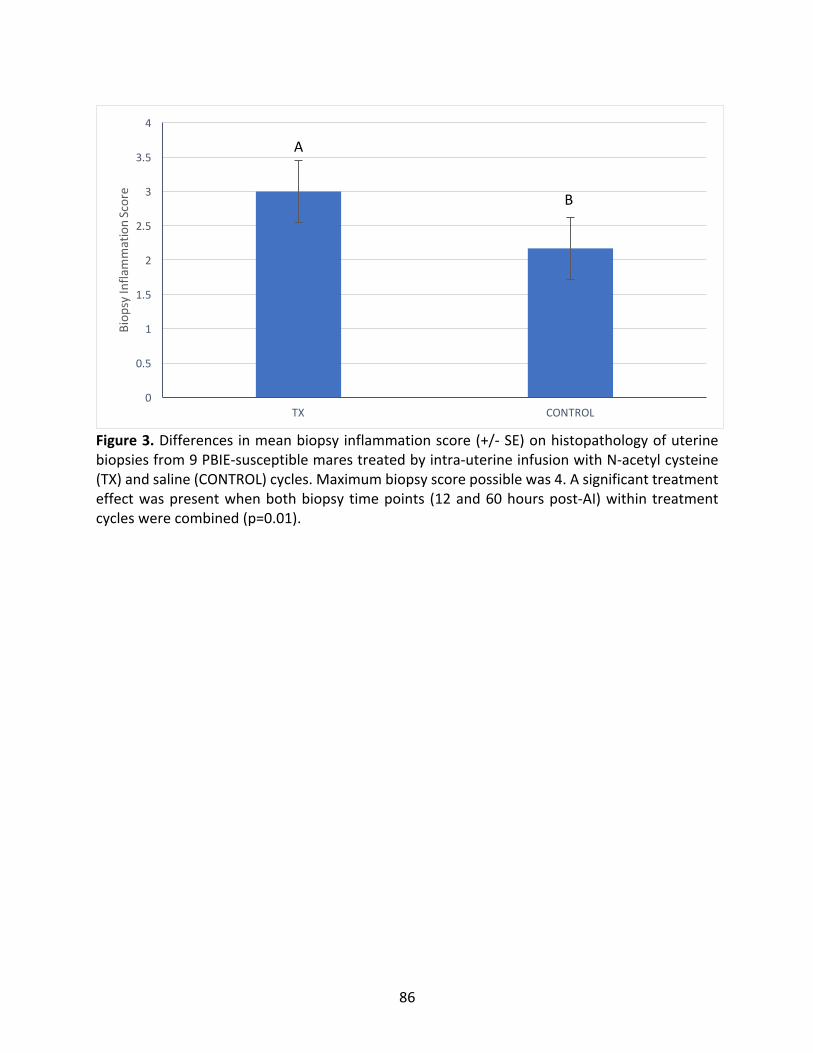
86
Figure3.Differencesinmeanbiopsyinflammationscore(+/-SE)onhistopathologyofuterinebiopsiesfrom9PBIE-susceptiblemarestreatedbyintra-uterineinfusionwithN-acetylcysteine(TX)andsaline(CONTROL)cycles.Maximumbiopsyscorepossiblewas4.Asignificanttreatmenteffectwaspresentwhenbothbiopsy timepoints (12and60hourspost-AI)within treatmentcycleswerecombined(p=0.01).
0
0.5
1
1.5
2
2.5
3
3.5
4
TX CONTROL
Biop
syInfla
mmationScore
A
B
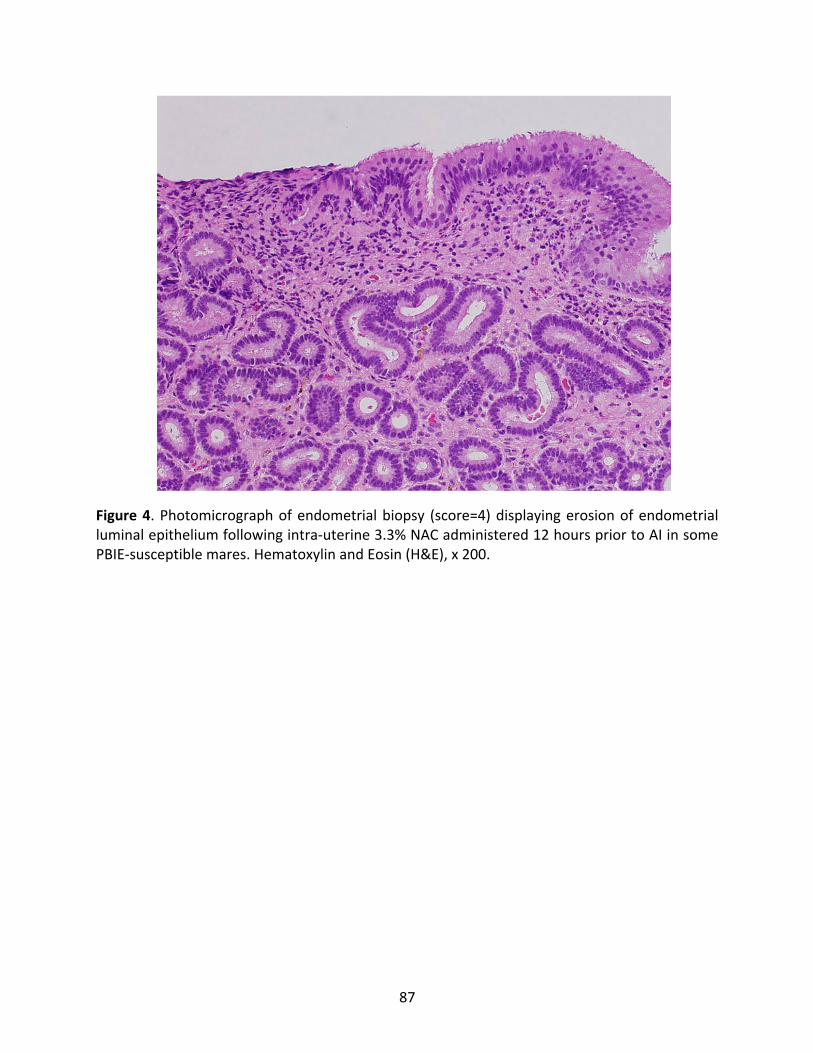
87
Figure4.Photomicrographofendometrialbiopsy (score=4)displayingerosionofendometrialluminalepitheliumfollowingintra-uterine3.3%NACadministered12hourspriortoAIinsomePBIE-susceptiblemares.HematoxylinandEosin(H&E),x200.
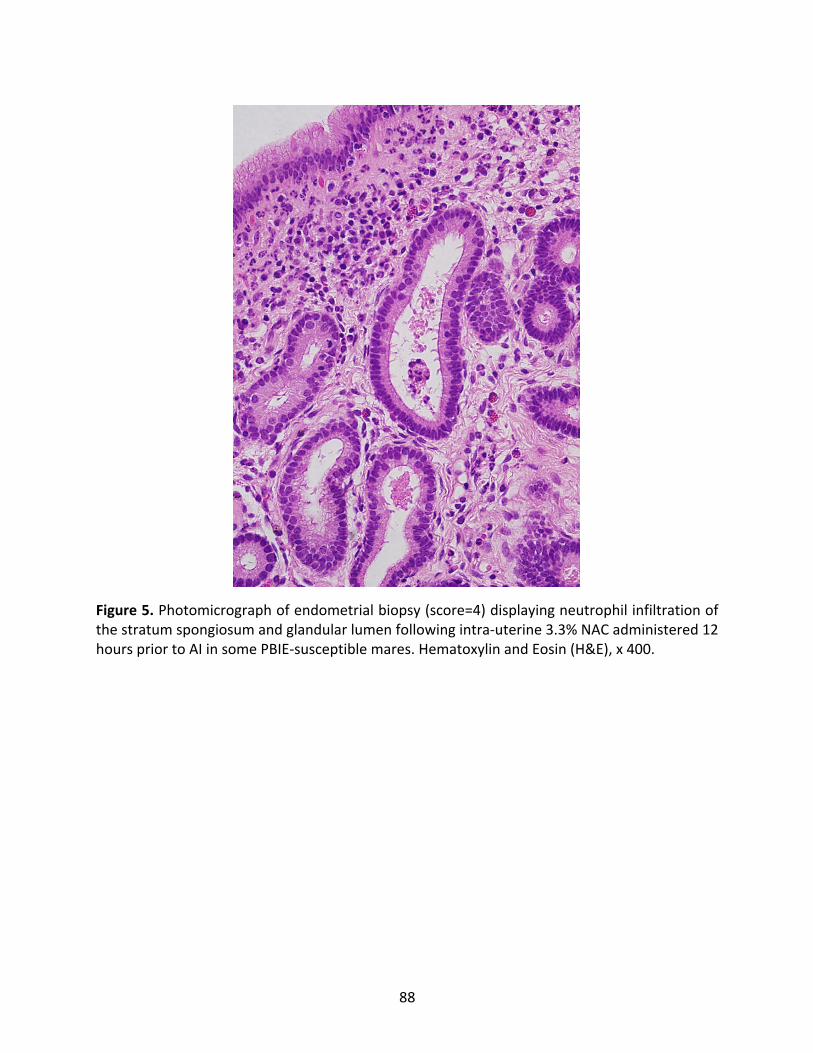
88
Figure5.Photomicrographofendometrialbiopsy(score=4)displayingneutrophilinfiltrationofthestratumspongiosumandglandularlumenfollowingintra-uterine3.3%NACadministered12hourspriortoAIinsomePBIE-susceptiblemares.HematoxylinandEosin(H&E),x400.
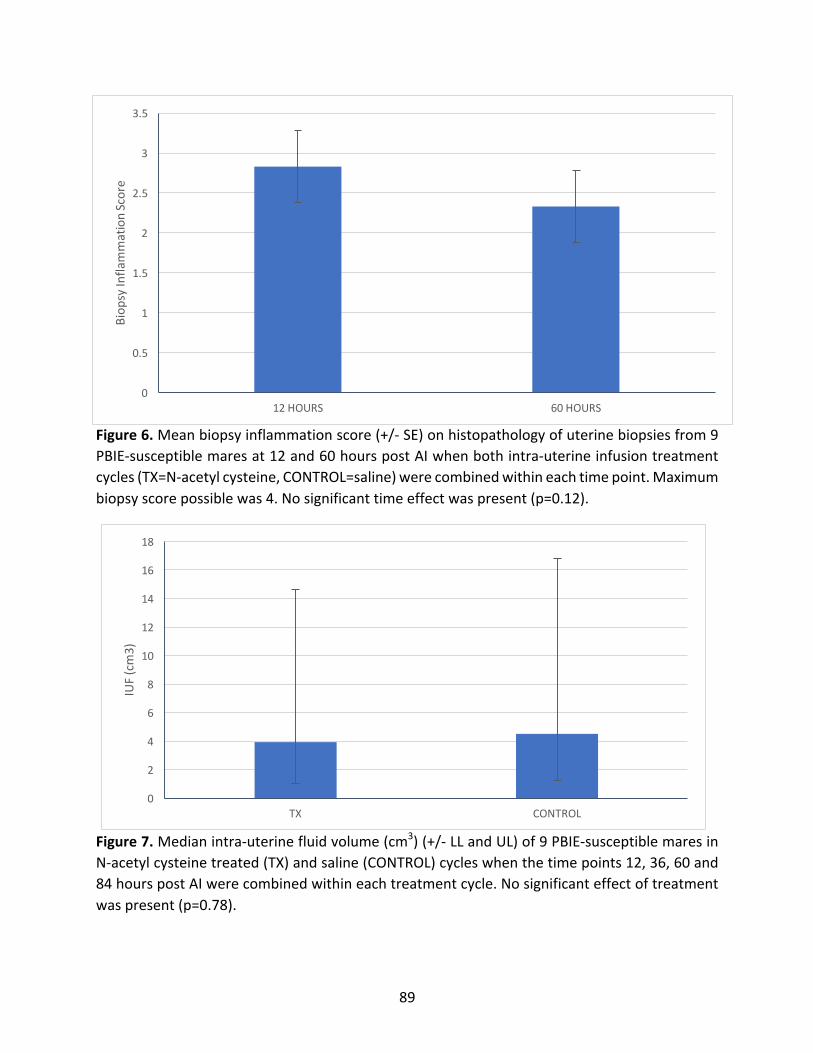
89
Figure6.Meanbiopsyinflammationscore(+/-SE)onhistopathologyofuterinebiopsiesfrom9PBIE-susceptiblemaresat12and60hourspostAIwhenbothintra-uterineinfusiontreatmentcycles(TX=N-acetylcysteine,CONTROL=saline)werecombinedwithineachtimepoint.Maximumbiopsyscorepossiblewas4.Nosignificanttimeeffectwaspresent(p=0.12).
Figure7.Medianintra-uterinefluidvolume(cm3)(+/-LLandUL)of9PBIE-susceptiblemaresinN-acetylcysteinetreated(TX)andsaline(CONTROL)cycleswhenthetimepoints12,36,60and84hourspostAIwerecombinedwithineachtreatmentcycle.Nosignificanteffectoftreatmentwaspresent(p=0.78).
0
0.5
1
1.5
2
2.5
3
3.5
12HOURS 60HOURS
Biop
syInfla
mmationScore
0
2
4
6
8
10
12
14
16
18
TX CONTROL
IUF(cm3)
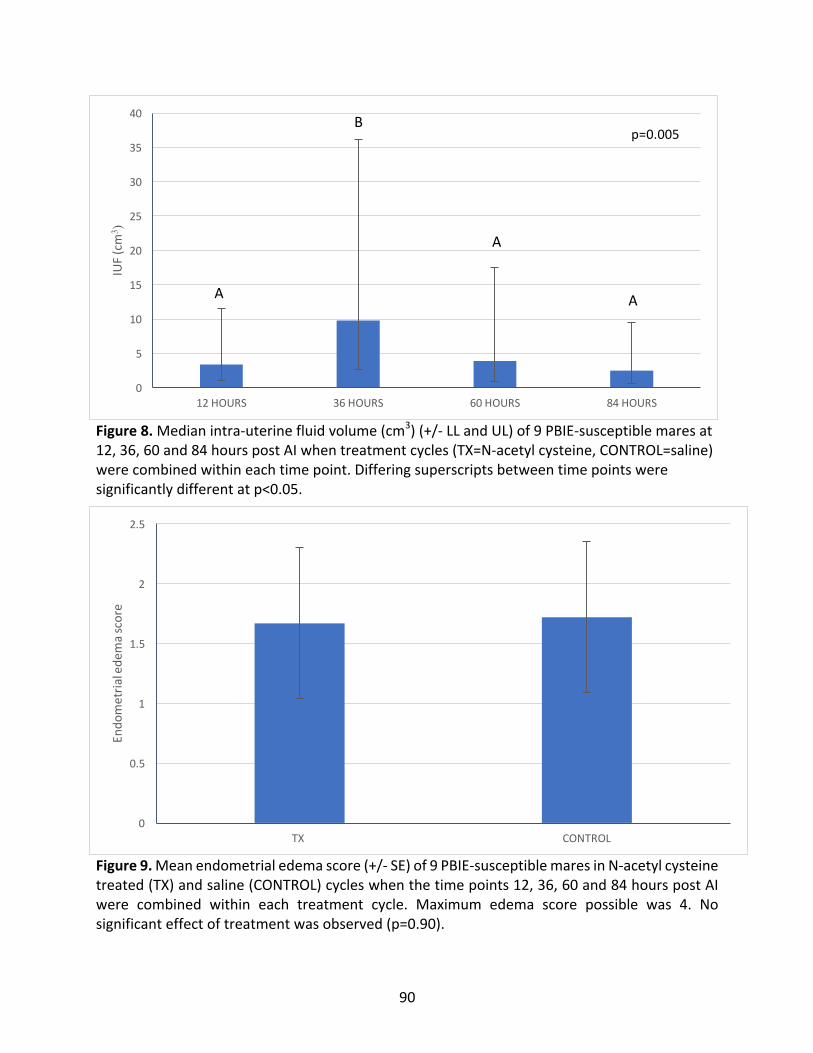
90
Figure8.Medianintra-uterinefluidvolume(cm3)(+/-LLandUL)of9PBIE-susceptiblemaresat12,36,60and84hourspostAIwhentreatmentcycles(TX=N-acetylcysteine,CONTROL=saline)werecombinedwithineachtimepoint.Differingsuperscriptsbetweentimepointsweresignificantlydifferentatp<0.05.
Figure9.Meanendometrialedemascore(+/-SE)of9PBIE-susceptiblemaresinN-acetylcysteinetreated(TX)andsaline(CONTROL)cycleswhenthetimepoints12,36,60and84hourspostAIwere combined within each treatment cycle. Maximum edema score possible was 4. Nosignificanteffectoftreatmentwasobserved(p=0.90).
0
5
10
15
20
25
30
35
40
12HOURS 36HOURS 60HOURS 84HOURS
IUF(cm
3 )p=0.005
A A
A
B
0
0.5
1
1.5
2
2.5
TX CONTROL
Endo
metria
lede
mascore
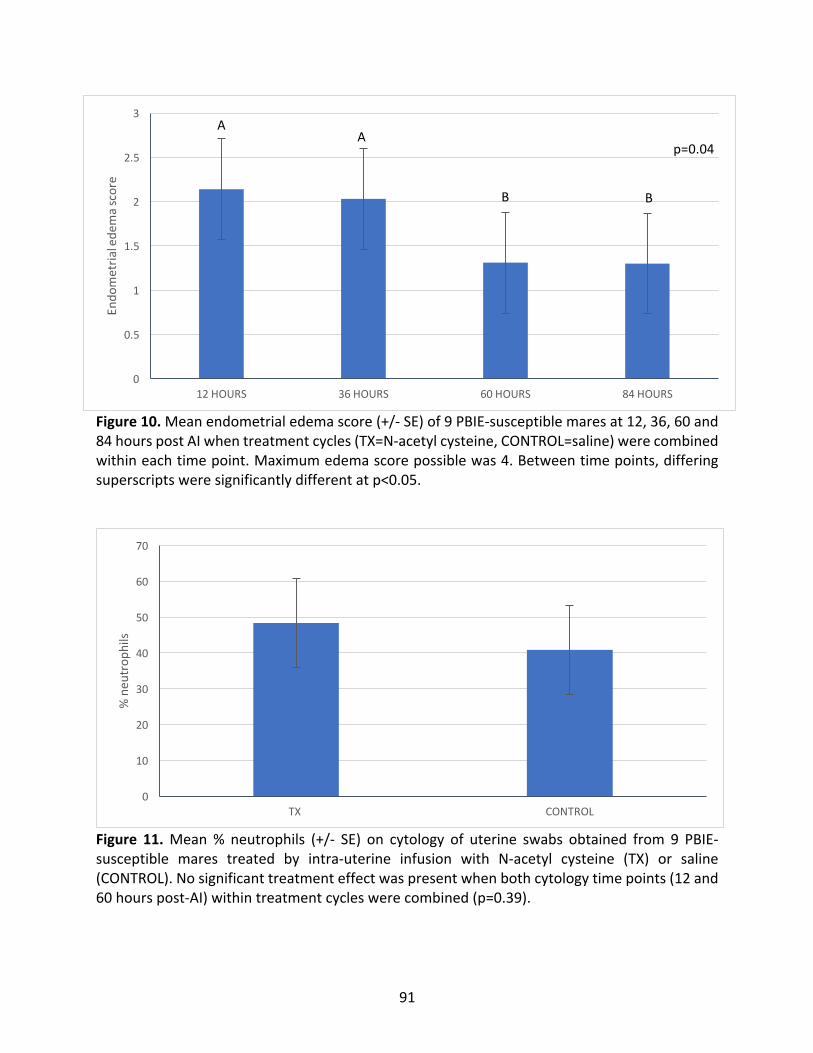
91
Figure10.Meanendometrialedemascore(+/-SE)of9PBIE-susceptiblemaresat12,36,60and84hourspostAIwhentreatmentcycles(TX=N-acetylcysteine,CONTROL=saline)werecombinedwithineachtimepoint.Maximumedemascorepossiblewas4.Betweentimepoints,differingsuperscriptsweresignificantlydifferentatp<0.05.
Figure 11.Mean% neutrophils (+/- SE) on cytology of uterine swabs obtained from 9 PBIE-susceptible mares treated by intra-uterine infusion with N-acetyl cysteine (TX) or saline(CONTROL).Nosignificanttreatmenteffectwaspresentwhenbothcytologytimepoints(12and60hourspost-AI)withintreatmentcycleswerecombined(p=0.39).
0
0.5
1
1.5
2
2.5
3
12HOURS 36HOURS 60HOURS 84HOURS
Endo
metria
lede
mascore
AA
B B
p=0.04
0
10
20
30
40
50
60
70
TX CONTROL
%neu
trop
hils
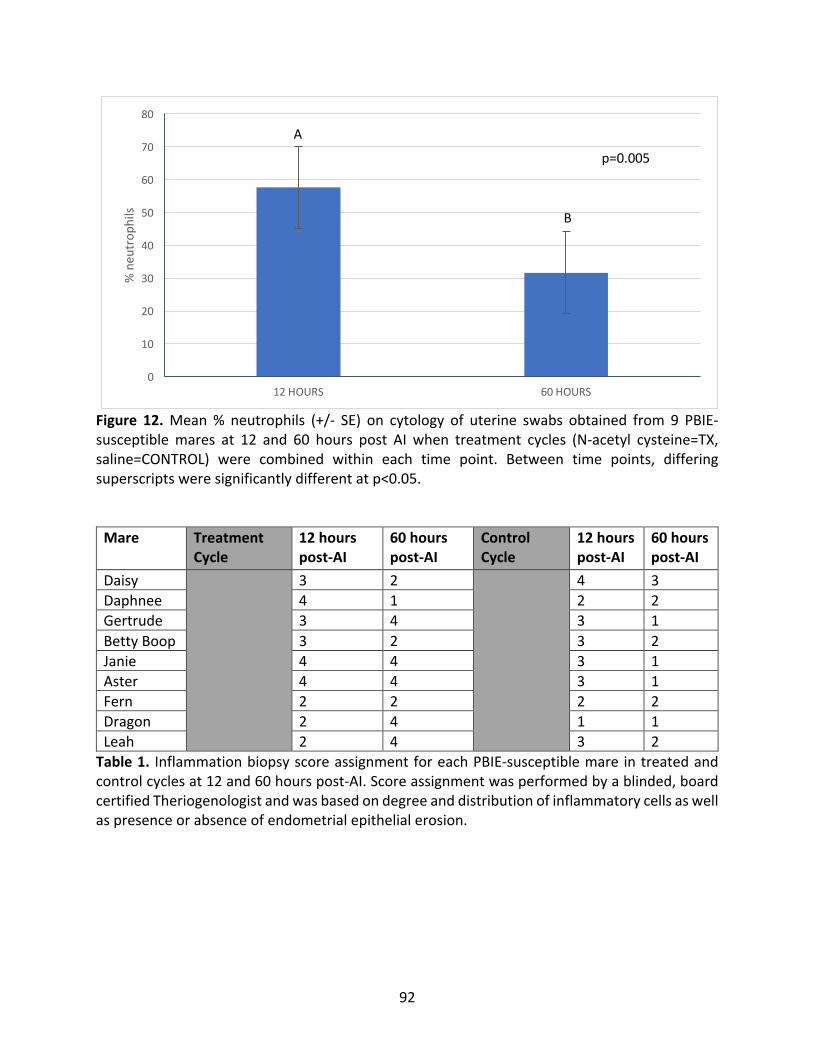
92
Figure 12.Mean% neutrophils (+/- SE) on cytology of uterine swabs obtained from 9 PBIE-susceptiblemares at 12 and 60 hours post AI when treatment cycles (N-acetyl cysteine=TX,saline=CONTROL) were combined within each time point. Between time points, differingsuperscriptsweresignificantlydifferentatp<0.05.Mare Treatment
Cycle12hourspost-AI
60hourspost-AI
ControlCycle
12hourspost-AI
60hourspost-AI
Daisy 3 2 4 3Daphnee 4 1 2 2Gertrude 3 4 3 1BettyBoop 3 2 3 2Janie 4 4 3 1Aster 4 4 3 1Fern 2 2 2 2Dragon 2 4 1 1Leah 2 4 3 2Table1. InflammationbiopsyscoreassignmentforeachPBIE-susceptiblemare intreatedandcontrolcyclesat12and60hourspost-AI.Scoreassignmentwasperformedbyablinded,boardcertifiedTheriogenologistandwasbasedondegreeanddistributionofinflammatorycellsaswellaspresenceorabsenceofendometrialepithelialerosion.
0
10
20
30
40
50
60
70
80
12HOURS 60HOURS
%neu
trop
hils
A
B
p=0.005

93
CHAPTERTHREE:THEEFFECTOFTREATMENTOFPBIESUSCEPTIBLEMARESWITHN-ACETYLCYSTEINEONNITRICOXIDELEVELSAND
INFLAMMATORYCYTOKINEPROFILES3.1ABSTRACT
Persistentbreeding-inducedendometritis(PBIE)isamajorcauseofinfertilityinmares.
Transient uterine inflammation is a normal response to breeding, however, PBIE susceptible
maresdonotclearthis inflammation inatimelyfashion.Uterine inflammationatthetimeof
embryonicdescent fromtheoviductsultimately results inearlyembryonicdeathand is seen
clinicallyasinfertility.N-acetylcysteine(NAC),amucolyticusedtotreatendometritisinmares,
hasanti-inflammatoryproperties,affectsinflammatorycytokinespost-treatmentandisamild
inhibitor of nitric oxide synthase. Increased nitric oxide (NO) levels and alterations in
inflammatorycytokineshavepreviouslybeendocumentedinPBIEmares.Theobjectiveofthis
studywastodetermineifNACtreatmentofPBIE-susceptiblemareswouldlowernitricoxideand
inflammatory cytokine levels. A randomized, blinded, cross-over design clinical trial was
performedutilizingPBIEsusceptiblemares (n=9).Mareswerescreenednegative forbacterial
endometritispriortoenrolment inthestudy. Intrauterineinfusionof180mlof3.3%NACwas
performed12hourspriortoinsemination,whenafollicle>35mmwaspresent.Interleukin-6
concentration (ELISA) andnitric oxide level (Colorimetric assay) in intra-uterine fluid samples
were determined at 12 and 60 hours post-insemination. Gene expression of inflammatory
cytokines (Interleukin-6, Interleukin-10, Interleukin-1β and Tumor Necrosis Factor-α) were
determined by qPCR from endometrial biopsy samples taken at 12 and 60 hours post-
insemination. Statistical analysis was performed using a repeated measures ANOVA. In this

94
clinical trial, pre-breeding intrauterine treatment with NAC did not affect NO levels, IL-6
concentrationorcytokinegeneexpression.Differencesacrosstimepointspost-AIwerenoted
forIL-6geneexpression,whichwashighestat12hourspost-AI(p=0.003).Expressionofother
cytokinesevaluateddidnotchangeovertime.WhiletreatmentwithNACpre-breedingdidnot
haveaneffectonthecytokinesevaluated,theassessmentofinflammatorycytokinesbothatthe
gene and protein level at different time points post artificial insemination will add to the
understandingofthealterationsininflammatorycytokinelevelsinmaressusceptibletoPBIE.
3.2INTRODUCTION
Persistentbreedinginducedendometritis(PBIE)inmaresischaracterizedbyaprolonged
inflammatory response following breeding, ultimately leading to early embryonic loss and
infertility.Anormalandtransient inflammatoryresponseoccurspost-breeding;however, ina
subset of susceptible mares, this inflammation persists (Troedsson, 2006; Woodward and
Troedsson2015;ChristoffersenandTroedsson,2017).SusceptibilitytoPBIEisassociatedwithan
altered inflammatory response as well as poor mechanical clearance of the uterus
(NikolakopoulosandWatson1999;Watson2000;Troedsson2006; LeBlancandCausey2009;
WoodwardandTroedsson2015;ChristoffersenandTroedsson,2017;Khanetal.2018).
Theinflammatoryresponseoftheendometriumcanbecharacterizedbyalterations in
cytokines.Baselinelevelsofpro-inflammatorycytokinesaredifferentbetweenPBIE-susceptible
versus resistant mares. Specifically, during estrus, PBIE-susceptible mares have elevated
endometrial mRNA expression of IL-1β, TNF-α, IL-6 and IL-8 compared to resistant mares
(Fumosoetal.2003;Fumosoetal.2007).Additionally,endometrialmRNAexpressionofthe

95
anti-inflammatorycytokineIL-10islowerduringestrusinPBIE-susceptiblemares,andhigherin
diestrus, compared to normal mares (Fumoso et al. 2007). Differences in the endometrial
inflammatoryresponsebetweennormalandPBIE-susceptiblemaresarealsoevidentfollowing
artificialinsemination(AI).ExpressionofIL-1β,TNF-α,IL-8andIL-10mRNAiselevatedsevendays
post-AIinPBIE-susceptiblemarescomparedtoresistantmares(Fumosoetal.2003;Fumosoet
al. 2007). Other differences include reduced IL-6, IL-1RN and IL-10 in susceptible mares
comparedtoresistantmares6hourspostAI(Woodwardetal.2013a).
Treatment approaches used in the management of mares with PBIE include ecbolic
therapy such as oxytocin, to enhance uterine clearance, and immunomodulators such as
dexamethasoneandmycobacterialcellwallextract(MCWE)tosuppressinflammation(LeBlanc
2010;WoodwardandTroedsson2013).TheeffectofimmunomodulationonmRNAexpression
ofinflammatorycytokineshasbeenexploredinbothsusceptibleandresistantmares(Fumosoet
al.2003;Fumosoetal.2007;Woodwardetal.2015).Itappearsthatcorrectionofthealtered
inflammatoryresponseassociatedwithsusceptibilitytoPBIEmaybeaneffectiveapproachto
treatment.
In addition to an altered inflammatory response post-breeding, researchers have
demonstratedthatelevatedlevelsofnitricoxide(NO),asmoothmusclerelaxant,maycontribute
to thedecreasedmechanical clearanceof theuterus reported inPBIEmares (Alghamdietal.
2005;Woodwardetal.2013b;Khanetal.2017).Induciblenitricoxidesynthase(iNOS)isthe
enzyme responsible for theproductionof inducedNO in theequineuterus. PBIE susceptible
mareshadelevatedNOlevelsintheiruterinesecretionsandincreasediNOSmRNAexpressionat
13hoursfollowingAI(Alghamdietal.2005).Adose-dependentinhibitionofNOonmyometrial

96
contractilitywasdemonstratedinvitro(Khanetal.2017),suggestingthatalteredNOlevelsin
PBIE-susceptiblemaresplayaroleinreducedmyometrialcontractilityanduterineclearance.
Given what is currently understood about the pathophysiology of PBIE, use of a
compoundexhibitingbothanti-inflammatoryandNO-suppressiveeffectsshouldbeaneffective
approachtotreatment.N-acetylcysteine(NAC)isamucolyticagentusedintreatmentofequine
bacterialendometritiswhereabiofilmissuspected,orincaseswhereexcessuterinemucusis
present (LeBlanc andCausey 2009;Gores-Lindholmet al. 2009; LeBlanc 2010). Intra-uterine
treatmentwitha3.3%NACsolutioninmareswithahistoryofendometritis,eitherduringthe
previouscycleor48hourspriortobreeding,leadstohigherthananticipatedfertilityrates(85%)
(Gores-Lindholmetal.2009;LeBlanc2010).N-acetylcysteine(NAC)reducesmucusviscosityby
disrupting disulphide bonds (LeBlanc 2010), and hasmild anti-oxidant and anti-inflammatory
effectsinmultiplemodelsofdisease(Stanislausetal.2005;Cazzolaetal.2017).Inhorses,NAC
inhibitedserum-derivedneutrophiloxidativeburstwheninfusedasa3%solutionanddidnot
haveanadverseeffectontheendometriumofeitherfertileorbarrenmares(Gores-Lindholmet
al.2013).Intra-uterineandoraladministrationofNACinmaresledtodecreasedneutrophiland
COX-2 staining on histologic endometrial samples from normal mares, confirming an anti-
inflammatoryeffect(Witteetal.2012;Melkusetal.2013).However,nostudiestodatehave
examinedeffectsofintrauterineinfusionofNAConNOlevels,cytokinemRNAexpression,oron
cytokineconcentrationinuterinefluid.
Theobjectiveofthisstudywastodetermineifintra-uterineinfusionofPBIE-susceptible
mareswitha3.3%NACsolutioninsalinewoulddecreaseNOlevelsandaltertheendometrial
cytokineprofilecomparedtocontrolcycles.

97
3.3MATERIALSANDMETHODS
AlltheproceduresperformedonanimalsinthisstudywereapprovedbytheUniversity
ofGuelphAnimalCareCommittee(AnimalUtilizationProtocolnumber3213),inaccordancewith
theCanadianCouncilonAnimalCareguidelines.
3.3.1MareSelection,StudyDesignandSampling
Arandomized,blinded,crossoverdesignedclinicaltrialwasperformedwithawashout
period(minimumoneestrouscycle)betweenexperimentalcycles.Theprimaryinvestigator,who
performedallclinicalevaluations,wasblindedtomareassignment(treatedversuscontrolcycles)
duringdatacollection.Inordertorandomizetheorderoftreatmentandcontrolcycles,mares
wereassignedanumber,withthosenumberssubsequentlyrandomlysortedintotreatedand
controlcycles.
MaressusceptibletoPBIE(n=10)wereselectedforinclusionintheclinicaltrialbasedon
an endometrial biopsy score of IIb or III, normal cytology and negative bacterial culture on
endometrial swab, and persistent accumulation of intra-uterine fluid 96 hours following
challengeby inseminationwithkilledsemen(Alghamdietal. 2005;Woodwardetal. 2013a;
Woodward et al. 2013b; Woodward et al. 2015). Mares were housed at the Arkell Equine
ResearchStationattheUniversityofGuelphforthedurationofthestudyandwereofvarious
breeds(Standardbredandmixedbreed)andages(7-19years).
Mares weremonitored for estrus by trans-rectal palpation and ultrasound.Mares in
diestrus,determinedbypresenceofacorpusluteum(CL),weregivenanintra-muscularinjection
ofcloprostenol(125mcg)(Estrumate®,IntervetCanadaCorp.,Kirkland,QC,Canada)toinduce
estrus.Onceafollicle>35mmwasidentifiedduringestrus,maresweregiven2,500IUofhuman

98
chorionic gonadotropin (hCG, Chorulon®, Intervet Canada Corp., Kirkland, QC, Canada)
intravenously,toinduceovulation.Usinga21-inchinseminationpipette(PartnarAnimalHealth,
Ilderton,ON,Canada),maresweretrans-cervically infusedwitheithera3.3%solutionofNAC
consisting of 30ml of N-acetyl cysteine (Acetylcysteine Injection 200mg/ml, Alveda
Pharmaceuticals Inc., Toronto, ON, Canada) added to 150ml of 0.9% sodium chloride (0.9%
SodiumChlorideInjectionUSP,BaxterCorporation,Mississauga,ON,Canada)(treatedcycle)or
180ml of 0.9% sodium chloride (0.9% Sodium Chloride Injection USP, Baxter Corporation,
Mississauga, ON, Canada) (control cycle). Twelve hours following infusion, the mares were
inseminatedwith1x109killedspermatozoain30mlofsemenextender,aspreviouslydescribed
(Woodwardetal.2013a;Woodwardetal.2013b;Woodwardetal.2015;Khanetal.2018).
Trans-rectalultrasonographywasperformedbeginning twelvehoursafterAI,andwas
repeatedevery24hoursthereafteruntil84hourspost-AI(12,36,60and84hours),toevaluate
endometrial edema grade (0-4; 0 = no edema; 4 = pathologic edema) and endometrial fluid
volume.FluidvolumewascalculatedusingtheformulaV=length*width*depth,eachmeasured
incm.Multiplelocationsweremeasuredintheuterinebodyandhorns,andanaverageof2-5
locationswereused in the calculation. Additional samples takenat12and60hourspost-AI
consistedofanendometrial swab forcytology,collectionofuterine fluidandanendometrial
biopsy. Cytology samples were aseptically obtained using a guarded uterine swab (Kalayjian
IndustriesInc.,SignalHill,CA,USA)aspreviouslydescribedbyKhanetal.(2018).Endometrial
biopsy specimens were procured in an aseptic manner using a Pilling-Weck biopsy punch
(JorgensenLaboratories,Loveland,CO,USA),asdescribedbyKenney(1978).Theendometrial
biopsy sample was immediately placed in 655µL of RNAlater (Sigma-Aldrich®, Oakville, ON,

99
Canada)andstoredovernightat4°C.Thefollowingmorning,itwasremovedfromtheRNAlater
andstoredat-80°Cuntilanalysis,asperthemanufacturer’sinstructions.
3.3.2Real-TimeQuantitativePCR
A pre-trial endometrial biopsy samplewas obtained from a subset ofmares (n=4), in
additiontoendometrialbiopsiesobtainedat12and60hourspost-AIforallmares.TotalRNA
fromendometrialtissuewasisolatedwiththeQiagenRNeasy™kit(©QIAGEN,Toronto,CAN),
accordingtothemanufacturer'sprotocol.FirststrandcDNAwassynthesizedfrom200ngofRNA
with50μmoligoprimerandreversetranscriptaseenzyme.ThereferencegenesSDHA,β-actin,
EeF2,RPS23andGAPDHwereamplifiedfromdifferentconcentrationsoftemplatecDNA.Most
reference genes were found to be inconsistent among mares, therefore, a specific set of
referencegeneswasoptimizedforeachmareandusedtodeterminetherelativeexpressionof
thegeneof interest foreachmare (Table1).Reverse transcriptionofmRNAwas initiated to
inactivate the reverse transcriptase and simultaneously activate the thermostable DNA
polymerase.cDNAwasamplifiedintriplicatebyqRT-PCRinaLightCycler480instrument(Roche
LifeScience,Laval,PQ)usingSYBRGreen(Roche)andprimerstoequineIL-1β, IL-6,IL-10,and
TNF-α (Table 2). Cycle threshold (CT) valueswere calculated and normalized to eachmare’s
specificreferencegenepair(calibratornormalizedrelativequantification).
3.3.3Intra-UterineFluidCollection
Intra-uterinefluidwascollectedinoneoftwoways.Inmareswithlow-to-moderatefluid
volume,aBivonacatheter(PartnarAnimalHealth)waspassedasepticallythroughthecervixand
into themare’suterus. After inflating the cuff, ahandheld suctionpump (NalgeneTM hand-
operatedvacuumpump,Sigma-Aldrich®Oakville,ON,Canada)wasattachedtothecatheterand

100
anassistantappliedexternalsuctionuntilfluidwasobtainedinthetube.Inmareswithalarge
volumeof intra-uterine fluid, a samplewas readilyobtainedby free-flowduringendometrial
swabsampling,throughtheouterplasticguard.Thefluidsamplewasimmediatelyplacedina
polypropylene centrifuge tube (Corning® Incorporated, Corning, NY, USA) and held at room
temperatureuntilprocessing.Oncereturnedtothelaboratory,thefluidwascentrifugedat1500
x g for 12minutes to remove cellular debris. The supernatant was aspirated, placed in 1ml
aliquots (Fisherbrand®, Cryogenic Vial 1.2mL, Fisher Scientific, Markham, ON, Canada) and
storedat-20°Cforfurtheranalyses.
3.3.4ELISADeterminationofInterleukin-6ConcentrationinUterineFluid
Intrauterine IL-6 concentration was determined using the Equine IL-6 DuoSet®ELISA
(R&DSystems™,Minneapolis,MN,USA)accordingtomanufacturerinstructions.Sampleswere
diluted1:20inreagentdiluent(1%BSAinPBS)andruninduplicate.Theintra-assaycoefficient
ofvariationwas7.4%andtheinter-assaycoefficientofvariationwas10.9%.Thekitwasvalidated
forequine intrauterine fluidbydeterminationofpercent recoveryand linearity,according to
R&DSystems™Spike,Recovery&LinearityProtocol.Thedetailedvalidationprocesscanbefound
inAppendix2.
3.3.5NitricOxideConcentration
Nitricoxideisunstableandhasashorthalf-lifeinvivo,necessitatingthemeasurementof
its metabolites, nitrate and nitrite, as alternatives. The concentration of nitrate and nitrite
presentinintrauterinefluidwasmeasuredtoprovideanestimateofNOconcentration,usingthe
Nitrate/NitriteColorimetricAssayKit(CaymanChemicalCo.,AnnHarbor,MI,USA),accordingto

101
the manufacturer’s instructions. Samples were diluted 1:5 with buffer solution, and run in
duplicate.
3.3.6StatisticalAnalyses
Statistical analyseswereperformedusing the SAS softwareprogram (version9.3, SAS
Institute,Inc.,Cary,NC,USA).OnlyestrouscyclesfittingthedefinitionofPBIEwereincludedin
theanalysis(n=9salinecontrol;n=9NACtreatmentcycles).Differences inendometrialmRNA
expressionofIL-6,IL-10,TNF-αandIL-1β,aswellasIL-6andNOconcentrationsinuterinefluid,
werecomparedbetweencontrolandtreatedcyclesovertime(12hand60hpost-AI)byrepeated
measuresANOVA.TheAkaikeInformationCriterion(AIC)wasutilizedtodeterminethebest-fit
model.TheShapiro-Wilktestdeterminedthatthedatawasnotnormallydistributedandrequired
logarithmictransformationpriortoanalysis,thereforeresultsarepresentedasmedians(back
transformed)+/-upperandlowerconfidencelimits.
Interactioneffectsbetweentreatmentandtimewereexaminedinthemodel,andthep
valuewas set at <0.05 for statistical significance.When therewas no statistically significant
treatment-by-time interaction seen, thedata fromboth treatmentgroupswere combined to
examinetheeffectoftimepost-AI,anddatafrombothtimepointswerecombinedwithineach
treatmentgrouptoevaluateforanymaineffectsoftreatment.
Forthesubsetofmareswithbaselinebiopsiesavailable(T=0;n=4),differencesinIL-6,IL-
10,TNF-αandIL-1βmRNAexpressionwerealsocomparedbetweenbaseline,12hoursand60
hourspost-AIusingrepeatedmeasuresANOVA.

102
3.4 RESULTS
3.4.1EffectofIntra-UterineTreatmentof3.3%NACinPBIESusceptibleMares
When both time points were combined within each treatment group, intrauterine
infusionwith3.3%NAC12hourspriortoAIdidnotaffectendometrialmRNAexpressionofIL-6
(p=0.86)(Appendix3:Figure1),IL-10(p=0.58)(Appendix3:Figure2),TNF-α(p=0.92)(Appendix
3:Figure3),orIL-1β(p=0.65)(Appendix3:Figure4)comparedtountreatedcycles(Table3).NAC
infusion had no effect on the concentration of IL-6 in uterine fluid as determined by ELISA
(p=0.083)(Figure1),nordiditaffecttheamountofNOinuterinefluid(p=0.46)(Figure2).
3.4.2ChangesinEndometrialInflammationinResponsetoAIOverTime
a) mRNAexpression–12versus60hours(n=9mares)
EndometrialIL-6mRNAexpressionwassignificantlygreaterat12hourspostAIcompared
to60hourspostAI,whenbothtreatmentgroupswerecombinedwithineachtimepoint(p=0.02)
(Figure3).Inthecontrolcycle,IL-6mRNAexpressionat12hourspostAIwassignificantlygreater
thanat60hourspostAI(p=0.02)(Appendix3:Figure5).EndometrialmRNAexpressiondidnot
changefrom12to60hoursforIL-10(p=0.73)(Appendix3;Figure6),TNF-α(p=0.86)(Appendix
3:Figure7)andIL-1β(p=0.6)(Appendix3:Figure8),whenbothtreatmentgroupswerecombined
withinineachtimepoint(Table4).
b) mRNAexpression–baselineversus12and60hourspost-AI(n=4)(Table5)
Forthesubsetoffourmareswithbaselinebiopsies,therewasanoverallsignificantmain
effectoftimeonIL-6mRNAexpression(p=0.0004)(Figure4)aswellasanoverallsignificanteffect
oftimeonIL-1βmRNAexpression(p=0.0030)(Figure5).Incontrast,mRNAexpressionofIL-10

103
(p=0.1779)(Figure6)andTNF-α(p=0.0744)(Figure7)didnotchangeovertime.Nosignificant
treatmentmaineffectwasseenforthesubsetofmareswithbaselinebiopsies.
3.4.3IL-6ConcentrationasMeasuredbyELISA
IL-6concentration in intrauterinefluid (ELISA)didnotchangeovertimepostAI,when
bothtreatmentgroupswerecombinedwithinineachtimepoint(p=0.42)(Figure8).Therewas
notimebytreatmentinteraction.
3.4.4Nitricoxide
AlthoughNOincreasedfrom12to60hourspost-AI,thechangeinNOconcentrationover
timedidnotreachsignificance,whenbothtreatmentgroupswerecombinedwithinineachtime
point(p=0.056)(Figure9).Therewasnotimebytreatmentinteraction.
3.5DISCUSSION
Theanti-inflammatoryeffectsofNAC,includingcytokinemodulation,havebeenshown
inmultiplemodels of animal andhumandisease. In a human chronic obstructivepulmonary
disease (COPD) in vitro study, NAC inhibited pro-inflammatory cytokines (IL-1β, IL-8, TNF-α)
(Cazzolaetal. 2017).Similarly, inamultiplesclerosis (MS)study, treatmentwithNAC led to
inhibitionofmonocyte/macrophageinducedTNF-α,IFN-γ,IL-1βproductionandstimulatedIL-10
production(Stanislausetal.2005).However,theresultsofthecurrentstudysuggestthatthe
anti-inflammatoryeffectsofintra-uterineinfusionofa3.3%NACsolutionweretoomildtoresult
inalterationsofcytokineexpression.Bycomparison,otherimmunomodulatorytherapieshave
demonstratedthepotentialtoaltertheinflammatorycytokineprofileofPBIE-susceptiblemares.
Some treatment modalities (lactoferrin, Mycobacterium Cell Wall Extract (MCWE) and

104
dexamethasone)lowerpro-inflammatorycytokinemRNAexpression,suchasTNF-α(Fedorkaet
al.2016)andIL-1β(Fumosoetal.2003;Woodwardetal.2015)andincreasemRNAexpression
of IL-10,ananti-inflammatorycytokine(Fumosoetal. 2007)to levelsmoretypicalofnormal
mares.However,manyoftheothercytokinesevaluatedinthesestudies(IL-6,IL-1RN,IL-8)were
unaffectedby these treatmentmethods. It is possible that a treatment effect followingNAC
infusionwasnotapparentduetothechoiceofcytokinesevaluatedinthecurrentstudy;further
studiesexaminingtheeffectsonotherendometrialcytokinesmayelicitadditionalinformation.
OtherpossiblereasonsforthelackoftreatmenteffectofNACincludetheselectionoftimepoints
forsampling,differencesinthedefinitionofPBIEmaresbetweenstudiesanddifferencesinqPCR
methodology,includingtheselectionofreferencegenes.
Methodologies used in qPCR differ across studies and may account for variability in
endometrialcytokinemRNAexpression.Specifically,thechoiceofreferencegene(s)appearsto
vary.Inmoststudies,onlyonereferencegeneisselectedtodeterminerelativegeneexpression,
andlimitedreportingofthevalidationprocessisgiven(Fumosoetal.2003;Fumosoetal.2007;
Christoffersenetal.2012a;Christoffersenetal.2012b;Woodwardetal.2013a;Woodwardet
al.2013b;Woodwardetal.2015;Fedorkaetal.2016;Fedorkaetal.2017;Fedorkaetal.2018).
Recently,ithasbeensuggestedthatmorethanonereferencegeneshouldbeselected,inorder
toincreasethevalidityoftheanalysis(Bustinetal.2009).Inaddition,referencegenespreviously
believedtobestablehavesincebeenshowntohavealargedegreeofvariationacrossstudies
(Kozeraet al. 2013). Therefore, validation of chosen reference genes specific to the species,
experimentaldesign,andtissueofinterestwouldimproverepeatabilityacrossstudies(Jacobet
al.2013;Kozeraetal.2013;Bustinetal.2017).Significantvariabilityisknowntooccurinequine

105
geneexpressionacrossdifferenttissues(Tessieretal.2017).Inthepresentstudy,twostable
referencegeneswereselectedandvalidatedforeachmare,andsevenoftheninemaresshared
atleastonegenewithinthereferencegenepairs(EeF2)(Table1).GiventhatcytokinemRNA
expressionismeasuredrelativetothatofselectedreferencegenes,itisthereforepossiblethat
someofthedifferencesnotedacrossstudieswithrespecttorelativecytokinegeneexpressionin
equineendometritis,areduetothedifferencesinresearchmethodologiesandtheselectionof
referencegenesbetweenstudies.
Intra-uterineinfusionwithNACdidnotalterNOlevelspost-AIinPBIE-susceptiblemares.
Elevated intra-uterine NO levels in PBIE susceptible mares are suspected to play a role in
decreaseduterinecontractilityanduterineclearance(Alghamdietal. 2005;Woodwardetal.
2013b;Khanetal.2017;Khanetal.2018).Alghamdietal.(2005)showedthatPBIEsusceptible
mareshadhigherNOintheiruterinesecretionsandhigheriNOSmRNAexpression13hourspost-
AIcomparedtoresistantmares.Inaddition,Woodwardetal.(2013b)showedthatsusceptible
mareshadelevatedmRNAexpressionofiNOSat2and6hourspost-AI.Nitricoxideproduction
andiNOSsynthesiswerereducedbytreatmentwithNACinratin-vitrostudies,murinepre-term
laborstudies,andmultiplesclerosisstudies(Pahanetal.1997;Bergaminietal.2001;Majano
etal.2004;Stanislausetal.2005;Paintliaetal.2008).Mycobacteriumcellwallextract(MCWE),
animmunomodulator,givenIV24hourspriortoAIdecreaseduterineNOinsusceptiblemares.
Inthesamestudy,administrationof50mgofdexamethasoneIV24hourspriortoAI,didnot
decreaseuterineNOconcentrations inPBIE-susceptiblemares (Woodwardetal. 2013b). In-
vitro treatment of endometrial tissue sampleswith 1400W, a highly specific iNOS inhibitor,
resultedindecreasedendometrialNOSactivityinbothsusceptibleandresistantmares(Khanet

106
al. 2018). ItappearsthatNACisaverymild iNOSinhibitor,andthedoseandtimingofNAC
administrationinthisstudymayexplainthelackofeffectsseen.
The preparation and dose of 3.3% NAC intra-uterine infusion was chosen for several
reasons.ThisdosewasfirstdescribedbyLeBlancandCausey(2009)asbeingmucolyticandable
todislodgeinspissateduterinesecretionsinmareswithahistoryofinfertility.Theeffectsofa
3.3%NACintra-uterineinfusionwerefurthercharacterizedbyGores-Lindolhmetal.(2009)and
LeBlanc (2010), who showed improved pregnancy rates (85%) in mares with a history of
endometritis.Further,Gores-Lindholmetal.(2013)demonstratedthattreatmentwithNACat
this dose led to decreased uterine mucus thickness, and had no adverse effects on the
endometrium in barren and healthymareswith Kenney biopsy scores ranging from I to IIb.
Pregnancy rates of 77% were attained when using this dose combined with uterine lavage,
oxytocintreatmentandas-neededintra-uterineantibiotics.Sincethattime,3.3%NAChasbeen
recommendedforcasesofmucushypersecretionorinmareswithsuspectedbiofilm(Lyle2012;
Troedsson2012;Buczkowskaetal.2015;Scoggin2016).Recently,aninvitrostudydemonstrated
that3.3%NACdisruptedE.colibiofilmandreadilykilledE.coliwithinthebiofilm(Ferrisetal.
2016).However,itispossiblethatthedosechosenwastoolowtoaffectcytokineexpressionand
NOconcentrations. Tirouvanziametal. (2006) showed thatonlyhighdosesofNACaffected
cytokine concentration in a human cystic fibrosis study. In addition, NO release from rat
peritoneal macrophages was significantly reduced when higher doses of NAC were utilized
(Pahanetal. 1997). Inmostofthesestudies,NACwasadministeredpre-breedingorpriorto
inductionof inflammation, justifying the timingofadministration inour study.However, it is
possible that treatment during an inflammatory responsemay bemore beneficial than pre-

107
treatment.Infact,Tirouvanziametal.(2006)showedclinicalimprovementinalreadyillpatients
treatedwithoralNAC.
Interleukin-6wastheonlycytokineshowingsignificantdifferencesinrelativeexpression
across timepoints. Increasedexpressionof IL-6at12hourscomparedto60hourspost-AI, is
similartothefindingsofWoodwardetal.(2013a),whoshowedelevatedIL-6mRNAexpression
between2and12hourspost–AI,andFumosoetal.(2003)whoshowedhigherexpressionofIL-
6mRNAat24hourscomparedto7dayspost-AI.Whilethesetimelinesarenotidenticaltothose
choseninourstudy,thetrendsnotedbetweenstudiesaresimilarinthatIL-6mRNAexpression
iselevatedattimepointsnearestAIanddecreasesovertime.
ThelackofdifferencesobservedforendometrialIL-10,TNF-αandIL-1βmRNAexpression
between12and60hourspost-AIisincontrasttootherstudies,wheresomeofthesecytokines
changedintheirexpressionovertime.Interleukin-10waselevatedinsusceptiblemaresat2and
24hours,andIL-1βmRNAexpressionwasincreasedat2and6hourscomparedtobaselinelevels,
followingAI(Woodwardetal.2013a).Interleukin-10mRNAexpressioninsusceptiblemareswas
lowerat24hourscomparedto7dayspost-AI(Fumosoetal.2007).Interestingly,thesestudies
alsoshowednochangesforothercytokines.NochangesinTNF-αmRNAexpressionwerenoted
inWoodwardetal.(2013a),nordidIL-1βmRNAexpressionchangebetween24hoursand7days
post-AI(Fumosoetal.2003).
Baseline cytokine mRNA expression is reportedly different in PBIE susceptible mares
comparedtonormalmares(Fumosoetal.2003;Fumosoetal.2007).Comparisonstocytokine
mRNAexpressionatbaselinehavealsobeenmadeinPBIEsusceptiblemaresfollowingAI,with
IL-1β, IL-6, IL-8, IL-10and IFN-γbeingelevated frombaselineatvarious timepoints following

108
insemination (Woodward et al. 2013a). In our study, although only a subset of mares had
baselinebiopsiestakenforqPCRanalysis,baselinemRNAexpressionwaslowerforallcytokines
tested,andincreasedat12hoursfollowingAI.Thiselevationincytokineexpressiondecreased
by60hourspost-AI,butdidnotreturntobaselinelevels.ThechangesobservedinIL-6andIL-1β
mRNAexpressionovertimearereflectiveoftheinflammatoryresponsetoAI.However,thelack
of significant changes across time for IL-10 and TNF-α mRNA expression from baseline was
unexpected.TumorNecrosisFactor-αispro-inflammatoryandwouldbeexpectedtorisenear
thetimeofAItoevokeendometrialinflammation.Interleukin-10isanti-inflammatoryandwould
beexpectedtorisesometimeafterAI,tomediatetheinflammatoryresponse.Ourresultsare
similartoWoodwardetal.(2013a)whofoundnodifferencesinTNF-αfollowingAIinsusceptible
mares and amuch lower expressionof IL-10 in susceptiblemares in comparison to resistant
mares.IL-10andTNF-αmayplaylessofaroleinPBIEthanpreviouslysuspected.
Despite an increase in IL-6 mRNA expression 12 hours post-AI, intra-uterine IL-6
concentrationasmeasuredbyELISA,wasnotsignificantlydifferentacrosstimepointspost-AIin
this study. However, IL-6 concentration was greatest at 12 hours post-AI, even though not
statisticallysignificant,demonstratingthatIL-6concentrationfollowsthesamepatternasmRNA
expression.Inthepresentstudy,considerablevariabilityinIL-6concentrationacrossmaresmade
detectionofdifferencesdifficult,and the lackof significancecouldbeattributed to the large
variabilityinourdata.
Inconclusion,treatmentwith3.3%NACasanintrauterineinfusion12hourspriortoAI
didnotaffectmRNAexpressionofIL-6,IL-10,TNF-αandIL-1β,nordiditaffectintra-uterineIL-6
concentration and NO levels, in PBIE-susceptible mares. In general, when comparing mRNA

109
expressiontobaselinevaluesforthesubsetofmareswithpre-trialbiopsies,itappearsthatAIin
itselfcausesanincreaseinmRNAexpressionacrosstimepointspost-AI.Subsequentdifferences
forallmaresbetween12and60hourspost-AIarelessevident,notablyforIL-10,TNF-αandIL-
1β,butareclearforIL-6. IL-6mRNAexpressionisgreatestat12hourscomparedto60hours
post-AI,implicatingitaslikelyplayingaroleinthemanifestationofPBIEinsusceptiblemares.
FuturestudiesshouldbeaimedatcharacterizingtheinflammatorycytokinemRNAexpression
patternpost-AIandhowitdiffersfromnormalinPBIEsusceptiblemares.Fromthere,whatis
knownoftheeffectofdifferenttreatmentmodalitiesonthesecytokinesatspecifictimepoints
couldbeutilizedaccordinglyintreatingPBIEsusceptiblemares.

110
3.6REFERENCES1. Alghamdi, A.S. and Foster, D.N. (2005). Seminal DNase frees spermatozoa entangled in
neutrophilextracellulartraps.Biol.Reprod.73,1174-1181.
2. Bergamini,S.,Rota,C.,Canali,R.,Staffieri,M.,Daneri,F.,Bini,A.,Giovannini,F,Tomasi,A.andIannone,A.(2001).N-acetylcysteineinhibitsinvivonitricoxideproductionbyinduciblenitricoxidesynthase.NitricOxide.5,349-360.
3. Buczkowska, J., Kozdrowski, R., Sikora, M., Dzieciol, M. and Matusz, A. (2015). Non-
traditionaltreatmentsforendometritisinmares.Bulg.J.Vet.Med.18,285-293.4. Bustin S. andNolan T. (2017). Talking the talk, but notwalking thewalk: RT-qPCR as a
paradigmforthelackofreproducibilityinmolecularresearch.Eur.J.Clin.Invest.47,756-774.
5. Bustin, S.A., Benes, V., Garson, J.A., Hellemans, J., Huggett, J., Kubista,M.,Mueller, R.,
Nolan,T.,Pfaffl,M.W.,Shipley,G.L.,Vandesompele,J.andWittwer,C.T.(2009).TheMIQEguidelines: Minimum Information for Publication of Quantitative Real-Time PCRExperiments.Clin.Chem.55,611-622.
6. Cazzola,M.,Calzetta,L.,Facciolo,F.,Rogliani,P.andMatera,M.G.(2017).Pharmacological
investigationontheanti-oxidantandanti-inflammatoryactivityofN-acetylcysteineinanexvivomodelofCOPDexacerbation.Resp.Res.18,1-10.
7. Christoffersen,M.andTroedsson,M.H.T. (2017). Inflammationand fertility in themare.
Reprod.Dom.Anim.Suppl52,14-20.8. Christoffersen,M.,WoodwardE.,Bojesen,A.M,Jacobsen,S.,Petersen,M.R.,Troedsson,
M.H.T. and Lehn-Jensen, H. (2012a). Inflammatory responses to induced infectiousendometritisinmaresresistantorsusceptibletopersistentendometritis.BMCVet.Res.8.
9. Christoffersen,M.,Woodward, E.M., Bojesen, A.M., Petersen,M.R., Squires, E.L., Lehn-
Jensen,H. and Troedsson,M.H.T. (2012b). Effect of immunomodulatory therapy on theendometrial inflammatory response to induced infectious endometritis in susceptiblemares.Theriogenology.78,991-1004.

111
10. Fedorka,C.E.,Scoggin,K.E.,Boakari,Y.L.,Hoppe,N.E.,Squires,E.L.,Ball,B.A.andTroedsson,M.H.T.(2018).Theanti-inflammatoryeffectofexogenouslactoferrinonbreeding-inducedendometritiswhenadministeredpost-breedinginsusceptiblemares.Theriogenology.114,63-69.
11. Fedorka,C.E.,Scoggin,K.E.,Woodward,E.M.,Squires,E.L.,Ball,B.A.,andTroedsson,M.H.T.(2016).TheeffectofseminalplasmaproteinsonendometrialmRNAcytokineexpressioninmaressusceptibletopersistentmating-inducedendometritis.Reprod.Dom.Anim.52,89-96.
12. Fedorka,C.E.,Woodward,E.M.,Scoggin,K.E.,Esteller-Vico,A.,Squires,E.L.,Ball,B.A.and
Troedsson,M.H.T.(2017).Theeffectofcysteine-richsecretoryprotein-3andlactoferrinonendometrialcytokinemRNAexpressionafterbreedinginthehorse.J.EquineVet.Sci.48,136-142.
13. Ferris,R.A.,McCue,P.M.,Borlee,G.I.,Loncar,K.D.,Hennet,M.L.andBorlee,B.R.(2016).In
Vitroefficacyofnonantibiotictreatmentsonbiofilmdisruptionofgram-negativepathogensandaninvivomodelofinfectiousendometritisutilizingisolatesfromtheequineuterus.J.Clin.Microbiol.54,631-639.
14. Fumoso,E.A.,Aguilar,J.,Giguere,S.,Rivulgo,M.,Wade,J.andRogan,D.(2007).Immune
parameters inmares resistant and susceptible topersistentpost-breedingendometritis:effectsofimmunomodulation.Vet.Immunol.Immunopathol.118,30-39.
15. Fumoso, E., Giguere, S.,Wade, J., Rogan, D., Videla-Dorna, I. and Bowden, R.A. (2003).
Endometrial IL-1β, IL-6andTNF-α,mRNAexpression inmaresresistantorsusceptibletopost-breeding endometritis. Effects of estrous cycle, artificial insemination andimmunomodulation.Vet.Immunol.Immunopathol.96,31-41.
16. Gores-Lindholm,A.,Causey,R.,Calderwood-Mays,M.andLeBlanc,M.M.(2009).Effectof
intra-uterineinfusionofdilutedN-acetylcysteineonequineendometrium.Proc.55th.Ann.Conv.Am.Assoc.EquinePract.326.
17. Gores-Lindholm,A.R.,LeBlanc,M.M.,Causey,R.,Hitchborn,A.,Fayrer-Hosken,R.A.,Kruger,
M., Vandenplas, M.L., Flores, P. and Ahlschwede, S. (2013). Relationships betweenintrauterine infusion of N-acetylcysteine, equine endometrial pathology, neutrophilfunction,post-breedingtherapy,andreproductiveperformance.Theriogenology.80,218-227.

112
18. Jacob, F., Guertler, R., Naim, S., Nixdorf, S., Fedier, A., Hacker, N.F. and Heinzelmann-Schwartz,V.(2013).PLoSOne.8.
19. Kenney,R.M.(1978).Cyclicandpathologicchangesofthemareendometriumasdetected
bybiopsy,withanoteonearlyembryonicdeath.J.Am.Vet.Med.Assoc.172,241–62.20. Khan,F.A.,Chenier,T.S.,Foster,R.A.,Hewson,J.andScholtz,E.L.(2018).Endometrialnitric
oxide synthase activity inmares susceptible or resistant to persistent breeding-inducedendometritisandtheeffectofaspecificiNOSinhibitorinvitro.Reprod.Dom.Anim.53,718-724.
21. Khan, F.A., Chenier, T.S.,Murrant, C.L., Foster, R.A.,Hewson, J. and Scholtz, E.L. (2017).
Dose-dependentinhibitionofuterinecontractilitybynitricoxide:Apotentialmechanismunderlyingpersistentbreeding-inducedendometritisinthemare.Theriogenology.90,59-64.
22. Kozera,B.andRapaczM.(2013).Referencegenesinreal-timePCR.J.Appl.Genetics.54,
391-406.23. LeBlanc,M.M.(2010).Advancesinthediagnosisandtreatmentofchronicinfectiousand
post-mating-inducedendometritisinthemare.Reprod.Dom.Anim.Suppl45,21-27.24. LeBlanc,M.M.andCausey,R.C. (2009).Clinicalandsubclinicalendometritis inthemare:
boththreatstofertility.Reprod.Dom.Anim.Suppl3,10-22.25. Lyle,S.K.(2012). Incorporatingnon-antibioticanti-infectiveagents intothetreatmentof
equineendometritis.ClinicalTheriogenology.4,386-391.
26. Majano,P.L.,Medina,J.,Zubia,I.,Sunyer,L.,Lara-Pezzi,E.,Maldonado-Rodriguez,A.,Lopez-Cabrera, M. and Moreno-Otero, R. (2004). N-acetyl-cysteine modulates inducible nitricoxidesynthasegeneexpressioninhumanhepatocytes.J.Hepatol.40,632-637.
27. Melkus,E.,Witte,T.,Walter,I.,Heuwieser,W.andAurich,C.(2013).Investigationsonthe
endometrialresponsetointrauterineadministrationofN-acetylcysteineinoestrousmares.Reprod.Dom.Anim.48,591-597.
28. Pahan,K.,Sheikh,F.G.,Namboodiri,A.M.S.andSingh,I.(1997).N-acetylcysteineinhibits
induction of NO production by endotoxin or cytokine stimulated rat peritonealmacrophages,C6glialcellsandastrocytes.FreeRadic.Biol.Med.24,39-48.

113
29. Paintlia, M.K., Paintlia, A.S., Singh, A.K. and Singh, I. (2008). Attenuation oflipopolysaccharide induced inflammatory responseandphospholipidsmetabolismat thefeto-maternalinterfacebyN-acetylcysteine.Pediatr.Res.64.
30. Palm,F.,Walter,I.,Budik,S.,Kolodziejek,J.,Nowotny,N.andAurich,C.(2008).Influenceof
differentsemenextendersandseminalplasmaonPMNmigrationandonexpressionofIL-1β,IL-6,TNF-αandCOX-2mRNAintheequineendometrium.Theriogenology.70,843-851.
31. Nikolakopoulos, E. and Watson, E.D. (1999). Uterine contractility is necessary for the
clearance of intrauterine fluid but not bacteria after bacterial infusion in the mare.Theriogenology.52,413-423.
32. Scoggin,C.F.(2016).EndometritisNontraditionaltherapies.Vet.Clin.Equine.32,499-511.
33. Stanislaus,R.,Gilg,A.G., Singh,A.K., Singh, I. (2005).N-acetyl-L-cysteineameliorates the
inflammatorydiseaseprocessinexperimentalautoimmuneencephalomyelitisinLewisrats.J.Autoimmun.2.
34. Tessier,L.,Cote,O.,Clark,M.E.,Viel,L.,Diaz-Mendez,A.,Ander,S.andBienzle,D.(2017).
Impairedresponseofthebronchialepitheliumtoinflammationcharacterizessevereequineasthma.BMCGenomics.18.
35. Tirouvanziam,R.,Conrad,C.K.,Bottiglieri,T.,Herzenberg,L.A.,Moss,R.B.andHerzenberg,
L.A.(2006).HighdoseoralN-acetylcysteine,aglutathioneprodrug,modulatesinflammationincysticfibrosis.Proc.Natl.Acad.Sci.103,4628-4633.
36. Troedsson,M.H.T. (2006).Breeding-inducedendometritis inmares.Vet.Clin.Equine.22,
705-712.37. Troedsson, M.H.T. (2012). Non-antibiotic treatment options in mares suffering from
endometritis.LeipzigerBlaueHefte.9,264-267.38. Watson,E.D.(2000).Post-breedingendometritisinthemare.Anim.Reprod.Sci.60-61,221-
232.39. Witte,T.S.,Melkus,E.,Walter,I.,Senge,B.,Schwab,S.,Aurich,C.andHeuwieser,W.(2012).
Theriogenology.78,1199-1208.

114
40. Woodward,E.M.andTroedsson,M.H.T. (2013)Equinebreeding-inducedendometritis.J.EquineVet.Sci.33,673–82.
41. Woodward,E.M.andTroedsson,M.H.T.(2015).Inflammatorymechanismsofendometritis.
EquineVet.J.47,384-389.42. Woodward,E.M.,Christoffersen,M.,Campos,J.,Betancourt,A.,Horohov,D.,Scoggin,K.E.,
Squires,E.L.andTroedsson,M.H.T.(2013a).Endometrialinflammatorymarkersoftheearlyresponse inmares susceptible or resistant to persistent breeding-induced endometritis.Reproduction.145,289-296.
43. Woodward,E.M.,Christoffersen,M.,Campos,J.,Horohov,D.W.,Scoggin,K.E.,Squires,E.
andTroedsson,M.H.T.(2013b).Aninvestigationofuterinenitricoxideproductioninmaressusceptibleand resistant topersistentbreeding-inducedendometritis and theeffectsofimmunomodulation.Reprod.Dom.Anim.48,554–561.
44. Woodward, E.M., Christoffersen, M., Horohow, D., Squires, E.L. and Troedsson, M.H.T.
(2015). The effect of treatment with immune modulators on endometrial cytokineexpressioninmaressusceptibletopersistentbreeding-inducedendometritis.EquineVet.J.47,235-239.
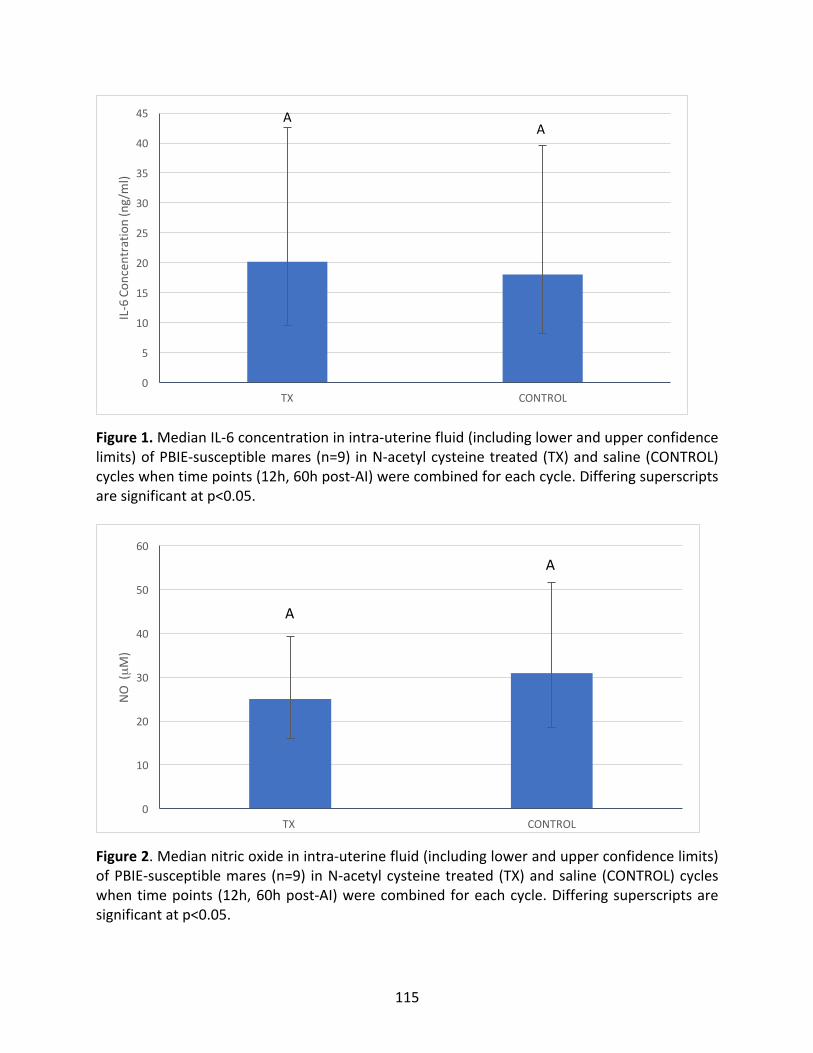
115
Figure1.MedianIL-6concentrationinintra-uterinefluid(includinglowerandupperconfidencelimits)ofPBIE-susceptiblemares(n=9) inN-acetylcysteinetreated(TX)andsaline(CONTROL)cycleswhentimepoints(12h,60hpost-AI)werecombinedforeachcycle.Differingsuperscriptsaresignificantatp<0.05.
Figure2.Mediannitricoxideinintra-uterinefluid(includinglowerandupperconfidencelimits)ofPBIE-susceptiblemares (n=9) inN-acetylcysteinetreated(TX)andsaline (CONTROL)cycleswhentimepoints (12h,60hpost-AI)werecombinedforeachcycle.Differingsuperscriptsaresignificantatp<0.05.
0
5
10
15
20
25
30
35
40
45
TX CONTROL
IL-6Con
centratio
n(ng/ml)
AA
0
10
20
30
40
50
60
TX CONTROL
NO(µM)
A
A
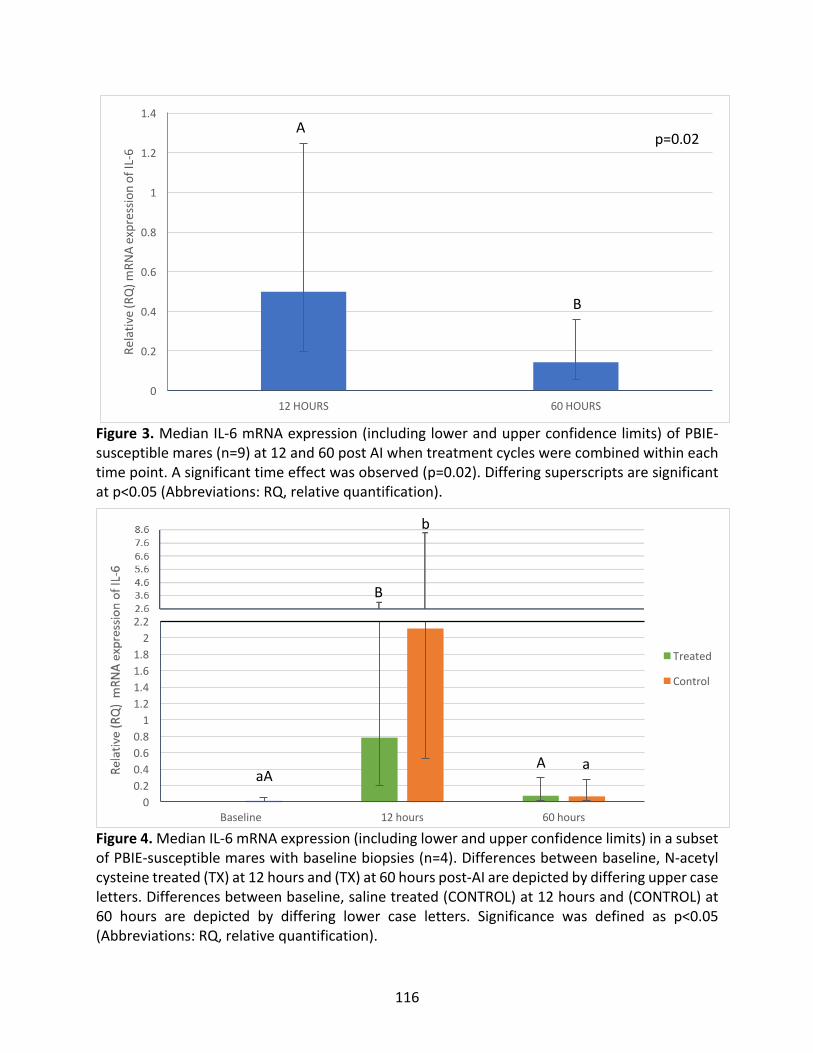
116
Figure3.MedianIL-6mRNAexpression(includinglowerandupperconfidencelimits)ofPBIE-susceptiblemares(n=9)at12and60postAIwhentreatmentcycleswerecombinedwithineachtimepoint.Asignificanttimeeffectwasobserved(p=0.02).Differingsuperscriptsaresignificantatp<0.05(Abbreviations:RQ,relativequantification).
Figure4.MedianIL-6mRNAexpression(includinglowerandupperconfidencelimits)inasubsetofPBIE-susceptiblemareswithbaselinebiopsies(n=4).Differencesbetweenbaseline,N-acetylcysteinetreated(TX)at12hoursand(TX)at60hourspost-AIaredepictedbydifferinguppercaseletters.Differencesbetweenbaseline,salinetreated(CONTROL)at12hoursand(CONTROL)at60 hours are depicted by differing lower case letters. Significance was defined as p<0.05(Abbreviations:RQ,relativequantification).
0
0.2
0.4
0.6
0.8
1
1.2
1.4
12HOURS 60HOURS
Relativ
e(RQ)m
RNAexpressio
nofIL-6
A
B
p=0.02
00.20.40.60.81
1.21.41.61.82
2.2
Baseline 12hours 60hours
Treated
Control
aA
B
A a
b
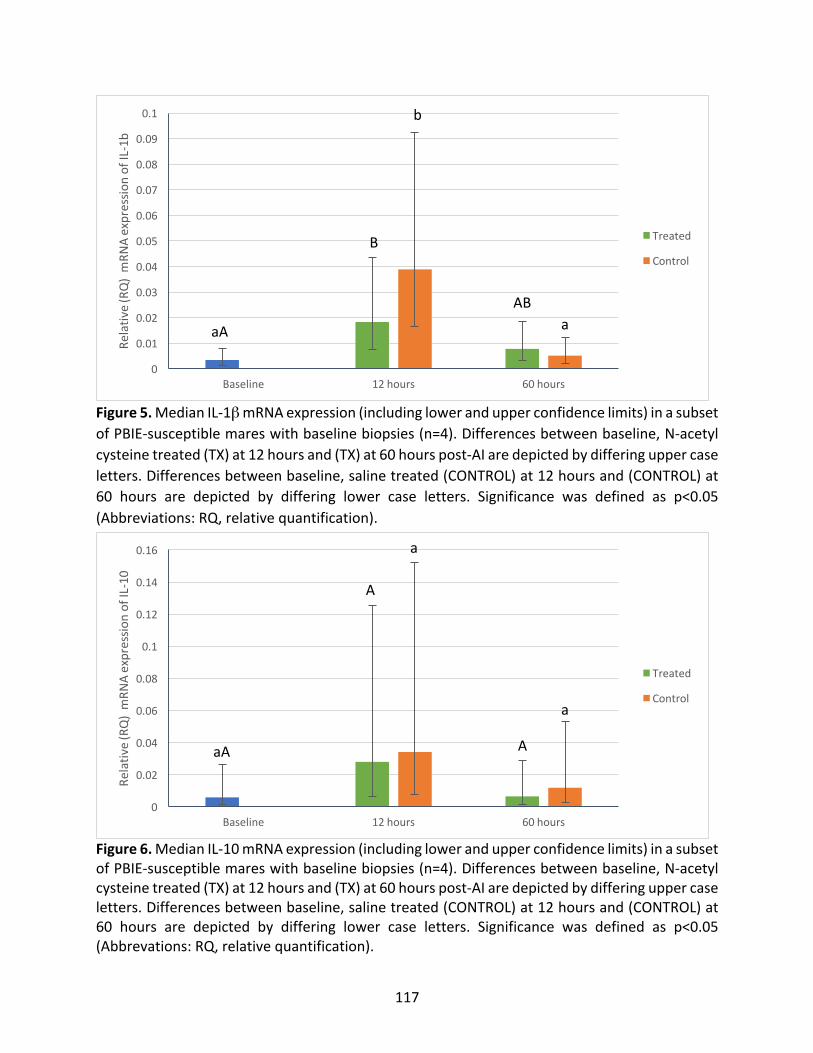
117
Figure5.MedianIL-1βmRNAexpression(includinglowerandupperconfidencelimits)inasubsetofPBIE-susceptiblemareswithbaselinebiopsies(n=4).Differencesbetweenbaseline,N-acetylcysteinetreated(TX)at12hoursand(TX)at60hourspost-AIaredepictedbydifferinguppercaseletters.Differencesbetweenbaseline,salinetreated(CONTROL)at12hoursand(CONTROL)at60 hours are depicted by differing lower case letters. Significance was defined as p<0.05(Abbreviations:RQ,relativequantification).
Figure6.MedianIL-10mRNAexpression(includinglowerandupperconfidencelimits)inasubsetofPBIE-susceptiblemareswithbaselinebiopsies(n=4).Differencesbetweenbaseline,N-acetylcysteinetreated(TX)at12hoursand(TX)at60hourspost-AIaredepictedbydifferinguppercaseletters.Differencesbetweenbaseline,salinetreated(CONTROL)at12hoursand(CONTROL)at60 hours are depicted by differing lower case letters. Significance was defined as p<0.05(Abbrevations:RQ,relativequantification).
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0.08
0.09
0.1
Baseline 12hours 60hours
Relativ
e(RQ)mRN
Aexpressio
nofIL-1b
Treated
Control
aA
B
b
ABa
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
Baseline 12hours 60hours
Relativ
e(RQ)mRN
Aexpressio
nofIL-10
Treated
Control
aA
A
a
A
a
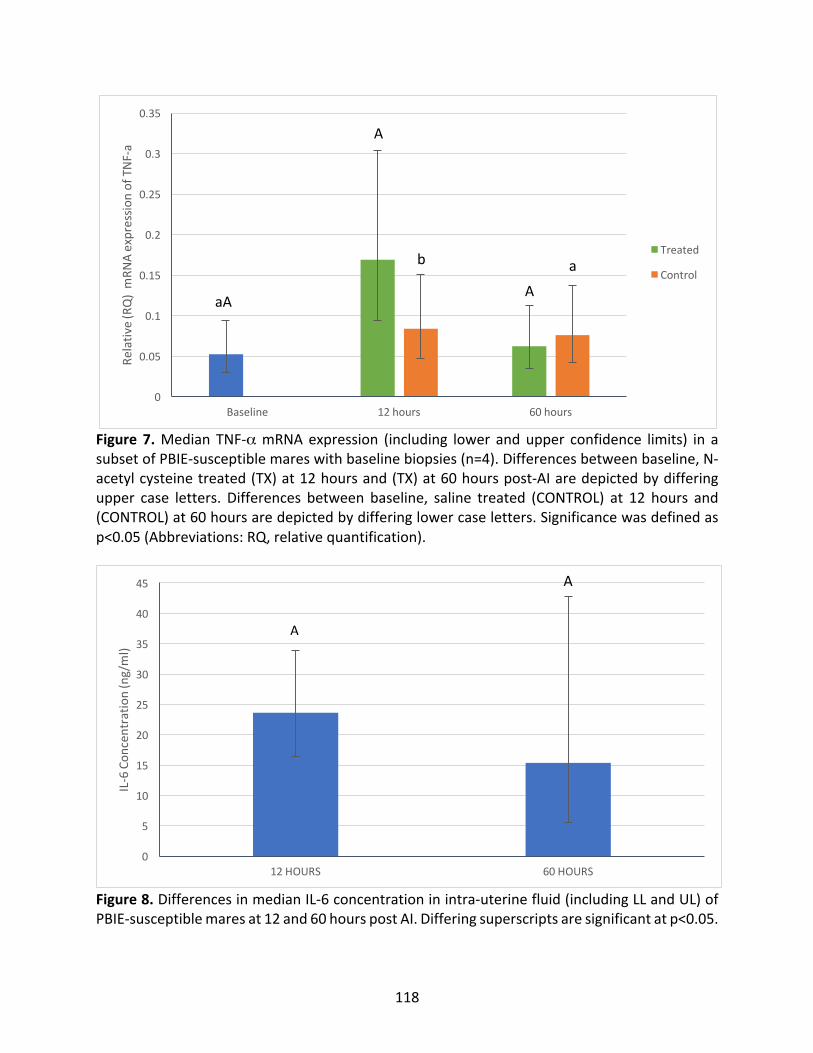
118
Figure7.Median TNF-αmRNAexpression (including lower andupper confidence limits) in asubsetofPBIE-susceptiblemareswithbaselinebiopsies(n=4).Differencesbetweenbaseline,N-acetylcysteinetreated(TX)at12hoursand(TX)at60hourspost-AIaredepictedbydifferingupper case letters. Differences between baseline, saline treated (CONTROL) at 12 hours and(CONTROL)at60hoursaredepictedbydifferinglowercaseletters.Significancewasdefinedasp<0.05(Abbreviations:RQ,relativequantification).
Figure8.DifferencesinmedianIL-6concentrationinintra-uterinefluid(includingLLandUL)ofPBIE-susceptiblemaresat12and60hourspostAI.Differingsuperscriptsaresignificantatp<0.05.
0
5
10
15
20
25
30
35
40
45
12HOURS 60HOURS
IL-6Con
centratio
n(ng/ml)
A
A
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
Baseline 12hours 60hours
Relativ
e(RQ)mRN
Aexpressio
nofTNF -a
Treated
Control
A
A
b a
aA
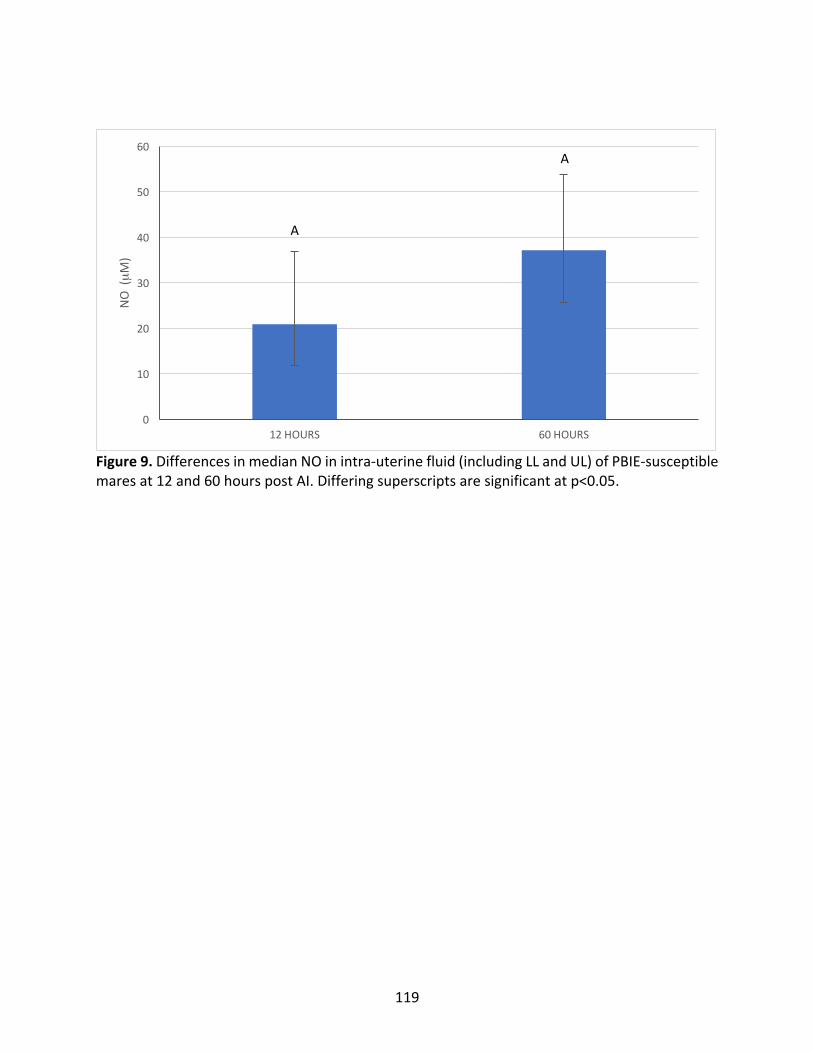
119
Figure9.DifferencesinmedianNOinintra-uterinefluid(includingLLandUL)ofPBIE-susceptiblemaresat12and60hourspostAI.Differingsuperscriptsaresignificantatp<0.05.
0
10
20
30
40
50
60
12HOURS 60HOURS
NO(µM)
A
A
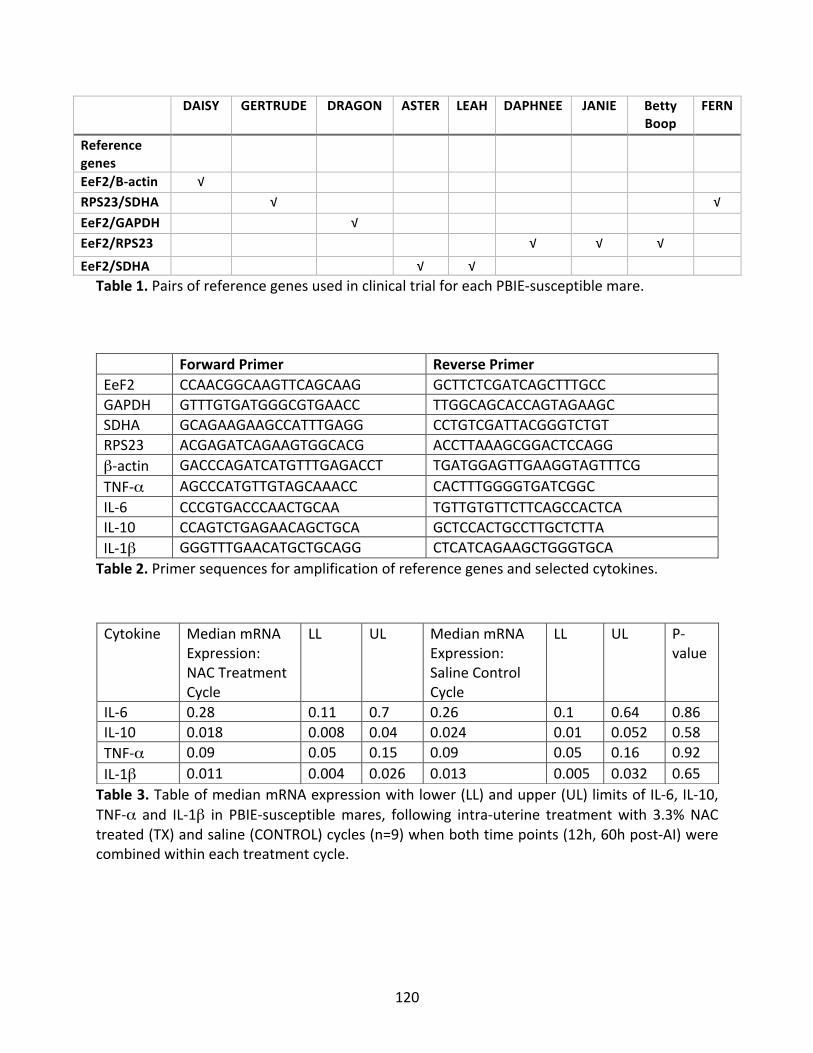
120
DAISY GERTRUDE DRAGON ASTER LEAH DAPHNEE JANIE BettyBoop
FERN
Referencegenes
EeF2/B-actin √
RPS23/SDHA √ √EeF2/GAPDH √
EeF2/RPS23 √ √ √
EeF2/SDHA √ √
Table1.PairsofreferencegenesusedinclinicaltrialforeachPBIE-susceptiblemare. ForwardPrimer ReversePrimerEeF2 CCAACGGCAAGTTCAGCAAG GCTTCTCGATCAGCTTTGCCGAPDH GTTTGTGATGGGCGTGAACC TTGGCAGCACCAGTAGAAGCSDHA GCAGAAGAAGCCATTTGAGG CCTGTCGATTACGGGTCTGTRPS23 ACGAGATCAGAAGTGGCACG ACCTTAAAGCGGACTCCAGGβ-actin GACCCAGATCATGTTTGAGACCT TGATGGAGTTGAAGGTAGTTTCG TNF-α AGCCCATGTTGTAGCAAACC CACTTTGGGGTGATCGGCIL-6 CCCGTGACCCAACTGCAA TGTTGTGTTCTTCAGCCACTCAIL-10 CCAGTCTGAGAACAGCTGCA GCTCCACTGCCTTGCTCTTAIL-1β GGGTTTGAACATGCTGCAGG CTCATCAGAAGCTGGGTGCATable2.Primersequencesforamplificationofreferencegenesandselectedcytokines.
Table3.TableofmedianmRNAexpressionwithlower(LL)andupper(UL)limitsofIL-6,IL-10,TNF-α and IL-1β in PBIE-susceptiblemares, following intra-uterine treatmentwith 3.3%NACtreated(TX)andsaline(CONTROL)cycles(n=9)whenbothtimepoints(12h,60hpost-AI)werecombinedwithineachtreatmentcycle.
Cytokine MedianmRNAExpression:NACTreatmentCycle
LL
UL MedianmRNAExpression:SalineControlCycle
LL
UL P-value
IL-6 0.28 0.11 0.7 0.26 0.1 0.64 0.86IL-10 0.018 0.008 0.04 0.024 0.01 0.052 0.58TNF-α 0.09 0.05 0.15 0.09 0.05 0.16 0.92IL-1β 0.011 0.004 0.026 0.013 0.005 0.032 0.65
121
Table4.TableofmedianmRNAexpressionwithlower(LL)andupper(UL)limitsofIL-6,IL-10,TNF-α and IL-1β at12and60hourspost-AI inPBIE-susceptiblemares (n=9)when treatmentcycleswerecombinedateachtimepoint.
CytokineBaseline
mRNAExpressionMean(LL,UL)
Cycle12hpost-AI
mRNAExpressionMean(LL,UL)
60hpost-AImRNAExpressionMean(LL,UL)
IL-6 0.013(0.003,0.052) CONTROL 2.11
(0.53,8.28)0.068
(0.017,0.27)
TX 0.79(0.2,3.11)
0.07(0.02,0.30)
IL-10 0.006(0.001,0.026) CONTROL 0.03
(0.008,0.15)0.012
(0.003,0.05)
TX 0.03(0.006,0.13)
0.006(0.001,0.03)
TNF-α 0.05(0.03,0.09) CONTROL 0.08
(0.05,0.15)0.08
(0.04,0.14)
TX 0.17(0.09,0.30)
0.06(0.03,0.11)
IL-1β 0.003(0.001,0.008) CONTROL 0.04
(0.02,0.09)0.005
(0.002,0.01)
TX 0.02(0.008,0.04)
0.008(0.003,0.02)
Table5.TableofmedianmRNAexpressionwithlower(LL)andupper(UL)confidencelimitsofIL-6,IL-10,TNF-αandIL-1βatbaseline,12and60hourspost-AIfortheN-acetylcysteinetreated(TX)andsaline(CONTROL)cyclesforasubsetofPBIE-susceptiblemares(n=4).
Cytokine MedianmRNAexpression12hourspost-AI
LL
UL MedianmRNAexpression60hourspost-AI
LL
UL Pvalue
IL-6 0.5 0.2 1.25 0.14 0.06 0.36 0.02IL-10 0.023 0.01 0.05 0.019 0.009 0.04 0.73TNF-α 0.091 0.05 0.16 0.086 0.05 0.15 0.86IL-1β 0.013 0.005 0.03 0.010 0.004 0.026 0.60

122
CHAPTERFOUR:GENERALDISCUSSION
4.1POSSIBLELINKSBETWEENMRNAEXPRESSIONANDPBIECLINICALSIGNS
Persistentbreeding inducedendometritis continues tobea sourceof infertility in the
equinebreedingindustry(Newcombe1997).PBIEiscausedbymechanicalfactors,suchaspoor
myometrialcontractility,andbydifferencesintheuterineimmuneresponsefollowingbreeding
(ChristoffersenandTroedsson2017).Altered inflammatorycytokineexpression isbelieved to
playaroleinPBIE,withsusceptiblemareshavingincreasedandpersistentmRNAexpressionof
pro-inflammatory cytokines and decreased mRNA expression of anti-inflammatory cytokines
followingAI(WoodwardandTroedsson2015;ChristoffersenandTroedsson2017).Manystudies
haveexaminedcytokinemRNAexpressioninbothPBIEresistantandsusceptiblemares,butnone
haverelatedmRNAexpressiontoclinicalsignsassociatedwithPBIE.N-acetylcysteine,usedasan
intra-uterinemucolyticinmares,hasalsobeenshowntohaveanti-inflammatoryproperties,such
asaffectingcytokinemRNAexpressioninothermodelsofdisease(Stanislausetal.2005;LeBlanc
2010;Cazzolaetal.2017).
TheclinicaltrialreportedinthisworkwasbasedonourhypothesisthattreatmentofPBIE
susceptiblemareswithNACwouldalterinflammatorycytokinemRNAexpression,leadingtothe
resolutionofPBIEclinicalsigns.Theresultsofourstudydidnotrevealanybeneficialeffectof
NACtreatmentoneitherPBIEclinicalsignsorcytokinemRNAexpression,makingitdifficultto
assess any improvement of clinical signs by means of altered mRNA expression, following
treatment.However,somespeculationscanstillbemaderegardingthisconceptbyexamining
patternsofcytokinemRNAexpressionandcomparingthemtotheclinicalsignsobservedatthe

123
different time points post-AI. IL-6 mRNA expression was significantly greater at 12 hours
compared to 60 hours post AI. This is similar to%neutrophil levels and endometrial edema
scores,bothofwhichweresignificantlyelevatedattimepointsnearestAI(12hourspostAIfor
% neutrophils; 12 and 36 hours post AI for uterine edema scores). Similar, though slightly
different trendswerenoted for intra-uterine fluid (IUF) volume,with significantly larger fluid
volumesat36hourscomparedto12,60and84hourspostAI.Thispatternofinflammationwas
expectedasneutrophilsare recruited followingbreeding (Kotilainenetal.1994) startingat4
hoursandpeakingat8hoursafterAI (Katila1995). InPBIEsusceptiblemares, thispattern is
similarbutisevenmorepronounced(Woodwardetal.2013).Fromthis,onecouldspeculatethat
IL-6playsaroleinpromotinginflammationearlyonintheuterineresponse,andthatthisinturn
results in the clinical signs observed in PBIE mares. Interleukin-6 has both pro- and anti-
inflammatoryproperties. It is involved inthe initiationof the inflammatoryresponsebutalso
controlsinflammationbymeansofdecreasingneutrophilrecruitment(Tizard2013).Theeffects
ofIL-6onneutrophilswereapparentinthisstudy,where%neutrophilsdecreasedsignificantly
by60hourspostAI,possiblyasaresultofearlierIL-6mRNAup-regulation.
ThepersistenceofclinicalsignsinourPBIEstudymares,albeitreducedat60hoursfrom
12hours,couldbetheresultofalackofIL-10mRNAup-regulation,asrelativeexpressionofIL-
10didnotchangebetween12and60hourspost-AI.Interleukin-6promotesexpressionofIL-10,
whichisananti-inflammatorycytokine(Tizard2013).ItispossiblethatalteredIL-6expression
also ledtoaberrations in IL-10expression.Wehypothesizethat the failureof increased IL-10
expressionneededtomodulatetheinflammatoryresponsetobreeding,couldaccountforsome
of the persistence of clinical signs seen with PBIE. One would expect that under normal

124
circumstances,levelsofIL-10mRNAexpressionwouldbehighestfollowinginflammation.Yet,if
thisincreaseinIL-10doesnotoccur,asissuspectedinPBIEmares,persistentinflammation,and
thereby persistence of clinical signs, results. This supports the notion that anti-inflammatory
cytokineexpressionisalteredinPBIEsusceptiblemares,andthattheremayinfactbelackofan
anti-inflammatoryresponse,notjustanexaggeratedpro-inflammatoryresponse,playingarole
inthiscondition.
Conclusionsontheeffectsofthepro-inflammatorycytokinesTNF-αandIL-1β,onclinical
signscannotbemadeduetoalackofchangenotedacrosstimepointsfollowingAI.Itispossible
thatIL-1βandTNF-αareinfactaffectedinPBIEsusceptiblemares,butthatthetimepointof
significant increase was not examined in this study. Further investigations are required to
determinehowthesecytokinesmayimpactclinicalsignsassociatedwithPBIE.Moreover,inan
attempttobetterlinkclinicaloutcomeswithcytokinemRNAexpression,futurestudiesshould
examinehow treatmentmethods known toalter cytokinemRNAexpression, suchasMCWE,
dexamethasone,plateletrichplasma(PRP)andlactoferrin,affectPBIEclinicalsigns.
4.2LIMITATIONS OneofthelargestlimitationsofourstudywastheinabilitytomeasureIL-10,TNF-α,and
IL-1β concentration in uterine fluid. While equine ELISA kits exist for these cytokines, no
validations have been published specifically for equine uterine fluid. As such, validationwas
attemptedforthesekitsbutwasunsuccessful.Themostcommonreasonforfailureofvalidation
wassuspectedtobematrixinterferenceduetoanunidentifiedcompoundorsubstancewithin
uterine fluid that caused either false increases or decreases of measured cytokine

125
concentrations.Many of the sampleswere viscous in nature, possibly contributing tomatrix
interferenceoftheassay.Theviscosityofthesamplewassuspectedtobeduetothehighprotein
contentinfluidobtainedfromaninflameduterus.SomeELISAkitscanbevalidateddespitethis
by simplydiluting the sample appropriatelywith the reagentdiluentprovided in the kit (MC
personalcommunication,R&Dsystemstechnicalsupport).ThiswasthecaseforIL-6,theonly
successfully validated cytokine, as the sample was diluted 1:20; however, the appropriate
dilutionfactorvariesforeachindividualcytokine.Thedilutionfactorshouldbegreatenoughto
overcomematrixinterference,butnotsomuchsothatthecytokineofinterestcannolongerbe
detected.VariousdilutionswereattemptedforIL-10,TNF-α,andIL-1β,asreportedinAppendix
4.
Although IL-6was successfully validated, the process itself had limitations, notably in
termsofthenegativecontrol.Forvalidation,apositiveandnegativecontrolwereutilizedand
consistedofPBIE-susceptiblemaresenrolledinthestudy(positivecontrols)andnormalmares
(negative controls) (Appendix 2). Undiluted uterine fluidwas readily available for PBIE-study
mares,however,normalmaresdidnothaveanyuterinefluid.Theprocurementofendometrial
fluidfromthesemareswasthroughalow-volumelavage,ultimatelydilutingthesample.While
theexactvolumeofinfusedfluidwasknownforthisprocedure,theamountrecoveredwasnot
100%, leading toanunknowndilution factor. Thismadecomparisonsbetweennegativeand
positivecontrolmaresdifficult. Sincefluidcouldnotbecollectedinanyotherwayinnormal
mares,thisrepresentedanotherlimitationforthevalidationprocess.
Othervalidationmethodsthatcouldhavebeenattemptedincludepurificationorwashing
ofthesamplewithspincolumnsandaddingfetalbovineserum(FBS)tothereagentdiluent.The

126
quantityofFBStobeaddedtothereagentdiluentvariesgreatlybasedonthetypeofsample,
andcanrangefrom10-50%(MCpersonalcommunication,R&Dsystemstechnicalsupport).FBS
andspincolumnscouldhaveeitherboundtheviscouscomponentofthesampleorfiltereditout.
Neitherofthesemethodswereperformedduetothelimitedvolumeofsamplefluidsavailable
fromthestudymares,sinceeachcytokinewouldrequireitsownvalidationattemptswitheither
FBS,spincolumnsorboth.Furtherstudiesshouldbeperformedtodetermineiftheadditionof
FBStoreagentdiluentcontributestothevalidationprocessandifso,whattheappropriateFBS
concentrationiswhenrunningthesekitsonequineuterinefluid.Inthesameregard,perfecting
themethodologyforendometrialfluidtobefilteredthroughspincolumnscouldalsobeatopic
ofresearch.
Theequine specieshasagreatdealof variability in its geneexpression (Tessieretal.
2017),makingtheselectionofreferencegenesachallenge.Consequently,eachstudymarehad
her own set of reference genes. Thismethod is valid as the reference genes selected were
expressed ina stablemanner foreach individualmare,allowing forcomparisons tobemade
acrossthem.Thismethodissuperiortochoosingtworeferencegenesforallmares,thatarenot
expressed in a stable manner. The latter could be accomplished by employing software
methodologysuchasGeNormorNormFinder,whicharedesignedtochoosethetwobestoverall
referencegenesforallsubjectsinastudy.Thiswasattemptedinourclinicaltrial,however,large
variabilityinthedatameantthatnotwobestreferencegenescouldbeidentified,leadingtothe
selectionofthebesttwoforeach individualmare.Toconfirmthatreferencegeneswerenot
affectingresultoutcomes,thevariable“referencegene”wasenteredintotherepeatedmeasures
ANOVAmodeltoconfirmthattheywerenon-significant.

127
Anadditionallimitationofthisstudywastherepeatedbiopsiesandprocurementoffluid
atmultipletimepointsduringeachestrouscycle.Theeffectofprevioustimepointsamplingon
subsequent samples is unknown.WatsonandSertich (1992) andChristoffersenetal. (2012),
demonstratedthatrepeatedbiopsieswerenotdetrimentaltouterinehealthorpregnancyrates;
however,nostudieshavespecificallyexaminedtheeffectsofprocurementoffluid,endometrial
cytologyandbiopsycombinedonthelevelofinflammation48hourslater.Ideally,onesampling
time pointwould have taken place per cycle,withmultiple cycles of treatment and control,
althoughthevariabilityacrosscyclesbecomesgreaterwiththisapproach.
4.3OVERALLCONCLUSIONSANDFUTUREDIRECTIONS
Despitesignificantamountsofresearch,themechanismsunderlyingthepathophysiology
ofPBIE inmaresremainpoorlyunderstood. Thisstudyprovidedmore informationonmRNA
expressionofIL-6,IL-10,TNF-αandIL-1βinPBIE-susceptiblemaresatboth12and60hourspost-
AI.BaselinemRNAexpressionofthesecytokineswasalsoobtainedforasubsetofmares.While
statistically significant differences were not often noted between these time points, the
informationandtrendsfoundinthestudycontributedtodefiningthegeneralcytokineprofile
and alterations in the immune response of PBIE-susceptible mares. Knowing more about
differentcytokineexpressionatvarioustimepointspost-AIandhowthesedifferfromnormal
maresmayhelptargetthedevelopmentofnewtreatmentmodalities.Furthermore,integrating
theimmuneresponsewiththeclinicalpictureofPBIE,byassessingbothcytokineexpressionand
clinicalsignsatvarioustimepoints,couldhelpcompleteourunderstandingofPBIE.Forexample,
PRPshowspromiseinitseffectsonIL-6,IL-8andIL-1βandiNOSmRNAexpression24hourspost-

128
AIinPBIE-susceptiblemares(Metcalfetal.2012).ElevatedNOisbelievedtoplayaroleindelayed
uterineclearance.OthertimepointscouldbeexaminedfollowingtreatmentwithPRPandclinical
impactscouldbeassessed,suchasfluidaccumulation,asithasbeenshownthatiNOSisaffected.
Furtherresearchisneededtodeterminehowothercytokineswouldbeaffectedatdifferenttime
pointsbyvarioustreatmentsandhowclinicalsignswouldtherebybeimpacted.
Woodwardetal.(2013)demonstratedthatmanycytokineschangesignificantlyintheir
mRNAexpressionat6hourspostAI.Thisisnottosayhowever,thatcytokinemRNAexpression
alsodoesnotchangeatothertimepointspostAI,orthatothercytokinesarealteredatdifferent
time-points. Up-regulation of mRNA expression of inflammatory cytokines can be extremely
short-livedandcouldeasilybemissediftheappropriatetimepointsarenotselectedforeach
cytokine of interest. Increased expression may also be extremely mild in nature, making
differencesdifficulttodetect,eveninextremeinflammatorymodels,suchassepticshock(MC
personalcommunication,Dr.Wood).Infuturestudies,additionaltimepointsbothimmediately
afterAIandlaterintheimmuneresponse,shouldbeexaminedinordertocapturechangesto
cytokine mRNA expression across a broader timeline of the immune response. Additional
cytokinesofinterestincludeIL-8,IL-1RAandIFN-γ.IL-8isinvolvedinneutrophilchemotaxisand
itiswell-knownthatneutrophilsareelevatedinPBIE-susceptiblemares.IL-1RA,encodedbyIL-
1RNgene,isananti-inflammatorycytokineinvolvedintheregulationofIL-1βwhereasIFN-γis
pro-inflammatory. IL-1RN and IFN-γ changed across certain time points in the study by
Woodward et al. (2013) and recently, PBIE-susceptible mares treated with CRISP-3 and
lactoferrin had altered endometrial IL-1RN and IFN-γ expression (Fedorka et al. 2018). The

129
additional examination of these cytokines in future studies would certainly add to our
understandingofthepathophysiologyofPBIE.
TreatmentwithNAChadnoeffectontheresolutionofclinicalsignsoronendometrial
cytokinemRNAexpression inPBIEsusceptiblemares. However,asignificantandunexpected
findingwastheseveredetrimentaleffectsoftreatmentwithNAContheendometriuminPBIE-
susceptible mares. Loss of endometrial luminal epithelium (erosion), diffuse, deep-seated
neutrophil infiltration and presence of neutrophils in the glandular lumenwere some of the
changesnotedinNACtreatedcycles,andaresimilartochangesnotedfollowingintra-uterine
irritantssuchaskerosene.TheseresultswereparticularlysurprisinggiventhatNACiswidelyused
inmares that could be susceptible to PBIE, such as thosewith bacterial endometritiswith a
biofilm or in mares with abnormal mucus accumulation. The results from this study should
prompt re-evaluation of the practice of NAC intrauterine infusion in mares with a history
suggestiveofPBIE.
ThisisthefirststudydemonstratingtheadverseeffectsofNAContheendometriumof
PBIE mares. Previous studies suggested NAC did not appear to have an effect on the
endometrium, however those studies included normal mares or mares with histories of
barrenness, but not necessarily PBIE. As was found for kerosene (Bracher et al. 1991), it is
possiblethatthedetrimentaleffectsofNACareexacerbatedinPBIEmaresorthosewithpoor
biopsygrades.Furthermore,itispossiblethatNACinitselfisnotdetrimentaltotheendometrium
histologically,butwhenitiscombinedwithAIinPBIE-susceptiblemares,theadverseeffectsare
severe,andthatsimilarchangescouldbeseenwithotherirritantscombinedwithNAC.Further
studiesareneededtoconfirmtheadverseeffectsofNAConmareswithdifferentendometrial

130
biopsygradesand/orwhengivenincombinationwithAIorothertreatments.Thiswouldallow
for clinicians to better target their patients when considering NAC infusion and allow for
appropriatemeasurestobetaken,suchasperforminguterinelavagebetweenNACinfusionand
breeding when NAC treatment is necessary. It is evident that more research needs to be
undertakenbeforetreatinganyPBIE-susceptiblemarewithNAC.
Finally,muchislefttolearnregardingtheoverallpathophysiologyofPBIE.Riskfactors
for PBIE-susceptiblemares, such as endometrial biopsy grade or extensive fibrosis, could be
evaluatedfortheireffectsontheexpressionof inflammatorygenes.Studieshaveshownthat
mareswith severeendometrial fibrosisassociatedwithendometrosishavealtereduterocalin
anduteroferrinexpressionpatternsaswellasalteredhormonereceptors,suggestingthatnormal
uterinefunctionalityisalteredincasesofendometrialpathology(Lehmannetal.2011;Rebordao
etal.2014).ItispossiblethatsomeofthesechangesalsoplayaroleinthedevelopmentofPBIE.
Specific treatments could then be developed for more targeted immunologic abnormalities.
Inflammationisatightlyregulatedprocessandbeginswithrecognitionofforeignmaterial,such
asspermatozoa.Thisisfollowedbythereleaseofinflammatorymediatorsforcellularsignaling
torecruitinflammatorycells.ItiswellknownthatsomeoftheseprocessesarealteredinPBIE-
susceptiblemares, howevermuch remainsunknown. Studiesmoving forward could focuson
differences in inflammatory gene sequencing in response to AI in PBIE-susceptible mares
compared to resistant mares. Marth et al. (2015, 2016) demonstrated the importance of
pathogenrecognitionandNOD-likereceptorsaswellasinductionofchemokinesinnormalmares
followingbacterialinoculationwithE.coli(3,12,24,48and72hours).Theinflammatoryresponse
wasveryrapid,occurringwithin3hourspost-inoculation(Marthetal.2015).Similarstudiesof

131
PBIEresistantandsusceptiblemaresatmultipleandearly(1-3hours)timepointsfollowingAI
wouldprovideuswithnewinsightintohowandwhyPBIEdevelops.Itispossiblethatsusceptible
maresalsohavealteredrecognitionofpathogens,inthiscasespermatozoa,leadingtotheup-
regulationofchemokines,cytokinesandothermediatorsofinflammation,suchasNO.Additional
informationonthealteredimmuneresponseofPBIEmares,includingtheinitialrecognitionof
pathogensandcellsignaling,willclarifythepathophysiologyofthisimportantcauseofinfertility.
Inconclusion,PBIEremainspoorlyunderstoodinmaresdespiteextensivepastresearch,
likelyduetothemultifacetedpathophysiologyofthisdiseaseprocessthatincludesriskfactors,
altered mechanical clearance mechanisms, potentially abnormal pathogen recognition,
persistent inflammationpost-breeding, and altered response to traditional uterine therapies.
Futureresearchtobetterunderstandtheearlypost-breedingresponseinparticularmayhelpto
identifykeytimepointsandmechanismsfortherapythatcanimpacttheimmuneresponseof
PBIE-susceptiblemares.

132
4.4REFERENCES1. Bracher,V.,Neuschaefer,A.andAllen,R.W.(1991).Theeffectofintra-uterineinfusionof
keroseneontheendometriumofmares.J.Reprod.Fert.Suppl44,706-707.2. Cazzola,M.,Calzetta,L.,Facciolo,F.,Rogliani,P.andMatera,M.G.(2017).Pharmacological
investigationontheanti-oxidantandanti-inflammatoryactivityofN-acetylcysteineinanexvivomodelofCOPDexacerbation.Resp.Res.18,1-10.
3. Christoffersen,M.andTroedsson,M.H.T.(2017). Inflammationandfertility inthemare.Reprod.Dom.Anim.Suppl52,14-20.
4. Christoffersen,M.,Woodward,E.,Bojesen,A.M.,Jacobsen,S.,Petersen,M.R.,Troedsson,M.H.T. and Lehn-Jensen, H. (2012). Inflammatory responses to induced infectiousendometritisinmaresresistantorsusceptibletopersistentendometritis.BMCVet.Res.8.
5. Fedorka, C.E., Scoggin, K.E., Boakari, Y.L., Hoppe, N.E., Squires, E.L., Ball, B.A. and
Troedsson, M.H.T. (2018). The anti-inflammatory effect of exogenous lactoferrin onbreeding-induced endometritis when administered post-breeding in susceptible mares.Theriogenology.114,63-69.
6. Katila, T. (1995). Onset and duration of uterine inflammatory response of mares afterinseminationwithfreshsemen.Biol.Reprod.Mono.1,515-517.
7. Kotilainen,T.,Huhtinen,M.andKatila,T.(1994).Sperm-inducedleukocytosisintheequineuterus.Theriogenology.41,629-636.
8. LeBlanc,M.M.(2010).Advancesinthediagnosisandtreatmentofchronicinfectiousandpost-mating-inducedendometritisinthemare.Reprod.Dom.Anim.Suppl45,21-27.
9. Lehmann,J.,Ellenberger,C.,Hoffman,C.,Bazer,F.W.,Klug,J.,Allen,W.R.,Sieme,H.andSchoon,H.-A.(2011).Morpho-functionalstudiesregardingthefertilityprognosisofmaressufferingfromequineendometrosis.Theriogenology.76,1326-1336.
10. Marth, C.D., Firestone, S.M., Glenton, L.Y., Browing, G.F., Young, N.D. and Krekeler, N.(2016).Oestrouscycle-dependentequineuterineimmuneresponsetoinducedinfectiousendometritis.Vet.Res.47.

133
11. Marth,C.D.,Young,N.D.,Glenton,L.Y.,Noden,D.M.,Browing,G.F.andKrekeler,N.(2015).Deepsequencingoftheuterineimmuneresponsetobacteriaduringtheequineoestrouscycle.BMCGenomics.16.
12. Metcalf,E.S.,Scoggin,K.andTroedsson,M.H.T.(2012).Theeffectofplatelet-richplasmaon endometrial pro-inflammatory cytokines in susceptible mares following semendeposition.J.EquineVet.Sci.32,498.
13. NewcombeJ.R.(1997).Theeffectoftheincidenceanddepthofintra-uterinefluidinearlydioestrusonpregnancyrateinmares.Pferdeheilkunde.13,545.
14. Rebordao,M.R.,Galvao,A.,Szostek,A.,Amaral,Mateus,L.,Skarynski,D.J.andFerreira-Dias,G. (2014). Physiopathologicmechanisms involved inmare endometrosis. Reprod.Dom.Anim.49;Suppl.4,82-87.
15. Stanislaus,R.,Gilg,A.G.,Singh,A.K.,Singh, I. (2005).N-acetyl-L-cysteineamelioratestheinflammatory disease process in experimental autoimmune encephalomyelitis in Lewisrats.J.Autoimmun.2.
16. Tessier,L.,Cote,O.,Clark,M.E.,Viel,L.,Diaz-Mendez,A.,Ander,S.andBienzle,D.(2017).Impairedresponseofthebronchialepitheliumtoinflammationcharacterizessevereequineasthma.BMCGenomics.18.
17. TizardI.R.(2013)Thedefenseofthebody.In:VeterinaryImmunology.9thedn.,I.R.Tizard,ElsevierSaunders,Missouri.pp1-10.
18. Watson,E.D.andSertich,P.L.(1992).Effectofrepeatedcollectionofmultipleendometrialbiopsyspecimensonsubsequentpregnancy inmares.J.Am.Vet.Med.Assoc.201,438-440.
19. WoodwardE.M.andTroedsson,M.H.T.(2015).Inflammatorymechanismsofendometritis.
EquineVet.J.47,384-389.
20. Woodward,E.M.,Christoffersen,M.,Campos,J.,Betancourt,A.,Horohov,D.,Scoggin,K.E.,Squires,E.L.andTroedsson,M.H.T.(2013).Endometrialinflammatorymarkersoftheearlyresponse inmares susceptibleor resistant topersistentbreeding-inducedendometritis.Reproduction.145,289-296.
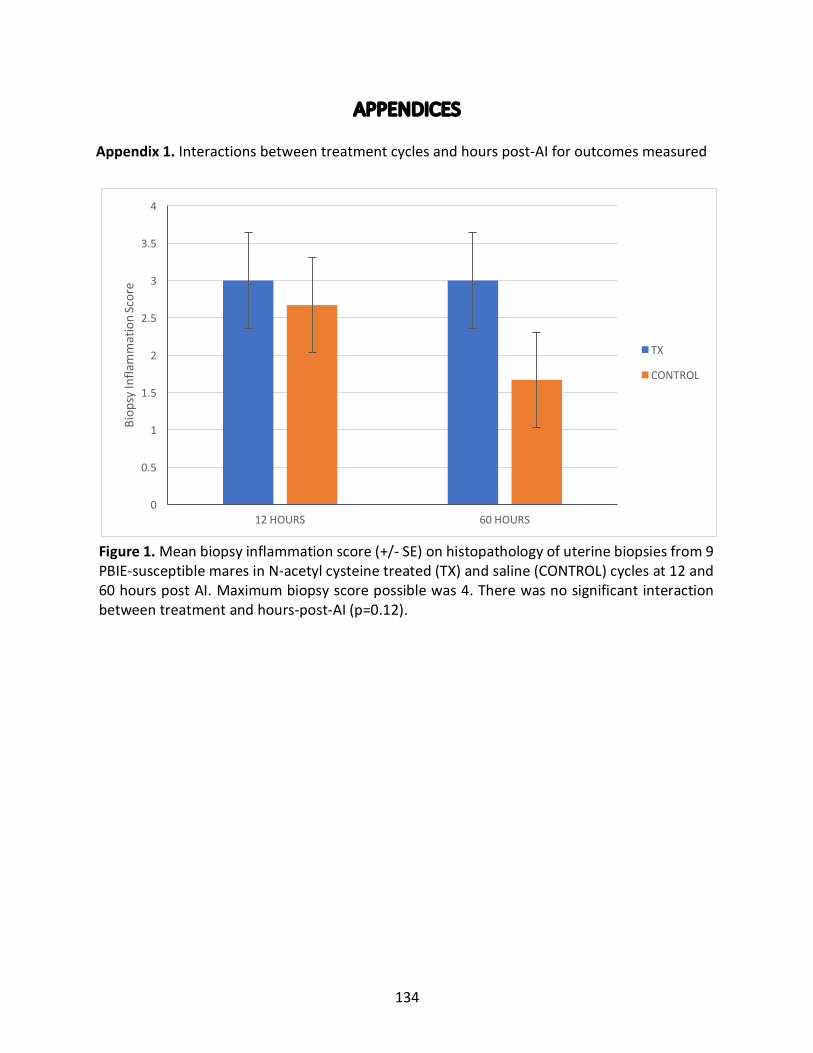
134
APPENDICESAppendix1.Interactionsbetweentreatmentcyclesandhourspost-AIforoutcomesmeasured
0
0.5
1
1.5
2
2.5
3
3.5
4
12HOURS 60HOURS
Biop
syInfla
mmationScore
TX
CONTROL
Figure1.Meanbiopsyinflammationscore(+/-SE)onhistopathologyofuterinebiopsiesfrom9PBIE-susceptiblemaresinN-acetylcysteinetreated(TX)andsaline(CONTROL)cyclesat12and60hourspostAI.Maximumbiopsyscorepossiblewas4.Therewasnosignificantinteractionbetweentreatmentandhours-post-AI(p=0.12).
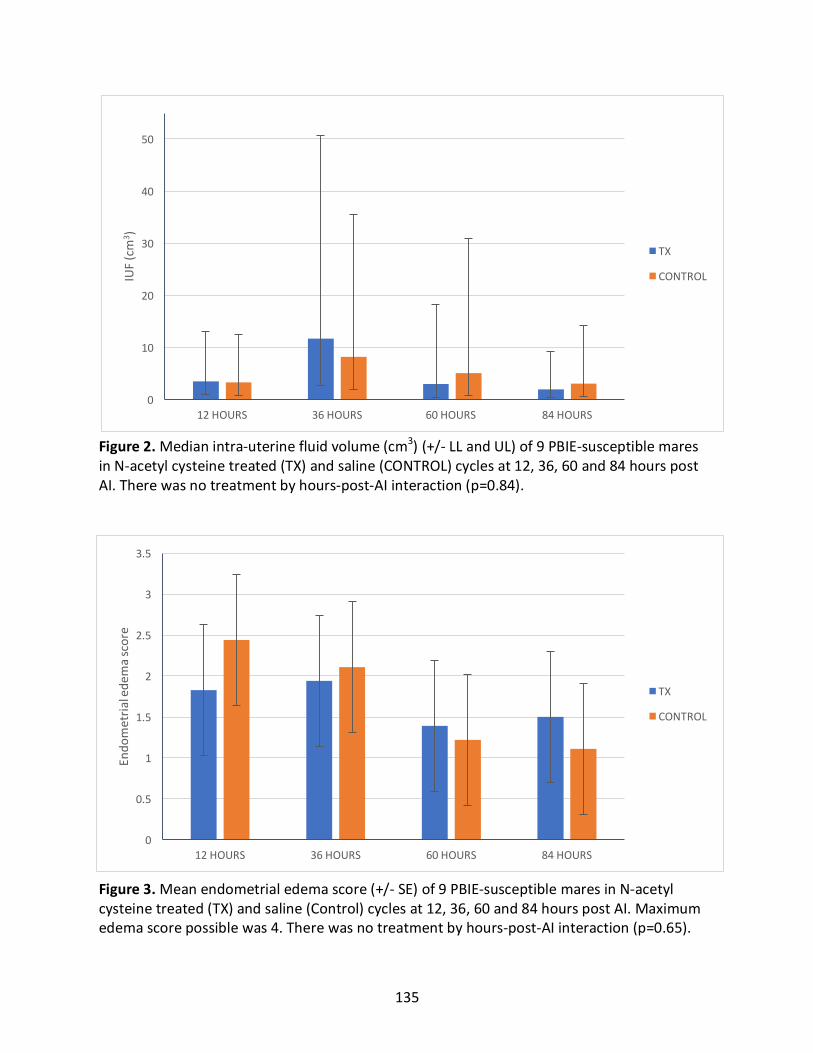
135
0
10
20
30
40
50
12HOURS 36HOURS 60HOURS 84HOURS
IUF(cm
3 )
TX
CONTROL
Figure2.Medianintra-uterinefluidvolume(cm3)(+/-LLandUL)of9PBIE-susceptiblemaresinN-acetylcysteinetreated(TX)andsaline(CONTROL)cyclesat12,36,60and84hourspostAI.Therewasnotreatmentbyhours-post-AIinteraction(p=0.84).
0
0.5
1
1.5
2
2.5
3
3.5
12HOURS 36HOURS 60HOURS 84HOURS
Endo
metria
lede
mascore
TX
CONTROL
Figure3.Meanendometrialedemascore(+/-SE)of9PBIE-susceptiblemaresinN-acetylcysteinetreated(TX)andsaline(Control)cyclesat12,36,60and84hourspostAI.Maximumedemascorepossiblewas4.Therewasnotreatmentbyhours-post-AIinteraction(p=0.65).
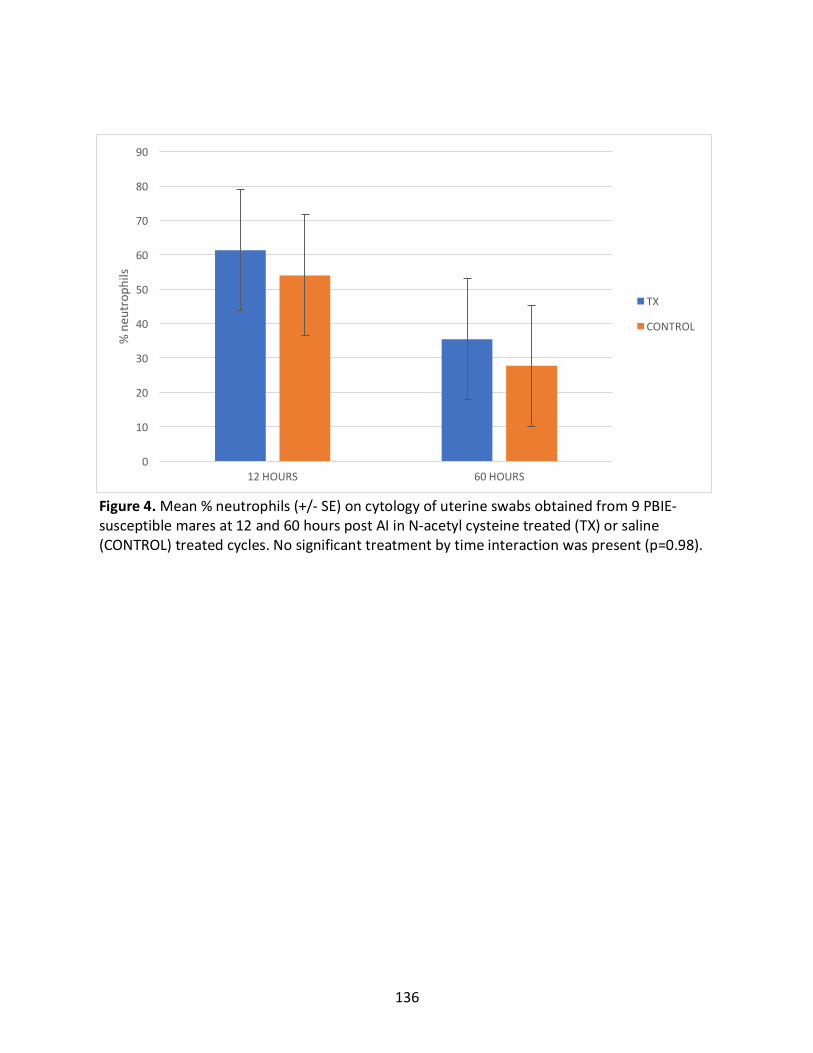
136
0
10
20
30
40
50
60
70
80
90
12HOURS 60HOURS
%neu
trop
hils
TX
CONTROL
Figure4.Mean%neutrophils(+/-SE)oncytologyofuterineswabsobtainedfrom9PBIE-susceptiblemaresat12and60hourspostAIinN-acetylcysteinetreated(TX)orsaline(CONTROL)treatedcycles.Nosignificanttreatmentbytimeinteractionwaspresent(p=0.98).

137
Appendix2.ValidationprocessforIL-6ELISAkit
ThevalidationprocessfortheEquineIL-6DuoSet®ELISA(R&DSystems™,Minneapolis,
MN, USA) was based on manufacturer instructions (Spike, Recovery, and Linearity & ELISA
DevelopmentGuidebyR&Dsystems).Aspikedneatcytokinesolution(A)waspreparedusingthe
cytokinestandardtoaconcentrationof45ng/ml (approximately10timestherecommended
highstandardofthestandardcurve).
Threebasesamplesof1000µLwereprepared:unspiked,spikedandcontrol.Theunspiked
sampleconsistedonlyof1000µLsamplematerial.Thespikedsampleconsistedof20µLofspiked
neatcytokinesolution(A)and980µLofsamplesolution.Thecontrolsamplehad20µLofspiked
neatcytokinesolution(A)and980µLofreagentdiluent(1%BSAinPBS),whichwasprovidedby
themanufacturer.
Serialdilutionsweremadefromtheneatsample:1:2,1:4and1:8withreagentdiluent.
Positiveandnegativecontrolsampleswereincludedinthevalidationprocess.Positivecontrols
consistedofstudymares(PBIEsusceptible)withexcessivefluid,allowingformultiplevalidation
attempts.Negativecontrolsconsistedofendometrialfluidobtainedbylow-volumelavagefrom
normalmares.Whileournegativecontrolsarenotidealduetodilutionofendometrialsamples
inorder tocollect fluid, therewasnoothermethodtocollectendometrial fluid fromnormal
maresthathavenointra-uterinefluid.
Percentrecoveryandlinearityofsamplesduringvalidationrunswerecalculatedasper
theSpike,RecoveryandLinearity&ELISADevelopmentGuideprovidedby themanufacturer
(R&D Systems Inc.) Methodology was validated when percent recovery and linearity of all
samplesfellwithin80-120%.

138
Severalvalidationrunswererequired. Thepositivecontrolmaresamples (spikedand
unspiked)wererunatdifferentdilutionfactors(1:20,1:30and1:50).The1:20dilutionfactor
allowed for percent recovery and linearity values to fall within 80-120%, validating this kit.
Samplesforthestudywerethereforeconductedata1:20dilutionwithreagentdiluent.
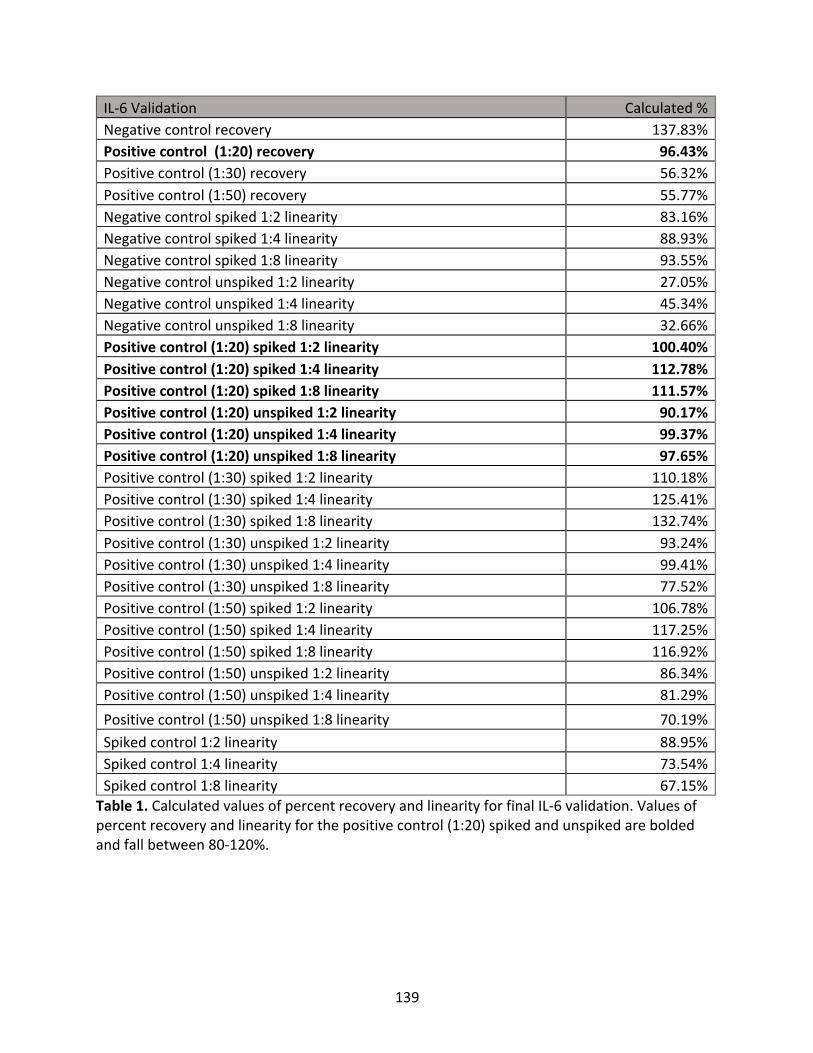
139
IL-6Validation Calculated%Negativecontrolrecovery 137.83%Positivecontrol(1:20)recovery 96.43%Positivecontrol(1:30)recovery 56.32%Positivecontrol(1:50)recovery 55.77%Negativecontrolspiked1:2linearity 83.16%Negativecontrolspiked1:4linearity 88.93%Negativecontrolspiked1:8linearity 93.55%Negativecontrolunspiked1:2linearity 27.05%Negativecontrolunspiked1:4linearity 45.34%Negativecontrolunspiked1:8linearity 32.66%Positivecontrol(1:20)spiked1:2linearity 100.40%Positivecontrol(1:20)spiked1:4linearity 112.78%Positivecontrol(1:20)spiked1:8linearity 111.57%Positivecontrol(1:20)unspiked1:2linearity 90.17%Positivecontrol(1:20)unspiked1:4linearity 99.37%Positivecontrol(1:20)unspiked1:8linearity 97.65%Positivecontrol(1:30)spiked1:2linearity 110.18%Positivecontrol(1:30)spiked1:4linearity 125.41%Positivecontrol(1:30)spiked1:8linearity 132.74%Positivecontrol(1:30)unspiked1:2linearity 93.24%Positivecontrol(1:30)unspiked1:4linearity 99.41%Positivecontrol(1:30)unspiked1:8linearity 77.52%Positivecontrol(1:50)spiked1:2linearity 106.78%Positivecontrol(1:50)spiked1:4linearity 117.25%Positivecontrol(1:50)spiked1:8linearity 116.92%Positivecontrol(1:50)unspiked1:2linearity 86.34%Positivecontrol(1:50)unspiked1:4linearity 81.29%Positivecontrol(1:50)unspiked1:8linearity 70.19%Spikedcontrol1:2linearity 88.95%Spikedcontrol1:4linearity 73.54%Spikedcontrol1:8linearity 67.15%Table1.CalculatedvaluesofpercentrecoveryandlinearityforfinalIL-6validation.Valuesofpercentrecoveryandlinearityforthepositivecontrol(1:20)spikedandunspikedareboldedandfallbetween80-120%.
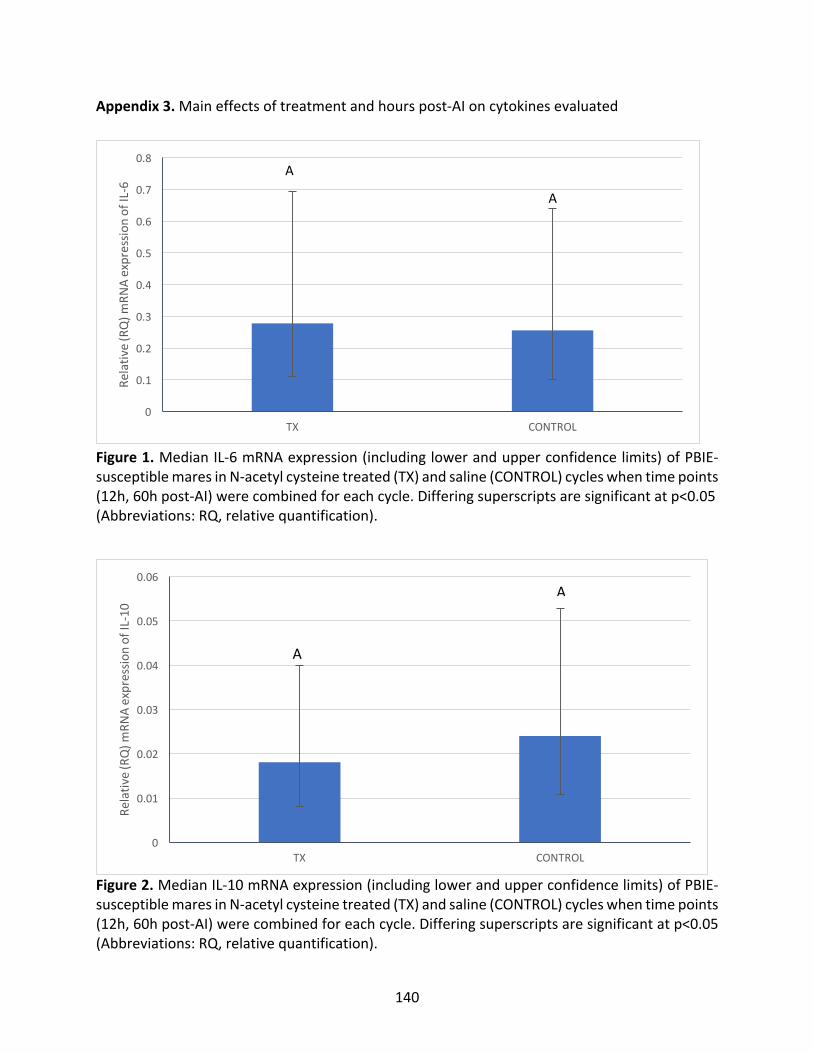
140
Appendix3.Maineffectsoftreatmentandhourspost-AIoncytokinesevaluated
Figure1.MedianIL-6mRNAexpression(includinglowerandupperconfidencelimits)ofPBIE-susceptiblemaresinN-acetylcysteinetreated(TX)andsaline(CONTROL)cycleswhentimepoints(12h,60hpost-AI)werecombinedforeachcycle.Differingsuperscriptsaresignificantatp<0.05(Abbreviations:RQ,relativequantification).
Figure2.MedianIL-10mRNAexpression(includinglowerandupperconfidencelimits)ofPBIE-susceptiblemaresinN-acetylcysteinetreated(TX)andsaline(CONTROL)cycleswhentimepoints(12h,60hpost-AI)werecombinedforeachcycle.Differingsuperscriptsaresignificantatp<0.05(Abbreviations:RQ,relativequantification).
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
TX CONTROL
Relativ
e(RQ)m
RNAexpressio
nofIL-6
A
A
0
0.01
0.02
0.03
0.04
0.05
0.06
TX CONTROL
Relativ
e(RQ)m
RNAexpressio
nofIL-10
A
A
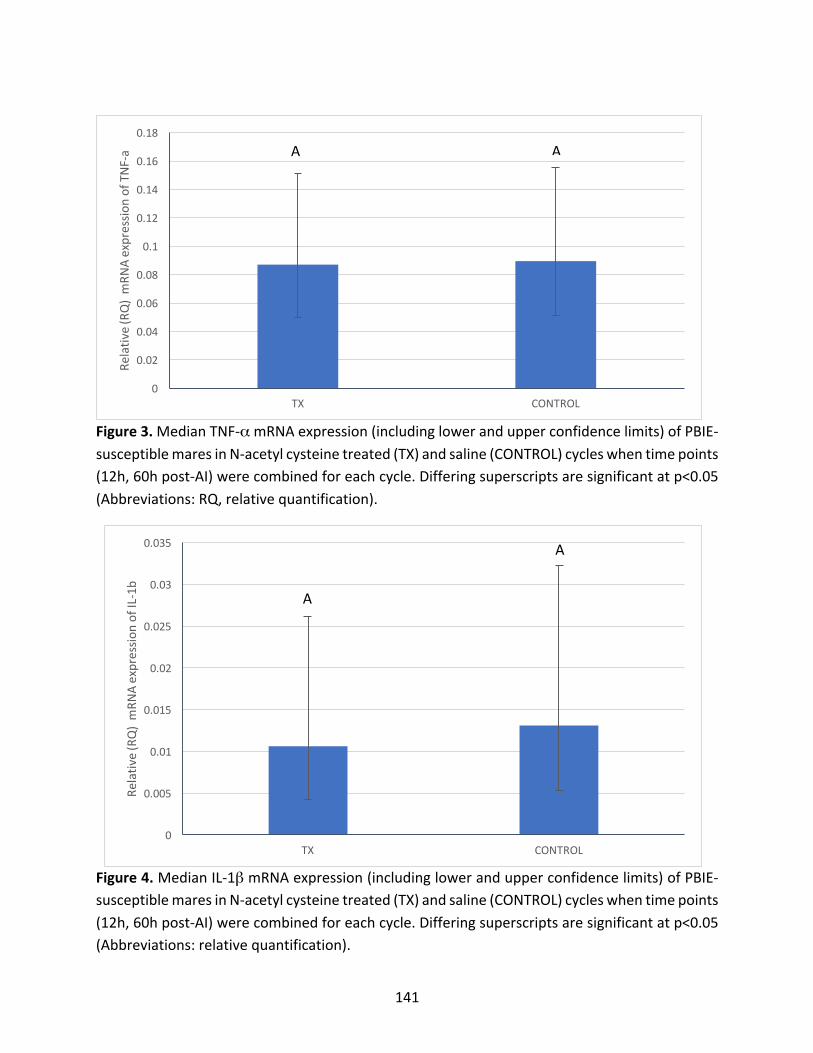
141
Figure3.MedianTNF-αmRNAexpression(includinglowerandupperconfidencelimits)ofPBIE-susceptiblemaresinN-acetylcysteinetreated(TX)andsaline(CONTROL)cycleswhentimepoints(12h,60hpost-AI)werecombinedforeachcycle.Differingsuperscriptsaresignificantatp<0.05(Abbreviations:RQ,relativequantification).
Figure4.MedianIL-1βmRNAexpression(includinglowerandupperconfidencelimits)ofPBIE-susceptiblemaresinN-acetylcysteinetreated(TX)andsaline(CONTROL)cycleswhentimepoints(12h,60hpost-AI)werecombinedforeachcycle.Differingsuperscriptsaresignificantatp<0.05(Abbreviations:relativequantification).
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
0.18
TX CONTROL
Relativ
e(RQ)mRN
Aexpressio
nofTNF-a A A
0
0.005
0.01
0.015
0.02
0.025
0.03
0.035
TX CONTROL
Relativ
e(RQ)mRN
Aexpressio
nofIL-1b
A
A
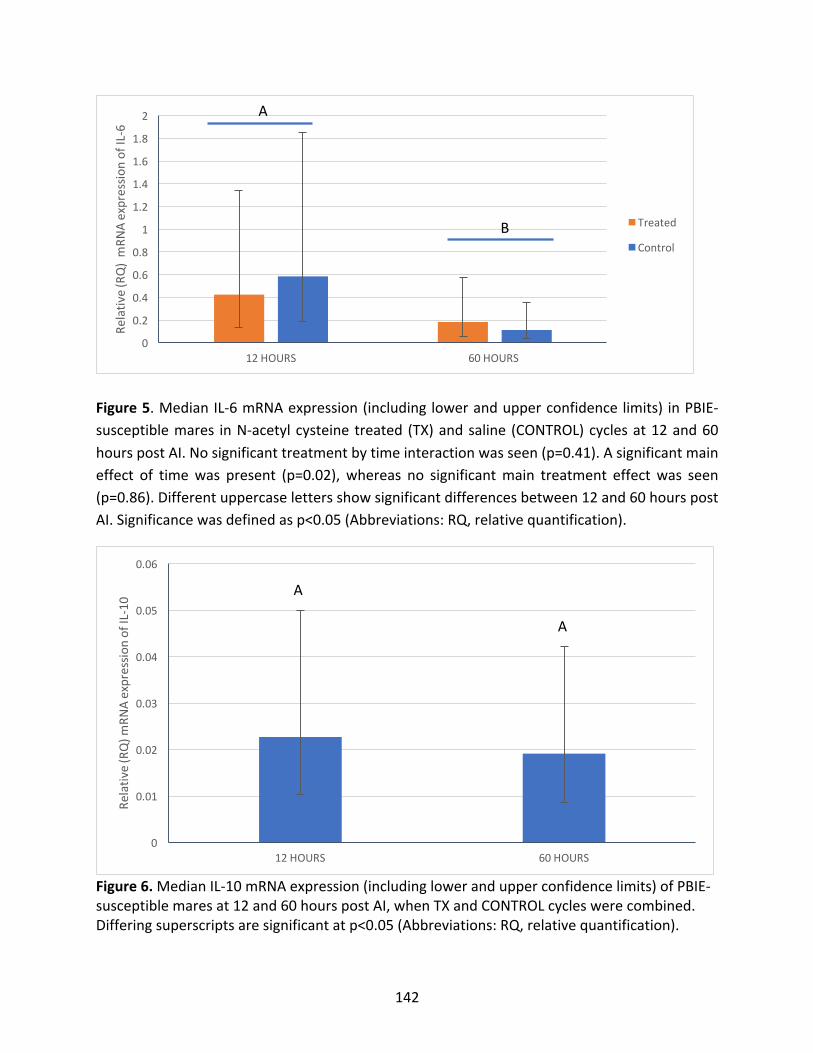
142
Figure5.MedianIL-6mRNAexpression(includinglowerandupperconfidencelimits) inPBIE-susceptiblemares inN-acetylcysteinetreated (TX)andsaline (CONTROL)cyclesat12and60hourspostAI.Nosignificanttreatmentbytimeinteractionwasseen(p=0.41).Asignificantmaineffect of timewas present (p=0.02), whereas no significantmain treatment effectwas seen(p=0.86).Differentuppercaselettersshowsignificantdifferencesbetween12and60hourspostAI.Significancewasdefinedasp<0.05(Abbreviations:RQ,relativequantification).
Figure6.MedianIL-10mRNAexpression(includinglowerandupperconfidencelimits)ofPBIE-susceptiblemaresat12and60hourspostAI,whenTXandCONTROLcycleswerecombined.Differingsuperscriptsaresignificantatp<0.05(Abbreviations:RQ,relativequantification).
0
0.01
0.02
0.03
0.04
0.05
0.06
12HOURS 60HOURS
Relativ
e(RQ)m
RNAexpressio
nofIL-10
A
A
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
12HOURS 60HOURS
Relativ
e(RQ)mRN
Aexpressio
nofIL-6
Treated
ControlB
A
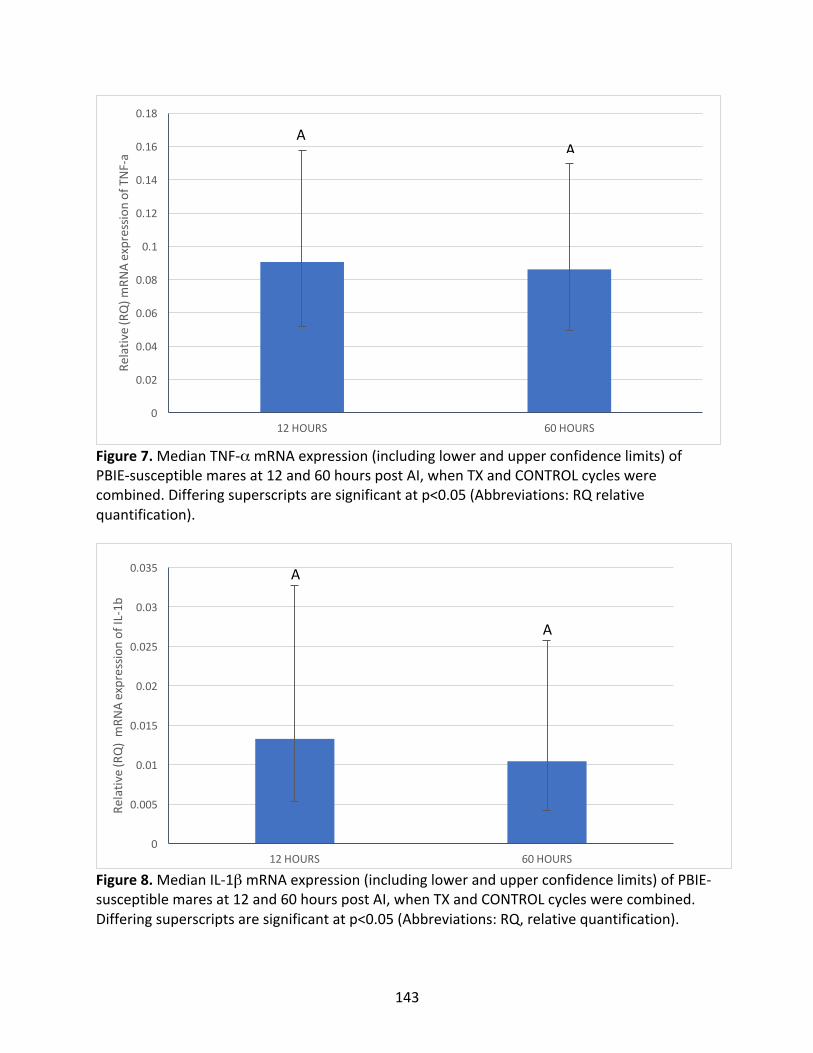
143
Figure7.MedianTNF-αmRNAexpression(includinglowerandupperconfidencelimits)ofPBIE-susceptiblemaresat12and60hourspostAI,whenTXandCONTROLcycleswerecombined.Differingsuperscriptsaresignificantatp<0.05(Abbreviations:RQrelativequantification).
Figure8.MedianIL-1βmRNAexpression(includinglowerandupperconfidencelimits)ofPBIE-susceptiblemaresat12and60hourspostAI,whenTXandCONTROLcycleswerecombined.Differingsuperscriptsaresignificantatp<0.05(Abbreviations:RQ,relativequantification).
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
0.18
12HOURS 60HOURS
Relativ
e(RQ)m
RNAexpressio
nofTNF-a
AA
0
0.005
0.01
0.015
0.02
0.025
0.03
0.035
12HOURS 60HOURS
Relativ
e(RQ)mRN
Aexpressio
nofIL-1b
A
A

144
Appendix4.ProcessesforvalidationattemptsforIL-10,TNF-αandIL-1βELISAkits
ThevalidationattemptsforIL-10,TNF-αandIL-1βELISAkits(R&DSystems)followedthe
sameprinciplesasthatdescribedinAppendix1forIL-6.
Briefly, four validation attemptswere conducted for IL-10. The spiked solution had a
concentrationof92.5ng/mlofIL-10.Forthefirstvalidation,positivecontrolswerediluted1:20,
1:30andneat,basedonsuccessfulresultsfromtheIL-61:20dilutionvalidation.Resultsfellbelow
the standardcurve, therefore in the secondattempt samples froma suspected trulypositive
controlmare(normalmarepost-breeding)wereutilized,yetresultscontinuedtofallbelowthe
standardcurve.Thestandardcurvewasthusshiftedtorangefrom5ng/mlto0.078ng/mlandall
neat positive control samples were utilized. However, the IL-10 kit was never successfully
validated.
For TNF-α, four validation attempts were conducted. The spiked solution had a
concentration of 37.5ng/ml of TNF-α.The first validation consisted of neat, 1:20 and 1:30
dilutionsofpositivecontrols.Resultsweremostpromisingwithdilutionsnear1:4,resultingin
three other validation attempts of 1:4 dilutions with different positive controls being used.
Attemptswereunsuccessful.
For IL-1β, one single validation attempt was conducted. The spiked solution had a
concentrationof45ng/mlofIL-1β.Thepositivecontrolmarewasdiluted1:10,1:20and1:30for
validation.Thiswasunsuccessful.Nootherattemptscouldbepursuedassamplesweredepleted.





























