Embed Size (px)
Citation preview

Samar Island is the third largest island in thePhilippines archipelago with an approximateland area of 13,107 square kilometers,extending between 10°45’ and 12°45’ Nlatitude and between 124°15’ and 125°45’ Elongitude (PhilGIS 2016). The island’s regionalgeomorphology includes (1) the volcanic hills
and mountains at the northwest tip, (2) theheavily dissected plateau in the north, (3) thelimestone area that rims the central portion,(4) the basement area in the central andsouthern parts, (5) the rolling hills in thesouthwest and (6) the Guiuan peninsula(Travaglia et al. 1978). Based on the modified
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
161
Palms ofSamarIsland,Philippines
JIRO T. ADORADOR
Plant Biology Division, Institute of Biological Sciences, College of Arts and Sciences, The University of the Philippines -Los Baños, College, 4031 Laguna, [email protected]
EDWINO S. FERNANDO
Department of Forest BiologicalSciences, College of Forestry and NaturalResources, The University of the Philippines -Los Baños, College, 4031 Laguna, Philippines
Surveys of palms (Arecaceae) on Samar Island in the Philippines were conducted
in view of the island’s diverse but extremely threatened forest ecosystems, its old
conglomerate geology, its significant biogeography and its poor palm collection
history. Inventories of palms were carried out in selected sites between 2013 and
2016. The inventories yielded 50 species of which about 70% are Philippine
endemics. Twelve are new island records, and there are 11 probable novel taxa
(five in Calamus, and one each for Areca, Oncosperma, Heterospathe, Orania,
Pinanga and the Ptychospermatinae) while one is a recollection. Overall, the
palm flora of Samar reflects its biogeographic affinity and diverse habitat types.
Notes on the observed morphological variations of most species will be utilized
in the future taxonomic re-evaluation of some Philippine palm species and their
complexes. The wealth of information generated in this study will ultimately be
used for the conservation of palms and their habitats on the island.
PALMS 61(4): 161–195
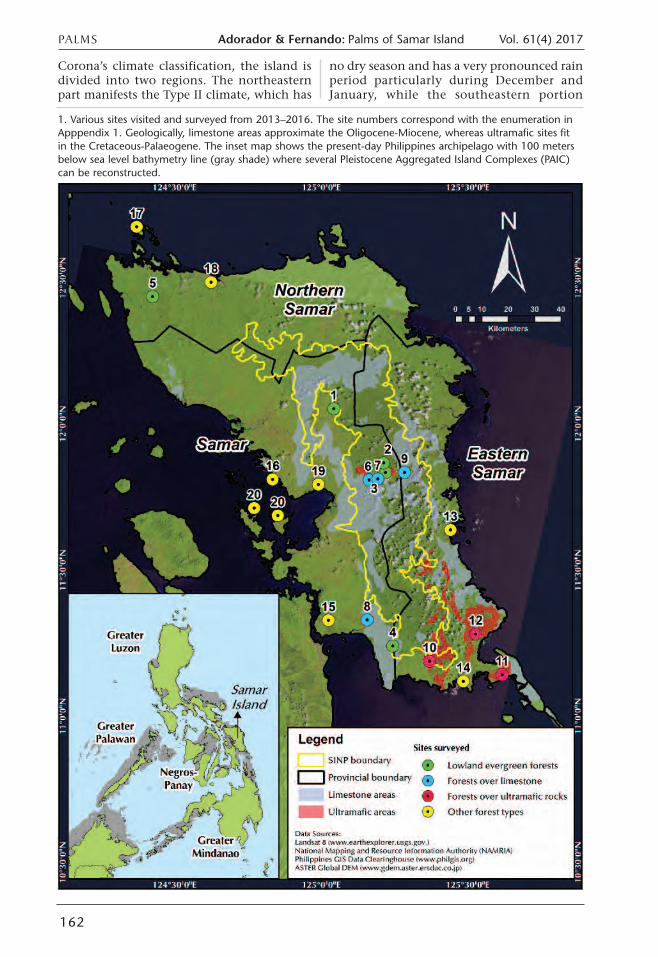
Corona’s climate classification, the island isdivided into two regions. The northeasternpart manifests the Type II climate, which has
no dry season and has a very pronounced rainperiod particularly during December andJanuary, while the southeastern portion
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
162
1. Various sites visited and surveyed from 2013–2016. The site numbers correspond with the enumeration inApppendix 1. Geologically, limestone areas approximate the Oligocene-Miocene, whereas ultramafic sites fitin the Cretaceous-Palaeogene. The inset map shows the present-day Philippines archipelago with 100 metersbelow sea level bathymetry line (gray shade) where several Pleistocene Aggregated Island Complexes (PAIC)can be reconstructed.

manifests the Type IV climate, which hasrainfall more or less evenly distributedthroughout the year.
Samar is part of the Greater Mindanaobiological region, which corresponds to theMindanao Pleistocence Aggregate Island
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
163
2 (top). The vast unexplored intact lowland forests viewed atop Mt. Huraw, San Jose de Buan, Samar. 3 (bottom). The contiguous lowland forests over rugged terrain towards the central mountains viewed atop akarst formation in Paranas, Samar. Photos by J.T. Adorador.

Complex (PAIC) that also includes Leyte,Bohol, Mindanao, Dinagat and Siargao(Heaney 1986) (see inset map in Fig. 1).Together with the eastern portion of Luzon(Sierra Madre range and Bicol peninsula), this
PAIC formed a continuous island arc intoHalmahera and New Guinea in the lateOligocene, about 25 Ma (Heaney 1986, Brown& Diesmos 2009, Vallejo 2011). This islandchain has been used by plants and animals to
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
164
4 (top). Secondary forests adjacent to local gold mining excavations in Guinmayuhan, Balangiga, E. Samar. 5 (bottom). A low karst forest along Sohoton River, Basey, Samar where Heterospathe intermedia abounds.Photos by J.T. Adorador.

migrate north and southwards (Lam 1945, Hall2001, Heads 2003, Vallejo 2011). The island isone of the centers of botanical diversity in thecountry and in the Malesian Region (Madulid2000). According to him there are at least sixforest formations found in the island, whichinclude (following the scheme of Fernando etal. 2008) lowland evergreen rainforest, lowermontane forest, forest over limestone, forestover ultramafic rocks, mangrove forest andbeach forest. Additionally, peat swamp forestwas hypothesized to occur in Dolongan, Basey(Davies 2006), whereas freshwater swampforests were observed by us along the seaward-flooded plains of Balangiga-Giporlos-Quinapondan. These two poorly documentedvegetation types in the island were found tobe degraded and converted into cultivatedareas or fishponds.
The contiguous forests found in centralportions of the island inevitably provide itsinhabitants with abundant water supply andrenewable forest products; likewise, it offerssanctuary to its rich flora and fauna. It wasdeclared a natural park (Samar Island NaturalPark or SINP) in 2003 through Presidential
Proclamation No. 422. The park comprises333,300 hectares of protected area and 125,400hectares of buffer zone – the largest terrestrialprotected area in the Philippines (UNDP-GEF2014). Despite this effort, the island’senvironment has been extensively degradedby logging and forest clearing for agriculturein the past 70 years (UNDP-GEF 2014).Likewise, the proliferation of small-scale illegallogging significantly contributes to forestdestruction. Coal and chromite miningactivities also pose a major threat to theisland’s biodiversity, while other threatsinclude unregulated quarrying of limestone,charcoal-making, over-harvesting of non-timber forest products (including rattans),pollution by industries and invasion of alienspecies (SEARCA 2004).
Although Samar Island is considered amongthe under-collected places for palms in thecountry (Fernando 1990a, c), an extensivereview of literature revealed about 35 speciesof indigenous palms listed for the island(Beccari 1919a, b, Pelser et al. 2011-onwards,Fernando 1990a, b, c, Madulid 2000, Gruezo& Harries 1984, SEARCA 2004, Baja-Lapis
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
165
6. (left). Calamus urdanetanus – recollected and rediscovered in Arizona, Hinabangan, Samar since itsdescription in 1919. 7 (right). Calamus daemonoropoides – note its densely bristly leaflets, sheath armed withdense partial whorls of spines, and unarmed sharply bulging knee. Photos by J.T. Adorador.

2010). Definitely, more taxa remain un-accounted since no taxonomic studies focusingon palms have been undertaken on the island.Preliminary site visits and subsequent researchand consultations, suggest several probablenew species of Pinanga and Calamus and apossible new genus in the Ptychospermatinae.
SINP’s diverse and conserved but extremelythreatened forest ecosystems, its oldconglomerate geology, its significantbiogeography and its poor palm collectionhistory altogether suggest the timeliness of thisendeavor and the likelihood of discoveringnovel palm taxa. A robust taxonomictreatment of palms and knowledge of theirecology essential for conservation were thegoals for this study.
Methods
Identification of potential sites for field surveyswas done through Geographic InformationSystem (GIS)-based processing techniquesthrough the aid of ASTER-DEM and variousthematic maps (e.g. forest cover, geology,administrative boundaries, and SINPboundary) (Fig. 1). All the eight forestformations (Fernando et al. 2008) that occur
in accessible and secured sites of the islandwere visited (Appendix 1, Figs. 2–5). Thenecessary gratuitous permit to carry out suchsurveys within SINP was granted by theDepartment and Natural Resources-Region 8(DENR-8) on July 9, 2015.
Complete palm specimens were collected(following Dransfield 1986), fertile if available,on several sites. Likewise, silica-dried leaf tissuesamples of most species were collected forfuture molecular studies. Oven-drying andmounting were executed in the Department ofForest Biological Sciences, College of Forestryand Natural Resources, University of thePhilippines Los Baños (UPLB). The first set ofpalm specimens are deposited at the ForestryHerbarium (LBC), UPLB Museum of NaturalHistory, while the other sets will be sent to thePhilippine National Herbarium (PNH) and theHerbarium of the Royal Botanic Gardens, Kew(K).
All available and relevant palm taxonomicliteratures were reviewed. Palm specimens werecarefully examined at the following herbaria:(1) LBC [Forestry Herbarium] and (2) CAHUP[College of Agriculture Herbarium–UP], bothmanaged by the UPLB Museum of Natural
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
166
8 (left). Habit of Calamus ochrolepis – note the characteristic elevated whorls of spines in the leaf sheath. 9 (right). Dense infructescence of Plectocomia elongata var. philippinensis. Photos by J.T. Adorador.

PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
167
10. The habit of Saribus rotundifolius – note its highly-branched infructescence. Photo by J.T. Adorador.

PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
168
11. Habit of the stilt-rooted Areca costulata – note the closely-set multifold leaflets and yellowish crownshaft.Photo by J.T. Adorador.

History and (3) EBL [ERDB BotanicalLaboratory and Herbarium (formerly known asFORI)] under the ERDB-DENR. Appropriateparts were measured in herbarium sheets andof specimens in situ.
The morphological species concept of differentPhilippine-occurring palm lineages providedthrough the works of Beccari (1919a, b), Baker(2015), Baker et al. (1999), Dransfield (1980,
1984, 1997), Dransfield et al. (2008), Fernando(1988a, b, 1990b, 2014), Baja-Lapis (1989),Madulid (1981), Lewis (2002), Hahn(unpublished), Heatubun et al. (2012, 2014),Dowe (2009), Bacon and Baker (2011), andKeim and Dransfield (2012), werecircumspectly analyzed and utilized in speciesdelimitation. The syntheses of data gatheredfrom numerous taxonomic articles, herbariummaterial and field observations generated the
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
169
12. The diminutive Pinanga sp. “tagibungang-liitan” – note its short coriaceous leaves and distichously-setfruits. Photo by J.T. Adorador.

PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
170
13. The spiny Oncosperma sp. “anibong-kinis” – note the glabrous prophyll and few-branched Infructescence.Photo by J.T. Adorador.

functional broad morphological speciesconcept used herein to delimit the indigenouspalms of the island.
Results and Discussions
The Palm Flora of Samar Island
The series of surveys across the island revealeda total of 50 palm species distributed in 16genera together comprising 26 calamoids, 16arecoids, seven coryphoids and the sole nypoid(Appendix 2). Calamus is the most species-richgenus on the island with 23 species, distantlyfollowed by Pinanga and Areca (each with 4species), then Heterospathe (3 species). Twenty-seven species were previously recorded fromthe island, while 12 are new records and 11 areprobable new taxa. Overall, it represents aquarter (39/139) of the currently known palmspecies (not including varieties and subspecies)in the country. Meanwhile, Philippineendemicity is about 70% (34/50), which islocally similar to the palm flora of the 16-haPalanan Forest Dynamics Plot (Yap 2010) andregionally consistent with the total vascularflora of the Philippine archipelago (Myers et al.
2010). Among the recognized taxa, only one(Pinanga samarana) is reported endemic to theisland, while most of the probable new speciesare believed to be similarly restricted. Despitethe island’s relative smaller geographic area,species richness is comparable with that ofSulawesi Java and Moluccas with 62, 46 and 40species, respectively (Baker & Couvreur 2012).The transitory biogeographic setting of theisland could partially explain the speciescompositional trend found there.
It is notable that Calamus urdanetanus (Fig. 6),previously known only from its type locality,Mt. Cabadbaran (Mt. Urdaneta) in Agusan delNorte, Mindanao island, was recollected forthe first time since its description by Beccariin 1919. For the new island records, sevenspecies were already predicted to occur sincethese palms were then known to be distributedacross Greater Mindanao (except Samar) andGreater Luzon island complexes (e.g. Calamusdaemonoropoides, C. ochrolepis, Plectocomiaelongata var. philippinensis (Figs. 7–9).Meanwhile, two species are recorded for thefirst time outside Mindanao, C. multinervis and
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
171
14 (left). The terminal portion of Calamus siphonospathus var. dransfieldii – note the dense brown sheathspines and indistinct ocrea. 15 (right). Habit of flowering Calamus moseleyanus – note its broad-ellipsoid dullgreen leaflets and diffusely yet stiff paniculate inflorescence. Photos by J.T. Adorador.

PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
172
16. The infructescence of Pinanga insignis – note its robust peduncle and distichously-set shortly-beaked fruits.Photo by J.T. Adorador.

the recollected rattan, whereas Saribus cf.merrillii and Livistona saribus are now foundoutside Luzon (Fernando et al. 2008). This
conspectus of updated geographic distributionresolves the palm disjunction of Luzon-Mindanao pattern (Fernando 1990a).
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
173
17. Habit of fruiting Orania decipiens – note highly branched inflorescence axes. Photo by J.T. Adorador.

Eleven are believed to be novel taxa but stillwarrant further investigation through studyof all available herbarium material and/orapplication of molecular techniques: five inCalamus, and one each for Areca, Oncosperma,Heterospathe, Orania, Pinanga and thePtychospermatinae. The species in the last four
genera are morphologically distinctive and arecurrently being formally described, while thosein the first three require more in-depth studies.The discovery of these new palm taxa is afunction of new collections from a poorly-known area, correspondence with several palmexperts and recent taxonomic updates.
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
174
18. Areca sp. “mudulan” showing its slender habit and immature Infructescence. Photo by J.T. Adorador.

The morphological characters that were foundto be very variable and diverse in herbariumspecimens and in situ include leaflet venation,the shape of the fruits and leaf sheathindumentum for the arecoids, while suchcharacters are sheath armature, inflorescencelength and number of fruits scales for thecalamoids. On the contrary, characters thatwere found to be consistent across differenthabitats and developmental stages for allpalms comprise general stem form, leafletbristles and covering, and overall inflorescencearchitecture.
Habitat and Ecology
There remain vast tracts of dense forestsscattered throughout the island, especiallythose growing over limestone areas and in thecentral portions where communist rebelsinexorably make camps. Among the island’sforest types, only three were surveyedcomprehensively on selected sites, whichinclude the lowland evergreen, over limestone,and over ultramafic rocks. Several palms werenoted to be widespread amidst the perceivedecological differences between the threevegetation types. Such rattans include Calamusmollis, C. ochrolepis, C. ornatus and Plectocomiaelongata var. philippinensis; on the other hand,
C. aidae and C. merrillii were rarely found overlimestone areas. Meanwhile, the widespreaderect palms include Saribus rotundifolius (Fig.10), Areca caliso, A. costulata (Fig. 11), Pinangasamarana, P. copelandii, P. sp. “tagibungang-liitan,” Caryota cumingii and C. rumphiana;Oncosperma sp. “anibong-kinis” was occa-sionally spotted over limestone-derived soils.
The widespread though sporadic occurrenceof a single new species of Pinanga (Fig. 12)across different forest types is rather surprising.It was first encountered near the summit of Mt.Huraw (800 m elevation) then atop towerkarsts of Tenani (400 m), ridges of Arizona (300m) and Vigan (200 m). Its several morpho-types were then regarded to be distinctivespecies but were dismissed to be just ecotypicvariations. Generally, this taxon could becharacterized by its clustering diminutive andvery slender habit, the typically pluricostulatecoriaceous leaflets, and short inflorescencewith few reflexed rachillae bearing two rows ofovoid-ellipsoidal fruits. This novel taxongenerally prefers ridges where vegetation isintact and light is abundant.
The probable new species of Oncosperma (Fig.13) shares morphological affinity with theLuzon-restricted O. gracilipes and the
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
175
19. Rattan sapling diversity in the lower slopes of Mt. Huraw – in the foreground (left to right) are Plectocomiaelongata var. philippinensis, Calamus mollis and C. filispadix, while in the background are C. aidae and C.daemonoropoides. Photo by J.T. Adorador.

Mindanao-widespread O. horridum and couldrepresent a distinctive race of hybrid origin.This clustering palm is characterized by itsmedian leaflets abaxially bristly on the distalportions (first report for the genus), unarmedprophyll and fruits developing only on thecentral portions of the rachillae. It grows wellin sheltered valleys but becomes spindlytowards the ridges.
Lowland evergreen forests
The lowland evergreen forests occupy thegreatest vegetation coverage within the park,which gradually merge with other forest typeshence creating ecotonal boundaries. Thosespecies observed to be restricted to this vege-tation include the rarities (e.g. Calamuspedicellaris, C. siphonosphathus var. dransfieldii(Fig. 14) and Saribus cf. merrillii) and a probablenew Calamus (C. sp. “tagsa-on”). Likewise,several species were shared with forests overultramafic areas, which comprise Calamuselmerianus, C. filispadix, C. megaphyllus, C.moseleyanus (Fig. 15), C. vidalianus, Pinangainsignis (Fig. 16) and Orania decipiens (Fig. 17).Such habitat sharing also occurs in the sus-pected new Areca sp. “mudulan” and Calamussp. “malubagacay-bilog.”
The inferred new Areca (Fig. 18) was observedto be sympatric (in some areas) with thelimestone-preferring A. caliso and the eurytopicA. costulata. It is more similar to the former butis distinctive in its much more slender habit,shorter leaves with generally 2-costulateleaflets and its much shorter inflorecsence withthe uppermost rachillae still bearing at leastone floral triad on its basal portions. Suchvariations in the vegetative traits could bedictated by the differences of the substrate butthose of the floral characteristics need to be re-evaluated (along with other Philippine Areca)– if this really represents a new species.
Meanwhile, the other inferred new taxa inCalamus approach the “C. mindorensis group”(including C. mindorensis, C. moseleyanus, C.multinervis and C. reyesianus), which localpeople generally refer to as malubagacay orlokuan. Beccari (1919a) recognized this artificialgroup (except C. reyesianus) but did not providea conspectus of it. Nevertheless, C. sp. “tagsa-on” (Back Cover) is distinctive in its glabrouscoriaceous leaflets typically in pairs and shortererect inflorescence. On the other hand, C. sp.“malubagacay-bilog” though only non-fertile
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
176
20 (left). Pinanga samarana – notice the peeled portion of the crownshaft, evidence of palm-heart extractionby monkeys. 21 (right). Calamus filispadix with a bird’s nest hanging from its flagellum. Photos by J.T. Adorador.

plants were seen, is rather distinctive in itsvery chartaceous broad-lanceolate leaflets andfine, continuous tertiary veins. These tworattans together with the “C. mindorensisgroup” need taxonomic reappraisal sinceseveral morphological heterogeneities wereobserved in the field, such as leaflet groupingsupon development or exposure to light andpersistence and/or absence of sheath armatureon mature individuals.
Among the intact lowland forests visited, theslopes of Mt. Huraw in San Jose de Buanoffered the highest palm diversity with 29species (17 rattans and 12 erect palms). Alongthe forest edges, there could be as much asfive different species of juvenile rattans in a 3m× 3m plot (Fig. 19) – not including theseedlings of Oncosperma and Areca. Theuniqueness of palm composition in thislocality could be due to the dense canopy,hence very little light reaching the understory.Pinanga samarana was uncommon along thetrails.
Meanwhile, the secondary forests of Arizona,Hinabangan and Ulot, Paranas possessed analmost similar palm floristic composition toMt. Huraw but differed in the abundance ofPinanga samarana, Calamus aidae, and C.filispadix. Thus, the proliferation of these palmscould be due to the increased light intensity
penetrating the forest floor of these relativelydisturbed forests. Interestingly, traces of palmheart extraction by monkeys were observed inP. samarana (Fig. 20) and the two rattansmentioned were recorded to provide suitableshelter for forest animals (Figs. 21 & 22).
Forests over limestone
The forests over limestone are arguably themost intact vegetation type on Samar Island.This is due to the difficulty of penetrating thethickets of its abruptly variable terrain; thus,illegal logging is simply not profitable. It islocally known as the kaigangan with karstformations, which reach up to 400 melevation. Across the sites surveyed,Heterospathe intermedia (Fig. 23), Areca caliso,Pinanga copelandii (Front Cover), Calamusdiscolor (Fig. 24), C. multinervis (Fig. 25) and C.ochrolepis, are the most characteristic specieswithin the valleys and mid-slopes of this forestformation. The palm floristic gradient thenchanges towards the peaks of karst towers.There abounds Pinanga samarana, morpho-logically different from the lowland counter-parts in having more slender stems, uni-costulate leaflets and more reflexed rachillaewith obovoid fruits. However, all these traitshifts could be attributed to environmentaldifferences of their habitats, especially lightand soil conditions.
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
177
22. Calamus aidae with its interwoven basal leaflets made as a nest by a forest rodent. Photo by J.T. Adorador.

PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
178
23. The habit of Heterospathe intermedia. Photo by J.T. Adorador.

Interstingly, four new taxa were discoveredatop this forest type. The novel rattans includetwo slender clustering Calamus, pudlos-uban(Fig. 26) and pudlos-liitan (Fig. 27) of San Rafael,Taft (Eastern Samar) and Tenani, Paranas(Samar), respectively. The former differs fromtypical C. discolor in having less spiny, glaucousgreen sheaths and epetiolate leaves with lessbristly leaflets. Meanwhile, the latter species isapparently allied to C. microsphaerion butultimately differs in the unarmed rachis andconcave short-lanceolate apiculate leaflets,which are grouped in pairs. This rattan wasrecorded to produce shoots on cut stems,which could be the reason for its abundanceon the summit. Aside from these traits, theirinflorescences details ultimately segregatethem from their respective analogouscounterparts.
The new species of slender Orania (Fig. 28)atop the calcareous mountain is rathersurprising since its counterparts thrive in areaswhere there is much thicker soil. Its closestmatch is O. littoralis of Papua New Guineausing the artificial key by Keim and Dransfield(2011). However, the inflorescence morpho-
logy is the most unusual among all knownOrania species, in developing an all-staminateinflorescence (without triads along therachillae) and a bisexual type (with floraltriads). Moreover, its fruit traits (color,mesocarp thickness and embryo shape) arerecorded to be different with those of O.decipiens when compared in fresh state – whichcould be used in re-annotating usefulcharacters for the Philippine species.
Perhaps, the most thrilling discovery of theentire palm flora of the island would be themember of Ptychospermatinae, known locallyas amuring (Fig. 29). It is morphologicallyembedded within the subtribe in its praemorseleaflet apices and bullet-shaped buds ofstaminate flowers. Moreover, after con-sultations with several experts on the subtribe(S. Zona, W. Baker and C. Heatubun), reviewof recent literature (Alapetite et al. 2014,Heatubun et al. 2012) and herbarium studies,we believe this taxon merits a unique genericplacement. It deviates from other members indetails of its inflorescence and fruit structure.It is restricted to the summits of karstformations in the central part of the island.
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
179
24 (left). Calamus discolor – note its dense brown spines and petiolate leaves with leaflets glaucous abaxially.24 (right). Habit of Calamus multinervis – note its brown sheath indumentum and plaited leaflets grouped inpairs which are observed in individuals under dense forest cover. Photos by J.T. Adorador.

The observed populations of amuring, whichcomprise several fruiting individuals, fewjuveniles but numerous seedlings generallysuggest its adaptability but slow growth andestablishment to the unique habitat. Fruitdoves (Ptilonopus spp.) were observed to feedon the thin but fleshy and bland-tastingmesocarps of this palm.
Forests over ultramafic rocks
In contrast, the remaining vegetation overultramafic areas on southeast portions of theisland were generally disturbed. Aside fromthe mining activities and illegal timberextraction of nearby communities, such areaswere devastated by one of the history’sstrongest typhoons (Haiyan) in November 3,2013 and most of them subsequentlysuccumbed to wild-(or man-induced) fires inthe summer of 2014. The forest surveys werecarried-out in the most accessible intact forestsoutside mining concession areas due to thedifficulty of acquiring permits from thecompanies.
Nevertheless, the fieldwork showed the generalpalm floristic make-up across the three metal-rich sites, which are dominated by thewidespread Pinanga samarana, Areca spp.,Calamus aidae and Plectocomia elongata var.
philippinensis. It is within these forest typesthat the extent of morphological variation ofP. samarana was observed and verified. Theencountered populations from these sites differfrom the typical lowland and limestonemorphotypes in having 3-pluricostulate leafletsand twisted rachillae with globose-ovoid fruits.Meanwhile, Orania decipiens, Calamuselmerianus, C. megaphyllus (Fig. 30), C.vidalianus (Fig. 31) and C. sp. “tagda” werelocally abundant in Guinmayuhan, Balangiga,whereas the new slender Pinanga, Calamusochrolepis, C. sp. “malubagacay-bilog” andHeterospathe sp. “saguisi-mina” were veryabundant in Vigan, Gen. MacArthur.
The sites’ unique edaphic conditionscharacterized by heavy metal concentrationand low litter-nutrient inputs paved way forthe niche specialization of certain plants suchas Nepenthes spp. This habitat differentiationmight also explain occurrence of the two newultramafic-restricted palms, Calamus sp.“tagda” (Fig. 32) and Heterospathe sp. “saguisi-mina” (Fig. 33). The novel flagellate rattan ischaracteristic in its long-petiolate leaves and itssheath armature, reminiscent of cirrate C.loherianus in Luzon. It is unique amongPhilippine species in having the proximal jointof the flagella ornamented with a crown of
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
180
26 (left). The slender Calamus sp. “pudlos-uban” – note its epetiolate discolorous leaves and less spiny sheath.27 (right). Calamus sp. “pudlos-liitan” with its short lanceolate leaflets grouped in pairs. Photos by J.T. Adorador.

PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
181
28. Orania sp. “banga-igang” with its very slender habit. Photo by J.T. Adorador.

PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
182
29. The habit of Ptychospermatinae or amuring with its broadly-truncate, apically praemorse leaflets andwhitish inflorescence axes. Photo by J.T. Adorador.

spines. Fertile collections would make suchcomparisons more complete. Meanwhile, thelatter slender clustering erect palm hassimilarities with wider-ranging H. philippinensisbut is different in the shorter leaf outline, thushaving fewer leaflets and purplishinflorescence axes. Across different sites, thispalm likewise exhibited noteworthy variationin inflorescence branching order and numberof its peduncular bracts and leaflets. These twopalms commonly prefer exposed ridges wherethe canopy is more open, but they are locallymore abundant on reddish soil.
Other habitats (freshwater swamp, beach, andmangrove)
The freshwater swamps along seaward coastalportions of the island were mainly convertedinto rice fields or coconut plantations.Remnants of natural vegetation in thesewaterlogged sites mainly comprisediscontinuous stands of Teminalia,Anthocephalus and Pandanus. The palmssporadically thriving here include Metroxylonsagu (Fig. 34) and Livistona saribus (Fig. 35).The latter fan palm is characterized by itsheavily armed petiole, drooping leaf apices
and bluish fruits, while the former clusteringpalm has erect dark-green leaves and trunks dieafter a massive apical flowering. Sago palmstands were not communally maintained andutilized since the fragmented clusters seemedto be only established by chance. Meanwhile,Nypa fruticans typically borders rice fieldswhere they border brackish water zonestowards the mangroves.
Beach forests of appreciable extent can only beencountered in uninhabited coasts or smallerislands where Heterospathe elata, Calamusdiscolor, C. usitatus and Caryota cumingiiflourish. These narrow coastal vegetation stripsare separated from the central lowland forestsby large expanses of cultivated lands orabandoned clearings where few plants ofArenga pinnata can be found.
Economic use of some palms
Among the rattans, at least five robust species(Calamus aidae, C. merrillii, C. moseleyanus, C.multinervis and C. ornatus) were recorded asused in furniture-making industry. Meanwhile,those slender-cane taxa used in handicrafts(such as weaving material and cordage) include
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
183
30 (left). Sapling of Calamus megaphyllus with its broad oblanceolate leaflets. 31 (right). Habit of clusteringCalamus vidalianus with its irregularly arranged leaflets. Photos by J.T. Adorador.

PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
184
32. The flagellate Calamus sp. “tagda” – note the dense brown spines of the sheath and short lamina outline.Photo by J.T. Adorador.

PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
185
33. Heterospathe sp. “saguisi-mina” – note its several undivided leaves and purplish inflorescence axes. Photoby J.T. Adorador.

C. daemonoropoides, C. discolor, C. filispadix, C.megaphyllus, C. mollis, C. sp. “tagda” and C.sp. “pudlos-liitan.” Some species provide foodfor inhabitants such as C. ornatus for its sourfruits and C. aidae for its bitter palm heart.
As for the erect palms, the leaves of severalspecies (Corypha utan, Livistona saribus andSaribus rotundifolius, Metroxylon sagu, Nypafruticans and Cocos nucifera) are used forthatching and roofing materials. Aside from
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
186
34 (top). Young stand of the clustering Metroxylon sagu. 35 (bottom). Remnant population of Livistona saribusin Basey, Samar with several cultivated Cocos nucifera in the background. Photos by J.T. Adorador.

the conventional uses of coconut, its fruit andfermented sap are commercially utilized forcopra (coco-meat) and lambanog, respectively.Local people seldom utilize the durable woodof Orania decipiens as house posts. On a lesserscale, the three forest Areca species wereobserved to serve as substitutes for A. catechufor their traditional mamâ. Likewise, Caryotarumphiana and Pinanga insignis are known fortheir palatable palm heart, while those of P.samarana are also said to be edible thoughbitter to taste.
Threats to their habitat
The never-ending small-scale timber poachingand slash-and-burn (kaingin) practices alwayspose significant threats to the island’s richforest biodiversity. Several widespread palmswere recorded to thrive in heavily disturbedlowland forests (such as Pinanga samarana andPlectocomia elongata var. philippinesis), but thisdoes not correspond to their general ecologicaltolerance as a lineage. These palms are naturalinhabitants of intact forests wherein their
population dynamics respond to naturaldisturbances such as canopy gaps and soilerosion. Thus, prolonged exposure to suchman-induced environmental stresses couldresult to the decrease of palm speciescomposition or even to the extirpation of theirrespective populations. Such is the case for thetwo annihilated populations of the newdiminutive Pinanga in the carbon-minedwasteland of Arizona, Hinabangan (Samar),wherein kaingin is frequently practiced forcassava cultivation (Fig. 36). Moreover, the1980s agricultural conversion of the entireDolongan peatlands at Basey (Samar) couldhave inflicted the same drastic trend to itspalm flora before we even knew what existedthere.
Over-harvesting of the rattan canes for thefurniture industry in past decades has resultedin the absence of mature rattans butabundance of juvenile plants in almost everyforest type visited. In Mt. Huraw, Arizona,Marabut, and Guinmayuhan, long canes of
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
187
36 (left). The extirpated cluster of Pinanga sp. “tagibungang-liitan” in Arizona, Hinabangan, Samar – note therolled and scorched leaves which could be due to extreme heat following kaingin practice. 37 (right). One ofthe numerous saplings of Calamus merrillii – a remnant of the over-collection of mature canes in Mt. Huraw,San Jose de Buan. Photos by J.T. Adorador.

Calamus merrillii (Fig. 37) and C. aidae werenot seen since local people had eagerlyharvested these prime species. The currenteconomic-paucity of the said industry couldpave the way to the re-establishment of thenatural populations, which must besustainably managed in the future.
Several sites on the entire island are declaredmineral reservations (e.g. laterite, bauxite,gold) – just waiting for any legal propositionsfor these ores to be extracted. In several sites,fuel-wood gathering and unproductive farmingpractices incessantly pressurize the spoiledvegetation. These destructive activities likelyjeopardize the habitats of ultramafic-restrictedpalms. Naturally, the rapidly transformedenvironment suppresses their respectiveecological niches and consequent reproductivestrategies. The surveyed burnt forests of theprevious laterite-exhumed concession area inCarapdapan, Salcedo (Eastern Samar) revealedthe sparse occurrence of Calamus sp. “tagda”and the disappearance of the new Heterospathe.
On the other hand, the palm species foundon the inaccessible interior rugged terrains offorests over limestone (e.g. Ptychospermatinae,Orania sp. “banga-igang,” Calamus sp. “pudlos-liitan”) are projected to be among the leastthreatened by human extracting activities.However, natural disasters such as typhoonsand prolonged droughts could bring the palmflora to its ecological tipping point in thislimiting environment.
Summary and Conclusions
A survey of palms on Samar Island has revealedthat its palm flora is very rich and particularlydiverse and unusual, comprising 50 specieswith 70% Philippine endemism, 12 newrecords and 11 probable new taxa. Acrossdifferent vegetation types and degree ofdisturbances, several palm species wereobserved to present morphological plasticity.Scrutiny of taxonomic literature, herbariummaterial and field observations required theapplication a broad morphological speciesconcept on the island’s palms. When taken ina broader context, this realization inevitablycalls for a thorough review of severalPhilippine palm species and their complexes.Despite most of the island’s remaining forestsbeing declared “protected,” human-inducedactivities constantly exert significantenvironmental pressures to the palms foundthere. The results of these surveys willultimately be used towards the conservation ofpalms and their habitats on the island.
Recommendations
Population surveys of the economicallyimportant rattans subject to commercialextraction should be carried out so thatsustainable management can be proposed.Thorough habitat surveys of the novel palmsand knowledge on their ecology andphenology are also recommended.
The interior swamps of Bermudahan inHinabangan, Samar, the highlands of thecentral portions of the island (Mts. Capotoanand Concord), and the volcanic soils in thenorthwestern part (Victoria-Calbayog) couldsupport unique palm flora, but these sites werenot included due to security reasons. Thus, ifrebel tensions dissipate and funds are available,surveys in these areas could produce furtherbotanical novelties.
Acknowledgments
The authors are indebted to Dr. WilliamGruezo and Dr. Nathaniel Bantayan for theirsound advice throughout the conduct of thestudy. To several palm experts who providedassistance in several ways, Drs. William Baker,Scott Zona, Charlie Heatubun, Fred Essig, AidaBaja-Lapis, Ary Keim, William Hahn, John L.Dowe and Christine Bacon. Dr. Pieter Pelserprovided access to certain publications. Wethank the keepers of three herbaria namely,LBC, CAHUP and EBL. JTA would also like toexpress gratitude to the DOST-ASTHRDP-NSCscholarship and the International Palm SocietyEndowment Fund for providing scholarshipand research grants. The SINP-PAMB, CENROand PENRO-Catbalogan, DENR-R8 are thankedfor providing assistance from processing thegratuitous and transport permits, especially tothe following; Zenaida Baisa (PASu), Eires Mate,Jovelyn Grovoso, Gene Tomas (EMS-Catbalogan), Marcialito Torres (CENRO-Catbalogan), and other members of the SINP-PAMB. We thank our field guides in variouslocalities; Titu, Michael, David, Rommel,Marlito, Marlon, Using, Leonardo, to theMamanwa tribe, Buchoy, Larry, Gloria familyand Lito. Most importantly JTA thanks hisdearest girlfriend, Zhereeleen Meneses, for herassistance and companionship on and offduring fieldwork.
LITERATURE CITED
ALAPETITE, E., W.J. BAKER AND S. NADOT. 2014.Evolution of stamen number inPtychospermatinae (Arecaceae): Insightsfrom a new molecular phylogeny of thesubtribe. Molecular Phylogenetics andEvolution 76: 277–240.
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
188

BACON, C.D. AND W.J. BAKER. 2011. Saribusresurrected. Palms 55: 109–116.
BAJA-LAPIS, A.C. 1989. An account of taxarelated to Calamus siphonospathus complex.Sylvatrop 2: 91–96.
BAJA-LAPIS, A.C. 2010. A field guide toPhilippine rattans. Asia Life SciencesSupplement 5: 1–214.
BAKER, W.J. 2015. A revised delimitation of therattan genus Calamus (Arecaceae). Phytotaxa197: 139–152.
BAKER, W.J. AND T.L.P. COUVREUR. 2012.Biogeography and distribution patterns ofSoutheast Asian palms. Pp. 164–190. In D.J.GOWER ET AL. (eds.). Biotic Evolution andEnvironmental Change in Southeast Asia.Cambridge University Press. .
BAKER, W.J., J. DRANSFIELD, M.M. HARLEY AND A.BRUNEAU. 1999. Morphology and cladisticanalysis of subfamily Calamoideae (Palmae).In A.J. HENDERSON AND F. BORCHSENIUS (eds.).Evolution, Variation and Classification ofPalms. Pp. 307–324. The New York BotanicalGarden Press, Bronx, NY.
BECCARI, O. 1919a. Palms of the PhilippineIslands. Leaflets of Philippine Botany 8:2997–3067.
BECCARI, O. 1919b. The palms of the PhilippineIslands. Philippine Journal of Science(Botany) 14: 295–362.
BROWN, R. AND A.C. DIESMOS. 2009. Philippines,biology. Pp. 723–732. In GILLESPIE, R.G AND
D.A. CLAGUE (eds). Encyclopedia of Islands.University of California Press, Berkeley.
DAVIES, J. 2006. National Action Plan forSustainable Use and Protection of PhilippinePeatlands. Pp. 39. Report for IFAD-GEFProject Proposal.
DOWE, J.L. 2009. A taxonomic account ofLivistona R.Br. (Arecaceae). Gardens BulletinSingapore 60: 185–344.
DRANSFIELD, J. 1980. On the identity of Sika inPalawan. Kalikasan, The Philippine Journalof Biology 9: 43–48.
DRANSFIELD, J. 1984. The Rattans of Sabah.Sabah Forest Record No. 13, ForestDepartment, Sabah. Pp. 182.
DRANSFIELD, J. 1986. A guide to collecting palms.Annals of the Missouri Botanical Garden 73:166–176.
DRANSFIELD, J. 1997. The Rattans of BruneiDarrussalam. Forestry Department Brunei
Darrusalam and The Royal botanic Gardens.Pp. 200.
DRANSFIELD, J., N.W. UHL, C.B. ASMUSSEN, W.J.BAKER, M.M. HARLEY AND C.E. LEWIS. 2008.Genera Palmarum – the Evolution andClassification of Palms, ed. 2. Royal BotanicGardens, Kew, Richmond, UK. Pp. 732.
FERNANDO, E.S. 1988a. The mottled-leavedspecies of Pinanga in the Philippines.Principes 32: 165–174.
FERNANDO, E.S. 1988b. Four new taxa ofPhilippine rattans (Palmae Calamoideae).Gardens’ Bulletin Singapore 41: 49–58.
FERNANDO, E.S. 1990a. A preliminary analysis ofthe palm flora of the Philippine islands.Principes 34: 28–45.
FERNANDO, E.S. 1990b. The genus Heterospathe(Palmae: Arecoideae) in the Philippines. KewBulletin 45: 219–234.
FERNANDO, E.S. 1990c. Diversity and con-servation status of the Philippine rattans.Rattan Information Centre Bulletin(Malaysia) 9: 7–21.
FERNANDO, E.S. 2007–present. A ProvisionalChecklist of the Palms of the Philippines.Unpublished.
FERNANDO, E.S. 2014. Three new species inCalamus sect. Podocephalus (Arecaceae:Calamoideae) from the Philippines,Indonesia, and Papua New Guinea.Phytotaxa 166: 069–076.
FERNANDO, E.S., M.H. SUH, J. LEE, AND D.K. LEE.2008. Forest Formations of the Philippines.ASEAN-Korea Environmental CooperationUnit (AKECU), Seoul National University,Korea. Pp. 232.
GRUEZO, W.S. AND H.C. HARRIES. 1984. Self-sown,wild-type coconuts in the Philippines.Biotropica 16: 140–147.
HAHN, W.J. (unpublished). Revision of Caryota.Pp. 195.
HALL, R. 2001. Cenozoic reconstruction of SEAsia and the SW Pacific; changing patternsof land and sea. In METCALFE, I., J.M.B. SMITH,M. MORWOOD AND I.D. DAVIDSON (eds.), Faunaland Floral Migrations and Evolution in SEAsia – Australia. Pp. 35-56. AA Balkema(Swets & Zeitlinger Publishers), Lisse.
HEADS, M.J. 2003. Ericaceae in Malesia:vicariance biogeography, terrane tectonicsand ecology. Telopea 10: 311–449.
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
189

HEANEY, L.R. 1986. Biogeography of mammalsin SE Asia: estimates of rates of colonization,extinction, and speciation. Biological Journalof the Linnean Society 28: 127–165.
HEATUBUN, C.D., J. DRANSFIELD, F. THOMAS, S.TJITROSOEDIRDJO, J.P. MOGEA AND W.J. BAKER.2012. A monograph of the betel but palms(Areca: Arecaceae) of East Malesia. BotanicalJournal of the Linnean Society 168: 147–173.
HEATUBUN, C.D., S. ZONA AND W.J. BAKER. 2014.Three new genera of arecoid palm(Arecaceae) from eastern Malesia. KewBulletin 69: e9525.
KEIM, A.P. AND J. DRANSFIELD. 2012. A mono-graph of the genus Orania (Arecaceae:Oranieae). Kew Bulletin 67: 127–190.
LAM, H.J. 1945. Notes on the historical phyto-geography of Celebes. Blumea 5: 600–640.
LEWIS, C.E. 2002. A phylogenetic analysis ofthe palm subtribe Oncospermatinae (Are-caceae) based on morphological characters.Brittonia 54: 78–91.
MADULID, D.A. 1981. A monograph ofPlectocomia (Palmae: Lepidocaryoideae).Kalikasan, Philippine Journal of Biology 10:1–94.
MADULID, D.A. 2000. A review and assessmentof the floristic knowledge of Samar Island:based on literature, PNH Records, andcurrent knowledge. USAID. Pp. 16.
MYERS, N, R.A. MITTERMEIER, C.G. MITTERMEIER,G.A. DA FONSECA AND J. KENT. 2000.
Biodiversity hotspots for conservationpriorities. Nature 403: 853–858.
PELSER, P.B., J.F. BARCELONA AND D.L. NICKRENT.2011-onwards. Co’s Digital Flora of thePhilippines. www.philippineplants.org.Accessed on January 13, 2016.
PHILIPPINES GIS DATA CLEARINGHOUSE (PhilGIS).2016. http://www.philgis.org.
SEAMEO REGIONAL CENTER FOR GRADUATE STUDY
AND RESEARCH IN AGRICULTURE (SEARCA). 2004.Biological Resources Assessment. SamarIsland Natural Park. Philippines. FinalReport. United Nations DevelopmentProgramme (UNDP).
TRAVAGLIA, C., A.F. BAES AND L.M. TOMAS. 1978.Geology of Samar Island. Bureau of Soils,Department of Agriculture – United NationsDevelopment Programme – Food andAgriculture Organization of the UnitedNations. Manila. Pp. 149.
UNITED NATIONS DEVELOPMENT PROGRAMME –GLOBAL ENVIRONMENT FACILITY (UNDP-GEF).2014. http://www.undp.org.
VALLEJO, B. 2011. The Philippines in Wallacea.Pp. 27–42. In Telnov, D. (ed.). Biodiversity,Biogeography and Nature Conservation inWallacea and New Guinea, Vol. I. TheEntomological Society of Latvia.
YAP, S.L. 2010. Phylogeography and Demo-graphy of Common Plant Species from thePhilippine Islands. PhD Dissertation. TheUniversity of Michigan.
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
190
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
191
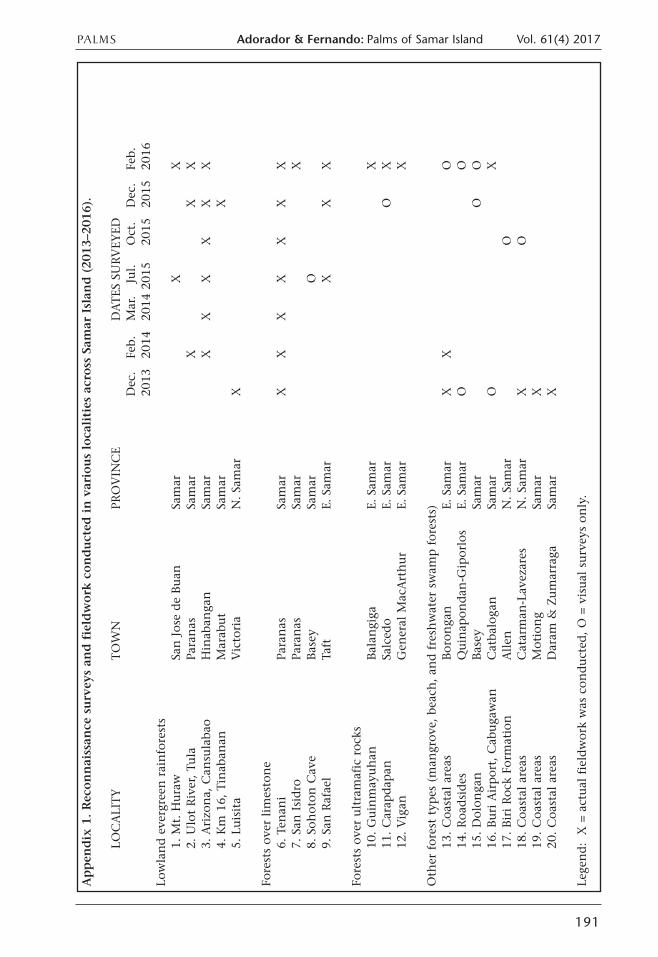
Appen
dix 1. Reconnaissan
ce survey
s an
d fieldwork
conducted
in various loca
lities across Sam
ar Islan
d (20
13–2
016).
LOC
ALI
TY
TO
WN
PRO
VIN
CE
DATES
SU
RV
EYED
Dec
. Fe
b.M
ar.
Jul.
Oct
.D
ec.
Feb.
2013
2014
2014
201
520
1520
1520
16Lo
wla
nd e
verg
reen
rai
nfo
rest
s1.
Mt. H
ura
wSa
n Jos
e de
Buan
Sam
arX
X2.
Ulo
t Riv
er, Tu
laPa
ranas
Sam
arX
XX
3. A
rizo
na,
Can
sula
bao
Hin
aban
gan
Sam
arX
XX
XX
X4.
Km
16,
Tin
aban
anM
arab
ut
Sam
arX
5. L
uis
ita
Vic
toria
N. Sa
mar
X
Fore
sts
over
lim
esto
ne
6. T
enan
iPa
ranas
Sam
arX
XX
XX
XX
7. S
an Isi
dro
Para
nas
Sam
arX
8. S
ohot
on C
ave
Bas
eySa
mar
O9.
San
Raf
ael
Taft
E. S
amar
XX
X
Fore
sts
over
ultra
maf
ic r
ocks
10. G
uin
may
uhan
Bal
angi
gaE.
Sam
arX
11. C
arap
dap
anSa
lced
oE.
Sam
arO
X12
. V
igan
Gen
eral
Mac
Art
hur
E. S
amar
X
Oth
er for
est ty
pes
(m
angr
ove,
bea
ch, an
d fre
shw
ater
sw
amp for
ests
)13
. C
oast
al a
reas
Bor
onga
nE.
Sam
arX
XO
14. Roa
dsi
des
Quin
apon
dan
-Gip
orlo
sE.
Sam
arO
O15
. D
olon
gan
Bas
eySa
mar
OO
16. Buri A
irpor
t, C
abuga
wan
Cat
balo
gan
Sam
arO
X17
. Biri Roc
k Fo
rmat
ion
Allen
N. Sa
mar
O18
. C
oast
al a
reas
Cat
arm
an-L
avez
ares
N. Sa
mar
XO
19. C
oast
al a
reas
Mot
iong
Sam
arX
20. C
oast
al a
reas
Dar
am &
Zum
arra
gaSa
mar
X
Lege
nd: X
= a
ctual
fie
ldw
ork
was
con
duct
ed, O
= v
isual
surv
eys
only
.
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
192
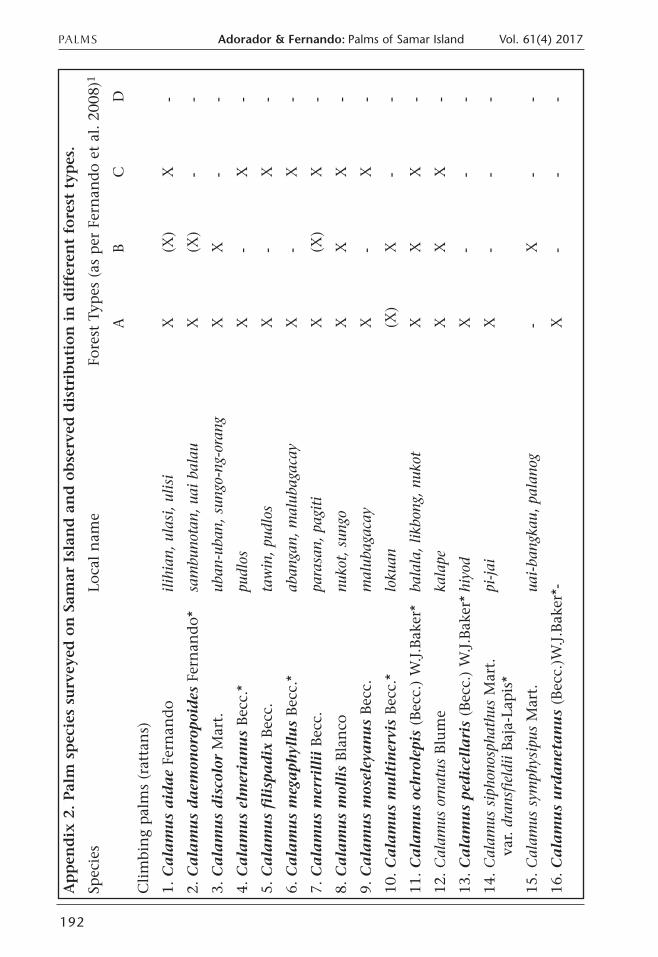
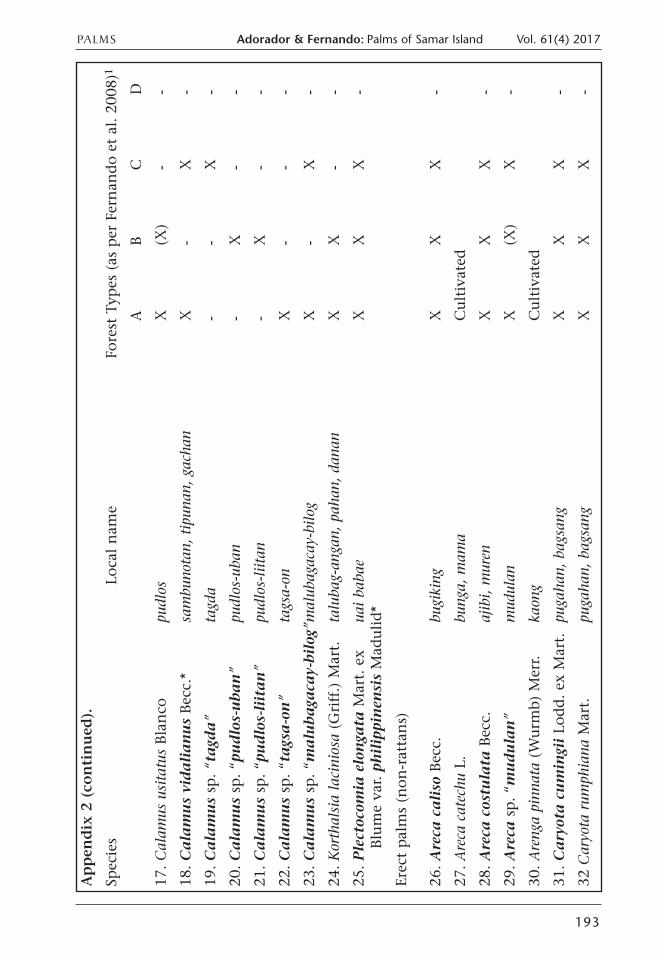
Appen
dix 2. Palm species su
rvey
ed on Sam
ar Islan
d and observed
distribution in different forest typ
es.
Spec
ies
Loca
l nam
eFo
rest
Typ
es (as
per
Fer
nan
do
et a
l. 2
008)
1
AB
CD
Clim
bing
pal
ms
(rat
tans)
1. C
ala
mu
s a
ida
eFe
rnan
do
ilihi
an, ul
asi,
ulisi
X(X
)X
-
2. C
ala
mu
s d
aem
onor
opoi
des
Fern
ando*
sam
buno
tan,
uai b
alau
X(X
)-
-
3. C
ala
mu
s d
isco
lor
Mar
t.ub
an-u
ban,
sun
go-n
g-oran
gX
X-
-
4. C
ala
mu
s el
mer
ian
us
Bec
c.*
pudlos
X-
X-
5. C
ala
mu
s fi
lisp
ad
ixBec
c.ta
win
, pu
dlos
X-
X-
6. C
ala
mu
s m
ega
phyl
lus
Bec
c.*
aban
gan,
malub
agac
ayX
-X
-
7. C
ala
mu
s m
erri
llii
Bec
c.pa
rasa
n, pag
iti
X(X
)X
-
8. C
ala
mu
s m
olli
sBla
nco
nuko
t, sun
goX
XX
-
9. C
ala
mu
s m
osel
eya
nu
sBec
c.m
alub
agac
ayX
-X
-
10.
Ca
lam
us
mu
ltin
ervi
sBec
c.*
loku
an(X
)X
--
11.
Ca
lam
us
och
role
pis
(Bec
c.) W
.J.B
aker
*ba
lala, lik
bong
, nu
kot
XX
X-
12. Calam
us orn
atus
Blu
me
kalape
XX
X-
13.
Ca
lam
us
ped
icel
lari
s(B
ecc.
) W
.J.B
aker
*hi
yod
X-
--
14. Calam
us sipho
nosp
hath
usM
art.
pi-jai
X-
--
var. d
rans
fieldiiBaj
a-La
pis
*
15. Calam
us sym
phys
ipus
Mar
t.ua
i-ba
ngka
u, palan
og-
X-
-
16.
Ca
lam
us
urd
an
eta
nu
s(B
ecc.
)W.J.B
aker
*-X
--
-
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
193
Appen
dix 2 (co
ntinued
).
Spec
ies
Loca
l nam
eFo
rest
Typ
es (as
per
Fer
nan
do
et a
l. 2
008)
1
AB
CD
17. Calam
us u
sita
tus
Bla
nco
pudlos
X(X
)-
-
18.
Ca
lam
us
vid
ali
an
us
Bec
c.*
sam
buno
tan,
tipun
an, ga
chan
X-
X-
19.
Ca
lam
us
sp. “t
agd
a”
tagd
a-
-X
-
20.
Ca
lam
us
sp. “p
ud
los-
uba
n”
pudlos
-uba
n-
X-
-
21.
Ca
lam
us
sp. “p
ud
los-
liit
an”
pudlos
-liit
an-
X-
-
22.
Ca
lam
us
sp. “t
ags
a-o
n”
tags
a-on
X-
--
23.
Ca
lam
us
sp. “m
alu
baga
cay-
bilo
g”m
alub
agac
ay-b
ilog
X-
X-
24. Kor
thalsia
lacini
osa
(Griff.) M
art.
talu
bag-an
gan,
pah
an, da
nan
XX
--
25.
Ple
ctoc
omia
elo
nga
taM
art. e
x ua
i ba
bae
XX
X-
Blu
me
var. p
hil
ippi
nen
sis
Mad
ulid*
Erec
t pal
ms
(non
-rat
tans)
26.
Are
ca c
ali
soBec
c.bu
giking
XX
X-
27. Areca
cat
echu
L.bu
nga, m
ama
Cultiv
ated
28.
Are
ca c
ostu
lata
Bec
c.ajibi,
mur
enX
XX
-
29.
Are
casp
. “m
ud
ula
n”
mud
ulan
X(X
)X
-
30. Areng
a pinn
ata
(Wurm
b) M
err.
kaon
gC
ultiv
ated
31.
Ca
ryot
a c
um
ingi
iLo
dd. ex
Mar
t.pu
gaha
n, b
agsa
ngX
XX
-
32 C
aryo
ta rum
phiana
Mar
t.pu
gaha
n, b
agsa
ngX
XX
-
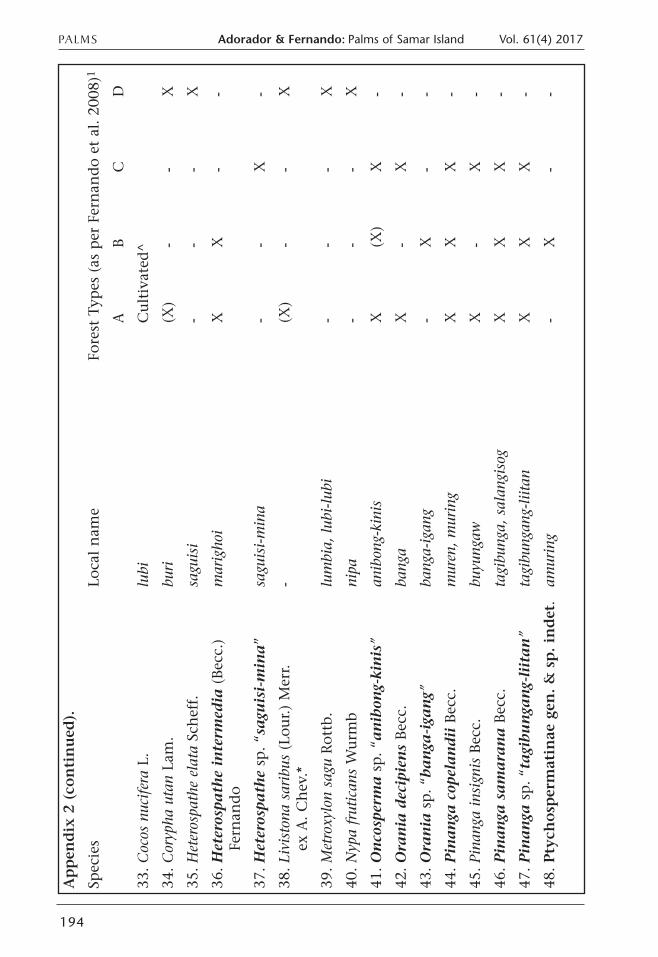
PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
194
Appen
dix 2 (co
ntinued
).
Spec
ies
Loca
l nam
eFo
rest
Typ
es (as
per
Fer
nan
do
et a
l. 2
008)
1
AB
CD
33. Coc
os n
ucifera
L.lu
biC
ultiv
ated
^
34. Cor
ypha
uta
nLa
m.
buri
(X)
--
X
35. Heteros
path
e elat
aSc
hef
f.sa
guisi
--
-X
36.
Het
eros
path
e in
term
edia
(Bec
c.)
mar
igho
iX
X-
-Fe
rnan
do
37.
Het
eros
path
esp
. “s
agu
isi-
min
a”
sagu
isi-m
ina
--
X-
38. Li
viston
a sa
ribu
s(L
our.) M
err.
-(X
)-
-X
ex A
. C
hev
.*
39. M
etroxy
lon
sagu
Rot
tb.
lum
bia, lub
i-lu
bi-
--
X
40. Nyp
a frut
ican
sW
urm
bni
pa-
--
X
41.
On
cosp
erm
asp
. “a
nib
ong-
kin
is”
anibon
g-kini
sX
(X)
X-
42.
Ora
nia
dec
ipie
ns
Bec
c.ba
nga
X-
X-
43.
Ora
nia
sp. “b
an
ga-i
gan
g”ba
nga-
igan
g-
X-
-
44.
Pin
an
ga c
opel
an
dii
Bec
c.m
uren
, m
urin
gX
XX
-
45. Pi
nang
a in
sign
isBec
c.bu
yung
awX
-X
-
46.
Pin
an
ga s
am
ara
na
Bec
c.ta
gibu
nga, salan
giso
gX
XX
-
47.
Pin
an
gasp
. “t
agi
bun
gan
g-li
ita
n”
tagibu
ngan
g-lii
tan
XX
X-
48. Ptych
osp
ermatinae
gen
. & sp. indet
.am
urin
g-
X-
-

PALMS Adorador & Fernando: Palms of Samar Island Vol. 61(4) 2017
195
Appen
dix 2 (co
ntinued
).
Spec
ies
Loca
l nam
eFo
rest
Typ
es (as
per
Fer
nan
do
et a
l. 2
008)
1
AB
CD
49.
Sari
bus
cf.
mer
rill
ii(B
ecc.
) Bac
on &
an
ahaw
, bigo
-buk
idX
--
-W
.J. Bak
er*^
^
50. Sa
ribu
s rotu
ndifoliu
s(L
am) Blu
me
anah
awX
XX
-
Lege
nd:
1 H
abitat
s: A
= low
land e
verg
reen
. B =
ove
r lim
esto
ne,
C =
ove
r ultra
maf
ic r
ocks
, D
= s
wam
py
(fre
sh o
r br
acki
sh)
Taxa
in bold
are
Philip
pin
e en
dem
ic s
pec
ies
(whic
h is
also
lik
ely
to a
pply
to
pos
sibl
y new
spec
ies)
. Thos
e w
ith a
ster
isk
(*) ar
e new
rec
ords
for
the
isla
nd.
The
sym
bol ‘(X
)’ indic
ates
that
the
spec
ies
was
enco
unte
red v
ery
rare
ly in s
uch
for
est ty
pe.
^The
repor
ted p
opula
tion
of nat
ive C. nu
cifera
(Gru
ezo
& H
arries
198
4) in the
Guiu
an p
enin
sula
was
not
vis
ited
.
^^This
pal
m s
uper
fici
ally
res
embl
es S. m
errilli
iba
sed o
n g
ross
veg
etat
ive
mor
phol
ogy
(Dow
e, p
ers. c
om.), bu
t fe
rtile
collec
tion
s ar
e nee
ded
to
verify
its
iden
tity
.





























