Embed Size (px)
Citation preview

Plant Molecular Biology 12:105-116 (1989) © Kluwer Academic Publishers, Dordrecht - Printed in the Netherlands 105
Organization and expression of polygalacturonase and other ripening related genes in Aiisa Craig "Neverripe" and "Ripening inhibitor" tomato mutants
Julie Knapp, l Philippe Moureau, 2,3 Wolfgang Schuch 2 and Don Grierson l* 1Department of Physiology and Environmental Science, University of Nottingham, School of Agriculture, Sutton Bonington, Loughborough LE12 5RD, U.K.; 2ICI Plant Biotechnology Group, P.O. Box 11, The Heath, Runcorn, Cheshire WA 7 4QE, UK.; 3Present address: University of Louvain, Avenue Hippocrate 54, UCL34.90, B-1200 Brussels, Belgium (*author for correspondence)
Received 13 June 1988; accepted in revised form 19 October 1988
Key words: mRNA, tomato ripening mutants, polygalacturonase
Abstract
The organization and expression of ripening-related genes were investigated in normal tomato (Lycopersicon esculentum cv. Ailsa Craig) and in Neverripe (Nr) and Ripening inhibitor (rin) mutants.
Hybridization studies with ripening-related cDNA clones showed that the gene for polygalacturonase (PG) is barely expressed in rin and expressed at a low level in Nr fruit. Four other genes were found to be expressed at reduced levels in rin. Exogenous ethylene was able to restore higher levels of expression of all the genes show- ing reduced expression in rin except that for PG. However, exogenous ethylene did not restore normal ripening in rin fruit. Analysis of chromosomal DNA by Southern blotting indicated that all the genes studied, including the PG gene, and also an upstream promoter of the PG gene, are present in the rin and Nr genomes and appear to be arranged in a similar way to those in normal tomatoes. The results are discussed in the light of the sugges- tion that these mutations may involve part of the regulatory apparatus leading to the expression of ripening
genes such as PG.
Introduction
Ripening of tomato and other climateric fruit is a coordinated process which involves alterations in a number of characteristics including colour, texture, flavour and aroma. These alterations are associated with changes in enzyme activity [29] which have been shown to be related to changes in translatable mRNAs [27, 33, 16, 1]. Ripening-related cDNA libraries have been produced from tomato and clones for a number of ripening-related mRNAs have been identified and characterized [32, 23]. Several clones have been shown to encode polygalac-
turonase (PG), the major cell wall softening enzyme [18, 8, 31]. The PG mRNA is found in ripening fruit [13, 21, 9] but not in immature fruit or leaves or roots [13, 24]. Genomic clones for a single PG gene have recently been isolated and sequenced, and 1449 bp of DNA immediately 5' to the PG gene has been shown to direct ripening-specific expression of the bacterial chloramphenicol acetyl transferase (CAT) gene in transgenic tomato plants [2].
Ethylene plays an important role in ripening of climacteric fruit. It stimulates colour formation, PG synthesis [17] and the production of ripening-related mRNAs including that for PG [13, 24]. However, the

106
regulatory mechanisms governing ethylene synthesis and the stimulation of gene expression in response to ethylene are not understood. These can be studied using ripening mutants which show specific altera- tions in gene expression [1, 14]. Ripening inhibitor (rin) mutant tomatoes mature very slowly, the fruit remaining green when normal fruit have turned red. Eventually, after several months, they develop a lem- on yellow colour but the pericarp remains firm [28] and there is little or no synthesis of PG, lycopene [34] or ethylene [19] during ripening. Neverripe (Nr) fruit [4] produce normal levels of ethylene but only low levels of PG, which leads to reduced softening dur- ing ripening. They contain low levels of lycopene and, even after months of ripening, only achieve an orange colour. Nr and rin fruit have been shown to be deficient in some mRNAs normally associated with ripening [15, 13, 1, 9].
The pleiotropic effects of the rin and Nr muta- tions can be explained in a number of ways. The lack of ethylene production in the rin fruit, for example, could lead to a failure to trigger correctly the ripen- ing response. Alternatively, the rin mutant could ac- tually lack the genes encoding a number of mRNAs normally involved in ripening or, as suggested previ- ously [13, 12], there could be a lack of a regulatory factor which normally acts on these genes. Altera- tions in gene organization or regulatory factors could also explain the Nr phenotype. Hence further investigation into the nature of these mutations may help elucidate the mechanism of ripening in climac- teric fruit. We used ripening-related cDNAs to study the organization and expression of PG and other genes in these mutants. As we previously reported, the PG gene is present but scarcely expressed in the rin mutant [12], even when supplied with exogenous ethylene.
Materials and methods
Plant material
Tomato seeds (Lycopersicon esculentum Mill. cv. Ailsa Craig) were surface-sterilized and grown in virus-free conditions as previously described [16] in a glasshouse with 16 h illumination (22°C)/8 h
dark (17 °C) cycles. Wild-type and mutant flowers from near-isogenic lines of Ailsa Craig homozygous for tin and Nr [6, 7] were hand-pollinated and tagged at anthesis. Fruit were picked at different stages of ripening or at various time points after anthesis and the pericarp was immediately frozen in liquid nitro- gen. rin fruit and controls for ethylene incubation were picked at the first sign of yellow colouration and placed in air in a humidified gas jar with or with- out the addition of 10/A/I ethylene and left in the light at 25 °C. Jars were vented every 48 h and fruit pericarp frozen in liquid nitrogen and stored at - 7 5 °C until required. Leaves were harvested from young axillary shoots and frozen as above.
Preparation of cloned DNA probes
Clones used were from a ripening related cDNA li- brary [32, 24]. Plasmids were prepared [3] and the cDNA inserts were excised from the vector using Pst I (BRL) and electroeluted from 0.80/o agarose gels [22]. cDNA inserts were radioactively labelled in the presence of [32p]dCTP (Amersham, 111 TBq/mmol, 1.85 MBq/50 ~1 reaction) using a BRL nick-translation kit.
Preparation of plant DNA
Chromosomal DNA was prepared [10] from wild- type, Nr and rin leaves.
Southern blot analysis of chromosomal DNA
5 ~g samples of DNA were incubated at 37 °C for 4 h with restriction endonucleases, and the resulting fragments were separated by 0.8% agarose gel elec- trophoresis [22]. The DNA was denatured by sub- merging the gel in 1.5 M NaC1, 0.5 M NaOH for 1 ¼ h. The surfaces of the gel were wiped dry and the DNA was transferred by blotting in 20× SSC (3 M NaC1, 0.3 M sodium citrate) overnight onto Hybond-N membrane (Amersham). The membrane was washed briefly in 2 × SSC, air dried, and the DNA was cross-linked to the membrane by 3.5

minutes irradiation on a UV transilluminator. The membranes were pre-hybridized at 65 °C for 4 h in 5 x SSPE (0.9 M NaC1, 0.05 M sodium phosphate pH 7.2, 5 mM Na 2 EDTA), 0.25% dried skimmed milk and subsequently hybridized overnight at 65 °C in fresh buffer solution containing the radioactively labelled D N A probes. Membranes were washed for 4 × 3 0 m i n u t e s in 2 x SSC, 0.1% SDS and 15 minutes in 0.1 x SSC, 0.1% SDS at 65 °C. Au- toradiography was carried out on either pre-flashed Kodak X-Omat or Fuji RX X-ray film at - 7 5 °C with intensifying screens.
Preparation of RNA
Total RNA was extracted and purified from tomato fruit [32]. Poly(A) + RNA was isolated from this us- ing Hybond Messenger Affinity Paper (Amersham), or oligo-dT-cellulose (Pharmacia) for Northern analysis.
RNA dot hybridizations
1/~g and 0.2 #g of each poly(A) + RNA sample was denatured with glyoxal and applied to the membrane as previously described [32] except that the mem- brane used was Hybond-N (Amersham). For the dot blot analysis of chronologically aged fruit, 10, 5, and 1 t~g of each total RNA sample were prepared in the same way. Insert cDNAs were denatured by incuba- tion for 15 minutes in 0.2 M NaOH, and control dots were applied to the membranes in a similar manner. After baking for 2 hours at 80 °C dot blots were pre- hybridized for 2 - 5 h at 60°C in 5× SSPE, 0.1% ficoll, 0.1% polyvinyl pyrollidone, 0.1% bovine se- rum albumin, 200 #g/ml denatured salmon sperm DNA, 1%0 SDS. After addition of radioactively labelled cDNA probes membranes were left to hy- bridize at 60°C for 16 h. Membranes were washed successively in 10x NTE (0.3 M NaC1, 0.06 M Tris- HC1 p H 8.0, 2 mM EDTA), 0.1% SDS for 15 minutes at room temperature, 10 × NTE, 0.1% SDS for 3 × 30 minutes, at 65 °C, and 1 × NTE, 0.1% SDS for 15 minutes at 65 °C. Autoradiography was carried out as above and autoradiographs were scanned with a
107
Joyce Loebl Chromoscan 3 densitometer and results were quantified [24].
Northern analysis o f mRNA
Poly(A) ÷ RNA samples and RNA size markers (BRL) were denatured using glyoxal as above and electrophoresed in a 1.5% agarose gel in 12.5 mM sodium phosphate, pH 6.5, with continuous recircu- lation of buffer. RNA was blotted in 20 x SSC for 16 h onto Hybond-N membrane which was baked, hybridized, washed and autoradiographed in the same way as dot blots above.
Results
Comparison of genomic DNA in normal and rin plants by hybridization to cDNA clones
To study the organization of the genes homologous to the cDNA probes used in this work, Southern blots of restriction enzyme digests o f normal, Nr and rin genomic D N A were probed with five ripening-related cDNA clones (Fig. 1). The results indicate that these genes are present in the rin and Nr genomes and appear to be arranged in a similar way to the normal genes. Similar results (not shown) were found for pTOM 75 and 137. This confirms that the mutant lines are isogenic for all seven genes inves- tigated. Also included on the pTOM 6 genomic blot were samples of the PG cDNA clone at concentra- tions equivalent to 1.14 and 0.57 copies per 0.75 pg of genomic DNA (the haploid content per tomato cell nucleus, (11)). A densitometer scan of the au- toradiograph gave values of 0.97, 0.99, and 1.08 co- pies of the PG gene per normal, Nr, and rin haploid genomes. This reconstruction experiment suggests that a single copy of the PG gene is present in both the normal and mutant genomes. The PG gene was found to be present in the rin genome (Fig. 3) al though we found virtually no expression of the corresponding m R N A in the rin fruit (Fig. 2).

108
Fig. 1. Southern analysis of tomato genomic DNA hybridized to ripening-related cDNAs. Track a contains lambda DNA restricted with Hind III, tracks b, c and d contain 5 txg normal, Nr and rin DNA respectively, restricted with Eco RI, tracks e, f and g contain normal, Nr and rin DNA restricted with Eco RI and Barn HI, tracks h, i and j contain normal Nr and rin DNA restricted with Bam HI, tracks k, 1 and m contain normal, Nr and rin DNA restricted with Hind III. Tracks n and o contain 12 and 6 pg pTOM 6 insert DNA which represent approximately 1 and 0.5 copies per haploid genome.
Compar i son o f the levels o f r ipening-related
m R N A s in normal, Nr and r in f r u i t by
hybridizat ion to c D N A clones
Using r ipening-related c D N A clones [32] the expres-
s ion dur ing r ipening of seven m R N A s was studied
by hybr id iza t ion to R N A extracted from no rma l and
m u t a n t fruit (Fig. 2). m R N A s homologous to all the
c D N A clones showed a substant ia l increase in
a m o u n t dur ing r ipening. Expression of those
homologous to clones pTOM 5, 6, 13, 36, 99 in-
creased greatly dur ing r ipening but these were not
expressed in immature fruit (except for low-level ex-
pression of pTOM 5 and 13). Two other mRNAs ,
homologous to pTOM 75 and 137, were present in
unr ipe fruit bu t increased several-fold dur ing r ipen-

109
E
E X
E
c 0
oO ¢/)
C~ X
z rr E
100
50
I00~
100
50
0
pTOM 5
a b c d e f g
pTOM 13
a b c d e f g
pTOM
a b c d e f
100
50
pTOM 137
a b c d e f g
100
50
100
100
pTOM 6
a b c d e f g
b
pTOM 36
50
a b c d e f g
pTOM 99
a b c d e f g
100 ~ . Relative maximal expression of
~ , cONAs
so Q
5 6 13 36 75 99 137
ing. In contrast to the results obtained with normal fruit, mRNAs homologous to pTOM 5, 13, 36 and 99 were reduced throughout ripening in the rin fruit. Furthermore, there was no detectable expression of the mRNA homologous to the PG clone (pTOM 6) either in green normal fruit or at any stage of de- velopment in rin fruit. Production of mRNAs homologous to pTOM 75 and 137 was not reduced in rin, in fact the pTOM 137 mRNA was slightly in- creased in the mutant. The mRNAs homologous to the cDNAs tested were found to be present in total RNA from ripening Nr fruit at levels similar to those found in normal fruit, except for pTOM 5 and pTOM 6 which were found at 44o70 and 31°70 of the maxi- mum levels in normal fruit (data not shown).
Ethylene restores expression of some ripening- related mRNAs in rin fruit
rin fruit were picked when they first began to change colour from green to yellow and were incubated in air with or without the addition of ethylene. RNA samples were prepared from poly(A) + RNA ex- tracted from fruit 1, 3, 7 and 9 days after picking and probed with the five ripening-related cDNAs which showed reduced expression in rin fruit ripened on the plant (Fig. 2). The concentration of mRNAs hybridizing to pTOM 5, 13, 36 and 99 increased in response to ethylene (Fig. 3) to levels approximately the same as those found during normal ripening of the wild-type fruit. The amount of mRNA hybridiz- ing to pTOM 6 showed no increase and remained un- detectable by this method throughout the incubation period indicating that expression of the PG gene cannot be restored to normal levels by supplying ethylene to detached rin fruit. No visible synthesis of lycopene was detected in the rin fruit after incuba- tion with ethylene.
Low-level production of PG mRNA in rin fruit
Fig. 2. Relative expression of ripening-related mR NAs at various stages of ripening in normal and rin tomatoes, a, b, c and d repre- sent levels of cDNA hybridization to m R N A from immature green, mature green, orange, and ripe red normal fruit, e, f and g represent levels o f hybridization to m R N A from green, turning yellow and full yellow rin fruit.
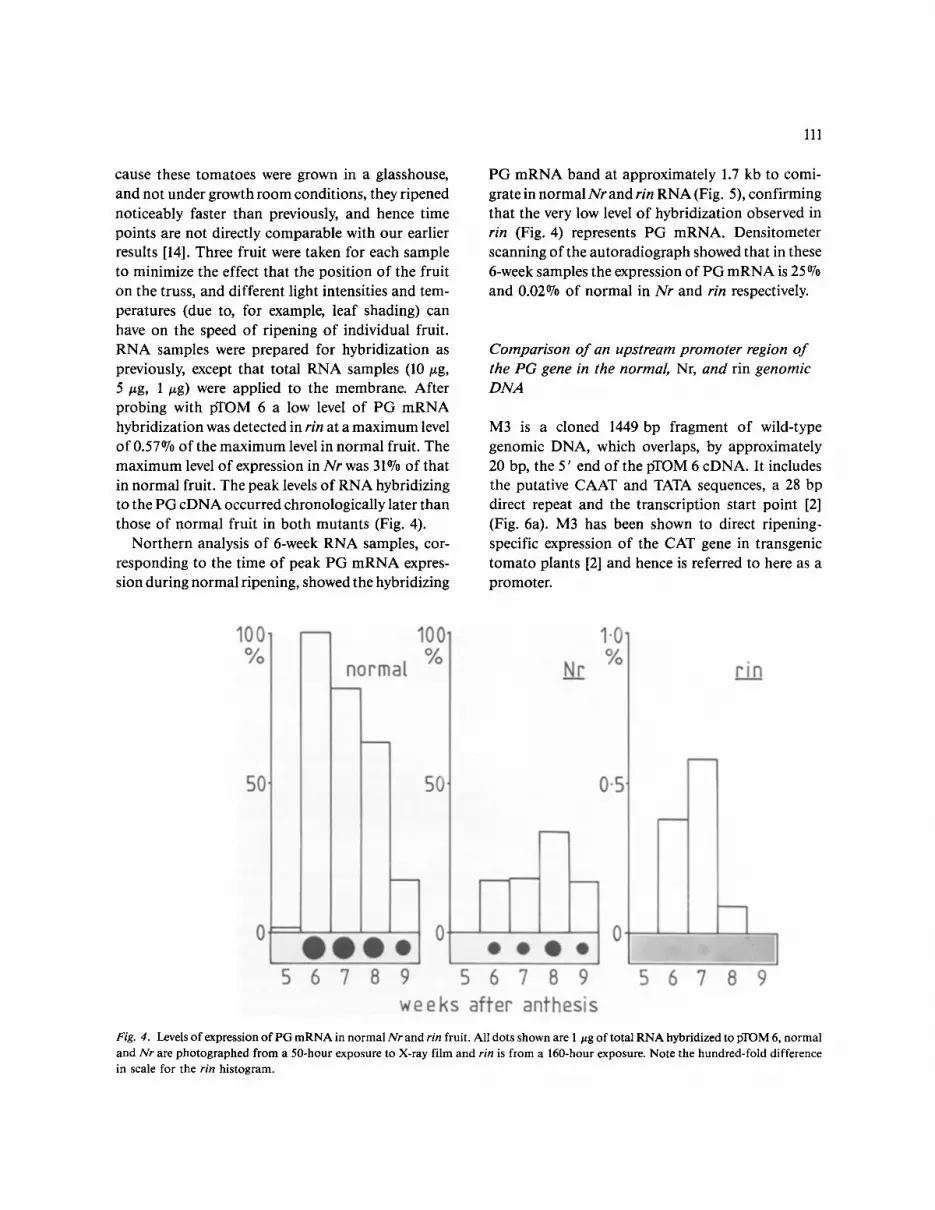
PG has been shown to be produced normally in the abscission zones of rin plants [36]. This led us to in- vestigate whether there is a low level of expression of PG mRNA in rin fruit. Normal, Nr, and rin fruit were picked 5, 6, 7, 8, and 9 weeks after anthesis. Be-

110
Fig. 3. Effect of ethylene on the expression of ripening-related genes in rin fruit. The m R N A dots are applied at 1 #g and 0.2/zg but, there is no 9 day rin + C2H 4 1 #g sample on the filter probed with pTOM 36.
111
cause these tomatoes were grown in a glasshouse, and not under growth room conditions, they ripened noticeably faster than previously, and hence time points are not directly comparable with our earlier results [14]. Three fruit were taken for each sample to minimize the effect that the position of the fruit on the truss, and different light intensities and tem- peratures (due to, for example, leaf shading) can have on the speed of ripening of individual fruit. RNA samples were prepared for hybridization as previously, except that total RNA samples (10 ~g, 5/zg, 1 ~g) were applied to the membrane. After probing with pTOM 6 a low level of PG m R N A hybridization was detected in rin at a max imum level of 0.57°70 of the max imum level in normal fruit. The max imum level of expression in Nr was 31°70 of that in normal fruit. The peak levels o f RNA hybridizing to the PG cDNA occurred chronologically later than those of normal fruit in both mutants (Fig. 4).
Northern analysis of 6-week RNA samples, cor- responding to the time of peak PG m R N A expres- sion during normal ripening, showed the hybridizing
PG m R N A band at approximately 1.7 kb to comi- grate in normal N r and rin RNA (Fig. 5), confirming that the very low level of hybridization observed in rin (Fig. 4) represents PG mRNA. Densitometer scanning of the autoradiograph showed that in these 6-week samples the expression of PG m R N A is 25 o70 and 0.02°70 of normal in N r and rin respectively.
Comparison o f an upstream promoter region o f the P G gene in the normal, Nr, and rin genomic D N A
M3 is a cloned 1449 bp fragment of wild-type genomic DNA, which overlaps, by approximately 20 bp, the 5 ' end of the pTOM 6 cDNA. It includes the putative CAAT and TATA sequences, a 28 bp direct repeat and the transcription start point [2] (Fig. 6a). M3 has been shown to direct ripening- specific expression of the CAT gene in transgenic tomato plants [2] and hence is referred to here as a promoter.
Fig. 4. Levels of expression of PG mRNA in normal Nr and rin fruit. All dots shown are 1 ttg of total RNA hybridized to pTOM 6, normal and Nr are photographed from a 50-hour exposure to X-ray film and rin is from a 160-hour exposure. Note the hundred-fold difference in scale for the rin histogram.

112
Fig. 5. Northern analysis of the PG mRNA expressed in 6-week- old normal Nr and rin fruit. This is a 10-hour exposure of the nor- mal (+) and Nr tracks containing 1 #g and 2.5 #g of normal and Nr poly(A) + mRNA, and a 250-hour exposure of the rin track containing 5 #g of rin poly(A) + mRNA. mRNA samples and RNA size markers (M) were denatured with glyoxal.
Normal, Nr, and rin genomic DNA was incubated with enzymes known to have restriction sites within the nucleic acid sequence of the M3 promoter clone (Fig. 6a) and genomic Southern blots probed with 32p-labelled M3 DNA to see if any differences could be observed in the PG promoters of the mutants. All three DNAs gave the same restriction patterns and the major bands were as predicted from the se- quence. There were also many other bands clearly visible (Fig. 6b). More stringent washing of the filter did not reduce hybridization to any of these bands (results not shown). To investigate whether some bands were due to partial digestion of the DNA, the Southern filter probed with the PG cDNA, on which all the hybridizing fragments were of the size expect- ed from the genomic sequence (Fig. 1), was re- probed with M3 (after removing the pTOM 6 probe by boiling in 0.1070 SDS and leaving to cool). Again the results showed bands of the expected size and many other hybridizing bands (Fig. 6c). The pTOM 6 D N A from the reconstruction experiment did not hybridize to M3 and was not visible. This eliminated the possibility of the extra bands being due to non- specific hybridization of the M3 probe.
Discussion
These results confirm that there are differences in mRNAs expressed in normal and mutant fruit dur- ing ripening [13, 12, 24, 1, 9]. It has been possible to investigate these differences quantitatively by hybridization of RNA from normal and rin fruit to ripening-related cDNA clones. The comparison was made on the basis of developmental stage and also, for the PG mRNA, chronological age. Thus the fully ripe normal fruit were studied approximately 8 weeks after anthesis whereas fully yellow rin fruit were approximately 6 months old. The results show that mRNAs homologous to cDNA clones pTOM 5, 13, 36, 99 are reduced in amount in rin fruit during the limited "ripening" that occurs in this mutant. The levels ranged from approximately 16°70 of nor- mal for pTOM 5 to approximately 74°70 for pTOM 13. mRNAs homologous to cDNA clones pTOM 75 and 137 were not significantly reduced in the mutant. This may be related to the fact that although their expression does increase during ripening of normal tomatoes they are not ripening-specific [13, 24]. There was a low level of accumulation of the mRNAs homologous to pTOM 5 and 13 in unripe fruit which was not detected in previous experiments [24]. The production of pTOM 13 m R N A noticed here may be a wound response due to picking, as expression of this sequence is known to occur very rapidly after wounding as well as during ripening [20]. mRNAs homologous to all these clones were found to be ex- pressed at at least 30°70 of their normal maximum level in N r (results not shown). Initially we found no production of PG m R N A homologous to pTOM 6 [13] in rin fruit. On further investigation we found low levels of expression of the PG gene, approxi- mately 0.5°70 of the level in normal ripening fruit. This is consistent with the traces of PG enzyme ac-
tivity found in rin fruit of the same tomato line [30]. Low levels of expression of PG m R N A and enzyme activity have also been reported for Rutgers rin [9]. In Nr, we found approximately 30°70 of the normal expression of the PG gene in ripening fruit (Fig. 4). We have previously shown that a reduced amount of PG enzyme is present in N r fruit [14] and that this is predominantly the PG1 isoenzyme [35, 5]. This has since been shown in Rutgers N r [9]. The reduc-

113
Fig. 6. Southern analysis to compare the upstream PG promoter in normal Nr and rin genomic DNA using the M3 probe. A. Restriction map of the PG gene derived from the genomic sequence, showing the origin of the M3 probe. B. Tracks a, b and c are 5/~g normal, Nr
and rin DNA restricted with Hae III, tracks d, e and f are restricted with Rsa I, tracks g, h and i are restricted with Taq I. C. Reprobe o f pTOM 6 blot (see Fig. 1). Normal , Nr and rin DNA restricted with Eco RI, Eco RI + Barn HI, Barn HI and Hind III. Tracks a to o are as in the pTOM 6 hybridization in Fig. 1.

114
tion in PG enzyme activity in these mutants cannot be due to a point mutat ion in the gene giving rise to expression of non-functional m R N A or an inactive translation product as such a transcript would have been detected by hybridization to the PG cDNA. Northern analysis showed the major PG m R N A transcript in both the mutant fruit to be the same size (approximately 1.7 kb) as that of normal fruit. This is 200 bp smaller than that reported in Rutgers [9]. Three bands are visible in the normal and Nr
hybridizing samples possibly corresponding to degradation products or other mRNAs with partial homology to the PG cDNA. They do not correspond to the expected sizes of mRNAs arising from the minor transcription start points for the PG gene [2]. The PG m R N A is expressed at too low a level in rin to observe whether these minor bands occur in this
mutant. Our estimates for the relative amounts of PG
m R N A in rin and Nr are very different to those of DellaPenna et al. [9], who found maximum values of 2.5°7o and 22°7o expression of PG m R N A in Rut- gers tomatoes homozygous for rin and Nr respec- tively. This contrasts with our values of 0.5°70 and 30°7o. The differences in PG m R N A expression be- tween normal, Nr and rin fruit respectively are thus three-fold and 200-fold for Ailsa Craig and five-fold and 40-fold for Rutgers. It was also reported [9] that peak expression of PG m R N A occurred at the same time point in both normal and rin fruit whereas, us- ing more time points, we found the peak expression to be delayed by both the Nr and rin mutations. These results suggest that there is a significant differ- ence between the levels and possibly the timing of PG expression when these mutations are incorporat- ed into the Ailsa Craig and Rutgers genomes. How- ever, our fruit were ripened on the plant, but the Rut- gers fruit were picked and ripened off the plant [9]. This may be significant as picking has been shown to affect ripening in the alc tomato ripening mutant [26].
Genes corresponding to the seven cDNA clones tested are all present singly or in a low copy number. No differences were found between the normal Nr and rin genomes. Three genes have now been found for pTOM 13 (M.J. Holdsworth, personal communi- cation), only one genomic sequence has been found
for pTOM 6 [2], and one gene and a pseudogene have been found for pTOM 5 (P. Moureau, personal com- munication). None of the genes studied, including the PG gene, shows any maj or rearrangements or de- letions in the rin and Nr genomes and within the limits of the method the genes appear identical to those of normal fruit, confirming that the lines are isogenic for these genes. DellaPenna et al. [9] showed that the structure of the Nr PG gene was significant- ly different to that in Rutgers tomatoes. This con- trasts with our results for Nr incorporated into an Ailsa Craig background and suggests that the Rut- gers Nr line may not be isogenic.
Incubating rin fruit in 10 td/1 ethylene for up to 9 days failed to stimulate any further significant PG m R N A production, although accumulation of the other four ripening-related mRNAs was greatly stimulated. During this time no visible synthesis of lycopene was detected. Thus rin fruit can respond to ethylene by increased accumulation of some mRNAs, but normal ripening is not restored and PG gene expression remains negligible. Therefore, the lack of ethylene production cannot be responsible for the lack of PG m R N A production in rin fruit. It has been suggested that rin is deficient in an ethy- lene receptor [25]. Since rin fruit can respond to ethylene by accumulating increased quantities of some ripening-related mRNAs, it must be concluded that there are functioning ethylene receptors in rin
fruit. However, since the PG gene is present but bare- ly expressed, it seems likely that part of the biochem- ical apparatus linking ethylene perception with gene expression is impaired in the rin mutant.
The less severe reduction of lycopene production, and expression of the PG gene in Nr fruit suggests that there may be a different control mechanism lacking in this mutant which affects the level of PG expression. As the rin and Nr mutations are known to be on chromosomes 5 and 9 [28, 7] and pTOM 6 has been mapped to chromosome 10 [26] it seems likely the mutations affect transcription of the PG gene indirectly. Thus, there may exist a trans-acting regulator of ripening which, as we previously sug- gested, is not functional in rin [13]. The results of the promoter Southern blots (Fig. 6) indicate that there are a number of sequences with a large degree of ho- mology to the M3 probe, present in the genome of

t h e n o r m a l a n d m u t a n t t o m a t o l ines. T h e P G p r o -
m o t e r is w i t h i n t he M 3 probe . Never the les s , M 3 m a y
c o n t a i n o t h e r n o n - p r o m o t e r e l ements , rin a n d N r
D N A a p p e a r s to c o n t a i n t he s a m e f r a g m e n t s
h y b r i d i z i n g to t he M 3 p r o b e as t h e n o r m a l t o m a t o
D N A , b u t it is still poss ib l e t h a t the re is a m u t a t i o n
w i t h i n t he rin P G p r o m o t e r . H o w e v e r th is w o u l d n o t
be expec t ed s ince the rin m u t a t i o n is k n o w n to m a p
to a d i f f e r en t c h r o m o s o m e to t he P G gene. N o
h y b r i d i z a t i o n was o b s e r v e d b e t w e e n the M3 p r o b e
a n d p T O M 6 c D N A a l t h o u g h they have app rox i -
m a t e l y 20 bp in c o m m o n . T h i s sugges ts t h a t t he
h y b r i d i z i n g b a n d s de t ec t ed by the P G p r o m o t e r f rag-
m e n t are n o t d u e to h o m o l o g y b e t w e e n s h o r t r eg ions
such as C A A T a n d T A T A boxes b u t t h a t the re are
o t h e r P G p r o m o t e r - l i k e s e q u e n c e s w i t h i n t h e t o m a t o
g e n o m e . I t is poss ib l e several genes r e q u i r e d fo r no r -
m a l r i p e n i n g share c o m m o n s e q u e n c e e l e m e n t s w i t h
t he P G p r o m o t e r a n d a f ac to r t h a t r ecogn izes these
s e q u e n c e s does n o t f u n c t i o n p r o p e r l y in rin.
Acknowledgements
J.K. was s u p p o r t e d by a r e sea rch s t u d e n t s h i p f r o m
t h e Sc i ence a n d E n g i n e e r i n g Resea rch C o u n c i l . P.M.
was a r ec ip i en t o f an E E C Tra in ing Fe l lowsh ip in Bi-
o m o l e c u l a r E n g i n e e r i n g .
References
1. Biggs MS, Harriman RW, Handa AK: Changes in gene ex- pression during tomato fruit ripening. Plant Physiol 81: 395-403 (1986).
2. Bird CR, Smith CJS, Ray JA, Moureau P, Bevan MW, Bird AS, Hughes S, Morris PC, Grierson D, Schuch W: The toma- to polygalacturonase gene and ripening-specific expression in transgenic plants. Plant Mol Biol 11:651-662 (1988).
3. Birnboim HC, Doly J: A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucl Acids Res 6: 1513-1523 (1979).
4. Clayberg CD, Butler L, Rick CM, Young PA: Second list of known genes in tomato..1. Hered 51" 167-174 (1960).
5. Crooks PR, Grierson D: Ultrastructure of tomato fruit ripen- ing and the role of polygalacturonase isoenzymes in cell wall degradation. Plant Physiol 72:1088-1093 (1983).
6. Darby LA: Isogenic lines of tomato fruit colour mutants. Hort Res 18:73-84 (1978).
115
7. Darby LA, Ritchie DB, Taylor IB: Isogenic lines of the toma- to 'Ailsa Craig'. In: Annual Report Glasshouse Crops Research Institute, pp. 168-184 (1978).
8. DellaPenna D, Alexander DC, Bennett AB: Proc Natl Acad Sci USA 83:6420-6424 (1986).
9. DellaPenna D, Kates DS, Bennett AB: Polygalacturonase gene expression in Rutgers, rin, nor, and Nr tomato fruits. Plant Physiol 85:502-507 (1987).
10. Dellaporta SL, Wood J, Hick JB: A plant DNA miniprepara- tion: Version II. Plant Mol Biol Rep 1:19-21 (1983).
11. Galbraith DW, Harkins KR, Maddox JM, Ayres NM, Shar- ma DP, Firoozabady E: Rapid flow cytometric analysis of the cell cycle in intact plant tissues. Science 220:1049-1051 (1983).
12. Grierson D, Maunders M J, Holdsworth M J, Ray J, Bird CR, Moureau P, Schuch W, Knapp JE, Tucker GA: Expression and function of ripening genes. In: Nevins D J, Jones RA (eds) Tomato Biotechnology, pp. 309- 323. Alan R Liss, New York (1987).
13. Grierson D, Maunders M J, Slater A, Ray J, Bird CR, Schuch W, Holdsworth M J, Tucker GA, Knapp JE: Gene expression during tomato fruit ripening. Phil Trans Roy Soc Lond B 314: 399- 410 (1986).
14. Grierson D, Purton ME, Knapp JE, Bathgate B: Tomato ripening mutants. In: Thomas H, Grierson D (eds) Develop- mental Mutants in Higher Plants, pp. 73-94. Cambridge University Press, Cambridge (1987).
15. Grierson D, Slater A, Maunders MJ, Crookes PR, Tucker GA, Schuch W, Edwards K: Regulation of the expression of toma- to fruit ripening genes: The involvement of ethylene. In." Roberts JA, Tucker GA (eds) Ethylene and Plant Develop- ment, pp 147-161. Butterworths, London (1985).
16. Grierson D, Slater A, Speirs J, Tucker GA: The appearance of polygalacturonase mRNA in tomatoes: one of a series of changes in gene expression during development and ripening. Planta 163:263-271 (1985).
17. Grierson D, TUcker GA: Timing of ethylene and polygalac- turonase synthesis in relation to the control of tomato fruit ripening. Planta 157:174-179 (1983).
18. Grierson D, Tucker GA, Keen J, Ray J, Bird CR, Schuch W: Sequencing and identification of a cDNA clone for tomato polygalacturonase. Nucl Acids Res 14:8595-8603 (1986).
19. Herner RC, Sink KC: Ethylene production and respiratory behaviour of the rin tomato mutant. Plant Physio152:38 - 42 (1973).
20. Holdsworth M J, Bird CR, Ray J, Schuch W, Grierson D: Structure and expression of an ethylene-related mRNA from tomato. Nucl Acids Res 15:731-739 (1987).
21. Lincoln JE, Cordes S, Read E, Fischer RL: Regulation of gene expression by ethylene during Lycopersicon esculentum (tomato) fruit development. Proc Natl Acad Sci USA 84: 2794-2797 (1986).
22. Maniatis T, Fritsch EF, Sambrook J: Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1983).
23. Mansson PE, Hsu D, Stalker D: Characterization of fruit

116
specific cDNAs from tomato. Mol Gen Genet 200: 356-361 (1985).
24. Maunders M J, Holdsworth M J, Slater A, Knapp JE, Bird CR, Schuch W, Grierson D: Ethylene stimulates the accumu- lation of ripening-related mRNAs in tomatoes. Plant Cell Environ 10" 177-184 (1987).
25. McGlasson WB: Ethylene and fruit ripening. Hort Sci 20" 51-54 (1985).
26. Mutschler M, Guttieri M, Kinzer S, Grierson D, Tucker GA: Changes in ripening-related processes in tomatoes condi- tioned by the alc mutant. Theor Appl Genet 76:285-292 (1988).
27. Rattanapanone N, Speirs J, Grierson D: Evidence for changes in messenger RNA content related to tomato fruit ripening. Phytochemistry 17" 1485-1486 (1978).
28. Robinson RW, Tomes ML: Ripening inhibitor a gene with multiple effects on ripening. Rept Tomato Genet Coop 18:36 (1968).
29. Sacher JA: Senescence and post harvest physiology. Ann Rev Plant Physiol 24:197-224 (1973).
30. Seymour GB, Harding SE, Taylor A J, Hobson GE, Tucker GA: Polyuronide solubilization during ripening of normal and mutant tomato fruit. Phytochem 26: 1981-1875 (1987).
31. Sheehy RE, Pearson J, Brady CJ, Hiatt WR: Molecular characterization of tomato fruit polygalacturonase. Mol Gen Genet 208:30-36 (1987).
32. Slater A, Maunders M J, Edwards K, Schuch W, Grierson D: Isolation and characterisation of cDNA clones for polygalac- turonase and other ripening-related proteins. Plant Mol Biol 5:137-147 (1985).
33. Speirs J, Brady C J, Grierson D, Lee E: Changes in ribosome organisation and messenger RNA abundance in ripening tomato fruits. Aust J Plant Phys 11:225-233 (1984).
34. Tichelaar EC, McGlasson WB, Buescher RW: Genetic regu- lation of tomato fruit ripening. Hort Sci 13: 508- 513 (1978).
35. Tucker GA, Robertson NG, Grierson D: Changes in poly- galacturonase isoenzymes during "ripening" of normal and mutant tomato fruit. Eur J Biochem 112:119-124 (1980).
36. Tucker GA, Schindler CB, Roberts JA: Flower abscission in mutant tomato plants. Planta 160:164-167 (1984).





























