Embed Size (px)
Citation preview

Orexin-A Augments Voltage-Gated Ca2� Currents andSynergistically Increases Growth Hormone (GH)Secretion with GH-Releasing Hormone in PrimaryCultured Ovine Somatotropes
RUWEI XU, QINLING WANG, MING YAN, MARIA HERNANDEZ, CHANGHONG GONG,WAH CHIN BOON, YOKO MURATA, YOICHI UETA, AND CHEN CHEN
Prince Henry’s Institute of Medical Research (R.X., Q.W., M.Y., M.H., C.G., W.C.B., Y.M., C.C.), Clayton, Victoria 3168,Australia; Department of Physiology, Monash University (R.X., C.C.), Clayton, Victoria 3168, Australia; and Department ofPhysiology, University of Occupational and Environmental Health (Y.U.), Yahatanishi-ku, 807-8555, Kitakyushu, Japan
Orexins are recently discovered neuropeptides that play animportant role in the regulation of hormone secretion, andtheir receptors have been recently demonstrated in the pitu-itary. The effects of orexin-A on voltage-gated Ca2� currentsand GH release in primary cultured ovine somatotropes wereexamined. The expression of orexin-1 receptor was demon-strated by RT-PCR in ovine somatotropes, from which Ca2�
currents were also isolated as L, T, and N currents. Applica-tion of orexin-A (100 nM) significantly and reversibly in-creased only the L current, and coadministration of orexin-Aand GHRH (10 nM) showed an additive effect on this current,but no effect of orexin-A was observed on either T or N current.Furthermore, the orexin-A-induced increase in the L currentwas completely abolished by the inhibition of protein kinase
C (PKC) activity using calphostin C (100 nM), phorbal 12,13-dibutyrate pretreatment (0.5 �M) for 16 h or specific PKC in-hibitory peptide PKC19–36 (1 mM). However, the increase in Lcurrent by orexin-A was sustained when cells were preincu-bated with a specific protein kinase A blocker H89 (1 �M) or aspecific intracellular Ca2� store depleting reagent thapsigar-gin (1 �M). Finally, orexin-A alone did not significantly in-crease GH release, but coadministration of orexin-A andGHRH showed a synergistic effect on GH secretion in vitro.Our results therefore suggest that orexin-A may play an im-portant role in regulating GHRH-stimulated GH secretionthrough the enhancement of the L-type Ca2� current and thePKC-mediated signaling pathway in ovine somatotropes. (En-docrinology 143: 4609–4619, 2002)
OREXIN-A AND -B (also known as hypocretin-1 andhypocretin-2) are two novel hypothalamic peptides
identified by two independent groups that act through twosubtypes of receptors (orexin 1-R and orexin 2-R) belongingto the G protein-coupled receptor superfamily (1, 2).Orexin-A is a peptide with 33 amino acids; orexin-B has 28amino acids. There is 46% sequence homology between thetwo. Orexin 1-R is strongly selective for orexin-A, whereasorexin 2-R has equal affinity for orexin-A and orexin-B (2). Ithas been well established that orexins play a significant rolein the control of energy homeostasis, e.g. food intake andsleep-wake cycle (3, 4). The wide distribution of orexin pep-tides, receptors, and their neuronal projections in the centralnervous system and pituitary revealed in recent studies hasstrongly suggested that the physiological functions of orex-ins reach much beyond the above activities (4–6). One ofsuch important features of orexins is the control of hormonalsecretion. Published studies have shown that orexinsstrongly and specifically activate both hypothalamus-pitu-itary-adrenal and hypothalamus-pituitary-gonadal axes andtherefore influence the secretion of a number of hormones,
including ACTH, LH, corticosterone, and PRL, both in vivoand in vitro (3, 7–11). In addition, the stimulatory effects oforexins on the secretion of insulin and TSH have been re-ported (12, 13).
The presence of orexin-A neuronal projections and its re-ceptors in the pituitary in different species, including hu-mans, indicates that orexins may regulate pituitary hormonesecretion (14, 15). However, the role of orexin-A in the reg-ulation of an important anabolic hormone in the pituitary,GH, is not clear. There have been studies of the possibleregulatory effects of orexins on GH secretion in vivo and invitro, but the results are still limited and controversial (3, 16).Given that GH is integrally linked to energy homeostasis andmetabolism and is closely related to the sleep-wake cycle, weproposed that orexins may play a role in the regulation of GHsecretion by modifying the membrane electrical properties,e.g. ion channels, in somatotropes. In fact, modification of ionchannels in somatotropes is one of the most important mech-anisms by which GH secretion is regulated by hypothalamicpeptides, including GHRH, GH-releasing peptide (GHRP),and somatostatin (17, 18). This hypothesis is also stronglysupported by several lines of evidence. First, orexins andtheir receptors were recently discovered in the anterior lobeof the pituitary in both rats and humans, indicating a possibleparacrine or autocrine effect on pituitary hormones, includ-ing GH (14, 15, 19). Second, orexins have been shown tochange the electrophysiological activities in a number of
Abbreviations: [Ca2�]i, Intracellular calcium concentration; Cal-C,calphostin C; CTX, �-conotoxin; GHRP, GH-releasing peptide; InsP3,inositol trisphosphate; NFD, nefidipine; orexin 1-R, orexin 1 receptor;orexin 2-R, orexin 2 receptor; PDBu, phorbal 12,13-dibutyrate; PKA,protein kinase A; PKC, protein kinase C; RT, reverse transcription; TTX,tetrodotoxin.
0013-7227/02/$15.00/0 Endocrinology 143(12):4609–4619Printed in U.S.A. Copyright © 2002 by The Endocrine Society
doi: 10.1210/en.2002-220506
4609

neuronal and nonneuronal cells (20, 21). For example, orexinshave been recently demonstrated to induce depolarization invagus neurons and reduce the K� currents in macrophages(22, 23). Finally, activation of orexin receptors may causeCa2� influx, and orexins may regulate hormone secretionthrough the adenylate cyclase signaling pathway (24–26).
The existence of Ca2� channels is ubiquitous in all neu-roendocrine cells. Upon appropriate stimulation, the move-ment of Ca2� ions across the cell membrane through Ca2�
channels not only transfers depolarizing charge into excit-able cells, but also initiates specific intracellular signaling tobe decoded by Ca2�-binding proteins, which subsequentlyleads to the operation of their diverse functions. It has longbeen recognized that voltage-gated Ca2� channels play a keyrole in the control of GH secretion from the pituitary (11, 17,27). Previous studies have shown that the modification ofother channels, including K�, Na�, Cl�, and cation channels,may also contribute to the depolarization of the somatotropein different species (28–30). Among these channels, voltage-gated Ca2� channels are particularly important, because theCa2� influx through these channels contributes predomi-nantly to the elevation of the intracellular calcium concen-tration ([Ca2�]i) in somatotropes, leading to GH release (29,31, 32). In ovine somatotropes, it has been shown that GHRHand GHRPs significantly influence GH secretion by modi-fying T- and L-type currents through activation of the ade-nylate cyclase-dependent cascade or protein kinase C (PKC)(17, 33).
In present study we tested the effect of orexin-A becauseit exhibits a stronger influence on the biosynthesis and se-cretion of anterior pituitary hormones than orexin-B (2, 7). Toinvestigate the possible role of orexin-A in the regulation ofGH secretion and the correlated mechanisms, we first stud-ied the effects of orexin-A on the L-type Ca2� current. Afterthe response was confirmed, the interaction betweenorexin-A and GHRH in the modification of L current wasexamined. We also assessed the influence of these two pep-tides on GH secretion by measuring the GH concentrationafter cultured somatotropes were treated with GHRH,orexin-A, or a combination of both. Furthermore, the in-volvement of second messenger systems in orexin-A-induced effects on Ca2� channels was determined.
Materials and MethodsChemicals
Orexin-A was purchased from American Peptide Co. (Sunnyvale,CA) and Peptide Institute (Osaka, Japan). GHRH1–44 was obtained fromAuspep (Parkville, Australia). DMEM, HEPES, and carbohydrate solu-tions were purchased from Trace Biosciences Pty. Ltd. (Noble Park,Australia). Medium 199 and collagenase type I were obtained fromWorthington Biochemical Corp. (Freehold, NJ). Tissue culture reagents(deoxyribonuclease, hyaluronidase, trypsin inhibitor, and pancreatin),sera, ATP, creatine phosphokinase, phosphocreatine, nefidipine (NFD),�-conotoxin (CTX), and all general salts for recording solutions andmolecular biological studies were purchased from Sigma (St. Louis, MO)or as otherwise specified in the text. Tetrodotoxin (TTX) was purchasedfrom Alomone Laboratories (Jerusalem, Israel). l-Glutamine was pur-chased from Life Technologies, Inc. (Gaithersburg, MD). PKC inhibitorypeptides (PKC19–36) and phorbal 12,13-dibutyrate (PDBu) were obtainedfrom Research Chemicals International (Natick, MA). H89 and calphos-tin C (Cal-C) were obtained from Calbiochem (Alexandria, Australia).
Preparation and culture of ovine somatotropes
Adult sheep pituitary glands were collected from a local abattoir andthen subjected to collagenase/pancreatin treatments to dissociate thecells as described previously (34). Briefly, whole pituitaries were di-vested of encapsulating neurohypophysis and pituitary stalk tissues.The anterior pituitaries were then minced and placed in calcium-freePBS with BSA. The tissue fragments were gently washed and incubatedwith deoxyribonuclease, hyaluronidase, trypsin inhibitor, pancreatinand collagenase (�3 mg/ml) for 30–40 min at 37 C in a shaking bath.After centrifugation at 1500 rpm, the cells were suspended in the me-dium 199 and optimally counted under a microscope. Cell yield wasnormally 3 � 107/pituitary gland, with more than 90% viability (trypanblue exclusion test). The cell suspension (3–5 ml) was placed, understerile conditions, above a layer of column of increasing density Percollsolutions. In our experimental conditions, seven Percoll dilutions wereprepared for a discontinuous density gradient as follows: 1.10, 1.074,1.071, 1.068, 1.063, 1.058, 1.040, and 1.029 g/ml. This Percoll gradient wasfurther calibrated using density marker beads (Pharmacia Biotech, Upp-sala, Sweden). Finally, a top layer of suspended cells was loaded gentlyover the Percoll gradient. Tubes then were centrifuged (J6-HC centri-fuge, Beckman, Palo Alto, CA) at 2500 rpm for 30 min at 4 C without abrake, and it was found that about 80% of cells in fractions 2 and 3, withdensity ranging from 1.063–1.071, were somatotropes after immunocy-tochemistry studies were performed (34). Cells were then removed fromeach layer, and the fractions with the most somatotropes were subse-quently seeded into 35-mm culture dishes for electrophysiological stud-ies. Six- and 48-well plates were used to culture cells for molecularbiological studies (RNA extraction and further RT-PCR) and incubationexperiments (GH RIA). Cells were grown in DMEM supplemented with10% fetal calf serum and 1% (vol/vol) l-glutamine (200 mm) in a hu-midified incubator (37 C, 5% CO2). The culture medium was changedevery 2–3 d, and electrophysiological recordings were performed after4–10 d in culture.
RNA isolation and RT-PCR
As the expression of orexin receptors was high in most regions in thebrain (35), we chose rat brain tissue as a positive control. SpragueDawley rats were purchased from Monash University Central AnimalServices. All animal experiments were approved by the ethical com-mittee of Monash Medical Center (Melbourne, Australia). A total of tworats were anesthetized with pentobarbital (100 mg/kg, ip) and killed bydecapitation. Brains were removed immediately and frozen quickly inliquid nitrogen. The total RNA from rat brain was extracted usingRNeasy Mini Kits fitted with a built-in column (QIAGEN, Hilden, Ger-many). Total RNA from cells cultured in petri dishes (�2 � 106 cells/dish) was isolated using the methods previously described (36). Briefly,culture medium from the culture dishes was removed, and cells werewashed with ice-cold PBS. Lysis buffer containing 2% sodium dodecylsulfate, 200 mm Tris-Cl, and 0.5 mm EDTA was added at about 50�l/cm2. After incubation for approximately 2 min, the lysate was trans-ferred to a sterile microfuge tube, and 150 �l potassium acetate solution(50 g potassium acetate, 11 ml glacial acetic acid, and water to 100 ml)were added. The tube was incubated on ice for 3–5 min and centrifugedat maximum speed in a microcentrifuge. The supernatant containing theRNA was further extracted with a mixture of chloroform/isomyl alcohol(24:1). The upper phase was removed and reextracted. The RNA wasfinally precipitated with an equal volume of ice-cold isopropanol on icefor 20–30 min and then pelleted after centrifugation and washing with70% ethanol. One microgram of total RNA extracted from each cellculture well was treated with deoxyribonuclease I (Roche, Indianapolis,IN) to eliminate possible contamination of genomic DNA.
One microgram of RNA from rat brain or ovine somatotropes wasthen reverse transcribed to cDNA in a 20-�l reverse transcription (RT)reaction system containing random primers and avian myeloblastosisvirus reverse transcriptase (Roche). The RT reactions were carried outat 46 C for 2 h. One microliter of the RT reaction products was used forsubsequent PCR amplification for 35 cycles. Primer for ovine orexin-1receptor was designed according to the rat cDNA templates with anexpected size of 310 bp (37). The set of primers for the amplification ofthe orexin-1 receptor was 5�-AATCGCACACGGCTCTTCTCTGTC(sense) and 5�-CACCATCAGCATCTTGGCAGTC (antisense). We used
4610 Endocrinology, December 2002, 143(12):4609–4619 Xu et al. • Orexin-A and Somatotropes

a denaturation step at 95 C for 1 min, an annealing step at 56 C for 1 min,and an extension step at 72 C for 1 min for a total of 35 cycles, followedby an additional extension step at 72 C for 10 min. Detection of PCRamplification products was carried out by electrophoresis (1.5% agarosegel containing ethidium bromide). The PCR products were extractedfrom the gel and sequenced in an automated DNA sequencer. Finally,the sequenced data were analyzed using Blast Nucleic Acid DatabaseSearches from the National Center for Biotechnology Information.
Electrophysiological recording
On the day of the recording, culture medium was replaced by patch-clamp bath solution through a gravity pressure perfusion system at least10 min before recording. Transmembrane Ca2� currents were recordedusing the gigaseal patch clamp technique in classic whole cell recordingconfiguration. Electrodes were pulled by a Sutter P-87 microelectrodepuller (Sutter Instrument Co., Novato, CA) from borosilicate micropi-pettes with inner filament and had an initial input resistance of 2–5 m�.All recordings were made using the Axopatch 200A amplifier (AxonInstruments, Foster City, CA). The bath solution was composed of thefollowing: 40 mm tetraethyl ammonium chloride, 90 mm NaCl, 5 mmCaCl2, 0.5 mm MgCl2, 10 mm glucose, and 10 mm HEPES (pH 7.4,adjusted with NaOH; osmolarity of 300 mosmol/liter with sucrose). Toexclude the contamination of Na� current, TTX was added to the bathsolution with a final concentration of 1 �m on the day of experimen-tation. The pipette solution was composed of the following: 120 mmCsAsp, 20 mm tetraethyl ammonium chloride, 10 mm EGTA, 10 mmglucose, and 10 mm HEPES. Just before recording, an ATP regenerativesystem (2 mm ATP, 5 mm Na2-phosphocreatine, and 20 U/ml creatinephosphokinase) plus 0.1 mm GTP were added to the pipette solution (pHadjusted to 7.4 and osmolarity to 300 mosmol/liter). PKC19–36 was alsoincluded in the pipette solution in studies of the involvement of the PKCpathway in orexin-A-induced effects in ovine somatotropes.
After obtaining the high resistance seal, the pipette potential was heldto �80 mV, and voltage pulses (10 mV, 200-msec duration) were de-livered periodically to monitor access resistance. Access to the cell in-terior was judged by the appearance of a membrane capacitative tran-sient current after a gentle, but quick, suction under our experimentalcondition. Whole cell capacitance (7.9 � 0.3 pF; n � 79) and seriesresistance were compensated (�80%) before experimentation, and leakcurrent was routinely subtracted using the option offered by theClampex 7.0 program (Axon Instruments). We also monitored thechange in series resistance over the course of each experiment, andrecordings with significant change in series resistance were terminatedor excluded from the final data analysis. The signals were filtered at 2kHz, and the sweeps were sampled at 1-msec intervals in our recordingprotocols.
Cell culture dishes were fixed on the stage of an Olympus Corp.inverted microscope (New Hyde Park, NY), and a gravity pressuresystem was used to perfuse the cells at a rate of approximately 1 ml/min.Protein kinase A (PKA)-cAMP and PKC blockers, including H89 andCal-C, were added by hand to the culture dishes containing the cells tobe recorded. Recordings started at least 5 min after these blockers wereadded to achieve an even distribution of these chemicals (the concen-trations cited in Results are the final concentrations when diluted in thebath solution). Orexin-A was mixed with bath solution and applied bygravity pressure perfusion system. Application of vehicle from the samesystem did not change Ca2� currents. All experiments were performedat room temperature (20–22 C).
Incubation procedures and RIA
Before each incubation experiment, the cells were washed three timeswith incubation medium (medium 199 containing 0.1% BSA) and thenpreincubated for 1 h with incubation medium as during the equilibrationperiod. The medium was subsequently discarded after 1-h incubation,and 2 ml/well fresh incubation medium, with or without the test sub-stances, were added for 30 min. Parallel incubations containing a solventof drugs alone were included in the incubation as a control. At the endof the incubation period, the conditioned medium was collected andstored at –20 C before the GH RIA.
The concentration of GH in the incubation medium was measured ina double antibody RIA using kits provided by the National Hormone
and Pituitary Program, NIH (ovine GH and ovine GH antisera). Allsamples were assayed in duplicate. The sensitivity of the assay was 0.3ng/ml. The inter- and intraassay coefficients of variation were less than15% and 8%, respectively (n � 4). All samples from one experiment weremeasured in the same assay, and GH values were expressed as nano-gram equivalents of ovine GH standard.
Data analysis
A pCLAMP 7.0 software (Axon Instruments) was used to acquire andanalyze the data. A paired t test was used as appropriate to evaluate thestatistical significance of differences between two group means of cur-rents, and the effects were considered significant at P 0.05. Group dataare expressed as the mean � sem in Results. The traces in the figures arerepresentative of at least four recordings under the same experimentalconditions or as indicated otherwise in the text. GH data in incubationstudies are also presented as the mean � sem. Statistical comparisonswere made by t test, and significance was taken as P 0.05.
ResultsPresence of orexin-1 receptor in purified ovine pituitary cells
RT-PCR and sequence analysis demonstrated that orexin1-R was strongly expressed in both ovine somatotropes andrat brain tissue. As shown in Fig. 1, no amplification wasdetected in the negative control, i.e. without RT, hence con-firming that the PCR product did not come from genomicDNA contamination. The PCR product was extracted, and apartial sequence for ovine pituitary cells was obtained. The
FIG. 1. Expression of orexin-1 receptor in ovine somatotropes.Ethidium bromide-stained 1.5% agarose gel showing cDNA amplifiedwith orexin 1-R receptor primer from RNA of ovine anterior pituitarycells (predominantly somatotropes) and rat brain tissue. The left lanewas loaded with molecular marker (Roche). The size of amplifiedfragments was 310 bp. No amplification was detectable in the neg-ative control, i.e. when avian myeloblastosis virus reverse transcrip-tase was omitted in the RT reaction mix.
Xu et al. • Orexin-A and Somatotropes Endocrinology, December 2002, 143(12):4609–4619 4611
ovine sequence showed 87% and 89% homology to rat andhuman orexin-1 receptor genes, respectively.
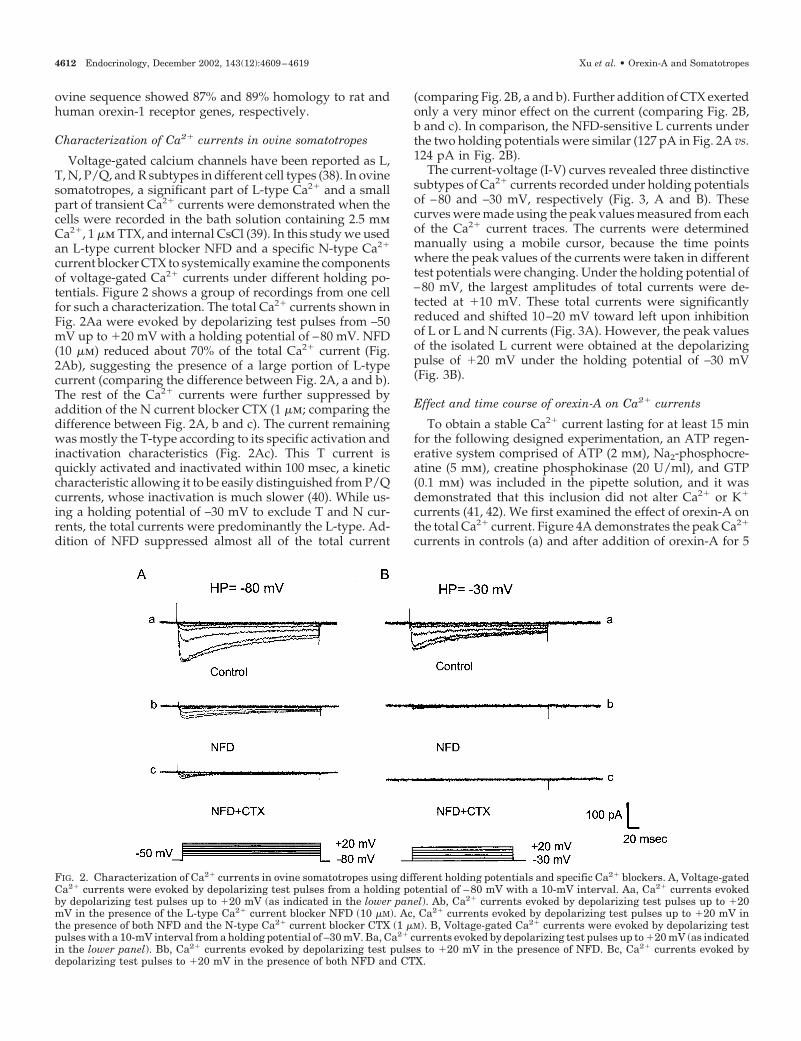
Characterization of Ca2� currents in ovine somatotropes
Voltage-gated calcium channels have been reported as L,T, N, P/Q, and R subtypes in different cell types (38). In ovinesomatotropes, a significant part of L-type Ca2� and a smallpart of transient Ca2� currents were demonstrated when thecells were recorded in the bath solution containing 2.5 mmCa2�, 1 �m TTX, and internal CsCl (39). In this study we usedan L-type current blocker NFD and a specific N-type Ca2�
current blocker CTX to systemically examine the componentsof voltage-gated Ca2� currents under different holding po-tentials. Figure 2 shows a group of recordings from one cellfor such a characterization. The total Ca2� currents shown inFig. 2Aa were evoked by depolarizing test pulses from –50mV up to �20 mV with a holding potential of –80 mV. NFD(10 �m) reduced about 70% of the total Ca2� current (Fig.2Ab), suggesting the presence of a large portion of L-typecurrent (comparing the difference between Fig. 2A, a and b).The rest of the Ca2� currents were further suppressed byaddition of the N current blocker CTX (1 �m; comparing thedifference between Fig. 2A, b and c). The current remainingwas mostly the T-type according to its specific activation andinactivation characteristics (Fig. 2Ac). This T current isquickly activated and inactivated within 100 msec, a kineticcharacteristic allowing it to be easily distinguished from P/Qcurrents, whose inactivation is much slower (40). While us-ing a holding potential of –30 mV to exclude T and N cur-rents, the total currents were predominantly the L-type. Ad-dition of NFD suppressed almost all of the total current
(comparing Fig. 2B, a and b). Further addition of CTX exertedonly a very minor effect on the current (comparing Fig. 2B,b and c). In comparison, the NFD-sensitive L currents underthe two holding potentials were similar (127 pA in Fig. 2A vs.124 pA in Fig. 2B).
The current-voltage (I-V) curves revealed three distinctivesubtypes of Ca2� currents recorded under holding potentialsof –80 and –30 mV, respectively (Fig. 3, A and B). Thesecurves were made using the peak values measured from eachof the Ca2� current traces. The currents were determinedmanually using a mobile cursor, because the time pointswhere the peak values of the currents were taken in differenttest potentials were changing. Under the holding potential of–80 mV, the largest amplitudes of total currents were de-tected at �10 mV. These total currents were significantlyreduced and shifted 10–20 mV toward left upon inhibitionof L or L and N currents (Fig. 3A). However, the peak valuesof the isolated L current were obtained at the depolarizingpulse of �20 mV under the holding potential of –30 mV(Fig. 3B).
Effect and time course of orexin-A on Ca2� currents
To obtain a stable Ca2� current lasting for at least 15 minfor the following designed experimentation, an ATP regen-erative system comprised of ATP (2 mm), Na2-phosphocre-atine (5 mm), creatine phosphokinase (20 U/ml), and GTP(0.1 mm) was included in the pipette solution, and it wasdemonstrated that this inclusion did not alter Ca2� or K�
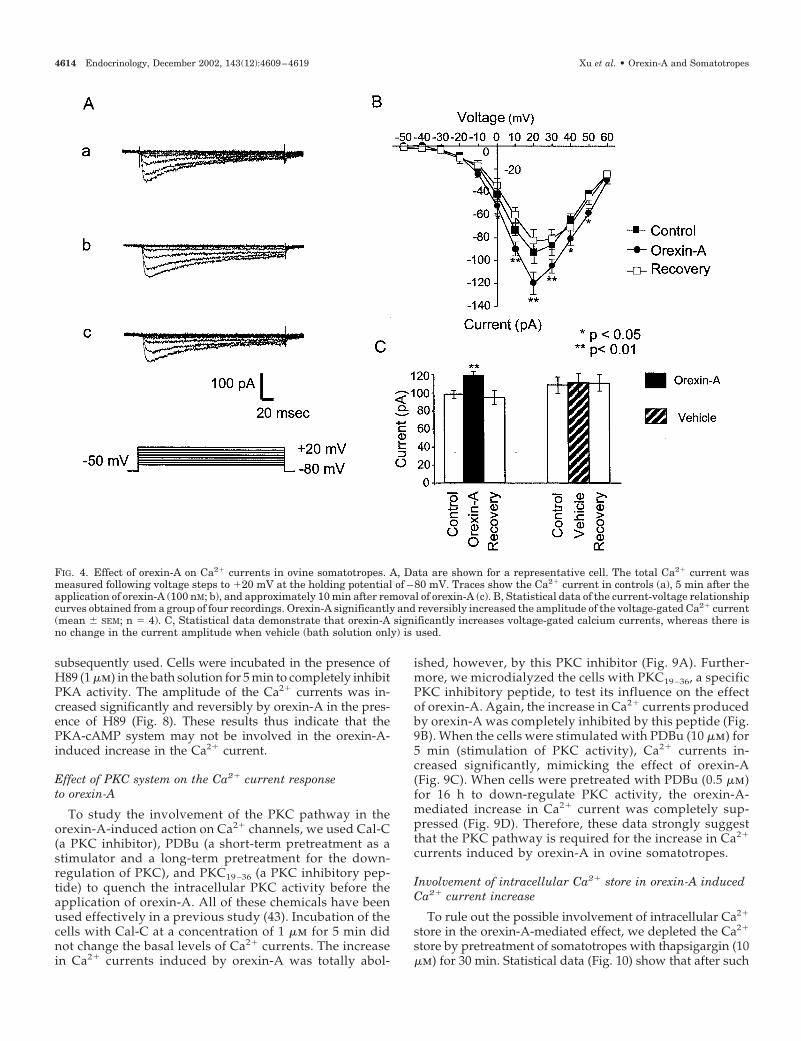
currents (41, 42). We first examined the effect of orexin-A onthe total Ca2� current. Figure 4A demonstrates the peak Ca2�
currents in controls (a) and after addition of orexin-A for 5
FIG. 2. Characterization of Ca2� currents in ovine somatotropes using different holding potentials and specific Ca2� blockers. A, Voltage-gatedCa2� currents were evoked by depolarizing test pulses from a holding potential of –80 mV with a 10-mV interval. Aa, Ca2� currents evokedby depolarizing test pulses up to �20 mV (as indicated in the lower panel). Ab, Ca2� currents evoked by depolarizing test pulses up to �20mV in the presence of the L-type Ca2� current blocker NFD (10 �M). Ac, Ca2� currents evoked by depolarizing test pulses up to �20 mV inthe presence of both NFD and the N-type Ca2� current blocker CTX (1 �M). B, Voltage-gated Ca2� currents were evoked by depolarizing testpulses with a 10-mV interval from a holding potential of –30 mV. Ba, Ca2� currents evoked by depolarizing test pulses up to �20 mV (as indicatedin the lower panel). Bb, Ca2� currents evoked by depolarizing test pulses to �20 mV in the presence of NFD. Bc, Ca2� currents evoked bydepolarizing test pulses to �20 mV in the presence of both NFD and CTX.
4612 Endocrinology, December 2002, 143(12):4609–4619 Xu et al. • Orexin-A and Somatotropes

min (b), and the recovery of Ca2� currents after orexin-A waswashed away (c). Application of orexin-A significantly andreversibly increased voltage-gated Ca2� currents. The stim-ulatory effect of orexin-A on total Ca2� currents was alsodemonstrated in the I-V curves (Fig. 4B). To exclude thesystemic variations, we performed the entire procedure us-ing bath solution as a placebo. The statistical data indicatethat orexin-A significantly and reversibly increased Ca2�
currents, whereas no influence from the perfusion systemwas evident (Fig. 4C). The time course of the response toorexin-A is also shown in Fig. 5. The stimulatory effect oforexin-A on the Ca2� currents took place gradually andreached a maximum in about 4 min. The complete recoveryof Ca2� currents required approximately 4–6 min after theremoval of orexin-A. Meanwhile, no significant “rundown”of Ca2� currents was observed during the experiment (Fig.5, control).
Effect of orexin-A on the L-type Ca2� current
As orexin-B increases only the L current in ovine soma-totropes (Xu, R., manuscript in preparation), we extendedour studies by examining the effect of orexin-A on this cur-rent. To isolate the L-type Ca2� current, the N-type Ca2�
current blocker CTX (10 �m) was added to bath solutionwhen the cells were recorded with a holding potential of –30mV to exclude the T current. The isolated L-type current wasthe largest portion of the total currents (�60% of the ampli-tude of the total currents) in ovine somatotropes, andorexin-A (100 nm) significantly and reversibly increased theL-type current (Fig. 6). No effect was observed in the N or Tcurrent from the addition of orexin-A at the same concen-tration (data not shown). As only the L-type current wasaltered by orexin-A, the total Ca2� current using the holdingpotential of –80 mV was used in the following experiments.Meanwhile, the Ca2� currents recorded under this holding
potential of –80 mV were found to be more stable than thoseunder the holding potential of –30 mV.
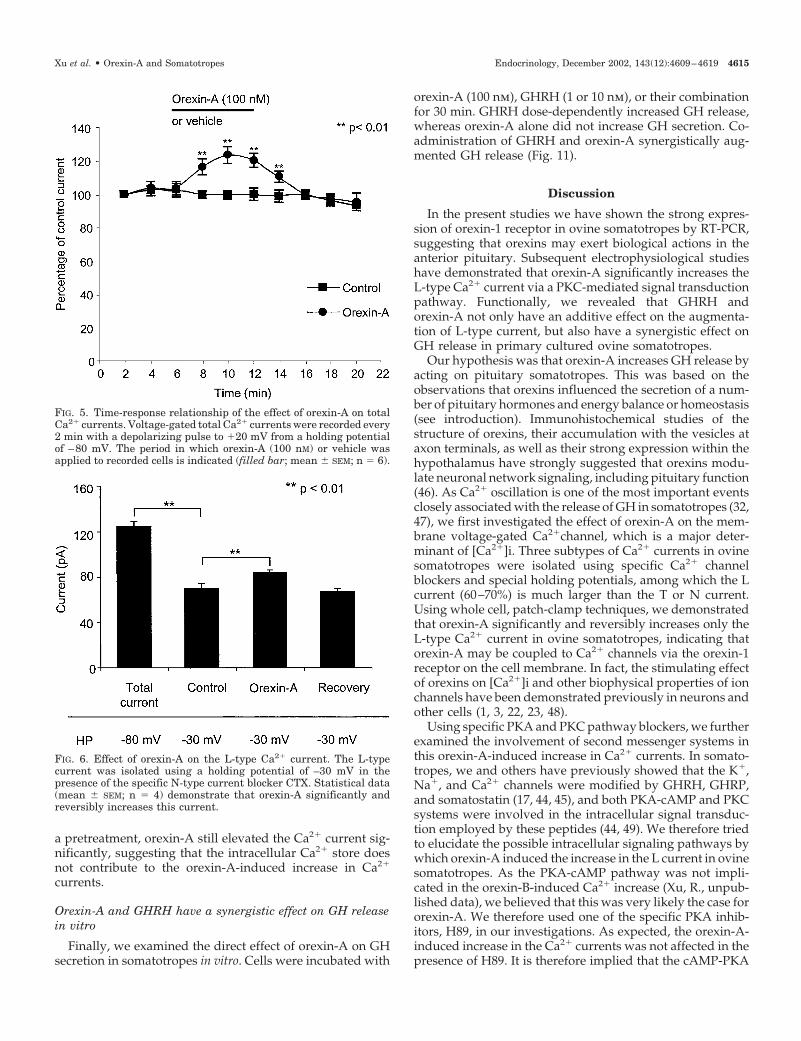
Orexin-A and GHRH additively increase Ca2� currents
As Ca2� currents have been previously demonstrated tobe increased by GHRH in ovine somatotropes, we testedthe interactions between orexin-A and GHRH (100 nm) inthe modification of the Ca2� current. To exclude the pos-sible cross-effect between the receptors of orexin-A andGHRH, we administered orexin-A (100 nm) and GHRH indifferent orders. In the first series of experiments GHRHwas delivered 8 min before orexin-A. GHRH increased theCa2� current significantly, and subsequent addition oforexin-A further elevated the level of Ca2� currents (Fig.7A). When orexin-A was added 8 min before GHRH,orexin-A alone significantly augmented the Ca2� currents,and addition of GHRH further enhanced the Ca2� currents(Fig. 7B). Complete recovery was achieved in both exper-imental groups. The total increase in Ca2� currents pro-duced by orexin-A and GHRH was similar between thetwo sets of experiments.
Involvement of the PKA-cAMP system in the Ca2� currentresponse to orexin-A
The involvement of second messenger systems in soma-totropes was investigated using specific PKA-cAMP block-ers. These blockers have been previously successfully em-ployed to examine the effect of a GHRH-induced change inthe Ca2� and K� currents in somatotropes (43–45). The spe-cific blocking effects of H89 and other inhibitors on the PKAsignaling pathway have been examined by us in a recentstudy of the effect of GHRP-2 on inward rectifying K� cur-rents (Xu, R., unpublished data) and by others (44). In thisstudy a cell-permeable, specific PKA inhibitor, H89, was
FIG. 3. Current-voltage (I-V) relation-ships of three subtypes of Ca2� currentsin ovine somatotropes. Currents mea-sured at the peak of each trace. I-Vcurves with different holding potentialscorrespond to the recording conditionsin Fig. 2, A and B, respectively (mean �SEM; n � 4). Note that under the holdingpotential of –80 mV (A), total Ca2� cur-rent (�), T- plus N-type (f), and T-typecurrents (indicated as Œ) were recorded,whereas under –30 mV (B), the majorityof the current was L-type (�).
Xu et al. • Orexin-A and Somatotropes Endocrinology, December 2002, 143(12):4609–4619 4613
subsequently used. Cells were incubated in the presence ofH89 (1 �m) in the bath solution for 5 min to completely inhibitPKA activity. The amplitude of the Ca2� currents was in-creased significantly and reversibly by orexin-A in the pres-ence of H89 (Fig. 8). These results thus indicate that thePKA-cAMP system may not be involved in the orexin-A-induced increase in the Ca2� current.
Effect of PKC system on the Ca2� current responseto orexin-A
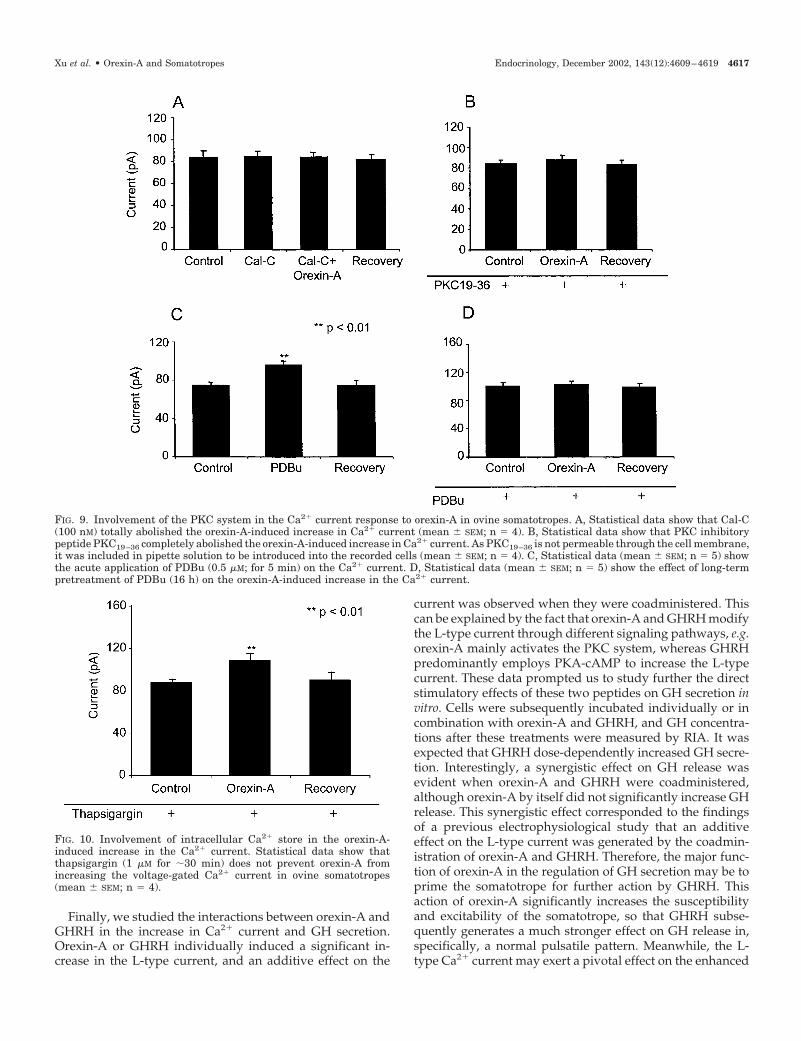
To study the involvement of the PKC pathway in theorexin-A-induced action on Ca2� channels, we used Cal-C(a PKC inhibitor), PDBu (a short-term pretreatment as astimulator and a long-term pretreatment for the down-regulation of PKC), and PKC19 –36 (a PKC inhibitory pep-tide) to quench the intracellular PKC activity before theapplication of orexin-A. All of these chemicals have beenused effectively in a previous study (43). Incubation of thecells with Cal-C at a concentration of 1 �m for 5 min didnot change the basal levels of Ca2� currents. The increasein Ca2� currents induced by orexin-A was totally abol-
ished, however, by this PKC inhibitor (Fig. 9A). Further-more, we microdialyzed the cells with PKC19 –36, a specificPKC inhibitory peptide, to test its influence on the effectof orexin-A. Again, the increase in Ca2� currents producedby orexin-A was completely inhibited by this peptide (Fig.9B). When the cells were stimulated with PDBu (10 �m) for5 min (stimulation of PKC activity), Ca2� currents in-creased significantly, mimicking the effect of orexin-A(Fig. 9C). When cells were pretreated with PDBu (0.5 �m)for 16 h to down-regulate PKC activity, the orexin-A-mediated increase in Ca2� current was completely sup-pressed (Fig. 9D). Therefore, these data strongly suggestthat the PKC pathway is required for the increase in Ca2�
currents induced by orexin-A in ovine somatotropes.
Involvement of intracellular Ca2� store in orexin-A inducedCa2� current increase
To rule out the possible involvement of intracellular Ca2�
store in the orexin-A-mediated effect, we depleted the Ca2�
store by pretreatment of somatotropes with thapsigargin (10�m) for 30 min. Statistical data (Fig. 10) show that after such
FIG. 4. Effect of orexin-A on Ca2� currents in ovine somatotropes. A, Data are shown for a representative cell. The total Ca2� current wasmeasured following voltage steps to �20 mV at the holding potential of –80 mV. Traces show the Ca2� current in controls (a), 5 min after theapplication of orexin-A (100 nM; b), and approximately 10 min after removal of orexin-A (c). B, Statistical data of the current-voltage relationshipcurves obtained from a group of four recordings. Orexin-A significantly and reversibly increased the amplitude of the voltage-gated Ca2� current(mean � SEM; n � 4). C, Statistical data demonstrate that orexin-A significantly increases voltage-gated calcium currents, whereas there isno change in the current amplitude when vehicle (bath solution only) is used.
4614 Endocrinology, December 2002, 143(12):4609–4619 Xu et al. • Orexin-A and Somatotropes
a pretreatment, orexin-A still elevated the Ca2� current sig-nificantly, suggesting that the intracellular Ca2� store doesnot contribute to the orexin-A-induced increase in Ca2�
currents.
Orexin-A and GHRH have a synergistic effect on GH releasein vitro
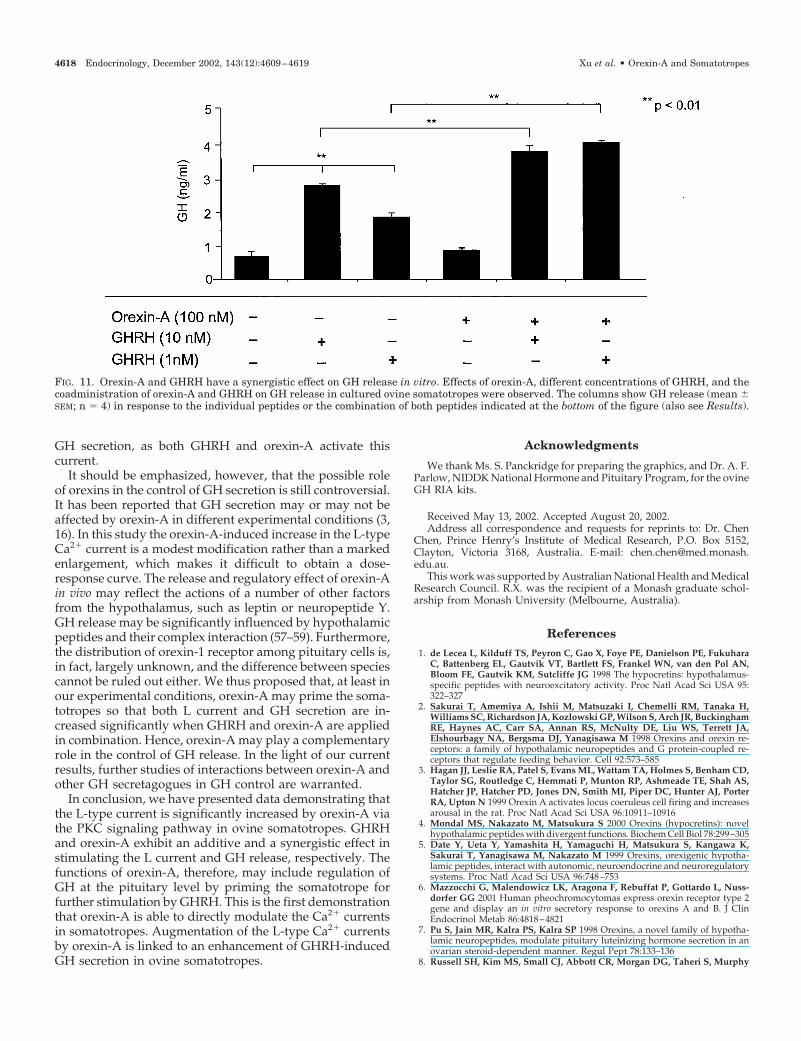
Finally, we examined the direct effect of orexin-A on GHsecretion in somatotropes in vitro. Cells were incubated with
orexin-A (100 nm), GHRH (1 or 10 nm), or their combinationfor 30 min. GHRH dose-dependently increased GH release,whereas orexin-A alone did not increase GH secretion. Co-administration of GHRH and orexin-A synergistically aug-mented GH release (Fig. 11).
Discussion
In the present studies we have shown the strong expres-sion of orexin-1 receptor in ovine somatotropes by RT-PCR,suggesting that orexins may exert biological actions in theanterior pituitary. Subsequent electrophysiological studieshave demonstrated that orexin-A significantly increases theL-type Ca2� current via a PKC-mediated signal transductionpathway. Functionally, we revealed that GHRH andorexin-A not only have an additive effect on the augmenta-tion of L-type current, but also have a synergistic effect onGH release in primary cultured ovine somatotropes.
Our hypothesis was that orexin-A increases GH release byacting on pituitary somatotropes. This was based on theobservations that orexins influenced the secretion of a num-ber of pituitary hormones and energy balance or homeostasis(see introduction). Immunohistochemical studies of thestructure of orexins, their accumulation with the vesicles ataxon terminals, as well as their strong expression within thehypothalamus have strongly suggested that orexins modu-late neuronal network signaling, including pituitary function(46). As Ca2� oscillation is one of the most important eventsclosely associated with the release of GH in somatotropes (32,47), we first investigated the effect of orexin-A on the mem-brane voltage-gated Ca2�channel, which is a major deter-minant of [Ca2�]i. Three subtypes of Ca2� currents in ovinesomatotropes were isolated using specific Ca2� channelblockers and special holding potentials, among which the Lcurrent (60–70%) is much larger than the T or N current.Using whole cell, patch-clamp techniques, we demonstratedthat orexin-A significantly and reversibly increases only theL-type Ca2� current in ovine somatotropes, indicating thatorexin-A may be coupled to Ca2� channels via the orexin-1receptor on the cell membrane. In fact, the stimulating effectof orexins on [Ca2�]i and other biophysical properties of ionchannels have been demonstrated previously in neurons andother cells (1, 3, 22, 23, 48).
Using specific PKA and PKC pathway blockers, we furtherexamined the involvement of second messenger systems inthis orexin-A-induced increase in Ca2� currents. In somato-tropes, we and others have previously showed that the K�,Na�, and Ca2� channels were modified by GHRH, GHRP,and somatostatin (17, 44, 45), and both PKA-cAMP and PKCsystems were involved in the intracellular signal transduc-tion employed by these peptides (44, 49). We therefore triedto elucidate the possible intracellular signaling pathways bywhich orexin-A induced the increase in the L current in ovinesomatotropes. As the PKA-cAMP pathway was not impli-cated in the orexin-B-induced Ca2� increase (Xu, R., unpub-lished data), we believed that this was very likely the case fororexin-A. We therefore used one of the specific PKA inhib-itors, H89, in our investigations. As expected, the orexin-A-induced increase in the Ca2� currents was not affected in thepresence of H89. It is therefore implied that the cAMP-PKA
FIG. 5. Time-response relationship of the effect of orexin-A on totalCa2� currents. Voltage-gated total Ca2� currents were recorded every2 min with a depolarizing pulse to �20 mV from a holding potentialof –80 mV. The period in which orexin-A (100 nM) or vehicle wasapplied to recorded cells is indicated (filled bar; mean � SEM; n � 6).
FIG. 6. Effect of orexin-A on the L-type Ca2� current. The L-typecurrent was isolated using a holding potential of –30 mV in thepresence of the specific N-type current blocker CTX. Statistical data(mean � SEM; n � 4) demonstrate that orexin-A significantly andreversibly increases this current.
Xu et al. • Orexin-A and Somatotropes Endocrinology, December 2002, 143(12):4609–4619 4615
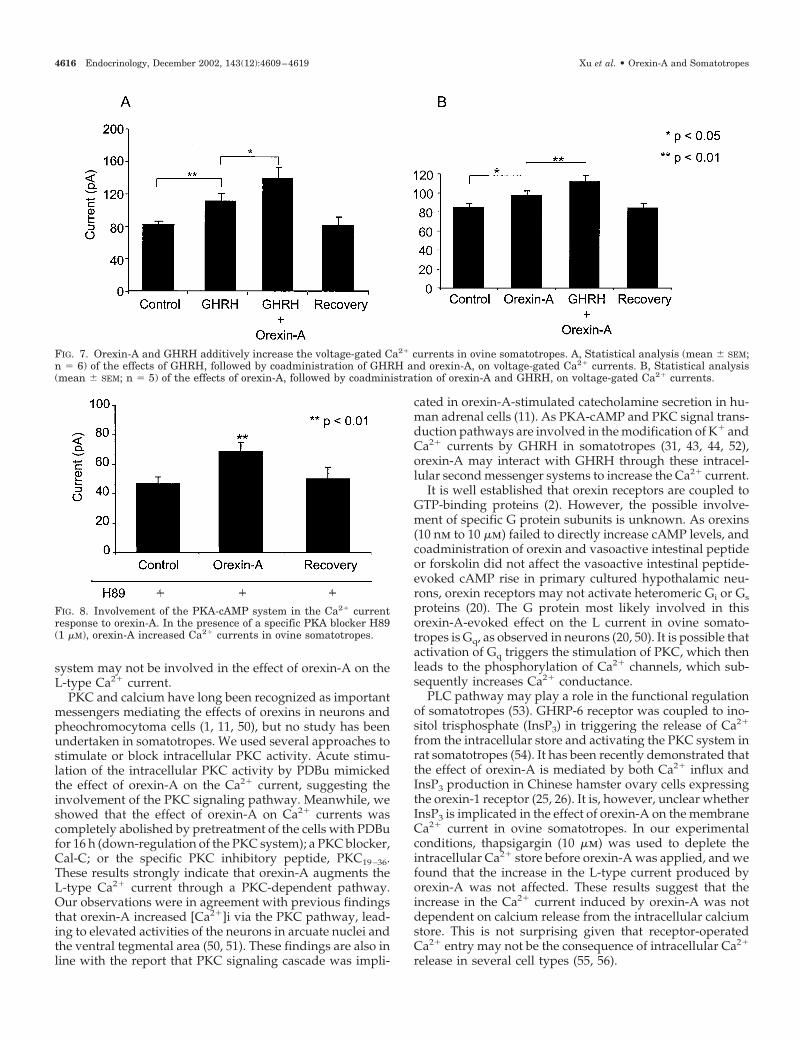
system may not be involved in the effect of orexin-A on theL-type Ca2� current.
PKC and calcium have long been recognized as importantmessengers mediating the effects of orexins in neurons andpheochromocytoma cells (1, 11, 50), but no study has beenundertaken in somatotropes. We used several approaches tostimulate or block intracellular PKC activity. Acute stimu-lation of the intracellular PKC activity by PDBu mimickedthe effect of orexin-A on the Ca2� current, suggesting theinvolvement of the PKC signaling pathway. Meanwhile, weshowed that the effect of orexin-A on Ca2� currents wascompletely abolished by pretreatment of the cells with PDBufor 16 h (down-regulation of the PKC system); a PKC blocker,Cal-C; or the specific PKC inhibitory peptide, PKC19–36.These results strongly indicate that orexin-A augments theL-type Ca2� current through a PKC-dependent pathway.Our observations were in agreement with previous findingsthat orexin-A increased [Ca2�]i via the PKC pathway, lead-ing to elevated activities of the neurons in arcuate nuclei andthe ventral tegmental area (50, 51). These findings are also inline with the report that PKC signaling cascade was impli-
cated in orexin-A-stimulated catecholamine secretion in hu-man adrenal cells (11). As PKA-cAMP and PKC signal trans-duction pathways are involved in the modification of K� andCa2� currents by GHRH in somatotropes (31, 43, 44, 52),orexin-A may interact with GHRH through these intracel-lular second messenger systems to increase the Ca2� current.
It is well established that orexin receptors are coupled toGTP-binding proteins (2). However, the possible involve-ment of specific G protein subunits is unknown. As orexins(10 nm to 10 �m) failed to directly increase cAMP levels, andcoadministration of orexin and vasoactive intestinal peptideor forskolin did not affect the vasoactive intestinal peptide-evoked cAMP rise in primary cultured hypothalamic neu-rons, orexin receptors may not activate heteromeric Gi or Gsproteins (20). The G protein most likely involved in thisorexin-A-evoked effect on the L current in ovine somato-tropes is Gq, as observed in neurons (20, 50). It is possible thatactivation of Gq triggers the stimulation of PKC, which thenleads to the phosphorylation of Ca2� channels, which sub-sequently increases Ca2� conductance.
PLC pathway may play a role in the functional regulationof somatotropes (53). GHRP-6 receptor was coupled to ino-sitol trisphosphate (InsP3) in triggering the release of Ca2�
from the intracellular store and activating the PKC system inrat somatotropes (54). It has been recently demonstrated thatthe effect of orexin-A is mediated by both Ca2� influx andInsP3 production in Chinese hamster ovary cells expressingthe orexin-1 receptor (25, 26). It is, however, unclear whetherInsP3 is implicated in the effect of orexin-A on the membraneCa2� current in ovine somatotropes. In our experimentalconditions, thapsigargin (10 �m) was used to deplete theintracellular Ca2� store before orexin-A was applied, and wefound that the increase in the L-type current produced byorexin-A was not affected. These results suggest that theincrease in the Ca2� current induced by orexin-A was notdependent on calcium release from the intracellular calciumstore. This is not surprising given that receptor-operatedCa2� entry may not be the consequence of intracellular Ca2�
release in several cell types (55, 56).
FIG. 7. Orexin-A and GHRH additively increase the voltage-gated Ca2� currents in ovine somatotropes. A, Statistical analysis (mean � SEM;n � 6) of the effects of GHRH, followed by coadministration of GHRH and orexin-A, on voltage-gated Ca2� currents. B, Statistical analysis(mean � SEM; n � 5) of the effects of orexin-A, followed by coadministration of orexin-A and GHRH, on voltage-gated Ca2� currents.
FIG. 8. Involvement of the PKA-cAMP system in the Ca2� currentresponse to orexin-A. In the presence of a specific PKA blocker H89(1 �M), orexin-A increased Ca2� currents in ovine somatotropes.
4616 Endocrinology, December 2002, 143(12):4609–4619 Xu et al. • Orexin-A and Somatotropes
Finally, we studied the interactions between orexin-A andGHRH in the increase in Ca2� current and GH secretion.Orexin-A or GHRH individually induced a significant in-crease in the L-type current, and an additive effect on the
current was observed when they were coadministered. Thiscan be explained by the fact that orexin-A and GHRH modifythe L-type current through different signaling pathways, e.g.orexin-A mainly activates the PKC system, whereas GHRHpredominantly employs PKA-cAMP to increase the L-typecurrent. These data prompted us to study further the directstimulatory effects of these two peptides on GH secretion invitro. Cells were subsequently incubated individually or incombination with orexin-A and GHRH, and GH concentra-tions after these treatments were measured by RIA. It wasexpected that GHRH dose-dependently increased GH secre-tion. Interestingly, a synergistic effect on GH release wasevident when orexin-A and GHRH were coadministered,although orexin-A by itself did not significantly increase GHrelease. This synergistic effect corresponded to the findingsof a previous electrophysiological study that an additiveeffect on the L-type current was generated by the coadmin-istration of orexin-A and GHRH. Therefore, the major func-tion of orexin-A in the regulation of GH secretion may be toprime the somatotrope for further action by GHRH. Thisaction of orexin-A significantly increases the susceptibilityand excitability of the somatotrope, so that GHRH subse-quently generates a much stronger effect on GH release in,specifically, a normal pulsatile pattern. Meanwhile, the L-type Ca2� current may exert a pivotal effect on the enhanced
FIG. 9. Involvement of the PKC system in the Ca2� current response to orexin-A in ovine somatotropes. A, Statistical data show that Cal-C(100 nM) totally abolished the orexin-A-induced increase in Ca2� current (mean � SEM; n � 4). B, Statistical data show that PKC inhibitorypeptide PKC19–36 completely abolished the orexin-A-induced increase in Ca2� current. As PKC19–36 is not permeable through the cell membrane,it was included in pipette solution to be introduced into the recorded cells (mean � SEM; n � 4). C, Statistical data (mean � SEM; n � 5) showthe acute application of PDBu (0.5 �M; for 5 min) on the Ca2� current. D, Statistical data (mean � SEM; n � 5) show the effect of long-termpretreatment of PDBu (16 h) on the orexin-A-induced increase in the Ca2� current.
FIG. 10. Involvement of intracellular Ca2� store in the orexin-A-induced increase in the Ca2� current. Statistical data show thatthapsigargin (1 �M for �30 min) does not prevent orexin-A fromincreasing the voltage-gated Ca2� current in ovine somatotropes(mean � SEM; n � 4).
Xu et al. • Orexin-A and Somatotropes Endocrinology, December 2002, 143(12):4609–4619 4617
GH secretion, as both GHRH and orexin-A activate thiscurrent.
It should be emphasized, however, that the possible roleof orexins in the control of GH secretion is still controversial.It has been reported that GH secretion may or may not beaffected by orexin-A in different experimental conditions (3,16). In this study the orexin-A-induced increase in the L-typeCa2� current is a modest modification rather than a markedenlargement, which makes it difficult to obtain a dose-response curve. The release and regulatory effect of orexin-Ain vivo may reflect the actions of a number of other factorsfrom the hypothalamus, such as leptin or neuropeptide Y.GH release may be significantly influenced by hypothalamicpeptides and their complex interaction (57–59). Furthermore,the distribution of orexin-1 receptor among pituitary cells is,in fact, largely unknown, and the difference between speciescannot be ruled out either. We thus proposed that, at least inour experimental conditions, orexin-A may prime the soma-totropes so that both L current and GH secretion are in-creased significantly when GHRH and orexin-A are appliedin combination. Hence, orexin-A may play a complementaryrole in the control of GH release. In the light of our currentresults, further studies of interactions between orexin-A andother GH secretagogues in GH control are warranted.
In conclusion, we have presented data demonstrating thatthe L-type current is significantly increased by orexin-A viathe PKC signaling pathway in ovine somatotropes. GHRHand orexin-A exhibit an additive and a synergistic effect instimulating the L current and GH release, respectively. Thefunctions of orexin-A, therefore, may include regulation ofGH at the pituitary level by priming the somatotrope forfurther stimulation by GHRH. This is the first demonstrationthat orexin-A is able to directly modulate the Ca2� currentsin somatotropes. Augmentation of the L-type Ca2� currentsby orexin-A is linked to an enhancement of GHRH-inducedGH secretion in ovine somatotropes.
Acknowledgments
We thank Ms. S. Panckridge for preparing the graphics, and Dr. A. F.Parlow, NIDDK National Hormone and Pituitary Program, for the ovineGH RIA kits.
Received May 13, 2002. Accepted August 20, 2002.Address all correspondence and requests for reprints to: Dr. Chen
Chen, Prince Henry’s Institute of Medical Research, P.O. Box 5152,Clayton, Victoria 3168, Australia. E-mail: [email protected].
This work was supported by Australian National Health and MedicalResearch Council. R.X. was the recipient of a Monash graduate schol-arship from Monash University (Melbourne, Australia).
References
1. de Lecea L, Kilduff TS, Peyron C, Gao X, Foye PE, Danielson PE, FukuharaC, Battenberg EL, Gautvik VT, Bartlett FS, Frankel WN, van den Pol AN,Bloom FE, Gautvik KM, Sutcliffe JG 1998 The hypocretins: hypothalamus-specific peptides with neuroexcitatory activity. Proc Natl Acad Sci USA 95:322–327
2. Sakurai T, Amemiya A, Ishii M, Matsuzaki I, Chemelli RM, Tanaka H,Williams SC, Richardson JA, Kozlowski GP, Wilson S, Arch JR, BuckinghamRE, Haynes AC, Carr SA, Annan RS, McNulty DE, Liu WS, Terrett JA,Elshourbagy NA, Bergsma DJ, Yanagisawa M 1998 Orexins and orexin re-ceptors: a family of hypothalamic neuropeptides and G protein-coupled re-ceptors that regulate feeding behavior. Cell 92:573–585
3. Hagan JJ, Leslie RA, Patel S, Evans ML, Wattam TA, Holmes S, Benham CD,Taylor SG, Routledge C, Hemmati P, Munton RP, Ashmeade TE, Shah AS,Hatcher JP, Hatcher PD, Jones DN, Smith MI, Piper DC, Hunter AJ, PorterRA, Upton N 1999 Orexin A activates locus coeruleus cell firing and increasesarousal in the rat. Proc Natl Acad Sci USA 96:10911–10916
4. Mondal MS, Nakazato M, Matsukura S 2000 Orexins (hypocretins): novelhypothalamic peptides with divergent functions. Biochem Cell Biol 78:299–305
5. Date Y, Ueta Y, Yamashita H, Yamaguchi H, Matsukura S, Kangawa K,Sakurai T, Yanagisawa M, Nakazato M 1999 Orexins, orexigenic hypotha-lamic peptides, interact with autonomic, neuroendocrine and neuroregulatorysystems. Proc Natl Acad Sci USA 96:748–753
6. Mazzocchi G, Malendowicz LK, Aragona F, Rebuffat P, Gottardo L, Nuss-dorfer GG 2001 Human pheochromocytomas express orexin receptor type 2gene and display an in vitro secretory response to orexins A and B. J ClinEndocrinol Metab 86:4818–4821
7. Pu S, Jain MR, Kalra PS, Kalra SP 1998 Orexins, a novel family of hypotha-lamic neuropeptides, modulate pituitary luteinizing hormone secretion in anovarian steroid-dependent manner. Regul Pept 78:133–136
8. Russell SH, Kim MS, Small CJ, Abbott CR, Morgan DG, Taheri S, Murphy
FIG. 11. Orexin-A and GHRH have a synergistic effect on GH release in vitro. Effects of orexin-A, different concentrations of GHRH, and thecoadministration of orexin-A and GHRH on GH release in cultured ovine somatotropes were observed. The columns show GH release (mean �SEM; n � 4) in response to the individual peptides or the combination of both peptides indicated at the bottom of the figure (also see Results).
4618 Endocrinology, December 2002, 143(12):4609–4619 Xu et al. • Orexin-A and Somatotropes

KG, Todd JF, Ghatei MA, Bloom SR 2000 Central administration of orexin Asuppresses basal and domperidone stimulated plasma prolactin. J Neuroen-docrinol 12:1213–1218
9. Jaszberenyi M, Bujdoso E, Pataki I, Telegdy G 2000 Effects of orexins on thehypothalamic-pituitary-adrenal system. J Neuroendocrinol 12:1174–1178
10. Irahara M, Tamura T, Matuzaki T, Saito S, Yasui T, Yamano S, Kamada M,Aono T 2001 Orexin-A suppresses the pulsatile secretion of luteinizing hor-mone via beta-endorphin. Biochem Biophys Res Commun 281:232–236
11. Mazzocchi G, Malendowicz LK, Gottardo L, Aragona F, Nussdorfer GG 2001Orexin A stimulates cortisol secretion from human adrenocortical cellsthrough activation of the adenylate cyclase-dependent signaling cascade. J ClinEndocrinol Metab 86:778–782
12. Mitsuma T, Hirooka Y, Mori Y, Kayama M, Adachi K, Rhue N, Ping J,Nogimori T 1999 Effects of orexin A on thyrotropin-releasing hormone andthyrotropin secretion in rats. Horm Metab Res 31:606–609
13. Nowak KW, Mackowiak P, Switonska MM, Fabis M, Malendowicz LK 2000Acute orexin effects on insulin secretion in the rat: in vivo and in vitro studies.Life Sci 66:449–454
14. Date Y, Mondal MS, Matsukura S, Ueta Y, Yamashita H, Kaiya H, KangawaK, Nakazato M 2000 Distribution of orexin/hypocretin in the rat medianeminence and pituitary. Brain Res Mol Brain Res 76:1–6
15. Blanco M, Lopez M, GarcIa-Caballero T, Gallego R, Vazquez-Boquete A,Morel G, SenarIs R, Casanueva F, Dieguez C, Beiras A 2001 Cellular local-ization of orexin receptors in human pituitary. J Clin Endocrinol Metab 86:1616–1619
16. Samson WK, Taylor MM 2001 Hypocretin/orexin suppresses corticotrophresponsiveness in vitro. Am J Physiol 281:R1140–R1145
17. Chen C, Vincent JD, Clarke IJ 1994 Ion channels and signal transductionpathways in the regulation of growth hormone secretion. Trends EndocrinolMetab 5:227–233
18. Chen C, Clarke IJ 1995 Modulation of Ca2� influx in the ovine somatotrophby growth hormone-releasing factor. Am J Physiol 268:E204–E212
19. Johren O, Neidert SJ, Kummer M, Dendorfer A, Dominiak P 2001 Prepro-orexin and orexin receptor mRNAs are differentially expressed in peripheraltissues of male and female rats. Endocrinology 142:3324–3331
20. van den Pol AN, Gao XB, Obrietan K, Kilduff TS, Belousov AB 1998 Pre-synaptic and postsynaptic actions and modulation of neuroendocrine neuronsby a new hypothalamic peptide, hypocretin/orexin. J Neurosci 18:7962–7971
21. Grudt TJ, van den Pol AN, Perl ER 2002 Hypocretin-2 (orexin-B) modulationof superficial dorsal horn activity in rat. J Physiol 538:517–525
22. Hwang LL, Chen CT, Dun NJ 2001 Mechanisms of orexin-induced depolar-izations in rat dorsal motor nucleus of vagus neurones in vitro. J Physiol537:511–520
23. Ichinose M, Asai M, Sawada M, Sasaki K, Oomura Y 1998 Induction ofoutward current by orexin-B in mouse peritoneal macrophages. FEBS Lett440:51–54
24. Malendowicz LK, Tortorella C, Nussdorfer GG 1999 Orexins stimulate cor-ticosterone secretion of rat adrenocortical cells, through the activation of theadenylate cyclase-dependent signaling cascade. J Steroid Biochem Mol Biol70:185–188
25. Lund PE, Shariatmadari R, Uustare A, Detheux M, Parmentier M, KukkonenJP, Akerman KE 2000 The orexin OX1 receptor activates a novel Ca2� influxpathway necessary for coupling to phospholipase C. J Biol Chem 275:30806–30812
26. Kukkonen JP, Akerman KE 2001 Orexin receptors couple to Ca2� channelsdifferent from store-operated Ca2� channels. Neuroreport 12:2017–2020
27. Holl RW, Thorner MO, Mandell GL, Sullivan JA, Sinha YN, Leong DA 1988Spontaneous oscillations of intracellular calcium and growth hormone secre-tion. J Biol Chem 263:9682–9685
28. Acs Z, Zsom L, Mergl Z, Makara GB 1993 Significance of chloride channelactivation in the �-aminobutyric acid induced growth hormone secretion in theneonatal rat pituitary. Life Sci 52:1733–1739
29. Naumov AP, Herrington J, Hille B 1994 Actions of growth-hormone-releasinghormone on rat pituitary cells: intracellular calcium and ionic currents.Pflugers Arch 427:414–421
30. Kato M 1996 Growth hormone-releasing hormone augments voltage-gatedNa� current in cultured rat pituitary cells. Am J Physiol 270:C125–C130
31. Lussier BT, French MB, Moor BC, Kraicer J 1991 Free intracellular Ca2�
concentration and growth hormone (GH) release from purified rat soma-totrophs. III. Mechanism of action of GH-releasing factor and somatostatin.Endocrinology 128:592–603
32. Kwiecien R, Tseeb V, Kurchikov A, Kordon C, Hammond C 1997 Growthhormone-releasing hormone triggers pacemaker activity and persistent Ca2�
oscillations in rat somatotrophs. J Physiol 499:613–62333. Chen C, Wu D, Clarke IJ 1996 Signal transduction systems employed by
synthetic GH-releasing peptides in somatotrophs. J Endocrinol 148:381–38634. Chen C, Heyward P, Zhang J, Wu D, Clarke IJ 1994 Voltage-dependent
potassium currents in ovine somatotrophs and their function in growth hor-mone secretion. Neuroendocrinology 59:1–9
35. Marcus JN, Aschkenasi CJ, Lee CE, Chemelli RM, Saper CB, Yanagisawa M,Elmquist JK 2001 Differential expression of orexin receptors 1 and 2 in the ratbrain. J Comp Neurol 435:6–25
36. Farrell RE 1993 Protocol: rapid isolation of RNA with SDS and potassiumacetate reagents. In: Farrell RE, ed. RNA methodologies: a laboratory guide forisolation and characterization. San Diego: Academic Press; 69–70
37. Lopez M, SenarIs R, Gallego R, GarcIa-Caballero T, Lago F, Seoane L,Casanueva F, Dieguez C 1999 Orexin receptors are expressed in the adrenalmedulla of the rat. Endocrinology 140:5991–5994
38. Tsien RW, Fox AP, Hess P, McCleskey EW, Nilius B, Nowycky MC, Rosen-berg RL 1987 Multiple types of calcium channel in excitable cells. Soc GenPhysiol Ser 41:167–187
39. Chen C, Clarke IJ 1995 Effects of growth hormone-releasing peptide-2(GHRP-2) on membrane Ca2� permeability in cultured ovine somatotrophs.J Neuroendocrinol 7:179–186
40. Glassmeier G, Hauber M, Wulfsen I, Weinsberg F, Bauer CK, Schwarz JR2001 Ca2� channels in clonal rat anterior pituitary cells (GH3/B6). PflugersArch 442:577–587
41. Chen C 1998 Gi-3 protein mediates the increase in voltage-gated K� currentsby somatostatin on cultured ovine somatotrophs. Am J Physiol 275:E278–E284
42. Chen C, Zhang J, Dayanithi G, Vincent JD, Israel JM 1989 Cation currentson identified rat gonadotroph cells maintained in primary culture. NeurochemInt 15:265–275
43. Xu R, Roh SG, Loneragan K, Pullar M, Chen C 1999 Human GHRH reducesvoltage-gated K� currents via a non-cAMP-dependent but PKC-mediatedpathway in human GH adenoma cells. J Physiol 520:697–707
44. Takei T, Takano K, Yasufuku-Takano J, Fujita T, Yamashita N 1996 En-hancement of Ca2� currents by GHRH and its relation to PKA and [Ca2�]i inhuman GH-secreting adenoma cells. Am J Physiol 271:E801–E807
45. Takei T, Yasufuku-Takano J, Takano K, Fujita T, Yamashita N 1998 Effect ofCa2� and cAMP on capacitance-measured hormone secretion in human GH-secreting adenoma cells. Am J Physiol 275:E649–E654
46. de Lecea L, Sutcliffe JG 1999 The hypocretins/orexins: novel hypothalamicneuropeptides involved in different physiological systems. Cell Mol Life Sci56:473–480
47. Kwiecien R, Hammond C 1998 Differential management of Ca2� oscillationsby anterior pituitary cells: a comparative overview. Neuroendocrinology 68:135–151
48. Ivanov A, Aston-Jones G 2000 Hypocretin/orexin depolarizes and decreasespotassium conductance in locus coeruleus neurons. Neuroreport 11:1755–1758
49. Adams EF, Lei T, Buchfelder M, Bowers CY, Fahlbusch R 1996 Protein kinaseC-dependent growth hormone releasing peptides stimulate cyclic adenosine3�,5�-monophosphate production by human pituitary somatotropinomas ex-pressing gsp oncogenes: evidence for crosstalk between transduction path-ways. Mol Endocrinol 10:432–438
50. Uramura K, Funahashi H, Muroya S, Shioda S, Takigawa M, Yada T 2001Orexin-a activates phospholipase C- and protein kinase C-mediated Ca2�
signaling in dopamine neurons of the ventral tegmental area. Neuroreport12:1885–1889
51. van den Pol AN 1999 Hypothalamic hypocretin (orexin): robust innervationof the spinal cord. J Neurosci 19:3171–3182
52. Chen C, Wu D, Clarke IJ 1996 Signal transduction systems employed bysynthetic GH-releasing peptides in somatotrophs. J Endocrinol 148:381–386
53. Lei T, Adams EF, Buchfelder M, Fahlbusch R 1996 Relationship betweenprotein kinase C and adenylyl cyclase activity in the regulation of growthhormone secretion by human pituitary somatotrophinomas. Neurosurgery39:569–575
54. Cheng K, Chan WW, Butler B, Barreto Jr A, Smith RG 1991 Evidence for arole of protein kinase-C in His-d-Trp-Ala-Trp-d-Phe-Lys-NH2-inducedgrowth hormone release from rat primary pituitary cells. Endocrinology 129:3337–3342
55. Felder CC, Poulter MO, Wess J 1992 Muscarinic receptor-operated Ca2� influxin transfected fibroblast cells is independent of inositol phosphates and releaseof intracellular Ca2�. Proc Natl Acad Sci USA 89:509–513
56. Zerbes M, Bunn SJ, Powis DA 1998 Histamine causes Ca2� entry via both astore-operated and a store-independent pathway in bovine adrenal chromaffincells. Cell Calcium 23:379–386
57. Rauch M, Riediger T, Schmid HA, Simon E 2000 Orexin A activates leptin-responsive neurons in the arcuate nucleus. Pflugers Arch 440:699–703
58. Ida T, Nakahara K, Kuroiwa T, Fukui K, Nakazato M, Murakami T, Mu-rakami N 2000 Both corticotropin releasing factor and neuropeptide Y areinvolved in the effect of orexin (hypocretin) on the food intake in rats. NeurosciLett 293:119–122
59. Komaki G, Matsumoto Y, Nishikata H, Kawai K, Nozaki T, Takii M, SogawaH, Kubo C 2001 Orexin-A and leptin change inversely in fasting non-obesesubjects. Eur J Endocrinol 144:645–651
Xu et al. • Orexin-A and Somatotropes Endocrinology, December 2002, 143(12):4609–4619 4619





























