Embed Size (px)
Citation preview

J. exp. Biol. (1981), 9a, 173-186JVith 8 figures
Printed in Great Britain
Na+ FLUXES ACROSSISOLATED PERFUSED GILLS OF THECHINESE CRAB ERIOCHEIR SINENSIS
BY A. PEQUEUX AND R. GILLES
Laboratory of Animal Physiology, University of LiSge,22 Quai Van Beneden, jB-4020 LUge, Belgium
{Received 5 August 1980)
SUMMARY
Sodium transport processes in the branchial epithelium of euryhalinecrustaceans have been investigated using a perfused preparation of gillsisolated from Chinese crabs Eriocheir sinensis acclimated to dilute (FW)and to concentrated (SW) media.
The results clearly establish the existence of a functional differencebetween the different pairs of branchiae with respect to their participationin the regulation of the blood Na+ content.
In FW-acclimated animals, the Na+ active uptake which counter-balances the salt loss along the concentration gradient is mostly achievedacross the three posterior pairs of gills. Conversely, the Na+ fluxes measuredin the three anterior paiis are essentially passive and carrier-mediated.
Further characterization of the Na+ uptake system present in the posteriorgills by means of inhibitors like ouabain and ethacrynic acid indicates theexistence of at least two spatially separated components of the Na+ carryingsystem.
It is shown that NH4+ may be used as co-ion for Na+ but that such a
coupling can only account for a very small part of the Na+ actively trans-ported inward. The existence of an electrogenic mechanism or of anothercoupled system has thus to be postulated but remains at present a matterof speculation.
To study FW-to-SW and SW-to-FW acclimation, Na+ fluxes weremeasured in isolated gills of SW-acclimated crabs and of FW crabs perfusedand incubated in SW conditions.
During acclimation to SW the Na+ active uptake in the posterior gillsis abolished primarily as a result of inhibition of the Na+ carrier activity.
INTRODUCTION
It is well established that the salt loss by euryhaline crustaceans when in dilutedmedia is counterbalanced by an active absorption of Na+ and Cl~ occurring essentiallythrough the gill tissue (for review, see for instance Krogh, 1939; Potts & Parry,1964; Gilles, 1975, 1979; Kirschner, 1979; Evans, 1980). Nevertheless, the infor-mation about the Na+ movements effectively taking place at that level is very scanty,most experiments having indeed been carried out on whole animals.

174 A. PEQUEUX AND R. GILLES
In an attempt to provide more complete information about the part played b jthe gills in the ionic anisosmotic regulation of crustacean body fluids, we recentlyperfected a perfused preparation of isolated gills from the euryhaline Chinese crabEriocheir sinensis. Preliminary results obtained with this preparation (Pe'queux &Gilles, 1978 a, b) show important differences in extracellular space, tissue and intra-cellular ionic contents as well as in transepithelial potential of the so-called 'anterior'and 'posterior' gills. The data indicate that the 'posterior' gills may show physio-logical specialization in iono-regulation. The purpose of the present work has beento test this hypothesis and to characterize the Na+ movements in both types of gills.
MATERIAL AND METHODSAnimals
Experiments were performed on gills isolated from Chinese crabs Eriocheir sinensisacclimated in the laboratory either to fresh water or to sea water of the followingNa+, K+ and Cl~ concentrations (m-equiv/1). FW: 0-63 Na+, 0-07 K+, 1-07 Ch; SW:474 Na+, 11 K-\ 3 i8 C1-.
Gills were cut off at their base and prepared for perfusion according to a methodalready described (Pe'queux & Gilles, 1978a). The so-called 'anterior' and 'posterior'gills correspond respectively to the three most anterior and the three most posteriorlocated pairs of large gills in the branchial chamber.
Salines
Gills isolated from FW-acclimated crabs were handled in and perfused by a'perfusion FW saline' corresponding to the blood composition of FW-acclimatedanimals. This saline contained 240 mM NaCl, 5 mM KC1, 5 mM MgCl2 and 12-5 mMCaClj. The pH was adjusted to 7-6, borate buffer (9 mM). Gills from SW crabs wereperfused by a 'perfusion SW saline' which was twice as concentrated. The pH waskept at 7-6 with borate buffer (9 mM).
The perfused gills from FW crabs were incubated at 22 °C in an ' incubationsaline' obtained by diluting the perfusion FW saline 250 times while keepingconstant its pH and buffer concentration. According to the experimental scheme,NaCl was added to this medium up to the required concentration. As far as SWcrabs are concerned, incubations were run either in a saline identical to the perfusionSW saline, or in the 'incubation saline' described above.
Transepithelial potential
Potential differences across the gill epithelium were measured by means of aKeithley electrometer and two Calomel electrodes respectively connected to theincubation and to the perfusion salines with agar-KCl bridges.
M./va+, uNa+ flux measurements
Na+ inward and outward movements were estimated by adding MNa+ (0-25 /iCi/ml)to the incubation saline or perfusion saline and by measuring its appearance on theother side in 0-25 or 1 ml samples; samples were collected each 15 min for 60-90 mi>

Na+ fluxes in Eriocheir sinensis 175400 -
n = 6
n= 12
Anterior gills
• • - Influx- 0 - 0 - Efflux
[Na+] in: 240 mM
100 200 300
[Na+] out mMFig. 1. Relation between external Na+ concentration (abscissa), Na+ influx and efflux(ordinate) in perfused anterior gills isolated from FW-acclimated Chinese crabs. Mean ofn experiments ± standard deviation (s.D.).
Sample radioactivity was measured with a y scintillator. At the end of the incubationperiod, the gills were blotted on filter paper and weighed. Flux measurements wereexpressed as /iequiv Na+/g wet wt.h.
Simultaneous influx-efflux experiments were performed according to the double-tracer method using "Na* and MNa+ and following the above described scheme.
NHt+ measurements
NH4+ concentrations and effluxes were determined by colorimetry according toNessler's reaction, using the ready-to-use Sigma ammonia Color reagent (Sigma14-2). Fluxes were expressed in /tequiv NH4+/g wet wt.h.
RESULTS
A. Na+ fluxes in isolated gills from FW-accUmated crabs
Fig. 1 gives Na+ influx and efflux data obtained with anterior gills and measuredas a function of the Na+ concentration in the incubation saline. The fluxes are ofthe same order of magnitude at each external Na+ level tested except for concentrationslower than 25 m-equiv/1. At these low concentrations the efflux becomes significantlylarger than the influx.
The situation is quite different in the posterior gills. In this tissue, the Na+ effluxremains undetectable, whatever the Na+ concentration in the incubation saline (range•om 0-96 to 250 m-equiv/1). In contrast, increasing external Na+ concentration

i 7 6 A. PEQUEUX AND R. GILLES
300 - i
Posterior gills
[Na+]in: 240 mM
« 200
<= 100 -
Na+ influx [fiequiv/g wet wt . h) "'
0050-
0025-
n = 6
fmtx '• 331 jiequiv/g wet wt . hA'm: 13-7 ITIM/1
0-5 1 [Na+] out
0r
100200 300
[Na+] out mMFig. z. Relation between external Na+ concentration (abscissa) and Na+ influx (ordinate)in perfused posterior gills isolated from FW-acclimated Chinese crabs. Mean values ofn experiments ± standard deviation (S.D.). The small figure shows the calculation of the/„„! and Km values of the carrier for the external Na+, derived from the linear regressionplot. (Correlation coefficient, 09866; abscissa, inverse of external Na+ concentration;ordinate, inverse of Na+ influx measurements.)
leads to an increase in Na+ influx. As in the case of the anterior gills, this processshows saturation kinetics with a maximum influx value reached for an outside Na+content of less than 50 m-equiv/1 (Fig. 2). The results allow calculation of an apparentKm for the Na"̂ carrier of 13-7 mM, with a maximum influx/max of 331 /iequiv/g wetwt.h (fig. 2).
B. Nature of the Na+ fluxes
(1) Comparison of measured and calculated flux ratio
From the results described above, it is clear that the influx recorded in the posteriorgills of FW-acclimated crabs must be of an active nature; no detectable efflux canbe shown in these gills, whatever the amplitude of the Na+ gradient applied acrossthe epithelium.
The nature of the fluxes observed in the anterior gills has been studied by double-tracer experiments following Ussing's views, according to which the passive natureof the Na+ movements can be established when the measured ratio of fluxes is equalto the ratio calculated from the relation
JoutJin
Na+inNa+out '
As shown in Table r, there is no significant difference between the measured andcalculated ratio of fluxes, whatever the Na+ gradient applied across the epithelium.The Na+ movements in the anterior gills appear thus to be essentially passive andonly governed by both concentration and electrical gradients.

Na+ fluxes in Eriocheir sinensis
Table i. Experimental and calculated ratio of fluxes in perfusedpreparation of anterior gills isolated from FW-acclimated Chinese crabs
177
FNa+ii
Out
IO
25
1 0 0
250
• / 1
A
In
240
240
240
240
Ratio
24
9-6
2-4
0-96
Na+
Influx Outfluxfiequiv/g wet wt. h
6936124-16178-584593826073481-063205134956
3493625301175-17453-5'227-0541476271-1832321
PD (raV,22 °C)
- S o *- 5 8- 5 2-48-5-22-5- 1 6
-2-5— I
RatioJ1
of fluxesoutfin"
(measured) (calculated)
503720380-98109870871086208460925
3-20723241-184136209701-26008680-922
• Sign refers to the internal medium.
0 15 45 75 0 15 45 75 105Time (min)
Fig. 3. Effects of ouabain io~* M on the Na+ influx of perfused anterior (A) and posterior (B)gills isolated from FW-acclimated Chinese crabs. Ouabain is applied in the outside (opencircles) or inside (black circles) medium at 45 min (arrow).
(2) Effect of inhibitors of the Na+ active transport
The effects of ouabain io~3 M on the transepithelial influx of Na+ have been testedon both the anterior and posterior gills from FW-acclimated crabs. The cardiacglycoside remains without any significant effect on the anterior gills. Fig. 3 showsthat ouabain severely lowers the Na+ influx of the posterior gills when added to theincubation saline only. Addition of the drug to the perfusion saline has no significanteffect.
Ethacrynic acid io~3 M has also been used in an attempt to identify other com-punents of the active Na+ influx in the posterior gills which could be insensitive
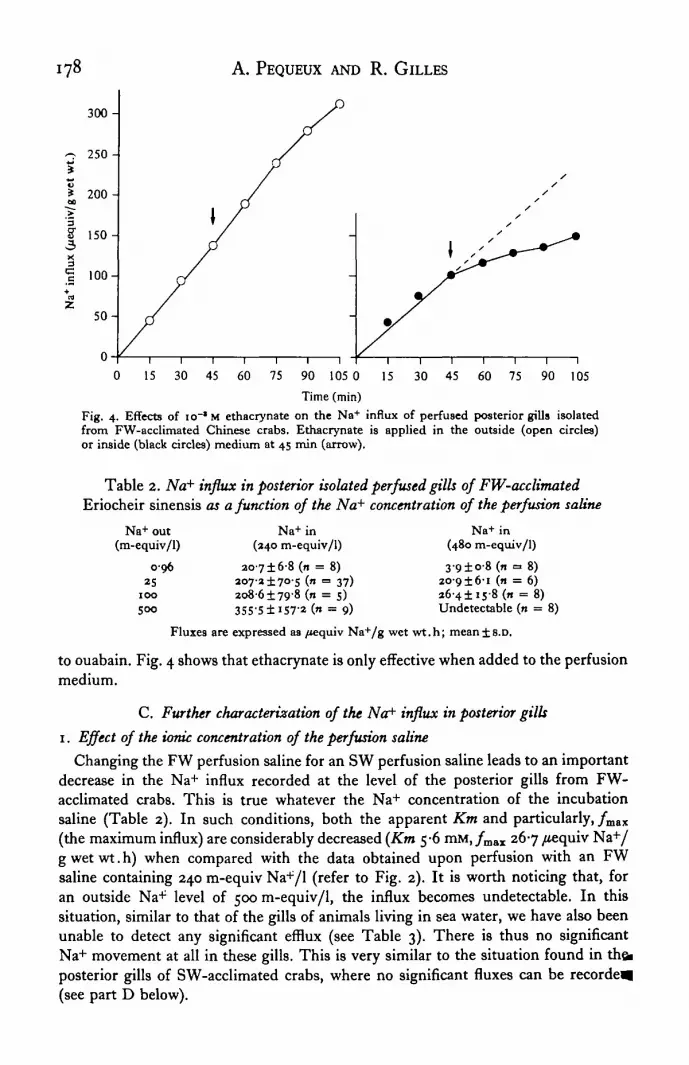
i78 A. PEQUEUX AND R. GILLES
300-
o 250 -
* 200
|
s150 -
100-
50-
0 15 30 45I
60
• ,-:
75l l
90 105 0 15 30 45 60 75 90 105
Time (min)
Fig. 4. Effects of IO-*M ethacrynate on the Na+ influx of perfused posterior gills isolatedfrom FW-acclimated Chinese crabs. Ethacrynate is applied in the outside (open circles)or inside (black circles) medium at 45 min (arrow).
Table 2. Na+ influx in posterior isolated perfused gills of FW-acclimatedEriocheir sinensis as a function of the Na+ concentration of the perfusion saline
Na+ out(m-equiv/1)
0-9625
IOO
5°o
Fluxes are expressed as /tequiv Na+/g wet wt.h; mean±s.D
Na+in(340 m-equiv/1)
207 ±6-8 (n = 8)307-2 ±70-5 (n = 37)2086±798 (n = 5)3S5'5 ± IS7-2 (» = 9)
Na+ in(480 m-equiv/1)
39 ± 0 8 (n = 8)2O-9±6-i (n = 6)264± 15-8 (n = 8)Undetectable (n = 8)
to ouabain. Fig. 4 shows that ethacrynate is only effective when added to the perfusionmedium.
C. Further characterization of the Na+ influx in posterior gills
1. Effect of the ionic concentration of the perfusion saline
Changing the FW perfusion saline for an SW perfusion saline leads to an importantdecrease in the Na+ influx recorded at the level of the posterior gills from FW-acclimated crabs. This is true whatever the Na+ concentration of the incubationsaline (Table 2). In such conditions, both the apparent Km and particularly, fmBX
(the maximum influx) are considerably decreased (Km 5-6 mM,/max 267 fiequiv Na+/g wet wt. h) when compared with the data obtained upon perfusion with an FWsaline containing 240 m-equiv Na+/1 (refer to Fig. 2). It is worth noticing that, foran outside Na+ level of 500 m-equiv/1, the influx becomes undetectable. In thissituation, similar to that of the gills of animals living in sea water, we have also beenunable to detect any significant efflux (see Table 3). There is thus no significantNa+ movement at all in these gills. This is very similar to the situation found in thaiposterior gills of SW-acclimated crabs, where no significant fluxes can be recorded(see part D below).

Na+ fluxes in Eriocheir sinensis 179
Table 3. Na+ fluxes in isolated perfused gills of SW-acclimated Eriocheir sinensis
as a function of the Na+ concentration of per fusion and incubation media
(Comparison with flux values obtained with gills fromFW-acclimated crabs incubated in SW conditions.)
SWCrabFWCrab
SWCrabFWCrabSWCrabSWCrabSW CrabSW CrabSW CrabSWCrab
Na+ (m-equiv/1)
Out
500500
500Soo1 0 0
«S0-96
1 0 0
250-96
In
480480
480480480480480240340240
Tnflur T?fflnv11111UJL u m i 1 n
(/tequiv/g wet wt. h)A. Anterior gills
34481960 (n = 3)635 ±195 (n =• 4)
B. Posterior gills65-0±75-8 (n = 12)Undetectable84±8-8 (n = 11)S'9±i3-2 (n = 18)
o-O3±o-o8 (n •= 12)5'S±8-3(n = 12)
i o 9 ± i 3 3 (n = 11)O'8±i-o (n = 12)
Mean ±8.1/. (n experiments).
2079 ±632 (n628±175 (n
UndetectableUndetectable——————
= 8)= 4)
x:
i
3
3X
i
.5
200 -
Na+
Posterior gills
[Na + ] in :240mM
[NH4+]in:
NH4+
I100
I200
I300
[Na+] out (mM)Fig. 5. Relation between external Na+ concentration (abscissa), Na+ influx and NH4"1" efflux(ordlnate) in perfused posterior gills isolated from FW-acclimated Chine»e crabs, the perfusionFW saline being free of added NH4+.
(2) Na* influx and ammonia efflux
In other communications (P6queux & Gilles, 19786; Gilles, 1978), we brieflyreported on a dependency of the ammonia efflux in posterior gills on the Na+ levelin the incubation saline, as well as on the fact that NH4+ can be a very good substitutefor K+ in stimulating the activity of the Na+/K+ ATPase extracted from these•ills. Such results are in agreement with the idea of a coupling between Na+ and
movements. This possibility is explored further in this section.
Fig. 5 confirms that the ammonia efflux in posterior gills depends on the

i8o A. PEQUEUX AND R. GILLES
Na* influx (^equiv/g wet wt . h)
50 100 150 200 250J I I i
300
Control
NH4C1, 10"3
Control
NH4Q, 5 . 10-*
Control
NH4C1, 10"4
B (
Control
NH4Q,5 . 10-*
Control
Proline 10"J
) 101
1-
1 i 1
I 1
1—
1
- i
20 30l l
hi
•H
Fig. 6. Effect of NH4C1 and proline on the Na+ influx of perfused posterior gills isolatedfrom FW-acclimated Chinese crabs. NH4C1 and proline are added to the perfusion saline(concentrations in mol/1). Results are mean values±S.D. A, incubation medium: artificialFW + 25 m-equiv Na+/1. B, incubation medium: artificial FW.
level in the incubation saline. However, there is a considerable discrepancy betweenthe magnitudes of Na+ influx and NH4+ efflux at any external Na+ level we havestudied. Even at an external Na+ concentration giving maximum NH4+ efflux, therecorded Na+ influx remains some 10 times larger. On the other hand, addition ofNH4C1 up to io~3 M to the perfusion saline fails to induce any significant modificationof the Na+ influx (Fig. 6). In the same way, addition of large amounts of proline,considered as a potent NH4+ donor in the gills after deamination, remains withoutsignificant effect on the Na+ influx (Fig. 6).
In another approach to a possible Na+/NH4+ coupling, the effects of ouabain andethacrynic acid on the ammonia efflux have been studied. As shown in Fig. 7,ouabain, shown to lower the Na^ influx when added to the incubation medium(section B, 2), induces a decrease in the efflux of ammonia when added on thatside of the epithelium. Conversely, ethacrynate when added to the perfusion salinefails to induce any significant effect on the NH4+ efflux while, as we have previouslyseen, it severely decreases the Na+ influx in the same conditions.

Na+ fluxes in Eriocheir sinensis 181.300 -
5 200 -
IK
I 100
30 - i
20 -
I3
I io H
Na+NH 4
+
I IFig. 7. Effects of ouabain io~a M (outside) and ethacrynate io"3 M (inside) on Na+ influxand NH4
+ efflux of posterior perfused gills isolated from FW-acclimated Chinese crabs.Flux data are expressed as /tequiv/g wet wt.h and represent mean values ±s.D.
D. Na+ fluxes in isolated gills from SW-acclimated crabs
The Na+ fluxes in gills of SW crabs incubated and perfused with salines havingNa+ levels similar to those found respectively in acclimation medium and blood arereported in Table 3. These results show that both influx and efflux are very largein the anterior gills. It is worth noticing that relatively high values can also berecorded in anterior gills from FW-acclimated crabs in similar conditions (Table 3).
In posterior gills, on the contrary, no significant Na+ efflux can be detected. As faras the Na+ influx is concerned, the limited specific activity of the incubation salinewe could prepare and the limited volume of the sampling allowed by the perfusiontechnique result in a low accuracy of measurement and thus in large standarddeviations. The obtained flux values can hardly be considered as statistically sig-nificant, however.
It is moreover interesting to notice here that the Na+ influx measured in posteriorgills of SW-acclimated crabs is not significant - not only at the high external Na+level corresponding to a normal SW medium (Table 3) but also at all the othermuch lower outside levels of Na+ we have tested (0-96, 25 and 100 m-equiv/1).Similarly, the Na+ influx in these gills remains not significant when the SW perfusionsaline is replaced by an FW one (Na+: 480 m-equiv/1 to Na+: 240 m-equiv/1 -Table 3).

182 A. PEQUEUX AND R. GILLES
DISCUSSION
Koch et al. (1954) suggested that the anterior and posterior gills of the euryhalinecrab Eriocheir sinensis may differ in their ability to transport ions. Further argumentscorroborating this view have been presented in only a few other studies (King &Schoffeniels, 1969; Schoffeniels & Gillies, 1970; Pequeux & Gilles, 1978a). Theresults of this paper clearly establish the existence of a functional difference betweenthese pairs of branchiae with respect to their participation in the blood osmoregulationthat this species is able to achieve when in diluted media. It must be rememberedthat the Chinese crab Eriocheir sinensis is a typical hyperosmoregulator, which canmaintain a pronounced blood hyperosmotic state when in diluted medium, andwhose blood remains isosmotic to the external medium when the animal is acclimatedto sea water (Berger, 1931; Conklin & Krogh, 1938; Krogh, 1939; Schoffeniels &Gilles, 1970). In the hyperosmoregulators, active Na+ uptake at the gill level appearsto be an essential mechanism in counterbalancing the salt loss in diluted media. Ourresults show that in E. sinensis the active Na+ influx is mostly achieved by the threeposterior pairs of gills. Considerations based on the comparison of the measuredratio of fluxes with those calculated according to the Ussing's equation for passiveion movements (Ussing, 1949), lead indeed to the conclusion that the fluxes ofNa+ in the anterior gills are essentially passive. The study of the Na+ fluxes as afunction of the external Na+ level in these gills further reveals saturation kineticsindicating that the movements of Na+, though passive, are carrier-mediated andnot the result of a simple diffusional process. It is also worth noticing that the Na+
efflux decreases as the external Na+ concentration decreases, in spite of the factthat the Na+ gradient across the epithelium is increasing: this can be explained byconsidering that the permeability of the anterior gills decreases at low external Na+
levels. Such a decrease in permeability has also been considered in fish gills (Maetz,1971; Kirschner, 1979) and is of interest, since it should decrease the salt lossoccurring in the animals acclimating to low salinities.
As far as the posterior gills are concerned we failed to demonstrate any significantNa+ efflux whatever the Na+ gradient applied across the epithelium. This showsthat the Na+ permeability of these branchiae is extremely low. Consequently, thelarge Na+ influx which can be measured must be of an active nature. On the otherhand, the magnitude of the Na+ influx is dependent on the external concentrationof Na+, revealing saturation kinetics which allows calculation of an apparent K^for the transepithelial Na+ carrying system(s) of 137 m-equiv/1. This value is ratherclose to the one (20 min) reported by Shaw (1961) when studying Na^ uptake inwhole specimens of another marine euryhaline decapod Carcinus maenas, but ishigh when compared to those available for freshwater crustaceans (0-2 mM Astacusfluviatilis, Shaw, 1959a; o-i mM, Potamon niloticus, Shaw, 19596; 0-15 mM, Gam'marus pulex, Shaw & Sutcliffe, 1961). It must however be considered that the/m a x
which can be calculated for these marine euryhaline decapods is higher than theone which can be measured in freshwater species (331 /iequiv/g wet wt.h E. sinensis,this study; iomM/kg.h, C. maenas, Shaw, 1961; 0-15 mM/kg.h Astacus fluviatilis,Shaw, 1959a). It may thus be that the rather low affinity of the carrier for Naifound in the marine euryhaline crustaceans is, at least partly, compensated by a higqnumber of pumping sites.

Na+ fluxes in Eriocheir sinensis 183
The study of the effects of ouabain and ethacrynic acid, inhibitors of two differentcomponents of Na+ transport in cell membranes, allows some further characterizationof the Na+ uptake system present in the posterior gills. It is interesting to considerthat the effect of these agents is dependent on the side of the epithelium on whichit is applied. Ouabain is effective only when added outside, in the incubation saline,while ethacrynate appears to be active only from the internal side of the epithelium.These results can be considered as indicating the existence of, at least, a two-component Na+ carrying system. One of these (the ouabain-sensitive one) would berestricted to the outside-facing membrane and would be linked to the activity ofan ATPase sensitive to this cardiac glycoside. Inhibition by ouabain is classicallyconsidered to be indicative of an Na+/K+ coupled carrier system. Such a couplingis however unlikely in our preparation, for uptake of Na+ in such conditions wouldimplicate loss of K+ to the environmental medium. Evidence has been provided inrecent years showing that NH4+ is a very good substitute for K+ in stimulating theouabain-sensitive ATPase in crustacean gill tissues (Towle, 1974; Mangum et al.1976; Pe"queux & Gilles, 1977; Gilles, 1978). The effect of ouabain on the Na+ influxin our preparation could thus be considered as an effect on an ATPase-mediatedNa+/NH4
+ coupled transport located on the outward-facing membranes of theepithelium. The fact that ouabain also inhibits the efflux of ammonia in the posteriorgills is in agreement with this idea. On the other hand, the obtained results do notexclude the possibility of the existence of other Na+ transport mechanisms on theexternal membrane. This point will be considered later on in the discussion. It isinteresting to consider here that ouabain does not induce any significant inhibitionof the transepithelial Na+ uptake when added to the perfusion saline, while ethacrynicacid is only effective when added to that saline. This might indicate that there is noNa+/K+ transport system on the inside-facing membrane of the epithelium. In thisview, the expulsion of Na+ from the cells would be achieved through another,ethacrynate-sensitive, Na"̂ transport system. This is however rather unlikely in thecontext of the regulation of the intracellular level of both K+ and Na+, which mustinvolve control of the movements of these ions between blood and the intracellularfluid. Another possibility is that the transepithelial Na+ uptake is achieved throughpathways different from those implicated in the expulsion of Na+ normally diffusingfrom blood to cells following its concentration gradient. In this view, a Na+/K+
coupled, ouabain-sensitive transport could be related to the expulsion from the cellsof the Na+ diffusing from the blood, while another pumping mechanism, ethacrynate-sensitive, would be involved in driving the Na+ implicated in the transepithelialmovement. In such a system, one could consider that the Na+ implicated in thetransepithelial movement is largely moving through a tissue pool different from thecellular one controlled by the Na+/K+ coupled transport. It is more likely that thecontribution of the Na+/K+ coupled, ouabain-sensitive transport to the total effluxfrom the cells is relatively small when compared to that of the transepithelial trans-port system. Both hypotheses may nevertheless account for the fact that ouabainhas no significant effect on the Na+ influx when added to the inside medium.Experiments are in progress in this laboratory to bring more light on this problem,
et us now come back to the uptake system located at the external side of the. As already discussed, the fact that the transepithelial flux of Na+ decreases

184 A. PEQUEUX AND R. GILLES
Incubation medium Gill Epithelium Blood
Na* 1 m-equiv/1 Na* 240 m-equiv/l
0
Ethacrynic acid
Fig. 8. Model of Na+ fluxes across the posterior gill epithelium of the Chinese crab E. sinenxisacclimated to diluted media. Details are described in the text. CA*: carbonic anhydrase.
when ouabain is added to the incubation medium led us to consider the possibilityof an Na+/NH4
+ coupled transport. The existence of such a process has alreadybeen postulated on the basis that it would account for the increase in ammoniaoutput observed in euryhaline crustaceans upon acclimation to diluted media (Shaw,i960; Gilles, 1975, 1978; Mangum et al. 1976). The fact that, in our preparation, theammonia efflux depended on the external level of Na+ is in agreement with the ideaof such a coupling. Various results, however, prompt us to minimize the importanceof this mechanism. As a matter of fact, though both Na+ influx and NH4
+ effluxdepended on the external Na+ level, the nature of this dependency is quite differentfor each of the ionic species considered (see Fig. 5). Furthermore, the Na+ influx ismuch larger than the ammonia efflux. This might be explained on the basis of anNa+:NH4+ stoichiometry that is not 1:1. The important difference between thefluxes, however, makes this hypothesis unlikely, and it is more reasonable to assumethat a large part of the active Na+ uptake is not coupled to NH4
+ excretion. Thisconclusion is further substantiated by results showing that increasing the concen-tration of NH4C1 in the perfusion saline fails to induce any significant modificationof the Na+ influx. Therefore, one has to consider that there is an Na+ pump on theexternal side of the epithelium, either working according to an electrogenic schemeor requiring a counter-ion different from NH4
+. In this view, an Na+/H+ coupledsystem, as postulated by different authors (Ehrenfeld, 1974; Evans, 1980), wouldappear as a good candidate. Actually, both possibilities remain essentially a matterof speculation and need further investigation. The main conclusions up to now Msummarized in Fig. 8.

Na+ fluxes in Eriocheir sinensis 185
Let us now consider the fluxes recorded in gills of SW-acclimated E. sinensis andof FW-acclimated ones perfused with salines, the Na+ level of which is similarto the one found in the blood of SW animals. These results may lead to a betterunderstanding of the processes involved in the FW-to-SW or SW-to-FW acclimation.
As shown in Table 3, the fluxes of Na+ reach a very high level in the anteriorgills of SW-acclimated animals while, as we have seen previously, they are muchlower in the FW-acclimated crabs and dependent on the external concentration ofNa+, a decrease in external Na+ resulting in a decrease in both influx and efflux.It thus appears that the passive, carrier-mediated fluxes of Na+ in the anterior gillsresult in high Na+ turnover when the animal is in SW and much lower exchange ofNa+ when in FW.
On the other hand, increasing the Na+ concentration in the perfusion saline ofposterior gills from FW animals to a level similar to that found in the blood of SWanimals leads to an immediate decrease in the Na+ influx, whatever the concentrationof Na+ in the incubation medium. As shown in Tables 2 and 3, the influx falls toundetectable values even when the concentration of Na+ in the incubation salinereaches a level similar to that of SW. In such conditions of high Na+ in both perfusionand incubation salines, the fluxes of Na+ in the posterior gills of SW-acclimatedcrabs are also barely significant. Acclimation to SW appears thus to result in animportant decrease, even in the virtually complete abolition of the active Na+uptake in the posterior gills; such an effect seems to be mainly related to the increasein Na+ blood level. From the important and instantaneous drop in /m a x observedupon perfusion of posterior gills from FW crabs with an SW perfusion saline it canbe reasonably concluded that the decrease in Na+ uptake activity might be mainlydue to a rapid decrease in the number of active carrier sites. This decrease beingextremely fast, it can be considered as resulting primarily from an inhibition of theNa+ carrier(s) activity rather than from a decrease in the synthesis of carrier(s)molecules. Whether or not such a repression of carrier(s) molecules synthesis mayoccur in long-term acclimation to sea water remains to be demonstrated. Experimentsare in progress in this laboratory in order to bring more insight to these questions.
This work has been supported by grants 'Credit aux Chercheurs' from the FNRSto the authors and by a grant no. 2.4511.76 from the FRFC to R. Gilles.
REFERENCES
BERCER, A. (1931). Uber die Anpassung eines Susswasserund eines Brackwasserkrebses an Medienvon verschiedenem Salzgehalt. PflUgert Arch. ges. Physiol. aa8, 790-807.
COUKLIN, R. & KROGH, A. (1938). A note on the osmotic behaviour of Eriocheir in concentrated andMytilui in dilute sea water. Z. vergl. Physiol. 26, 239—241.
EHRKNFELD, J. (1974). Aspects of ionic transport mechanisms in crayfish Astacus leptodactylus. J. exp.Biol. 61, 57-70.
EVANS, D. H. (1980). Salt transport mechanisms in branchial epithelia. In Animals and EnvironmentalFitneis, vol. 1 (ed. R. Gilles), pp. 61-78. Oxford-New York: Pergamon Press.
GILLES, R. (1975). Mechanisms of ion and osmoregulation. In Marine Ecology, vol. 2, part 1 (ed.O. Kinne), pp. 250-347.
GILLES, R. (1978). Intracellular free amino acids and cell volume regulation during osmotic stresses.In Osmotic and Volume Regulation - A. Benzon Symposium XI (ed. C. Barker Jergensen andE. Skadhauge), pp. 470-491. Copenhagen: Munksgaard.ILLES, R. (1979). Intracellular organic osmotic effectors. In Mechanisms of Osmoregulation in Animals(ed. R. Gilles), pp. m-153 . Chichester-New York: Wiley-Interscience.

186 A. PEQUEUX AND R. GILLES
K I N G , E. N. & SCHOFFENIELS, E. (1969). ' I n vitro' preparation of crab gill for use in ion transp^studies. Archs int. Phytiol. Biochim. 77, 105-m.
KIRSCHNER, L. B. (1979). Control mechanisms in crustaceans and fishes. In Mechanism! of Osmo-regulation in Animals (ed. R. Gilles), pp. 157—222. Chichester-New York: Wiley-Interscience.
KOCH, H. J., EVANS, J. & SCHICKS, E. (1954). The active absorption of ions by the isolated gills ofthe crab Eriocheir sinensis (M.Edw.). Meded. Vlaam. Acad. K. Wet. 16, no. 5, 3-16.
KROGH, A. (1939). Osmotic Regulation in Aquatic Animals. Cambridge University Press. 242 pp.MAETZ, J. (1971). Fish gills: mechanisms of salt transfer in fresh water and sea water. Phil. Trans.
R. Soc. B a6a, 209-249.MANGUM, C. P., SILVBRTHORN, S. V., HARRIS, J. L., TOWLE, D. W. & KRALL, A. R. (1976). The
relationship between blood pH, ammonia excretion and adaptation to low salinity in the blue crabCallinectes sapidus. J. exp. Zoo!. 195, 129-136.
PEQUEUX, A. & GILLES, R. (1977). Osmoregulation of the Chinese crab Eriocheir sinensis as related tothe activity of the (Na+ + K+) ATPase. Archs int. Physiol. Biochim. 85, 426-428.
PEQUEUX, A. & GILLES, R. (1978a). Osmoregulation of the euryhaline Chinese crab Eriocheir sinensis.Ionic transport across isolated perfused gills as related to the salinity of the environment. Proceedingsof the 12th EMBS, Stirling, Scotland. Physiology and Behaviour of marine organisms, pp. 105-111.Oxford-New York: Pergamon Press.
PEQUEUX, A. & GILLES, R. (19786). Na+/NH4+ co-transport in isolated perfused gills of the Chinese
crab Eriocheir sinensis acclimated to fresh water. Experientia 34, 1593-1594.POTTS, W. T. W. & PARRY, G. (1964). Osmotic and Ionic Regulation in Animals, pp. 423. London:
Pergamon Press.SCHOFFENIELS, E. & GILLES, R. (1970). Osmoregulation in aquatic arthropods. In Chemical Zoology,
vol. v, part A (ed. M. Florkin and B. T. Scheer), pp. 255-286. New York: Academic Press.SHAW, J. (1959a). The absorption of sodium ions by the crayfish Astacus pallipes Lereboullet. I. The
effect of external and internal sodium concentrations. J. exp. Biol. 36, 126-144.SHAW, J. (19596). Salt and water balance in the East African freshwater crab, Potamon niloticus
(M.Edw.). J. exp. Biol. 36, 157-176.SHAW, J. (i960). The absorption of sodium ions by the crayfish. II. The effect of external anion.
J. exp. Biol. 37, 534-547-SHAW, J. (1961). Studies on the ionic regulation in Carcinus maenas L. I. Sodium balance. J. exp. Biol.
38, 135-153-SHAW, J. & SUTCLIFFE, D. W. (1061). Studies on sodium balance in Gammarus duebeni Lilljeborg and
G. pulex pulex (L.). J. exp. Biol. 38, 1-15.TOWLE, D. W. (1974). Equivalence of gill (Na++ K+)ATPase from blue crabs acclimated to high
and low salinity. Am. Zool. 14, 1259.USSING, H. H. (1949). The distinction by means of tracers between active transport and diffusion.
Acta physiol. scand. 19, 43-56.















![[Possible existence of tetrodotoxin-sensitive, potential-dependent chlorine conductivity in the membrane of perfused cardiomyocytes]](https://img.pdfslide.net/doc/110x75/6348b53f031992cdcf022cb8/possible-existence-of-tetrodotoxin-sensitive-potential-dependent-chlorine-conductivity.jpg)



![Investigation of factors affecting fluorometric quantitation of cytosolic [Ca2+] in perfused hearts](https://img.pdfslide.net/doc/110x75/634ddcabd38be601b805ed3b/investigation-of-factors-affecting-fluorometric-quantitation-of-cytosolic-ca2.jpg)









