Embed Size (px)
Citation preview

The novel putative transporter NPT1 plays a critical role inearly stages of Plasmodium berghei sexual developmentmmi_7767 1343..1357
Bertrand Boisson,1*† Céline Lacroix,1
Emmanuel Bischoff,2 Pascale Gueirard,1‡
Daniel Y. Bargieri,1‡ Blandine Franke-Fayard,3
Chris J. Janse,3 Robert Ménard1 andPatricia Baldacci1**1Institut Pasteur, Biologie et Génétique du Paludisme,75724 Paris, France.2Institut Pasteur, Plateforme Puces à ADN Génopole,75724 Paris, France.3Leiden Malaria Research Group, Department ofParasitology, Leiden University Medical Centre, Leiden,The Netherlands.
Summary
Transmission of Plasmodium species from a mamma-lian host to the mosquito vector requires the uptake,during an infected blood meal, of gametocytes, theprecursor cells of the gametes. Relatively little isknown about the molecular mechanisms involved inthe developmental switch from asexual developmentto sexual differentiation or the maturation and sur-vival of gametocytes. Here, we show that a genecoding for a novel putative transporter, NPT1, playsa crucial role in the development of Plasmodiumberghei gametocytes. Parasites lacking NPT1 areseverely compromised in the production of gameto-cytes and the rare gametocytes produced are unableto differentiate into fertile gametes. This is the earliestblock in gametocytogenesis obtained by reversegenetics and the first to demonstrate the role of aprotein with a putative transport function in sexualdevelopment. These results and the high degreeof conservation of NPT1 in Plasmodium speciessuggest that this protein could be an attractive targetfor the development of novel drugs to block thespread of malaria.
Introduction
The Plasmodium parasite is the causative agent ofmalaria, a disease that kills approximately 780 000 peopleper year (WHO, 2010). Whereas the pathology of malariais caused by the asexual replication cycles of Plasmodiumin erythrocytes, sexual reproduction of the parasite takesplace in a mosquito vector. Mature female and malegametocytes, the precursor cells of the gametes, circulatein the blood inside erythrocytes and are taken up by amosquito during a bite. In the mosquito gut, the gameto-cytes rapidly exit erythrocytes and transform intogametes. The gametes then fertilize, giving rise to a motilezygote, the ookinete, which actively penetrates the mos-quito midgut epithelium and transforms into an oocyst.Thousands of sporozoites are formed inside an oocyst,which migrate to the salivary glands and are transmittedduring a blood meal to a new host. The few parasitestransmitted by the mosquito first multiply inside hepato-cytes, where erythrocyte-infective forms of the parasiteare generated.
Gametocytogenesis, the developmental pathwayleading to the production of mature gametocytes, firstrequires a developmental switch (known as commitment)from the asexual to sexual cycle, followed by severalstages of maturation of the gametocytes (Lobo andKumar, 1998). In P. falciparum, the Plasmodium speciesmost lethal to humans, it has been shown that this switchoccurs in the cycle before gametocytes are observed.Moreover, all the merozoites released from a singleschizont are either asexual or sexual (Bruce et al., 1990;Talman et al., 2004), and in the latter case produce eithermale or female gametocytes (Silvestrini et al., 2000;Smith et al., 2000).
In P. falciparum, gametocytogenesis takes several days(Thomson, 1914) and the different stages of gametocytematuration have been well defined by their morphologyand the expression of sexual-stage-specific antigens(Carter and Graves, 1988; Alano, 2007). Transcriptomeanalyses have shown that a cluster of about 200 genes isexpressed during sexual development (Silvestrini et al.,2005; Young et al., 2005). However, less than 10 genesare significantly upregulated in early P. falciparum game-tocytes, indicating that there may be only a few genesrequired for the initial stages of sexual development
Accepted 30 June, 2011. For correspondence. *E-mail [email protected]; Tel. (+33) 144389459; Fax (+33) 140613089. **[email protected]; Tel. (+33) 144389459; Fax (+33) 140613089.†Present address: Laboratory of Human Genetics of Infectious Dis-eases, Rockefeller University, New York, NY 10065, USA. ‡Theseauthors equally contributed to the study.
Molecular Microbiology (2011) 81(5), 1343–1357 � doi:10.1111/j.1365-2958.2011.07767.xFirst published online 4 August 2011
© 2011 Blackwell Publishing Ltd

(Silvestrini et al., 2005). Furthermore, the proteins fromonly six of these upregulated genes are known to beexpressed in early gametocytes: Pfs16, present in theparasitophorous vacuole membrane (PVM) (Bruce et al.,1994); Pfg27 (Carter et al., 1989); Pfmdv-1/peg3, presentin the parasite plasma membrane, PVM and cleft struc-tures (Furuya et al., 2005; Lanfrancotti et al., 2007);Pfg14.744, located in the parasitophorous vacuole andexported to the erythrocytic cytoplasm (Eksi et al., 2005);and Pfg14.748 and Pfpeg4, associated with the PVM(Eksi et al., 2005; Silvestrini et al., 2005; Alano, 2007).The precise functions of these proteins are still unknown.Gene disruption of Pfs16 resulted in a four to fivefoldreduction in the number of gametocytes, which despitehaving a normal appearance were unable to infect mos-quitoes (Kongkasuriyachai et al., 2004). An important rolefor Pfg27 in gametocytogenesis was initially reported(Lobo et al., 1999), however, a more recent study sug-gests that it is required for the maintenance of gameto-cytes (Olivieri et al., 2009). Gene inactivation of Pfmdv-1/peg3 caused a severe reduction (95%) in the numbers ofmature gametocytes due to a block in development atstage I. The mutant gametocytes presented abnormalmembrane structures and infection of mosquitoes wascompromised but not abolished (Furuya et al., 2005).
In P. berghei, a species that infects rodents, it has beenreported that mature gametocytes develop more rapidly(26–30 h) than in P. falciparum and that commitment tosexual development may occur 12–16 h after merozoiteinvasion (Mons, 1986). Morphologically, young gameto-cytes can only be distinguished from trophozoites at20 h and male gametocytes are detectable after 24 h. Aproteome analysis of P. berghei gametocytes identi-fied 236 male-specific and 101 female-specific proteins(Khan et al., 2005), most of which have orthologues inP. falciparum. However, there are very few markers forearly P. berghei gametocytes and, with the exception ofPfmdv-1/peg3, there are no obvious orthologues in theP. berghei genome of the genes expressed in earlyP. falciparum gametocytes. The inactivation of mdv-1/peg3 in P. berghei has been reported by two groups andresulted in parasites that produced significantly fewerzygotes, ookinetes and oocysts due to a reported role infemale gametocyte activation (Lal et al., 2009) or egressof both male and female gametes from the erythrocyte(Ponzi et al., 2009).
Although the molecular processes leading to sexualcommitment are poorly understood, the developmentalswitch from asexual to sexual development and sex deter-mination (i.e. female/male sex ratio) have been shown tobe sensitive to environmental stimuli, involving signallingmechanisms between the environment and the parasite(Paul et al., 2000; Drakeley et al., 2006; Dixon et al.,2008; Doerig et al., 2009). Analyses of the proteomes of
P. berghei (Khan et al., 2005) and P. falciparum gameto-cytes (Lasonder et al., 2002) identified several gameto-cyte specific kinases and phosphatases that might beinvolved in these pathways and one MAP kinase has beenshown to be specifically expressed in gametocytes (Dorinet al., 1999). Although several protein kinases (Billkeret al., 2004; Rangarajan et al., 2005; McRobert et al.,2008; Kato et al., 2009) and a phosphodiesterase (Tayloret al., 2008) have been shown to play a role in (cGMP)signalling that regulate gamete formation, the signallingcascades operating during gametocytogenesis per sehave yet to be characterized.
We have identified a P. berghei gene,PBANKA_020830, which shows a significant increase oftranscripts in liver stages. This gene is the orthologue of aP. falciparum gene, PFA0245w, encoding a protein with aputative transport function (Martin et al., 2005). By apply-ing reverse genetics, we examined the role of this novelputative transporter, named NPT1, during the life cycle ofP. berghei and found an early and essential role of thisprotein in Plasmodium sexual development.
Results
Identification, genomic organization and expression ofthe npt1 locus in P. berghei
We identified the gene PBANKA_020830 (previous iden-tifier PB001038.01.0) in a systematic screen based onan in silico and quantitative RT-PCR analysis of geneexpression (Ishino et al., 2009) as being more highlyexpressed in liver stages compared with sporozoites ofP. berghei. When expression of PBANKA_020830 wascompared in P. berghei mixed erythrocytic stages, puri-fied gametocytes, sporozoites collected from mosquitosalivary glands, and liver stages collected after 5, 17, 40and 50 h maturation inside HepG2 cells, the highestlevels of PBANKA_020830 mRNA were detected in liverstages (Fig. 1A).
The gene encodes a protein of 505 amino acids thatis predicted to contain 12 transmembrane segments(Figs 1B and S1). It is well conserved across Plasmodiumspecies with orthologues present in P. falciparum(PFA0245w), P. vivax (PVX_081515), P. knowlesi(PKH_020840) and two rodent malaria species, P.chabaudii (PCAS_020670) and P. yoelii (PY07333) (seeTable 1, Fig. 1C). PFA0245w was previously reportedto belong to an uncharacterized family of five novelputative transporters (NPTs) presenting some features ofthe major facilitating superfamily (MFS) (Martin et al.,2005). We thus renamed PBANKA_020830 as npt1.Using the HMM profile of the MFS obtained from thePfam database (PF07690), we performed an in silicoanalysis of 18 genomes from Apicomplexa and Ciliate
1344 B. Boisson et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 81, 1343–1357

organisms to identify all MFS members and to assesstheir phylogenetic relationship. This analysis (Fig. S2)showed that the five NPTs are present in all Plasmodiumspecies analysed and that the NPT family is notPlasmodium- or Apicomplexa-specific.
npt1 is located on chromosome 2 (Kooij et al., 2005) andits synteny is conserved with that of its P. falciparumorthologue, PFA0245w. In the P. falciparum genome,PFA0245w is located on chromosome 1, downstream ofPFA0240w, a paralogue encoding another member of the
NPT family (see Fig. 1D and Table 1). When this studywas initiated, the two P. berghei orthologous genes werelocated on separate contigs (RP1871 and RP1234,PlasmoDB 2005). We amplified by PCR and sequencedthe region between these contigs (GenBank # GU907667),which demonstrated the conservation of the genomic orga-nization in P. berghei and P. falciparum (Fig. 1D), asalso confirmed by the Wellcome Trust Sanger Institute(PBANKA_020830 and PBANKA_020840, the orthologueof PFA0240w, GeneDB). Furthermore, we prepared a
Fig. 1. Expression, phylogeny and genomic organization of Pbnpt1.A. Histogram representation of Pbnpt1 relative gene expression in P. berghei gametocytes (gam), sporozoites (spz), HepG2 cells (liver stage,LS) 5, 17, 40 and 50 h post infection and mixed blood stages (mBS) as determined by quantitative RT-PCR. The values were normalizedrelative to the expression of hsp70 mRNA in each sample. Error bars are SD; The asterisk indicates statistically significant compared with spz(P < 0.05).B. Schematic representation of the prediction of 12 transmembrane helices in NPT1 using TMHMM2.0. Predicted inside and outside regionsare depicted in blue and green, respectively.C. Phylogenetic tree of the NPT family of proteins belonging to the MFS in P. falciparum and P. berghei. Bootstrap values are indicated oneach branch.D. Schematic representation of the Pbnpt1 locus. Black arrows indicate the contigs RP1234 and RP1871. The dashed black line shows thesequence amplified between the two contigs. ORFs are indicated as thick arrows: npt1 (blue); PBANKA_020840, a close paralogue of npt1(pale blue); predicted ORFs (black). The 5′ and 3′ UTRs determined by RACE are shown as thin lines (green and red, respectively).
Table 1. Members of the novel putative transporter family identified in P. falciparum and P. berghei.
P. falciparum PFA0240w PFA0245w PFC0530w PFI0720w PF11_0310P. berghei orthologuea PBANKA_020840 PBANKA_020830 PBANKA_041050 PBANKA_081570 PBANKA_091830Identity 47 60 51 60 60Similarity 58 73 68 70 70Syntenic locationa Yes Yes Yes Yes YesChromosome P. bergheib chr 2 chr 2 chr 4 chr 8 chr 9
a. Data obtained from PlasmoDB.b. Data obtained from Kooij et al., 2005.
NPT1 is essential for Plasmodium gametocytogenesis 1345
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 81, 1343–1357
RACE library from total RNA of P. berghei NK65 sporozoi-tes isolated from mosquito salivary glands in order todetermine the transcription start and polyadenylation sitesof npt1 transcripts. The longest 5′ and 3′ UTRs isolatedwere 712 bp and 783 bp, respectively (see Fig. 1D).Finally, the 2.5 kb region upstream of the open readingframe (ORF) was analysed for the presence of sequencemotifs that might be involved in gene regulation. Two motifsthat have been described upstream of genes upregulatedin sexual development and sporozoites (Young et al.,2008) were found: ATATAGA, at position -1931 relative tothe npt1 translation start, and AAGACA, at positions -1324and -2087. Interestingly, alignment of the genomicsequences 2 kb after the npt1 ORF with CLUSTALW soft-ware revealed a highly conserved region with stretches ofGC in the 3′ UTR (between +950 and 1200) of all Plasmo-dium species (Fig. S3), suggesting that this region may befunctionally important.
Expression of npt1 was previously reported in Plas-modium pre-erythrocytic stages. A transcript of theP. yoelii orthologue, PY07333, was found in an ESTlibrary of salivary gland sporozoites (Kappe et al., 2001)and the protein was detected in the proteomic analysisof P. yoelii liver stages collected at 40 h maturity froman infected liver (Tarun et al., 2008). BLAST-n analysisof Pbnpt1 sequence against P. berghei EST librariesgave alignments with one EST from erythrocytic sch-izonts, two from 31 h liver stages, and one from ooki-netes. In P. falciparum, PFA0245w mRNA was found toincrease 5–10 times in sporozoites incubated with hepa-tocytes (Siau et al., 2008), while in erythrocytic stagesexpression was very low but increased 20 h post inva-sion and reached a maximum at 32 h, corresponding tothe trophozoite stage (Martin et al., 2005). PFA0245wmRNA levels were also reported to increase moderatelyfrom day 2 to day 12 during P. falciparum gametocyto-genesis (Young et al., 2005). Therefore, the availableexpression data, together with our qPCR analysis, indi-cate that npt1 transcription occurs in various stages ofthe Plasmodium life cycle, in both the vertebrate andmosquito hosts, with evidence for upregulation duringliver stages.
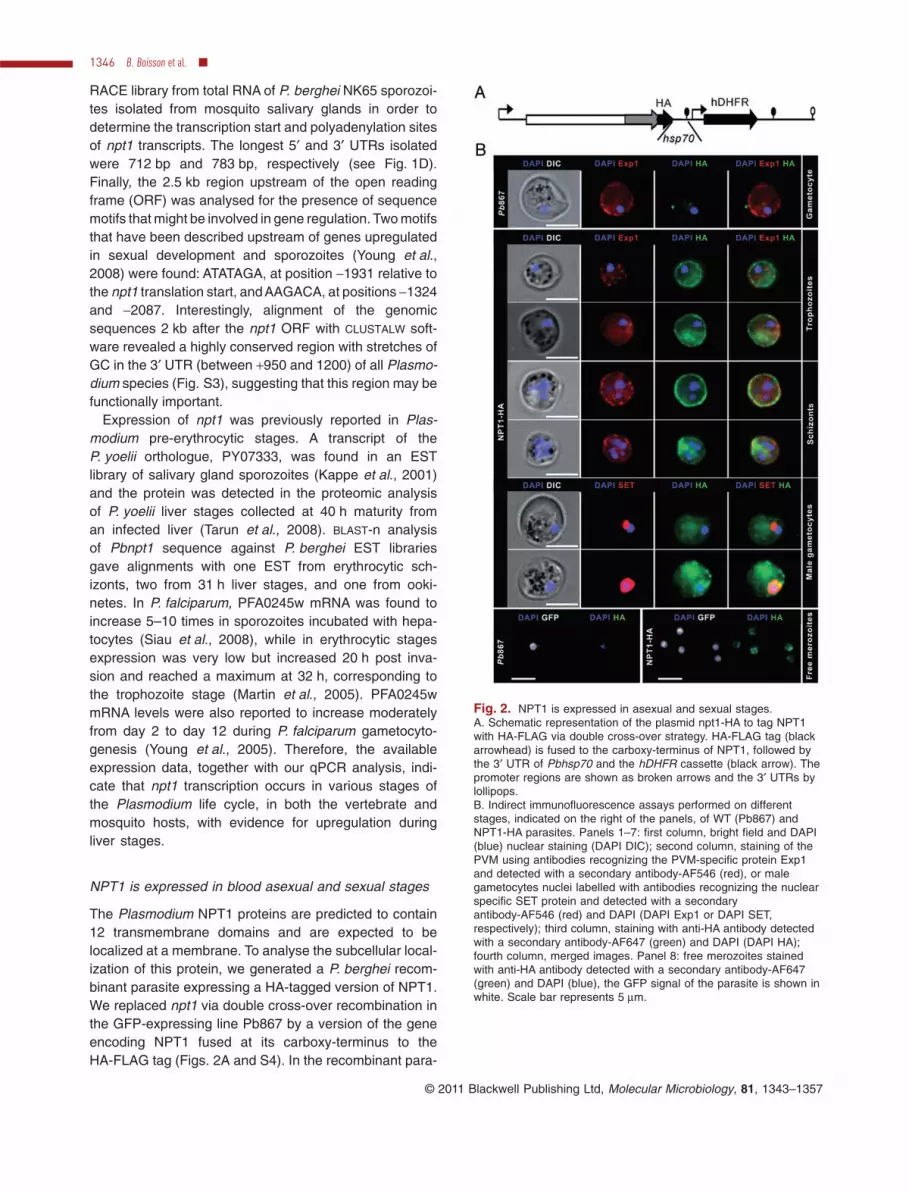
NPT1 is expressed in blood asexual and sexual stages
The Plasmodium NPT1 proteins are predicted to contain12 transmembrane domains and are expected to belocalized at a membrane. To analyse the subcellular local-ization of this protein, we generated a P. berghei recom-binant parasite expressing a HA-tagged version of NPT1.We replaced npt1 via double cross-over recombination inthe GFP-expressing line Pb867 by a version of the geneencoding NPT1 fused at its carboxy-terminus to theHA-FLAG tag (Figs. 2A and S4). In the recombinant para-
Fig. 2. NPT1 is expressed in asexual and sexual stages.A. Schematic representation of the plasmid npt1-HA to tag NPT1with HA-FLAG via double cross-over strategy. HA-FLAG tag (blackarrowhead) is fused to the carboxy-terminus of NPT1, followed bythe 3′ UTR of Pbhsp70 and the hDHFR cassette (black arrow). Thepromoter regions are shown as broken arrows and the 3′ UTRs bylollipops.B. Indirect immunofluorescence assays performed on differentstages, indicated on the right of the panels, of WT (Pb867) andNPT1-HA parasites. Panels 1–7: first column, bright field and DAPI(blue) nuclear staining (DAPI DIC); second column, staining of thePVM using antibodies recognizing the PVM-specific protein Exp1and detected with a secondary antibody-AF546 (red), or malegametocytes nuclei labelled with antibodies recognizing the nuclearspecific SET protein and detected with a secondaryantibody-AF546 (red) and DAPI (DAPI Exp1 or DAPI SET,respectively); third column, staining with anti-HA antibody detectedwith a secondary antibody-AF647 (green) and DAPI (DAPI HA);fourth column, merged images. Panel 8: free merozoites stainedwith anti-HA antibody detected with a secondary antibody-AF647(green) and DAPI (blue), the GFP signal of the parasite is shown inwhite. Scale bar represents 5 mm.
1346 B. Boisson et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 81, 1343–1357

site line, NPT1-HA, expression of the tagged gene wasdriven by the 5′ UTR of npt1 and the 3′UTR of hsp70. Thepresence of the HA-FLAG did not affect the function ofNPT1 during gametocytogenesis, as assessed by thepresence of gametocytes in blood smears of infectedmice or by the prevalence of oocysts in the midguts ofmosquitoes on days 7 and 14 post infection (NPT1-HA76 � 21% compared with Pb867 73 � 15%).
The expression and cellular localization of NPT1 wasinvestigated by indirect immunofluorescence on enrichedblood stages and purified schizonts of Pb867 andNPT1-HA parasites. As shown in Fig. 2B, NPT1-HA wasdetected in both asexual stages (panels 2–5) and sexualstages (panel 6–7). The absence of anti-HA labelling inthe blood stages of line Pb867 samples (panel 1) dem-onstrated the specificity of the antibody staining. NPT1was detected essentially at the periphery of the devel-oping trophozoites and schizonts. This distribution wassimilar to that of Exp-1, a membrane protein associatedwith the PVM (Gunther et al., 1991). In male gameto-cytes, identified by strong SET staining (Pace et al.,2006), the NPT1 labelling was more diffuse (panels6–7). Attempts to perform immuno-TEM to localizeNPT1-HA at a specific membrane (PVM, plasmalemmaor other) were inconclusive. Nevertheless, the specificlabelling of NPT1-HA at the periphery of free merozoites(lower panel) strongly suggests a predominantly parasiteplasma membrane localization of NPT1, although alocalization to the inner membrane complex underneaththe plasma membrane cannot be excluded.
NPT1 is not essential for multiplication of asexualblood stages
To study the function of NPT1 during the parasite lifecycle, we disrupted npt1 in P. berghei NK65. We gener-ated a first targeting construct, named pNPT1-I, which,after double cross-over homologous recombination,would insert the hDHFR selectable cassette within thenpt1 coding sequence at amino acid 197 (Fig. 3A). Aftertransfection of the linearized plasmid pNPT1-I into NK65merozoites, recombinant parasites were selected andcloned. Southern blot analysis of genomic DNA con-firmed the predicted npt1 disruption in the NPT1-I clone(Fig. S5A). Unexpectedly, when transcription of npt1 inthe blood stages of the mutant NPT1-I was analysed byRT-PCR, we were able to amplify transcripts of the npt1gene. Primers PA and PB, which hybridize in the 3′ partof the npt1 ORF downstream of the hDHFR cassette,amplified a fragment of the expected size (Fig. 3B,upper panel, lanes 3 and 4). In addition, primers PC andPD, which hybridize on either side of the cassette, alsoamplified a fragment whose size corresponded to thefusion of the (wild-type) WT fragment to the 1.6-kb-long
hDHFR cassette (itself composed of 0.6 kb of the eef1apromoter sequences, 0.6 kb of the hDHFR ORF and0.4 kb of 3′ untranslated sequences of P. berghei DHFR-TS) (Fig. 3B, middle panel, lanes 3 and 4). These resultsshowed the presence of a long ‘chimeric’ transcriptencompassing the entire disrupted locus in the NPT1-Iparasites. Since we have not been able to generate spe-cific antibodies against PbNPT1, it was not possible toanalyse whether these transcripts would give rise to aNPT1 product.
We therefore constructed a second targeting construct,named pNPT1-D, which, after double cross-over homolo-gous recombination, would replace the entire npt1 codingsequence by the hDHFR selectable cassette (Fig. 3A).After electroporation of linearized pNPT1-D into NK65merozoites, a recombinant parasite clone, namedNPT1-D, was selected. Genotype analysis by Southern
Fig. 3. Targeted gene disruption of npt1.A. Schematic representation of npt1 locus in NPT1-I and NPT1-Dmutants. Grey boxes indicate the regions of homology used fordouble cross-over recombination; black arrow indicates theselectable marker hDHFR. PA/PB and PC/PD are primers used forRT-PCR analysis (arrow heads).B. Detection of npt1 transcripts by RT-PCR in mixed blood stagesof WT, NPT1-I and NPT1-D parasites. Middle panel shows thechimeric transcript detected in NPT1-I parasites. Primers PA/PB andPC/PD are as in (A). ‘+’ and ‘-’ indicate RT-PCR performed with orwithout reverse transcriptase, respectively.C. Normal multiplication of NPT1-I and NPT1-D parasites. Mice(n = 5 for each parasite line) were injected with mixed blood stagesto obtain an initial parasitemia of 0.001%. Graph shows theparasitemia of mice 3 and 4 days post infection. No statisticaldifference found between the parasitemia in mice infected with thedifferent lines (P = 0.55 and 0.15 for NPT1-I and NPT1-D,respectively compared with WT).
NPT1 is essential for Plasmodium gametocytogenesis 1347
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 81, 1343–1357

blot of genomic DNA confirmed the allelic replacement(Fig. S5B). In this mutant, RT-PCR analysis of total RNAof mixed blood stages demonstrated the absence of npt1transcript (Fig. 3B, lanes 5 and 6, Fig. S5C).
The asexual multiplication rate of NPT1-I and NPT1-Dparasites in the blood as determined during the cloningprocedure (Spaccapelo et al., 2010) was comparable toWT parasites, since all mice infected with a single para-site showed a parasitemia of 0.5–2% at day 8 afterinfection. Furthermore, when mice were infected byintravenous injection of similar numbers of WT, NPT1-Iand NPT1-D parasites, the developments of parasitemiaat days 3 and 4 after infection were comparable in allmice (Fig. 3C). These results demonstrate that the lackof NPT1 does not affect asexual blood stage multiplica-tion of P. berghei. Since the NPT1-I clone, despitegenerating a chimeric transcript at the npt1 locus, wasnot expected to produce a functional NPT1 protein, wecharacterized both the NPT1-I and NPT1-D mutantclones.
NPT1 is essential for production of mature and fertilegametocytes
To analyse parasite infectivity to mosquitoes, we allowedAnopheles stephensi mosquitoes to feed on mice infectedwith NK65 (WT), NPT1-I or NPT1-D parasites and analy-sed oocyst production at day 8 after the infectious bloodmeal. Oocysts were easily detectable in mosquitoesinfected with the WT parasites but were not found inmosquitoes fed on mice infected with the NPT1-I orNPT1-D clones (Fig. 4A), which also lacked sporozoites intheir salivary glands at days 21–24 (Fig. S5D). Bothmutants NPT1-I and NPT1-D showed the same pheno-type, confirming that parasites of the NPT1-I clone did notexpress a functional NPT1, despite the presence of the‘chimeric’ transcript.
We then analysed Giemsa-stained smears of tail-bloodof infected mice. While 0.5–5% gametocytes were consis-tently observed in the blood smears of WT-infected mice,we were unable to detect mature male gametocytes in
Fig. 4. NPT1-defective parasites are unable to produce fertile female and male gametes.A. Absence of oocysts in midguts of mosquitoes infected with NPT1-I or NPT1-D parasites compared with WT at days 8 or 14 post infection.Circles correspond to individual mosquitoes. The panel represents one of the two or more infections performed with three infected mice perparasite clone and per infection; number of mosquitoes analysed per infection: NK65 (WT) n = 20, 18, 46, 66; NPT1-I n = 20, 19, 22; NPT1-Dn = 64, 48.B. Exflagellation of male gametocytes from blood infected with WT, NPT1-I or NPT1-D parasites. Results obtained from five independentexperiments for the comparison WT/NPT1-I; two experiments for the comparison WT/NPT1-D; two infected mice were analysed per parasiteclone.C. Production of ookinetes in cross-fertilizations between WT, NPT1-I, NPT1-D, p47- and p45/48- parasites. Results obtained from at least twoindependent experiments: NK65 (WT) n = 5; NPT1-I n = 4; NPT1-D n = 2; p47- and p45/48- parasites n = 4; two infected mice were analysedper parasite clone. The WT cross-fertilizations gave 14–66% conversion of female gametocytes. The cross between aphidicolin treated WT(WT*, NK65) and p47- (ANKA) was performed as control (n = 1).
1348 B. Boisson et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 81, 1343–1357

smears of mutant-infected mice. In addition, male gameteformation could not be detected in standardized in vitroassays for exflagellating male gametocytes (Ponzi et al.,2009) using NPT1-I or NPT1-D parasites (Fig. 4B). InGiemsa-stained slides, low numbers of parasites weredetected that resembled female gametocytes. These wereuninuclear parasites, nearly filling the host cell with hemo-zoin granules scattered throughout the cytoplasm. Sincematuring, asexual trophozoites of P. berghei have a com-parable morphology and are difficult to distinguish fromfemale gametocytes (Mons et al., 1985), we performedstandard cross-fertilization assays (Ponzi et al., 2009) toinvestigate whether mutant parasites were able to producefertile female gametes. Cross-fertilization was performedwith a P. berghei mutant line, p47-, that produces fertilemale, but not female gametes (Khan et al., 2005). As acontrol line, we used a P. berghei mutant line, p45/48-,which produces fertile female, but not male gametes (vanDijk et al., 2001). Since the p47- and p45/48- mutationswere introduced in P. berghei ANKA while our npt1- muta-tions were generated in P. berghei NK65, and cross-fertilization between gametes of these lines has neverbeen reported before, we first verified that these two strainswere capable of cross-fertilization. For this, we co-infectedmice with equal numbers of parasites of a P. bergheiANKAreporter line expressing RFP via the eef1a promoter andparasites of a NK65 reporter line expressing GFP via theCS promoter. These mice were used to infect mosquitoesto allow for cross-fertilization between gametes of the twodifferent strains. Cross-fertilization was determined byanalysis of fluorescence of both mature oocysts and indi-vidual sporozoites, in which both the eef1a and cs promot-ers are active. The presence of oocysts and sporozoitesexpressing both GFP and RFP confirmed that cross-fertilization occurred between gametes of the NK65 andANKA strains (Fig. S6 and Movie S1).
Next, blood from mice infected with NPT1-I or NPT1-Dparasites was mixed with that of mice infected with eitherp47- or p45/48- parasites, and fertilization was scoredby the in vitro ookinete conversion rate as previouslydescribed (van Dijk et al., 2001). We verified that ooki-netes emerged after crossing the p47- with the p45/48-line, as well as after mixing the ANKA p47- line and WTNK65 parasites pre-treated with aphidicolin, an inhibitor ofpolymerase-a that specifically blocks the formation ofmale gametes. In all control experiments fertilization wassuccessful, as determined by the formation of ookinetes(Fig. 4C). In contrast, ookinetes were not observed afterself-fertilization of the NPT1-I or NPT1-D clones, or aftercross-fertilization of each of these mutants with either thep47- or the p45/48- lines (Fig. 4C). These results indi-cated that mutant parasites did not produce fertile femalegametes and confirmed the absence of male gameteproduction.
NPT1 deficiency leads to an early block ingametocytogenesis
To assess if gametocytogenesis occurred in the mutantparasites, the transcription of p28 (also known as pos28or pbs21), p25 (also known as pbs25) (Siden-Kiamoset al., 2000) and the translational regulator dozi (Mairet al., 2006), which are known to be transcribed in game-tocytes, were analysed by both non-quantitative andquantitative RT-PCR on RNA from mixed blood stages(Fig. S7A and B). These transcripts were detected in themutant parasites, indicating that they were capable ofundergoing some degree of sexual differentiation. Wethen analysed, by quantitative RT-PCR, the expressionof npt1 and p28 in samples enriched for erythrocyticschizonts and merozoites (asexual stages) or gameto-cytes (sexual stages) of the ANKA gametocyte producerline Pb871 and the gametocyte defective line Pb872. Theresults showed that npt1 is expressed in both asexual andsexual stages, in sharp contrast to the profile obtainedwith the gametocyte-transcribed gene p28 (Fig. 5A).
In order to quantify more precisely the reduction in thenumber of male and female gametocytes and to deter-mine when the defective phenotype appears, we intro-duced the npt1- mutations in a P. berghei reporter line thatexpresses GFP and RFP in male and female gameto-cytes, respectively. This reporter line, Fluo-frmg, containsa gfp and a rfp gene under the control of a male- andfemale-specific promoter, respectively, and allows forunequivocal detection of developing and mature gameto-cytes both by fluorescence microscopy and quantitativeFACS analysis (Ponzi et al., 2009). Recombinant para-sites lacking NPT1 expression were generated by trans-fection with the pNPT1-I or pNPT1-D constructs. Selectionand cloning resulted in the Fluo/NPT1-I and Fluo/NPT1-Dclones, respectively, whose correct genotypes were con-firmed by PCR and Southern blot analysis of genomicDNA (Fig. S5A and B). We analysed these parasites forthe presence of npt1 transcripts and observed, asdescribed above, a chimeric transcript in Fluo/NPT1-I butnot in Fluo/NPT1-D parasites (Fig. S5C).
Gametocyte production was analysed in standardizedsynchronous infections of rats after intravenous injectionof purified schizonts (Janse et al., 2006a). In these con-ditions, 15–25% of WT merozoites that have invadederythrocytes develop into mature gametocytes and 40 hafter infection these gametocytes are present in additionto rings and trophozoites from the second asexual cycle.The very low numbers of fluorescent cells excluded theirreliable quantification by manual counting. We thereforepurified gametocytes from the blood of rats with synchro-nous infections 40 h post infection and counted the fluo-rescent cells by FACS. As controls, we also analysed thePb871 (gametocyte producer) and Pb872 (gametocyte
NPT1 is essential for Plasmodium gametocytogenesis 1349
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 81, 1343–1357

non-producer) ANKA non-fluorescent parasite lines. Asshown in Fig. 5B, the numbers of mature male and femalegametocytes were greatly reduced (90–95%) in theFluo/NPT1-I (P = 0.11) and Fluo/NPT1-D (P = 0.03) linescompared with the WT. This 90–95% decrease in thegametocyte preparations was independently confirmedby quantitative RT-PCR of p28 transcripts (Fig. 5C).Moreover, > 90% of the cells from the Fluo/NPT1-I andFluo/NPT1-D lines presented an abnormal morphologywith vacuolization of the cytoplasm (Figs 5D and F andS8A). The abnormal morphology of mutant gametocyteswas confirmed by analysis by transmission electronmicroscopy. The rare gametocytes observed presentedunusual membrane structures (whorls) and large vacu-oles in their cytoplasm (Figs 5E and S8B), indicatinggeneral features of degeneration. Together, these obser-
vations demonstrated that NPT1-deficient parasiteswere severely compromised in the production of maturegametocytes.
Complementation of NPT1-defective parasites restoresgametocytogenesis and transmission to mosquitoes
In order to verify that the phenotype of mutant parasiteswas due solely to the genetic modifications introduced inthe npt1 locus, and not to other mutations that may haveoccurred during the selection of the parasites or to aneffect on neighbouring genes, we complemented theNPT1-I clone with a WT copy of npt1. A plasmid, c-npt1-gfp, containing 500 bp of the 5′ UTR, the entire ORF and1000 bp of the 3′ UTR of npt1, together with gfp drivenfrom the hsp70 promoter, was inserted by single cross-
Fig. 5. NPT1-defective parasites are severely impaired in gametocytogenesis.A. Histogram representation of npt1 and p28 relative gene expression in P. berghei asexual and sexual stages. The Pb871 line producesgametocytes and Pb872 does not produce gametocytes. The values were normalized relative to the expression of hsp70 mRNA in eachsample. Error bars are SD (n = 2).B. Flow cytometry analysis on purified gametocytes in Fluo-frmg (Fluo) lines. Male gametocytes are shown in green and female in red. Twonon-fluorescent lines Pb871 and Pb872 were used as controls for gating cell size. Right panels show box-plots of the number of gametocytescounted per 105 events at 42 h; four independent experiments with one infected rat per parasite clone. Box limits represent the first and thirdquantiles and median is shown as a bar. The asterisk indicates statistical significance, P-value < 0.05.C. Histogram representation of p28 relative gene expression in Fluo purified gametocytes. The values were normalized relative to theexpression of hsp70 mRNA in each sample.D. Micrographs of purified gametocytes stained with Giemsa from Fluo and Fluo/NPT1-defective parasites. Bar scale: 10 mm.E. Micrographs of ultrastructural sections of Fluo and Fluo/NPT1-I gametocytes obtained by TEM. : haemozin, : osmiophilic bodies, N:nucleus and V: vacuole. Scale bar represents 1 mm.F. Histogram representation of the numbers of morphologically normal (white box) versus abnormal (black box) gametocytes. White numbersin black boxes indicate the percentage of abnormal gametocytes; three independent experiments with one infected rat per parasite clone.
1350 B. Boisson et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 81, 1343–1357

over recombination into the npt1-I locus. This insertiongenerated a full-length npt1 ORF driven by the nativepromoter and 1000 bp of the endogenous 3′ UTR(Fig. 6A). The complemented NPT1-I clone, c-NPT1-GFP,was selected after two rounds of cell sorting for GFP-expressing parasites, as previously described (Janseet al., 2006a). RT-PCR analysis performed on c-NPT1-GFP mixed blood stages parasites showed that npt1expression was indeed restored (Fig. 6B).
Analysis of Giemsa-stained blood smears of c-NPT1-GFP infected mice revealed the presence of morphologi-cally mature gametocytes, and the gametocytaemia wascomparable to NK65 infected mice (see Fig. S9A). Asshown in Fig. 6C (left panel), c-NPT1-GFP parasites wereable to produce mature fertile gametes and ookinetesin vitro in numbers comparable to the WT. Moreover,c-NPT1-GFP parasites were able to infect the midgut(Fig. 6C, middle panel) and salivary glands of mosquitoes
(Fig. 6C, right panel) like WT parasites. These resultsdemonstrate the essential role of NPT1 in gametocytoge-nesis and parasite transmission to mosquitoes.
Discussion
We show here that a novel putative transporter, NPT1,plays a critical role in early stages of P. berghei sexualdevelopment. Parasites lacking NPT1 were severelyimpaired in gametocytogenesis, unable to produce fertilegametes and consequently non-infectious to mosqui-toes, and complementation of the defective parasiteswith a WT copy of npt1 restored gametocyte formationand mosquito infection. We provide evidence thatNPT1 is located at the parasite plasma membrane inasexual blood stages, including free merozoites. Theexpression of npt1 is not significantly upregulated insexual stages and the detection of the protein in asexual
Fig. 6. Complementation of NPT1-I parasitesA. Schematic representation of single cross-over strategy used to complement NPT1-I parasites using GFP as a selectable marker. Thelinearized plasmid containing the WT npt1 locus (grey arrow) and GFP cassette (green arrow) was transfected into NPT1-I parasites. Thepromoter region (broken arrow) and the 3′ UTR region (lollipop) are indicated. The triple line represents the plasmid backbone and black barthe probe used for Southern blot analysis.B. RT-PCR analysis of npt1 transcript (indicated by arrow) in mixed blood stages of WT, NPT1-I and c-NPTI-GFP. Primers used are as inFig. 2B.C. Top panels: GFP expression in c-NPTI-GFP ookinetes, oocysts and sporozoites. The salivary gland sporozoites were immunostained withanti-CS antibody coupled to an Alexa Fluor 568 antibody. Bottom panels: Histogram representation of numbers of ookinetes, oocysts andsporozoites in the c-NPTI-GFP clone compared with WT and NPT1-I parasites; two independent infections.
NPT1 is essential for Plasmodium gametocytogenesis 1351
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 81, 1343–1357

blood stages, in addition to gametocytes, suggests thatit may be required in the precursors of sexual stages.This is the first example of the role of a putative trans-port protein in P. berghei sexual development. Consider-ing the high degree of conservation of NPT1 in allPlasmodium species, it likely plays a similar role ingametocytogenesis of other malaria parasites includingP. falciparum.
Membrane transport proteins are integral membraneproteins that mediate the passage across the membranebilayer of specific molecules and ions. In Plasmodium, asin other organisms, they serve a diverse range of importantphysiological roles including uptake of nutrients, elimina-tion of waste products and maintenance of transmembraneelectrochemical gradients (see Martin et al., 2005; 2009and references therein). More than 100 transport proteinshave been identified in P. falciparum but only a few havebeen cloned and functionally characterized. These pro-teins are increasingly recognized as valid drug targets(Staines et al., 2010). They play a key role in the growthand replication of blood stages of the parasite, as well as inthe development of drug resistance against antimalarials(Kirk, 2004; Sanchez et al., 2010). However, it is largelyunknown whether the different stages of Plasmodiumparasites use similar or different transport machineriesduring their complex life cycle. For example, it is unknownwhether the uptake of nutrients for growth and multiplica-tion of blood stages is mediated by the same transportersas those used by mosquito or liver stage parasites.
NPT1 belongs to the MFS of transmembrane solutetransporters. This huge superfamily contains 29 families,each transporting a specific substrate, for examplesugars, nucleosides, metabolites, organic anions andcations, etc. (for reviews, see Pao et al., 1998; Saieret al., 1999). At present, the function of NPT1 cannot bepredicted since despite having features of the MFS oftransport proteins, it lacks key amino acid signaturesthat could indicate the molecule(s) it might transport(Martin et al., 2005; 2009). It could play a role as asensor for a signal leading to sexual differentiation (com-mitment) and/or during the maturation/homeostasis/survival of gametocytes. The presence of a few earlystage gametocytes in NPT1-defective parasites togetherwith the presence of some p28 transcripts suggests thatgametocytogenesis might still initiate, albeit inefficiently.On the other hand, the drastic reduction in gametocytenumbers and their abnormal morphology suggests NPT1plays a very early role in gametocytogenesis, possiblycommitment itself. It is interesting to note that acarboxylate-transporter (PAD) belonging to the MFS hasrecently been shown to play an essential role in trypa-nosome differentiation (Dean et al., 2009).
We have shown that the levels of Pbnpt1 transcriptsare increased in liver stages. Furthermore, npt1 is
upregulated when P. falciparum sporozoites are placedin contact with hepatocytes (Siau et al., 2008) and theP. yoelii orthologous protein has been detected in liverstages (Tarun et al., 2008). These data suggest thatNPT1 may also play a role in the development ofPlasmodium liver stages. Since NPT1-deficient para-sites do not produce gametocytes and cannot infectmosquitoes, we were unable to test this hypothesisdirectly. However, we have indirect evidence that thismay be the case. When c-NPT1-GFP sporozoites wereinjected into rats, a delay in the emergence of bloodinfection was observed and the emerging parasites werefound to have undergone a recombination event thatreverted to the WT npt1 allele (see Fig. S9B). The npt1locus in c-NPT1-GFP parasites is interrupted in the 3′UTR region before the sequence that is highly con-served in all Plasmodium species. Several other npt1recombinant parasites have been generated in the labo-ratory that have modifications downstream of the npt1ORF and that also present an impaired pre-erythrocyticphase, as determined by the prepatent period of bloodinfection. This leads to the intriguing possibility thatNPT1 not only plays a role in gametocytes but also inliver stages.
It is now widely accepted that meeting the new chal-lenge of parasite eradication will require tools, includingdrugs, which not only target parasite multiplication in theblood but also inhibit transmission of the parasite betweenthe two hosts. If conditional mutagenesis of npt1 were toprovide evidence for an essential role of NPT1 in liverstages, then this protein would be a highly attractive drugtarget, since blocking NPT1 activity would prevent bothsteps of parasite transmission.
Experimental procedures
Rodents, parasite strains and mosquito infections
Swiss and C57BL/6 mice and female Wistar rats wereobtained from Janvier (France).
The reference ‘wild-type’ Plasmodium berghei strains areNK65 and ANKA cl15cy1 strain (obtained from MR4, MRA-871:Pb871). The ANKA non-gametocyte producer cloneHPEcy1m50cl1 is MR4-872 (Pb872) and the GFP ANKAline (Janse et al., 2006b) (MR4-867:Pb867). The ANKA lineexpressing RFP from the eef1a promoter (ANKA L733)is described in Sturm et al. (2009). The NK65 derivativeexpressing GFP via the CS promoter and inserted at CSlocus is described in Natarajan et al. (2001). The Fluo-frmg(Fluo) line expressing RFP in female gametocytes and GFPin male gametocytes has been described in Ponzi et al.(2009). It expresses RFP in female gametocytes at 30 h postinvasion, via the PBANKA_131950 (coding for CCP2) pro-moter and GFP in male gametocytes at 16 h, via thePBANKA_041610 (coding for dynein heavy chain, putative)promoter. The ANKA p47- line that does not produce femalegametes and the p45/48- line that does not produce male
1352 B. Boisson et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 81, 1343–1357

gametes are described in Khan et al. (2005) and van Dijket al. (2001), respectively.
Plasmodium berghei parasites were maintained in Swissmice and transmitted by A. stephensi, strain SD500, asdescribed previously (Thiberge et al., 2007). The course ofinfections and gametocyte production were monitored onGiemsa-stained blood smears and mosquitoes fed for 2 con-secutive days on mice with 1–2% parasitemia. The number ofoocysts per midgut on days 8–12 post blood meal and thenumber of sporozoites per salivary glands on days 21–24were counted.
All studies on animals complied with the guidelines on theethical use of animals from the European CommunitiesCouncil Directive of 24 November 1986 (86/609/EEC).
Primers
See Table S1 in Supporting Information.
Quantitative reverse transcription polymerasechain reaction
RNA was collected in two independent experiments from thefollowing stages/parasite samples of the NK65 strain: mixedblood stages, purified gametocytes, salivary gland sporozoi-tes (day 21 post infection) and infected HepG2 cells (at 5, 17,40 and 50 h post infection). After reverse transcription, serialdilutions of cDNAs were made in triplicate. PCR conditionswere one cycle at 95°C for 10 min, 40 cycles at 95°C for 15 s,55°C for 15 s and 60°C for 45 s. npt1pt1 mRNA was normal-ized to the hsp70 (PB001074.01.0) mRNA in each sample.
Identification of 5� and 3� UTRs by RACE
A RACE library was made from total RNA isolated from2.106 salivary gland sporozoites with the Marathon cDNAAmplification kit-Clontech according to manufacturer’srecommendations. The PCR fragments were cloned in TOPOTA vector and sequenced.
Gene inactivation by double cross-over strategy
Plasmids for generating NPT1-I and NPT1-D parasites wereconstructed by cloning about 600 bp of the 5′ and 3′ genomicsequences of npt1 on either side of the hDHFR cassette inthe pBC vector (Fig. S10A). NPT1-I contains an insertion of1601 bp at position amino acid 197 of the predicted ORF andNPT1-D a complete deletion of the predicted ORF. See Sup-porting Information for Southern blot analyses.
Generation and selection of c-NPT1-I complementedparasites by single cross-over strategy
The entire ORF of npt1 and the UTRs, corresponding tonucleotides 100–3000 of the contig RP1871, were amplifiedfrom NK65 genomic DNA with primers RP1234-F andRP1871-R. This fragment was cloned into the pBC vectorcontaining the GFP cassette (Fig. S10B). The resulting
plasmid, c-npt1I-gfp, was linearized with PstI and transfectedinto the NPT1-I (clone c24). Recombinant parasites wereisolated as described in Janse et al. (2006a). Briefly, aftertransfection parasites were injected into two C57BL/6 miceand 3 days later, GFP-positive parasites from both mice weresorted on a FACSAria (BD Biosciences) and injected into fivemice (109–225 parasites injected per mouse). One mousebecame infected with GFP-expressing parasites. This popu-lation was sorted and about 2000 GFP- positive parasiteswere injected into two naïve mice. Both became positive andwere determined to be pure by the correlation between para-sitemia measured by GFP+ infected erythrocytes andGiemsa-stained blood smears.
Generation of NPT1-HA parasites by doublecross-over strategy
The HA-FLAG sequence was cloned in pBC between HindIIIand EcoRI sites and the 3′ UTR of hsp70 (737 bp) wasinserted downstream. The hDHFR cassette (see above) wasintroduced between the SmaI and NotI sites. The C-terminussequence (959 bp) of the npt1 coding region was clonedbetween ApaI and ClaI and the 3′ UTR (866 bp) between NotIand AscI. The resulting plasmid, pBC-npt1-HA, was digestedwith ApaI and AscI before transfection (Fig. S4A). Duringcloning procedure (Spaccapelo et al., 2010), NPT1-HAgrowth was comparable to WT parasites (parasitemia of0.5–2% at day 8 after infection).
Exflagellation and cross-fertilization
Exflagellation and cross-fertilization experiment were per-formed as described in Ponzi et al. (2009). Briefly, two orthree drops of blood from the tail of infected mice were mixedwith 2 ml of heparin (Sigma; 5000 units ml-1 in PBS). Fivemicrolitres were placed on a glass slide, covered with acoverslip and incubated for 10 min in a humid chamber at20°C. Samples were immediately observed on a Axiostarmicroscope (Zeiss) with 40¥ objective. The number of exfla-gellation events observed per homogenous field, containingabout 3000 red blood cells, were noted. In parallel, a bloodsmear was made from the tail of infected mice and the para-sites revealed by Giemsa staining.
Ookinetes were obtained from the in vitro culture ofgametocyte-infected mouse blood as described in Janseet al. (1985). Blood of infected mice was collected (seebelow) and added to RPMI 1640 media (Gibco) supple-mented with 10% FCS adjusted to pH 8.0. After 22 h incuba-tion at 20°C the number of ookinetes was determined bymicroscopic observation (¥40). Aphidicolin treatment wasperformed as described in Janse et al. (1986). The drug wasremoved from the culture medium after 12 min by centrifuga-tion of gametocytes/gametes, by discarding the supernatantand replacing by fresh media.
Purification of gametocytes and FACS analysis
Synchronized infections of rats and purification of gameto-cytes were performed as described in Janse and Waters
NPT1 is essential for Plasmodium gametocytogenesis 1353
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 81, 1343–1357

(1995). Briefly, parasites were injected into rats treated 4days earlier with 35 mg kg-1 phenylhydrazine (SIGMA-Aldrich). When parasitemia reached 1–3% cardiac puncturewas performed and blood transferred to RPMI mediumsupplemented with 20% FCS, HEPES 25 mM, neomycin 50units ml-1. Cultures were maintained at 37°C, in 5%CO2 + 10% O2 for 16 h with gentle agitation. Schizonts werepurified on a 55% Nycodenz (Axis-Shield PoC, Norway) gra-dient and injected into the tail vein of rats. Twenty-six hourslater gametocytes were separated from ring stages on a 49%Nycodenz gradient. Immature gametocytes were analysedimmediately by FACS or allowed to mature in culture for upto 48 h. RFP and GFP gametocytes were analysed on aFACsCalibur (Becton Dickinson).
Indirect Immunofluorescence assay
Mixed blood stages of NPT1-HA or Pb867 were enriched ona CS column using a VarioMacs (Miltenyi Biotec) and sch-izonts were purified from cultures on Nycodenz gradients.Samples were washed with PBS and fixed with 4% PFA +0.0075% glutaraldehyde for 30 min as described by Tonkinet al. (2004). Cells were then permeabilized with 0.001%Triton X-100 for 10 min. Samples were incubated with a ratanti-HA high affinity monoclonal antibody (clone 3FC10,Roche) and a chicken anti-exp1 antibody [kindly provided byDr V. Heussler (Sturm et al., 2009)] diluted in PBS + 3% BSAfor 1 h at RT or rabbit anti-SET antibody [kindly provided byDr M. Ponzi (Pace et al., 2006)]. After several washes,samples were incubated with Alexa Fluor conjugated second-ary antibodies (Molecular probes) and DAPI. Samples weremounted in Prolong (Molecular probes) for observation on aZeiss Observer Z1 microscope. Images were acquired usingAxiovision software.
Transmission electron microscopy
Mixed blood stages of WT (Pb867), NPT1-I and NPT1-Dparasites were enriched on a CS column using a VarioMacs(Miltenyi Biotec). Samples were processed as described inOlivieri et al. (2009). Contrasted ultra-thin sections (60 nm)were observed in a JEM 1010 Transmission Electron Micro-scope (Jeol, Tokyo, Japan).
Statistical analysis
The quantitative reverse transcription polymerase chain reac-tion data in Fig. 1 were analysed by Wilcox test.
The FACs analysis in Fig. 4B was analysed by Kruskal–Wallis test.
Phylogenetic analysis of MFS
The Pfam HMM profile MFS_1 (PF07690) and the Hmmersoftware were used to scan 16 Apicomplexa genomes(Babesia bovis, Crytosporidium hominis, Crytosporidiummuris, Crytosporidium parvum, Neospora caninum, Plasmo-dium berghei, Plasmodium chabaudi, Plasmodium falci-parum, Plasmodium gallinaceum, Plasmodium knowlesi,Plasmodium reichenowi, Plasmodium vivax, Plasmodium
yoelii, Theileria annulata, Theileria parva, Toxoplasmagondii ) and two Cilata genomes (Paramecium tetraurelia,Tetrahymena thermophyla). The Saccharomyces cerevisiaeprotein NP_009739.1 was used as an out-group. This analy-sis identified 744 members in the 18 Alveolata genomes. Theamino acid sequences were then aligned using CLUSTAL
2.0.12. Proteins sequences from the NPT family of Plasmo-dium species were also aligned separately and the tree wasconstructed using the Neighbour joining algorithm with 1000bootstrap trials (MEGA 4.0). The phylogenetic tree was drawnusing Figtree 1.3.1.
Software and website references
WHO, Geneva, Switzerland: http://www.who.int/malaria/world_malaria_report_2010/world_malaria_report_2010.pdfPlasmoDB: http://plasmodb.org/plasmo/GeneDB: http://www.genedb.org/TMHMM server: http://www.cbs.dtu.dk/services/TMHMM/POLYAH: http://linux1.softberry.com/CLUSTALW: http://www.ebi.ac.uk/clustalw/Hmmer: http://hmmer.org/Figtree 1.3.1.: http://tree.bio.ed.ac.uk/software/figtree/
Data deposition
Sequences were deposited in the GenBank database. 5′UTRs: # GU907662, GU907663, GU907664; 3′UTRs: #GU907665, GU907666; 2 kb region between contigs RP1871and RP1234: # GU907667.
Acknowledgements
We thank members of Biologie and Génétique du Paludisme,the Leiden Malaria Research Group, Drs G. Milon, P. David,R. Paul for stimulating discussions. We are extremely gratefulto Dr Lucia Bertuccini for providing advice on TEM prepara-tions and observations. Dr M.C. Prevost of the Plate-forme deMicroscopie Ultrastructurale (Institut Pasteur) is thanked forhelp with TEM. Drs C. Bourgouin, I. Thiéry and the othermembers of the ‘Centre de Production et d’ Infection desAnophèles’ (Institut Pasteur) are thanked for mosquitorearing. B.B. was supported by the Fonds dédiés ‘Combattreles maladies Parasitaires’ financed by the Ministère de laRecherche and Sanofi-Aventis. This work was supported byfunds from the Institut Pasteur, the Howard Hughes MedicalInstitute, Natixis and the European Commission (FP6 BioMa-lPar Network of Excellence and FP7/2007–2013 under grantagreement N°242095).
References
Alano, P. (2007) Plasmodium falciparum gametocytes: stillmany secrets of a hidden life. Mol Microbiol 66: 291–302.
Billker, O., Dechamps, S., Tewari, R., Wenig, G., Franke-Fayard, B., and Brinkmann, V. (2004) Calcium and acalcium-dependent protein kinase regulate gamete forma-tion and mosquito transmission in a malaria parasite. Cell117: 503–514.
Bruce, M.C., Alano, P., Duthie, S., and Carter, R. (1990)Commitment of the malaria parasite Plasmodium falci-
1354 B. Boisson et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 81, 1343–1357

parum to sexual and asexual development. Parasitology100 (Part 2): 191–200.
Bruce, M.C., Carter, R.N., Nakamura, K., Aikawa, M., andCarter, R. (1994) Cellular location and temporal expressionof the Plasmodium falciparum sexual stage antigen Pfs16.Mol Biochem Parasitol 65: 11–22.
Carter, R., and Graves, P. (1988) Gametocytes. In Malaria:Principles and Practice of Malariology. Wernsdorfer, W.H.,and McGregor, I. (eds). Edinburgh: Churchill Livingstone,pp. 253–305.
Carter, R., Graves, P.M., Quakyi, I.A., and Good, M.F. (1989)Restricted or absent immune responses in human popula-tions to Plasmodium falciparum gamete antigens that aretargets of malaria transmission-blocking antibodies. J ExpMed 169: 135–147.
Dean, S., Marchetti, R., Kirk, K., and Matthews, K.R. (2009)A surface transporter family conveys the trypanosome dif-ferentiation signal. Nature 459: 213–217.
van Dijk, M.R., Janse, C.J., Thompson, J., Waters, A.P.,Braks, J.A., Dodemont, H.J., et al. (2001) A central role forP48/45 in malaria parasite male gamete fertility. Cell 104:153–164.
Dixon, M.W., Thompson, J., Gardiner, D.L., and Trenholme,K.R. (2008) Sex in Plasmodium: a sign of commitment.Trends Parasitol 24: 168–175.
Doerig, C., Baker, D., Billker, O., Blackman, M.J., Chitnis, C.,Dhar Kumar, S., et al. (2009) Signalling in malariaparasites. The MALSIG consortium. Parasite 16: 169–182.
Dorin, D., Alano, P., Boccaccio, I., Ciceron, L., Doerig, C.,Sulpice, R., and Parzy, D. (1999) An atypical mitogen-activated protein kinase (MAPK) homologue expressed ingametocytes of the human malaria parasite Plasmodiumfalciparum. Identification of a MAPK signature. J Biol Chem274: 29912–29920.
Drakeley, C., Sutherland, C., Bousema, J.T., Sauerwein,R.W., and Targett, G.A. (2006) The epidemiology ofPlasmodium falciparum gametocytes: weapons of massdispersion. Trends Parasitol 22: 424–430.
Eksi, S., Haile, Y., Furuya, T., Ma, L., Su, X., and Williamson,K.C. (2005) Identification of a subtelomeric gene familyexpressed during the asexual-sexual stage transition inPlasmodium falciparum. Mol Biochem Parasitol 143:90–99.
Furuya, T., Mu, J., Hayton, K., Liu, A., Duan, J., Nkrumah, L.,et al. (2005) Disruption of a Plasmodium falciparum genelinked to male sexual development causes early arrest ingametocytogenesis. Proc Natl Acad Sci USA 102: 16813–16818.
Gunther, K., Tummler, M., Arnold, H.H., Ridley, R., Goman,M., Scaife, J.G., and Lingelbach, K. (1991) An exportedprotein of Plasmodium falciparum is synthesized as anintegral membrane protein. Mol Biochem Parasitol 46:149–157.
Ishino, T., Boisson, B., Orito, Y., Lacroix, C., Bischoff, E.,Loussert, C., et al. (2009) LISP1 is important for the egressof Plasmodium berghei parasites from liver cells. CellMicrobiol 11: 1329–1339.
Janse, C.J., and Waters, A.P. (1995) Plasmodium berghei:the application of cultivation and purification techniques tomolecular studies of malaria parasites. Parasitol Today 11:138–143.
Janse, C.J., Mons, B., Rouwenhorst, R.J., Van der Klooster,P.F., Overdulve, J.P., and Van der Kaay, H.J. (1985) In vitroformation of ookinetes and functional maturity ofPlasmodium berghei gametocytes. Parasitology 91:19–29.
Janse, C.J., van der Klooster, P.F., van der Kaay, H.J., vander Ploeg, M., and Overdulve, J.P. (1986) DNA synthesis inPlasmodium berghei during asexual and sexualdevelopment. Mol Biochem Parasitol 20: 173–182.
Janse, C.J., Franke-Fayard, B., and Waters, A.P. (2006a)Selection by flow-sorting of genetically transformed, GFP-expressing blood stages of the rodent malaria parasite,Plasmodium berghei. Nat Protoc 1: 614–623.
Janse, C.J., Franke-Fayard, B., Mair, G.R., Ramesar, J.,Thiel, C., Engelmann, S., et al. (2006b) High efficiencytransfection of Plasmodium berghei facilitates novel selec-tion procedures. Mol Biochem Parasitol 145: 60–70.
Kappe, S.H., Gardner, M.J., Brown, S.M., Ross, J., Matus-chewski, K., Ribeiro, J.M., et al. (2001) Exploring the tran-scriptome of the malaria sporozoite stage. Proc Natl AcadSci USA 98: 9895–9900.
Kato, K., Sudo, A., Kobayashi, K., Sugi, T., Tohya, Y., andAkashi, H. (2009) Characterization of Plasmodium falci-parum calcium-dependent protein kinase 4. Parasitol Int58: 394–400.
Khan, S.M., Franke-Fayard, B., Mair, G.R., Lasonder, E.,Janse, C.J., Mann, M., and Waters, A.P. (2005) Proteomeanalysis of separated male and female gametocytesreveals novel sex-specific Plasmodium biology. Cell 121:675–687.
Kirk, K. (2004) Channels and transporters as drug targets inthe Plasmodium-infected erythrocyte. Acta Trop 89: 285–298.
Kongkasuriyachai, D., Fujioka, H., and Kumar, N. (2004)Functional analysis of Plasmodium falciparum parasito-phorous vacuole membrane protein (Pfs16) duringgametocytogenesis and gametogenesis by targeted genedisruption. Mol Biochem Parasitol 133: 275–285.
Kooij, T.W., Carlton, J.M., Bidwell, S.L., Hall, N., Ramesar, J.,Janse, C.J., and Waters, A.P. (2005) A Plasmodium whole-genome synteny map: indels and synteny breakpoints asfoci for species-specific genes. PLoS Pathog 1: e44.
Lal, K., Delves, M.J., Bromley, E., Wastling, J.M., Tomley,F.M., and Sinden, R.E. (2009) Plasmodium male develop-ment gene-1 (mdv-1) is important for female sexualdevelopment and identifies a polarised plasma mem-brane during zygote development. Int J Parasitol 39: 755–761.
Lanfrancotti, A., Bertuccini, L., Silvestrini, F., and Alano, P.(2007) Plasmodium falciparum: mRNA co-expression andprotein co-localisation of two gene products upregulated inearly gametocytes. Exp Parasitol 116: 497–503.
Lasonder, E., Ishihama, Y., Andersen, J.S., Vermunt, A.M.,Pain, A., Sauerwein, R.W., et al. (2002) Analysis of thePlasmodium falciparum proteome by high-accuracy massspectrometry. Nature 419: 537–542.
Lobo, C.A., and Kumar, N. (1998) Sexual differentiation anddevelopment in the malaria parasite. Parasitol Today 14:146–150.
Lobo, C.A., Fujioka, H., Aikawa, M., and Kumar, N. (1999)Disruption of the Pfg27 locus by homologous recombina-
NPT1 is essential for Plasmodium gametocytogenesis 1355
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 81, 1343–1357

tion leads to loss of the sexual phenotype in P. falciparum.Mol Cell 3: 793–798.
McRobert, L., Taylor, C.J., Deng, W., Fivelman, Q.L., Cum-mings, R.M., Polley, S.D., et al. (2008) Gametogenesis inmalaria parasites is mediated by the cGMP-dependentprotein kinase. PLoS Biol 6: e139.
Mair, G.R., Braks, J.A., Garver, L.S., Wiegant, J.C., Hall, N.,Dirks, R.W., et al. (2006) Regulation of sexual develop-ment of Plasmodium by translational repression. Science313: 667–669.
Martin, R.E., Henry, R.I., Abbey, J.L., Clements, J.D., andKirk, K. (2005) The ‘permeome’ of the malaria parasite: anoverview of the membrane transport proteins of Plasmo-dium falciparum. Genome Biol 6: R26.
Martin, R.E., Ginsburg, H., and Kirk, K. (2009) Membranetransport proteins of the malaria parasite. Mol Microbiol 74:519–528.
Mons, B. (1986) Intra erythrocytic differentiation of Plasmo-dium berghei. Acta Leiden 54: 1–124.
Mons, B., Janse, C.J., Boorsma, E.G., and Van der Kaay,H.J. (1985) Synchronized erythrocytic schizogony andgametocytogenesis of Plasmodium berghei in vivo and invitro. Parasitology 91: 423–430.
Natarajan, R., Thathy, V., Mota, M.M., Hafalla, J.C., Menard,R., and Vernick, K.D. (2001) Fluorescent Plasmodiumberghei sporozoites and pre-erythrocytic stages: a new toolto study mosquito and mammalian host interactions withmalaria parasites. Cell Microbiol 3: 371–379.
Olivieri, A., Camarda, G., Bertuccini, L., van de Vegte-Bolmer, M., Luty, A.J., Sauerwein, R., and Alano, P. (2009)The Plasmodium falciparum protein Pfg27 is dispensablefor gametocyte and gamete production, but contributes tocell integrity during gametocytogenesis. Mol Microbiol 73:180–193.
Pace, T., Olivieri, A., Sanchez, M., Albanesi, V., Picci, L.,Siden Kiamos, I., et al. (2006) Set regulation in asexualand sexual Plasmodium parasites reveals a novel mecha-nism of stage-specific expression. Mol Microbiol 60: 870–882.
Pao, S.S., Paulsen, I.T., and Saier, M.H., Jr (1998) Majorfacilitator superfamily. Microbiol Mol Biol Rev 62: 1–34.
Paul, R.E., Coulson, T.N., Raibaud, A., and Brey, P.T. (2000)Sex determination in malaria parasites. Science 287: 128–131.
Ponzi, M., Siden-Kiamos, I., Bertuccini, L., Curra, C., Kroeze,H., Camarda, G., et al. (2009) Egress of Plasmodiumberghei gametes from their host erythrocyte is mediatedby the MDV-1/PEG3 protein. Cell Microbiol 11: 1272–1288.
Rangarajan, R., Bei, A.K., Jethwaney, D., Maldonado, P.,Dorin, D., Sultan, A.A., and Doerig, C. (2005) A mitogen-activated protein kinase regulates male gametogenesisand transmission of the malaria parasite Plasmodiumberghei. EMBO Rep 6: 464–469.
Saier, M.H., Jr, Beatty, J.T., Goffeau, A., Harley, K.T., Heijne,W.H., Huang, S.C., et al. (1999) The major facilitatorsuperfamily. J Mol Microbiol Biotechnol 1: 257–279.
Sanchez, C.P., Dave, A., Stein, W.D., and Lanzer, M. (2010)Transporters as mediators of drug resistance in Plasmo-dium falciparum. Int J Parasitol 40: 1109–1118.
Siau, A., Silvie, O., Franetich, J.F., Yalaoui, S., Marinach, C.,
Hannoun, L., et al. (2008) Temperature shift and host cellcontact up-regulate sporozoite expression of Plasmodiumfalciparum genes involved in hepatocyte infection. PLoSPathog 4: e1000121.
Siden-Kiamos, I., Vlachou, D., Margos, G., Beetsma, A.,Waters, A.P., Sinden, R.E., and Louis, C. (2000) Distinctroles for pbs21 and pbs25 in the in vitro ookinete to oocysttransformation of Plasmodium berghei. J Cell Sci 113 (Part19): 3419–3426.
Silvestrini, F., Alano, P., and Williams, J.L. (2000) Commit-ment to the production of male and female gametocytes inthe human malaria parasite Plasmodium falciparum. Para-sitology 121 (Part 5): 465–471.
Silvestrini, F., Bozdech, Z., Lanfrancotti, A., Di Giulio, E.,Bultrini, E., Picci, L., et al. (2005) Genome-wide identifica-tion of genes upregulated at the onset of gametocytogen-esis in Plasmodium falciparum. Mol Biochem Parasitol143: 100–110.
Smith, T.G., Lourenco, P., Carter, R., Walliker, D., andRanford-Cartwright, L.C. (2000) Commitment to sexual dif-ferentiation in the human malaria parasite, Plasmodiumfalciparum. Parasitology 121: 127–133.
Spaccapelo, R., Janse, C.J., Caterbi, S., Franke-Fayard, B.,Bonilla, J.A., Syphard, L.M., et al. (2010) Plasmepsin4-deficient Plasmodium berghei are virulence attenuatedand induce protective immunity against experimentalmalaria. Am J Pathol 176: 205–217.
Staines, H.M., Derbyshire, E.T., Slavic, K., Tattersall, A., Vial,H., and Krishna, S. (2010) Exploiting the therapeutic poten-tial of Plasmodium falciparum solute transporters. TrendsParasitol 26: 284–296.
Sturm, A., Graewe, S., Franke-Fayard, B., Retzlaff, S., Bolte,S., Roppenser, B., et al. (2009) Alteration of the parasiteplasma membrane and the parasitophorous vacuole mem-brane during exo-erythrocytic development of malariaparasites. Protist 160: 51–63.
Talman, A.M., Domarle, O., McKenzie, F.E., Ariey, F., andRobert, V. (2004) Gametocytogenesis: the puberty of Plas-modium falciparum. Malar J 3: 24.
Tarun, A.S., Peng, X., Dumpit, R.F., Ogata, Y., Silva-Rivera,H., Camargo, N., et al. (2008) A combined transcriptomeand proteome survey of malaria parasite liver stages. ProcNatl Acad Sci USA 105: 305–310.
Taylor, C.J., McRobert, L., and Baker, D.A. (2008) Disruptionof a Plasmodium falciparum cyclic nucleotide phosphodi-esterase gene causes aberrant gametogenesis. Mol Micro-biol 69: 110–118.
Thiberge, S., Blazquez, S., Baldacci, P., Renaud, O., Shorte,S., Menard, R., and Amino, R. (2007) In vivo imaging ofmalaria parasites in the murine liver. Nat Protoc 2: 1811–1818.
Thomson, D. (1914) The origin and development of gametes(crescents) in malignant tertian malaria: some observa-tions on flagellation etc. Ann Trop Med Parasitol 8: 85–104.
Tonkin, C.J., van Dooren, G.G., Spurck, T.P., Struck, N.S.,Good, R.T., Handman, E., et al. (2004) Localization oforganellar proteins in Plasmodium falciparum using a novelset of transfection vectors and a new immunofluorescencefixation method. Mol Biochem Parasitol 137: 13–21.
WHO (2010) World Malaria Report. Geneva, Switzerland:World Health Organisation.
1356 B. Boisson et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 81, 1343–1357

Young, J.A., Fivelman, Q.L., Blair, P.L., Vega, P., Le Roch,K.G., Zhou, Y., et al. (2005) The Plasmodium falciparumsexual development transcriptome: a microarray analysisusing ontology-based pattern identification. Mol BiochemParasitol 143: 67–79.
Young, J.A., Johnson, J.R., Benner, C., Yan, S.F., Chen, K.,Le Roch, K.G., et al. (2008) In silico discovery of transcrip-tion regulatory elements in Plasmodium falciparum. BMCGenomics 9: 70.
Supporting information
Additional supporting information may be found in the onlineversion of this article.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting materials suppliedby the authors. Any queries (other than missing material)should be directed to the corresponding author for the article.
NPT1 is essential for Plasmodium gametocytogenesis 1357
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 81, 1343–1357





























