Embed Size (px)
Citation preview

J TTISSN 0974-7907 (Online) ISSN 0974-7893 (Print)
26 December 2018 (Online & Print)Vol. 10 | No. 15 | 12907–13046
Building evidence for conservation globally Journal of Threatened Taxa
10.11609/jott.2018.10.15.12907-13046www.threatenedtaxa.org
OPEN ACCESS

EDITORSFounder & Chief EditorDr. Sanjay Molur, Coimbatore, India
Managing EditorMr. B. Ravichandran, Coimbatore, India
Associate EditorsDr. B.A. Daniel, Coimbatore, India Dr. Ulrike Streicher, Wildlife Veterinarian, Danang, Vietnam Ms. Priyanka Iyer, Coimbatore, IndiaDr. Manju Siliwal, Dehra Dun, IndiaDr. Meena Venkataraman, Mumbai, India
Editorial AdvisorsMs. Sally Walker, Coimbatore, India Dr. Robert C. Lacy, Minnesota, USA Dr. Russel Mittermeier, Virginia, USA Dr. Thomas Husband, Rhode Island, USA Dr. Jacob V. Cheeran, Thrissur, India Prof. Dr. Mewa Singh, Mysuru, IndiaMr. Stephen D. Nash, Stony Brook, USA Dr. Fred Pluthero, Toronto, Canada Dr. Martin Fisher, Cambridge, UK Dr. Ulf Gärdenfors, Uppsala, Sweden Dr. John Fellowes, Hong Kong Dr. Philip S. Miller, Minnesota, USA Prof. Dr. Mirco Solé, Brazil
Editorial Board Subject Editors 2015–2017Aaron Bauer, Villanova University, Villanova, USAAmazonas Chagas Júnior, Universidade Federal de Mato Grosso, Cuiabá, Brasil Anders G.J. Rhodin, Chelonian Research Foundation, Lunenburg, USA A. Biju Kumar, University of Kerala, Thiruvananthapuram, IndiaA.J. Solomon Raju, Andhra University, Visakhapatnam, IndiaA.J.T. Johnsingh, Nature Conservation Foundation, Mysuru, IndiaAlbert G. Orr, Griffith University, Nathan, AustraliaAlessandre Pereira Colavite, Universidade Federal da Paraíba, BrazilAlexi Popov, National Museum of Natural History, Sofia, BulgariaAlexander Ereskovsky, IMBE, Marseille, FranceAndreas Köhler, Universidade de Santa Cruz do, BrazilAngela R. Glatston, Rotterdam Zoo, The Netherlands Anjana Silva, Rajarata University of Sri Lanka, Saliyapura, Sri LankaAnkita Gupta, ICAR-NBAIR, Bengaluru, IndiaAnnemarie Ohler, Muséum national d’Histoire naturelle, Paris, FranceAnsie Dippenaar-Schoeman, University of Pretoria, Queenswood, South AfricaAntonio D. Brescovit, Instituto Butantan, BrasilAntonio A. Mignucci-Giannoni, Universidad Interamericana de Puerto Rico, Puerto RicoAnwaruddin Chowdhury, The Rhino Foundation for nature in North East India, Guwahati, IndiaAparna Watve, Tata Institute of Social Sciences, Osmanabad, IndiaArthur Y.C. Chung, Sabah Forestry Department, Sandakan, Sabah, MalaysiaAsheesh Shivam, Nehru Gram Bharti University, Allahabad, India Ashwin Naidu, University of Arizona, Tucson, USA B.C. Choudhury (Retd.), Wildlife Institute of India, Dehradun, India. B. Ravi Prasad Rao, Sri Krishnadevaraya University, Anantpur, IndiaB. Shivaraju, Bengaluru, Karnataka, IndiaB.S. Kholia, Botanical Survey of India, Gangtok, Sikkim, IndiaBolívar R. Garcete-Barrett, FACEN, Universidad Nacional de Asunción, ParaguayBrett C. Ratcliffe, University of Nebraska, Lincoln, USABrian Fisher, California Academy of Sciences, USAC. Raghunathan, Zoological Survey of India, Andaman and Nicobar IslandsC. Srinivasulu, Osmania University, Hyderabad, IndiaCarl Ferraris, Smithsonian Institution, Portland, USACarol Inskipp, Bishop Auckland Co., Durham, UKCecília Kierulff, Victorville , CaliforniaCecilia Volkmer Ribeiro, Porto Alegre, Brazil.Chris Bowden, Royal Society for the Protection of Birds, Sandy, UK
Christoph Kueffer, Institute of Integrative Biology, Zürich, SwitzerlandChristoph Schwitzer, University of the West of England, Clifton, Bristol, BS8 3HAChristopher L. Jenkins, The Orianne Society, Athens, Georgia Cleofas Cervancia, Univ. of Philippines Los Baños College Laguna, PhilippinesColin Groves, Australian National University, Canberra, AustraliaCrawford Prentice, Nature Management Services, Jalan, MalaysiaC.T. Achuthankutty, Scientist-G (Retd.), CSIR-National Institute of Oceanography, Goa Dan Challender, University of Kent, Canterbury, UK D.B. Bastawade, Maharashtra, IndiaD.J. Bhat, Retd. Professor, Goa University, Goa, IndiaDale R. Calder, Royal Ontaro Museum, Toronto, Ontario, CanadaDaniel Brito, Federal University of Goiás, Goiânia, BrazilDavid Mallon, Manchester Metropolitan University, Derbyshire, UKDavid Olson, Zoological Society of London, UKDavor Zanella, University of Zagreb, Zagreb, CroatiaDeepak Apte, Bombay Natural Hisotry Society, Mumbai, IndiaDiana Doan-Crider, Texas A&M University, Texas, USA Dietmar Zinner, German Primate Center, Göttingen, GermanyDunston P.Ambrose, St. Xavier’s College, Palayamkottai, IndiaE. Vivekanandan, Central Marine Fisheries Research Institute, Kochi, IndiaEduard Vives, Museu de Ciències Naturals de Barcelona, Terrassa, SpainEric Smith, University of Texas, Arlington, USAErin Wessling, Max Planck Institute for Evolutionary Anthropology, GermanyErrol Vela, University of Montpellier, Montpellier, France Farkhanda Manzoor Dugal, Lahore College for Women University, PakistanF.B. Vincent Florens, University of Mauritius, MauritiusFerdinando Boero, Università del Salento, Lecce, Italy Francesco Dal Grande, Senckenberg Gesellschaft für Naturforschung, FrankfurtFrederic H. Martini, University of Hawaii at Manoa, Hanolulu, Hawaii. George Mathew, Kerala Forest Research Institute, Peechi, IndiaGernot Vogel, Heidelberg, GermanyGiovanni Amori, CNR - Institute of Ecosystem Studies, Rome, Italy Gombobaatar Sundev, Professor of Ornithology, Ulaanbaatar, MongoliaG.P. Mandal, Zoological Survey of India, Kolkata, India G.P. Sinha, Botanical Survey of India, Allahabad, India Hari Balasubramanian, EcoAdvisors, Nova Scotia, CanadaHayrünisa Baş Sermenli, Muğla University, Kötekli, Turkey H.C. Nagaveni, Institute of Wood Science and Technology, Bengaluru, IndiaH.C. Paulo Corgosinho, Bairro Universitário, Frutal, BrazilH.N. Kumara, Salim Ali Centre for Ornithology and Natural History, Anaikatty, India H. Raghuram, The American College, Madurai, IndiaHector Barrios-Garrido, James Cook University, Townsville, Australia Heidi S. Riddle, Riddle’s Elephant and Wildlife Sanctuary, Arkansas, USAHem Sagar Baral, Charles Sturt University, NSW AustraliaHemant V. Ghate, Modern College, Pune, IndiaHeok Hee Ng, National University of Singapore, Science Drive, SingaporeHimender Bharti, Punjabi University, Patiala, IndiaHui Xiao, Chinese Academy of Sciences, Chaoyang, ChinaIan J. Kitching, Natural History Museum, Cromwell Road, UKIan Redmond, UNEP Convention on Migratory Species, Lansdown, UKIndraneil Das, Sarawak, MalaysiaIvana Karanovic, Hanyang University, Seoul, Korea J. Jerald Wilson, King Abdulaziz University, Jeddah, Saudi ArabiaJ.W. Duckworth, IUCN SSC, Bath, UKJack Tordoff, Critical Ecosystem Partnership Fund, Arlington, USAJan Zima, Institute of Vertebrate Biology, Brno, Czech Republic James M. Carpenter, American Museum of Natural History, New York, USA James Young, Hong Kong Lepidopterists’ Society, Hong KongJean-Pierre Boudot, University of Lorraine, Nancy, France Jeff McNeely, IUCN, Gland, SwitzerlandJesse Leland, Southern Cross University, New South Wales, AustraliaJill Pruetz, Iowa State University, Ames, USA Jim Sanderson, Small Wild Cat Conservation Foundation, Hartford, USA Jodi L. Sedlock, Lawrence University, Appleton, USA John C. Morse, Clemson University, Long Hall, Clemson, USAJohn Huber, Canadian National Collection of Insects, Ontario, Canada. John Noyes, Natural History Museum, London, UKJohn Veron, Coral Reef Foundation, Townsville, Australia
ISSN 0974-7907 (Online); ISSN 0974-7893 (Print)
Published by Typeset and printed at Wildlife Information Liaison Development Society Zoo Outreach Organization
No. 12, Thiruvannamalai Nagar, Saravanampatti - Kalapatti Road, Saravanampatti, Coimbatore, Tamil Nadu 641035, India
Ph: 0 938 533 9863Email: [email protected], [email protected]
www.threatenedtaxa.org
continued on the back inside cover
Caption: Owls are amazing animals that are severely threatened by superstitions. This pen illustration by Priyanka Iyer, Zoo Outreach Organisation, is to celebrate not only their beauty but also all other threatened species.
Caption: Diverse! Colourful! Almost Magical! Beetles are all around us from ancient cultures, art and entertainment to being considered pests, food, medicine, biodiversity indicators and even inspiration for technologies. This pencil and pen sketch by Priyanka Iyer, Zoo Outreach Organization is to celebrate Beetles.

12907
Arti
cle
DOI: https://doi.org/10.11609/jott.4347.10.15.12907-12915
Editor: Mewa Singh, University of Mysore, Mysuru, India Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 4347 | Received 20 June 2018 | Final received 04 December 2018 | Finally accepted 19 December 2018
Citation: Ganguly, I & N.S. Chauhan (2018). Dietary preference and feeding patterns of the urban Rhesus Macaque Macaca mulatta (Mammalia: Primates: Cerco-pithecidae) in Asola-Bhatti Wildlife Sanctuary in India. Journal of Threatened Taxa 10(15): 12907–12915; https://doi.org/10.11609/jott.4347.10.15.12907-12915
Copyright: © Ganguly & Chauhan 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, repro-duction and distribution by providing adequate credit to the authors and the source of publication.
Funding: This study was funded by World Wide Fund for Nature - Small Grant Programme (2016–2018), India.
Competing interests: The authors declare no competing interests.
Author Details: Ishita Ganguly, B.Sc (Hons.), MSc, MPhil in Zoology and presently pursuing doctoral degree in Wildlife Science from Amity Institute of Forestry and Wildlife, Amity University Noida, India. I have been working on ecology of Rhesus Macaque and human-macaque conflict in urban landscape since 2015. My project is funded by WWF Small grant programme (2016–2018). Currently, writing my thesis and also preparing for attending students conference (SSCS Cambridge, UK) 2019 in United Kingdom. Dr. N.P.S. Chauhan, MSc & PhD, Zoology from Delhi University, actively involved in teaching, training and research in Delhi University colleges, North-Eastern Hill University, Shillong, Wildlife Institute of India, Dehradun and now in Amity University, Noida and presently serving as Director of Amity Institute of Forestry and Wildlife. He is the main supervisor of many PhD students in Amity University.
Author Contribution: IG wrote the project, raised funding, completed field research, worked on data analysis, writing manuscript and communication. NSC contributed in planning of research, writing the manuscript and revising.
Acknowledgements: We thank Mr. A.K. Shukla, Chief Wildlife Warden,Delhi Forest and Wildlife Department, for issuing the permit to work in the field area and Mr. S.K. Muan Giete, Deputy Conservator of Forest, South Division, Delhi, for providing support for the smooth conduct of the field research. We also thank Prof. Dr. Praful Singh and Ms. Pradipika Verma, Amity Institute of GIS and Remote Sensing,for designing the maps though GIS software and data imaging and Dr. G.S. Rawat, Wildlife Institute of India, for identifying the plant species specimens.
Dietary preference and feeding patterns of the urban Rhesus Macaque Macaca mulatta (Mammalia: Primates: Cercopithecidae) in Asola-Bhatti Wildlife Sanctuary in India
Ishita Ganguly 1 & Netrapal Singh Chauhan 2
1,2 Amity Institute of Forestry and Wildlife, Amity University Campus, Sector 125, Gautam Buddha Nagar, Noida, Uttar Pradesh 201303, India1 [email protected] (corresponding author), 2 [email protected]
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12907–12915
Abstract: We studied the feeding patterns and discrete spatio-temporal food habitsof16 groups of the urban Rhesus Macaque Macaca mulatta following their relocation in Asola-Bhatti Wildlife Sanctuary near Delhi, India. We observed that the macaques fed on 31 plant species, with Prosopis juliflora and P. cineraria appearing in most scans. We classified the food consumed by the species into six main categories the species and recorded the average time spent on each of these throughout the year. The maximum time was spent on supplementary feeding provided by the forest department and the minimum on natural plant resources. There was a significant difference in the consumption of different food categories from morning to evening but there were no significant seasonal variations. This study showed that Rhesus Macaque adopted different foraging strategies based on the availability of resources in their new environment and that variety in food resources buffered seasonality in their diet. Information on their feeding patterns and food habits will help in developing management protocols for the primates in urban environments.
Keywords: Feeding ecology, Delhi NCR, management, opportunistic feeding, primates, relocation, urban landscape.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12907–12915
Dietary preference and feeding patterns of the urban Rhesus Macaque Ganguly & Chauhan
12908
INTRODUCTION
Rhesus Macaque Macaca mulatta is the most common non-human primate in the forested and urban areas of Asia (Hasan et al. 2013). It is found throughout India in its peninsular (Madhya Pradesh, West Bengal, and Assam), northern (Jammu & Kashmir, Himachal Pradesh, Punjab, Haryana, Uttar Pradesh, Rajasthan, and Gujarat) (Seth et al. 2001), and northeastern (Assam, Meghalaya, and Arunachal Pradesh; (Molur et al. 2003) regions. Information on the feeding ecology of a species provides the detailed dietary specialization necessary for its survival and is an important part of its natural life history (Harcourt et al. 2002). Flexibility in diet patterns plays a pivotal role in the survival of non-human primate species in urban and peri-urban ecosystems through resource sharing and competition and has evolutionary implications in the long-term. Primates are known to adopt several foraging strategies (Fleagle & Gilbert 2006). Natural diet of forest Rhesus Macaques includes fruits, seeds, inflorescences, flowers, buds, leaves, young shoots, twigs, barks, roots, and pith and resin of gymnosperms, angiosperms, and fungi (Fooden 2000). Macaques are also known to consume animal food items that such as insects, spiders, worms, termites, grasshoppers, lizards, ants, beetles, molluscs, crayfish, shellfish, honeycombs, crabs, and bird eggs (Mandal 1964; Lindburg 1971; Malik 1983). In marine coastal areas, the species is mostly known to rely on seeds and fruits (Hanya et al. 2003) and also catch live fish as in the Sunderban (Majumder et al. 2012). In forested habitats,primates consume 25% to 40% of the total frugivore biomass (Chapman et al. 1995). Ingestion of fruits or young leaves with sugar and insects with protein content help to balance their diet (Janson & Chapman 1999). Feeding patterns are also associated with human-macaque conflict—crop raiding by macaques in villages near forest areas has increased the level of negative association of the species by farmers (Air 2015). The dependency of urban macaques on anthropogenic food resources and their behaviours associated with food utilisation from urban areas often increase the risk of undesirable interactions with human beings (Sha & Hanya 2013).
In urban landscapes, the nutrition required for primates becomes highly questionable. Urban macaques largely share human food resources (Gupta 2001) and depend on cultivated crops, plants, and even garbage (Lee et al. 1986). In some cases, macaques depend on humans for being fed (Strum 1994)—the feeding patterns of primates that live in tourist sites and temples
are often influenced by provisioning of food by humans. Urban macaques have also acquired behavioural adaptations in food-acquisition techniques (Mangalam & Singh 2013). Urban habitats, in contrast to natural ones, have a more direct influence on primate behaviours associated with competitive resource utilizations and foraging techniques. Several anthropogenic barriers and disturbances interfere with the feeding ecology of primates in urban environments.
Although Rhesus Macaques were assessed as a Least Concern (LC) species by IUCN (2018), primates are threatened globally by human-wildlife negative interactions, habitat loss and fragmentation, and several other anthropogenic factors (Strum 1994, 2001; Mittermeier & Konstant 1996, 1997; Kemf & Wilson 1997; Cowlishaw & Dunbar 2000; Peterson 2003; Hill 2005). The Negative interactions between humans and macaques due to food provisioning and other anthropogenic drivers possess major challenges for the survival and persistence of the species. The need for translocation of Rhesus Macaques and the consequences were due to its proliferation in urban areas of India was suggested and studied earlier (Malik & Johnson 1991, 1994; Southwick et al. 1998). Translocation or relocation is a widely used conservation tool but it is known to induce stress, as evident in the higher level of stress hormones in females of the species during the translocation process (Aguilar-Cucurachi et al. 2010).
The reproductive capacity, inter-birth interval, and the size of social groups in primates are often determined by the amount of food they consume (Air 2015). Again, the availability of different food resources can reduce seasonal fluctuations in diet and provisioning of food regularly to urban primates may have adverse effects on their behaviour, social organization, and conservation (Sinha&Vijayakrishnan 2017). The aim of this study was to investigate the food habits and feeding patterns of urban macaques in and around (0–1 km) Asola-Bhatti Wildlife Sanctuary in Delhi. The characteristic features of this sanctuary such as the availability of food resources infringe villages and food provisioning by the public have played an influential roles in the feeding ecology of its Rhesus Macaques. Information of dietary patterns of urban macaques will enhance the knowledge of its natural history and survival and that will help in the management of the species in urban ecosystems.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12907–12915
Dietary preference and feeding patterns of the urban Rhesus Macaque Ganguly & Chauhan
12909
MATERIALS AND METHODS
Study area
Asola-Bhatti Wildlife Sanctuary is situated in South Delhi District (28.410–28.490 N and 77.190N–77.270 E) and covers a total area of about 6,874ha and there is a high density of Acacia pendula,scrub forests, and trees with a short diameters (Kushwaha et al. 2014) (Fig. 1). The forest area is located at the foothills of the Aravalli range and is about 16km long and 4.3km wide, with elevation ranging from 235–288 m. The forest is surrounded by hilly areas with shrubs, stunted trees, and moderate density forest cover and is adjacent to the urban areas of Delhi-Haryana interstate border region, Sangam Vihar, Faridabad (Surajkund Road), Pali Village, Satberi, Deragaon, Fatepurberi, and Anangpur. This protected area has semi-arid vegetation with xerophytic plants and several
large, deep pits. The largest water-filled pit is Neeli-Jheel, situated 3km from Gate No. 7, where urban monkeys are intermittently released after capture since 2007.
The dominant tree species in Asola-Bhatti Wildlife Sanctuary are Prosopis juliflora, a native species introduced to counter forest degradation (Burkart 1976; Pasiecznik et al. 2001), which and present at high (7.68%), moderate (16.03%), and low (47.90%) densities,and Anogeissus pendula and Acacia nilotica, present in forest plantation, scrub vegetation (12.04%), water bodies (0.16%), and human settlements (2.92%) (Kushwaha et al. 2014). There were no Rhesus Macaques in the area before translocation began and the present population is derived entirely from relocated animals alone. This area exhibits extreme fluctuation in annual temperature, with summer highs in May (43–47 0C) and winter lows in January (60C). June to September is the wet season with an average annual rainfall of about 617mm. This sanctuary is composed of
Figure 1. Asola-Bhatti Wildlife Sanctuary in Delhi, India
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12907–12915
Dietary preference and feeding patterns of the urban Rhesus Macaque Ganguly & Chauhan
12910
Asola Village in the north and Bhatti area in the south. It is a man-made sanctuary and the only protected area in Delhi. Most of the area of the sanctuary is degraded with the prevalence of xerophytic plants (Khanna & Sati 2003). Vegetation shows remarkable dominance of shrubs and stunted trees (Naithani et al. 2006).
Study groupsWe sighted a total of 16 groups of translocated
Rhesus Macaques in the area. Observations were made from a close distance of <10m and data on demography and food habits were collected from 06.00–18.00 hr daily from May 2016 to June 2017. Group size varied from 14 to 63 individuals per group (30.57±2.67) and a total of 492 (n) individuals were observed (Table 1).
Food categoriesWe classified the food consumed by the relocated
Rhesus Macaques into categories based on variations in resource availability:
i) Natural plant species: The natural plants, trees, herbs, and shrubs available in the sanctuary.
ii) Supplementary foods: Food given by the forest department daily in this sanctuary for feeding the Rhesus Macaques only (seasonal vegetables and fruits; 2500kg/day).
iii) Provisioned food: Banana and roadside food (bread and chick-pea) thrown by the public to the macaques daily; the macaques often travel to the boundary walls, cross it, and sit on the highway it to have these itemsthe the.
iv) Anthropogenic food: Garbage and human food resources (Indian bread, oily fries, and potato chips) snatched by the macaques daily from human settlements situated within 0.5km of the protected area.
v) Water: Water from channelsmade in the sanctuary exclusively for Rhesus Macaques (a total of 36 in number).
vi) Others: Insects, soil, lizards, and bird eggs.
Scan sampling techniqueInstantaneous or scan sampling (Altmann 1974) was
used to gather information on the feeding habits and food items of the macaques. Group scans were taken on all visible members of the group for 5min at every 10-minute interval. We recorded 13,740 scan samples and the type of food items eaten (young leaves,mature leaves, roots, stems, flowers, fruits, shoots, gum, bark, or animal prey). We collected the unidentified species for taxonomic identification (leaves, stem, and fruits) through herbarium in the Wildlife Institute of India,
Dehradun.
Focal sampling techniqueWe focused on individuals (adult male/ adult female/
juvenile/ infant) and made 12 entries per hour of theiractivitiesins. We recorded the time spent by that focal individualson each food plant and the parts eaten along with thetime spent at different feeding sites. We recorded 13,874 focal samples and categorized the different plant parts eaten by the macaques.
We estimated the time spent feeding on different food items in a day as per theformula by Gupta &Kumar (1994):
Ta = Na / N x 100,where Ta is the percentage of time spent on an
activity a, Na is the number of records with activity a, andN is the total number of records for the day.
Analysis: Analysis of variance (ANOVA) was used to compare the feeding time on food categories and the number of food plants eaten monthly and seasonally (Simpson et al. 1960). Independent sample t-test was used to analyse the difference in the average time spent on each category. Chi square test was performed to compare the association between groups. Microsoft Excel 2010 was used to summarize the data and Minitab version 17.0 software and web tool were used to calculate
Table 1. Group composition of the relocated Rhesus Macaques followed for studying feeding habitsin Asola-Bhatti Wildlife Sanctuary in Delhi
AM AF SAM SAF JUV INF Total
1 3 4 7 8 5 2 33
2 2 3 8 7 6 4 37
3 2 3 2 3 3 1 14
4 3 4 5 9 5 4 33
5 2 3 7 9 3 1 32
6 3 4 6 8 4 2 33
7 4 6 7 9 4 2 36
8 3 5 5 7 5 3 28
9 7 11 14 13 11 7 63
10 5 7 6 9 3 1 32
11 2 3 7 11 2 3 34
12 3 5 5 8 2 3 26
13 3 4 4 5 4 5 25
14 2 5 4 6 2 2 21
15 3 4 3 5 2 3 20
16 2 3 6 7 4 3 25 AM - adult male, AF - adult female, SAM - sub-adult male, SAF - sub-adult female, JUV - juvenile, INF - infant (N=492).
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12907–12915
Dietary preference and feeding patterns of the urban Rhesus Macaque Ganguly & Chauhan
12911
descriptive statistics. Landsat data imageries 2016 and ArcGIS software were used to map the study area using coordinates collected during the data sampling though Garmin GPS 72H.
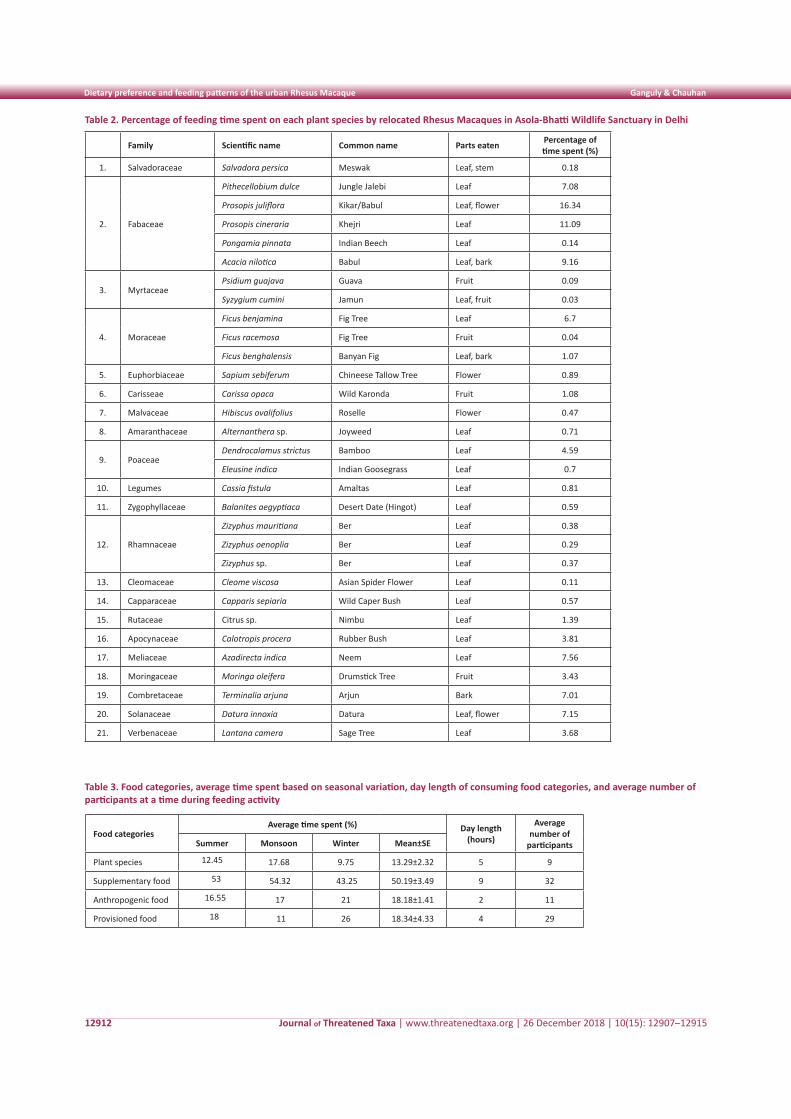
RESULTS
Food categoriesFood plants, plant families, parts eaten, and the
average percentage of time spent feeding on each plant species are given in Table 2. The macaques were mostly found to feed on Fabaceae (8.76±2.64), Moraceae (2.60±2.06), Rhamnaceae (0.34±0.02), and Mytraceae (0.06±0.03) families. Among the plant parts (nature food items) eaten, 34.65% of feeding time was spent on leaves, followed by 31% on bark and piths, 22.90% on flowers, and 11.01% on fruits. The macaques were mostly found in the lower canopy and bottom of trees in summer (39.13%), in the upper canopy in monsoon (31.26%), and in the middle to lower canopy in the winter (19%). The species was observed to spend 79% time on the ground and only 21% time on the trees.
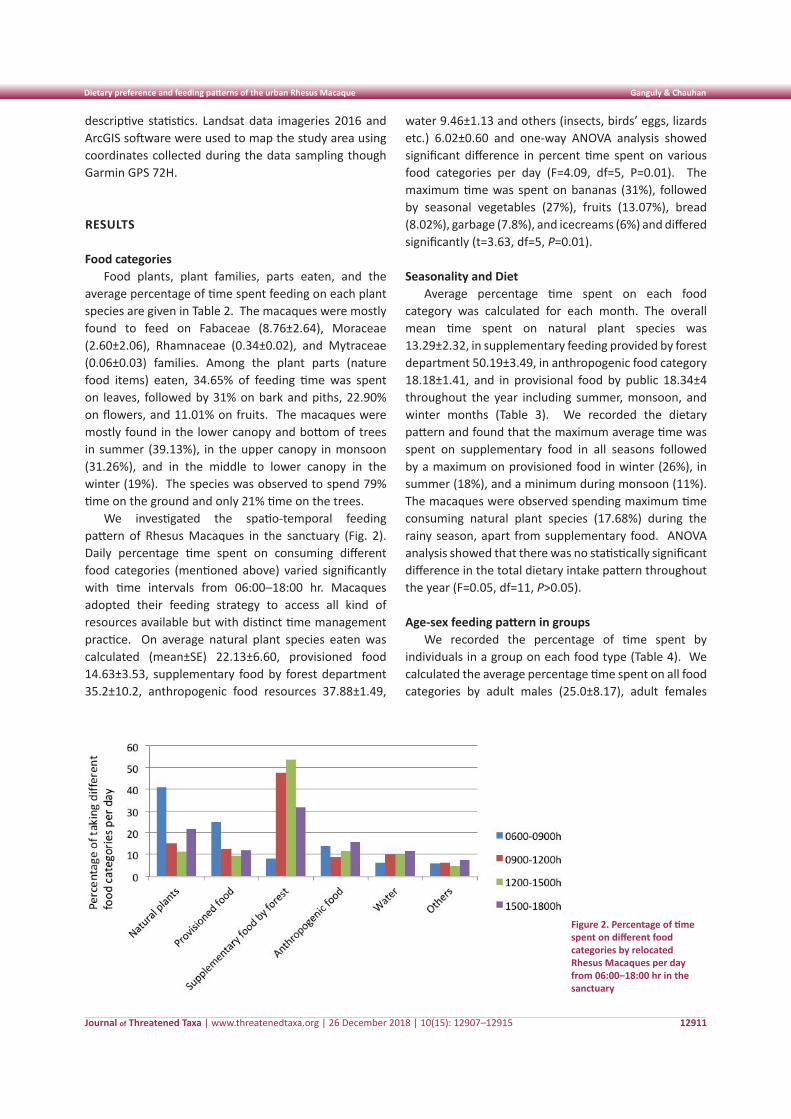
We investigated the spatio-temporal feeding pattern of Rhesus Macaques in the sanctuary (Fig. 2). Daily percentage time spent on consuming different food categories (mentioned above) varied significantly with time intervals from 06:00–18:00 hr. Macaques adopted their feeding strategy to access all kind of resources available but with distinct time management practice. On average natural plant species eaten was calculated (mean±SE) 22.13±6.60, provisioned food 14.63±3.53, supplementary food by forest department 35.2±10.2, anthropogenic food resources 37.88±1.49,
water 9.46±1.13 and others (insects, birds’ eggs, lizards etc.) 6.02±0.60 and one-way ANOVA analysis showed significant difference in percent time spent on various food categories per day (F=4.09, df=5, P=0.01). The maximum time was spent on bananas (31%), followed by seasonal vegetables (27%), fruits (13.07%), bread (8.02%), garbage (7.8%), and icecreams (6%) and differed significantly (t=3.63, df=5, P=0.01).
Seasonality and DietAverage percentage time spent on each food
category was calculated for each month. The overall mean time spent on natural plant species was 13.29±2.32, in supplementary feeding provided by forest department 50.19±3.49, in anthropogenic food category 18.18±1.41, and in provisional food by public 18.34±4 throughout the year including summer, monsoon, and winter months (Table 3). We recorded the dietary pattern and found that the maximum average time was spent on supplementary food in all seasons followed by a maximum on provisioned food in winter (26%), in summer (18%), and a minimum during monsoon (11%). The macaques were observed spending maximum time consuming natural plant species (17.68%) during the rainy season, apart from supplementary food. ANOVA analysis showed that there was no statistically significant difference in the total dietary intake pattern throughout the year (F=0.05, df=11, P>0.05).
Age-sex feeding pattern in groupsWe recorded the percentage of time spent by
individuals in a group on each food type (Table 4). We calculated the average percentage time spent on all food categories by adult males (25.0±8.17), adult females
Figure 2. Percentage of time spent on different food categories by relocated Rhesus Macaques per day from 06:00–18:00 hr in the sanctuary
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12907–12915
Dietary preference and feeding patterns of the urban Rhesus Macaque Ganguly & Chauhan
12912
Table 2. Percentage of feeding time spent on each plant species by relocated Rhesus Macaques in Asola-Bhatti Wildlife Sanctuary in Delhi
Family Scientific name Common name Parts eaten Percentage of time spent (%)
1. Salvadoraceae Salvadora persica Meswak Leaf, stem 0.18
2. Fabaceae
Pithecellobium dulce Jungle Jalebi Leaf 7.08
Prosopis juliflora Kikar/Babul Leaf, flower 16.34
Prosopis cineraria Khejri Leaf 11.09
Pongamia pinnata Indian Beech Leaf 0.14
Acacia nilotica Babul Leaf, bark 9.16
3. MyrtaceaePsidium guajava Guava Fruit 0.09
Syzygium cumini Jamun Leaf, fruit 0.03
4. Moraceae
Ficus benjamina Fig Tree Leaf 6.7
Ficus racemosa Fig Tree Fruit 0.04
Ficus benghalensis Banyan Fig Leaf, bark 1.07
5. Euphorbiaceae Sapium sebiferum Chineese Tallow Tree Flower 0.89
6. Carisseae Carissa opaca Wild Karonda Fruit 1.08
7. Malvaceae Hibiscus ovalifolius Roselle Flower 0.47
8. Amaranthaceae Alternanthera sp. Joyweed Leaf 0.71
9. PoaceaeDendrocalamus strictus Bamboo Leaf 4.59
Eleusine indica Indian Goosegrass Leaf 0.7
10. Legumes Cassia fistula Amaltas Leaf 0.81
11. Zygophyllaceae Balanites aegyptiaca Desert Date (Hingot) Leaf 0.59
12. Rhamnaceae
Zizyphus mauritiana Ber Leaf 0.38
Zizyphus oenoplia Ber Leaf 0.29
Zizyphus sp. Ber Leaf 0.37
13. Cleomaceae Cleome viscosa Asian Spider Flower Leaf 0.11
14. Capparaceae Capparis sepiaria Wild Caper Bush Leaf 0.57
15. Rutaceae Citrus sp. Nimbu Leaf 1.39
16. Apocynaceae Calotropis procera Rubber Bush Leaf 3.81
17. Meliaceae Azadirecta indica Neem Leaf 7.56
18. Moringaceae Moringa oleifera Drumstick Tree Fruit 3.43
19. Combretaceae Terminalia arjuna Arjun Bark 7.01
20. Solanaceae Datura innoxia Datura Leaf, flower 7.15
21. Verbenaceae Lantana camera Sage Tree Leaf 3.68
Table 3. Food categories, average time spent based on seasonal variation, day length of consuming food categories, and average number of participants at a time during feeding activity
Food categoriesAverage time spent (%) Day length
(hours)
Average number of
participantsSummer Monsoon Winter Mean±SE
Plant species 12.45 17.68 9.75 13.29±2.32 5 9
Supplementary food 53 54.32 43.25 50.19±3.49 9 32
Anthropogenic food 16.55 17 21 18.18±1.41 2 11
Provisioned food 18 11 26 18.34±4.33 4 29

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12907–12915
Dietary preference and feeding patterns of the urban Rhesus Macaque Ganguly & Chauhan
12913
(22.01±7.13), sub-adult males (7.92±4.02), sub adult females (6.06±3.19), juveniles (0.65±0.20,) and infants (0). Adult males dominated the pattern and used up the maximum amount of food provided to them and spent the maximum time on it. Adult females were much protective and did not allow their infants to feed on artificial foods. Infants compensated their nutritional requirement through lactation only.
DISCUSSION
Non-human primates compete with human beings for resource utilisation and space, which can lead to negative interactions (Priston & Underdown 2009), especially in urban areas (Lee & Priston 2005). In India, Rhesus Macaques often co-exist with human populations and are highly dependent on them for food (Southwick et al. 1976). The high feeding dependency on anthropogenic food resources is, however, not correlated with natural resource scarcity. While natural resources such as fruits are highly variable over the year, anthropogenic food resources are potentially more stable and easily available. A study on Long-tailed Macaques Macaca fascicularis showed that the main drivers for exploitation of anthropogenic foods were natural food plant resource scarcity or an overt dependence on anthropogenic foods (Sha & Hanya 2013). Utilization of anthropogenic food resources lowered preferences of macaques on fruits and natural plants in another study (Hambali et al. 2014). The consequences of the dependency of macaques on human food resources can include food stealing, which may lead to negative interactions with humans. In our study, the relocated Rhesus Macaques were more inclined towards anthropogenic, supplementary, and provisioned food resources than natural plant resources in the forest. As this sanctuary is situated in a human-dominated landscape and human settlements are located close by (less than 50m away), Rhesus Macaques disperse from the sanctuary and consume food from nearby households, markets, and temple areas. The forest department of Delhi Government was also assigned to provide supplementary food to the rehabilitated macaques for the maintenance of a viable population in the newly introduced environment.
Our results showed that the macaques fed on natural plant species in the early morning between 06:00– 09:00 hr (40.8%), after which their tendency to consume natural resources declined before rising in the late afternoon (21.75%). Food provisioning by the public was recorded mostly in the early morning (25%) and continued
throughout the day in the fringes of the sanctuary. Between 09:00hr and 11:00hr, macaques gathered at feeding stations within the sanctuary near the Bhatti Range Office, reaching a peak number between 12:00hr and 15:00hr (53.75%). The macaques were reportedly given 2,500kg food per day by the forest department and this feeding pattern had a large influence on their daily activity and movement. During supplementary feeding, the macaques did not consume natural plants within the forest area. The relocated Rhesus Macaques were highly inclined towards human food outside the sanctuary and often entered nearby houses or snatched bread and vegetables from open markets in the nearby Sanjay Colony (Bhatti Mines).
Our results showed that the macaques mostly preferred leaves and stems of Prosopis juliflora (16.34%) and P. cineraria (11.09%), which were reported to be beneficial for their health. The heartwood of these two plant species contains ample antioxidants such as flavonol and mesquitol (Sirmah et al. 2009). Though numerous species of medicinal plants and fruiting trees are available in the sanctuary, the macaques did not spend much time in natural foraging but mostly depended on artificial feeding. Our results showed a high consumption of supplementary food items throughout the year with no seasonal differences and low average time spent on natural food plants. The macaques showed dependency on anthropogenic and provisioned food over natural fruit. The former included bananas, seasonal fruits and vegetables, bread, chickpeas, fried snacks, and ice-creams; the macaques were even reported to steal cold water from refrigerators of houses in nearby localities at a 0.25–5 km distance (USA Today 2017).
Roadside food provisioning is a common practice across cities and villages in India. Southwick et al. (1976) documented the impact of artificial feeding on the ecology and behaviour of macaques. Our study provides information on feeding practices of Rhesus Macaques after translocation to an area containing various types of natural and anthropogenic food resources in a human-dominated landscape. An understanding of the basic natural history of primates is essential for their conservation (Caro 2007; Fashing 2007). The primary threat primates face today is habitat destruction (Wieczkowski 2004; Chapman et al. 2006). By reducing forest size and quality, habitat destruction leads to the reduction of food sources for forest-dwelling primates and, in some cases, threatens them with local extinction (Lee &Hauser 1998; Muoria et al. 2003). The increasing population of Rhesus Macaques living in proximity to human habitations has become a major issue in

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12907–12915
Dietary preference and feeding patterns of the urban Rhesus Macaque Ganguly & Chauhan
12914
India. Rapid urbanisation, deforestation, and habitat fragmentation altered the natural living spaces of animals and their natural behaviour in the wild. Most primate species were severely affected by threats in anthropogenic landscapes (Sinha & Vijayakrishnan 2017). The translocation of Rhesus Macaques from city areas to forest situated at close proximity with human settlementsmight not reduce the conservation threats for the taxa. The step, however, altered their feeding strategies as the species was observed to become more dependent on supplementary and anthropogenic food resources than on natural foraging. Though artificial feeding of fruits and vegetables might increase overall nutrition, their natural frugivorous behaviour seem to be lost. The macaques were seen to snatch and steal even those anthropogenic food resources that had no health benefits, as they were used to such behaviour in human habitats (Ganguly et al. 2018). The dietary habits of Rhesus Macaques were totally different in a human-dominated forest land. Previous studies showed that the species thrived in eight diverse habitats (temple, urban, village, village-cum-pond, pond, roadside, canal sides, and forest) having varying degrees of human interactions in India (Seth et al. 1986). The feeding practice seemed to increase the urban threats, diseases, and anthropogenic stress in the Rhesus Macaque population. In our study, the species was observed to spend maximum time on the ground instead of on the trees and their dependency on supplementary, provisioned, and anthropogenic food sources did not indicate conservation success. Understanding the feeding ecology in this sanctuary would help in planning the management of macaques in other urban areas.
REFERENCES
Aguilar-Cucurachi, M.A., P.A.D. Dias, A. Rangel-Negrín, R. Chavira, L. Boeck & D. Canales-Espinosa (2010). Preliminary evidence of accumulation of stress during translocation in mantled howlers. American Journal of Primatology 72(9): 805–10; https://doi.org/10.1002/ajp.20841
Air, A. (2015). Crop Raiding and Conflict: Study of Rhesus Macaque-human Conflict in Shivapuri-Nagarjun National Park, Kathmandu Nepal. Natural Resources Management, Norwegian University of Science and Technology, 32pp.
Altmann, J. (1974). Observational study of behavior: sampling methods. Behavior 49(3/4): 227–267.
Begum, S. & M.M. Feeroz (2013). Distribution of Rhesus Macaques (Macaca mulatta) in Bangladesh: Interpopulation Variation in Group Size and Composition. Primate Conservation 26: 125–132.
Burkart, A. (1976). A monograph of the genus Prosopis (Leguminosae: Mimosoideae) (Part 1 and 2). Catalogue of the recognized species of Prosopis. Journal of the Arnold Arboretum 57: 219–249, 450–525.
Caro, T. (2007). Behavior and conservation: a bridge too far? Trends in Ecology & Evolution 22(8): 394–400.
Chapman, C.A., M.J. Lawes & H.A.C. Eeley (2006). What hope for African primate diversity? African Journal of Ecology 44: 116–133.
Chapman, C.A., R.W. Wrangham & L.J. Chapman (1995).Ecological constraints on group-size: an analysis of spider monkey and chimpanzee subgroups. Behavioral Ecology and Sociobiology 36(1): 59–70.
Cowlishaw, G. & R. I. Dunbar (2000). Primate Conservation Biology. University of Chicago Press, Chicago, 498pp; https://doi.org/10.1017/S1367943001221337
Fashing, P.J. (2007). Behavior, ecology and conservation of colobine monkeys: an introduction. International Journal of Primatology 28: 507–511.
Fleagle, J.G. & C.C. Gilbert (2006). The biogeography of primate evolution: the role of plate tectonics, climate and chance, pp. 375–418. In: Lehman, S.M. & J.G. Fleagle (eds.). Primate Biogeography. Springer, New York.
Fooden, J. (2000). Systematic Review of the Rhesus Macaque, Macaca mulatta (Zimmermann 1780).Field Museum of Natural History, Chicago, USA, 180pp.
Ganguly, I., N.P.S. Chauhan & P. Verma (2018).Assessment of human-macaque conflict and possible mitigation strategies in and around Asola-Bhatti Wildlife Sanctuary. Environment & Ecology 36(3): 823–827.
Goldstein, S.J. & A.F. Richard (1989). Ecology of Rhesus Macaques (Macaca mulatta) in northwest Pakistan. International Journal of Primatology 10(6): 531–567.
Gupta, A.K. & A. Kumar (1994). Feeding ecology and conservation of the Phayre’s Leaf Monkey Presbytis phayrei in northeast India. Biological Conservation 69(3): 301–306
Gupta, A.K. (2001). Status of primates in Tripura. Envis Bulletin 1(1): 127–135.
Hambali, K., A. Ismail, B.M. Md-Zain, A. Amir & F.A. Karim (2014). Diet of Long Tailed Macaques (Macaca fascicularis) at the Entrance of Kuala Selangor Nature Park (Anthropogenic Habitat): Food Selection that Leads to Human-Macaque Conflict. Acta Biologica Malaysiana 3(2): 58–68.
Hanya, G., N. Noma & N. Agetsuma (2003). Altitudinal and seasonal variations in the diet of Japanese Macaques in Yakushima. Primates 44: 51–59.
Harcourt, A.H., S.A. Coppeto & S.A. Parks (2002). Rarity, specialization and extinction in primates. Journal of Biogeography 29(4): 445–456; https://doi.org/10.1046/j.1365-2699.2002.00685.x
Hasan, K., M.A. Aziz, S.M.R. Alam, Y. Kawamoto, Y. Lisa, Jones-Engel, R.C. Kyes, S. Akhtar & C.M. Hill (2005). People, crops, and primates: a conflict of interests, pp. 41–59. In: Paterson, J.D.& J. Wallis (eds.). Commensalism and Conflict: The Human–Primate Interface. American Society of Primatologists, Norman.
IUCN (2018). The IUCN Red List of Threatened Species, Version 2018-1. http://www.iucnredlist.org. Downloaded on 05 July 2018.
Janson, C.H. & C.A. Chapman (1999). Resources and primate community structure, pp. 237–268. In: Fleagle, J.G., C. Janson & K.E. Reed (eds.). Primate Communities. Cambridge University Press, Cambridge.
Kemf, E. & A. Wilson (1997). Great Apes in the Wild: Species Status Report. World Wide Fund for Nature, Gland, 36pp.
Khanna, V. & J.P. Sati (2003). Some Selected Fauna of Asola-Bhatti Wildlife Sanctuary (Delhi), pp. 1–70. In: Fauna of Conservation Area Vol 16. Zoological Survey of India, Kolkata.
Kushwaha, S.P.S., S. Nandy & M. Gupta (2014). Growing stock & woody biomass assessment in Asola-Bhatti Wildlife Sanctuary, Delhi, India. Environment Monitoring Assessment 186(9): 5911–5920; https://doi.org/10.1007/s10661-014-3828-0
Lee P.C. & N.E.C. Priston (2005). Human attitudes to primates: Perceptions of pests, conflict and consequences for primate conservation, pp. 1–23. In: Paterson JD, Wallis J, editors. Commensalism and conflict: the human-primate interface. Norman, Oklahama, American Society of Primatology; http://doi.org/10.1002/047001539X.ch1
Lee, P.C. & M.D. Hauser (1998). Long-term consequence of changes

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12907–12915
Dietary preference and feeding patterns of the urban Rhesus Macaque Ganguly & Chauhan
12915
in territory quality on feeding and reproductive strategies of Vervet Monkeys. Journal of Animal Ecology 67: 347–358.
Lee, P.C., E.J. Brennan, J.G. Else & J. Altmann (1986). Ecology and behavior of vervet monkeys in a tourist lodge habitat, pp. 229–235. In: Else, J.G. & P.C. Lee (eds.). Primate Ecology and Conservation. Cambridge, Cambridge University Press.
Lindburg, D.G. (1971). The Rhesus Monkey in north Indian ecological and behavioral study, pp. 1–106. In: Rosenblum, L.A. (ed.). Primate Behavior 2. New York, Academic Press.
Majumder, J., R. Lodh & B.K. Agarwala (2012). Fish feeding adaptation by Rhesus Macaque Macaca mulatta (Cercopithecidae) in the Sundarban mangrove swamps, India. Journal of Threatened Taxa 4(4): 2539–2540; https://doi.org/10.11609/JoTT.o2884.2539-40
Malik, I. & R.L. Johnson (1991). Trapping and conservation: development of a translocation in India, pp. 63–64. In: Ehara, A., T. Kimura & M. Iwamoto (eds.). Primatology Today. Elsevier, Amsterdam.
Malik, I. & R.L. Johnson (1994). Commensal Rhesus in India: the need and cost of translocation. Revue d’Ecologie (La Terre et la Vie) 49: 233–243.
Malik, I. (1983). A study of selected behavioural traits of Rhesus Monkeys (Macaca mulatta) in free-ranging environments. PhD Thesis. University of Meerut.
Mandal, A.K. (1964). The behaviour of the Rhesus Monkeys (Macaca mulatta Zimmermann) in the Sundarbans. Journal of the Bengal Natural History Society 33: 153–165.
Mangalam, M. & M. Singh (2013). Flexibility in food extraction techniques in urban free-ranging Bonnet Macaques, Macaca radiata. PLoS ONE 12: e85497; https://doi.org/10.1371/ journal.pone.0085497
Mittermeier, R. & W. Konstant (1996/1997). Primate conservation: a retrospective and a look into the 21stCentury. Primate Conservation 17: 7–17.
Molur, S., D. Brandon-Jones, W. Dittus, A. Eudey, A. Kumar, M. Singh, M.M. Feeroz, M. Chalise, P. Priya & S. Walker (2003). Status of South Asian Primates: Conservation Assessment and Management Plan (C.A.M.P.) Workshop Report, 2003. Zoo Outreach Organisation / CBSG-South Asia, Coimbatore, India, viii+432pp
Muoria, P.K., G.M. Karere, N.N. Moinde & M.A. Suleman (2003). Primate census and habitat evaluation in the Tana delta region, Kenya.African Journal of Ecology 41(2): 157–163.
Naithani, H.B., S.S. Negi, M. Pal, S. Chandra & V.P. Khanduri (2006). Vegetation Survey and Inventorisation of Species in the Ridge Forest of Delhi. Department of Forest and Wildlife, Government of Delhi, Delhi.
Pasiecznik, N.M., P. Felker, P.J.C. Harris, L.N. Harsh, G. Cruz, J.C. Tewari, K. Cadoret & L.J. Maldonado (2001). The Prosopis juliflora-Prosopis pallida Complex. Coventry, HDRA, UK, 172pp.
Peterson, D. (2003). Eating Apes. University of California Press, Berkeley, 320pp.
Priston, N.E.C. & S.J. Underdown (2009). A simple method for calculating the likelihood of crop damage by primates: an epidemiological approach. International Journal of Pest Management 55(1): 51–56; https://doi.org/10.1080/09670870802450268
Seth, P.K. & N.K. Pokwal (2001).Behaviour and ecology of primates. Journal of the Bombay Natural History Society 14(3): 172–180.
Seth, P.K., P.K. Chopra & S. Seth (2001). Indian Rhesus Macaque: habitat, ecology and activity patterns of naturally occurring populations, pp. 68–80. In: Gupta, A.K. (ed.). Non-human Primates of India, Vol 1(1). ENVIS Bulletin: Wildlife & Protected Areas. Wildlife Institute of India, Dehradun (India).
Seth, P.K. & S. Seth (1986). Ecology and behavior of rhesus monkeys in India, pp. 89–103. In: lse, J.G. & P.C. Lee (eds.). Primate Ecology And Conservation. Cambridge, Cambridge University Press.
Sha, J.C.M. & G. Hanya (2013). Temporal food resource correlates to the behavior and ecology of food-enhanced Long-Tailed Macaques (Macaca fascicularis). Mammal Study 38(3): 163–175; https://doi.org/10.3106/041.038.0305
Simpson, G.G., A. Roe & R.C. Lewontin (1960). Quantitative Zoology. Harcourt, Brace & Company, New York. American Journal of Science 258(2): 300–311.
Sinha, A. & S. Vijayakrishnan (2017). Primates in Urban Settings, pp. 1–8. In: Fuentes, A. (ed.). The International Encyclopedia of Primatology. John Wiley & Sons, Inc. Published 2017 by John Wiley & Sons, Inc.; https://doi.org/10.1002/9781119179313.wbprim0458
Sirmah, P., S. Dumarcay, E. Masson & P. Gerardin (2009). Unusual amount of (-) mesquitol from the heartwood of Propopis juliflora. Natural Product Research 23(2): 183–189.
Southwick, C.H., I. Malik & M.F. Siddiqi (1998). Translocations of Rhesus Monkeys in India: prospects and outcomes. American Journal of Primatology 45(2): 209–210.
Southwick, C.H., M.F. Siddiqi, M.Y. Farooqui & B.C. Pal (1976). The effects of artificial feeding on aggressive behaviour of Rhesus Monkeys in India. Animal Behaviour 24(1): 11–15.
Strum, S.C. (1994). Prospects for management of primate pests. Revue d’Ecologie (La Terre et la Vie) 49: 295–306.
Strum, S.C. (2001). Almost Human. University of Chicago Press, Chicago, 308pp.
USA Today (2017). Why India is going bananas over birth control for monkeys. https://www.usatoday.com/story/news/world/2017/05/11/monkeys-india
Wieczkowski, J. (2004). Ecological correlates of abundance in the Tana Mangabey (Cercocebus galeritus). American Journal of Primatology 63: 125–138.
Wilson, E.O. (2007). Rhesus Monkey (Macaca mulatta) feeding behaviour and diet of Rhesus Monkey in Shivalik forest in north India. IUCN Red List of Threatened Species 1(8): 1–11.
Threatened Taxa

Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12916
Arti
cle
DOI: https://doi.org/10.11609/jott.4125.10.15.12916-12932
Editor: Ruth H. Carmichael, University of South Alabama, Dauphin Island, USA. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 4125 | Received 14 March 2018 | Final received 03 August 2018 | Finally accepted 02 December 2018
Citation: Kaiser, D. & S. Schoppe (2018). Postembryonic development of the Tri-spine Horseshoe Crab Tachypleus tridentatus (Merostomata: Xiphosura) in a nursery habitat in the Philippines. Journal of Threatened Taxa 10(15): 12916–12932; https://doi.org/10.11609/jott.4125.10.15.12916-12932
Copyright: © Kaiser & Schoppe 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduc-tion and distribution by providing adequate credit to the authors and the source of publication.
Funding: Mainly self-funded. The ‘Studienstiftung des Deutschen Volkes’ provided a scholarship for the first author in 2001.
Competing interests: The authors declare no competing interests.
For Author Details and Author Contribution see end of this article.
Acknowledgements: We are especially grateful for the cooperation of the Western Philippine University in Puerto Princesa City on Palawan, Philippines. We extend our special thanks to Prof. Günter Arlt from the University of Rostock, Germany. We further thank Sharah Barredo for her contribution to the collection of juveniles in 2017. The authors are much obliged to the two anonymous reviewers for their constructive comments that considerably improved the manuscript.
Postembryonic development of the Tri-spine Horseshoe Crab Tachypleus tridentatus (Merostomata: Xiphosura)
in a nursery habitat in the Philippines
Dorkas Kaiser 1 & Sabine Schoppe 2
1 Institute of Biosciences, University of Rostock, Rostock 18051, Germany 2 Western Philippines University, Puerto Princesa City, Aborlan, Palawan 5302, Philippines
1,2, Katala Foundation Inc., Puerto Princesa City, Santa Monica, 5300 Palawan, Philippines 1 [email protected] (corresponding author), 2 [email protected]
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Abstract Populations of the Tri-spine Horseshoe Crab Tachypleus tridentatus have dramatically decreased over their distribution range and conservation efforts are now crucial. The implementation of appropriate management strategies and stock assessment rely on accurate growth-rate estimates. The postembryonic development of the species in the tropics, however, is not elucidated. To provide the information needed to assess the demographics of juvenile populations and to judge the status of T. tridentatus in the Philippines, we conducted a mark-recapture experiment in a nursery habitat on Palawan Island. The results obtained during the 10-month period provide the first consecutive data on the stepwise growth of the species in the Philippines and the first near comprehensive dataset collected within a single population of juveniles in the tropics. By analyzing size-frequency (prosomal width) distributions of 853 individuals and by using 94 juveniles that molted during the study, 13 molt stages were differentiated. Based on the intermolt periods of six instars, we estimated the growth curve of T. tridentatus following two models (non-linear and power function). The data support the assumption that growth continues year-round in the tropics and also indicate that the average age of mature male and female T. tridentatus in the Philippines ranges from three to four years. The agreement with a field study in Japan suggests that 14 postembryonic stages may be characteristic for the development of natural populations throughout the range of the species. Though more data are needed to validate these results, the study provides a sound baseline for future studies in the tropics.
Keywords: Tri-spine Horseshoe Crab, juveniles, development stages, intertidal zone, morphometry, allometry, size-age relationship, growth curve.
Abbreviations: AA - Distance between the anal angles; BL - Body length (PL+OL+TL); BM beach - Bernardo Marcelo Beach (study site); CL - Carapace length (PL+OL); DS - Development stage; Eyes - Distance between the compound eyes; IMP - Intermolt period; OL - Opisthosomal length; OW1–3 - Opisthosomal width 1–3 ; PES - Postembryonic stage; PL - Prosomal length; PW - Prosomal width; SPSS - Statistical Package for the Social Sciences; TL - Telson length; Statistics: CI - 95% confidence interval; df - Degrees of freedom ; M - Mean value; Min / Max - Minimum and maximum values; n - Sample number ; p - Significance level; r - Pearson’s correlation coefficient; SD - Standard deviation; SE - Standard error of the mean; T - Statistics of the one-sample t test; U - Statistics of the Mann-Whitney U test; W - Statistics of the Shapiro-Wilk Test ; Z - Statistics of the Kolmogorov-Smirnov test.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12917
INTRODUCTION
The Tri-spine Horseshoe Crab Tachypleus tridentatus (Leach, 1819) (Xiphosura: Chelicerata) is the largest of four extant species of ancient marine arthropods, the origin of which can be traced back to 445 million years (Shuster et al. 2003; Rudkin et al. 2008). The information about horseshoe crabs in the Philippines is especially scarce. Waterman (1958) recorded T. tridentatus and Carcinoscorpius rotundicauda (Latreille, 1802) for western and southern Philippines, respectively; based on a picture, both species were listed as occurring in the Province of Palawan (Sekiguchi 1988). The first survey conducted from northern to central Palawan confirmed the presence of T. tridentatus in the area (Schoppe 2002).
Recent harvest pressures and habitat loss prompted the need for management actions to protect horseshoe crabs (Berkson et al. 2009). Although still listed in the Data Deficient category by the IUCN (2017), observations in Taiwan, Japan, Hong Kong, Singapore, Malaysia, Borneo, and Thailand indicate that populations of T. tridentatus have dramatically decreased (Itow 1993; Hsieh & Chen 2009, 2015; Chen et al. 2015; Kwan et al. 2016; Lee & Morton 2016; Wada et al. 2016; Manca et al. 2017). Unlike other regions in Asia, in the Philippines, there is no commercial exploitation of T. tridentatus for the production of amebocyte lysate or food, but habitat loss due to land reclamation, sea sand mining, and coastal development are destroying its natural breeding beaches and nursery habitats. As a result, T. tridentatus faces a high risk of extinction without management efforts to conserve its viable populations and habitats (Schoppe 2002).
The implementation of effective management strategies and stock assessment relies on accurate estimations of growth rates (size at age, size at maturity, and age at first capture), size distributions, and population size (Froese et al. 2008; Chang et al. 2012; Cunningham & Darnell 2015; Cao et al. 2016). A lot of effort was made during the last few decades to understand the growth biology of horseshoe crabs, including laboratory studies on the influence of environment factors such as water temperature and salinity (Jegla & Costlow 1982; Chen et al. 2010; Zaleha et al. 2011), pH (Tanacredi & Portilla 2015), sediment type (Hong et al. 2009; Hieb et al. 2015), tank size (Hieb et al. 2015; Chen et al. 2016), and food quantity and quality (Carmichael et al. 2009; Schreibman & Zarnoch 2009; Hu et al. 2013). Due to the lack of calcified structures persisting through the molt, however, the age of the different instars was not
unambiguously determined for any horseshoe crab species (Carmichael et al. 2003; Chen et al. 2010).
Specifications for T. tridentatus vary strongly in the literature, although there is a consensus that the females molt once more than the males to reach maturity. Based on laboratory studies, Sekiguchi et al. (1988) calculated that the females spent 14 years to molt 16 times before attaining maturity, while Chen et al. (2010) estimated that the females mature in stage 15 after four years when reared in warm water. Goto & Hattori (1929) identified 14 postembryonic stages by measuring individuals in their natural habitats in Japan. Kawahara (1984), on the other hand, suggested that the females in Japan molt 15 times in 10 years to reach maturity, while Asano (1942, cited in Lee & Morton 2005) estimated that they molt 18 times in 16 years. To our knowledge, the postembryonic development of T. tridentatus in their tropical environment is not yet determined, but laboratory studies indicate that the growth of juveniles could continue throughout the year when the temperature is greater than 28°C (Lee & Morton 2005; Chen et al. 2010). Most studies concerning the life history of T. tridentatus are from Japan, China, or Hong Kong — countries where ecdysis and spawning appear to stop during colder seasons (for instance, Sekiguchi et al. 1988; Chiu & Morton 2004; Zhou & Morton 2004; Lee & Morton 2005; Hu et al. 2009, 2015; Kwan 2015; Kwan et al. 2015) — while fewer studies were conducted in the tropics (Robert et al. 2014; Mohamad et al. 2016; Manca et al. 2017; Mashar et al. 2017).
In this study, we characterized the postembryonic development of T. tridentatus in a nursery habitat on Palawan to provide sound baseline data needed for conservation, particularly in southeastern Asia (Berkson et al. 2009; Shuster & Sekiguchi 2009). In 2001, the population in the study site was estimated to comprise of 298 individuals with a male-to-female ratio of 1.2:1 (Kaiser 2002). The main objectives of this study were 1) to identify the number of instars until maturity, 2) to describe the post-embryonic growth patterns, and 3) to estimate the size-age relationship of T. tridentatus in the Philippines. These data represent the first dataset of this type for the tropics.
MATERIALS AND METHODS
Study site The growth of juvenile T. tridentatus was studied in
a nursery habitat located close to the Puerto Princesa City on the eastern coast of Palawan in the Philippines

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12918
(9.7620N & 118.7720E) (Fig. 1). The study region is characterized by a rainy season that lasts from June to November and a dry season from December to May (PAGASA 2017). The annual rainfall averages at 1,684mm, while the mean annual temperature is 27.2°C. The area has a mixed semidiurnal tidal cycle (National Ocean Service 2008); the tidal range during spring tide may rise up to 1.9m. The mean water temperature in the study area is 29°C and 31°C during high and low tides, respectively (Table 1). The average pH ranges between 7.6 and 8 and the mean salinity between 30 and 31‰ (Table 1). The study site reported upon herein is the Bernardo Marcelo Beach (hereinafter BM beach) (Image 1), a 130m-wide and 600m-long seagrass meadow. About 65% of the sampling area (50,700m²) is covered with seagrass, while the rest comprises sandy patches. The mean grain size is 0.103±0.02 mm (n = 33). It comprises a zone between the mean low and mean high water levels and the upper sub-tidal range. The local residents use the northern part of the beach for recreation in the weekends. Shellfish are gathered and fishermen position bottom-set gillnets seaward of the sampling area in a seagrass meadow without larger sand patches and an adjacent riff (about 600m wide).
Sampling strategy (capture-mark-recapture)The juvenile population of T. tridentatus at BM beach
was assessed during daytime at low tide from May to December 2001 for 84 days and between April and May 2017 for 23 days. Following Rudloe (1983), the assessment started about two to three hours before the lowest water level was reached and lasted for four to five hours. By walking barefoot in a zigzag course, the
Figure 1. The geographic location of the study site. a - Puerto Princesa City on the eastern coast of Palawan in the Philippines;b - Bernardo Marcelo Beach
Image 1. The study site BM beach at low tide
© Dorkas Kaiser

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12919
surface of the entire sampling area was systematically searched for juveniles and exuviae. The majority of juveniles were found with the help of their feeding trails and mostly in the sandy-muddy substrate; some larger individuals were sensed by foot. Additionally, several adult horseshoe crabs were handed over by fishermen and found at the market; they were measured as well.
Morphometric parameters (prosomal length, opisthosomal length, telson length, prosomal width, opisthosomal widths 1–3, the distance between the anal angles, the distance between the compound eyes) were measured to the nearest 0.1mm using a Vernier caliper (Fig. 2). Sex was identified based on the size and shape of the genital papillae (Bonaventura et al. 1982). Superglue was used to fix a number on the prosoma of each individual. For identification after ecdysis, the lateral mobile spines of the opisthosoma were shortened
(Kawahara 1982) following a coding system (Appendix 1). The identification mark of the adults was modified following Sokoloff (1978). All juveniles were released where they were found. The individuals found during or shortly after ecdysis were kept under an ambient temperature of 25–32 °C in a tank with air supply and the water of BM beach; they were measured and marked when the carapace had sufficiently hardened. A few individuals supplied by fishermen or shellfish gatherers were marked and released at BM beach.
Data analysesUnless otherwise stated, the significance threshold
was set to α = 0.05 and the significance was tested at a two-tailed level. Statistical Package for the Social Sciences (SPSS), version 15.0 (SPSS, Inc., Chicago) and Microsoft Excel 2010 were used to carry out the
Figure 2. Measurements taken. a - prosomal length (PL), opisthosomal length (OL), prosomal width (PW), opisthosomal width (OW1–3), and the distance between the anal angles (AA) and between the compound eyes (Eyes); b - telson length (TL). The carapace length (CL) equals PL+OL; the body length (BL) equals PL+OL+TL
Table 1. Mean values (M) ± standard deviation (SD) of the hydrographic conditions prevailing in the nursery habitat of Bernardo Marcelo Beach on Palawan, with sample size (n) and minimum and maximum values (Range)
Tide Temperature (oC) pH Salinity (‰) Dissolved oxygen (%)
High Mean ± SD 29.2 ± 1.2 7.6 ± 0.05 31.0 ± 0.82 71.5 ± 34.3
Range 26.5–32.0 7.5–7.7 29.0–32.4 25.8–111.9
n 50 17 59 35
Low Mean ± SD 31.1 ± 0.9 8.0 ± 0.15 30.0 ± 1.05 94.2 ± 45.3
Range 29–32.5 1 7.8–8.3 27.4–32.0 48.3–228.0
n 54 10 67 46
1 On hot days, the water temperature during low tide was up to 41°C.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12920
statistical tests.
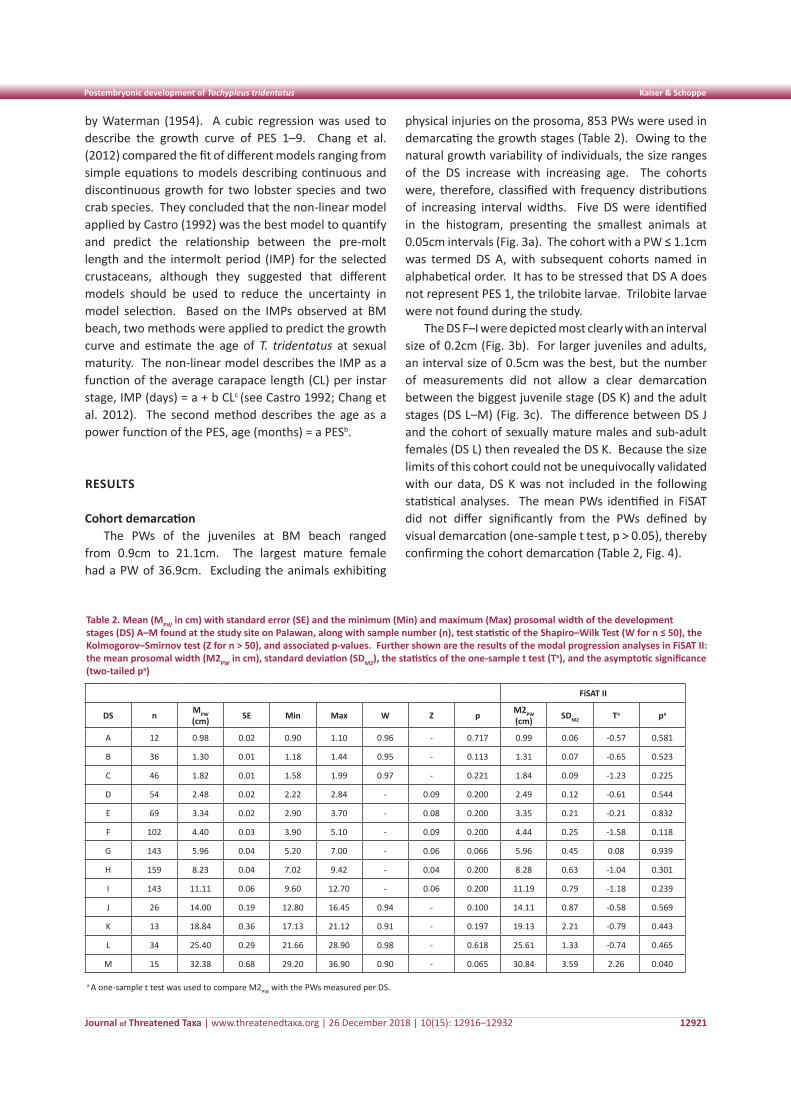
Cohort demarcationThe prosomal width (PW) was used to mark the
instar stages that characterize the postembryonic development of T. tridentatus on Palawan. The PW of all individuals and exuviae were grouped into specified size intervals and size-frequency histograms were plotted. The resulting histograms were multimodal and indicated different cohorts or development stages (DS) that differed in abundance. The visual cohort demarcations were adjusted by comparing the data with 94 individuals that molted during the study period or whose individual exuviae was measured (hereinafter referred to as molting individuals). The measured adults allowed the defining of the size limits for the cohort of sexually mature males and sub-adult females, as well as the stage of sexually mature females.
The resulting size ranges of the DS were ascertained by conducting a modal progression analysis of the size-frequency distributions with the Fish Stock Assessment Tool, FiSAT II (FAO 2006–2017, http://www.fao.org/). FiSAT II applies the maximum likelihood concept to separate the normally distributed components of the data, thus allowing the demarcation of the cohorts from the multimodal distributions. Thereafter, a one-sample t test was used to compare the means calculated in FiSAT with the DS defined by the visual demarcation and the molting individuals.
Body measuresThe Shapiro-Wilk test (n ≤ 50) and the Kolmogorov-
Smirnov test (with Lilliefors correction) (n > 50) were used to examine whether the established size ranges and the body parameters were normally distributed. The homogeneity of variances was tested using the Levene’s test (Field 2009). Because some data showed a non-normal distribution when differentiating the juveniles’ sex, the non-parametric Kruskal-Wallis test was used to compare the body measures of females and males. Subsequently, pair-wise comparisons for each DS were performed using the Mann-Whitney procedure. A Bonferroni correction for multiple comparisons was applied to the significance level (differences were regarded as significant at α < 0.006). The Mann-Whitney procedure was further used to compare various morphometric ratios between juvenile and adult T. tridentatus (males, females, and individuals with unknown sex were pooled). A one-sample t test was used to compare the carapace lengths per DS with the mean values reported by Goto & Hattori (1929).
Hiatt growth modelAccording to Hiatt (1948), a linear growth model can
be used to fit the post- and pre-molt size of crustaceans and this model was applied to assess horseshoe crab growth (Carmichael et al. 2003; Hu et al. 2013). The model describes the growth under natural conditions as PWn+1 = a + b PWn, where PWn is the pre-molt PW at instar n and PWn+1 is the post-molt PW at instar n+1.
The y-intercept indicates whether the size increment increases (a < 0) or decreases (a > 0) with an increase in the size of the animals or whether it stays constant during the development (a = 0). Slope b represents the growth coefficient, allowing conclusions on the variations between the size increments of the consecutive molts. The Hiatt growth model was applied first to illustrate the size increment observed for the 94 molting individuals. Additionally, the model was applied based on the data of the cohort demarcation by using the average PW of the instars. A Student’s t test was used to compare the slopes of the regression lines in the two Hiatt diagrams to allow conclusions to be drawn regarding the demarcation of the DS identified at BM beach.
Allometric growthThe allometric growth of each morphometric
parameter (y) was expressed as a power function of the PW (x) with the equation y = a xb (Fuiman 1983). The relative growth coefficient was estimated by linear regression of the log-transformed allometric growth curve of the type log y = log a + b log x, where a is the intercept and b the growth coefficient (Gould 1966). The growth coefficient identifies whether the growth pattern indicates isometric growth (b = 1 for the lengths or widths and b = 3 for weight), positive allometry (b > 1 for lengths or widths and b > 3 for weight), or negative allometry (b < 1 for lengths or widths and b < 3 for weight). If the value 1.0 (or 3.0 in the case of the weight) is outside the 95% confidence interval of b, the difference is regarded as significant. A Student’s t test was used to compare the slopes of the regression lines to assess whether males and females differed significantly. To illustrate a specific characteristic of the growth of the horseshoe crabs’ telson, a linear regression for the relation between the PW and the telson length (TL) was again carried out graphically.
Growth curveThe first postembryonic stage (PES), the trilobite
larvae, was not found during the study. The PW of the trilobite larvae was estimated following the Dyar’s rule (Ln PW = a PES – b), which was already applied
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12921
by Waterman (1954). A cubic regression was used to describe the growth curve of PES 1–9. Chang et al. (2012) compared the fit of different models ranging from simple equations to models describing continuous and discontinuous growth for two lobster species and two crab species. They concluded that the non-linear model applied by Castro (1992) was the best model to quantify and predict the relationship between the pre-molt length and the intermolt period (IMP) for the selected crustaceans, although they suggested that different models should be used to reduce the uncertainty in model selection. Based on the IMPs observed at BM beach, two methods were applied to predict the growth curve and estimate the age of T. tridentatus at sexual maturity. The non-linear model describes the IMP as a function of the average carapace length (CL) per instar stage, IMP (days) = a + b CLc (see Castro 1992; Chang et al. 2012). The second method describes the age as a power function of the PES, age (months) = a PESb.
RESULTS
Cohort demarcationThe PWs of the juveniles at BM beach ranged
from 0.9cm to 21.1cm. The largest mature female had a PW of 36.9cm. Excluding the animals exhibiting
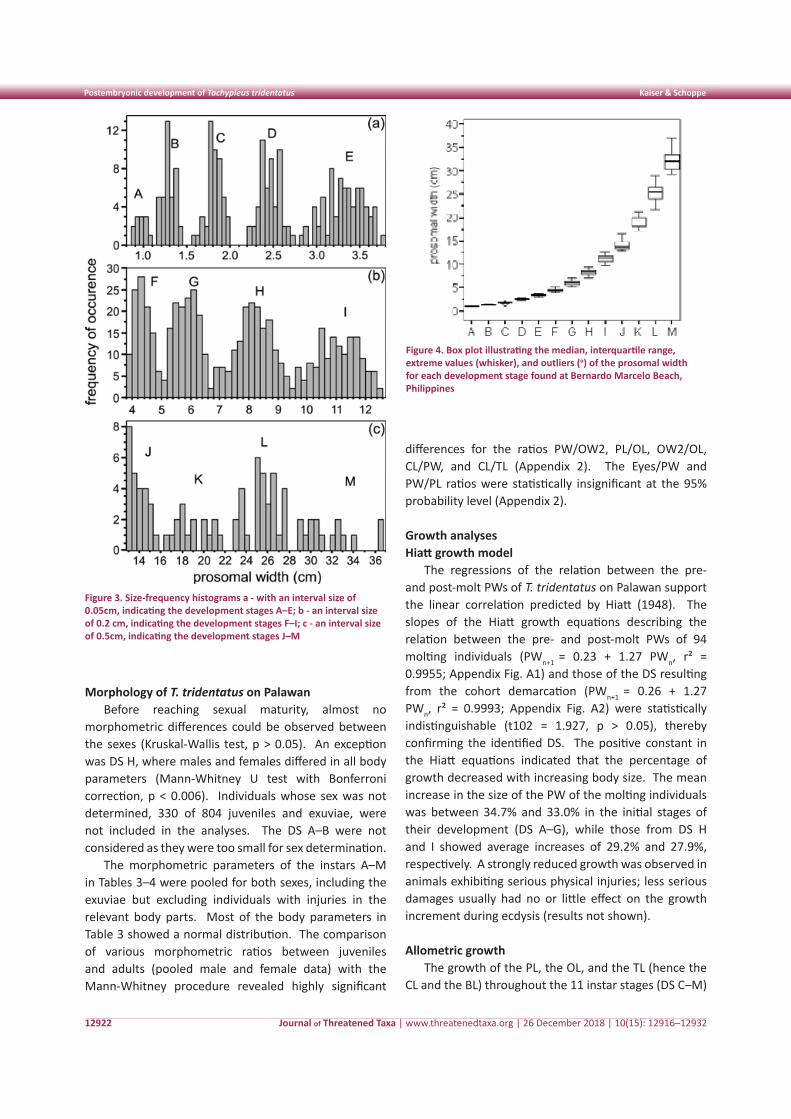
physical injuries on the prosoma, 853 PWs were used in demarcating the growth stages (Table 2). Owing to the natural growth variability of individuals, the size ranges of the DS increase with increasing age. The cohorts were, therefore, classified with frequency distributions of increasing interval widths. Five DS were identified in the histogram, presenting the smallest animals at 0.05cm intervals (Fig. 3a). The cohort with a PW ≤ 1.1cm was termed DS A, with subsequent cohorts named in alphabetical order. It has to be stressed that DS A does not represent PES 1, the trilobite larvae. Trilobite larvae were not found during the study.
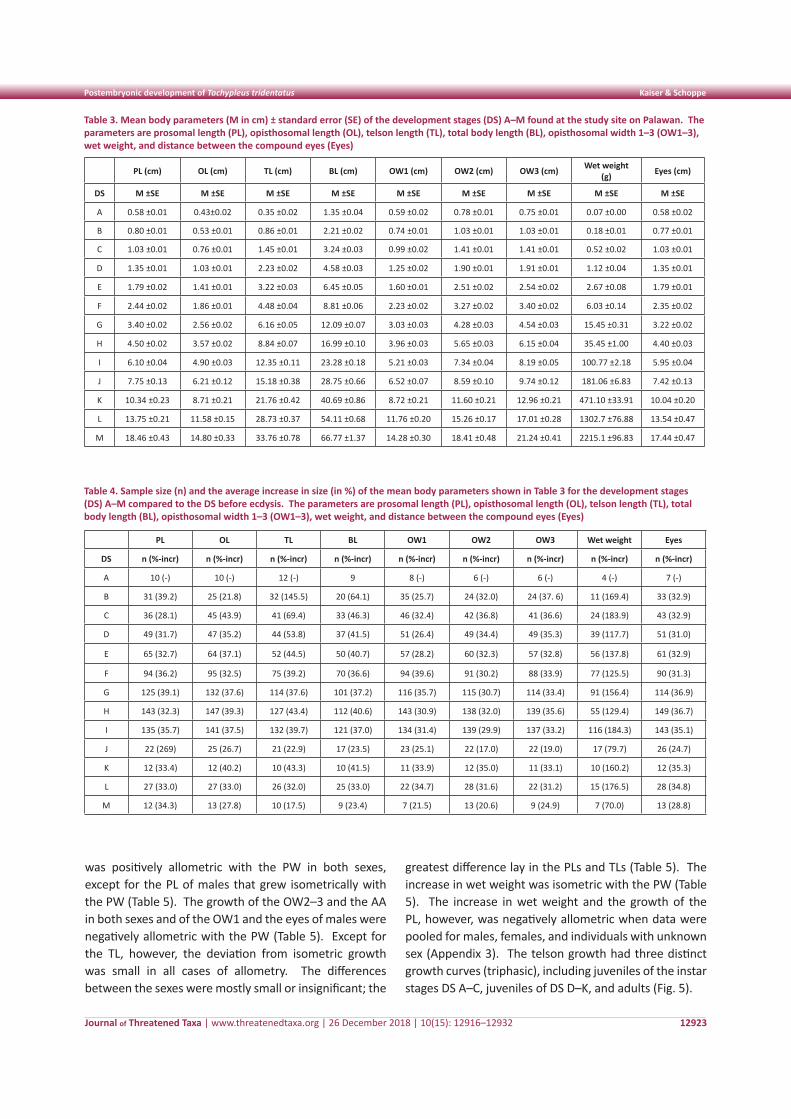
The DS F–I were depicted most clearly with an interval size of 0.2cm (Fig. 3b). For larger juveniles and adults, an interval size of 0.5cm was the best, but the number of measurements did not allow a clear demarcation between the biggest juvenile stage (DS K) and the adult stages (DS L–M) (Fig. 3c). The difference between DS J and the cohort of sexually mature males and sub-adult females (DS L) then revealed the DS K. Because the size limits of this cohort could not be unequivocally validated with our data, DS K was not included in the following statistical analyses. The mean PWs identified in FiSAT did not differ significantly from the PWs defined by visual demarcation (one-sample t test, p > 0.05), thereby confirming the cohort demarcation (Table 2, Fig. 4).
Table 2. Mean (MPW in cm) with standard error (SE) and the minimum (Min) and maximum (Max) prosomal width of the development stages (DS) A–M found at the study site on Palawan, along with sample number (n), test statistic of the Shapiro–Wilk Test (W for n ≤ 50), the Kolmogorov–Smirnov test (Z for n > 50), and associated p-values. Further shown are the results of the modal progression analyses in FiSAT II: the mean prosomal width (M2PW in cm), standard deviation (SDM2), the statistics of the one-sample t test (Ta), and the asymptotic significance (two-tailed pa)
FiSAT II
DS n MPW (cm) SE Min Max W Z p M2PW
(cm) SDM2 Ta pa
A 12 0.98 0.02 0.90 1.10 0.96 - 0.717 0.99 0.06 -0.57 0.581
B 36 1.30 0.01 1.18 1.44 0.95 - 0.113 1.31 0.07 -0.65 0.523
C 46 1.82 0.01 1.58 1.99 0.97 - 0.221 1.84 0.09 -1.23 0.225
D 54 2.48 0.02 2.22 2.84 - 0.09 0.200 2.49 0.12 -0.61 0.544
E 69 3.34 0.02 2.90 3.70 - 0.08 0.200 3.35 0.21 -0.21 0.832
F 102 4.40 0.03 3.90 5.10 - 0.09 0.200 4.44 0.25 -1.58 0.118
G 143 5.96 0.04 5.20 7.00 - 0.06 0.066 5.96 0.45 0.08 0.939
H 159 8.23 0.04 7.02 9.42 - 0.04 0.200 8.28 0.63 -1.04 0.301
I 143 11.11 0.06 9.60 12.70 - 0.06 0.200 11.19 0.79 -1.18 0.239
J 26 14.00 0.19 12.80 16.45 0.94 - 0.100 14.11 0.87 -0.58 0.569
K 13 18.84 0.36 17.13 21.12 0.91 - 0.197 19.13 2.21 -0.79 0.443
L 34 25.40 0.29 21.66 28.90 0.98 - 0.618 25.61 1.33 -0.74 0.465
M 15 32.38 0.68 29.20 36.90 0.90 - 0.065 30.84 3.59 2.26 0.040
a A one-sample t test was used to compare M2PW with the PWs measured per DS.
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12922
Morphology of T. tridentatus on PalawanBefore reaching sexual maturity, almost no
morphometric differences could be observed between the sexes (Kruskal-Wallis test, p > 0.05). An exception was DS H, where males and females differed in all body parameters (Mann-Whitney U test with Bonferroni correction, p < 0.006). Individuals whose sex was not determined, 330 of 804 juveniles and exuviae, were not included in the analyses. The DS A–B were not considered as they were too small for sex determination.
The morphometric parameters of the instars A–M in Tables 3–4 were pooled for both sexes, including the exuviae but excluding individuals with injuries in the relevant body parts. Most of the body parameters in Table 3 showed a normal distribution. The comparison of various morphometric ratios between juveniles and adults (pooled male and female data) with the Mann-Whitney procedure revealed highly significant
differences for the ratios PW/OW2, PL/OL, OW2/OL, CL/PW, and CL/TL (Appendix 2). The Eyes/PW and PW/PL ratios were statistically insignificant at the 95% probability level (Appendix 2).
Growth analyses Hiatt growth model
The regressions of the relation between the pre- and post-molt PWs of T. tridentatus on Palawan support the linear correlation predicted by Hiatt (1948). The slopes of the Hiatt growth equations describing the relation between the pre- and post-molt PWs of 94 molting individuals (PWn+1 = 0.23 + 1.27 PWn, r² = 0.9955; Appendix Fig. A1) and those of the DS resulting from the cohort demarcation (PWn+1 = 0.26 + 1.27 PWn, r² = 0.9993; Appendix Fig. A2) were statistically indistinguishable (t102 = 1.927, p > 0.05), thereby confirming the identified DS. The positive constant in the Hiatt equations indicated that the percentage of growth decreased with increasing body size. The mean increase in the size of the PW of the molting individuals was between 34.7% and 33.0% in the initial stages of their development (DS A–G), while those from DS H and I showed average increases of 29.2% and 27.9%, respectively. A strongly reduced growth was observed in animals exhibiting serious physical injuries; less serious damages usually had no or little effect on the growth increment during ecdysis (results not shown).
Allometric growth The growth of the PL, the OL, and the TL (hence the
CL and the BL) throughout the 11 instar stages (DS C–M)
Figure 3. Size-frequency histograms a - with an interval size of 0.05cm, indicating the development stages A–E; b - an interval size of 0.2 cm, indicating the development stages F–I; c - an interval size of 0.5cm, indicating the development stages J–M
Figure 4. Box plot illustrating the median, interquartile range, extreme values (whisker), and outliers (o) of the prosomal width for each development stage found at Bernardo Marcelo Beach, Philippines
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12923
was positively allometric with the PW in both sexes, except for the PL of males that grew isometrically with the PW (Table 5). The growth of the OW2–3 and the AA in both sexes and of the OW1 and the eyes of males were negatively allometric with the PW (Table 5). Except for the TL, however, the deviation from isometric growth was small in all cases of allometry. The differences between the sexes were mostly small or insignificant; the
greatest difference lay in the PLs and TLs (Table 5). The increase in wet weight was isometric with the PW (Table 5). The increase in wet weight and the growth of the PL, however, was negatively allometric when data were pooled for males, females, and individuals with unknown sex (Appendix 3). The telson growth had three distinct growth curves (triphasic), including juveniles of the instar stages DS A–C, juveniles of DS D–K, and adults (Fig. 5).
Table 3. Mean body parameters (M in cm) ± standard error (SE) of the development stages (DS) A–M found at the study site on Palawan. The parameters are prosomal length (PL), opisthosomal length (OL), telson length (TL), total body length (BL), opisthosomal width 1–3 (OW1–3), wet weight, and distance between the compound eyes (Eyes)
PL (cm) OL (cm) TL (cm) BL (cm) OW1 (cm) OW2 (cm) OW3 (cm) Wet weight (g) Eyes (cm)
DS M ±SE M ±SE M ±SE M ±SE M ±SE M ±SE M ±SE M ±SE M ±SE
A 0.58 ±0.01 0.43±0.02 0.35 ±0.02 1.35 ±0.04 0.59 ±0.02 0.78 ±0.01 0.75 ±0.01 0.07 ±0.00 0.58 ±0.02
B 0.80 ±0.01 0.53 ±0.01 0.86 ±0.01 2.21 ±0.02 0.74 ±0.01 1.03 ±0.01 1.03 ±0.01 0.18 ±0.01 0.77 ±0.01
C 1.03 ±0.01 0.76 ±0.01 1.45 ±0.01 3.24 ±0.03 0.99 ±0.02 1.41 ±0.01 1.41 ±0.01 0.52 ±0.02 1.03 ±0.01
D 1.35 ±0.01 1.03 ±0.01 2.23 ±0.02 4.58 ±0.03 1.25 ±0.02 1.90 ±0.01 1.91 ±0.01 1.12 ±0.04 1.35 ±0.01
E 1.79 ±0.02 1.41 ±0.01 3.22 ±0.03 6.45 ±0.05 1.60 ±0.01 2.51 ±0.02 2.54 ±0.02 2.67 ±0.08 1.79 ±0.01
F 2.44 ±0.02 1.86 ±0.01 4.48 ±0.04 8.81 ±0.06 2.23 ±0.02 3.27 ±0.02 3.40 ±0.02 6.03 ±0.14 2.35 ±0.02
G 3.40 ±0.02 2.56 ±0.02 6.16 ±0.05 12.09 ±0.07 3.03 ±0.03 4.28 ±0.03 4.54 ±0.03 15.45 ±0.31 3.22 ±0.02
H 4.50 ±0.02 3.57 ±0.02 8.84 ±0.07 16.99 ±0.10 3.96 ±0.03 5.65 ±0.03 6.15 ±0.04 35.45 ±1.00 4.40 ±0.03
I 6.10 ±0.04 4.90 ±0.03 12.35 ±0.11 23.28 ±0.18 5.21 ±0.03 7.34 ±0.04 8.19 ±0.05 100.77 ±2.18 5.95 ±0.04
J 7.75 ±0.13 6.21 ±0.12 15.18 ±0.38 28.75 ±0.66 6.52 ±0.07 8.59 ±0.10 9.74 ±0.12 181.06 ±6.83 7.42 ±0.13
K 10.34 ±0.23 8.71 ±0.21 21.76 ±0.42 40.69 ±0.86 8.72 ±0.21 11.60 ±0.21 12.96 ±0.21 471.10 ±33.91 10.04 ±0.20
L 13.75 ±0.21 11.58 ±0.15 28.73 ±0.37 54.11 ±0.68 11.76 ±0.20 15.26 ±0.17 17.01 ±0.28 1302.7 ±76.88 13.54 ±0.47
M 18.46 ±0.43 14.80 ±0.33 33.76 ±0.78 66.77 ±1.37 14.28 ±0.30 18.41 ±0.48 21.24 ±0.41 2215.1 ±96.83 17.44 ±0.47
Table 4. Sample size (n) and the average increase in size (in %) of the mean body parameters shown in Table 3 for the development stages (DS) A–M compared to the DS before ecdysis. The parameters are prosomal length (PL), opisthosomal length (OL), telson length (TL), total body length (BL), opisthosomal width 1–3 (OW1–3), wet weight, and distance between the compound eyes (Eyes)
PL OL TL BL OW1 OW2 OW3 Wet weight Eyes
DS n (%-incr) n (%-incr) n (%-incr) n (%-incr) n (%-incr) n (%-incr) n (%-incr) n (%-incr) n (%-incr)
A 10 (-) 10 (-) 12 (-) 9 8 (-) 6 (-) 6 (-) 4 (-) 7 (-)
B 31 (39.2) 25 (21.8) 32 (145.5) 20 (64.1) 35 (25.7) 24 (32.0) 24 (37. 6) 11 (169.4) 33 (32.9)
C 36 (28.1) 45 (43.9) 41 (69.4) 33 (46.3) 46 (32.4) 42 (36.8) 41 (36.6) 24 (183.9) 43 (32.9)
D 49 (31.7) 47 (35.2) 44 (53.8) 37 (41.5) 51 (26.4) 49 (34.4) 49 (35.3) 39 (117.7) 51 (31.0)
E 65 (32.7) 64 (37.1) 52 (44.5) 50 (40.7) 57 (28.2) 60 (32.3) 57 (32.8) 56 (137.8) 61 (32.9)
F 94 (36.2) 95 (32.5) 75 (39.2) 70 (36.6) 94 (39.6) 91 (30.2) 88 (33.9) 77 (125.5) 90 (31.3)
G 125 (39.1) 132 (37.6) 114 (37.6) 101 (37.2) 116 (35.7) 115 (30.7) 114 (33.4) 91 (156.4) 114 (36.9)
H 143 (32.3) 147 (39.3) 127 (43.4) 112 (40.6) 143 (30.9) 138 (32.0) 139 (35.6) 55 (129.4) 149 (36.7)
I 135 (35.7) 141 (37.5) 132 (39.7) 121 (37.0) 134 (31.4) 139 (29.9) 137 (33.2) 116 (184.3) 143 (35.1)
J 22 (269) 25 (26.7) 21 (22.9) 17 (23.5) 23 (25.1) 22 (17.0) 22 (19.0) 17 (79.7) 26 (24.7)
K 12 (33.4) 12 (40.2) 10 (43.3) 10 (41.5) 11 (33.9) 12 (35.0) 11 (33.1) 10 (160.2) 12 (35.3)
L 27 (33.0) 27 (33.0) 26 (32.0) 25 (33.0) 22 (34.7) 28 (31.6) 22 (31.2) 15 (176.5) 28 (34.8)
M 12 (34.3) 13 (27.8) 10 (17.5) 9 (23.4) 7 (21.5) 13 (20.6) 9 (24.9) 7 (70.0) 13 (28.8)
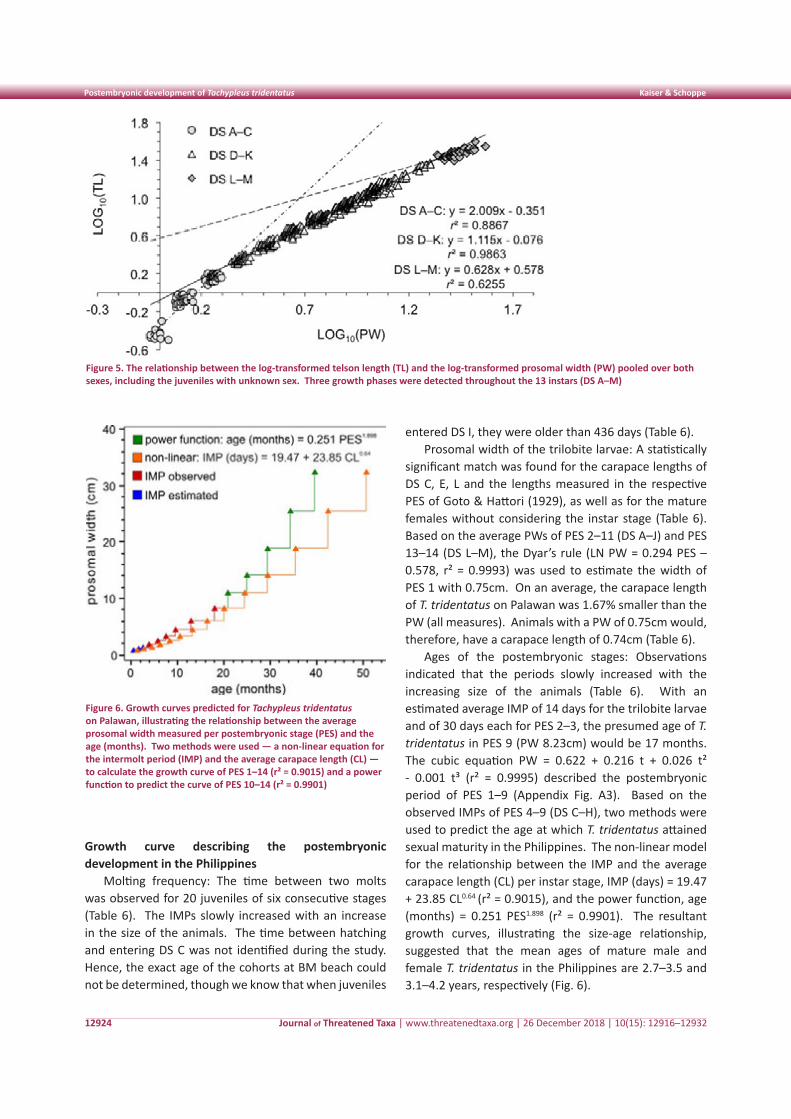
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12924
Growth curve describing the postembryonic development in the Philippines
Molting frequency: The time between two molts was observed for 20 juveniles of six consecutive stages (Table 6). The IMPs slowly increased with an increase in the size of the animals. The time between hatching and entering DS C was not identified during the study. Hence, the exact age of the cohorts at BM beach could not be determined, though we know that when juveniles
entered DS I, they were older than 436 days (Table 6). Prosomal width of the trilobite larvae: A statistically
significant match was found for the carapace lengths of DS C, E, L and the lengths measured in the respective PES of Goto & Hattori (1929), as well as for the mature females without considering the instar stage (Table 6). Based on the average PWs of PES 2–11 (DS A–J) and PES 13–14 (DS L–M), the Dyar’s rule (LN PW = 0.294 PES – 0.578, r² = 0.9993) was used to estimate the width of PES 1 with 0.75cm. On an average, the carapace length of T. tridentatus on Palawan was 1.67% smaller than the PW (all measures). Animals with a PW of 0.75cm would, therefore, have a carapace length of 0.74cm (Table 6).
Ages of the postembryonic stages: Observations indicated that the periods slowly increased with the increasing size of the animals (Table 6). With an estimated average IMP of 14 days for the trilobite larvae and of 30 days each for PES 2–3, the presumed age of T. tridentatus in PES 9 (PW 8.23cm) would be 17 months. The cubic equation PW = 0.622 + 0.216 t + 0.026 t² - 0.001 t³ (r² = 0.9995) described the postembryonic period of PES 1–9 (Appendix Fig. A3). Based on the observed IMPs of PES 4–9 (DS C–H), two methods were used to predict the age at which T. tridentatus attained sexual maturity in the Philippines. The non-linear model for the relationship between the IMP and the average carapace length (CL) per instar stage, IMP (days) = 19.47 + 23.85 CL0.64 (r² = 0.9015), and the power function, age (months) = 0.251 PES1.898 (r² = 0.9901). The resultant growth curves, illustrating the size-age relationship, suggested that the mean ages of mature male and female T. tridentatus in the Philippines are 2.7–3.5 and 3.1–4.2 years, respectively (Fig. 6).
Figure 6. Growth curves predicted for Tachypleus tridentatus on Palawan, illustrating the relationship between the average prosomal width measured per postembryonic stage (PES) and the age (months). Two methods were used — a non-linear equation for the intermolt period (IMP) and the average carapace length (CL) — to calculate the growth curve of PES 1–14 (r² = 0.9015) and a power function to predict the curve of PES 10–14 (r² = 0.9901)
Figure 5. The relationship between the log-transformed telson length (TL) and the log-transformed prosomal width (PW) pooled over both sexes, including the juveniles with unknown sex. Three growth phases were detected throughout the 13 instars (DS A–M)
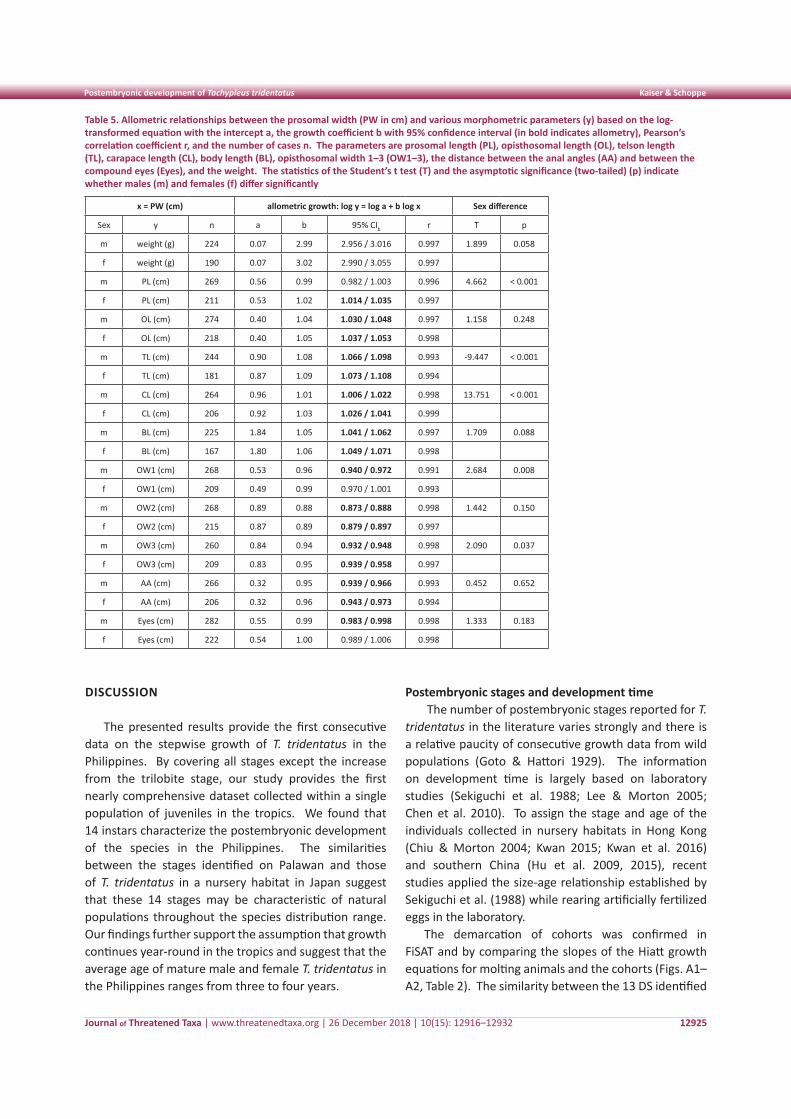
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12925
DISCUSSION
The presented results provide the first consecutive data on the stepwise growth of T. tridentatus in the Philippines. By covering all stages except the increase from the trilobite stage, our study provides the first nearly comprehensive dataset collected within a single population of juveniles in the tropics. We found that 14 instars characterize the postembryonic development of the species in the Philippines. The similarities between the stages identified on Palawan and those of T. tridentatus in a nursery habitat in Japan suggest that these 14 stages may be characteristic of natural populations throughout the species distribution range. Our findings further support the assumption that growth continues year-round in the tropics and suggest that the average age of mature male and female T. tridentatus in the Philippines ranges from three to four years.
Postembryonic stages and development time The number of postembryonic stages reported for T.
tridentatus in the literature varies strongly and there is a relative paucity of consecutive growth data from wild populations (Goto & Hattori 1929). The information on development time is largely based on laboratory studies (Sekiguchi et al. 1988; Lee & Morton 2005; Chen et al. 2010). To assign the stage and age of the individuals collected in nursery habitats in Hong Kong (Chiu & Morton 2004; Kwan 2015; Kwan et al. 2016) and southern China (Hu et al. 2009, 2015), recent studies applied the size-age relationship established by Sekiguchi et al. (1988) while rearing artificially fertilized eggs in the laboratory.
The demarcation of cohorts was confirmed in FiSAT and by comparing the slopes of the Hiatt growth equations for molting animals and the cohorts (Figs. A1–A2, Table 2). The similarity between the 13 DS identified
Table 5. Allometric relationships between the prosomal width (PW in cm) and various morphometric parameters (y) based on the log-transformed equation with the intercept a, the growth coefficient b with 95% confidence interval (in bold indicates allometry), Pearson’s correlation coefficient r, and the number of cases n. The parameters are prosomal length (PL), opisthosomal length (OL), telson length (TL), carapace length (CL), body length (BL), opisthosomal width 1–3 (OW1–3), the distance between the anal angles (AA) and between the compound eyes (Eyes), and the weight. The statistics of the Student’s t test (T) and the asymptotic significance (two-tailed) (p) indicate whether males (m) and females (f) differ significantly
x = PW (cm) allometric growth: log y = log a + b log x Sex difference
Sex y n a b 95% CIb r T p
m weight (g) 224 0.07 2.99 2.956 / 3.016 0.997 1.899 0.058
f weight (g) 190 0.07 3.02 2.990 / 3.055 0.997
m PL (cm) 269 0.56 0.99 0.982 / 1.003 0.996 4.662 < 0.001
f PL (cm) 211 0.53 1.02 1.014 / 1.035 0.997
m OL (cm) 274 0.40 1.04 1.030 / 1.048 0.997 1.158 0.248
f OL (cm) 218 0.40 1.05 1.037 / 1.053 0.998
m TL (cm) 244 0.90 1.08 1.066 / 1.098 0.993 -9.447 < 0.001
f TL (cm) 181 0.87 1.09 1.073 / 1.108 0.994
m CL (cm) 264 0.96 1.01 1.006 / 1.022 0.998 13.751 < 0.001
f CL (cm) 206 0.92 1.03 1.026 / 1.041 0.999
m BL (cm) 225 1.84 1.05 1.041 / 1.062 0.997 1.709 0.088
f BL (cm) 167 1.80 1.06 1.049 / 1.071 0.998
m OW1 (cm) 268 0.53 0.96 0.940 / 0.972 0.991 2.684 0.008
f OW1 (cm) 209 0.49 0.99 0.970 / 1.001 0.993
m OW2 (cm) 268 0.89 0.88 0.873 / 0.888 0.998 1.442 0.150
f OW2 (cm) 215 0.87 0.89 0.879 / 0.897 0.997
m OW3 (cm) 260 0.84 0.94 0.932 / 0.948 0.998 2.090 0.037
f OW3 (cm) 209 0.83 0.95 0.939 / 0.958 0.997
m AA (cm) 266 0.32 0.95 0.939 / 0.966 0.993 0.452 0.652
f AA (cm) 206 0.32 0.96 0.943 / 0.973 0.994
m Eyes (cm) 282 0.55 0.99 0.983 / 0.998 0.998 1.333 0.183
f Eyes (cm) 222 0.54 1.00 0.989 / 1.006 0.998
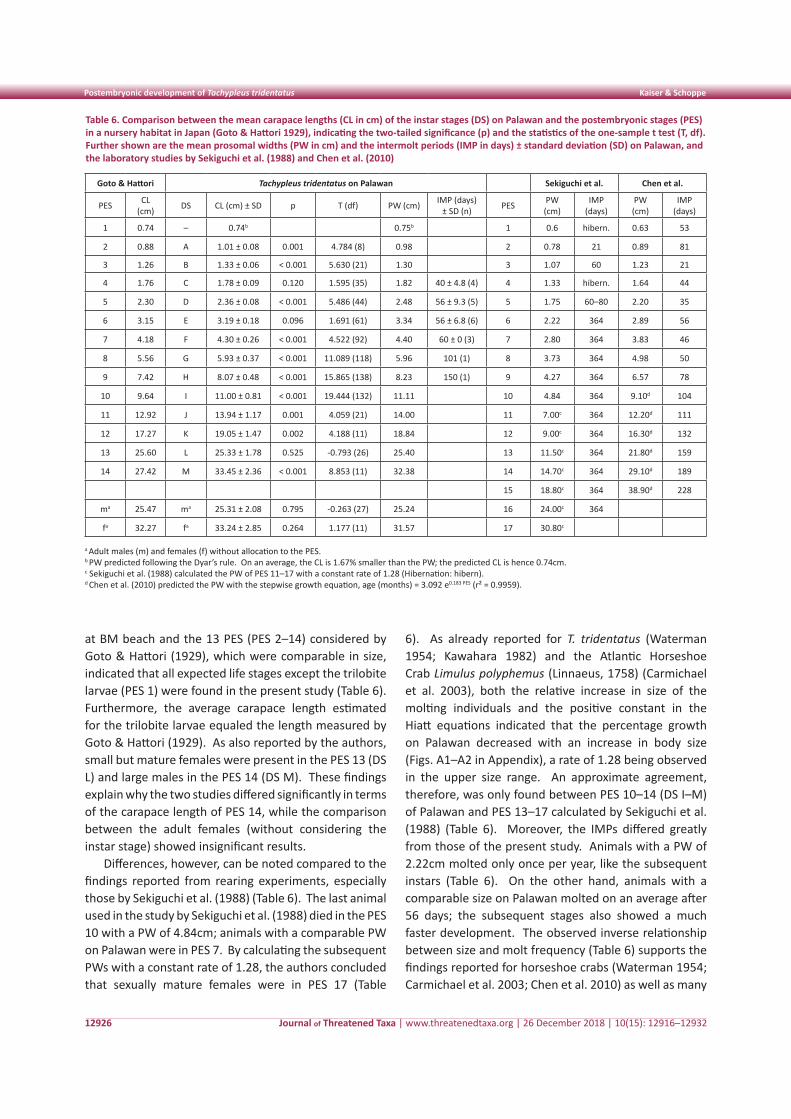
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12926
at BM beach and the 13 PES (PES 2–14) considered by Goto & Hattori (1929), which were comparable in size, indicated that all expected life stages except the trilobite larvae (PES 1) were found in the present study (Table 6). Furthermore, the average carapace length estimated for the trilobite larvae equaled the length measured by Goto & Hattori (1929). As also reported by the authors, small but mature females were present in the PES 13 (DS L) and large males in the PES 14 (DS M). These findings explain why the two studies differed significantly in terms of the carapace length of PES 14, while the comparison between the adult females (without considering the instar stage) showed insignificant results.
Differences, however, can be noted compared to the findings reported from rearing experiments, especially those by Sekiguchi et al. (1988) (Table 6). The last animal used in the study by Sekiguchi et al. (1988) died in the PES 10 with a PW of 4.84cm; animals with a comparable PW on Palawan were in PES 7. By calculating the subsequent PWs with a constant rate of 1.28, the authors concluded that sexually mature females were in PES 17 (Table
6). As already reported for T. tridentatus (Waterman 1954; Kawahara 1982) and the Atlantic Horseshoe Crab Limulus polyphemus (Linnaeus, 1758) (Carmichael et al. 2003), both the relative increase in size of the molting individuals and the positive constant in the Hiatt equations indicated that the percentage growth on Palawan decreased with an increase in body size (Figs. A1–A2 in Appendix), a rate of 1.28 being observed in the upper size range. An approximate agreement, therefore, was only found between PES 10–14 (DS I–M) of Palawan and PES 13–17 calculated by Sekiguchi et al. (1988) (Table 6). Moreover, the IMPs differed greatly from those of the present study. Animals with a PW of 2.22cm molted only once per year, like the subsequent instars (Table 6). On the other hand, animals with a comparable size on Palawan molted on an average after 56 days; the subsequent stages also showed a much faster development. The observed inverse relationship between size and molt frequency (Table 6) supports the findings reported for horseshoe crabs (Waterman 1954; Carmichael et al. 2003; Chen et al. 2010) as well as many
Table 6. Comparison between the mean carapace lengths (CL in cm) of the instar stages (DS) on Palawan and the postembryonic stages (PES) in a nursery habitat in Japan (Goto & Hattori 1929), indicating the two-tailed significance (p) and the statistics of the one-sample t test (T, df). Further shown are the mean prosomal widths (PW in cm) and the intermolt periods (IMP in days) ± standard deviation (SD) on Palawan, and the laboratory studies by Sekiguchi et al. (1988) and Chen et al. (2010)
Goto & Hattori Tachypleus tridentatus on Palawan Sekiguchi et al. Chen et al.
PES CL (cm) DS CL (cm) ± SD p T (df) PW (cm) IMP (days)
± SD (n) PES PW (cm)
IMP (days)
PW (cm)
IMP (days)
1 0.74 – 0.74b 0.75b 1 0.6 hibern. 0.63 53
2 0.88 A 1.01 ± 0.08 0.001 4.784 (8) 0.98 2 0.78 21 0.89 81
3 1.26 B 1.33 ± 0.06 < 0.001 5.630 (21) 1.30 3 1.07 60 1.23 21
4 1.76 C 1.78 ± 0.09 0.120 1.595 (35) 1.82 40 ± 4.8 (4) 4 1.33 hibern. 1.64 44
5 2.30 D 2.36 ± 0.08 < 0.001 5.486 (44) 2.48 56 ± 9.3 (5) 5 1.75 60–80 2.20 35
6 3.15 E 3.19 ± 0.18 0.096 1.691 (61) 3.34 56 ± 6.8 (6) 6 2.22 364 2.89 56
7 4.18 F 4.30 ± 0.26 < 0.001 4.522 (92) 4.40 60 ± 0 (3) 7 2.80 364 3.83 46
8 5.56 G 5.93 ± 0.37 < 0.001 11.089 (118) 5.96 101 (1) 8 3.73 364 4.98 50
9 7.42 H 8.07 ± 0.48 < 0.001 15.865 (138) 8.23 150 (1) 9 4.27 364 6.57 78
10 9.64 I 11.00 ± 0.81 < 0.001 19.444 (132) 11.11 10 4.84 364 9.10d 104
11 12.92 J 13.94 ± 1.17 0.001 4.059 (21) 14.00 11 7.00c 364 12.20d 111
12 17.27 K 19.05 ± 1.47 0.002 4.188 (11) 18.84 12 9.00c 364 16.30d 132
13 25.60 L 25.33 ± 1.78 0.525 -0.793 (26) 25.40 13 11.50c 364 21.80d 159
14 27.42 M 33.45 ± 2.36 < 0.001 8.853 (11) 32.38 14 14.70c 364 29.10d 189
15 18.80c 364 38.90d 228
ma 25.47 ma 25.31 ± 2.08 0.795 -0.263 (27) 25.24 16 24.00c 364
fa 32.27 fa 33.24 ± 2.85 0.264 1.177 (11) 31.57 17 30.80c
a Adult males (m) and females (f) without allocation to the PES.b PW predicted following the Dyar’s rule. On an average, the CL is 1.67% smaller than the PW; the predicted CL is hence 0.74cm.c Sekiguchi et al. (1988) calculated the PW of PES 11–17 with a constant rate of 1.28 (Hibernation: hibern).d Chen et al. (2010) predicted the PW with the stepwise growth equation, age (months) = 3.092 e0.183 PES (r² = 0.9959).

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12927
crustacean species (Kurata 1962; Caddy 1987; Chang et al. 2012).
Considering the high water temperatures in the study region throughout the year and the increasing length of the IMPs, we assumed that the trilobite larvae in the Philippines molt after two weeks and the PES 2–3 after 30 days. Following our estimations, the mean age of juveniles entering PES 10 (mean PW of 11.1cm) was 14.5 months (Fig. 6). The observed IMPs and our age estimations for adults were similar to the four years estimated by Chen et al. (2010), although an additional stage was observed in their experiment (Table 6). The authors reared juvenile T. tridentatus under conditions better comparable to those prevailing at BM beach. The temperature was 28–30 °C, seawater salinity was 30‰, water flow was maintained, more substrate was provided for digging, and the tank was much larger than that in Japan. The least time to hatch and greatest survival of the Asian species were observed in water temperatures of around 29°C (Carmichael & Brush 2012), and Chen et al. (2004) suggested 28–31 °C as the optimal seawater temperature for the year-round growth of the juvenile T. tridentatus. These observations were in line with the results of Yeh (1999, cited in Chen et al. 2010) and Lee & Morton (2005), who reported that ecdysis in T. tridentatus continues when the temperature remains at >28°C but stops at <22°C. Our findings support the assumption that growth continues year-round in the tropics and suggest that T. tridentatus in the Philippines attain sexual maturity at the age of three to four years. The similarity with the instars from the Inland Sea in Japan (Goto & Hattori 1929), where the water temperature drops down to 13°C in winter, suggests that 14 stages may be characteristic of the postembryonic development of natural populations throughout the distribution range of T. tridentatus. Temperature seems to have less influence on the molt increment than on the IMP. To confirm these findings, however, further investigations should be carried out in habitats located at the limits of the distribution range of the species.
In light of the fact that the growth increments per molt of T. tridentatus in the two nursery habitats were comparable, it is likely that the caging operations are responsible for the deviations observed in the laboratory studies. Significantly reduced growth increments at each molt and longer IMPs in the laboratory were observed in horseshoe crabs (Carmichael & Brush 2012) and various crustacean species (e.g., Hiatt 1948; Harms et al. 1994; Bonilla-Gómez et al. 2013). The effects of the holding time at the laboratory and the space available in the holding tank were reported (e.g., González-Gurriarán
et al. 1998). The development of L. polyphemus in the laboratory was considerably slower than that in their natural habitat — the postembryonic growth time was nearly halved when the embryonic development took place in nature (Jegla & Costlow 1982). As shown for horseshoe crabs (Carmichael & Brush 2012; Hu et al. 2013) and several crustacean species (Hartnoll 2001; Chang et al. 2012), the IMP increased and the rate of increase in size decreased when the quantity or quality of food in culture was suboptimal. The diet of horseshoe crabs in nature is broad and highly mixed and they move up the food webs as they age and grow (Carmichael et al. 2004; Zhou & Morton 2004). The most commonly offered diet in culture was brine shrimp (Artemia spp.) or dietary supplements that were not part of their known natural diet (see Carmichael & Brush 2012). Moreover, recent studies with horseshoe crabs show the importance of the sediment type for growth and survival (Hong et al. 2009; Hieb et al. 2015), while others found shorter hatching times and higher molting frequency and molt increments with increased water circulation or increased dissolved oxygen concentrations (Carmichael & Brush 2012). The relationships with environmental factors and the potential interactions among different variables were not resolved (Carmichael & Brush 2012). These reports, however, illustrate that variations in the growth rate can be considerable, depending on the abiotic and biotic parameters, which might explain the large differences observed with the number of stages and the size-age relationship reported by Sekiguchi (1988) and the comparable development time, with only one additional instar stage reported by Chen et al. (2010). These findings may further imply the similarity between BM beach and the nursery habitat of Goto & Hattori (1929) in terms of environment conditions (apart from water temperature) and quality of food supply. Further studies are needed to characterize the habitat of different populations and to identify the key drivers for growth and survival in nature and in rearing experiments.
Growth patternOur finding that the relationship between the pre-
and post-molt PWs of juvenile horseshoe crabs could be fitted with the Hiatt growth model is consistent with the results of previous studies (Carmichael et al. 2003; Hu et al. 2015). Moreover, the Hiatt equations were almost identical to the equation for a juvenile population assessed at a nursery habitat in southern China (Hu et al. 2015). As also reported by Sekiguchi et al. (1988), three distinct growth curves described the relative

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12928
growth of the TL and the PW throughout the 13 stages. Accordingly, Chen et al. (2010) reported two phases for the nine juvenile stages considered by them. Deviations from isometric growth in the other body parts were fairly small on Palawan — the shape of the horseshoe crab at different development stages was therefore similar. Isometric growth was reported by Chen et al. (2010) for most of the body parts of the juveniles in their study. Lee & Morton (2005), in contrast, reported a positive growth allometry for the weight of juveniles collected in their natural habitat but reared in the laboratory. On Palawan, significant differences between juveniles and adults were indicated in most of the analysed body ratios. Most findings agree with those of Chiu & Morton (2003), although they reported insignificant differences for the CL/PW and CL/TL ratios. Several studies (for instance, Yamasaki et al. 1988; Chiu & Morton 2003; Mohamad et al. 2016) revealed that morphometric parameters, body ratios, and the allometric relationships of adult and juvenile T. tridentatus differed significantly between populations and geographic regions, a characteristic that might explain the differences between the studies. The differences might also be due to the different size ranges considered and whether or not sex was differentiated.
LimitationsThe absence of the trilobite larvae might suggest
that no recent spawning activity occurred at BM beach. Because the subsequent stages were present and the smallest instar stages are difficult to detect, however, they may have been simply overlooked. The newly hatched larvae do not need to feed because they can subsist on the yolk of the embryo (Botton et al. 1992). The low occurrence of PES 12 (DS K) at BM beach supports earlier observations that larger juveniles were moving further down in the intertidal zone, with subadults at the seaward limit (Rudloe 1981; Kaiser 2002; Hu et al. 2009; Morton & Lee 2011). It might also imply a greater mortality of juveniles in PES 12, though laboratory studies revealed a decrease in mortality rates with increasing age (Carmichael & Brush 2012).
It was recognized that mark-recapture experiments were often the only method to validate the growth of natural populations (González-Gurriarán et al. 1998; Lee & Morton 2005; Hu et al. 2015), but several authors emphasized that caution should be used when extrapolating from tagged animals to natural populations (Kurata 1962; Caddy 1987). Although handling during measuring and marking may have disturbed the juveniles in the present study, we doubt that cutting the mobile spines for identification would have harmed the animals
to an extent that their growth was no longer natural — we observed that animals with injuries showed lower rather than higher molt increments. Given a recapture rate of 57% in 2001 (Kaiser 2002), the labels glued on the prosoma also seemed to pose no adverse impacts on health.
CONCLUSIONS
Determining when a horseshoe crab reaches sexual maturity is of extreme importance for interpreting the population dynamics to enable their management and conservation (Carmichael et al. 2015). By presenting the first consecutive data on the postembryonic development of T. tridentatus in the Philippines, the present study adds to the fragmentary background knowledge about the species in southeastern Asia and provides the information needed to assess the demographics of juvenile populations and judge the status of T. tridentatus in the Philippines. With populations having decreased dramatically over their distribution range, it is clear that these data are now in demand. Because of the small number of IMPs observed and the absence of the trilobite larvae, however, more data are needed to validate the present findings. The relative paucity of consecutive growth data from wild populations and the lack of comparable data for the tropics made it difficult to judge the universal applicability of the dataset. Nevertheless, the reported results provide sound quantitative and qualitative baseline data for future assessment and monitoring studies in the tropics. To define a wider range of applicability, future investigations should aim to determine the size-age relationship of natural T. tridentatus populations from different geographic regions, preferably in habitats located at the limits of the species distribution range. These studies should also characterize the abiotic and biotic parameters of habitats because the causes of variable growth and survival rates have important implications for conservation and aquaculture efforts that are aimed at restoring depleted populations of horseshoe crabs.
REFERENCES
Asano, U. (1942). On the life history of Tachypleus tridentatus. Botany and Zoology 10: 120–124 (in Japanese).
Berkson, J., C.-P. Chen, J. Mishra, P. Shin, B. Spear & J. Zaldívar-Rae (2009). A discussion of horseshoe crab management in five countries: Taiwan, India, China, United States, and Mexico, pp465–475. In: Tanacredi, J.T., M.L. Botton & D. Smith (eds.). Biology and Conservation of Horseshoe Crabs. Springer, Boston, MA, USA,

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12929
662pp.Bonaventura, J., C. Bonaventura & S. Tesh (eds.) (1982). Physiology and
Biology of Horseshoe Crabs: Studies on Normal and Environmental Stressed Animals. Alan R. Liss, Inc., New York, 316pp.
Bonilla-Gómez, J.L., X. Chiappa-Carrara, C. Galindo, G. Cuzón & G. Gaxiola (2013). Effects of adaptation to laboratory conditions on growth, molting, and food consumption of juvenile Farfantepenaeus duorarum (Decapoda: Penaeidae). Journal of Crustacean Biology 33(2): 191–197.
Botton, M.L., P.J. Loveland & T.R. Jacobsen (1992). Overwintering by trilobite larvae of the horseshoe crab Limulus polyphemus on a sandy beach of Delaware Bay (New Jersey, USA). Marine Ecology Progress Series 88: 289–292.
Caddy, J. (1987). Size-frequency analysis for Crustacea: moult increment and frequency models for stock assessment. Kuwait Bulletin of Marine Science 9: 43–61.
Cao, J., W. Guan, S. Truesdell, Y. Chen & S. Tian (2016). An individual-based probabilistic model for simulating fisheries population dynamics. Aquaculture and Fisheries 1(Supplement C): 34–40; https://doi.org/10.1016/j.aaf.2016.10.001
Carmichael, R.H., D. Rutecki & I. Valiela (2003). Abundance and population structure of the Atlantic Horseshoe Crab Limulus polyphemus in Pleasant Bay, Cape Cod. Marine Ecology Progress Series 246: 225–239.
Carmichael, R.H., D. Rutecki, B. Annett, E. Gaines & I. Valiela (2004). Position of horseshoe crabs in estuarine food webs: N and C stable isotopic study of foraging ranges and diet composition. Journal of Experimental Marine Biology and Ecology 299(2): 231–253; https://doi.org/10.1016/j.jembe.2003.09.006
Carmichael, R.H., E. Gaiues, Z. Sheller, A. Tong, A. Clapp & I. Valiela (2009). Diet composition of juvenile horseshoe crabs: implications for growth and survival of natural and cultured stocks, pp521–534. In: Tanacredi, J.T., M.L. Botton & D. Smith (eds.). Biology and Conservation of Horseshoe Crabs. Springer, Boston, MA, USA, 662pp.
Carmichael, R.H. & E. Brush (2012). Three decades of horseshoe crab rearing: a review of conditions for captive growth and survival. Reviews in Aquaculture 4(1): 32–43; https://doi.org/10.1111/j.1753-5131.2012.01059.x
Carmichael, R.H., E.E. Hieb, G. Gauvry & C.N. Shuster (2015). Examination of large exuviae with mating scars: do female American Horseshoe Crabs, Limulus polyphemus, molt after sexual maturity? pp353–366. In: Carmichael, R.H., M.L. Botton, P.K.S. Shin & S.G. Cheung (eds.). Changing Global Perspectives on Horseshoe Crab Biology, Conservation and Management. Springer International Publishing, Cham, 599pp.
Castro, M. (1992). A methodology for obtaining information on the age structure and growth rates of the Norway lobster, Nephrops norvegicus (L.) (Decapoda, Nephropoidea). Crustaceana 63(1): 29–43.
Chang, Y.-J., C.-L. Sun, Y. Chen & S.-Z. Yeh (2012). Modelling the growth of crustacean species. Reviews in Fish Biology and Fisheries 22(1): 157–187; https://doi.org/10.1007/s11160-011-9228-4
Chen, C.-P., H.-Y. Yeh & P.-F. Lin (2004). Conservation of the horseshoe crab at Kinmen, Taiwan: strategies and practices. Biodiversity & Conservation 13(10): 1889–1904; https://doi.org/10.1023/b:bioc.0000035868.11083.84
Chen, C.-P., M.C. Yang, L.F. Fan, G. Qiu, Y.Y. Liao & H.L. Hsieh (2015). Co-occurrence of juvenile horseshoe crabs Tachypleus tridentatus and Carcinoscorpius rotundicauda in an estuarine bay, southwestern China. Aquatic Biology 24(2): 117–126.
Chen, C.-P., R.F. Chen, P.H. Chen, H.C. Liu & H.L. Hsieh (2016). Intermediate culture of juvenile horseshoe crab (Tachypleus tridentatus) mixed with juvenile spotted babylon (Babylonia areolata) for restocking horseshoe crab populations. AACL Bioflux 9(3): 623–633.
Chen, Y., C.W. Lau, S.G. Cheung, C.H. Ke & P.K.S. Shin (2010). Enhanced growth of juvenile Tachypleus tridentatus (Chelicerata: Xiphosura) in the laboratory: a step towards population restocking
for conservation of the species. Aquatic Biology 11(1): 37–46.Chiu, H.M.C. & B. Morton (2003). The morphological differentiation
of two horseshoe crab species, Tachypleus tridentatus and Carcinoscorpius rotundicauda (Xiphosura), in Hong Kong with a regional Asian comparison. Journal of Natural History 37(19): 2369–2382; https://doi.org/10.1080/00222930210149753
Chiu, H.M.C. & B. Morton (2004). The behaviour of juvenile horseshoe crabs, Tachypleus tridentatus (Xiphosura), on a nursery beach at Shui Hau Wan, Hong Kong. Hydrobiologia 523(1–3): 29–35; https://doi.org/10.1023/b:hydr.0000033085.71861.63
Cunningham, S.R. & M.Z. Darnell (2015). Temperature-dependent growth and molting in early juvenile blue crabs Callinectes sapidus. Journal of Shellfish Research 34(2): 505–510; https://doi.org/10.2983/035.034.0246
FAO (2006–2017). Fisheries and aquaculture software. FISAT II – FAO-ICLARM Stock Assessment Tool. In: FAO Fisheries and Aquaculture Department. FAO, Rome. Downloaded on 02 November 2017; http://www.fao.org/fishery/topic/16072/en
Field, A.P. (ed.) (2009). Discovering Statistics using SPSS (and sex and drugs and rock ’n’ roll), 3rd Edition. SAGE Publications, London, UK, 822pp.
Froese, R., A. Stern-Pirlot, H. Winker & D. Gascuel (2008). Size matters: How single-species management can contribute to ecosystem-based fisheries management. Fisheries Research 92(2–3): 231–241; http://doi.org/10.1016/j.fishres.2008.01.005
Fuiman, L.A. (1983). Growth gradients in fish larvae. Journal of Fish Biology 23(1): 117–123; https://doi.org/10.1111/j.1095-8649.1983.tb02886.x
González-Gurriarán, E., J. Freire, A.C. Fariña & A. Fernández (1998). Growth at moult and intermoult period in the Norway Lobster Nephrops norvegicus from Galician waters. ICES Journal of Marine Science 55(5): 924–940; https://doi.org/10.1006/jmsc.1998.0365
Goto, S. & O. Hattori (1929). Notes on the spawning habits and growth stages of the Japanese king-crab. X Congrès International de Zoologie 2: 1147–1155.
Gould, S.J. (1966). Allometry and size in ontogeny and phylogeny. Biological Reviews 41(4): 587–640; https://doi.org/10.1111/j.1469-185X.1966.tb01624.x
Harms, J., B. Meyer-Harms, R.R. Dawirs & K. Anger (1994). Growth and physiology of Carcinus maenas (Decapoda, Portunidae) larvae in the field and in laboratory experiments. Marine Ecology Progress Series 108: 107–118.
Hartnoll, R.G. (2001). Growth in Crustacea - twenty years on. Hydrobiologia 449(1–3): 111–122; https://doi.org/10.1023/a:1017597104367
Hiatt, R.W. (1948). The biology of the Lined Shore Crab, Pachygrapsus crassipes Randall. Pacific Science 2(3): 135–213.
Hieb, E.E., J.D. Baggett, A.M. Aven & R.H. Carmichael (2015). Effects of sediment type and tank shape on horseshoe crab (Limulus polyphemus) growth and survival in culture, pp289–302. In: Carmichael, R.H., M.L. Botton, P.K.S. Shin & S.G. Cheung (eds.). Changing Global Perspectives on Horseshoe Crab Biology, Conservation and Management. Springer International Publishing, Cham, 599pp.
Hong, S., X. Zhang, Y. Zhao, Y. Xie, Y. Zhang & H. Xu (2009). Effect of sediment type on growth and survival of juvenile horseshoe crabs (Tachypleus tridentatus), pp535–540. In: Tanacredi, J.T., M.L. Botton & D. Smith (eds.). Biology and Conservation of Horseshoe Crabs. Springer, Boston, MA, USA, 662pp.
Hsieh, H.-L. & C.-P. Chen (2009). Conservation program for the Asian horseshoe crab Tachypleus tridentatus in Taiwan: characterizing the microhabitat of nursery grounds and restoring spawning grounds, pp417–438. In: Tanacredi, J.T., M.L. Botton & D. Smith (eds.). Biology and Conservation of Horseshoe Crabs. Springer, Boston, MA, USA, 662pp.
Hsieh, H.-L. & C.-P. Chen (2015). Current status of Tachypleus tridentatus in Taiwan for Red List assessment, pp383–396. In: Carmichael, R.H., M.L. Botton, P.K.S. Shin & S.G. Cheung (eds.). Changing Global Perspectives on Horseshoe Crab Biology,

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12930
Conservation and Management. Springer International Publishing, Cham, 599pp.
Hu, M., Y. Wang, Y. Chen, S.G. Cheung, P.K.S. Shin & Q. Li (2009). Summer distribution and abundance of juvenile Chinese horseshoe crabs Tachypleus tridentatus along an intertidal zone in southern China. Aquatic Biology 7(1–2): 107–112.
Hu, M., Y. Wang, S.G. Cheung & P.K.S. Shin (2013). Comparison of different frozen natural foods on survival and growth of juvenile Chinese horseshoe crab Tachypleus tridentatus (Leach, 1819): implications on laboratory culture. Aquaculture Research 44(4): 567–573; https://doi.org/10.1111/j.1365-2109.2011.03059.x
Hu, M., B.K.Y. Kwan, Y. Wang, S.G. Cheung & P.K.S. Shin (2015). Population structure and growth of juvenile horseshoe crabs Tachypleus tridentatus and Carcinoscorpius rotundicauda (Xiphosura) in southern China, pp167–180. In: Carmichael, H.R., L.M. Botton, K.S.P. Shin & G.S. Cheung (eds.). Changing Global Perspectives on Horseshoe Crab Biology, Conservation and Management. Springer International Publishing, Cham, 599pp.
Itow, T. (1993). Crisis in the Seto Inland Sea: the decimation of the horseshoe crab. EMECS Newsletter 3: 10–11.
IUCN (2017). The IUCN Red List of Threatened Species, Version 2017-3. http://www.iucnredlist.org. Downloaded on 07 December 2017.
Jegla, T.C. & J.D. Costlow (1982). Temperature and salinity effects on developmental and early posthatch stages of Limulus, pp103–113. In: Bonaventura, J., C. Bonaventura & S. Tesh (eds.). Physiology and Biology of Horseshoe Crabs: Studies on Normal and Environmentally Stressed Animals. Alan R. Liss, Inc., New York, 316pp.
Kaiser, D. (2002). Zur Ökologie und Morphologie von Tachypleus tridentatus (Leach, 1819) (Chelicerata: Merostomata) im Gezeitenwatt der Ostküste von Palawan (Philippinen). Diploma Thesis. Faculty of Biology, University of Rostock, vii+131pp.
Kawahara, D. (1982). Investigation on ecology of horseshoe crab larvae. Aquabiology 4: 380–382.
Kawahara, H. (1984). Growth and diurnal rhythm of horseshoe crabs. Nature and Animals 14: 22–26.
Kurata, H. (1962). Studies on the age and growth of crustacea. Bulletin of the Hokkaido Regional Fisheries Research Laboratory 24: 1–115.
Kwan, B.K.Y. (2015). Ecological and health perspectives of juvenile Asian horseshoe crabs. PhD Thesis. Department of Biology and Chemistry, City University of Hong Kong, xxx+226pp.
Kwan, B.K.Y., S.G. Cheung & P.K.S. Shin (2015). A dual stable isotope study for diet composition of juvenile Chinese horseshoe crab Tachypleus tridentatus (Xiphosura) on a seagrass-covered intertidal mudflat. Marine Biology 162(5): 1137–1143; https://doi.org/10.1007/s00227-015-2647-3
Kwan, B.K.Y., H.-L. Hsieh, S.G. Cheung & P.K.S. Shin (2016). Present population and habitat status of potentially threatened Asian horseshoe crabs Tachypleus tridentatus and Carcinoscorpius rotundicauda in Hong Kong: a proposal for marine protected areas. Biodiversity and Conservation 25(4): 673–692; https://doi.org/10.1007/s10531-016-1084-z
Lee, C.N. & B. Morton (2005). Experimentally derived estimates of growth by juvenile Tachypleus tridentatus and Carcinoscorpius rotundicauda (Xiphosura) from nursery beaches in Hong Kong. Journal of Experimental Marine Biology and Ecology 318(1): 39–49; https://doi.org/10.1016/j.jembe.2004.12.010
Lee, C.N.-W. & B. Morton (2016). Changes in the distributions of juvenile horseshoe crabs (Arthropoda: Chelicerata) (2002–2014) related to environmental perturbations at Pak Nai and Ha Pak Nai, Deep Bay, Hong Kong SAR, China. Marine Pollution Bulletin 108(1–2): 134-146; https://doi.org/10.1016/j.marpolbul.2016.04.037
Manca, A., F. Mohamad, A. Ahmad, M.F.A.M. Sofa & N. Ismail (2017). Tri-spine Horseshoe Crab, Tachypleus tridentatus (L.) in Sabah, Malaysia: the adult body sizes and population estimate. Journal of Asia-Pacific Biodiversity 10(3): 355–361; http://doi.org/10.1016/j.japb.2017.04.011
Mashar, A., N.A. Butet, B. Juliandi, Y. Qonita, A.A. Hakim & Y. Wardiatno (2017). Biodiversity and distribution of horseshoe crabs in northern coast of Java and southern coast of Madura. IOP
Conference Series: Earth and Environmental Science 54(1): e012076; https://doi.org/10.1088/1755-1315/54/1/012076
Mohamad, F., A. Manca, A. Ahmad, M. Fawwaz, A.M. Sofa, A. Asyraf & I. Noraznawati (2016). Width-weight and length-weight relationships of the Tri-spine Horseshoe Crab, Tachypleus tridentatus (Leach 1819) from two populations in Sabah, Malaysia: implications for population management. Journal of Sustainability Science and Management 11(1): 1–13.
Morton, B. & C.N. Lee (2011). Spatial and temporal distributions of juvenile horseshoe crabs (Arthropoda: Chelicerata) approaching extirpation along the northwestern shoreline of the New Territories of Hong Kong SAR, China. Journal of Natural History 45(3-4): 227–251; https://doi.org/10.1080/00222933.2010.522263
National Ocean Service (2008). NOAA Ocean Service Education. http://oceanservice.noaa.gov/education/kits/tides/media/supp_tide07a.html. Electronic version accessed 10 February 2017.
PAGASA (2017). The Philippine Atmospheric, Geophysical and Astronomical Services Administration. http://www.pagasa.dost.gov.ph. Electronic version accessed 10 March 2017.
Robert, R., S.H.M. Ali & P.F. Amelia-Ng (2014). Demographics of horseshoe crab populations in Kota Kinabalu, Sabah, Malaysia, with emphasis on Carcinoscorpius rotundicauda and some aspects of its mating behaviour. Pertanika Journal of Tropical Agricultural Science 37(3): 375–388.
Rudkin, D.M., G.A. Young & G.S. Nowlan (2008). The oldest horseshoe crab: a new Xiphosurid from late Ordovician Konservat-Lagerstätten deposits, Manitoba, Canada. Palaeontology 51(1): 1-9; https://doi.org/10.1111/j.1475-4983.2007.00746.x
Rudloe, A. (1981). Aspects of the biology of juvenile horseshoe crabs, Limulus polyphemus. Bulletin of Marine Science 31(1): 125–133.
Rudloe, A. (1983). The effect of heavy bleeding on mortality of the horseshoe crab, Limulus polyphemus, in the natural environment. Journal of Invertebrate Pathology 42(2): 167–176; https://doi.org/10.1016/0022-2011(83)90059-9
Schoppe, S. (2002). First data on the distribution of horseshoe crabs (Chelicerata: Merostomata) in Palawan, with notes on their status in the Philippines. UPV Journal of Natural Sciences - UP Visayas 7(1–2): 198–206.
Schreibman, M.P. & C.B. Zarnoch (2009). Aquaculture methods and early growth of juvenile horseshoe crabs (Limulus polyphemus), pp501–511. In: Tanacredi, J.T., M.L. Botton & D. Smith (eds.). Biology and Conservation of Horseshoe Crabs. Springer, Boston, MA, USA, 662pp.
Sekiguchi, K., H. Seshimo & H. Sugita (1988). Post-embryonic development of the horseshoe crab. The Biological Bulletin 174(3): 337–345; https://doi.org/10.2307/1541959
Sekiguchi, K. (ed.) (1988). Biology of Horseshoe Crabs. Science House, Tokyo, Japan, 428pp.
Shuster, C.N., R.B. Barlow & H.J. Brockmann (eds.) (2003). The American Horseshoe Crab. Harvard University Press, Cambridge, 427pp.
Shuster, C.N. & K. Sekiguchi (2009). Basic habitat requirements of the extant species of horseshoe crabs (Limulacea), pp115–129. In: Tanacredi, J.T., M.L. Botton & D. Smith (eds.). Biology and Conservation of Horseshoe Crabs. Springer US, Boston, MA, 662pp.
Sokoloff, A. (1978). Observations on populations of the horseshoe crab Limulus (=Xiphosura) polyphemus. Researches on Population Ecology 19(2): 222–236; https://doi.org/10.1007/bf02518829
Tanacredi, J.T. & S. Portilla (2015). Preliminary investigation of effects of seawater pH on early development of Limulus polyphemus, pp281–287. In: Carmichael, R.H., M.L. Botton, P.K.S. Shin & S.G. Cheung (eds.). Changing Global Perspectives on Horseshoe Crab Biology, Conservation and Management. Springer International Publishing, Cham, 599pp.
Wada, T., T. Mitsushio, S. Inoue, H. Koike & R. Kawabe (2016). Movement patterns and residency of the Critically Endangered horseshoe crab Tachypleus tridentatus in a semi-enclosed bay determined using acoustic telemetry. PLoS ONE 11(2): 1–15; https://doi.org/10.1371/journal.pone.0147429
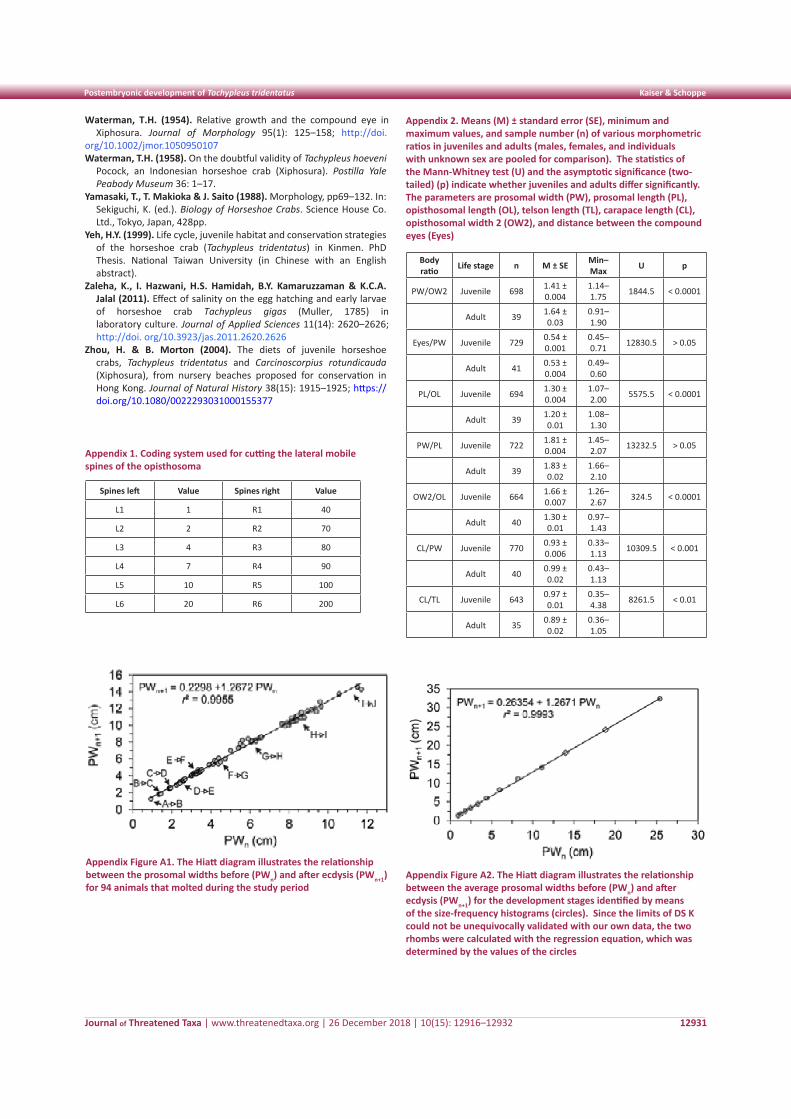
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12931
Appendix 1. Coding system used for cutting the lateral mobile spines of the opisthosoma
Spines left Value Spines right Value
L1 1 R1 40
L2 2 R2 70
L3 4 R3 80
L4 7 R4 90
L5 10 R5 100
L6 20 R6 200
Appendix 2. Means (M) ± standard error (SE), minimum and maximum values, and sample number (n) of various morphometric ratios in juveniles and adults (males, females, and individuals with unknown sex are pooled for comparison). The statistics of the Mann-Whitney test (U) and the asymptotic significance (two-tailed) (p) indicate whether juveniles and adults differ significantly. The parameters are prosomal width (PW), prosomal length (PL), opisthosomal length (OL), telson length (TL), carapace length (CL), opisthosomal width 2 (OW2), and distance between the compound eyes (Eyes)
Body ratio Life stage n M ± SE Min–
Max U p
PW/OW2 Juvenile 698 1.41 ± 0.004
1.14–1.75 1844.5 < 0.0001
Adult 39 1.64 ± 0.03
0.91–1.90
Eyes/PW Juvenile 729 0.54 ± 0.001
0.45–0.71 12830.5 > 0.05
Adult 41 0.53 ± 0.004
0.49–0.60
PL/OL Juvenile 694 1.30 ± 0.004
1.07–2.00 5575.5 < 0.0001
Adult 39 1.20 ± 0.01
1.08–1.30
PW/PL Juvenile 722 1.81 ± 0.004
1.45–2.07 13232.5 > 0.05
Adult 39 1.83 ± 0.02
1.66–2.10
OW2/OL Juvenile 664 1.66 ± 0.007
1.26–2.67 324.5 < 0.0001
Adult 40 1.30 ± 0.01
0.97–1.43
CL/PW Juvenile 770 0.93 ± 0.006
0.33–1.13 10309.5 < 0.001
Adult 40 0.99 ± 0.02
0.43–1.13
CL/TL Juvenile 643 0.97 ± 0.01
0.35–4.38 8261.5 < 0.01
Adult 35 0.89 ± 0.02
0.36–1.05
Appendix Figure A1. The Hiatt diagram illustrates the relationship between the prosomal widths before (PWn) and after ecdysis (PWn+1) for 94 animals that molted during the study period
Appendix Figure A2. The Hiatt diagram illustrates the relationship between the average prosomal widths before (PWn) and after ecdysis (PWn+1) for the development stages identified by means of the size-frequency histograms (circles). Since the limits of DS K could not be unequivocally validated with our own data, the two rhombs were calculated with the regression equation, which was determined by the values of the circles
Waterman, T.H. (1954). Relative growth and the compound eye in Xiphosura. Journal of Morphology 95(1): 125–158; http://doi.
org/10.1002/jmor.1050950107Waterman, T.H. (1958). On the doubtful validity of Tachypleus hoeveni
Pocock, an Indonesian horseshoe crab (Xiphosura). Postilla Yale Peabody Museum 36: 1–17.
Yamasaki, T., T. Makioka & J. Saito (1988). Morphology, pp69–132. In: Sekiguchi, K. (ed.). Biology of Horseshoe Crabs. Science House Co. Ltd., Tokyo, Japan, 428pp.
Yeh, H.Y. (1999). Life cycle, juvenile habitat and conservation strategies of the horseshoe crab (Tachypleus tridentatus) in Kinmen. PhD Thesis. National Taiwan University (in Chinese with an English abstract).
Zaleha, K., I. Hazwani, H.S. Hamidah, B.Y. Kamaruzzaman & K.C.A. Jalal (2011). Effect of salinity on the egg hatching and early larvae of horseshoe crab Tachypleus gigas (Muller, 1785) in laboratory culture. Journal of Applied Sciences 11(14): 2620–2626; http://doi. org/10.3923/jas.2011.2620.2626
Zhou, H. & B. Morton (2004). The diets of juvenile horseshoe crabs, Tachypleus tridentatus and Carcinoscorpius rotundicauda (Xiphosura), from nursery beaches proposed for conservation in Hong Kong. Journal of Natural History 38(15): 1915–1925; https://doi.org/10.1080/0022293031000155377
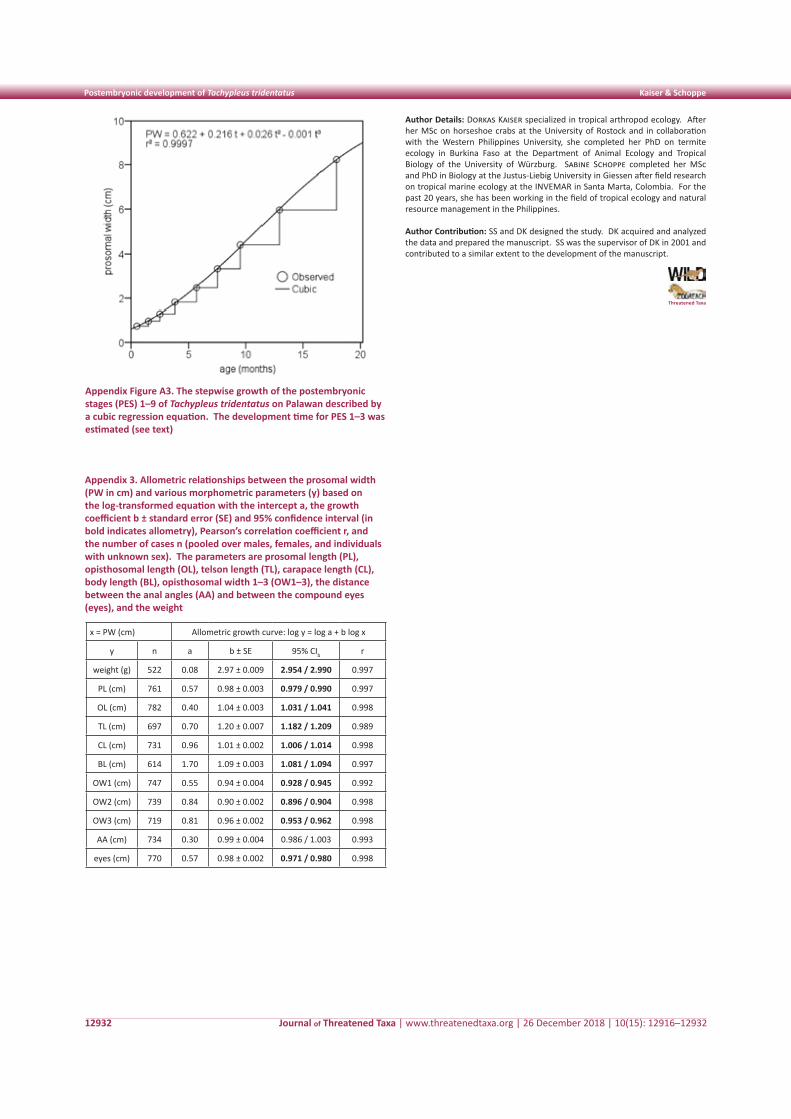
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12916–12932
Postembryonic development of Tachypleus tridentatus Kaiser & Schoppe
12932
Appendix Figure A3. The stepwise growth of the postembryonic stages (PES) 1–9 of Tachypleus tridentatus on Palawan described by a cubic regression equation. The development time for PES 1–3 was estimated (see text)
Threatened Taxa
Author Details: Dorkas Kaiser specialized in tropical arthropod ecology. After her MSc on horseshoe crabs at the University of Rostock and in collaboration with the Western Philippines University, she completed her PhD on termite ecology in Burkina Faso at the Department of Animal Ecology and Tropical Biology of the University of Würzburg. Sabine Schoppe completed her MSc and PhD in Biology at the Justus-Liebig University in Giessen after field research on tropical marine ecology at the INVEMAR in Santa Marta, Colombia. For the past 20 years, she has been working in the field of tropical ecology and natural resource management in the Philippines. Author Contribution: SS and DK designed the study. DK acquired and analyzed the data and prepared the manuscript. SS was the supervisor of DK in 2001 and contributed to a similar extent to the development of the manuscript.
Appendix 3. Allometric relationships between the prosomal width (PW in cm) and various morphometric parameters (y) based on the log-transformed equation with the intercept a, the growth coefficient b ± standard error (SE) and 95% confidence interval (in bold indicates allometry), Pearson’s correlation coefficient r, and the number of cases n (pooled over males, females, and individuals with unknown sex). The parameters are prosomal length (PL), opisthosomal length (OL), telson length (TL), carapace length (CL), body length (BL), opisthosomal width 1–3 (OW1–3), the distance between the anal angles (AA) and between the compound eyes (eyes), and the weight
x = PW (cm) Allometric growth curve: log y = log a + b log x
y n a b ± SE 95% CIb r
weight (g) 522 0.08 2.97 ± 0.009 2.954 / 2.990 0.997
PL (cm) 761 0.57 0.98 ± 0.003 0.979 / 0.990 0.997
OL (cm) 782 0.40 1.04 ± 0.003 1.031 / 1.041 0.998
TL (cm) 697 0.70 1.20 ± 0.007 1.182 / 1.209 0.989
CL (cm) 731 0.96 1.01 ± 0.002 1.006 / 1.014 0.998
BL (cm) 614 1.70 1.09 ± 0.003 1.081 / 1.094 0.997
OW1 (cm) 747 0.55 0.94 ± 0.004 0.928 / 0.945 0.992
OW2 (cm) 739 0.84 0.90 ± 0.002 0.896 / 0.904 0.998
OW3 (cm) 719 0.81 0.96 ± 0.002 0.953 / 0.962 0.998
AA (cm) 734 0.30 0.99 ± 0.004 0.986 / 1.003 0.993
eyes (cm) 770 0.57 0.98 ± 0.002 0.971 / 0.980 0.998

12933
Com
mun
icat
ion
DOI: https://doi.org/10.11609/jott.4218.10.15.12933-12939
Editor: Caroline Leuchtenberger, Instituto Federal de Educação, Farroupilha, Brazil. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 4218 | Received 24 April 2018 | Final received 198 December 2018 | Finally accepted 20 December 2018
Citation: Jorge-Neto, P.N., C.S. Pizzutto, G.R. de Araujo, T. de Deco-Souza, L.C. da Silva, J.A. Salomão Jr. & H. Baldassare (2018). Copulatory behavior of the Jaguar Panthera onca (Mammalia: Carnivora: Felidae). Journal of Threatened Taxa 10(15): 12933–12939; https://doi.org/10.11609/jott.4218.10.15.12933-12939
Copyright: © Jorge-Neto et al. 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001.
Competing interests: The authors declare no competing interests.
For Author Details, Author Contribution and Acknowledgments see end of this article.
Ethics statement: This study has been approved by the Ethics Committee of the School of Veterinary Medicine and Animal Science, University of São Paulo, under protocol number 2072180118.
Copulatory behavior of the Jaguar Panthera onca (Mammalia: Carnivora: Felidae)
Pedro Nacib Jorge-Neto 1 , Cristiane Schilbach Pizzutto 2 , Gediendson Ribeiro de Araujo 3 , Thyara de Deco-Souza 4 , Leanes Cruz da Silva 5 , Jorge Aparecido Salomão Jr. 6 & Hernan Baldassare 7
1,2,6 Department of Animal Reproduction, Faculty of Veterinary Medicine and Animal Science, University of São Paulo (USP) / Av. Prof. Dr. Orlando Marques de Paiva, 87 - Cidade Universitária, São Paulo / SP, 05508-270, Brazil3,4 Faculty of Veterinary Medicine and Animal Science, Federal University of Mato Grosso do Sul (UFMS) / Rua Senador Filinto Muller, 2443 - Vila Ipiranga Campo Grande / MS, 79070-900, Brazil5 Federal University of Viçosa (UFV) / Avenida Peter Henry Rolfs, s/n - Campus Universitário, Viçosa / MG, 36570-900, Brazil7 Department of Animal Science, Faculty of Agricultural and Environmental Sciences, McGill University, 21111 Lakeshore Rd., Ste. Anne de Bellevue, Quebec H9X 3V9, Canada 1 [email protected], 2 [email protected] (corresponding author), 3 [email protected], 4 [email protected], 5 [email protected], 6 [email protected], 7 [email protected]
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12933–12939
Abstract: The relevance of the Jaguar in Brazilian fauna is incompatible with the lack of literature regarding its reproductive behavior, showing that research in this area should be intensified. The knowledge of its basic reproductive behavior is extremely important for understanding the fertility factors of the species and the role it plays in its ecosystem. In this study, we analyzed 210 sequences of sexual behaviors of Jaguars Panthera onca starting from proceptivity of the female and ending with copulation; this sequence is called the copulatory behavior. Behavioral sequences were filmed, and the observed behaviors were analyzed and recorded including occurrence frequency. Different behaviors were observed in association with two types of copulation, it was understood that copulation occurs with and without penile penetration. The information found in the present study is valuable for the reproductive management of Jaguars.
Keywords: Copulation, receptiveness, reproduction, sexual behavior.
Abstract / Resumo: Toda a relevância que onça pintada tem para a fauna brasileira é incompatível com a ausência de literatura sobre seu comportamento reprodutivo, mostrando que precisamos cada vez mais intensificar as pesquisas nesta esfera. O conhecimento dos comportamentos reprodutivos básicos é extremamente importante para o entendimento da fecundidade de uma espécie e todo o papel que ela representa dentro do seu ecossistema. Neste trabalho analisamos 210 sequências de comportamentos sócio-sexuais de onça pintada (Panthera onca) a partir da proceptividade da fêmea até a finalização da cópula; a esta sequência chamados de comportamento copulatório. As sequências comportamentais foram registradas por filmagens e os comportamentos observados foram registrados por frequência de ocorrência. Diferentes comportamentos foram observados em associação com dois tipos de cópulas, o que nos permitiu concluir que a cópula ocorre com e sem penetração peniana. As informações encontradas serão de grande valia para o manejo reprodutivo da onça pintada.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12933–12939
Copulatory behavior of the Jaguar Jorge-Neto et al.
12934
INTRODUCTION
The Jaguar Panthera onca is one of the most important top predators among Brazilian fauna. This animal’s population is decreasing each year due to anthropogenic action, such as fragmentation of its habitat, farming, hunting and slaughter (Cullen et al. 2016; Olsoy et al. 2016; Espinosa et al. 2018). Today, this species is considered as Near Threatened by the IUCN Red List (Quigley et al. 2017) and many research projects involving environmental education, conservation and reproduction with the species have been developed in Brazil, such as NEX No Extinction (Corumbá de Goiás - GO), Projeto Onças do Rio Negro (Aquidauana - MS), Projeto Onçafari (Miranda - MS), Pantera Brasil (Poconé - MT) and Projeto Amigo da Onça (Parque Nacional Boqueirão da Onça - BA), in the attempt to minimize population decrease and negative consequences to Brazilian biodiversity (Tortato et al. 2017; Araujo et al. 2018; Diniz et al. 2018).
Basic information regarding the reproductive behavior of Jaguars is scarce, although this issue is of extreme importance for the development of assisted reproduction and conservation actions. According to Holt et al. (2003) and Owen et al. (2010), reproductive behaviors and subjacent mechanisms associated with reproductive success are particularly important, because fitness is fundamentally a fertility function. For these authors, the study of reproduction is, therefore, crucial for the conservation of species, populations and, indirectly, for the vitality of the entire ecosystem.
Ovulation in this species is usually induced through coitus (Wildt et al. 1979), however, recent studies involving captive females (Gonzalez et al. 2017) reported luteal activity in non-pregnant females that were allocated in individual enclosures, suggesting that spontaneous ovulation occurs occasionally. Interestingly, through hormonal analysis of fecal steroids, Barnes et al. (2016) reported spontaneous ovulations in females housed with males and non-occurrence of spontaneous ovulation in females maintained without the presence of males, demonstrating that the Jaguar is a polyestrous species with induced ovulation.
Stehlik (1971) reported copulatory behavior of the Jaguar at Ostrava Zoo that was briefly described later in 1976 by Lanier and Dewsbury. These authors studied and technically described the copulatory behavior of four Panthera species (P. pardus, P. uncia, P. tigris and P. onca), concluding that behavioral patterns were qualitatively similar among them, with little variation; however, no study with greater description of such
behavior was conducted for the Jaguar. Limited information regarding copulatory behavior
of Jaguars may be justified due to their solitary habits, where interaction with partners is done only during reproductive periods, which makes behavioral studies very difficult with wild animals. On the other hand, the majority of Jaguars kept in zoos and Brazilian rescue centers, present particularities that prevent proper studies with this species, such as animals being too old, castrated, treated with contraceptives or without a reproductive history. One observation of opportunistic courtship behavior, in Pantanal, showed the female’s receptiveness and how the male marks the territory before copulatory behavior (Leuchtenberger et al. 2009).
In light of this lack of information concerning the copulatory behavior of Jaguars and its crucial importance for the development of reproductive strategies and assisted reproduction projects, the objective of this study was to analyze a substantial number of sexual interactions of one adult jaguar couple and thereby describe and characterize the reproductive behavior of the captive jaguar.
MATERIAL AND METHODS
We monitored a couple of captive adult Jaguars in the Peter Crawshaw Rescue Center, in southern Pantanal of Brazil. The male Jaguar was four years old and vasectomized; the female was eight years old at the time of the study. The animals’ enclosure measured 39m in width and 49m in length. Animals were paired during the entire year and were monitored through cameras Intelbras VDH 5040 VF G2, 24 hours.
A total of 210 films recorded four consecutive natural estrus. Sexual behaviors were registered using the continuous focal method (Martin & Bateson 2007). Moreover, for this study, the proceptivity of the female until the effectiveness of copulation was considered. This sequence of behavioral events was entitled copulatory behavior. A large portion of the methodology for behavioral evaluation was adapted from the classification described by Lanier & Dewsbury (1976), and can be found in Table 1.
Statistical AnalysesEach copulatory behavior sequence, which
encompassed the period between female proceptivity and the end of copulation, was considered as one film event. In each film, each behavior (as specified in Table 1) was considered as one registered occurrence.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12933–12939
Copulatory behavior of the Jaguar Jorge-Neto et al.
12935
At the end of each film, a quantitative analysis of occurrence frequency (%) was performed for each behavior presented in relation to the total occurrences in all filmed sequences. Further analysis of copulatory behaviors between event ending with vs. without penile penetration was conducted by contingency analysis and Fisher’s exact test. Differences were considered to be statistically significant at the 95% confidence level (P<0.05).
RESULTS
The duration of each estrus period based on female receptivity were nine, eight, eight and 10 days for the four estrus cycles evaluated and the estrus-to-estrus interval was 34, 39 and 30 days. The visualization and consequent confirmation of penile introduction during copulation occurred in 10 episodes. All behavior sequences observed during female receptivity (210 events) until the end of the male´s pelvic movement behavior were similar. The analysis of the male’s pelvic movement behavior showed two different behavioral sequences, characterized as copulatory behavior with penile penetration (122 events) and copulatory behavior without penile penetration (88 events).
In the sequence of copulatory behavior without penile penetration, the male left the female after finishing his pelvic movements in 42% of the sequences observed. In turn, in the sequence with penile penetration, additional copulatory behaviors were observed from this moment
on, such as the male biting or licking the female’s nape, and female rocking and rolling in lateral-dorsal decubitus in 58% of observations. The frequencies of sexual behavior occurrences that involved both Jaguars’ copulatory behavior sequences can be observed in Figure 1:
- The positioning of the female (squatting) was verified in 100% of our observations;
- Male vocalization was observed in only 1/88 copulatory events without penetration (P<0.001);
- High copulation frequency could be partly explained by the fact that penile penetration occurred only in 42% of the mounts;
- Female vocalization during proceptivity occurred at practically the same frequency preceding copulations with and without penile penetration (55% and 45%, respectively; P>0.05);
- During copulation, female vocalization was more frequent when there was penetration (95.1% vs. 79.6%, P<0.01);
When penile penetration could be confirmed, the copulatory behavior presented the following sequence of events:
- Proceptivity of the female with tail movement and presentation to the male (Figure 2.1)
- The female lies in ventral decubitus, deviating the tail to the side and exposing the anal-genital region to the male; the thoracic limbs of the female are, in general, elongated and the pelvic limbs are flexed next to the body (Figure 2.2)
- The male mounts from the back and on top
Table 1. Behavioral catalog used for the description of the copulatory behavior of the couple of Jaguars Panthera onca at the Peter Crawshaw Rescue Center, in southern Pantanal
Sexual behavior Definition
Pre-copulatory vocalization of the female When the female vocalizes during proceptiveness
Copulatory vocalization of the female When the female vocalizes during the copula
Vocalization of the male When the male vocalizes during the copula
Attractiveness of the male When the male approaches the female and initiates the interaction that may lead to the copula
Proceptiveness of the female When the female requests the male, approaching and turning to him, with presentation of the anal-genital region;
Receptiveness of the female When the female accepts mount from the male
Squatting of the female When the female squats in ventral decubitus, in sexual receptiveness posture
Biting or licking of the male on the females nape When the male licks or bites the females nape during the copula
Rocking of the female When the female, after the copula, turns around and hits the male with one paw
Rolling of the female When the female rolls into lateral dorsal decubitus after the copula
Copula without penial introduction When the pelvic movement of the male occurs during the mount on the female, however, without introduction of the penis
Copula with penial introduction When the pelvic movement of the male occurs during the mount on the female, followed by the introduction of the penis.
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12933–12939
Copulatory behavior of the Jaguar Jorge-Neto et al.
12936
of the female, keeping her between his front paws. He then approximates his genital region to the female’s, squatting with the pelvic limbs
- The female deviates the tail and the male initiates pelvic impulse – the penis is introduced – the male bites/licks the female’s nape three or four times (Figure 2.3) – the female may or may not emit a low growling – the male roars, presumptively indicating ejaculation
- Female rocking – rolling into lateral dorsal decubitus (Figure 2.4).
DISCUSSION
The results found in this study represent the first complete descriptions of the copulatory behavior specific to the Jaguar Panthera onca since the 1970s, which is information of great relevance for studies involving biology and even reproductive biotechnologies. In this context, the understanding of reproductive behaviors and aptitude of any species is critical for the understanding fertility (Owen et al. 2010).
One opportunist observation (Leuchtenberger et al. 2009) and previous studies from Lanier & Dewsbury (1976) and Stehlik (1971) reported rudimentary
information regarding the reproductive behavior of the genus Panthera, but only part of these reports involving copulation in the genus Panthera can be considered specifically for the Jaguar.
By initiating observations from the moment the female shows herself as proceptive to the male, it was clear that the percentage of approach from the male to the female corresponded exactly to the percentage of female proceptivity, thereby indicating that female signalization for possible receptiveness is highly effective in triggering male attractiveness and initiation of courting.
From the moment when the female was receptive to the male and male pelvic movements began, in 42% of the sequences observed, the male finished the copulatory behavior and left the female. This supported the conclusion that this would be a copulatory behavior without penile penetration. In 58% of observations, the male remained in the act of copulation and started biting or licking the female’s nape. In turn, the female started rocking and rolling in lateral-dorsal decubitus. In this situation, we concluded that penile penetration occurred. When penile penetration occurred, the male vocalized in 100% of the events before lightly biting or licking the female’s nape, as described by Hancock (2000) for leopards and by Lanier & Dewsbury (1976)
Figure 1. Frequencies of occurrences of the behavioral events found in the copulatory behavior of the Jaguar Panthera onca.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12933–12939
Copulatory behavior of the Jaguar Jorge-Neto et al.
12937
Figure 2. Schematic diagram of copulatory behavior of the Jaguar Panthera onca: 2.1 (Proceptivity of the female with tail movement and presentation to the male), 2.2 (Proceptivity of the female with tail movement and presentation to the male), 2.3 (The female deviates the tail and the male initiates pelvic impulse – the penis is introduced – the male bites/licks the female’s nape), 2.4 (Female rocking – rolling into lateral dorsal decubitus) – Illustrator: Pedro Busana

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12933–12939
Copulatory behavior of the Jaguar Jorge-Neto et al.
12938
for Jaguars. In contrast, male vocalization was observed in only 1/88 copulatory events without penetration (P<0.001). These findings are consistent with the study by Lanier & Dewsbury (1976), who described that the males’ roar during copulation was verified in every observation of copulation with penile penetration, signalizing success in ejaculation.
It is interesting to point out that the elevated number of copulations in felid species, according to Wildt et al. (2010), has been proposed as a method to induce multiple ovulations among females, and also, in species with high incidence of teratospermia, to ensure deposition of an adequate amount of normal sperm in the vagina, thereby increasing the chance of pregnancy.
According to other comparative in situ and ex situ studies, when compared with others wild felid species such as Cheetahs (Crosier et al. 2009), Clouded Leopards (Wildt et al. 1986), and Oncillas (Swanson & Brown 2004) teratospermia (>60% of defective spermatozoids) is not common in the Jaguar (Morato et al. 2001; Araujo et al. 2018; Gonzales et al. 2017) . Nevertheless, multiple copulations were observed in the present study. High copulation frequency could be partly explained by the fact that penile penetration occurred only in 42% of the mounts, so the male continued attempting to mount until completing ejaculation. Nevertheless, we believe that future studies must be conducted in a format allowing the confirmation of semen deposition in the vagina.
Every positioning of the female, described by Lanier & Dewsbury (1976), such as squatting (elongated anterior limbs and flexed posterior limbs next to the body) was verified in 100% of our observations, both for copulations considered as “with” and “without” penetration. This suggests that the success of penile penetration is not related to female posture, because the female was found in the same position in both situations.
Female vocalization during proceptivity did not influence the result of copulation since it occurred at practically the same frequency preceding copulations with and without penile penetration. During copulation, however, female vocalization was more frequent when there was penetration. Only when copulation involved penile penetration was the female rocking followed by rolling into lateral dorsal decubitus. These findings are consistent with observations by Stehlik (1971) but in disagreement with report by Lanier & Dewsbury (1976), who did not describe female rolling into lateral dorsal decubitus as characteristic behavior after copulation.
CONCLUSIONS
- Basic information regarding the reproductive behavior of Jaguars is of extreme importance for the management, development of assisted reproduction and conservation projects.
- Copulatory behavior of the Jaguar was described in a qualitative manner starting from first signs of female proceptivity and until the finalization of copulation.
- We found that close to half of the copulation events of Panthera onca might occur without penile penetration and, in these cases, ejaculation is believed not to occur.
- Numerous copulations occured during female estrous. Consistent with thoughts reported by others, we believe this behavior may be necessary to promote multiple ovulations and to ensure sufficient number of successful penile penetrations with ejaculation, thereby ensuring proper numbers of normal fertilizing sperm are deposited in the vagina.
REFERENCES
Araujo, G.R., T.A.R. Paula, T. Deco-Souza, R.G. Morato, L.C.F. Bergo, L.C. Silva, D.S. Costa & C. Braud (2018). Comparision of sêmen samples collect from wild and captive Jaguars (Panthera onca) by urethral catheterization after pharmacological induction. Animal Reproduction Science 195: 1–7; https://doi.org/10.1016/j.anireprosci.2017.12.019
Barnes, S.A., J.A. Teare, S. Staaden, L. Metrione & L.M. Penfold (2016). Characterization and manipulations of reproductive cycles in the Jaguar (Panthera onca). General and Comparative Endocrinolology 225: 95-103; https://10.1016/j.ygcen.2015.09.012
Crosier, A.E., J.N. Henghali, J. Howard, B.S. Pukazhenthi, K.A. Terrell, L.L. Marker & D.E. Wildt (2009). Improved quality of cryopreserved Cheetah (Acinonyx jubatus) spermatozoa after centrifugation through accudenz. Jounal of Andrology 30: 298–308; https://10.2164/jandrol.108.006239
Cullen, L. Jr., J.C. Stanton, F. Lima, A. Uezu, M.L.L. Perilli & H.R. Akçakaya (2016). Implications of fine-grained habitat fragmentation and road mortality for Jaguar conservations in the Atlantic forest, Brazil. PLoS One 11(12): e0167372; https://doi.org/10.1371/journal.pone.0167372
Diniz, M.F., R.B. Machado, A.A. Bispo & D. Brito (2018). Identifying key sites for connecting Jaguar populations in the Brazilian Atlantic Forest. Animal Conservation 21: 201-210; https://10.1111/acv.12367
Espinosa, S., G. Celis & L.C. Branch (2018). When roads appear jaguars decline: Increased access to an Amazonian wilderness area reduces potential for Jaguar conservation. PLoS One 13(1): e0189740; https://doi.org/10.1371/journal.pone.0189740
Gonzalez, S.J., J.G. Howard, J. Brown, H. Grajales, J. Pinzón, H. Monsalve, M.A. Moreno & C.J. Escobar (2017). Reproductive analysis of male and female captive Jaguars (Panthera onca) in a Colombian zoological park. Theriogenology 89: 192-200; https://10.1016/j.theriogenology.2016.09.049
Hancock, D. (2000). A time with leopards. Black Eagle Publishing, Cape Town, South Africa.
Holt, W.V., A.R. Pickard, J.C. Rodger & D.E. Wildt (2003). Reproductive Science and Integrated Conservation. Cambridge University Press, Cambridge, United Kingdom, 20pp.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12933–12939
Copulatory behavior of the Jaguar Jorge-Neto et al.
12939
Lanier, D.L. & D.A. Dewsbury (1976). A quantitative study of copulatory behaviour of large felidae. Behavioural Processes 1(4): 327–333; https://doi.org/10.1016/0376-6357(76)90014-0
Leuchtenberger, C., P. Carwshaw, G. Mourão & C. Lehn (2009). Courtship behavior by Jaguars in the Pantanal of Mato Grosso do Sul. Natureza & Conservação 7(1): 218–222.
Martin, P. & P. Bateson (2007). Measuring Behavior: An Introductory Guide. United States of America: Cambridge University Press, 176pp.
Morato, R.G., V.A. Conforti, F.C. Azevedo, A.T. Jacomo, L. Silveira, D. Sana, A.L. Nunes, M.A. Guimaraes & R.C. Barnabe (2001). Comparative analyses of semen and endocrine characteristics of free-living versus captive Jaguars (Panthera onca). Reproduction 122(5): 745–751.
Olsoy, P.J., K.A. Zeller, J.A. Hicke, H.B. Quigley, A.R. Rabinowitz & D.H. Thornton (2016). Quantifying the effects of deforestation and fragmentation on a range-wide conservation plan for Jaguars. Biological Conservation 203: 8–16; https://doi.org/10.1016/j.biocon.2016.08.037
Owen, C., S. Niemann & R. Slotow (2010). Copulatory parameters and reproductive success of wild Leopards in South Africa. Journal of Mammology 91(5): 1178–1187; https://doi.org/10.1644/09-MAMM-A-256.1
Quigley, H., Foster, R., Petracca, L., Payan, E., Salom, R. & Harmsen, B. (2017). Panthera onca (errata version published in 2018). The IUCN Red List of Threatened Species 2017: e.T15953A123791436. Downloaded on 20 December 2018; https://doi.org/10.2305/IUCN.UK.2017-3.RLTS.T15953A50658693.en
Stehlik, J. (1971). Breeding Jaguars at Ostrava Zoo. International Zoo Yearbook 11(1): 116–118; https://doi.org/10.1111/j.1748-1090.1971.tb01871.x
Swanson, W.F. & J.L. Brown (2004). International training programs in reproductive sciences for conservation of Latin American felids. Animal Reproduction Science 82–83: 21–34; https://doi.org/10.1016/j.anireprosci.2004.05.008
Tortato, F.R., T.J. Izzo, R. Hoogesteijn & C.A. Peres (2017). The numbers of the beast: Valuation of Jaguar (Panthera onca) tourism and cattle depradation in the Brazilian Pantanal. Global Ecology and Conservation 11: 106–114; https://doi.org/10.1016/j.gecco.2017.05.003
Wildt, D.E., C.C. Platz, P.K. Chakraborty & S.W. Seager (1979). Oestrous and ovarian activity in a female Jaguar (Panthera onca). Journal of Reproduction and Fertility 56(2): 555–558; https://doi.org/10.1530/jrf.0.0560555
Wildt, D.E., J.G. Howard, L.L. Hall & M. Bush (1986). Reproductive physiology of the clouded leopard: I. Electroejaculates contain high proportions of pleiomorphic spermatozoa throughout the year. Biology Reproduction 34: 937–47; https://doi.org/10.1095/biolreprod34.5.937
Wildt, D.E., W.F. Swanson, J.L. Brown, A. Sliwa, & A. Vargas (2010). Felids ex situ for managed programmes, research, and species recovery, pp. 217–235. In: MacDonald, D.W. & A. Loveridge (eds.). Biology and Conservation of Wild Felids. Oxford: Oxford University Press.
Threatened Taxa
Author Details: Pedro Nacib Jorge Neto, DVM, MBA. Actually master’s degree student (PPGRA-FMVZ / USP) and Technical-Commercial Director of IMV Technologies Brazil. Member of REPROCON research group. Cristiane Schilbach Pizzutto, DVM, MSc, PhD, Postdoc. Professor at FMVZ/USP. Chairman of the Animal Welfare Committee of CRMV - SP. Member of the International Environmental Enrichment Conference Committee and of REPROCON research group. Gediendson Ribeiro de Araujo, DVM, MSc, PhD, Postdoc. Veterinarian at UFMS. Expert in scientific capture of free-living felids. Member of REPROCON research group. Thyara de Deco-Souza, DVM, MSc, PhD. Professor at FAMEZ/UFMS. Expert in morphophysiology of reproduction of wild and domestic animals. Member of REPROCON research group. Leanes Cruz da Silva, DVM, MSc, PhD. Expert in carnivore conservation. Jorge Aparecido Salomão Jr., DVM. Actually master’s degree student (PPGRA-FMVZ / USP). Works with clinic and surgery of wild and exotic animals in private clinics. Member of REPROCON research group. Hernan Baldassarre, DVM, PhD. Professor at McGill University (Canada). Expert in small ruminants reproduction. Global consultant for animal breeding companies. Member of REPROCON research group.
Author Contribution: PNJN, CSP, GRA, TDS and HB were responsible for wrote and review the manuscript; LCS and JASJr were responsible for collect of the data and analyze of the films.
Acknowledgments: We would like to thank CAPES by post-doctoral fellowship to Dr. Gediendson Ribeiro Araújo and master fellowship to Pedro Nacib Jorge Neto; Peter Crawshaw Rescue Center, Pantanal and all its employees for the support and the realization of this study; Pedro Busana for the magnificent drawings.

Amphibians of Dibang River Basin, Arunachal Pradesh Roy et al.
12940
Com
mun
icat
ion
DOI: https://doi.org/10.11609/jott.4249.10.15.12940-12952 | ZooBank: urn:lsid:zoobank.org:pub:0D3499C5-AEDA-4987-A8BF-E5C287F89C09
Editor: S.K. Dutta, Retired Professor of Zoology, Bhubaneswar, India. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 4249 | Received 13 May 2018 | Final received 23 July 2018 | Finally accepted 04 December 2018
Citation: Roy, J.K., R.H. Begum & M.F. Ahmed (2018). Amphibians of the Dibang River Basin, Arunachal Pradesh: an annotated checklist with distribution records. Journal of Threatened Taxa 10(15): 12940–12952; https://doi.org/10.11609/jott.4249.10.15.12940-12952
Copyright: © Roy et al. 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: Aaranyak, Rufford Small Grant Foundation (RSGF), KWEF (Kurita Water and Environment Foundation), and the Science and Engineering Research Board of Department of Science and Technology, Government of India.
Competing interests: The authors declare no competing interests.
Author Details: Jayanta Kumar Roy is a biologist with Herpetological Research and Conservation Division of Aaranyak and has been working on amphibians of northeast India for last seven years. Currently he is focusing on ecology of amphibians of Dibang River Basin in the Eastern Himalaya and completed his PhD research from Assam University, Diphu Campus. He has recorded several rare and threatened amphibian species from the region. Ramie H, Begum is a Biomedical Scientist working in the field of animal disease monitoring and surveillance for more than 14 years. A DBT overseas associate and a visiting professor at University of California, USA, she currently Heads the Department of Life Science and Bioinformatics at Assam University Diphu Campus. M Firoz Ahmed has been involved in conservation research on herpetofauna and tigers in India since 1998. He has been working on herpetofauna and reported new species to science. He has carried out herpetofaunal inventory and taxonomy work in northeast India with specialization in Rhacophorids. He currently heads the Herpetological Research and Conservation Division of Aaranyak.
Author Contribution: JKR - study design, field data collection, analysis and manuscript writing. RHB - manuscript writing and guided JKR. MFA - study design and manuscript writing.
Acknowledgements: The authors are grateful to Aaranyak, Rufford Small Grant Foundation (RSGF), and KWEF (Kurita Water and Environment Foundation) for financial assistance to conduct the study. We sincerely thank Science and Engineering Research Board, the Department of Science and Technology, Government of India for its grant to Aaranyak that helped carry out additional field surveys in 2017. We are thankful to the Department of Environment and Forests, Arunachal Pradesh for providing necessary permission. We thank Prof. S.K. Dutta, Annemarie Ohler and Dr. Saibal Sengupta for help in species identification. We are thankful to Dr. Karthikeyan Vasudevan, Dr. Kanto Nishikawa and Dr. Abhijit Das for their advice and providing key literature. Our thanks to Naba Jibi Pulu for local guidance and logistics help. We thank Gapo Miuli, Rigon Menda, Ahi Mihu, Emkey Tayu, Naba Durga Tayu, Nabaliya Rasup Tayu for their support in the field.
Amphibians of the Dibang River Basin, Arunachal Pradesh: an annotated checklist with distribution records.
Jayanta K. Roy 1 , Ramie H. Begum 2 & M. Firoz Ahmed 3
1,2 Department of Life Science and Bioinformatics, Assam University, Diphu Campus, Karbi Anglong, Assam 782460, India
1,3 Herpetofauna Research and Conservation Division, Aaranyak, 13 Tayab Ali Byelane, Bishnu Rabha Path, Beltola Tinali, Beltola, Guwahati, Assam 781028, India
1 [email protected], 2 [email protected], 3 [email protected] (corresponding author)
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12940–12952
Abstract: The present study across the Dibang River Basin is being presented as an annotated checklist from Arunachal Pradesh. A systematic survey was conducted during 2014–2017 by visual encounter surveys, as well as opportunistic records across the basin. Thirty-eight species of amphibians belonging to 17 genera in six families were recorded. Five new distribution records for Arunachal Pradesh, and one genus Oreolalax was recorded for the first time from India. Further, a rare report on Theloderma moloch and Rhacophorus tuberculatus from northeastern India provided significant information on species microhabitat and updated the amphibian distribution records from Arunachal Pradesh.
Keywords: Five new state records, microhabitat, one new genus record for India, Oreolalax, Rhacophorus tuberculatus, Theloderma moloch.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12940–12952
Amphibians of Dibang River Basin, Arunachal Pradesh Roy et al.
12941
INTRODUCTION
Arunachal Pradesh, a part of the eastern Himalayan Biodiversity Hotspot is still poorly inventoried for flora and fauna. Studies carried out in the state till now have reported 39 amphibians species from Arunachal Pradesh (Sarkar & Ray 2006). Although, the first and most extensive survey on amphibians was carried out by Annandale (1912), where 25 species were documented during the Abor Hill expedition. Subsequently, 22 species from Arunachal Pradesh were reported by Chanda (1994), 28 species from East Kameng (Pakhui Tiger Reserve), Changlang (Namdapha National Park) & Upper Siang (Mouling National Park) District by Pawar & Birand (2001), 50 species from Dihang–Dibang Biosphere Reserve by Borah & Bordoloi (2003), and 35 species from Eaglenest Wildlife Sanctuary by Athreya (2006).
Systematic studies and empirical observations on amphibian species distribution in northeastern region was found to be seriously lacking, although several studies recorded amphibian species from the region (Annandale 1912; Chanda 1994; Pawar & Birand 2001; Bordoloi et al. 2002; Athreya 2006; Sarkar & Ray 2006; Ahmed et al. 2009; Mathew & Sen 2010). In the present study, we presented an updated amphibian species distribution and detailed microhabitat characteristics from Arunachal Pradesh.
MATERIALS AND METHODS
The Dibang River Basin (Fig. 1) is situated in the foothills flanked by the eastern Himalaya. It covers two districts of Arunachal Pradesh: the Lower Dibang Valley and Dibang Valley with a total geographic area of 13,029km2 are situated between 27.99–28.98 0N and 95.78–95.81 0E. The Dibang River flows from the southern flank of the eastern Himalaya and joins the Brahmaputra River in eastern Assam near Tinsukia Town. The entire basin is a mountainous tract and altitude ranges from 200m to 4900m; the annual rainfall varies from 3500–5000 mm (CGWB 2013). The rocky headwater streams with thick canopy cover is the characteristic habitat features of the study area that provide a suitable habitat for rare and range restricted amphibian species (Morse et al. 1993; Meyer & Wallace 2001).
We conducted an extensive survey across the Dibang River Basin covering altitude from 200m to 3500m from 2014–2017. Systematic surveys were conducted along different stream and in forested habitat. The amphibian survey includes two hour (1830–2030 h) visual encounter survey (VES) (Crump & Scott 1994) followed by opportunistic observations during the study period. We monitored amphibian breeding pools located in very remote places in the valley in deep forest during short expeditions of 10–15 days. All amphibian encounters
Figure 1. Map of the study area showing sampling sites for amphibian species distribution across the Dibang River Basin, Arunachal Pradesh, India during 2014–2017.
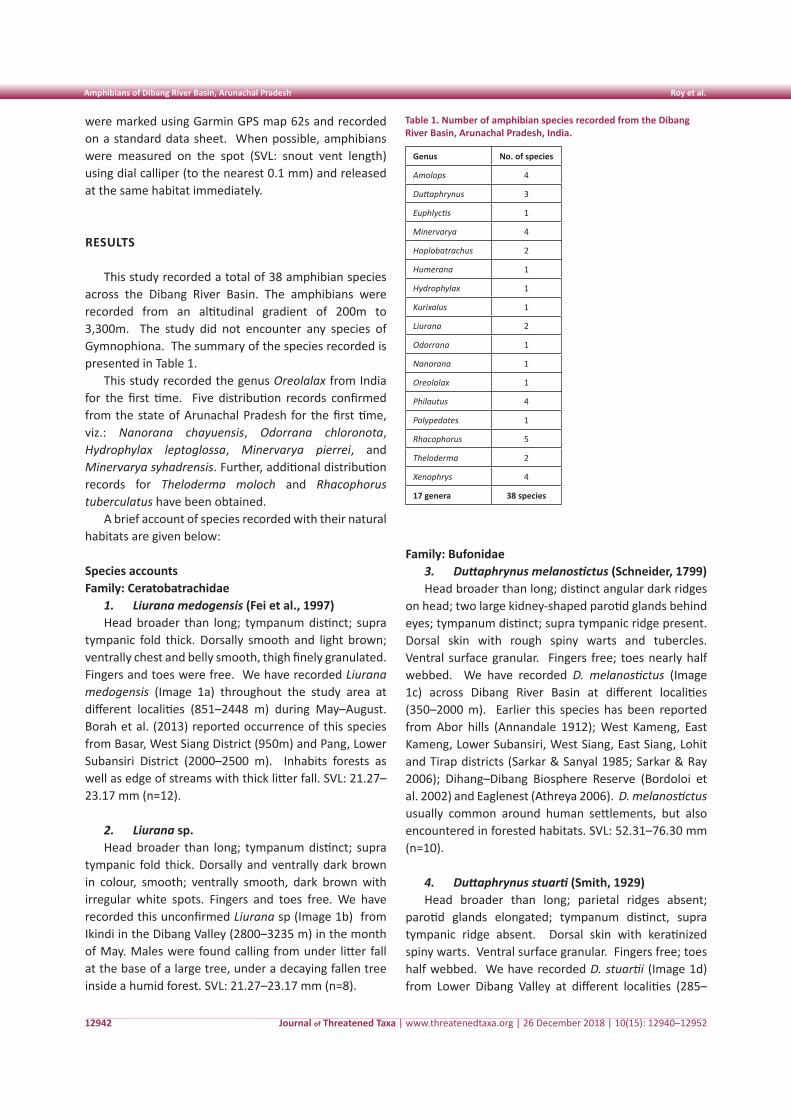
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12940–12952
Amphibians of Dibang River Basin, Arunachal Pradesh Roy et al.
12942
were marked using Garmin GPS map 62s and recorded on a standard data sheet. When possible, amphibians were measured on the spot (SVL: snout vent length) using dial calliper (to the nearest 0.1 mm) and released at the same habitat immediately.
RESULTS
This study recorded a total of 38 amphibian species across the Dibang River Basin. The amphibians were recorded from an altitudinal gradient of 200m to 3,300m. The study did not encounter any species of Gymnophiona. The summary of the species recorded is presented in Table 1.
This study recorded the genus Oreolalax from India for the first time. Five distribution records confirmed from the state of Arunachal Pradesh for the first time, viz.: Nanorana chayuensis, Odorrana chloronota, Hydrophylax leptoglossa, Minervarya pierrei, and Minervarya syhadrensis. Further, additional distribution records for Theloderma moloch and Rhacophorus tuberculatus have been obtained.
A brief account of species recorded with their natural habitats are given below:
Species accounts Family: Ceratobatrachidae
1. Liurana medogensis (Fei et al., 1997)Head broader than long; tympanum distinct; supra
tympanic fold thick. Dorsally smooth and light brown; ventrally chest and belly smooth, thigh finely granulated. Fingers and toes were free. We have recorded Liurana medogensis (Image 1a) throughout the study area at different localities (851–2448 m) during May–August. Borah et al. (2013) reported occurrence of this species from Basar, West Siang District (950m) and Pang, Lower Subansiri District (2000–2500 m). Inhabits forests as well as edge of streams with thick litter fall. SVL: 21.27–23.17 mm (n=12).
2. Liurana sp.Head broader than long; tympanum distinct; supra
tympanic fold thick. Dorsally and ventrally dark brown in colour, smooth; ventrally smooth, dark brown with irregular white spots. Fingers and toes free. We have recorded this unconfirmed Liurana sp (Image 1b) from Ikindi in the Dibang Valley (2800–3235 m) in the month of May. Males were found calling from under litter fall at the base of a large tree, under a decaying fallen tree inside a humid forest. SVL: 21.27–23.17 mm (n=8).
Family: Bufonidae3. Duttaphrynus melanostictus (Schneider, 1799) Head broader than long; distinct angular dark ridges
on head; two large kidney-shaped parotid glands behind eyes; tympanum distinct; supra tympanic ridge present. Dorsal skin with rough spiny warts and tubercles. Ventral surface granular. Fingers free; toes nearly half webbed. We have recorded D. melanostictus (Image 1c) across Dibang River Basin at different localities (350–2000 m). Earlier this species has been reported from Abor hills (Annandale 1912); West Kameng, East Kameng, Lower Subansiri, West Siang, East Siang, Lohit and Tirap districts (Sarkar & Sanyal 1985; Sarkar & Ray 2006); Dihang–Dibang Biosphere Reserve (Bordoloi et al. 2002) and Eaglenest (Athreya 2006). D. melanostictus usually common around human settlements, but also encountered in forested habitats. SVL: 52.31–76.30 mm (n=10).
4. Duttaphrynus stuarti (Smith, 1929)Head broader than long; parietal ridges absent;
parotid glands elongated; tympanum distinct, supra tympanic ridge absent. Dorsal skin with keratinized spiny warts. Ventral surface granular. Fingers free; toes half webbed. We have recorded D. stuartii (Image 1d) from Lower Dibang Valley at different localities (285–
Table 1. Number of amphibian species recorded from the Dibang River Basin, Arunachal Pradesh, India.
Genus No. of species
Amolops 4
Duttaphrynus 3
Euphlyctis 1
Minervarya 4
Hoplobatrachus 2
Humerana 1
Hydrophylax 1
Kurixalus 1
Liurana 2
Odorrana 1
Nanorana 1
Oreolalax 1
Philautus 4
Polypedates 1
Rhacophorus 5
Theloderma 2
Xenophrys 4
17 genera 38 species

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12940–12952
Amphibians of Dibang River Basin, Arunachal Pradesh Roy et al.
12943
1486 m) during April–August. Agarwal & Mistry (2008) recorded this species from Eaglenest Wildlife Sanctuary (1250–2100 m) from western Arunachal Pradesh. They were present along edge of stream, forest trails with litter fall. SVL 36.42–73.13 mm (n=12).
5. Duttaphrynus sp.Head broader than long. parietal ridge absent.
Tympanum distinct; parotid glands wider and elongated. Dorsal skin with smooth warts and large tubercles. Ventral surface granular with large tubercles. Fingers free, toes half webbed. We have recorded this unconfirmed typical Indo-China species of Bufo (Image 1e) from Anini, Angrim Valley and Mipi (1403–1678 m) during February–March. They are early breeders in the area (early February–mid March). Males were observed calling from temporary roadside water pools as well as permanent pool of water after a heavy thundershower. SVL: 81.25–91.12 mm (n=4).
Family: Megophryidae6. Oreolalax sp.Head broader than long. Tympanum not distinct;
supra tympanic fold thick. Dorsally olive to greenish-grey with numerous longitudinal bars. Ventrally smooth, dark brown with two blotch or outgrowths on chest. Fingers and toes free. A single individual of this species was recorded from Ikindi at 3235m in May. This record of Oreolalax sp (Image 1f) from the study area is a new distribution record of the genus to the country. It was observed in the night on an elevated tree trunk covered with mosses in a rhododendron forest. SVL: 39.21mm (n=1).
7. Xenophrys robusta (Boulenger, 1908)Head as broad as long. V–shape marked behind head.
Tympanum distinct; supra tympanic fold thick. Dorsally smooth or finely granulated, reddish brown to dark brown. Ventrally smooth, dark grey in colour. Fingers free; toes rudimentarily webbed. We have recorded X. robusta (Image 1g) species during March–September across the Dibang River Basin (297–1612 m). Previously known from the Dihang–Dibang Biosphere Reserve (Bordoloiet al. 2002) and from Namdapha and Mouling National Park (Pawar & Bindra 2001). They inhabit along the edges of gently flowing streams with large boulders and thick riparian vegetation cover. SVL: 80.79–120.12 mm (n=6).
8. Xenophrys sp1.Head longer than wide. Tympanum distinct and
concealed by supra tympanic fold. Greenish to brown dorsum with benzene ring marked on dorsum. Ventrally smooth; dark grey in colour. Fingers and toes free. We have recorded this unconfirmed Xenophrys sp1 (Image 1h) across the Dibang River Basin (679–2541 m) during March–July. Males were found calling from shrubs and large boulders near stream banks. They were abundant along the edges of narrow stream reaches as well as road side wall with thick and wet vegetation. This was the smallest among all the Xenophrys sp. recorded during this study. SVL: 28.98–34.37 mm (n=5).
9. Xenophrys sp2.Head broader than long. V–shaped preorbital
ridge and a second inverted V–shaped marked on the mid–dorsum. Tympanum distinct; supra tympanic fold present. Dorsally dark brown with fine tubercles, smooth. Ventral surface smooth, grey with black blotches. Fingers free; toes rudimentarily webbed. We have recorded this unconfirmed species of Xenophrys sp. 2 (Image 1i) (947–2079 m) from Anini during May–July. The microhabitat was same as the X. robusta; however distribution in mid elevation. The SVL measured for one individual of the species was 60.34mm.
10. Xenophrys sp3.Head broader than long. V–shaped preorbital
ridge and a second inverted V–shaped marked on the mid–dorsum. Tympanum distinct and concealed by supra tympanic fold. Dorsally reddish brown, smooth. Ventrally smooth, grey with dark orange blotches. Fingers free; toes rudimentarily webbed. We have recorded this unconfirmed species of Xenophrys sp 3 (Image 1j) from high altitude in Ikindi (2184–3060 m) in May. They were observed along the edge of a stream as well as on forest litter fall near a water pool at a high elevation. SVL: 45.2–47.6 mm (n=5).
Family: Dicroglossidae11. Euphlyctis cyanophlyctis (Schneider, 1799)Head slightly broader than long. Tympanum distinct.
Dorsum greyish to brownish, smooth with dark, round spots uniformly distributed; small tubercles and warts present on dorsum. Ventral surface almost white and smooth. Fingers free; toes fully webbed. We have recorded E. cyanophlyctis (Image 1k) across the Dibang River Basin at all the sampling sites (232–1738 m) during the year. It has a wide distribution range in Arunachal Pradesh (Annandale 1912; Chanda 1994; Pawar & Bindra 2001; Bordoloi et al. 2002; Sarkar & Ray 2006). Euphlyctis cyanophlyctis locally common and abundant

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12940–12952
Amphibians of Dibang River Basin, Arunachal Pradesh Roy et al.
12944
Image 1 (a-o): Amphibian species recorded from the Dibang River Basin of Arunachal Pradesh during 2014–17.a) Liurana medogensis; b) Liurana sp; c) Duttaphrynus melanostictus; d) Duttaphrynus stuarti; e) Duttaphrynus sp.; f) Oreolalax sp.; g) Xenophrys robusta; h) Xenophrys sp1; i) Xenophrys sp2; j) Xenophrys sp3; k) Euphlyctis cyanophlyctis; l) Minervarya nepalensis; m) Minervarya pierrei; n) Minervarya syhadrensis; o) Minervarya teraiensis. © Image i - M. Firoz Ahmed; rest of the images - Jayanta K. Roy.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12940–12952
Amphibians of Dibang River Basin, Arunachal Pradesh Roy et al.
12945
in temporary and permanent water bodies, paddy fields, marshy areas in plain grassland. SVL=44.5–66.5 mm (n=10).
12. Minervarya nepalensis (Dubois, 1975) Head longer than wide. Tympanum distinct; supra
tympanic fold narrow. Dorsally greyish-brown with dark irregular spots, narrow mid dorsal line present. Ventrally smooth and grey in colour. Fingers free; toes half webbed. We have recorded Minervarya nepalensis (Image 1l) from lowland areas in the Lower Dibang Valley (224–796 m asl). Bordoloi et al. (2002) recorded this species from the Dihang–Dibang Biosphere Reserve of Arunachal Pradesh without mentioning any specific locality. M. nepalensis is present in different habitats such as agricultural land, temporary or permanent pools and grassland etc. SVL=33–37 mm (n=12).
13. Minervarya pierrei (Dubois, 1975)Head slightly longer than wide. Tympanum distinct;
supra tympanic fold narrow. Dorsum olive to brownish with serrated longitudinal bars; broad mid–dorsal line present on the dorsal side of body. Ventrally smooth, grey in colour. Fingers free; toes half webbed. We have recorded M. pierrei (Image 1m) from lowland Nijamghat areas in the Lower Dibang Valley (232m) in June. It inhabits agricultural land, small temporary water bodies and grassland. Previously not reported from Arunachal Pradesh. SVL=40.20–44.62 mm (n=4).
14. Minervarya syhadrensis (Annandale, 1919)Head slightly broader than length. Tympanum
distinct; supra tympanic fold narrow. Dorsally greyish- to brownish-olive with black irregular spots; very narrow mid–dorsal line present on dorsum. Ventrally smooth, grey to dark grey in colour. Fingers free; toes half webbed. We have recorded M. syhadrensis (Image 1n) from lowland areas of Nizamghat in the Lower Dibang Valley (274–387 m) during April–July. It inhabits small temporary water pools; paddy field; grassland. There is no earlier report of this species from Arunachal Pradesh. SVL=25.10–29.33 mm (n=16).
15. Minervarya teraiensis (Dubois, 1984)Head broader than long. Tympanum distinct; supra
tympanic fold present. Dorsally greyish to brownish with fine granules; narrow mid–dorsal line present or absent. Ventrally smooth, dark grey in colour. Fingers free; toes half webbed. Nine individuals of M. teraiensis (Image 1o) recorded; eight from lowland areas of Nizamghat (232masl) and one individual from Chisindo (795m)
during April–May. Earlier, Bordoloi et al. (2002) reported this species from Dehang Debang Biosphere Reserve of Arunachal Pradesh without any specific locality mention. F. teraiensis was observed in temporary or permanent water bodies, paddy field, shallow pool in forest. SVL=44.6–47.54 mm (n=9).
16. Hoplobatrachus tigerinus (Daudin, 1802) Head longer than broad. Tympanum distinct; supra
tympanic fold very prominent and thick. Dorsally yellowish to olive green, marked with large irregular blackish spots, irregular glandular folds on dorsum; mid–dorsal line present. Ventrally smooth and white. Fingers free; toes fully webbed. We have recorded H. tigerinus (Image 2p) from the lowland areas of Nizamghat in Lower Dibang Valley (215–500 m) in April. Earlier records from Arunachal Pradesh (Annandale 1912; Chanda 1994; Pawar & Bindra 2001; Bordoloi et al. 2002; Sarkar & Ray 2006) suggested a wide distribution of the species in the region. H. tigerinus inhabits paddy fields, marshy grassland and also near large water bodies in floodplain. SVL: 82.30–118.30 mm (n=25).
17. Hoplobatrachus crassus (Jerdon, 1853)Head broader than long. Tympanum distinct; supra
tympanic fold thick. Dorsum greenish or olive, granulate with prominent warts and irregular longitudinal glandular folds; mid-dorsal line absent. Ventrally smooth, grey in colour. Fingers free; toes fully webbed. We have recorded a single individual of H. crassus (Image 2.q) in a temporary roadside pool at New Chidu (297m) in July. Previously reported by Pawar & Bindra (2001) from Pakke Tiger Reserve followed by Borah & Bordoloi (2003) in Chessa and Papumpare (500m). This is also the first record of H. crassus from the Dibang Valley as well as eastern Arunachal Pradesh. It is also the easternmost distribution of the species in the Brahmaputra Valley since it was reported by Saikia et al. (2000). H. crassus inhabits marshy and shrub wetlands, paddy field, temporary or permanent water bodies. SVL=70.81mm (n=1).
18. Nanorana chayuensis (Ye, 1977) Head broader than long. Tympanum distinct; supra
tympanic fold thick. Dorsally olive brown to dark brown with irregular warts on dorsum. Ventrally grey, two oval patches of spine groups with 33 to 56 spines on each patch on male chest during breeding season. Fingers free; toes fully webbed. We have recorded N. chayuensis (Image 2r) across the Dibang River Basin (816–2539 m) during April–July. This report is a first

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12940–12952
Amphibians of Dibang River Basin, Arunachal Pradesh Roy et al.
12946
record of the species from northeast India as well as new distribution record of this species from Arunachal Pradesh. Earlier Deuti & Ayyaswamy (2008) reported this species from Darjeeling District of West Bengal (1860m). N. chayuensis inhabits fast flowing first order streams with slippery boulders and steep bank angle. SVL=62.11–84.11 mm (n=18).
Family: Ranidae 19. Amolops marmoratus (Blyth, 1855)
Head broader than long. Tympanum distinct; supra tympanic fold not distinct. Dorsally brown with olive green to brown gray irregular spots, granulated. Ventrally yellowish-white and granulated. Fingers free; toes fully webbed. We have recorded A. marmoratus (Image 2s) from the Lower Dibang Valley at different locations (273–1294 m) during March–August. Previously recorded from the Upper Renging (655m) (Annandale 1912) followed by Bordoloi et al. (2002) from Namdapha and Boleng, and Dihang–Dibang Biosphere Reserve without any specific locality (500–2000 m). A. marmoratus inhabits perennial streams; rocky streams bed (riffles). SVL 27.10–72.9 mm (n=15).
20. Amolops viridimaculatus (Jiang, 1983)Head slightly longer than broad. Tympanum not
distinct; supra tympanic fold narrow. Dorsally smooth, brown with numerous greenish blotch on dorsum. Ventrally smooth, grey greenish. Fingers free; toes fully webbed. Observed A. viridimaculatus (Image 2t) common across the Dibang River Basin at different localities (679–2538 m) during April–July. Previously reported from the Mouling National Park (Pawar & Bindra 2001) and from the Eaglenest Wildlife Sanctuary (Athreya 2006) at Bompu (2200m) and New Khellong (1250m). A. viridimaculatus inhabits undisturbed rocky perennial streams with thick canopy cover. SVL 65.20–78.20 mm (n=12).
21. Amolops cf. chunganensis (Pope, 1929) Head slightly longer than broad. Tympanum distinct;
supra tympanic fold not clear. Dorsally Olive green to gray brown or reddish brown; dorsolateral line running from posterior eye to vent. Ventrally smooth, pale yellow. Fingers free; toes fully webbed. Recorded A. chunganensis (Image 2u) from Etalin (720 m) during May–July. Athreya (2006) recorded this species as unconfirmed from Sessni (1250m), Eaglenest Wildlife Sanctuary. A. chunganensis inhabits at the edges of large streams with large boulders and thick canopy cover. SVL 29.07–33.80 mm (n=3).
22. Amolops sp.Head broader than long. Tympanum distinct; supra
tympanic fold narrow or indistinct. Dorsally brown with numerous spiny granules serrated on abdominal side; thick dorsolateral line present, Ventrally smooth, grey in colour. A single individual of this unconfirmed species of Amolops (Image 2v) was recorded from Riyali (1468m) in July. It inhabits wet and slippery boulders along the edge of a fast flowing stream. SVL: 51.3mm (n=1).
23. Humerana humeralis (Boulenger, 1887)Head longer as broad. Tympanum distinct; supra
tympanic fold not distinct. Dorsally light brown to bright green; uniformly tuberculated; thick dorsolateral line (Ahmed et al. 2009). Ventrally whitish, smooth. Fingers free; toes two-third webbed. We have recorded H. humeralis (Image 2w) from lowland areas in Nizamghat (232–420 m) during March–July. Previously recorded from the Pakke Tiger Reserve (Hussain et al. 2007). It inhabits paddy field; marshy areas with thick vegetation and tall grasses. SVL: 53.71–63.78 mm (n=4).
24. Hydrophylax leptoglossa (Cope, 1868)Head long as broad. Tympanum distinct; supra
tympanic fold indistinct or absent. Dorsally brown with small to large black spots or markings; dorsolateral line from posterior eye to vent. Ventrally smooth, white spotted brown. Fingers free; toes two–third webbed. We have recorded H. leptoglossa (Image 2x) from the Sally Lake (488m) in June. This species was known to occur at low elevations in Assam and Mizoram, (Chanda 1994; Lalremsanga et al. 2007a; Ahmed et al. 2009; Bortamuli et al. 2010). Recorded two males calling from thick bushes by the lake. Also inhabits slow flowing forest streams and swampy habitats with thick vegetation (Ahmed et al. 2009). SVL: 49.37–50.34 (n=2).
25. Odorrana chloronota (Gunther, 1876)Head as long as broad. Tympanum distinct; supra
tympanic fold not present. Dorsally bright green with 5–6 dark spots. A prominent golden streak present on the upper jaw. Ventrally smooth, dark grey. Fingers free; toes fully webbed. We have recorded O. chloronata (Image 2y) from the lowland areas of Nizamghat (252m) and Etalin (680m) during March–July. This is the first record of O. chloronata from Arunachal Pradesh. O. chloronata has been originally reported from Darjeeling (Gunther 1876) and later from Mizoram (Lalremsanga et al. 2007b); Meghalaya and Assam (Mathew & Sen 2010) with no specific location. The microhabitat recorded for O. chloronata was near small and fast flowing streams

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12940–12952
Amphibians of Dibang River Basin, Arunachal Pradesh Roy et al.
12947
Image 2 (p-ad): Amphibian species recorded from the Dibang River Basin of Arunachal Pradesh during 2014–17. p) Hoplobatrachus tigerinus; q) Hoplobatrachus crassus; r) Nanorana chayuensis; s) Amolops marmoratus; t) Amolops viridimaculatus; u) Amolops cf. chunganensis; v) Amolops sp; w) Humerana humeralis; x) Hydrophylax leptoglossa; y) Odorrana chloronota; z) Kurixalus naso; aa) Polypedates himalayensis; ab) Rhacophorus bipunctatus; ac) Rhacophorus burmanus ad) Rhacophorus maximus. © Images 2s, 2ab - M. Firoz Ahmed; rest of the images - Jayanta K. Roy.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12940–12952
Amphibians of Dibang River Basin, Arunachal Pradesh Roy et al.
12948
with thick canopy cover. SVL: 70.81mm (n=1).
Family: Rhacophoridae26. Kurixalus naso (Annandale, 1912)Head broader than long. Tympanum distinct;
supra tympanic fold present, narrow. Dorsally light to dark brown in colour with prominent pustules and folds including dorsal part of limbs. Ventrally whitish and granular. K. naso (Image 2z) was recorded from Nizamghat (252–1631 m) during April–June. Earlier, Annandale (1912) reported this species from the Egar stream between Renging and Rotung and later from the Mouling National Park (Pawar & Bindra 2001); Doimara in Eaglenest Wildlife Sanctuary (Athreya 2006). K. naso lives near the edge of forest and stream with thick understory vegetation or bushes. SVL: 33.4–35.3 mm (n=8).
27. Polypedates himalayensis (Gray, 1830) Head broader than long. Tympanum distinct; supra
tympanic fold thick. Dorsally smooth, brownish with light darker spots. Ventrally dull whitish and granular. Fingers free; toes three–fourth webbed. P. himalayensis (Image 2aa) was recorded from different localities (297–1486 m) during May–July. Annandale (1912) recorded this species from Arunachal Pradesh as a subspecies of P. maculatus himalayensis from the collection of the Abor Hill Expedition followed by Pawar & Bindra (2001) from the Mouling National Park. Sarkar & Ray (2006) reported it from the West Kameng, East Siang and the Tirap districts without mention of any specific locality. P. himalayensis inhabits near stagnant water bodies temporary or permanent with thick vegetation (herbs and shrubs). Males were found calling from thick grasses and shrubs in shallow pools of water as well as from terrace paddy. SVL: 37.50–44.39 mm (n=10).
28. Rhacophorus bipunctatus (Ahl, 1927)Head broader than long. Tympanum not distinct;
supra tympanic fold narrow. Dorsally light green to green with fine black dots. Ventrally white, granular. Dark characteristic spots on side by arm and groin. Fingers two–third webbed; toes fully webbed. R. bipunctatus (Image 2ab) was recorded from the Maruli (1290m) and Tiwarigaon (1486m) in May and July respectively. Earlier, Annandale (1912) recorded this species from the Rotung and consequently from the Tirap (Sarkar & Sanyal 1985), and the Siang (Chanda 1994; Borah & Bordoloi 2003), without any mention of specific locality; and from the Sessni in Eaglenest Wildlife Sanctuary (Athreya 2006). Males were found calling from branches of shrubs at the
edge of forest as well as roadside water pool and marshy areas. SVL: 40.80–55 mm (n=12).
29. Rhacophorus burmanus (Andersson, 1939)Head slightly longer than broad. Tympanum distinct;
supra tympanic fold very thick. Dorsally green with dark brown blotches; milky white blotches on lateral sides of the body. Ventrally granulated, grey in colour. Fingers one-fourth webbed; toes two-third webbed. R. burmanus (Image 2ac) was recorded from Aropo (1367m) and Maruli (1408m) in May. This is the first record of the species from Arunachal Pradesh which was recently reported from Nagaland and Manipur (Sengupta et al. 2017). Individuals were observed on the ground at the edge of forest and roadside with thick shrubs near a small water pool respectively. SVL: 56.21–52.30 mm (n=2).
30. Rhacophorus maximus (Gunther, 1858) Head slightly broader than long. Tympanum distinct;
supra tympanic fold present, narrow. Dorsally green, smooth. Ventral and lateral sides of body granulated. Fingers two–third webbed; toes fully webbed. R. maximus (Image 2ad) was recorded across the Dibang River Basin at different localities (297–2000 m) during April–July. It has been reported widely from Arunachal Pradesh; the upper Rotung (Annandale 1912); Pakke, Namdapha and the Mouling National Park (Pawar & Birand 2001); Siang, Namdapha, Itanagar and the Dihang-Dibang Biosphere Reserve (Borah & Bordoloi 2003); Lower Subansiri (Sarkar & Ray 2006). During the breeding season R. maximus is generally encountered near stagnant water bodies, temporary or permanent, marshy area and roadside water bodies. SVL: 38.55–93.00 mm (n=16).
31. Rhacophorus translineatus (Wu, 1977)Head longer than broad. Tympanum distinct; supra
tympanic fold present, thick. Dorsally reddish-brown to light brown in color; very fine granules on dorsum with narrow 9–12 transverse dark brown line from snout to vent. Ventrally whitish with thin network markings. Fingers two–third webbed; toes fully webbed. R. translineatus (Image 3ae) was recorded from Tiwarigaon (1480m) and the Ahini Ango (920m) in July and August respectively. Previously, it was reported from the Dihang–Dibang Biosphere Reserve (Bordoloi et al. 2002) without any specific locality or voucher specimens, and from Eaglenest Wildlife Sanctuary (Athreya 2006). R. translineatus inhabits marshy pools under thick canopy and males were found calling from nearby vegetation, bushes and trees. SVL: 58.25–59.68 mm (n=4).

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12940–12952
Amphibians of Dibang River Basin, Arunachal Pradesh Roy et al.
12949
32. Rhacophorus tuberculatus (Anderson, 1871) Head slightly longer than broad. Tympanum distinct;
supra tympanic fold present, narrow. Dorsally deep brown with numerous fine black spots on dorsum. Ventrally whitish, granular mixed with large tubercles. R. tuberculatus (Image 3af) was recorded from Nizamghat (395m) in July. R. tuberculatus has been reported from Janakmukh (183m), Rotung (396m) and Kalek (1158m) during the Abor hill expedition (Annandale 1912). We observed a single R. tuberculatus sitting on a branch along the edge of a narrow stream covered by thick bushes. SVL: 41.8 mm (n=1).
33. Theloderma asperum (Boulenger, 1886)Head broader than long. Tympanum distinct; supra
tympanic fold indistinct. Dorsally dark gray to brown with small to large spinules. Ventrally smooth, dark. Fingers free; toes half webbed. T. asperum (Image 3ag) was recorded from New Chidu (344m) and Elopa (851m) in May. Annandale (1912) recorded this species from the Egar stream between the Renging and the Rotung in East Siang District; subsequently, from Namdapha and the Mouling National Park (Pawar & Birand 2001). Males were found calling from water accumulated in a tree trunk hole inside a tropical humid forest. Also found in an artificial water tank near agricultural land close to the forest. SVL: 28.9–30.8 mm (n=4).
34. Theloderma moloch (Annandale, 1912) Head broader than long. Tympanum distinct; supra
tympanic fold indistinct. Dorsally greyish-brown with prominent ridge more or less serrated warts on dorsum. Ventrally black, granulated, tubercles around vent. Fingers free; toes three–fourth webbed. T. moloch (Image 3ah) was recorded from Elopa (780m) and Chisindo (910m) in May and July, respectively (Roy et al. 2017). Annandale (1912) originally described this species from the Upper Rottung, East Siang District and later it was reported from the Eaglenest Wildlife Sanctuary (Athreya 2006). It is also reported from Namdapha by Biju et al. (2016). The microhabitat and other natural history notes are reported by Roy et al. (2017). SVL 36.10–39.46 mm (n=4).
35. Philautus sp1Head broader than long. Both tympanum and supra
tympanic fold indistinct. Dorsally smooth, reddish-brown. Ventrally granular, whitish. Fingers free; toes one–third webbed. Recorded this unconfirmed Philautus sp.(Philautus sp1, Image 3ai) from moist forest areas of Nizamghat, New Chidu and Ejengo (232–947 m)
during March–July. Inhabits along the edge of forest and males were observed calling from thick bushes under a moderate canopy cover. Also observed near human habitations. SVL 19.5–20.2 mm (n=3).
36. Philautus sp2Head broader than long. Tympanum fairly distinct;
supra tympanic fold present, narrow. Dorsally smooth, grey brown; 2–3 dark brown spots on flanks near to ventrum. Ventrally granular, grey in colour. Fingers free; toes one–third webbed. Recorded this unconfirmed species of Philautus (Philautus sp 2., Image 3aj) from the Nizamghat, Elopa, Chisindo, Tiwarigaon, Challis (40 Kilo), Etalin and Riyali (329–2071 m) during March–July. Inhabits along the edge of streams with thick riparian vegetation cover. SVL: 22.81–25.10 mm (n=6).
37. Philautus sp3Head broader than long. Tympanum fairly distinct;
supra tympanic fold present; narrow. Dorsally brown, slightly granular with irregular dark bands on dorsum. Ventrally granular, grey with irregular dark spots. Fingers free; toes one–third webbed. Recorded this unconfirmed species of Philautus (Philautus sp3, Image 3ak) from nearby Tiwarigaon (1545m) in July. A single male individual of this species was observed calling from shrubs by a roadside wall with thick moist vegetation grown on it. SVL: 17.59 mm (n=1).
38. Philautus sp4Head broader than long. Tympanum fairly distinct;
supra tympanic fold present, thick. Dorsally brown with irregular fine granules. Ventrally granular, grey mixed with brown spots. Fingers free; toes one–third webbed. Recorded this unconfirmed species of Philautus (Philautus sp4, Image 3al) from the Etalin (752m) in May. A single male individual of this species was observed calling from a stem of Alocasia sp. approximately three feet above ground along the edge of a stream. SVL: 21.20mm (n=1).
DISCUSSION
This study presented the distribution record of 38 amphibian species from across the Dibang River Basin for the first time from an altitudinal gradient of 224m to 3,235m of a Himalayan river. Previously, Annandale (1912) and Bordoloi et al. (2002) reported amphibian diversity from Abor Hills (Siang River Basin) and the Dihang–Dibang Biosphere Reserve (Siang and Dibang

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12940–12952
Amphibians of Dibang River Basin, Arunachal Pradesh Roy et al.
12950
Figure 2. IUCN red list status for 38 amphibian species recorded across the Dibang River Basin, Arunachal Pradesh, India during 2014–17.
Image 3 (ae-al): Amphibian species recorded from the Dibang River Basin of Arunachal Pradesh during 2014–17.ae) Rhacophorus translineatus; af) Rhacophorus tuberculatus; ag) Theloderma asperum; ah) Theloderma moloch; ai) Philautus sp1; aj) Philautus sp2; ak) Philautus sp3; al) Philautus sp4. © Jayanta K. Roy.
river basins), respectively. The species described as new to science by Annandale (1912) has been encountered during this study for the first time since it was described. This study further confirms some record of species previously described by other authors (Bordoloi et al. 2002; Sarkar & Ray 2006; Mathew & Sen 2010; Borah et al. 2013) and at the same time we could record altitude, microhabitats and geolocate the occurrences from the river basin.
As the inventories of amphibians are very few in the state, this study reports five new distribution records of amphibians for Arunachal Pradesh. In addition, one species is recorded for the first time from India (Orelalax sp), however, species level identity of the species is yet to be confirmed and is in progress. In this study, we have comparatively described the present species distribution with regards to their previous known distribution records from Arunachal Pradesh, their microhabitat characteristics and specific morphological characteristics.
Biogeographically, when compared to biogeographic
regions (Duellman 1999) we found that the amphibian (unconfirmed species excluded) distribution from the study area shows an overlapping and sharing of species from India/Sri Lanka (42.9%), southern Himalaya (85.7%), northeastern montane (78.6%), and China (32.1%).
The conservation status (IUCN 2017) of the amphibians encountered (Fig. 2) include, two Near

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12940–12952
Amphibians of Dibang River Basin, Arunachal Pradesh Roy et al.
12951
Threatened (NT) 5% anurans: Amolops viridimaculatus and Rhacophorus burmanus; one vulnerable (VU) 2.5%: Theloderma moloch; Least Concern (LC) 44.5% (N=17); Data deficient (DD) 16% (N=6) and Not Evaluated (NE) 31.5% (N=12). It is interesting to note that nearly 50% of the 38 species are data deficient and not evaluated yet. This study would further help in assessing the conservation status of those species that needs evaluation and reevaluation. This study also observed that unscientific developments including many large hydroelectric projects at various stages of implementations as well as various roads planned within the study area are most likely to have irreversible effects on the ecology in the river basin during the next few decades.
The river basin approach of this study has helped in planning long term ecological study on amphibians including patterns of distributions along an altitudinal gradient from the plains of Assam to the high Himalayas. Long term ecological studies in the river basin would help in understanding factors that influence and limit distribution of species to understand the possible effect of weather change on species that are restricted by the Himalayan mountain ranges and mid elevation foothills. Amphibians, being indicators of the health of the environment, could help understand the impact of changing weather on ecosystems if monitored in the river basin.
REFERENCES
Ahmed, M.F., A. Das, & S.K. Dutta (2009). Amphibians and Reptiles of Northeast India. A Photographic Guide. Aaranyak, Guwahati, India 169 pp.
Agarwal, I. & V.K. Mistry (2008). Bufo stuarti from western Arunachal Pradesh, India. Russian Journal of Herpetology 15: 166–168.
Ahl, E. (1927). Zur Systematik der asiatischen Arten der Froschgattung Rhacophorus. Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin 1927: 35–47.
Anderson, J. (1871). A list of the reptilian accession to the Indian Museum, Calcutta from 1865 to 1870, with a description of some new species. Journal of the Asiatic Society of Bengal 40: 12–39.
Andersson, L.G. (1939). Batrachians from Burma collected by Dr. R. Malaise, and from Bolivia and Ecuador collected by Dr. C. Hammarlund. Arkiv for Zoologi 30 (23): 1–24.
Annandale, N. (1912). Zoological Results of the Abor Expedition, 1911–1912. Record Indian Museum 8 (1): 7–59; https://doi.org/10.5962/bhl.part.1186
Annandale, N. (1919). The fauna of certain small streams in the Bombay Presidency: Some frogs from streams in the Bombay Presidency. Records of the Indian Museum 16: 109–161; https://doi.org/10.5962/bhl.part.25917
Athreya, R. (2006). Eaglenest Biodiversity Project (2003–2006): Conservation Resources for Eaglenest Wildlife Sanctuary, Kaati Trust, Pune, web–version. url: http://www.iiserpune.ac.in/~rathreya/Downloads/athreya2006_ebp.pdf. Accessed on 05 December 2015.
Biju, S.D., G. Senevirathne, S. Garg, S. Mahony, R.G. Kamei, A. Thomas,
Y. Shouche, C.J. Raxworthy, M. Meegaskumbura & I. van–Bocxlaer (2016). Frankixalus, a new Rhacophorid genus of tree hole breeding frogs with oophagous tadpoles. PLoS ONE 11(1), e0145727; https://dx.doi.org/10.1371/journal.pone.0145727
Blyth, E. (1855). Report of the Curator; Zoological Department, for March meeting. Journal of the Asiatic Society of Bengal 24: 187–188.
Borah, M.M. & S. Bordoloi (2003). Altitudinal distribution pattern of Amphibian fauna of Arunachal Pradesh with special reference to Dehang Debang Biosphere Reserve, Arunachal Pradesh, India, Himalayan Biosphere Reserve Bulletin 5 (1–2), 51–55.
Borah, M.M., S. Bordoloi, J. Purkayastha, M. Das, A. Dubois & A. Ohler (2013). Limnonectes (Taylorana) medogensis (Fei, Ye & Huang, 1997) from Arunachal Pradesh (India), and on the identity of some diminutive ranoid frogs (Anura: Dicroglossidae, Occidozygidae). Herpetozoa 26: 39–48.
Bordoloi, S., M.M. Borah, P.K. Sarmah & J. Sharma. (2002). Amphibian and insect fauna of amphibian habitats of Dehang–Debang Biosphere Reserve, Arunachal Pradesh. Himalayan Biosphere Reserves Bulletin 4: 33–38.
Bortamuli, T., S. Bordoloi, A. Ohler & S. Grosjean (2010). External morphology, Buccopharyngeal anatomy and development rate of the tadpoles of two Asian Ranidae (Amphibia: Anura), Hylarana humeralis (Boulenger, 1887) and Hylarana leptoglossa (Cope, 1868). Journal of Natural History 44(7–8): 421–445; https://doi.org/10.1080/00222930903359651
Boulenger, G.A. (1886). First report on additions to the batrachian collection in the Natural–History Museum. Proceedings of the Zoological Society of London 1886: 411–416.
Boulenger, G.A. (1887). An account of the batrachians obtained in Burma by M.L. Fea of the Genoa Civic Museum. Annali del Museo Civico di Storia Naturale di Genova. Serie 2, 5: 418–424.
Boulenger, G.A. (1908). A revision of the Oriental pelobatid batrachians (genus Megalophrys). Proceedings of the Zoological Society of London 1908: 407–430.
CGWB (Central Ground Water Board 2013). North Eastern region, Guwahati: http://www.cgwb.gov.in/Regions/NER/stateprofiles.html.
Chanda, S.K. (1994). Anuran (Amphibia) fauna of northeast India. Zoological Survey of India 18 (2), 1–143.
Cope, E.D. (1868). An examination of the Reptilia and Batrachia obtained by the Orton Expedition to Equador and the Upper Amazon, with notes on other species. Proceedings of the Academy of Natural Sciences of Philadelphia 20: 96–140.
Daudin, F.M. (1802). “An. XI”. Histoire Naturelle des Rainettes, des Grenouilles et des Crapauds. Quarto version. Paris: Levrault.
Deuti, K., & A.K. Ayyaswamy (2008). Geographic distribution: Nanorana chayuensis. Herpetological Review 39: 234.
Dubois, A. (1975). Une nouveau complexe d’espèces jumelles distinguées par le chant: les grenouilles de Nepal voisines de Rana limnocharis Boie (Amphibienes, Anoures). Comptes Rendus Hebdomadaires des Seances de l’Academie des Sciences. 281: 1717–1720.
Dubois, A. (1984). Note preliminaire sur le groupe de Rana limnocharis Gravenhorst, 1829 (Amphibiens, Anoures). Alytes 3: 143–159.
Duellman, W.E. (1999). Distribution patterns of amphibians in South America. Patterns of distribution of amphibians: a global perspective (ed. by W.E. Duellman), pp. 445–471. The John Hopkins University Press, Baltimore, MD.
Fei, L., C.Y. Ye & Y.Z. Huang (1997). Taxonomic studies of the genus Liurana of China including descriptions of a new species (Amphibia:Ranidae). Herpetologica 6–7: 75–80.
Gray, J.E. (1830). Illustrations of Indian Zoology; Chiefly Selected from the Collection of Major–General Hardwicke, F.R.S. Volume 1. Part 3. London: Treuttel, Wurtz, Treuttel, Jun. and Richter; https://doi.org/10.5962/bhl.title.95127
Günther, A.C.L.G. (1858). Neue Batrachier in der Sammlung des britischen Museums. Archiv fur Naturgeschichte. 24: 319–328.
Günther, A.C.L.G. (1876). Third report on collections of Indian reptiles obtained by the British Museum. Proceedings of the Zoological

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12940–12952
Amphibians of Dibang River Basin, Arunachal Pradesh Roy et al.
12952
Society of London 567–577.Hussain, B., J. Gogoi, N.K. Choudhury, B.K. Baruah, D.K. Sharma, J.
Kalita & S. Sengupta (2007). Rana humeralis—a new record from Assam and Arunachal Pradesh, India, with notes on its morphometry and natural history. Russian Journal of Herpetology 14: 167–171.
IUCN (2017). The IUCN Red List of Threatened Species. Version 2017-3. Downloaded on 05 December 2017.
Jerdon, T.C. (1853). Catalogue of reptiles inhabiting the Peninsula of India. Journal of the Asiatic Society of Bengal 22: 522–534.
Jiang, Y.M. (1983). A new species of the genus Staurois (Ranidae), Staurois viridimaculatus. Acta Herpetologica Sinica/Liangqi baxing dongwu yanjiu. New Series. Chengdu 2(3): 71.
Lalremsanga, H.T., S. Sailo, B. Kharbuli & R.N.K. Hooroo (2007a). Record of Sylvirana leptoglossa (Cope, 1868) (Anura: Ranidae) from Kolasib district, Mizoram, Northeastern India. Frog Leg 13: 9–10.
Lalremsanga, H.T., S. Sailo & R.N.K. Hooroo (2007b). Record of Rana chloronota (Gunther, 1875) (Anura: Ranidae) from Mizoram, northeastern India. Hamadryad 31: 359.
Mathew, R. & N. Sen (2010). Pictorial Guide to Amphibians of North East India. Zoological Survey of India, Kolkata, India: 1–144.
Meyer, J.L. & J.B. Wallace (2001). Lost linkages and lotic ecology: Rediscovering small streams. Blackwell Scientific, Oxford (United Kingdom).
Morse, J.C, B.P. Stark & W.P. McCafferty (1993). Southern Appalachian streams at risk: implications for mayflies, stoneflies, caddisflies, and other aquatic biota. Aquatic Conservation–Marine and Freshwater Ecosystems 3: 293–303; https://doi.org/10.1002/aqc.3270030404
Pawar, S. & A. Birand (2001). A survey of amphibians, reptiles and birds in northeast India. CERC Technical Report–6; Center for Ecological Research and Conservation, Mysore.
Roy, J.K., R.H. Begum & M.F. Ahmed (2017). Report on poorly known Theloderma moloch (Annandale, 1912) from Arunachal Pradesh, Northeast India. Russian Journal of Herpetology 24(3): 248–250.
Saikia, S., N.K. Choudhury, B. Hussain & S. Sengupta (2000). Size analysis and distribution of Jerdon’s Bull Frog Hoplobatrachus crassus (Jerdon, 1853) in Assam. Journal of the Bombay Natural History Society 97: 286–288.
Sarkar, A.K. & D.P. Sanyal. (1985). Amphibia. Records of the Zoological Survey of India 81: 285–295.
Sarkar, A.K. & S. Ray (2006). Amphibia. Alfred, J. R. B. ed., Fauna of Arunachal Pradesh. Zoological Survey of India 13(Part 1): 1–196.
Schneider, J.G. (1799). Historia Amphibiorum Naturalis et Literarariae. Fasciculus Primus. Continens Ranas, Calamitas, Bufones, Salamandras et Hydros in Genera et Species Descriptos Notisque suis Distinctos. Jena: Friederici Frommanni.
Sengupta, S., Das. A. & Ahmed, F. (2017). Distribution of Rhacophorus burmanus (Anderson 1939) with notes on its natural history. Herpetozoa 29 (3/4).
Wu In Sichuan Institute of Biology Herpetology Department. (1977). A survey of amphibians in Xizang (Tibet). Acta Zoologica 23: 54–63.
Smith, M.A. (1929). On a collection of amphibians and reptiles from the upper reaches of the Brahmaputra. Records of the Indian Museum 31: 77–80.
Threatened Taxa

12953
Com
mun
icat
ion
DOI: https://doi.org/10.11609/jott.3891.10.15.12953-12968 | ZooBank: urn:lsid:zoobank.org:pub:05A3C45B-0203-4FCA-A791-C9F909DFE18C
Editor: R.K. Avasthi (Retd. Associate Professor/Head), Vaish College, Rohtak, India. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 3891 | Received 14 November 2017 | Final received 14 September 2018 | Finally accepted 20 October 2018
Citation: Khan, M.I., M.K. Usmani, S. Usmani & H. Naz (2018). Taxonomic studies on the gaudy grasshoppers (Orthoptera: Pyrgomorphoidea: Pyrgomorphidae) from the northeastern states of India. Journal of Threatened Taxa 10(15): 12953–12968; https://doi.org/10.11609/jott.3891.10.15.12953-12968
Copyright: © Khan et al. 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: The University Grants Commission, New Delhi, for providing financial assistance during the tenure of a major research project (Ref. no. 33-33/2007 (SR)).
Competing interests: The authors declare no competing interests.
Author Details: MIK has completed PhD in Zoology, interested in taxonomy, biodiversity and ecology of insects. He has successfully completed a research project funded by UGC entitled with “Studies on the taxonomy and diversity of Acridoidea (Orthoptera) in North-Eastern States of India” (1st April, 2008 – 31st March, 2011). MKU is a Professor, Section of Entomology, research interest lies in the taxonomy, biology, ecology and biodiversity of insect pests of agricultural importance and in their biological control through the use of insect natural enemies. SU has completed PhD in Zoology, interested in taxonomy, biodiversity and ecology of insects. HN is a PhD student, interested in taxonomy, biodiversity and ecology of insects.
Author Contribution: All authors contributed equally.
Acknowledgements: We wish to extend our gratitude to the University Grants Commission, New Delhi for providing financial assistance during the tenure of a major research project (Ref. no. 33-33/2007 (SR)) being carried out on “Studies on taxonomy and diversity of northeastern States of India”.
Taxonomic studies on the gaudy grasshoppers (Orthoptera: Pyrgomorphoidea: Pyrgomorphidae) from the northeastern states of India
M. Imran Khan 1 , M. Kamil Usmani 2 , Shahnila Usmani 3 & Hira Naz 4
1,2,3,4 Section of Entomology, Department of Zoology, Aligarh Muslim University, Aligarh, Uttar Pradesh 202002, India 1 [email protected] (corresponding author), 2 [email protected], 3 [email protected], 4 [email protected]
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12953–12968
Abstract: A survey of the northeastern states of India recorded 10 species representing five genera belonging to four tribes of the family Pyrgomorphidae. For identification, in addition to conventional morphological characters, the detailed structures of male and female genitalia were also included. All the genera studied are described. Morphometry and distribution of each species are also given.
Keywords: Caelifera, Pyrgomorphidae, tribes, species, key.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12953–12968
Taxonomic studies on gaudy grasshoppers Khan et al.
12954
INTRODUCTION
Pyrgomorphidae is a family of the order Orthoptera under the suborder Caelifera. Its members are commonly known as Gaudy Grasshoppers in the superfamily Pyrgomorphoidea. Some members (tribes Atractomorphini and Psednurini) are green in colour while others (tribe Monistrini) have bright warning colours. The present study was based on conventional morphological characters along with a detailed study of genital structures of Pyrgomorphidae for a better understanding of the significance of morphological structures. A comparative study was done on the genitalia with reference to supra-anal plate and cerci, subgenital plate, epiphallus, and aedeagus of the male and the supra-anal plate, subgenital plate, ovipositor, and spermatheca of the female.
In the present study, Pyrgomorphidae is recognized as a family of superfamily Pyrgomorphoidea. Kirby (1910, 1914) prepared a catalogue for Acrididae (including this group) of the world and a volume of Fauna of British India including the fauna of Pakistan, Bangladesh, Sri Lanka, and Burma. Later, Chopard (1924a), Uvarov (1925, 1929), and Ayyar (1940) also worked on the Indian species of Pyrgomorphidae.
Kevan et al. (1970) worked on the taxonomy of the oriental taxa of Pyrgopmorphidae. Bhowmik (1964), Tandon & Shishodia (1969, 1989), Tandon (1976), Bhowmik & Halder (1983), and Usmani & Shafee (1985) worked on the taxonomy of various genera and species of Pyrgomorphinae from different parts of India.
The system of classifying grasshoppers by earlier workers was mainly based on easily recognizable externally visible characters. Slifer & King (1936), Slifer (1939, 1940, 1943), Dirsh (1957a,b), and Meinodas et al. (1982) showed the taxonomic significance of spermatheca in Acrididae.
Dirsh & Uvarov (1953) studied the apical valve of aedeagus in three species of Anacridium. Dirsh (1956a,b) showed the importance of aedeagus in classifying and grouping various families of Acridoidea.
Herrera & Schnidrig (1983) described the male genitalia of 64 species of Orthoptera from Navarra. Usmani & Ajaili (1993) showed the taxonomic significance of aedeagus in some Libyan species of Acridoidea. Ajaili & Usmani (1994) made comparative studies on the male subgenital and supra-anal plates and cerci in some Libyan species of Acridoidea.
Kumar et al. (2014) conducted a comprehensive study on the male supra-anal plate, cerci, and subgenital plate in 12 species of grasshoppers representing six genera under
four tribes belonging to the family Pyrgomorphidae.
MATERIAL AND METHODS
A survey for collection of Pyrgomorphid specimens during 2008–2011 from grasslands and agricultural fields of northeastern states was carried out. Specimens were handpicked or collected using sweeping nets. All the collected specimens were preserved in 70% ethyl alcohol. Specimens were stretched and photographed. For genitalic studies, the apical tip of the abdomen was cut and boiled in 10% KOH solution and the genital structures were isolated. All drawings were prepared under camera lucida (Nikon SMZ 25). Descriptions of male genitalia follow the terminologies used in Dirsh (1956a). All observations on morphometry are given in millimetres.
Family: Pyrgomorphidae Brunner von Wattenwyl, 1874Brunner von Wattenwyl. 1874. Ver. Der. Zool. Ges.
24:22.Diagnosis: Body of variable shapes, head acutely
conical, fastigial furrow present. Prosternal process present. Elytra and wings fully developed, reduced, or absent. Tympanam normally present. The lower basal lobe of hind femur longer than the upper one. Brunner’s organ present except in a few genera, with thin, almost cursorial hind legs. External apical spine of hind tibia present or absent. Ectophallus differentiated; cingulum capsule-like; valves of penis paired, undivided; spermatophore sac in dorsal position. Epiphallus bridge-shaped, with dorso-lateral appendices; ancore absent; lophi hook-like. Oval sclerites absent. No stridulatory mechanism known. The Pyrgomorphidae is a very well defined family, with a peculiar phallic complex which is rather uniform through the family. The relationship with other families is rather obscure and no close affinities exist. They have some common features with Lentulidae, such as undivided paired valves of the penis and the dorsal position of the spermatophore sac, and others with Ommexechidae, such as the presence of a fastigial forrow and the paired undivided valves of the penis. All other characters, however, are so distinct that the relationship is a very remote one. Pyrgomorphidae is represented in all the tropical and subtropical parts of the world by a large number of genera, a list of which appears unnecessary.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12953–12968
Taxonomic studies on gaudy grasshoppers Khan et al.
12955
Key to species of Atractomorpha Saussure, 1862
1. Eyes comparatively long, elongate oval ................................................................................................................................. 2- Eyes comparatively short, round oval or void ..…….............................................................................…...………………………….. 3
2. Build rather short and moderatey stout; head and pronotum relatively short; fastigium of vertex shorter; lateral pronotal lobe fairly deep, without a membranous area in metazona; aedeagal valve long and slender and curved upward in lateral view …..……...……………………..............................................................................................................................………… burri Bolivar, 1905-- Build very slender; head and pronotum relatively long; fastigium of vertex arrower and longer; lateral pronotal lobe shallower, sometimes with a small membranous area in the metazona; aedeagal valves longer and more strongly curved ................................................................................................................................................................................ psittacina (Haan, 1842)
3. Generally rather small; fastigium of vertex often comparatively short; membranous area in metazona of lateral pronotal lobe usually very distinct in female and well indicated in male; hind wings normally tyrian pink to light mallow purple or pale magenta at base, but quite often rather havily infumated; aedaegal valve small and short .................... himalayica (Bolivar, 1905)-̶ Size range variable; fastigium of vertex usually comparatively somewhat longer than above; membranous area in metazona of lateral pronotal lobe variably developed, often less distinct than above; hind wings pinkish to rose red or rose at base, less frequently infumated; aedeagal valve slightly shorter or slightly longer ……..………………............................……………………………….. 4
4. Size as a rule a little smaller, body length often less than 20mm in male and 30mm in female; fastigium of vertex generally narrower apically and less flat dorsally; hind wings rose red, not infrequently infumated, at least basally; aedeagal sclerites and aedeagal valves slightly shorter and less gradually tapered …………………..….....................…........……..……….. angusta Karsch, 1888̶ Size generally a little larger, body length usually more than 20mm in male and 30mm in female; fastigium of vertex usually rather broad and generally very flat dorsally; hind wings rose red or rose, not frequently infumated; aedaegal sclerites and aedaegal valves slightly longer and more gradually tapered …………......................................................……. sinensis Bolivar, 1905
Key to the tribes of the family Pyrgomorphidae Brunner (After Kevan & Akbar 1964)
1. Body usually depressed and usually rather strongly rugose (sometimes with plicate, longitudinal tubercles); coloration mottled brown or grayish; fastigium of vertex usually (but not always) short, blunt, broad; terminal segments of antennae incrassate, fused or partly so, often pitted; tegmina (when present) usually with small nodules on main veins; hind wings hyaline (sometimes faintly bluish) or infumated only; prosternum with reflexed, collar-like anterior margin and “double” tubercle; [phallic structures rather unspecialized; epiphallus with lateral plates having wide, basal, externo-lateral expansions, very strong, laterally directed lophi, and widely divergent appendices; ectophallus with wide basal emargination, short, broad ventral process, and central membrane of cingulum rather extensive, subquadrate or sub-rectangular; aedeagal scterites rather slender and acute, gonopore basal or mesal] …........……................................................................................... Chrotogonini Bolivar, 1904̶ Note as above ………………............................................................................................................................……………………..... 2
2. Head acutely conical, fastigium of vertex usually long, acute; antennae inserted distinctly in front of ocelli; tegmina usually fully developed, usually very tapered and pointed, brachypterous forms somewhat depressed (especially in female), strongly micropterous or apterous forms unknown; inferior margin of lateral pronotal lobe very straight and usually beset with small, fine, even, granular tubercles; infero-external area of hind femur expanded, often considerably so, and displaced subventrally; posterior tibial spines rather long and sharp;[epiphallus anchor-like, or with lateral plates widely expanded and lophi bifid (when aedeagal valves are strongly decurved apically), or of more orthodox form (when aedeagal valves bear large, prominent dorsal processes and aedeagal sclerites are slightly decurved apically)] ……..................................................................................………… 3̶ Not combining the above characters; antennae not usually inserted distinctly in front of ocelli, sometimes even behind them; tegmina, if fully developed, not usually very tapered and pointed, strongly micropterous and apterous forms frequent, brachypterous forms seldom (and, if body depressed, never) with the inferior margin of the lateral pronotal lobe remarkably straight and granular; infero-external area of hind femur not, or but little, expanded and displaced; posterior tibial spines usually shorter and blunter; [phallic structures without the above special features, if somewhat anchor-like, then very broadly so and aedeagal valves greatly enlarged (small apterous species)] ........................................................................ Tagastini Bolivar, 1905
3. Body rather heavy, somewhat depressed (especially in female), antennae inserted only a short distance in front of ocelli; infero-posterior angle of lateral pronotal lobe strongly acute; hind tarsal segments not elongate; male cerci rather specialized; [epiphallus with broad, wing-like lateral plates and lophi bifid; ectophallus with ventral process very short and pointed; endopallic apodemes with very long, forwardly directed ventral processes, aedeagal valves and sclerites very long and strongly decurved]; Southeast Asia only ................................................................................. Pseudomorphacridini Kevan & Akbar, 1964̶ Body less robust, not depressed, or, if slightly so (some females), hind tarsal segments elongate; antennae usually inserted well in advance of ocelli; infero-posterior angle of lateral pronotal lobe not strongly acute; male cerci unspecialized; [phallic structures not as above; ventral process of ectophallus moderately long, very broad and subtruncated apically]; widespread in Old World ……...……..………..…........................................................................................................... Atractomorphini Bolivar, 1905

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12953–12968
Taxonomic studies on gaudy grasshoppers Khan et al.
12956
Tribe Atractomorphini Bolivar, 1905Dignosis: Body slightly robust, not depressed, or,
if so slightly (some females), hind tarsal segments elongate; head elongated, acutely conical, fastigium of vertex usually long, acute, a row of fine, distinct granular tubercles extending from behind the eye across the inferior margin of the lateral pronotal lobe; infero-posterior angle of lateral pronotal lobe not strongly acute; antennae usually inserted well in advance of ocelli; tegmina usually fully developed, usually very tapered and pointed. Brachypterous forms, somewhat depressed (especially in females), strongly micropterous or apterous forms unknown; hind femur trigonal in cross section; posterior tibial spines rather long and sharp; male cerci unspecialized; ventral process of ectophellus moderately long, very broad and subtruncated apically.
Genus Atractomorpha Saussure, 1862 Saussure. 1862. Ann. Soc. Ent. Fr. 41: 474. Brunner von Wattenwyl. 1898. Abh. Senckenb.
Natforsch. Ges. 24(2): 234. Diagnosis: Small to medium size; integument finely
rugose; antennae slightly compressed, shorter than head and pronotum together, their bases located in front of lateral ocelli, fastigium of vertex elongated, flat, horizontal or slightly upcurved with parabolic or angular apex; apical
fastigial areolae poorly developed; head narrow, acutely conical, with a row of postocular tubercles; frons strongly oblique; frontal ridge incurved, narrow and low, shallowly sulcate, with obtuse lateral carinulae. Pronotum elongated, sub-cylindrical, slightly widening backwards; dorsum slightly flattened, crossed by three fine sulci; median carina and lateral carinar weak; metazoan much shorter than prozona, its posterior margin widely obtuse-angular, almost rounded; lateral lobe with row of low marginal tubercles; prosternal process cuneiform; elytra and wings fully developed, apex of elytron acutely attenuate; tympanal organ well developed; hind femur narrow, with external lower marginal area narrow, displaced ventrally to external medial area; lower lobes of hind knee much shorter than upper one; hind tibiae in apical part with expanded margins; external apical spine present; arolium of moderate size; male supra anal plate elongate angular; cercus subconical, straight with subacute apex; Subgenital plate short, with rounded apex.
The genus is represented by five species from this region.
Atractomorpha burri Bolívar, 1905
(Image 1; Fig. 1)Bolivar, I. 1905. Bol. R. Soc. Esp. Hist. Nat. 5: 197.
Image 1. Atractomorpha burri male & female

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12953–12968
Taxonomic studies on gaudy grasshoppers Khan et al.
12957
Kirby, W.F. 1901. 3 (2): 332. Banerjee, S.K. and Kevan, D.K. Mc E. 1960. Treubia,
25: 165-189.Kevan & Chen. 1969. Zool. J. Linn. Soc. 48: 193. Morphological Characters: Build rather short and
moderately stout; head and pronotum relatively short; fastigium of vertex shorter; pronotal lobe fairly deep, without a membranous area in metazona; outer face of hind femur rather strongly convexed and keeled; hind wings relatively long, their apices not falling much short of those of the tegmina at rest, extensively tyrian pink to light mallow purple at base.
Male genitalia: Supra anal plate elongate angular, epiproct long and narrow with obtusely conical apex, cerci as long as or slightly shorter than the epiproct, and conical at apex, paraproct smaller. Subgenital plate triangular with conical apex. Epiphallus anchor shaped bridge small, anterior projections not prominent, lateral plates broad and fused medially, broader at the base, middle piece of epiphallus much wider, lophi hooked with conical apex, appendices rod shaped, slightly expanded at apex, reaching up to the apex of lophi. Apical valve
long anad narrow, apical tip pointed, longer tanan basal valve, basal valve wide at base.
Female genitalia: Supra anal plate elongated, longer than wide, apex obtusely rounded. Cercus short, longer than wide and incurved. Subgeniatal plate: posterior margin without setae, egg-guide wide at base; narrowing apically, one and half time longer than wide. Spermatheca with apical and pre-apical diverticula, pre-apial diverticulum moderately long tubular, apical diverticulum long, tubular with protuberance. Ovipositor: dorsal valve long, wide with apex pointed, slightly longer than apodeme and more than twice as long as wide, ventral valve uniformly wide, apex elongated and obtusely rounded.
Material Examined: Reg. no. 133, 9 females, 5 males, 10-x-2009, on grasses, Meghalaya, East Khasi Hills, coll. M.I. Khan.
Morphometry: (length in mm)Male: Body 19.77, tegmina 18.61, pronotum 5.35,
hind femur 11.25.Female: Body 22.47, tegmina 19.97, pronotum 6.65,
Figure 1. Atractomorpha burri male (A–D) and female (E–H). A - supra anal plate, B - sub genital plate, C - epiphallus, D - aedeagus, E - supra anal plate, F - sub genital plate, G - spermatheca, H - ovipositor.
Image 2. Ataractomorpha psittacina male (A–D) and female (E–H). A - supra anal plate, B - sub genital plate, C - epiphallus, D - aedeagus, E - supra anal plate, F - sub genital plate, G - spermatheca, H - ovipositor.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12953–12968
Taxonomic studies on gaudy grasshoppers Khan et al.
12958
hind femur 12.39.Distribution: Bangladesh, Bhutan, China, Cambodia,
India (Arunachal Pradesh, Assam, Manipur, Meghalaya, Nagaland, Mizoram, Odisha, Sikkim, Tripura & West Bengal), Laos, Malaysia, Myanmar, Nepal, Thailand, and Veitnam.
Atractomorpha psittacina psittacina (Haan, 1842)(Image 2; Fig. 2)Haan. 1842. 16/18: 143, 146Morphological Characters: Build very slender;
head and pronotum relatively long; fastigium of vertex narrower and longer; lateral pronotal lobe shallow, sometimes with a small membranous area in the metazona; outer face of hind femur not strongly convex or keeled; hind wings normally comparatively long in relation to tegmina, not falling much short of them when at rest, usually dull magenta purple, often rather pale, at extreme base, often with dark veins and somewhat infumated, sometimes colourless.
Male genitalia: Supra anal plate elongate angular, epiproct triangular, long and broad at base with obtusely conical apex, cerci shorter than the epiproct and conical at apex, paraproct smaller. Subgenital plate rounded at apex. Epihallus anchor shaped, bridge much narrow, anterior projections not prominent with rounded apex,
lateral plates fused medially, middle piece of epiphallus narrow, lophi hooked with pointed apex, appendices rod-shaped, expanded at apex, slightly far from the apex of lophi. Apical valve narrow as long as basal valve, apex tubular, basal valve wide at base.
Female genitalia: Supra anal plate triangular, longer than wide, apex obtusely rounded, cercus short, wide at base, longer than wide, narrowing apically. Subgenital plate, posterior margin smooth without setae, notched in the middle, egg-guide narrowing apically, twice as long as wide, tip pointed. Spermatheca: apical and pre-apical diverticulum long, both are long, tubular and narrow. Ovipositor: dorsal valve broad, long, slightly longer than apodeme with apical tip pointed, ventral valve uniformly broad with tip pointed.
Material Examined: Reg. no. 134, 19 females, 15 males, 21-x-2008, on grasses, Meghalaya, Rai Bhoi, coll. M.I. Khan.
Morphometry: (length in mm)Male: Body 25.34, tegmina 21.35, pronotum 5.45,
hind femur 11.49.Female: Body 28.94, tegmina 22.50, pronotum 6.16,
hind femur 12.37. Distribution: Borneo, India (Arunachal Pradesh,
Assam, Meghalaya, Rajasthan, Tripura, & West Bengal), and Malaysia.
Figure 2. Atractomorpha psittacina male & female

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12953–12968
Taxonomic studies on gaudy grasshoppers Khan et al.
12959
Atractomorpha himalayica Bolívar, 1905(Image 3; Fig. 3)Bolivar, I. 1905. Bol. R. Soc. Esp. Hist. Nat. 5: 198, 204.Navas, 1905. Bol. Soc. Arag. Cienc. Nat. 4: 273.Kirby, 1910. 3(2): 332.Bey-Bienko & Mistshenko 1951. 1: 275.Morphological Characters: Generally rather small;
eyes generally shorter and rather convex; fastigium of vertex often comparatively short; inter-ocular space generally slightly convex; membranous area in metazona of lateral pronotal lobe usually very distinct in female and well indicated in male; hind wings normally tyrian pink to light mallow purple or pale magenta at base, but quite often rather heavily infumated.
Male genitalia: Supra anal plate elongate angular, epiproct triangular, long and slightly wider at base with obtusely conical apex, cerci shorter than the epiproct, and conical at apex, paraproct smaller. Subgenital plate oval at apex. Epihallus anchor shaped, bridge broader, anterior projections prominent with pointed apex, lateral plates fused medially, middle piece of epiphallus with subparallel margins, lophi hooked with pointed apex, appendices rod shaped, slightly expanded at apex, slightly far from the apex of lophi. Aedeagus: apical valve long, narrow, upcurved, distinctly longer than basal valve, apical tip pointed, basal valve narrow, broad at base.
Female genitalia: Supra anal plate elongated, slightly
Figure 3. Atractomorpha himalayica male (A–D) and female (E–H). A - supra anal plate, B - sub genital plate, C - epiphallus, D - aedeagus, E - supra anal plate, F - sub genital plate, G - spermatheca, H - ovipositor.
Image 3. Atractomorpha himalayica male & female

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12953–12968
Taxonomic studies on gaudy grasshoppers Khan et al.
12960
longer than wide, apex rounded, cercus short, broad, one and half time as long as wide. Subgenital plate: posterior margin smooth without setae, with a notch in the middle, egg-guide broad basally, narrowing apically, twice as long as wide. Spermatheca: apical diverticulum long, narrow and tubular, pre-apical diverticulum narrow basally, broad apically. Ovipositar, dorsal valve elongated, broad medially, apical tip pointed, longer than apodeme, ventral valve uniformally broad, apex pointed.
Material Examined: Reg. no. 135, 13 females, 10 males, 11-ii-2009, on grasses, Mizoram, Aizwal, coll. M.I. Khan.
Morphometry: (length in mm)Male: Body 18.45, tegmina 13.92, pronotum 4.24,
hind femur 9.05.Female: Body 21.53, tegmina 14.81, pronotum 5.93,
hind femur 10.39. Distribution: Bangladesh, Bhutan, China,
India(Arunachal Pradesh, Assam, Nagaland, Sikkim, & West Bengal), Nepal, and southern Malaya.
Atractomorpha angusta Karsch, 1888(Image 4; Fig. 4)Karsch. 1888. Entom. Nachricht. 14(21): 333.Bolivar, I. 1905. Bol. R. Soc. Esp. Hist. Nat. 5: 198, 207.Kirby, 1910. 3(2): 332.Kevan, 1963. Ark. Zool. 16(4): 80.Morphological Characters: Size a little smaller; eyes
generally a little longer and less convex; fastigium of vertex generally narrower apically and less flat dorsally; interocular space generally flatter; membranous area in metazona of lateral pronotal lobe variably developed, often less distinct than above; hind wings rose red, not infrequently infumated, at least basally.
Male genitalia: Supra anal plate broad basally, moderately narrowing apically, as long as wide, apex rounded, cercus elongated, broad basally, narrowing apically, more than one and half time as long as wide, apex obtusely rounded. Epiphallus anchor shaped, anchorae elongated, narrow basally, broad apically. Aedeagus, apical valve narrowing apically with apex pointed, twice as long as wide, basal valve short and wide at apex.
Female genitalia: Supra anal plate broad at base, slightly narrow at apex, as long as wide, apex obtusely rounded, cercus short, broad, narrow apex, one and half time as long as wide. Subgenital plate smooth without setae, notched in the middle, egg-guide short, broad at base, narrowing apically, twice as long as wide. Spermatheca, apical diverticulum long, tubular and narrow. Pre-apical diverticulum uniformly tubular, S-shaped. Ovipositor: dorsal valve elongated, broad,
slightly longer than apodeme, apical tip pointed, ventral valve uniformly broad, apical tip obtusely rounded.
Morphometry: (length in mm)Female: Body 20.97, tegmina 14.37, pronotum 5.71,
hind femur 11.93Material examined: Reg. no. 136, 15 females, 21-IX-
2008, on grasses, Manipur, Ukhrul, coll. M.I. Khan.
Image 4. Atractomorpha angusta (female)
Figure 4. Atractomorpha angusta (female).A - Supra anal plate, B - Sub genital plate; C - Spermatheca, D - Ovipositor

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12953–12968
Taxonomic studies on gaudy grasshoppers Khan et al.
12961
Distribution: Bhutan, Cambodia, India (Andaman & Nicobar Islands, Assam, Arunachal Pradesh, Manipur, Meghalaya, & West Bengal), Indonesia, Laos, Myanmar, Malaysia, Nepal, Singapore, Thailand, and Vietnam.
Atractomorpha sinensis Bolívar, 1905(Image 5, Fig. 5)Bolivar, I. 1905. Bol. R. Soc. Esp. Nat. 5: 198, 205.Morphological Characters: Size generally a little larger;
fastigium of vertex usually rather broad and generally very flat dorsally; pronotum with distinct carinae; lateral pronotal lobes without membranous area; hind wings rose red or rose, not frequently infumated.
Male genitalia: Supra anal plate elongate angular, epiproct triangular, long and wider at base with obtusely conical apex, cerci shorter than the epiproct and conical at apex, paraproct smaller. Subgenital plate flat and globular at apex. Epihallus anchor shaped, bridge wider, anterior projections prominent with hook like apex, lateral plates fused medially, broader at base, middle piece of epiphallus slightly narrow, lophi hooked with obtusely conical apex, appendices rod shaped, much expanded at apex, slightly far from the apex of lophi. Aedeagus apical valve elongated, uniformly tubular, upcurved, apex obtusely rounded, basal valve short and broad.
Female genitalia: Supra anal plate broad at base, slightly narrowing apically, as long as wide, apex rounded, cercus short, broad, narrowing apically, apex blunt, one
Image 5. Atractomorpha sinensis male & female
Plate 6: Ataractomorpha sinensis : Male (A-D) Female (E-H) A. Supra anal plate, B. Sub genital plate, C. Epiphallus, D. Aedeagus E. Supra anal plate F. Sub genital plate, G. Spermatheca, H. Ovipositor
1.0 mm
A
1.0 mm
B
0.5 mm
C
0.5 mm
D
1.0 mm
E
1.0 mm
F
1.0 mm
H 0.5 mm
G
Figure 5. Atractomorpha sinensis: male (A–D) & female (E–H).A - Supra anal plate, B - Sub genital plate, C - Epiphallus, D - Aedeagus, E - Supra anal plate, F - Sub genital plate, G - Spermatheca, H - Ovipositor

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12953–1296812962
than basal tarsal segment arolium large; valve of
cingulum broad ……...…… Tenuitarsus Bolivar, 1904
Key to species of Chrotogonus Serville, 1838
1. Tegmina slightly reaching near apex of hind knee ; wings hyaline, slightly shorter than tegmina …. ......................................... armatus Steinmann, 1965̶ Tegmina surpassing the apex of hind knee; much shorter than the length of tegmina ; aedeagus rather broad with sclerites and valves rather blunt apically
…….................…………….... oxypterus Blanchard, 1836
Genus Chrotogonus Serville, 1838Serville. 1838. Hist. nat. des insectes. Orthoprteres
702.Navas. 1904. Bol. Soc. Arag. Cienc. Nat. 3: 133.Diagnosis : Small and robust; body depressed,
integument strongly tuberculate; antennae thick, slightly widening in apical half, shorter than head and pronotum together; fastigium of vertex short, angular, concave; apical fastigial areolae large, with sharp marginal carinulae; occipital carina present; frontal ridge between antennae strongly compressed and protruding forwards, with slit-like sulcus, below almost obliterated. Pronotum above flattened, with posterior angle of lateral lobes spread sidewise, strongly tuberculate, with irregular, interrupted, median and lateral carinae, crossed by three sulci; metazona longer than prozona, its posterior margin angular; anterior margin of prosternum strongly expanded, collar like, covering lower part of mouth with a pair of posterior lateral tubercles; elytra and wings fully developed, shortened and vestigial; tympanum absent or vestigial; hind femur moderately slender; hind tibiae depressed and expanded towards apex; external apical spine absent; internal pair of spurs much longer than external; arolium of medium size; male supra anal plate angular; cersus short, obtusely conical; subgenital plate short, subconical, with obtuse apex; epiphallus with large, strongly curved, acute lophi.
Chrotogonus (Chrotogonus) oxypterus (Blanchard, 1836) (Image 6; Fig. 6)
Blanchard, 1836. Ann. Soc. Ent. Fr. 5: 622.Walker, F. 1870. 4: 793.Bolivar, I. 1902. Ann. Soc, ent. Fr. 70: 603.Kirby, 1901. 3 (2): 301.Morphological Characters: Small to medium
sized insect. Body becomes robust and compressed. Antennae filliform, nine segmented, shorter than head and pronotum together. Head and pronotum more tuberculated. Tegmina surpassing the tip of hind femur, veins with small tubercles. Hind femur stout, reaching up
and half time as long as wide. Subgenital plate, posterior margin smooth without setae, notched in the middle, egg-guide short, broad basally, narrowing apically, one and half time as long as wide. Spermatheca, apical and pre-apical diverticulum, narrow, tubular, diverticulum shorter than pre-apical diverticulum. Ovipositor, dorsal valve broad, with apical tip obtuse, slightly shorter than apodeme, ventral valve narrow, uniformally broad, apical tip bluntly rounded.
Material Examined: Reg. no. 137, 23 females, 18 males, 28-x-2008, on paddy field, Nagaland, Dimapur, coll. M.I. Khan.
Morphometry: (length in mm)Male: Body 22.43, Tegmina 17.54, Pronotum 3.90,
Hind femur 10.92Female: Body 25.49, Tegmina 18.74, Pronotum 4.90,
Hind femur 11.82 Distribution: Bhutan, India (Assam, Jammu & Kashmir,
Meghalaya, and West Bengal), Indonesia, Myanmar, Sri Lanka
Tribe Chrotogonini I. Bolivar, 1904Bolivar, I. 1904. Bol. R. Soc. Esp. Hist. Nat. 4:90.Diagnosis : Body usually depressed and usually rather
strongly rugose (sometimes with plicate, longitudinal tubercles); coloration mottled brown or greyish; fastigium of vertex usually (but not always) short, blunt, broad; terminal segments of antennae incrassate, fused or partly so, often pitted; tegmina (when present) usually with small nodules on main veins; hind wings hyaline (sometimes faintly bluish) or infumated only; prosternum with reflexed, collar like anterior margin and double tubercle; epiphallus with lateral plates having wide, basal, externo-lateral expansions, very strong, laterally directed lophi, and widely divergent appendices; ectophallus with wide basal emargination, short, broad ventral process, and central membrane of cingulum rather extensive subquadrate or sub-rectangular; aedeagal sclerites rather slender and acute, gonopore basal or medial.
Key to genera of the tribe Chrotogonini I. Bolivar
1. Body strongly depressed; dorsum of pronotum strongly tuberculate; middle femur short, much shorter than head and pronotum together; hind femur with lower basal lobe longer than upper lobe; hind tibial spurs shorter than basal tarsal semment; arolium large; valve of cingulum narrow ……………………..........…… Chrotogonus Serville, 1838 ̶ - Body slightly depressed; dorsum of pronotum never strongly tuberculate; middle femur thin and strongly elongated, as long as or longer than head and pronotum together; hind femur with lower basal lobe shorter than upper lobe; hind tibial spurs longer
Taxonomic studies on gaudy grasshoppers Khan et al.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12953–12968 12963
to the tip of abdomen. Hind tibiae nearly equal to hind femur with eight outer and nine internal spines.
Male genitalia: Supra anal plate angular with broad and short epiproct, cerci as long as or slightly longer than the epiproct, obtusely conical at apex, paraproct broad and slightly longer than epiproct. Subgenital plate triangular and conical at the end. Epiphallus bridge shaped, bridge moderately slender, slightly wider and long; anterior projection prominent with angular tip; lateral plates separated medially, lophi with curved apices, apex acute; lateral appendices rod shaped and hooked at the end, apex pointed and reaching the tip of lophi. Aedeagus, apical valve slightly upcurved, apex pointed, longer than basal vave. Basal valve broad, much wide at base.
Material Examined: Reg. no. 138, 8 males, 3.x.2011, on grasses, Arunachal Pradesh, East Siang, Pasighat, coll. M.I. Khan.
Morphometry: (length in mm)Male: Body 15.23, Tegmina 5.31, Pronotum 4.34,
Hind femur 8.31Distribution: Bangladesh, India (Andhra Pradesh,
Bihar, Chhattisgarh, Goa, Karnataka, Kerala, Maharashtra, Madhya Pradesh, Odisha, Tamil Nadu, Uttar Pradesh & West Bengal), and Sri Lanka
Chrotogonus (Chrotogonus) armatus Steinmann, 1965(Image 7; Fig. 7)Steinmann. 1965. Mus. Praze. 36: 293.Kevan, 1977. 16: 539.Morphological Characters: Body yellowish brown
with metazona of pronotum and mid of hind femur outer and upper surface white. Antennae eleven segmented, shorter than head and pronotum together. Lateral carina of pronotum represented by weak lines only in metazona. Tegmina slightly reaching up to the apex of hind femur. Hind tibiae with seven outer and eight inner spines.
Male genitalia: Supra anal plate angular with broad and long epiproct, cerci shorter than the epiproct, broad and obtusely rounded at apex, paraproct is also broader and as long as or slightly longer than the epiproct. Subgenital plate broad and rounded at the apex. Epihallus bridge shaped, bridge more slender, slightly narrow and elongated; anterior projection less prominent; lateral plates separated medially, lophi with curved apices, apex acute; lateral appendices rod shaped and hooked at the end, apex expanded and crossing the tip of lophi. Aeadegus apical valve narrow, tubular, as long as basal valve, basal valve broad, wide.
Material Examined: Reg. no. 139, 5 males, 2.x.2011, on grasses, Arunachal Pradeh, East Siang, Pasighat, coll.
M.I. Khan.Morphometry: (length in mm)Male: Body 15.12, Tegmina 5.29, Pronotum 4.43,
Hind femur 8.21Distribution: Afghanistan, Bangladesh, India (Andhra
Pradesh, Assam, Bihar, Jammu & Kashmir, Uttar Pradesh & West Bengal), Nepal and Pakistan.
Genus Tenuitarsus Blivar, 1904Bolivar, I. 1904. Bol. R. Soc. Esp. Hist. Nat. 4: 90.Bey- Bienko & Mistshenko. 1951. 1: 279 [296].
Image 6. Chrotogonus (Chrotogonus) oxypterus
Plate 5: Chrotogonus (Chrotogonus) oxypterus (Male) A. Supra anal plate, B. Sub genital plate, C. Epipallus D. Aedeagus
1.0 mm
A
0.5 mm
C
1.0 mm
B
0.5 mm
D
Figure 6. Chrotogonus (Chrotogonus) oxypterus (male)A - Supra anal plate, B - Sub genital plate, C - Epiphallus, D - Aedeagus
Taxonomic studies on gaudy grasshoppers Khan et al.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12953–12968
Taxonomic studies on gaudy grasshoppers Khan et al.
12964
Diagnosis: Small and slender; body slightly depressed, integument rugose and hairy; antennae in apical half thickened, with fused segments, shorter than head and pronotum together; fastigium of vertex short, sloping forwards, slightly concave, with obtuse angular apex and deep; large apical fastigial areolae and sharp marginal carinulae; weak occipital carina present; frontal ridge between antennae compressed
and protruding forwards, with slit-like sulcus, below almost obliterated. Pronotum subcylindrical, widening backwards, tuberculate, with weak linear median carina, crossed by three sulci, lateral carinae absent, metazona about as long as prozona, its posterior margin rounded; anterior margin of prosternum strongly expanded, collar like, covering lower part of mouth; elytra and wings fully developed; tympanum absent; middle femur and tibiae elongated, thin; hind femur slender, with lower basal lobe slightly shorter than upper one; hind tibiae slightly expanded in apicl half; external apical spine absent; spurs of hind tibiae strongly elongated, longer than basal tarsal segment, thin, internal pair longer than external; all tarsi thin, slightly elongated; arolium very small; male supra anal plate angular; cercus short, obtusely conical; subgenital plate short, widely subconical; epiphallus with large lophi, moderately curved at apices.
The genus is represented by a single species from this region.
Tenuitarsus orientalis Kevan, 1959 (Image 8; Fig. 8)Kevan, 1959. Publ. Cult. Comp. Diamant. Angola. 43:
21.Kevan. 1977. 16: 532. Shishodia, Chandra & Gupta. 2010. Rec. Zool. Surv.
India, Misc. Pub., 314: 135.Kumar, Usmani & Kumari. 2014. J. Entomol. Res. Soc.
16(1): 23.Morphological Characters: Body slightly depressed,
integument rugose and hairy; antennae shorter than head and pronotum together; fastigium of vertex short, slightly concave; large apical fastigial areolae and sharp marginal carinulae. Pronotum subcylindrical, widening backwards, tuberculate, with weak linear median carina, crossed by three sulci, lateral carinae absent, metazona about as long as prozona, its posterior margin rounded; elytra and wings fully developed; middle femur and tibiae elongated, thin; hind femur slender, with lower basal lobe slightly shorter than upper one; hind tibiae slightly expanded in apical half.
Male genitalia: Supra anal plate angular with broad and small triangular epiproct, cerci as long as or slightly longer than the epiproct and obtusely conical at apex, paraproct also broader and equally or smaller than the epiproct. Subgenital plate broad and rounded at the apex. Epiphallus bridge shaped, bridge small and slightly wider; anterior projection less prominent with conical apex; lateral plates separated medially, lophi with curved apices, apex obtusely conical; lateral appendices rod shaped and pointed at the apex and crossing the tip of lophi. Aedeagus, apical valve much narrower, apex
Figure 7. Chrotogonus (Chrotogonus) armatus male
Plate 7: Chrotogonus (Chrotogonus) armatus (male) A. Supra anal plate, B. Sub genital plate, C. Epiphallus, D. Aedeagus
1.0 mm
A
1.0 mm
B
0.5 mm
C
0.5 mm
D
Figure 7. Chrotogonus (Chrotogonus) armatus (male)A - Supra anal plate, B - Sub genital plate, C - Epiphallus, D - Aedeagus

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12953–12968
Taxonomic studies on gaudy grasshoppers Khan et al.
12965
Image 8. Tenuitarsus orientalis male & female
Image 8. Tenuitarsus orientalis male (A–D) and female (E–H). A - supra anal plate, B - subgenital plate, C - epiphallus, D - aedeagus, E - supra anal plate, F - sub genital plate, G - spermatheca, H - ovipositor.
obtuse, as long as basal valve, basal valve uniformaly broad, wide at base.
Female genitalia: Supra anal plate short, broad, wider than long, apex rounded, cercus short, broad, slightly longer than wide, apex obtuse, much shorter than supra anal plate. Subgenital plate, posterior margin smooth, round, without setae, egg-guide cone shaped, longer than wide, apex obtuse. Spermatheca, single apical diverticulum which is S- shaped, tubular and uniformly broad. Ovipositor, dorsal valve short, broad, apical tip rounded, external edge serrated, much shorter than lateral apodeme, ventral valve short, uniformly broad, apical tip obtusely rounded.
Material Examined: Reg. no. 140, 6 females, 31.i.2009, in paddy field, Assam, Guwahati, Bongra, 15 males, 28-X-2008, on paddy field, Assam, Lakhimpur, North Lakhimpur, coll. M.I. Khan.
Morphometry: (length in mm)Male: Body 20.43, Tegmina 5.16, Pronotum 4.19,
Hind femur 12.17Female: Body 23.64, Tegmina 6.32, Pronotum 5.32,
Hind femur 13.75 Distribution: Bhutan, India (Assam, Arunachal
Pradesh, West Bengal & Rajasthan), Myanmar, and Pakistan.
Genus Tagasta Bolívar, 1905Bolivar, I. 1905. Bol. R. Soc. Esp. Hist. Nat. 5: 111.Willemse, 1928. Zool. Mededelingen (Leiden). 11(1):
5.Diagnosis: Body slightly compressed. Head conical,

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12953–12968
Taxonomic studies on gaudy grasshoppers Khan et al.
12966
Image 9. Tagasta indica female. A - supra anal plate, B - subgenital plate, C - spermatheca, D - ovipositor.
shorter than the pronotum, tempora widened in front, only separated by a short suture, front very oblique, frontal ridge much flattened, hardly sulcated, shortly compressed between the antennae; the latter concolorous, filiform and inserted between the eyes. Eyes rounded; ocelli distinct, cheeks granulated. Pronotum pubescent, roundly truncate in front, obtusely angulated, with the median carina very slightly indicated, lateral carinae obsolete; the prozona considerably longer then the metazona; the lower margin oblique, subsinuate bordered with whitish, the anal angle obtuse nearly rectangular. Tegmina not or scarcely longer than the hind femora, with costal area considerably expanded near the base. Wings distinctly shorter than the tegmina, red or hyaline. Legs long and slender; hind tibiae with rounded spines. Female. Subgenital plate smooth, without setae.
The genus is represented by a single species from this region.
Tagasta indica Bolívar, 1905 (Image 9; Fig. 9)Bolivar, I. 1905. Bol. R. Soc. Esp. Hist. Nat. 5: 112-114.Morphological Characters: Oilvaceous in colour;
fastigium of vertex equilaterally triangular; antennae inserted near the eyes; pronotum rounded in front and obtusely angulated behind; median carina almost and lateral carinae wholly absent; tegmina as long as hind femora, with a brown spot at the base; hind wings one-fifth shorter than tegmina.
Female genitalia: Supra anal plate broad, as long as wide, apex rounded, cercus short, broad basally, narrow apically, one and half time as long as wide, apex obtusely rounded. Subgenital plate, smooth without setae, egg-guide broad basally, narrowing apically, slightly less than two times as long as wide, apex rounded. Spermatheca with single diverticulum, long, uniformly broad, S-shaped. Ovipositor, dorsal valve long, broad, slightly longer than lateral apodeme, external edge serrated, apical tip pointed, ventral valve long, uniformly broad, apical tip long, obtusely rounded.
Material Examined: Reg. no. 141, 13 females, 21.x.2008, on paddy field, Meghalaya, Rai Bhoi, Umran, coll. M.I. Khan.
Morphometry: (length in mm)Female: Body 33.57, tegmina 21.91, pronotum 8.54,
hind femur 18.71.Distribution: Bhutan, India (Andaman & Nicobar
Islands, Arunachal Pradesh, Meghalaya, Nagaland. Sikkim, Tripura, & West Bengal), and Myanmar.
Genus Pseudomorphacris Carl, 1916Carl, 1916. Revue Suisse de Zool. 24(6): 465.
Kevan, 1963. Ent. Monthly mag. 98: 208.Kevan, 1977. 16: 345.Diagnosis: Body of medium size, somewhat
depressed; integument rugose, antennae slightly compressed, shorter than head and pronotum together, inserted only a short distance in front of ocelli. Fastigium of vertex flattened, usually long acute. Apical areolae poorly developed. Head subconical, elongated, shorter than pronotum. Frons oblique. Frontal ridge sulcate with obtuse lateral carinae. Pronotum elongated,
Figure 9. Tagasta indica female

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12953–12968
Taxonomic studies on gaudy grasshoppers Khan et al.
12967
slightly widening backwards, dorsum slightly flattened, crossed by three cerci, median carina and lateral carina weak; metazona shorter than prozona, posterior margin obtuse angular; infero-posterior angle of lateral pronotal lobe strongly acute. Prosternum with anterior margin thickened, completely lacking tubercles. Prosternal process cylindrical. Tegmina fully developed, usually tapered and pointed. Posterior tibia round and have superior obtuse lobes, posterior tibial spine rather sharp and long; hind tarsal segment not elongate, external apical spine present, arolium of moderate size. Male supra anal plate elongate, apex obtusely rounded, subgenital plate elongate, apex rounded.
The genus is represented by a single species from this region.
Pseudomorphacris notata (Brunner von Wattenwyl, 1893) (Image 10; Fig. 10)
Brunner von Wattenwyl. 1893. Ann. Mus. Civ. Stor. Nat. Genova. 213(33): 130.
Bolivar, I. 1905. Bol. R. Soc. Esp. Hist. Nat. 5: 112.Kirby, 1910. 3(2): 330.Morphological Characters: Body-form more slender
and stout than Atractomorpha Saussure and Tagasta Bolivar. Head somewhat narrower at base; tegmina at rest reaching approximately to hind knee, with a black spot with yellow tinge at the base; hind wings distinctly purplish, not falling far short of tegmina when at rest;
Figure 10. Pseudomorphacris notata male & female
hind tibiae pink; male cerci, in lateral view, less strongly curved or bent.
Male genitalia: Supra anal plate elongate, one and half times long as wide, apex obtusely rounded, cercus elongate, twice as long as wide, broad basally, narrowing apically, apex obtusely rounded. Subgenital plate elongated, apex rounded. Epiphallus, anchore short, lophi triangular, dorso-lateral appandages tubular. Aedeagus, apical valve narrow, tubular, slightly excurved, apex obtuse, basal valve uniformaly broad, as long as apical valve.
Female genitalia: Supra anal plate short, broad, wider than long, apex obtusely rounded, cercus short, broad basally, narrowing apically, longer than wide, apex obtuse. Subgenital plate, posterior margin straight, smooth without setae, egg-guide elongate, narrowing apically, twice as long as wide, apex pointed. Spermatheca with single apical diverticulum, uniformaly broad, curved. Ovipositor, dorsal valve broad, external edge slightly serrated, three times as long as wide, distinctly shorter than lateral apodeme, apical tip pointed, ventral valve uniformaly broad, apical tip obtuse.
Material Examined: Reg. no. 142, 23 females, 18 males, 21.x.2011, on grasses, Tripura, Pencharthal (North Tripura), 13 females, 9 males, 25-X-2011, on grasses, Mizoram, Aizwal, coll. M.I. Khan.
Morphometry: (length in mm)Male: Body 25.42, tegmina 8.18, pronotum 5.27, hind

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12953–12968
Taxonomic studies on gaudy grasshoppers Khan et al.
12968
femur 6.71.Female: Body 28.73, tegmina 9.27, pronotum 6.71,
hind femur 7.83.Distribution: Bangladesh, India (Assam, Mizoram, &
Tripura), Myanmar, and Thailand.
References
Ajaili, A.A. & M.K. Usmani (1994). Taxonomic significance of male subgenital plate in some Libyan Grasshoppers (Orthoptera: Acridoidea). Sebha University Journal, Part B 1: 5–12.
Ayyar, T.V.R. (1940). Handbook of Entomology for South India. Government Press, Madras, 1pl+xx+528pp.
Bhowmik, H.K. (1964). On a new species of Eyprepocnemis (Insecta: Orthoptera: Acrididae) from India. Journal of the Bombay Natural History Society 32: 89–96.
Bhowmik, H.K. & P. Halder (1983). Preliminary distributional records with remarks on little known species of Acrididae (Orthoptera: Insecta) from the western Himalayas (Himachal Pradesh). Records of the Zoological Survey of India 81: 167–191.
Chopard, L. (1924a). On some cavernicolous Orthoptera and Dermaptera from Assam & Burma. Records of the Indian Museum 26: 81–92.
Chopard, L. (1924b). The fauna of an island in the Chilka Lake: the Dermaptera and Orthoptera of Barkuda Islands. Records of the
Indian Museum 26: 168–191.Dirsh, V.M. (1956a). Preliminary revision of the genus Catantops
Schaun and review of the group Catantopini (Orthoptera: Acrididae). Companhia de Diamantes de Angola, Serviços Culturais Dundo-Lunda-Angola, Lisbon 28: 11–150.
Dirsh, V.M. (1956b). The phallic complex in Acridoidea (Orthoptera) in relation to taxonomy. Transactions of the Royal Entomological Society of London 108: 223–356.
Dirsh, V.M. (1957a). The spermatheca as a taxonomic character in Acridoidea (Orthoptera). Proceedings of the Entomological Society of London (A) 32: 107–114.
Dirsh, V.M. (1957b). Two new genera of Acridoidea (Orthoptera). Annals & Magazine of Natural History 10: 860–862.
Dirsh, V.M. & B.P. Uvarov (1953). Tree locusts of the genus Anacridium (Orthoptera: Acrididae). Eos Madrid 29(1): 7–69.
Herrera, L. & S. Schnidrig (1983). Andropigios de los Ortópteros de Navarra (Orthoptera). Boletín de la Real Sociedad Española de Historia Natural (Sección Biológica) 10: 1–52.
Kevan, D.K. McE., S.S. Akbar & Y.C. Chang (1970). The concealed copulatory structures of the Pyrgomorphidae (Orth. Acridoidea), Part II - Tribes Fijipyrgini, Verduliini, Brumiiellini, Psednurini, Mitricephalini, Geloimi, Saggittacridini, Gymnohippini and Malagasphenini. Eos Madrid 45: 173–228.
Kirby, W.F. (1910). A Synonymic Catalogue of Orthoptera (Orthoptera Saltatoria). London, ix+674pp.
Kirby, W.F. (1914). The Fauna of British India, including Ceylon and Burma, Orthoptera (Acridiidae) 1. London, ix+276pp.
Kumar, H., M.K. Usmani & R. Kumari (2014). Taxonomic significance of male supra-anal plate, cerci and subgenital plate in the classification of Indian Pyrgomorphidae (Orthoptera: Pyrgomorphoidea). Journal of the Entomological Research Society 16(1): 21–26.
Meinodas, K., S.A. Shafee & M.K. Usmani (1982). Taxonomic significance of spermatheca in some Indian grasshoppers (Orthoptera: Acrididae). Journal of the Bombay Natural History Society 79(2): 331–335.
Slifer, E.H. & R.L. King (1936). An internal structure in the Crytacanthacridinae (Orthoptera, Acrididae) of possible taxonomic value. Journal of the New York Entomological Society 44: 345–348.
Slifer, E.H. (1939). The internal genetalia of female Acridinae, Oedipodinae and Pauliniinae (Orthoptera, Acrididae). Journal of Morphology 65: 437–470.
Slifer, E.H. (1940). The internal genetalia of female Ommexechinae and Crytacanthacridinae. Journal of Morphology 67: 199–240.
Slifer, E. H. (1943). The internal genetalia of some previous unstudied species of female Acrididae (Orthoptera). Journal of Morphology 72: 225–237.
Tandon, S.K. & M.S. Shishodia (1969). On a collection of Acridoidea (Orthoptera) from the Nagarjuna Sagar Dam Area. Oriental Insects 3(3): 265–267.
Tandon, S.K. (1976). A check-list of the Acridoidea. Records of Zoological Survey of India, Occasional Paper No. 3: 1–48
Tandon, S.K. & M.S. Shishodia (1989). Fauna of Orissa, State Fauna Series 1, Part 2. Zoological Survey of India, 93–145pp.
Usmani, M.K. & A.A. Ajaili (1993). Taxonomic significance of Aedeagus in some Libyan Grasshoppers (Orthoptera: Acridoidea). Indian Journal of Systematic Entomology 10(1): 5–10.
Usmani, M.K. & S.A. Shafee (1985). A new species of the genus Ramakrishnaia I. Bolivar (Orthoptera: Pyrgomorphidae) from India. International Journal of Entomological Research 27(3): 204–207.
Uvarov, B.P. (1925). Grasshoppers (Orthoptera: Acrididae) from the Mount Everest. Annals and Magazine of Natural History 16(9): 165–173.
Uvarov, B.P. (1929). Acrididen (Orthoptera) aus Sud-Indien. Revue suisse de Zoologie 36: 533–563.
Threatened Taxa
Image 10. Pseudomorphacris notate male (A–D) and female (E–H). A - supra anal plate, B - sub genital plate, C - epiphallus, D - aedeagus, E - supra anal plate, F - sub genital plate, G - spermatheca, H - ovipositor.

12969
Com
mun
icat
ion
DOI: https://doi.org/10.11609/jott.3463.10.15.12969-12978 | ZooBank: urn:lsid:zoobank.org:pub:57B7ABD1-A0FA-4BAF-A7D4-24121B224CC6
Editor: K.A. Subramanian, Zoological Survey of India, Chennai, India. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 3463 | Received 24 March 2017 | Final received 13 December 2018 | Finally accepted 16 December 2018
Citation: Debata, S. & K.K. Swain (2018). Odonata (Insecta) diversity of Kuldiha Wildlife Sanctuary and its adjoining areas, Odisha, eastern India. Journal of Threatened Taxa 10(15): 12969–12978; https://doi.org/10.11609/jott.3463.10.15.12969-12978
Copyright: © Debata & Swain 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: None.
Competing interests: The authors declare no competing interests.
Author Details: Subrat Debata is a wildlife researcher and naturalist with special interest on small mammals and waterbirds. Kedar Kumar Swain is presently working as Divisional Forest Officer in Chandaka Wildlife Division, Forest and Environment Department, Government of Odisha.
Author Contribution: Both the authors contributed equally in field work. SD designed and wrote the paper.
Acknowledgements: We are thankful to the Forest Range Officers of Kuldiha Wildlife Range and Soro Wildlife Range of Balasore Wildlife Division for the logistics and other field support during the study. We are thankful to Udit Pratap Das and Arajush Payra for allowing us to use some of their photographs.
Odonata (Insecta) diversity of Kuldiha Wildlife Sanctuary and its adjoining areas, Odisha, eastern India
Subrat Debata ¹ & Kedar Kumar Swain 2
1 Aranya Foundation, Plot No-625/12, Mars Villa, Panchasakha Nagar, Dumduma, Bhubaneswar, Odisha 751019, India2 Office of the Divisional Forest Officer, Chandaka Wildlife Division, Gaja Vihar, Bhubaneswar, Odisha 751003, India1 [email protected] (corresponding author), 2 [email protected]
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12969–12978
Abstract: A study was carried out to assess the Odonata fauna of Kuldiha Wildlife Sanctuary, Odisha, eastern India from November 2012 to October 2013. During the study a total of 54 species of odonates including 37 species of dragonflies (Anisoptera) and 17 species of damselflies (Zygoptera) were recorded. Among the dragonflies, the family Libellulidae was well represented with 30 species whereas among the damselflies, Coenagrionidae was well represented with seven species. Overall, the odonate fauna of Kuldiha Wildlife Sanctuary accounted for 49.09% of the odonate species known from Odisha and 10.73% of India. Therefore, further long-term studies on these lesser-known insect fauna in Kuldiha Wildlife Sanctuary will be useful in understanding their status over time.
Keywords: Anisoptera, Coenagrionidae, damselflies, dragonflies, Libellulidae, Zygoptera.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12969–12978
Odonates of Kuldiha Wildlife Sanctuary Debata & Swain
12970
INTRODUCTION
The order Odonata comprising both dragonflies and damselflies are believed to have evolved some 250 million years ago (Subramanian 2005). These aquatic insects being predators in both larval and adult stages are an important and widespread component of freshwater ecosystems (Adarsh et al. 2015) as well as valuable indicators of water quality and landscape disturbance (Watson et al. 1982; Castella 1987; Varghese et al. 2014). Globally around 5,952 species of odonates have been described; of which 503 species have been reported within the geographic limits of India so far (Joshi et al. 2017).
Odisha is one of the eastern coastal states of India and being situated along the amalgamation zone of Chhotanagpur Plateau, Eastern Ghats Highlands, Lower Gangetic Plain and the Eastern Coastal Plain’s biogeographic provinces (Ray 2005), represents a mixture of both Indo-Malayan and Afro-Mediterranean biodiversity elements (Das et al. 2015). Odonata research in Odisha dates back to the early 1900s when Laidlaw (1915) and Fraser & Dover (1922) studied the faunal diversity of Chilika Lake. Afterwards, as part of faunal expeditions, several collections were made from different parts of Odisha and the results of 58 species were documented in the state fauna series (Srivastava & Das 1987). Some of the recent published works from Odisha include: Mitra (2000) who reported 69 species of odonates throughout Odisha; Sethy & Siddiqi (2007) reported 16 species from Simlipal Biosphere Reserve; Das et al. (2010, 2011) reported 31 species from Baripada Forest Division, 26 species from Nandankanan Zoological
Park and 58 species within the buffer area of Simlipal Tiger Reserve, respectively; Nair (2011) reported 110 species throughout Odisha and eastern India and 92 species from Simlipal Biosphere Reserve; Debata et al. (2013) reported 55 species from Hadgarh Wildlife Sanctuary; Payra et al. (2014) reported 56 species from Athagarh Forest Division; Sajan & Mohapatra (2014) reported the occurrence of Lesser Blue Wing (Rhyothemis triangularis Kirby, 1889) in Odisha from Kotgarh Wildlife Sanctuary and recently Pandey & Mohapatra (2017) reported 24 species from the Regional Institute of Education campus, Bhubaneswar. The vital information on diversity and distribution of odonates, however, is still missing from different parts of Odisha. Moreover, the increasing biotic pressure, deforestation and disappearance of wetlands are becoming major threats to odonates today. Therefore, documentation of Odonata from different geographic regions and habitats of Odisha is crucial for establishing baseline data for future comparison (Nair 2011). In this study, we summarize our findings of odonate fauna of Kuldiha Wildlife Sanctuary (KWS) in Odisha.
MATERIALS AND METHODS
Study AreaThe KWS (Fig. 1) is situated along the tropic zone
between 21.333–21.5000N and 86.500–86.7500E covering an area of 272.75km2 in northern Odisha region. The landscape is characterized by undulating terrain and altitude ranges between 169–682 m. The climate is seasonal, with summer season between March to
Figure 1. Map showing location of Kuldiha Wildlife Sanctuary, Odisha, eastern India
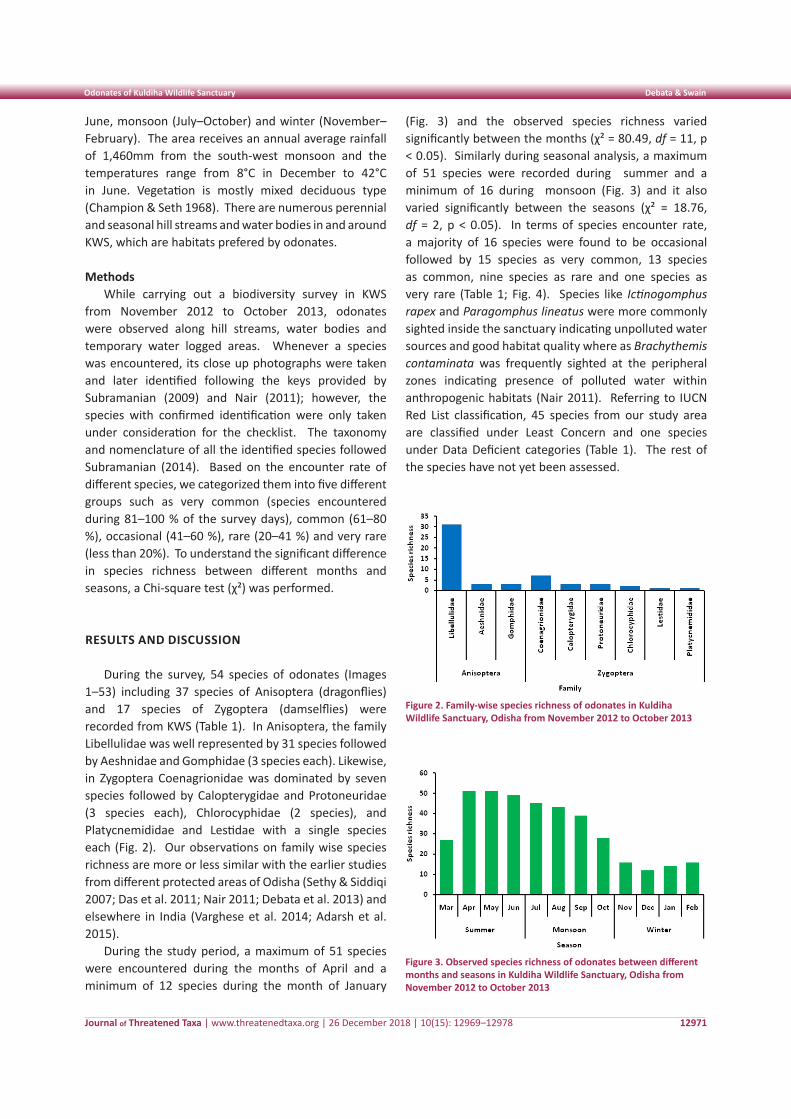
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12969–12978
Odonates of Kuldiha Wildlife Sanctuary Debata & Swain
12971
June, monsoon (July–October) and winter (November–February). The area receives an annual average rainfall of 1,460mm from the south-west monsoon and the temperatures range from 8°C in December to 42°C in June. Vegetation is mostly mixed deciduous type (Champion & Seth 1968). There are numerous perennial and seasonal hill streams and water bodies in and around KWS, which are habitats prefered by odonates.
MethodsWhile carrying out a biodiversity survey in KWS
from November 2012 to October 2013, odonates were observed along hill streams, water bodies and temporary water logged areas. Whenever a species was encountered, its close up photographs were taken and later identified following the keys provided by Subramanian (2009) and Nair (2011); however, the species with confirmed identification were only taken under consideration for the checklist. The taxonomy and nomenclature of all the identified species followed Subramanian (2014). Based on the encounter rate of different species, we categorized them into five different groups such as very common (species encountered during 81–100 % of the survey days), common (61–80 %), occasional (41–60 %), rare (20–41 %) and very rare (less than 20%). To understand the significant difference in species richness between different months and seasons, a Chi-square test (χ²) was performed.
RESULTS AND DISCUSSION
During the survey, 54 species of odonates (Images 1–53) including 37 species of Anisoptera (dragonflies) and 17 species of Zygoptera (damselflies) were recorded from KWS (Table 1). In Anisoptera, the family Libellulidae was well represented by 31 species followed by Aeshnidae and Gomphidae (3 species each). Likewise, in Zygoptera Coenagrionidae was dominated by seven species followed by Calopterygidae and Protoneuridae (3 species each), Chlorocyphidae (2 species), and Platycnemididae and Lestidae with a single species each (Fig. 2). Our observations on family wise species richness are more or less similar with the earlier studies from different protected areas of Odisha (Sethy & Siddiqi 2007; Das et al. 2011; Nair 2011; Debata et al. 2013) and elsewhere in India (Varghese et al. 2014; Adarsh et al. 2015).
During the study period, a maximum of 51 species were encountered during the months of April and a minimum of 12 species during the month of January
(Fig. 3) and the observed species richness varied significantly between the months (χ² = 80.49, df = 11, p < 0.05). Similarly during seasonal analysis, a maximum of 51 species were recorded during summer and a minimum of 16 during monsoon (Fig. 3) and it also varied significantly between the seasons (χ² = 18.76, df = 2, p < 0.05). In terms of species encounter rate, a majority of 16 species were found to be occasional followed by 15 species as very common, 13 species as common, nine species as rare and one species as very rare (Table 1; Fig. 4). Species like Ictinogomphus rapex and Paragomphus lineatus were more commonly sighted inside the sanctuary indicating unpolluted water sources and good habitat quality where as Brachythemis contaminata was frequently sighted at the peripheral zones indicating presence of polluted water within anthropogenic habitats (Nair 2011). Referring to IUCN Red List classification, 45 species from our study area are classified under Least Concern and one species under Data Deficient categories (Table 1). The rest of the species have not yet been assessed.
Figure 2. Family-wise species richness of odonates in Kuldiha Wildlife Sanctuary, Odisha from November 2012 to October 2013
Figure 3. Observed species richness of odonates between different months and seasons in Kuldiha Wildlife Sanctuary, Odisha from November 2012 to October 2013
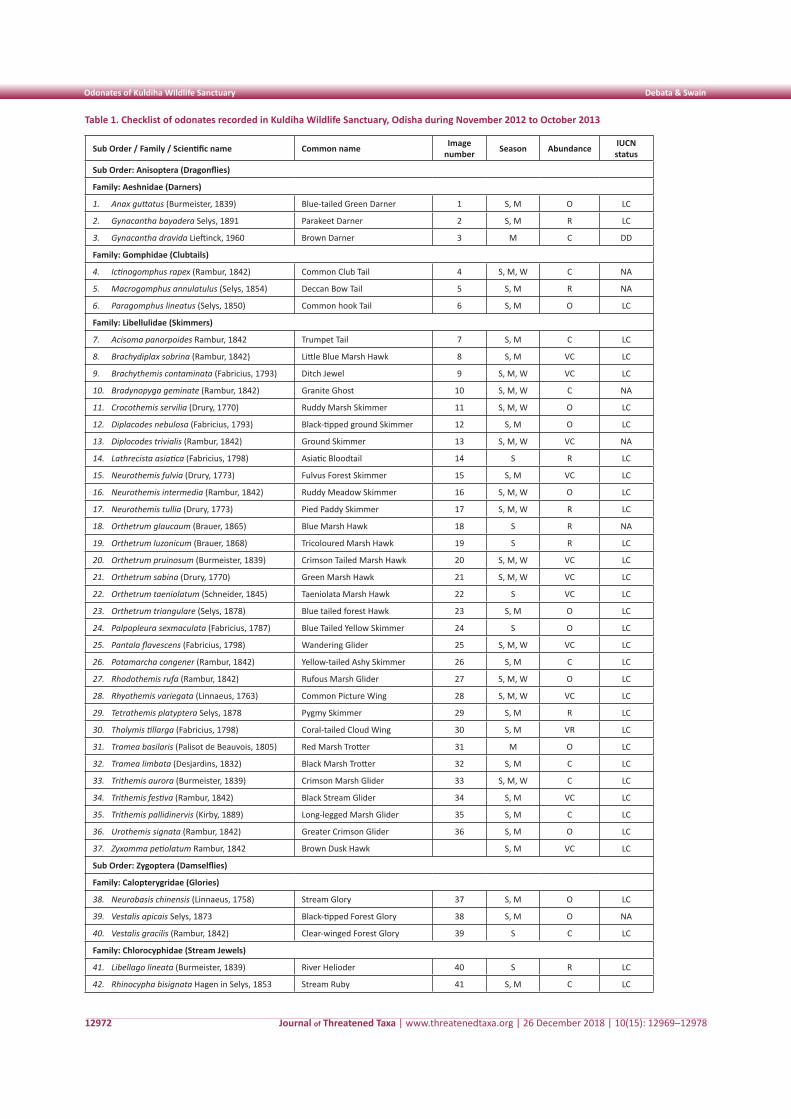
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12969–12978
Odonates of Kuldiha Wildlife Sanctuary Debata & Swain
12972
Table 1. Checklist of odonates recorded in Kuldiha Wildlife Sanctuary, Odisha during November 2012 to October 2013
Sub Order / Family / Scientific name Common name Image number Season Abundance IUCN
status
Sub Order: Anisoptera (Dragonflies)
Family: Aeshnidae (Darners)
1. Anax guttatus (Burmeister, 1839) Blue-tailed Green Darner 1 S, M O LC
2. Gynacantha bayadera Selys, 1891 Parakeet Darner 2 S, M R LC
3. Gynacantha dravida Lieftinck, 1960 Brown Darner 3 M C DD
Family: Gomphidae (Clubtails)
4. Ictinogomphus rapex (Rambur, 1842) Common Club Tail 4 S, M, W C NA
5. Macrogomphus annulatulus (Selys, 1854) Deccan Bow Tail 5 S, M R NA
6. Paragomphus lineatus (Selys, 1850) Common hook Tail 6 S, M O LC
Family: Libellulidae (Skimmers)
7. Acisoma panorpoides Rambur, 1842 Trumpet Tail 7 S, M C LC
8. Brachydiplax sobrina (Rambur, 1842) Little Blue Marsh Hawk 8 S, M VC LC
9. Brachythemis contaminata (Fabricius, 1793) Ditch Jewel 9 S, M, W VC LC
10. Bradynopyga geminate (Rambur, 1842) Granite Ghost 10 S, M, W C NA
11. Crocothemis servilia (Drury, 1770) Ruddy Marsh Skimmer 11 S, M, W O LC
12. Diplacodes nebulosa (Fabricius, 1793) Black-tipped ground Skimmer 12 S, M O LC
13. Diplocodes trivialis (Rambur, 1842) Ground Skimmer 13 S, M, W VC NA
14. Lathrecista asiatica (Fabricius, 1798) Asiatic Bloodtail 14 S R LC
15. Neurothemis fulvia (Drury, 1773) Fulvus Forest Skimmer 15 S, M VC LC
16. Neurothemis intermedia (Rambur, 1842) Ruddy Meadow Skimmer 16 S, M, W O LC
17. Neurothemis tullia (Drury, 1773) Pied Paddy Skimmer 17 S, M, W R LC
18. Orthetrum glaucaum (Brauer, 1865) Blue Marsh Hawk 18 S R NA
19. Orthetrum luzonicum (Brauer, 1868) Tricoloured Marsh Hawk 19 S R LC
20. Orthetrum pruinosum (Burmeister, 1839) Crimson Tailed Marsh Hawk 20 S, M, W VC LC
21. Orthetrum sabina (Drury, 1770) Green Marsh Hawk 21 S, M, W VC LC
22. Orthetrum taeniolatum (Schneider, 1845) Taeniolata Marsh Hawk 22 S VC LC
23. Orthetrum triangulare (Selys, 1878) Blue tailed forest Hawk 23 S, M O LC
24. Palpopleura sexmaculata (Fabricius, 1787) Blue Tailed Yellow Skimmer 24 S O LC
25. Pantala flavescens (Fabricius, 1798) Wandering Glider 25 S, M, W VC LC
26. Potamarcha congener (Rambur, 1842) Yellow-tailed Ashy Skimmer 26 S, M C LC
27. Rhodothemis rufa (Rambur, 1842) Rufous Marsh Glider 27 S, M, W O LC
28. Rhyothemis variegata (Linnaeus, 1763) Common Picture Wing 28 S, M, W VC LC
29. Tetrathemis platyptera Selys, 1878 Pygmy Skimmer 29 S, M R LC
30. Tholymis tillarga (Fabricius, 1798) Coral-tailed Cloud Wing 30 S, M VR LC
31. Tramea basilaris (Palisot de Beauvois, 1805) Red Marsh Trotter 31 M O LC
32. Tramea limbata (Desjardins, 1832) Black Marsh Trotter 32 S, M C LC
33. Trithemis aurora (Burmeister, 1839) Crimson Marsh Glider 33 S, M, W C LC
34. Trithemis festiva (Rambur, 1842) Black Stream Glider 34 S, M VC LC
35. Trithemis pallidinervis (Kirby, 1889) Long-legged Marsh Glider 35 S, M C LC
36. Urothemis signata (Rambur, 1842) Greater Crimson Glider 36 S, M O LC
37. Zyxomma petiolatum Rambur, 1842 Brown Dusk Hawk S, M VC LC
Sub Order: Zygoptera (Damselflies)
Family: Calopterygridae (Glories)
38. Neurobasis chinensis (Linnaeus, 1758) Stream Glory 37 S, M O LC
39. Vestalis apicais Selys, 1873 Black-tipped Forest Glory 38 S, M O NA
40. Vestalis gracilis (Rambur, 1842) Clear-winged Forest Glory 39 S C LC
Family: Chlorocyphidae (Stream Jewels)
41. Libellago lineata (Burmeister, 1839) River Helioder 40 S R LC
42. Rhinocypha bisignata Hagen in Selys, 1853 Stream Ruby 41 S, M C LC

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12969–12978
Odonates of Kuldiha Wildlife Sanctuary Debata & Swain
12973
Sub Order / Family / Scientific name Common name Image number Season Abundance IUCN
status
Family: Coenagrionidae (Marsh Darts)
43. Agriocnemis lecteola Selys, 1877 Milky Dartlet 42 S, M VC NA
44. Agriocnemis pygmaea (Rambur, 1842) Pygmy Dartlet 43 S, M, W VC LC
45. Amphiallagma parvum (Selys, 1876) Azure Dartlet 44 S, M O LC
46. Ceriagrion coromandelium (Fabricius, 1798) Coromandel Marshdart 45 S, M, W VC NA
47. Ischnura aurora (Brauer, 1865) Golden Dartlet 46 S, M, W C LC
48. Pseudagrion decorum (Rambur, 1842) Three lined Dart 47 S R LC
49. Pseudagrion rubriceps Selys, 1876 Saffron Faced Blue Dart 48 S C LC
Family: Lestidae (Spread Wings)
50. Lestes viridulus Rambur, 1842 Emerald Striped Spreadwing 49 M O LC
Family: Platycnemididae (Bush Darts)
51. Copera vittata Selys, 1863 Blue Bush Dart 50 S, M O LC
Family: Protoneuridae (Bamboo Tails)
52. Caconeura ramburi (Fraser, 1922) Coorg Bambootail 51 S, M VC DD
53. Disparoneura quadrimaculata (Rambur, 1842) Black-winged Bambootail 52 S, M O LC
54. Prodasineura verticalis (Selys, 1860) Black Bambootail 53 S, M C LC
S - Summer; M - Monsoon; W - Winter; VC - Very Common; C - Common; O - Occasional; R - Rare; VR - Very Rare; LC - Least Concern; DD - Data Deficient; NT - Near Threatened; NA - Not Assessed
Figure 4. Observed encountered rate of different Odonata species in Kuldiha Wildlife Sanctuary, Odisha from November 2012 to October 2013
Although KWS represents around 0.17 % of the total geographic area and 3.31 % of the total protected areas network of Odisha, it contributes around 49.09 % of the Odonata species richness of the state and 10.73 % of India. Yet, the present study gives a preliminary observation on Odonata fauna of KWS as part of multi taxa inventory. Therefore, more detailed and targeted long term studies on these lesser-known insect fauna will be useful in understanding their status and monitoring the change over time in the study area.
REFERENCES
Adarsh, C.K., R. Arunraj & P.O. Nameer (2015). Odonata (Insecta) diversity of Chinnar Wildlife Sanctuary, the southern Western Ghats, India. Journal of Threatened Taxa 7(2): 6910–6919; https://doi.org/10.11609/JoTT.o3771.6910-9
Castella, E. (1987). Larval Odonata distribution as a describer of fluvial ecosystems: the Rhone and Ain rivers, France. Advances in Odonatology 3: 23–40.
Champion, H.G. & S.K. Seth (1968). The Forest Types of India. Government of India Press, Nasik, India, 404pp.
Das, P.K., P.P. Mohapatra & A.K. Biswal (2015). Glimpses of Biodiversity of Odisha. Odisha
Biodiversity Board, Bhubaneswar, Odisha, India, 84pp.Das, S.K., H.K. Sahu & S.D. Rout (2010). Odonates of Baripada Division
of Simlipal Biosphere Reserve, including North Orissa University Campus, Orissa, India. Tiger Paper 37(2): 13–15.
Das, S.K., B. Baruah, N. Dash, S. Singhnaik & H.K. Sahu (2011). Diversity of Odonates in Nandankanan Zoological Park with range extension notes of White Dartlet (Agriocnemis pieris) in Orissa, India. Tiger Paper 38(2): 29–32
Debata, S., H.K. Sahu, S.D. Rout & R.K. Mishra (2013). An observation on Odonata diversity in Hadgarh Wildlife Sanctuary, Odisha, India. Tiger Paper 40(2): 10–13.
Fraser, F.C. & C. Dover (1922). The fauna of island in Chilika Lake - Dragonflies. Records of the Indian Museum 24(3): 303–311.
Joshi, S., P. Koparde, K.A. Subramanian & P. Roy (eds.) (2017). Odonata of India, v. 1.00. Indian Foundation for Butterflies. http:// http://www.indianodonata.org/. Electronic version accessed 22 April 2017.
Laidlaw, E.F. (1915). Fauna of the Chilika lake. No.2 Odonata. Memoirs of Indian Museum 5: 177–188.
Mitra, T.R. (2000). A note on an Odonata collection from Orissa, India. Notul Odonatol 5: 60–61.
Nair, M.V. (2011). Dragonflies & Damselflies of Orissa and Eastern India. Wildlife Organisation, Forest & Environment Department, Government of Orissa, 252pp.
Pandey, P. & A.K. Mohapatra (2017). Diversity of two families Libellulidae and Coenagrionidae (Odonata) in Regional Institute of Education Campus, Bhubaneswar, Odisha, India. Journal of

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12969–12978
Odonates of Kuldiha Wildlife Sanctuary Debata & Swain
12974
Image 1. Anax guttatus mating
Image 4. Ictinogomphus rapex Male
Image 7. Acisoma panorpoides Male
Image 10. Bradynopyga geminate Male
Image 2. Gynacantha bayadera Male
Image 5. Macrogomphus annulatulus Male
Image 8. Brachydiplax sobrina Female
Image 11. Crocothemis servilia Male
Image 3. Gynacantha dravida Male
Image 6. Paragomphus lineatus Male
Image 9. Brachythemis contaminata Male
Image 12. Diplacodes nebulosa Male
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12969–12978
Odonates of Kuldiha Wildlife Sanctuary Debata & Swain
12975
Image 13. Diplocodes trivialis Female
Image 16. Neurothemis intermedia Female
Image 19. Orthetrum luzonicum Male
Image 22. Orthetrum taeniolatum Male
Image 14. Lathrecista asiatica Female
Image 17. Neurothemis tullia Male
Image 20. Orthetrum pruinosum Mating
Image 23. Orthetrum triangulare Male
Image 15. Neurothemis fulvia Male
Image 18. Orthetrum glaucaum Male
Image 21. Orthetrum sabina Male
Image 24. Palpopleura sexmaculata Male
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata © Subrat Debata
© Subrat Debata© Arajush Payra

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12969–12978
Odonates of Kuldiha Wildlife Sanctuary Debata & Swain
12976
Image 25. Pantala flavescens Male
Image 28. Rhyothemis variegata Male
Image 31. Tramea basilaris Male
Image 34. Trithemis festiva Male
Image 26. Potamarcha congener Male
Image 29. Tetrathemis platyptera
Image 32. Tramea limbata
Image 35. Trithemis pallidinervis Male
Image 27. Rhodothemis rufa Male
Image 30. Tholymis tillarga Male
Image 33. Trithemis aurora Male
Image 36. Urothemis signata Male
© Subrat Debata
© Subrat Debata
© Subrat Debata © Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Arajush Payra

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12969–12978
Odonates of Kuldiha Wildlife Sanctuary Debata & Swain
12977
Image 37. Neurobasis chinensis Male
Image 40. Libellago lineata Male
Image 43. Agriocnemis pygmaea Male
Image 46. Ischnura aurora Male
Image 38. Vestalis apicais Male
Image 41. Rhinocypha bisignata Male
Image 44. Amphiallagma parvum Male
Image 47. Pseudagrion decorum Male
Image 39. Vestalis gracilis
Image 42. Agriocnemis lecteola Male
Image 45. Ceriagrion coromandelium Male
Image 48. Pseudagrion rubriceps Egg laying
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Udit Pratap Das
© Udit Pratap Das

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12969–12978
Odonates of Kuldiha Wildlife Sanctuary Debata & Swain
12978
Image 49. Lestes viridulus Male
Image 52. Disparoneura quadrimaculata Male
Image 50. Copera vittata Male
Image 53. Prodasineura verticalis Male Image 51. Caconeura ramburi Mating
© Subrat Debata
© Subrat Debata
© Subrat Debata
© Subrat Debata © Subrat Debata
Threatened Taxa 9(2): 9851–9857; https://doi.org/10.11609/jott.2547.9.2.9851-9857
Payra, A., S.K. Dash, A.K. Mishra, H.S. Palei, R.K. Mishra & S.D. Rout (2014). A preliminary study on Odonata (Dragonfly and Damselfly) diversity in Athagarh forest division, Odisha, India. e-planet 12(2): 43–49.
Ray, G.C. (2005). Geography of Orissa. Kitab Mahal, Cuttack, 308pp.Sajan, S.K. & P.P. Mohapatra (2014). New Record of Lesser Blue Wing
Rhyothemis triangularis Kirby, 1889 (Odonata: Libellulidae) from Odisha, India. Journal of Bombay Natural History Society 111(1): 60; https://doi.org/10.17087/bnhs/2014/v11i1/56544
Sethy, P.G.S & S.Z. Siddiqui (2007). Observations on Odonates in Simlipal Biosphere Reserve, Orissa. Zoos’ Print Journal 22(11): 2893–2894.
Srivastava, V.K. & S. Das (1987). Insecta: Odonata, pp. 135–159. In: Director (ed.). Fauna of Orissa. State Fauna Series 1 (Part 1). Zoological Survey of India Publication, Calcutta, India.
Subramanian, K.A. (2005). Dragonflies and Damselflies of Peninsular India- A Field Guide. 1st Edition. Project Lifescape Series. Indian Academy of Science, Bangaore, 118pp.
Subramanian, K.A. (2009). India- A Lifescape, Dragonflies of India - A Field Guide. Vigyan Prasar, India Offset Press, New Delhi, 180pp.
Subramanian, K.A. (2014). A Checklist of Odonata of India. Zoological Survey of India, Kolkata, 31pp.
Varghese, A.P., P.R. Nikesh & J. Mathew (2014). Odonata (Insecta) diversity of Salim Ali Bird Sanctuary and its adjacent areas in Thattekkad, Kerala, India. Journal of Threatened Taxa 6(6): 5887–5893; https://doi.org/10.11609/JoTT.o3395.5887-93
Watson, J.A.L., A.H. Arthington & D.L. Conrick (1982). Effect of sewage effluent on dragonflies (Odonata) of Bulimba Creek, Brisbane. Australian Journal of Marine and Freshwater Research 33: 517–528; https://doi.org/10.1071/MF9820517
Threatened Taxa

12979
Shor
t Co
mm
unic
atio
nDOI: https://doi.org/10.11609/jott.4102.10.15.12979-12985 | ZooBank: urn:lsid:zoobank.org:pub:57FFC118-0D59-4450-9D4B-052061CAFBC1
Editor: L.A.K. Singh, Bhubaneswar, Odisha, India. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 4102 | Received 02 March 2018 | Final received 22 November 2018 | Finally accepted 04 December 2018
Citation: Chattopadhyay, S,. S. Dey & U.S. Roy (2018). On the diversity of the vertebrate fauna (excluding fishes) of Panchet Hill (Garh Panchkot), Purulia, West Bengal, India. Journal of Threatened Taxa 10(15): 12979–12985; https://doi.org/10.11609/jott.4102.10.15.12979-12985
Copyright: © Chattopadhyay et al. 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, repro-duction and distribution by providing adequate credit to the authors and the source of publication.
Funding: University Grants Commission (Minor Research Project, Sanction No. PSW–119/13–14, ERO ID No. SKB–005, Dated: 18 March 2014), New Delhi, India.
Competing interests: The authors declare no competing interests.
Acknowledgements: This work was a part of a minor research project and the authors thankfully acknowledge the University Grants Commission, New Delhi, India for providing the necessary funds. The authors are thankful to the local inhabitants of the village adjacent to Panchet Hill for their immense and unconditional help and cooperation. The authors thankfully acknowledge the help and cooperation extended by the Director of Public Instruction, Government of West Bengal, Kolkata. Dr. Somenath Dey is thankful to Dr. P.C. Lama, Officer-in-Charge, Darjeeling Government College, Darjeeling, West Bengal, India while Dr. Utpal Singha Roy is thankful to Dr. P. Pramanik, Principal, Durgapur Government College, West Bengal for their keen interest in the present work. The authors thankfully acknowledge the help and support provided by the District Forest Officer, Panchet - I Division, Purulia.
On the diversity of the vertebrate fauna (excluding fishes) of Panchet Hill (Garh Panchkot), Purulia, West Bengal, India
Sanjib Chattopadhyay 1 , Somenath Dey 2 & Utpal Singha Roy 3
1 Panchakot Mahavidyalaya, Sarbari, Neturia, Purulia, West Bengal 723121, India2 P.G. Department of Zoology, Darjeeling Government College, 19, Lebong Cart Road, Darjeeling, West Bengal 734101, India3 Department of Zoology, Durgapur Government College, J.N. Avenue, Durgapur, Paschim Bardhaman, West Bengal 713214, India1 [email protected], [email protected], [email protected] (corresponding author)
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12979–12985
Abstract: The present study was conducted at Panchet Hill (Garh Panchkot), Purulia, West Bengal between June 2013 and May 2015. Multiple methods were used for making a consolidated checklist and comments on the relative abundance of vertebrate diversity, excluding fishes. The methods included hand capturing, extensive searches in micro habitats, opportunistic spotting and information collection from the local people. A total of 106 different vertebrate species were recorded during the study span of two years. Aves was recorded as the Class with the highest diversity (63 species) while Amphibia was recorded as the Class with the lowest diversity (9 species). Most of the species recorded during the present study belong to ‘Least Concern’ category as designated by IUCN. The Black-headed Ibis Threskiornis melanocephalus and Striped Hyaena Hyaena hyaena belong to ‘Near Threatened’ category while the White-rumped Vulture Gyps bengalensis belongs to ‘Critically Endangered’ category. The present study location is facing pressures from the usual anthropogenic interventions and needs attention from the concerned authorities.
Keywords: Amphibia, aves, biodiversity, Garh Panchkot, mammalia, Panchet Hill, Purulia, reptilia, vertebrates.
Over geological time scale biodiversity has followed the trend towards net increase; however, a marked decline in global biodiversity occurred during the late Quaternary period as a consequence of both direct and indirect human activities (Gaston & Spicer 2004). Hughes et al. (1997) reported that in tropical forests on an average 1,800 populations are being destroyed per hour while 16 million annually. An ever-increasing human population with huge demands on the natural resources have imposed a worldwide burden and consequently have depleted biological diversity. India with a burgeoning human population is no exception in this regard (Marcot & Nyberg 2005). Despite the tremendous pressure over the natural resources, India which covers about 2.4% of the world’s land area, harbours about 8% of the world’s total species (UNEP 2001). The rich tradition and culture of India since ancient times have set high values to protect its sacred

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12979–12985
Vertebrate fauna of Panchet Hill, West Bengal Chattopadhyay et al.
12980
biota (Bhagwat et al. 2005). Currently, India has about 21.34% of its geographical area classified as forest which includes 764 protected areas covering about 4.93% of the total land area (WII ENVIS 2017).
As of 2015 West Bengal has 18.96% of the state’s geographical area designated as forest of which 59.4% has been classified as reserve forest (WBFD 2017). Panchet Hill (Garh Panchkot) in West Bengal is a protected forest located at Raghunathpur sub–division of Purulia District, with the highest elevation of about 650m (Fig. 1). Few research articles are published from this area including those of Raha & Mallick (2016) and recent report on “biodiversity conservation plan of Panchet hill (Garh Panchkot)” by EMTRC (2016). Over the last few decades, a large number of studies have enlisted the diversity and distribution of vertebrate taxa from different protected areas of the country. To the best of our knowledge, however, no such studies have ever been done/ reported from Panchet Hill. This
was the primary motivation behind the present work with the objective of enlisting all the vertebrate fauna, excluding fishes, from Panchet Hill protected forest.
Materials and MethodsStudy area: Panchet Hill (23.60N & 86.70E) is a hillock
with an elevation of about 650m, and of hard rock present amidst undulating topography of laterite, gravel mixed red soil of district Purulia, West Bengal (Mandal 2012). The Damodar River marks the northern boundary of this region while Panchet Dam is located adjacent to it (Fig. 1). Prevailing environmental conditions of this region are extreme where summer temperature rises up to 400C while in winter the temperature drops down to 70C. Annual average rainfall measures about 170cm. According to the biogeographic zone given by Rodgers et al. (2002) Panchet hill lies in the bio-geographic zone 6 (Deccan Peninsula). The vegetation of the present study location is dominated by Butea monosperma
Figure 1. Map showing the study site under present investigation from Panchet hill (Garh Panchkot) of Purulia District, West Bengal, India. (Satellite image source Google Map; Panchet Hill © Utpal Singha Roy).
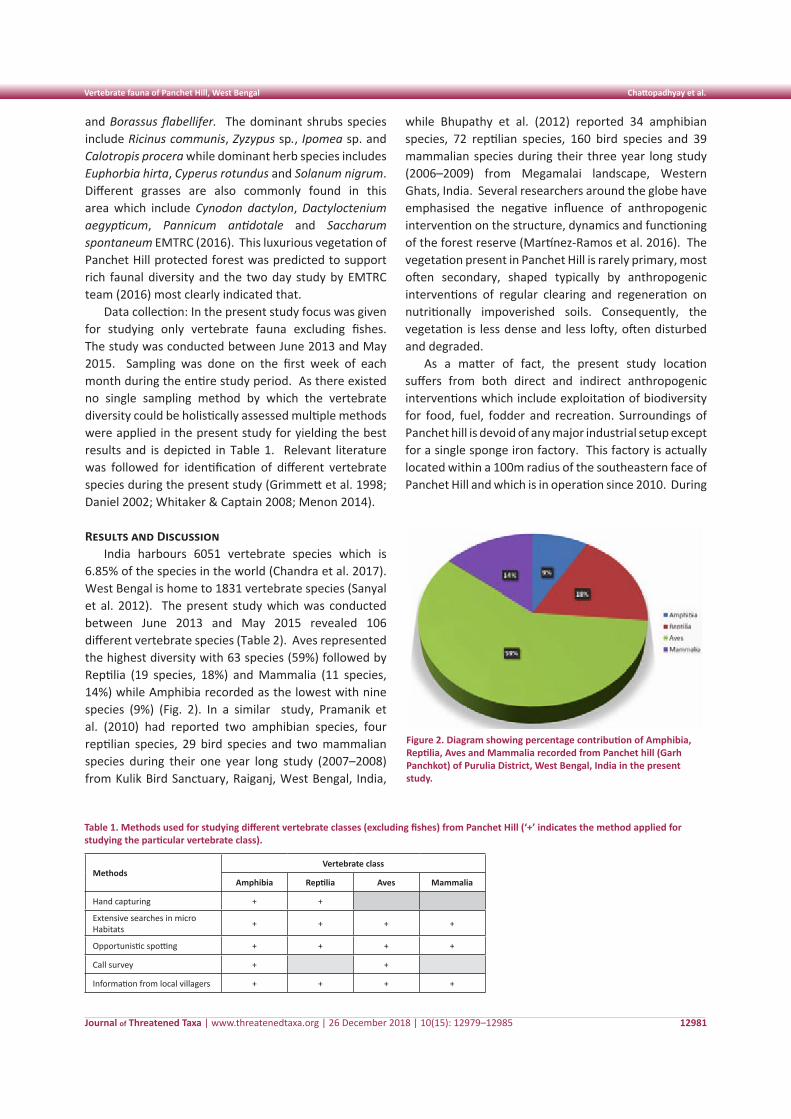
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12979–12985
Vertebrate fauna of Panchet Hill, West Bengal Chattopadhyay et al.
12981
and Borassus flabellifer. The dominant shrubs species include Ricinus communis, Zyzypus sp., Ipomea sp. and Calotropis procera while dominant herb species includes Euphorbia hirta, Cyperus rotundus and Solanum nigrum. Different grasses are also commonly found in this area which include Cynodon dactylon, Dactyloctenium aegypticum, Pannicum antidotale and Saccharum spontaneum EMTRC (2016). This luxurious vegetation of Panchet Hill protected forest was predicted to support rich faunal diversity and the two day study by EMTRC team (2016) most clearly indicated that.
Data collection: In the present study focus was given for studying only vertebrate fauna excluding fishes. The study was conducted between June 2013 and May 2015. Sampling was done on the first week of each month during the entire study period. As there existed no single sampling method by which the vertebrate diversity could be holistically assessed multiple methods were applied in the present study for yielding the best results and is depicted in Table 1. Relevant literature was followed for identification of different vertebrate species during the present study (Grimmett et al. 1998; Daniel 2002; Whitaker & Captain 2008; Menon 2014).
Results and DiscussionIndia harbours 6051 vertebrate species which is
6.85% of the species in the world (Chandra et al. 2017). West Bengal is home to 1831 vertebrate species (Sanyal et al. 2012). The present study which was conducted between June 2013 and May 2015 revealed 106 different vertebrate species (Table 2). Aves represented the highest diversity with 63 species (59%) followed by Reptilia (19 species, 18%) and Mammalia (11 species, 14%) while Amphibia recorded as the lowest with nine species (9%) (Fig. 2). In a similar study, Pramanik et al. (2010) had reported two amphibian species, four reptilian species, 29 bird species and two mammalian species during their one year long study (2007–2008) from Kulik Bird Sanctuary, Raiganj, West Bengal, India,
Table 1. Methods used for studying different vertebrate classes (excluding fishes) from Panchet Hill (‘+’ indicates the method applied for studying the particular vertebrate class).
MethodsVertebrate class
Amphibia Reptilia Aves Mammalia
Hand capturing + +
Extensive searches in microHabitats + + + +
Opportunistic spotting + + + +
Call survey + +
Information from local villagers + + + +
Figure 2. Diagram showing percentage contribution of Amphibia, Reptilia, Aves and Mammalia recorded from Panchet hill (Garh Panchkot) of Purulia District, West Bengal, India in the present study.
while Bhupathy et al. (2012) reported 34 amphibian species, 72 reptilian species, 160 bird species and 39 mammalian species during their three year long study (2006–2009) from Megamalai landscape, Western Ghats, India. Several researchers around the globe have emphasised the negative influence of anthropogenic intervention on the structure, dynamics and functioning of the forest reserve (Martínez-Ramos et al. 2016). The vegetation present in Panchet Hill is rarely primary, most often secondary, shaped typically by anthropogenic interventions of regular clearing and regeneration on nutritionally impoverished soils. Consequently, the vegetation is less dense and less lofty, often disturbed and degraded.
As a matter of fact, the present study location suffers from both direct and indirect anthropogenic interventions which include exploitation of biodiversity for food, fuel, fodder and recreation. Surroundings of Panchet hill is devoid of any major industrial setup except for a single sponge iron factory. This factory is actually located within a 100m radius of the southeastern face of Panchet Hill and which is in operation since 2010. During
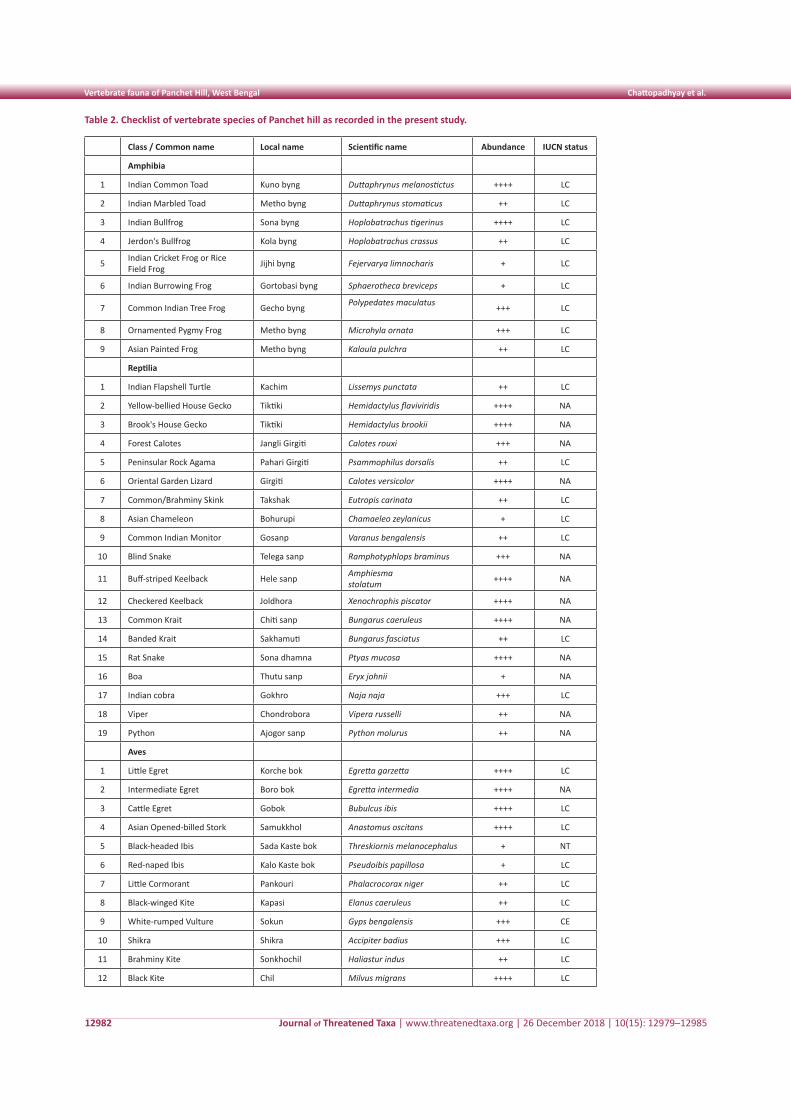
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12979–12985
Vertebrate fauna of Panchet Hill, West Bengal Chattopadhyay et al.
12982
Table 2. Checklist of vertebrate species of Panchet hill as recorded in the present study.
Class / Common name Local name Scientific name Abundance IUCN status
Amphibia
1 Indian Common Toad Kuno byng Duttaphrynus melanostictus ++++ LC
2 Indian Marbled Toad Metho byng Duttaphrynus stomaticus ++ LC
3 Indian Bullfrog Sona byng Hoplobatrachus tigerinus ++++ LC
4 Jerdon's Bullfrog Kola byng Hoplobatrachus crassus ++ LC
5 Indian Cricket Frog or Rice Field Frog Jijhi byng Fejervarya limnocharis + LC
6 Indian Burrowing Frog Gortobasi byng Sphaerotheca breviceps + LC
7 Common Indian Tree Frog Gecho byng Polypedates maculatus +++ LC
8 Ornamented Pygmy Frog Metho byng Microhyla ornata +++ LC
9 Asian Painted Frog Metho byng Kaloula pulchra ++ LC
Reptilia
1 Indian Flapshell Turtle Kachim Lissemys punctata ++ LC
2 Yellow-bellied House Gecko Tiktiki Hemidactylus flaviviridis ++++ NA
3 Brook's House Gecko Tiktiki Hemidactylus brookii ++++ NA
4 Forest Calotes Jangli Girgiti Calotes rouxi +++ NA
5 Peninsular Rock Agama Pahari Girgiti Psammophilus dorsalis ++ LC
6 Oriental Garden Lizard Girgiti Calotes versicolor ++++ NA
7 Common/Brahminy Skink Takshak Eutropis carinata ++ LC
8 Asian Chameleon Bohurupi Chamaeleo zeylanicus + LC
9 Common Indian Monitor Gosanp Varanus bengalensis ++ LC
10 Blind Snake Telega sanp Ramphotyphlops braminus +++ NA
11 Buff-striped Keelback Hele sanp Amphiesmastolatum ++++ NA
12 Checkered Keelback Joldhora Xenochrophis piscator ++++ NA
13 Common Krait Chiti sanp Bungarus caeruleus ++++ NA
14 Banded Krait Sakhamuti Bungarus fasciatus ++ LC
15 Rat Snake Sona dhamna Ptyas mucosa ++++ NA
16 Boa Thutu sanp Eryx johnii + NA
17 Indian cobra Gokhro Naja naja +++ LC
18 Viper Chondrobora Vipera russelli ++ NA
19 Python Ajogor sanp Python molurus ++ NA
Aves
1 Little Egret Korche bok Egretta garzetta ++++ LC
2 Intermediate Egret Boro bok Egretta intermedia ++++ NA
3 Cattle Egret Gobok Bubulcus ibis ++++ LC
4 Asian Opened-billed Stork Samukkhol Anastomus oscitans ++++ LC
5 Black-headed Ibis Sada Kaste bok Threskiornis melanocephalus + NT
6 Red-naped Ibis Kalo Kaste bok Pseudoibis papillosa + LC
7 Little Cormorant Pankouri Phalacrocorax niger ++ LC
8 Black-winged Kite Kapasi Elanus caeruleus ++ LC
9 White-rumped Vulture Sokun Gyps bengalensis +++ CE
10 Shikra Shikra Accipiter badius +++ LC
11 Brahminy Kite Sonkhochil Haliastur indus ++ LC
12 Black Kite Chil Milvus migrans ++++ LC
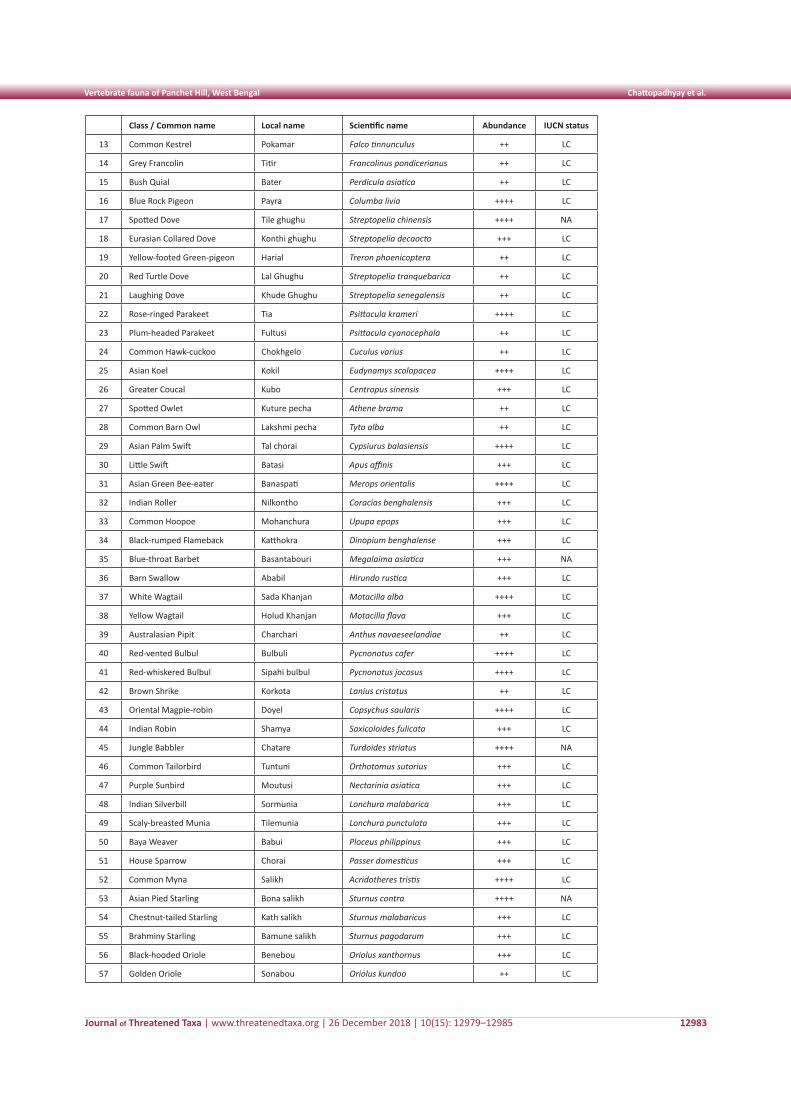
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12979–12985
Vertebrate fauna of Panchet Hill, West Bengal Chattopadhyay et al.
12983
Class / Common name Local name Scientific name Abundance IUCN status
13 Common Kestrel Pokamar Falco tinnunculus ++ LC
14 Grey Francolin Titir Francolinus pondicerianus ++ LC
15 Bush Quial Bater Perdicula asiatica ++ LC
16 Blue Rock Pigeon Payra Columba livia ++++ LC
17 Spotted Dove Tile ghughu Streptopelia chinensis ++++ NA
18 Eurasian Collared Dove Konthi ghughu Streptopelia decaocto +++ LC
19 Yellow-footed Green-pigeon Harial Treron phoenicoptera ++ LC
20 Red Turtle Dove Lal Ghughu Streptopelia tranquebarica ++ LC
21 Laughing Dove Khude Ghughu Streptopelia senegalensis ++ LC
22 Rose-ringed Parakeet Tia Psittacula krameri ++++ LC
23 Plum-headed Parakeet Fultusi Psittacula cyanocephala ++ LC
24 Common Hawk-cuckoo Chokhgelo Cuculus varius ++ LC
25 Asian Koel Kokil Eudynamys scolopacea ++++ LC
26 Greater Coucal Kubo Centropus sinensis +++ LC
27 Spotted Owlet Kuture pecha Athene brama ++ LC
28 Common Barn Owl Lakshmi pecha Tyto alba ++ LC
29 Asian Palm Swift Tal chorai Cypsiurus balasiensis ++++ LC
30 Little Swift Batasi Apus affinis +++ LC
31 Asian Green Bee-eater Banaspati Merops orientalis ++++ LC
32 Indian Roller Nilkontho Coracias benghalensis +++ LC
33 Common Hoopoe Mohanchura Upupa epops +++ LC
34 Black-rumped Flameback Katthokra Dinopium benghalense +++ LC
35 Blue-throat Barbet Basantabouri Megalaima asiatica +++ NA
36 Barn Swallow Ababil Hirundo rustica +++ LC
37 White Wagtail Sada Khanjan Motacilla alba ++++ LC
38 Yellow Wagtail Holud Khanjan Motacilla flava +++ LC
39 Australasian Pipit Charchari Anthus novaeseelandiae ++ LC
40 Red-vented Bulbul Bulbuli Pycnonotus cafer ++++ LC
41 Red-whiskered Bulbul Sipahi bulbul Pycnonotus jocosus ++++ LC
42 Brown Shrike Korkota Lanius cristatus ++ LC
43 Oriental Magpie-robin Doyel Copsychus saularis ++++ LC
44 Indian Robin Shamya Saxicoloides fulicata +++ LC
45 Jungle Babbler Chatare Turdoides striatus ++++ NA
46 Common Tailorbird Tuntuni Orthotomus sutorius +++ LC
47 Purple Sunbird Moutusi Nectarinia asiatica +++ LC
48 Indian Silverbill Sormunia Lonchura malabarica +++ LC
49 Scaly-breasted Munia Tilemunia Lonchura punctulata +++ LC
50 Baya Weaver Babui Ploceus philippinus +++ LC
51 House Sparrow Chorai Passer domesticus +++ LC
52 Common Myna Salikh Acridotheres tristis ++++ LC
53 Asian Pied Starling Bona salikh Sturnus contra ++++ NA
54 Chestnut-tailed Starling Kath salikh Sturnus malabaricus +++ LC
55 Brahminy Starling Bamune salikh Sturnus pagodarum +++ LC
56 Black-hooded Oriole Benebou Oriolus xanthornus +++ LC
57 Golden Oriole Sonabou Oriolus kundoo ++ LC

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12979–12985
Vertebrate fauna of Panchet Hill, West Bengal Chattopadhyay et al.
12984
Class / Common name Local name Scientific name Abundance IUCN status
58 Black Drongo Finge Dicrurus macrocercus ++++ LC
59 Small Blue Kingfisher Choto machranga Alcedo atthis +++ LC
60 White-breasted Kingfisher Dholabuk Machranga Halcyon smyrnensis +++ LC
61 Rufous Treepie Harichacha Dendrocitta vagabunda +++ LC
62 House Crow Kak Corvus splendens ++++ LC
63 Large-billed Crow Darkak Corvus macrorhynchos ++ LC
Mammalia
1 Indian Hare Khorgosh Lepus nigricollis ++ LC
2 Indian Crested Porcupine Sojaru Hystrix indica + LC
3 Northern Plains Gray Langur Hanuman Semnopithecus entellus +++ LC
4 Indian Grey Mongoose Neul Herpestes edwardsii ++ LC
5 Common Palm Civet Gondhogokul Paradoxurus hermaphroditus + LC
6 Indian Flying Fox Badur Pteropus giganteus ++++ LC
7 Indian Pygmy Bat Chamchike Pipistrellus tenuis ++++ LC
8 Common Palm Squirrel Kathbirali Funambulus palmarum ++++ LC
9 House Rat Idur Rattus rattus +++ LC
10 House Mouse Nengti idur Mus musculus +++ LC
11 House Shrew Chucho Suncus murinus +++ LC
12 Indian Mole-rat Metho idur Bandicota bengalensis +++ LC
13 Striped Hyaena Lakra Hyaena hyaena + NT
14 Bengal Fox Khaksial Vulpes bengalensis ++ LC
15 Jungle Cat Bonbiral Felis chaus + LC
Abbreviations used: Relative abundance expressed as ‘+’ means less abundant; ‘++’ means more abundant and so on. CE - Critically Endangered, LC - Least Concern, NA - This taxon has not yet been assessed for the IUCN Red List, NT - Near Threatened.
Image 1. Pollutants released from sponge iron factory located within 100m radius of southeastern face of Panchet Hill (A–C) and black soot over plant leaves (D) as found in the present study.© Utpal Singha Roy

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12979–12985
Vertebrate fauna of Panchet Hill, West Bengal Chattopadhyay et al.
12985
the present study pollutants released from the factory caused the ground to be covered with ash and slag while flying ash was found to leave black soot over plant leaves even at the heaight above 50m (Image 1). EMTRC (2016) have reported occurrence of pollution resistant invasive plant species like Lantana camara, Parthenium sp., and Tridax procumbens with an overall decrease in native plant species diversity from the polluted site. Lower vertebrate diversity was noted from the polluted southeastern face of Panchet Hill in comparison to all the other sites as well. Most of the species recorded during the present study belong to ‘Least Concern’ category as designated by IUCN (2017); however, Black-headed Ibis Threskiornis melanocephalus and Striped Hyaena Hyaena hyaena belong to ‘Near Threatened’ category while White-rumped Vulture Gyps bengalensis belongs to ‘Critically Endangered’ category. Striped Hyaena Hyaena hyaena was observed only once during the present study but reports by local villagers suggest that they were spotted at least five times during the present study duration. White-rumped Vulture Gyps bengalensis was recorded three times during the present study.
Our record of 106 different vertebrate species from Panchet Hill (Garh Panchkot), Purulia in West Bengal forms the base line information. Additional studies including multiple plant and animal taxa will enrich our knowledge about diversity of wild species from this ecoregion. Such studies will help in assessing the spatial and temporal distribution pattern and population status, which are vital for preparing a conservation plan to support sustainable development.
References
Bhagwat, S.A., C.G. Kushalappa, P.H. Williams & N.D. Brown (2005). A landscape approach to biodiversity conservation of sacred groves in the Western Ghats of India. Conservation Biology 19: 1853–1862.
Bhupathy, S., G. Srinivas, N. Sathishkumar, M. Murugesan, S. Babu, R. Suganthasakthivel & P. Sivakumar (2012). Diversity and conservation of selected biota of the Megamalai landscape, Western Ghats, India. Current Science 102(4): 590–595.
Chandra, K., S. Sheela & D. Das (2017). Animal Discoveries 2016: New species and new records. Zoological Survey of India, Ministry of
Environment, Forest and Climate Change, Kolkata, 108pp.Daniel, J.C. (2002). The Book of Indian Reptiles and Amphibians. 1st
Edition. Oxford University Press, Bombay, 252pp.EMTRC (2016). Biodiversity conservation plan of Panchet hill
(Garh Panchkot): Near Village: Parbatpur, Tehsil: Raghunathpur, District: Purulia, West Bengal. EMTRC Consultants Pvt. Ltd., Delhi & PG Department of Conservation Biology Durgapur Government College, West Bengal, 41pp; http://environmentclearance.nic.in/writereaddata/FormB/EC/Additional_Report/04022017QJ7F473FBiodiversityConservationplan.pdf
Gaston, K.J. & J.I. Spicer (2004). Biodiversity: An Introduction. 2nd Edition. Blackwell Publishing, Oxford, UK, 191pp.
Grimmett, R., C. Inskipp & T. Inskipp (1998). Birds of the Indian Subcontinent. Oxford University Press, Delhi, 888pp.
Hughes, J.B., G.C. Daily & P.R. Ehrlich (1997). Population diversity: its extent and extinction. Science 278: 689–692.
IUCN (2017). The IUCN Red List of Threatened Species. Version 2017–3. <www.iucnredlist.org>. Downloaded on 26 February 2018.
Mandal, S.K. (2012). Survey of wetlands in Puruliya District, West Bengal, with special emphasis on their macrophytes. PhD Thesis. Department of Botany, The University of Burdwan, 377pp.
Marcot, B.G. & J.B. Nyberg (2005). The future of forest biodiversity conservation amidst development: reflection and vision. International Forestry Review 7: 21.
Martínez-Ramos, M., I.A.O. Rodrígueza, D. Piñerob, R. Dirzoc & J. Sarukhán (2016). Anthropogenic disturbances jeopardize biodiversity conservation within tropical rainforest reserves. Proceedings of the National Academy of Sciences 113(19): 5323–5328.
Menon, V. (2014). Indian Mammals - A Field Guide. Hachette Book Publishing Indian Pvt. Ltd., 528pp.
Pramanik, A.K., K.B. Santra & C.K. Manna (2010). Abundance and diversity of plants and animals in the Kulik bird sanctuary, Raiganj, West Bengal, India. Journal of Biodiversity 1(1): 13–17.
Raha, S. & H. Mallick (2016). Piscicidal and fish stupefying plants used by tribal people living around Panchakot hills, Purulia, West Bengal, India. International Journal of Life Sciences 5(1): 27–30.
Rodgers, W.A., H.S. Panwar & V.B. Mathur (2002). Wildlife Protected Area Network in India: A Review (Executive summary). Wildlife Institute of India. Dehradun, 44pp.
Sanyal, A.K., J.R.B. Alfred, K. Venkataraman, S.K. Tiwari & S. Mitra (2012). Status of biodiversity of West Bengal. Zoological Survey of India, Kolkata, 969pp+35pls.
UNEP (2001). India: state of the environment 2001. United Nations Environment Programme, Regional Resource Centre for Asia and the Pacific, Pathumthani, Thailand.
WBFD (2017). West Bengal Forest Department; http://www.westbengalforest.gov.in/history.php. Accessed on 29.09.2017
Whitaker, R. & A. Captain (2008). Snakes of India - The Field Guide. Draco Books, Chennai, 385pp.
WII ENVIS (2017). ENVIS Centre on Wildlife & Protected Areas; http://www.wiienvis.nic.in/Database/Protected_Area_854.aspx. Accessed on 29.09.2018
Threatened Taxa

First record of rare Furry Lobster from Arabian Sea Idreesbabu et al.
12986
Shor
t Co
mm
unic
atio
n
DOI: https://doi.org/10.11609/jott.4166.10.15.12986-12989 | ZooBank: urn:lsid:zoobank.org:pub:3EE67903-F0B3-4643-8850-455D3ECE9A90
Editor: Kareen Schnabel, National Institute of Water & Atmospheric Research Ltd. (NIWA), New Zealand. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 4166 | Received 30 March 2018 | Final received 05 November 2018 | Finally accepted 23 November 2018
Citation: Idreesbabu, K.K., C.P.R. Shanis & S. Sureshkumar (2018). First record of the rare Furry Lobster Palinurellus wieneckii (De Man, 1881) (Decapoda: Palinu-ridae) from the Arabian Sea. Journal of Threatened Taxa 10(15): 12986–12989; https://doi.org/10.11609/jott.4166.10.15.12986-12989
Copyright: © Idreesbabu et al. 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduc-tion and distribution by providing adequate credit to the authors and the source of publication.
Funding: The present study was supported as part of its ongoing Marine Biodiversity Documentation program at the Department of Science & Technology, Lakshadweep Administration, India and a grant (Grant-In-Aid General-39).
Competing interests: The authors declare no competing interests.
Acknowledgments: The authors are indebted to Mr. Saheer M.C. and Mr. Khaleel C.K. who assisted in collecting the lobster specimens. The authors would like to sincerely thank the administration of the Department of Science & Technology, Union Territory of Lakshadweep, for providing permission to carry out this study We are thankful to Dr. Peter K.L. Ng and Mr. Lee Kong Chian from the Natural History Museum, National University of Singapore, for their support in confirming the identification of the species and for sending valuable publications required for the preparation of the manuscript. The authors gratefully acknowledge the anonymous reviewer for the critical comments, viich substantially improved the quality of this article.
First record of the rare Furry Lobster Palinurellus wieneckii (De Man, 1881) (Decapoda: Palinuridae) from the Arabian Sea
K.K. Idreesbabu 1 , C.P. Rajool Shanis 2 & S. Sureshkumar 3
1 Department of Science and Technology, Kavaratti, Union Territory of Lakshadweep 682555, India 2 PG and Research Department of Aquaculture and Fishery Microbiology, MES Ponnani College, Ponnani,
Kerala 679586, India3 School of Ocean Science and Technology, Kerala University of Fisheries and Ocean Studies, Panangad, Kochi,
Kerala 682506, India1 [email protected] (corresponding author), 2 [email protected], 3 [email protected]
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12986–12989
Furry Lobster or Coral Lobster of the genus Palinurellus Von Martens, 1878 belonging to the family Palinuridae Latreille, 1802 was recorded from the Indo-West Pacific and the western Atlantic. It is rare throughout its range and descriptions were typically based on only a few specimens. The numerous short setae covering its body give the animal its common name, Furry Lobster. It is comparatively smaller in size than other palinurids and
Abstract: Two female specimens of the Furry Lobster Palinurellus wieneckii (De Man, 1881) with a total length of 118mm and 114mm, respectively, were obtained from the coral reefs off Kavaratti Island, Laccadive Islands, west of India. Only two species are currently recognized in this genus, which were described from a small number of specimens. As P. wieneckii is very rare, the present report from the Lakshadweep Archipelago provides a valuable new distribution point, which is the first record for the Arabian Sea. Illustrations and photographs are provided for this rare lobster.
Keywords: Distribution, taxonomy, Indian Ocean, Lakshadweep, Laccadive Islands.
its systematic placement was uncertain until recently. Due to its peculiar appearance, the genus Palinurellus was previously regarded as belonging to a separate family, the Synaxidae Bate, 1888. Recent phylogenetic analyses using molecular tools, however, showed Synaxidae to be an invalid family and, subsequently, the genus Palinurellus was placed in the family Palinuridae (Holthuis 1966; Palero et al. 2009; Tsang et al. 2009; Chan 2010; Chien et al. 2013).
Only two species are currently recognized in the genus Palinurellus, P. gundlachi (Von Martens, 1878) from the western Atlantic and P. wieneckii (De Man, 1881) from the Indo-West Pacific (Chan 2010). The definitions of these two species, however, remain somewhat unclear because of the limited number of specimens available (Holthuis 1966). We report P. wieneckii for the first time from the Arabian Sea and the entire Indian coastline, providing an intermediate report of the species in the wider Indo-West Pacific.
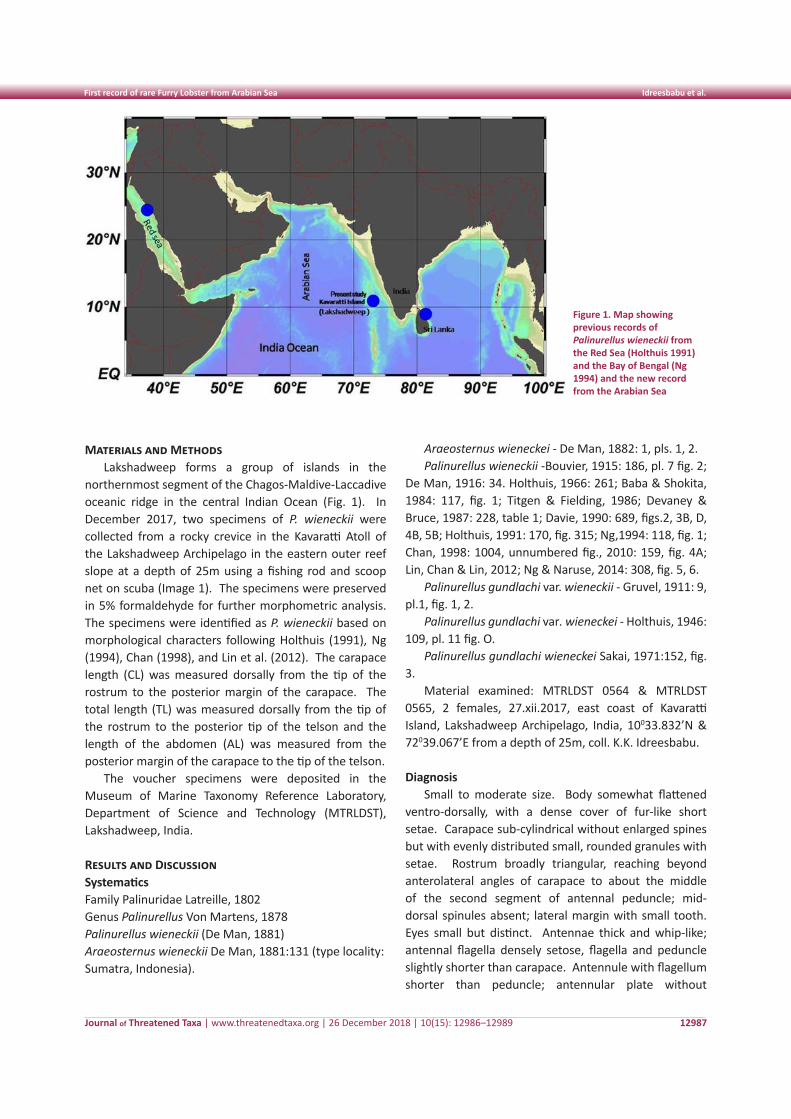
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12986–12989
First record of rare Furry Lobster from Arabian Sea Idreesbabu et al.
12987
Materials and MethodsLakshadweep forms a group of islands in the
northernmost segment of the Chagos-Maldive-Laccadive oceanic ridge in the central Indian Ocean (Fig. 1). In December 2017, two specimens of P. wieneckii were collected from a rocky crevice in the Kavaratti Atoll of the Lakshadweep Archipelago in the eastern outer reef slope at a depth of 25m using a fishing rod and scoop net on scuba (Image 1). The specimens were preserved in 5% formaldehyde for further morphometric analysis. The specimens were identified as P. wieneckii based on morphological characters following Holthuis (1991), Ng (1994), Chan (1998), and Lin et al. (2012). The carapace length (CL) was measured dorsally from the tip of the rostrum to the posterior margin of the carapace. The total length (TL) was measured dorsally from the tip of the rostrum to the posterior tip of the telson and the length of the abdomen (AL) was measured from the posterior margin of the carapace to the tip of the telson.
The voucher specimens were deposited in the Museum of Marine Taxonomy Reference Laboratory, Department of Science and Technology (MTRLDST), Lakshadweep, India.
Results and Discussion SystematicsFamily Palinuridae Latreille, 1802Genus Palinurellus Von Martens, 1878Palinurellus wieneckii (De Man, 1881) Araeosternus wieneckii De Man, 1881:131 (type locality: Sumatra, Indonesia).
Araeosternus wieneckei - De Man, 1882: 1, pls. 1, 2.Palinurellus wieneckii -Bouvier, 1915: 186, pl. 7 fig. 2;
De Man, 1916: 34. Holthuis, 1966: 261; Baba & Shokita, 1984: 117, fig. 1; Titgen & Fielding, 1986; Devaney & Bruce, 1987: 228, table 1; Davie, 1990: 689, figs.2, 3B, D, 4B, 5B; Holthuis, 1991: 170, fig. 315; Ng,1994: 118, fig. 1; Chan, 1998: 1004, unnumbered fig., 2010: 159, fig. 4A; Lin, Chan & Lin, 2012; Ng & Naruse, 2014: 308, fig. 5, 6.
Palinurellus gundlachi var. wieneckii - Gruvel, 1911: 9, pl.1, fig. 1, 2.
Palinurellus gundlachi var. wieneckei - Holthuis, 1946: 109, pl. 11 fig. O.
Palinurellus gundlachi wieneckei Sakai, 1971:152, fig. 3.
Material examined: MTRLDST 0564 & MTRLDST 0565, 2 females, 27.xii.2017, east coast of Kavaratti Island, Lakshadweep Archipelago, India, 10033.832’N & 72039.067’E from a depth of 25m, coll. K.K. Idreesbabu.
DiagnosisSmall to moderate size. Body somewhat flattened
ventro-dorsally, with a dense cover of fur-like short setae. Carapace sub-cylindrical without enlarged spines but with evenly distributed small, rounded granules with setae. Rostrum broadly triangular, reaching beyond anterolateral angles of carapace to about the middle of the second segment of antennal peduncle; mid-dorsal spinules absent; lateral margin with small tooth. Eyes small but distinct. Antennae thick and whip-like; antennal flagella densely setose, flagella and peduncle slightly shorter than carapace. Antennule with flagellum shorter than peduncle; antennular plate without
Figure 1. Map showing previous records of Palinurellus wieneckii from the Red Sea (Holthuis 1991) and the Bay of Bengal (Ng 1994) and the new record from the Arabian Sea

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12986–12989
First record of rare Furry Lobster from Arabian Sea Idreesbabu et al.
12988
stridulating organ. All walking legs without pincers; first pair setose and much more massive than others. First pleopod present. Abdomen and tail fan robust; posterior half of tail fan soft, flexible; dorsal surface of abdomen setose with rounded tubercles, lined with a longitudinal low smooth keel along dorsal midline; transverse groove absent.
Size: TL about 200mm, corresponding to CL of about 80mm (Holthuis 1991; Chan 1998). The TL of specimens collected from Lakshadweep were 118mm and 114mm, CL were 53mm and 47.8mm, and AL were 62mm and 61mm. Carapace was partially damaged in one specimen.
Colouration: Uniformly bright orange or orange-red. Eyes dark brown as reported by Ng & Naruse (2014) (Fig. 2).
Distribution: Widely distributed in the Indo-West Pacific. The species was reported from Natal in South Africa, Mauritius, Thailand, Vietnam, Malaysia, Indonesia, Papua New Guinea, the Solomon Islands, the Ryukyu Islands in Japan, the Caroline Islands, Guam, the Marshall Islands, New Caledonia, Hawaii, the Tuamotu Islands in French Polynesia, and Australia (Devaney & Bruce 1987; Holthuis 1991; Ng 1994; Nguyen & Pham 1995; Chan 1998; Debelius 1999; Paulay et al. 2003: Ng & Naruse 2014). It was also reported from Sri Lanka and the Red
Sea (Holthuis 1991; Ng 1994; Chan 2010). Usually, it is associated with coral reefs at depth ranges of 9–27 m and is probably nocturnal, inhabiting deep caves (Holthuis 1991; Chan 1998).
Remarks The diagnostic characters to differentiate between
the two species of Palinurellus are not well-defined, partly due to the rarity of these lobsters (Holthuis 1991). Biogeographically, the two species are separated as Palinurellus wieneckii is found in the Indo-West Pacific while P. gundlachi occurs in the western Atlantic. Several carcinologists (Gruvel 1911; Holthuis 1946; Sakai 1971) treated P. wieneckii as a subspecies. The carapace is sub-cylindrical with evenly distributed, small, and rounded granules with setae in P. wieneckii but is long and rounded with short setae and rounded nodules in P. gundlachi. In P. wieneckii, the rostrum is described as broadly triangular (Lin et al. 2012), reaching beyond the anterolateral angles of the carapace and while same is described as a small, triangular rostrum between
Image 1. Female specimen of Palinurellus wieneckii (De Man, 1881) collected off Kavaratti Island (MTRLDST 0564). a - dorsal view, b - ventral view
Figure 2. Palinurellus wieneckii (De Man, 1881), female (MTRLDST 0564). a - dorsal view of carapace, b - ventral view of antenna, antennules with peduncles, c - dorsal view of abdominal segments d - dorsal view of lateral carapace and pereopods

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12986–12989
First record of rare Furry Lobster from Arabian Sea Idreesbabu et al.
12989
the eyes in P. gundlachi (Williams & Williams 2010). The supra-orbital spine is prominent and pointed in P. gundlachi but is inconspicuous in P. wieneckii (De Man 1916; Holthuis 1946). According to these characters, the material examined here matches the diagnosis of P. wieneckii (Fig. 2).
Holthuis (1966) observed that the pleopods on the first abdominal somite are generally present in females but absent in males, though this character appears to be variable. In the present study, the specimens collected from the Arabian Sea were females and had pleopods on the first abdominal somite. The transverse grove is absent in the abdominal somites, which is prominent in the genus Palinurus as reported by Groeneveld et al. (2006).
There were no previous records of this species from the Arabian Sea. The record provided here fills a gap in the known distribution range of P. wieneckii based on collections in the atolls of the Lakshadweep Archipelago in the, north-central Indian Ocean, documenting the occurrence and distribution of the genus Palinurellus from the Indian waters and the Arabian Sea. The present observation confirms its intermediary distribution of the species between the eastern Indian Ocean and the Red Sea.
References
Baba, K. & S. Shokita (1984). Palinurellus wieneckii, a rare species of spiny lobster found in Okinawa-Jima of the Ryukyus (Cmstacea: Decapoda: Palinuddae). Galaxea 3: 117–118.
Bate, C.S. (1881). On Synaxes, a new genus of Crustacea. Annals and Magazine of Natural History 7(5): 220–228.
Bate, C.S. (1888). Report on the Crustacea Macrura collected by H.M.S. Challenger during the years 1873–76, pp. i–xc. In: Thomson, C. & J. Murray (eds.). Report on the Scientific Results of the Voyage of H.M.S. Challenger during the Years 1873–76. Neill & Company, Edinburgh, xc+942pp.
Chan, T.Y. (1998). Lobsters, pp. 973–1043. In: Carpenter, K.E. & V.H. Niem (eds.) FAO Species Identification Guide for Fishery Purposes. The Living Marine Resources of the Western Central Pacific, Vol. 2. Cephalopods, Crustaceans, Holothurians and Sharks. FAO, Rome, 1396pp.
Chan, T.Y. (2010). Annotated checklist of the world’s marine lobsters (Crustacea: Decapoda: Astacidea, Glypheidea, Achelata, Polychelida). Raffles Bulletin of Zoology, Supplement 23: 153–181.
Chien, H.Y., L. Yuan, C. Zhaoxia & T.Y. Chan (2013). Complete mitochondrial genome of the furry lobster Palinurellus wieneckii (De Man, 1881) (Decapoda, Achelata, Palinuridae). Mitochondrial DNA 25(4): 295–297; https://doi.org/10.3109/19401736.2013.796521
Davie, P.J. (1990). A new genus and species of marine crayfish, Palibythus magnificus, and new records of Palinurellus (Decapoda, Palinuridae) from the Pacific Ocean. Invertebrate Taxonomy 4(4): 685–695.
De Man, J.G. (1881). Carcinological studies in the Leyden Museum, No. 1. Notes from the Leyden Museum 3: 121–144.
De Man, J.G. (1916). The Decapoda of the Siboga expedition, part III: families Eryonidae, Palinuridae, Scyllaridae and Nephropsidae. Siboga Expedition Monograph 39(a2): 1–122.
Debelius, H. (1999). Crustacea Guide of the World: Shrimps, Crabs, Lobsters, Mantis Shrimps, Amphipods. IKAN, Frankfurt, 321pp.
Devaney, D.S. & A.J. Bruce (1987). Crustacea Decapoda (Penaeidea, Stenopodidea, Caridea, and Palinura) of Enewetak atoll, pp221–233. In: Devaney, D.M., E.S. Reese, B.L. Burch & P. Helfrich (eds.). The Natural History of Enewetak Atoll, Vol. II. Biogeography and Systematics. U.S. Department of Energy, Oak Ridge, 348pp.
Groeneveld, J.C., C.L. Griffiths & A.P. van Dalsen (2006). A new species of Spiny Lobster, Palinurus barbarae (Decapoda, Palinuridae) from Walters Shoals on the Madagascar Ridge. Crustaceana 79(7): 821–833.
Gruvel, A. (1911). Contribution à l’étude systématique des Palinuridae. Comptes Rendus de l’ Académie des Sciences, Paris 152: 1350–1352.
Holthuis, L.B. (1946). Biological results of the Snellius expedition, XIV. The Decapoda Macrura of the Snellius Expedition, I. The Stenopodidae, Nephropsidae, Scyllaridae and Palinuridae. Temminckia 7: 1–178.
Holthuis, L.B. (1966). On spiny lobsters of the genera Palinurellus, Linuparus and Puerulus (Crustacea, Decapoda, Palinuridae). Symposium on Crustacea, Marine Biological Association of India 1: 260–278.
Holthuis, L.B. (1991). Marine Lobsters of the World: An Annotated and Illustrated Catalogue of Species of Interest to Fishes known to date, FAO Fisheries Synopsis No. 125, Vol. 13. FAO, Rome, 292pp.
Latreille, P.A. (1802). Histoire naturelle, Générale et particulière des Crustacés et des Insectes. Ouvrage faisant suite à l’histoire naturelle Générale et particulière, composée par Leclerc de Buffon, et rédigée par C.S. Sonnini, membre de plusieurs societies savantes. Familles naturelles des Genres 3. F. Du Fart, Paris, 467pp.
Lin, C.C., T.Y. Chan & C.W. Lin (2012). First record of the rare lobster Palinurelles wieneckii (de Man, 1881) (Decapoda, Achelata, Palinuridae) from Taiwan. Crustaceana 85(11): 1385–1391.
Ng, P.K.L. (1994). First record of the synaxid lobster, Palinurellus wieneckii (de Man, 1881) (Crustacea, Decapoda, Palinuridae) from Sri Lanka. Journal of South Asian Natural History 1(1): 117–118.
Ng, P.K.L. & T. Naruse (2014). The lobsters of Christmas Island and Cocos (Keeling) Island, with new records of Palinurellus wieneckii (De Man, 1881) and Enoplometopus viigtmanni Türkay, 1989 (Crustacea: Decapoda: Palinuridae, Scyllaridae, Enoplometopidae). Raffles Bulletin of Zoology, Supplement 30: 305–312.
Nguyen, V.C. & T.D. Pham (1995). Checklist of Marine Shrimps and Lobster in Vietnam. Science & Technics Publishing House, Nhatrang, 170pp.
Palero, F., K.A. Crandall, P. Abelló, E. Macpherson & M. Pascual (2009). Phylogenetic relationships between spiny, slipper and coral lobsters (Crustacea, Decapoda, Achelata). Molecular Phylogenetics and Evolution 50(1): 152–162; https://doi.org/10.1016/j.ympev.2008.10.003
Paulay, G., R. Kropp, P.K.L. Ng & L.G. Eldredge (2003). The crustaceans and pycnogonids of the Mariana Islands. Micronesica 35–36: 456–513.
Sakai, T. (1971). Notes from the carcinological fauna of Japan (IV). Research on Crustacea 4.5: 138–156.
Titgen, R.H. & A. Fielding (1986). Occurrence of Palinurellus wieneckii (deMan, 1881) in the Hawaiian Islands (Decapoda, Palinura: Synaxidae). Journal of Crustacean Biology 6(2): 294–296.
Tsang, L.M., T.Y. Chan, M.K. Cheung & K.H. Chu (2009). Molecular evidence for the southern hemisphere origin and deep-sea diversification of spiny lobsters (Crustacea: Decapoda: Palinuridae). Molecular Phylogenetics and Evolution 51(2): 304–311.
Von Martens, E. (1878). Ueber einige Crustaceen und Mollusken, welche das zoologische Museum in letzter Zeit erhalten hat. Sitzungsgberichte der Gesellschaft naturforschender Freunde zu Berhn 1878: 131–135.
Williams, E.H., Jr. & L. Williams (2010). First record of the Caribbean Furry Lobster, Palinurellus gundlachi (Decapoda, Synaxidae) in Puerto Rico and as a food item of the Red Hind, Epinephelus guttatus (Perciformes, Serranidae). Crustaceana 83(7): 893–895.
Threatened Taxa

12990
Shor
t Co
mm
unic
atio
n
LOGOs
DOI: https://doi.org/10.11609/jott.3935.10.15.12990-12994 | ZooBank: urn:lsid:zoobank.org:pub:3C6C9BAF-F36F-493E-A2F6-FC68979D5426
Editor: B.K. Agarwala, Tripura University, West Tripura, India. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 3935 | Received 02 December 2017 | Final received 29 October 2018 | Finally accepted 05 December 2018
Citation: Khadakkar, S.S., A.D. Tiple & A.M. Khurad (2018). Description of life stages of dung beetle Scaptodera rhadamistus (Fabricius, 1775) (Coleoptera: Scarabaei-dae: Scarabaeinae) with notes on nesting and biology. Journal of Threatened Taxa 10(15): 12990–12994; https://doi.org/10.11609/jott.3935.10.15.12990-12994
Copyright: © Khadakkar et al. 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: None.
Competing interests: The authors declare no competing interests.
Acknowledgements: We are thankful to Dr. M. M. Rai, Cordinator and Dr. M. K. Rathod, Scientist, Centre for Sericulture and Biological Pest Management Research, Nagpur for providing necessary facilities to conduct the following studies and encouragement. The authors are also thankful to State Biodiversity Board, Maharashtra for providing permission to collect scarab beetles. First author acknowledges the help of Dr Sandhya Kranthi, Head of Plant Protection division, and Dr. Vishlesh Nagrare, Principle Scientist, Central Institute for Cotton Research, Nagpur, MS for permitting use of Binoculor for morphometric studies.
Description of life stages of dung beetle Scaptodera rhadamistus (Fabricius, 1775) (Coleoptera:
Scarabaeidae: Scarabaeinae) with notes on nesting and biology
Suvarna S. Khadakkar 1 , Ashish D. Tiple 2 & Arun M. Khurad 3
1,3 Centre for Sericulture and Biological Pest Management Research (CSBR), RTM Nagpur University, North Ambazari Road, Nagpur, Maharashtra 440022, India
2 Department of Zoology, Vidyabharti College, Seloo, Wardha, Maharashtra 442104, India1 [email protected] (corresponding author), 2 [email protected], 3 [email protected]
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12990–12994
Beetles belonging to family Scarabaeidae are commonly called as ‘Scarabs’ and their larvae are known as white grubs. Arrow (1931) provided detailed account of Indian Scarabaeidae. The monotypic Scaptodera rhadamistus (Fabricius, 1775), was previously cited as Liatongus (Paraliatongus) Reitter under tribe Oniticellini of subfamily Scarabaeinae (Hanski & Cambefort 1991; Philips 2016). Larvae of different stages and adults
forage by clearing excrement (Arrow 1931). Adult males of S. rhadamistus are attractive owing to the coloration and structure of pronotum.
Much of the literature available relating to scarab beetles are on adult taxonomy. Information regarding their immature forms and nest-building behaviour is deficient (Ritcher 1966; Veeresh 1980; Sreedevi & Tyagi 2014). Studies on natural history of dung beetles of the subfamily Scarabaeinae lack the information on S. rhadamistus (Halffter & Matthews 1966). The objective of this study is to present an account of larval morphology and nest-building behavior of S. rhadamistus, a commonly found scarab in central Indian region, based on a study conducted in and around Nagpur-Wardha forest areas.
Species diagnosis: In life, adults are yellowish-orange with metallic green colored patches present on dorsal and lateral regions, elongate, oval; 13–15 mm in length and 6–8 mm in width. Males have a prominent pronotum with elevated margins forming deep cavity at
Abstract: Immature stages of Scaptodera rhadamistus (Fabricius) are described for the first time along with notes on nidification and biology. The larvae differ from other Scarabaeinae species in the structure of raster on tenth sternum with two irregular bunches of serrations ventrally one on either half. Pupae with pronotum transverse having rounded margins resemble adults, and consist of four lateral, single caudal and single pteronotal support projection. Adult males and females differ in coloration, structure of pronotum, presence of spine like process on mesosternum and, in the structure of male and female genitalia.
Keywords: Description, immatures, nesting, scarab beetle, Scaptodera rhadamistus.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12990–12994
Life stages of Scaptodera rhadamistus Khadakkar et al.
12991
middle, anteriorly forming a short process that reflexes backwards distinguishing this species from other species of the same genus. Males and females differ in size and structure of pronotum, and in structure of spine-like process on mesosternum. Females are smaller, with simple pronotum.
The species is native to Oriental region (India, Laos, Sri Lanka, and Thailand) (Schoolmeesters 2017). In India, the species occurs in the states of Gujarat, Madhya Pradesh, Maharashtra and Tamil Nadu (Chandra & Ahirwar 2005; Chandra et al. 2011; Mittal & Jain 2015).
Materials and MethodsAdults and brood balls of S. rhadamistus from
open grazing fields of Kavdas Village and nearby areas of Wardha District, Maharashtra were collected. This region lies on the periphery of Bor Wildlife Sanctuary. Field-collected brood balls were maintained in the laboratory at temperature and relative humidity of 270C ± 20C and 65–75 %, respectively (Hayes 1929). Morphology was described from five specimens, each of III Instar larvae, pupae, and adult males and females. Larvae were kept in boiling water for about three minutes, followed by cooling, and preservation in 70% alcohol or glycerol to prevent shrinking (Ritcher 1966). Leica S8APO stereoscopic microscope was used to study the morphological characters, for photography, and for morphometry. Terminology of Edmonds & Haffler (1978) was followed. Adult specimens were washed in soap water to clear dirt, rinsed in clear water, air-dried to remove the moisture, oven-dried for about two hours, labeled, and studied. Arrow (1931) was followed for morphological characterization.
Male genitalia were dissected out from the adults under stereoscopic microscope. Individual male genitalia was further boiled in 10% KOH solution, washed in water, treated with glacial acetic acid for two to three minutes, and rinsed in distilled water. The dissected genitalia were stored in 1ml eppendorf filled with glycerol for future reference.
ResultsBrood Balls
A pair of adults construct nest in cow dung pats with an average diameter of 25–30 cm, and 5-10 cm thick. Each nest contains 3–7 brood balls per brood chamber with an average size of 4.7±1.41 cm and usually guarded by females. Each brood ball has a diameter of 18.9±0.74 mm (n = 20, range 18– 20 mm) (Images 1 & 2).
A. Eggs: Lemon yellow when freshly laid, oblong, average length 4.5 ± 0.1 mm (n = 10, range 4.4 to 4.6
mm); average maximum width in the middle 2.3 ± 0.11 mm (n = 10, range 2.3–2.5 mm); one egg is laid per brood ball, the egg is held vertically and attached to the substratum at the basal end with apical end bearing a small hump (Image 3).
B. Larva (III Instar): Larvae translucent in appearance; body covered with very fine setae; humped in the middle giving a ‘V’ shaped appearance to the body; average length 19 ± 0.20 mm (n = 5, range 18.8–19.3 mm), average width at abdominal hump 7 ± 0.15 mm (n = 5, range 6.8– 7.2 mm) (Image 4). Clypeus wider than long, rectangular, lateral margins straight, posterior margin bilobed; irregular row of seven to nine setae in middle; cranium surface smooth; frons with one seta each at anterior angle to the dorsal surface of head, three to four anterior frontal setae on each side, posterior frontal setae absent; remaining cranial surface with two para-ocellar setae, and with a row of eight dorso-epicranial setae (Image 5); antennae four segmented each; labrum symmetrical, trilobed, broadly oval; maxillary stridulatory area with a row of 8–10 teeth; epipharynx chaetoparie each with 9–11 setae (Image 7); mandibles stout, with single median seta, incisor lobes with three teeth on left mandible and two on right mandible (Image 8). Three pairs of thoracic legs, each two segmented, with 10–12 fine setae, claws absent (Image 9). Raster on 10th sternum with two irregular bunches of serrations ventrally one on either half distinct.
C. Pupa: Pupa exarate; pronotum transverse with rounded margins resembling adults; four thumb like tergal support projections, one per segment present laterally on abdominal segments three to six (Image 10). Pronotal support projection absent. One pteronotal support projection present. Caudal projection callus-like (Image 11).
D. Adult: Male - body yellowish-orange, with metallic green patches; elongate, oval; length 14–15 mm with an average of 14.46 ± 0.4 mm, width 7 to 8 mm with an average of 7.52 ± 0.3 mm (Image 12); head semicircular, elevated area between eyes, clypeus metallic green, lateral margins blackish; antenna eight segmented; pronotum with elevated margins forming deep cavity at middle, anteriorly forming a short process that reflexes backwards, laterally blunt and angular downward, with circular black spots (Image 13); scutellum very small, finely punctuated; a large spine-like process with broad base and rounded on sides present on meso-sternum (Image 14); elytra striate with a broad black central median suture and a central dark spot shared by both sides of elytra, with one ventral spherical dark spot and two oval median

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12990–12994
Life stages of Scaptodera rhadamistus Khadakkar et al.
12992
dark spots present transversally; femora of fore-, mid- and hind legs orange; fore-tibia setose, tridentate, with elongate tooth; tibia, tarsi and claws of fore-, mid- and hind legs brownish-black; abdominal sclerites metallic greenish-black, pygidium dark, with fine, evenly spread punctuations.
Male genitalia: It consists of parameres and phallobase. Phallobase is broad and curved at the apical end, and about 2.5 times the length of parameres. Parameres bend at the junction of phallobase and parameres (Image 15).
Specimens examined: Col/47/M, 04.iv.2017, 03 ex., Kavdas, Hingna, Nagpur, Maharashtra, Suvarna Khadakkar (location: 21.060N & 78.860E, elevation: 310m); Col/47/M/(a), 17.iii.2017, 01 ex., Seloo, Wardha, Maharashtra, Suvarna Khadakkar (location: 20.830N & 78.700E, elevation: 265m); Col/47/M/(b), 10.ii.2017, 01 ex., Zilpi, Hingna, Nagpur, Suvarna Khadakkar (location: 21.060N & 78.860E, elevation: 366m)
Female - body yellowish-orange, oval; head: clypeus dark metallic green, black on margins; length, 13–14 mm; width, 6–7 mm (Image 16); antenna eight segmented; head semicircular; pronotum (Image 17) yellowish-orange with irregular central metallic green patch, wider than long, narrowed towards head and broader towards base, with single dark metallic green spots laterally, without any process, bare, with lateral margins rounded than angular; with a large spine-
like process, slender compared with males with broad base, present on meso-sternum (Image 18); elytra with a broad black central median suture and a dark spot shared by both sides of elytra at the middle; each half of elytra with one ventral spherical dark spot and two oval median dark spots present transversally; abdominal sclerites metallic greenish-black; femora of fore-, mid- and hind legs orange; tibia, tarsi and claws of fore-, mid- and hind legs brownish-black; pygidium dark; otherwise similar to males.
Specimens examined: Col/47/F, 17.iii.2017, 03 ex., Kavdas, Hingna, Nagpur Maharashtra, Suvarna Khadakkar (location: 21.060N & 78.860E, elevation: 310m); Col/47/F/(a), 17.iii.2017, 01 ex., Seloo, Wardha, Maharashtra, Suvarna Khadakkar (location: 20.830N & 78.700E, elevation: 265m); Col/47/F/(b), 10.ii.2017, 01 ex., Zilpi, Hingna, Nagpur, Suvarna Khadakkar (location: 21.060N & 78.860E, elevation: 366m).
Behaviour of larvaeBrood balls of smaller diameter and dried from
outside contained malformed and/or dead larvae whereas those moist from outside contained healthy larvae and eggs. When a hole is made in moist brood
Images 1–4. Brood balls of Scaptodera rhadamistus.1 - hatched brood balls; 2 - unhatched brood balls; 3 - egg; 4 - IIIrd instar larva. (Scale 1 & 2 = 1cm; 3 & 4 = 1mm). © Suvarna S Khadakkar
Images 5–9. Head and mouth parts of IIIrd instar larva of Scaptodera rhadamistus.5 - cranium; 6 - maxilla; 7 - epiphrynx; 8 - mandibles; 9 - prothoracic leg (Scale 5 & 9 = 1mm; 6 & 8 = 0.5mm). © Suvarna S Khadakkar
1
4
2
3
5
6
7
8
9

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12990–12994
Life stages of Scaptodera rhadamistus Khadakkar et al.
12993
ball with the help of a needle, the larva present inside eat the dung present in ball and seal the hole with the excrement. Larvae were unable to fill the holes when the punctures were bigger. Centipedes were found to occupy the brood balls after the adult emergence.
DiscussionFully grown III instar larvae of S. rhadamistus can be
characterized by the presence of a prominent hump in the middle of body, two segmented legs, and raster on 10th sternum with two irregular bunches of serrations ventrally one on either half distinct from ovate fields of stout setae. Pupal support projections present in scarabaeinae pupae play a major role in taxonomic studies. Four thumb-like tergal support projections, one per segment laterally on abdominal segments three to six, one pteronotal support projection, callus like caudal projection, and absence of pronotal support projection are other prominent characters of S. rhadamistus.
S. rhadamistus build nests in large undisturbed dung pats. Brood count of 3–7 brood balls at a time, per pair of adults of S. rhadamistus is considered less in comparison to other scarabs. Investment of energy in parental care may be a reason for smaller brood. Brood balls as well as females guarding the brood balls signifies the importance of parental care in this species. Diameter of the brood balls as well as the moisture
Images 10–11. Pupa of Scaptodera rhadamistus.10 - lateral tergal projections on segments (3-6); 11 - caudal projection (Scale = 1mm). © Suvarna S Khadakkar
10 11
Images 12–15. Scaptodera rhadamistus (male)12 - dorsal view; 13 - lateral view; 14 - large posterior process; 15 - aedegus with line (Scale = 1mm). © Suvarna S Khadakkar
12 13
14
15

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12990–12994
Life stages of Scaptodera rhadamistus Khadakkar et al.
12994
content plays important role in the development of S. rhadamistus. These aspects of life history are important for conservation of the species.
References
Arrow, G.J. (1931). Fauna of British India Coleoptera, Lamellicornia Part III (Coprinae). Taylor and Francis, London, 374pp.
Chandra, K. & S.C. Ahirwar (2005). Scarabaeid beetles of Bandhavgarh National Park, Madhya Pradesh. Zoos’ Print Journal 20: 1961–1964.
Chandra, K., S. Khan, D. Gupta & S.P. Singh (2011). Additional records of scarab fauna (Coleoptera:Scarabaeidae) of Pachmarhi Biosphere Reserve, Madhya Pradesh, India. National Journal of Life Sciences 8: 65–68.
Edmonds, W.D. & G. Halffter (1978). Taxonomic review of immature dung beetles of the subfamily Scarabaeinae (Coleoptera: Scarabaeidae). Systematic Entomology 3: 307–331.
Halffter, G. & E.G. Matthews (1966). The natural history of dung beetles of the subfamily Scarabaeinae (Coleoptera: Scarabaeidae). Folia Entomológica Mexicana 12-14: 1–312.
Hanski, I. & Y. Cambefort (1991). Dung Beetle Ecology. Princeton University Press, Princeton, xiii+481pp. Threatened Taxa
Hayes, W.P. (1929). Morphology, Taxonomy and Biology of larval Scarabaeoidea. Illinois Biological Monographs. Vol. XII, No. 2. University of Illinois, Illinois, 166pp.
Mittal, I.C. & R.A. Jain (2015). checklist of Indian dung beetles (Coleoptera : Scarabaeidae). Indian journal of Entomology 77: 383–404.
Philips, T.K. (2016). Phylogeny of the Oniticellini and Onthophagini dung beetles (Scarabaeidae, Scarabaeinae) from morphological evidence. Zookeys 579: 9–57.
Ritcher, P.O. (1966). White Grubs and Their Allies. Oregon State University Press, No. 4, 220pp.
Schoolmeesters, P. (2017). Catalogue of life. Available from: http://www.catalogueoflife.org/col/details/species/id/4edb5905b3181bb4df7c9c8ccc049ee1. Downloaded on 27 March 2017.
Sreedevi, K. & S. Tyagi (2014). Diagnostics in larvae of white grubs (Scarabaeidae: Coleoptera) - important characters. Indian Journal of Entomology 76(2) 169–172.
Veeresh, G.K. (1980). Larval taxonomy of white grubs, with special reference to Melolonthine beetles. Final Report of ICAR Ad-hoc Project, 16–18pp.
Images 16–18. Scaptodera rhadamistus (female)16 - dorsal view; 17 - pronotum; 18 - large posterior process on mesosternum (Scale = 1mm).© Suvarna S Khadakkar
16 17
18

12995
Shor
t Co
mm
unic
atio
n
DOI: https://doi.org/10.11609/jott.4031.10.15.12995-13001 | ZooBank: urn:lsid:zoobank.org:pub:40BA8EB3-3FCE-48FE-8D0A-4AA766878ADD
Editor: K.A. Subramanian, Zoological Suryve of India, Chennai, India. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 4031 | Received 26 January 2018 | Final received 15 October 2018 | Finally accepted 16 November 2018
Citation: Tuhin, M.S.H. & M.K. Khan (2018). An updated list of Odonata of southwestern Bangladesh. Journal of Threatened Taxa 10(15): 12995–13001; https://doi.org/10.11609/jott.4031.10.15.12995-13001
Copyright: © Tuhin & Khan 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: The project was supported by The Rufford Foundation (Grant number 18697-1).
Competing interests: The authors declare no competing interests.
Acknowledgements: We thank Payal Barua for her comments on the initial version of the manuscript. We are also thankful to Rupa Saha for helping with manuscript preparation
An updated list of Odonata of southwestern Bangladesh
M. Sajjad Hossain Tuhin 1 & M. Kawsar Khan 2
1 Forestry and Wood Technology Discipline, Khulna University, Khulna 9208, Bangladesh2 Department of Biochemistry and Molecular Biology, Shahjalal University of Science and Technology, Sylhet 3114, Bangladesh1 [email protected], 2 [email protected] (corresponding author)
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12995–13001
Odonates (dragonflies and damselflies) were one of the earliest winged insects that evolved in the Permian period (Kalkman et al. 2008) and distributed all over the world except in Antarctica (Silsby 2001; Grimaldi & Engel 2005; Trueman 2007). Although odonates are highly distributed in diverse ecological niches, they are sensitive to the alteration of their habitats. Hence, odonates are considered as indicators of the status of freshwater ecosystems (Watson et al. 1982; Brown 1991; Martin & Maynou 2016). Odonates are also extensively studied in evolutionary and ecological research (Córdoba-Aguilar
Abstract: An odonate survey was conducted throughout the southwestern region of Bangladesh, concentrating on eight districts and the Sundarban, from August 2014 to August 2016. A total of 50 species under 30 genera belonging to six families was recorded during the study period. Among these, 31 species belonged to Anisoptera and 19 to Zygoptera suborders. Libellulidae and Coenagrionidae were the most dominant anisopteran and zygopteran families with 28 and 17 species, respectively. One Zygoptera species Mortonagrion varalli was newly added to the odonate fauna of Bangladesh.
Keywords: Damselflies, dragonflies, habitat associations, Mortonagrion varalli, Odonata, species richness, Sundarban.
2008). At present, 5,740 species of odonates are known from the world (Subramanian 2009).
Odonates are considered freshwater insects as the females lay eggs on water or submerged plants and the larval development occurs underwater (Hornung & Rice 2003). Unlike the larva, the adults are aerial. Their foraging and reproductive success, however, depends heavily on the availability of freshwater resources. Hence, odonate assemblage is higher in aquatic habitats (Oppel 2005). Odonate diversity also varies in different climatic zones (Balzan 2012). Similar to other insect orders, the majority of the dragonfly species inhabits tropical and subtropical climate zones (Dumont 1991). The Indo-Malayan region is one of the most diverse habitats of highly endangered odonates (Clausnitzer et al. 2009). Bangladesh, being located in the Indo-Burma biodiversity hotspot zone, is expected to possess high odonate diversity (Chowdhury & Mohiuddin 2011). Along with geographic variation, seasonal variation such as temperature, humidity, and rainfall influences species richness of odonates. Bangladesh has six seasons with warm and wet summer, monsoon, and autumn from April to September. The temperature starts to fall after

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12995–13001
Odonata of southwestern Bangladesh Tuhin & Khan
12996
September and late monsoon, winter, and spring are dry and cold although the temperature barely drops below 10OC.
Bangladesh is a rich habitat for odonate diversity because of its geographic location and abundance of water bodies (Chowdhury & Mohiuddin 2011). Very few studies, however, were carried out to annotate the odonate fauna of the country. Chowdhury & Mohiuddin (2011) listed 96 species of odonates from the eastern region while Khan (2015b) reported 76 species from the northeastern region of Bangladesh. In recent years, a few species were added to the odonate fauna of Bangladesh and at present 108 species are known from the country (Khan 2015a, in press). The odonate survey till date, however, focused mainly on the eastern region while surveys are lacking in other parts of Bangladesh, especially in the southwestern region, which has diverse freshwater resources.
The southwestern region of Bangladesh is administratively mainly under Khulna Division. This division consists of 10 administrative districts and covers a large area of 22,285km2. The largest tract of mangrove forest of the world, the Sundarban, is also situated under this division and is distributed over three districts, namely, Khulna, Bhagerhat, and Satkhira. Many rivers,
canals, ponds, and lakes occur in this part of the country. These freshwater resources are excellent habitats for odonates (Khan 2015b). Biswas et al. (1980) took the first approach to annotate the odonates of this region; however, their study was limited to Bagerhat District. Since this last odonatological survey in the region, there were no attempts to study odonates. In the present study, we conducted a broad survey in the southwestern region of Bangladesh to document odonates of the area.
Materials and MethodsStudy Site
Khulna Division lies between 21.6430N to 24.1810N and 88.5610E to 89.9420E (Fig. 1). The study area has a tropical climate with a mild winter from October to March, a hot and humid summer from March to June, and a humid, warm, rainy monsoon from June to October. Temperature varies all the year round: the temperature falls to the lowest in January and December at 12–15 0C and reaches the highest in April–June at 41–45 0C. Daily relative humidity fluctuates between 50% and 90%, which is the lowest in the evening and highest in the morning. The maximum precipitation is experienced in July with 20–25 days of rain with 368mm precipitation (Bangladesh Bureau of Statistics 2014).
Figure 1. Reference map of southwestern region (Khulna Division) of Bangladesh marked with surveyed sites
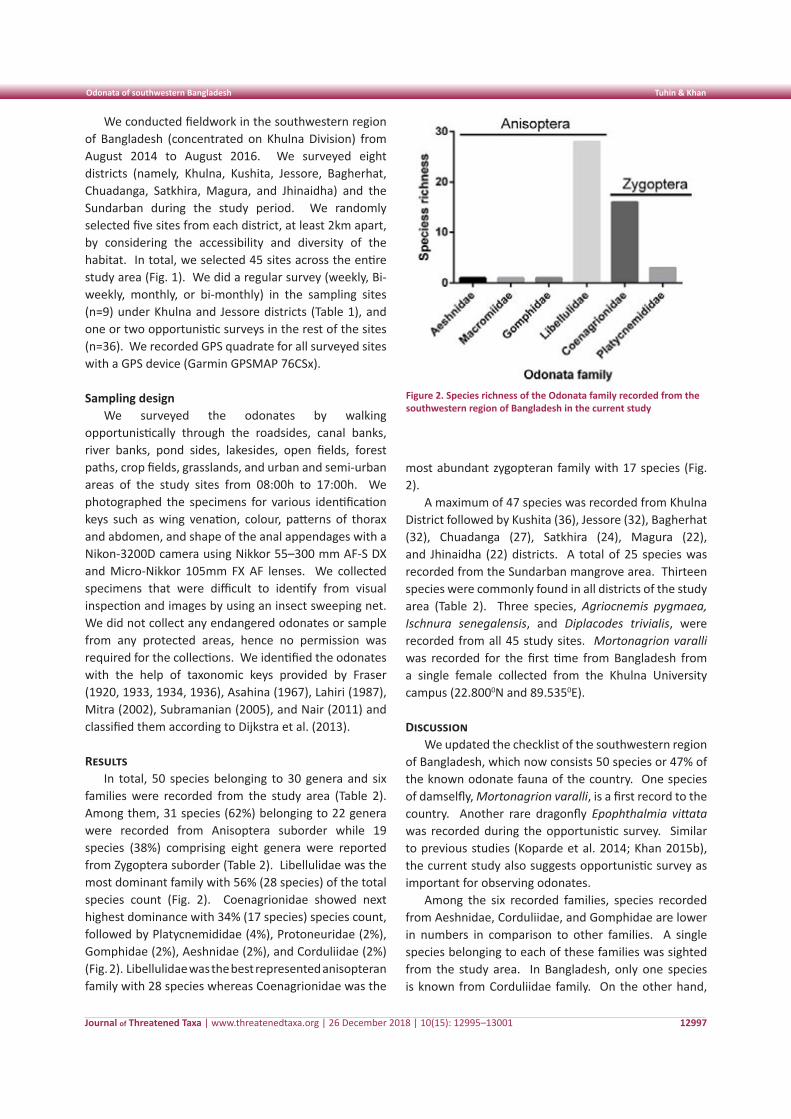
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12995–13001
Odonata of southwestern Bangladesh Tuhin & Khan
12997
We conducted fieldwork in the southwestern region of Bangladesh (concentrated on Khulna Division) from August 2014 to August 2016. We surveyed eight districts (namely, Khulna, Kushita, Jessore, Bagherhat, Chuadanga, Satkhira, Magura, and Jhinaidha) and the Sundarban during the study period. We randomly selected five sites from each district, at least 2km apart, by considering the accessibility and diversity of the habitat. In total, we selected 45 sites across the entire study area (Fig. 1). We did a regular survey (weekly, Bi-weekly, monthly, or bi-monthly) in the sampling sites (n=9) under Khulna and Jessore districts (Table 1), and one or two opportunistic surveys in the rest of the sites (n=36). We recorded GPS quadrate for all surveyed sites with a GPS device (Garmin GPSMAP 76CSx).
Sampling designWe surveyed the odonates by walking
opportunistically through the roadsides, canal banks, river banks, pond sides, lakesides, open fields, forest paths, crop fields, grasslands, and urban and semi-urban areas of the study sites from 08:00h to 17:00h. We photographed the specimens for various identification keys such as wing venation, colour, patterns of thorax and abdomen, and shape of the anal appendages with a Nikon-3200D camera using Nikkor 55–300 mm AF-S DX and Micro-Nikkor 105mm FX AF lenses. We collected specimens that were difficult to identify from visual inspection and images by using an insect sweeping net. We did not collect any endangered odonates or sample from any protected areas, hence no permission was required for the collections. We identified the odonates with the help of taxonomic keys provided by Fraser (1920, 1933, 1934, 1936), Asahina (1967), Lahiri (1987), Mitra (2002), Subramanian (2005), and Nair (2011) and classified them according to Dijkstra et al. (2013).
Results In total, 50 species belonging to 30 genera and six
families were recorded from the study area (Table 2). Among them, 31 species (62%) belonging to 22 genera were recorded from Anisoptera suborder while 19 species (38%) comprising eight genera were reported from Zygoptera suborder (Table 2). Libellulidae was the most dominant family with 56% (28 species) of the total species count (Fig. 2). Coenagrionidae showed next highest dominance with 34% (17 species) species count, followed by Platycnemididae (4%), Protoneuridae (2%), Gomphidae (2%), Aeshnidae (2%), and Corduliidae (2%) (Fig. 2). Libellulidae was the best represented anisopteran family with 28 species whereas Coenagrionidae was the
most abundant zygopteran family with 17 species (Fig. 2).
A maximum of 47 species was recorded from Khulna District followed by Kushita (36), Jessore (32), Bagherhat (32), Chuadanga (27), Satkhira (24), Magura (22), and Jhinaidha (22) districts. A total of 25 species was recorded from the Sundarban mangrove area. Thirteen species were commonly found in all districts of the study area (Table 2). Three species, Agriocnemis pygmaea, Ischnura senegalensis, and Diplacodes trivialis, were recorded from all 45 study sites. Mortonagrion varalli was recorded for the first time from Bangladesh from a single female collected from the Khulna University campus (22.8000N and 89.5350E).
Discussion We updated the checklist of the southwestern region
of Bangladesh, which now consists 50 species or 47% of the known odonate fauna of the country. One species of damselfly, Mortonagrion varalli, is a first record to the country. Another rare dragonfly Epophthalmia vittata was recorded during the opportunistic survey. Similar to previous studies (Koparde et al. 2014; Khan 2015b), the current study also suggests opportunistic survey as important for observing odonates.
Among the six recorded families, species recorded from Aeshnidae, Corduliidae, and Gomphidae are lower in numbers in comparison to other families. A single species belonging to each of these families was sighted from the study area. In Bangladesh, only one species is known from Corduliidae family. On the other hand,
Figure 2. Species richness of the Odonata family recorded from the southwestern region of Bangladesh in the current study
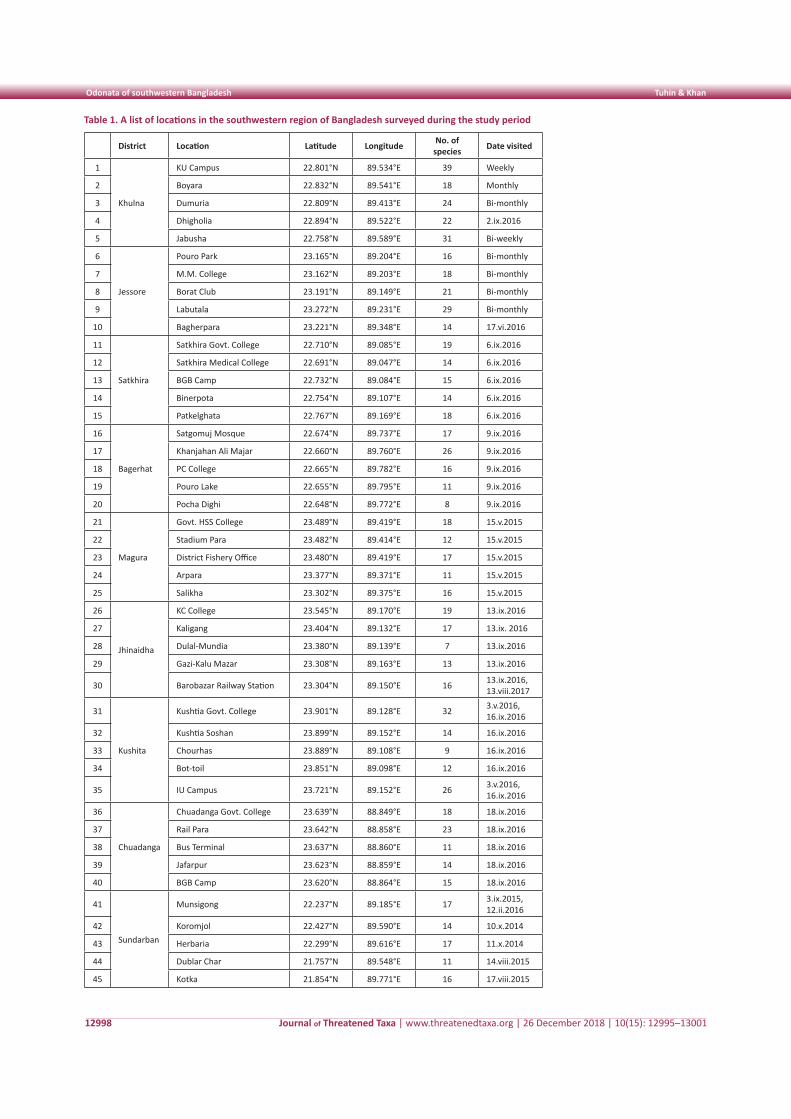
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12995–13001
Odonata of southwestern Bangladesh Tuhin & Khan
12998
Table 1. A list of locations in the southwestern region of Bangladesh surveyed during the study period
District Location Latitude Longitude No. of species Date visited
1
Khulna
KU Campus 22.801°N 89.534°E 39 Weekly
2 Boyara 22.832°N 89.541°E 18 Monthly
3 Dumuria 22.809°N 89.413°E 24 Bi-monthly
4 Dhigholia 22.894°N 89.522°E 22 2.ix.2016
5 Jabusha 22.758°N 89.589°E 31 Bi-weekly
6
Jessore
Pouro Park 23.165°N 89.204°E 16 Bi-monthly
7 M.M. College 23.162°N 89.203°E 18 Bi-monthly
8 Borat Club 23.191°N 89.149°E 21 Bi-monthly
9 Labutala 23.272°N 89.231°E 29 Bi-monthly
10 Bagherpara 23.221°N 89.348°E 14 17.vi.2016
11
Satkhira
Satkhira Govt. College 22.710°N 89.085°E 19 6.ix.2016
12 Satkhira Medical College 22.691°N 89.047°E 14 6.ix.2016
13 BGB Camp 22.732°N 89.084°E 15 6.ix.2016
14 Binerpota 22.754°N 89.107°E 14 6.ix.2016
15 Patkelghata 22.767°N 89.169°E 18 6.ix.2016
16
Bagerhat
Satgomuj Mosque 22.674°N 89.737°E 17 9.ix.2016
17 Khanjahan Ali Majar 22.660°N 89.760°E 26 9.ix.2016
18 PC College 22.665°N 89.782°E 16 9.ix.2016
19 Pouro Lake 22.655°N 89.795°E 11 9.ix.2016
20 Pocha Dighi 22.648°N 89.772°E 8 9.ix.2016
21
Magura
Govt. HSS College 23.489°N 89.419°E 18 15.v.2015
22 Stadium Para 23.482°N 89.414°E 12 15.v.2015
23 District Fishery Office 23.480°N 89.419°E 17 15.v.2015
24 Arpara 23.377°N 89.371°E 11 15.v.2015
25 Salikha 23.302°N 89.375°E 16 15.v.2015
26
Jhinaidha
KC College 23.545°N 89.170°E 19 13.ix.2016
27 Kaligang 23.404°N 89.132°E 17 13.ix. 2016
28 Dulal-Mundia 23.380°N 89.139°E 7 13.ix.2016
29 Gazi-Kalu Mazar 23.308°N 89.163°E 13 13.ix.2016
30 Barobazar Railway Station 23.304°N 89.150°E 16 13.ix.2016, 13.viii.2017
31
Kushita
Kushtia Govt. College 23.901°N 89.128°E 32 3.v.2016, 16.ix.2016
32 Kushtia Soshan 23.899°N 89.152°E 14 16.ix.2016
33 Chourhas 23.889°N 89.108°E 9 16.ix.2016
34 Bot-toil 23.851°N 89.098°E 12 16.ix.2016
35 IU Campus 23.721°N 89.152°E 26 3.v.2016, 16.ix.2016
36
Chuadanga
Chuadanga Govt. College 23.639°N 88.849°E 18 18.ix.2016
37 Rail Para 23.642°N 88.858°E 23 18.ix.2016
38 Bus Terminal 23.637°N 88.860°E 11 18.ix.2016
39 Jafarpur 23.623°N 88.859°E 14 18.ix.2016
40 BGB Camp 23.620°N 88.864°E 15 18.ix.2016
41
Sundarban
Munsigong 22.237°N 89.185°E 17 3.ix.2015, 12.ii.2016
42 Koromjol 22.427°N 89.590°E 14 10.x.2014
43 Herbaria 22.299°N 89.616°E 17 11.x.2014
44 Dublar Char 21.757°N 89.548°E 11 14.viii.2015
45 Kotka 21.854°N 89.771°E 16 17.viii.2015
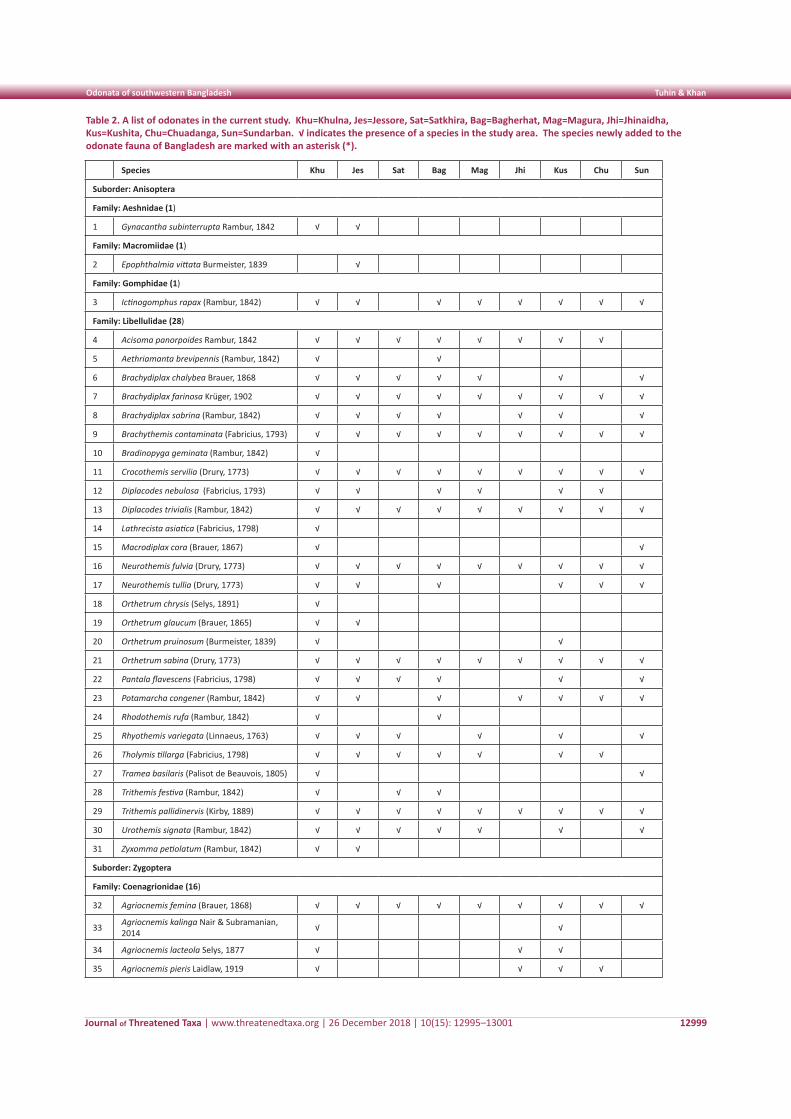
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12995–13001
Odonata of southwestern Bangladesh Tuhin & Khan
12999
Table 2. A list of odonates in the current study. Khu=Khulna, Jes=Jessore, Sat=Satkhira, Bag=Bagherhat, Mag=Magura, Jhi=Jhinaidha, Kus=Kushita, Chu=Chuadanga, Sun=Sundarban. √ indicates the presence of a species in the study area. The species newly added to the odonate fauna of Bangladesh are marked with an asterisk (*).
Species Khu Jes Sat Bag Mag Jhi Kus Chu Sun
Suborder: Anisoptera
Family: Aeshnidae (1)
1 Gynacantha subinterrupta Rambur, 1842 √ √
Family: Macromiidae (1)
2 Epophthalmia vittata Burmeister, 1839 √
Family: Gomphidae (1)
3 Ictinogomphus rapax (Rambur, 1842) √ √ √ √ √ √ √ √
Family: Libellulidae (28)
4 Acisoma panorpoides Rambur, 1842 √ √ √ √ √ √ √ √
5 Aethriamanta brevipennis (Rambur, 1842) √ √
6 Brachydiplax chalybea Brauer, 1868 √ √ √ √ √ √ √
7 Brachydiplax farinosa Krüger, 1902 √ √ √ √ √ √ √ √ √
8 Brachydiplax sobrina (Rambur, 1842) √ √ √ √ √ √ √
9 Brachythemis contaminata (Fabricius, 1793) √ √ √ √ √ √ √ √ √
10 Bradinopyga geminata (Rambur, 1842) √
11 Crocothemis servilia (Drury, 1773) √ √ √ √ √ √ √ √ √
12 Diplacodes nebulosa (Fabricius, 1793) √ √ √ √ √ √
13 Diplacodes trivialis (Rambur, 1842) √ √ √ √ √ √ √ √ √
14 Lathrecista asiatica (Fabricius, 1798) √
15 Macrodiplax cora (Brauer, 1867) √ √
16 Neurothemis fulvia (Drury, 1773) √ √ √ √ √ √ √ √ √
17 Neurothemis tullia (Drury, 1773) √ √ √ √ √ √
18 Orthetrum chrysis (Selys, 1891) √
19 Orthetrum glaucum (Brauer, 1865) √ √
20 Orthetrum pruinosum (Burmeister, 1839) √ √
21 Orthetrum sabina (Drury, 1773) √ √ √ √ √ √ √ √ √
22 Pantala flavescens (Fabricius, 1798) √ √ √ √ √ √
23 Potamarcha congener (Rambur, 1842) √ √ √ √ √ √ √
24 Rhodothemis rufa (Rambur, 1842) √ √
25 Rhyothemis variegata (Linnaeus, 1763) √ √ √ √ √ √
26 Tholymis tillarga (Fabricius, 1798) √ √ √ √ √ √ √
27 Tramea basilaris (Palisot de Beauvois, 1805) √ √
28 Trithemis festiva (Rambur, 1842) √ √ √
29 Trithemis pallidinervis (Kirby, 1889) √ √ √ √ √ √ √ √ √
30 Urothemis signata (Rambur, 1842) √ √ √ √ √ √ √
31 Zyxomma petiolatum (Rambur, 1842) √ √
Suborder: Zygoptera
Family: Coenagrionidae (16)
32 Agriocnemis femina (Brauer, 1868) √ √ √ √ √ √ √ √ √
33 Agriocnemis kalinga Nair & Subramanian, 2014 √ √
34 Agriocnemis lacteola Selys, 1877 √ √ √
35 Agriocnemis pieris Laidlaw, 1919 √ √ √ √

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12995–13001
Odonata of southwestern Bangladesh Tuhin & Khan
13000
Species Khu Jes Sat Bag Mag Jhi Kus Chu Sun
36 Agriocnemis pygmaea (Rambur, 1842) √ √ √ √ √ √ √ √ √
37 Ceriagrion cerinorubellum (Brauer, 1865) √ √ √ √ √ √ √ √
38 Ceriagrion coromandelianum (Fabricius, 1798) √ √ √ √ √ √ √ √ √
39 Ceriagrion olivaceum Laidlaw, 1914 √ √ √
40 Amphiallagma parvum (Selys, 1876) √
41 Ischnura aurora (Brauer, 1865) √ √ √ √ √ √ √ √ √
42 Ischnura rufostigma Selys, 1876 √ √
43 Ischnura senegalensis (Rambur, 1842) √ √ √ √ √ √ √ √ √
44 Mortonagrion aborense (Laidlaw, 1914) √ √
45 Mortonagrion varalli Fraser, 1920 * √
46 Pseudagrion microcephalum (Rambur, 1842) √ √ √ √ √ √
47 Pseudagrion rubriceps Selys, 1876 √ √ √ √ √
Family: Platycnemididae (3)
48 Copera marginipes (Rambur, 1842) √ √ √ √
49 Onychargia atrocyana Sélys, 1865 √ √ √ √ √
50 Pseudocopera ciliata (Selys, 1863) √ √ √ √ √ √ √ √ √
Aeshnidae and Gomphidae represent six species each. Hence, the lower species number recorded from these families is a representation of the fewer known species of the families in Bangladesh. The present study recorded three species from Platycnemididae family of which Pseudocopera ciliata was frequently sighted in different study sites. All anisopteran species excluding Lathrecista asiatica and Bradinopyga geminata were frequently observed. Similarly, all Coenagrionidae species except mortonagrion species were frequently recorded.
Species diversity varies with change in habitat and microclimate. Some of the species were found only in distinct habitats whereas others adapted to broader landscapes were found in diverse habitats (Corbet 1999). Agriocnemis femina, Agriocnemis kalinga, Agrioscemis pygmaea, Agriocnemis lecteola, Agriocnemis pieris, and Diplacodes trivialis were found restricted to grassland habitats. Ischnura senegalensis, Pseudagrion microcephalum, and Pseudagrion rubriceps were observed mainly in water bodies. On the other hand, Epophthalmia vittata, Lathrecista asiatica, and Gynacantha subinterrupta were found only at the higher canopy of certain places. These observations provide evidence of habitat diversification and specialization of different species.
In the present study, 25 odonate species were recorded from the Sundarban mangrove forest. Due to high salinity and lack of freshwater, Sundarban is not well known for its odonate diversity. Previously,
26 species were recorded from the Indian part of the Sundarban region (Mitra & Mitra 2009). Among those, 17 species were also recorded in the current study from the Bangladeshi part of the Sundarban region. Also, eight species, which were unknown from the Sundarban region before, were recorded in the present study.
Water salinity is high in the southwestern region in comparison to the other areas of Bangladesh. Salinity is even higher in the mangrove areas and varies between 5ppt and 25ppt (Joshi & Ghose 2003). Pure, non-polluted water is important for odonate breeding (Carchini et al. 2005) and saline water has a negative influence on the odonate diversity (Needham & Westfall 1955). Hence, the lower odonate diversity in the southwestern region of Bangladesh (50 species in comparison to 96 species in the eastern region) can be a result of water salinity. The southwestern region also lacks the diversity of freshwater resources like waterfalls, streams, and tropical forest that are present in the eastern region.
In conclusion, we recorded 50 species of odonates in the current survey and updated the checklist of the southwestern region of Bangladesh. The present study is the first documentation of the odonate diversity in the Sundarban region of Bangladesh. Regional checklists are important to understand the diversity and conservation needs of a species and our study will fulfill that demand for the odonates of the region. Further long-term studies are required to understand the biology, population structure, threats, and conservation action needs of the

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 12995–13001
Odonata of southwestern Bangladesh Tuhin & Khan
13001
odonates of this region.
References
Asahina, S. (1967). A revision of the Asiatic species of the damselflies of the genus Ceriagrion (Odonata, Agrionidae). Japanese Journal of Zoology 15(3): 255–334.
Balzan, M.V. (2012). Associations of dragonflies (Odonata) to habitat variables within the Maltese Islands: a spatio-temporal approach. Journal of Insect Science 12(1): 87, https://doi.org/10.1673/031.012.8701
Bangladesh Bureau of Statistics (2014). District Statistics 2011 Khulna. Bangladesh Bureau of Statistics, Statistics Division, Dhaka, Bangladesh, 128pp.
Biswas, V., M.A. Bashar & A. Begum (1980). On a collection of Odonata from Bagerhat District, Khulna, Bangladesh. Indian Odonatology 3: 65–66
Brown, K.S. (1991). Conservation of neotropical environments: insects as indicators, pp. 349–404. In: Collins, N.M. (eds.). The Conservation of Insects and their Habitats. Academic Press, London, 493pp.
Carchini, G., A.G. Solimini & A. Ruggiero (2005). Habitat characteristics and Odonate diversity in mountain ponds of central Italy. Aquatic Conservation 15(6): 573–581.
Chowdhury, S.H. & M. Mohiuddin (2011). A check-list of the Odonata from the eastern region of Bangladesh with some taxonomic notes. University Journal of Zoology, Rajshahi University 30: 61–66; https://doi.org/10.3329/ujzru.v30i0.10755
Clausnitzer, V., V.J. Kalkman, M. Ram, B. Collen, J.E. Baillie, M. Bedjanič & H. Karube (2009). Odonata enter the biodiversity crisis debate: the first global assessment of an insect group. Biological Conservation 142(8): 1864–1869.
Corbet, P.S. (1980). Biology of Odonata. Annual Review of Entomology 25(1): 189–217.
Corbet, P.S. (1999). Dragonflies Behavior and Ecology of Odonata. Harley Books, Colchester, Essex, U.K. 830pp, ISBN 0 946589 64 X.
Córdoba-Aguilar, A. (ed.) (2008). Dragonflies and Damselflies: Model Organisms for Ecological and Evolutionary Research. Oxford University Press, London.
Dijkstra, K.D.B., V.J. Kalkman, R.A. Dow, F.R. Stokvis & J. van Tol (2013). Redefining the damselfly families: a comprehensive molecular phylogeny of Zygoptera (Odonata). Systematic Entomology 39(1): 68–96; https://doi.org/10.1111/syen.12035
Dumont, H.J. (1991). Fauna Palaestina (Insecta). Israel Academy of Sciences & Humanities, Jerusalem, 297pp.
Fraser, F.C. (1920). Mortonagrion varalli. Journal of Bombay Natural History Society 27: 148.
Fraser, F.C. (1933). The Fauna of British India including Ceylon and Burma: Odonata, Vol. 1. Taylor & Francis Ltd., London, 423pp.
Fraser, F.C. (1934). The Fauna of British India including Ceylon and Burma: Odonata, Vol. 2. Taylor & Francis Ltd., London, 423pp.
Fraser, F.C. (1936). The Fauna of British India including Ceylon and Burma: Odonata, Vol. 3. Taylor & Francis Ltd., London, 461pp.
Grimaldi, D. & M.S. Engel (2005). Evolution of the Insects. Cambridge University Press, New York, 763pp.
Hornung, J.P. & C.L. Rice (2003). Odonata and wetland quality in southern Alberta, Canada: a preliminary study. Odonatologica 32(2): 119–129.
Joshi, H. & M. Ghose (2003). Forest structure and species distribution along soil salinity and pH gradient in mangrove swamps of the Sundarbans. Tropical Ecology 44(2): 195–204.
Kalkman, V.J., V. Clausnitzer, K.D.B. Dijkstra, A.G. Orr, D.R. Paulson & J. van Tol (2008). Global diversity of dragonflies (Odonata) in freshwater. Hydrobiologia 595(1): 351–363; https://dx.doi.org/10.1007/s10750-007-9029-x
Khan, M.K. (2015a). Gynacantha subinterrupta Rambur, 1842: an addition to the odonates (Insecta: Odonata: Aeshnidae) of Bangladesh. Journal of Threatened Taxa 7(10): 7704–7705; https://doi.org/10.11609/JoTT.o4276.7704-5
Khan, M.K. (2015b). Dragonflies and damselflies (Insecta: Odonata) of the northeastern region of Bangladesh with five new additions to the Odonata fauna of Bangladesh. Journal of Threatened Taxa 7(11): 7795–7804; https://doi.org/10.11609/JoTT.o4314.7795-804
Khan, M.K. (2018). Odonata of Bangladesh with three new records for the country. Journal of Threatened Taxa 10(13): 12821-12827; https://doi.org/10.11609/jott.3819.10.13.12821-12827
Koparde, P., P. Mhaske & A. Patwardhan (2014). New records of dragonflies and damselflies (Insecta: Odonata) from the Western Ghats of Maharashtra, India. Journal of Threatened Taxa 6(5): 5744–5754; https://doi.org/10.11609/JoTT.o3402.5744-54
Lahiri, A.R. (1987). Studies on the odonate fauna of Meghalaya. Records Zoological Survey India, Occasional Paper 99: 1–402.
Martín, R. & X. Maynou (2016). Dragonflies (Insecta: Odonata) as indicators of habitat quality in Mediterranean streams and rivers in the province of Barcelona (Catalonia, Iberian Peninsula). International Journal of Odonatology 19(3): 107–124; https://doi.org/10.1080/13887890.2016.1172991
Mitra, T.R. (2002). Geographical distribution of Odonata (Insecta) of Eastern India. Memoirs of Zoological Survey of India 19(1): 1–208.
Mitra, A. & B. Mitra (2009). Pictorial Handbook of Common Dragonflies and Damselflies (Insecta: Odonata) of Mangroves of Sundarbans, India. Zoological Survey of India, Kolkata, 56pp.
Nair, M.V. (2011). Dragonfles & Damselfles of Orissa and Eastern India. Wildlife Organisatin, Forest & Environment Department, Government of Orissa, Bhubaneshwar, 254pp.
Needham, J.G. & M.J. Westfall, Jr. (1955). A Manual of the Dragonflies of north America (Anisoptera) including the Greater Antilles and the Provinces of the Mexican Border. University of California Press, Berkeley, California, 615pp.
Oppel, S. (2005). Habitat associations of an Odonata community in a lower montane rainforest in Papua New Guinea. International Journal of Odonatology 8(2): 243–257.
Silsby, J. (2001). Dragonflies of the World. Smithsonian Institution Press, Washington, D.C., 216pp.
Silva, D., P., de Marco & D.C. Resende (2010). Adult odonate abundance and community assemblage measures as indicators of stream ecological integrity: a case study. Ecological Indicators 10(3): 744–752.
Subramanian, K.A. (2005). Dragonflies and Damselflies of Peninsular India - A Field Guide. Indian Academy of Sciences, Bangalore, India, 118pp.
Subramanian, K.A. (2009). A Checklist of Odonata of India. Zoological Survey of India, Kolkata, 36pp.
Trueman, W.H.J. (2007). A brief history of the classification and nomenclature of Odonata. Zootaxa 1668(1): 381–394; https://dx.doi.org/10.11646/zootaxa.1668.1.20
Watson, J.A.L., A.H. Arthington, & D.L. Conrick (1982). Effect of sewage effluent on dragonflies (Odonata) of Bulimba Creek, Brisbane. Marine and Freshwater Research 33(3): 517–528.
Threatened Taxa

Reproductive biology of Salacia fruticosa Subin et al.
13002
Shor
t Co
mm
unic
atio
n
DOI: https://doi.org/10.11609/jott.3069.10.15.13002-13005
Editor: V. Sampath Kumar, Botanical Survey of India, Howrah, India. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 3069 | Received 03 January 2017 | Final received 30 August 2018 | Finally accepted 07 December 2018
Citation: Subin, K., P.A. Jose & T.V. Sarath (2018). On the reproductive biology of Salacia fruticosa Wall. ex M.A. Lawson - an endemic medicinal plant of the Western Ghats, india. Journal of Threatened Taxa 10(15): 13002–13005; https://doi.org/10.11609/jott.3069.10.15.13002-13005
Copyright: © Subin et al. 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: Kerala Forest Research Institute Plan Grant received through Kerala State Council for Science,Technology and Environment, Thiruvananthapuram.
Competing interests: The authors declare no competing interests.
Acknowledgements : The authors thank the Director, Kerala Forest Research Institute, Peechi for approval of the plant grant and constant encouragement provided throughout the study.
On the reproductive biology of Salacia fruticosa Wall. ex M.A. Lawson - an endemic medicinal plant of the
Western Ghats, india
K. Subin 1 , P.A. Jose 2 & T.V. Sarath 3
1,2,3 Sustainable Forest Management Division, Kerala Forest Research Institute, Peechi, Kerala 680653, India1 [email protected], 2 [email protected] (corresponding author), 3 [email protected]
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13002–13005
Salacia L. belonging to the family Celastraceae, consists of about 200 species worldwide, distributed in tropical America, Africa and Asia (Mabberley 2008) of which 21 species are reported from India (Ramamurthy & Naithani 2000). Out of this, 15 species are reported from peninsular India and eight species from Kerala itself (Udayan et al. 2012). The true raw drug, Ekanayakam (Malayalam) / Pitika (Sanskrit) is extracted from Salacia reticulata Wight, however, the species is so rare and sparse for raw drug collection in situ. The
Abstract: Salacia fruticosa Wall. ex M.A. Lawson, an endemic species was studied for the reproductive biology as this species showed reduced fruit set and natural regeneration. The stigma-anther proximity, an extremely low number of pollen grains, a short period of pollen viability, a sparse incidence of pollinators, protandrous and facultative autogamous nature of the flowers and a low percent in fruit set were identified as biological constraints for the species. The incidence of seed pest was added to the poor seed and seedling bank and accelerated rarity process of the species.
Keywords: Endemic, ex situ conservation, insect-pest, medicinal plant, overexploitation, reproductive biology, Salacia fruticosa.
Salacia oblonga Wall. ex Wight & Arn., S. fruticosa Wall. ex M.A.Lawson and S. chinensis L. are the substitutes used and all of them are overexploited and facing a high threat in their habitats (Chithra et al. 2010).
Salacia fruticosa is a woody climbing shrub, endemic to Western Ghats, distributed in the evergreen and semi evergreen forests and also in the plains. It has anti hyperglycemic properties (Venkateshwarlu et al. 2009). Endemism and fragmented distribution, over exploitation, poor fruit set and seed infestation have led to the study of reproductive biology of the plant in order to understand the reproductive constraints of the species.
Material and MethodsPeriodic dynamic changes in reproductive
phenological phases of the species Salacia fruticosa were monitored and recorded on a day to day basis at KFRI, Peechi with respect to bud initiation, development, anthesis, pollination behavior etc. as per the methods suggested by different authors (Faegri & Pijl 1979; Armstrong & Drummond 1984; Sreekala et al. 2008;

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13002–13005
Reproductive biology of Salacia fruticosa Subin et al.
13003
Jose & Pandurangan 2012, 2013). Stigma receptivity was determined by the physical appearance of stigma such as turgidity, shine and oily appearance and it is confirmed by using hydrogen peroxide (H2O2) (bubble formation). The pollen-ovule ratio was worked out as per the method suggested by Cruden (1977). Pollen fertility test was carried out using Acetocarmine staining method (Sharma & Sharma 1980). Pollen germination was carried out with 15% sucrose solution. Bagging experiment was conducted for evaluating the pollination behavior.
Study area
The study was carried out in a population of S. fruticosa growing in the medicinal garden of the Kerala Forest Research Institute, Peechi, situated between 10.5290N & 76.3480E (Fig.1).
Results
In Salacia fruticosa, the flower bud takes one week to reach the full bloom stage. The opening of the flower takes place from 11.00–12.00 hr. The flowers are protandrous as the anther dehisces at 10.00hr while the stigma is receptive only at 11.00hr. The stigma was found to be receptive for about 30 hours. Each flower has three anthers and 3-celled ovaries with 1 (rarely 2) ovules in each cell. Pollen grains are liberated through longitudinal slits of the anther. A single anther contains ~105 pollen grains, thus one flower comprises around ~315 pollen grains. Hence, pollen-ovule ratio was worked out as 105 pollens per ovule (105:1). The pollen grains are globose in nature and having 13µm in diameter. Nearly 92% pollen grains are found viable
and 87% pollen germination was recorded at the time of anthesis. The pollen viability and germination was found to decrease and a drastic decline was recorded after three hours from anthesis though the stigma was found receptive for 30 hours. After around one-&-half months the fruits attain maturity and the percentage of fruit set was found to be 25% (detailed floral characters are given in Table 1 & Image 1). The emasculated flowers with artificial pollination as well as flowers which are polybagged (with big holes) were found to be inefficient
Table 1. Reproductive characters of Salacia fruticosa
Floral Characters Findings
Flowering period Throughout the year
Flower type Pentamerous, bisexual, actinomorphic
Flower colour Yellow
Flower opening time (anthesis) 11.00–12.00 hr
Floral nature Protandrous
Anther dehiscence mode Longitudinal slit
Anther dehiscence time 10.00hr
Average no. of pollens/anther 105
Mean no. of pollen grains/ flower 315
Mean no. of ovules/flower 3
Pollen shape Globular, smooth
Stigma receptive time and period 11.00hr onwards (up to 30 hours)
Pollen - Ovule ratio 105:1
Pollen diameter 13 µm
Pollen fertility 92.3%
Fruit development period 45 days
Percentage of fruit set 25%
Figure 1. Map showing the study site

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13002–13005
Reproductive biology of Salacia fruticosa Subin et al.
13004
for fruit set, that underlines the autogamous nature of the species. Majority of the fruits on maturity were found to be infested and the seeds were consumed by the caterpillars of the adult butterfly, Bindahara moorei Fruhstorfer. About 70–80 % matured fruits were infested by this insect.
Discussion and ConclusionKnowledge of reproductive biology particularly
the anthesis, pollen and stigma viability, nature of pollination and fruit set are essential to understand the causes of rarity of the species. The dehiscence of anthers and release of pollen grains prior to the receptivity of stigma is considered as an indicator for promoting facultative autogamy in the species. The low number of pollen grains, i.e., 105 pollen/anther was found to promote cross pollination through insects but the incidence of pollinators during blooming time was
negligibly sparse. A low count of pollens and a sparse incidence of pollinators are limiting the species from both anemophily and entomophily. According to Cruden (1977), plants with pollen–ovule ratio lying between 31.9 to 396 support facultative autogamy and signifies the above observations in the species.
Depending upon the population history and reproductive features of the species, reduced pollinator service may have several negative impacts on the plant population including reproductive failure (Jennersten 1988) or decreased effective population size through reduced gene flow and increased selfing (Bawa 1990). These altered reproductive patterns may cause less of genetic diversity and/or reduced progeny fitness due to inbreeding depression (Jain 1976; Barrett & Kohn 1991). Self fertile individuals, however, may be at a selective advantage in some particular habitats, if outcrossing is disfavored because plant density is low or if pollinators
Image 1 . Reproductive biology of Salacia fruticosa. A - A twig with fruit; B–C - Flower development; D - Fruit and Seed infestation by Caterpillars; E - Infested fruit cut open showing consumed seeds and pulp. © K. Subin
A C
B
D E

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13002–13005
Reproductive biology of Salacia fruticosa Subin et al.
13005
are scarce or cross pollination is inadequate (Jain 1976; Lloyd 1980).
Even though the stigma receptive for 30 hours, the drastic decline of pollen viability within three hours after anthesis reduces chances for effective pollination. The bagging experiment also underlines the chances for self pollination in the species. The low rate of pollination reduced fruit set to 25% in the species. The insect-pest incidence and its extent of damage on the seed and seedling output accelerate endangerment of the species in the near future.
References
Armstrong, E.J. & B.A. Drummond (1984). Floral biology of Myristica fragrans Houtt. (Myristicaceae) the nutmeg of commerce, Biotropica 32–38
Barrett, S.C.H. & J.R. Kohn (1991). Genetic and evolutionary consequences of small population size in plants: implications for conservation. In: Falk, D. & K.E. Holsinger, (eds.), Genetics and Conservation of Rare Plants. Oxford University Press, New York. pp. 3–30.
Bawa, K.S. (1990). Plant-pollinator interactions in tropical rain forest. Annual Reviews of Ecology and Systematics 21: 399–422.
Chithra, M., P.S. Udayan & I. Balachandran (2010). Systematic study of the genus Salacia L. occurring in Kerala, pp. 881–882. In: Nair, C.T.S. (ed.). Proceedings of the 22nd Kerala Science Congress, 28–31 January. Kerala Forest Research Institute, Peechi.
Cruden, R.W. (1977). Pollen-ovule ratios- A conservative indicator of breeding system in flowering plants. Evolution 31: 32–46.
Faegri, K. & L. Pijl (1979). The Principles of Pollination Ecology. Pergamon Press, London, 248pp.
Jain, S.K. (1976). The evolution of inbreeding in plants. Annual Review of Ecology and Systematics 10: 469–495.
Jennersten, O. (1988). Pollination in Dianthus deltoides (Caryophyllaceae): effects of habitat fragmentation on visitation and seed set. Conservation Biology 2: 359–366.
Jose, P.A. & A.G. Pandurangan (2012). Reproductive biology of Ochreinauclea missionis (Wall. Ex G. Don) Ridsd. An endemic and vulnerable tree from the Western Ghats, India. Annals of Forestry 20(2) :161-167.
Jose, P.A. & A.G. Pandurangan (2013). Reproductive biology of Gluta travancorica Bedd.. An endemic and threatened tree from the Southern Western Ghats, India. Annals of Forestry 21(2) :165–171.
Lloyd, D.G. (1980). Demographic factors and mating patterns in Angiosperms, pp. 677–688. In: Solbrig, O.T. (ed.) Demography and Evolution in Plant Populations. Blackwell, Oxford.
Mabberley, D.J. (2008). Mabberley’s Plant- Book: A Portable Dictionary of the Vascular Plants, their classification and uses. Cambridge University Press, 1021pp.
Ramamurthy, K. & B. D. Naithani (2000). Hippocrataceae, pp. 138–162. In: Singh, N.P., J.N., Vohra, P.K. Hajra & D.K. Singh (eds.). Flora of India 5. Botanical Survey of India, Calcutta.
Sharma, A. & A. Sharma (1980). Chromosome techniques – Theory and Practice. Butterworth & Co. (Pub.) Ltd., London, 724 pp.
Sreekala, A.K., A.G.,Pandurangan, R. Ramasubbu & S.K. Kulloli (2008). Reproductive Biology of Impatience coelotropis Fischer, a critically endangered balsam from the Southern Western Ghats. Current Science 95: 386–388.
Udayan, P.S., R. Yohannan, M.S. Devipriya, V. Devipriya & A.K. Pradeep (2012). A new species of Salacia (Hippocrateaceae) from south India. Edinburgh Journal of Botany 69: 255–258.
Venkateshwarlu, E., R. Narsimha, A. Reddy, A.S. Sunder, G. Kiran, J.V. Rao & S. Madhusudhan (2009). Anti-hyperglycemic activity of methanolic extract of Salacia fruticosa leaves in alloxan induced diabetic rats. Drug Invention Today 1(2): 95–97.
Threatened Taxa

Contribution to the Macromycetes of West Bengal Saha et al.
13006
Shor
t Co
mm
unic
atio
n
DOI: https://doi.org/10.11609/jott.4188.10.15.13006-13013
Editor: R.K. Verma, Tropical Forest Research Institute, Jabalpur, India. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 4188 | Received 10 April 2018 | Final received 28 September 2018 | Finally accepted 21 November 2018
Citation: Saha, R., A.K. Dutta, S. Paloi, A. Roy & K. Acharya (2018). Contribution to the Macromycetes of West Bengal, India: 28–33. Journal of Threatened Taxa 10(15): 13006–13013; https://doi.org/10.11609/jott.4188.10.15.13006-13013
Copyright: © Saha et al. 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: Department of Environment, the Government of West Bengal, India.
Competing interests: The authors declare no competing interests.
Acknowledgements: The authors would like to acknowledge the Department of Environment, the Government of West Bengal, India, for providing financial assistance. The facilities provided by the department of Botany (UGC-CAS Phase VI, VII), University of Calcutta, and infrastructural support by DST-FIST are also duly acknowledged.
Contribution to the Macromycetes of West Bengal, India: 28–33
Rituparna Saha 1 , Arun Kumar Dutta 2 , Soumitra Paloi 3 , Anirban Roy 4 & Krishnendu Acharya 5
1,2,3,5 Molecular and Applied Mycology and Plant Pathology Laboratory, Department of Botany, University of Calcutta, Kolkata, West Bengal 700019, India
1,2,3,4 West Bengal Biodiversity Board, Prani Sampad Bhawan, 5th floor, LB 2, Sector III, Salt Lake City, Kolkata, West Bengal 700106, India
1 [email protected], 2 [email protected], 3 [email protected], 4 [email protected], 5 [email protected] (corresponding author)
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13006–13013
West Bengal has a diverse range of biogeographical and ecological conditions due to the presence of the coastal region of the Bay of Bengal on one side and the subalpine mountains of the eastern Himalayan region on the other. This wide array of phyto-topographical features facilitates the luxuriant growth of macrofungi belonging to several types and stature like poroid, dentate, and gilled. The habitat of these fungi varies from saprophytic to humicolous and mycorrhizal (Pradhan et al. 2016).
The genus Ganoderma (Basidiomycota: Polyporales) is a woody, poroid group of saprotrophic fungi. The
Abstract: The present paper deals with the report of six poroid woody macro fungi belonging to the family Ganodermataceae from West Bengal in India. The taxonomic account of these collected fungi is represented herein with detailed macro- and micro-morphological features.
Keywords: Basidiomycota, Ganoderma species, Polyporales, taxonomy.
distinguishing features of this genus include the presence of truncate basidiospores and the colour reaction of pileus and pore surface, which never turn permanently dark in 10% KOH solution (Sharma 2012).
The present study reports six poroid fungi with their detailed morpho-taxonomic enumeration from the state.
Material and MethodsMacrofungal specimens were collected during the
rainy seasons (June–October) of 2010–2017. Colour photographs were taken and macro-morphological features of each specimen were studied in the field. A small part of each fresh specimen was cut and its reaction with chemicals was observed. Each collection was then wrapped in tissue papers and kept separately in a box to avoid contamination. Finally, the collected specimens were withered in a hot air drier until the moisture was totally removed. Microscopic features were observed with a Carl Zeiss AX10 Imager A1 phase contrast microscope from thin handmade sections

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13006–13013
Contribution to the Macromycetes of West Bengal Saha et al.
13007
of the dried basidiocarps stained with congo red and Melzer’s reagent. Identification and colour terminology followed Kornerup & Wanscher (1978), Bhosle et al. (2010), and Sharma (2012). Thirty measurements of basidiospores were taken from each sample for calculating the dimensions of the basidiospore. Length/breadth ratio denotes the Q value. Mean Q value (Qm) was measured by dividing the total sum of Q value by the total number of spores observed. Hand drawings of different identifying characters were obtained with a camera lucida and a 0.1mm rotring pen, which was used to trace the lines. The voucher specimens were preserved following Pradhan et al. (2015) and were deposited in the Calcutta University Herbarium (CUH), Kolkata, India.
TaxonomyGanoderma lucidum (Curtis) P. Karst.
Revue mycol., Toulouse 3(no. 9): 17 (1881) (Fig. 1; Image 1)
Basidiocarp annual, laterally stipitate. Pileus applanate when young and funnel-shaped at maturity, 45–111 mm in length and 36–70 mm in breadth, 17mm thick near the base. Pileus upper surface reddish-brown (8D6, 8E6), shiny, glabrous, laccate. Margin lobed, whitish (1A1), thin. Pore surface whitish (1A1), greyish-orange (5B3) on bruising. Pores 4–6 per mm, circular. Tubes not stratified, brown (7E5), 3–6 mm deep. Context soft, double layered, upper reddish golden brownish-orange (6C3) and lower light brown (7D4) near the tubes, 4–10 mm thick near the base. Stipe cylindrical, central when young and lateral at maturity, 70–95 mm in length and 10–25 mm in breadth, reddish-brown (8E7), laccate, glabrous, shiny.
Hyphal system trimitic. Generative hyphae of context greyish-yellow (4C3), thin walled, clamped at septa, 2.5–3.5 µm in breadth; binding hyphae branched, interwoven, 0.7–2 µm wide, hyaline; skeletal hyphae unbranched at base and branched towards apex, 3.5–4.5 µm wide, greyish-yellow (4C3), interwoven. Generative hyphae of the tube layer greyish-yellow (4C3), thin-walled, clamped at septa, difficult to observe, 2.5–3.5 µm in breadth; binding hyphae branched, 0.5–2 µm wide, hyaline; skeletal hyphae unbranched at base and branched towards apex, 3.5–5 µm wide, greyish-yellow (4C3). Cuticular hyphae regular, closely packed with clavate end cells, sometimes cells are cylindrical or tubular, golden (4C6) to brownish-orange (6C8), thick, 6–12 µm wide. Basidiospores truncate, (7–)7.5–8(–8.5) × (3.5–)4–4.5(–5) µm, Q = 1.6–2.0, Qm = 1.84, double
walled, exospores hyaline, thin and endospore thick, brownish-orange (7C5).
Habit and habitat: Solitary to gregarious, grown on dead wooden log of angiosperm.
Specimen examined: CUH AM532, 26.vii.2017, 22.3490N & 88.0690E, elevation 12m, Uluberia, Kolkata, West Bengal, India, coll. A.K. Dutta & S. Paloi.
Remarks: Ganoderma lucidum is well characterized
Image 1. Ganoderma lucidum. Scale = 10mm
© A.K. Dutta
Figure 1. Ganoderma lucidum. a - hyphae of the cutis, b - basidiospores, c - context hyphae, d - tube layer hyphae. Bars = 5µm. Drawing by R. Saha.
a
c
b
d

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13006–13013
Contribution to the Macromycetes of West Bengal Saha et al.
13008
by its laccate-shaped, laterally stipitate, reddish-brown coloured basidiocarp with whitish margin, distinctly double layered context differentiated by reddish golden brownish-orange upper and light brown (7D4) lower near the tubes, presence of regular, closely packed clavate end cells at the cutis, and basidiospores measuring 7–8.5 × 3.5–5 µm in diameter with Qm of 1.84.
The taxon has a worldwide distribution and was previously reported from India (Sharma 2012). The description of the morphological features of the collected specimen matches that of the earlier report. The present collection does not reveal the presence of gasterospores and differs from the material reported from Portugal (Steyaert 1975). The collection reported from East Anglia by Corner (2009), however, varies a bit from the present collection with regard to the size of the basidiospores (9.5–12 × 6–6.5 µm vs 7–8.5 × 3.5–5 µm), which may be attributed to climatic and geographic variations.
Among macro-microscopically related taxa, Ganoderma flexipes differs by having a reddish-brown margin, chlamydospore in the context and trama, and larger basidiospores (7.5–10.5 × 6–7.5 µm); G. curtisii differs by having purplish-brown to black coloured pileus and larger spores (8.5–11 × 5–7 µm; Sharma 2012).
Ganoderma applanatum (Pers.) Pat. Hyménomyc. Eur. (Paris): 143 (1887) (Fig. 2; Image 2)
Basidiocarp sessile, dull, non-laccate, glabrous, perennial, woody, applanate, 50–86 × 30–35 mm in diameter, 35–44 mm thick. Upper surface of pileus greyish to reddish-brown (8D3, 9E4) with concentric zonation, sulcate. Margin 6–15 mm thick, round, reddish-brown (9E4) in colour. Pore surface whitish (1A1), pore 4–5 per mm, round. Tube multilayered (3–4 layered), dark brown (7F7). Context thick, single layered, dark brown (7F7).
Hyphal system trimitic. Generative hyphae of context thin walled, 4.5–5.5 µm wide, yellowish-brown (6D4, 5D7), clamp connection present at septa, difficult to observe; skeletal hyphae dominant, 4.5–6 µm wide, dark brown (9F4), branched; binding hyphae branched, hyaline, 1.7–2.5 µm wide. Generative hyphae of tube layer thin walled, 4–5.5 µm wide, yellowish-brown (6D4, 5D7), clamp connection present at septa, difficult to observe; skeletal hyphae dominant, 4.5–6.5 µm wide, dark brown (9F4), branched; binding hyphae branched, hyaline, 1.7–2.5 µm wide. Cutis irregular, trichodermis type with spores embedded within the gelatinous layer, cutis hyphae 1.5–5.5 µm wide, brown (7E5).
Basidiospore truncate, (6.5–)7.5–9(–10) × 3.5–4.5(–6) µm in diameter, Q = 1.6–2.1, Qm = 1.8, double walled, exospores hyaline, thin, endospore pale yellow to dark
Image 2. Ganoderma applanatum. Scale = 10mm
© K. Acharya
Figure 2. Ganoderma applanatum. a - hyphae of the cutis, b - basidiospores, c - context hyphae, d - tube layer hyphae. Bars = 5µm. Drawing by R. Saha.
a
c
b
d
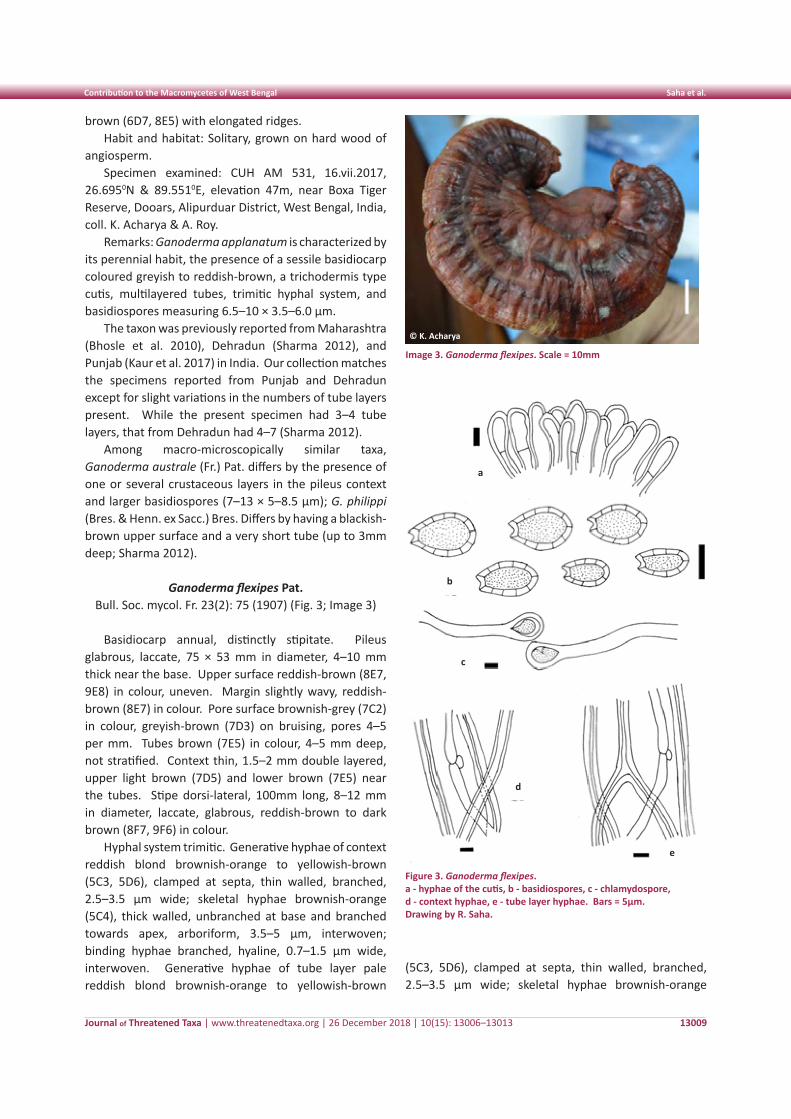
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13006–13013
Contribution to the Macromycetes of West Bengal Saha et al.
13009
brown (6D7, 8E5) with elongated ridges.Habit and habitat: Solitary, grown on hard wood of
angiosperm.Specimen examined: CUH AM 531, 16.vii.2017,
26.6950N & 89.5510E, elevation 47m, near Boxa Tiger Reserve, Dooars, Alipurduar District, West Bengal, India, coll. K. Acharya & A. Roy.
Remarks: Ganoderma applanatum is characterized by its perennial habit, the presence of a sessile basidiocarp coloured greyish to reddish-brown, a trichodermis type cutis, multilayered tubes, trimitic hyphal system, and basidiospores measuring 6.5–10 × 3.5–6.0 µm.
The taxon was previously reported from Maharashtra (Bhosle et al. 2010), Dehradun (Sharma 2012), and Punjab (Kaur et al. 2017) in India. Our collection matches the specimens reported from Punjab and Dehradun except for slight variations in the numbers of tube layers present. While the present specimen had 3–4 tube layers, that from Dehradun had 4–7 (Sharma 2012).
Among macro-microscopically similar taxa, Ganoderma australe (Fr.) Pat. differs by the presence of one or several crustaceous layers in the pileus context and larger basidiospores (7–13 × 5–8.5 µm); G. philippi (Bres. & Henn. ex Sacc.) Bres. Differs by having a blackish-brown upper surface and a very short tube (up to 3mm deep; Sharma 2012).
Ganoderma flexipes Pat. Bull. Soc. mycol. Fr. 23(2): 75 (1907) (Fig. 3; Image 3)
Basidiocarp annual, distinctly stipitate. Pileus glabrous, laccate, 75 × 53 mm in diameter, 4–10 mm thick near the base. Upper surface reddish-brown (8E7, 9E8) in colour, uneven. Margin slightly wavy, reddish-brown (8E7) in colour. Pore surface brownish-grey (7C2) in colour, greyish-brown (7D3) on bruising, pores 4–5 per mm. Tubes brown (7E5) in colour, 4–5 mm deep, not stratified. Context thin, 1.5–2 mm double layered, upper light brown (7D5) and lower brown (7E5) near the tubes. Stipe dorsi-lateral, 100mm long, 8–12 mm in diameter, laccate, glabrous, reddish-brown to dark brown (8F7, 9F6) in colour.
Hyphal system trimitic. Generative hyphae of context reddish blond brownish-orange to yellowish-brown (5C3, 5D6), clamped at septa, thin walled, branched, 2.5–3.5 µm wide; skeletal hyphae brownish-orange (5C4), thick walled, unbranched at base and branched towards apex, arboriform, 3.5–5 µm, interwoven; binding hyphae branched, hyaline, 0.7–1.5 µm wide, interwoven. Generative hyphae of tube layer pale reddish blond brownish-orange to yellowish-brown
(5C3, 5D6), clamped at septa, thin walled, branched, 2.5–3.5 µm wide; skeletal hyphae brownish-orange
Image 3. Ganoderma flexipes. Scale = 10mm
© K. Acharya
Figure 3. Ganoderma flexipes. a - hyphae of the cutis, b - basidiospores, c - chlamydospore, d - context hyphae, e - tube layer hyphae. Bars = 5µm. Drawing by R. Saha.
a
c
b
d
e

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13006–13013
Contribution to the Macromycetes of West Bengal Saha et al.
13010
(5C4), thick walled, unbranched at base and branched towards apex, arboriform, 3.5–5.5 µm, interwoven; binding hyphae branched, hyaline, 0.7–1.5 µm wide, interwoven. Cuticular hyphae regular, closely packed with clavate end cells, light brown (6D5) in colour, 4–7 µm wide. Basidiospores truncate, 7–9(–10) × 4–5.5(–6.5) µm in diameter, Q = 1.4–2.2, Qm = 1.7, bitunicate, exospores hyaline, thin walled, endospore light brown (7D6). Chlamydospore present in the context and trama, 9.5–10.5 µm in diameter, ovoid, reddish-blond brownish-orange (5C3) in colour.
Habit and habitat: solitary, grown on dead dicotyledonous wood.
Specimen examined: CUH AM534, 09.viii.2015, 22.5950N & 88.2630E, elevation 4m, Howrah, West Bengal, India, coll. K. Acharya.
Remarks: The present specimen is characterized by a laccate pileus coloured reddish-brown, distinctly double layered context (upper light brown and lower brown) near the tubes, a dorsi-lateral stipe, presence of closely packed clavate end cells at the cutis, basidiospores measuring 7–10 × 4–6.5 µm in diameter (Qm = 1.7), and presence of chlamydospore in the pileus context and trama.
Based on the artificial key proposed by Sharma (2012), the presence of chlamydospores characterises the specimen to be Ganoderma flexipes. This taxon has a worldwide distribution and was reported from China (Zhou et al. 2015), India (Uttarakhand) (Sharma 2012), and Vietnam (Steyaert 1972). Our present specimen mostly matches the description of that from India except for having a slightly larger chlamydospore (9.5–10.5 µm vs 6–9.5 µm). The specimen from China differs from the present collection with regard to slightly longer basidiospores (8.5–11 × 5–7 µm vs 7–10 x 4–6.5 µm).
The collection from Vietnam had slightly larger sized basidiocarps and comparatively larger basidiospores (8–13 × 5.5–8 µm).
Among morphologically related species, Ganoderma lucidum differs by the presence of white coloured pileus margin and the absence of chlamydospore; G. curtisii (Berk.) Murrill differs by the presence of purplish-brown to black pileus and the absence of chlamydospores; G. ahmadii differs by the presence of uniform coloured context and the absence of chlamydospores (Sharma 2012).
Ganoderma ahmadii Steyaert Persoonia 7(1): 91 (1972) (Fig. 4; Image 4)
Basidiocarp annual, laterally stipitate. Pileus glabrous, laccate, flabelliform, 45–55 × 60–75 mm in diameter, 4–8 mm thick near the base. Upper surface of pileus glabrous, laccate, shiny, dark brown (9F6). Margin lobed, whitish (1A1) to straw (4B1) coloured, thick. Pore surface whitish (1A1) to straw (4B1) in colour, greyish-brown (8D3) on bruising, pores 4–7 per mm. Tubes reddish-brown (9E4) in colour, 2–7 mm deep, not
Image 4. Ganoderma ahmadii. Scale = 10mm
© K. AcharyaFigure 4. Ganoderma ahmadii. a - hyphae of the cutis, b - basidiospores, c - context hyphae, d - tube layer hyphae. Bars = 5µm. Drawing by R. Saha.
a
c
b
d

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13006–13013
Contribution to the Macromycetes of West Bengal Saha et al.
13011
stratified. Context 1–5 mm thick, single layered, reddish-brown (8E4) in colour. Stipe dorsi-lateral, 100–125 mm long and 8–10 mm broad, laccate, shiny, glabrous, dark brown (8F5, 9F5) in colour.
Hyphal system trimitic. Generative hyphae of context greyish-brown (6D3), thin walled, branched, clamped at septa, difficult to observe, 3–4.5 µm wide; skeletal hyphae dominant, light brown (6D4), thick walled, unbranched at base and branched towards apex, arboriform, 4.5–6 µm, interwoven; binding hyphae branched, greyish-brown (6D3), septate, 1–2 µm wide, interwoven. Generative hyphae of tube layer greyish-brown (6D3), thin walled, branched, clamped at septa, difficult to observe, 3–4.5 µm wide; skeletal hyphae light brown (6D4), thick walled, unbranched at base and branched towards apex, arboriform, 4.5–5.5 µm, interwoven; binding hyphae branched, greyish-brown (6D3), septate, 0.7–1.5 µm wide, interwoven. Cuticular hyphae regular, closely packed with clavate end cells, light brown (6D4, 6D5) in colour, 7–11 µm wide. Basidiospores truncate, brown, bitunicate, (7.5–)8–9(–10.5) × 4–5(–6.5) µm in diameter, Q = 1.4–2.2, Qm = 1.7, exospores hyaline, thin walled, endospore thick, brown (6E6) with elongated ridges.
Habit and habitat: solitary, growing on dead and rotten Bambusa bambos wood.
Specimen examined: CUH AM530, 18.vii.2017, 26.3880N & 89.5260E, elevation 43m, Kaljani, Coochbehar District, West Bengal, India, coll. K. Acharya & A. Roy.
Remarks: Ganoderma ahmadii possesses characteristic features of an annual habit; laterally stipitate, laccate basidiocarp coloured reddish-brown to dark brown with white to straw coloured pileus margin; single layered context coloured reddish-brown; presence of a dorsi-lateral stipe measuring 100–125 mm long; pileus surface consisting of regular, closely packed clavate end cells; and truncate, double walled basidiospores measuring 7.5–10.5 × 4–6.5 µm in diameter.
Ganoderma ahmadii was previously reported from Uttarakhand, India (Sharma 2012). Our collection matches the previous report. The specimen reported from Pakistan differs by having double layered context coloured cinnamon buff towards the cutis and Verona brown near the tubes and larger spores (8–11 × 5.5–7 µm; Steyaert 1972).
Among the macro- and micro-morphologically related species, Ganoderma flexipes differs from G. ahmadii on the basis of the presence of chlamydospore in the context and trama. Ganoderma ahmadii is also morphologically different from G. applanatum on the
basis of size, shape, and colour of the basidiocarp. The basidiocarp of G. applanatum has sessile, non-laccate pileus with stratified tube, and trichodermis type cutis (Sharma 2012).
Ganoderma lucidum var. lucidum (Curtis) P. Karst. Revue mycol., Toulouse 3(no. 9): 17 (1881)
(Fig. 5; Image 5)
Basidiocarp annual, laterally stipitate. Pileus reniform, up to 90mm in diameter, 10–12 mm thick near the base. Upper surface reddish-brown (8D6, 8D7), shiny, glabrous, laccate. Margin thin, obtuse, white to orange grey (5A1, 5B2) in colour. Pore surface white (1A1), light brown (7D4) on bruising, pore sub orbicular, 4–5 per mm. Tubes not stratified, reddish-brown (8E5) in colour, 3–6 mm deep. Context soft, spongy, double layered, yellowish-brown (6E5) at upper side and brown (7E5) lower near to the tubes, 10–11 mm thick near the base. Stipe lateral, shiny, glabrous, laccate, reddish-brown (8E6), 50–70 mm in length and 10–20 mm broad.
Hyphal system trimitic. Generative hyphae of context light brown (6D5), thin walled, branched, 3.5–5 µm wide, clamped at septa, difficult to observe; skeletal hyphae dominant, brown (7E5), thick walled, unbranched at base and branched towards apex, 4.5–7 µm in breadth, interwoven; binding hyphae colourless, branched, 0.7–1.8 µm wide, interwoven. Cuticular hyphae regular, closely packed with clavate end cells, yellowish-brown (5D6) to light brown (7E5) in colour, thick walled, 4–11.5 µm wide. Basidiospores truncate, 5–6.5(–7.5) × 3.5–4.5(–5.5) µm in diameter, Q = 1.0–1.8, Qm = 1.5, bitunicate, exospores hyaline, thin-walled and endospore with elongated ridges, light brown (6D5) to brown (7E5) in colour.
Habit and habitat: Cespitose, grown on dead wooden log of angiosperm.
Specimen examined: CUH AM529, 25.vii.2016, 22.9920N & 88.5310E, elevation 10m, near Chandirampur, Birohi, Nadia, West Bengal, India, coll. K. Acharya.
Remarks: The present variety is easily recognized by the smaller size of the basidiospores (5.3–7.5 × 3.5–5.3 µm in diameter with Qm = 1.5). Ganoderma lucidum var. lucidum was previously reported from India (Maharastra, Bhosle et al. 2010). According to the key of Bhosle et al. (2010), under G. lucidum complex, if the spore index is 1.5, then the specimen is considered as G. lucidum var. lucidum. In the case of our collection, the spore index was 1.5. Hence, it could be identified as G. lucidum var. lucidum. Though the specimen from Maharashtra had a few larger spores (7–8.5 × 5–6 µm), the spore index was
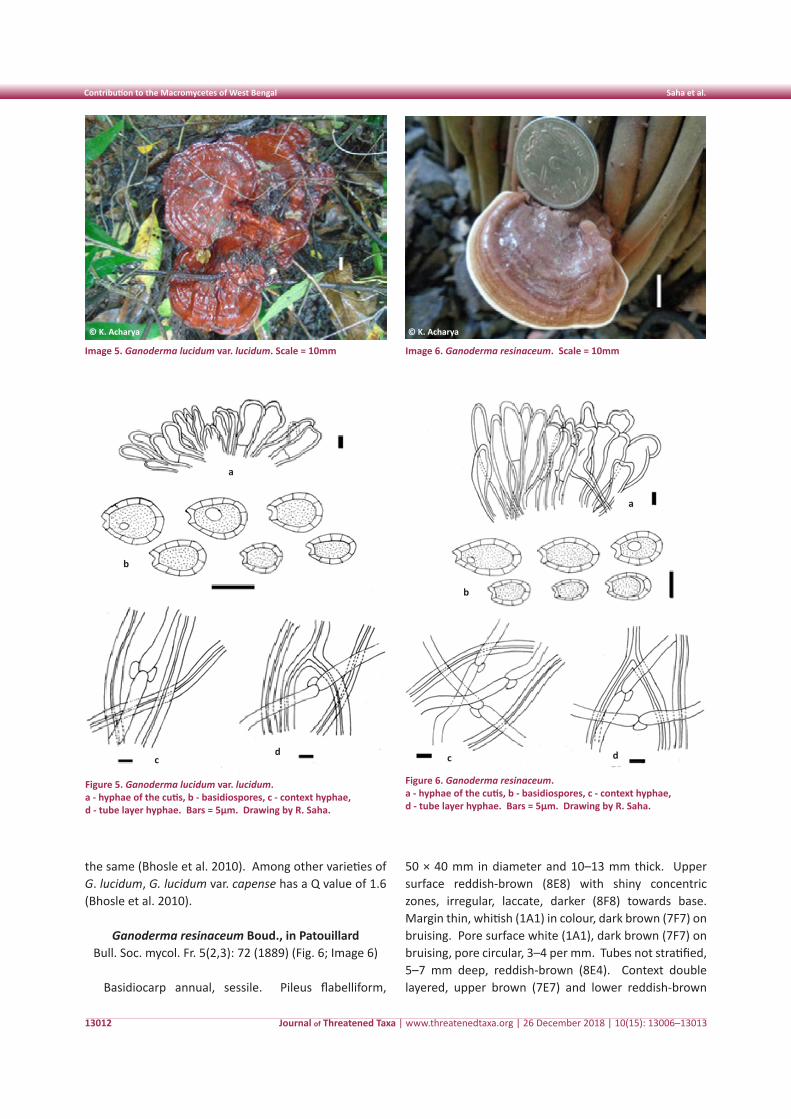
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13006–13013
Contribution to the Macromycetes of West Bengal Saha et al.
13012
Image 5. Ganoderma lucidum var. lucidum. Scale = 10mm
the same (Bhosle et al. 2010). Among other varieties of G. lucidum, G. lucidum var. capense has a Q value of 1.6 (Bhosle et al. 2010).
Ganoderma resinaceum Boud., in Patouillard Bull. Soc. mycol. Fr. 5(2,3): 72 (1889) (Fig. 6; Image 6)
Basidiocarp annual, sessile. Pileus flabelliform,
Image 6. Ganoderma resinaceum. Scale = 10mm
50 × 40 mm in diameter and 10–13 mm thick. Upper surface reddish-brown (8E8) with shiny concentric zones, irregular, laccate, darker (8F8) towards base. Margin thin, whitish (1A1) in colour, dark brown (7F7) on bruising. Pore surface white (1A1), dark brown (7F7) on bruising, pore circular, 3–4 per mm. Tubes not stratified, 5–7 mm deep, reddish-brown (8E4). Context double layered, upper brown (7E7) and lower reddish-brown
© K. Acharya© K. Acharya
Figure 5. Ganoderma lucidum var. lucidum. a - hyphae of the cutis, b - basidiospores, c - context hyphae, d - tube layer hyphae. Bars = 5µm. Drawing by R. Saha.
Figure 6. Ganoderma resinaceum. a - hyphae of the cutis, b - basidiospores, c - context hyphae, d - tube layer hyphae. Bars = 5µm. Drawing by R. Saha.
a
c
b
d
a
c
b
d

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13006–13013
Contribution to the Macromycetes of West Bengal Saha et al.
13013
(8E5) near the tubes, up to 10mm thick near the base. Stipe absent.
Hyphal system trimitic. Generative hyphae of context brown (7E6) to dark brown (7F7), thin walled, clamped at septa, difficult to observe, 3–4.5 µm in breadth; binding hyphae interwoven, 2.5–3.5 µm wide, hyaline; skeletal hyphae unbranched at base and branched towards apex, 3.5–5 µm wide, dark brown (7F7), interwoven. Generative hyphae of tube layer brown (7E6) to dark brown (7F7), thin walled, clamped at septa, difficult to observe, 3.5–4.5 µm in breadth; binding hyphae branched, 2.5–3.5 µm wide, hyaline; skeletal hyphae unbranched at base and branched towards apex, 3.5–5 µm wide, dark brown (7F7). Cuticular hyphae regular, closely packed with clavate end cells, brown (7E8) in colour, thick, 7–10 µm wide. Basidiospores truncate, (7–)9–12.5(–13.5) × 4–5.5(–7) µm in diameter, Q = 1.3–2.3, Qm = 1.7, bitunicate, exospores hyaline, thin and endospore thick, brown (7E8).
Habit and habitat: Solitary, grown on root of Areca catechu.
Specimen examined: CUH AM562, 13.xi.2017, 22.5270N & 88.3620E, elevation 13m, Ballygunge Science College campus, Kolkata, West Bengal, India, coll. K. Acharya.
Remarks: Ganoderma resinaceum is characterized by features like a sessile, laccate basidiocarp coloured reddish-brown with white margin, double layered context separated by brownish upper and reddish-brown lower side near the tubes, the presence of closely packed clavate end cells at the cutis, and truncate basidiospores measuring 7–13.5 × 4–7 µm.
Ganoderma resinaceum was earlier reported from Italy (Corner 1983) and India (Bhosle et al. 2010; Sharma 2012). Our collection matches those in earlier reports.
Among morphologically related species, Ganoderma lucidum has a distinct stipe, G. flexipes has chlamydospores, and G. applanatum differs by the presence of trichodermis type cutis (Sharma 2012).
References
Bhosle, S., K. Ranadive, G. Bapat, S. Garad, G. Deshpande & J. Vaidya (2010). Taxonomy and diversity of Ganoderma from the western parts of Maharastra (India). Mycosphere 1(3): 249–262.
Corner, E.J.H. (1983). Ad Polyporaceas I. Amauroderma and Ganoderma. Beihefte zur Nova Hedwigia 75: 1–182.
Kaur, G., A.P. Singh & G.S. Dhingra (2017). Diversity of the genus Ganoderma in Punjab (India). Mycobiota 7: 25–49; https://doi.org/10.12664/mycobiota.2017.07.05
Kornerup, A. & J.H. Wanscher (1978). Methuen Handbook of Colour. Methuen Publishing Ltd, London, 256pp.
Pradhan, P., A.K. Dutta & K. Acharya (2015). A low cost long term preservation of macromycetes for fungarium. Protocol Exchange. Available online 17 March 2015; https://doi.org/10.1038/protex.2015.026
Pradhan, P., A.K. Dutta, S. Paloi, A. Roy & K. Acharya (2016). Diversity and distribution of macrofungi in the eastern Himalayan ecosystem. EurAsian Journal of BioSciences 10: 1–12.
Sharma, J.R. (2012). Aphylloporales of Himalaya. Botanical Survey of India, 590pp.
Steyaert, R.L. (1972). Species of Ganoderma and related genera mainly of the Bogor and Leiden herbaria. Persoonia 7(1): 55–118.
Steyaert, R.L. (1975). Ganoderma lucidum. CMI Descriptions of Pathogenic Fungi and Bacteria 445: 1–2.
Zhou, L.W., Y. Cao, S.H. Wu, J. Vlasak, D.W. Li, M.J. Li & Y.C. Dai (2015). Global diversity of the Ganoderma lucidum complex (Ganodermataceae, Polyporales) inferred from morphology and multilocus phylogeny. Phytochemistry 114: 7–15.
Key to the species Ganoderma reported in the study
1. Pileus upper surface laccate; hyphae of the cutis regular, closely packed with clavate end cells ........... 21a. Pileus upper surface non-laccate; hyphae of the cutis irregular, clavate end cells absent ..... G. applanatum2. Pileal context double layered, upper yellowish-brown and lower brown to dark brown near the tubes ...... 32a. Pileal context uniformly coloured brown to dark brown .......................................................... G. ahmadii3. Basidiocarp containing a well-developed stipe ...................................................................................... 43a. Basidiocarp sessile ............................................................................................................... G. resinaceum4. Chlamydospore present in the context and trama; fruitbody reddish-brown with concolorous margin ............................................................................................................................................................... G. flexipes4a. Chlamydospore absent; fruit body reddish brown with discolorous, white margin …......................…… 55. Basidiospores measuring 7–8.5 × 3.5–5 µm with Qm > 1.5 …………………...........................………. G. lucidum5a. Basidiospores smaller, measuring 5–7.5 × 3.5–5.5 µm with Qm of 1.5 ................... G. lucidum var. lucidum
Threatened Taxa

13014
Not
e
DOI: https://doi.org/10.11609/jott.3357.10.15.13014-13016 | ZooBank: urn:lsid:zoobank.org:pub:585E1C6D-259B-42DD-8BD2-64DD9EA517DF
Editor: Nishith A. Dharaiya, HNG University, Patan, India. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 3357 | Received 21 February 2017 | Final received 07 December 2018 | Finally accepted 10 December 2018
Citation: Kamalakannan, M. (2018). The identification of Takin Budorcas taxicolor (Mammalia: Bovidae) through dorsal guard hair. Journal of Threatened Taxa 10(15): 13014–13016; https://doi.org/10.11609/jott.3357.10.15.13014-13016
Copyright: © Kamalakannan 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: Ministry of Environment, Forest and Climate Change, Govt. of India.
Competing interests: The author declares no competing interests.
Acknowledgments: I sincerely thank Dr. Kailash Chandra, director, Dr. C. Venkatraman, officer-in-charge of the Mammal & Osteology Section, ,and Dr. J.K. De, former scientist, of the Zoological Survey of India ,for providing the facilities and encouragement to accomplish this work.
The identification of Takin Budorcas taxicolor (Mammalia: Bovidae) through
dorsal guard hair
Manokaran Kamalakannan
Mammal & Osteology Section, Zoological Survey of India, M-Block, New Alipore, Kolkata, West Bengal 700053, India
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13014–13016
Takin Budorcas taxicolor Hodgson (1850) is a heavily built and clumsy-looking animal, native to northeastern India, Bhutan, China, and northern Myanmar (Salter 1997). Budorcas taxicolor is usually identified by its external morphology, i.e., long, shaggy coat that varies from golden yellow to deep dark brown, a dark stripe
running on the dorsal side from head to tail, and a dark brown face (Menon 2014). In the present study, however, it was found that B. taxicolor can also be identified with the help of its hair samples. Mammalian hairs have certain advantages from the viewpoint of taxonomy and systematics (Sarkar et al. 2011). There are many works worldwide on hair identification of different species of mammals (Mayer 1952; Brunner & Coman 1974; Moore et al. 1974; Teerink 1991). In India, studies on the hair of mammals were carried out on Artiodactyla (Koppikar & Sabins 1976), Rodentia (Sarkar 2012), Carnivora (Chakraborty & De 2010), and Primates (Sarkar et al. 2011). This study attempts to find out the hair characteristics of B. taxicolor, which was hitherto unknown.
A total of five tufts of dorsal guard hair was collected from the five preserved skins of B. taxicolor housed at the National Zoological Collections of the Zoological Survey of India, Kolkata, India, and was processed by following the method of Teerink (1991). Subsequently, the morphological characteristics of hair such as total length, diameter, and profile (n=20) were recorded using a dial calliper and hand lens. The cuticular scale
characteristics were studied according to the standard methodology (Brunner & Coman 1974; Teerink 1991) and the scale pattern, margin, margin distance, and count of the hair were studied by moulding the hair in clear varnish overnight and observing the impressions of its cuticular scales. The medullary characteristics such as composition, structure, and margins were recorded from the hair cleaned and mounted in a solution of xylene and DPX (50:50), after Chakraborty et al. (1996). The transverse section of hair too was performed as per Chakraborty et al. (1996). Different terminologies were followed according to Brunner & Coman (1974) and Teerink (1991). The photomicrographs were taken using a camera fitted with an optical light microscope (Olympus BX41) and scanning electron microscope (ZEISS Evo18 - special edition).
The hair of B. taxicolor can be easily identified by its morphologic and microscopic characteristics (Table 1). The dorsal guard hair of B. taxicolor studied is bicoloured, with alternated bands of earth yellow and coffee colours. The profile of the hair is undulated. The total length of hair varied greatly from 13.6mm to 51.6mm (30.4±12.8mm); similarly, the diameter of hair varied
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13014–13016
Identification of Takin through dorsal guard hair Kamalakannan
13015
from 84.9µm to 258.8µm (215.6±25.2µm). The cuticular characteristics were recorded as follows: scale position - transversal, scale patterns - irregular wave, the structure of scale margins - rippled, and the distance between scale margins - near (Images 1a,b). All the measurement values of cuticular scales varied greatly and the average values were recorded as follows: cuticular scale count (per mm length of hair): 146.4±15.8µm, length of cuticular scale 98.8±12.1µm, and width of cuticular scale 11.1±1.1µm. The medullary characteristics were recorded as follows: the composition of medulla - unicellular irregular, the structure of medulla - uniserial ladder, and form of the medulla margins - scalloped (Image 1c); the average width of the medulla was recorded as 65.6±3.9µm. The shape of the transverse section was observed as circular (Image 1d).
A study by Kamalakannan (2015) on a total of 17 species of artiodactyls (11 bovids, four cervids, one suid, and one mouse deer) was found that the microscopic characteristics of hair of all the species were nearly the same, except in B. taxicolor. The dorsal guard hair of B. taxicolor possesses completely unique microscopic characteristics, especially that of the medulla (Image 1c) that differs from other species of mammals. According to the study, the unique structure of the medulla, uniserial ladder, was found only in B. taxicolor and was not reported earlier. The irregular wave of scale patterns, the rippled scale margins of the cuticle, and the circular shape of a transverse section of hair also determined the species identity of B. taxicolor, as these characteristics are infrequent in other species of mammals.
Methods of hair identification need exact identification keys (Brunner & Coman 1974; Teerink
Table 1. Morphologic and microscopic characteristics of dorsal guard hair of Budorcas taxicolor
Hair characteristics Result Hair characteristics Result
Coat colour Brownish-grey Distance between cuticular scale margins Near
Colour of hair Bicoloured; base: earth yellow, tip: coffee
Cuticular scale count/mm length of hair 123–167 (146.4±15.8)
Number of colour bands Two Length of cuticular scale (µm) 84.6–120 (98.8±12.1)
Profile Undulated Width of cuticular scale (µm) 9.1–12.5 (11.1±1.1)
Length (mm) 13.6–51.6 (30.4±12.8) Composition of medulla Unicellular irregular
Diameter (µm) 84.9–258.8 (215.6±25.2) Structure of medulla Uniserial ladder
Cuticular scale position Transversal Margins of medulla Scalloped
Cuticular scale patterns Irregular wave Width of medulla (µm) 56.3–70.3
Cuticular scale margins Rippled Transverse section Circular
1991) as they have some similarities between the species. Hair identification keys of the family Bovidae are much required in the field of forensic science and predator diet analysis for species identification (Sahajibal et al. 2010; Dharaiya & Soni 2012). Budorcas taxicolor is a Vulnerable species as per the IUCN Red List of Threatened Species (2018) and is listed under the Schedule-I of the Indian Wildlife (Protection) Act, 1972, and Appendix-II of CITES (Song et al. 2008). It is trafficked for its meat, which is consumed locally, its skin, and other derivatives (Menon & Kumar 1999). On the other hand, it is also the chief prey of large carnivores. Therefore, the identification keys along with the photomicrographs presented here can be used in animal forensic science as well as in predator diet analysis as an appropriate reference for species identification of B. taxicolor.
References
Brunner, H. & B.J. Coman (1974). The Identification of Mammalian Hair. Inkata Press, Melbourne, Australia, 176pp.
Chakraborty, R. & J.K. De (2010). Atlas on Hairs of Indian Mammals, Part I: Carnivora. Zoological Survey of India, Kolkata,141pp.
Chakraborty, R., J.K. De & S. Chakraborty (1996). Identification of dorsal guard hairs of Indian species of the genus Panthera Oken (Carnivora: Felidae). Mammalia 60(3): 473–480.
Dharaiya, N. & V.C. Soni (2012). Identification of hairs of some mammalian prey of large cats in Gir Protected Area, India. Journal of Threatened Taxa 4(9): 2928–2932.
Kamalakannan, M. (2015). Tricho-taxonomic studies of Indian mammal species belonging to the orders Artiodactyla and Lagomorpha. PhD Thesis. University of Kalyani, West Bengal, India.
Koppikar, B.R. & J.H. Sabins (1976). Identification of hairs of some Indian mammals. Journal of the Bombay Natural History and Society 73: 5–20.
Mayer, W.V. (1952). The hair of California mammals with keys to the dorsal guard hairs of California mammals. American Midland Naturalist 38: 480–512.
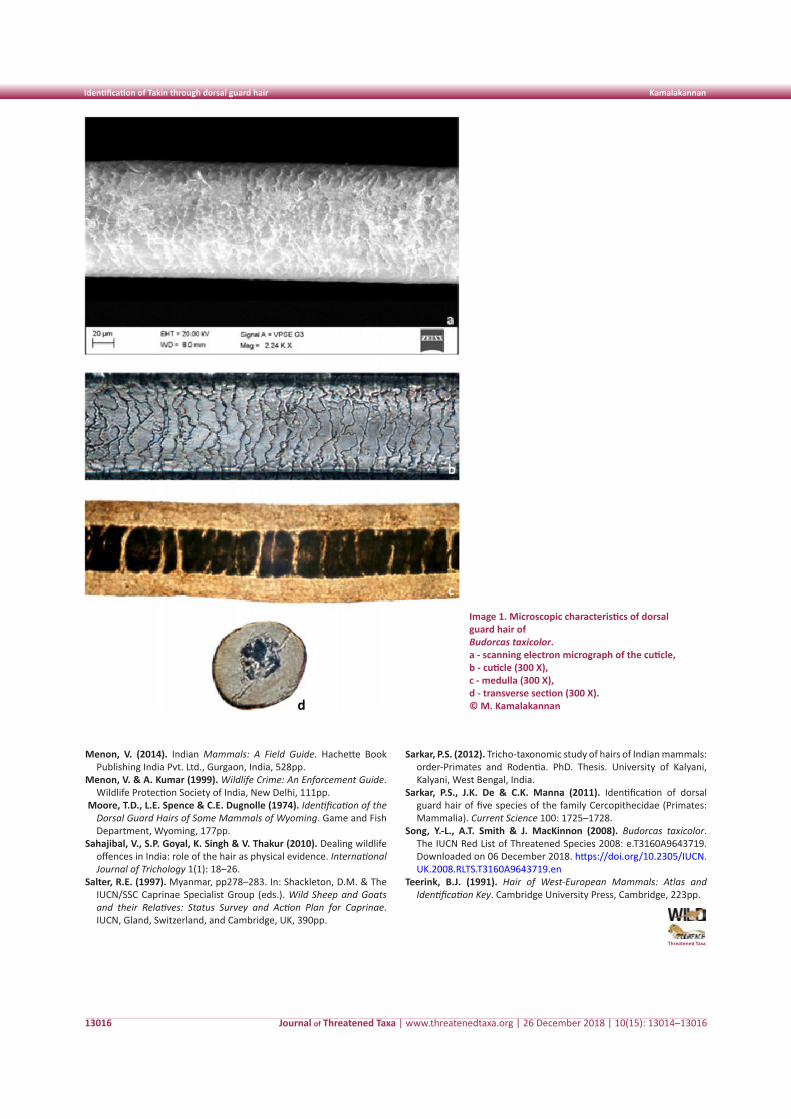
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13014–13016
Identification of Takin through dorsal guard hair Kamalakannan
13016
Menon, V. (2014). Indian Mammals: A Field Guide. Hachette Book Publishing India Pvt. Ltd., Gurgaon, India, 528pp.
Menon, V. & A. Kumar (1999). Wildlife Crime: An Enforcement Guide. Wildlife Protection Society of India, New Delhi, 111pp.
Moore, T.D., L.E. Spence & C.E. Dugnolle (1974). Identification of the Dorsal Guard Hairs of Some Mammals of Wyoming. Game and Fish Department, Wyoming, 177pp.
Sahajibal, V., S.P. Goyal, K. Singh & V. Thakur (2010). Dealing wildlife offences in India: role of the hair as physical evidence. International Journal of Trichology 1(1): 18–26.
Salter, R.E. (1997). Myanmar, pp278–283. In: Shackleton, D.M. & The IUCN/SSC Caprinae Specialist Group (eds.). Wild Sheep and Goats and their Relatives: Status Survey and Action Plan for Caprinae. IUCN, Gland, Switzerland, and Cambridge, UK, 390pp.
Threatened Taxa
Image 1. Microscopic characteristics of dorsal guard hair of Budorcas taxicolor. a - scanning electron micrograph of the cuticle, b - cuticle (300 X), c - medulla (300 X), d - transverse section (300 X). © M. Kamalakannan
Sarkar, P.S. (2012). Tricho-taxonomic study of hairs of Indian mammals: order-Primates and Rodentia. PhD. Thesis. University of Kalyani, Kalyani, West Bengal, India.
Sarkar, P.S., J.K. De & C.K. Manna (2011). Identification of dorsal guard hair of five species of the family Cercopithecidae (Primates: Mammalia). Current Science 100: 1725–1728.
Song, Y.-L., A.T. Smith & J. MacKinnon (2008). Budorcas taxicolor. The IUCN Red List of Threatened Species 2008: e.T3160A9643719. Downloaded on 06 December 2018. https://doi.org/10.2305/IUCN.UK.2008.RLTS.T3160A9643719.en
Teerink, B.J. (1991). Hair of West-European Mammals: Atlas and Identification Key. Cambridge University Press, Cambridge, 223pp.

13017
Not
e
DOI: https://doi.org/10.11609/jott.3555.10.15.13017–13019 | ZooBank: urn:lsid:zoobank.org:pub:34410CCD-B11B-40E8-A56C-D6DF78EA5867
Editor: Merwyn Fernandes, TRAFFIC, New Delhi, India. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 3555 | Received 10 June 2017 | Final received 21 August 2018 | Finally accepted 05 November 2018
Citation: Maurya, V., J.P. Singh, K. Naseem, S. Mehra, P.M. Dhakate, N. Verma & A.G. Ansari (2018). Photographic evidence of Striped Hyena Hyaena hyaena (Mammalia: Carnivora: Hyaenidae) in Ramnagar forest division, Uttarakhand, India. Journal of Threatened Taxa 10(15): 13017–13019; https://doi.org/10.11609/jott.3555.10.15.13017–13019
Copyright: © Maurya et al. 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: None.
Competing interests: The authors declare no competing interests.
Acknowledgements: We are grateful to the additional principal conservator chief of forests (APCCF), Mr. Paramjit Singh, for his constant guidance and encouragement. We are thankful to the staff of the Ramnagar Forest Division for their valuable support during the field survey. Also, I thank my colleagues Dr. Shankar Kumar and Dr. Chaitenya Verma for providing the necessary information and support.
Photographic evidence of Striped Hyena Hyaena hyaena (Mammalia: Carnivora: Hyaenidae) in Ramnagar forest division, Uttarakhand, India
Vipul Maurya 1 , Jai Pratap Singh 2 , Kahkashan Naseem 3 , Surender Mehra 4 , Parag M. Dhakate 5 , Neha Verma 6 & A.G. Ansari 7
1 School of Forestry and Environment, Sam Higginbottom Institute of Agriculture, Technology and Sciences, Allahabad, Uttar Pradesh 211007, India2 Resham Majri Village, Dehradun, Uttarakhand 248140, India 3 Divisional Forest Officer, Tarai West Forest Division, Uttarakhand 244715, India 4 Director, Corbett Tiger Reserve, Ramnagar, Nainital District, Uttarakhand 244715, India 5 Conservator of Forests, Western Circle, Haldwani, Uttarakhand 263139, India6 Divisional Forest Officer, Ramnagar Forest Division, Uttarakhand 244715, India 7 Conservationist, Village-Mohan, Ramnagar, Nainital, Uttarakhand 244715, India1 [email protected] (corresponding author), 2 [email protected], 3 [email protected], 4 [email protected],5 [email protected], 6 [email protected], 7 [email protected]
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13017–13019
The Striped Hyena Hyaena hyaena has a large but inconsistent range extending from eastern Africa through the Middle East to India. The Striped Hyena is classified as Near Threatened on the IUCN Red List because of persecution, depletion of prey, and its habitat being converted to agricultural lands (Arumugam et al. 2008). It is also a known fact that hyenas are scavengers in their
habitat (Kruk 1976; Prater 1980; MacDonald 1984; Boitani & Bartoli 1986; Hofer 1998; Menon & Daniel 2003) but there were accounts of them feeding on insects, reptiles, rodents, birds, vegetables, and livestock (Heptner & Sludsky 1972; Rieger 1981; Mills & Hofer 1998; Lukarevsky 2001; Singh et al. 2010). The species is known to exist around human settlements and to survive by consuming dried bones, carcasses, and fruits (Kruk 1976; Hofer 1998). In India, Striped Hyenas occur in arid and semi-arid ecosystems (Alam et al. 2014) and are also known to occur sympatrically with tigers (Prater 1971; Menon 2003; Harihar et al. 2010).
There is considerable evidence of the presence of hyenas in the Terai region of northern India — the first estimation of their population carried out in a moist mixed deciduous forest in Rajaji National Park indicated the presence of the species in low density, as these areas are highly disturbed. These habitats have a notable population of tigers and are similar to that of Ramnagar Forest Division (Harihar et al. 2010). Hyenas are one of the least studied large scavengers/carnivore species in India due to the dearth of its observed records. For instance, the Striped Hyena was last reported in Ramnagar Division, Uttarakhand, in the late 1970s (Working Plan Ramnagar Forest Division 1977).
This study was carried out in the Ramnagar Forest Division located in Uttarakhand (Fig. 1). The geographic extent of the area is from 78.10′–79.10 0E and 29.56–

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13017–13019
Striped Hyena in Ramnagar forest division, Uttarakhand Maurya et al.
13018
29.55 0N and it covers a total of 48,736.90ha. The Ramnagar Forest Division is a forested landscape on the eastern boundary of the Corbett Tiger Reserve and northwestern boundary of the Terai Central Forest Division and Terai West Forest Division. It falls within the western circle of the Kumaun Forest and constitutes five forest ranges, namely, Kosi, Kota, Dechauri, Fatehpur, and Kaladhungi, which are interconnected by a trail of dense mixed forests from the eastern boundary of the Corbett Tiger Reserve up to Kaladhungi. The area is rich in faunal diversity, namely, tigers, elephants, leopards, ungulates, reptiles, and fish. The study targeted three of the forest ranges, namely, Dechauri, Kaladhungi, and Kota, which were identified on the basis of the presence of Hyena signs during a preliminary survey conducted from March to December 2015. These ranges were further divided into grids of 3km×3km and transects were laid in each grid. Confirmed signs for the species were recorded from 11 grids; these signs formed the basis for sites of camera traps, which provided photographic evidence of the species.
Here we report the first photographic image of a Striped Hyena in Ramnagar Forest Division, Uttarakhand (Image 1a captured on 12-07-2015 at 10:40hr). During camera trap survey from March to December 2015, a single image of a Striped Hyena was captured on camera. A total of 26 signs (Image 1b representing fresh pug-mark sign captured on 06-07-2015 at 07:15hr) were recorded during the survey at 11 different grids. The overall encounter rate was 1.39/km. The camera point where the image was captured was near Haathi Galiyaar Forest Barrier in Ramnagar Forest Division (29.550N / 79.250E) from which the nearest human settlement and water body (Dabka River) are 2km and 1km away, respectively. During the previous surveys conducted for monitoring Tigers, co-predators, prey and their habitats conducted in 2010, 2012, 2014, and 2016, there was no indirect sign/photographic evidence to confirm the presence of hyenas in the area. The only information regarding its presence was mentioned in the Working Plan Ramnagar Forest Division (1977). In the working plans published after 1977, there was no mention of this particular
Figure 1. Map showing grids surveyed for the presence of Striped Hyena and locations of camera traps in Ramnagar Forest Division, Uttarakhand

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13017–13019
Striped Hyena in Ramnagar forest division, Uttarakhand Maurya et al.
13019
Image 1. a - Camera trap image of Striped Hyena Hyaena hyaena recorded near Haathi Galiyaar Forest Barrier, b - pug mark of hyena found in Ramnagar Forest Division, Uttarakhand
species. The substantial proof of the presence of the Striped Hyena in Ramnagar Forest Division presented in this study can create opportunities for further research on the ecology, behaviour, and population estimation specific for the species in this region.
References
Alam, M.S., J.A. Khan, S.P. Kushwaha, R. Agrawal, B.J. Pathak & S. Kumar (2014). Assessment of suitable habitat of Near Threatened Striped Hyena (Hyaena hyaena Linnaeus, 1758) using remote sensing and geographic information system. Asian Journal of Geoinformatics 14(2): 860–866.
Arumugam, R., A. Wagner & G. Mills (2008). Hyaena hyaena. In: IUCN Red List of Threatened Species, Version 2. http://www.iucnredlist. org. Downloaded on 10 september 2017.
Boitani, L. & S. Bartoli (1986). The MacDonald Encyclopedia of Mammals. MacDonald & Co. Ltd., London, 157–164pp.
Harihar, A., M. Ghosh, M. Fernandes, B. Pandav & S.P. Goyal (2010). Use of photographic capture-recapture sampling to estimate density of Striped Hyena (Hyaena hyaena): implications for conservation. Mammalia 74(1): 83–87; https://doi.org/10.1515/mamm.2009.072
Heptner, V.G. & A.A. Sludsky (1972). Mammals of the Soviet Union: Carnivores (Hyenas and Cats), Vol. 2, Part 2. Vysshaya Shkola, Moskva, 784pp.
Hofer H. (1998). Species accounts, pp. 18–38. In: Mills G. & H. Hofer (eds.). Status Survey and Conservation Action Plan Hyenas. IUCN/SSC Hyaena Specialist Group, Oxford, U.K.
Kruuk, H. (1976). Feeding and social behavior of the Striped Hyaena (Hyaena vulgaris Desmarest). African Journal of Ecology 14(2): 91–111.
Lukarevsky, V.S. (2001). The Leopard, Striped Hyena, and Wolf in Turkmenistan. Signar, Moscow, 128pp.
MacDonald, D.W. (Ed.) (1984). The Encyclopaedia of Mammals. Greenwich Editions, London, 154–159pp.
Menon, V. (2003). Field Guide to Indian Mammals. Dorling Kindersley & Penguin Books, India, 201pp.
Mills, G. & H. Hofer (1998). Status Survey and Conservation Action Plan Hyenas. IUCN/SSC Hyaena Specialist Group, Oxford, U.K., 109–130pp.
Prater, S.H. (1971). The Book of Indian Animals. Oxford University Press, Oxford, 347pp.
Prater, S.H. (1980). The Book of Indian Animals. Bombay Natural History Society, Bombay & Oxford University Press, London, 324pp.
Rieger, I. (1981). Hyaena hyaena. Mammal Species 150: 1–5.Singh, P., A.M. Gopalaswamy & K.U. Karanth (2010). Factors
influencing densities of Striped Hyenas (Hyaena hyaena) in arid regions of India. Journal of Mammalogy 91(5): 1152–1159.
Working Plan Ramnagar Forest Division (1977). Western Circle, Uttar Pradesh.
Threatened Taxa

13020
Not
e
LOGOs
DOI: https://doi.org/10.11609/jott.4434.10.15.13020-13023 | ZooBank: urn:lsid:zoobank.org:pub:C102DEC2-A2D5-44E0-BCF4-962E18FA7560
Editor: Paul Racey, University of Exeter, UK. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 4434 | Received 26 July 2018 | Final received 29 October 2018 | Finally accepted 05 November 2018
Citation: Kamalakannan, M., C. Venkatraman, T.H. Dar & K. Chandra (2018). Range extension of the Least Leaf-nosed Bat Hipposideros cineraceus Blyth, 1853 (Mam-malia: Chiroptera: Hipposideridae): to central India. Journal of Threatened Taxa 10(15): 13020–13023; https://doi.org/10.11609/jott.2964.10.15.13020-13023
Copyright: © Kamalakannan et al. 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, repro-duction and distribution by providing adequate credit to the authors and the source of publication.
Funding: Ministry of Environment, Forest and Climate Change, Govt. of India.
Competing interests: The authors declare no competing interests.
Range extension of the Least Leaf-nosed Bat Hipposideros cineraceus Blyth, 1853
(Mammalia: Chiroptera: Hipposideridae): to central India
M. Kamalakannan 1 , C. Venkatraman 2 , Tauseef Hamid Dar 3 & Kailash Chandra 4
1,2,3,4 Zoological Survey of India, Prani Vigyan Bhawan, Block M, New Alipore, Kolkata, West Bengal 700053, India
1 [email protected] (corresponding author), 2 [email protected], 3 [email protected],
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13020–13023
The Least Leaf-nosed Bat Hipposideros cineraceus Blyth, 1853 is small in size and one of the nine species of leaf-nosed bats (Family: Hipposideridae) of India (Bates & Harrison 1997; Wilson & Reeder 2005) and 83 species of the world (Murray et al. 2012). It is wide ranging but sparsely distributed from South Asia to Southeast Asia,
and it has been recorded in only a few locations in India, viz., Uttarakhand (Scully 1887), Meghalaya (Hinton & Lindsay 1926), West Bengal, Assam, and Arunachal Pradesh (Bates & Harrison 1997) at an elevation ranging from 62–1,480 m (Bates & Harrison 1997; Molur et al. 2002). There is little information available on the natural history of this species; it roosts in hollows of trees in forests (Bhat & Jacob 1990; Bates & Harrison 1997; Molur et al. 2002).
On 24 December 2004, three bat specimens were collected using a mist net by a survey team of the Zoological Survey of India (ZSI) from N.P. Kailash Cave in Kanger Khati National Park (18.77870N & 81.99710E; Fig. 1), Jagadalpur District of Chhattisgarh. The wet preserved specimens were misidentified as male specimens of H. cineraceus and deposited in the National Zoological Collections (NZC) of Mammal & Osteology section, ZSI, Kolkata under the registration numbers 25794, 25795 & 25796 (Image 1a). The authors have recently re-examined the specimens in NZC and found that they are female specimens of H. cineraceus and were not as reported earlier by the collector. The specimens were identified as H. cineraceus based on the keys provided by Bates & Harrison (1997) and Douangboubpha et al. (2010).
Identification characteristics: Hipposideros cineraceus can easily be diagnosed through its internarial septum and the anterior leaf (Image 1b) from related species such as H. ater and H. durgadasi (bicolor species). In H. cineraceus, the internarial septum is slightly triangular in shape and its tip is blunt, and there is a slight emargination on the anterior leaf without any supplementary leaflets (Image 2a). In H. ater, the internarial septum is clearly triangular in shape and its tip is pointed, and there is no emargination on the anterior leaf, with one pair of rudimentary supplementary leaflets (Image 2b). In H. durgadasi, the internarial septum is bulbous in shape and
Abbreviations used: External measurements: FA: Forearm length; HB: Head Body length; T: Tail Length; Tail tip: Tail tip length; E: Ear Length; HF: Hindfoot length; Hw: Horseshoe width; Tib: Tibia length; 3mt: Length of the third metacarpal; 4mt: Length of the fourth metacarpal; 5mt: Length of the fifth metacarpal; 1st ph3rd D: First phalanx of the third Digit; 2nd ph3rdD: Second phalanx of the third Digit; 1st ph4th D: Length of the first phalanx of the fourth digit; 2nd ph4thD: Length of the second phalanx of the fourth Digit.
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
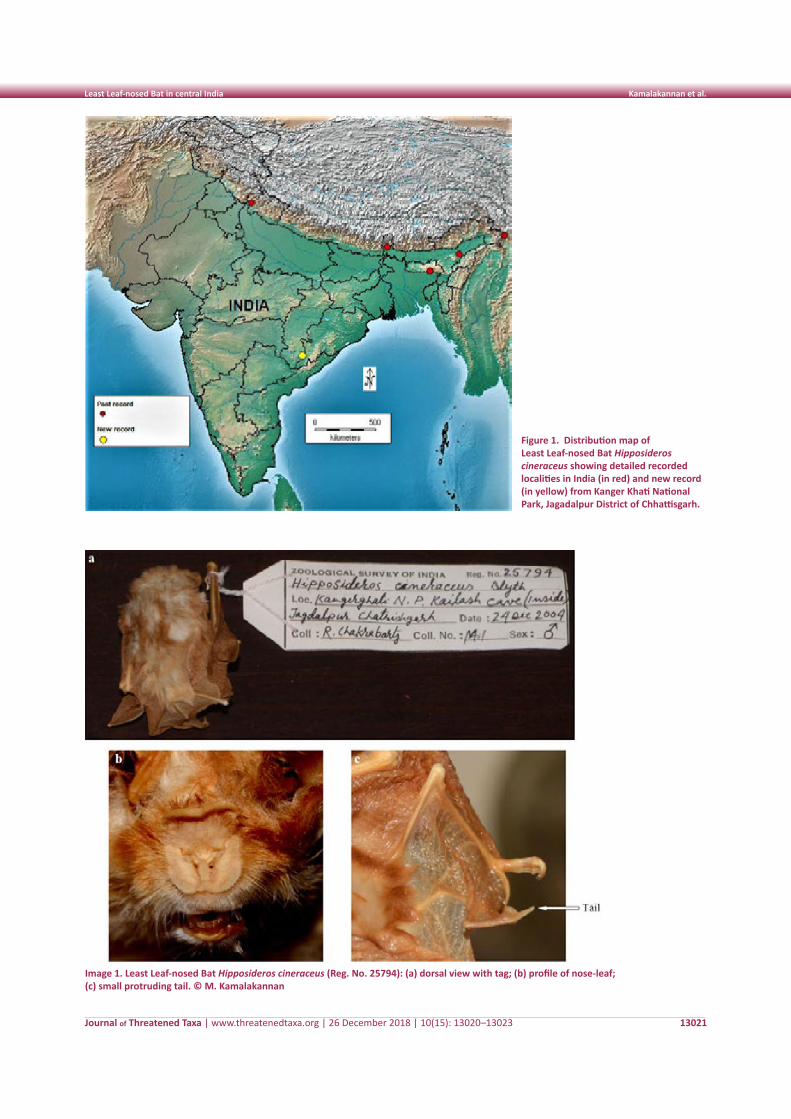
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13020–13023
Least Leaf-nosed Bat in central India Kamalakannan et al.
13021
Figure 1. Distribution map of Least Leaf-nosed Bat Hipposideros cineraceus showing detailed recorded localities in India (in red) and new record (in yellow) from Kanger Khati National Park, Jagadalpur District of Chhattisgarh.
Image 1. Least Leaf-nosed Bat Hipposideros cineraceus (Reg. No. 25794): (a) dorsal view with tag; (b) profile of nose-leaf; (c) small protruding tail. © M. Kamalakannan
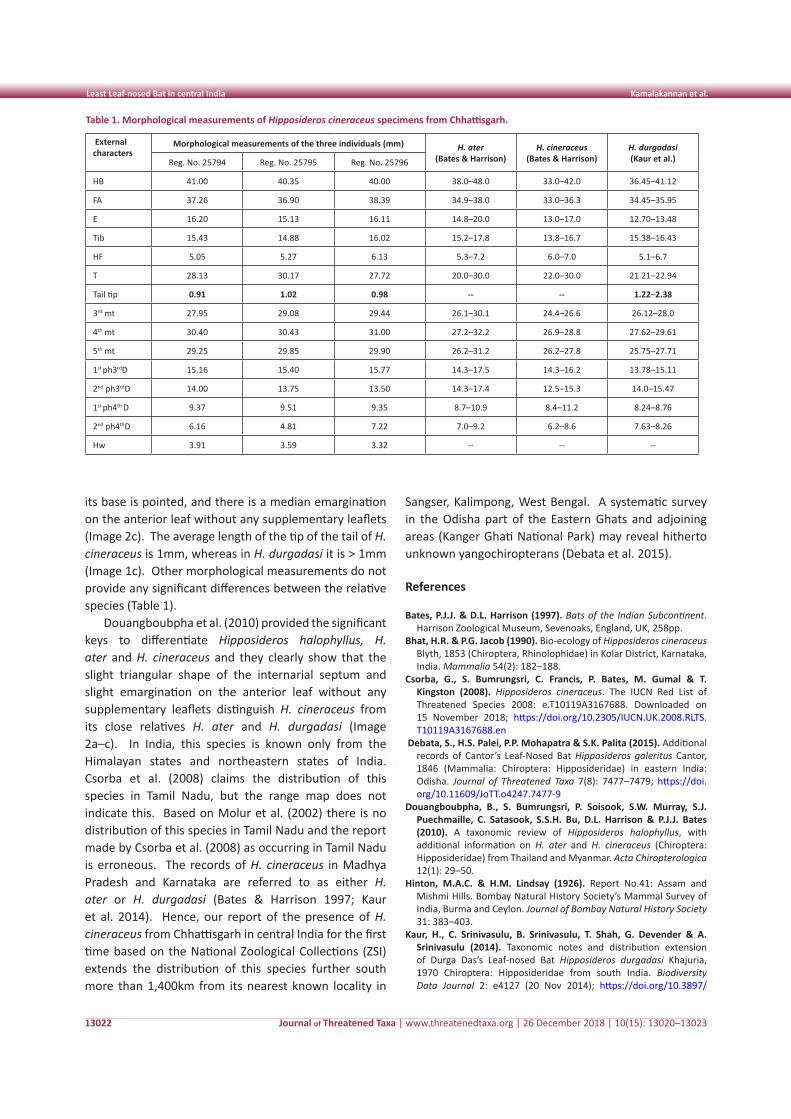
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13020–13023
Least Leaf-nosed Bat in central India Kamalakannan et al.
13022
Table 1. Morphological measurements of Hipposideros cineraceus specimens from Chhattisgarh.
External characters
Morphological measurements of the three individuals (mm) H. ater(Bates & Harrison)
H. cineraceus(Bates & Harrison)
H. durgadasi(Kaur et al.)Reg. No. 25794 Reg. No. 25795 Reg. No. 25796
HB 41.00 40.35 40.00 38.0–48.0 33.0–42.0 36.45–41.12
FA 37.26 36.90 38.39 34.9–38.0 33.0–36.3 34.45–35.95
E 16.20 15.13 16.11 14.8–20.0 13.0–17.0 12.70–13.48
Tib 15.43 14.88 16.02 15.2–17.8 13.8–16.7 15.38–16.43
HF 5.05 5.27 6.13 5.3–7.2 6.0–7.0 5.1–6.7
T 28.13 30.17 27.72 20.0–30.0 22.0–30.0 21.21–22.94
Tail tip 0.91 1.02 0.98 -- -- 1.22–2.38
3rd mt 27.95 29.08 29.44 26.1–30.1 24.4–26.6 26.12–28.0
4th mt 30.40 30.43 31.00 27.2–32.2 26.9–28.8 27.62–29.61
5th mt 29.25 29.85 29.90 26.2–31.2 26.2–27.8 25.75–27.71
1st ph3rdD 15.16 15.40 15.77 14.3–17.5 14.3–16.2 13.78–15.11
2nd ph3rdD 14.00 13.75 13.50 14.3–17.4 12.5–15.3 14.0–15.47
1st ph4th D 9.37 9.51 9.35 8.7–10.9 8.4–11.2 8.24–8.76
2nd ph4thD 6.16 4.81 7.22 7.0–9.2 6.2–8.6 7.63–8.26
Hw 3.91 3.59 3.32 -- -- --
its base is pointed, and there is a median emargination on the anterior leaf without any supplementary leaflets (Image 2c). The average length of the tip of the tail of H. cineraceus is 1mm, whereas in H. durgadasi it is > 1mm (Image 1c). Other morphological measurements do not provide any significant differences between the relative species (Table 1).
Douangboubpha et al. (2010) provided the significant keys to differentiate Hipposideros halophyllus, H. ater and H. cineraceus and they clearly show that the slight triangular shape of the internarial septum and slight emargination on the anterior leaf without any supplementary leaflets distinguish H. cineraceus from its close relatives H. ater and H. durgadasi (Image 2a–c). In India, this species is known only from the Himalayan states and northeastern states of India. Csorba et al. (2008) claims the distribution of this species in Tamil Nadu, but the range map does not indicate this. Based on Molur et al. (2002) there is no distribution of this species in Tamil Nadu and the report made by Csorba et al. (2008) as occurring in Tamil Nadu is erroneous. The records of H. cineraceus in Madhya Pradesh and Karnataka are referred to as either H. ater or H. durgadasi (Bates & Harrison 1997; Kaur et al. 2014). Hence, our report of the presence of H. cineraceus from Chhattisgarh in central India for the first time based on the National Zoological Collections (ZSI) extends the distribution of this species further south more than 1,400km from its nearest known locality in
Sangser, Kalimpong, West Bengal. A systematic survey in the Odisha part of the Eastern Ghats and adjoining areas (Kanger Ghati National Park) may reveal hitherto unknown yangochiropterans (Debata et al. 2015).
References
Bates, P.J.J. & D.L. Harrison (1997). Bats of the Indian Subcontinent. Harrison Zoological Museum, Sevenoaks, England, UK, 258pp.
Bhat, H.R. & P.G. Jacob (1990). Bio-ecology of Hipposideros cineraceus Blyth, 1853 (Chiroptera, Rhinolophidae) in Kolar District, Karnataka, India. Mammalia 54(2): 182–188.
Csorba, G., S. Bumrungsri, C. Francis, P. Bates, M. Gumal & T. Kingston (2008). Hipposideros cineraceus. The IUCN Red List of Threatened Species 2008: e.T10119A3167688. Downloaded on 15 November 2018; https://doi.org/10.2305/IUCN.UK.2008.RLTS.T10119A3167688.en
Debata, S., H.S. Palei, P.P. Mohapatra & S.K. Palita (2015). Additional records of Cantor’s Leaf-Nosed Bat Hipposideros galeritus Cantor, 1846 (Mammalia: Chiroptera: Hipposideridae) in eastern India: Odisha. Journal of Threatened Taxa 7(8): 7477–7479; https://doi.org/10.11609/JoTT.o4247.7477-9
Douangboubpha, B., S. Bumrungsri, P. Soisook, S.W. Murray, S.J. Puechmaille, C. Satasook, S.S.H. Bu, D.L. Harrison & P.J.J. Bates (2010). A taxonomic review of Hipposideros halophyllus, with additional information on H. ater and H. cineraceus (Chiroptera: Hipposideridae) from Thailand and Myanmar. Acta Chiropterologica 12(1): 29–50.
Hinton, M.A.C. & H.M. Lindsay (1926). Report No.41: Assam and Mishmi Hills. Bombay Natural History Society’s Mammal Survey of India, Burma and Ceylon. Journal of Bombay Natural History Society 31: 383–403.
Kaur, H., C. Srinivasulu, B. Srinivasulu, T. Shah, G. Devender & A. Srinivasulu (2014). Taxonomic notes and distribution extension of Durga Das’s Leaf-nosed Bat Hipposideros durgadasi Khajuria, 1970 Chiroptera: Hipposideridae from south India. Biodiversity Data Journal 2: e4127 (20 Nov 2014); https://doi.org/10.3897/
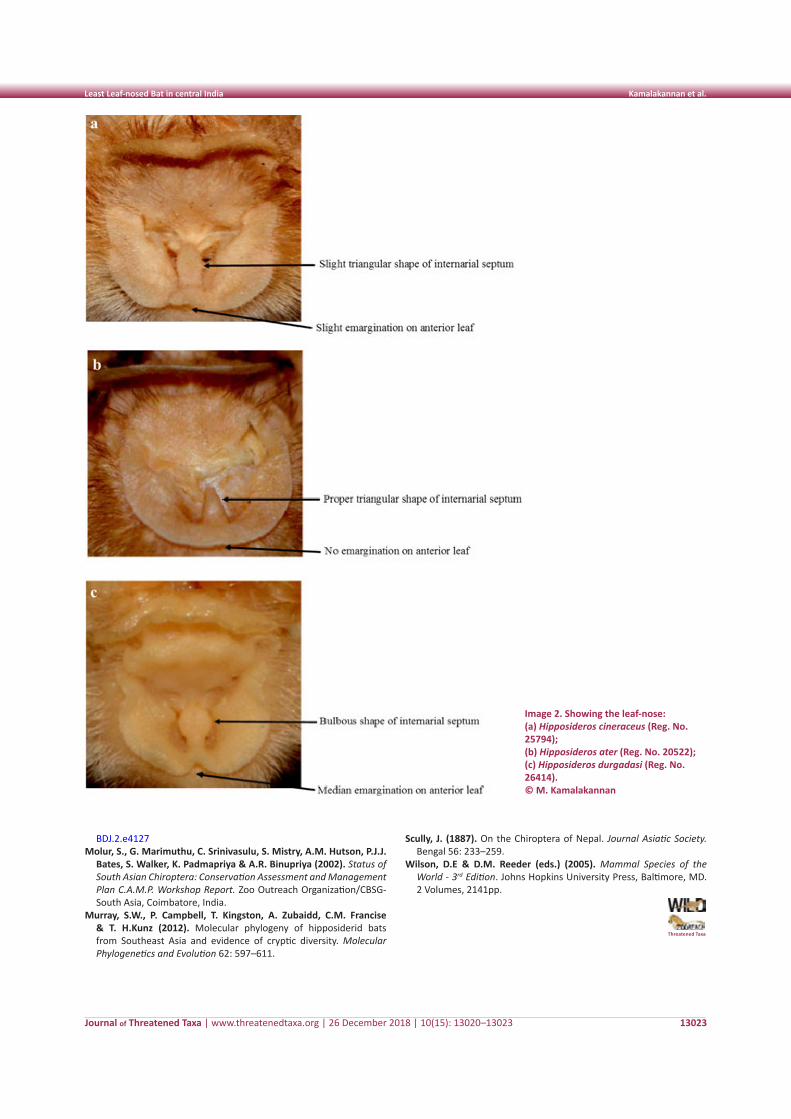
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13020–13023
Least Leaf-nosed Bat in central India Kamalakannan et al.
13023
Image 2. Showing the leaf-nose: (a) Hipposideros cineraceus (Reg. No. 25794); (b) Hipposideros ater (Reg. No. 20522); (c) Hipposideros durgadasi (Reg. No. 26414). © M. Kamalakannan
BDJ.2.e4127 Molur, S., G. Marimuthu, C. Srinivasulu, S. Mistry, A.M. Hutson, P.J.J.
Bates, S. Walker, K. Padmapriya & A.R. Binupriya (2002). Status of South Asian Chiroptera: Conservation Assessment and Management Plan C.A.M.P. Workshop Report. Zoo Outreach Organization/CBSG-South Asia, Coimbatore, India.
Murray, S.W., P. Campbell, T. Kingston, A. Zubaidd, C.M. Francise & T. H.Kunz (2012). Molecular phylogeny of hipposiderid bats from Southeast Asia and evidence of cryptic diversity. Molecular Phylogenetics and Evolution 62: 597–611.
Scully, J. (1887). On the Chiroptera of Nepal. Journal Asiatic Society. Bengal 56: 233–259.
Wilson, D.E & D.M. Reeder (eds.) (2005). Mammal Species of the World - 3rd Edition. Johns Hopkins University Press, Baltimore, MD. 2 Volumes, 2141pp.
Threatened Taxa

13024
Not
e
DOI: https://doi.org/10.11609/jott.3995.10.15.13024-13028 | ZooBank: urn:lsid:zoobank.org:pub:DB10328E-C069-481A-A4C3-71B2AE049517
Editor: Justus Joshua, Green Future Foundation, Trichy, India. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 3995 | Received 03 January 2018 | Final received 10 September 2018 | Finally accepted 20 November 2018
Citation: Thomas, K., D.K. Vinodkumar, J.M. John, M. Shaji & P.O. Nameer (2018). A report on the possible interbreeding between Grizzled Giant Squirrel Ratufa macroura and Indian Giant Squirrel Ratufa indica from Chinnar Wildlife Sanctuary in the southern Western Ghats, India. Journal of Threatened Taxa 10(15): 13024–13028; https://doi.org/10.11609/jott.3995.10.15.13024-13028
Copyright: © Thomas et al. 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: Kerala Agricultural University (KAU).
Competing interests: The authors declare no competing interests.
Acknowledgements: The authors would like to thank the Chief Wildlife Warden, Kerala State Forest Department, for the study permit (WL10-947/2013). We also thank the Kerala Agricultural University (KAU) for the financial support for the conduct of the research. The Dean, College of Forestry, KAU, is also thanked for the encouragement and support.
A report on the possible interbreeding between Grizzled Giant Squirrel
Ratufa macroura and Indian Giant Squirrel Ratufa indica from
Chinnar Wildlife Sanctuary in the southern Western Ghats, India
Kiran Thomas 1 , D.K. Vinodkumar 2 , Jomals Mathews John 3 , M. Shaji 4 & P.O. Nameer 5
1,4,5 Centre for Wildlife Studies, College of Forestry, Kerala Agricultural University, Thrissur 680656, Kerala, India
2 UNESCO C2C for Asia & Pacific Region, Wildlife Institute of India, Chandrabani, Dehradun, Uttarakhand 248001, India
3 GIS Analyst, Lincoln GIS District Office, 105 W Texas Ave, Ruston, Louisiana, 71270 USA
1 [email protected], 2 [email protected], 3 [email protected], 4 [email protected],
5 [email protected] (corresponding author)
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13024–13028
Natural hybridisation events are recorded in many mammalian orders across the world. Schwartz (1980) reported more than 400 hybrids in mammals and many of them were fertile. Natural hybridisation among mammals, however, is less understood than that in other vertebrates. The possible evolutionary consequences
of hybridisation between sympatric species are scarcely studied as such hybridisation was thought to be extremely rare until recently (Ermakov et al. 2002). The role of hybridisation and introgression in determining plant diversity was widely studied but little information is available on the effects of these on animal diversification (Dowling & Secor 1997).
Hybrid animal taxa are rarely reported worldwide. Careful testing of the reported hybrid should be done to understand its population viability. According to Dowling & Secor (1997), hybridisation leads to instantaneous creation of several unique complexes of polyploid and unisexual animals.
Two species of the giant squirrels are seen in the Western Ghats, Ratufa indica and R. macroura. The former is more widespread in distribution (Borges 2015) while the latter is confined only to a few sites (Joshua 1992; Babu & Kalaimani 2014). Though the distribution of these two giant squirrels mostly does not overlap, they do occur in close proximity at a few places. One such place is the Palani Hills in Tamil Nadu where they occur at different elevations (Moore & Tate 1965;
Agarwal & Chakraborty 1979).
Morphology of the two species of giant squirrels in the Western Ghats
Indian Giant Squirrel Ratufa indica: The Indian or the Malabar Giant Squirrel is a squirrel of varying pelages whose back is a mixture of maroon and black with cream or buff underparts. Two subspecies occur in the Kerala part of the Western Ghats, namely R. i. indica seen north of the Palakkad Gap and R. i. maxima (Image 1) that is seen south of the Palakkad Gap. Ratufa indica indica is completely maroon on its back and ears, with
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13024–13028
Possible hybridization between Indian and Grizzled Giant Squirrels Thomas et al.
13025
a pale cream venter and pale face and tail tip. Ratufa indica maxima is similar to R. i. indica except for its black saddle across its shoulders, darker maroon on its dorsal side, and uniformily black and brown tail with no pale tip (Menon 2014; Borges 2015).
Grizzled Giant Squirrel Ratufa macroura: Also known as the Sri Lankan Giant Squirrel, this species is brownish-grey in colour. Its pale hair tips give it a grizzled look. The underside is dirty white and the ears, crown, and dorsal midline are dark brown or black (Image 2). It has three subspecies of which R. m. dandolena, the one that occurs in the Western Ghats, is the smallest of the giant squirrels in India. The ears are short and brown and the tail has long pale hairs making it look greyish compared to the tails of the other subspecies (Menon 2014). The cheeks are buff with small dark areas corresponding to the blackish cheek patches in the other two Indian Ratufa species. It has prominent black to grey or dark brown patches on the forehead and shoulders. The nose is pinkish and the ears are devoid of tufts of hair as in R. indica (Joshua & Johnsingh 2015).
We present here two instances of possible hybridisation between R. indica and R. macroura, observed in 2007 and 2014 from the Anjanad Valley in the Kerala part of the Western Ghats. The Anjanad Valley forms part of the Anamalai Hills of the Western Ghats and is one of the three locations in Kerala where the eastern slopes of the ghats fall within the Kerala State. It has a rain shadow effect, as some of the highest reaches and the broadest stretches of the Western Ghats lie immediately to its west (Nair 1992).
Along the eastern edge of the Anjanad Valley lies the Chinnar Wildlife Sanctuary. It is located 18km north of Marayur in the Idukki District of Kerala State. It is located between 10.25–10.35 0N & 77.08–77.26 0E and has a
total area of 90.44km² (Fig. 1). The Munnar-Udumalpet Road, SH 17, passes through the sanctuary for 16km and bisects it into almost equal portions. It is contiguous with Eravikulam National Park to its south and Indira Gandhi Wildlife Sanctuary to its north. It forms an integral part of the 1,187km2 block of protected forests straddling the Kerala-Tamil Nadu border in the Anamalai Hills.
The terrain is undulating with hills and hillocks of varying heights. The altitude ranges from 400m at Chinnar to 2,372m at Nandamalai. The major peaks in the sanctuary are Varayattumalai (1,845m), Thengamalai (1,422m), Vellakkalmalai (1,883m), Jambumalai (1,395m), Aralipana (1,494m). The area is drained by two perennial rivers passing through the sanctuary, namely Chinnar and Pambar. During the northeastern monsoon that brings the major rains in the Anjanad Valley, a few ephemeral water sources take origin from the higher mountains and drain the area. The entire valley remains dry for the rest of the season.
The vegetation varies from sub-temperate sholas on the mountains to dry scrub in the arid plains. The vegetation of the sanctuary can be broadly classified into the following types according to Champion & Seth (1968): southern tropical thorn forest (scrub jungle), southern dry mixed deciduous forest (dry deciduous forest), southern moist mixed deciduous forest (moist deciduous forest), tropical riparian fringing forest (riparian forest), southern montane wet temperate forest (hill shola forest), and southern montane wet grassland (grassland). The dominant vegetation is dry deciduous forest followed by scrub forest. Together they constitute about 50% of the total forest area, which is located in the low altitude areas. The riparian fringing forests are linearly distributed along the hill folds and occupy a small area. Shola forests occupy a tiny fraction
Image 1. Indian Giant Squirrel Ratufa indica Image 2. Grizzled Giant Squirrel Ratufa macroura
© Kiran Thomas© Dileep Anthikad

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13024–13028
Possible hybridization between Indian and Grizzled Giant Squirrels Thomas et al.
13026
of the total area.First instance: On 13 May 2007, we came across an
interesting interaction between R. indica and R. macroura near the forest office of Marayur Forest Division. In this area, R. i. maxima has been seen beside a dried stream in a dry deciduous forest patch adjacent to the Chinnar WS. Interestingly, Marayur Reserve Forest is a place where R. macroura is only occasionally seen.
Two male R. indica were found combating for a female R. macroura. During the course of the combat, one of the individuals of the R. indica fell to the ground from a height of about 5m. The other R. indica then followed the R. macroura and was observed mounting her after a while. This event happened on a Terminalia bellirica tree at a height of about 10m. The copulation lasted only for a few seconds and when separated, they stared at each other for some time (Image 3). The R. indica male then continued following the R. macroura, presumably for another attempt to mount. The subsequent attempts, however, were unsuccessful as the R. macroura resisted the attempts and chased the R. indica away. Later, both chased each other and were seen running around for about 21/2 hours, almost non-stop (Image 4).
Second instance: During our studies on R. macroura at Chinnar WS in 2013–14, we came across at least three to four different individuals with aberrant coat colour, which appeared to be the hybrids between R. indica and R. macroura (Images 5–7), which is about 17% of the total population of Grizzled Giant Squirrel
Image 3. The Indian Giant Squirrel and the Grizzled Giant Squirrel staring each other after the copulation at Marayur Forest Division in Kerala, southern India
© Jomals Mathews John
Figure 1. Study area - Chinnar Wildlife Sanctuary
© D.K. Vinodkumar
Image 4. The Indian Giant Squirrel chasing the Grizzled Giant Squirrel at Marayur RF

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13024–13028
Possible hybridization between Indian and Grizzled Giant Squirrels Thomas et al.
13027
Table 1. The details on the encounter of the suspected hybrids of Ratufa indica and R. macroura at Chinnar Wildlife Sanctuary in the southern Western Ghats
Date of observation Location
1 10.viii.2013 Chinnar checkpost
2 9.xi.2013 Churulipetti
3 23.i.2014 Along the stretch of Chinnar to Churulipetti streams
4 14.iii.2014 Koottar
5 1.iv.2014 Churulipetti
6 13.v.2007 Near the District Forest Office of Marayur Forest Department
Image 6–7. 6 - Hybrid individuals between GGS and IGS showing varying pelage colour; 7 - Hybrid individual between GGS and IGS
(Thomas & Nameer 2018). There were five instances between August 2013 and April 2014 (Table 1) when we encountered the suspected hybrid individuals at the
sanctuary, the locations of which were mapped (Fig. 2). The general structure of the pelage colour of these hybrids was a mixture of both R. indica and R. macroura. We could, however, observe three different pelage patterns among the hybrid individuals. In one case, the pelage looked similar to that of R. indica, excepting the creamy white underparts and the cheeks, which were brown to chocolate brown in colour and the tail has chestnut tinge (Image 5). The second type of pelage looked similar to that of R. macroura, excepting that the grizzled upper part of the individual appeared brown to black in color (Image 6). The third type of pelage encountered was similar to that of the darker form of R. indica but the tail, instead of being completely black, had a pale tip (Image 7), a character similar to that of R. i. indica.
There was a previous instance of interbreeding between R. indica and R. macroura where seven hybrid
© Kiran Thomas
Figure 2. The locations where the hybrid individuals were sighted in Chinnar WS, southern India
6
© Kiran Thomas7
Image 5. A hybrid between Indian and Grizzled Giant Squirrel in the Churulipetti area of Chinnar WS, Kerala, southern India
© Kiran Thomas

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13024–13028
Possible hybridization between Indian and Grizzled Giant Squirrels Thomas et al.
13028
individuals were reported at Srivilliputhur Grizzled Giant Squirrel Sanctuary in Tamil Nadu, southern India (Joshua 1992). Joshua (1996) opined that the R. macroura was pushed towards the foothills of the Ayyanarkoil Valley in Grizzled Giant Squirrel Sanctuary, where R. indica exists. He further noted that the habitat shift in R. macroura was due to habitat degradation.
Ratufa indica was historically unknown in the Chinnar WS. The nearest known distribution of R. indica is in the Marayur RF, located towards the southwestern part of the Chinnar WS. Similarly, R. macroura was also not known from any habitat outside the riverine forests of Chinnar and Pambar rivers and their tributaries in Kerala. Over the last one decade or so, however, there were occasional sightings of R. macroura from the Marayur RF. These sightings became more frequent and regular since then. These range expansions of R. macroura to hitherto unknown sites could be due to the disturbances in its riverine habitats at Chinnar WS (Thomas & Nameer 2018). The expansion of the range of R. macroura into that of R. indica enable them to interact closely, thus leading to interbreeding.
It would be interesting to know whether these hybrids are fertile or not. Detailed investigation on the status of the hybrid individuals, exact reasons for hybridisation, and a study on the genetics of the R. macroura need to be undertaken at the Chinnar WS to find out the genetic purity of this species. The mixup and hybridisation between R. macroura and R. indica should be monitored to find out its effect on the long-term survival of the R. macroura, which has a restricted distribution and a Near Threatened species (Joshua et al. 2008). The interbreeding could be a challenge to the long-term conservation of R. macroura at Chinnar WS. At this juncture, it is noteworthy to recall the local extinction of the native Red Squirrel Sciurus vulgaris by the introduced Grey Squirrel Sciurus carolinensis in southern and northern England (Lloyd 1983; Reynolds 1985).
References
Agarwal, V.C. & S. Chakraborty (1979). Taxonomic notes on some Oriental squirrels. Mammalia 43(2): 161–172.
Babu, S. & A. Kalaimani (2014). New site record of Grizzled Giant Squirrel Ratufa macroura from Thiruvannamalai Forest Division, Eastern Ghats, Tamil Nadu, India. Journal of Threatened Taxa 6(2): 5492–5493; https://doi.org/10.11609/JoTT.o3680.5492-3
Borges, R.M. (2015). Indian Giant Squirrel (Ratufa indica), pp483–500. In: Johnsingh, A.J.T & N. Manjrekar (eds.). Mammals of South Asia, Vol. II. The University Press (India) Private Ltd., Hyderabad, India, 720pp.
Champion, H.G. & S.K. Seth (1968). A Revised Survey of the Forest Types of India. Government of India Press, Delhi, 404pp.
Dowling, T.E. & C.L. Secor (1997). The role of hybridization and introgression in the diversification of animals. Annual Review of Ecology and Systematics 28: 593–619.
Ermakov, O.A., V.L. Surin, S.V. Titov, A.F. Tagiev, A.V. Luk’yanenko & N.A. Formozov (2002). A molecular genetic study of hybridization in four species of ground squirrels (Spermophilus: Rodentia, Sciuridae). Russian Journal of Genetics 38(7): 796–809.
Joshua, J. (1992). Ecology of the Endangered Grizzled Giant Squirrel (Ratufa macroura) in Tamil Nadu, south India. Ph.D Thesis. Bharathidasan University, Tiruchirapalli, Tamil Nadu, 191pp.
Joshua, J. (1996). Interbreeding between Grizzled Giant Squirrel (Ratufa macroura Pennant) and Malabar Giant Squirrel (R. indica Erxleben). Journal of the Bombay Natural History Society 93(1): 82–83.
Joshua, J., de A. Goonatilake, W.L.D.P.T.S. & S. Molur (2008). Ratufa macroura. In: The IUCN Red List of Threatened Species. Downloaded on 23 December 2017.
Justus, J. & A.J.T. Johnsingh (2015). Grizzled Giant Squirrel, pp501–512. In: Johnsingh, A.J.T. & N. Manjrekar (eds.). Mammals of South Asia, Vol 2. University Press, Hyderabad, India, 720p.
Lloyd, H.G. (1983). Past and present distribution of Red and Grey squirrels. Mammal Review 13(2–4): 69–80.
Menon, V. (2014). Indian Mammals: A Field Guide. Hachette Book Publishing India Pvt. Ltd., Guragon, India, 528pp.
Moore, J.C. & G.H.H. Tate (1965). A study of the diurnal squirrels, Sciurinae, of the Indian and Indo-Chinese subregions. Fieldiana Zoology 48: 1–351.
Nair, S.C. (1992). The sSsouthern Western Ghats: A Biodiversity Conservation Plan. Indian National Trust for Art and Cultural Heritage, Indraprastha Press, New Delhi, 91pp.
Reynolds, J.C. (1985). Details of geographical replacement of the Red Squirrel (Sciurus vulgaris) by the Grey Squirrel (Sciurus carolinensis) in Eastern England. Journal of Animal Ecology 54: 149–162.
Schwartz, S.S. (1980). Ekologicheskie zakonomernosti evolyutsii (Ecological Regularities at Evolution). Nauka Publishers, Moscow.
Thomas, K & P.O. Nameer (2018). Alarming population status of the Grizzled Giant Squirrel (Ratufa macroura) (Mammalia: Rodenta: Sciuridae) in Chinnar Wildlife Sanctuary, the Western Ghats, India. Journal of Threatened Taxa 10(10): 12350–12356; https://doi.org/10.11609/jott.3536.10.10.12350-12356
Threatened Taxa

13029
Not
e
DOI: https://doi.org/10.11609/jott.4664.10.15.13029-13031 | ZooBank: urn:lsid:zoobank.org:pub:FA557694-487F-4769-98FB-E491175A0D2E
Editor: K.A. Subramanian, Zoological Survey of India, Chennai, India. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 4664 | Received 20 September 2018 | Final received 30 October 2018 | Finally accepted 23 November 2018
Citation: Cowan, E.M. & P.J. Cowan (2018). Ischnura fountaineae (Insecta: Odonata: Zygoptera) in Oman, eastern Arabia. Journal of Threatened Taxa 10(15): 13029–13031; https://doi.org/10.11609/jott.4664.10.15.13029-13031
Copyright: © Cowan & Cowan 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduc-tion and distribution by providing adequate credit to the authors and the source of publication.
Funding: None.
Competing interests: The authors declare no competing interests.
Acknowledgements: We thank the moderator of the All Odonata website gallery. David Chelmick gave us the URL for the Fons Peels dragonfly website.
Ischnura fountaineae (Insecta: Odonata: Zygoptera) in Oman, eastern Arabia
Elaine Mary Cowan 1 & Peter John Cowan 2
1 School of Education, University of Aberdeen, AB24 3FX, Scotland, UK2Department of Biological Sciences and Chemistry, University of Nizwa, Birkat Al Mawz, Sultanate of Oman1 [email protected], 2 [email protected] (corresponding author)
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13029–13031
We considered Ischnura fountaineae Morton, 1905 to occur, in Oman, only in the western Hajar mountains of northern Oman (fairly close to the United Arab Emirates) based on records in the Mahadah area by the late Bob Reimer (Cowan & Cowan 2017; Reimer et al. 2009). We had not knowingly recorded the species at that time, however, the author Elaine M. Cowan (EMC) remembered that she had photographed a very blue eyed individual with pale blue thorax, without a hint of green, at ‘Hoota’ Wadi (Arabic: valley) pool (Cowan & Cowan 2013, 2015). This pool (23.0710N & 57.3680E, elevation 680m) is near the Al Hoota cave tourist complex in the southern foothills of the Jebel Akhdar range (the central and highest part of the Hajar mountains). In February 2018, EMC submitted the photograph (Image 1) to the expert-moderated All Odonata website gallery (www.allodonata.com) for an ID opinion. The moderator replied that it was indeed I. fountaineae and the photo is on display there as such.
Confident identification of Ischnura fountaineae from photographs had seemed a challenge. Grunwell (2010) felt the best distinguishing feature for males from a male I. evansi (the commonest Ischnura in Oman,
Cowan & Cowan 2017) was the downwards sloping to the front of the lower edge of the black on the upperside of segment 2 of the abdomen. Dijkstra & Lewington (2006) considered the diagnostic field characters of mature male I. fountaineae to be the sky blue ground colour of the thorax and abdomen base, without a hint of green, and their shining black markings. The postocular spots are small. In the hand, males are reliably identified by the anal appendages (Dijkstra & Lewington 2006; contra Reimer et al. 2009). The best gallery of online photos of I. fountaineae is perhaps that on the Fons Peels website (www.dragonflypix.com). Immature males do not have any trace of green but are whitish with black markings (Galliani et al. 2017). Reimer et al. (2009) stated that I. evansi and I. fountaineae can be most easily distinguished in the field by the colouration of the thorax and first few segments of the abdomen and the antehumeral stripes on the thorax. In I. evansi the colour is greenish-blue and there are always clearly visible stripes of the same colour on the shoulders. Ischnura fountaineae is sky-blue without a hint of green, the same colour as the blue that occurs on the eighth abdominal segment (i.e., the ‘tail-light’). The antehumeral stripe may be missing, interrupted or very narrow. Our understanding of the diagnostic field characters (i.e., visible on a digital photograph) is that male I. fountaineae have a pale blue thorax, the same colour as the tail-light, and blue eyes (the anterior of the eyes can sometimes appear greenish). The positive identification of females may be difficult under field

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13029–13031
Ischnura fountaineae in Oman Cowan & Cowan
13030
conditions (Dijkstra & Lewington 2006).Boudot et al. (2015) presented a map of the world
distribution of I. fountaineae. According to them, the species occurs across northern Africa from eastern Morocco to Egypt, east to Kazakhstan and westernmost China. They show a southern outlier, in eastern Saudi Arabia, Qatar, United Arab Emirates and northern Oman south to about 22.50N. Reimer et al. (2009) presented the occurrence of I. fountaineae in the United Arab Emirates, the Mahadah area of the western Hajar, Oman and interestingly mentioned that photographs by Gary
Image 1. Ischnura fountaineae, 19 January 2016, at the Hoota Wadi pool in the southern foothills of the Jebel Akhdar range, northern Oman. Note the pale blue thorax and blue eyes.
Image 2. Ischnura fountaineae, 11 April 2014, Wadi Qtm, Saiq plateau, Jebel Akhdar mountain range, northern Oman. Small blue postocular spots.
Image 3. Ischnura fountaineae, 25 August 2014, Wadi Qtm, Saiq plateau, Jebel Akhdar mountain range, northern Oman. Small blue postocular spots. Image 4. Ischnura fountaineae, 25 August 2014, Wadi Qtm, Saiq
plateau, Jebel Akhdar mountain range, northern Oman, showing green anteriorly on eyes and pale blue thorax.
Feulner of Ischnura males and females along irrigation channels among cultivation on the Saiq plateau at c. 2000m elevation in the Jebel Akhdar range of Oman appeared to show I. fountaineae. The species has not been reported by other observers in Oman (Cowan & Cowan 2017, 2018; Lambret et al. 2017).
Reviewing our archive of photographs from Oman, we now have eight records of I. fountaineae at Hoota Wadi pool. These are: 11 September 2013, 11, 19 January, 19 August, 9, 16 September 2015 and 19 (Image
© E.M. Cowan
© E.M. Cowan
© E.M. Cowan
© E.M. Cowan

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13029–13031
Ischnura fountaineae in Oman Cowan & Cowan
13031
Image 5. Ischnura evansi, 3 October 2016 at the Hoota Wadi pool in the southern foothills of the Jebel Akhdar mountain range, Oman. Note the green eyes and green thorax.
Image 6. Ischnura senegalensis, Khor Taqa, Dhofar governorate, southern Oman, 17 October 2013 (Cowan & Cowan 2018). Note the green eyes and green thorax, black ‘saddle’ on S2 of abdomen separating two patches of surrounding green colour, and bicoloured pterostigma.
1), 25 January 2016. We also have three records from Wadi Qtm (23.0720N & 57.6270E, 1,970m), which is on the Saiq plateau of Jebel Akhdar and has an irrigation channel (falaj) and pools. These three records are: 11 April (Image 2), 25 August (Images 3, 4) 2014 and 18 April 2016. The records of 11 April 2014 and 18 April 2016, at Wadi Qtm, were erroneously reported as I. evansi in Cowan & Cowan (2017). The apparent status of I. fountaineae in Oman should be “uncommon Western Hajar and Jebel Akhdar regions” (see Cowan & Cowan 2017 for further information about these regions and sites).
Ischnura evansi (Image 5) which is clearly differentiated by green eyes and greenish thorax was photographed at the Hoota Wadi pool on most visits there (including the 8 dates when I. fountaineae was recorded). Ischnura senegalensis is found elsewhere in Oman but is mainly coastal. Ischnura senegalensis has a deep black ‘saddle’ on S2 of the abdomen separating two patches of surrounding colour (Image 6).
References
Boudot, J-P., E. Riservato & S. Hardersen (2015). Ischnura fountaineae Morton, 1905, pp. 127–128. In: Boudot, J-P. & V.J. Kalkman (eds.). Atlas of the European Dragonflies and Damselflies. Netherlands: KNNV Publishing, 381pp.
Cowan, E.M. & P.J. Cowan (2013). The dragonflies and damselflies of a wadi pool near Nizwa, northern Oman, 2012–2013. Tribulus 21: 14–23.
Cowan, E.M. & P.J. Cowan (2015). Odonata (Insecta) at a wadi pool near Nizwa, northern Oman. Journal of Threatened Taxa 7(9): 7538–7546; https://doi.org/10.11609/JoTT.o4207.7538-46
Cowan, E.M. & P.J. Cowan (2017). The Odonata (Insecta) of northern and central Oman. Journal of Threatened Taxa 9(10): 10776–10791; https://doi.org/10.11609/jott.3230.9.10.10776-10791
Cowan, E.M. & P.J. Cowan (2018). The Odonata (Insecta) of Dhofar, southern Oman. Journal of Threatened Taxa 10(11): 12499–12514; https://doi.org/10.11609/jott.4161.10.11.12499-12514
Dijkstra, K-D.B. & R. Lewington (2006). Field Guide to the Dragonflies of Britain and Europe. British Wildlife Publishing, UK, 320pp.
Galliani, C., R. Scherini & A. Piglia (2017). Dragonflies and Damselflies of Europe. WBA Handbooks, Verona, Italy, 349pp.
Grunwell, M.J. (2010). Dragonflies and damselflies in the state of Qatar. Journal of Qatar Natural History Group 3: 2–15.
Reimer, R.W., G.R. Feulner & R.J. Hornby (2009). Errata and addenda: updated illustrated checklist of dragonflies of the UAE - including a third species of Ischnura damselfly. Tribulus 18: 28–36.
Threatened Taxa
© E.M. Cowan
© E.M. Cowan

13032
Not
e
DOI: https://doi.org/10.11609/jott.4036.10.15.13032-13036 | ZooBank: urn:lsid:zoobank.org:pub:0F5E646D-99AD-49FC-AFB9-060480B6E560
Editor: Brian Fisher, California Academy of Sciences, San Francisco, USA. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 4036 | Received 29 January 2018 | Final received 11 October 2018 | Finally accepted 30 November 2018
Citation: Rasheed, M.T., I. Bodlah, A.G. Fareen & X. Huang (2018). First record of Leptogenys hysterica Forel, 1900 (Hymenoptera: Formicidae: Ponerinae) from Pakistan. Journal of Threatened Taxa 10(15): 13032–13036; https://doi.org/10.11609/jott.4036.10.15.13032-13036
Copyright: © Rasheed et al. 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: Pakistan Science Foundation Islamabad (PSF/NSLP/P-UAAR 313); Pir Mehr Ali Shah Arid Agriculture University, Rawalpindi.
Competing interests: The authors declare no competing interests.
Acknowledgements: We are greatly thankful to Pakistan Science Foundation for the financial support provided under the project PSF/NSLP/P-UAAR (313) to perform these studies. Anonymous reviewers are also greatly acknowledged for their helpful comments and suggestions about the manuscript.
First record of Leptogenys hysterica Forel, 1900 (Hymenoptera: Formicidae:
Ponerinae) from Pakistan
Muhammad Tariq Rasheed 1 , Imran Bodlah 2 , Ammara Gull e Fareen 3 & Xiaolei Huang 4
1,2,3 Insect Biodiversity and Conservation Group, Department of Entomology, Pir Mehr Ali Shah Arid Agriculture University,
Shamsabad, Muree Road, Rawalpindi, Punjab 46000, Pakistan 4 State Key Laboratory of Ecological Pest Control for Fujian and Taiwan
Crops, College of Plant Protection, Fujian Agriculture and Forestry University, Fuzhou 350002, China
1 [email protected], 2 [email protected] (corresponding author), 3 [email protected], 4 [email protected]
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13032–13036
The ant genus Leptogenys Roger, 1861 belongs to the subfamily Ponerinae and is considered to be one of the most speciose genera throughout tropical and subtropical regions with 308 species, 25 subspecies, and one fossil record (AntWeb 2018). These ants commonly reproduce by ergatogynes or gamergates (Ito
1997; Ito & Ohkawara 2000; Peeters 2012; Bharti & Wachkoo 2013). Individuals of this genus prefer rotten or dead wood, leaf litter, and surfaces under stones as habitats; a few species are also considered subarboreal (Bolton 1975; Rakotonirina & Fisher 2014). These ants mainly predate on termites and terrestrial isopods (Bolton 1975; Lattke 2011).
Significant contributions to the knowledge of this genus from southeastern Asia include Wu & Wang (1995), Xu (2000), Zhou (2001), Terayama (2009), and Arimoto (2017). Bharti & Wachkoo (2013) provided keys to the Leptogenys in India with descriptions of two new species. Xu & He (2015) reviewed the Oriental species of this genus, provided an identification key to the ant fauna in China, and described and added two new records to the country. Other noteworthy studies include that of Bolton (1975) and Lattke (2011), which remarkably contributed to the species of Leptogenys in the Afrotropical region and the New World, respectively.
Very limited work on the exploration of ants in Pakistan was undertaken till date (Umair et al. 2012; Bodlah et al. 2016; Bodlah et al. 2017a,b). Twenty-seven species of Leptogenys were recorded from India, one of
its neighbouring countries (Bharti & Wachkoo 2013). Here we report Leptogenys hysterica for the first time from the country with diagnostic note and illustrations.
Materials and MethodsAs a result of extensive surveys during 2016–2017,
workers of the genus Leptogenys were collected from different forest areas of Margalla Hills in Rawalpindi, Islamabad, Pakistan. Specimens of this species were found under rocks near leaf litter and were hand collected. The collected specimens were placed in potassium cyanide killing jars. A few specimens were also preserved in 75% ethanol. Taxonomic analysis of the collected specimens was performed under Labomed microscope using keys by Bharti & Wachkoo (2013). Identifications were made by observing the metanotal groove, the width and length of the petiolar node, and comparison of the mesosomal length with the
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13032–13036
First record of Leptogenys hysterica from Pakistan Rasheed et al.
13033
abdominal length. Images were prepared by a digital camera (Nikon DS-
Fi3) attached with a Nikon 1500 SMZ stereo microscope and cleaned using Adobe Photoshop CS6 software. Measurements (in millimeters) and indices were calculated with the help of a stage and ocular micrometer. Identified specimens were tagged with the valid names, localities, dates of collection, and microhabitats. After taxonomic treatment, the specimens were deposited in the Department of Entomology, Pir Mehr Ali Shah Arid Agriculture University, Rawalpindi, Pakistan.
Morphometric terminology (in millimeters) and indices are given as: HL - In full face view, the maximum length of head from clypeus to vertex in a straight line; HW - Maximum width of the head in frontal view; SL - Total length of scape without the neck; EL - Maximum eye length; EW - Maximum width of eye in oblique view; PW - Maximum pronotal width in dorsal view; WL - Maximum length of mesosoma from anterior to posterior margin of pronotum except for collar in lateral view; PL - Maximum length of petiole from anterior to the posterior margin of tergite; PH - Maximum height of petiole from apex to the dorsal point in lateral view; PDW - Maximum width of node in dorsal view; GL - Maximum abdominal length; CI - Cephalic index = HW/HL×100; OI - Ocular index = EL/HW×100; SI - Scape index = SL/HW×100; LPI - Lateral petiole index = PH/PL×100; DPI - Dorsal petiole index = PDW/PL×100
Results and DiscussionLeptogenys hysterica Forel, 1900
(Images 1–9)Leptogenys (Lobopelta) hysterica Forel, 1900: 311
(w.) SRI LANKA. Imai et al., 1984 (k.) 5. Status as species: Bingham, 1903: 64; Bharti & Wachkoo, 2013: 17 (in key); Xu & He, 2015: 156 (in key).
Worker morphometrics (in millimetres): HL 0.98–1.0, HW 0.62–0.65, EL 0.16–0.18, EW 0.13–0.15, PW 0.52–0.55, SL 0.98–1.0, WL 0.53–0.6, PL 0.4–0.6, PW 0.41–0.43, PH 0.52–0.55, GL 1.66–1.68, TL 5.25–5.47. Indices: CI 63.41–65, SI 153.84–157.05, OI 24.61–28.84, LPI 91.66–132, DPI 68.33–108 (n=5).
Worker description: Head densely punctate, rectangular in full face view, likewise narrowed anteriorly and posteriorly, posterior and lateral margin convex, carinate medially, exceeding nearly the level of eyes; eyes convex, placed laterally just below the cephalic mid length; frontal grove narrow hardly touching the level of eyes; clypeus triangular, medial clypeal lobe rounded, apex bluntly rounded with minute setae, strongly carinate at middle, thin and translucent, anterior and
lateral margin longitudinally carinate; mandible long, narrow at anterior, thicker at apex, carinate longitudinally in lateral view, basal tooth absent, apical tooth present, masticatory margin without teeth, smooth; scape exceeding the lateral cephalic margin.
Mesosoma in dorsal view densely punctate (Image 5), deeply impressed at metanotal grove; mesothorax shorter than pro- and metathorax; metathorax rugose laterally (Image 6); gaster one and half times longer than mesosomal length (Image 8), pronotum width more than rest of mesosomal width, metanotum length more than rest of mesosomal length; propodeum declivity transversely striated (Image 9).
Posterior petiolar margin slightly wider than anterior petiolar margin, longer in length than width, forming smooth convexity.
Gaster’s length more than weber’s length (Image 8); basal tergite shining and densely punctate; base of second tergite cross-ribbed, remaining tergite after basal tergite smooth and shiny.
Mandible smooth dorsally, narrow, sparsely punctate laterally; clypeus triangular with strong medial carinae, longitudinally striated; scape covered with piligerous and become dense at apex; pro pleuron rugose longitudinally and meso-meta pleuron transversally rugose.
Distribution: Indo-Malayan region in Borneo, India, Indonesia, Laos, Malaysia, Sri Lanka, and Thailand (Bharti et al. 2017; AntWeb 2018).
Material examined Pakistan: FOR-001, 10 workers, 05.iii.2016, Pakistan,
Islamabad, Trail 5, 33.818°N, 73.123°E, 630m, coll. M.T. Rasheed; 7 workers, 08.iv.2016, Pakistan, Islamabad, Pir Sohawa, 35.872°N, 73.185°E, 1,046m, coll. I. Bodlah; 10 workers, 25.v.2016, Pakistan, Rawalpindi, Murree, 33.293°N, 73.368°E, 1,158m, coll. A.G. Fareen; 10 workers, 08.vi.2017, Pakistan, Islamabad, Daman e koh, 34.243°N, 73.185°E, 722m, coll. M.T. Rasheed.
Differential diagnosis: Leptogenys hysterica most resembles L. punctiventris (Mayr, 1879) from which it can be distinguished by its distinct metanotal groove (Image 4) and broader than long petiole in dorsal view (Image 7). The DPI in L. hysterica is 68.00–108.00 mm while in L. punctiventris the metanotal groove is obsolete with longer than wide petiolar node and DPI < 90.00mm.
Ecology: Members of the genus Leptogenys prefer to make their nests in the soil or under stone surfaces, logs of trees, tree bark, dead wood, and leaf litter in mountainous areas having humid forests (Bolton 1975; Lattke 2011; Bharti & Wachko 2013). We found L. hysterica nesting in loose soil on a stone embankment

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13032–13036
First record of Leptogenys hysterica from Pakistan Rasheed et al.
13034
near leaf litter and dried vegetation in the forest areas of Margalla Hills in Rawalpindi.
Remarks The discovery of this species from the mountainous
forests of Pakistan extends its known distribution reported by Bharti & Wachkoo (2013) from neighbouring country, India. The current study indicates that this species is also distributed in the foothills of the Himalayan region. Extensive deforestation, development of housing societies, and an increase in public activities in national park trails in Margalla Hills in Islamabad, Murree and Rawalpindi are causing forest fragmentation and degradation, which may threaten the existence of L. hysterica in the country in the future. National policies are needed for the conservation of this species and its
natural habitats. Most of the forest areas of Pakistan are still unexplored for ants and efforts need to be made for the further exploration of the species in the country.
References
AntWeb (2018). Bolton World Catalog. https://www.antweb.org/project.do?name=worldants. (Accessed date: 10 Feburary, 2018).
Arimoto, K. (2017). Taxonomy of the Leptogenys modiglianii species group from southeast Asia (Hymenoptera, Formicidae, Ponerinae). ZooKeys 651: 79–106; https://doi.org/10.3897/zookeys.651.10336
Bharti, H. & A.A. Wachkoo (2013). Two new species of the ant genus Leptogenys (Hymenoptera: Formicidae) from India, with description of a plesiomorphic ergatogyne. Asian Myrmecology 5: 11–19.
Bharti, H., A.A. Wachkoo & R. Kumar (2017). First inventory of ants (Hymenoptera: Formicidae) in northwestern Shivalik, India. Halteres 8: 33–68; https/doi.org/10.5281/zenodo.582706
Bingham, C.T. (1903). The Fauna of British India, including Ceylon and
Images 1–3. Leptogenys hysterica Forel, 1900, worker. 1 - head, full-face view; 2 - body, dorsal view; 3 - body, lateral view
1 2
3
© Muhammad Tariq Rasheed © Muhammad Tariq Rasheed
© Muhammad Tariq Rasheed

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13032–13036
First record of Leptogenys hysterica from Pakistan Rasheed et al.
13035
Burma. Hymenoptera: Ants and Cuckoo Wasps, Vol. II. Taylor and Francis, London, 506pp.
Bodlah, I., M.T. Rasheed, A.G. Fareen, M.S. Ajmal & M.A. Bodlah (2016). First record of two new species of genus Tetraponera (Hymenoptera: Pseudomyrmecinae: Formicidae) from Pakistan. Journal of Entomology and Zoology Studies 4(4): 1028–1030.
Bodlah, I., M.T. Rasheed & M.A. Bodlah (2017a). New distributional
records of Tetraponera rufonigra (Jerdon) from Gilgit Baltistan, Pakistan. Asian Journal of Agriculture & Biology 5(2): 56–59.
Bodlah, I., M.A. Bodlah, M.T. Rasheed, T. Akhter, A. Aihetasham & M. Yousaf (2017b). New distributional records of psyllid, Trioza fletcheri minor Crawford, 1912 and record of its first association with two ant’s species in Pothwar. Asian Journal of Agriculture & Biology 5(1): 1–6.
Images 8–9. Leptogenys hysterica Forel, 1900, worker. 8 - gaster length is one half than mesosomal length; 9 - propodeum declivity transversely striated
8 9
Images 4–7. Leptogenys hysterica Forel, 1900, worker. 4 - distinct metanotal groove; 5 - mesosoma in dorsal view densely punctate; 6 - metathorax rugose laterally; 7 - petiolar node broader than long dorsally
6 7
4 5
© Imran Bodlah
© Imran Bodlah
© Imran Bodlah
© Imran Bodlah
© Imran Bodlah
© Imran Bodlah

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13032–13036
First record of Leptogenys hysterica from Pakistan Rasheed et al.
13036
Bolton, B. (1975). A revision of the ant genus Leptogenys Roger (Hymenoptera: Formicidae) in the Ethiopian region with a review of the Malagasy species. Bulletin of the British Museum (Natural History) Entomology 31: 235–305.
Forel, A. (1900). Les Formicides de l’Empire des Indes et de Ceylan, part VII. The Journal of the Bombay Natural History Society 13: 303–332.
Imai, H.T., U.C. Baroni, M. Kubota, G.P. Sharma, M.H. Narasimhanna, B.C. Das, A.K. Sharma, A. Sharma, G.B. Deodikar, V.G. Vaidya & M.R. Rajasekarashetty (1984). Karyological survey of Indian ants. The Japanese Journal of Genetics 59(1): 1–32; https://doi.org/10.1266/jjg.59.1
Ito, F. (1997). Colony composition and morphological caste differentiation between ergatoid queens and workers in the ponerine ant genus Leptogenys in the Oriental tropics. Ethology, Ecology and Evolution 9(4): 335–343.
Ito, F. & K. Ohkawara (2000). Production and behavior of ergatoid queens in two species of the Indonesian ponerine ant genus Leptogenys (diminuta-group) (Hymenoptera: Formicidae). Annals of the Entomological Society of America 93(4): 869–873.
Lattke, J.E. (2011). Revision of the New World species of the genus Leptogenys Roger (Insecta: Hymenoptera: Formicidae: Ponerinae). Arthropod Systematics and Phylogeny 69(3): 127–264.
Peeters, C. (2012). Convergent evolution of wingless reproductives across all subfamilies of ants and sporadic loss of winged queens (Hymenoptera: Formicidae). Myrmecological News 16: 75–91.
Rakotonirina, J.C. & B.L. Fisher (2014). Revision of the Malagasy ponerine ants of the genus Leptogenys Roger (Hymenoptera: Formicidae). Zootaxa 3836(1): 1–163; http://doi.org/10.11646/zootaxa.3836.1.1
Terayama, M. (2009). A synopsis of the Family Formicidae of Taiwan (Insecta, Hymenoptera). Liberal Arts, Research Bulletin of Kanto Gakuen University 17: 81–266.
Umair, M., A. Zia, M. Naeem & M.T. Chaudhry (2012). Species composition of ants (Hymenoptera: Formicidae) in Pothwar plateau of Punjab Province, Pakistan. Pakistan Journal of Zoology (44): 699–705.
Wu, J. & C. Wang (1995). The Ants of China. China Forestry Publishing House, Beijing, 214pp.
Xu, Z. (2000). Five new species and one new record species of the ant genus Leptogenys Roger (Hymenoptera: Formicidae) from Yunnan Province, China. Entomologia Sinica 7: 117–126.
Xu, Z.H. & Q.J. He (2015). Taxonomic review of the Ponerinae ant genus Leptogenys Roger, 1861 (Hymenoptera: Formicidae) with a key to the Oriental species. Myrmecological News 21: 137–161.
Zhou, S.Y. (2001). Ants of Guangxi. Guangxi Normal University Press, Guilin, China, 255pp.
Threatened Taxa

13037
Not
e
DOI: https://doi.org/10.11609/jott.4302.10.15.13037-13038 | ZooBank: urn:lsid:zoobank.org:pub:D07A8377-FAB0-4E5A-B43A-43B1BEF3C040
Editor: Sabu K. Thomas, St. Joseph’s College, Kozhikode, India. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 4302 | Received 29 May 2018 | Final received 01 November 2018 | Finally accepted 24 November 2018
Citation: Hegde, V.D., D. Vasanthakumar & S.V. Manthen (2018). First report of darkling beetle Blaps orientalis Solier, 1848 (Coleoptera: Tenebrionidae) From India. Journal of Threatened Taxa 10(15): 13037–13038; https://doi.org/10.11609/jott.4302.10.15.13037-13038
Copyright: © Hegde et al. 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: None.
Competing interests: The authors declare no competing interests.
Acknowledgements: We are thankful to Dr. Kailash Chandra, director, Zoological Survey of India (ZSI), Kolkata, for the facilities provided. We are indebted to all the staff of the Western Regional Centre, ZSI, Pune, for their constant encouragement. Special thanks are due to Dr. Sameer Kumar Pati, ZSI, Pune, for the images.
First report of darkling beetle Blaps orientalis Solier, 1848 (Coleoptera: Tenebrionidae) from India
V.D. Hegde 1 , D. Vasanthakumar 2 & S.V. Manthen 3
1,2,3 Western Regional Centre, Zoological Survey of India, Vidyanagar, Ravet Road, Akurdi, Pune, Maharashtra 411044, India1 [email protected] (corresponding author), 2 [email protected], 3 [email protected]
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13037–13038
The darkling beetles belonging to the tribe Blaptini Leach, 1815 are large, oblong-elongate beetles, usually measuring more than 20mm in length (Aalbu et al. 2002). This tribe consists of 500 species under 28 genera worldwide (Medvedev 2001, 2007; Medvedev & Merkl 2002), which includes 23 species of six genera from India (unpublished data). The genus Blaps Fabricius, 1775 is one of the most diverse genera of darkling beetles in this tribe (Soldati et al. 2017). The species belonging to this genus are generally flightless and well adapted to semi-arid and arid conditions owing to several specific behavioural and morphological adaptations (Condamine et al. 2011). This genus includes more than 250 species worldwide (Löbl et al. 2008), 15 of which were reported from India. The details and distribution of all Blaps species earlier reported from northern and northeastern India are given in Table 1. The first report of B. orientalis Solier, 1848 from India after its earlier report from Bela in Balochistan (earlier Belutschistan), Pakistan (Schuster 1930), is provided here. Interestingly, this collection from Pune in Maharashtra shows the distribution of
Blaps species from peninsular India for the first time.
Systematic Account (As per Bouchard et al. 2005)Family: Tenebrionidae Latreille, 1802Subfamily: Tenebrioninae Latreille, 1802Tribe: Blaptini Leach, 1815Subtribe: Blaptina Leach, 1815Genus: Blaps Fabricius, 1775Species: orientalis Solier, 1848
Material examined: Ent-1/56A/162, 1 ex., male, 07.vii.1961, National Chemical Laboratory campus, Pune (18054156 N & 73 081155 E altitude 594m), Maharashtra, India, coll. S.M. Ketkar.
Diagnostic characters: Body black, length 38mm, width 17mm, elongate-oval (Image 1A); head widest at eye level; antennae medium size, reaching close to the base of pronotum when directed backwards and apical antennomeres bearing only simple setiform sensoria; punctation of head not coarse, moderately dense.
Pronotum longer than wide, transverse, convex, narrowed backward and arched towards the base; sparsely and smoothly punctated. Pronotum 1.9 times as wide as head; anterior margin rounded, lateral margin rounded in anterior half and straight in basal half; basal margin truncate, closely embracing and overlapping basal aspect of elytra.
Elytra elongate (1.9 times as long as wide), with epipleura narrow throughout its length, broadest around its middle; 2.5 times as long and 1.3 times as wide as

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13037–13038
First report of Blaps orientalis from India Hegde et al.
13038
pronotum, 2.7 times as wide as head; elytra 3.9 times as long as mucro; mucro narrow and elongate. Length of caudal extension at the apex of elytra (mucro) is 6.26mm. Abdomen wrinkled and presence of punctures in fourth and fifth abdominal ventrites between irregular wrinkles. Presence of hair brush between first and second abdominal ventrites (Image 1B).
Table 1. List of all Blaps species reported from India
Species Distribution
Blaps apicecostata Blair, 1922 Sikkim
B. breiti Reitter, 1913 Himachal Pradesh
B. crassicornis (Fairmaire, 1891) Jammu & Kashmir
B. gentilis Fairmaire, 1887 The northern part of India
B. himalyaica Blair, 1923 Sikkim
B. indicola Bates, 1879 Jammu & Kashmir
B. marginicollis (Fairmaire, 1891) Jammu & Kashmir
B. moerens Allard, 1880 Uttarakhand and Himachal Pradesh
B. nadaii Medvedev, 2004 Uttarakhand
B. nathani Kulzer, 1956 Rajasthan
B. perlonga F. Bates, 1879 Jammu & Kashmir
B. socia Seidlitz, 1898 Uttarakhand and Sikkim
B. srinagaricus Kaszab, 1975 Jammu & Kashmir
B. tristiciae Bogatchev, 1949 Jammu & Kashmir
B. urophora Fairmaire, 1891 Jammu & Kashmir
Image 1. Full habitus of Blaps orientalis. A - dorsal view, B - ventral view
References
Aalbu, R.L., C.A. Triplehorn, J.M. Campbell, K.W. Brown, R. Somerby & D.B. Thomas (2002). 106. Tenebrionidae, pp. 463–509. In: Arnett, R.H. Jr., M.C. Thomas, P.E. Skelley & J.H. Frank (eds.). The Beetles of the United States, Vol. 2. CRC Press, New York, 850pp.
Bouchard, P., J.F. Lawrence, A.E. Davies & A.F. Newton (2005). Synoptic classification of the world Tenebrionidae (Insecta: Coleoptera) with a review of family-group names. Annales Zoologici 55(4): 499–530.
Condamine, F.L., L. Soldati, J.Y. Rasplus & G.J. Kergoat (2011). New insights on systematics and phylogenetics of Mediterranean Blaps species (Coleoptera: Tenebrionidae: Blaptini), assessed through morphology and dense taxon sampling. Systematic Entomology 36: 340–361; https://doi.org/10.1111/j.1365-3113.2010.00567.x
Löbl, I., M. Nabozhenko & O. Merkl (2008). Family Tenebrionidae Latreille, 1802, pp. 219–238. In: Löbl, I. & A. Sme-tana (eds.). Catalogue of Palaearctic Coleoptera, Tenebrionoidea, Vol. 5. Apollo Books, Stenstrup, Denmark, 670pp.
Medvedev, G.S. (2001). Evolution and system of darkling beetles of the tribe Blaptini (Coleoptera, Tenebrionidae). Chteniya Pamyati Nikolaya Aleksandrovicha Kholodkovskogo 53: 1–332.
Medvedev, G.S. (2007). A contribution to the taxonomy and morphology of the tribe Blaptini (Coleoptera, Tenebrionidae). Entomological Review 87: 181–214.
Medvedev, G.S. & O. Merkl (2002). Viettagona vietnamensis gen. et sp. n. from Vietnam (Coleoptera, Tenebrionidae: Blaptini). Acta Zoologica Academiae Scientiarum Hungaricae 48(4): 317–332.
Solier, A.J.J. (1848). Essai sur les Collapterides. 14e tribu - Blapsites, pp. 149–370. In: Baudi di Selve, F. (ed.). Studi Entomologici, Tom. I. Stamporio Degli Artisti Tipografi, Torino, Italy, 376pp.
Soldati, L., F.L. Condamine, A.L. Clamens & G.J. Kergoat (2017). Documenting tenebrionid diversity: progress on Blaps Fabricius (Coleoptera, Tenebrionidae, Tenebrioninae, Blaptini) systematics, with the description of five new species. European Journal of Taxonomy 282: 1–29.
Schuster, A. (1930). Neue Tenebrioniden (Coleopt.) aus Belutschistan. Koleopterologische Rundschau XV: 235–239.
Threatened Taxa

LOGOs
13039
Not
e
DOI: https://doi.org/10.11609/jott.4071.10.15.13039-13043
Editor: K. Ravikumar, I-AIM (FRLHT), Bengaluru, India. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 4071 | Received 10 February 2018 | Final received 14 November 2018 | Finally accepted 30 November 2018
Citation: Chowlu, K., A. Bhattcharjee & P. Kumar (2018). Notes on the occurrence of orchids Bulbophyllum medioximum, Herminium edgeworthii and H. macrophyl-lum (Orchidaceae) in Arunachal Pradesh, India. Journal of Threatened Taxa 10(15): 13039–13043; https://doi.org/10.11609/jott.4071.10.15.13039-13043
Copyright: © Chowlu et al. 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, reproduction and distribution by providing adequate credit to the authors and the source of publication.
Funding: Botanical Survey of India.
Competing interests: The authors declare no competing interests.
Acknowledgements: We are thankful to the Director, Paramjit Singh, Botanical Survey of India, Kolkata. We would like to thank Dr. Luvkush, ex research scholar of BSI, Dehradun for his suggestions. The first author would like to thank Mr. Sunil Soniyal who accompanied her to the forest during the field survey.
Notes on the occurrence of orchids Bulbophyllum medioximum, Herminium edgeworthii and H. macrophyllum (Orchidaceae) in Arunachal Pradesh, India
Krishna Chowlu 1 , Avishek Bhattacharjee 2 & Pankaj Kumar 3
1 Botanical Survey of India, Arunachal Pradesh Regional Centre, Senki View, Papum Pare, Itanagar, Arunachal Pradesh 791111, India2 Central National Herbarium, Botanical Survey of India, Acharya Jagadish Chandra Bose Indian Botanic Garden, Howrah, West Bengal 711103, India3 Kadoorie Farm and Botanic Garden, Lam Kam Road, Lam Tsuen, Tai Po, New Territories, Hong Kong S.A.R., China 1 [email protected], 2 [email protected], 3 [email protected] (corresponding author)
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13039–13043
Arunachal Pradesh, the northeastern most state of India, is a part of the eastern Himalaya and considered as one of the biodiversity rich regions of the Eastern Himalayan Biodiversity Hotspot (CEPF 2018). Out of 1,350 species of orchids recorded from India, about 560 species are reported from Arunachal Pradesh (Rao 2006). Arunachal Pradesh may be botanically termed as the ‘Orchid Paradise of India’ because of the maximum concentration of orchid species (about 40% of the country) in the state when compared to the other states of the country (Rao 2010). During a routine floristic survey in West Kameng and Tawang districts of Arunachal Pradesh, a few orchid specimens (in flowering) belonging to the genera Bulbophyllum and Herminium were collected by the first author and brought for cultivation to the botanical garden of the Botanical Survey of India,
Arunachal Pradesh Regional Centre, Itanagar for further studies. After dissection, a necessary perusal of the literature (Pearce & Cribb 2002; Luksom 2007; Chen et al. 2009) and consultation of herbarium material deposited at APFH, ARUN ASSAM, K and BM the specimens have been identified as Bulbophyllum medioximum J.J.Verm., Schuit. & de Vogel, Herminium macrophyllum (D.Don) Dandy. and Herminium edgeworthii (Hook. f. ex Collett) X.H.Jin, Schuit., Raskoti & Lu Q. Huang of which the former is a new record for Indian flora while the latter two have not been reported so far from Arunachal Pradesh, constituting new records for the state. An updated taxonomy of the species has been provided following Govaerts et al. (2018).
New Record for India 1. Bulbophyllum medioximum J.J.Verm., Schuit. & de Vogel, Phytotaxa 166: 104. 2014 (Image 1)
Ione annamensis Ridl., J. Nat. Hist. Soc. Siam 4: 115. 1921.
Sunipia annamensis (Ridl.) P.F.Hunt, Kew Bull. 26: 183. 1971.
Type: Vietnam, Langbian Province, South Annam, Langbian Peaks, C. Boden Kloss s.n.
(BM000525247) (BM!). Plant epiphytic herb, 18–21 cm high including
the inflorescence. Rhizome creeping, 5–11 mm long. Pseudobulbs 1.3–1.8 × 0.5–0.8 cm, ovoid, 1.2–1.7 cm apart on rhizome. Leaves solitary, 7–8 × 1.0–1.2 cm,

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13039–13043
Occurrence of Bulbophyllum medioximum, Herminium edgeworthii & H. macrophyllum in Arunachal Pradesh Chowlu et al.
13040
oblong-lanceolate, minutely bilobed at apex, coriaceous, petiole c. 0.4 cm long. Inflorescence 18–21 cm long, erect; peduncle 15–17 cm long, arising from the base of pseudobulbs, covered by the brown sheathing bracts at the base, upper portion with c. 5 sterile bracts, terete, greenish-black; rachis 2.5–3.0 cm long, up to four flowers per inflorescence. Flower c. 5mm across (petal to petal), widely opening, pale purple with deep purple linings, lip with pale purple with deep purple linings and yellow in epichile. Pedicellate ovary 6–7 mm long, light green. Floral bracts 4–5 × 2.3–2.5 mm, lanceolate, acute at apex, translucent creamy, prominent. Dorsal sepal 5–6 × 3–4 mm, ovate-lanceolate, acute at apex,
pale purple with deep purple lining, 5 nerved, margins minutely recurved. Lateral sepals 8–10 × 3.5–4.0 mm, oblong-lanceolate, bifid at apex, two sepals fused to form a single structure or sepal (synsepalum) but free at apex, lying perpendicular to the pedicellate ovary. Petals 2.5–3.0 × 2.0–2.5 mm, 1-nerved, triangular-ovate, acute, minutely serrate along the margin. Lip simple, 4–5 × 3.5 mm, cup shaped, keeled, pale purple with deep purple linings with epichile yellow, obtuse at apex, base thickened and connate with very short foot of column. Column 3.5–4 mm long, greenish-yellow, rostellum with 2 processes; foot c. 1mm long. Anther cap 0.7–0.8 × 0.5–0.6 mm, pale greenish-white.
Image 1. Bulbophyllum medioximum J.J.Verm., Schuit. & de Vogel. A - Habit; B - Inflorescence; C - Flower; D - Flower; E - Dissected parts; F - Lip; G - Column with ovary plus pedicel. © K. Chowlu

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13039–13043
Occurrence of Bulbophyllum medioximum, Herminium edgeworthii & H. macrophyllum in Arunachal Pradesh Chowlu et al.
13041
Flowering. November–December. Habitat: Commonly found growing in open grasslands
at around 1,071m elevation. Distribution: China, India, Thailand and Vietnam. Specimen examined: 40107 (ARUN), 02.x.2015, India,
Arunachal Pradesh, Kameng District, Orchid Sanctuary Sessa, 1,071m, coll. Krishna Chowlu.
New record for Arunachal Pradesh 1. Herminium macrophyllum (D. Don) Dandy. J. Bot. 70: 328. 1932 (Image 2)
Neottia macrophylla D.Don, Prodr. Fl. Nepal.: 27. 1825.
Image 2. Herminium macrophyllum (D. Don) Dandy. A & C - Habitat; B - Inflorescence; D - Habit; E - Bract; F - Flower; G - Flower (side view); H - Flower; I - floral parts; J - Column with ovary plus pedicel. © K. Chowlu
Spiranthes macrophylla (D.Don) Spreng., Syst. Veg. 3: 708. 1826.
Peristylus macrophyllus (D.Don) Lawkush, V.Kumar & Bankoti, Indian J. Forest. 36: 388. 2013.
Type: Nepal, 1819, N. Wallich s.n. (BM000034355) (BM!)
Plants 7–20 cm tall. Bulbs ovoid or ellipsoid. Stem green, terete with 2 or 3 leaved. Leaves basal, narrowly oblongelliptic, 4–10 × 0.7–2.2 cm, apex acute to subacute. Inflorescence 515cm long, green; peduncle terete; rachis 2.5–7 cm long, densely many flowered; floral bracts lanceolate, 1–2 mm long, apex acute, shorter than ovary. Flowers 2–3 mm across, yellowish-green or

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13039–13043
Occurrence of Bulbophyllum medioximum, Herminium edgeworthii & H. macrophyllum in Arunachal Pradesh Chowlu et al.
13042
Image 3. Herminium edgeworthii (Hook.f. ex Collett) X. H. Jin et. A & C - Part of Inflorescnce; B - Leaf; D - Habit; E - Inflorescence; F - Flower; G - Flower (side view); H - Bract; I - Floral parts; J - Lip with column; K - Lip (side view) with spur & pedicel ovary; L - Column with ovary plus pedicel. © K. Chowlu
green; ovary distinctly hooked toward apex. Dorsal sepal ovate, 1.3 –2.4 × 0.6–1.2 mm, obtuse, yellowish-green to green; lateral sepals oblong-lanceolate, 1.5–2.4 × 0.6–1.2 mm, acute. Petals spreading, ovate-lanceolate, 1.2–2.0 × 0.8–1.2 mm, acuminate, yellowish-green or green; lip ovate-lanceolate, 1.8–2.2 × 0.8–1 mm, 3lobed; Column less than 1mm.
Flowering: June–August. Habitat: Commonly found growing in open grasslands
at 3,670m elevation.
Distribution: China, India, Nepal and Pakistan. Specimen examined: 40930 (ARUN), 21.ix.2016,
India, Arunachal Pradesh, Tawang District, Sela Pass, 3,670m, coll. Krishna Chowlu.
2. Herminium edgeworthii (Hook.f. ex Collett) X.H.Jin, Schuit., Raskoti & Lu Q. Huang in Cladistics 32: 32. 2015 (Image 3)
Habenaria edgeworthii Hook. f. ex Collett in Fl. Siml.: 504, t. 166. 1902.

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13039–13043
Occurrence of Bulbophyllum medioximum, Herminium edgeworthii & H. macrophyllum in Arunachal Pradesh Chowlu et al.
13043
Habenaria acuminata sensu T.A. Rao in Bull. Bot. Surv. India 2: 89. 1960.
Platanthera edgeworthii (Hook. f. ex Collett) R.K. Gupta Fl. Nainital.: 349. 1968.
Type: India, Banasar, August 1834, M.P. Edgeworth s.n. (K000247468) (K!).
Plant fusiform. Stem leafy base sheathed. Leaves 3–4, 5–9 × 2–4.5 cm, lower broadly ovate– lanceolate, base sheathing, acute, margins undulate; Stem bracts lanceolate, acute. Inflorescence 14–26 cm long, subdensely many flowered; rachis erect, c. 20cm long, terete. Floral bracts lanceolate, acute to acuminate, 0.8–1.5 × 0.2–0.4 cm, green. Flower 1.5–1.8 cm long, green. Sepals broadly ovate, 5.5–6.5 × 6.0–7.0 mm, obtuse, concave, margins minutely hairy. Lateral sepals 6–8 × 5 mm, oblong–ovate, apex acute, margins minutely hairy, apex folded externally. Petals 7 × 4 mm, lanceolate base broad, apex obtuse, hooked with the dorsal sepal. Lip simple, 7–8 × 3–4 mm, oblong, obtuse, yellowish-green; spur 1.7–2.1 cm long, green, much longer than the pedicel ovary. Column 2–3 mm long; stigma oblong.
Flowering: August–September. Habitat: Found growing in open grassland along with
other grasses at around 2,915m elevation.
Distribution: China, India, Nepal and Pakistan. Specimens examined: 40936 (ARUN), 18.ix.2017,
India, Arunachal Pradesh, Tawang District, 2,915m, coll. Krishna Chowlu.
References
CEPF (2018). Biodiversity Hotspot maps. Conservation International Foundation. Published on the Internet; http://www.cepf.net/Pages/default.aspx. Accessed 20 January 2018.
Chen, Q., Z.J. Liu, G.H. Zhu, K.Y. Lang, Z.H. Ji, Y.B. Luo, X.B. Jin, P.J. Cribb, J.J. Wood, S.W. Gale, P. Ormerod, J.J. Vermeulen, H.P. Wood, D. Clayton & A. Bell (2009). Orchidaceae, Pp. 236–245. In: Wu, Z.Y., P.H. Raven & D.Y. Hong (eds.). Flora of China - Vol. 25. Science Press, Beijing & Missouri Botanical Garden Press, St. Louis.
Govaerts, R., M.A. Campacci, D.H. Baptista, P.J. Cribb, A. George, K. Kreutz & J.J. Wood (2018). World Checklist of Orchidaceae. The Board of Trustees of the Royal Botanic Gardens, Kew. Published on the Internet: https://www.kew.org/wcsp/monocots/ (accessed on 14 November 2018)
Luksom, S.Z. (2007). The Orchids of Sikkim and North East Himalaya. Concept, Siliguri. 984pp.
Pearce, N.R. & P.J. Cribb (2002). Orchids of Bhutan. Royal Botanic Garden Edinburg and Royal Government of Bhutan, 221–233pp.
Rao, A.N. (2006). Alpine orchids of Arunachal Pradesh. Bulletin of Arunachal Forest Research 22(1&2): 27–32.
Rao, A.N. (2010). Orchid Flora of Arunachal Pradesh an update. Bulletin of Arunachal Forest Research 26(1&2): 82–110.
Threatened Taxa

13044
Not
e
DOI: https://doi.org/10.11609/jott.4321.10.15.13044-13045
Editor: Mandar N. Datar, MACS-Agharkar Research Institute, Pune, India. Date of publication: 26 December 2018 (online & print)
Manuscript details: Ms # 4321 | Received 07 June 2018 | Final received 17 December 2018 | Finally accepted 20 December 2018
Citation: Shameer, M.C. & V.K. Sreenivas (2018). Lectotypification of two names in the genus Gymnostachyum (Acanthaceae). Journal of Threatened Taxa 10(15): 13044–13045; https://doi.org/10.11609/jott.4321.10.15.13044-13045
Copyright: © Shameer & Sreenivas 2018. Creative Commons Attribution 4.0 International License. JoTT allows unrestricted use of this article in any medium, repro-duction and distribution by providing adequate credit to the authors and the source of publication.
Funding: None.
Competing interests: The authors declare no competing interests.
Acknowledgements: Authors are grateful to the personnel of the following herbaria: CAL, MH and K, for permitting to access their digital images to do this research; Dr. K.M. Prabhukumar for his suggestions and directions for improving the manuscript. Critical comments received from the anonymous reviewers helped us to improve the manuscript.
Lectotypification of two names in the genus Gymnostachyum (Acanthaceae)
M.C. Shameer 1 & V.K. Sreenivas 2
1,2 Department of Botany, Sri Vyasa N.S.S. College, Vyasagiri P.O., Wadakkanchery, Thrissur District, Kerala 680652, India
1 Research and Development Centre, Bharathiar University, Maruthamalai Road, Coimbatore, Tamil Nadu 641046, India
1 [email protected] (corresponding author), 2 [email protected]
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13044–13045
The genus Gymnostachyum Nees, belongs to the family Acanthaceae, comprises about 52 species (www.plantsoftheworldonl ine.org) , distributed mainly in tropical Asia (www.efloras.org). In India, 11 species and four varieties have been recorded to date from various geographical regions (Karthikeyan
et al. 2009); of which, 12 taxa are recorded so far from peninsular India (Gamble 1924; Prabhukumar et al. 2015). As part of the ongoing taxonomic revision in the genus Gymnostachyum in India, the need for lectotypification of two names was identified, viz., G. glabrum (Dalzell) T. Anderson (1850: 338, 1867: 506) and G. glabrum var. denticulatum C.B. Clarke (1885: 509). Clarke (1885) delineates var. denticulatum from latter by its distinctly toothed and glabrous nature of lamina. The lectotypes are designated here according to ICN Shenzhen Code Art. 9.3 (Turland et al. 2018).
Gymnostachyum glabrum (Dalzell) T. Anderson (1867: 506)
Cryptophragmium glabrum Dalzell (1850: 338) Type (lectotype, designated here): India,
Maharashtra, Bombay Ghats, N.A. Dalzell s.n., (K000885669) [K; digital image!] Image 1.
Residual syntypes: India, Maharashtra, South Concan, M Law s.n., (K000885668) [digital image!]
Nomenclatural notes: Dalzell (1850) proposed the name Cryptophragmium glabrum based on his collections from Bombay Ghats and Law’s collection
from South Concan (present day Konkan) region, India. Later, Anderson (1867) transferred the name to the genus Gymnostachyum, as G. glabrum. During the study, we have traced two specimens, which represent duplicates from a heterogenous collection (Dalzell’s and Law’s) with one sheet each. Both the sheets are well preserved and bear flowers and fruits. According to ICN Art. 9.3 (Turland et al. 2018), the specimen collected by the author and kept in K (K000885669) fits the description, and is preserved very well with locality and collector name, which is considered as the best choice and designated here as the lectotype.
Apart from this, we have also traced one more specimen from CAL (CAL0000019998), unfortunately, the collector’s name, date of collection and locality were missing in the sheet, hence not considered as type.
Gymnostachyum glabrum (Dalzell) T. Anderson var. denticulatum C.B. Clarke (1884: 509).
Type (lectotype, designated here): India, Maharashtra, Concan Ghat, Kala Naddi, December 1852, D Ritchie 1211 (K000885672) [digital images!] Image 2.
ISSN 0974-7907 (Online)ISSN 0974-7893 (Print)
OPEN ACCESS

Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018 | 10(15): 13044–13045
Lectotypification of two names in the genus Gymnostachyum Shameer & Sreenivas
13045
Residual syntypes: India, Maharashtra, Ram Ghat, February 1853, D Ritchie 1211 (K barcode K000885670) [digital image!]; N.A. Dalzell s.n., (K000885671) [digital image!]
Nomenclatural notes: Clarke (1884) proposed a new variety of G. glabrum, var. denticulatum based on the collections of Dalzell & Ritchie from Concan Ghats of India. Earlier both the collectors had included their collections under the name Cryptophragmium glabrum. In the protologue, the author mentioned about two collections and we have traced three sheets, all are housed at K. We found that Ritchie collected the plants in two seasons, viz., December 1852 (K000885672) and February 1853 (K000885670) and both the specimens were mounted on a single sheet with a single collection number. All the sheets are well preserved and bear flowers and fruits. In Dalzell’s collection, he has not mentioned the locality, collection number and date of collection. According to ICN Art. 9.3 of Shenzhen code (Turland et al. 2018), the specimen collected by Ritchie and mounted on the right side of the sheet with locality, date and collection number kept at K (K000885672) fits the description, which is considered as the best choice and designated here as the lectotype.
References
Anderson, T. (1867). An enumeration of the Indian species of Acanthaceae. Botanical Journal of Linnean Society 7: 425–526.
Clarke, C.B. (1885). Acanthaceae, pp. 387–558. In: Hooker, J.D. (ed.) The Flora of British India - 4. Reeve & Co., Ltd., London.
Dalzell, N.A. (1850). Contributions to the Botany of western India. Hooker’s Journal of Botany and Kew Garden Miscellany 2: 336–344.
Karthikeyan, S., M.Sanjappa & S.Moorthy (2009). Flowering Plants of India - Dicotyledons, Volume 1 (Acanthaceae – Avicenniaceae). Botanical Survey of India, Kolkata, 365pp.
Turland, N.J., J.H. Wiersema, F.R. Barrie, W. Greuter, D.L. Hawksworth, P.S. Herendeen, S. Knapp, W.-H. Kusber, D.-Z. Li, K. Marhold, T.W. May, J. McNeill, A.M. Monro, J. Prado, M.J. Price & G.F. Smith (eds.) (2018). International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017. Regnum Vegetabile 159. Koeltz Botanical Books, Glashütten: https://doi.org/10.12705/Code.2018
Prabhukumar, K.M., B. Indira, V.B. Sreekumar, K.S. Ramya, S. George, K.J. Dantas & A.D. Suma (2015). Gymnostachyum warrieranum (Acanthaceae), a new species from Western Ghats of India with morphological and molecular data. Kew Bulletin 70(3): 1–9; https://doi.org/10.1007/s12225-015-9590-9
www.efloras.org (accessed on 14 April 2018)www.plantsoftheworldonline.org (accessed on 14 December 2018)
Image 1. Digital Image of proposed lectotype housed at K (barcode K000885669).
Image 2. Digital Image of proposed lectotype housed at K (barcode K000885672).
Threatened Taxa

National Biodiversity Authoritywww.nbaindia.org
Journal of Threatened Taxa | www.threatenedtaxa.org | 26 December 2018
Logo Design Contest for National Biodiversity Authorityrt
National Biodiversity Authority (NBA), a statutory body under Ministry of Environment Forest & Climate Change was established by the Central Government in 2003 to implement the provisions of the Biological Diversity Act, 2002. Its head-quarters is situated in Chennai, Tamil Nadu. As a Statutory Body, NBA performs facilitative, regulatory and advisory func-tions for the Government of India on issues of conservation, sustainable use of biological resources and fair and equitable sharing of benefits arising out of the use of biological resourc-es. NBA regulates activities like research, commercial utiliza-tion, bio-survey and bio-utilization of biological resources and/or associated knowledge occurring in India. It also grants ap-proval for persons seeking Intellectual property rights for inventions based on Indian biological resources or associated knowl-edge. Thus, companies, institutions and individuals that access biological resources and/or associated knowledge for developing products and obtaining patents shall seek NBA approval before undertaking such activities. At the time of granting approval, NBA would insist that a certain percentage of the profit that the company or individual reaps shall be shared with NBA. The benefits shared in such a manner would be used for conservation and promotion of biodiversity, incentivising communities involved in conservation of the resource and socio economic development of areas from where the resource was accessed. NBA is now plan-ning to develop a logo/ mark which the companies or individuals can affix on their product for which they have obtained approval from NBA. Further details with respect to National Biodiversity Authority may be seen at the website http://nbaindia.org/ In this regard, NBA invites entries for the logo contest.
Participants are required to conceptualize and design a mark/logo such that-
1. The mark be distinctive and may convey the three objectives of the Biological Diversity Act, 2002, namely – conserva-tion of biological diversity, sustainable use of its components and fair and equitable sharing of benefits arising out of its utilization.2. The mark shall dignify the concept of ‘Access and Benefit Sharing (ABS)’ which denotes the equitable sharing of benefits by the users (Companies/Institutions/Industry) to the providers (Communities/Farmers/Traditional Knowledge Holders) of biological resources.3. The mark can be used by companies to advertise that their product is based on a biological resource and/ or associated knowledge from India.4. The mark will be certifying that the particular product on which it is affixed is complying with ‘Access and Benefit Shar-ing (ABS)’ requirements under Indian law.5. Consumers can choose products with this mark and can be assured that the product was manufactured using biological resources and/or associated knowledge from India and that it has met the ABS requirements under Indian law.
The Designer of the finally selected Logo will get a prize of Rs. 50,000/- (Rupees fifty thousand only) and would be required to give copyright of the design to National Biodiversity Authority. (The prize money will be payable after deduction of TDS).
The last date for accepting entries is 21st January, 2019 till 06.00PM.
Click here to read Terms and Condition <https://static.mygov.in/rest/s3fs-public/mygov_154511177051553221.pdf>
For any queries write to: Mr. T. Narendran, Technical Officer (IPR) Tel: 044-22542777/1075 E-mail: [email protected]

Jorg Freyhof, Leibniz Institute of Freshwater Ecoloy and Inland Fisheries, Berlin, GermanyJózsef Lanszki, University of West Hungary, Sopron, HungaryK. Haridasan, FRLHT, Bengaluru, IndiaK. Karthigeyan, Botanical Survey of India, Howrah, IndiaKlaus Ruetzler, NMNHistory, Smithsonian Institution, Washington, USA. K. Ravikumar, FRLHT, Bengaluru, Karnataka, IndiaK. Veenakumari, NBAIR, Bengaluru, India. K.A. Subramanian, Zoological Survey of India, New Alipore, Kolkata, IndiaKees Rookmaaker, Rhino Resource Center, United KingdomKelly B. Miller, University of New Mexico, USA K.S. Gopi Sundar, International Crane Foundation, Baraboo, USAK.S. Negi, NBPGR-ICAR, Nainital District, Uttarakhand, IndiaK.R. Sasidharan, Institute of Forest Genetics and Tree Breeding, Coimbatore, IndiaKailash Chandra, Zoological Survey of India, Jabalpur, Madhya Pradesh, IndiaKareen Schnabel, NIWA, Wellington, New Zealand Karin Schwartz, George Mason University, Fairfax, VirginiaKarol Bucsek, Witt Museum, München, Germany Kevin Smith, IUCN, Cambridge, UKKlaus Ruetzler, Smithsonian Institution, Washington, DCKristin Leus, Copenhagen Zoo, Annuntiatenstraat, Merksem, BelgiumKurt R. Arnold, North Dakota State University, Saxony, Germany L.D. Singla, Guru Angad Dev Veterinary and Animal Sciences University, Ludhiana, India Lala A.K. Singh, Bhubaneswar, Orissa, IndiaLarry R. Noblick, Montgomery Botanical Center, Miami, USA Lional Monod, Natural History Museum of Geneva, Genève, Switzerland Llewellyn D. Densmore, Texas Tech University, Lubbock, USALukas Rüber, Department of Vertebrates, Natural History Museum, SwitzerlandM. Afzal Khan, Department of Zoology, Aligarh Muslim University, Aligarh, IndiaMandar N. Datar, Agharkar Research Institute, Pune, India Mandar S. Paingankar, University of Pune, Pune, Maharashtra, IndiaMarc W. Holderied, University of Bristol, Bristol, UK Mario Cohn-Haft, Instituto Nacional de Pesquisas da Amazônia (INPA), Brasil Martin B.D. Stiewe, The Natural History Museum, UKMary K. Wicksten, Texas A&M University, College Station, USA Matheus dos Santos Rocha, Universidade do Vale do Rio dos Sinos, Brasil Merlin Franco, Curtin University, MalaysiaMichael J. Parr, American Bird Conservancy, Washington, USA Mewa Singh, Mysore University, Mysuru, IndiaMohammad Hayat, Aligarh Muslim University, Aligarh, IndiaMohilal Meitei, Manipur University, Camchipur, Manipur, IndiaM. Nithyanandan, Environmental Department, La Ala Al Kuwait Real Estate. Co. K.S.C., KuwaitM. Mike Kerry, Seaford, East Sussex, UK M. Sabu, University of Calicut, Malappuram, India M.K. Vasudeva Rao, Shiv Ranjani Housing Society, Pune, IndiaN.P. Balakrishnan, Ret. Joint Director, BSI, Coimbatore, IndiaNancy van der Poorten, Toronto, CanadaNathalie Yonow, Swansea University, Swansea, UK Nguyen Thi Phuong Lien, Vietnam Academy of Science and Technology, Hanoi, Vietnam Neelesh Dahanukar, IISER, Pune, Maharashtra, IndiaNorbert Delahaye, Colombo, Sri LankaNoor Azhar Mohamed Shazili, Universiti Malaysia Terrengganu, MalaysiaOguz Turkozan, Adnan Menderes University, Aydın, Turkey Okan Külköylüoğlu, Abant Izzet Baysal University, Bolu, TurkeyOlivier S.G. Pauwels, Royal Belgian Institute of Natural Sciences, Belgium Pankaj Kumar, Kadoorie Farm and Botanic Garden Corporation, Hong Kong S.A.R., ChinaPartha Pratim Bhattacharjee, Tripura University, Suryamaninagar, IndiaPaul A. Racey, University of Exeter, Devon, UK Paul J.J. Bates, Harrison Institute, Kent, UKPenelope Greenslade, Federation University, Ballarat, Australia Peter Boveng, NOAA Alaska Fisheries Science Center, Seattle, USAPhyllis C. Lee, University of Stirling, Stirling, UK Pierre Moulet, Museum Requien, Avignon, France Pritpal S. Soorae, Environment Agency, Abu Dhabi, UAEPriya Davidar, Pondicherry University, Kalapet, Puducherry, IndiaP. Lakshminarasimhan, Botanical Survey of India, Pune, IndiaP.M. Sureshan, Zoological Survey of India, Kozhikode, Kerala, IndiaP.O. Nameer, Kerala Agricultural University, Thrissur, Kerala, IndiaP.S. Easa, Kerala Forest Research Institute, Peechi, India Purnendu Roy, London, UK Qiang Liu, Xishuangbanna Tropical Botanical Garden, Yunnan, ChinaR. Sundararaj, Institute of Wood Science & Technology, Bengaluru, IndiaR. Varatharajan, Manipur University, Imphal, Manipur, IndiaR.K. Avasthi, Rohtak University, Haryana, India
R.K. Verma, Tropical Forest Research Institute, Jabalpur, IndiaR.M. Sharma, (Retd.) Scientist, Zoological Survey of India, Pune, IndiaRainer Hutterer, Zoological Research Museum Alexander Koenig, Bonn, GermanyRagnar Kinzelbach, University of Rostock, Rostock, GermanyRajah Jayapal, SACON, Coimbatore, Tamil Nadu, IndiaRajashekhar K. Patil, Mangalore University, Mangalore, IndiaRajeev Raghavan, Kerela University of Fisheries and Ocean Studies (KUFOS), Kochi, India Rajiv S. Kalsi, M.L.N. College, Yamuna Nagar, Haryana, IndiaRaju Vyas, Vadodara, IndiaRaymond Henry, Auburn University, Auburn, USARenkang Peng, Charles Darwin University, Darwin, Australia Reuven Yosef, Ben Gurion University of the Negev, Eilat Campus, Israel. Richard Corlett, Xishuangbanna Tropical Botanical Garden, Yunnan, ChinaRichard Gallon, llandudno, North Wales, LL30 1UPRichard Kiprono Mibey, Vice Chancellor, Moi University, Eldoret, Kenya Robert W. Sites, University of Missouri, Columbia, USA. Robert D. Sluka, Chiltern Gateway Project, A Rocha UK, Southall, Middlesex, UKRobin Wen Jiang Ngiam, National Parks Board, Singapore Robin Wilson, Museum Victoria, Melbourne, AustraliaRoland Wirth, Zoologische Gesellschaft für Arten-und Populationsschutz, Germany Rory Dow, National Museum of natural History Naturalis, The NetherlandsRosana Moreira da Rocha, Universidade Federal do Paraná, Curitiba, Brasil S. Ajmal Khan, Annamalai University, Parangipettai, India S. Arularasan, Annamalai University, Parangipettai, India S. Balachandran, Bombay Natural History Society, Mumbai, India S.C. Verma, Professor Emeritus, Panjab University, Chandigarh, India S. Gombobaatar, National University of Mongolia, Ulaanbaatar, Mongolia Saito Motoki, The Butterfly Society of Japan, Tokyo, JapanSanjay Sondhi, Titli Trust, Dehradun, IndiaSanjeeva Nayaka, CSIR-National Botanical Research Institute, Lucknow, India Shomita Mukherjee, SACON, Coimbatore, Tamil Nadu, IndiaShonil A. Bhagwat, The Open University, UKSpartaco Gippoliti, Societa Italiana per la Storia della Fauna ‘G. Altobello’, Roma, ItalyStephen D. Cairns, Smithsonian Institution, Washington, USA Stephen D. Nash, Scientific Illustrator, State University of New York, NY, USAStephen C. Weeks, The University of Akron, Ohio, USASushil K. Dutta, Centre for Ecological Sciences, IISc, Bengaluru, Karnataka, IndiaTadashi Kawai, Wakkanai Fisheries Research Institute, Hokkaido, JapanTaej Mundkur, Wetlands International, Wageningen, The NetherlandsTim New, La Trobe University, Melbourne Victoria, AustraliaTony Whitten, Fauna & Flora International, Cambridge, UK Topiltzin Contreras MacBeath, Universidad Autónoma del estado de Morelos, MéxicoUllasa Kodandaramaiah, IISER-TVM, Thiruvananthapuram, India Uwe Braun, Martin-Luther-Universität, Neuwerk, Germany Vatsavaya S. Raju, Kakatiya University, Warangal, IndiaV.B. Hosagoudar, Bilgi, Karnataka, India V. Irudayaraj, St. Xavier’s College, Palayamkottai, Tamil Nadu, IndiaV. Gokula, National College, Tiruchirappalli, India V. Sampath Kumar, Botanical Survey of India, Howrah, IndiaV. Santharam, Institute of Bird Studies & Natural History, Chittoor, IndiaVijayasankar Raman, University of Mississippi, USAW. Vishwanath, Manipur University, Imphal, IndiaWiebke Herding, Amsterdam, The NetherlandsWioletta Tomaszewska, Museum and Institute of Zoology, Wilcza, PolandXiaoli Tong, South China Agricultural University, Guangzhou, China
Reviewers 2015–2017 Due to pausity of space, the list of reviewers for 2015-2017 is available online.
English EditorsMrs. Mira Bhojwani, Pune, India Dr. Fred Pluthero, Toronto, Canada Mr. P. Ilangovan, Chennai, India
English/Copy EditorVidya Mary George, Bengaluru, India
Web Design Latha G. Ravikumar, Coimbatore, India
Typesetting Arul Jagadish, Coimbatore, IndiaS. Radhika, Coimbatore, IndiaK. Geetha, Coimbatore, IndiaK. Ravindran, Coimbatore, India
Journal of Threatened Taxa is indexed/abstracted in Bibliog-raphy of Systematic Mycology, Biological Abstracts, BIOSIS Previews, CAB Abstracts, EBSCO, Google Scholar, Index Copernicus, Index Fungorum, JournalSeek, National Academy of Agricultural Sciences, NewJour, OCLC WorldCat, SCOPUS, Stanford University Libraries, Virtual Library of Biology, Zoo-logical Records.
NAAS rating (India) 5.10
Print copies of the Journal are available at cost. Write to the Managing Editor, JoTT, c/o Wildlife Information Liaison Development, No. 12, Thiruvannamalai Nagar, Saravanampatti - Kalapatti Road, Saravanampatti, Coimbatore, Tamil Nadu 641035, [email protected]

Contribution to the Macromycetes of West Bengal, India: 28–33-- Rituparna Saha, Arun Kumar Dutta, Soumitra Paloi, Anirban Roy & Krishnendu Acharya, Pp. 13006–13013
Notes
The identification of Takin Budorcas taxicolor (Mammalia: Bovidae) through dorsal guard hair-- Manokaran Kamalakannan, Pp. 13014–13016
Photographic evidence of Striped Hyena Hyaena hyaena (Mammalia: Carnivora: Hyaenidae) in Ramnagar forest division, Uttarakhand, India-- Vipul Maurya, Jai Pratap Singh, Kahkashan Naseem, Surender Mehra, Parag M. Dhakate, Neha Verma & A.G. Ansari, Pp. 13017–13019
Range extension of the Least Leaf-nosed Bat Hipposideros cineraceus Blyth, 1853 (Mammalia: Chiroptera: Hipposideridae): to central India-- M. Kamalakannan, C. Venkatraman, Tauseef Hamid Dar & Kailash Chandra, Pp. 13020–13023
A report on the possible interbreeding between Grizzled Giant Squirrel Ratufa macroura and Indian Giant Squirrel Ratufa indica from Chinnar Wildlife Sanctuary in the southern Western Ghats, India-- Kiran Thomas, D.K. Vinodkumar, Jomals Mathews John, M. Shaji & P.O. Nameer, Pp. 13024–13028
Ischnura fountaineae (Insecta: Odonata: Zygoptera) in Oman, eastern Arabia-- Elaine Mary Cowan & Peter John Cowan, Pp. 13029–13031
First record of Leptogenys hysterica Forel, 1900 (Hymenoptera: Formicidae: Ponerinae) from Pakistan-- Muhammad Tariq Rasheed, Imran Bodlah, Ammara Gull e Fareen & Xiaolei Huang, Pp. 13032–13036
First report of darkling beetle Blaps orientalis Solier, 1848 (Coleoptera: Tenebrionidae) from India-- V.D. Hegde, D. Vasanthakumar & S.V. Manthen, Pp. 13037–13038
Notes on the occurrence of orchids Bulbophyllum medioximum, Herminium edgeworthii and H. macrophyllum (Orchidaceae) in Arunachal Pradesh, India-- Krishna Chowlu, Avishek Bhattacharjee & Pankaj Kumar, Pp. 13039–13043
Lectotypification of two names in the genus Gymnostachyum (Acanthaceae)-- M.C. Shameer & V.K. Sreenivas, Pp. 13044–13045
Miscellaneous
National Biodiversity Authority
Partners
Threatened Taxa
Member
Publisher & Host
www.threatenedtaxa.org
The Journal of Threatened Taxa is dedicated to building evidence for conservation globally by publishing peer-reviewed articles online every month at a reasonably rapid rate at www.threatenedtaxa.org. All articles published in JoTT are registered under Creative Commons Attribution 4.0 International License unless otherwise mentioned. JoTT allows unrestricted use of articles in any medium, reproduction, and distribution by providing adequate credit to the authors and the source of publication.
OPEN ACCESS
ISSN 0974-7907 (Online); ISSN 0974-7893 (Print)
December 2018 | Vol. 10 | No. 15 | Pages: 12907–13046Date of Publication: 26 December 2018 (Online & Print)
DOI: 10.11609/jott.2018.10.15.12907-13046
Articles
Dietary preference and feeding patterns of the urban Rhesus Macaque Macaca mulatta (Mammalia: Primates: Cercopithecidae) in Asola-Bhatti Wildlife Sanctuary in India-- Ishita Ganguly & Netrapal Singh Chauhan, Pp. 12907–12915
Postembryonic development of the Tri-spine Horseshoe Crab Tachypleus tridentatus (Merostomata: Xiphosura) in a nursery habitat in the Philippines-- Dorkas Kaiser & Sabine Schoppe, Pp. 12916–12932
Communications
Copulatory behavior of the Jaguar Panthera onca (Mammalia: Carnivora: Felidae)-- Pedro Nacib Jorge-Neto, Cristiane Schilbach Pizzutto, Gediendson Ribeiro de Araujo, Thyara de Deco-Souza, Leanes Cruz da Silva, Jorge Aparecido Salomão Jr. & Hernan Baldassare, Pp. 12933–12939
Amphibians of the Dibang River Basin, Arunachal Pradesh: an annotated checklist with distribution records-- Jayanta K. Roy, Ramie H. Begum & M. Firoz Ahmed, Pp. 12940–12952
Taxonomic studies on the gaudy grasshoppers (Orthoptera: Pyrgomorphoidea: Pyrgomorphidae) from the northeastern states of India-- M. Imran Khan, M. Kamil Usmani, Shahnila Usmani & Hira Naz, Pp. 12953–12968
Odonata (Insecta) diversity of Kuldiha Wildlife Sanctuary and its adjoining areas, Odisha, eastern India-- Subrat Debata & Kedar Kumar Swain, Pp. 12969–12978
Short Communications
On the diversity of the vertebrate fauna (excluding fishes) of Panchet Hill (Garh Panchkot), Purulia, West Bengal, India-- Sanjib Chattopadhyay, Somenath Dey & Utpal Singha Roy, 12979–12985
First record of the rare Furry Lobster Palinurellus wieneckii (De Man, 1881) (Decapoda: Palinuridae) from the Arabian Sea-- K.K. Idreesbabu, C.P. Rajool Shanis & S. Sureshkumar, Pp. 12986–12989
Description of life stages of dung beetle Scaptodera rhadamistus (Fabricius, 1775) (Coleoptera: Scarabaeidae: Scarabaeinae) with notes on nesting and biology-- Suvarna S. Khadakkar, Ashish D. Tiple & Arun M. Khurad, Pp. 12990–12994
An updated list of Odonata of southwestern Bangladesh-- M. Sajjad Hossain Tuhin & M. Kawsar Khan, Pp. 12995–13001
On the reproductive biology of Salacia fruticosa Wall. ex M.A. Lawson - an endemic medicinal plant of the Western Ghats, india-- K. Subin, P.A. Jose & T.V. Sarath, Pp. 13002–13005





























