Embed Size (px)
Citation preview
Journal of Structural Biology 154 (2006) 232–245
www.elsevier.com/locate/yjsbi
Observing growth steps of collagen self-assembly by time-lapse high-resolution atomic force microscopy
David A. Cisneros, Carlos Hung, Clemens M. Franz, Daniel J. Muller ¤
Biotechnology Center, University of Technology Dresden, 01307 Dresden, Germany
Received 19 October 2005; received in revised form 1 February 2006; accepted 2 February 2006Available online 20 March 2006
Abstract
Insights into molecular mechanisms of collagen assembly are important for understanding countless biological processes and at thesame time a prerequisite for many biotechnological and medical applications. In this work, the self-assembly of collagen type I moleculesinto Wbrils could be directly observed using time-lapse atomic force microscopy (AFM). The smallest isolated Wbrillar structures initiatingWbril growth showed a thickness of t1.5 nm corresponding to that of a single collagen molecule. Fibrils assembled in vitro established anaxial D-periodicity of t67 nm such as typically observed for in vivo assembled collagen Wbrils from tendon. At given collagen concentra-tions of the buVer solution the Wbrils showed constant lateral and longitudinal growth rates. Single Wbrils continuously grew and fusedwith each other until the supporting surface was completely covered by a nanoscopically well-deWned collagen matrix. Their thickness oft3 nm suggests that the Wbrils were build from laterally assembled collagen microWbrils. Laterally the Wbrils grew in steps of t4 nm, indi-cating microWbril formation and incorporation. Thus, we suggest collagen Wbrils assembling in a two-step process. In a Wrst step, collagenmolecules assemble with each other. In the second step, these molecules then rearrange into microWbrils which form the building blocks ofcollagen Wbrils. High-resolution AFM topographs revealed substructural details of the D-band architecture of the Wbrils forming thecollagen matrix. These substructures correlated well with those revealed from positively stained collagen Wbers imaged by transmissionelectron microscopy.© 2006 Elsevier Inc. All rights reserved.
Keywords: AFM; Collagen molecules; Growth steps; MicroWbrils; Molecular interactions; Self-assembly
1. Introduction connective tissue morphogenesis and to the dynamics that
Collagen represents the most abundant structural pro-tein in vertebrates. Its speciWc structural role to maintaintissue and cellular shape, strength, and integrity is carriedout by more than 20 specialized collagens, which eitherform Wbrillar or other assemblies (Kadler et al., 1996b;Myllyharju and Kivirikko, 2004). Besides shaping andorganizing the extracellular matrix (ECM) collagen pro-vides a scaVold, which is then functionalized by other extra-cellular proteins (Hohenester and Engel, 2002). Cellsembedded in the ECM undergo reciprocal and adaptiveinteractions, which are essential to the regulation of
* Corresponding author. Fax: +49 351 463 40342.E-mail address: [email protected] (D.J. Muller).
1047-8477/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.jsb.2006.02.006
characterizes tissue homeostasis and wound repair (Grin-nell, 2003). Besides their involvement in tissue structuringand cell attachment (Akiyama et al., 1990; Grinnell, 2000,2003), collagens receive considerable interest because theyare involved in various human diseases (Kadler, 1993;Kunicki, 2002; Myllyharju and Kivirikko, 2001, 2004;Ortiz-Urda et al., 2005; Prockop, 1998, 1999; Yuspa andEpstein, 2005). Two- and three-dimensional synthetic colla-gen matrices (Bishop, 2000; Coombes et al., 2002; Guidryand Grinnell, 1985; Lee et al., 2001; Thomas et al., 2003)serve as platforms for cell biological and tissue engineeringapplications. Furthermore, collagen is frequently used tocoat non-biological surfaces, thereby enhancing their bio-compatibility (Coombes et al., 2002; Lee et al., 2001; Lloyd,2002) or to pattern functionalized surfaces (Demers et al.,2002; Lee et al., 2002; Sato et al., 2000). The most abundant

D.A. Cisneros et al. / Journal of Structural Biology 154 (2006) 232–245 233
collagen, type I, readily forms Wbrils in a self-assembly pro-cess for which the exact axial and lateral organization ofcollagen molecules is crucial. However, the process of howcollagen molecules pack laterally to form large supramolec-ular arrays has never been observed at high-resolutionin vitro or in vivo.
1.1. Assembly of collagen Wbrils
Although the structure of collagen has been extensivelystudied in detail, major steps of the self-assembly process ofcollagen molecules into microWbrils and Wbrils remain to beanswered (Kadler et al., 1996a). Ultrastructural informa-tion provided by electron microscopy (EM) and X-raydiVraction indicate that tendon Wbers consists of buildingblocks separated by average periodicities of around 4–5 nm(Hulmes et al., 1981; Hulmes and Miller, 1981; Orgel et al.,2001; Prockop and Fertala, 1998; Squire and Freundlich,1980). From these results a model of collagen assemblinginto microWbrils could be deduced. In a Wrst step, three pep-tide chains of collagen I wind into a triple-helical collagenmolecule of t300 nm in length (L) and t1.5 nm in diameter(Kadler et al., 1996b). It was suggested that Wve of these col-lagen molecules form one microWbril (Hulmes, 2002; Orgelet al., 2001; Ottani et al., 2002). The microWbril cross-sec-tion of Wve stranded collagen molecules exhibit a dimensionof t3£ 5 nm2, which is consistent with current models sug-gesting that the microWbrils are the building blocks oflarger Wbrils (Fig. 1). EM (Holmes et al., 2001; Meek et al.,1979), X-ray diVraction (Orgel et al., 2001), and atomicforce microscopy (AFM) (Baselt et al., 1993; Gale et al.,1995) data of collagen microWbrils show a 67 nm periodicity(D-period) along the longitudinal axis. This periodicity isdue to the stagger of two collagen molecules in the longitu-dinal direction by 67 nm with respect to each other. Sincethe length of a single collagen molecule corresponds toabout 4.46 D (t300 nm), a gap of 0.54 D is found betweenthe ends of adjacent collagen molecules in the collagen Wbril[Fig. 1; (Hulmes, 2002; Orgel et al., 2001; Wess et al., 1998)].
Current models describe the formation of collagen Wbrilsas a self-assembly process of microWbrils (Hulmes, 2002;Kadler et al., 1996a). It is further assumed that microWbrilsinterlink, thereby assembling into corrugated sheets ofmicroWbrils that establish and stabilize native Wbrils in tis-sue (Orgel et al., 2001). Such interWbrillar cross-linkingwould explain on one side the extreme mechanical strengthand on the other the high elastic resilience of collagenWbrils. At the same time the model supports structuralobservations suggesting that concentric packing arrange-ments of microWbrillar sheets or layers form the Wnal Wbril.
AFM has contributed to the understanding of collagenstructure. An advantage of AFM over EM is that it can beused to investigate biological systems in aqueous solution,thereby preserving their physiological conformations andfunctions (Drake et al., 1989). At the same time, the highsignal-to-noise ratio, superior to any other microscopictechnique, can permit resolution of surface structural
details down to <1 nm (Engel and Muller, 2000; Frederixet al., 2003; Müller et al., 2002). AFM has been used tostudy collagen assembly and network formation in diVerenttypes of biological systems (Bigi et al., 1997; Gayatri et al.,2001; Jurvelin et al., 1996; Marshall et al., 2001; Melleret al., 1997; Raspanti et al., 2001; Thalhammer et al., 2001),dynamic processes associated with enzymatic collagen deg-radation (Fantner et al., 2004; Lin et al., 1999; Sun et al.,2000), and the interaction of collagens with other moleculessuch as proteoglycans (Chen and Hansma, 2000; Fantneret al., 2004; Raspanti et al., 1997, 2005) or the discoidinreceptor (Agarwal et al., 2002). Several workers have usedAFM to image the structure of collagen polymerizedin vitro (ChernoV and ChernoV, 1992), in rat tendon Wber(Gutsmann et al., 2003; Raspanti et al., 2002; Revenkoet al., 1994), in dentin collagen Wbrils (Habelitz et al., 2002;Ho et al., 2005; Yang et al., 2005), in bone (Hassenkamet al., 2004), and in bovine humeral articular cartilage(Jurvelin et al., 1996). Recently, the eVect of pH and electro-lytes on the self-assembly of collagen and the exposure ofthe typical D-periodicity was characterized (Jiang et al.,2004a). The AFM stylus could then be applied as a nanotweezer to remodel the collagen matrix on the nanometerscale. Structural details ranging from Wve nanometer to afew hundred micrometer in size could be written into the
Fig. 1. Model of the structural assembly of collagen type I molecules intoWbrils. Description beginning from the top. The collagen molecule assem-bles from three collagen monomers forming a 294.8 nm long triple helix.Five collagen molecules stagger laterally shaping one microWbril exhibit-ing a cross-section of t3 £ 5 nm2. The smallest possible lateral shiftbetween two collagen molecules corresponds to 0.54 times the D-bandperiodicity of t67 nm. Oriented microWbrils assemble into a unipolar col-lagen Wbril showing a pronounced D-band periodicity. High-resolutionAFM topograph showing D-band periodicity and characteristic substruc-tures of a hydrated collagen Wbril (taken from Fig. 7).

234 D.A. Cisneros et al. / Journal of Structural Biology 154 (2006) 232–245
collagen matrix (Jiang et al., 2004b). These collagen matri-ces could be used to direct attachment, orientation, andmotility of Wbroblasts (Poole et al., 2005).
In this work, we directly observe the self-assembly of col-lagen type I molecules into higher ordered structures bytime-lapse AFM. Collagen molecules assembled laterallyand fused into broad sheet-like Wbrillar structures, whichwe recently named microribbons (Jiang et al., 2004a). Thislateral growth and fusion of collagen continued until theWnal matrix was completed. The smallest Wbrillar unitimaged corresponded to the dimensions of laterally assem-bled single layered collagen molecules. The collagen Wbrils,however, exhibited the thickness of microWbrils and showedgrowth steps corresponding to the width of a single microW-bril. Thus, we suggest a two-step process to describe the col-lagen Wbril assembly. High-resolution AFM topographsshowed detailed substructures of the self-assembled colla-gen. The characteristic D-band of collagen exhibited sub-structural details that reXected the staggering of collagenmolecules. The comparison with transmission electronmicroscopy (TEM) images suggested that the observed col-lagen Wbrils exhibited structural features observed ofin vivo assembled collagen Wbers from tendon.
2. Materials and methods
2.1. Sample preparation
Chemical reagents of analytical grade and ultrapurewater (18 M�/cm) were taken to prepare all aqueous solu-tions used. Solubilized bovine dermal collagen was pre-pared and puriWed as described (Bell et al., 1979),concentrated to 3 mg/ml and stored at pH »2 and at a tem-perature of 4 °C. The Wnal composition of the sample was97% collagen type I and 3% collagen type III. As a support-ing surface, a mica disc of 6.5 mm in diameter was used.After being freshly cleaved, a drop of buVer solution(t30 �l) was placed on to the mica surface. The sample andbuVer solution was placed into an AFM equipped with aXuid cell. After thermal relaxation, a small amount of buVersolution containing collagen at diVerent concentrations wasinjected into the buVer solution of the Xuid cell while time-lapse AFM imaging was continuously performed. After theWrst collagen molecules have been aligned by the hydrody-namic Xow created by the injection the self-assembly pro-cess of collagen Wbrils continued in this direction (Jianget al., 2004a).
2.2. AFM
The AFM (MultiMode, di-Veeco, CA) was operated inbuVer solution using a Xuid cell. The piezoelectric scannerhad a scan range of »100£ 100 �m2. Oxide sharpenedSi3N4 cantilevers having a nominal force constant of0.08 N/m (OMCL TR400PS) were employed and purchasedfrom Olympus Ltd (Tokyo). Imaging was performed intapping-mode (TM) with a drive frequency close to the
resonance frequency of the cantilevers (8–9.5 kHz) mountedto the Xuid cell immersed in buVer solution. The driveamplitude of the cantilever was set to a root mean square(RMS) value of 18–30 nm. All samples were prepared andimaged in buVer solution at room temperature (21 °C).
2.3. Image analysis
Image analysis and processing was carried out usingImage SXM (Barret, 2004) and Image J (AbramoV et al.,2004). A mask was created with the threshold option ofImage J to measure the ratio of collagen coating the mica ofan area of 1 �m2. Lateral and longitudinal growth was char-acterized by measuring the Wbril width and length at diVer-ent assembly times on surface areas of 15£ 15 �m2, whichwere randomly selected on the supporting surface having adiameter of 6.5 mm. In the case of the longitudinal growthmeasurement, the position of the Wbril end was measuredand the growth was derived from the diVerence to the Wbrilend of the following frame. The lateral Wbril growth (takenat diVerent areas) was derived from the increasing Wbrilwidth, which was determined using the “Wnd edge” functionof Image J from contour masks of the Wbrils. The thermallyinduced lateral drift of the topographs was compensatedmanually to allow determining Wbril widths at the same ref-erence points. Growth steps of individual collagen Wbrilscould be observed after increasing the temporal resolutionby scanning only a few lines over the Wbril, with each lineexhibiting the maximum number of 4096 pixels. The widthincrease was determined from the full-width at half-maxi-mum (FWHM) of the height proWle along a cross-sectionof 20 nm. Fibrils were considered exhibiting the same widthbetween two diVerent frames if their diVerence in width wasless than 2 pixels. To enhance ultrastructural details of col-lagen 13 D-periods of a selected topographical area wereaveraged. The averaged height proWle was cross-correlatedto the EM density proWle (Fig. 8) using the correlationfunction of Igor Pro (Wavemetrics, Inc.).
3. Results
3.1. Observing the lateral and longitudinal assembly of collagen
Jiang et al. established conditions (pH and electrolyte)for which collagen molecules adsorb and align onto micaunder an externally applied hydrodynamic Xow (Jianget al., 2004a). The collagen molecules assembled into Wbrilsthat formed a two-dimensional (2D) ultrathin (t3 nm)matrix. Characteristic nanoscopic structures of the matrixsuch as collagen Wbril spacing and orientation could be pro-duced at high accuracy and reproducibility (>95%). Colla-gen Wbrils of the matrix only established their characteristicD-band periodicity (t67 nm) in the presence of K+ ions. Ata pH close to the pI of collagen (9.3) (Hattori et al., 1999)individual collagen Wbrils establishing the collagen matrixcould be clearly resolved (Jiang et al., 2004a). To investigate

D.A. Cisneros et al. / Journal of Structural Biology 154 (2006) 232–245 235
the self-assembly of individual collagen molecules into aWbrillar network, we performed our experiments in anaqueous solution buVered at pH 9.2 in the presence of K+
ions.Injecting collagen molecules into the buVer solution ini-
tiated the growth of collagen Wbrils (Jiang et al., 2004a),which was monitored using time-lapse tapping-mode (TM)AFM (Fig. 2). The buVer solution (pH 9.2, 50 mM glycine,200 mM KCl) of the experiment contained collagen mole-cules at a concentration of 12 �g/ml. The Wrst two topo-graphs revealed the adsorption of a single collagen Wbril
onto the mica surface (Figs. 2A and B), which was thencontinuously growing into the lateral and longitudinaldirection (Figs. 2B and C; arrows). Repeated imaging of thesame sample area revealed that the single Wbril continu-ously grew while another Wbril grew into the scanningframe (Figs. 2D–F). The longitudinal axes of both Wbrilswere parallel to each other. However, the growth directionsof both Wbril ends were opposite to each other (Figs. 2B, F).Imaging both ends of a single Wbril showed that they grewinto opposite directions (data not shown). From theseexperiments, it could be concluded that single collagen
Fig. 2. Observing collagen self-assembly by time-lapse TM-AFM. (A–G) Topographs of laterally and longitudinally growing collagen Wbrils (arrows). TheD-band periodicity of single Wbrils can be clearly resolved independently of the Wbrillar width. (D–F) A Wbril growing longitudinally into the center of thetopograph while the upper Wbril grows laterally. (G–I) The growing collagen Wbrils assemble into a matrix that almost covers the entire supporting surface.Defects, such as formed by colliding interphases between two formerly separated Wbrils [circle in (I)] disappear after a certain time (J–L). Eventually thesupporting surface is almost completely covered and almost all defects are annealed or minimized. Topographs were recorded in buVer solution (pH 9.2,50 mM glycine, 200 mM KCl), at a collagen concentration of 12 �g/ml at 27 °C and exhibit a full gray level corresponding to a vertical scale of 5 nm.

236 D.A. Cisneros et al. / Journal of Structural Biology 154 (2006) 232–245
Wbrils simultaneously grew on both ends and sides. Afterseveral minutes the width of both Wbrils signiWcantlyincreased until they came into contact (Figs. 2F–I). A thirdWbril was laterally approaching from the bottom of thetopograph. Soon after this third Wbril joined the two previ-ously fused Wbrils as well. Apparently, the growing Wbrilsjoined each other laterally (Figs. 2I–L) thereby forming apart of the 2D matrix that later on covered the entire sup-porting surface. The D-band periodicity established showedthe same orientation throughout the collagen matrix.
The presence of the 67 nm periodicity (D-band) indi-cated that collagen molecules assembled into their nativeWbrillar structure beginning from the very Wrst stage of theirassembly. As directly revealed from the equally gray shadedstructures of the topography, the thickness of the collagenmatrices did not further increase after reaching their Wnalheight of 2.9§0.3 nm (nD34). Thus, we conclude thatcollagen microWbrils assembled into Xat Wbrils. These band-like structures, which Wnally established the ultraXatcollagen matrix, were recently named microribbons (Jianget al., 2004a).
In the early stage of Wbril fusion it appeared that occa-sionally defects occurred in the 2D collagen matrix (Fig. 2I;circle). After a certain time period about 50% of thesedefects disappeared, suggesting that collagen moleculesfrom the solution have Wlled in the gaps. Some gaps how-ever exhibited some other particles insides besides elon-gated collagen molecules (Fig. 2J; circles), which were mostprobably contaminants from the buVer solution. Thesegaps were minimized with time. Another interesting featureobserved was that the D-periodicities of neighboring Wbrilswere slightly shifted against each other before Wbril fusion.
However, as two Wbrils approached each other, theirD-bands could register with each other thereby preventinga mismatch.
The smallest single Wbrils observed (Fig. 3A) exhibited alength of t800 nm and a width of 21.4§ 3.7 nm (nD9) mea-sured at full-width at half-maximum (FWHM). It shouldbe noted here that the width of isolated Wbrillar structuresand other isolated objects imaged by AFM could appearsigniWcantly broadened (Allen and Goldberg, 1992; Sch-warz et al., 1994). This broadening is due to non-linear con-volution between the AFM stylus and the sample. Incontrast, measuring the periodicities of structural featuresby AFM is not subject to stylus-sample superimposition(Engel et al., 1997), and thus, the determination of the D-band periodicity reXected the true dimensions of the sam-ple. However, the time-lapse AFM topographs oVeredinsights into the growth behavior of single early Wbrillarstructure into larger Wbrils exhibiting lengths and widthssuperseding the dimensions of the initial Wbrils severaltimes (Figs. 3A–C). Initially, these early collagen Wbrils hada height of 1.4§0.4 nm, and in subsequent assembly stepsthey increased their height to t3 nm (see above). Subse-quently, the growing Wbril laterally joined the thicker colla-gen Wbril at the bottom of the topograph (Fig. 3C). Thelateral growth rate of early Wbrillar structures did not diVerfrom those measured for longer and broader Wbrils(4.9§ 0.1 nm/min).
3.2. Observing tips of collagen Wbrils fusing
Fig. 4 documents the fusion of the tip of an early Wbrillarstructure with that of a larger collagen Wbril. Initially
Fig. 3. Lateral and longitudinal growth of Wbrillar collagen structures at an early stage. TM-AFM topographs (top panel) and phase images (bottompanel) show single Wbrils at diVerent growth stages, each one exposing the typical D-band periodicity (t65 nm) of collagen. The initial Wbril continuouslygrew over the time. Images were recorded in buVer solution (pH 9.2, 50 mM glycine, 200 mM KCl), at a collagen concentration of 6 �g/ml at 27 °C andexhibited a full gray level corresponding to vertical scale of 5 nm for height and 15° for phase.

D.A. Cisneros et al. / Journal of Structural Biology 154 (2006) 232–245 237
(Fig. 4A), the longitudinal axes of both Wbrils were shiftedin relation to each other and spatially separated (circle).Three minutes later (Fig. 4B), this gap has been bridgedallowing both Wbril tips to fuse with each other. The Wbril-lar structure bridging the gap was somewhat bent to con-nect both Wbril tips. This demonstrates again that collagenWbrils, although exceptionally well suited to withstand ten-sile stresses, have a certain elastic resilience (Jiang et al.,2004a). As revealed from the vertical scale of the topogra-phy, the broad Wbrillar structures protruded higher (t3 nm)compared to the tips of the growing collagen Wbrils1.4§ 0.4 nm (nD16). The lateral growth rates of two fusedWbrils were the same as measured for each Wbril beforefusion. Further growth of collagen tips to broad Wbersincreased their height to 3 nm. It may be speculated that theXexible collagen tips have not been perfectly contoured bythe scanning AFM stylus. Therefore, their apparent heightwould appear reduced. However, after stabilizing the colla-gen Wbril tips by glutaraldehyde we observed that indeedtheir height was only t1.5 nm (data not shown).
3.3. Lateral vs. longitudinal growth kinetics
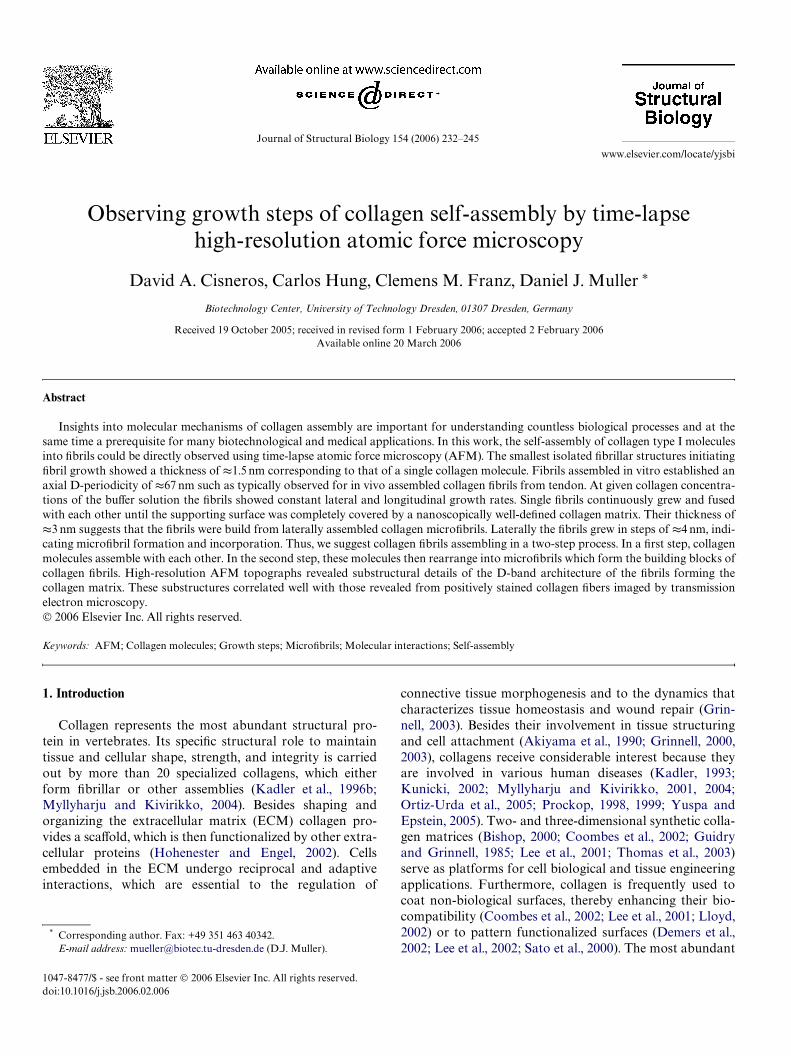
The growth of the collagen matrix was characterized inbuVer solutions containing collagen concentrations rangingfrom 6 to 24 �g/ml. Recording one TM-AFM topographrequired about 4 min. The growth rates of the collagenmatrix (Fig. 5) were determined by the percentage of the
supporting surface covered (see Section 2). As expected(Kadler et al., 1996a), growth curves vs. time showed a typi-cal sigmoidal behavior. The slope during the lag phase var-ied on the collagen concentration of the buVer solution(Fig. 5A). The relative growth rates of individual Wbrils at6 �g/ml were determined in longitudinal and lateral direc-tion (Movie 1, supplementary material) (Figs. 5B–C). Lin-ear Wts allowed us estimating the slopes of average lateraland longitudinal growth rates, which were 4.9§ 0.1 nm/minand 60.6§1.3 nm/min, respectively. Both grow rates wereconstant for a given collagen concentration until the sup-porting surface was fully covered. A second collagen layergrowing on top of the Wrst one could not be observed underthe conditions used in this work (Jiang et al., 2004a). It wasdiYcult to monitor the Wbril growth at higher collagen con-centrations. Under these conditions the collagen grew tofast to allow tracking individual Wbrils by the relativelyslow imaging process of the TM-AFM (Fig. 2).
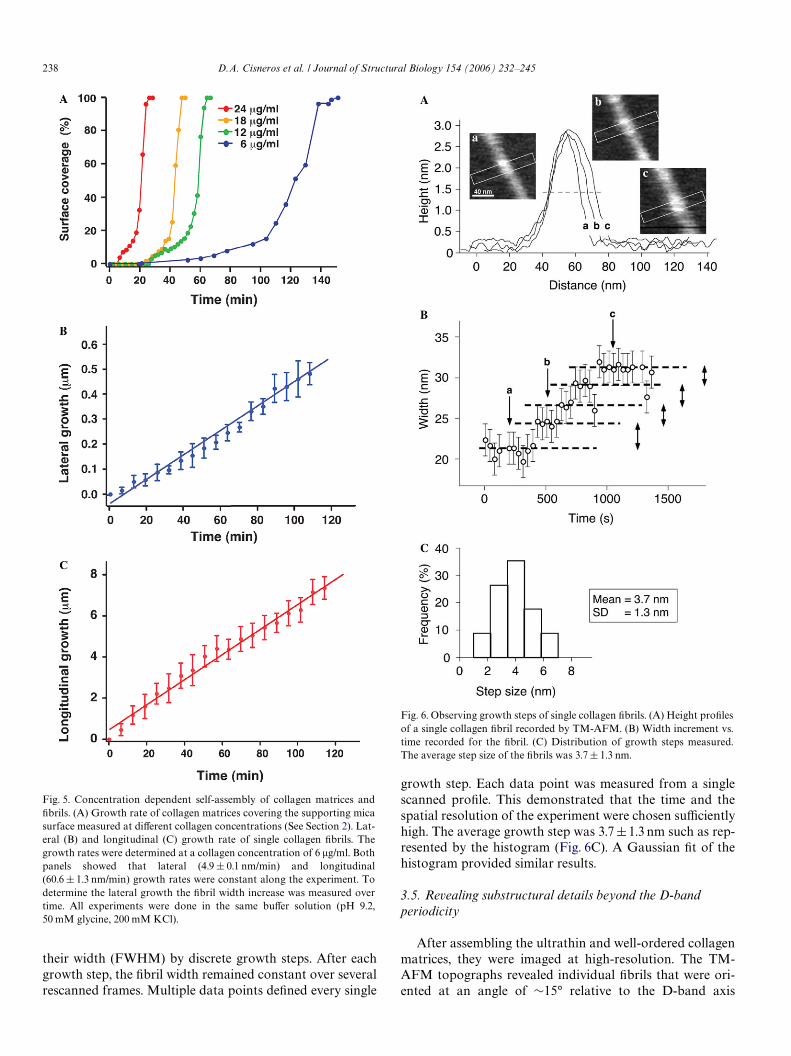
3.4. Revealing growth steps of single collagen Wbrils
To enhance the temporal resolution of the TM-AFM,we scanned individual collagen Wbrils by only a few heightlines. Each height line exhibiting 1024 pixels was scannedperpendicular to a selected collagen Wbril (Fig. 6A). Thepixel size of the scanned proWles was smaller than 0.5 nmyielding a spatial resolution of »1 nm. Analysis of the time-lapse height proWles (Fig. 6B) showed that Wbrils increased
Fig. 4. Fusion of two Wbril ends. (A) Collagen Wbril prior to the fusion event. Top, topographs recorded using TM-AFM. Bottom, TM-AFM phase imagesrecorded of the same area. (B) A collagen early Wbrillar structure fusing with the thicker collagen Wbril. Topographs were recorded in buVer solution (pH9.2, 50 mM glycine, 200 mM KCl), a collagen concentration of 6 �g/ml at 27 °C and exhibit a full gray level corresponding to vertical scale of 5 nm forheight and 15° for phase.
238 D.A. Cisneros et al. / Journal of Structural Biology 154 (2006) 232–245
their width (FWHM) by discrete growth steps. After eachgrowth step, the Wbril width remained constant over severalrescanned frames. Multiple data points deWned every single
Fig. 5. Concentration dependent self-assembly of collagen matrices andWbrils. (A) Growth rate of collagen matrices covering the supporting micasurface measured at diVerent collagen concentrations (See Section 2). Lat-eral (B) and longitudinal (C) growth rate of single collagen Wbrils. Thegrowth rates were determined at a collagen concentration of 6 �g/ml. Bothpanels showed that lateral (4.9 § 0.1 nm/min) and longitudinal(60.6 § 1.3 nm/min) growth rates were constant along the experiment. Todetermine the lateral growth the Wbril width increase was measured overtime. All experiments were done in the same buVer solution (pH 9.2,50 mM glycine, 200 mM KCl).
growth step. Each data point was measured from a singlescanned proWle. This demonstrated that the time and thespatial resolution of the experiment were chosen suYcientlyhigh. The average growth step was 3.7§1.3 nm such as rep-resented by the histogram (Fig. 6C). A Gaussian Wt of thehistogram provided similar results.
3.5. Revealing substructural details beyond the D-band periodicity
After assembling the ultrathin and well-ordered collagenmatrices, they were imaged at high-resolution. The TM-AFM topographs revealed individual Wbrils that were ori-ented at an angle of »15° relative to the D-band axis
Fig. 6. Observing growth steps of single collagen Wbrils. (A) Height proWlesof a single collagen Wbril recorded by TM-AFM. (B) Width increment vs.time recorded for the Wbril. (C) Distribution of growth steps measured.The average step size of the Wbrils was 3.7§ 1.3 nm.

D.A. Cisneros et al. / Journal of Structural Biology 154 (2006) 232–245 239
(Fig. 7A). This observation was in excellent agreement withthe 15° tilt observed for microWbrils forming corneal Wbrils(Holmes et al., 2001), tendon Wbers (Gutsmann et al., 2005;Raspanti et al., 1989; Wess et al., 1998), and to recentlypublished models of the collagen microWbril assembly(Engel and Prockop, 1998; Hulmes, 2002; Wess, 2005).AFM topographs recorded at higher resolution, revealeddistinct substructures within the D-periodicities (Fig. 7B).The corrugations resulted in banded patterns that were ori-ented in parallel to the D-bands. Averaging topographicsegments (Fig. 7B; rectangle) enhanced the characteristiccorrugations, which were repeated throughout the heightproWle of the D-bands. The averaged asymmetric heightproWle could be correlated to the features of the unpro-cessed AFM topograph (Fig. 7B; lower panel).
4. Discussion
4.1. Collagen assembly observed by tapping-mode AFM
Collagen Wbril formation is mainly an entropy drivenself-assembly process (Engel and Prockop, 1998; Kadleret al., 1987, 1996b; McBride et al., 1992), which is driven bythe loss of solvent molecules from the protein surface. Theformation of circular cross-sections between collagen mole-cules minimizes surface area and volume ratio of the Wnalstructure. Although many publications have contributed tothe general understanding of the collagen Wbril assembly(Christiansen et al., 2000; Holmes et al., 2001; Paige and
Goh, 2001; Prockop and Fertala, 1998; Silver et al., 2003),the underlying self-assembly process could never beenimaged directly.
Here, we have developed an assay to characterize theself-assembly process of collagen into Wbrils and highlyordered Wbrillar networks. The exceptionally high spatialresolution and signal-to-noise ratio of the AFM allowed usto follow the growth of single collagen Wbrils and their inte-gration into higher ordered structures. Single Wbrils showedconstant growth rates depending on the collagen concen-tration of the buVer solution (Fig. 5). The longitudinalgrowth direction of the Wbrils was determined by the hydro-dynamic Xow which was created by injecting the collagenmolecules into the buVer solution covering the AFM sup-port (Jiang et al., 2004a). This injection was initiated at thestarting point of the experiments. All collagen moleculesassembled after this injection grew in the same lateral direc-tion. Certain buVer conditions were shown to guide thegrowth of a second collagen layer on top of the Wrst one(Jiang et al., 2004a; and unpublished results). However, inall experiments performed in this work, the collagen Wbrilsdid not grow into vertical direction to the supporting sur-face. This suggests that the support formed an interfacethat guided the self-assembly of collagen into two dimen-sions. Thus, the growth kinetics was determined by the lat-eral and longitudinal growth rates of the Wbrils. Time-lapseAFM movies showed that single Wbrils grew independentfrom each other until they fused with adjacent Wbrils(Fig. 4B). At the end of the assembly the Wbrils formed a
Fig. 7. High-resolution TM-AFM topographs of self-assembled collagen matrices. (A) Single collagen Wbrils are slightly tilted (»15°) in respect to the nor-mal axis of the D-band periodicity. The D-bands exhibited height modulations of t0.4 § 0.1 nm (compare Table 1). The rectangle outlines an interface oftwo overlapping collagen Wbrils. (B) Substructural details and height proWle beyond the D-band periodic structure. Unprocessed AFM topograph (top)
and averaged height proWle (lower panel). The height proWle of a selected area of 980 £ 70 nm2 was averaged line for line. The collage of the lower panelallows the correlation of the averaged height proWle and the unprocessed topograph. Correlation of the height proWle with the TEM data (Fig. 8) alloweddetermining the microWbril polarity establishing the matrix. The arrow points from the C- to the N-terminal end of the unipolar microWbrils. Topographswere recorded in buVer solution (pH 9.2, 50 mM glycine, 200 mM KCl) at 27 °C and exhibit a full gray level corresponding to vertical scale of 5 nm (A) and3 nm (B) for height.
240 D.A. Cisneros et al. / Journal of Structural Biology 154 (2006) 232–245
continuous 2D matrix, which covered the entire supportingsurface.
When assembled under certain buVer solutions a furthercollagen matrix can be grown on the Wrst one (Jiang et al.,2004a). Instead of investigating the self-assembly of such athree-dimensional collagen matrix, we have chosen a buVersolution in which the self-assembled Wbrils were clearly dis-tinguishable from each other. This criterion allowed us toresolve steps of the lateral and longitudinal self-assembly ofcollagen Wbrils at high-resolution. We assume that thecollagen Wbrils self-assembled into their native structure,which is demonstrated by the D-periodicity and the sub-structural features observed at high-resolution (see below).Thus what we characterized in this work is the self-assem-bly of collagen molecules into their native structure. It maybe, however, possible that collagen Wbrils are able to self-assemble in alternative ways. This possibility may be under-lined by the recent observation that collagen can assembleinto diVerent matrices (Jiang et al., 2004a). To elucidateother potential self-assembly pathways of collagen mayform an interesting research project with medical, biotech-nological, and biological relevance.
Under the experimental conditions studied here, wecould not observe any self-regulation of the Wbril width.After completed assembly the collagen matrix covered theentire supporting surface, which could be several centimeterin diameter (Jiang et al., 2004a). The absence of self-regula-tion may be directly related to the absence of proteogly-cans, which have been suggested to regulate the lateralfusion of Wbrils (Graham et al., 2000; Kadler et al., 1996b,2000) or to other enzymes controlling the collagen shapewithin organisms (Holmes et al., 1996; Myllyharju andKivirikko, 2004). However, it has been previously shown,that under certain environmental conditions the pH andthe electrolyte composition and concentration of the buVersolution can determine the spacing and width of the Wbrilsas well (Jiang et al., 2004a).
4.2. Minimum unit of Wbril growth
In embryonic chicken, early Wbrils have been describedto have a length of t1 �m (Kadler et al., 2000). These Wbrilsexpose a particular axial mass distribution (AMD) in whichthe mass of the Wbril increases from the Wbril tip to a maxi-mum point and then decreases without any plateau region(Graham et al., 2000; Kadler et al., 2000). Other studies onthe self-assembly of type I collagen suggest characteristicunits that range in length from 4-D staggered dimers(t570 nm) (Kobayashi et al., 1985; Silver et al., 1979; Wardet al., 1986) to units that were about 700 nm long (Bernengoet al., 1978). In both cases the unit lengths were between 2and 3 collagen molecules long, while the number of mole-cules that formed such a unit may range between 5 and 17.Estimates of the unit diameter range between 2 and 6 nm(Gale et al., 1995). The smallest isolated Wbrillar structuresobserved exhibited lengths of t800 nm, widths of t20 nmand heights of t1.5 nm (Fig. 3). The height of the Wnal
collagen matrix was t3 nm. This suggests that the thicknessof these early Wbrillar collagen structures establishing thecollagen matrix changed from single layered collagen mole-cules to a Wbril exhibiting a thickness being equivalent totwo collagen molecules. However, the width of an objectdetermined by TM-AFM is often subject to non-linear sty-lus-sample convolution artifacts and thus appear broader(Allen and Goldberg, 1992; Schwarz et al., 1994). It may befurther assumed that isolated Wbrils exhibit an enhancedmobility and Xexibility on the supporting mica surface andthus appear broader when imaged. This assumption thatthe width of isolated Wbrils appeared thicker in the topo-graph would be supported by recent observations that thesmallest Wbrillar units embedded in a self-assembled colla-gen matrix exhibited a width of t5–6 nm (Jiang et al.,2004a). In embedding matrices the width of the Wbril can beestimated with much higher accuracy since the AFM stylusconvolution on one side broadens isolated structures butdoes not inXuence their distances relative to each other(Engel et al., 1997). While the diameter of such a smallestunit t5£ 3 nm would Wt that of a single collagen microW-bril, Wbrillar structures exhibiting a height corresponding tothat of a single collagen molecule do not match currentmodels of microWbrils (Holmes et al., 2001; Hulmes, 2002;Ottani et al., 2002; Wess et al., 1998). Hence we suggest thatthe collagen matrix is formed by collagen microWbrils, butthat the smallest isolated Wbrillar structures observed havenot been assembled into microWbrils yet. While characteriz-ing the height of Wbrillar tips it was observed that they wereonly about one collagen molecule thick. Time-lapse AFMobserved that the Wbril tips grew laterally into broad struc-tures and vertically into a Wnal thickness of t3 nm. Thus,we assume, that the Wnal assembly of collagen moleculesinto microWbrils took place while assembling into Wbrillarstructures. A two-step mechanism may explain the collagenself-assembly into Wbrils. First the collagen molecules asso-ciated with each other or with Wbrils. Then the collagenmolecules assemble into microWbrils, which form the build-ing blocks of collagen Wbrils.
4.3. Growth steps of collagen Wbrils
In our experiments, the smallest Wbril units grew at thesame rate as longer and wider Wbrils. Recording proWles ofindividual collagen Wbrils at improved temporal resolu-tion (Fig. 6) showed that single Wbrils increased theirwidth stepwise. The spatial resolution of these experi-ments (t1 nm) was about the width of one collagen mole-cule. However, on average the growth steps observed wereabout 4 nm, which corresponds well to the width of a sin-gle microWbril (Holmes et al., 2001; Hulmes, 2002; Ottaniet al., 2002; Wess et al., 1998). The constant lateral growthrates measured (Fig. 5B) suggested that the proposedstepwise growth process did not change during the entireself-assembly process of the collagen matrix. This datasupport the idea that the smallest unit of Wbril growth is amicroWbril, suggesting that microWbrils could form the

D.A. Cisneros et al. / Journal of Structural Biology 154 (2006) 232–245 241
units or “building blocks” of the collagen matrix assem-bly. Apparently this interpretation is contradicted by theobservation that the smallest isolated Wbrillar structuresshow only a height of 1.5 nm. The two-step process of col-lagen assembly assumes that collagen molecules bind witha certain rate constant to the growing Wbril. In a secondstep, these molecules rearrange to form a microWbril. Theobserved step size of Wbril growth (t4 nm) correspondswell to the average diameter (t3£ 5 nm) of a microWbril.As mentioned, convolution eVects occurring between theAFM stylus and the collagen Wbril broaden the proWle ofthe Wbril width. Structures protruding less than the Wbrilwill not be imaged unless they laterally extend far enoughto be detected by the AFM stylus (Allen and Goldberg,1992; Engel et al., 1997; Schwarz et al., 1994). Single colla-gen molecules laterally associating with a higher Wbrilcannot be detected by the AFM stylus until these mole-cules have increased their height by the formation of amicroWbril. Thus, what we observed were the growth stepsof microWbril formation by collagen molecules associatedto the collagen Wbril.
4.4. How do Wbrils fuse?
TM-AFM topographs showed that adjacent collagenWbrils could either fuse laterally or by their ends. In about50% of all cases, the regions of laterally fused Wbrils showedno or only very little structural mismatch of their D-period-icity. However, in other cases a mismatch between the join-ing Wbrils was observed as a shift in their D-bandperiodicities (Fig. 7A). It was previously shown, that Wbrilsassembled from acid-soluble collagen, such as used in thiswork (see Section 2), are unipolar, D-periodic and have twosmoothly tapered ends (Graham et al., 2000; Kadler et al.,1996b). Such unipolar Wbrils can fuse in a number of ways.Unipolar Wbrils containing collagen molecules withoriented amino N- to carboxyl C-ends could fuse in side-by-side register resulting in Wbril thickening. Fibrils of thesame orientation could also fuse in end-on-end registerresulting in a longer Wbril. End-on-end fusion of two oppo-sitely oriented unipolar Wbrils is also possible. Such a fusionof one Wbril oriented N- to C- from left to right and theother Wbril oriented N- to C- approaching from right to leftwould yield a bipolar Wbril. Side-by-side fusion of twooppositely directed unipolar Wbrils is, however, not allowedaccording to current models describing the collagen Wbrilfusion.
Analysis of our experimental data shows that two Wbrilsends that connected by end-to-end fusion never exhibited amismatch of their D-periodicity. This supports the abovemodel that oppositely oriented unipolar collagen moleculescan approach and fuse with each other. However, theremust be a certain Xexibility of the collagen molecules toestablish the conditions to fuse. Approaching collagenWbrils cannot recognize each other until they can interactby molecular mechanisms. Since molecular interactionsoccurring between two Wbril ends may be to low to physi-
cally move the entire Wbrils immobilized to the supportingsurface, their ends must exhibit a certain Xexibility whichenables their rearrangement for coordinated fusion. Long-range interactions may also assist in aligning the Wbril endsand staggering them in the right manner. The need andexistence for such Xexibility to fuse two approaching colla-gen ends was observed by the example shown (Fig. 4). Thisobservation agrees well with that of collagen matrices inwhich individual collagen Wbers could be mechanicallyrearranged during their assembly process (Jiang et al.,2004a).
Adjacent Wbrils that grew towards each other were fre-quently observed to fuse laterally (side-by-side). In about50% of all cases, the periodic D-bands of fused Wbrilsshowed no mismatch. Because two independently growingcollagen matrices cannot stagger their D-bands beforeinteracting with each other, we assume that long-rangeinteractions may align approaching Wbrils so that they canstagger in the right manner to facilitate their fusion. Asobserved previously, the collagen matrices exhibit a highXexibility to be rearranged during the assembly process(Jiang et al., 2004b).
The molecular basis for the Xexibility of type I collagenresults from sequences lacking amino acids proline andhydroxyproline. Electron microscopy data (Hofmannet al., 1984), modeling studies (Paterlini et al., 1995), andstereochemical maps (Silver et al., 2002) suggest thatregions lacking of proline and hydroxyproline exhibit anincreased Xexibility and thus can adopt more conforma-tions than other regions of the triple helix. In contrastregions with the sequence Gly-Pro-Hyp are very rigid.Hence, the collagen triple helix can be considered a com-posite of regions with varying degrees of stiVness (Silveret al., 2003). Furthermore, it has been recently postulated,that the type III collagen molecule is more Xexible thantype I molecule and that mixing of both collagen typeswould increase the elastic modulus of collagen Wbrils (Sil-ver et al., 2002). Since we used a mixture of t97% type Iand of t3% type III collagen in this work, we assume thatboth the molecular Xexibilities contributed by the colla-gen molecule itself and by the mixture of both collagentypes may facilitate the collagen self-assembly into thehighly ordered Wbrillar matrix.
In all other cases (50%) when collagen Wbrils fused, weobserved a mismatch between the D-bands of two fusedcollagen Wbrils (Fig. 7). Apparently, these fusing collagenWbrils could not stagger at their interface. We do notknow how the collagen molecules were rearranged at thisinterface. AFM topographs recorded at subnanometerresolution may in future reveal the precise arrangement ofsingle collagen molecules at such interfaces. It may beassumed that fusing collagen Wbrils may be unipolar hav-ing diVerent ends (C- and N-terminal ends). Side-by-sidefusion of two unipolar Wbrils may result in a perfect matchof their D-bands. In contrast, side-by-side fusion of twooppositely oriented molecules would result in a mismatchsuch as observed here. High-resolution TM-AFM

242 D.A. Cisneros et al. / Journal of Structural Biology 154 (2006) 232–245
topographs clearly showing a directionality of the D-bandstructure may further support this hypothesis (see below).
4.5. Self-healing process of collagen
Time-lapse TM-AFM topographs showed that fusing col-lagen Wbrils could leave structural gaps in the growing colla-gen matrix. However, in about 50% of all cases, the gapsdisappeared after a certain time period (Fig. 2). It wasrecently observed that after mechanical removal of collagenmolecules from rat-tail tendon the molecules could bind backto the Wbril (Gutsmann et al., 2005). It was suggested that thisself-organization and self-healing signiWcantly contributes totoughness and strength of tendon Wbrils. During wound heal-ing, myoWbroblasts increase the synthesis of the ECM anddecrease its degradation (Grinnell, 2000, 2003). Such self-organizing capabilities of collagen to heal out defects couldcontribute to a faster and more eYcient tissue repair.
4.6. Collagen matrices assembled in vitro posses structural characteristics of tendon
Collagen Wbrils self-assembled under the conditionsemployed here established the characteristic D-band period-icity of t67 nm such as observed by EM or AFM of in vivoassembled collagen Wbers. High-resolution TM-AFM topo-graphs (Fig. 7) of hydrated Wbrils clearly resolved a multi-banded substructure of the D-periodic collagen. Afteraligning the height proWle with the TEM density proWle(Fig. 7B) by cross-correlation, these structural details of theD-band could be correlated to bands observed by TEM ofpositively stained collagen Wbrils (Chapman et al., 1990;Meek et al., 1979). The height of each protrusion of the D-banded substructure is given in Table 1.
TEM images of collagen exhibited a more detailed sub-structural information if the collagen Wbers were positivelystained instead of the more frequently used negative stain-ing (Chapman et al., 1990; Graham et al., 2000; Kadleret al., 2000; Meek et al., 1979). The structural information
Table 1Average height of collagen bands substructuring the D-periodicity
A topographic area of »68 �m2 (980£ 70 nm2) of the collagen matrix wasaveraged line by line. All protrusions could be assigned to the collagenbands by comparing the TM-AFM proWle to the TEM data (Chapmanet al., 1990).
Band Average height (nm) Standard deviation (nm)
C2 0.32 0.08C3 0.12 0.05D 0.31 0.07E1 0.23 0.05E2 0.25 0.05A1 0.32 0.06A2 0.35 0.07A3 0.34 0.08A4 ND NDB1 0.43 0.07B2 0.48 0.06C1 0.40 0.10
obtained from positively stained collagen Wbrils showed anexcellent agreement to AFM topographs revealed fromnative unstained collagen Wbrils down to a lateral resolu-tion of t2 nm (Chapman et al., 1990). So far, such excellentstructural correlations were only obtained between nega-tively stained specimen observed by TEM and the AFMdata revealed from native biological samples (Karraschet al., 1994; Schabert et al., 1995). It is assumed, that posi-tive staining contrasts the biological specimen in a diVerentway than negative stain, which can be easily recognized bycomparing TEM images of collagen Wbrils stained by eitherone of both approaches (Chapman et al., 1990; Grahamet al., 2000; Kadler et al., 2000; Meek et al., 1979). Positivestain contrasts mainly electrostatic information of a biolog-ical sample and the uptake of stain by collagen Wbers istherefore assumed to be determined by the nature andabundance of charged side-chains on the collagen surface(Chapman et al., 1990). Hence, TEM data obtained frompositively stained Wbrils may be directly correlated withchemical information of the collagen. In contrast, AFMtopographs normally contour protein surface structures(Müller et al., 2002). However, it was recently shown, thatunder certain circumstances AFM topographs could be acomposite of structural and electrostatic information of theprotein surface (Philippsen et al., 2002). These electrostaticinteractions, are suppressed at suYciently high electrolyteconcentrations such as used to record the high-resolutiontopographs shown here (data available upon request).Thus, we may assume that both topographical and chemi-cal information of collagen Wbrils are identical up to a reso-lution of t2 nm.
Fig. 8. Comparing an averaged TM-AFM height proWle of collagen withthe TEM density proWle. The top panel shows the averaged AFM heightproWle of Fig. 7. The lower panel shows the density pattern revealed frompositively stained collagen Wbrils by TEM (Chapman et al., 1990; Meeket al., 1979). Labeling of the pattern follows the conventional notation inwhich groups of closely spaced bands are denoted by a, b, c, d, and e. Indi-vidual bands within a group have numerical preWxes a1, a2, etc. TM-AFMheight and TEM density proWles were aligned by correlation analysis. Allheight protrusions of the TM-AFM proWle correlated well with the den-sity of the bands obtained by positive stain TEM.

D.A. Cisneros et al. / Journal of Structural Biology 154 (2006) 232–245 243
4.7. Determining the orientation of collagen molecules within the matrix
From density proWles revealed by TEM the polarity (ori-entation) of collagen molecules within tendon can be deter-mined (Chapman et al., 1990). The correlation betweenAFM height and TEM density proWles (Fig. 8) allowed usto determine the orientation of collagen molecules assem-bled into the matrix (Fig. 7).
5. Conclusions
We have established an assay to observe the growth andfusion of individual collagen Wbrils. The lateral and longitu-dinal fusion of single Wbrils could be directly followed untilthey formed the Wnal collagen matrix. The kinetic parame-ters of the collagen assembly suggest that both the longitudi-nal and lateral growth of the collagen Wbrils at a lowconcentration were mainly determined by a two-step pro-cess describing the assembly of collagen molecules followedby the formation of microWbrils. The collagen Wbrils withinthis matrix possessed the typical longitudinal 67 nm periodicstructures such as observed for collagen Wbers assembledin vivo. The structure of these matrices corresponds to theone found in vivo and could therefore be used as scaVoldsfor the construction of biocompatible materials. AFM topo-graphs of native Wbrils within this collagen matrix revealedsubstructural details of the D-band such as observed byhigh-resolution TEM on in vivo assembled collagen. Besideshaving a great potential to be used in various biotechnologi-cal and medical applications, we propose that these ultrathincollagen matrices can be used as assays to characterize colla-gen binding domains or proteins, to characterize factors thatinXuence cell adhesion and migration on collagen or tostudy how proteoglycans regulate collagen Wbrillogenesis.
Acknowledgments
We thank Jonathon Howard and Pierre-Henri Puech forvaluable discussions and critical comments. This work wassupported by the deutsche Volkswagenstiftung, the Bun-desministerium für Bildung und Forschung (BMBF), andthe Arbeitskreis Osteoporose (AO) Davos, Switzerland.
Appendix A. Supplementary material
Supplementary data associated with this article can befound, in the online version, at doi:10.1016/j.jsb.2006.02.006.
References
AbramoV, M.D., Magelhaes, P.J., Ram, S.J., 2004. Image processing withImageJ. Biophotonics. Int. 11, 36–42.
Agarwal, G., Kovac, L., Radziejewski, C., Samuelsson, S.J., 2002. Bindingof discoidin domain receptor 2 to collagen I: an atomic force micros-copy investigation. Biochemistry 41, 11091–11098.
Akiyama, S.K., Nagata, K., Yamada, K.M., 1990. Cell surface receptors forextracellular matrix components. Biochim. Biophys. Acta 1031, 91–110.
Allen, D.B., Goldberg, B.D., 1992. Stimulation of collagen synthesis andlinear growth by growth hormone in glucocorticoid-treated children.Pediatrics 89, 416–421.
Barret, S.D., 2004. Image SXM, <http://www.ImageSXM.org.uk/>.Baselt, D.R., Revel, J.P., Baldeschwieler, J.D., 1993. SubWbrillar structure
of type I collagen observed by atomic force microscopy. Biophys. J. 65,2644–2655.
Bell, E., Ivarsson, B., Merrill, C., 1979. Production of a tissue-like structureby contraction of collagen lattices by human Wbroblasts of diVerentproliferative potential in vitro. Proc. Natl. Acad. Sci. USA 76, 1274–1278.
Bernengo, J.C., Herbage, D., Marion, C., Roux, B., 1978. Intermolecularinteraction studies on native and enzyme-treated acid-soluble collagen.Biochim. Biophys. Acta 532, 305–314.
Bigi, A., GandolW, M., Roveri, N., Valdre, G., 1997. In vitro calciWedtendon collagen: an atomic force and scanning electron microscopyinvestigation. Biomaterials 18, 657–665.
Bishop, P.N., 2000. Structural macromolecules and supramolecular orga-nisation of the vitreous gel. Prog. Retin Eye Res. 19, 323–344.
Chapman, J.A., Tzaphlidou, M., Meek, K.M., Kadler, K.E., 1990. The col-lagen Wbril—a model system for studying the staining and Wxation of aprotein. Electron Microsc. Rev. 3, 143–182.
Chen, C.H., Hansma, H.G., 2000. Basement membrane macromolecules:insights from atomic force microscopy. J. Struct. Biol. 131, 44–55.
ChernoV, E.A.G., ChernoV, D.A., 1992. Atomic force microscope imagesof collagen-Wbers. J. Vac. Sci. Tech. 10, 596–599.
Christiansen, D.L., Huang, E.K., Silver, F.H., 2000. Assembly of type I col-lagen: fusion of Wbril subunits and the inXuence of Wbril diameter onmechanical properties. Matrix Biol. 19, 409–420.
Coombes, A.G., Verderio, E., Shaw, B., Li, X., GriYn, M., Downes,S., 2002. Biocomposites of non-crosslinked natural and synthetic poly-mers. Biomaterials 23, 2113–2118.
Demers, L.M., Ginger, D.S., Park, S.J., Li, Z., Chung, S.W., Mirkin, C.A.,2002. Direct patterning of modiWed oligonucleotides on metals andinsulators by dip-pen nanolithography. Science 296, 1836–1838.
Drake, B., Prater, C.B., Weisenhorn, A.L., Gould, S.A.C., Albrecht, T.R.,Quate, C.F., Cannell, D.S., Hansma, H.G., Hansma, P.K., 1989. Imag-ing crystals, polymers, and processes in water with the atomic forcemicroscope. Science 243, 1586–1588.
Engel, A., Muller, D.J., 2000. Observing single biomolecules at work withthe atomic force microscope. Nat. Struct. Biol. 7, 715–718.
Engel, A., Schoenenberger, C.A., Muller, D.J., 1997. High resolution imag-ing of native biological sample surfaces using scanning probe micros-copy. Curr. Opin. Struct. Biol. 7, 279–284.
Engel, J., Prockop, D.J., 1998. Does bound water contribute to the stabilityof collagen? Matrix Biol. 17, 679–680.
Fantner, G.E., Birkedal, H., Kindt, J.H., Hassenkam, T., Weaver, J.C.,Cutroni, J.A., Bosma, B.L., Bawazer, L., Finch, M.M., Cidade, G.A.,Morse, D.E., Stucky, G.D., Hansma, P.K., 2004. InXuence of the degra-dation of the organic matrix on the microscopic fracture behavior oftrabecular bone. Bone 35, 1013–1022.
Frederix, P.L., Akiyama, T., Staufer, U., Gerber, C., Fotiadis, D., Muller,D.J., Engel, A., 2003. Atomic force bio-analytics. Curr. Opin. Chem.Biol. 7, 641–647.
Gale, M., Pollanen, M.S., Markiewicz, P., Goh, M.C., 1995. Sequentialassembly of collagen revealed by atomic force microscopy. Biophys. J.68, 2124–2128.
Gayatri, R., Sharma, A.K., Rajaram, R., Ramasami, T., 2001. Chro-mium(III)-induced structural changes and self-assembly of collagen.Biochem. Biophys. Res. Commun. 283, 229–235.
Graham, H.K., Holmes, D.F., Watson, R.B., Kadler, K.E., 2000. IdentiWca-tion of collagen Wbril fusion during vertebrate tendon morphogenesis.The process relies on unipolar Wbrils and is regulated by collagen–pro-teoglycan interaction. J. Mol. Biol. 295, 891–902.
Grinnell, F., 2000. Fibroblast-collagen-matrix contraction: growth-factorsignalling and mechanical loading. Trends Cell Biol. 10, 362–365.

244 D.A. Cisneros et al. / Journal of Structural Biology 154 (2006) 232–245
Grinnell, F., 2003. Fibroblast biology in three-dimensional collagen matri-ces. Trends Cell Biol. 13, 264–269.
Guidry, C., Grinnell, F., 1985. Studies on the mechanism of hydratedcollagen gel reorganization by human skin Wbroblasts. J. Cell Sci.79, 67–81.
Gutsmann, T., Fantner, G.E., Venturoni, M., Ekani-Nkodo, A., Thompson,J.B., Kindt, J.H., Morse, D.E., Fygenson, D.K., Hansma, P.K., 2003.Evidence that collagen Wbrils in tendons are inhomogeneously struc-tured in a tubelike manner. Biophys. J. 84, 2593–2598.
Gutsmann, T., Hassenkam, T., Cutroni, J.A., Hansma, P.K., 2005. SacriW-cial bonds in polymer brushes from rat tail tendon functioning asnanoscale velcro. Biophys. J. 89, 536–542.
Habelitz, S., Balooch, M., Marshall, S.J., Balooch, G., Marshall Jr., G.W.,2002. In situ atomic force microscopy of partially demineralizedhuman dentin collagen Wbrils. J. Struct. Biol. 138, 227–236.
Hassenkam, T., Fantner, G.E., Cutroni, J.A., Weaver, J.C., Morse, D.E.,Hansma, P.K., 2004. High-resolution AFM imaging of intact and frac-tured trabecular bone. Bone 35, 4–10.
Hattori, S., Adachi, E., Ebihara, T., Shirai, T., Someki, I., Irie, S., 1999.Alkali-treated collagen retained the triple helical conformation and theligand activity for the cell adhesion via alpha 2 beta 1 integrin. J. Bio-chem. 125, 676–684.
Ho, S.P., Sulyanto, R.M., Marshall, S.J., Marshall, G.W., 2005. The cemen-tum–dentin junction also contains glycosaminoglycans and collagenWbrils. J. Struct. Biol. 151, 69–78.
Hofmann, H., Voss, T., Kuhn, K., Engel, J., 1984. Localization of Xexible sites inthread-like molecules from electron micrographs. Comparison of intersti-tial, basement membrane and intima collagens. J. Mol. Biol. 172, 325–343.
Hohenester, E., Engel, J., 2002. Domain structure and organisation inextracellular matrix proteins. Matrix Biol. 21, 115–128.
Holmes, D.F., Gilpin, C.J., Baldock, C., Ziese, U., Koster, A.J., Kadler,K.E., 2001. Corneal collagen Wbril structure in three dimensions: struc-tural insights into Wbril assembly, mechanical properties, and tissueorganization. Proc. Natl. Acad. Sci. USA 98, 7307–7312.
Holmes, D.F., Watson, R.B., Chapman, J.A., Kadler, K.E., 1996. Enzymiccontrol of collagen Wbril shape. J. Mol. Biol. 261, 93–97.
Hulmes, D.J., 2002. Building collagen molecules, Wbrils, and supraWbrillarstructures. J. Struct. Biol. 137, 2–10.
Hulmes, D.J., Jesior, J.C., Miller, A., Berthet-Colominas, C., WolV, C.,1981. Electron microscopy shows periodic structure in collagen Wbrilcross sections. Proc. Natl. Acad. Sci. USA 78, 3567–3571.
Hulmes, D.J., Miller, A., 1981. Molecular packing in collagen. Nature 293,239–240.
Jiang, F., Horber, H., Howard, J., Muller, D.J., 2004a. Assembly of colla-gen into microribbons: eVects of pH and electrolytes. J. Struct. Biol.148, 268–278.
Jiang, F., Khairy, K., Poole, K., Howard, J., Muller, D.J., 2004b. Creatingnanoscopic collagen matrices using atomic force microscopy. MicroscRes. Tech. 64, 435–440.
Jurvelin, J.S., Müller, D.J., Wong, M., Studer, D., Engel, A., Hunziker, E.B.,1996. Surface and sub-surface morphology of bovine humeral articularcartilage as assessed by atomic force- and transmission electronmicroscopy. J. Struct. Biol. 117, 45–54.
Kadler, K.E., 1993. Learning how mutations in type I collagen genes causeconnective tissue disease. Int. J. Exp. Pathol. 74, 319–323.
Kadler, K.E., Hojima, Y., Prockop, D.J., 1987. Assembly of collagen Wbrilsde novo by cleavage of the type I pC-collagen with procollagen C-pro-teinase. Assay of critical concentration demonstrates that collagen self-assembly is a classical example of an entropy-driven process. J. Biol.Chem. 262, 15696–15701.
Kadler, K.E., Holmes, D.F., Graham, H., Starborg, T., 2000. Tip-mediatedfusion involving unipolar collagen Wbrils accounts for rapid Wbril elon-gation, the occurrence of Wbrillar branched networks in skin and thepaucity of collagen Wbril ends in vertebrates. Matrix Biol. 19, 359–365.
Kadler, K.E., Holmes, D.F., Trotter, J.A., Chapman, J.A., 1996a. CollagenWbril formation. Biochem. J. 316 (Pt. 1), 1–11.
Kadler, K.E., Holmes, D.F., Trotter, J.A., Chapman, J.A., 1996b. CollagenWbril formation. Biochem. J. 316, 1–11.
Karrasch, S., Hegerl, R., Hoh, J.H., Baumeister, W., Engel, A., 1994.Atomic force microscopy produces faithful high-resolution images ofprotein surfaces in an aqueous environment. Proc. Natl. Acad. Sci.USA 91, 836–838.
Kobayashi, K., Ito, T., Hoshino, T., 1985. Electron microscopic demon-stration of acid-labile, 4D-staggered intermolecular association of col-lagen formed in vitro. Coll. Relat. Res. 5, 253–260.
Kunicki, T.J., 2002. The inXuence of platelet collagen receptor polymor-phisms in hemostasis and thrombotic disease. Arterioscler. Thromb.Vasc Biol. 22, 14–20.
Lee, C.H., Singla, A., Lee, Y., 2001. Biomedical applications of collagen.Int. J. Pharm. 221, 1–22.
Lee, K.B., Park, S.J., Mirkin, C.A., Smith, J.C., Mrksich, M., 2002. Pro-tein nanoarrays generated by dip-pen nanolithography. Science 295,1702–1705.
Lin, H., Clegg, D.O., Lal, R., 1999. Imaging real-time proteolysis of singlecollagen I molecules with an atomic force microscope. Biochemistry 38,9956–9963.
Lloyd, A.W., 2002. Interfacial bioengineering to enhance surface biocom-patibility. Med. Device Technol. 13, 18–21.
Marshall, G.W., Yucel, N., Balooch, M., Kinney, J.H., Habelitz, S., Mar-shall, S.J., 2001. Sodium hypochlorite alterations of dentin and dentincollagen. Surf Sci. 491, 444–455.
McBride Jr., D.J., Kadler, K.E., Hojima, Y., Prockop, D.J., 1992. Self-assemblyinto Wbrils of a homotrimer of type I collagen. Matrix 12, 256–263.
Meek, K.M., Chapman, J.A., Hardcastle, R.A., 1979. The staining patternof collagen Wbrils. Improved correlation with sequence data. J. Biol.Chem. 254, 10710–10714.
Meller, D., Peters, K., Meller, K., 1997. Human cornea and sclera studiedby atomic force microscopy. Cell Tissue Res. 288, 111–118.
Müller, D.J., Janovjak, H., Lehto, T., Kuerschner, L., Anderson, K., 2002.Observing structure, function and assembly of single proteins by AFM.Prog. Biophys. Mol. Biol. 79, 1–43.
Myllyharju, J., Kivirikko, K.I., 2001. Collagens and collagen-related dis-eases. Ann. Med. 33, 7–21.
Myllyharju, J., Kivirikko, K.I., 2004. Collagens, modifying enzymes andtheir mutations in humans, Xies and worms. Trends Genet. 20, 33–43.
Orgel, J.P., Miller, A., Irving, T.C., Fischetti, R.F., Hammersley, A.P., Wess,T.J., 2001. The in situ supermolecular structure of type I collagen.Structure (Camb) 9, 1061–1069.
Ortiz-Urda, S., Garcia, J., Green, C.L., Chen, L., Lin, Q., Veitch, D.P.,Sakai, L.Y., Lee, H., Marinkovich, M.P., Khavari, P.A., 2005. Type VIIcollagen is required for Ras-driven human epidermal tumorigenesis.Science 307, 1773–1776.
Ottani, V., Martini, D., Franchi, M., Ruggeri, A., Raspanti, M., 2002. Hier-archical structures in Wbrillar collagens. Micron 33, 587–596.
Paige, M.F., Goh, M.C., 2001. Ultrastructure and assembly of segmental longspacing collagen studied by atomic force microscopy. Micron 32, 355–361.
Paterlini, M.G., Nemethy, G., Scheraga, H.A., 1995. The energy of forma-tion of internal loops in triple-helical collagen polypeptides. Biopoly-mers 35, 607–619.
Philippsen, A., Im, W., Engel, A., Schirmer, T., Roux, B., Muller, D.J., 2002.Imaging the electrostatic potential of transmembrane channels: atomicprobe microscopy of OmpF porin. Biophys. J. 82, 1667–1676.
Poole, K., Khairy, K., Friedrichs, J., Franz, C., Cisneros, D.A., Howard, J.,Mueller, D., 2005. Molecular-scale topographic cues induce the orien-tation and directional movement of Wbroblasts on two-dimensionalcollagen surfaces. J. Mol. Biol. 349, 380–386.
Prockop, D.J., 1998. What holds us together? Why do some of us fallapart? What can we do about it? Matrix Biol. 16, 519–528.
Prockop, D.J., 1999. Hopkins Memorial Medal lecture. Pleasant surprisesen route from the biochemistry of collagen to attempts at gene therapy.Biochem. Soc. Trans. 27, 15–31.
Prockop, D.J., Fertala, A., 1998. The collagen Wbril: the almost crystallinestructure. J. Struct. Biol. 122, 111–118.
Raspanti, M., Alessandrini, A., Ottani, V., Ruggeri, A., 1997. Direct visual-ization of collagen-bound proteoglycans by tapping-mode atomicforce microscopy. J. Struct. Biol. 119, 118–122.

D.A. Cisneros et al. / Journal of Structural Biology 154 (2006) 232–245 245
Raspanti, M., Binaghi, E., Gallo, I., Manelli, A., 2005. A vision-based, 3Dreconstruction technique for scanning electron microscopy: directcomparison with atomic force microscopy. Microsc. Res. Tech. 67, 1–7.
Raspanti, M., Congiu, T., Guizzardi, S., 2001. Tapping-mode atomic forcemicroscopy in Xuid of hydrated extracellular matrix. Matrix Biol. 20,601–604.
Raspanti, M., Congiu, T., Guizzardi, S., 2002. Structural aspects of theextracellular matrix of the tendon: an atomic force and scanning elec-tron microscopy study. Arch. Histol. Cytol. 65, 37–43.
Raspanti, M., Ottani, V., Ruggeri, A., 1989. DiVerent architectures of thecollagen Wbril: morphological aspects and functional implications. Int.J. Biol. Macromol. 11, 367–371.
Revenko, I., Sommer, F., Minh, D.T., Garrone, R., Franc, J.M., 1994. Atomicforce microscopy study of the collagen Wbre structure. Biol. Cell 80, 67–69.
Sato, K., Ebihara, T., Adachi, E., Kawashima, S., Hattori, S., Irie, S., 2000.Possible involvement of aminotelopeptide in self-assembly and thermalstability of collagen I as revealed by its removal with proteases. J. Biol.Chem. 275, 25870–25875.
Schabert, F.A., Henn, C., Engel, A., 1995. Native Escherichia coliOmpF porin surfaces probed by atomic force microscopy. Science268, 92–94.
Schwarz, U.D., Haefke, H., Reimann, P., Guntherodt, H.J., 1994. Tip arte-facts in scanning force microscopy. J. Microsc. 173, 183–197.
Silver, F.H., Ebrahimi, A., Snowhill, P.B., 2002. Viscoelastic properties ofself-assembled type I collagen Wbers: molecular basis of elastic and vis-cous behaviors. Connect. Tissue Res. 43, 569–580.
Silver, F.H., Freeman, J.W., Seehra, G.P., 2003. Collagen self-assemblyand the development of tendon mechanical properties. J. Biomech.36, 1529–1553.
Silver, F.H., Langley, K.H., Trelstad, R.L., 1979. Type I collagen Wbrillo-genesis: initiation via reversible linear and lateral growth steps. Bio-polymers 18, 2523–2535.
Squire, J.M., Freundlich, A., 1980. Direct observation of a transverse peri-odicity in collagen Wbrils. Nature 288, 410–413.
Sun, H.B., Smith, G.N., Hasty, K.A., Yokota, H., 2000. Atomic forcemicroscopy-based detection of binding and cleavage site of matrixmetalloproteinase on individual type II collagen helices. Anal. Bio-chem. 283, 153–158.
Thalhammer, S., Heckl, W.M., Zink, A., Nerlich, A.G., 2001. Atomic forcemicroscopy for high resolution imaging of collagen Wbrils—a new tech-nique to investigate collagen structure in historic bone tissues. J. Arch.Sci. 28, 1061–1068.
Thomas, A.C., Campbell, G.R., Campbell, J.H., 2003. Advances in vasculartissue engineering. Cardiovasc. Pathol. 12, 271–276.
Ward, N.P., Hulmes, D.J., Chapman, J.A., 1986. Collagen self-assemblyin vitro: electron microscopy of initial aggregates formed during thelag phase. J. Mol. Biol. 190, 107–112.
Wess, T.J., 2005. Collagen Wbril form and function. Adv. Protein Chem. 70,341–374.
Wess, T.J., Hammersley, A.P., Wess, L., Miller, A., 1998. Molecular pack-ing of type I collagen in tendon. J. Mol. Biol. 275, 255–267.
Yang, W., Lu, Y., Kalajzic, I., Guo, D., Harris, M.A., Gluhak-Heinrich, J.,Kotha, S., Bonewald, L.F., Feng, J.Q., Rowe, D.W., Turner, C.H.,Robling, A.G., Harris, S.E., 2005. Dentin matrix protein 1 gene cis-reg-ulation: use in osteocytes to characterize local responses to mechanicalloading in vitro and in vivo. J. Biol. Chem. 280, 20680–20690.
Yuspa, S.H., Epstein Jr., E.H., 2005. Cancer. An anchor for tumor cellinvasion. Science 307, 1727–1728.





























