Embed Size (px)
Citation preview

www.elsevier.com/locate/jchemneu
Journal of Chemical Neuroanatomy 32 (2006) 143–158
Organization of brain extracellular matrix in the Chilean fat-tailed
mouse opossum Thylamys elegans (Waterhouse, 1839)
Gert Bruckner a,*, Sanja Pavlica a, Markus Morawski a,Adrian G. Palacios b, Andreas Reichenbach a
a Paul Flechsig Institute for Brain Research, Department of Neurochemistry, University of Leipzig, Jahnalle 59, D-04109 Leipzig, Germanyb Centro de Neurociencia de Valparaıso (CNV), Facultad de Ciencias, Universidad de Valparaıso, Gran Bretana 1111, Playa Ancha, Valparaıso, Chile
Received 13 July 2006; received in revised form 8 August 2006; accepted 15 August 2006
Available online 25 September 2006
Abstract
We investigated the structural and molecular organization of the extracellular matrix in Thylamys elegans, a marsupial representative of the
mammalian order Didelphimorphia. Perineuronal nets (PNs) associated with distinct types of neurons were visualized by detection of chondroitin
sulfate proteoglycans and hyaluronan, and by labeling with Wisteria floribunda agglutinin (WFA), a marker for PNs in the mammalian brain. In the
neocortex of Thylamys, these methods revealed PNs on pyramidal cells. In contrast, parvalbumin-immunoreactive interneurons in the neocortex
and hippocampal formation (displaying robust, WFA-labeled PNs in placental mammals) were ensheathed only with a delicate rim of hyaluronan
and proteoglycans not detectable with WFA. The absence of WFA staining was characteristic also of some subcortical regions which contained PNs
intensely labeled for chondroitin sulfate proteoglycan and hyaluronan. However, corresponding to placental mammals, numerous subcortical
nuclei showed clearly WFA-stained PNs. Similar as in placental mammals, cholinergic basal forebrain neurons and tyrosine hydroxylase-
immunoreactive neurons of the substantia nigra and locus coeruleus were devoid of PNs. Together with our earlier study on Monodelphis, the
present results reveal that South American opossums show either a particular ‘‘marsupial’’ or ‘‘Didelphid’’ type of extracellular matrix
chemoarchitecture, supporting the view that these components may vary phylogenetically as integral parts of neuronal physiology at the systems
and single cell level.
# 2006 Elsevier B.V. All rights reserved.
Keywords: Perineuronal nets; Chondroitin sulfate proteoglycans; Hyaluronan; Cerebral cortex; Brain evolution; Marsupials
1. Introduction
The extracellular matrix specifically contributes to func-
tional characteristics of cells, tissues and organs in develop-
mental and adult stages (for reviews see Comper, 1996). In the
mammalian brain, the spatial and molecular organization of the
extracellular matrix shows a great diversity that obviously
mirrors the complexity of neuronal functions at the single cell
and systemic level (Bruckner et al., 1996; Carlson and
Hockfield, 1996; Matthews et al., 2002). It can be assumed
that brain evolution is accompanied by a corresponding
evolution of the extracellular matrix.
Since perineuronal nets (PNs) have been shown to be a basic
form of extracellular matrix organization in the central nervous
* Corresponding author. Tel.: +49 341 9725 732; fax: +49 341 9725 749.
E-mail address: [email protected] (G. Bruckner).
0891-0618/$ – see front matter # 2006 Elsevier B.V. All rights reserved.
doi:10.1016/j.jchemneu.2006.08.002
system (Brauer et al., 1984; Hendry et al., 1988; Delpech et al.,
1989; Bruckner et al., 1993, 1994, 1996; Murakami et al., 1994;
for reviews see Celio and Blumcke, 1994; Carlson and
Hockfield, 1996; Celio et al., 1998; Yamaguchi, 2000), they
appear as appropriate subjects for studies focused on
evolutionary aspects. In the various vertebrate species
investigated so far, PNs are composed of aggregating
chondroitin sulfate proteoglycans, connected with hyaluronan,
link proteins and tenascin glycoproteins (Yamaguchi, 2000;
Bruckner et al., 2003; Dityatev and Schachner, 2003; Carulli
et al., 2006). These major components may form complexes in
variable proportions resulting in distinct molecular properties,
such as hydration and viscosity, or electrical load of the cellular
microenvironment. Especially, the proteoglycans are assumed
to have a great potential of molecular variation, by formation of
splice variants of their core proteins and by variable
glycosylation by the attached glycosaminoglycan chains
(Matthews et al., 2002; Rauch, 2004). It is conceivable that

G. Bruckner et al. / Journal of Chemical Neuroanatomy 32 (2006) 143–158144
distinct characteristics of the extracellular matrix persist from
ancient to recent animal species in association with neuronal
systems conserved during brain evolution. Correspondingly, the
extracellular matrix may undergo evolutionary alterations
related to functional changes of anatomically defined neuronal
systems. However, the great variability of extracellular matrix
components also inherits the spontaneous formation of multiple
new molecular combinations, which are endowed with
analogous functional properties.
The present study was undertaken to investigate the
association of PNs with defined types of neurons in the South
American mouse opossum (Thylamys elegans), a marsupial
representative of the phylogenetically ancient mammalian
order Didelphimorphia (Steiner et al., 2005). Complementary
to our previous study focussed on the cerebral cortex of the
related species Monodelphis domestica (Bruckner et al., 1998),
we performed a detailed analysis of subcortical regions in the
present investigation. Since, we used cytochemical methods
established in extracellular matrix research, the results are
comparable with data obtained from phylogenetically distant
mammalian, including the human, brain. Our data suggest that
there exist unique marsupial as well as common mammalian
features of extracellular matrix organization.
2. Materials and methods
2.1. Animals and tissue processing
Four adult male mouse opossums (T. elegans) were captured in the wild and
kept in a standard animal facility at the Universidad de Valparaiso (Chile).
Permission to work on collected specimens was under authorization #3014 from
Chilean Servicio Agricola y Ganadero (SAG). Animal care and experimental
procedures complied with international regulations (NIH publications No. 80-
23). For histochemical procedures, animals were euthanized by intraperitoneal
lethal dose of ketamine hydrochloride (1 g/kg body weight) and xylazine
hydrochloride (0.5 g/kg body weight). The brains were removed from the skull
and placed in 4% paraformaldehyde in 0.1 M phosphate buffered saline (PBS),
pH 7.4 for 14 days, cryoprotected in 30% sucrose, frozen and cut into a series of
30 mm thick sections. All sections were collected and extensively rinsed in
0.1 M Tris-buffered saline, pH 7.4 (TBS).
2.2. Anatomical mapping of brain regions and associated
extracellular matrix
Every sixth section of one series of frontal and one series of sagittal sections
were stained with a modified Nissl technique using 0.1% toluidine blue stem
solution diluted 1:200 in 0.1 M acetic acid buffer, pH 4.6, overnight at room
temperature (RT). Alternate sections of these series were used for extracellular
matrix staining with biotinylated Wisteria floribunda agglutinin (Bio-WFA;
Table 1
Cytochemical detection of extracellular matrix components
Detected components Marker Dilution
N-acetylgalactosamine Bio-WFAa 2.5 mg/ml
Chondroitin sulfate
proteoglycan core
protein
Rabbit anti-CSPGb 1:800
Hyaluronan BHABPc 10 mg/ml
a Biotinylated Wisteria floribunda agglutinin, reduced form.b Antigen from chondroitinase ABC-digested bovine nasal cartilage proteoglycac Biotinylated hyaluronic acid binding protein, isolated from bovine nasal cartila
Sigma, Deisenhofen) that was visualized with a standard streptavidin/perox-
idase technique and nickel-enhanced diaminobenzidine as a chromogen. The
treatment of the tissue and the staining followed the previously published
protocol (Hartig et al., 1992, 1994).
Since no cytoarchitectonic brain atlas is available for Thylamys, brain
regions were identified using the stereotaxic atlas of the brain of the opossum
Didelphis marsupialis (Oswaldo-Cruz and Rocha-Miranda, 1968). Structural
details of individual brain regions were identified with the help of studies
describing the anatomy of the cholinergic basal forebrain nuclei (Semba, 2004),
midbrain catecholaminergic regions (Hazlett et al., 1991), red nucleus (King
et al., 1971), cerebellar nuclei (Martin et al., 1974), as well as cortical areas and
hippocampus (Benevento and Ebner, 1971; Hamel, 1967, 1982; Rowe, 1990;
Beck et al., 1996; Huffman et al., 1999; Frost et al., 2000) in American
marsupials.
Most of structural abbreviations used in the present paper were adopted
from the nomenclature applied for mice (Franklin and Paxinos, 1997) and rats
(Paxinos and Watson, 1998).
2.3. Cytochemistry of extracellular matrix components
The reagents used for the fluorescence microscopic analysis of extracellular
matrix components in the present study are specified in Table 1. Tissue sections
showed no labeling when staining procedures were performed without the use
of primary antibodies or WFA.
Antibodies proved to be non-reactive for Thylamys brain were mouse anti-
tenascin-R (Mab 619 and Mab 596; M. Schachner, Hamburg) and rabbit anti-
mouse aggrecan (AB1031; Chemicon, Temecula).
Lectin staining of N-acetylgalactosamine-containing components: Preced-
ing the labeling with Bio-WFA, free-floating sections were treated for 1 h with a
blocking solution consisting of 5% normal goat serum in 0.1 M Tris-buffered
saline with 0.3% Triton X-100 (NGS-TBS-T). The sections were incubated with
Bio-WFA (2.5 mg/ml NGS-TBS-T), rinsed with TBS and processed for 1 h with
a second solution containing streptavidin conjugated to the red fluorescent Cy3
or green fluorescent Cy2 (Dianova, Hamburg; 20 mg/ml TBS containing 2%
bovine serum albumin, BSA).
Immunoreaction for chondroitin sulfate proteoglycan components: Non-
specific binding sites for subsequently applied immunoreagents were blocked
with NGS-TBS-T for 1 h. Free-floating sections were then incubated overnight
at room temperature with solutions containing the primary anti-chondroitin
sulfate proteoglycan (CSPG) antibodies specified in Table 1. The sections were
then rinsed with TBS and processed for 1 h with a solution containing Cy3-goat
anti-rabbit IgG or Cy3-goat anti-mouse IgG (Dianova, Hamburg; 20 mg/ml TBS
containing 2% BSA) as secondary antibodies.
Detection of hyaluronan: The presence and distribution of hyaluronan was
detected by using biotinylated hyaluronic acid-binding protein (BHABP,
1 mg/ml TBS containing 2% BSA) overnight at room temperature. The
labeling was then visualized by Cy3-streptavidin as described for WFA-
staining.
To test the specificity of the BHABP binding, free-floating sections were
pretreated with hyaluronidase from Streptomyces hyalurolyticus (50 U/ml
0.1 M PBS, pH 5.0; Sigma H1136; Koppe et al., 1997) for 4 and 16 h at
37 8C. Hyaluronidase treatment of the tissue sections for 4 h resulted in
decreased staining intensity. Binding of BHABP was at the background level
after 16 h of enzymatic treatment.
Source References
Sigma (Deisenhofen) Hartig et al. (1992, 1994)
Quartett (Berlin) Bertolotto et al. (1986)
Seikagaku America
n.
ge proteoglycan.

G. Bruckner et al. / Journal of Chemical Neuroanatomy 32 (2006) 143–158 145
Table 2
Association of different types of neurons with perineuronal nets revealed by
WFA staining, CSPG immunoreaction and detection of hyaluronan using
BHABP in Thylamys elegans
WFA CSPG BHABP
Cortex, primary sensory areas
Pyramidal cells, layer 5 + + +
Interneurons, layers 2–6a � � �
Subiculum
Pyramidal cells, deep layers + + +
Interneurons, deep layersa �/� ++ +/�
Hippocampus, CA1 region
SO and SP interneuronsa � � �
2.4. Cytochemistry of neuronal markers
Parvalbumin (PARV): Immunoreaction was performed by incubation of
sections after blocking in 5% GNS and 0.3% Triton for 1 h with rabbit anti-
parvalbumin (PV 28; antigen from rat muscle; Swant, Belinzona, dilution
1:250) overnight at RT. After rinsing, sections were processed for 1 h with a
second solution containing Cy2-goat anti-rabbit IgG (Dianova, Hamburg;
20 mg/ml) and streptavidin conjugated to Cy3 or Cy2 (Dianova, Hamburg;
20 mg/ml TBS containing 2% bovine serum albumin, BSA).
Choline acetyltransferase (ChAT): The sections were incubated in blocking
solution containing 5% donkey normal serum in TBS (D-TBS) and 0.3% Triton,
and were then incubated with goat anti-choline acetyltransferase (ChAT)
antibody (Chemicon AB144P; dilution 1:50) in 5% D-TBS and 0.1% Triton,
overnight at room temperature. Thereafter, sections were rinsed in TBS and
incubated with Cy3-donkey anti-goat IgG (20 mg/ml) in TBS supplemented
with 2% BSA for 60 min.
Tyrosine hydroxylase (TH): After blocking in 5% normal goat serum in
TBS containing 0.3% Triton (NGS-TBS-T), the sections were incubated
with rabbit anti-tyrosine hydroxylase (AB152, Chemicon, Temecula; dilu-
tion 1:200). The sections were further processed as described for PARV
immunoreaction.
2.5. Double labeling of neurons and extracellular matrix components
The immunoreactions for the detection of PARV, ChAT and TH were used
for double labeling with markers detecting extracellular matrix components.
Mixtures of primary and secondary reagents were applied according to the
reagents and the subsequent treatment of sections as described for single
labeling.
2.6. Light microscopy, confocal laser scanning microscopy and
image processing
After carbocyanine staining, the sections were extensively washed with
TBS, briefly placed in distilled water, mounted on fluorescence-free slides, air-
dried and coverslipped with Entellan (dissolved in toluene; Merck, Darmstadt).
Tissue sections were examined with a Zeiss Axioplan-AxioVision micro-
scope and a Zeiss confocal laser scanning microscope (LSM 510). Confocal
images of Cy2 fluorescence were obtained with the Argon laser (488 nm) and
emission filter BP 505-530. The HeNe 1 laser (543 nm) and the emission filter
BP 560–615 were used to detect Cy3 fluorescence. Photoshop 5.0 (Adobe
Systems, Mountain View, CA) was used to process the confocal images with
minimal alterations to the contrast and background.
Abbreviations
Subcortical forebrain
aca A nterior commissure M o M olecular layerMedial septum/diagonal band (ChAT�) + + +
Aq A quaeduct (Sylvius) M o5 M otor trigeminal nuGlobus pallidus, external parta + + +
Au P rimary auditory cortex M S M edial septal nuReticular thalamic nua � + +/++
BMAB asomedial amygdala, anterior n u N ucleusZona incerta + + +
BLP B asolateral amygdala, posterior O B O lfactory bulbCpu C
audate putamen o MolO uter molecular layer Brainstem CA1 C A1 field, hippocampus o pt O ptic tractSubstantia nigra, reticular part (TH�)a + + +
CA3 C A3 field, hippocampus p c P osterior commissure Red nu ++ ++ ++ CbC C erebellar cortex P e P eriventricular hypothalamic area Medial nu trapezoid bodya � +/++ ++ CbN C erebellar nu P ir P iriform cortex Gigantocellular reticular nu ++ ++ ++ Cg C ingulate cortex P o P olymorph layer Motor trigeminal nu �/� + ++ Cl C laustrum R MCR ed nu, magnocellular partCerebellar cortex
cp C erebral peduncle R S R etrosplenial cortexGolgi neurons + � �
DG D entate gyrus R t R eticular thalamic nuBasket neuronsa � + +
DLG D orsolateral geniculate nu S S ubiculumEnt E
ntorhinal cortex S 1 P rimary somatosensory cortex Deep cerebellar nuclei � ++ ++ Fr F rontal cortex S C S uperior colliculusa Clearly immunoreactive for parvalbumin in WFA or BHABP double labeling
GP G lobus pallidus S N S ubstantia nigraGr G
ranule cell layer S O S tratum oriens experiments. Symbols indicate staining intensity of perineuronal nets:�/� = nearH H
ippocampus S P S tratum pyramidale background level, � = very low, + = low, +/++ = moderate, ++ = high; � = no�
hc H ippocampal commissure T h T halamus staining detectable; ChAT = choline acetyltransferase-immunonegative neu-�
HDB N u horizontal limb diagonal bandT u O lfactory tubercleic I
rons;
nternal capsule T
TH = tyrosine hydroxylase-immu
z N
noneg
u trapezoid body
Icj I
sland of Caleja V 1 P rimary visual cortexiMol I
nner molecular layer V DBN u vertical limb diagonal bandlo L
ateral olfactory tract V LG V entrolateral geniculate nuLC L
ocus coeruleus V TA V entral tegmental areaLV L
ateral ventricle Z I Z ona incertaMed M
edial cerebellar nu 3 O culomotor nuMG M
edial geniculate nu 4 V F ourth ventricle7 F
acial nu3V T
hird ventricle3. Results
The results presented in this study were selected with respect
to anatomical regions (Table 2) that express clear patterns of
extracellular matrix organization in M. domestica, the only
marsupial investigated so far (Bruckner et al., 1998), as well as
regional patterns known from the rat (Seeger et al., 1994) and
murine brain (Bruckner et al., 2000, 2003). The location of the
analyzed regions in the Thylamys brain is indicated in selected
Nissl-stained sections in Fig. 1. Since our study is focussed on
PNs, the neuropil zones are described only if particular region-
specific properties were apparent.
ative neurons.

G. Bruckner et al. / Journal of Chemical Neuroanatomy 32 (2006) 143–158146
Fig. 1. Structural features of the brain of Thylamys elegans shown in Nissl-stained sections. (A) Parasagittal plane. (B–F) Frontal sections selected approximately at the
rostro-caudal position indicated in (A). For explanation of abbreviations see list of abbreviations. Scale bars: 1 mm in (A); 1 mm in (B) and (F), also applies to (C)–(E).
3.1. Phenotypes of perineuronal nets
WFA-staining of PNs revealed a great divergence in the
staining intensity between different regions, and even within
individual nuclei. Three major structural phenotypes may be
defined (Fig. 2). In the cerebral cortex, WFA-labeled PNs were
associated exclusively with pyramidal cells and showed a very
faint morphology; this will be called the ‘‘pyramidal type’’ of
PNs (Fig. 2A). By contrast, the well-known robust PN
phenotype expressing a clearly contoured, lattice-like structure
was widely distributed in subcortical regions (Fig. 2B). The
soma and the proximal parts of dendrites as well as the axon
initial segment of the subcortical neurons were covered by
matrix aggregates. The third PN type is characterized by a
diffuse perineuronal accumulation of WFA-stained extracel-
lular matrix components (Fig. 2C). Such ‘‘diffuse’’ PNs

G. Bruckner et al. / Journal of Chemical Neuroanatomy 32 (2006) 143–158 147
Fig. 1. (Continued ).

G. Bruckner et al. / Journal of Chemical Neuroanatomy 32 (2006) 143–158148
Fig. 2. Different structural phenotypes of perineuronal nets revealed by WFA-DABNi staining in a frontal section located close to section C in Fig. 1. (A) Pyramidal cell
type frequently seen in layer 5 of primary sensory areas and the retrosplenial cortex. WFA-binding extracellular matrix faintly covers the soma, proximal parts of apical
dendrites and the axon initial segments of pyramidal cells in the S1 cortex. (B) Robust lattice-like type of PNs, predominating in many subcortical regions. The image
shows perineuronal nets in the magnocellular preoptic nucleus of the basal forebrain. (C) Diffuse type of PNs, characteristic of the periventricular hypothalamic area.
Extracellular matrix components infiltrate the adjacent pericellular neuropil. Arrows in (A) and (B) indicate the axon initial segments. Scale bar: 20 mm (applies to all).
(Wegner et al., 2003) were frequently found in the periven-
tricular area of the hypothalamus.
In addition to the variable structural appearance, a great
heterogeneity in the molecular composition of PNs is indicated
by combined application of WFA staining, CSPG immunor-
eaction and detection of hyaluronan by BHABP. Perineuronal
matrix components were labeled with BHABP and CSPG
antibodies in the cerebral cortex and many subcortical nuclei.
However, PNs detected with these stainings were devoid of
WFA labeling in several other regions (Table 2), thereby
contrasting with the patterns known from other mammalian
species.
3.2. Cerebral cortex and hippocampal formation
In the neocortex, clearly stained, robust PNs were not
detected. WFA-staining revealed PNs associated with layer 5
pyramidal cells in primary sensory areas (Fig. 3A and A0) and
with layer 2/3 pyramidal cells in the perirhinal cortex located
dorsal to the rhinal fissure (not shown). The pyramidal PNs
were also labeled with the CSPG (Fig. 3B0) immunoreaction.
Non-pyramidal neurons frequently showed a CSPG-immunor-
eactive perisomatic rim (Fig. 3B0). BHABP staining usually
revealed the perisomatic rim around layer 5 pyramidal cells and
some PARV-positive interneurons, however, its intensity barely
exceeded that of the surrounding neuropil (Fig. 3C and C0).WFA-labeled PNs associated with PARV-negative neurons
were found in the claustrum (not shown).
The hippocampus was characterized by intensely CSPG-
immunoreactive neuropil in the CA1–3 regions (Fig. 4A)
indicating a sharp border to the retrosplenial cortex. In the
subiculum, interneurons were associated with PNs clearly
positive for CSPG (Fig. 3A and A0). WFA-stained pyramidal
PNs were located in a particular cell layer extending from the
subiculum into the CA1 region (Fig. 3A and A0). PARV-positive
interneurons were negative for WFA staining in the hippo-
campus and dentate gyrus, such as found in the neocortex.
However, BHABP-staining (Fig. 4B, B0 and B00) revealed
perisomatic rims around PARV-positive neurons in the dentate
gyrus (Fig. 4B00).
3.3. Subcortical forebrain
The medial septum-diagonal band nuclei, well differentiated
in marsupials (Semba, 2004), contained densely packed PNs
embedded in a diffusely stained neuropil (Fig. 5A). ChAT-
positive neurons in the medial septum-diagonal band complex,
ventral pallidum and substantia innominata, as well as striatal
interneurons were not associated with PNs. However, clearly
contoured, intensely WFA-stained PNs were distributed in the
ventral striatopallidal region of the forebrain (not shown). In the
globus pallidus, the net-associated neurons expressed PARV
immunoreactivity (not shown). The distribution pattern of
WFA-stained PNs was largely corresponding with the patterns
detected by CSPG immunoreaction and BHABP staining in the
basal forebrain. As in other subcortical regions, BHABP
indicated an ubiqitous occurrence of hyaluronan (Fig. 5C), as
well as clearly contoured PNs (Fig. 5C0). In the hypothalamus
diffuse WFA-stained PNs occurred in the close proximity to the
third ventricle (Fig. 5B).
Special features of extracellular matrix organization were
detected in the ventral thalamus. In contrast to the pattern
known from placental mammals, the reticular thalamic nucleus
failed to display WFA staining (Fig. 6A, A00 and B) and was
only weakly labeled by the CSPG immunoreaction (Fig. 6B).
The PARV-positive neurons of this nucleus were ensheathed by

G. Bruckner et al. / Journal of Chemical Neuroanatomy 32 (2006) 143–158 149
Fig. 3. Characteristics of extracellular matrix in the neocortex. Parvalbumin (PARV) expressing interneurons are labeled by immunofluorescence in the S1
somatosensory cortex. For correlation of anatomical regions see Fig. 1A–C. (A) WFA-staining reveals faint PNs only around layer five pyramidal cells (L5). (A0)Subcellular domains of WFA-stained pyramidal PNs include proximal parts of apical dendrites, somata and axon initial segments (arrows). (B and B0) Weak CSPG
immunoreactivity reveals PNs associated with an interneuron (arrow head) devoid of WFA staining whereas pyramidal PNs are double labeled. (C) Detection of
hyaluronan with BHABP shows uniform distribuion pattern of neuropil staining at low magnification. (C0) Staining intensity of BHABP-labeled perisomatic rims
around layer 5 pyramidal cells (asterisks) is similar to neuropil staining. Scale bars: 100 mm in (A) and (C); 20 mm in (A0), (B), (B0) and (C0).
a faint rim of CSPG immunoreactivity and BHABPstaining
(Fig. 6B and C). However, PNs in the zona incerta (Fig. 6A, A0
and B) and the ventral lateral geniculate nucleus were clearly
stained by WFA.
In the dorsal thalamus the primary relay nuclei were
virtually devoid of clearly labeled PNs (not shown).
3.4. Brainstem
The basic distribution patterns of PNs in the brainstem
largely corresponded with the patterns known from placental
mammals. Typically, the red nucleus was the subcortical region
most prominently stained by WFA, CSPG immunoreaction and
BHABP (Fig. 7A, B, B0 and C). In contrast, tyrosine
hydroxylase-immunoreactive neurons in the substantia nigra,
ventral tegmental area and locus coeruleus were devoid of PNs
(Fig. 7A, A0, C, C0 and D).
As an example of motor nuclei, Fig. 8 shows the motor
trigeminal nucleus (Mo5). Motor neurons immunoreactive for
choline acetyltransferase were ensheathed by PNs immunor-
eactive for CSPG, whereas WFA binding was not detectable. In
contrast to CSPG, the intense BHABP staining of PNs was a
G. Bruckner et al. / Journal of Chemical Neuroanatomy 32 (2006) 143–158150
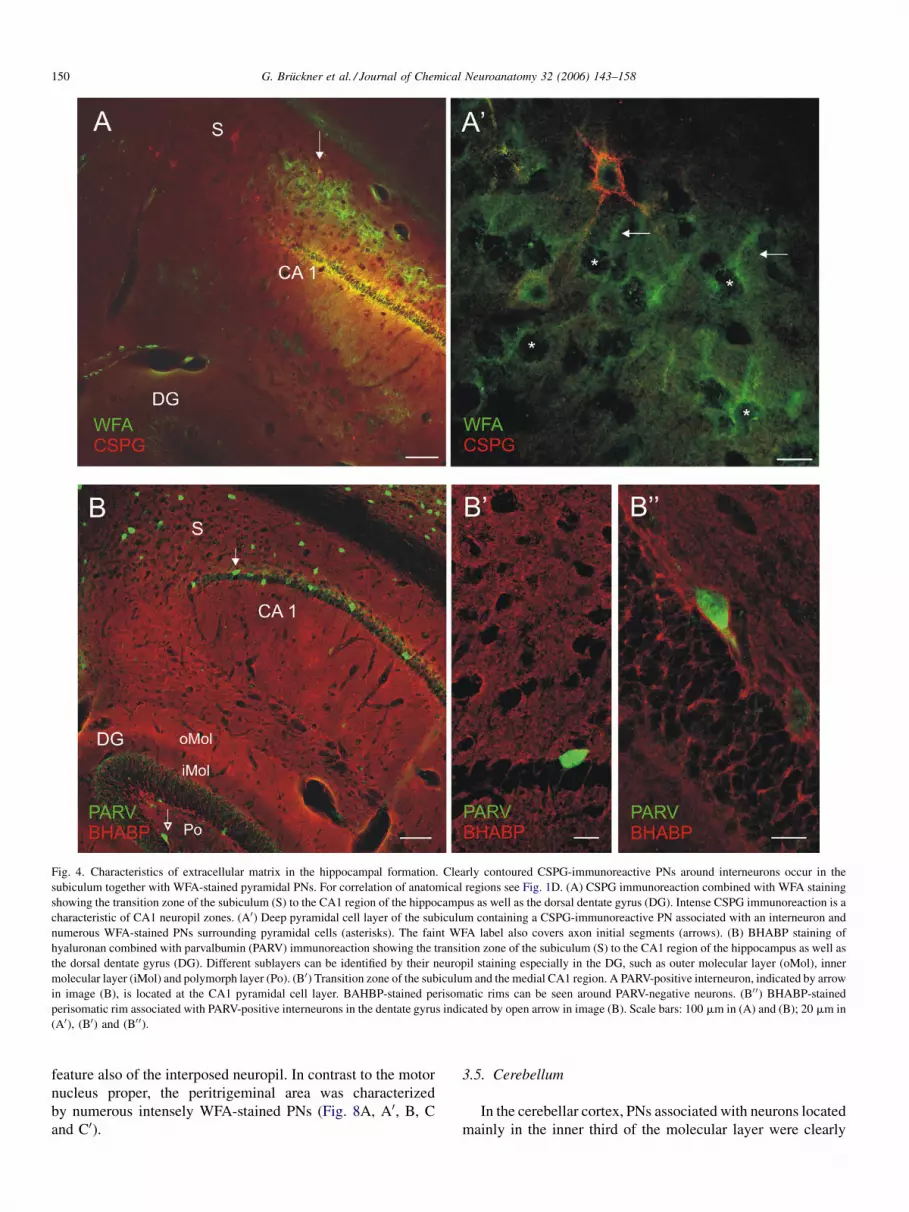
Fig. 4. Characteristics of extracellular matrix in the hippocampal formation. Clearly contoured CSPG-immunoreactive PNs around interneurons occur in the
subiculum together with WFA-stained pyramidal PNs. For correlation of anatomical regions see Fig. 1D. (A) CSPG immunoreaction combined with WFA staining
showing the transition zone of the subiculum (S) to the CA1 region of the hippocampus as well as the dorsal dentate gyrus (DG). Intense CSPG immunoreaction is a
characteristic of CA1 neuropil zones. (A0) Deep pyramidal cell layer of the subiculum containing a CSPG-immunoreactive PN associated with an interneuron and
numerous WFA-stained PNs surrounding pyramidal cells (asterisks). The faint WFA label also covers axon initial segments (arrows). (B) BHABP staining of
hyaluronan combined with parvalbumin (PARV) immunoreaction showing the transition zone of the subiculum (S) to the CA1 region of the hippocampus as well as
the dorsal dentate gyrus (DG). Different sublayers can be identified by their neuropil staining especially in the DG, such as outer molecular layer (oMol), inner
molecular layer (iMol) and polymorph layer (Po). (B0) Transition zone of the subiculum and the medial CA1 region. A PARV-positive interneuron, indicated by arrow
in image (B), is located at the CA1 pyramidal cell layer. BAHBP-stained perisomatic rims can be seen around PARV-negative neurons. (B0 0) BHABP-stained
perisomatic rim associated with PARV-positive interneurons in the dentate gyrus indicated by open arrow in image (B). Scale bars: 100 mm in (A) and (B); 20 mm in
(A0), (B0) and (B0 0).
feature also of the interposed neuropil. In contrast to the motor
nucleus proper, the peritrigeminal area was characterized
by numerous intensely WFA-stained PNs (Fig. 8A, A0, B, C
and C0).
3.5. Cerebellum
In the cerebellar cortex, PNs associated with neurons located
mainly in the inner third of the molecular layer were clearly

G. Bruckner et al. / Journal of Chemical Neuroanatomy 32 (2006) 143–158 151
Fig. 5. Characteristics of extracellular matrix in the basal forebrain. Cholinergic neurons detected by ChAT immunoreaction are devoid of PNs. For correlation of
anatomical regions see Fig. 1B and C. (A) The medial septum (MS)-diagonal band (DB) region is characterized by neighboured clusters of WFA-stained PNs and
ChAT-immunoreactive neurons. (A0) Higher magnification of the DB region. (B) Periventricular hypothalamic region (Pe) characterized by diffuse PNs and
interposed WFA-stained neuropil. ChAT-immunoreactive neurons are devoid of PNs. (C) Distribution pattern of hyaluronan revealed by BHABP staining. The
external part of the globus pallidus (GP) shows moderate staining intensity related to numerous PNs. ChAT-immunoreactive neurons in the ventral striatopallidal
region (VP) are intermingled with PN-associated neurons. (C0) BHABP-stained PNs in the ventral pallidum are associated with non-cholinergic neurons. Scale bars:
100 mm in (A)–(C); 20 mm in (A0) and (C0).

G. Bruckner et al. / Journal of Chemical Neuroanatomy 32 (2006) 143–158152
Fig. 6. Characteristics of extracellular matrix in the ventral thalamus. Ventral to the zona incerta (ZI) the parvalbumin immunoreactive neurons delineate the oval
reticular thalamic nucleus (Rt) located in close proximity to cerebral peduncle (cp). For correlation of anatomical regions see Fig. 1D. (A and A0) WFA-stained PNs
occur only in the ZI. (B) CSPG immunoreaction combined with WFA staining reveals double-labeling and molecular heterogeneity of the ZI. Rt is CSPG-
immunoreactive with low intensity. (C) Moderate BHABP staining of PNs is shown in ZI and Rt. Scale bars: 100 mm in (A)–(C); 20 mm in (A0).
labeled by CSPG immunoreaction (Fig. 9A). The position of
these neurons is suggestive of deep stellate cells or basket cells
(Palay and Chan-Palay, 1974). BHABP staining revealed a
perineuronal rim around PARV-immunoreactive neurons in the
inner molecular layer (Fig. 9B). PNs were not detectable with
WFA in the molecular layer and around Purkinje cells. Golgi
neurons in the granule cell layer were ensheathed by extremely
faintly WFA-binding PNs virtually not detectable by CSPG
immunoreaction (not shown). PNs of Golgi neurons could not
be differentiated from adjacent BHABP-stained matrix
components.
PNs in the cerebellar nuclei (lateral, interposed, medial)
were clearly labeled by CSPG immunoreaction (Fig. 9C and C0)and BHABP (Fig. 9D and D0) but were not stained by WFA. The
net-associated neurons were densely covered by PARV-positive
synaptic profiles (Fig. 9D0).
4. Discussion
The spatial and molecular organization of extracellular
matrix components was revealed in the present study by lectin
(WFA) staining, CSPG immunoreaction and detection of

G. Bruckner et al. / Journal of Chemical Neuroanatomy 32 (2006) 143–158 153
Fig. 7. Characteristics of extracellular matrix in the substantia nigra and locus coeruleus. Dopaminergic and noradrenergic neurons detected by tyrosine hydroxylase
immunoreaction are devoid of PNs. For correlation of anatomical regions see Fig. 1A (sagittal plane) and Fig. 1F. (A) Overview showing the red nucleus (RMC)
associated with intensely WFA-stained PNs, the substantia nigra (SN) and the ventral tegmental area (VTA) crossed by the oculomotor nerve (3n). (A0) Lateral part of
the reticular substantia nigra. Clearly contoured PNs ensheath the soma, proximal parts of dendrites and the axon initial segment (arrows) around tyrosine hydroxylase
immunonegative neurons. (B, B0 and B0 0) CSPG immunoreaction combined with WFA staining reveal a large degree of double labeling of PNs in the midbrain.
Molecular heterogeneity of PNs is indicated by different proportions of red and green fluorescence. (C) Detection of hyaluronan by BHABP indicates the red nucleus
as a prevalently stained midbrain region corresponding with the distribution patterns of PNs. (C0) In the substantia nigra PNs appear as BHABP-stained perisomatic
rims around tyrosine hydroxylase-immunonegative neurons. (D) Tyrosine hydroxylase immunoreaction showing the locus coeruleus (LC) and the subcoeruleus area
in the close proximity to brainstem neurons associated with intensely WFA-stained PNs in a sagittal section. Caudal is to the right. Scale bars: 100 mm in (A), (B),
(B0), (B0 0), (C), (D); 20 mm in (A0), (C0).

G. Bruckner et al. / Journal of Chemical Neuroanatomy 32 (2006) 143–158154
Fig. 8. Characteristics of extracellular matrix in the motor trigeminal nucleus (Mo5). Cholinergic neurons were labeled by ChAT immunoreaction. The nucleus
is characterized by CSPG-immunoreactive and BHABP-stained perisomatic rims and neuropil zones. Clearly contoured WFA-stained PNs only occur in the
peritrigeminal region. For correlation of anatomical regions see Fig. 1G. (A and A0) WFA-staining reveals the motor neurons devoid of label. (B and B0) CSPG-

G. Bruckner et al. / Journal of Chemical Neuroanatomy 32 (2006) 143–158 155
hyaluronan by affinity binding of BHABP in the brain of the
South American mouse opossum T. elegans. This indicated the
existence of important specific characteristics, in parallel with
features shared with many placental mammalian species. Thus,
two basic trends were identified: (i) there are clear changes
during the evolution of cortical and subcortical neuronal
systems, as revealed by the quantity of perineuronal deposition
and by the molecular properties of extracellular matrix
components and (ii) however, distinct architectural patterns
of extracellular matrix organization at the systemic and single
cell level, as well as the structural characteristics of subcellular
PN domains, appear to be conserved during mammalian
phylogeny for millions of years.
4.1. Modified versus conserved properties of perineuronal
nets
On the one hand, there emerges the picture of several
‘‘marsupial features’’ of the extracellular matrix. One feature
clearly differentiating the extracellular matrix in the brain of
South American marsupials (Thylamys, present study, Mono-
delphis, Bruckner et al., 1998) from that of placental mammals
represents a quantitative phenomenon, indicated especially by
the absence of robust PNs around parvalbumin-immunoreactive
interneurons in the neocortex and hippocampus. These PNs
only weakly accumulate CSPGs and hyaluronan, and are
virtually not detectable with WFA known to label PNs of
placental mammalian cortical interneurons with high intensity
(Hartig et al., 1992). A similar phenomenon is apparent also in
the reticular thalamic nucleus characterized by strong
accumulation of CSPGs in placental mammalian species
(Bruckner et al., 1996).
As another hallmark of the extracellular matrix in Thylamys,
clearly CSPG-immunoreactive PNs are devoid of WFA-
staining in a number of subcortical regions that are
characterized by strong WFA-binding in placental mammals,
e.g. the cerebellar nuclei (Seeger et al., 1994; Bruckner et al.,
2000). This observation confirms the view that extracellular
matrix organization in the mammalian brain can be varied not
only quantitatively but also qualitatively (Matthews et al.,
2002). There is more than ‘‘presence/lack’’ or ‘‘dominant/
faint’’; rather, the molecular composition of CSPGs is
differentially regulated, and appears to play a major role in
phylogenetic and/or functional adaptation.
On the other hand, features indicating conserved patterns of
extracellular matrix organization are expressed in different
subcortical regions in Thylamys, e.g. the red nucleus, sharing
major structural and molecular properties with regions mapped
in the rat and murine brain (Seeger et al., 1994; Bruckner et al.,
2000). Likewise, the absence of PNs around certain types of
neurons, such as cholinergic neurons in the basal forebrain
(Brauer et al., 1993; Adams et al., 2001) and aminergic neurons
immunoreactive PNs appear as perisomatic rims around ChAT-positive neurons.
double labeling. PNs in the peritrigeminal regions show molecular diversity in
Detection of hyaluronan by BHABP. The motor nucleus is intensely labeled by sta
20 mm in (A0)–(D0).
of the substantia nigra and the locus coeruleus (Hobohm et al.,
1998) also appears to be a principle conserved during
mammalian evolution.
4.2. Functional significance of phylogenetic differences in
the extracellular matrix
Although the functional role of PNs has not yet been
elucidated, more than a single function may be considered.
Their activity-dependent formation during postnatal devel-
opment (Guimaraes et al., 1990), as well as the reactivation of
ocular dominance plasticity (Pizzorusso et al., 2002) and the
induction of axonal sprouting (Corvetti and Rossi, 2005) after
enzymatic decomposition of extracellular matrix CSPGs
indicate an involvement in the regulation of PNs in neuronal
plasticity (Hockfield, 1993; Rhodes and Fawcett, 2004). The
polyanionic character of many PNs is suggestive of a role in
regulation of the ionic micromilieu in analogy to the
perinodal gap substance at the nodes of Ranvier (Bruckner
et al., 1993, 2006; Hartig et al., 1999; Horn et al., 2003).
Macromolecular matrix complexes may significantly modify
the hydration and the diffusion parameters of the extracelluar
space (Sykova, 1997). Finally, supplementing rather than
replacing other roles, a neuroprotective effect of perineuronal
matrix components is reasonable to assume (Bruckner et al.,
1999; Morawski et al., 2004).
Considering the special pattern of extracellular matrix
organization in South American marsupials, notably the
absence of robust PNs around interneurons in the cerebral
cortex, it may be concluded that the discussed functions of PNs
are not relevant in the cerebral cortex of these species.
However, since numerous PNs occurred in association with
layer 5 pyramidal cells in an area-specific distribution in the
two opossum species investigated so far, the extracellular
matrix appears to mirror a special type of cortical structure and
function in marsupials, contrasting with that of placental
mammals (Bruckner et al., 1998). To our knowledge, studies
investigating electrophysiological characteristics of the par-
valbumin-expressing interneurons have not been performed in
South American opossums. It would be interesting to know
whether these neurons belong to the fast-spiking type of
cortical interneurons, which is predominantly associated with
PNs in placental mammals (Kawaguchi et al., 1987; Kosaka
and Heizmann, 1989; Naegele and Barnstable, 1989; Hartig
et al., 1992, 1994; Celio, 1993). Regarding the possible
involvement of PNs in the regulation of synaptic plasticity,
especially the critical period-plasticity of local cortical circuits
(Hensch, 2005), the cortex of opossums may be used as an
alternative model. Studies in Monodelphis showed that massive
cross-modal cortical plasticity occurs after enucleation at early
developmental stages (Kahn and Krubitzer, 2002), whereas
such plasticity was previously demonstrated in macaque
Neuropil zones are stained with moderate intensity. (C and C0) WFA/CSPG
dicated by different proportions of red and green fluorescence. (D and D0)ining of perisomatic rims and neuropil zones. Scale bars: 100 mm in (A)–(D);

G. Bruckner et al. / Journal of Chemical Neuroanatomy 32 (2006) 143–158156
Fig. 9. Characteristics of extracellular matrix in the cerebellar cortex and deep cerebellar nuclei. Parvalbumin immunoreaction (PARV) indicates clearly labeled
Purkinje cells inclusive axonal projections in sagittal sections (for comparison see Fig. 1A). (A) CSPG/WFA double staining reveals CSPG-immunoreactive PNs in
the lower molecular layer (m) devoid of WFA staining. Asterisks indicate the Purkinje cell layer. (B) Detection of hyaluronan by BHABP shows PNs as perisomatic

G. Bruckner et al. / Journal of Chemical Neuroanatomy 32 (2006) 143–158 157
monkeys (Dehay et al., 1996) and rodents (Olavarria and Li,
1995) to occur at later stages of brain development.
Acknowledgements
The authors wish to thank Mrs. Margit Schmidt for excellent
technical assistance, her last contribution to our research after
three decades of intelligent, accurate and patient professional
work. This work was supported by the Deutsche Forschungs-
gemeinschaft, grant number BR 1208/3-3 and the Interdiszi-
plinares Zentrum fur Klinische Forschung (IZKF) in the course
of the MD/PhD program at the University of Leipzig.
References
Adams, I., Brauer, K., Arelin, C., Hartig, W., Fine, A., Mader, M., Arendt, T.,
Bruckner, G., 2001. Perineuronal nets in the rhesus monkey and human
basal forebrain including basal ganglia. Neuroscience 108, 285–298.
Beck, P.D., Pospichal, M.W., Kaas, J.H., 1996. Topography, architecture, and
connections of somatosensory cortex in opossums: evidence for five
somatosensory areas. J. Comp. Neurol. 366, 109–133.
Benevento, L.A., Ebner, F.F., 1971. The areas and layers of corticocortical
terminations in the visual cortex of the Virginia opossum. J. Comp. Neurol.
141, 157–190.
Bertolotto, A., Palmucci, L., Gagliano, A., Mongini, T., Tarone, G., 1986.
Immunohistochemical localization of chondroitin sulfate in normal and
pathological human muscle. J. Neurol. Sci. 73, 233–244.
Brauer, K., Bruckner, G., Leibnitz, L., Werner, L., 1984. Structural and
cytochemical features of perineuronal glial nets in the rat brain. Acta
Histochem. 74, 53–60.
Brauer, K., Hartig, W., Bigl, V., Bruckner, G., 1993. Distribution of parvalbu-
min-containing neurons and lectin-binding perineuronal nets in the rat basal
forebrain. Brain Res. 631, 167–170.
Bruckner, G., Brauer, K., Hartig, W., Wolff, J.R., Rickmann, M.J., Derouiche,
A., Delpech, B., Girard, N., Oertel, W.H., Reichenbach, A., 1993. Perineur-
onal nets provide a polyanionic, glia-associated form of microenvironment
around certain neurons in many parts of the rat brain. Glia 8, 183–200.
Bruckner, G., Grosche, J., Hartlage-Rubsamen, M., Schmidt, S., Schachner, M.,
2003. Region and lamina-specific distribution of extracellular matrix pro-
teoglycans, hyaluronan and tenascin-R in the mouse hippocampal forma-
tion. J. Chem. Neuroanat. 26, 37–50.
Bruckner, G., Grosche, J., Schmidt, S., Hartig, W., Margolis, R.U., Delpech, B.,
Seidenbecher, C.I., Czaniera, R., Schachner, M., 2000. Postnatal develop-
ment of perineuronal nets in wild-type mice and in a mutant deficient in
tenascin-R. J. Comp. Neurol. 428, 616–629.
Bruckner, G., Hartig, W., Kacza, J., Seeger, J., Welt, K., Brauer, K., 1996.
Extracellular matrix organization in various regions of rat brain grey matter.
J. Neurocytol. 25, 333–346.
Bruckner, G., Hartig, W., Seeger, J., Rubsamen, R., Reimer, K., Brauer, K.,
1998. Cortical perineuronal nets in the gray short-tailed opossum (Mono-
delphis domestica): a distribution pattern contrasting with that shown in
placental mammals. Anat. Embryol. 197, 249–262.
Bruckner, G., Hausen, D., Hartig, W., Drlicek, M., Arendt, T., Brauer, K., 1999.
Cortical areas abundant in extracellular matrix chondroitin sulfate proteo-
glycans are less affected by cytoskeletal changes in Alzheimer’s disease.
Neuroscience 92, 791–805.
Bruckner, G., Seeger, G., Brauer, K., Hartig, W., Kacza, J., Bigl, V., 1994.
Cortical areas are revealed by distribution patterns of proteoglycan com-
ponents and parvalbumin in the Mongolian gerbil and rat. Brain Res. 658,
67–86.
rims around neurons located in the lower molecular layer (arrows). Neuropil zones ar
PNs in the medial (Med) cerebellar nucleus selectively immunoreactive for CSPG
nucleus. (D) BHABP staining in the medial cerebellar nucleus reveals clearly contou
associated with PNs in the medial cerebellar nucleus. Scale bars: 100 mm in (A)–
Bruckner, G., Szeoke, S., Pavlica, S., Grosche, J., Kacza, J., 2006. Axon initial
segment ensheathed by extracellular matrix in perineuronal nets. Neu-
roscience 138, 365–375.
Carlson, S.S., Hockfield, S., 1996. Central nervous system. In: Comper, W.D.
(Ed.), Extracellular Matrix. Tissue Function, vol. 1. Harwood Academic
Publishers, Amsterdam, pp. 1–23.
Carulli, D., Rhodes, K.E., Brown, D.J., Bonnert, T.P., Pollack, S.J., Oliver, K.,
Strata, P., Fawcett, J.W., 2006. Composition of perineuronal nets in the adult
rat cerebellum and the cellular origin of their components. J. Comp. Neurol.
494, 559–577.
Celio, M.R., 1993. Perineuronal nets of extracellular matrix around parvalbu-
min-containing neurons of the hippocampus. Hippocampus 3, 55–60.
Celio, M.R., Blumcke, I., 1994. Perineuronal nets—a specialized form of
extracellular matrix in the adult nervous system. Brain Res. Rev. 19,
128–145.
Celio, M.R., Spreafico, R., De Biasi, S., Vitellaro-Zuccarello, L., 1998.
Perineuronal nets: past and present. Trends Neurosci. 21, 510–515.
Comper, W.D., 1996. Extracellular Matrix. Tissue Function, vol. 1. Harwood
Academic Publishers, Amsterdam.
Corvetti, L., Rossi, F., 2005. Degradation of chondroitin sulfate proteoglycans
induces sprouting of intact Purkinje axons in the cerebellum of the adult rat.
J. Neurosci. 25, 7150–7158.
Dehay, C., Giroud, P., Berland, M., Killackey, H., Kennedy, H., 1996. Con-
tribution of thalamic input to the specification of cytoarchitectonic cortical
fields in the primate: effects of bilateral enucleation in the fetal monkey on
the bounderies, dimensions, and gyrification of striate and extrastriate
cortex. J. Comp. Neurol. 367, 70–89.
Delpech, B., Delpech, A., Bruckner, G., Girard, N., Maingonnat, C., 1989.
Hyaluronan and hyaluronectin in the nervous system. In: Evered, D.,
Whelan, J. (Eds.), The Biology of Hyaluronan. Ciba Foundation Sympo-
sium, vol. 143. John Wiley, Chichester, pp. 208–232.
Dityatev, A., Schachner, M., 2003. Extracellular matrix molecules and synaptic
plasticity. Nat. Rev. Neurosci. 4, 456–468.
Franklin, K.B.J., Paxinos, G., 1997. The Mouse Brain in Stereotaxic Coordi-
nates. Academic Press, San Diego.
Frost, S.B., Milliken, G.W., Plautz, E.J., Masterton, R.B., Nudo, R.J., 2000.
Somatosensory and motor representations in cerebral cortex of a primitive
mammal (Monodelphis domestica): a window into the early evolution of
sensory motor cortex. J. Comp. Neurol. 421, 29–51.
Guimaraes, A., Zaremba, S., Hockfield, S., 1990. Molecular and morphological
changes in the cat lateral geniculate nucleus and visual cortex induced by
visual deprivation are revealed by monoclonal antibodies Cat-304 and Cat-
301. J. Neurosci. 10, 3014–3024.
Hamel, E.G., 1967. A study of the hippocampal formation in the opossum,
Didelphis virginiana. In: Hassler, R., Stephan, H. (Eds.), Evolution of the
Forebrain. Plenum Press, New York, pp. 81–91.
Hamel, E.G., 1982. Telencephalon of marsupials. In: Crosby, E.C., Schnitzlein,
H.N. (Eds.), Comparative Correlative Neuroanatomy of the Vertebrate
Telencephalon. MacMillan, New York, pp. 317–337.
Hartig, W., Brauer, K., Bigl, V., Bruckner, G., 1994. Chondroitin sulfate
proteoglycan-immunoreactivity of lectin-labeled perineuronal nets around
parvalbumin-containing neurons. Brain Res. 635, 307–311.
Hartig, W., Brauer, K., Bruckner, G., 1992. Wisteria floribunda agglutinin-
labelled nets surround parvalbumin-containing neurons. NeuroReport 3,
869–872.
Hartig, W., Derouiche, A., Welt, K., Brauer, K., Grosche, J., Mader, M.,
Reichenbach, A., Bruckner, G., 1999. Cortical neurons immunoreactive
for the potassium channel Kv3.1b subunit are predominantly surrounded by
perineuronal nets presumed as a buffering system for cations. Brain Res.
842, 15–29.
Hazlett, J., Ho, R.H., Matin, G.F., 1991. Organization of midbrain catechola-
mine-containing nuclei and their projections to the striatum in the North
American opossum, Didelphis virginiana. J. Comp. Neurol. 306, 585–601.
e labeled in the granule cell layer (Gr). (C) CSPG/WFA double staining showing
. (C0) CSPG-immunoreactive, clearly contoured PNs in the medial cerebellar
red PNs. (D0) Parvalbumin immunoreaction showing perisomatic axonal profiles
(D); 20 mm in (C0) and (D0).

G. Bruckner et al. / Journal of Chemical Neuroanatomy 32 (2006) 143–158158
Hendry, S.H.C., Jones, E.G., Hockfield, S., McKay, R.D.G., 1988. Neuronal
populations stained with the monoclonal antibody CAT-301 in the mam-
malian cerebral cortex and thalamus. J. Neurosci. 8, 518–542.
Hensch, T.K., 2005. Critical period plasticity in local cortical circuits. Nat. Rev.
Neurosci. 6, 877–888.
Hobohm, C., Hartig, W., Brauer, K., Bruckner, G., 1998. Low expression of
extracellular matrix components in rat brain stem regions containing
modulatory aminergic neurons. J. Chem. Neuroanat. 15, 135–142.
Hockfield, S., 1993. Molecular correlates of activity-dependent development
and synaptic plasticity. In: Baudry, M., Thompson, R.F., Davis, J.L. (Eds.),
Synaptic Plasicity. Molecular, Cellular, and Functional Aspects. The MIT
Press, Cambridge, pp. 1–11.
Horn, A.K., Bruckner, G., Hartig, W., Messoudi, A., 2003. Saccadic omnipause
and burst neurons in monkeys and human are ensheathed by perineuronal
nets but differ in their expression of calcium-binding proteins. J. Comp.
Neurol. 455, 341–352.
Huffman, K., Nelson, J., Clarey, J., Krubitzer, L., 1999. Organization of
somatosensory cortex in three species of marsupials, Dasyurus hallucatus,
Dactylopsila trivirgata, and Monodelphis domestica: Neural correlates of
morphological specializations. J. Comp. Neurol. 403, 5–32.
Kahn, D.M., Krubitzer, L., 2002. Massive cross-modal cortical plasticity and
the emergence of a new cortical area in developmentally blind mammals.
Proc. Natl. Acad. Sci. 99, 11429–11434.
Kawaguchi, Y., Katsumaru, H., Kosaka, T., Heizmann, C.W., Hama, K., 1987.
Fast spiking cells in rat hippocampus (CA1 region) contain the calcium-
binding protein parvalbumin. Brain Res. 416, 369–374.
King, J.S., Bowman, M.H., Martin, G.F., 1971. The red nucleus of the opossum
(Didelphis marsupialis virginiana): a light and electron microscopic study.
J. Comp. Neurol. 143, 157–184.
Koppe, G., Bruckner, G., Hartig, W., Delpech, B., Bigl, V., 1997. Character-
ization of proteoglycan-containing perineuronal nets by enzymatic treat-
ments of rat brain sections. Histochem. J. 29, 1–10.
Kosaka, T., Heizmann, C.W., 1989. Selective staining of a subpopulation of
parvalbumin-containing GABAergic neurons in the rat cerebral cortex by
lectins with specific affinity for terminal N-acetylgalactosamine. Brain Res.
483, 158–163.
Martin, G.F., King, J.S., Dom, R., 1974. The projections of the deep cerebellar
nuclei of the opossum, Didelphis marsupialis virginiana. J. Hirnforsch. 15,
545–573.
Matthews, R.T., Kelly, G.M., Zerillo, C.A., Gray, G., Tiemeyer, M., Hockfield,
S., 2002. Aggrecan glycoforms contribute to the molecular heterogeneity of
perineuronal nets. J. Neurosci. 22, 7536–7547.
Morawski, M., Bruckner, M.K., Riederer, P., Bruckner, G., Arendt, T., 2004.
Perineuronal nets potentially protect against oxidative stress. Exp. Neurol.
188, 309–315.
Murakami, T., Tsubouchi, M., Tubouchi, Y., Taguchi, T., Ohtsuka, A., 1994.
The occurrence of neurons with strongly negatively charged surface coats in
mammalian, avian, reptilian, amphibian and piscine brains. Acta Med.
Okayama 48, 195–197.
Naegele, J.R., Barnstable, C.J., 1989. Molecular determinants of GABAergic
local-circuit neurons in the visual cortex. Trends Neurosci. 12, 28–34.
Olavarria, J.F., Li, C.P., 1995. Effects of neonatal enucleation on the organiza-
tion of callosal linkages in striate cortex of rats. J. Comp. Neurol. 361, 138–
151.
Oswaldo-Cruz, E., Rocha-Miranda, C.E., 1968. The Brain of the Opossum
(Didelphis marsupialis). A Cytoarchitectonic Atlas in Stereotaxic Coordi-
nates. Instituto de Biofisica, Universidade Federal do Rio de Janeiro, Rio de
Janeiro.
Palay, S.L., Chan-Palay, V., 1974. Cerebellar Cortex. Cytology and Organiza-
tion. Berlin, Springer Verlag.
Paxinos, G., Watson, C., 1998. The Rat Brain in Streotaxic Coordinates, fourth
ed. Academic Press, New York.
Pizzorusso, T., Medini, P., Berardi, N., Chierzi, S., Fawcett, J.W., Maffei, L.,
2002. Reactivation of ocular dominance plasticity in the adult visual cortex.
Science 298, 1187–1189.
Rauch, U., 2004. Extracellular matrix components associated with remodeling
processes in brain. Cell Mol. Life Sci. 61, 2031–2045.
Rhodes, K.E., Fawcett, J.W., 2004. Chondroitin sulphate proteoglycans: pre-
venting plasticity or protecting the CNS? J. Anat. 204, 33–48.
Rowe, M., 1990. Organization of the cerebral cortex in monotremes and
marsupials. In: Jones, E.G., Peters, A. (Eds.), Cerebral Cortex. Comparative
Structure and Evolution of Cerebral Cortex, vol. 8B. Plenum Press, New
York, pp. 263–334.
Seeger, G., Brauer, K., Hartig, W., Bruckner, G., 1994. Mapping of perineuronal
nets in the rat brain stained by colloidal iron hydroxide histochemistry and
lectin cytochemistry. Neuroscience 58, 371–388.
Semba, K., 2004. Phylogenetic and ontogenetic aspects of the basal forebrain
cholinergic neurons and their innervation of the cerebral cortex. Prog. Brain
Res. 145, 3–43.
Steiner, C., Tilak, M., Douzery, J.P., Catzeflis, F.M., 2005. New DNA data from
a transthyretin nuclear intron suggest an Oligocene to Miocene diversifica-
tion of living South America opossums (Marsupialia: Didelphidae). Mol.
Phylogen. Evol. 35, 363–379.
Sykova, E., 1997. The extracellular space in the CNS: its regulation, volume and
geometry in normal and pathological neuronal function. Neuroscientist 3,
28–41.
Wegner, F., Hartig, W., Bringmann, A., Grosche, J., Wohlfarth, K., Zus-
chratter, W., Bruckner, G., 2003. Diffuse perineuronal nets and modified
pyramidal cells immunoreactive for glutamate and the GABAA receptor
a1 subunit form a unique entity in rat cerebral cortex. Exp. Neurol. 184,
705–714.
Yamaguchi, Y., 2000. Chondroitin sulfate proteoglycans in the nervous system.
In: Iozzo, R.V. (Ed.), Proteoglycans. Structure, Biology, and Molecular
Interactions. Marcel Dekker, New York, pp. 379–402.



























