Embed Size (px)
Citation preview

21PROASELLUS SPECIES GROUP FROM LAKE OHRID
BIOLOGICAL LETT. 2006, 43(1): 21�32Available online at http://www.biollett.amu.edu.pl
Notes on genetic differentiation within the endemic Proasellusspecies group from Lake Ohrid (Balkan Peninsula)
ADRIANNA KILIKOWSKA1* , ANNA WYSOCKA1, JERZY SELL1,MAREK ZIÊTARA2 and TADEUSZ SYWULA
1Department of Genetics and Cytology, University of Gdañsk, 80-822 Gdañsk, Poland2Biological Station, University of Gdañsk, 80-680 Gdañsk, Poland
(Received on 20th June 2005; Accepted on 25th March 2006)
This study is part of the last project designed by Professor Tadeusz Sywula.The authors dedicate the present paper to his memory.
Abstract: The Balkan relict Lake Ohrid is inhabited by 4 isopod species representing the freshwaterfamily Asellidae. Three of them are endemic and belong to the genus Proasellus: P. remyi, P. arnautovici,P. gjorgjevici; the fourth species, Asellus aquaticus, is widespread in Europe. The endemic species aredivided into morphological forms related to the lake�s vertical zones. An electrophoretic survey of allozymevariation in 2 polymorphic loci (Pgm and Gpi) was conducted in the 3 endemic species and their subspeciesor forms. The observed patterns of morphological differentiation within examined species are supportedby genetic data. Thus the differentiation probably results from a process of intralacustrine speciation,which could be based on the mechanism of ecological isolation by habitat. Moreover, the data suggestthat all examined taxa may have been reproductively isolated.
Key words: Asellidae, Proasellus, allozymes, endemic species, genetic relationships, population genetics
INTRODUCTION
Lake Ohrid, surrounded by high karstic mountain ranges, is situated on theBalkan Peninsula in the eastern part of the Mediterranean region (Fig. 1). It is a typicaloligotrophic, tectonic lake that was created in the Tertiary period. With its 348 km2
of surface and maximum depth of 286 m, the lake is one of the biggest among theancient lakes (STANKOVIÈ 1960, MARTENS 1997).
The lake is an ideal place to study evolutionary processes of intralacustrinespeciation because its fauna is characterized by an exceptionally high level of ende-mism and diversity caused by the long isolation from other water systems. The high-est degree of endemicity is observed within gastropods (about 86% of the 50 knownspecies), fish (60%) and flatworms (79%) (KRSTANOVSKI 1994, KRSTANOVSKI &
* All corespondence please send to: A. Kilikowska, Department of Genetics and Cytology, University of Gdañsk, ul. K³adki 24,80-822 Gdañsk, Poland; Tel: + 48 58 301 22 41 ex: 352; e-mail: [email protected]
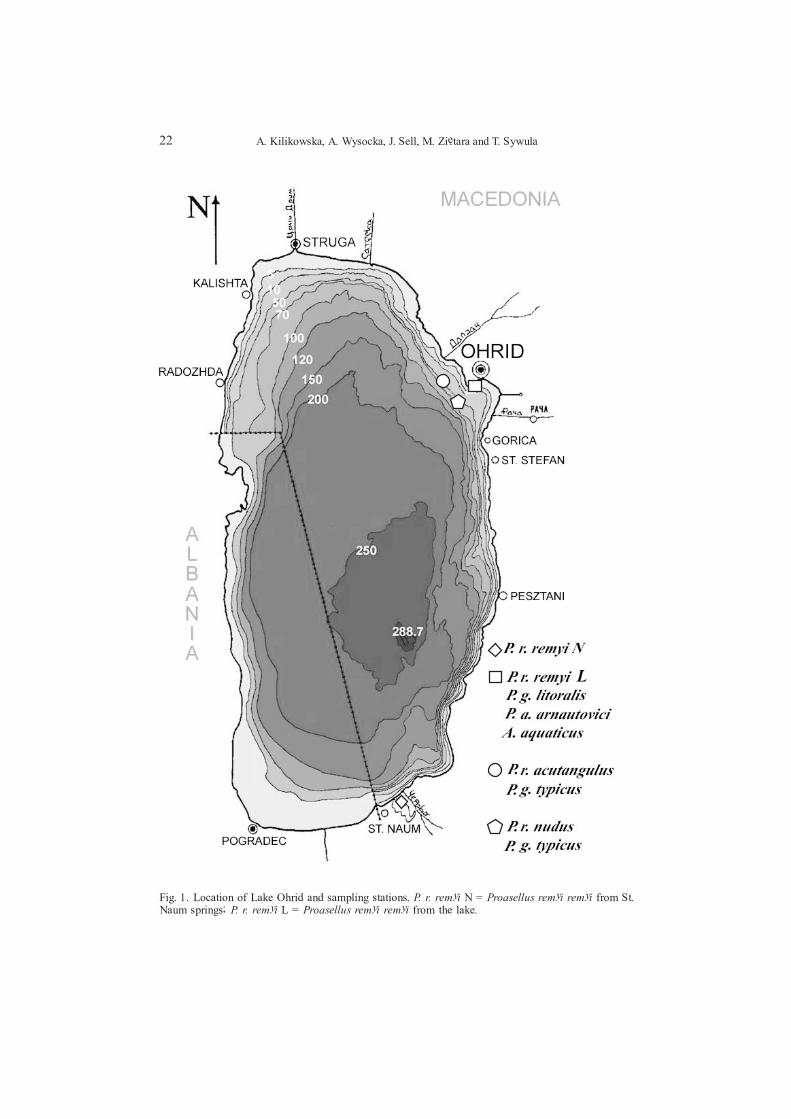
22 A. Kilikowska, A. Wysocka, J. Sell, M. Ziêtara and T. Sywula
Fig. 1. Location of Lake Ohrid and sampling stations. P. r. remyi N = Proasellus remyi remyi from St.Naum springs; P. r. remyi L = Proasellus remyi remyi from the lake.

23PROASELLUS SPECIES GROUP FROM LAKE OHRID
TRAJANOVSKI 2002, SPIRKOVSKI et al. 2002). There are also many other examplesof endemic fauna within the Oligochaeta and Hirudinea (STANKOVIÈ 1960). Crusta-ceans are also subject to adaptive radiation in ancient lakes. It was argued thatthe percentage of crustacean endemicity in ancient lakes is higher than in otherlong-lived habitats (MARTENS & SCHÖN 1999). Undoubtedly, the most remarkableexample of crustacean radiation are gammaroidean amphipods from Lake Baikal(MARTENS & SCHÖN 1999, SHERBAKOV 1999, SHERBAKOV et al. 1999, VÄINÖLÄ &KAMALTYNOV 1999, MACDONALD et al. 2005). Besides, radiations of the Ostracodaand Isopoda in ancient lakes have also been reported (MARTENS & SCHÖN 1999, SA-LEMAA & KAMALTYNOV 1994).
Lake Ohrid is inhabited by 4 isopod species representing the freshwater fam-ily Asellidae. Three of them are endemic and belong to the genus Proasellus Du-dich, 1925: P. remyi (Monod, 1932), P. arnautovici (Remy, 1932), P. gjorgjevici(Karaman, 1933). The fourth asellid species recorded in Lake Ohrid is Asellus aquati-cus Linnaeus, 1758, which is widespread in Europe and represents the nominategenus Asellus (KARAMAN 1953, STANKOVIÈ 1960). Taxonomic studies conducted byKARAMAN (1953) have led to the description of the new subspecies P. arnautovicielongatus (different from the littoral P. arnautovici arnautovici) and several morpho-logical forms: P. remyi f. remyi, P. remyi f. acutangulus, P. remyi f. nudus,P. gjorgjevici f. typicus, and P. gjorgjevici f. litoralis.
The taxonomy of asellids has focused on the morphology of the male pleopodII, characterized by specific copulatory appendages. Besides, all these forms arecharacterized by a complex of different morphological features. They differ in numberof segments on the antennal flagella, development and pigmentation of eyes, pro-portions of the body, shape of the head and of the pleotelson, and appearance ofchitinous bristles on the epimerites (KARAMAN 1953). Moreover, each form is lim-ited to the particular vertical zone characterized by specific conditions of life andthe bottom type. The most diverse littoral zone is inhabited by very numerous P. remyif. remyi, and relatively rare P. gjorgjevici f. litoralis and P. arnautovici arnautovici.The bottom of the littoral zone is being constantly washed by the moving waters. Itis bare or covered by different types of rocks � from great blocks to coarse sand.Littoral isopods can be found adhered to the stones mainly. In places with macro-phytic vegetation, where the bottom is even covered by mud with organic detritus,asellids can also be found, but not very often. Besides, in this zone young individ-uals of Dreissena sp. attained its maximum and their shells are very often inhabitedby Proasellus species. The annual amplitude of temperature in the littoral is greaterthan in other zones (STANKOVIÈ 1960). In the sublittoral, P. remyi f. acutangulus isthe most common asellid, but P. remyi f. remyi, P. arnautovici elongatus and P.gjorgjevici f. typicus can be found there as well. The sublittoral bottom is mainlycovered by finer sands mixed with mud and detritus and the continuous mollusc shellzone is observed there. Besides, the population of adult individuals of Dreissena sp.has its maximum density in the sublittoral zone. The deposits of shells serve as amicrohabitat for sublittoral Asellidae, especially for P. remyi f. acutangulus (STAN-KOVIÈ 1960). The profundal zone is mainly taken by P. gjorgjevici f. typicus but P.remyi f. nudus and P. arnautovici elongatus could be also found there, although theyare rather rare. The deepest zone of Lake Ohrid is characterised by constancy of

24 A. Kilikowska, A. Wysocka, J. Sell, M. Ziêtara and T. Sywula
ecological factors. The stagnation of water is almost complete, there is total lack oflight, and the oscillation of temperature does not exceed 0.6°C. The bottom is cov-ered by mud (STANKOVIÈ 1960).
A very high morphological variation among morphological forms of Proasel-lus in Lake Ohrid raises a question whether all these differences are only a pheno-typic response to different life conditions or are the results of a reproductive isola-tion and divergent selection between environments and niches.
Two alternative hypotheses may be proposed to explain these high morpho-logical variations among endemic morphological forms of Proasellus in Lake Ohrid:(1) the different forms result from speciation events and are reproductively isolatedfrom each other, (2) the different forms are an effect of the high morphological plas-ticity of each species and are not genetically isolated. The aim of this study was toevaluate the above scenarios. Two polymorphic enzymes, glucose-phosphate isomer-ase (GPI) and phosphoglucomutase (PGI) were selected as nuclear genetic markersto gain insight into the issue of reproductive isolation.
MATERIAL AND METHODS
Samples were collected in August 2003 and 2004 from the littoral, sublittoraland profundal zones of the lake (along the line Ohrid � Radozhda) (Fig. 1, Table 1)and identified to species and form. Only four specimens of P. arnautovici elongatus
Proasellus species Subspecies/form Sampling station Sample
size
P. r. f. remyi N St. Naum spring; (depth 1 m), under stones 165
P. r. f. remyi L
Lake Ohrid; littoral (depth 1−15 m), under stones, on Chara sp., on shells of live zebra mussels; sublittoral (depth 20−30 m), empty shells of zebra mussels
233
P. r. f. acutangulus Lake Ohrid; sublittoral (depth 20−30 m; 40−50 m); empty shells of zebra mussels
202
P. remyi
P. r. f. nudus Lake Ohrid; profundal (depth 60−70 m; 90−100 m); muddy bottom
96
P. a. arnautovici Lake Ohrid; littoral (depth 10−15 m), under stones, on Chara sp., on shells of live zebra mussels
80
P. arnautovici
P. a. elongatus Lake Ohrid; sublittoral (depth 40−50 m), sandy bottom
4
P. g. f. litoralis Lake Ohrid; littoral (depth 10−15 m), under stones, on Chara sp., on shells of live zebra mussels
98
P. gjorgjevici
P. g. f. typicus Lake Ohrid; profundal (depth 90−100 m), muddy bottom
133
Table 1. Sampling stations, depth, type of bottom, and sample size (i.e. number of specimens) of studypopulations

25PROASELLUS SPECIES GROUP FROM LAKE OHRID
were collected from the profundal, therefore this form was excluded from furtheranalysis. All forms were distinguished on the basis of the description given by KA-RAMAN (1953).
The specimens for the study were taken from under the stones by hand or fromthe deeper part of the lake by a dredge. The number of specimens in sample, depthand type of bottom are presented in Table 1. The mean sample size was 126; indi-vidual sample size varied according to the abundance of particular forms. The ani-mals were brought alive to the laboratory, where they were identified and immedi-ately frozen in liquid nitrogen. Next, they were transferred to Poland, where theywere stored at -80°C until allozyme analyses.
For the allozyme analyses, specimens were homogenized in a grinding solu-tion (1 mg/ml NADP, 1 µl/ml $-mercaptoethanol, 1 µl/ml Triton X-100 in H
2O) (RI-
CHARDSON et al. 1986). Small animals were homogenized whole, while larger ani-mals were halved.
Cellulose acetate electrophoresis was applied to study the glucose-phosphateisomerase (GPI, EC 5.3.1.9) locus Gpi and the phosphoglucomutase (PGM, EC2.7.5.1) locus Pgm.
The procedures and staining methods by HEBERT and BEATON (1989) provedto be useful in case of all species studied. Each specimen was examined with re-spect to both enzyme systems. Loci and alleles were designated according to thedecreasing electrophoretic mobilities of the corresponding proteins. The identity ofalleles (mobility classes) was assessed by direct comparison. BIOSYS-1 software(SWOFFORD & SELANDER 1989) was used for the statistical analysis of the electro-phoretic data. An allele was regarded as rare when its frequency was lower than 0.05(CHAKRABORTY et al. 1980). Contingency P2 analysis at both loci was performed.Representative samples of each morphological form were tested for conformity withHardy-Weinberg expectations by using the P2 test with pooling of genotypes for qualityof fit. The agreement with Hardy-Weinberg expectations was also tested by meansof the goodness-of-fit P2 test for combined populations: (1) according to species rank(a = P. arnautovici; b = P. remyi; c = P. gjorgjevici); (2) according to the vertical zoneof the lake (a = littoral populations: P. arnautovici arnautovici, P. remyi f. remyi andP. gjorgjevici f. litoralis; b = sublittoral populations: P. remyi f. remyi and P. remyi f.acutangulus; c = profundal populations: P. remyi f. nudus and P. gjorgjevici f. typi-cus); (3) all taxa together. WRIGHT�s (1978) F-statistics were used to determine thedegree of inbreeding within the combined populations.
RESULTS
The obtained electrophoretic patterns are in accordance with the model predict-ed for dimeric proteins in the case of GPI and monomeric proteins in the case of PGM,with 5 codominant alleles at both loci. A total of 9 electrophoretic phenotypes of GPIwere identified in the populations screened. PGM gels revealed 7 electrophoretic phe-notypes with a variably weak anodal secondary band. The analysed enzyme systemswere coded by several loci but only one could be genetically interpreted.
The allele frequencies and P2 values connected with Hardy-Weinberg expecta-tions for each of the examined populations are presented in Table 2. Not all of the 2electrophoretic phenotypes could be interpreted genetically in some individuals and
26 A. Kilikowska, A. Wysocka, J. Sell, M. Ziêtara and T. Sywula
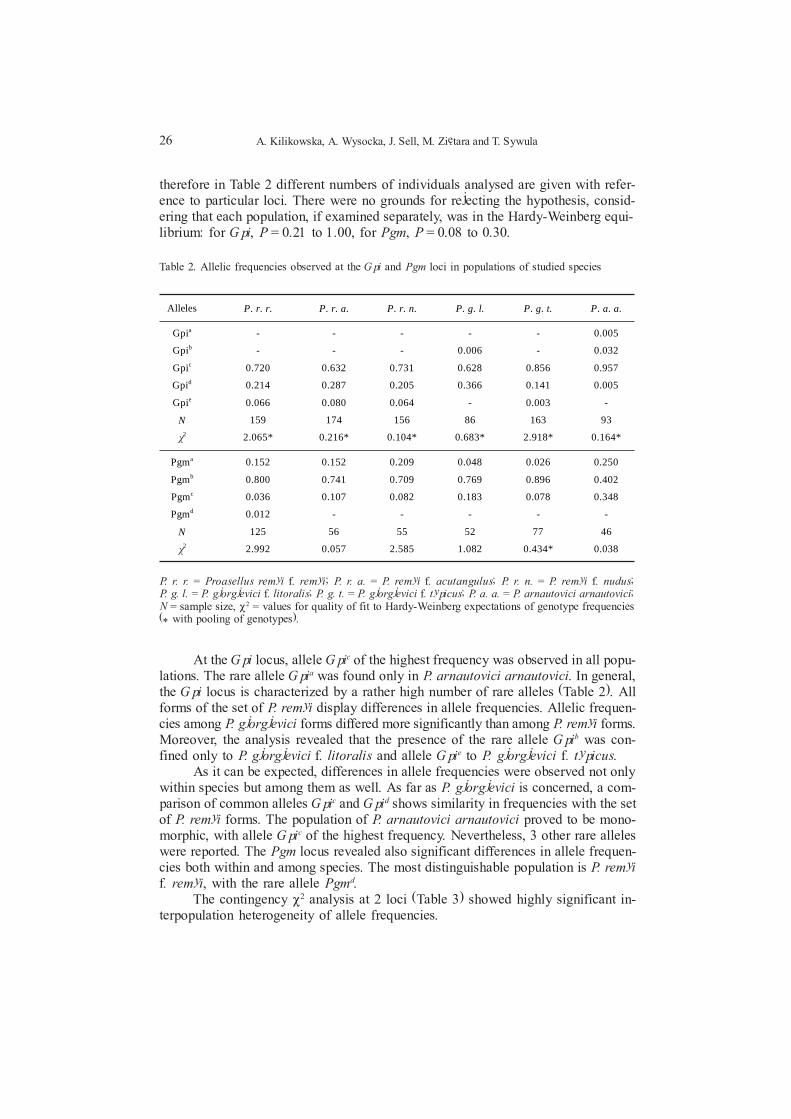
therefore in Table 2 different numbers of individuals analysed are given with refer-ence to particular loci. There were no grounds for rejecting the hypothesis, consid-ering that each population, if examined separately, was in the Hardy-Weinberg equi-librium: for Gpi, P = 0.21 to 1.00, for Pgm, P = 0.08 to 0.30.
Alleles P. r. r. P. r. a. P. r. n. P. g. l. P. g. t. P. a. a.
Gpia - - - - - 0.005
Gpib - - - 0.006 - 0.032
Gpic 0.720 0.632 0.731 0.628 0.856 0.957
Gpid 0.214 0.287 0.205 0.366 0.141 0.005
Gpie 0.066 0.080 0.064 - 0.003 -
N 159 174 156 86 163 93
÷2 2.065* 0.216* 0.104* 0.683* 2.918* 0.164*
Pgma 0.152 0.152 0.209 0.048 0.026 0.250
Pgmb 0.800 0.741 0.709 0.769 0.896 0.402
Pgmc 0.036 0.107 0.082 0.183 0.078 0.348
Pgmd 0.012 - - - - -
N 125 56 55 52 77 46
÷2 2.992 0.057 2.585 1.082 0.434* 0.038
At the Gpi locus, allele Gpic of the highest frequency was observed in all popu-lations. The rare allele Gpia was found only in P. arnautovici arnautovici. In general,the Gpi locus is characterized by a rather high number of rare alleles (Table 2). Allforms of the set of P. remyi display differences in allele frequencies. Allelic frequen-cies among P. gjorgjevici forms differed more significantly than among P. remyi forms.Moreover, the analysis revealed that the presence of the rare allele Gpib was con-fined only to P. gjorgjevici f. litoralis and allele Gpie to P. gjorgjevici f. typicus.
As it can be expected, differences in allele frequencies were observed not onlywithin species but among them as well. As far as P. gjorgjevici is concerned, a com-parison of common alleles Gpic and Gpid shows similarity in frequencies with the setof P. remyi forms. The population of P. arnautovici arnautovici proved to be mono-morphic, with allele Gpic of the highest frequency. Nevertheless, 3 other rare alleleswere reported. The Pgm locus revealed also significant differences in allele frequen-cies both within and among species. The most distinguishable population is P. remyif. remyi, with the rare allele Pgmd.
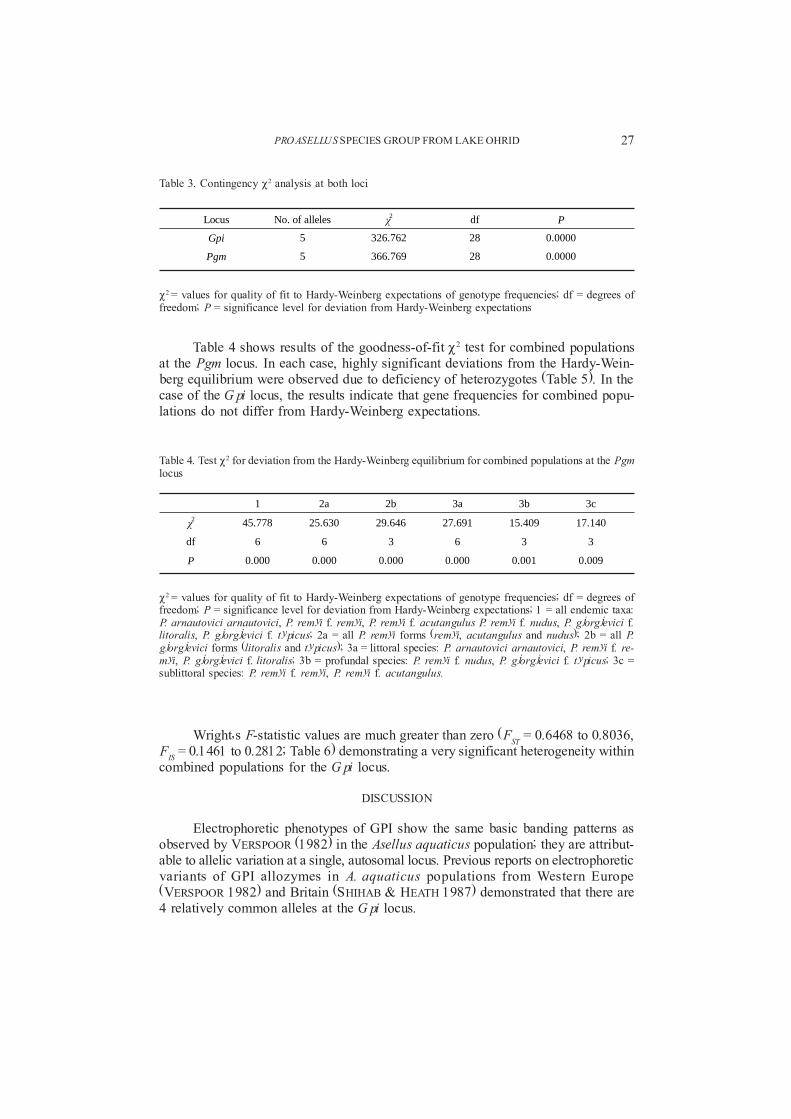
The contingency P2 analysis at 2 loci (Table 3) showed highly significant in-terpopulation heterogeneity of allele frequencies.
Table 2. Allelic frequencies observed at the Gpi and Pgm loci in populations of studied species
P. r. r. = Proasellus remyi f. remyi; P. r. a. = P. remyi f. acutangulus; P. r. n. = P. remyi f. nudus;P. g. l. = P. gjorgjevici f. litoralis; P. g. t. = P. gjorgjevici f. typicus; P. a. a. = P. arnautovici arnautovici;N = sample size, P2 = values for quality of fit to Hardy-Weinberg expectations of genotype frequencies(* with pooling of genotypes).
27PROASELLUS SPECIES GROUP FROM LAKE OHRID
Locus No. of alleles ÷2 df P
Gpi 5 326.762 28 0.0000
Pgm 5 366.769 28 0.0000
P2 = values for quality of fit to Hardy-Weinberg expectations of genotype frequencies; df = degrees offreedom; P = significance level for deviation from Hardy-Weinberg expectations; 1 = all endemic taxa:P. arnautovici arnautovici, P. remyi f. remyi, P. remyi f. acutangulus P. remyi f. nudus, P. gjorgjevici f.litoralis, P. gjorgjevici f. typicus; 2a = all P. remyi forms (remyi, acutangulus and nudus); 2b = all P.gjorgjevici forms (litoralis and typicus); 3a = littoral species: P. arnautovici arnautovici, P. remyi f. re-myi, P. gjorgjevici f. litoralis; 3b = profundal species: P. remyi f. nudus, P. gjorgjevici f. typicus; 3c =sublittoral species: P. remyi f. remyi, P. remyi f. acutangulus.
Table 3. Contingency P2 analysis at both loci
P2 = values for quality of fit to Hardy-Weinberg expectations of genotype frequencies; df = degrees offreedom; P = significance level for deviation from Hardy-Weinberg expectations
Table 4. Test P2 for deviation from the Hardy-Weinberg equilibrium for combined populations at the Pgmlocus
Table 4 shows results of the goodness-of-fit P2 test for combined populationsat the Pgm locus. In each case, highly significant deviations from the Hardy-Wein-berg equilibrium were observed due to deficiency of heterozygotes (Table 5). In thecase of the Gpi locus, the results indicate that gene frequencies for combined popu-lations do not differ from Hardy-Weinberg expectations.
1 2a 2b 3a 3b 3c
÷2 45.778 25.630 29.646 27.691 15.409 17.140
df 6 6 3 6 3 3
P 0.000 0.000 0.000 0.000 0.001 0.009
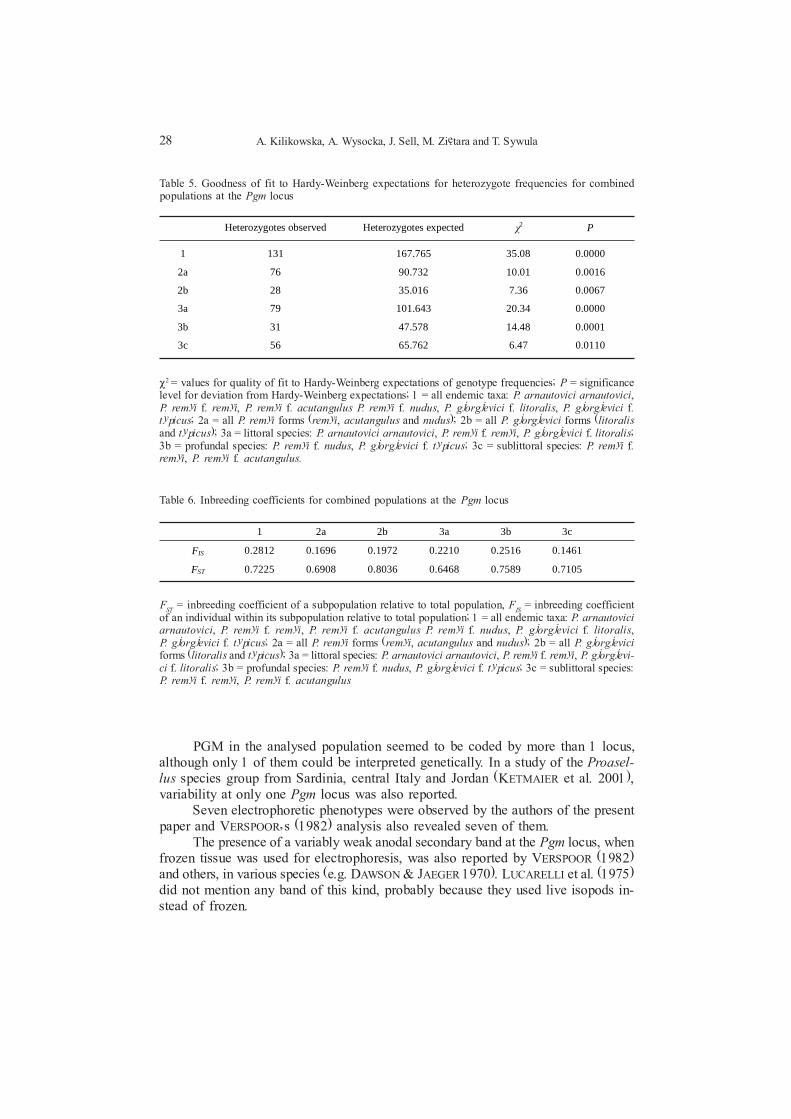
Wright�s F-statistic values are much greater than zero (FST
= 0.6468 to 0.8036,F
IS = 0.1461 to 0.2812; Table 6) demonstrating a very significant heterogeneity within
combined populations for the Gpi locus.
DISCUSSION
Electrophoretic phenotypes of GPI show the same basic banding patterns asobserved by VERSPOOR (1982) in the Asellus aquaticus population; they are attribut-able to allelic variation at a single, autosomal locus. Previous reports on electrophoreticvariants of GPI allozymes in A. aquaticus populations from Western Europe(VERSPOOR 1982) and Britain (SHIHAB & HEATH 1987) demonstrated that there are4 relatively common alleles at the Gpi locus.
28 A. Kilikowska, A. Wysocka, J. Sell, M. Ziêtara and T. Sywula
PGM in the analysed population seemed to be coded by more than 1 locus,although only 1 of them could be interpreted genetically. In a study of the Proasel-lus species group from Sardinia, central Italy and Jordan (KETMAIER et al. 2001),variability at only one Pgm locus was also reported.
Seven electrophoretic phenotypes were observed by the authors of the presentpaper and VERSPOOR�s (1982) analysis also revealed seven of them.
The presence of a variably weak anodal secondary band at the Pgm locus, whenfrozen tissue was used for electrophoresis, was also reported by VERSPOOR (1982)and others, in various species (e.g. DAWSON & JAEGER 1970). LUCARELLI et al. (1975)did not mention any band of this kind, probably because they used live isopods in-stead of frozen.
Heterozygotes observed Heterozygotes expected ÷2 P
1 131 167.765 35.08 0.0000
2a 76 90.732 10.01 0.0016
2b 28 35.016 7.36 0.0067
3a 79 101.643 20.34 0.0000
3b 31 47.578 14.48 0.0001
3c 56 65.762 6.47 0.0110
P2 = values for quality of fit to Hardy-Weinberg expectations of genotype frequencies; P = significancelevel for deviation from Hardy-Weinberg expectations; 1 = all endemic taxa: P. arnautovici arnautovici,P. remyi f. remyi, P. remyi f. acutangulus P. remyi f. nudus, P. gjorgjevici f. litoralis, P. gjorgjevici f.typicus; 2a = all P. remyi forms (remyi, acutangulus and nudus); 2b = all P. gjorgjevici forms (litoralisand typicus); 3a = littoral species: P. arnautovici arnautovici, P. remyi f. remyi, P. gjorgjevici f. litoralis;3b = profundal species: P. remyi f. nudus, P. gjorgjevici f. typicus; 3c = sublittoral species: P. remyi f.remyi, P. remyi f. acutangulus.
Table 5. Goodness of fit to Hardy-Weinberg expectations for heterozygote frequencies for combinedpopulations at the Pgm locus
1 2a 2b 3a 3b 3c
FIS 0.2812 0.1696 0.1972 0.2210 0.2516 0.1461
FST 0.7225 0.6908 0.8036 0.6468 0.7589 0.7105
FST
= inbreeding coefficient of a subpopulation relative to total population, FIS = inbreeding coefficient
of an individual within its subpopulation relative to total population; 1 = all endemic taxa: P. arnautoviciarnautovici, P. remyi f. remyi, P. remyi f. acutangulus P. remyi f. nudus, P. gjorgjevici f. litoralis,P. gjorgjevici f. typicus; 2a = all P. remyi forms (remyi, acutangulus and nudus); 2b = all P. gjorgjeviciforms (litoralis and typicus); 3a = littoral species: P. arnautovici arnautovici, P. remyi f. remyi, P. gjorgjevi-ci f. litoralis; 3b = profundal species: P. remyi f. nudus, P. gjorgjevici f. typicus; 3c = sublittoral species:P. remyi f. remyi, P. remyi f. acutangulus
Table 6. Inbreeding coefficients for combined populations at the Pgm locus

29PROASELLUS SPECIES GROUP FROM LAKE OHRID
An analysis of goodness of fit to the Hardy-Weinberg equilibrium in separatepopulations reveals that the genotypic frequencies at both loci confirm the expecta-tions. These results indicate that the mating is random within each population of P.arnautovici arnautovici, P. remyi f. remyi, P. gjorgjevici f. litoralis, P. remyi f. nudus,P. gjorgjevici f. typicus, and P. remyi f. acutangulus.
The allelic composition of the populations demonstrates that there are differ-ences in allele frequencies both within and among species. It was confirmed by thecontingency P2 analysis at 2 loci (Table 3).
Differences in allele frequencies among P. gjorgjevici forms are higher than inthe case of P. remyi. Moreover, at the Gpi locus the presence of rare alleles (Gpib
and Gpie) was confined to either the littoral or the profundal form of P. gjorgjevici.Higher genetic differences among P. gjorgjevici forms seem to follow KARAMAN�s(1953) conclusions drawn on the basis of morphological characters and the highestabundance of P. gjorgjevici in the profundal, that this species is the oldest one amongthe asellids in Lake Ohrid. Moreover, P. gjorgjevici f. typicus seems to be the bestadapted to living conditions in the profundal zone. Its body is depigmented, pale pinkor yellowish. The degree of eye reduction is higher than in other profundal inhabit-ants: P. remyi f. nudus and P. arnautovici elongatus. The body and appendages aresignificantly longer than in the littoral form. According to morphological analysis, itseems that profundal populations of isopods differentiated from littoral forms in thelake (KARAMAN 1953, STANKOVIÈ 1960). The highest value of Wright�s F-statistics(F
ST = 0.8036) in the combined population of both forms: P. gjorgjevici f. litoralis
and P. gjorgjevici f. typicus, may also support this idea. This result indicates subdi-vision of the combined population and limited gene flow between those taxa.
Less significant differences in allele frequencies among P. remyi forms inhabit-ing the lake may reflect a shorter evolutionary history. Besides, some individualsof mixed morphological characters of P. remyi f. remyi and P. remyi f. acutanguluswere found by KARAMAN (1953) as well as during our study. Although we have noknowledge of viability or sterility of such specimens, the disequilibrium in Hardy-Weinberg expectations in combined populations seems to confirm that also betweenthese forms some efficient reproductive barriers, which prevent effective hybridiza-tion, should exist. No hybrids between the other species forms were found.
On the basis of characteristics of pleopods IV, KARAMAN (1953) assumed thatP. remyi and P. arnautovici arnautovici are more closely related to each other thanto P. gjorgjevici. Differences in allele frequencies at both loci, and the presence ofprivate rare alleles at the Gpi locus do not show any such relationship. P. arnauto-vici arnautovici gave an impression of being relatively distinct from the rest of theendemic species. However, at present, the number of loci analysed is not sufficientto draw any definite conclusions.
The assumption that there is no reproductive isolation within the analysedcombined populations was tested by calculating goodness of fit to the Hardy-Wein-berg equilibrium for each locus. In the case of the Gpi locus, the results indicate thatgene frequencies for combined populations do not differ from Hardy-Weinberg ex-pectations. This may be due to the fact that this locus was reported to be under se-lection (SHIHAB & HEATH 1987, CÁRDENAS et al. 2001, WATT et al. 2003).

30 A. Kilikowska, A. Wysocka, J. Sell, M. Ziêtara and T. Sywula
At the Pgm locus, all combined populations were out of equilibrium due toheterozygote deficiency (Table 4). The deficit could result from a subdivision of onepopulation into isolated and differentiated reproductive units (Wahlund effect) orselection. Besides, inbreeding may contribute to the deficit in heterozygotes (WRIGHT
1978).The results are not unexpected in the case of fusing all analysed populations
into one (Table 3). This indicates that they could represent reproductively isolatedunits in sympatry. The heterozygote deficiency seems to be due to the Wahlund ef-fect occurring when sympatric populations with limited genetic interchange aresampled as a single population. When populations were combined according to thehabitat shared, the results were similar (Table 3). High values of F
ST and F
IS(Table 5) in both cases may indicate a high level of subdivision within the combinedpopulations, restriction of gene flow and inbreeding within the real populations. It isinteresting that even within the sublittoral group consisting of two P. remyi forms(remyi and acutangulus), which seem to be relatively close to one another, the in-breeding coefficient F
IS (F
IS = 0.1461) shows the lowest value of all combined popu-
lations.The deviations from the Hardy-Weinberg equilibrium (Table 3) caused by het-
erozygote deficiency (Table 4) and very high values of F-statistics (Table 5) for fusedpopulations of the nominate species are very intriguing. There could also be otherpossibilities beside those mentioned above, which can account for this deficit, e.g.incorrect scoring of polymorphism, X-lineage, presence of null alleles, or non-ran-dom sampling. However, because of the fact that each population fits the Hardy-Weinberg equilibrium when analysed separately, and that there were no differencesin genotype frequencies between sexes, such explanations do not apply to our study.So, the heterozygote deficiency seems also to be caused by the Wahlund effect. Veryhigh values of F
ST (Table 5) demonstrate that there is very little gene flow between
the 6 taxa despite the fact that officially they are believed to belong to only 3 spe-cies. Asellids have limited swimming abilities and therefore are regarded as slowcolonizers of new habitats, even within a lake scale, and that is supposed to limittheir potential for adaptive radiation (HARGEBY 1990, HIDDING et al. 2003,HARGEBY et al. 2004). Besides, isopods lack the larval stage that usually accountsfor a high rate of dispersal (WETZER 2001). Thus, the migration rate of particularforms is probably limited, especially that they spend their lives flattened against stonesor empty shells of the zebra mussels. This may contribute to a high level of inbreed-ing (F
IS;
Table 5) that also results in heterozygote deficiency. In the case of our study,
different selection pressures are also likely to play a significant role in the divergenceamong populations, since they live in different types of habitat (the littoral, sublitto-ral and profundal zones of the very deep lake).
According to the biological species concept of MAYR (1963), 2 populationsmay be considered as biological species only when reproductive isolation is com-plete. In our study, it is difficult to say whether the isolation is complete, especiallyin the case of P. remyi f. remyi and P. remyi f. acutangulus where some individualsof mixed morphological characters were observed. Some mating experiments shouldbe carried out to throw light on the matter. Still, according to the latter definition ofspecies (BIGELOW 1965, MAYR 1982), the isolation of gene pools of different spe-

31PROASELLUS SPECIES GROUP FROM LAKE OHRID
cies does not need to be complete as long as they �remain reproductively isolated inthe sense that they do not fuse into a single population� (MAYR 1982, p. 298). Thiscase appears to apply to the examined populations.
To sum up, it seems that the observed patterns of morphological differentia-tion within examined species are supported by genetic data and this is a process ofintralacustrine speciation based on the mechanism of ecological isolation by habitat(STANKOVIÈ 1960) but its rate, manner and detailed mechanisms should be moreclosely established. Although the present data suggest congruence of morphologicaldifferences with genetic differentiation, a further study is necessary to give a betterpicture of the population structure and phylogenetic relationships among and withinspecies.
REFERENCES
BIGELOW R. S. 1965. Hybrid zones and reproductive isolation. Evolution 19: 449�458.CÁRDENAS E., MUNSTERMANN L. E., MARTÍNEZ O., CORREDOR D., FERRO C. 2001. Genetic vari-
ability among populations of Lutzomyia (Psathyromyia) shannoni (Dyar 1929) (Diptera: Psy-chodidae: Phlebotominae) in Colombia. Mem. Inst. Oswaldo Cruz, Rio de Janeiro 96: 189�196.
CHAKRABORTY R., FUERST P. A., NEI M. 1980. Statistical studies on protein polymorphism in naturalpopulations. III. Distribution of allele frequencies and the number alleles per locus. Genetics94: 1039�1063.
DAWSON D. M., JAEGER S. 1970. Heterogeneity of phosphoglucomutase. Biochem. Genet. 4: 1.HARGEBY A. 1990. Macrophyte associated invertebrates and effect of habitat permanence. Oikos
57: 338�346.HARGEBY A., JOHANSSON J., AHNESJO J. 2004. Habitat-specific pigmentation in a freshwater
Isopod: Adaptive evolution over a small spatiotemporal scale. Evolution 58: 81�94.HEBERT P. D. N., BEATON M. J. 1989. Methodologies for alloenzyme analysis using cellulose ac-
etate electrophoresis. Helena Lab., Beaumont.HIDDING B., MICHEL E., NATAYAGANOVA A. V., SHERBAKOV D. Y. 2003. Molecular evidence re-
veals a polyphyletic origin and chromosomal speciation of Lake Baikal�s endemic asellid iso-pods. Mol. Ecol. 12: 1509�1514.
KARAMAN S. 1953. Die Aselliden des Ohridsees. Period. Biol. Soc. Sci. Nat. Croat. Zagreb. Ser.II/B 4�6: 46�76.
KETMAIER V., ARGANO R. COBOLLI M., DE MATTHAEIS E., MESSANA G. 2001. A systematic andbiogeographical study of epi- and hypogean populations of Proasellus species group fromSardinia, central Italy and Jordan: allozyme insights. J. Zool. Sys. Evol. Res. 9: 51�61.
KRSTANOVSKI Z. 1994. Biosystematical and ecological investigations of Planarians in Lakes Ohrid,Prespa and Dojran with their costal waters. PhD Thesis. Sts. Cyril and Methodius University,Skopje (in Macedonian).
KRSTANOVSKI Z., TRAJANOVSKI S. 2002. Lake Ohrid bottom fauna. The benthic fauna of Lake Ohrid.In: Limnological Investigations of Lake Ohrid (MITIC V., Ed.), pp. 109�128, Hydrobiologi-cal Institute, Ohrid.
LUCARELLI P., FANO E. A., VITAGLIANO G. T. 1975. Existence of phosphoglucomutase enzymepolymorphism in Asellus aqaticus (L.) (Crustacea, Isopoda). Rendiconti 584: 647.
MACDONALD III K. S., YAMAPOLSKY L., DUFFY E.. 2005. Molecular and morphological evolutionof the amphipod radiation of Lake Baikal. Mol. Phylogenet. Evol. 35: 323�343.
MARTENS K. 1997. Speciation in ancient lakes. Trends Ecol. Evol. 12: 5.MARTENS K., SCHÖN I. 1999. Crustacean biodiversity in ancient lakes: A review. Crustaceana 72:
899�910.MAYR E. 1963. Animal species and evolution. Harvard University Press, Cambridge, Massachu-
setts.

32 A. Kilikowska, A. Wysocka, J. Sell, M. Ziêtara and T. Sywula
MAYR E. 1982. The growth of biological thought: diversity, evolution and inheritance. Belknap,Cambridge, Massachusetts.
RICHARDSON B. J., BAVERSTOCK P. R., ADAMS M. 1986. Allozyme electrophoresis: A Handbookfor Animal and Systematics and Population Studies. Academic Press, Sydney.
SALEMAA H., KAMALTYNOW R. 1994. Chromosomal relationships of the endemic Amphipoda(Crustacea) in the ancient lakes Ohrid and Baikal. In: Genetic and Evolution of Aquatic Or-ganisms (BEAUMONT A. R., Ed.), pp. 405�414, Chapman & Hall, London.
SHERBAKOV D. YU. 1999. Molecular phylogenetic studies on the origin of biodiversity in LakeBaikal. Trends Ecol. Evol. 14: 92�95.
SHERBAKOV D. YU., KAMALTYNOV R. M., OGARKOV O. B., VÄINÕLÄ R., VAINIO J. K., VERHEY-EN E. 1999. On the phylogeny of Lake Baikal amphipods in the light of mitochondrial andnuclear DNA sequence data. Crustaceana 72: 911�919.
SHIHAB A. F., HEATH D. J. 1987. Components of fitness and the PGI polymorphism in the fresh-water isopod Asellus aquaticus (L.). 1. Fecundity selection. Heredity 58: 69�73.
SPIRKOVSKI Z., TALEVSKI T., STOJANOVSKI T., VELKOVA-JORDANOSKA L. 2002. Lake Ohrid fish.In: Limnological Investigations of Lake Ohrid (MITIC V., Ed.), pp. 130�214, Hydrobiologi-cal Institute, Ohrid.
STANKOVIÈ S. 1960. The Balkan Lake Ohrid and its living world. Uitgeverij Dr. W. Junk, Den Haag.SWOFFORD D. L., SELANDER R. B. 1989. BIOSYS 1, release 1.7. University of Illinois Press, Urbana,
IL.VÄINÖLÄ R., KAMALTYNOV R. M. 1999. Species diversity and speciation in the endemic amphi-
pods of Lake Baikal: Molecular evidence. Crustaceana 72: 945�956.VERSPOOR E. 1982. Description and Genetics of Glucose Phosphate Isomerase (GPI) and Phos-
phoglucomutase (PGM) polymorphism in Asellus aquaticus (L.). Biochem. Genet. 20: 891�905.
WATT W. B., WHEAT C. W., MEYER E. H., MARTIN J. F. 2003. Adaptation at specific loci. VII.Natural selection, dispersal and the diversity of molecular�functional variation patterns amongbutterfly species complexes (Colias: Lepidoptera, Pieridae). Mol. Ecol. 12: 1265�1275.
WETZER R. 2001. Hierarchical analysis of mtDNA variation and the use of mtDNA for isopod(Crustacea: Peracarida: Isopoda) systematics. Contrib. Zool. 70: 23�29.
WRIGHT S. 1978. Evolution and genetics of populations. Volume 2. The theory of gene frequen-cies. University of Chicago Press, Chicago.





























