Embed Size (px)
Citation preview

Nanos downregulates transcription and modulates CTD phosphorylation
in the soma of early Drosophila embryos
Girish Deshpande, Gretchen Calhoun, Timothy M. Jinks1,Alexandros D. Polydorides2, Paul Schedl*
Department of Molecular Biology, Princeton University, Princeton, NJ 08540, USA
Received 3 August 2004; received in revised form 17 December 2004; accepted 17 December 2004
Available online 26 January 2005
Abstract
nanos (nos) specifies posterior development in the Drosophila embryo by repressing the translation of maternal hb mRNA. In addition to
this somatic function, nos is required in the germline progenitors, the pole cells, to establish transcriptional quiescence. We have previously
reported that nos is required to keep the Sex-lethal establishment promoter, Sxl-Pe, off in the germline of both sexes. We show here that nos
also functions to repress Sxl-Pe activity in the surrounding soma. Sxl-Pe is inappropriately activated in the soma of male embryos from nos
mothers, while Sxl-Pe can be repressed in female embryos by ectopic Nos protein. nos appears to play a global role in repressing transcription
in the soma as the effects of nos on promoter activity are correlated with changes in the phosphorylation status of the carboxy terminal
domain (CTD) repeats of the large RNA polymerase II subunit. Finally, we present evidence indicating that the suppression of transcription
in the soma by Nos protein is important for normal embryonic development.
q 2004 Elsevier Ireland Ltd. All rights reserved.
Keywords: nanos; C-terminal domain (CTD); CTD phosphorylation; Sex-lethal; Transcriptional repression
1. Introduction
Abdominal fate in the Drosophila embryo is specified by
nanos (nos) mRNA localized in the posterior pole plasm
(St Johnston, 1993). Translation of this mRNA generates a
Nos gradient that extends anteriorly (Gavis and Lehmann,
1992). The Nos gradient represses the translation of
uniformly distributed maternal Hunchback (Hb) mRNA
(Wharton and Struhl, 1991; Wang and Lehmann, 1991).
This posterior repression, together with the transcriptional
activation of the hb gene by Bicoid in the anterior, generates
a Hb gradient at the anterior (Struhl et al., 1992; Gavis and
Lehmann, 1992). Translational repression by Nos requires
Pumilio (Pum), and it is thought that Pum interacts with Nos
0925-4773/$ - see front matter q 2004 Elsevier Ireland Ltd. All rights reserved.
doi:10.1016/j.mod.2004.12.009
* Corresponding author. Tel.: C1 609 258 4979; fax: C1 609 258 1028.
E-mail address: [email protected] (P. Schedl).1 Present address: Division of Developmental Neurobiology, National
Institute for Medical Research, Mill Hill, London NW7 1AA, UK.2 Present address: Laboratory of Molecular Neuro-oncology, Rockefeller
University, New York, NY, USA.
Response Elements (NREs) in the hb mRNA 3 0UTR
(Murata and Wharton, 1995). In the absence of either Nos
or Pum, Hb is translated from maternal mRNA in the
posterior and blocks abdominal segmentation.
In addition to its role in the soma, nos is required for the
proper development of the germline (Kobayashi et al., 1996;
Forbes and Lehmann, 1998). Pole cells in embryos from nos
mutant mothers (referred to as either nosmK embryos or pole
cells) are not correctly determined. Unlike wild type, newly
formed nosmK pole cells fail to downregulate transcription
and attenuate the cell cycle (Asaoka et al., 1998, 1999;
Deshpande et al., 1999). Later in embryogenesis they
exhibit a range of migration defects and fail to associate
with the somatic gonadal precursor cells.
One gene that is inappropriately expressed in nosmK pole
cells is Sex-lethal (Sxl) (Deshpande et al., 1999). Sxl
controls somatic sexual development, and is switched on in
females but not males (Cline and Meyer, 1996). During
most of development on/off regulation is post-transcrip-
tional; however, when sexual identity is determined Sxl is
regulated at the transcriptional level. Sexual identity is
Mechanisms of Development 122 (2005) 645–657
www.elsevier.com/locate/modo

G. Deshpande et al. / Mechanisms of Development 122 (2005) 645–657646
chosen at the blastoderm stage by counting the
X chromosome to autosome ratio. Counting depends upon
the expression of X-linked numerators at a level pro-
portional to their dose. The known numerators include the
transcription factors scute (sc), sisterless-a (sis-a) and runt
(rt), and the JAK/STAT ligand, unpaired (upd) (Cline and
Meyer, 1996). In 2X/2A (female) nuclei, the concentration
of numerators is sufficient to activate the Sxl-Pe establish-
ment promoter (Keyes et al., 1992). In contrast, in 1X/2A
nuclei (male), these positive factors are unable to overcome
the negative effects of autosomal denominators and Sxl-Pe
remains off. The Sxl proteins produced by the Sxl-Pe
transcripts in 2X/2A embryos activate the Sxl autoregula-
tory feedback loop by directing the female splicing of the
first transcripts from the Sxl-Pm maintenance promoter.
Translation of these female spliced Sxl-Pm mRNAs
produces Sxl proteins which autoregulate their own
expression and ensure a commitment to female identity
during the remainder of development. In males, the Sxl-Pm
pre-mRNAs are spliced in the non-productive default
pattern ensuring a commitment to male identity. While Sxl
is activated in XX soma at the blastoderm stage, it is not
turned on in the germline until much later in development
(Horabin et al., 1995). Moreover, though germline acti-
vation is sex specific, the mechanism is distinct from that in
the soma (Cline and Meyer, 1996). In fact, the available
evidence suggests that Sxl-Pe is not active in the germline.
Consequently, it is surprising that Sxl-Pe is turned on in
nosmK pole cells at the blastoderm stage in both sexes.
The ectopic activation of Sxl-Pe in nosmK pole cells
raises the question of what role, if any, the nos gene plays in
somatic sex-determination and more generally in regulating
somatic transcription. We now show that nos functions to
repress Sxl-Pe activity in the soma and that the Sxl-Pe
promoter is inappropriately turned on in the somatic 1X/2A
nuclei of nosmK embryos while it is upregulated in 2X/2A
nosmK embryos. The effects of nos on Sxl-Pe appear to be
due to a general upregulation in RNA polymerase II activity
in nosmK embryos. Transcription by RNA polymerase II is
correlated with the phosphorylation of serine residues ser2
and ser5 in the 7 amino acid (aa) repeats in the CTD domain
of the largest subunit. These two serines are thought to be
phosphorylated sequentially in the transcriptional cycle
(Dahmus, l996; Komarnitsky et al., 2000). Ser5 phos-
phorylation occurs first and is correlated with promoter
clearance, while ser2 phosphorylation occurs after clearance
and is correlated with elongation. Studies in Caenorhabditis
elegans indicate that high levels of phospho-ser2 and ser5
are present in transcriptionally active somatic nuclei. In
contrast, in quiescent germline nuclei there is little phospho-
ser2, and although polymerase with phospho-ser5 is
detected, there is much less than in the soma. In pie-1
mutants, which are unable to silence germ cell transcription,
the levels of phospho-ser2 and ser5 approach that in somatic
nuclei (Mello et al., 1996; Seydoux et al., 1996; Seydoux
and Dunn, 1997). We show here that ser2 and ser5
phosphorylation is upregulated in the germline and soma
of nosmK embryos, while both modifications can be
downregulated in the soma by ectopic Nos expression.
2. Results
2.1. Sxl-Pe is ectopically activated in the soma of nos
mutant 1X/2A embryos
In wild type, newly formed pole cells shut down RNA
polymerase II and remain transcriptionally inactive until
midway through embryogenesis (Seydoux and Dunn, 1997;
Van Doren et al., 1998). Maternally derived Nos is required
to establish/maintain this transcriptional quiescence. In
embryos from nosK mothers genes like even-skipped (eve)
and fushi tarzu (ftz) that are normally active only in the soma
are inappropriately expressed in pole cells at the syncytial
blastoderm stage. Though pole cells have the highest
concentration of Nos, there is a Nos protein gradient
extending to near the middle of the syncytial blastoderm
embryo. Consequently, the inhibitory effects of Nos in the
germline suggest that it may also suppresses transcription in
the soma.
Consistent with this idea we found that both eve and ftz
are inappropriately expressed in the posterior soma of
nosmK blastoderm embryos (not shown). However, since
translation of maternal hb mRNA drastically alters seg-
mentation in nosmK embryos, the activation of eve and ftz
could be due to excess Hb protein rather than to a failure in
downregulating transcription. Since hb does not have a
positive role in Sxl-Pe regulation (not shown) we examined
Sxl expression in the soma of nosmK embryos. In wild-type
blastoderm embryos Sxl expression is driven from the Sxl-
Pe promoter. As illustrated in Fig. 1, it directs the
production of Sxl protein in female but not male embryos.
Whereas Sxl is only observed in 50% of wild-type embryos,
it is found in the soma of all blastoderm stage nosm embryos.
This finding indicates that Sxl must be expressed in both
female and male nosmK embryos.
The nosm embryos could nevertheless be divided into two
equal classes based on protein levels. Embryos in the first
class have high levels of Sxl distributed uniformly
throughout the soma (Fig. 1). As embryos in this
class appear to have even more Sxl than wild-type
females, a reasonable presumption is that they correspond
to nosmK females. Embryos in the second class have lower
levels of Sxl than wild-type females (see Fig. 1) and thus are
likely to correspond to nosmK males. The ‘male’ embryo
shown in Fig. 1 has a relatively sharp posterior to anterior
gradient; however, the gradient in other male embryos can
be quite shallow and extend to the very anterior. Never-
theless, even these embryos can be readily distinguished
from wild-type males which have no detectable Sxl, and
from either nosmK ‘females’ or wild-type females which
have considerably more Sxl.

Fig. 1. Sxl protein is ectopically expressed from Sxl-Pe in the soma and pole cells of nosmK embryos. Panels in the top half show syncytial blastoderm embryos
stained with Sxl antibody. 0–4 h old embryos laid by white1 (w1) and nosBN mothers were probed with a monoclonal antibody directed against Sxl and the
staining pattern visualized by standard DAB immunohistochemical detection. As described in the text, wild-type embryos can be divided into two equal classes
based on the Sxl antibody-staining pattern in the soma. Embryos in the first class are darkly stained and previous studies demonstrate that these correspond to
females. Embryos in the second class are unstained. These are male embryos. Embryos from nosBN mothers can also be divided into two equal classes. Embryos
in the first class are darkly stained. As these embryos represent 50% of the population and resemble wild-type female embryos, we presume that they
correspond to nosBN female embryos (see also Fig. 2). Embryos in the second class are lightly stained (and sometimes exhibit a posterior to anterior gradient).
As these embryos also represent 50% of the population and resemble neither wild-type females, nor wild-type males, we believe that they correspond to nosBN
male embryos. To rule out the possibility that the low level of Sxl protein seen in these nosmK ‘male’ embryos is due to background staining, we mixed wild
type and nosmK before staining. As expected two classes of male embryos were observed in the mixture. The first class resembled wild-type males and had no
detectable Sxl. The second class resembled nosmK male embryos and had a low level of Sxl, often distributed in a shallow gradient. Panels in top half: WT
female: w1 female embryo showing a high level of Sxl protein in somatic nuclei and no protein in pole cells. WT male: w1 male embryo lacks Sxl protein. nos
‘female’: nosmK female embryo has Sxl in soma and pole cells. nosK male: nosmK male embryo has a posterior to anterior gradient of Sxl. Bottom panels show
blastoderm embryos stained with b-galactosidase antibody. nosBN or w1 virgin females were mated with males homozygous for Sxl-Pe3.0 kbLac-Z (Keyes et al.,
1992). 0–4 h old embryos were collected and stained with b-galactosidase antibody. WT male: no b-galactosidase is observed in either the soma or pole cells.
The nosmK embryos fell into two classes. The first class resembles wild-type females, and has high levels of b-galactosidase in the soma. However, unlike wild
type, b-galactosidase is also observed in the pole cell of these embryos. Based on their similarity to wild-type females and the fact that w50% of the nosmK
embryos fall into this class, we believe that these embryos are female. Embryos in the second class differ from wild type of either sex in that there is a low level
of b-galactosidase in both germline and soma. As observed for Sxl, there is often a posterior gradient. Since these embryos also represent 50% of the population
they are likely to correspond to nosmK males. Two nosmK male embryos are shown.
G. Deshpande et al. / Mechanisms of Development 122 (2005) 645–657 647
Since Nos is known to repress translation, the Sxl protein
in male progeny of nosK mothers could arise from the
translation of maternally derived Sxl mRNA rather than
from the activation of Sxl-Pe. To demonstrate that Sxl-Pe is
inappropriately turned on, we crossed nosBN females with
males carrying the full length Sxl-Pe3.0 kbLac-Z reporter.
In wild-type embryos, the Sxl-Pe reporter mimics
the endogenous Sxl-Pe promoter, driving b-galactosidase
expression in 2X/2A but not 1X/2A animals (Estes et al.,
l995; Table 1A; see also WT male in Fig. 1). In contrast,
since b-galactosidase can be detected in nearly 100% of
the nosmK embryos (Table 1A), Sxl-Pe3.0 kb must be
activated in both sexes. As is the case for Sxl protein, two
different b-galactosidase expression patterns are observed.

Table 1A
Sxl-Pe3.0 kb
Staining
Maternal genotype Dark Med.–
light
Negative Total
nosBN 64 (57%) 43 (38%) 5 (4%) 112
w1 156 (45%) 194 (55%) 350
FRT hbFBnosBN 47 (39%) 66 (56%) 8 (7%) 121
P-nos; FRT
hbFBnosBN
29 (47%) 7 (11%) 26 (42%) 62
nosBN, hbFBnosBN (females carrying germline clones for hbFBnosBN), P-nos;
FRT hbFBnosBN and w1 virgin females were mated with males carrying two
copies of Sxl-Pe3.0 kb transgene. Embryos derived from each cross were
stained for b-galactosidase activity. Based on the intensity of staining
embryos were classified into several categories.
Fig. 2. Expression of Sxl in wild type and nosK embryos. Wild type and
nosBN females were mated to wild-type males carrying an X-linked
ftz:hsp70LacZ transgene (Hagstrom et al., 1996). The UPS enhancer in this
transgene drives b-galactosidase expression in stripes in germ band
extending embryos. In this experiment, only female embryos carry the
ftz:hsp70LacZ transgene. Embryos collected from each cross were stained
with Sxl and b-galactosidase antibodies and examined by confocal
microscopy. Wild-type blastoderm stage embryos could be divided into
two equal classes based on Sxl antibody staining. Embryos in the first class
are female and express Sxl in the soma, but not the pole cells. Embryos in
the second class are male and Sxl cannot be detected. Unlike wild type, Sxl
can be detected in pole cells of all nosmK blastoderms. Based on the amount
of Sxl in the soma, the nosmK embryos can be divided into two equal
classes. Embryos in the first class express very high levels of Sxl protein
and as illustrated in Fig. 2 much greater amounts of Sxl accumulate in these
embryos than in wild-type females. These nosmK embryos are thought to be
female. Embryos in the second class also express Sxl; however, the amount
of Sxl is considerably less than wild type or nosmK ‘females’, and is often
most highly concentrated apically. These nosmK embryos are thought to be
males. The two panels at the bottom show germ band extended wild type
and nosmK embryos. At this stage all of the wild type and nosmK embryos
that had high levels of Sxl also expressed b-galactosidase and hence are
female. Note that as was observed at the blastoderm stage, nosmK female
embryos have much higher levels of Sxl than wild-type females. The
expected number (w50%) of male wild type and nosmK embryos lacking
b-galactosidase were also observed (not shown). While the wild-type males
had no Sxl, many of the nosmK male embryos had some residual Sxl.
G. Deshpande et al. / Mechanisms of Development 122 (2005) 645–657648
In slightly more than half of the blastoderm embryos high
levels of b-galactosidase are present throughout the soma
(see example in Fig. 1 and Table 1A). These nosmK embryos
resemble wild-type females. The remaining embryos have
lower levels of b-galactosidase (Fig. 1 and Table 1A).
To confirm these findings we examined Sxl expression
using confocal microscopy. In the experiment shown in
Fig. 2, males carrying an X-linked ftz(UPS):hsp70-LacZ
transgene (Hagstrom et al., 1996) were crossed to wild type
or nosBN females. The ftz UPS stripe enhancer activates the
hsp70 promoter and b-galactosidase expression can be
detected in gastrulating female embryos; however, it is not
observed prior to gastrulation. As illustrated in Fig. 2, Sxl
protein is present in the soma of wild-type female, but not
male embryos, while it cannot be detected in the pole cells
of either sex. Though Sxl in females localizes in the nucleus,
there are also substantial amounts in the apical and basal
cytoplasm at this stage. Unlike wild type, Sxl is present in
the soma and pole cells of all nosmK embryos. As found
with DAB staining, the nosmK blastoderm embryos could be
divided into two equal classes based on the level and pattern
of Sxl accumulation in the soma. Embryos in ‘Class I’ had
high levels of Sxl protein in the nucleus and surrounding
cytoplasm. Though the distribution of protein in these
embryos is similar to wild-type females, the amount of
Sxl, as judged by antibody staining, is higher (compare wild
type and nosmK female embryos in Fig. 2). Embryos in
‘Class II’ had much lower levels of Sxl than wild-type
females and much of the protein is localized in the apical
cytoplasm (Fig. 2).
We also examined later stages of development when
b-galactosidase expression from the ftz(UPS):hsp70-LacZ
transgene can be reproducibly detected. As illustrated at the
bottom of Fig. 2, for both wild type and nosmK, the germ
band extended embryos that expressed high levels of Sxl
were b-galactosidase positive. Consequently, these Sxl
expressing embryos must be female. Moreover, as was
observed at the blastoderm stage, the level of Sxl protein, as
judge by antibody staining, in older nosmK females is
consistently higher than in older wild-type females.
As expected, the male, b-galactosidase negative, germ
band extended embryos in the wild-type collection had no
Sxl. In the nosmK collection, residual Sxl protein could be
detected in some of the b-galactosidase negative males

Table 1B
Sxl-Pe0.4 kb
Maternal
genotype
Very
dark
Dark Medium Light Negative
nosBN 35 (20%) 35 (20%) 47 (28%) 49 (29%) 2
hbFBnosBN 23 (21%) 31 (29%) 23 (21%) 24 (21%) 5
w1 205 (45%) 0 0 259 (55%)
G. Deshpande et al. / Mechanisms of Development 122 (2005) 645–657 649
(not shown); however, the level of Sxl was considerably
lower than in the blastoderm ‘Class II’ nosmK embryos.
These findings indicate that the amount of Sxl expressed
from Sxl-Pe in nosmK male blastoderm embryos is generally
not sufficient to stably activate the autoregulatory feedback
loop.
2.2. Activation of the minimal promoter, Sxl-Pe0.4 kb
To further localize the nos responsive sequences we
tested a reporter containing the minimal Sxl-Pe promoter,
Sxl-Pe0.4 kb (Estes et al., 1995). While Sxl-Pe0.4 kb
exhibits the appropriate sex-specificity in wild type (see
Table 1B), it drives considerably less b-galactosidase
expression than Sxl-Pe3.0 kb and unlike the larger
promoter whose activity is nearly uniform, Sxl-Pe0.4 kb
is much more active in the anterior than the posterior. As
was the case for Sxl-Pe3.0 kb, Sxl-Pe0.4 kbLac-Z must be
inappropriately turned-on in males in the absence of Nos
function since it is active in nearly 100% of the nosmK
embryos (Fig. 3 and Table 1B). In addition, the level of
b-galactosidase in nosmK ‘female’ embryos is increased
relative to wild type. Notice that the effects of nos on
this promoter are not strictly limited to the posterior of
the embryo.
2.3. Ectopic expression of Nos protein downregulates
Sxl-Pe activity
If Sxl-Pe is upregulated in the absence of Nos, it should
be repressed by excess Nos. To test this prediction we used
nos transgenes in which the nos 3 0UTR is replaced by
a tubulin (tub) or a bicoid (bcd) 3 0UTR. In progeny from
nos-tub 3 0UTR mothers ectopic Nos protein is produced
throughout the embryo, while in progeny from nos-bcd
3 0UTR mothers ectopic Nos is translated from transgene
mRNA localized at the anterior.
We first examined the effects of excess Nos on Sxl-Pe
activity in 2X/2A embryos. Though about 50% of the
blastoderm embryos produced by nos-tub 3 0UTR or nos-bcd
3 0UTR mothers still express Sxl protein, most show striking
abnormalities in the pattern of protein accumulation. In the
case of nos-tub 3 0UTR, Sxl expression is often reduced in
the posterior (see arrows in Fig. 4). However, we also
observe embryos in which there is a general reduction in
Sxl protein throughout or show a patchy pattern of Sxl
protein accumulation several different regions (not shown).
In the case of nos-bcd 3 0UTR, female embryos in which Sxl
protein is reduced in the anterior are often observed (Fig. 4).
These findings argue that ectopic Nos downregulates Sxl-
Pe. Consistent with this idea, Nos from the nos-bcd 3 0UTR
(Fig. 3) or nos-tub 3 0UTR (not shown) transgenes represses
the Sxl-Pe3.0 kb reporter.
Since activation of the normally silent Sxl-Pe in 1X/2A
embryos is one of the most striking effects of reduced Nos,
we wished to determine whether excess Nos downregulates
Sxl-Pe in male embryos as it does in female embryos.
However, this experiment is not possible with the wild type
Sxl-Pe because it is already off in nosC 1X/2A embryos. For
this reason, we took advantage of a ‘gain-of-function’
promoter, Sxl-PeGOF, that has extra numerator binding sites
and is active in both sexes (Jinks et al., 2000; Kramer et al.,
1999). As shown in Fig. 3, Sxl-PeGOF is repressed in 1X/2A
embryos by excess Nos.
2.4. Phosphorylation of the RNA polymerase CTD domain
is altered in nos embryos
The upregulation of transcription in nosmK embryos
suggested that it would be of interest to examine ser2 and
ser5 phosphorylation of the CTD domain since these
modifications of the largest RNA polymerase II subunit
are correlated with transcriptionally engaged polymerases.
As reported by Seydoux and Dunn (1997), phospho-ser2 is
detected in somatic nuclei of wild-type blastoderm embryos,
but not in pole cell nuclei (Fig. 5). This is not due to a
germline specific reduction in the amount of polymerase II
as pole cell nuclei are labeled to the same extent as somatic
nuclei by an antibody that recognizes epitopes in exon 2 of
the large subunit (not shown). Unlike wild type, the
phospho-ser2 CTD modification is readily detected in
nosmK pole cells (Fig. 5, see legend). In stage 4 embryos
roughly 50% of the pole cells are stained have the CTD
phospho-ser2 modification while the number and level
increases in stage 5 embryos. We also probed for the
phospho-ser5 modification (not shown). As observed in the
C. elegans germline, only a low level of phospho-ser5 is
found in wild-type pole cells, while it is elevated in a subset
of nosmK pole cells.
The effects of nos on CTD phosphorylation are not
limited to the germline. The level of phospho-ser2 antibody
staining in somatic nuclei of syncytial blastoderm nosmK
embryos is upregulated compared to syncytial wild-type
embryos stained in parallel (Figs. 5 and 6). This increase is
readily detected in the posterior of the embryo where Nos
protein levels are maximum. Since nosmK and wild-type
embryos are labeled to the same extent by the exon2-
specific polymerase antibody (Fig. 7), the increase in
phospho-ser2 in the soma of nosmK embryos is not due to
differences in the amount of the large subunit. We also
found that phospho-ser5 appears to be increased in nosmK
(not shown).
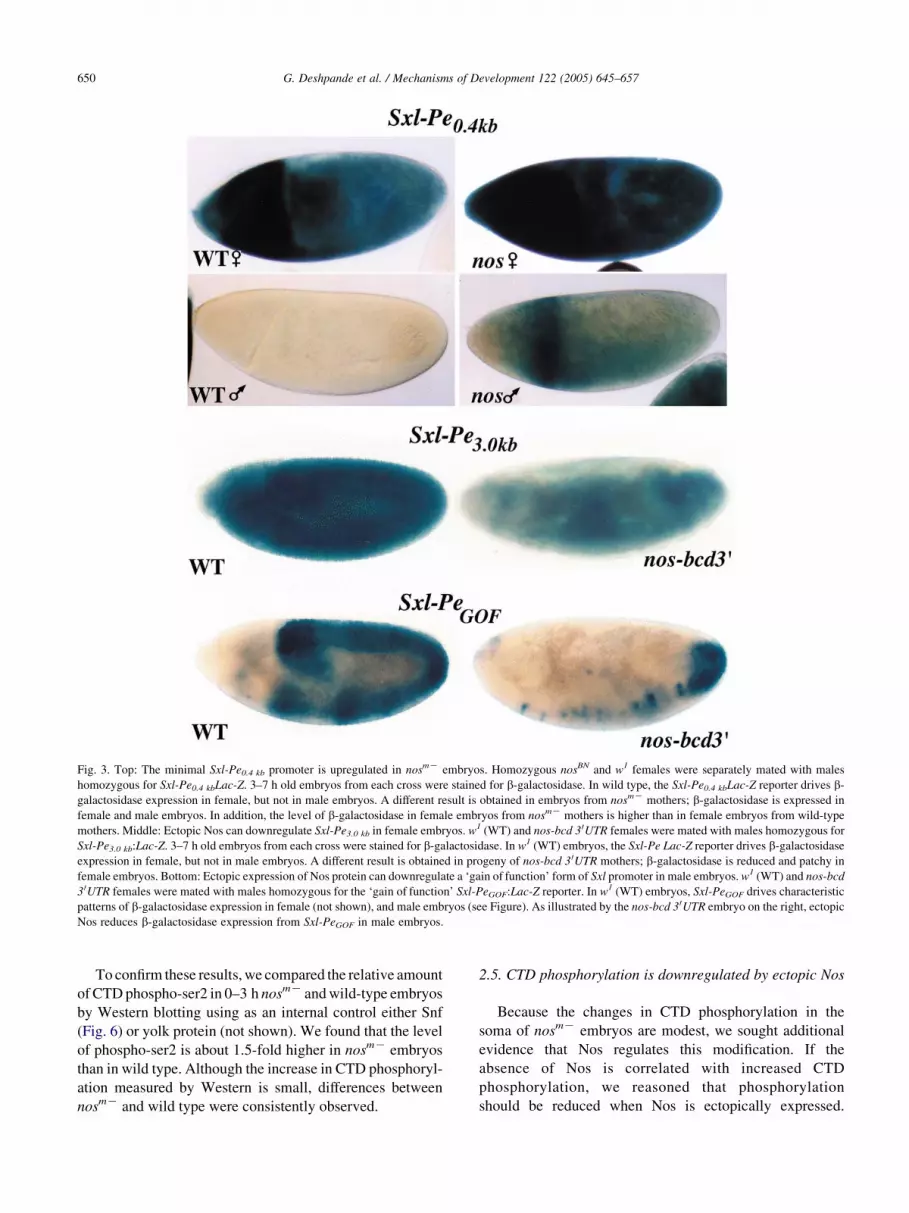
Fig. 3. Top: The minimal Sxl-Pe0.4 kb promoter is upregulated in nosmK embryos. Homozygous nosBN and w1 females were separately mated with males
homozygous for Sxl-Pe0.4 kbLac-Z. 3–7 h old embryos from each cross were stained for b-galactosidase. In wild type, the Sxl-Pe0.4 kbLac-Z reporter drives b-
galactosidase expression in female, but not in male embryos. A different result is obtained in embryos from nosmK mothers; b-galactosidase is expressed in
female and male embryos. In addition, the level of b-galactosidase in female embryos from nosmK mothers is higher than in female embryos from wild-type
mothers. Middle: Ectopic Nos can downregulate Sxl-Pe3.0 kb in female embryos. w1 (WT) and nos-bcd 3 0UTR females were mated with males homozygous for
Sxl-Pe3.0 kb:Lac-Z. 3–7 h old embryos from each cross were stained for b-galactosidase. In w1 (WT) embryos, the Sxl-Pe Lac-Z reporter drives b-galactosidase
expression in female, but not in male embryos. A different result is obtained in progeny of nos-bcd 3 0UTR mothers; b-galactosidase is reduced and patchy in
female embryos. Bottom: Ectopic expression of Nos protein can downregulate a ‘gain of function’ form of Sxl promoter in male embryos. w1 (WT) and nos-bcd
3 0UTR females were mated with males homozygous for the ‘gain of function’ Sxl-PeGOF:Lac-Z reporter. In w1 (WT) embryos, Sxl-PeGOF drives characteristic
patterns of b-galactosidase expression in female (not shown), and male embryos (see Figure). As illustrated by the nos-bcd 3 0UTR embryo on the right, ectopic
Nos reduces b-galactosidase expression from Sxl-PeGOF in male embryos.
G. Deshpande et al. / Mechanisms of Development 122 (2005) 645–657650
To confirm these results, we compared the relative amount
of CTD phospho-ser2 in 0–3 h nosmK and wild-type embryos
by Western blotting using as an internal control either Snf
(Fig. 6) or yolk protein (not shown). We found that the level
of phospho-ser2 is about 1.5-fold higher in nosmK embryos
than in wild type. Although the increase in CTD phosphoryl-
ation measured by Western is small, differences between
nosmK and wild type were consistently observed.
2.5. CTD phosphorylation is downregulated by ectopic Nos
Because the changes in CTD phosphorylation in the
soma of nosmK embryos are modest, we sought additional
evidence that Nos regulates this modification. If the
absence of Nos is correlated with increased CTD
phosphorylation, we reasoned that phosphorylation
should be reduced when Nos is ectopically expressed.
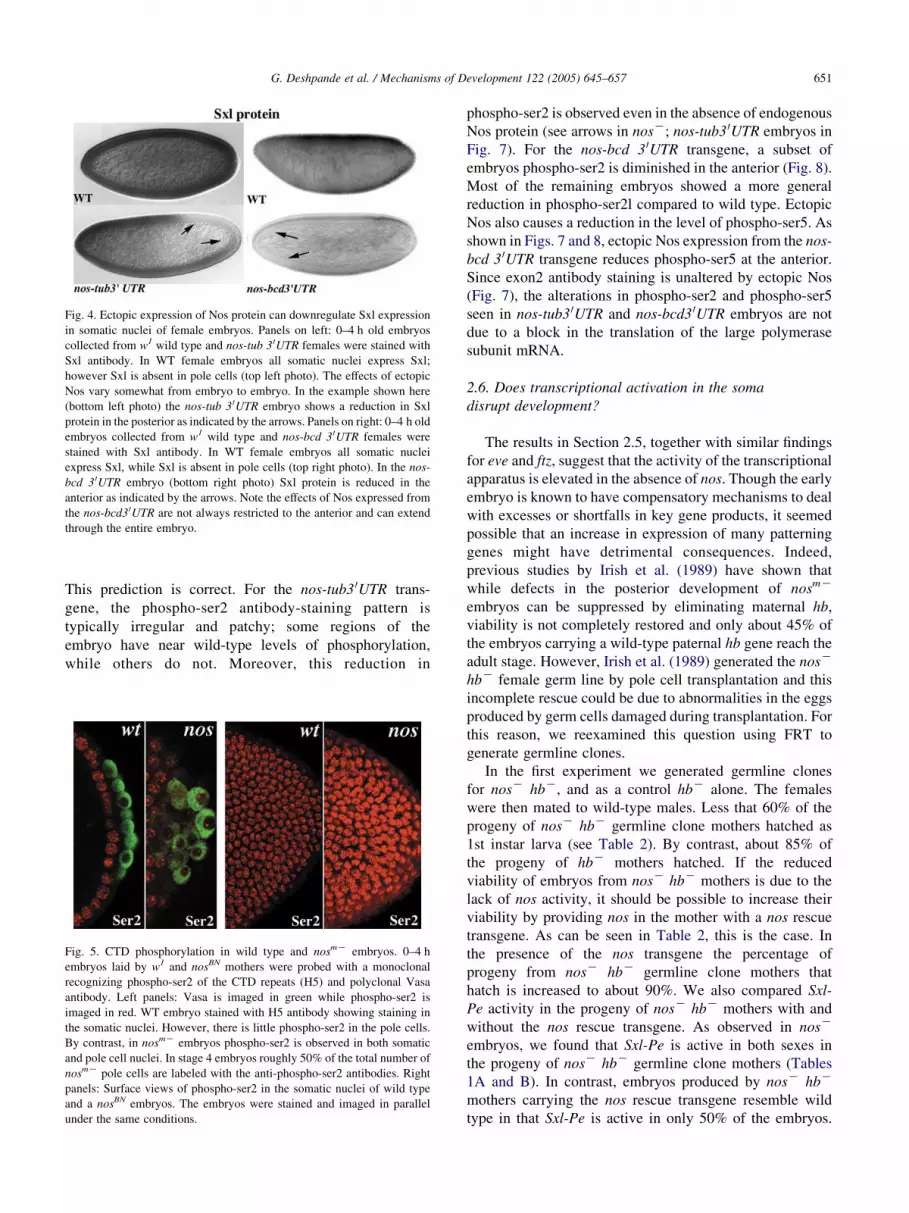
Fig. 4. Ectopic expression of Nos protein can downregulate Sxl expression
in somatic nuclei of female embryos. Panels on left: 0–4 h old embryos
collected from w1 wild type and nos-tub 3 0UTR females were stained with
Sxl antibody. In WT female embryos all somatic nuclei express Sxl;
however Sxl is absent in pole cells (top left photo). The effects of ectopic
Nos vary somewhat from embryo to embryo. In the example shown here
(bottom left photo) the nos-tub 3 0UTR embryo shows a reduction in Sxl
protein in the posterior as indicated by the arrows. Panels on right: 0–4 h old
embryos collected from w1 wild type and nos-bcd 3 0UTR females were
stained with Sxl antibody. In WT female embryos all somatic nuclei
express Sxl, while Sxl is absent in pole cells (top right photo). In the nos-
bcd 3 0UTR embryo (bottom right photo) Sxl protein is reduced in the
anterior as indicated by the arrows. Note the effects of Nos expressed from
the nos-bcd30UTR are not always restricted to the anterior and can extend
through the entire embryo.
G. Deshpande et al. / Mechanisms of Development 122 (2005) 645–657 651
This prediction is correct. For the nos-tub3 0UTR trans-
gene, the phospho-ser2 antibody-staining pattern is
typically irregular and patchy; some regions of the
embryo have near wild-type levels of phosphorylation,
while others do not. Moreover, this reduction in
Fig. 5. CTD phosphorylation in wild type and nosmK embryos. 0–4 h
embryos laid by w1 and nosBN mothers were probed with a monoclonal
recognizing phospho-ser2 of the CTD repeats (H5) and polyclonal Vasa
antibody. Left panels: Vasa is imaged in green while phospho-ser2 is
imaged in red. WT embryo stained with H5 antibody showing staining in
the somatic nuclei. However, there is little phospho-ser2 in the pole cells.
By contrast, in nosmK embryos phospho-ser2 is observed in both somatic
and pole cell nuclei. In stage 4 embryos roughly 50% of the total number of
nosmK pole cells are labeled with the anti-phospho-ser2 antibodies. Right
panels: Surface views of phospho-ser2 in the somatic nuclei of wild type
and a nosBN embryos. The embryos were stained and imaged in parallel
under the same conditions.
phospho-ser2 is observed even in the absence of endogenous
Nos protein (see arrows in nosK; nos-tub3 0UTR embryos in
Fig. 7). For the nos-bcd 3 0UTR transgene, a subset of
embryos phospho-ser2 is diminished in the anterior (Fig. 8).
Most of the remaining embryos showed a more general
reduction in phospho-ser2l compared to wild type. Ectopic
Nos also causes a reduction in the level of phospho-ser5. As
shown in Figs. 7 and 8, ectopic Nos expression from the nos-
bcd 3 0UTR transgene reduces phospho-ser5 at the anterior.
Since exon2 antibody staining is unaltered by ectopic Nos
(Fig. 7), the alterations in phospho-ser2 and phospho-ser5
seen in nos-tub3 0UTR and nos-bcd3 0UTR embryos are not
due to a block in the translation of the large polymerase
subunit mRNA.
2.6. Does transcriptional activation in the soma
disrupt development?
The results in Section 2.5, together with similar findings
for eve and ftz, suggest that the activity of the transcriptional
apparatus is elevated in the absence of nos. Though the early
embryo is known to have compensatory mechanisms to deal
with excesses or shortfalls in key gene products, it seemed
possible that an increase in expression of many patterning
genes might have detrimental consequences. Indeed,
previous studies by Irish et al. (1989) have shown that
while defects in the posterior development of nosmK
embryos can be suppressed by eliminating maternal hb,
viability is not completely restored and only about 45% of
the embryos carrying a wild-type paternal hb gene reach the
adult stage. However, Irish et al. (1989) generated the nosK
hbK female germ line by pole cell transplantation and this
incomplete rescue could be due to abnormalities in the eggs
produced by germ cells damaged during transplantation. For
this reason, we reexamined this question using FRT to
generate germline clones.
In the first experiment we generated germline clones
for nosK hbK, and as a control hbK alone. The females
were then mated to wild-type males. Less that 60% of the
progeny of nosK hbK germline clone mothers hatched as
1st instar larva (see Table 2). By contrast, about 85% of
the progeny of hbK mothers hatched. If the reduced
viability of embryos from nosK hbK mothers is due to the
lack of nos activity, it should be possible to increase their
viability by providing nos in the mother with a nos rescue
transgene. As can be seen in Table 2, this is the case. In
the presence of the nos transgene the percentage of
progeny from nosK hbK germline clone mothers that
hatch is increased to about 90%. We also compared Sxl-
Pe activity in the progeny of nosK hbK mothers with and
without the nos rescue transgene. As observed in nosK
embryos, we found that Sxl-Pe is active in both sexes in
the progeny of nosK hbK germline clone mothers (Tables
1A and B). In contrast, embryos produced by nosK hbK
mothers carrying the nos rescue transgene resemble wild
type in that Sxl-Pe is active in only 50% of the embryos.

Fig. 6. Phosphorylation of serine 2 in CTD repeats of RNA Polymerase II in wild type and nosmK mutant embryos. Panel on left: 0–4 h embryos from w1 and
nosKBN mothers were stained with monoclonal (H5) specific for CTD repeats that are phosphorylated on serine 2. Top: WT embryo probed with the H5
monoclonal. Wild-type embryos show near-uniform staining of somatic nuclei and little or no staining of pole cell nuclei. The two embryos labeled nos are
nosmK embryos probed with H5 antibody. Note that the staining appears to be stronger in the somatic nuclei of nosmK embryos than in the wild-type control.
(H5 staining of somatic nuclei in wild type and nosmK embryos sometimes appears stronger on the dorsal side.) Panels on right: Embryonic extracts prepared
from 0 to 2.5 h old embryos were resolved using 7% SDS polyacrylamide gel electrophoresis, blotted onto nitrocellulose and probed with H5 antibody. After
visualization of the H5 signal, the same filter was probed with Snf antibody. Left panel: Extracts from embryos laid by WT mothers. Right panel: nosmK
embryo extracts. These embryos were laid by nosBN mothers. Individual band intensities were measured and averaged using NIH image analysis. (119.89G
31.60 for the control samples and 187.20G33.06 for the experimental samples). The normalization was performed using corresponding band intensity values
obtained using Snf as a control. The normalized enrichment was estimated to be 1.48-fold.
G. Deshpande et al. / Mechanisms of Development 122 (2005) 645–657652
In a third experiment we followed embryos produced from
mating nosK hbK germline clone mothers to wild-type
males from hatching to the adult stage. In this experiment,
only about 55% of the 457 progeny hatch as first instar
larva. Of the unhatched embryos, approximately 25% had
knirps-like posterior defects typical of that produced by
Hb misexpression (see Fig. 9). A range of defects was
observed in the remaining embryos. Some had near wild-
type cuticles, while others formed only scraps of cuticle
or exhibited a variety of segmentation defects such as
fusions or deletions of multiple segments. Only about
42% of the progeny survived to the adult stage. As might
be expected from the Sxl expression pattern at mid-
embryogenesis, activation of Sxl-Pe in male nosK
blastoderm embryos had no lasting consequences, and
ratio of male to female adults was almost 50/50.
3. Discussion
During the rapid nuclear division cycles in cleavage
stage Drosophila embryos RNA polymerase II transcription
is largely shut down and only a few genes are actively
transcribed. RNA polymerase II transcription in somatic
nuclei is upregulated soon after they migrate to
the periphery of the embryo at stage 9 and by nuclear
cycle 10 and 11 many of the key segmentation genes are
already actively transcribed (Lamb and Laird, l976;
Zalokar, l976). While RNA polymerase II activity is
substantially augmented when the nuclei reach the periph-
ery of the embryo in the soma, the opposite occurs in the
germline pole cell nuclei. When these nuclei migrate into
the posterior pole plasm and pole cells are formed,
transcription is shut down rather than activated (Williamson

Fig. 7. Ectopic Nos downregulates Ser2 and Ser5 CTD phosphorylation. Ser2 CTD Phosphorylation: 0–4 h old embryos from w1 and nosBN; nos-tub 3 0UTR
mothers were stained with a monoclonal (H5) recognizing phosphorylated serine at position 2 in the CTD repeats of the large RNA polymerase II subunit. The
H5 antibody-staining pattern in a wild-type blastoderm embryo is in the top panel, while the two lower panels have embryos from nosBN; nos-tub 3 0UTR
mothers processed in parallel. Note that staining is reduced and patchy in the embryos from transgene mothers (see arrows). Ser5 CTD Phosphorylation: 0–4 h
old stage embryos laid by w1 and nos-bcd 3 0UTR mothers were stained with a monoclonal antibody (H14) recognizing phosphorylated serine at position 5 in the
CTD repeats of the large RNA polymerase II subunit. The H14 antibody-staining pattern in a wild-type blastoderm embryo is in the top panel, while the two
lower panels have embryos from nos-bcd 3 0UTR mothers processed in parallel. Note that staining is reduced and/or patchy in the anterior of embryos from
transgene mothers (see arrows). Note also that unlike H5, the H14 antibody labels wild-type pole cells, indicating that there is a low level of ser5
phosphorylation in wild-type germ cells. Exon 2: The polyclonal ‘ex2’ antibody recognizes epitopes encoded by exon 2 of the large RNA polymerase II
subunit. Shown on the far right is ex2 antibody staining of embryos from wild type, nosBN and nos-tub3 0UTR mothers. Note that the pattern and intensity of
staining in embryos from both nosBN and transgene mothers is indistinguishable from wild type. Though not readily visible at the magnification shown in these
photographs, the pole cells of all embryos are labeled by the ex2 antibody.
G. Deshpande et al. / Mechanisms of Development 122 (2005) 645–657 653
and Lehmann, 1996). Previous studies have implicated the
posterior determinants nos and pum in establishing/main-
taining transcriptional quiescence in pole cells (Deshpande
et al., 1999; Asaoka et al., 1998, 1999). In embryos derived
from mothers mutant for either nos or pum, RNA
polymerase II transcription is not properly downregulated
in the pole cells and several genes which are normally active
only in somatic nuclei are ectopically expressed (Parisi and
Lin, 2000).
Since only nos mRNA localized at the posterior pole is
translated, pole cells have the highest levels of Nos.
However, translation of the localized message generates a
Nos gradient that extends to the center of the embryo. An
obvious question is whether this Nos gradient also affects
RNA polymerase II activity in somatic nuclei. Indeed,
transcription of Sxl-Pe is upregulated in somatic nuclei
when Nos protein is removed, and is repressed when Nos
protein is ectopically expressed. The role of Nos protein
in repressing transcription is not restricted to the sex
determination pathway since the activity of other
promoters also appears to be increased in the absence of
nos function.
3.1. Mechanism of activation
Several mechanisms could potentially explain the
ectopic activation of Sxl-Pe in the soma and germline of
nos mutant embryos. The most obvious is that this
promoter is turned-on by maternal Hb expressed in the
absence of nos. However, Sxl-Pe was upregulated in nosK
embryos even when we eliminated maternal Hb. In
addition, ectopic expression of Hb from a transgene
lacking NREs seemed to repress rather than activate Sxl-
Pe (not shown). Another possibility is that the zygotic
expression of one or more of the X-linked numerators is
elevated in nos embryos, upsetting X chromosome to
autosome counting. However, as none of the known
numerators has a recognizable NRE in the 3 0UTR of its
message, it seems unlikely that these genes are direct
targets for translational repression by Nos protein.
In addition, it is not at all clear why numerator genes
(which are transcribed in the zygote) would be subject
to translational repression by Nos, while autosomal
denominator genes such as deadpan (which turns off
Sxl-Pe) would not.

Fig. 8. Downregulation of Ser2 and Ser5 CTD Phosphorylation in the anterior in embryos from nos-bcd 3 0UTR mothers. Top panels: confocal images of
anterior and posterior halves of a blastoderm stage embryo from nos-bcd 3 0UTR mother probed with antibodies against CTD repeats containing the phospho-
ser2 modification. Bottom panels: confocal images of the anterior half of a blastoderm embryo from nos-bcd 3 0UTR mothers stained with the DNA dye Hoechst
(blue) and antibodies against CTD repeats containing the phospho-ser5 modification (green).
Table 2
Germline clone analysis
Total #Hatched % Hatched
Experiment I
FRT hbFBnosBN!w1 1079 625 58
FRT hbFB!w1 1580 1339 85
Experiment II
FRT hbFBnosBN!w1 69 39 57
P-nos; FRT hbFBnosBN!w1 421 388 92
G. Deshpande et al. / Mechanisms of Development 122 (2005) 645–657654
For this reason, we favor the idea that Sxl-Pe is activated
in nosmK embryos at least in part because RNA polymerase
II activity is upregulated. Support for this idea comes from
analysis of CTD phosphorylation. When RNA polymerase
is transcriptionally engaged the CTD domain is phosphory-
lated on serine 2 and 5. In wild-type pole cells, phospho-ser2
cannot be detected, while there is only little phospho-ser5.
In contrast, phospho-ser2 is found in nosmK pole cells, while
the level of phospho-ser5 is increased. nos-dependent
alterations in CTD phosphorylation are also evident in the
soma. When Nos is absent, the level of ser2 and ser5 CTD
phosphorylation is elevated, while both types of CTD
phosphorylation are reduced by ectopic Nos protein.
Additional evidence that nos has a global effect on
transcription comes from the finding that nos regulates the
methylation of histone H3 in the germline of worms and
flies (Schaner et al., 2003). In both organisms, the
methylation of histone H3 on lysine 4 (H3meK4) is
upregulated in the soma when zygotic transcription
commences in early embryogenesis. In contrast, little or
no methylation H3 K4 is observed in the transcriptionally
quiescent germline. Inhibition of H3 K4 methylation in
germ cells requires nos and H3meK4 is markedly
upregulated in nosK germ cells. In light these findings, we
examined K4 methylation in the soma of nosmK embryos.

Fig. 9. Cuticular defects observed in embryos derived from females
carrying germline clones for hbFBnosBN. Females carrying germline clones
for hbFBnosBN were mated with wild-type males. Embryos derived from
this cross were allowed to develop (see Table 2). The photos show four
different examples of cuticles. (A) Embryo displaying a near normal
segmentation pattern. (B) Embryo with fusions resembling a weak ‘knirps-
like’ phenotype. (C) Embryo with more severe segmentation defects.
(D) Embryo that lacks several segments.
G. Deshpande et al. / Mechanisms of Development 122 (2005) 645–657 655
As might be expected from the effects of nos on CTD
phosphorylation, somatic H3meK4 is elevated compared to
wild type (Desphande, unpublished data).
Since phosphorylation of serines 2 and 5 are correlated
with transcription, the nos-dependent alterations in CTD
phosphorylation are consistent with the idea that nos has
a global impact on RNA polymerase II activity. If this is
the case, an important question is whether CTD
phosphorylation is the cause or the consequence of nos
induced changes in the activity of the transcriptional
apparatus. Because actively transcribing RNA polymerase
has a hyperphosphorylated CTD domain, any mechanism,
which leads to a general increase (or decrease) in
transcription, would likely alter the level of CTD
phosphorylation. This makes it difficult to distinguish
between cause and effect. On the other hand, besides
being a characteristic feature of elongating polymerase,
CTD phosphorylation has been linked to the last steps in
the initiation process, promoter clearance and the
formation of an elongation competent RNA polymerase
complex. Moreover, there is growing evidence that these
steps in the transcription cycle are subject to regulation
(Lee and Lis, l998; Batchelder et al., 1999; Nissen and
Yamamoto, 2000; Shim et al., 2002).
The fact that CTD phosphorylation may be a key
control point in the transcriptional cycle raises the
possibility that nos exerts its effects on polymerase
activity by inhibiting the translation of some factor
which promotes CTD phosphorylation. In nos mutants,
the level of this factor would increase, leading to a
general derepression of transcription. Conversely, the
level of this factor would decrease by ectopic Nos,
reducing overall transcription.
3.2. The Nos gradient and activation of Sxl-Pe
in nosmK embryos
While our results clearly show that Sxl-Pe is inappropri-
ately turned on in the soma of male embryos and
upregulated in the soma of female embryos in the absence
of nos activity, it was initially surprising to find that there is
usually not a very pronounced posterior–anterior activation
gradient. In fact, the smaller Sxl-Pe0.4 kb promoter is clearly
activated not only in the posterior but also in the anterior of
nos embryos, while the larger Sxl-Pe3.0, usually shows at
most only a very shallow posterior–anterior gradient of b-
galactosidase expression. Since the Nos gradient does not
extend beyond the midpoint of the embryo, and the
repressive effects of Nos on hb mRNA translation are
restricted to this posterior domain, one might have expected
that the activation of Sxl-Pe in would be tightly restricted to
the posterior half of nos mutant embryos. However, proteins
of average size would be expected to diffuse (in water or
even in cytoplasm) through the volume of a fly embryo over
a time scale of minutes, and the establishment of gradients
like those seen for Nos or Bcd are likely to require special
mechanisms including a localize source of product, as well
as the sequestration (e.g. nuclear localization) and degra-
dation of the product. If the Nos target for inhibiting general
RNA Pol II activity is translated from a uniformly
distributed maternal mRNA and is able to equilibrate
through the embryo during the time between the onset of the
very rapid nuclear divisions and the formation of the cellular
blastoderm, only a shallow gradient of this factor in the
soma might be expected at any one time in the presence or
absence of Nos. In contrast, in pole cells, where repression
of this factor by Nos would presumably be required to
impose transcriptional quiescence, the formation of the cell
membrane would prevent factor synthesized in the soma
from influencing polymerase activity. This would enable
Nos in the pole cells to reduce the level of this factor below
the threshold required for transcriptional activation.
3.3. Transcriptional repression in the soma
As observed in many species, establishing transcriptional
quiescence in the newly formed pole cells during early
embryogenesis is a critical step in the development of the
Drosophila germline. However, it is not immediately
obvious what role nos mediated down regulation of
polymerase activity would have in the development of the
soma. Obviously, hyperactivation of Sxl-Pe in nosmK
embryos could inappropriately switch on the Sxl autoregu-
latory feedback loop in males. However, our analysis of Sxl
accumulation in post-blastoderm stages suggests that only
very few nosmK 1X/2A embryos actually make the wrong
choice in sexual identity. There is also little evidence of a
sex-bias in adult progeny of nosK hbK germline clone
mothers. The fact that the Sxl autoregulatory loop is usually
not activated in male nosmK embryos, which are

G. Deshpande et al. / Mechanisms of Development 122 (2005) 645–657656
hemizygous for Sxl, is not altogether surprising. In females
that have only a single wild type Sxl gene, activation of the
autoregulatory loop is severely compromised by conditions
which diminish Sxl-Pe activity (Cline, 1988). Since the
amount of Sxl produced by Sxl-Pe in nosmK male embryos
is much less than that in wild-type females, the auto-
regulatory loop should be activated infrequently.
While nosmK males largely escape the effects of
activating Sxl-Pe, the increased polymerase activity appears
to have other consequences. Previous studies have shown
that removal of maternal hb suppresses the posterior defects
of nosmK embryos (Hulskamp et al., 1989; Irish et al., 1989;
Struhl, 1989). However, in the experiments of Irish et al.
(1989) only about 40% of the hbC/hbK embryos from nosK
hbK mothers survive to adults. Likewise, we have found
that only 60% of the embryos produced by hbK nosK
mothers hatch as first instar larva, and that an even lower
number survive to the adult stage. Taken together, these
experiments argue that nos has important functions in the
soma besides blocking translation of maternal hb mRNA. It
seems possible that the segmentation/developmental defects
evident in progeny of hbK nosK clone mothers could arise
from the upregulation of various patterning genes in the
absence of nos activity. We presume that in wild-type
embryos the activity of the transcriptional apparatus and of
target zygote promoters is appropriately adjusted to
compensate for the repressive effects exerted by Nos.
Because the transcriptional apparatus is hyperactivated in
the absence of Nos function, this balance is perturbed and
many genes are overexpressed.
4. Experimental procedures
4.1. Strains and culturing
Flies were grown on a standard medium at 25 8C unless
otherwise noted.
4.2. Germline clonal analysis
Females carrying hbFBnosBN or hbFBnosBN; nosC
transgene germline clones were generated essentially as
described previously (Forbes and Lehmann, 1998).
4.3. Immunohistochemistry
Embryos were probed with antibodies essentially as
described in Deshpande et al. (1995). In order to confirm
any differences seen in the staining patterns between
embryos with different maternal nos genetic backgrounds
when they were stained independently, we mixed the
experimental and control samples together and then stained
the combined samples. In all cases, we observed the
expected mixture of ‘wild type’ and ‘mutant’ staining
phenotypes. Figures only show embryos stained
independently. Anti-Sxl antibody (Bopp et al., 1991) is a
mouse monoclonal and was used at 1:10 dilution. H5 and
H14, the antibodies against phosphorylated CTD repeats of
the large RNA polymerase II subunit were purchased from
Research Diagnostics Inc. Both the antibodies were pre-
absorbed against WT embryonic samples and were sub-
sequently used at 1:250 dilution. Anti-exon 2, a goat
polyclonal antibody (1:100) against the large subunit of
PolII, was a generous gift of Dr Arno Greenleaf.
4.4. Western blot analysis
The Western blot analysis was performed essentially as
described in Deshpande et al. (1995). Embryonic extracts,
prepared from 0 to 2.5 h old embryos, were resolved using
7% SDS polyacrylamide gel electrophoresis, blotted onto
nitrocellulose and probed with H5. Typically, total embryo-
nic extract made out of 2 embryos was loaded per lane. To
compare the total protein loaded onto each lane, the blots
were subsequently reprobed with Snf antibody (1:10). The
intensity of individual bands was measured using NIH
image program and the ratios were calculated to estimate the
fold increase.
Acknowledgements
We thank Gary Struhl, Paul Macdonald, Paul Lasko and
Arno Greenleaf for antibodies, Claude Desplan, Liz Gavis,
Judith Lengyel, and Robin Wharton for fly stocks. We
would like to thank Trudi Schupbach, Eric Wieschaus, Liz
Gavis and Daryl Gohl for advice. We also thank Radhika
Mohan for help in preparing the manuscript. This work was
supported by a grant from the N.I.H. (P.S.).
References
Asaoka, M., Sano, H., Obara, Y., Kobayashi, S., 1998. Maternal Nanos
regulates zygotic gene expression in germline progenitors of Droso-
phila melanogaster. Mech. Dev. 78, 153–158.
Asaoka, M., Yamada, M., Nakamura, A., Hanyu, K., Kobayashi, S., 1999.
Maternal pumilio acts together with Nanos in germline development in
Drosophila embryos. Nat. Cell Biol. 1, 431–437.
Batchelder, C., Dunn, M.A., Choy, B., Suh, Y., Cassie, C., Shim, E.Y.,
et al., 1999. Transcriptional repression by the Caenorhabditis elegans
germ-line protein PIE-1. Genes Dev. 3, 202–212.
Bopp, D., Bell, L., Cline, T.W., Schedl, P., 1991. Developmental
distribution of female-specific Sex-lethal proteins in Drosophila
melanogaster. Genes Dev. 5, 403–415.
Cline, T.W., 1988. Evidence that sisterless-a and sisterless-b are two of
several discrete ‘numerator elements’ of the X/A sex determination
signal in Drosophila that switch Sxl between two alternative stable
expression states. Genetics 119, 829–862.
Cline, T.W., Meyer, B.M., 1996. Vive la difference, males vs females in
flies vs worms. Annu. Rev. Genet. 30, 637–702.
Dahmus, M.E., 1996. Reversible phosphorylation of the C-terminal domain
of RNA polymeraseII. J. Biol. Chem. 265, 19185–19191.

G. Deshpande et al. / Mechanisms of Development 122 (2005) 645–657 657
Deshpande, G., Stukey, J., Schedl, P., 1995. scute (sis-b) function in
Drosophila sex determination. Mol. Cell Biol. 15, 4430–4440.
Deshpande, G., Calhoun, G., Yanowitz, J., Schedl, P., 1999. Novel
functions of nanos in downregulating mitosis and transcription during
the development of Drosophila germline. Cell 99, 271–281.
Estes, P.A., Keyes, L.N., Schedl, P., 1995. Multiple response elements in
the Sex-lethal early promoter ensure its female-specific expression
pattern. Mol. Cell Biol. 15, 904–917.
Forbes, A., Lehmann, R., 1998. Nanos and Pumilio have critical roles in the
development and function of Drosophila germline stem cells.
Development 125, 679–690.
Gavis, E.R., Lehmann, R., 1992. Localization of nanos RNA controls
embryonic polarity. Cell 71, 301–313.
Hagstrom, K., Muller, M., Schedl, P., 1996. Fab-7 functions as a chromatin
domain boundary to ensure proper segment specification by the
Drosophila bithorax complex. Genes Dev. 10, 3202–3215.
Horabin, J., Bopp, D., Waterbury, J., Schedl, P., 1995. Selection and
maintenance of sexual identity in the Drosophila germline. Genetics
141, 1521–1535.
Hulskamp, M., Schroder, C., Pfeifle, C., Jackle, H., Tautz, D., 1989.
Posterior segmentation of the Drosophila embryo in the absence of a
maternal posterior organizer gene. Nature 338, 629–632.
Irish, V., Lehmann, R., Akam, M., 1989. The Drosophila posterior group
gene nanos functions by repressing hunchback activity. Nature 338,
646–648.
Jinks, T.M., Polydorides, A.D., Calhoun, G., Schedl, P., 2000. The
JAK/STAT signaling pathway is required for the initial choice of sexual
identity in Drosophila melanogaster. Mol. Cell 5, 581–587.
Keyes, L.N., Cline, T.W., Schedl, P., 1992. The primary sex determination
signal of Drosophila acts at the level of transcription. Cell 68, 933–943.
Kobayashi, S., Yamada, M., Asaoka, M., Kitamura, T., 1996. Essential role
of the posterior morphogen nanos for germline development in
Drosophila. Nature 380, 708–711.
Komarnitsky, P., Cho, E.J., Buratowski, S., 2000. Different phosphorylated
forms of RNA polymerase II and associated mRNA processing factors
during transcription. Genes Dev. 14, 2452–2460.
Kramer, S.G., Jinks, T.M., Schedl, P., Gergen, J.P., 1999. Direct activation
of Sex-lethal transcription by the Drosophila runt protein. Development
126, 191–200.
Lamb, M.M., Laird, C.D., 1976. Increase in nuclear poly (A) containing
RNA at syncytial blastoderm in Drosophila melanogaster embryos.
Dev. Biol. 54, 31–42.
Lee, D., Lis, J.T., 1998. Transcriptional activation independent of TFIIH
kinase and the RNA polymerase II mediator in vivo. Nature 393,
389–392.
Mello, C.C., Schubert, C., Draper, B., Zhang, W., Lobel, R., Priess, J.R.,
1996. The PIE-1 protein and germline specification in C. elegans
embryos. Nature 382, 710–712.
Murata, Y., Wharton, R.P., 1995. Binding of Pumilio to maternal
hunchback mRNA is required for posterior patterning in Drosophila
embryos. Cell 80, 747–756.
Nissen, R.M., Yamamoto, K.R., 2000. The glucocorticoid receptor inhibits
NFkappaB by interfering with serine-2 phosphorylation of the RNA
polymerase II carboxy-terminal domain. Genes Dev. 14, 2314–2329.
Parisi, M., Lin, H., 2000. Translational repression: a duet of nanos and
pumilio. Curr. Biol. 10, 81–83.
Schaner, C.E., Deshpande, G., Schedl, P., Kelly, W.G., 2003. A conserved
chromatin architecture marks and maintains the restricted germ cell
lineage in worms and flies. Dev. Cell. 5, 747–757.
Seydoux, G., Dunn, M.A., 1997. Transcriptionally repressed germ cells
lack a subpopulation of phosphorylated RNA polymerase II in early
embryos of Caenorhabditis elegans and Drosophila melanogaster.
Development 124, 2191–2201.
Seydoux, G., Mello, C.C., Pettitt, J., Wood, W.B., Priess, J.R., Fire, A.,
1996. Repression of gene expression in the embryonic germ lineage of
C. elegans. Nature 382, 713–716.
Shim, E.Y., Walker, A.K., Shi, Y., Blackwell, T.K., 2002. CDK-9/cyclin T
(P-TEFb) is required in two post-initiation pathways for transcription in
the C. elegans embryo. Genes Dev. 16, 2135–2146.
St Johnston, D., 1993. Pole Plasm and the Posterior Group Genes. From the
Development of Drosophila melanogaster. Cold Spring Harbor
Laboratory.
Struhl, G., 1989. Differing strategies for organizing anterior and posterior
body pattern in Drosophila embryos. Nature 338, 741–744.
Struhl, G., Johnston, P., Lawrence, P.A., 1992. Control of Drosophila body
pattern by the hunchback morphogen gradient. Cell 69, 237–249.
Van Doren, M., Williamson, A.L., Lehmann, R., 1998. Regulation of
zygotic gene expression in Drosophila primordial germ cells. Curr.
Biol. 8, 243–246.
Wang, C., Lehmann, R., 1991. Nanos is the localized posterior determinant
in Drosophila. Cell 66, 637–647.
Wharton, R.P., Struhl, G., 1991. RNA regulatory elements mediate control
of Drosophila body pattern by the posterior morphogen nanos. Cell 67,
955–967.
Williamson, A., Lehmann, R., 1996. Germ cell development in Drosophila.
Annu. Rev. Cell Dev. Biol. 12, 365–391.
Zalokar, M., 1976. Autoradiographic study of protein and RNA formation
during early development of Drosophila eggs. Dev. Biol. 49, 425–437.





























