Embed Size (px)
Citation preview

Is adult neurogenesis essential forolfaction?Francoise Lazarini1,2 and Pierre-Marie Lledo1,2
1 Institut Pasteur, Laboratory for Perception and Memory, 75015 Paris, France2Centre National de la Recherche Scientifique (CNRS) Unite de Recherche Associee 2182, 75015 Paris, France
In mammals, new neurons are recruited into restrictedbrain areas throughout life. Adult-born neurons pro-duced in the subventricular zone of the lateral ventriclemigrate rostrally towards the olfactory bulb. Althoughthousands of neurons reach this central structure everyday, the functional impact of their integration into ma-ture circuits remains a matter of debate. Recent inves-tigations have revealed no striking sensory deficits perse when adult bulbar neurogenesis is challenged. How-ever, some cognitive functions, such as perceptual learn-ing and olfactory memory, are clearly impaired. In thisreview we highlight the role of network activity in shap-ing ongoing neurogenesis and, in turn, how the integra-tion of adult-born neurons refines pre-existing networkfunction, and consequently olfactory behavior.
IntroductionNew neurons are continuously generated in two discreteareas of the adult brain: the subventricular zone (SVZ) ofthe lateral ventricles and the subgranular zone of thehippocampus [1]. The latter gives rise to new granuleneurons, whichmature locally in the dentate gyrus, where-as the former produces neuroblasts that migrate along therostral migratory stream (RMS) en route to the olfactorybulb (OB), the first olfactory relay in the CNS. Neuralprogenitors finish rostral migration in the core of the OBwhere they begin radial migration and mature into inter-neurons [2]. Despite extensive cellular characterization ofindividual adult-born neurons, the impact of adult neuro-genesis on OB circuit function and olfactory behavior isstill unclear. It has been argued that ongoing adult neu-rogenesis is essential for structural maintenance of the OBcircuit because blocking adult neurogenesis depletes thebulb of interneurons [3]. This review focuses on newinsights indicating that the function of adult bulbar neu-rogenesis goes beyond the mere maintenance of neuronalcircuits. In particular, we detail the functional character-istics of new neurons and how they shape mitral cellfunction and olfactory behavior.
Functions for old and new neuronsIn rodents, olfaction is a key chemosensory modality thatenables diverse essential functions such as food selection,danger detection and conspecific interactions. To fulfill thisrepertoire of functions the olfactory system has to detectand discriminate between odorants from a rich and variedolfactory environment, and then preserve this information
Review
Glossary
Bottom-up pathway: since Aristotle, the classical view of corticalprocessing proposes that sensory information evolves through afeed-forward hierarchy. In the first step of the olfactory system, thebottom-up pathway includes sensory signals from the olfactoryepithelium to the OB.Classical conditioning: first demonstrated by Pavlov [90], thisinvolves repeatedly pairing an unconditioned stimulus (which reli-ably evokes a reflexive response) with another previously neutralstimulus (which does not normally evoke the response). Followingconditioning, the response occurs both to the unconditioned stimu-lus and to the other, unrelated stimulus.Coincidence detection: a process by which a neuronal element canencode information by detecting the incidence of simultaneous yetspatially-segregated events. In general, coincidence detectionenables a combinatorial code to achieve outcomes that cannotbe generated by isolated stimuli and to increase the number ofresults attainable with a limited stimulus set. In relation to neuro-science, coincidence detection, long appreciated to play a role instrengthening synaptic connections in the mature CNS, can beimportant for establishing synaptic connections during neuralmaturation.Long-term memory: memory lasting several days to months.Long-term potentiation: an increase in size of a synaptic responsethat indicates plasticity in synaptic connections, lasting one hour ormore.Non-operant associative olfactory task: passive pairing of a contextwith an odorant stimulation that does not require the learning of aspecific behavior in order to obtain a reward.Olfactory associative memory: the memory of an association be-tween a given odorant stimulus and a reward.Olfactory enrichment: repeated exposure to novel odorants for sev-eral days.Olfactory fear-conditioning: learning an association between a neu-tral stimulus (odorant) and an aversive stimulus. The animal cannotchoose to escape the conditioned stimulus upon testing.Olfactory habituation/spontaneous discrimination task: assessmentof non-associative short-term memory (ie. spontaneous discrimina-tion among dissimilar or similar odorants, unaltered by previousreinforcement).Olfactory habituation: progressive decline in investigating repeatedsimilar odorant stimuli.Operant associative olfactory learning: a form of learning consistingof the acquisition of a particular behavior by pairing the neutralodorant stimulus with a reward.Operant conditioning: involves a learning paradigm that leads tothe modification of the occurrence and expression of a givenbehavior. Operant conditioning is distinguished from classicalconditioning in that operant conditioning concerns changes ofvoluntary behavior.Pattern segregation: the separation of neuronal cell discharge pat-terns allowing divergence from similar input patterns.Perceptual learning: a form of implicit memory that can be defined asan increased sensitivity to stimulus parameters that improves per-ceptual acuity due to experience.
Corresponding author: Lledo, P.-M. ([email protected])
20 0166-2236/$ – see front matter ! 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.tins.2010.09.006 Trends in Neurosciences, January 2011, Vol. 34, No. 1
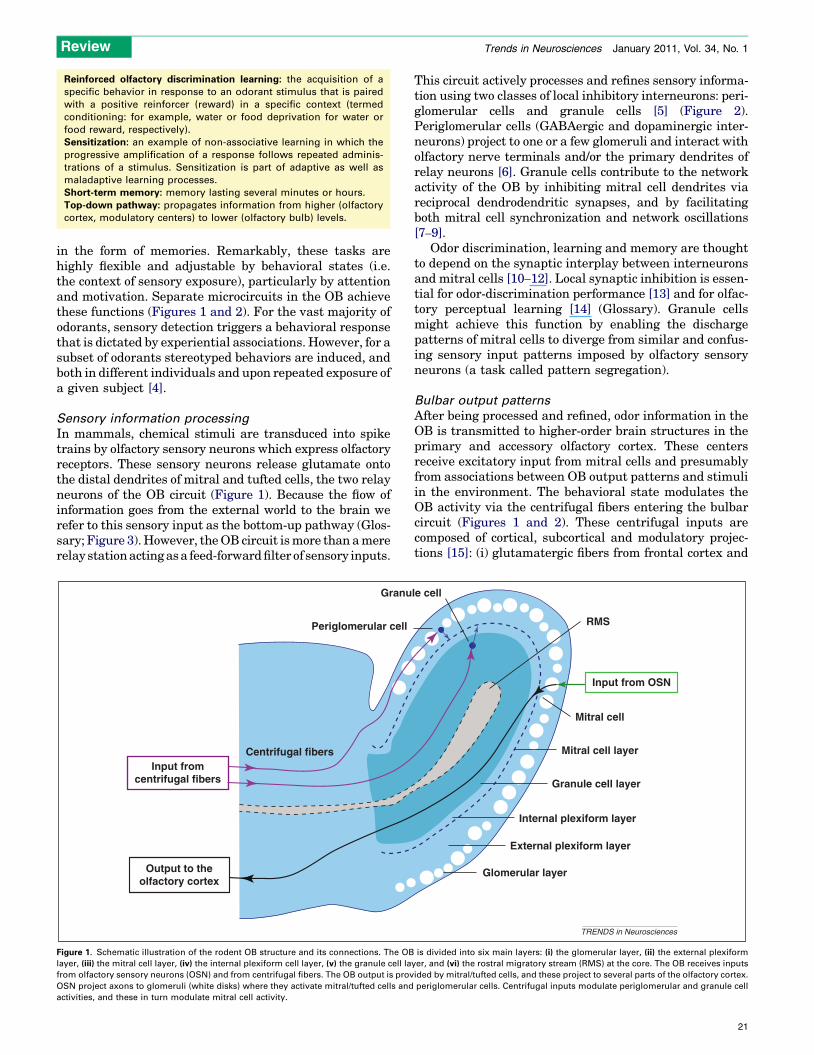
in the form of memories. Remarkably, these tasks arehighly flexible and adjustable by behavioral states (i.e.the context of sensory exposure), particularly by attentionand motivation. Separate microcircuits in the OB achievethese functions (Figures 1 and 2). For the vast majority ofodorants, sensory detection triggers a behavioral responsethat is dictated by experiential associations. However, for asubset of odorants stereotyped behaviors are induced, andboth in different individuals and upon repeated exposure ofa given subject [4].
Sensory information processingIn mammals, chemical stimuli are transduced into spiketrains by olfactory sensory neurons which express olfactoryreceptors. These sensory neurons release glutamate ontothe distal dendrites of mitral and tufted cells, the two relayneurons of the OB circuit (Figure 1). Because the flow ofinformation goes from the external world to the brain werefer to this sensory input as the bottom-up pathway (Glos-sary; Figure 3).However, theOB circuit ismore than amererelay stationactingasa feed-forwardfilter of sensory inputs.
This circuit actively processes and refines sensory informa-tion using two classes of local inhibitory interneurons: peri-glomerular cells and granule cells [5] (Figure 2).Periglomerular cells (GABAergic and dopaminergic inter-neurons) project to one or a few glomeruli and interact witholfactory nerve terminals and/or the primary dendrites ofrelay neurons [6]. Granule cells contribute to the networkactivity of the OB by inhibiting mitral cell dendrites viareciprocal dendrodendritic synapses, and by facilitatingboth mitral cell synchronization and network oscillations[7–9].
Odor discrimination, learning and memory are thoughtto depend on the synaptic interplay between interneuronsand mitral cells [10–12]. Local synaptic inhibition is essen-tial for odor-discrimination performance [13] and for olfac-tory perceptual learning [14] (Glossary). Granule cellsmight achieve this function by enabling the dischargepatterns of mitral cells to diverge from similar and confus-ing sensory input patterns imposed by olfactory sensoryneurons (a task called pattern segregation).
Bulbar output patternsAfter being processed and refined, odor information in theOB is transmitted to higher-order brain structures in theprimary and accessory olfactory cortex. These centersreceive excitatory input from mitral cells and presumablyfrom associations between OB output patterns and stimuliin the environment. The behavioral state modulates theOB activity via the centrifugal fibers entering the bulbarcircuit (Figures 1 and 2). These centrifugal inputs arecomposed of cortical, subcortical and modulatory projec-tions [15]: (i) glutamatergic fibers from frontal cortex and
Reinforced olfactory discrimination learning: the acquisition of aspecific behavior in response to an odorant stimulus that is pairedwith a positive reinforcer (reward) in a specific context (termedconditioning: for example, water or food deprivation for water orfood reward, respectively).Sensitization: an example of non-associative learning in which theprogressive amplification of a response follows repeated adminis-trations of a stimulus. Sensitization is part of adaptive as well asmaladaptive learning processes.Short-term memory: memory lasting several minutes or hours.Top-down pathway: propagates information from higher (olfactorycortex, modulatory centers) to lower (olfactory bulb) levels.
[()TD$FIG]
Input fromcentrifugal fibers
Output to theolfactory cortex
Input from OSN
Centrifugal fibers
Granule cell
Periglomerular cell RMS
Mitral cell
Mitral cell layer
Granule cell layer
Glomerular layer
External plexiform layer
Internal plexiform layer
TRENDS in Neurosciences
Figure 1. Schematic illustration of the rodent OB structure and its connections. The OB is divided into six main layers: (i) the glomerular layer, (ii) the external plexiformlayer, (iii) the mitral cell layer, (iv) the internal plexiform cell layer, (v) the granule cell layer, and (vi) the rostral migratory stream (RMS) at the core. The OB receives inputsfrom olfactory sensory neurons (OSN) and from centrifugal fibers. The OB output is provided by mitral/tufted cells, and these project to several parts of the olfactory cortex.OSN project axons to glomeruli (white disks) where they activate mitral/tufted cells and periglomerular cells. Centrifugal inputs modulate periglomerular and granule cellactivities, and these in turn modulate mitral cell activity.
Review Trends in Neurosciences January 2011, Vol. 34, No. 1
21

olfactory cortex [16,17], and (ii) modulatory projectionsfrom the locus cœruleus (norepinephrine), the horizontallimb of the diagonal band of Broca (acetylcholine), and thedorsal raphe nucleus (serotonin) [18,19]. Centrifugalinputs shape the function of the bulbar network on varioustimescales (from milliseconds to days) and in differentcontexts (see below). Interestingly, most of the centrifugalconnections target granule cells [20] and tune the dendro-dendritic inhibition of mitral cells [21,22]. On the otherhand, centrifugal serotoninergic fibers preferentially tar-get periglomerular cells and modulate odor input at thebeginning of odor information processing, acting indirectlyon olfactory sensory neuron terminals by modulating peri-glomerular inhibitory cells [23].
By modulating the activity of local interneurons, cen-trifugal inputs shape the spatial and temporal firing pat-terns of mitral cell activity, and thus are important forproducing dynamic odor representation in the context of
different behavioral states [23–27]. Much of the sensoryinput that enters the brain does so because we activelyseek sensory information through sniffing and othermeans, and because sensory investigation can often betriggered by environmental cues. In this case, centrifugalinputs to the OB provide the contextual information re-quired for sensory formatting of odor representations,learning and memory. In short, this top-down pathwayrepresents amechanism bywhich the brainmakes sense ofthe olfactory content (Figure 3). Experiments that specifi-cally manipulate the top-down pathway have revealed itsunique influence on olfaction. For instance, inhibition ofcentrifugal cholinergic inputs to the OB circuit affects theability of rodents to differentiate between similar odorants[28,29]. Similarly, experimentalmanipulation of noradren-ergic activity in the OB has resulted in changes in olfactoryperception and memory formation [30] and in olfactoryhabituation memory [31]. Decreasing glutamatergic feed-back from the olfactory peduncle to the OB has revealed akey role for centrifugal inputs in changing the formation ofodor–reward associations, but not primary bulbar odorrepresentations [18]. In this context it is noteworthy thatthe first connections of adult-born neurons are establishedwith centrifugal fibers, and these can provide excitation tothese young neurons soon after they arrive and integratewithin the OB circuit [2,32,33]. Thus, centrifugal afferentsoffer a pathway by which new neuron activity, and thusolfactory function, can be shaped by the behavioral stateand olfactory experience, including learning. An importantaim of future studies will be to characterize the detailedsynaptic mechanisms that link adult-generated neurons tocentrifugal projections.
[()TD$FIG]
Input from OSN(Sensory experience)
Centrifugal fibers (CF)(Behavioral state)
Mitral cell
PG
GC
Ouput to the olfactory cortex
TRENDS in Neurosciences
Figure 2. A canonical OB circuit. The olfactory system includes the olfactorysensory neurons (OSN) located in the sensory epithelium, the relay neurons (mitraland tufted cells) of the OB, and cortical neurons in downstream structures,comprising the primary and accessory olfactory cortex. These centers include theanterior olfactory nucleus, which connects the OB through a portion of the anteriorcommissure, the olfactory tubercle, the piriform cortex (considered to be theprimary olfactory cortex), the cortical nucleus of the amygdala, and the entorhinalarea [16,21,77]. The OB is the first relay of olfactory processing and is encoding bythree main types of neurons: (i) OSN detect odorants and transmit actionpotentials to the OB, where they form glutamatergic synapses with the dendritesof (ii) relay neurons (called mitral cells), and (iii) local interneurons in the targetarea (called glomeruli). Mitral/tufted cells are glutamatergic neurons which projectto the olfactory cortex. The activity of mitral/tufted cells is modulated byglumatergic input from OSNs, GABAergic input from interneurons called granulecells (GC), from GABAergic and dopaminergic input from interneurons calledperiglomerular cells (PG), as well as by centrifugal innervations (centrifugal fibers:CF). Unlike most CNS regions, inhibitory interneurons in the OB far outnumberprincipal cells by at least 50:1 [105]. The granule cells account for about 90% of allbulbar interneurons [105]. Periglomerular and granule cells are the primary targetsof the centrifugal inputs to the OB. Sensory experience modulates the activity ofglomeruli, mitral/tufted cells and of granule cells. Centrifugal inputs play a key rolein odor detection, discrimination and learning/memory by regulating sensoryinformation. Newly generated interneurons continuously integrate into theexisting neuronal circuit of the adult OB, and here they provide a source ofplasticity that could allow behavioral adaptation to an ever-changing environment.Adult-born neurons target mitral cells in the OB, thus providing an inhibitoryfunction. Learning modulates the survival of newborn granule cells probably viacentrifugal inputs. In this way, newborn granule cells could support long-termmemory after learning.
[()TD$FIG]
Figure 3. Regulation of OB activity. The neuronal activity of the OB circuit dependson three intermingled processes: (i) sensory inputs from the olfactory epithelium(the bottom-up pathway), (ii) centrifugal fibers (the top-down pathway), and (iii)adult neurogenesis. The olfactory epithelium provides sensory information to theOB circuit from two different behavioral contexts: passive odorant exposure oractive sensing. Centrifugal fibers contribute to the context-dependent modulationof the circuit activity (i.e. attention, reward, learning, and memory). ChallengingOB network activity influences the recruitment of new neurons, and this in turn willaffect the ongoing activity of the host circuit. Note that adult neurogenesis is alsosensitive to olfactory experience (i.e. sensory deprivation or odor-enrichedenvironment) and to the behavioral state (i.e. learning to discriminate betweentwo odorants).
Review Trends in Neurosciences January 2011, Vol. 34, No. 1
22

As discussed below, both bottom-up and top-down path-ways regulate adult neurogenesis in a tightly orchestratedmanner following temporal constraints defined by theprogression of neuroblast maturation. According to thishypothesis, new neurons could act as coincidence detectorsby appearing at the nexus between two key events: thespatial dimensionwithin the network and the developmen-tal stage of the new neurons (Figure 4).
Wiring adult-generated neurons into pre-existingcircuitsThousands of adult-born neurons arrive at theOB each dayand integrate into the existing neural circuitry within justa few days [34]. About 95% of the new cells differentiateinto granule cells, less than 3% become periglomerularcells [35], and a very small proportion give rise to gluta-matergic juxtaglomerular neurons [36]. Each subtype ofnew bulbar interneuron originates from specific neuralstem cells located in distinct regions of the SVZ–RMSsystem [37–39]. Adult-born neurons express GABAA andglutamate receptors as soon as they start migrating [40]and when entering the deeper layers of the OB [33,41,42],before gradually increasing their responsiveness to bothGABA and glutamate. Two studies on the dynamic patternof synaptogenesis onto newborn neurons entering the OBhave revealed an unexpected contribution of centrifugalafferents to new neuron maturation [33,43]. The findingthat proximal synaptic structures on the granule cell somaand dendrites form much earlier than dendrodendriticcontacts with resident mitral cells has shed light on theearly function of centrifugal fibers in adult neurogenesis.The source of the centrifugal inputs that form early con-nections remains to be determined. Revealing their iden-tity will further increase our understanding of thecontribution of top-down processing in controlling OBadult neurogenesis.
The development of input synapses in adult-formed OBneurons is largely complete by four weeks after cell birth,
and is sensitive to neuronal activity during a restrictedperiod [32,44]. One half of new neurons die during theirprocess of dendritic branching and synaptogenesis at be-tween 15 and 40 days after cell birth [44–50]. Most adult-generated neurons that survive this period of cell deathwill survive for several months and become indistinguish-able from pre-existing neurons [45,47,48,51–53], but see[46].
Having established that not only thousands of adult-born neurons integrate into the existing OB circuit eachday, but also about half of them survive, it was important todetermine if the integration of new neurons influences OB-related behaviors. The ablation of adult neurogenesisusing systemic or genetic techniques is a popular strategyfor addressing this issue. Unfortunately, no availabletechnique can specifically reduce the population of adult-generated OB neurons. All reported ablation techniquespresent both advantages and undesired off-target effects,and this could explain why discrepancies have emergedbetween these efforts (Box 1). Further studies using selec-tive and reversible ablation of adult neurogenesis shouldhelp to elucidate the respective roles of neurogenic systemsin animal behavior.
Interactions between sensory experience and adultneurogenesisTo elucidate the role of sensory inputs on adult neurogen-esis, many studies have used passive stimulation para-digms [54–56]. Passive olfactory experiences include eithertransient odorant exposure or olfactory enrichment lastingseveral days. As described below, recent findings demon-strate that some passive olfactory experiences, but not all,impact on bulbar neurogenesis (Box 2).
Effects of passive odorant exposureActivity-dependent factors regulatemany aspects of bulbarneurogenesis including synapse formation and the survivalof new neurons [44,46,49,54,55,57,58]. Whereas passive
[()TD$FIG]
Networkactivation
Spatial
Networkmaturation
Input from CFcontext-dependent
modulation
Input from OSNinformation
content
Output to the olfactory cortex
OB Coincident detection
Adult-generatedneurons
Recruitment
EliminationDevelop-mental
TRENDS in Neurosciences
Figure 4. Coincidence detection in the OB. Within the OB network the relay neurons (mitral and tufted cells) are spatially activated by sensory inputs from both the olfactorysensory neurons (OSN) and by the centrifugal fibers (CF). Soon after their arrival within the OB, adult-born neurons receive their first inputs from CF before establishingsynapses with afferent inputs (mediated by the mitral cells). Thus, newborn neurons are able to associate a context to the information content at a particular stage of theirmaturation (developmental/temporal dimension). Information processing and refinement within the OB circuitry is therefore likely to depend on both spatial anddevelopmental timing dimensions as a result of coincidence detection by adult-generated neurons. This process might participate in the encoding of the odorrepresentation, which is then sent to the olfactory cortex.
Review Trends in Neurosciences January 2011, Vol. 34, No. 1
23

short-term exposure to odorants does not change the sur-vival of new neurons [49,57,59,60], passive long-term expo-sure to an odorant-enriched environment enhances bothSVZ proliferation [39] and the survival of new granule cells[54,58]. Conversely, long-term sensory deprivation reducesthe survival of adult-born granule cells [44,55,58,61]. Inter-estingly, new cells that survive after sensory deprivationdisplay an increased density of proximal input synapses intheir unbranched apical dendrites [32]. It remains to beestablished whether adult-born granule cells survive in theabsence of sensory input because they are able to compen-sate for their excitatory drive by enhancing their synapsedensity. Similarly, adult-born periglomerular neurons dis-play activity-dependent survival. Sensory deprivationdecreases [55], whereas olfactory enrichment increases[54], the survival of distinct populations of adult-born peri-glomerular neurons, supporting the hypothesis that theirintegration rate is dependent on the magnitude of sensoryactivity [58]. Also, sensory enrichment accelerates the for-mation of glutamatergic input synapses impinging onto thenewcomers as visualized by labeling clusters of a postsyn-aptic marker [postsynaptic density (PSD) protein PSD95]genetically targeted to adult-born neurons [62].
Impact of olfactory learningOdor sampling behavior reflects not only current percep-tion but also expectation following learning rules[24,63,64], highlighting once again the importance of thetop-down pathway in processing sensory information in theOB circuit. Thus, olfactory perception is highly dependenton experience, and repeated exposures to similar odorantscan improve olfactory discrimination (a task called olfac-tory perceptual learning) in rodents and humans [65,66].Not surprisingly, adult neurogenesis also constitutes asubstrate for context-dependent modulation of odorresponses (Box 2). For example, bulbar neurogenesis
increases following olfactoryperceptual learning [67].More-over, adult-generated neurons are preferentially involved inthe processing of already experienced odorants compared tonaıve odorants [67]. Similarly, bulbar neurogenesis hasbeenobserved to increase inmice followinganactive sensoryexperience such as associative olfactory learning, but not bysimple passive exposure to odorants [49,57,59,60]. More-over, olfactory learning in mice induces clustering of new-born cells in specific regions of the granule cell layer, 5 dayspost-conditioning, in an odorant-specific manner [60]. Inthe time window when learning increases cell survival(i.e. 18–30 days after neuronal birth), new granule cellsbegin to receive glutamatergic synaptic contacts. This sug-gests that the survival of adult-generatedgranule cells couldbe sensitive to experience during the period of early synap-togenesis. Interestingly, olfactory learning induces spatialredistribution of newborn neurons, and this is conservedafter 30 days but not after 90 days post-conditioning, sug-gesting that cell death might occur in 30 to 90 day-oldnewborn cells [60]. Because learning to discriminate be-tween two odorants reduces the survival of 38/45-day-oldgranule cells [49,68], but not 65-day-old neurons [49], it ispossible that olfactory learning promotes the survival ofspecific 18–30-day-old neurons and simultaneously inducescell death of other 38–45-day-old neurons. This would ex-plain why the previous olfactory learning tasks observe nonet increase in granule cell survival [49,68].
Recent experiments demonstrate that adult-born gran-ule cell synaptogenesis is sensitive to changes in synapticinput [2,32,69]. These studies have suggested that bothsynaptogenesis and neuronal survival take place after theneuron receives a minimum threshold of excitation fromlocal glutamatergic inputs or centrifugal fibers. Furtherstudies will be needed to clarify the respective functions ofsensory-driven inputs and centrifugal fibers on the regu-lation of newborn neuron survival and synaptogenesis.
Box 1. Strategies employed to challenge adult neurogenesis
Most of the experimental approaches currently employed to studyimpaired neurogenesis use generic anti-mitotic [91] or anti-prolifera-tive [92] drug treatments, radiation, genetic manipulation [93,94], oraging mice models [95].
Treatments with antimitotic drugs, such as SVZ infusion of theantimitotic drug cytosine arabinosine (Ara-C), are useful for haltingadult SVZ neurogenesis [60,67,75]. However, these treatments areunspecific because they target all dividing cells, including neuro-vascular cell types and oligodendrocyte precursor cells [96]. Inaddition, Ara-C is also toxic for non-dividing cells such asoligodendrocytes [96] and post-mitotic neurons [97]. Although noadverse side-effects were observed at low doses [67,75], slightlyhigher doses were observed to cause weight loss, bone marrowsuppression, cerebellar neurotoxicity and reduction in motorcoordination. In addition, antimitotic treatment probably reducescell division throughout a large portion of the brain, even when thedrug is focally infused into the parenchyma of the SVZ or into thecerebrospinal fluid of the lateral ventricle. For instance, a 30–50%reduction in hippocampal neurogenesis was recently observed afterinfusion of Ara-C into the ventricles, thereby demonstrating that itseffects are not limited to the SVZ, [60,75]. Remarkably, strongcognitive deficits have been reported to result from a similarreduction in hippocampal neurogenesis [98]. Finally, treatment withantimitotic agents might disrupt the continuous production ofolfactory sensory neurons which are necessary for olfactoryfunction perception [99].
An alternative way to disrupt adult neurogenesis uses focalirradiation. Region-restricted irradiation is more specific than focalinfusion of antimitotic drugs because only the irradiated area, and notneighboring areas, exhibits a decrease in neurogenesis [50,72,89,100].However, irradiation affects all dividing cells, including oligodendro-cyte cell precursors [101] and some subtypes of neurovascular cells[102]. It also induces strong microglial activation, a hallmark ofneuroinflammation, lasting for two months [103]. It is thereforecrucial to perform behavioral tests several months after irradiation.
Genetic manipulations are often used to study postnatal neurogen-esis. Studies using traditional knockout of genes essential forneurogenesis are interesting, but often result in conflicting data onolfactory behavior [84,93–95]. Their interpretations are often limited byabnormalities of the whole brain structure or compensatory effectselicited during development. A better way to suppress neurogenesisgenetically is by inducible recombination [98] or conditional transgenicablation of neural stem cells [3,104,106] or of neuronal precursor cells[107]. These genetic approaches are less specific than focal irradiationor antimitotic drug treatment because bulbar and hippocampalneurogenesis are both ablated, thus limiting the interpretation ofbehavioral analyses.
Mouse models of aging have also been used to study decreasedneurogenesis [95]. However, it is difficult to interpret unambiguouslywhether sensory deficits found in these models result from a specificdecrease of bulbar neurogenesis or to one (or more) of the manyother impairments observed in the aging brain.
Review Trends in Neurosciences January 2011, Vol. 34, No. 1
24

In particular, it will be important to decipher the mecha-nism by which some adult-generated neurons are ‘primed’to survive following a learning task that involves the top-down pathway.
Functions of adult-born neurons in the olfactory bulbcircuitThe potential functions of adult-born neurons in the OBcan be summarized into fourmain tasks: (i) maintenance of
[()TD$FIG]
Operantassociative
olfactory learning
Non operantassociative
olfactory learning
Acquisitionof a specific behaviorfor getting the reward
Specific context:pairing of a neutralodorant stimulus
with a reward
Effect on bulbar neurogenesis:increased survival of newborn neuronsselection of newly generated neurons
supporting long term memory
Simple exposureto odorants
Specific context:lasting minutes to hours
Olfactory enrichment
Specific context:lasting several days
with repeated exposuresof novel odorants
Effect on bulbar neurogenesis:increased survival of newborn neurons
increased short term memory
No effect on bulbar neurogenesis
No effect on bulbar neurogenesis
Olfactory perceptuallearning
Specific context:lasting several days
with repeated exposuresof similar odorants
Effect on bulbar neurogenesis:increased survival of newborn neurons
improved odorant discrimination
TRENDS in Neurosciences
Figure I. Consequences of sensory experience on adult OB neurogenesis. Some olfactory experiences, but not all, impact upon bulbar neurogenesis. In particular,cumulative evidence suggests that both the associative and non-associative forms of olfactory learning require adult neurogenesis.
Box 2. Operant associative olfactory learning versus non-operant olfactory learning
Learning can be divided into distinct categories, including non-associative forms such as habituation and sensitization, in which ananimal learns about the properties of a single stimulus, andassociative forms in which an animal learns about the relationshipbetween two stimuli (classical conditioning) or between a stimulusand the animal’s own behavior (operant conditioning).
If when an animal first responds to a stimulus it is neither rewardingnor harmful, the animal then reduces subsequent responses. Forexample, a short-term exposure of an adult rat to an odorant, in theabsence of any paired reinforcement, leads to a reduction in theproportion of mitral cells responding to that odorant with increasedfiring rates. This process, called habituation, reflects the progressivediminution of behavioral response probability with repetition stimulus.
A central question is whether these different forms of learningdepend on adult neurogenesis. When applied to olfaction, do theassociative and non-associative forms of learning require the contin-uous arrival of newborn olfactory neurons? This indeed seems to be thecase (Figure I). For example, increasing newborn neuron survival in theOB by non-associative olfactory experience leads to stronger short-term olfactory memory [54]. Similarly, enrichment to odors improvesolfactory discrimination in adult rats [108] through a non-associativeprocess called perceptual learning that requires intact adult neurogen-
esis [67]. Also, an olfactory memory deficit was reported in aspontaneous short-termmemory paradigm after suppression of bulbarneurogenesis [75].
Adult-born neurons are also at play in associative olfactory learning.When adult neurogenesis is suppressed in the adult cricket botholfactory learning and memory tested in an operant olfactoryconditioning are impaired [82]. Similarly, correlative studies performedin rodents indicate a function of adult neurogenesis in associativelearning [49,57,68]. Also, blocking of neurogenesis by irradiation oranti-mitotic treatment in mice impairs long-term retention of anassociative olfactory task but not its acquisition [50,60]. Once selectedto survive by learning, newborn neurons could change the neuralrepresentation of the learned odorants by reshaping the physiology ofthe OB network. Following this view, it can be postulated that newbornneurons might be able to retain a memory trace of previous olfactoryexperience during an associative learning task. Finally, no deficit inolfactory memory was reported when adult neurogenesis was reducedby genetic ablation [3]. This discrepancy might result from technicalissues (specific context) and/or from the degree of difficulty of thediscrimination tasks that were used. Further experiments are requiredto clarify the relationship between the degree of adult neurogenesisand the complexity of discrimination learning tasks.
Review Trends in Neurosciences January 2011, Vol. 34, No. 1
25

OB circuitry, (ii) shaping sensory information processing,(iii) supporting learning processes, and/or (iv) mediatingodor memory. Below we discuss these non-exclusiveoptions.
The contribution of adult neurogenesis to circuitmaintenanceIn the developing OB most newborn granule cells reside inthe superficial granule cell layer and can survive for thelifetime of the animal [47]. This phenomenon is reversedduring adulthood, although the precise rate of neuronalturnover is a matter of debate. Some studies estimate thatthere is turnover of only a small proportion (!15%) of thetotal population of interneurons in the granule cell layer[70,71]. Others suggest that most granule cells locateddeeper in the granule cell layer and half of the superficialones are subject to continuous turnover [3,72]. Theseinconsistencies could result from different methodologiesand should be clarified in the future using differentmodels.Interestingly, cell survival seems to depend on the quantityof newly produced neurons that reach the OB: when bulbarneurogenesis is reduced after SVZ irradiation the survivalrate of the newborn neurons reaching the bulb is signifi-cantly prolonged [50] (but see also [72]). This observationcould reflect competition between adult-generated neuronsfor survival factors released by the neural targets, just asthey compete with existing neurons for innervation viasynaptic projections.
Impact of adult neurogenesis on shaping sensoryinformation processingAdult-born neurons target mitral and tufted cells in theOB, providing an additional inhibitory function to thebulbar circuit. However, a preliminary study suggests thatadult-born neurons can also target local inhibitory inter-neurons, yielding disinhibitory function in the OB circuitry[73]. Thus, the contribution of adult-generated neurons tobulbar physiology remains unclear and conflicting resultshave been reported regarding the functional outcomes ofadult neurogenesis for olfaction [72,74,75]. One studyfound that neurogenesis ablation by irradiation had noimpact on bulbar activity [72]. By contrast, another studybased on intraventricular infusion of the antimitotic agentAra-C reported strong changes in mitral and tufted cellactivity induced by depleting adult-born neurons, leadingto impairment of short-term olfactory memory [75]. Thecontrasting results obtained in these two studies highlightthe need for further investigations to decipher the role ofadult-generated neurons inmodulatingmitral cell activity.Finally, it is worth noting that a correlation between theturnover rate of adult bulbar neurogenesis and olfactorydiscrimination time was recently established [74]. Thisstudy used a pan-caspase inhibitor to impair the naturalelimination of adult-born granule cells, and found that thetime required to discriminate between two similar odor-ants was subsequently increased. Therefore, the continu-ous recruitment of adult-born neurons can lead to changesin the strength of the temporal binding of signals originat-ing from different odorant receptors. This might be crucialnot only for adjusting odor discrimination, but also forperceptual learning as described below [76].
Impact of adult neurogenesis on learning processesPrevious studies have demonstrated that active sensoryprocessing results in perceptual learning, associativelearning, and memory formation, through changes in OBnetwork activity [59]. Adult-generated neurons integratedinto the OB circuitry seem not to be involved in perceptionper se (Box 3), but instead in cognitive function (Box 2). Forinstance, manipulation of bulbar neurogenesis revealedthat the supply of new neurons is not necessary for odorantdetection, discrimination or associative olfactory learning[3,50,60]. On the other hand, adult OB neurogenesis isrequired for olfactory fear-conditioning [72], olfactory per-ceptual learning [67] and long-term olfactory memory ofassociative olfactory learning [50,60].
Developing adult-generated interneurons are uniquein that their connectivity adapts to the degree of neuro-nal activity in the host circuit [2,32,51,77,78]. They alsotrigger unique responses during odorant familiarization[79], suggesting a key role in olfactory processing andplasticity. This idea has received recent support from anelectrophysiological study [80]. It was found that, soonafter synaptic integration, adult-born neurons displayrobust long-term potentiation not found in pre-existingneurons. This ability is progressively lost as the cellsmature over time. These findings are similar to thosereported for adult-generated dentate gyrus neuronswhich exhibit synaptic plasticity that fades with time[81]. Interestingly, the synaptic plasticity of young adult-born OB interneurons occurs during a time-windowwhen new neurons are initially added to the circuit,indicating that they might play a role in circuit plasticityrequired for learning. Although synaptic development ismostly completed by four weeks after cell birth[42,43,46,48,51], spines can still undergo rearrangementafter this period, indicating that mature OB interneur-ons still maintain some capacity for synaptic modifica-tion [51]. Thus, the precise stages at which new OBneurons play an important role in olfaction remain tobe characterized.
Box 3. Role of adult neurogenesis in olfactory
discrimination
Olfactory discrimination is a primary function of OB granule cells[13]. Because adult-generated granule cells are a significantcomponent of the bulbar interneuron population, it was notsurprising to find a link between the turnover rate of adult bulbarneurogenesis and olfactory reaction time [74]. But, surprisingly,most experiments aimed at manipulating adult bulbar neurogenesishave failed to demonstrate a direct role of adult neurogenesis inodorant discrimination [3,50,72,75]. Olfactory discrimination isclearly impaired in animals in which bulbar neurogenesis has beenaltered during embryogenesis [84,93,94], but not later duringadulthood [3,50,72,75], suggesting that this task is not mediatedby adult-generated neurons. The function of newborn neurons inodorant discrimination might be difficult to unveil because of theirrelatively small proportion compared to the population size ofmature interneurons (but see [3]). This difficulty could also be due tothe eventual development of compensatory effects such as synapticchanges in mature neurons, or to the current methods used tochallenge adult neurogenesis (Box 1). Designing experiments thatselectively activate or inhibit new neurons within the adult OBduring odorant discrimination tasks of differing complexity will berequired to solve this conundrum.
Review Trends in Neurosciences January 2011, Vol. 34, No. 1
26

Role of adult neurogenesis in olfactory memoryThe recruitment of new neurons in the OB circuitry mightallow the association of otherwise temporally distinctinput signals originating from bottom-up and/or top-downconnections (Figure 4). Interestingly, newly-formed neu-rons receive first synapses from centrifugal fibers beforeestablishing contact with afferent inputs frommitral cells[33,43]. Through these early contacts with the top-downpathway, newborn neurons are positioned to provide atop-down context to olfactory information. In addition,these early contacts could support memory based on inte-grating recursive signaling from the olfactory cortex andtheOB. In linewith this hypothesis, a correlation has beenfound between the degree of bulbar neurogenesis andshort-term olfactory memory [54,82,83]. Moreover, block-ing bulbar neurogenesis before and during odor enrich-ment prevents olfactory perceptual learning, and thusmemory formation of experienced odorants in mice [67].Early contacts with top-down fibers offer a mechanism bywhich odor experience increases newborn granule cellresponsiveness and improves olfactory discriminationlearning [14,57,67]. Overall, mounting evidence supportsthe hypothesis that adult-generated neurons are requiredin the formation and/or shaping of olfactory memorycircuits.
Halting adult neurogenesis by radiation or Ara-C treat-ment also modifies long-term odor associative memory[50,60]. By contrast, long-term memory has been shownto be unaffected in mice producing a neuron-specific eno-lase–diphtheria toxin [3], and in mice treated with Ara-C[75], despite the significant neurogenic reduction in the twolatter studies. Details of the experimental design couldunderlie these discrepancies, including the strains of ani-mal models used, off-target effects of the ablationmethods,and the behavioral paradigms used. Considering the latteroption it is worth noting that some studies have employednon-operant tasks [3,75] whereas other studies have usedoperant tasks [50,60]. Operant conditioning differs fromnon-operant conditioning in that the animal is motivatedto acquire a particular behavior to complete its task. Non-operant and operant conditioning paradigms for investi-gating olfactory abilities often lead to different conclusions,even from the same animal model, indicating they mightsupport different olfactory functions [84–86]. The discrep-ancy between the two learning paradigms could arise fromthe participation of top-down centrifugal fibers broughtinto play only during operant conditioning.
Together, these data indicate that adult-born neuronsare required for long-term memory of associative olfactorytasks involving active learning [50,60], but not for non-operant associative training [3,75] (but see [72]). Thesefindings also indicate that the need for adult-born neuronscould depend on the nature of the task. A recent studyshows that the procerebrum of invertebrates, which isanalogous to the mammalian OB, represents the storagesite for associative olfactory memories and is directlyinvolved in the memory retrieval process [87]. In theOB, adult-generated neurons could be the primary targetsfor learning-induced changes originating from sensoryinputs (the sensory space) and centrifugal fibers (the in-ternal state), and could act as key elements in the recall of
memory traces (Figure 4). Similarly, it was proposed thatadult hippocampal neurogenesis in rodents plays a role inmemory storage, rather than during the learning process[88]. Interestingly, as in the OB, developing immaturehippocampal neurons might serve as pattern separators[89]. This function is particularly important for the accu-racy of memory encoding. Pattern separation is also essen-tial for the formation and use of memories derived fromstimuli that are close in space or time.
Concluding remarksOlfactory sensory processing begins in sensory neuronsthat transform chemical signaling into electrical patterns.This stage is unlikely to change over time during ananimal’s experience. Mitral cells then distribute this neu-ral activity widely to downstream structures, and areinfluenced by a combination of sensory input and behav-ioral state. This second transformation step constitutes atarget not only for adult neurogenesis but also for top-downconnections. Because the ability to process sensory infor-mation depends on the functional architecture and synap-tic connectivity of the OB, adult neurogenesis has aparticular impact on information processing. Understand-ing the precise functions of adult bulbar neurogenesis willrequire elucidation of the mechanisms that control thedevelopment of both synaptic inputs and outputs of theadult-born OB interneurons, and how these precisesequences impact on olfactory behavior.
The activity-dependent regulation of neurogenesis andexperience-dependent participation of newneurons in olfac-tory information processing both highlight the relationshipbetween adult neurogenesis and olfactory abilities. In thisreview we have highlighted the potential functions of newneurons as follows: (i),maintaining OB circuitry – newbornneurons could replace dying interneurons in the OB, thusplaying a structural role in network maintenance. (ii) Tun-ing information processing – adult-born neurons targetmitral cells in the OB, thus providing further inhibitoryactivity necessary for sharp pattern-segregation. In somecircumstances itmightbevital todecorrelateandmodify theodor representation that will ultimately be sent to down-stream brain structures. (iii) Role in learning – becauseadult-born neurons express unique synaptic plasticity,these neurons could be more suited than mature neuronsto encoding new information. (iv) Role in memory – asproposed here, newborn neurons could provide significantcontext to the information content due to their role ascoincidencedetectors.Even in theabsence of sensory inputs,newborn neurons could be driven by centrifugal fibers, afeature that could represent part of a memory process.
Accumulating evidence suggests that modifications toadult neurogenesis alter olfactory processing in diverseways. It will be of great interest to explore precisely at whatstage of development the adult-generated OB neuronscontribute to olfactory perception, odor memory, or fearresponses. Are new neurons only functionally importantwhen they are young and more excitable? Or are they mostimportant later when they are mature and are fully inte-grated into the existing network? Also, future studies willneed to selectively target distinct populations of adult-borninterneurons (e.g. periglomerular cells versus granule
Review Trends in Neurosciences January 2011, Vol. 34, No. 1
27

cells). So far the role of new periglomerular cells in olfac-tory information processing remains unknown. Finally,further studies on the relationship/cross-talk betweenadult-generated neurons and centrifugal projections willbe helpful in unraveling the cellular and molecularmechanisms of odor discrimination and those involved informing olfactory traces during learning and reconstitut-ing them during memory recall. Clarification of theseissues will not only highlight how OB neurogenesisimpacts upon olfaction but will guide the future develop-ment of cell therapies for neurological disorders (Box 4).
AcknowledgmentsOur lab is supported by the Agence Nationale de la Recherche (ANR-2007SEST-01411), the life insurance company ‘AG2R-La Mondiale’, the Ecoledes Neurosciences de Paris (ENP) and the Fondation pour la RechercheMedicale ‘Equipe FRM’. We thank Beatrice de Cougny for technicalassistance with Figure 1, and Matthew Valley and Gabriel Lepousez forcomments on the manuscript.
References1 Kriegstein, A. and Alvarez-Buylla, A. (2009) The glial nature of
embryonic and adult neural stem cells. Annu. Rev. Neurosci. 32,149–184
2 Kelsch, W. et al. (2010) Watching synaptogenesis in the adult brain.Annu. Rev. Neurosci. 33, 131–149
3 Imayoshi, I. et al. (2008) Roles of continuous neurogenesis in thestructural and functional integrity of the adult forebrain. Nat.Neurosci. 11, 1153–1161
4 Stowers, L. and Logan, D.W. (2010) Olfactory mechanisms ofstereotyped behavior: on the scent of specialized circuits. Curr.Opin. Neurobiol. 20, 274–280
5 Wilson, R.I. (2008) Neural and behavioral mechanisms of olfactoryperception. Curr. Opin. Neurobiol. 18, 408–412
6 Kosaka, K. and Kosaka, T. (2005) Synaptic organization of theglomerulus in the main olfactory bulb: compartments of theglomerulus and heterogeneity of the periglomerular cells. Anat.Sci. Int. 80, 80–90
7 Bathellier, B. et al. (2006) Circuit properties generating gammaoscillations in a network model of the olfactory bulb. J. Neurophysiol.95, 2678–2691
8 Lagier, S. et al. (2007) GABAergic inhibition at dendrodendriticsynapses tunes gamma oscillations in the olfactory bulb. Proc.Natl. Acad. Sci. U. S. A. 104, 7259–7264
9 Geffen, M.N. et al. (2009) Neural encoding of rapidly fluctuating odors.Neuron 61, 570–586
10 Yokoi, M. et al. (1995) Refinement of odor molecule tuning bydendrodendritic synaptic inhibition in the olfactory bulb. Proc.Natl. Acad. Sci. U. S. A. 92, 3371–3375
11 Laurent, G. (2002) Olfactory network dynamics and the coding ofmultidimensional signals. Nat. Rev. Neurosci. 3, 884–895
12 Lledo, P.M. and Lagier, S. (2006) Adjusting neurophysiologicalcomputations in the adult olfactory bulb. Semin. Cell Dev. Biol. 17,443–453
13 Abraham, N.M. et al. (2010) Synaptic inhibition in the olfactory bulbaccelerates odor discrimination in mice. Neuron 65, 399–411
14 Mandairon, N. et al. (2008) Odor enrichment increases interneuronsresponsiveness in spatially defined regions of the olfactory bulbcorrelated with perception. Neurobiol. Learn. Mem. 90, 178–184
15 Shipley, M.T. and Ennis, M. (1996) Functional organization ofolfactory system. J. Neurobiol. 30, 123–176
16 Shepherd, G.M. et al. (2004) Olfactory Bulb. In The SynapticOrganization of the Brain (Shepherd, G.M., ed.), pp. 165–217,Oxford University Press
17 Willhite, D.C. et al. (2006) Viral tracing identifies distributedcolumnar organization in the olfactory bulb. Proc. Natl. Acad. Sci.U. S. A. 103, 12592–12597
18 Kiselycznyk, C.L. et al. (2006) Role of centrifugal projections to theolfactory bulb in olfactory processing. Learn. Mem. 13, 575–579
19 Mouret, A. et al. (2009) Centrifugal drive onto local inhibitoryinterneurons of the olfactory bulb.Ann. N. Y. Acad. Sci. 1170, 239–254
20 Macrides, F. et al. (1981) Cholinergic and catecholaminergic afferentsto the olfactory bulb in the hamster: a neuroanatomical, biochemical,and histochemical investigation. J. Comp. Neurol. 203, 495–514
21 Wilson, R.I. and Mainen, Z.F. (2006) Early events in olfactoryprocessing. Annu. Rev. Neurosci. 29, 163–201
22 Strowbridge, B.W. (2009) Role of cortical feedback in regulatinginhibitory microcircuits. Ann. N. Y. Acad. Sci. 1170, 270–274
23 Petzold, G.C. et al. (2009) Serotonergic modulation of odor input to themammalian olfactory bulb. Nat. Neurosci. 12, 784–791
24 Kay, L.M. and Laurent, G. (1999) Odor- and context-dependentmodulation of mitral cell activity in behaving rats. Nat. Neurosci.2, 1003–1009
25 Buonviso, N. and Chaput, M. (2000) Olfactory experience decreasesresponsiveness of the olfactory bulb in the adult rat. Neuroscience 95,325–332
26 Martin, C. et al. (2004) Learning modulation of odor-inducedoscillatory responses in the rat olfactory bulb: a correlate of odorrecognition? J. Neurosci. 24, 389–397
27 Restrepo, D. et al. (2009) From the top down: flexible reading of afragmented odor map. Trends Neurosci. 32, 525–531
28 Linster, C. et al. (2001) Selective loss of cholinergic neurons projectingto the olfactory system increases perceptual generalization betweensimilar, but not dissimilar, odorants. Behav. Neurosci. 115, 826–833
29 Mandairon, N. et al. (2006) Cholinergic modulation in the olfactorybulb influences spontaneous olfactory discrimination in adult rats.Eur. J. Neurosci. 24, 3234–3244
30 Doucette, W. et al. (2007) Adrenergic modulation of olfactory bulbcircuitry affects odor discrimination. Learn. Mem. 14, 539–547
31 Guerin, D. et al. (2008) Noradrenergic neuromodulation in theolfactory bulb modulates odor habituation and spontaneousdiscrimination. Behav. Neurosci. 122, 816–826
32 Kelsch, W. et al. (2009) A critical period for activity-dependentsynaptic development during olfactory bulb adult neurogenesis. J.Neurosci. 29, 11852–11858
33 Panzanelli, P. et al. (2009) Early synapse formation in developinginterneurons of the adult olfactory bulb. J. Neurosci. 29, 15039–15052
34 Alvarez-Buylla, A. et al. (2001) A unified hypothesis on the lineage ofneural stem cells. Nat. Rev. Neurosci. 2, 287–293
35 Lledo, P.M. et al. (2006) Adult neurogenesis and functional plasticityin neuronal circuits. Nat. Rev. Neurosci. 7, 179–193
36 Brill, M.S. et al. (2009) Adult generation of glutamatergic olfactorybulb interneurons. Nat. Neurosci. 12, 1524–1533
37 Florian, T. et al. (2007) Mosaic organization of neural stem cells in theadult brain. Science 317, 381–384
38 Young, K.M. et al. (2007) Subventricular zone stem cells areheterogeneous with respect to their embryonic origins and
Box 4. Outstanding questions
" At what stage of development do adult-generated OB neuronscontribute to olfactory perceptual learning, odor memory, andfear responses?
" How does the orchestrated development of the synaptic inputsonto adult-born bulbar neurons impact upon olfactory abilities?
" Are new neurons functionally important when they are young andmore excitable? Or are they most important later when they aremature and fully integrated into the existing network?
" Is there any role for new periglomerular cells in forming odormemories?
" What is the relationship/cross-talk between adult-generatedneurons and centrifugal projections?
" What are the cellular and molecular mechanisms underlyingolfactory perceptual learning and odor discrimination?
" What are the mechanisms involved in forming olfactory tracesduring learning and in reconstituting them during recall?
" What is the contribution of adult neurogenesis to pheromone-related behaviors such as mating, social recognition and maternalbehavior?
" What is the contribution of the constitutive adult bulbar neuro-genesis to brain repair occurring during brain trauma, ischemia,and neurodegenerative or inflammatory diseases?
Review Trends in Neurosciences January 2011, Vol. 34, No. 1
28

neurogenic fates in the adult olfactory bulb. J. Neurosci. 27, 8286–8296
39 Alonso, M. et al. (2008) Turning astrocytes from the rostral migratorystream into neurons: a role for the olfactory sensory organ. J.Neurosci. 28, 11089–11102
40 Platel, J.C. et al. (2010) NMDA receptors activated by subventricularzone astrocytic glutamate are critical for neuroblast survival prior toentering a synaptic network. Neuron 65, 859–7275
41 Belluzzi, O. et al. (2003) Electrophysiological differentiation of newneurons in the olfactory bulb. J. Neurosci. 23, 10411–10418
42 Carleton, A. et al. (2003) Becoming a new neuron in the adult olfactorybulb. Nat. Neurosci. 6, 507–518
43 Whitman, M.C. and Greer, C.A. (2007) Synaptic integration of adult-generated olfactory bulb granule cells: basal axodendritic centrifugalinput precedes apical dendrodendritic local circuits. J. Neurosci. 27,9951–9961
44 Yamaguchi, M. and Mori, K. (2005) Critical period for sensoryexperience-dependent survival of newly generated granule cells inthe adult mouse olfactory bulb. Proc. Natl. Acad. Sci. U. S. A. 102,9697–9702
45 Winner, B. et al. (2002) Long-term survival and cell death of newlygenerated neurons in the adult rat olfactory bulb.Eur. J. Neurosci. 16,1681–1689
46 Petreanu, L. and Alvarez-Buylla, A. (2002) Maturation and deathof adult-born olfactory bulb granule neurons: role of olfaction. J.Neurosci. 22, 6106–6113
47 Lemasson, M. et al. (2005) Neonatal and adult neurogenesis providetwo distinct populations of newborn neurons to the mouse olfactorybulb. J. Neurosci. 25, 6816–6825
48 Kelsch, W. et al. (2008) Sequential development of synapses indendritic domains during adult neurogenesis. Proc. Natl. Acad. Sci.U. S. A. 105, 16803–16808
49 Mouret, A. et al. (2008) Learning and survival of newly generatedneurons: when time matters. J. Neurosci. 28, 11511–11516
50 Lazarini, F. et al. (2009) Cellular and behavioral effects of cranialirradiation of the subventricular zone in adult mice. PLoS One 4,e7017
51 Mizrahi, A. (2007) Dendritic development and plasticity of adult-bornneurons in the mouse olfactory bulb. Nat. Neurosci. 10, 444–452
52 Whitman, M.C. and Greer, C.A. (2007) Adult-generated neuronsexhibit diverse developmental fates. Dev. Neurobiol. 67, 1079–1093
53 Grubb, M.S. et al. (2008) Functional maturation of the first synapse inolfaction: development and adult neurogenesis. J. Neurosci. 28, 2919–2932
54 Rochefort, C. et al. (2002) Enriched odor exposure increases thenumber of newborn neurons in the adult olfactory bulb andimproves odor memory. J. Neurosci. 22, 2679–2689
55 Mandairon, N. et al. (2006) Long-term fate and distribution ofnewborn cells in the adult mouse olfactory bulb: influences ofolfactory deprivation. Neuroscience 141, 443–451
56 Escanilla, O. et al. (2008) Odor–reward learning and enrichment havesimilar effects on odor perception. Physiol. Behav. 94, 621–626
57 Alonso, M. et al. (2006) Olfactory discrimination learning increasesthe survival of adult-born neurons in the olfactory bulb. J. Neurosci.26, 10508–10513
58 Bovetti, S. et al. (2009) Olfactory enrichment influences adultneurogenesis modulating GAD67 and plasticity-related moleculesexpression in newborn cells of the olfactory bulb. PLoS One 4, e6359
59 Mandairon, N. and Linster, C. (2009) Odor perception and olfactorybulb plasticity in adult mammals. J. Neurophysiol. 101, 2204–2209
60 Sultan, S. et al. (2010) Learning-dependent neurogenesis in theolfactory bulb determines long-term olfactory memory. FASEB J.24, 2355–2363
61 Mandairon, N. et al. (2003) Deprivation of sensory inputs to theolfactory bulb up-regulates cell death and proliferation in thesubventricular zone of adult mice. Neuroscience 119, 507–516
62 Livneh, Y. et al. (2009) Sensory input enhances synaptogenesis ofadult-born neurons. J. Neurosci. 29, 86–97
63 Rinberg, D. et al. (2006) Sparse odor coding in awake behavingmice. J.Neurosci. 26, 8857–8865
64 Kepecs, A. et al. (2007) Rapid and precise control of sniffing duringolfactory discrimination in rats. J. Neurophysiol. 98, 205–213
65 Davis, R.L. (2004) Olfactory learning. Neuron 44, 31–48
66 Wilson, D.A. and Stevenson, R.J. (2003) The fundamental role ofmemory in olfactory perception. Trends Neurosci. 26, 243–247
67 Moreno, M.M. et al. (2009) Olfactory perceptual learning requiresadult neurogenesis. Proc. Natl. Acad. Sci. U. S. A. 106, 17980–17985
68 Mandairon, N. et al. (2006) Neurogenic correlates of an olfactorydiscrimination task in the adult olfactory bulb. Eur. J. Neurosci.24, 3578–3588
69 Lin, C.W. et al. (2010) Genetically increased cell-intrinsic excitabilityenhances neuronal integration into adult brain circuits. Neuron 65,32–39
70 Ninkovic, J. et al. (2007) Distinct modes of neuron addition in adultmouse neurogenesis. J. Neurosci. 27, 10906–10911
71 Lagace, D.C. et al. (2007) Dynamic contribution of nestin-expressingstem cells to adult neurogenesis. J. Neurosci. 27, 12623–12629
72 Valley, M.T. et al. (2009) Ablation of mouse adult neurogenesis altersolfactory bulb structure and olfactory fear conditioning. Front.Neurogenet. 3, 51
73 Bardy, C., et al. (2009) Deciphering the output of adult-born neuronsin the olfactory bulb circuit with channelrhodopsin2. Program No.12.6, 2009 Neuroscience Meeting Planner, Society for Neuroscience
74 Mouret, A. et al. (2009) Turnover of newborn olfactory bulb neuronsoptimizes olfaction. J. Neurosci. 29, 12302–12314
75 Breton-Provencher, V. et al. (2009) Interneurons produced inadulthood are required for the normal functioning of the olfactorybulb network and for the execution of selected olfactory behaviors. J.Neurosci. 29, 15245–15257
76 Wilson, D.A. and Stevenson, R.J. (2003) Olfactory perceptuallearning: the critical role of memory in odor discrimination.Neurosci. Biobehav. Rev. 27, 307–328
77 Lledo, P.M. and Saghatelyan, A. (2005) Integrating new neurons intothe adult olfactory bulb: joining the network, life-death decisions, andthe effects of sensory experience. Trends Neurosci. 28, 248–254
78 Adam, Y. and Mizrahi, A. (2010) Circuit formation and maintenance–perspectives from the mammalian olfactory bulb. Curr. Opin.Neurobiol. 20, 134–140
79 Magavi, S.S. et al. (2005) Adult-born and preexisting olfactory granuleneurons undergo distinct experience-dependent modifications of theirolfactory responses in vivo. J. Neurosci. 25, 10729–10739
80 Nissant, A. et al. (2009) Adult neurogenesis promotes synapticplasticity in the olfactory bulb. Nat. Neurosci. 12, 728–730
81 Ge, S. et al. (2007) A critical period for enhanced synaptic plasticity innewly generated neurons of the adult brain. Neuron 54, 559–566
82 Scotto-Lomassese, S. et al. (2003) Suppression of adult neurogenesisimpairs olfactory learning andmemory in an adult insect. J. Neurosci.23, 9289–9296
83 Rochefort, C. and Lledo, P.M. (2005) Short-term survival of newbornneurons in the adult olfactory bulb after exposure to a complex odorenvironment. Eur. J. Neurosci. 22, 2863–2870
84 Gheusi, G. et al. (2000) Importance of newly generated neurons inthe adult olfactory bulb for odor discrimination. Proc. Natl. Acad. Sci.U. S. A. 97, 1823–1828
85 Linster, C. et al. (2002) Spontaneous versus reinforced olfactorydiscriminations. J. Neurosci. 22, 6842–6845
86 Schellinck, H.M. et al. (2004) Neural cell adhesion molecule (NCAM)null mice do not show a deficit in odour discrimination learning.Behav. Brain Res. 152, 327–334
87 Matsuo, R. et al. (2010) Spontaneous recovery of the injured higherolfactory center in the terrestrial slug limax. PLoS One 5, e9054
88 Deng, W. et al. (2009) Adult-born hippocampal dentate granule cellsundergoing maturation modulate learning and memory in the brain.J. Neurosci. 29, 13532–13542
89 Clelland, C.D. et al. (2009) A functional role for adult hippocampalneurogenesis in spatial pattern separation. Science 325, 210–213
90 Pavlov, I.P. (1927/1960).Conditional Reflexes, Dover Publications (the1960 edition is an unaltered republication of the 1927 translation byOxford University Press)
91 Doetsch, F. et al. (1999) Regeneration of a germinal layer in theadultmammalian brain.Proc.Natl. Acad. Sci.U.S.A.96, 11619–11624
92 Crandall, J. et al. (2004) 13-cis-retinoic acid suppresses hippocampalcell division and hippocampal-dependent learning in mice. Proc. Natl.Acad. Sci. U. S. A. 101, 5111–5116
93 Kim, W.R. et al. (2007) Impaired migration in the rostral migratorystream but spared olfactory function after the elimination of
Review Trends in Neurosciences January 2011, Vol. 34, No. 1
29

programmed cell death inBax knock-outmice. J. Neurosci. 27, 14392–14403
94 Bath, K.G. et al. (2008) Variant brain-derived neurotrophic factor(Val66Met) alters adult olfactory bulb neurogenesis and spontaneousolfactory discrimination. J. Neurosci. 28, 2383–2393
95 Enwere, E. et al. (2004) Aging results in reduced epidermal growthfactor receptor signaling, diminished olfactory neurogenesis, anddeficits in fine olfactory discrimination. J. Neurosci. 24, 8354–8365
96 Dietrich, J. et al. (2006) CNS progenitor cells and oligodendrocytes aretargets of chemotherapeutic agents in vitro and in vivo. J. Biol. 5, e22
97 Wallace, T.L. and Johnson, E.M., Jr (1989) Cytosine arabinoside killspostmitotic neurons: evidence that deoxycytidine may have a role inneuronal survival that is independent of DNA synthesis. J. Neurosci.9, 115–124
98 Zhang, C.L. et al. (2008) A role for adult TLX-positive neural stem cellsin learning and behaviour. Nature 451, 1004–1007
99 Harding, J.W. andWright, J.W. (1979) Effects of intranasal irrigationwithmitotic inhibitors on olfactory behavior and biochemistry inmice.Brain Res. 168, 31–41
100 Santarelli, L. et al. (2003) Requirement of hippocampal neurogenesisfor the behavioral effects of antidepressants. Science 301, 805–809
101 Panagiotakos, G. et al. (2007) Long-term impact of radiation on thestem cell and oligodendrocyte precursors in the brain. PLoS One 2,e588
102 Coderre, J.A. et al. (2006) Late effects of radiation on the centralnervous system: role of vascular endothelial damage and glial stemcell survival. Radiat. Res. 166, 495–503
103 Monje, M.L. et al. (2002) Irradiation induces neural precursor-celldysfunction. Nat. Med. 8, 955–962
104 Imura, T. et al. (2003) The predominant neural stem cell isolated frompostnatal and adult forebrain but not early embryonic forebrainexpresses GFAP. J. Neurosci. 23, 2824–2832
105 Shepherd, G.M. et al. (2007) The olfactory granule cell: from classicalenigma to central role in olfactory processing.Brain Res. Rev. 55, 373–382
106 Singer, B.H. et al. (2009) Conditional ablation and recovery offorebrain neurogenesis in the mouse. J. Comp. Neurol. 514, 567–582
107 Jin, K. et al. (2010) Transgenic ablation of doublecortin-expressingcells suppresses adult neurogenesis and worsens stroke outcome inmice. Proc. Natl. Acad. Sci. U. S. A. 107, 7993–7998
108 Mandairon, N. et al. (2006) Enrichment to odors improves olfactorydiscrimination in adult rats. Behav. Neurosci. 120, 173–179
Review Trends in Neurosciences January 2011, Vol. 34, No. 1
30





























