Embed Size (px)
Citation preview

Nestin-Immunoreactive Cells in Rat Pituitary Areneither Hormonal nor Typical Folliculo-Stellate Cells
Olga Krylyshkina, Jianghai Chen, Liese Mebis, Carl Denef, and Hugo Vankelecom
Laboratory of Cell Pharmacology, Department of Molecular Cell Biology, University of Leuven School of Medicine, CampusGasthuisberg, B-3000 Leuven, Belgium
Nestin is an intermediate filament protein that has originallybeen identified as a marker of neuroepithelial stem/progeni-tor cells. The present study explored whether nestin immu-noreactivity (nestin-ir) is present in the rat pituitary and inwhich cell type(s). Nestin-ir was observed in scattered cells inthe anterior, intermediate, and neural lobes. Nestin-ir cellswere predominantly of stellate shape and were more numer-ous in immature than in adult animals. Nestin-ir did not co-localize with any pituitary hormone, and did not colocalize oronly very sporadically with the folliculo-stellate cell markerS100. In the intermediate lobe, nestin-ir cells containedglial fibrillary acidic protein in an age-dependent manner.Nestin-ir cells were closely associated with endothelial andfibronectin-ir cells, but did mostly not coincide. Nestin-ir wasnot found in �-smooth muscle actin-ir myofibroblasts or inmicroglial cells. Regardless of age, nestin-ir was detected insome unidentifiable cells that border the pituitary cleft. Nes-
tin-ir remained present in pituitary cultured as three-dimen-sional aggregates. Treatment with basic fibroblast growth fac-tor or leukemia inhibitory factor increased the number ofnestin-ir cells. Starting from anterior lobe cell monolayer cul-tures, nestin-ir cells could be selected and propagated to avirtually pure population. These nestin-ir cells displayed re-markable motility and proliferative activity, and did not ex-press hormones, glial fibrillary acidic protein, or S100, butcontained vimentin-, fibronectin-, and �-smooth muscle actin-ir. In conclusion, nestin-ir is present in the pituitary in cellsthat are neither hormonal nor typical folliculo-stellate. Theexpression pattern depends on age and lobe examined. Peri-capillar localization suggests a pericyte phenotype for some ofthem. Whether the heterogeneous nestin-ir population alsocontains pituitary progenitor cells remains to be explored.(Endocrinology 146: 2376–2387, 2005)
THE MATURE PITUITARY is an endocrine organ thatdynamically changes its hormone output in response
to a variety of physiological and pathological triggers to meetthe endocrine needs of the organism. Several of these con-ditions are associated with rapid changes in the number ofcertain cell types. Although cell proliferation is believed tooccur within the various cell type populations, it remainsunknown whether the expansion of a certain population mayalso result from cell development and differentiation fromstem cells or progenitor cells. It also is not known whethersteady cell turnover in the gland, estimated to be approxi-mately 1.5%/d in the young adult male rat (1), is driven froma stem/progenitor cell pool.
Nestin is a class VI intermediate filament protein that hasfirst been identified in the rat embryonic central nervoussystem, where it is abundantly expressed (2, 3). During post-natal development, nestin expression in the brain becomes
more restricted and is particularly found in neuroepithelialstem and proliferative progenitor cells present in the (sub)ventricular zone (3–6). Nestin has also been detected in pu-tative stem/progenitor cells of a number of other tissues,such as dermis (7), pancreas (8–10), retina (11), dental tissue(12), and hair follicle (13).
Although nestin expression has attracted much attentionas a marker in stem/progenitor cells, it is also present in othercell types, such as in newly developing vascular endothelialcells (14–20) and in activated stellate cells of the brain, pan-creas, and liver (15, 21–23). In the adult human gastrointes-tinal tract, nestin expression has been found in CD34-positivecells, enteroglial cells, and interstitial cells of Cajal (24).Also Leydig and Sertoli cells have been reported to containnestin (25). Furthermore, a number of findings do notsupport nestin as an in situ stem/progenitor cell marker inthe pancreas (15–19, 26, 27). Rather, pancreatic nestin-positive cells are proposed to act as niche cells that induceor support endocrine cell neogenesis (17, 20). Thus, nestin-expressing cells may also behave as supporting cells intissue homeostasis.
Nestin-positive stem/progenitor cells appear to behavepeculiarly in culture. Neural stem/progenitor cells can bepropagated in suspension cultures as floating colonies (neu-rospheres) when treated with basic fibroblast growth factor(bFGF) and/or epidermal growth factor (EGF) (28–30). Vir-tually all of the cells within a neurosphere express nestin.Nestin-positive putative precursor cells have also been ob-tained and multiplied in monolayer cultures. A monolayer ofcells expressing nestin grows out from pancreatic islets of
First Published Online January 27, 2005Abbreviations: AL, Anterior lobe; AMCA, 7-amino-4-methylcouma-
rin-3-acetyl; bFGF, basic fibroblast growth factor; BrdU, 5-bromo-2�-deoxyuridine; DCX, doublecortin; DNase, deoxyribonuclease; EGF, epi-dermal growth factor; FCS, fetal calf serum; FS, folliculo-stellate; GFAP,glial fibrillary acidic protein; IL, intermediate lobe; -ir, immunoreactiv-ity; LIF, leukemia inhibitory factor; M-MuLV, Moloney murine leukemiavirus; NIL, neurointermediate lobe; NIP cell, nestin-positive islet-derived progenitor cell; NL, neural lobe; PECAM-1, platelet endothelialcell adhesion molecule-1, also called CD31; PRL, prolactin; Sap, saponin;SM, serum-free medium; �SMA, �-smooth muscle actin; vWF, von Wil-lebrand factor.Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
0013-7227/05/$15.00/0 Endocrinology 146(5):2376–2387Printed in U.S.A. Copyright © 2005 by The Endocrine Society
doi: 10.1210/en.2004-1209
2376

Langerhans [nestin-positive islet-derived progenitor cells(NIP)] (8). NIP cells are able to differentiate in vitro intomultiple liver and pancreatic cell phenotypes.
To date, stem/progenitor cells in the postnatal pituitarygland have not been identified. Pinpointing these cells maybe important to better unravel lineage relationships and celltype renewal and maintenance in the postnatal pituitary.Inspired by the expression profiles of nestin and its potentialrelationship to the stem/progenitor cell phenotype, thepresent investigation was intended to examine whether nes-tin is present in immature and adult pituitary glands,whether its presence is restricted to a certain cell type(s), andhow nestin-ir cells behave in culture.
Materials and MethodsImmunofluorescence of pituitary Vibratome sections andanalysis by confocal laser scanning microscopy
Female Wistar rats (5 and 14 d old or 8 wk old, random cycling) werepurchased from Elevage Janvier (Schaijk, The Netherlands). Rats werekilled by CO2 asphyxiation and decapitation. All animal experimentswere approved by the university ethical committee and conducted inaccordance with The Endocrine Society ethical guidelines. Pituitarieswere carefully isolated under the stereomicroscope and immediatelyfixed for 3 h with 4% paraformaldehyde (Riedel-deHaen, Seelze, Ger-many) in PBS (Invitrogen Life Technologies, Inc., Paisley, UK) or for 48 hwith IHC Zinc Fixative (BD Biosciences, Erembodegem, Belgium) asneeded for CD31/platelet endothelial cell adhesion molecule-1, alsocalled CD31 (PECAM-1) immunostaining. Sometimes rats were per-fused with PBS and 4% paraformaldehyde before separation of thegland. Pituitaries were embedded in agarose (ICN Biomedicals, Aurora,OH; 2% in PBS) before sectioning with the Vibratome (Leica VT 1000 S,Leica Microsystems, Wetzlar, Germany). Pituitaries were attentivelypositioned to cut 50-�m sections through all lobes and cleft. Sections werestored in PBS at 4 C until additional processing by immunofluorescence.
Each staining was performed on duplicate Vibratome sections atdifferent, evenly distributed locations of the pituitary gland. Permeabi-lization of the sections was achieved with two 30-min incubations insaponin (Sigma-Aldrich Corp., St. Louis, MO; 0.5% in PBS; PBS-Sap).Aspecific binding sites were preadsorbed with 20% normal goat ornormal donkey serum (DakoCytomation, Glostrup, Denmark). Sectionswere incubated overnight at room temperature with one or two of thefollowing primary antibodies: mouse antirat nestin IgG1 (Rat-401, BDBiosciences, Mountain View, CA) (2) at a final dilution of 1:100 to 1:200in PBS-Sap; rabbit anti-nestin (no. 130, provided by Dr. R. McKay, NIH,National Institute for Neurological Disorders and Stroke, Bethesda, MD;recognizes rat, mouse, and human nestin) at 1:2,000 dilution; mouseantirat PECAM-1/CD31 (BD Biosciences) at 1:20 dilution; mouse antiratOx-42 (CD11b, Serotec, Oxford, UK) at 1:100 dilution; mouse antihuman�-smooth muscle actin (�SMA; DakoCytomation) at 1:100 dilution; rab-bit antirat GH (UCB Bioproducts, Brussels, Belgium) at 1:2,500 dilution;rabbit anticow S100 at 1:100 dilution (from DakoCytomation or ZymedLaboratories, San Francisco, CA), anticow glial fibrillary acidic protein(GFAP) at 1:10,000 dilution (DakoCytomation); rabbit antirat vWF at1:100 dilution (Chemicon Biognost, Heule, Belgium); mouse anti-pancadherin at 1:200 dilution (Sigma-Aldrich Corp., Bornem, Belgium); goatantihuman vimentin at 1:20 dilution (Sigma-Aldrich Corp.); rabbit an-tihuman fibronectin at 1:200 dilution (Sigma-Aldrich Corp.); goat anti-human doublecortin (DCX; Santa Cruz Biotechnology, Inc., Santa Cruz,CA) at 1:400 dilution; and rabbit antihuman ACTH at 1:7,500 dilution,antirat prolactin (anti-rPRL-IC5) at 1:2,500 dilution, antirat LH� at1:10,000 dilution, antirat LH� at 1:50,000 dilution, antihuman FSH� at1:1,250 dilution, antirat TSH� at 1:12,500 dilution (all provided by Dr.A. F. Parlow through the National Hormone and Pituitary Program;NIDDK, Torrance, CA). As negative controls, primary antibodies wereomitted, or mouse nonsense IgG and normal rabbit or goat serum (bothfrom DakoCytomation) were substituted for the primary antibodies atcomparable dilutions. After thorough washing in PBS-Sap, sections wereincubated for 1.5 h with the appropriate secondary antibodies: Alexa
Fluor 488- or 555-conjugated goat antimouse or goat antirabbit (Molec-ular Probes, Eugene, OR; all at 1:1,000 dilution in PBS-Sap) or Cy3-conjugated donkey antigoat at 1:500 dilution (Jackson ImmunoResearchLaboratories, West Grove, PA). Nuclei were counterstained with theDNA-binding dye ToPro-3 (Molecular Probes). Sections were mountedon glass slides using Vectashield (Vector Laboratories, Inc., Burlingame,CA), gently covered with glass coverslips, and stored at �80 C untilanalysis by confocal laser microscopy.
Sections were scanned using a confocal laser scanning microscope(LSM 510, Zeiss, Zaventem, Belgium) in multitrack mode. Specificationswere as follows: for Alexa Fluor 488, excitation at 488 nm and emissionat 505–550 nm; for Alexa Fluor 555 and Cy3, excitation at 543 nm andemission at 560–615 nm through a NFT545; and for ToPro-3, excitationat 633 nm and emission above 650 nm. Bleedthrough of a fluorophorein a nonappropriate channel was not observed. For all stainings, at leastone of the duplicate sections at different positions in the pituitary wasthoroughly scanned, and stainings were repeated two or more times.Figures were prepared using Zeiss LSM Image Examiner or Browser andMicrosoft PowerPoint.
RT-PCR analysis of nestin expression in the pituitary
Anterior (AL) and neurointermediate (NIL) lobes from adult ratswere carefully dissected, and total RNA was prepared with TriPurereagent (Invitrogen Life Technologies, Inc.) according to the manufac-turer’s recommendations. RT was carried out in the following reactionmixture: 2 �l RNA (2–20 ng), 4 �l MgCl2 (25 mm), 2 �l 10� PCR Goldbuffer, 0.8 �l deoxy-NTPs (100 mm), 1 �l random hexamers (50 �m; allfrom Applied Biosystems, Lennik, Belgium), 1 �l Moloney murine leu-kemia virus (M-MuLV) reverse transcriptase (200 U/�l; Invitrogen LifeTechnologies, Inc.), 1 �l ribonuclease inhibitor (40 U/�l; RNasin, Pro-mega Corp., Leiden, The Netherlands), and 8.2 �l H2O. RT was per-formed using the following temperature cycle: 10 min at 25 C, 50 minat 42 C, and 10 min at 95 C. For PCR, 1 �l of the RT reaction was addedto 9 �l of a mixture containing 0.9 �l MgCl2 (25 mm), 1.1 �l 10� PCRGold buffer, 0.1 �l deoxy-NTPs (100 mm), 0.15 �l AmpliTaq Gold DNApolymerase (5 U/�l; Applied Biosystems), and 0.1 �l of each oligonu-cleotide primer (100 �m; Invitrogen Life Technologies, Inc.). For ampli-fication of nestin cDNA, the following primers were used: 5�-AAG-CAGGGTCTACAGAGTCAGATCG-3� (sense) and 5�-GCTGTCACAG-GAGTCTCAAGGGTAT-3� (antisense), yielding an amplified fragmentof 299 bp. Samples were subjected to 7 min at 95 C, 40 cycles of 10 secat 95 C, 20 sec at 58.4 C, and 25 sec at 72 C, and finally 7 min at 72 C ina GeneAmp PCR System 2400 (Applied Biosystems). To excludegenomic DNA amplification, primers were selected to span introns. Inaddition, RT was carried out without M-MuLV reverse transcriptase.As a negative control for PCR, H2O was used instead of RT reactionproduct. As a positive control, RT-PCR was performed for L19, aribosomal protein constitutively expressed in all cells (data not shown).Amplified DNA products were electrophoresed in a 2% agarose gelcontaining 0.5 �g/ml ethidium bromide. Amplicons were cut from thegel, and their identities were confirmed by nucleotide sequence ana-lysis (performed by Lark Technologies, Essex, UK).
Aggregate culture of pituitary cells and analysis by whole-aggregate immunofluorescence or FACS
Pituitary cells were cultured as aggregates as described in detailpreviously (31). In brief, pituitary glands (or the separately dissected ALor NIL) were dissociated into single cells using successive incubationswith trypsin, deoxyribonuclease (DNase), trypsin inhibitor, and EDTA.Finally, tissue pieces were mechanically triturated, and cells were trans-ferred to DNase-containing medium before harvesting by centrifuga-tion. Cells were seeded at 2 � 106/2 ml in 35-mm petri dishes in chem-ically defined serum-free medium (SM; produced by Invitrogen LifeTechnologies, Inc.) supplemented, or not, with human recombinantbFGF (20–100 ng/ml), EGF (20 ng/ml; both from R&D Systems, Min-neapolis, MN), rat leukemia inhibitory factor (LIF; 10 ng/ml; ChemiconBiognost), or a combination of these growth factors. Dishes were placedon a gyratory shaker in a 1.5% CO2 incubator (37 C), resulting in theformation of well characterized aggregates (31). On d 2, aggregates weretransferred to 55-mm petri dishes in a final volume of 6 ml. Medium wasrenewed twice a week. Live aggregates were viewed and photographed
Krylyshkina et al. • Nestin in the Pituitary Endocrinology, May 2005, 146(5):2376–2387 2377

using an inverted microscope (Nikon TMS-F, Analis, Namen, Belgium)and a digital camera (Nikon DXM 1200) connected to a computer withACT-1 image capturing software (Nikon).
Immunofluorescent analysis of whole aggregates was performed asdescribed above for Vibratome sections, except for the fixation time withparaformaldehyde (2 h). From all stainings, at least five aggregates werethoroughly scanned in the confocal setting, and stainings were repeatedtwo or more times.
Quantification of nestin-immunopositive cells in the aggregates aswell as in the pituitary ex vivo was performed by FACS. Pituitary glandsand aggregates (after a 6-wk culture period) were dispersed into singlecells as described above. Cells were fixed on ice for 1 h in 4% parafor-maldehyde, permeabilized at room temperature during 10 min withPBS-Sap, and then consecutively incubated on ice with normal goatserum (20%; 5 min), mouse antirat nestin antibody (1:200; 1 h), andphycoerythrin-labeled goat antimouse Ig (1:20; 45 min; BD Pharmingen,San Diego, CA). Cells were analyzed on a FACSort (BD Biosciences),and nestin-positive cells were gated to calculate their proportion. Datawere statistically analyzed using a one-way ANOVA with Tukeymultiple comparison test (P � 0.05; statistical software from StatsDirect,Cheshire, UK).
Monolayer culture of pituitary cells and immuno-fluorescent analysis
ALs from adult (8-wk-old) random cycling female Wistar rats weredissociated into single cells as described above. Cells were plated in SMin six- or 24-well plates on coverslips coated with collagen (5 �g/cm2;mouse collagen type IV, BD Biosciences) at a density of 0.3 � 106/cm2.Cultures were kept at 37 C in a 1.5% CO2 incubator. Other coatings, suchas fibronectin and laminin, were also tested, but did not allow properselection of nestin-ir cells. After 2 d in culture, strongly attached cellswere selected by removing the less adhering cells with trypsin. Differentprotocols were tried and resulted in the following strict selectionmethod. Coverslips were placed for 80 sec in trypsin solution (1% inDMEM/0.3% BSA; prewarmed at 37 C), then intensively rinsed with astream of sterile prewarmed (37 C) PBS and placed for 5–10 min in a dishcontaining trypsin inhibitor solution (0.1% in DMEM/0.3% BSA). Thissolution was replaced with SM supplemented with 10% fetal calf serum(FCS; HyClone, Logan, UT). Selected cells (which mainly representednestin-ir cells) were cultured in 1.5% CO2 at 37 C for 4–5 d until theyreached 80–90% confluence. Then coverslips were processed for immu-nofluorescent analysis or cells were replated for additional passages.Passaging was performed as follows. Cells were trypsinized (0.5% tryp-sin in DMEM/0.3% BSA, containing 1 mm EDTA) for 10 min, spun downthrough a 3% BSA layer containing trypsin inhibitor, resuspended infresh SM supplemented with 10% FCS, and plated at a dilution of 1:4 oncollagen-coated coverslips if immunostaining was to be performed or oncollagen-coated plastic dishes for propagation. Live cultures wereviewed and photographed as described above for aggregates. Videorecording of live cell cultures was performed using a Leica AS MDWworkstation connected to the computer with Leica Deblur software. Cellseparation by gravity sedimentation through a BSA gradient was per-formed as previously described (32). For immunofluorescent analysis,cells were fixed on coverslips with 4% paraformaldehyde in PBS for 15min, permeabilized for 5 min in 4% paraformaldehyde with 0.4% TritonX-100 (Sigma-Aldrich Corp.), and incubated with primary antibodies(diluted in 1% BSA/PBS) for 1 h and with secondary antibodies for 50min. The antibodies and dilutions used were as described above forVibratome sections. For actin visualization, Alexa 488-conjugated phal-loidin (Molecular Probes) was added to antibody mixtures at a dilutionof 1:50. When necessary, positive control cells were simultaneouslyprocessed: rat heart vessel preparations for CD31 staining, rat brainmicroglial cell preparations for Ox-42 staining, and fetal rat brain forDCX staining. Finally, coverslips were mounted on glass slides usingVectashield containing 0.5 �g/ml 4�,6-diamido-2-phenylindole hydro-chloride (Roche, Mannheim, Germany) for nuclear counterstaining.
In some experiments the dipeptide �-Ala-Lys-N�-AMCA (7-amino-4-methylcoumarin-3-acetyl) was used as a marker for folliculo-stellate(FS) cells (33). �-Ala-Lys-N�-AMCA (provided by Dr. K. Bauer, MaxPlanck Institute, Hannover, Germany) was added to the culture mediumat a concentration of 100 �m for 2 h at 37 C. Incubation was stopped onice. Cells were rinsed three times with ice-cold Hanks’ balanced salt
solution (Invitrogen Life Technologies, Inc.) and once with ice-cold PBS.Cells were then fixed for 20 min in 3% paraformaldehyde/1% glutar-aldehyde (Merck, Darmstadt, Germany) at room temperature andtreated three times for 5 min each time with 0.5 mg/ml NaBH4 (Sigma-Aldrich Corp.) on ice. Before immunostaining, performed as describedabove, cells were briefly permeabilized with 0,1% Triton X-100/PBS for2 min.
To visualize cell proliferation, the nucleotide analog 5-bromo-2�-deoxyuridine (BrdU; BD Biosciences) was added to the cell cultures for6–24 h at a final concentration of 10 �m. Cells were fixed, treated withDNase (100 U/ml DNase I in PBS; amplification grade, Invitrogen LifeTechnologies, Inc.) for 30 min at 37 C, incubated with mouse anti-BrdU(1:5; BD Biosciences), and processed for immunostaining as describedabove.
Slides with immunofluorescent samples were examined using a LeicaDMRB epifluorescence microscope with appropriate filter sets. Pictureswere taken with a digital camera (Nikon DXM 1200), connected to acomputer with ACT-1 image-capturing software (Nikon). Figures andmovies were prepared using ImageJ (1.30v, freely available at http://rsb.info.nih.gov/ij/), Adobe Photoshop (Adobe Systems, San Jose, CA),and PowerPoint (Microsoft, Redmond, CA).
ResultsNestin-ir in the pituitary gland of immature and adult rats
Pituitary Vibratome sections from immature (5 and 14 dold) and adult rats were immunofluorescently labeled usingthe monoclonal antirat nestin antibody Rat-401 (2) and an-alyzed by confocal laser scanning microscopy. Nestin-ir waspresent scattered over the three lobes and was associatedwith distinct cells as clear from nuclear counterstaining (Fig.1). Simultaneous staining using the monoclonal Rat-401 anda polyclonal antinestin antiserum (gift from Dr. R. McKay)showed evident overlap in the vast majority of immunopo-sitive cells, confirming the identity of the immunoreactivematerial (Fig. 1D). No staining was observed when the mono-clonal Rat-401 was replaced with nonsense mouse IgG1 (datanot shown).
Most nestin-ir cells had a stellate shape with long pro-cesses; some appeared more rounded or polygonal. Nestin-irmarkedly declined between immature and adult ages (Fig. 1,A vs. C). About 0.5% (0.49 � 0.19%; average � sem of threeindependent measurements) of freshly dispersed pituitarycells from 14-d-old rats were scored nestin-positive by flowcytometric analysis.
RT-PCR revealed the presence of nestin mRNA in the ALand NIL (Fig. 2). The amplified fragment was identified bynucleotide sequencing as part of rat nestin cDNA.
Identification of nestin-ir cells within pituitary lobes
First, double immunofluorescence was performed for nes-tin and pituitary hormones. At the ages analyzed, no colo-calization of nestin-ir with ACTH, GH, PRL, LH�, FSH�,TSH�, or LH� was detected (Fig. 1, E–I, for 14-d-old pitu-itary; adult pituitary data not shown). In the adult AL,nestin-ir did only sporadically colocalized with S100, amarker ofFS cells (34, 35) (Fig. 1J). Furthermore, colocalizationof nestin- and S100-ir was not observed in the neural lobe(NL) or in the 14-d-old pituitary (data not shown).
In the immature IL, some of the nestin-ir cells expressedGFAP (Fig. 1K). No colocalization was found in the immatureNL, where GFAP is abundantly expressed in pituicytes (Fig.1K). GFAP-ir was only very sporadically detected in the AL,
2378 Endocrinology, May 2005, 146(5):2376–2387 Krylyshkina et al. • Nestin in the Pituitary
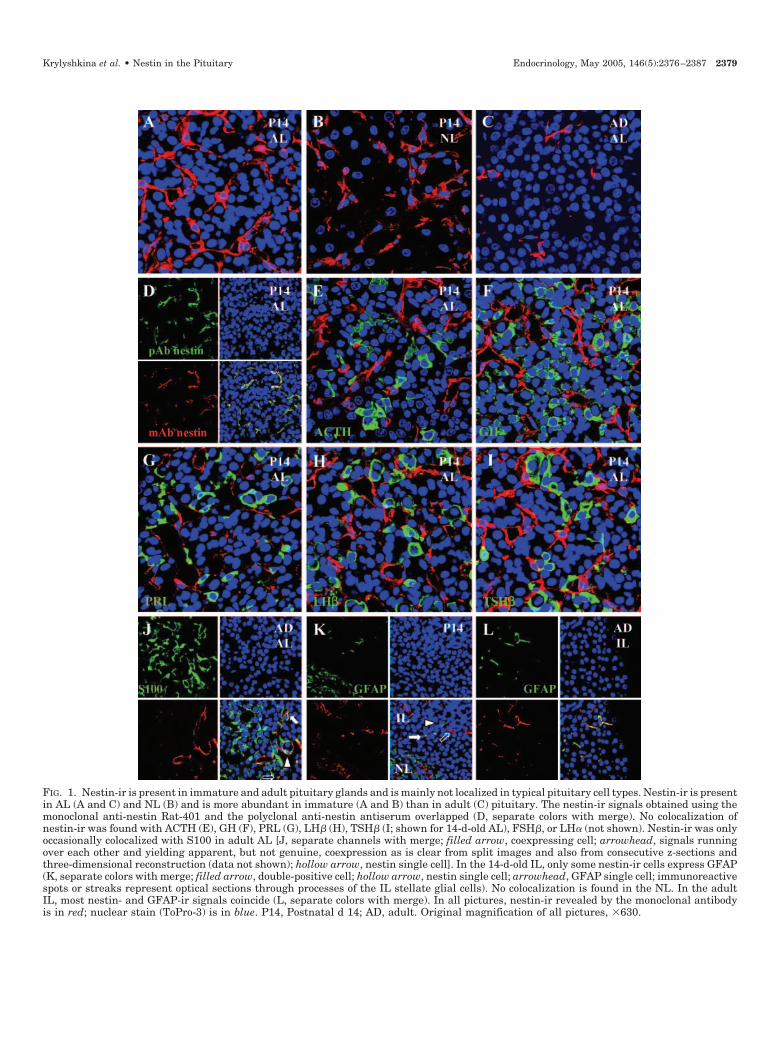
FIG. 1. Nestin-ir is present in immature and adult pituitary glands and is mainly not localized in typical pituitary cell types. Nestin-ir is presentin AL (A and C) and NL (B) and is more abundant in immature (A and B) than in adult (C) pituitary. The nestin-ir signals obtained using themonoclonal anti-nestin Rat-401 and the polyclonal anti-nestin antiserum overlapped (D, separate colors with merge). No colocalization ofnestin-ir was found with ACTH (E), GH (F), PRL (G), LH� (H), TSH� (I; shown for 14-d-old AL), FSH�, or LH� (not shown). Nestin-ir was onlyoccasionally colocalized with S100 in adult AL [J, separate channels with merge; filled arrow, coexpressing cell; arrowhead, signals runningover each other and yielding apparent, but not genuine, coexpression as is clear from split images and also from consecutive z-sections andthree-dimensional reconstruction (data not shown); hollow arrow, nestin single cell]. In the 14-d-old IL, only some nestin-ir cells express GFAP(K, separate colors with merge; filled arrow, double-positive cell; hollow arrow, nestin single cell; arrowhead, GFAP single cell; immunoreactivespots or streaks represent optical sections through processes of the IL stellate glial cells). No colocalization is found in the NL. In the adultIL, most nestin- and GFAP-ir signals coincide (L, separate colors with merge). In all pictures, nestin-ir revealed by the monoclonal antibodyis in red; nuclear stain (ToPro-3) is in blue. P14, Postnatal d 14; AD, adult. Original magnification of all pictures, �630.
Krylyshkina et al. • Nestin in the Pituitary Endocrinology, May 2005, 146(5):2376–2387 2379

and not together with nestin (data not shown). In the adultpituitary, colocalization of nestin- and GFAP-ir was detectedin the great majority of nestin-positive IL cells (Fig. 1L), butnot, or only very occasionally, in nestin-ir cells of the AL andNL (data not shown).
To examine whether nestin expression was associatedwith endothelial cells, sections were simultaneously stainedfor nestin and the endothelial cell markers von Willebrandfactor (vWF) and CD31/PECAM-1. In the 14-d-old pituitary,nestin-ir often codistributed with vWF- and CD31-ir, butconvincing colocalization in cells was only sporadically ob-served (Fig. 3, A and B). In the adult pituitary, nestin-ir cellsalso closely associated with CD31-ir cells, but did not overlap(Fig. 3C). vWF was not detected in adult pituitary, in contrastto CD31, which is considered to be a more general marker ofendothelial cells (36).
Nestin-ir cells did not represent microglial cells, becausenestin-ir was not colocalized with Ox-42 (CD11b), a markerfor macrophage/microglial cells in the pituitary (37) (datanot shown).
To examine whether nestin-ir cells displayed mesenchy-mal features, colocalization with fibronectin, vimentin, and�SMA was tested. Again, a great part of nestin-ir closelyassociated, but not wholly overlapped, with fibronectin-ir(Fig. 3D) that was mainly found in or around endothelial cellsin both immature and adult AL. In IL and NL, no colocal-ization of nestin- and fibronectin-ir was observed (data notshown). �SMA-ir cells, present in small numbers aroundcapillary structures, were also in close proximity with nes-tin-ir cells in all lobes at both ages, but signals did not overlap(Fig. 3E). �SMA-ir cells, in contrast to nestin-ir cells, werelower in number in immature than adult pituitaries. Vimen-tin-ir was only sporadically detected in the AL (and not in theIL or NL), but no colocalization with nestin-ir was found(data not shown).
To address the question of whether some nestin-ir cellsrepresented neuronal progenitor cells and whether the pi-tuitary, in particular the NL, could function as a secondarygerminal center for neurons, as has recently been demon-strated for the hypothalamus (38), sections were stained for
DCX, a marker of migrating and differentiating neuronalcells during embryonic and postnatal development (39). NoDCX-ir was found in the various lobes of the pituitary gland,at either early postnatal or adult ages (data not shown).
Nestin-ir cells in the layers bordering pituitary cleft
Nestin-ir was also detected in the cell layers lining the cleft,the remnant of Rathke’s pouch. Nestin-ir cells were observedat all ages analyzed (Fig. 3, F–H; postnatal d 5 not shown) inboth the anterior wall (bordering the AL) and the posteriorwall (flanking the IL), and either occurred as solitary cells orin clusters. No expression of hormones, S100, vWF, CD31,GFAP, fibronectin, vimentin, �SMA, and Ox-42 (Fig. 3H anddata not shown) was detected in these nestin-ir cells. Some-times, hormone-, S100-, fibronectin-, vimentin-, and �SMA-ircells were also found in the cell layers bordering the cleft (Fig.3H and data not shown).
Nestin-ir in pituitary cell aggregate cultures
As analyzed by whole-aggregate immunofluorescenceand confocal laser scanning microscopy, nestin-ir was de-tected in aggregates formed from immature or adult AL cellsand persisted in these aggregates up to at least 7 wk inserum-free culture (Fig. 3, I–J). Similar to that in vivo, nestin-irwas not detected in hormone-positive cells (Fig. 3I) and wasnot or only sporadically detected in GFAP- and S100-expressing cells, respectively (Fig. 3J and data not shown).vWF-, CD31-, Ox-42-, �SMA-, and vimentin-ir were notfound in pituitary cell aggregate cultures. In contrast, ag-gregates made from NIL cells alone showed colocalization ofGFAP- and nestin-ir (data not shown).
bFGF (20 ng/ml) induced a 10-fold rise in the number ofnestin-positive cells after a 6-wk treatment period (from0.29% in untreated aggregates to 2.69% in bFGF-treated ag-gregates), and LIF (10 ng/ml) induced a 5-fold increase (from0.29% to 1.44%; Fig. 4A). EGF (20 ng/ml) did not affect thenumber of nestin-ir cells (0.35% vs. 0.29% in controls), nor didEGF add to the effect of bFGF (2.45% for EGF plus bFGF vs.2.69% for bFGF alone). bFGF and LIF together exerted apartially additive effect (3.72% for bFGF plus LIF vs. 2.69%for bFGF). The stimulatory effect of bFGF was dose depen-dent; a 10-fold increase in nestin-positive cells was obtainedwith 20 and 50 ng/ml bFGF, and a 20-fold rise was obtainedwith 100 ng/ml bFGF (Fig. 4B).
A remarkable finding in the bFGF-treated, but not in theuntreated, EGF-treated, or LIF-treated, AL cell aggregateswas the development in time of hollow bulges (Fig. 3K).Some of the cells bordering the hollow bulges containednestin-ir (Fig. 3L), but no hormones, S100, or GFAP (data notshown).
Nestin-ir cells in AL monolayer cultures
Dissociated cells from adult AL were cultured on collagen-coated coverslips in chemically defined SM without any sup-plements. In primary (1- to 4-d) cultures (Fig. 5A), nestin-ircells were present and showed distinct morphological char-acteristics. Nestin-ir cells were large, fully spread cells withwell developed actin structure and a large nucleus that was
FIG. 2. The nestin gene is expressed in the pituitary gland. Nes-tin mRNA is detected by RT-PCR in adult AL and NIL, yieldingan amplified fragment of 299 bp. M, 100-bp marker; � and �, RTwith and without M-MuLV, respectively; �P, negative PCR con-trol.
2380 Endocrinology, May 2005, 146(5):2376–2387 Krylyshkina et al. • Nestin in the Pituitary
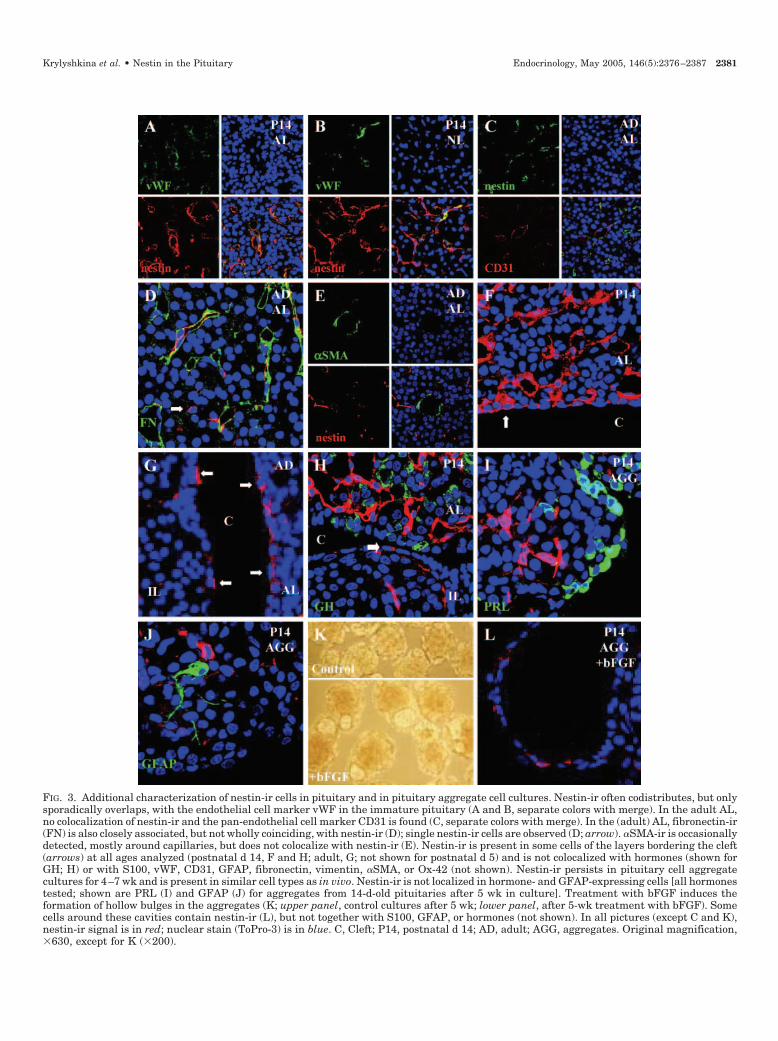
FIG. 3. Additional characterization of nestin-ir cells in pituitary and in pituitary aggregate cell cultures. Nestin-ir often codistributes, but onlysporadically overlaps, with the endothelial cell marker vWF in the immature pituitary (A and B, separate colors with merge). In the adult AL,no colocalization of nestin-ir and the pan-endothelial cell marker CD31 is found (C, separate colors with merge). In the (adult) AL, fibronectin-ir(FN) is also closely associated, but not wholly coinciding, with nestin-ir (D); single nestin-ir cells are observed (D; arrow). �SMA-ir is occasionallydetected, mostly around capillaries, but does not colocalize with nestin-ir (E). Nestin-ir is present in some cells of the layers bordering the cleft(arrows) at all ages analyzed (postnatal d 14, F and H; adult, G; not shown for postnatal d 5) and is not colocalized with hormones (shown forGH; H) or with S100, vWF, CD31, GFAP, fibronectin, vimentin, �SMA, or Ox-42 (not shown). Nestin-ir persists in pituitary cell aggregatecultures for 4–7 wk and is present in similar cell types as in vivo. Nestin-ir is not localized in hormone- and GFAP-expressing cells [all hormonestested; shown are PRL (I) and GFAP (J) for aggregates from 14-d-old pituitaries after 5 wk in culture]. Treatment with bFGF induces theformation of hollow bulges in the aggregates (K; upper panel, control cultures after 5 wk; lower panel, after 5-wk treatment with bFGF). Somecells around these cavities contain nestin-ir (L), but not together with S100, GFAP, or hormones (not shown). In all pictures (except C and K),nestin-ir signal is in red; nuclear stain (ToPro-3) is in blue. C, Cleft; P14, postnatal d 14; AD, adult; AGG, aggregates. Original magnification,�630, except for K (�200).
Krylyshkina et al. • Nestin in the Pituitary Endocrinology, May 2005, 146(5):2376–2387 2381
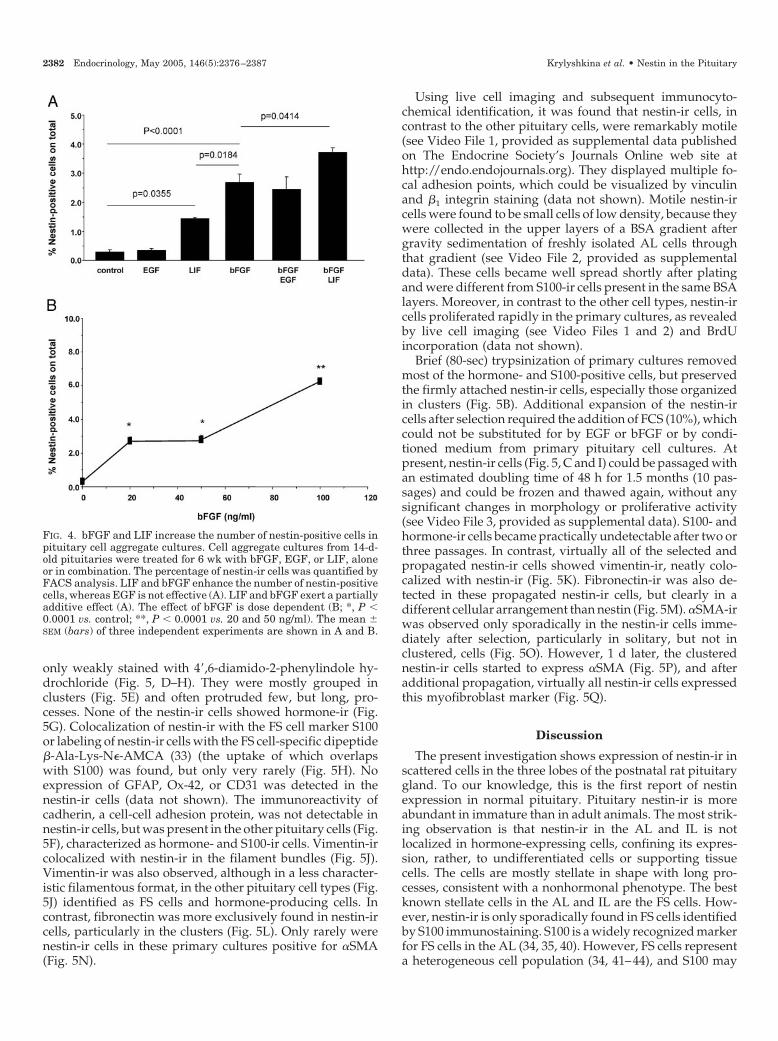
only weakly stained with 4�,6-diamido-2-phenylindole hy-drochloride (Fig. 5, D–H). They were mostly grouped inclusters (Fig. 5E) and often protruded few, but long, pro-cesses. None of the nestin-ir cells showed hormone-ir (Fig.5G). Colocalization of nestin-ir with the FS cell marker S100or labeling of nestin-ir cells with the FS cell-specific dipeptide�-Ala-Lys-N�-AMCA (33) (the uptake of which overlapswith S100) was found, but only very rarely (Fig. 5H). Noexpression of GFAP, Ox-42, or CD31 was detected in thenestin-ir cells (data not shown). The immunoreactivity ofcadherin, a cell-cell adhesion protein, was not detectable innestin-ir cells, but was present in the other pituitary cells (Fig.5F), characterized as hormone- and S100-ir cells. Vimentin-ircolocalized with nestin-ir in the filament bundles (Fig. 5J).Vimentin-ir was also observed, although in a less character-istic filamentous format, in the other pituitary cell types (Fig.5J) identified as FS cells and hormone-producing cells. Incontrast, fibronectin was more exclusively found in nestin-ircells, particularly in the clusters (Fig. 5L). Only rarely werenestin-ir cells in these primary cultures positive for �SMA(Fig. 5N).
Using live cell imaging and subsequent immunocyto-chemical identification, it was found that nestin-ir cells, incontrast to the other pituitary cells, were remarkably motile(see Video File 1, provided as supplemental data publishedon The Endocrine Society’s Journals Online web site athttp://endo.endojournals.org). They displayed multiple fo-cal adhesion points, which could be visualized by vinculinand �1 integrin staining (data not shown). Motile nestin-ircells were found to be small cells of low density, because theywere collected in the upper layers of a BSA gradient aftergravity sedimentation of freshly isolated AL cells throughthat gradient (see Video File 2, provided as supplementaldata). These cells became well spread shortly after platingand were different from S100-ir cells present in the same BSAlayers. Moreover, in contrast to the other cell types, nestin-ircells proliferated rapidly in the primary cultures, as revealedby live cell imaging (see Video Files 1 and 2) and BrdUincorporation (data not shown).
Brief (80-sec) trypsinization of primary cultures removedmost of the hormone- and S100-positive cells, but preservedthe firmly attached nestin-ir cells, especially those organizedin clusters (Fig. 5B). Additional expansion of the nestin-ircells after selection required the addition of FCS (10%), whichcould not be substituted for by EGF or bFGF or by condi-tioned medium from primary pituitary cell cultures. Atpresent, nestin-ir cells (Fig. 5, C and I) could be passaged withan estimated doubling time of 48 h for 1.5 months (10 pas-sages) and could be frozen and thawed again, without anysignificant changes in morphology or proliferative activity(see Video File 3, provided as supplemental data). S100- andhormone-ir cells became practically undetectable after two orthree passages. In contrast, virtually all of the selected andpropagated nestin-ir cells showed vimentin-ir, neatly colo-calized with nestin-ir (Fig. 5K). Fibronectin-ir was also de-tected in these propagated nestin-ir cells, but clearly in adifferent cellular arrangement than nestin (Fig. 5M). �SMA-irwas observed only sporadically in the nestin-ir cells imme-diately after selection, particularly in solitary, but not inclustered, cells (Fig. 5O). However, 1 d later, the clusterednestin-ir cells started to express �SMA (Fig. 5P), and afteradditional propagation, virtually all nestin-ir cells expressedthis myofibroblast marker (Fig. 5Q).
Discussion
The present investigation shows expression of nestin-ir inscattered cells in the three lobes of the postnatal rat pituitarygland. To our knowledge, this is the first report of nestinexpression in normal pituitary. Pituitary nestin-ir is moreabundant in immature than in adult animals. The most strik-ing observation is that nestin-ir in the AL and IL is notlocalized in hormone-expressing cells, confining its expres-sion, rather, to undifferentiated cells or supporting tissuecells. The cells are mostly stellate in shape with long pro-cesses, consistent with a nonhormonal phenotype. The bestknown stellate cells in the AL and IL are the FS cells. How-ever, nestin-ir is only sporadically found in FS cells identifiedby S100 immunostaining. S100 is a widely recognized markerfor FS cells in the AL (34, 35, 40). However, FS cells representa heterogeneous cell population (34, 41–44), and S100 may
FIG. 4. bFGF and LIF increase the number of nestin-positive cells inpituitary cell aggregate cultures. Cell aggregate cultures from 14-d-old pituitaries were treated for 6 wk with bFGF, EGF, or LIF, aloneor in combination. The percentage of nestin-ir cells was quantified byFACS analysis. LIF and bFGF enhance the number of nestin-positivecells, whereas EGF is not effective (A). LIF and bFGF exert a partiallyadditive effect (A). The effect of bFGF is dose dependent (B; *, P �0.0001 vs. control; **, P � 0.0001 vs. 20 and 50 ng/ml). The mean �SEM (bars) of three independent experiments are shown in A and B.
2382 Endocrinology, May 2005, 146(5):2376–2387 Krylyshkina et al. • Nestin in the Pituitary
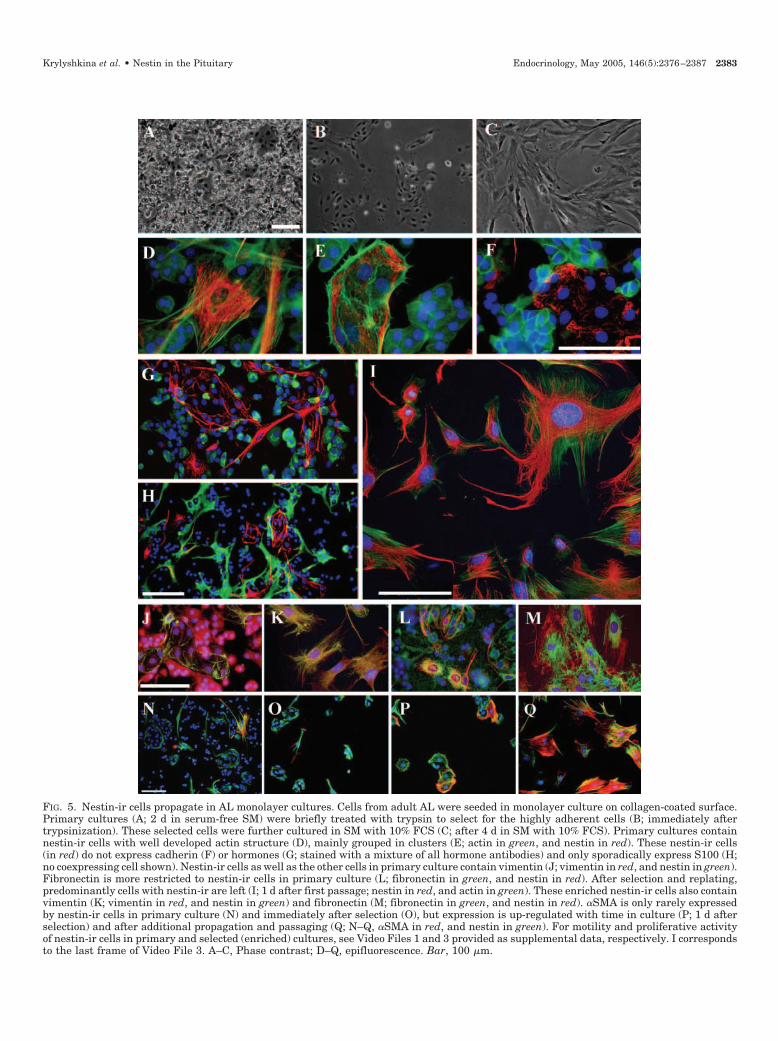
FIG. 5. Nestin-ir cells propagate in AL monolayer cultures. Cells from adult AL were seeded in monolayer culture on collagen-coated surface.Primary cultures (A; 2 d in serum-free SM) were briefly treated with trypsin to select for the highly adherent cells (B; immediately aftertrypsinization). These selected cells were further cultured in SM with 10% FCS (C; after 4 d in SM with 10% FCS). Primary cultures containnestin-ir cells with well developed actin structure (D), mainly grouped in clusters (E; actin in green, and nestin in red). These nestin-ir cells(in red) do not express cadherin (F) or hormones (G; stained with a mixture of all hormone antibodies) and only sporadically express S100 (H;no coexpressing cell shown). Nestin-ir cells as well as the other cells in primary culture contain vimentin (J; vimentin in red, and nestin in green).Fibronectin is more restricted to nestin-ir cells in primary culture (L; fibronectin in green, and nestin in red). After selection and replating,predominantly cells with nestin-ir are left (I; 1 d after first passage; nestin in red, and actin in green). These enriched nestin-ir cells also containvimentin (K; vimentin in red, and nestin in green) and fibronectin (M; fibronectin in green, and nestin in red). �SMA is only rarely expressedby nestin-ir cells in primary culture (N) and immediately after selection (O), but expression is up-regulated with time in culture (P; 1 d afterselection) and after additional propagation and passaging (Q; N–Q, �SMA in red, and nestin in green). For motility and proliferative activityof nestin-ir cells in primary and selected (enriched) cultures, see Video Files 1 and 3 provided as supplemental data, respectively. I correspondsto the last frame of Video File 3. A–C, Phase contrast; D–Q, epifluorescence. Bar, 100 �m.
Krylyshkina et al. • Nestin in the Pituitary Endocrinology, May 2005, 146(5):2376–2387 2383

not stain all of the FS cells (43), suggesting that nestin-ir islocalized in an S100-negative FS cell subpopulation. In thisrespect it should be noted that we have previously shownthat the cytokine interferon-� induces the expression of in-ducible nitric oxide synthase not only in some S100-ir cells,but also in a large number of unidentifiable cells character-ized as small cells of low density (32). These cells in whichinducible nitric oxide synthase is induced may overlap withthe nestin-ir cells. In addition, we previously detected trans-ferrin-like immunoreactivity in a considerable number ofcells in the AL and IL different from hormone- and S100-ircells (45). Unfortunately, transferrin immunostaining failedin vibratome sections and whole aggregates, and doublestaining with nestin was not possible. On the other hand,colocalization of nestin and S100 has recently been reportedin human pituitary adenomas (46).
A remarkable characteristic of some of the nestin-ir stain-ing is its occurrence in close proximity to endothelial cells.The pericapillar localization and stellate shape of these nes-tin-ir cells are reminiscent of a pericyte phenotype (15, 47, 48).Pericytes were also found to be nestin positive in the pan-creas (15). As pericytes, nestin-ir cells may regulate capillaryperfusion in the pituitary by their contractile movements(47). Pericytes have also recently been demonstrated to beimportant in angiogenesis (48). Alternatively, the close en-dothelial association of nestin-ir cells may be interpreted inthe context of recent findings that nestin-expressing stemcells can give rise to endothelial cells (49), and that endo-thelial cells seem to be essential players in the stem cell niche(12, 50, 51). In addition to the close proximity of nestin-ir cellsto endothelial cells, we found some endothelial cells thatcontained nestin-ir in the immature (but not adult) pituitary.The expression of nestin in endothelial cells has also beenreported in other tissues during both embryonic and earlypostnatal ages (14, 15, 18, 21, 24) and is strongly reduced inadult organs (15).
In the IL, part of the nestin-ir overlaps with GFAP-ir in theimmature pituitary, whereas virtually all nestin-ir coincideswith GFAP in the adult IL. GFAP in the IL marks stellate glialcells (52–54). The function of these cells is not clear (52), butthey undergo dynamic changes under certain experimentalconditions (53, 54). Nestin may be expressed in these stellateglial cells depending on the activation state. Indeed, nestinexpression has also been observed in stellate cells of othertissues when activated (15, 21–23). In contrast, expression ofGFAP in the developing IL may indicate maturation of theseglial-like (nestin-ir) cells. It has been reported that glial-likeIL cells shift from vimentin to GFAP expression during post-natal development (55).
In the NL, the great majority of nestin-ir cells do notexpress GFAP, a marker for astrocyte-like pituicytes (56), andonly sporadically is nestin-ir localized to endothelial cells. NoDCX was detected within the NL, suggesting that the neu-rohypophysis does not contain neuronal progenitor cells,which, in contrast, have been found in the hypothalamus(38). Because the nucleated cells in the NL mainly representpituicytes, we suggest that nestin is expressed in pituicyteprecursors, and that during differentiation, nestin is replacedby filament proteins of mature pituicytes, such as GFAP.Stage-dependent expression of nestin and GFAP has also
been reported in the developing brain (2, 3). The sporadiccolocalization of nestin and GFAP in (adult) NL cells mayreflect transient overlap of both proteins en route to differ-entiation, which is different from the apparently more per-manent coexpression in (adult) IL cells. The considerablylower expression of nestin in adult vs. immature NL is rec-oncilable with full maturation of pituicytes of adult animals.Also in the AL, GFAP is only very sporadically found to-gether with nestin. GFAP in the AL labels a subset of FS cells(34, 53).
The expression of nestin was also observed in a number ofcells in the layers bordering the pituitary cleft, the lumenremaining from Rathke’s pouch (57). This finding was in-dependent of postnatal age examined. These cells also didnot belong to known cell types; they did not express any ofthe pituitary hormones and markers analyzed. Hormone-,S100-, vimentin-, fibronectin-, and �SMA-positive cells wereintermingled between these bordering cells. The marginalcell layer indeed is not a homogeneous structure. It containscells with different morphologies and ultrastructural fea-tures (57, 58), and S100-ir in the layer has also been reportedby others (35, 59). The marginal cell layers around the clefthave in the past repeatedly been proposed to embody astem/progenitor cell compartment in the postnatal pituitary(57, 60–63), and the presence of nestin may be consistent withsuch an assumption. Compelling evidence, however, is stilllacking.
In our well-characterized aggregate culture system (31),nestin-ir was detected in similar cell types as in the ex vivosections, and most nestin-ir cells also did not belong toknown pituitary cell lineages. Growth factors such as bFGFand LIF, which are well documented to support maintenanceand proliferation of stem/progenitor cells in culture (7, 8, 11,28–30, 64), augmented the number of nestin-ir cells in theaggregates. The partially additive effect of bFGF and LIF maysuggest a partly overlapping, partly different target popu-lation. A remarkable observation was that treatment withbFGF resulted in the formation of hollow bulges in the ag-gregates. These cavities were bordered by a layer of cells,some of which showed nestin-ir, reminiscent of the nestin-ircells around the cleft.
When examined in monolayer culture, nestin-ir also didnot colocalize with hormones or S100. A striking character-istic is that in these cultures, cells that express nestin-ir werefound to move actively and proliferate rapidly. On the basisof their firm attachment to collagen-coated dishes (as appar-ent from the flattened morphology and the prominent focaladhesion points), nestin-ir cells could be cleared from the lessadhering hormone- and S100-positive cells. Adhesive prop-erties have previously been applied by others to isolate pro-genitor cells from certain tissues (65, 66). Nestin-ir cells couldbe additionally enriched because of their proliferative activ-ity, which was much higher than the division rate of the fewremaining hormone- and S100-ir cells. High proliferativeactivity has also been reported in monolayer cell culturesoriginating from pancreatic islets that principally consistedof NIP cells with multipotential capacity (8). The morphologyof the enriched pituitary nestin-ir cells strikingly resemblesthat of NIP cells, with wave-like structures formed in theculture dish, and is also reminiscent of the morphological
2384 Endocrinology, May 2005, 146(5):2376–2387 Krylyshkina et al. • Nestin in the Pituitary

appearance of pluripotent bone marrow stromal cells (mes-enchymal stem cells) in culture, also purified on the basis oftight adhesion (65). Conspicuous structural similarity wasalso found with the rat fetal brain stem cell monolayer cul-tures as used by U et al. (67). Of note, the nestin-ir cellsobtained in culture did not represent activated microglialcells or some motile endothelial cells, because no Ox-42 orvWF/CD31 was detected in these cells, respectively.
Motility is an outstanding characteristic of pituitarynestin-ir cells in culture, which first may be related to theircontractile properties as pericytes. On the other hand, mo-tility and, accordingly, expression of specific adhesion mol-ecules have in the last few years recurrently been linked toa stem/progenitor cell phenotype, allowing such cells to beanchored in a niche and to migrate when needed for forma-tion of new cells during physiological cell turnover or tissuerepair (68, 69). In epidermal stem cells, high levels of �1integrin and low levels of E-cadherin were found to be typ-ical (70). We report here a similar expression pattern of �1integrin and cadherin in cultured nestin-ir cells from the AL,characteristics not present in the other AL cells. Whethernestin-ir cells also exhibit high motility in the pituitary in vivois at present not clear. We found motility in cells shortly afterisolation from the AL and further characterized these cells byBSA gradient separation as small cells of low density. To ourknowledge, no data are available on motile cells within thepituitary gland in situ. It is tempting to speculate that motilecells in the pituitary gland, if present, play a role in formationof new cells under physiological conditions and during tis-sue repair as they do in the brain (68, 69).
The propagated nestin-ir cells in culture contain vimentin,fibronectin, and �SMA and may thus represent mesenchy-mal cells of the pituitary or, alternatively, adopt mesenchy-mal features in culture. In support of the latter hypothesis, weobserved no convincing colocalization of fibronectin, vimen-tin, and �SMA in nestin-ir cells in pituitary sections. More-over, �SMA expression is not present in the nestin-ir cellsafter selection, but only gradually appears in the propagatednestin-ir cells. Mesenchymal features and markers have beenfound in nestin-positive cells in other tissues. For instance, anumber of researchers described nestin-ir cells in the pan-creas as mesenchymal-like cells with no stem/progenitor cellcharacteristics (15, 16, 26). In contrast, nestin-ir cells derivedfrom skin displaying multipotential capacity also containedfibronectin (7). The class III intermediate filament proteinvimentin, although generally considered a mesenchymalmarker, has been reported to be an obligatory partner ofnestin for proper assembly of intermediate filaments, at leastin astrocytes (71). Nestin and vimentin coexpression has beenfound in putative stem/progenitor cells, such as neuro-sphere cells (6), neuroepithelial and radial glial cells (2), andpancreatic islet precursor cells (10), as well as in other celltypes, such as pancreatic mesenchymal and endothelial cells(16, 18, 27). �SMA, a recognized marker of tissue myofibro-blasts, has been described as a partner for nestin in putativepancreatic progenitor cells (72) and has been found in cul-tured papillary cells, which were proposed to play a role asadult kidney stem cells (73). Last, but not least, coexpressionof vimentin and nestin in motile cells may point toward a rolein repair processes, as reported in regenerating skeletal mus-
cle (74). Considering all of these data, it would be temptingto speculate that the enriched nestin-ir cells represent mul-tipotential progenitor cells of the pituitary. However, at-tempts to date to drive differentiation of these cells intopituitary cell phenotypes failed (Krylyshkina, O., and H.Vankelecom, unpublished observations).
Finally, it is at present unclear how our enriched nestin-ircell cultures relate to the pituitary follicular cell culturesreported by Ferrara et al. (75), seminal cell cultures of FS cellsthat have led to the identification and isolation of the growthfactors bFGF and vascular endothelial growth factor. Thereare some striking differences in the culture protocols used,but their impact on the final outcome is unknown. Ferrara etal. (75) immediately seeded cells in high serum conditions.Follicular cell cultures were obtained by natural overgrowthof proliferating cells and did not require extracellular matrixcomponents to be established. In contrast, collagen (not fi-bronectin or laminin) and cell culture during the first days(primary culture) without serum was in our hands essentialto allow selective (dis)attachment for obtaining the almosthomogeneous nestin-ir cell cultures. Moreover, we foundthat the FS cell markers S100 (not checked by Ferrara et al.)and �-Ala-Lys-N�-AMCA do not label nestin-ir cells. In con-trast, FS cells have also been shown to express fibronectin (76)and vimentin (77). However, we did not find substantialcolocalization of vimentin and fibronectin with S100 or�-Ala-Lys-N�-AMCA in pituitary sections (unpublishedobservations).
In conclusion, nestin is expressed in the pituitary gland aswell as in pituitary cell cultures. Within the different lobes,the level of expression and the colocalization profile changewith age. Nestin-ir is not present in hormone and typical FScells, but is present in a variety of other not well definedpituitary cell types, including putative pericytes and someuncharacterized cells around the pituitary cleft. Nestin-ircells rapidly propagate and are strikingly motile in mono-layer cell cultures; at least these nestin-ir cells show somemesenchymal characteristics. The biological significance ofthese enriched nestin-positive cells as well as of the nestin-expressing cells in the pituitary in vivo, particularly in termsof a stem/progenitor cell function or a supportive role, re-mains to be explored.
Acknowledgments
We thank Dr. A. F. Parlow (Harbor-University of California-Los An-geles Medical Center, Torrance, CA) to supply us with the hormoneantisera. We are also very grateful to Dr. R. McKay (National Institutesof Health, Bethesda, MD) for generously providing the polyclonal an-tinestin antiserum, and to Dr. K. Bauer (Max Planck Institute, Hannover,Germany) for supplying the �-Ala-Lys-N�-AMCA dipeptide. We ap-preciate the kind sharing of the Leica live imaging workstation by Drs.P. Zimmermann and G. David (Department of Human Genetics, Uni-versity of Leuven, Leuven, Belgium). We acknowledge the help of NicoleHersmus (own laboratory) with RT-PCR analysis, of Nathalie De Vos(student in Pharmacy, University of Leuven) for confirming the reli-ability of the nestin staining, and of Dr. J. van den Oord (Department ofMorphology and Medical Imaging, University of Leuven) for help in-terpreting the immunocytochemical data.
Received September 10, 2004. Accepted January 20, 2005.Address all correspondence and requests for reprints to: Dr. Hugo
Vankelecom, Laboratory of Cell Pharmacology, Department of Molec-
Krylyshkina et al. • Nestin in the Pituitary Endocrinology, May 2005, 146(5):2376–2387 2385

ular Cell Biology, University of Leuven, Campus Gasthuisberg (O&N),Herestraat 49, B-3000 Leuven, Belgium. E-mail: [email protected].
This work was supported by grants from the Fund for ScientificResearch (Flanders, Belgium) and the Flemish Ministry of Science Policy(Concerted Research Actions).
References
1. Nolan LA, Kavanagh E, Lightman SL, Levy A 1998 Anterior pituitary cellpopulation control: basal cell turnover and the effects of adrenalectomy anddexamethasone treatment. J Neuroendocrinol 10:207–215
2. Hockfield S, Mckay RDG 1985 Identification of major cell classes in thedeveloping mammalian nervous system. J Neurosci 5:3310–3328
3. Lendahl U, Zimmerman LB, Mckay RDG 1990 CNS stem cells express a newclass of intermediate filament protein. Cell 60:585–595
4. Johansson CB, Momma S, Clarke DL, Risling M, Lendahl U, Frisen J 1999Identification of a neural stem cell in the adult mammalian central nervoussystem. Cell 96:25–34
5. Morshead CM, van der Kooy D 2004 Disguising adult neural stem cells. CurrOpin Neurobiol 14:1–7
6. Lobo MVT, Alonso FJM, Redondo C, Lopez-Toledano MA, Caso E, HerranzAS, Paino CL, Reimers D, Bazan E 2003 Cellular characterization of epidermalgrowth factor-expanded free-floating neurospheres. J Histochem Cytochem51:89–103
7. Toma JG, Akhavan M, Fernandes KJL, Barnabe-Heider F, Sadikot A, KaplanDR, Miller FD 2001 Isolation of multipotent adult stem cells from the dermisof mammalian skin. Nat Cell Biol 3:778–784
8. Zulewski H, Abraham EJ, Gerlach MJ, Daniel PB, Moritz W, Muller B,Vallejo M, Thomas MK, Habener JF 2001 Multipotential nestin-positive stemcells isolated from adult pancreatic islets differentiate ex vivo into pancreaticendocrine, exocrine, and hepatic phenotypes. Diabetes 50:521–533
9. Kim SY, Lee SH, Kim BM, Kim EH, Min BH, Bendayan M, Park IS 2004Activation of nestin-positive duct stem (NPDS) cells in pancreas upon neogenicmotivation and possible cytodifferentiation into insulin-secreting cells fromNPDS cells. Dev Dynam 230:1–11
10. Wu FD, Jagir MR, Powell JS 2003 Long-term correction of hyperglycemia indiabetic mice after implantation of cultured human cells derived from fetalpancreas. Blood 102:942A
11. Tropepe V, Coles BLK, Chiasson BJ, Horsford DJ, Elia AJ, McInnes RR, vander Kooy D 2000 Retinal stem cells in the adult mammalian eye. Science287:2032–2036
12. Miura M, Gronthos S, Zhao MR, Lu B, Fisher LW, Robey PG, Shi ST 2003SHED: stem cells from human exfoliated deciduous teeth. Proc Natl Acad SciUSA 100:5807–5812
13. Li LN, Mignone J, Yang M, Matic M, Penman S, Enikolopov G, Hoffman RM2003 Nestin expression in hair follicle sheath progenitor cells. Proc Natl AcadSci USA 100:9958–9961
14. Mokry J, Nemecek S 1999 Cerebral angiogenesis shows nestin expression inendothelial cells. Gen Physiol Biophys 18:25–29
15. Lardon J, Rooman I, Bouwens L 2002 Nestin expression in pancreatic stellatecells and angiogenic endothelial cells. Histochem Cell Biol 117:535–540
16. Humphrey RK, Bucay N, Beattie GM, Lopez A, Messam CA, Cirulli V,Hayek A 2003 Characterization and isolation of promoter-defined nestin-positive cells from the human fetal pancreas. Diabetes 52:2519–2525
17. Treutelaar MK, Skidmore JM, Dias-Leme CL, Hara M, Zhang LZ, SimeoneD, Martin DM, Burant CF 2003 Nestin-lineage cells contribute to the micro-vasculature but not endocrine cells of the islet. Diabetes 52:2503–2512
18. Klein T, Ling ZD, Heimberg H, Madsen OD, Heller RS, Serup P 2003 Nestinis expressed in vascular endothelial cells in the adult human pancreas. J His-tochem Cytochem 51:697–706
19. Delacour A, Nepote V, Trumpp A, Herrera PL 2004 Nestin expression inpancreatic exocrine cell lineages. Mech Dev 121:3–14
20. Joanette EA, Reusens B, Arany E, Thyssen S, Remacle RC, Hill DJ 2004Low-protein diet during early life causes a reduction in the frequency of cellsimmunopositive for nestin and CD34 in both pancreatic ducts and islets in therat. Endocrinology 145:3004–3013
21. Frisen J, Johansson CB, Torok C, Risling M, Lendahl U 1995 Rapid, wide-spread, and long-lasting induction of nestin contributes to the generation ofglial scar tissue after CNS injury. J Cell Biol 131:453–464
22. Clarke SR, Shetty AK, Bradley JL, Turner DA 1994 Reactive astrocytes ex-press the embryonic intermediate neurofilament nestin. Neuroreport 5:1885–1888
23. Niki T, Pekny M, Hellemans K, De Bleser P, Van den Berg K, Vaeyens F,Quartier E, Schuit F, Geerts A 1999 Class VI intermediate filament proteinnestin is induced during activation of rat hepatic stellate cells. Hepatology29:520–527
24. Vanderwinden JM, Gillard K, De Laet MH, Messam CA, Schiffmann SN2002 Distribution of the intermediate filament nestin in the muscularis propriaof the human gastrointestinal tract. Cell Tissue Res 309:261–268
25. Lobo MV, Arenas MI, Alonso FJ, Gomez G, Bazan E, Paino CL, Fernandez
E, Fraile B, Paniagua R, Moyano A, Caso E 2004 Nestin, a neuroectodermalstem cell marker molecule, is expressed in Leydig cells of the human testis andin some specific cell types from human testicular tumours. Cell Tissue Res316:369–376
26. Gao R, Ustinov J, Pulkkinen MA, Lundin K, Korsgren O, Otonkoski T 2003Characterization of endocrine progenitor cells and critical factors for theirdifferentiation in human adult pancreatic cell culture. Diabetes 52:2007–2015
27. Street CN, Lakey JRT, Seeberger K, Helms L, Rajotte RV, Shapiro AMJ,Korbutt GS 2004 Heterogenous expression of nestin in human pancreatictissue precludes its use as an islet precursor marker. J Endocrinol 180:213–225
28. Reynolds BA, Weiss S 1992 Generation of neurons and astrocytes from iso-lated cells of the adult mammalian central nervous system. Science 255:1707–1710
29. Morshead CM, Reynolds BA, Craig CG, Mcburney MW, Staines WA, Mo-rassutti D, Weiss S, Vanderkooy D 1994 Neural stem cells in the adultmammalian forebrain: a relatively quiescent subpopulation of subependymalcells. Neuron 13:1071–1082
30. Gritti A, Parati EA, Cova L, Frolichsthal P, Galii R, Wanke E, Faravelli L,Morassutti DJ, Roisen F, Nickel DD, Vescovi AL 1996 Multipotential stemcells from the adult mouse brain proliferate and self-renew in response to basicfibroblast growth factor. J Neurosci 16:1091–1100
31. Denef C, Maertens P, Allaerts W, Mignon A, Robberecht W, Swennen L,Carmeliet P 1989 Cell-to-cell communication in peptide target cells of anteriorpituitary. Methods Enzymol 168:47–71
32. Vankelecom H, Matthys P, Denef C 1997 Inducible nitric oxide synthase inthe anterior pituitary gland: induction by interferon-� in a subpopulation offolliculostellate cells and in an unidentifiable population of non-hormone-secreting cells. J Histochem Cytochem 45:847–857
33. Otto C, Dieck T, Bauer K 1996 Dipeptide uptake by adenohypophysealfolliculo-stellate cells. Am J Physiol 271:C210–C217
34. Allaerts W, Carmeliet P, Denef C 1990 New perspectives in the function ofpituitary folliculo-stellate cells. Mol Cell Endocrinol 71:73–81
35. Cocchia D, Miani N 1980 Immunocytochemical localization of the brain-specific S-100 protein in the pituitary gland of adult rat. J Neurocytol 9:771–782
36. Muller AM, Hermanns MI, Skrzynski C, Nesslinger M, Muller KM, Kirk-patrick CJ 2002 Expression of the endothelial markers PECAM-1, vWf, andCD34 in vivo and in vitro. Exp Mol Pathol 72:221–229
37. Sato T, Inoue K 2000 Dendritic cells in the rat pituitary gland evaluated by theuse of monoclonal antibodies and electron microscopy. Arch Histol Cytol63:291–303
38. Markakis EA, Palmer TD, Randolph-Moore L, Rakic P, Gage FH 2004 Novelneuronal phenotypes from neural progenitor cells. J Neurosci 24:2886–2897
39. Gleeson JG, Lin PT, Flanagan LA, Walsh CA 1999 Doublecortin is a micro-tubule-associated protein and is expressed widely by migrating neurons. Neu-ron 23:257–271
40. Tsuchida T, Nagao S, Ohmoto T 1991 The fine-structure of the S-100 proteinpositive cells in the rat pituitary gland: an immunoelectron microscopic study.Brain Res 564:164–166
41. Vankelecom H, Denef C 1997 Paracrine communication in the anterior pi-tuitary as studied in reaggregate cell cultures. Microsc Res Technol 39:150–156
42. Soji T, Mabuchi Y, Kurono C, Herbert DC 1997 Folliculo-stellate cells andintercellular communication within the rat anterior pituitary gland. MicroscRes Technol 39:138–149
43. Allaerts W, Jeucken PHM, Debets R, Hoefakker S, Claassen E, Drexhage HA1997 Heterogeneity of pituitary folliculo-stellate cells: implications for inter-leukin-6 production and accessory function in vivo. J Neuroendocrinol 9:43–53
44. Inoue K, Couch EF, Takano K, Ogawa S 1999 The structure and function offolliculo-stellate cells in the anterior pituitary gland. Arch Histol Cytol 62:205–218
45. Tilemans D, Vande Vijver V, Verhoeven G, Denef C 1995 Production oftransferrin-like immunoreactivity by rat anterior pituitary and intermediatelobe. J Histochem Cytochem 43:657–664
46. Osamura Y, Ishii Y, Messam CA, Umeoka K, Sanno N, Teramoto A, Ex-pression of neural stem cell marker nestin in human pituitary glands andpituitary adenomas. Program of the 84th Annual Meeting of The EndocrineSociety, San Francisco, CA, 2002, Poster 2–597
47. Kawamura H, Kobayashi M, Li Q, Yamanishi S, Katsumura K, Minami M,Wu DM, Puro DG 2004 Effects of angiotensin II on the pericyte-containingmicrovasculature of the rat retina. J Physiol 561:671–683
48. Gerhardt H, Betsholtz C 2003 Endothelial-pericyte interactions in angiogen-esis. Cell Tissue Res 314:15–23
49. Amoh Y, Li L, Yang M, Moossa AR, Katsuoka K, Penman S, Hoffman RM2004 Nascent blood vessels in the skin arise from nestin-expressing hair-folliclecells. Proc Natl Acad Sci USA 101:13291–13295
50. Palmer TD, Willhoite AR, Gage FH 2000 Vascular niche for adult hippocam-pal neurogenesis. J Comp Neurol 425:479–494
51. Shen Q, Goderie SK, Jin L, Karanth N, Sun Y, Abramova N, Vincent P,Pumiglia K, Temple S 2004 Endothelial cells stimulate self-renewal and ex-pand neurogenesis of neural stem cells. Science 304:1338–1340
52. Boyd WH 1987 Nonsecretory neuronal elements in the bovine pituitary in-termediate lobe. Anatomischer Anzeiger 164:117–128
53. Gary KA, Chronwall BM 1995 Regulation of GFAP expression in glial-like
2386 Endocrinology, May 2005, 146(5):2376–2387 Krylyshkina et al. • Nestin in the Pituitary

cells of the rat pituitary intermediate lobe by lactation, salt-loading, and ad-renalectomy. Glia 13:272–282
54. Redecker P, Morgenroth C 1989 Comparative immunohistochemical study ofthe presence of glial fibrillary acidic protein in the pituitary of several verte-brates. Anatomischer Anzeiger 168:37–47
55. Gary KA, Sands SA, Chronwall BM 1995 Glial-like cells of the rat pituitaryintermediate lobe change morphology and shift from vimentin to GFAP ex-pression during development. Int J Dev Neurosci 13:555–565
56. Suess U, Pliska V 1981 Identification of the pituicytes as astroglial cells byindirect immunofluorescence staining for the glial fibrillary acidic protein.Brain Res 221:27–33
57. Correr S, Motta PM 1981 The rat pituitary cleft: a correlated study by scanningand transmission electron microscopy. Cell Tissue Res 215:515–529
58. Soji T, Yashiro T, Herbert DC 1989 Granulated ‘marginal cell layer’ in the ratanterior pituitary gland. Tissue Cell 21:849–856
59. Shirasawa N, Kihara H, Yamaguchi S, Yoshimura F 1983 Pituitary folliculo-stellate cells immunostained with S-100 protein anti-serum in postnatal, cas-trated and thyroidectomized rats. Cell Tissue Res 231:235–249
60. Yoshimura F, Soji T, Sato S, Yokoyama M 1977 Development and differen-tiation of rat pituitary follicular cells under normal and some experimentalconditions with special reference to an interpretation of renewal cell system.Endocrinol Jpn 24:435–449
61. Gon G, Nakamura F, Ishikawa H 1987 Cystlike structures derived from themarginal cells of Rathke’s cleft in rat pituitary grafts. Cell Tissue Res 250:29–33
62. Wilson DB 1986 Distribution of 3H-thymidine in the postnatal hypophysis ofthe C57BL mouse. Acta Anat 126:121–126
63. Carbajo S, Hernandez JL, Carbajoperez E 1992 Proliferative activity of cellsof the intermediate lobe of the rat pituitary during the postnatal period. TissueCell 24:829–834
64. Williams RL, Hilton DJ, Pease S, Willson TA, Stewart CL, Gearing DP,Wagner EF, Metcalf D, Nicola NA, Gough NM 1988 Myeloid leukemia in-hibitory factor maintains the developmental potential of embryonic stem cells.Nature 336:684–687
65. Hung SC, Chen NJ, Hsieh SL, Li H, Ma HL, Lo WH 2002 Isolation andcharacterization of size-sieved stem cells from human bone marrow. Stem Cells20:249–258
66. Lim DA, Alvarez-Buylla A 1999 Interaction between astrocytes and adult
subventricular zone precursors stimulates neurogenesis. Proc Natl Acad SciUSA 96:7526–7531
67. U HS, Alilain W, Saljooque F 2002 Fetal brain progenitor cells transdiffer-entiate to fates outside the nervous system. Mol Endocrinol 16:2645–2656
68. Imitola J, Raddassi K, Park KI, Mueller FJ, Nieto M, Teng YD, Frenkel D,Li J, Sidman RL, Walsh CA, Snyder EY, Khoury SJ Directed migration ofneural stem cells to sites of CNS injury by the stromal cell-derived factor1�/CXC chemokine receptor 4 pathway. Proc Natl Acad Sci USA 101:18117–18122
69. Prestoz L, Relvas JB, Hopkins K, Patel S, Sowinski P, Price J, ffrench-Constant C 2001 Association between integrin-dependent migration capacityof neural stem cells in vitro and anatomical repair following transplantation.Mol Cell Neurosci 18:473–484
70. Moles JP, Watt FM 1997 The epidermal stem cell compartment: variation inexpression levels of E-cadherin and catenins within the basal layer of humanepidermis. J Histochem Cytochem 45:867–874
71. Eliasson C, Sahlgren C, Berthold CH, Stakeberg J, Celis JE, Betsholtz C,Eriksson JE, Pekny M 1999 Intermediate filament protein partnership inastrocytes. J Biol Chem 274:23996–24006
72. Taguchi M, Otsuki M 2004 Co-localization of nestin and PDX-1 in smallevaginations of the main pancreatic duct in adult rats. J Mol Histol 35:785–789
73. Oliver JA, Maarouf O, Cheema FH, Martens TF, Al Awqati Q 2004 The renalpapilla is a niche for adult kidney stem cells. J Clin Invest 114:795–804
74. Vaittinen S, Lukka R, Sahlgren C, Hurme T, Rantanen J, Lendahl U, ErikssonJE, Kalimo H 2001 The expression of intermediate filament protein nestin asrelated to vimentin and desmin in regenerating skeletal muscle. J NeuropatholExp Neurol 60:588–597
75. Ferrara N, Schweigerer L, Neufeld G, Mitchell R, Gospodarowicz D 1987Pituitary follicular cells produce basic fibroblast growth factor. Proc Natl AcadSci USA 84:5773–5777
76. Liu YC, Tanaka S, Inoue K, Kurosumi K 1989 Localization of fibronectin inthe folliculo-stellate cells of the rat anterior pituitary by the double bridgeperoxidase-antiperoxidase method. Histochemistry 92:43–45
77. Marin F, Boya J, Lopez-Carbonell A 1989 Immunocytochemical localizationof vimentin in stellate cells (folliculo-stellate cells) of the rat, cat and rabbitpituitary pars distalis. Anat Embryol 179:491–495
Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
Krylyshkina et al. • Nestin in the Pituitary Endocrinology, May 2005, 146(5):2376–2387 2387





























