Embed Size (px)
Citation preview

i
Parasites and cleaning behaviour in damselfishes
Derek Sun
BMarSt, Honours I
A thesis submitted for the degree of Doctor of Philosophy at
The University of Queensland in 2015
School of Biological Sciences

ii
Abstract
Pomacentrids (damselfishes) are one of the most common and diverse group of marine fishes found
on coral reefs. However, their digenean fauna and cleaning interactions with the bluestreak cleaner
wrasse, Labroides dimidiatus, are poorly studied. This thesis explores the digenean trematode fauna
in damselfishes from Lizard Island, Great Barrier Reef (GBR), Australia and examines several
aspects of the role of L. dimidiatus in the recruitment of young damselfishes.
My first study aimed to expand our current knowledge of the digenean trematode fauna of
damselfishes by examining this group of fishes from Lizard Island on the northern GBR. In a
comprehensive study of the digenean trematodes of damselfishes, 358 individuals from 32 species
of damselfishes were examined. I found 19 species of digeneans, 54 host/parasite combinations, 18
were new host records, and three were new species (Fellodistomidae n. sp., Gyliauchenidae n. sp.
and Pseudobacciger cheneyae). Combined molecular and morphological analyses show that
Hysterolecitha nahaensis, the single most common trematode, comprises a complex of cryptic
species rather than just one species. This work highlights the importance of using both techniques in
conjunction in order to identify digenean species. The host-specificity of digeneans within this
group of fishes is relatively low. Most of the species possess either euryxenic (infecting multiple
related species) or stenoxenic (infecting a diverse range of hosts) specificity, with only a handful of
species being convincingly oioxenic (only found in one host species). Overall, the trematode
richness in pomacentrid fishes is relatively low compared with that of many other coral reef fish
families. The richness is probably best explained by the small size of most pomacentrids. In terms
of beta diversity, of the total 23 trematode species having now been reported for the GBR in
pomacentrids, just 14 are shared between Heron and Lizard Island. Five species have been recorded
only from Lizard Island and four only from Heron Island. A total of 14 species reported from the
two sites can be classified as core pomacentrid-specific parasites.
I then examined whether the abundance and diversity of damselfish recruits differed on reefs
relative to reefs from which cleaner wrasse had been removed for 12 years. Past studies have shown
that cleaner wrasse enhance abundance and diversity of adult resident damselfishes (species that are
site-attached), however, it is not known whether this effect occurs at recruitment or is driven by
post-settlement events such as migration or differential mortality. I characterised the abundance of
damselfish recruits on reefs with and without cleaner wrasse for five days in each of three lunar
cycles, beginning four days after the new moon (November, December and January). The total
abundance of damselfish recruits and specifically two species, Chrysiptera rollandi and P.
amboinensis, was higher on reefs with cleaner wrasse than on reefs without. However, overall

iii
species diversity of damselfish recruits did not differ. This study shows that the effects of the
presence of cleaner wrasse begin at recruitment.
Although the presence of cleaner wrasse increases the abundance of recruiting damselfish, it
is unclear what mechanism(s) are involved in this process. I conducted aquarium experiments to
examine whether young P. amboinensis preferentially select a microhabitat that is adjacent to a
cleaner wrasse. The study showed that both settlement-stage and juvenile P. amboinensis spend a
greater proportion of time in a microhabitat adjacent to a cleaner wrasse than in one adjacent to a
control wrasse (a non-cleaner wrasse, Halichoeres melanurus) or where no fish was present. More
time was spent next to juvenile than adult cleaner wrasse. I then conducted field observations to
determine if recruits that are in a 1 m radius of a juvenile cleaner wrasse receive any direct benefits
(cleaning) and whether juvenile cleaner wrasse preferentially cleaned larger individuals. The size
and identity of all fish that were cleaned within a 1 m radius of a cleaner were recorded, along with
nearby non-cleaned conspecifics. I showed that only 2 % of the clients that a juvenile cleaner
wrasse clean per 20 min observation were small recruits (< 20 mm, TL) whereas larger
damselfishes comprised 58 % of clients. Juvenile cleaner wrasse also preferred to clean larger
individuals than the median size of the surrounding nearby non-cleaned conspecifics. These results
show that cleaner wrasse serve as a positive cue during microhabitat selection but that direct
benefits to a small recruit are not obvious. The possibilities of indirect rather than direct (cleaning)
benefits are discussed.
I then used sentinel traps to investigate the cumulative number of gnathiid isopods infecting
the ambon damselfish, Pomacentrus amboinensis, on patch reefs with and without cleaner wrasse.
Previous studies may have under surveyed the number of gnathiids on a host related to the presence
or absence of cleaner fish, due to the sampling techniques used (wired cages or handnets). As
gnathiids are temporary parasites, they can drop off a host and return to the benthos before the fish
is sampled; sentinel traps are able to retain such individuals. Nocturnal collections produced almost
three times more gnathiids than diurnal collections. These results are consistent with previous
studies. There was no difference in the number of gnathiids found in sentinel traps placed on reefs
with and without a cleaner wrasse. These results appear initially to be at variance with the many
advantages to fishes reported as being associated with the presence of L. dimidiatus. These
observations lead to the novel hypothesis that the advantage to clients from cleaners is in the
reduction in the length of parasitism rather than the number of infections being reduced.
This research has provided important information regarding the parasites of damselfishes,
from the starting point of the characterisation of the parasite fauna to how cleaning behaviour
affects the ecology of recruiting fish, thus increasing our understanding of recruitment processes on
coral reefs. My study has demonstrated that the richness of damselfishes exceeds that of their

iv
trematode parasites on the GBR although the parasite diversity (11 families) is remarkable. The
work has contributed insight into both host range and species richness, having uncovered both novel
trematode species, and novel hosts for known species. Regarding cleaning interactions, I have
shown that 1) the abundance of damselfish recruits is higher on reefs with cleaner wrasse than on
those without; 2) damselfish recruits preferentially select a microhabitat that is adjacent to a cleaner
wrasse; and 3) that the benefits for young fish to be near cleaners are likely to be indirect rather than
direct. These aspects of my study demonstrates the importance of cleaning mutualism not only for
adult fishes, but for juvenile fishes during the recruitment period.

v
Declaration by author
This thesis is composed of my original work, and contains no material previously published or
written by another person except where due reference has been made in the text. I have clearly
stated the contribution by others to jointly-authored works that I have included in my thesis.
I have clearly stated the contribution of others to my thesis as a whole, including statistical
assistance, survey design, data analysis, significant technical procedures, professional editorial
advice, and any other original research work used or reported in my thesis. The content of my thesis
is the result of work I have carried out since the commencement of my research higher degree
candidature and does not include a substantial part of work that has been submitted to qualify for
the award of any other degree or diploma in any university or other tertiary institution. I have
clearly stated which parts of my thesis, if any, have been submitted to qualify for another award.
I acknowledge that an electronic copy of my thesis must be lodged with the University Library and,
subject to the policy and procedures of The University of Queensland, the thesis be made available
for research and study in accordance with the Copyright Act 1968 unless a period of embargo has
been approved by the Dean of the Graduate School.
I acknowledge that copyright of all material contained in my thesis resides with the copyright
holder(s) of that material. Where appropriate I have obtained copyright permission from the
copyright holder to reproduce material in this thesis.

vi
Publications during candidature
Sun D, Bray RA, Yong R, QY, Cutmore SC, Cribb TH (2014) Pseudobacciger cheneyae n. sp.
(Digenea: Gymnophalloidea) from Weber’s chromis (Chromis weberi Fowler & Bean)
(Perciformes: Pomacentridae) at Lizard Island, Great Barrier Reef, Australia. Systematic
Parasitology 88:141-152
Sun D, Blomberg SP, Cribb TH, McCormick MI, Grutter AS (2012) The effects of parasites on the
early life stages of a damselfish. Coral Reefs 31:1065-1075
Publications included in this thesis
Incorporated as part of Chapter 2: Sun D, Bray RA, Yong R, QY, Cutmore SC, Cribb TH (2014)
Pseudobacciger cheneyae n. sp. (Digenea: Gymnophalloidea) from Weber’s chromis (Chromis
weberi Fowler & Bean) (Perciformes: Pomacentridae) at Lizard Island, Great Barrier Reef,
Australia. Systematic parasitology 88:141-152
TH Cribb, RA Bray and SC Cutmore provided feedback and comments on the manuscript. R. Q-Y
Yong provided editorial advice and assistance with specimen measurements. SC Cutmore assisted
with phylogenetic analyses. D Sun collected and analysed the data and wrote the manuscript.

vii
Contributions by others to the thesis
Chapter 1: TH Cribb provided feedback and comments. D Sun was responsible for the remainder
of the work.
Chapter 2: TH Cribb, RA Bray and R. Q-Y Yong provided feedback and comments on the
manuscript. SC Cutmore assisted with phylogenetic analyses. D Sun collected and analysed the data
and wrote the manuscript.
Part of Chapter 2: TH Cribb, RA Bray and SC Cutmore provided feedback and comments on the
manuscript. R. Q-Y Yong provided editorial advice and assistance with specimen measurements.
SC Cutmore assisted with phylogenetic analyses. D Sun collected and analysed the data and wrote
the manuscript.
Chapter 3: KL Cheney and AS Grutter provided advice on experiment design. KL Cheney, J
Werminghausen, TH Cribb and AS Grutter provided feedback and comments on the manuscript. D
Sun designed and conducted the study, collected and analysed the data and wrote the manuscript.
Chapter 4: KL Cheney and AS Grutter provided advice on experimental design and statistical
procedures. J Werminghausen assisted with data collection. KL Cheney, MI McCormick, MG
Meekan, TH Cribb and AS Grutter provided with feedback and comments on the manuscript. D Sun
designed and conducted the study, collected and analysed the data and wrote the manuscript.
Chapter 5: KL Cheney and AS Grutter provided advice on experimental design and statistical
procedures. J Werminghausen and EC McClure assisted with field data collection. KL Cheney, J.
Werminghausen, EC McClure, MI McCormick, MG Meekan, TH Cribb and AS Grutter provided
editorial and feedback on the manuscript. D Sun designed and conducted the study, collected and
analysed the data and wrote the manuscript.
Chapter 6: TH Cribb provided feedback and comments. D Sun was responsible for the remainder
of the work.
Statement of parts of the thesis submitted to qualify for the award of another degree
None

viii
Acknowledgements
There are many people who need be thanked for their contribution to this project. However, first I
would like to thank my two supervisors, Tom Cribb and Karen Cheney. Tom, your enthusiasm and
knowledge in regards to parasites is remarkable. Throughout the years, including my honours years,
you have guided and supported me and for that I am thankful. Karen, thank you for being
supportive, sharing your knowledge and providing me with invaluable advice. I particularly want to
acknowledge the support from Alexandra Grutter. Your encouragement, passion and enthusiasm for
anything that I did, whether it be small or large made my candidature enjoyable. I’ve learnt so much
and I can’t thank you enough for all of your guidance and advice. Words cannot express my
gratitude to you for everything you have done for me. I would also like to thank both Mark
McCormick and Mark Meekan for reading drafts, providing me with advice and for allowing me to
use their light traps. Also a thank you to both Simon Blomberg and Jeff Hanson for helping me with
all of my questions about statistics.
I am truly grateful for the staff at the Lizard Island Research Station. I thank you for the
support and the friendship that you have all showed me throughout the years. To Lyle, Anne,
Marianne, Lance, Tania and Bob I can’t thank you all enough for helping me with this research, you
have all gone way beyond what is required. Thank you all for the memorable events that we have
all shared. They will never be forgotten.
I am deeply thankful to all the people who have helped me throughout my candidature;
Johanna, Eva, Fabio, Genevieve, Maarten, Russell, Scott, Anne and Andy. I would especially like
to thank Johanna, Eva and Fabio for assisting me in the field, all without ever once complaining for
having to get up at ungodly hours to help me with the light traps or the sentinel traps. Your
assistance is one of the main reasons that I am able to complete this thesis. Genevieve and Maarten
thank you for all of your help in the field, which includes helping me get out of that nally-bin on the
boat. A special thank you goes to Russell, who throughout this PhD has read countless drafts and
has taught me so much about marine parasitology. Scott for helping me with molecular analysis, as
I’m utterly hopeless at it; Matt for teaching me how to put out the light traps and Anne and Andy
for keeping me sane with our regular need for coffee and for your helpful comments and support. I
am also extremely thankful to the members of the Marine Parasitology Lab, Coral Reef Ecology
Lab and the people who occupy room 113 of Goddard throughout my candidature.
I would like to thank the organisations that have provided me with financial support for the
research conducted throughout my candidature. These include the Australian Research Council
Discovery Grant (ARC grant to A. S. G. et al. and K. L. C. and T. H. C. et al.) and Sea World
Research and Rescue Foundation, Australia (SWRRFI) for project funding. I would also like to

ix
thank the International Society for Behavioural Ecology for the student travel award that has
allowed me the opportunity to present this research at the conference in New York.
Finally, I would like to thank my family and friends for always being supportive. Mum and
Dad, thank you for all of your love and financial support. I can’t thank you enough. To my brothers
and sister, William, Henry and Michelle, thank you for always being there for me, even though you
thought that all I did was swim with fishes. I am also grateful to my close friends Karina, Glen,
Justin and Simon for their constant support and kindness. I could always count on you guys to put a
smile on my face. Love all of you guys. Thanks.

x
Keywords
gnathiid isopods, cleaning mutualism, recruitment, trematodes, damselfish, ectoparasites, host-
specificity, Great Barrier Reef, beta diversity
Australian and New Zealand Standard Research Classifications (ANZSRC)
ANZSRC code: 060201, Behavioural Ecology, 50%
ANZSRC code: 060205, Marine and Estuarine Ecology, 20%
ANZSRC code: 060301, Animal Systematics and Taxonomy, 30%
Fields of Research (FoR) Classification
FoR code: 0602, Ecology, 70%
FoR code: 0699, Other Biological Sciences, 30%

xi
Table of Contents
Abstract ............................................................................................................................................... ii Declaration by author ........................................................................................................................ v Publications during candidature ..................................................................................................... vi Publications included in this thesis .................................................................................................. vi
Contributions by others to the thesis.............................................................................................. vii Statement of parts of the thesis submitted to qualify for the award of another degree ............ vii Acknowledgements.......................................................................................................................... viii Keywords ............................................................................................................................................ x Australian and New Zealand Standard Research Classifications (ANZSRC) ............................. x Fields of Research (FoR) Classification ........................................................................................... x
Table of Contents .............................................................................................................................. xi List of Figures .................................................................................................................................. xiii
List of Tables ................................................................................................................................... xiv Abbreviations ................................................................................................................................... xv Chapter 1: General Introduction...................................................................................................... 1 1.1 Preamble 2
1.2 Pomacentrids 2
1.3 Parasitism on coral reefs 2
1.4 Parasite diversity on coral reefs 3
1.5 Ecological importance of parasites 5
1.6 Cleaning mutualism 8
1.7 Aims and Thesis structure 11
Chapter 2: Trematodes of northern Great Barrier Reef Pomacentridae: richness, host-
specificity and beta diversity ........................................................................................................... 14 Abstract 15
Introduction 15
Materials and methods 17
Results 19
Discussion 34
Acknowledgments 41
Pseudobacciger cheneyae n. sp. (Digenea: Gymnophalloidea) from Weber’s chromis (Chromis
weberi Fowler & Bean) (Perciformes: Pomacentridae) at Lizard Island, Great Barrier Reef,
Australia ............................................................................................................................................ 42 Abstract 43
Introduction 43
Materials and methods 44
Gymnophalloidea Odhner, 1905 incertae familiae 47
Discussion 51
Acknowledgements 56
Chapter 3: Presence of cleaner wrasse increases the recruitment of fishes to coral reefs ........ 57 Abstract 58
Introduction 58
Material and methods 59
Results 60
Discussion 64
Acknowledgements 65

xii
Chapter 4: Cleaner wrasse influence habitat selection of young damselfish .............................. 66 Abstract 67
Introduction 68
Materials and methods 69
Results 74
Discussion 78
Acknowledgements 82
Chapter 5: Diel patterns of infection by gnathiid isopods on the ambon damsel, Pomacentrus
amboinensis, and the influence of the presence of cleaner wrasse ............................................... 83 Abstract 84
Introduction 84
Materials and methods 88
Results 93
Discussion 95
Acknowledgements 98
Chapter 6: General Discussion ....................................................................................................... 99 References ....................................................................................................................................... 103 Appendices ...................................................................................................................................... 113

xiii
List of Figures
Chapter 2: Trematodes of northern Great Barrier Reef Pomacentridae: richness, host-
specificity and beta diversity
Figure 1 Body length and ventral sucker length of the three morphological types of Hysterolecitha
nahaensis 24
Figure 2 Neighbour joining tree based on ITS2 rDNA sequences of Hysterolecitha nahaensis
complex from Lizard Island. Samples are identified by host and dissection number. 25
Pseudobacciger cheneyae n. sp. (Digenea: Gymnophalloidea) from Weber’s chromis (Chromis
weberi Fowler & Bean) (Perciformes: Pomacentridae) at Lizard Island, Great Barrier Reef,
Australia
Figures 12 Pseudobacciger cheneyae n. sp. ex Chromis weberi from off Lizard Island 49
Figure 3 Relationships between Pseudobacciger cheneyae n. sp. and other Gymnophalloidea,
Bucephalidae and Faustulidae taxa based on Bayesian inference and Maximum Likelihood analyses
of the 28S rDNA dataset. 51
Chapter 3: Presence of cleaner wrasse increases the recruitment of fishes to coral reefs
Figure 1 Total abundance of common damselfishes on experimental reefs. 61
Figure 2 Mean (± SE) abundance of recruiting damselfish species. 62
Figure 3 Diversity of all damselfish recruits per reef. 63
Chapter 4: Cleaner wrasse influence habitat selection of young damselfish
Figure 1 Front view of the laboratory experimental tank design. 71
Figure 2 The percentage of time spent on microhabitat adjacent to fish treatment by damselfish
client Pomacentrus amboinensis. 75
Figure 3 Cleaning interactions of juvenile cleaner wrasse, Labroides dimidiatus and damselfishes.
77
Figure 4 The probability of an individual recruit (< 20 mm) being cleaned by a juvenile cleaner
wrasse, Labroides dimidiatus, in relation to the abundance of a) other fish species and b) abundance
of recruits per 20 min observations. 78
Chapter 5: Diel patterns of infection by gnathiid isopods on the ambon damsel, Pomacentrus
amboinensis, and the influence of the presence of cleaner wrasse
Figure 1 Photograph of the sentinel trap used in the experiment. 91
Figure 2 Individual subcages used to hold Pomacentrus amboinensis in the sentinel traps. 92

xiv
List of Tables
Chapter 2: Trematodes of northern Great Barrier Reef Pomacentridae: richness, host-
specificity and beta diversity
Table 1 Host-parasite associations for pomacentrid fishes and digenean trematodes from Lizard
Island. 32
Table 2 Comparison between data collected from Heron Island (Barker et al. 1994) and Lizard
Island (present study) for digeneans from pomacentrid fishes. 38
Table 3 Core pomacentrid specific digeneans from Heron and Lizard Island. 40
Pseudobacciger cheneyae n. sp. (Digenea: Gymnophalloidea) from Weber’s chromis (Chromis
weberi Fowler & Bean) (Perciformes: Pomacentridae) at Lizard Island, Great Barrier Reef,
Australia
Table 1 Collection data and GenBank accession numbers for taxa sequenced during this study 46
Table 2 28S rDNA sequence data from GenBank included in phylogenetic analyses 50
Chapter 4: Cleaner wrasse influence habitat selection of young damselfish
Table 1 Ontogenetic stage and size of fish used in each part for laboratory experiment.
Size is mean ± SE, TL. 71
Chapter 5: Diel patterns of infection by gnathiid isopods on the ambon damsel, Pomacentrus
amboinensis, and the influence of the presence of cleaner wrasse
Table 1 Number of traps (with and with no fish) sampled per sampling day, site, reef pair cleaner
presence treatment, and day and night. Total n per host = 382 traps. 90
Table 2 Number of sentinel traps with gnathiids, day and night trap samples and the number of
gnathiids per trap collected from the sentinel trap experiment on Lizard Island 94

xv
Abbreviations
GBR: Great Barrier Reef
PCO: Principle coordinate analysis
TK-HSD: Tukey-Kramer HSD
GLM: General linear model
LMER: Linear mixed-effect models
GLMER: Generalized linear mixed effect model
LRT: Likelihood ratio test
REML: Restricted maximum likelihood
PERMANOVA: Permutational multivariate analysis of variance
NJ: Neighbour joining
PVC: Polyvinyl chloride
SE: Standard error
TL: Total length
ITS2: Internal transcribed spacer 2
rDNA: Ribosomal Deoxyribonucleic acid
mm: Millimetre
h: Hours
s: Seconds
min: Minute
n: Number
m: Metre

1
Chapter 1: General Introduction

2
Chapter 1: General Introduction
1.1 Preamble
This study explores the complex interactions between three key faunal components on the Great
Barrier Reef (GBR): the parasites (both endo- and ectoparasitics) that infect reef fishes, cleaner
wrasse (Labroides dimidiatus) that feed on, and hence remove parasites from fishes, and
pomacentrids fishes (damselfishes), a large and abundant reef fish family. In these interactions,
pomacentrids are contextualised as both hosts to parasites, and clients to the cleaner wrasse. The
fish and their parasites are abundant and diverse; the parasites remain poorly known. The effects of
the parasites are subtle – we never see a pomacentrid dying because of its parasites – but real. The
cleaner wrasse are keystone species whose parasite-eating behaviour has remarkable implications
for the ecosystems overall. Here I review these components and conclude by outlining the studies
that I have completed to further our understanding of them.
1.2 Pomacentrids
The family Pomacentridae (damselfishes) is one of the most abundant and diverse group of marine
fishes that inhabit coral reefs (Randall et al. 1997). Currently, there are 387 species of pomacentrids
recognised globally of which more than 89 are reported for the GBR (Grutter, in review). They
have been placed into five subfamilies: Abudefdufinae, Chrominae, Lepidozyginae (monospecific),
Pomacentrinae and Stegastinae (Cooper et al. 2009). Damselfishes are known to occupy a wide
range of habitats (e.g. rubble, coral and algae substrates) (Medeiros et al. 2010) and collectively
have a rather broad diet, including planktivory, herbivory and omnivory (Meekan et al. 1995).
These traits most likely have contributed to the success of this family on coral reefs. Damselfishes
are thus an important family of fishes that are a major part of and directly influence the structure of
benthic communities (Hixon and Brostoff 1983; Ceccarelli 2007).
1.3 Parasitism on coral reefs
Parasitism is a non-mutualistic symbiotic relationship and is considered one of the most successful
modes of life given the number of parasite species that exist (Poulin and Morand 2000). All groups
of plants and animals are susceptible to parasitism and many groups have adopted parasitism as a
biological strategy. The abundance of parasitism in the marine environment is demonstrated by the
estimate of Rohde (2002) that there are 100,000 species of protistan and metazoan parasites of
marine fishes globally.

3
Coral reefs are the most diverse and complex marine ecosystems and an important
component of the diversity is comprised of parasites. Multiple parasite groups are abundant on coral
reefs including Protista (multiple phyla), Myxozoa, Monogenea, Digenea, Cestoda, Nematoda,
Acanthocephala, Copepoda, and Isopoda (Justine et al. 2010). Justine et al. (2010) predicted that the
1,700 fish species of New Caledonia harbour 17,000 parasite species in total. For trematode worms,
Cribb et al. (2014b) predicted a total of 1,100 ̶ 1,800 species for the GBR and for monogeneans
Whittington (1998) predicted there is at least one species for each GBR fish species. Despite these
high estimates of richness, it is thought, by all estimates, that only a small fraction of species have
been discovered or described (Cribb et al. 2014b). The parasite fauna of many vertebrate groups
(e.g. many fish families) has not been surveyed. In addition, little is known about the ecology or
ecological role of parasites on coral reefs. Relatively few ecological studies have focused on coral
reef parasites relative to the volume of terrestrial and freshwater studies.
1.4 Parasite diversity on coral reefs
Parasitological studies have demonstrated that parasites are highly abundant and diverse in the
organisms that inhabit coral reefs, especially among coral reef fishes (Justine 2010). Here I focus on
one group, the digenean trematodes. These parasites comprise one of the largest groups of internal
metazoan parasites, comprising approximately 18,000 nominal species worldwide (Cribb et al.
2001). So far, 326 species have been described from the GBR, with a total of 1,1001,800 species
predicted for the region (Cribb et al. 2014b), making digeneans a dominant group of internal
parasites of fishes on the GBR. Although studies examining the trematode fauna of GBR fishes
span more than a century, it has only been in the last three decades that the group has been accorded
detailed attention; the present total of 326 described species known now grew from only 30 species
known in 1988 (Cribb et al. 2014b). The majority of known species of trematodes infect the gut,
with a minority specialized in other organs, e.g. Aporocotylidae in the circulatory system (Smith
2002), Gorgoderidae in the urinary bladder (Ho et al. 2014) and Transversotrematidae under the
scales (Hunter and Cribb 2012). They have complex life cycles that require multiple hosts (Poulin
and Cribb 2002). In the most common life cycle form, infection of a molluscan first intermediate
host by a miracidial larva leads to the asexual production of cercariae, which emerge from the
mollusc and infect either the final host directly (Aprorocotylidae and Transversotrematidae), encyst
in the open (e.g. Haploporidae and Gyliauchenidae), or infect a second intermediate host (e.g.
Cryptogonimidae and Bucephalidae). In the second intermediate host the cercaria develops into a
metacercaria, which has the biological goal of being consumed, with its host by the final host. This
mode of infection is called trophic transmission (Brown et al. 2001).

4
The digenean fauna of GBR fishes has been examined systematically, with many of the
major families of fish having been extensively examined. Understanding the host specificity of
digeneans is a critical aspect in understanding transmission in this group. Host specificity among
parasites has been classified into three categories; oioxenicity, where parasites infect a single host
species (e.g. Lepotrema adlardi known only from Abudefduf bengalensis); stenoxenicity, where
species of parasites infect related host species (e.g. Derogenes pearsoni from damselfishes) and
euryxenicity where parasites infect a wide range of hosts (e.g. Aponurus laguncula known from 67
species from 29 fish families) (Miller et al. 2011b).
Most of the survey effort for digeneans of GBR fishes has been concentrated at two
locations, Heron Island on the southern end of the GBR, and Lizard Island in the north, with the
reefs in between little surveyed. The explanation for this is the presence of dedicated research
stations at the two locations, allowing for systematic collection and processing of samples. The first
fish digeneans to be described from the GBR were two species from teleost fishes (Tetraodontidae)
and two from elasmobranchs (Dasyatidae) by Johnston (1913). Of the 326 currently-known species,
just 66 (20 %) species have been reported from both Heron and Lizard Island. However, many fish
families have been examined more thoroughly at one location than the other. An important example
is the Pomacentridae, which have been extensively surveyed at Heron Island, but not at Lizard
Island.
Studies on pomacentrids of the southern GBR (Cribb et al. 1992; Bray et al. 1993b; Bray et
al. 1993a; Barker et al. 1994; Cribb et al. 1994a; Bray and Cribb 1998) demonstrated a fauna of
seven families and 18 species of adult trematodes. These findings were based on a survey at Heron
Island, comprising 312 individuals from 39 species. For Lizard Island on the northern GBR, where
the present studies were based, nine species of trematodes from five families have been reported
(Bray et al. 1993b; Grutter 1994; Cribb et al. 1998; Grutter et al. 2010; Sun et al. 2012), two of
which (Lepotrema oyabitcha and Transversotrema damsella) are not known from Heron Island
(Bray et al. 1993b; Hunter and Cribb 2012). Despite the recording of nine species at Lizard Island,
the fauna there has not been studied systematically and the limited data available suggested that
there are significant differences between the northern and southern GBR. The extent to which the
parasite fauna of the northern and southern GBR is shared by and between fish species is poorly
understood. There has been no systematic study of matched sets of statistically informative samples
of the same species of fishes between the two sites.

5
1.5 Ecological importance of parasites
The importance of parasites to fish health is a complex issue. Although a high proportion of
individual fish everywhere are infected with parasites, pathogenic effects are rarely immediately
evident and are only rarely studied in wild fish. It has been speculated that many parasitic infections
are relatively benign (Candolin and Voigt 2001; Carrassón and Cribb 2014). When closely studied,
however, fish parasites have often been shown to have a variety of negative effects on their host.
They have been shown to affect critical life factors such as host growth rate (Adlard and Lester
1994; Jones and Grutter 2008), body condition (Lemly and Esch 1984), reproduction (Fogelman et
al. 2009; Mensink et al. 2014), population dynamics (Scott and Dobson 1989), and behaviour
(Barber et al. 2000; Roche et al. 2013b).
For coral reef fishes, most parasite related research has been heavily biased towards
diversity studies of the parasites of adult coral reef fishes (Cribb et al. 1994b; Grutter 1994; Morand
et al. 2000) and little is known about the ecological role of most parasites. Few studies have
examined the ecology of internal parasites on coral reef fish (Muñoz et al. 2006; Muñoz et al.
2007). Studies by Grutter et al. (2010) and Sun et al. (2012) showed that there was a difference in
parasite (mostly worms) prevalence with age group in the young damselfishes. These studies also
demonstrated that the presence of internal parasites may have an effect on the growth rate of an
individual at or during settlement. One possible explanation for the limited amount of study of the
effects of internal parasite on the ecology and behaviour of a host is that they are hidden within a
host. There has been more work done on the ecology of ectoparasites, including copepods (e.g.
Finley and Forrester 2003) and especially cymothoids and gnathiids. Adlard and Lester (1994)
showed that barrier reef chromis, Chromis nitida, that were infected with the cymothoid isopod
Anilocra pomacentri had lower growth rate and higher mortality than non-parasitised fish. Mortality
was caused by several factors, including erratic swimming behaviour upon parasitic attachment
leading to increased susceptibility to predation, and gross pathology leading to deterioration in body
condition, particularly in juvenile fishes. Östlund-Nilsson et al. (2005) recorded higher oxygen
consumption, an increase in pectoral fin beat frequency and reduced swimming speed for an
apogonid fish parasitised with Anilocra apogonae. Fogelman and Grutter (2008) showed that three
apogonid species infected by A. apogonae had reduced growth and survival.
Gnathiid isopods are arguably the most ecologically important and certainly the most
carefully studied of the ectoparasites of coral reef fishes. They are abundant in both temperate and
tropical waters (Sikkel et al. 2009) and are common on coral reef fishes (Grutter 1994). They do not
possess a pelagic phase like most other marine invertebrates (Smit and Davies 2004). The life cycle
incorporates three juvenile (larval) stages - three zuphea stages (unfed phases), three praniza stages
(fed phases) and the non-feeding adult (Smit and Davies 2004). These mobile ectoparasites emerge

6
from the benthos as the zuphea phase and locate a host fish. Once engorged on blood and body
fluids, as the praniza phase, they return to the benthos and moult into the next larval phase.
Gnathiids thus exhibit protelian parasitism - parasitic as juveniles and free-living as adults (Smit
and Davies 2004). Unlike other ectoparasites such as copepods and isopods which may be
permanent parasites, gnathiids isopods are temporary parasites and are also often referred to as
micropredators (Lafferty and Kuris 2002).
The mouthparts of gnathiids are highly modified enabling them to attach and feed on the
surfaces of fishes (Monod 1926). Gnathiids attach themselves to the body, fins, eye rims, gill
chambers, nares, lips and the buccal cavity of their fish host (Monod 1926). In feeding on the blood
and tissue fluids from their hosts they can cause either tissue damage or, when present in large
numbers, death (Marino et al. 2004; Hayes et al. 2011). The amount of time required for gnathiids
to become fully engorged varies among both species and developmental stages, with feeding period
being longer for larger species and later juvenile stages (Grutter 2003; Smit et al. 2003; Smit and
Davies 2004). Gnathiids can become engorged in less than 10 minutes (Grutter 2003) but can attach
for up to 10 h (Grutter 2003; Smit et al. 2003; Grutter et al. 2011).
Previous studies on the ecology of gnathiids have demonstrated their importance on coral
reefs. They can have a wide range of effects on adult fish. They can cause tissue damage (Heupel
and Bennett 1999) and gill mucus shedding (Hayes et al. 2011), reduce haematocrit value (Jones
and Grutter 2005), are probable vectors of haemogregarine blood parasites (Davies and Smit 2001;
Davies et al. 2004; Curtis et al. 2013) and viral diseases (Davies et al. 2009), and can even cause
mortality due to blood loss when numbers are high (Mugridge and Stallybrass 1983; Hayes et al.
2011). Infection by gnathiids also influences the behavioural interactions between cleaners and their
clients (Grutter 2001; Sikkel et al. 2004).
In contrast to the relatively detailed knowledge of the role/importance of gnathiids in adult
fish biology, the nature of the interaction between gnathiids and young fish is poorly known. What
is already known about gnathiids on adult fishes (i.e. prevalence, and feeding ecology) (Grutter
1995a,1996a,2003; Grutter 2008) suggests that they may also significantly affect young reef fish.
However, only a handful of studies have investigated their effects on young reef fish. Jones and
Grutter (2008) found that settling Dischistodus perspicillatus damselfish infected with three
gnathiids each night for one week had slower growth, measured by a percent increase in size, than
those infected with one or none. Grutter et al. (2008) recorded mortality (12 % and 16 %) for
settling Neopomacentrus azysron damselfish infected with one or three gnathiids, respectively.
Penfold et al. (2008) found that in the laboratory, gnathiid isopods killed 6.3 % of juvenile
Acanthochromis polyacanthus damselfish that were < 10 mm SL compared with 0 % for uninfected
individuals. Grutter et al. (2011) showed that settling P. amboinensis previously infected with a

7
gnathiid swam more slowly, had higher oxygen consumption, and had reduced survival relative to
uninfected individuals. In the wild, 3.5 % of a population of juvenile P. amboinensis caught at dawn
were infected with a gnathiid and in the laboratory the infection was found to last for up to 10 h
(Grutter et al. 2011). These studies highlight the potential importance of gnathiids in the early life
stages of coral reef fishes and how they may affect successful recruitment to the adult population.
Although numerous studies have examined the emergence of gnathiids in both the
Caribbean and on the GBR in regards to diurnal variation (Jacoby and Greenwood 1988; Grutter
and Hendrikz 1999; Sikkel et al. 2006; Jones and Grutter 2007), little is known as to whether the
presence of cleaner wrasse has a significant effect on the number of gnathiids on reef fish.
Estimating the number of gnathiids on a fish is complicated. As gnathiids are temporary parasites,
they are able to detach themselves from the host when it is disturbed (e.g. host is captured) or can
be eaten by a cleaner fish before the host has been examined for gnathiids. Therefore, studies that
have examined the number of gnathiids in relation to cleaner fish presence may have
underestimated the number of gnathiids on a host. Previous studies that have examined the number
of gnathiids on reef fishes have used sampling methods that may also have overlooked the possible
interaction of cleaner fish and the behaviour of gnathiids. Grutter (1999a) showed that individual
caged thicklip wrasse, Hemigymnus melapterus, had a 4.5-fold reduction in the number of gnathiid
isopods on individuals when on reefs with a cleaner wrasse relative to those on reefs without.
Cheney and Côté (2003) showed that longfin damselfish, Stegastes diencaeus, that were captured
from territories that had access to a cleaning station had lower numbers of gnathiids than
individuals that did not. Although these studies demonstrated that the presence of cleaner fish can
affect the number of gnathiid isopods on reef fish, the findings only provide information in regards
to the gnathiid load on the fish at a given time (time of host captured) and do not take into
consideration the issues raised above in regards to the potential loss of gnathiids.
Sikkel et al. (2011) reported a trap design that is able to quantify the cumulative number of
gnathiids on a host over time. These ‘sentinel’ traps use live ‘bait’ fish to attract gnathiids that are
seeking a host. However, unlike wire cages which have been used in the past (e.g. Grutter 1999a;
Grutter 1999b) from which gnathiids can escape if disturbed or fed, these traps retain gnathiids once
they have entered the trap. The traps also prevent any interaction between cleaner fish and the live
fish inside the traps, thus preventing loss of gnathiids due to removal by cleaner fish. These traps
thus have the capacity to provide an accurate representation of the cumulative number of gnathiids
attracted to a fish over a period of time.

8
1.6 Cleaning mutualism
Positive interspecific interactions are encounters between organisms that occur where at least one
individual benefits while the other is not harmed (Bronstein 1994). Such interactions have been
classified as either mutualism or facilitation/commensalism, with the outcome of the interaction
often creating a more favourable environment, either directly or indirectly, for one or multiple
species (Stachowicz 2001; Bronstein et al. 2004). Mutualistic interactions have been shown to be of
great ecological importance within terrestrial (Bascompte and Jordano 2007), freshwater (Brown et
al. 2012) and marine ecosystems (Knowlton and Rohwer 2003). Some mutualistic relationships can
extend beyond the two species involved in the initial interaction to other species within the
community, and sometimes the overall structure and function of ecosystems (Stachowicz 2001).
Some organisms within mutualistic relationships can have a disproportionate effect on the
success of other species and are known as “keystone” mutualists (Bond 1994). The presence of
keystone mutualists may be pivotal in shaping the structure of communities. Keystone mutualists
are known in both terrestrial (Harms et al. 2000) and aquatic environments (Caley et al. 1996). This
is seen, for example, in the relationship between frugivores (notably mammals and birds) and plants
via seed dispersal, which can lead to enhanced biodiversity of plants (Jordano 2000). On coral reefs,
the bluestreak cleaner wrasse, Labroides dimidiatus, is considered a key stone mutualist because
their presence affects coral reef fish biodiversity (Bshary 2003; Grutter et al. 2003; Waldie et al.
2011) through cleaning symbioses.
Cleaning symbiosis is the removal and subsequent ingestion of ectoparasites, and diseased
and injured tissue by cleaning organisms (Losey 1972; Côté 2000); it occurs in both the terrestrial
and aquatic environment (Poulin and Grutter 1996; Weeks 2000; Cheney and Côté 2001; Becker et
al. 2005; Biani et al. 2009). It has long been held that cleaning symbioses in the marine
environment are mutualistic interactions in which cleaners benefit from the removal of parasites and
tissue from their fish clients (Grutter 1996a; Arnal and Côté 2000; Arnal and Morand 2001) and
clients benefit from a reduced ectoparasite load (Losey 1979; Gorlick et al. 1987; Grutter 1999b). In
the marine environment cleaning behaviour is performed by narrow range of fish and crustaceans
(see list in Poulin and Grutter 1996). Typically cleaners have defined territories, ‘cleaning stations’,
which are visited by organisms, the ‘clients’ (Côté 2000). There is strong evidence to show that
clients do obtain net benefits from the interaction (Bshary et al. 2007; Clague et al. 2011; Ros et al.
2011; Soares et al. 2011). Several studies have shown that the presence of cleaner fish affects the
size and abundance of crustacean parasites on fish (Gorlick et al. 1987; Grutter 1999a; Cheney and
Côté 2001; Grutter and Lester 2002; Clague et al. 2011), increases the size distribution of fish
(Clague et al. 2011; Waldie et al. 2011), and affects the composition of client fish communities
(Bshary 2003; Grutter et al. 2003; Waldie et al. 2011).

9
Labroides dimidiatus is the most common cleaner fish species on the GBR (Randall et al.
1997). At Lizard Island, some individual client fish visit a cleaning station up to 144 times a day
(Grutter 1995a). Individual cleaner wrasses have been shown to consume an average of 1,218 ± 118
parasites, mostly gnathiids, from 2,297 ± 83 fish inspected each day (Grutter 1996a). These studies
suggest that cleaner wrasse presence plays an important role in reducing the number of parasites,
which may ultimately lead to fish choosing to be near a cleaner fish.
On patch reefs around Lizard Island, GBR, the species richness and abundance of visitors
(fish species that move between patch reefs) were reduced when L. dimidiatus individuals were
removed over an 18-month period (Grutter et al. 2003). After 8.5 years of maintaining these same
reefs free of L. dimidiatus, Waldie et al. (2011) found a 23 % and 37 % reduction in species
richness and abundance of all resident fishes (species that are site-attached) relative to reefs that had
cleaners. Furthermore, there was a 66 % reduction in the abundance of juvenile (< 70 mm TL)
visitor fishes on reefs without L. dimidiatus. This reduction in juvenile abundance could explain the
23 % lower abundance and 33 % lower species richness of visitor fishes on these reefs. Although
the benefits of L. dimidiatus are thus well documented for adults and/or visiting reef fishes, its
importance in settlement processes is largely unknown.
The ecological determinants of settlement choice for pelagic coral reef fish larvae are
complex, but dramatically affect population dynamics and the size of future adult populations
(Hoey and McCormick 2004; Heinlein et al. 2010). Selecting an optimum habitat can maximise the
growth and survival of coral reef fish during and after settlement (McCormick and Hoey 2004;
Lecchini et al. 2007a). Factors affecting microhabitat choice include water temperature (Grorud-
Colvert and Sponaugle 2011; Lienart et al. 2014), habitat structure (Feary et al. 2007; Coker et al.
2012b), coral type (Jones 1988; Danilowicz 1996), position on the reef (Srinivasan 2003; Jordan et
al. 2012), and presence of conspecifics (Ohman et al. 1998; Adam 2010). There has been only
limited research on the significance of the presence of heterospecifics in influencing the
microhabitat choice of young reef fish.
A few studies have demonstrated a positive or negative correlation between presence of
heterospecifics and fish settlement. The abundance of some wrasses (Labridae) is highest in the
algal territories of herbivorous damselfishes, possibly due to increased algal cover providing shelter
and food resources at settlement (Green 1994; Green 1998), suggesting correlation rather cause.
Resident piscivorous groupers and moray eels inhibit levels of settlement of damselfish and
acanthurids, but increase the settlement of a labrid, Thalassoma bifasciatum (Almany 2003). As
mortality from predation is often low for cleaners (Côté 2000; Grutter 2004b), it was hypothesised
that juvenile T. bifasciatum may be immune to piscivory due to the species being a cleaner (Almany
2003). In contrast, the presence of two resident damselfishes in the absence of a resident piscivore

10
increases the abundance of acanthurid settlers (Almany 2003). Given the high fish diversity of coral
reefs (Allen 2008), the potential for complex interactions that influence settlement processes is
considerable.
The importance of gnathiids and other external parasites to fish health has also been
demonstrated indirectly by studies such as that of Grutter et al. (2003), which showed a positive
association between cleaner wrasse presence and fish richness and abundance. This relationship
suggested that mobile reef fish actively seek cleaner wrasses in order to have gnathiids and other
parasites removed. Clague et al. (2011) showed that on reefs with cleaner fish, older P. moluccensis
were larger at a certain age than those from reefs with no cleaner fish. They also found that copepod
abundance was higher on fish from reefs with no cleaner fish, but only for larger host fish.
Gnathiids have also been demonstrated to be of importance during settlement, in that even a single
gnathiid infection can decrease the settlement success of P. amboinensis larvae (Grutter et al. 2011).
Thus, if gnathiids are not removed from fish by cleaner fish or gnathiid population densities are
higher on reefs without cleaner fish, there could be a reduction in juvenile fish abundance. It would
thus be an advantage for young reef fish to settle near cleaning stations. However, the role of
cleaning behaviour with respect to influencing the settlement behaviour of juvenile fish remains
relatively unstudied.
As cleaner fish are known to reduce their clients’ ectoparasite loads, proximity to a cleaner
station should be advantageous for resident fish species. Waldie et al. (2011) found a 66 % increase
in juveniles of visiting fish species on reefs associated with a cleaner fish relative to those without
them. However, no study has examined the effect of cleaner wrasse presence on the juveniles of
resident fish species, most of which consist of damselfishes. Previous studies have shown that the
ability to locate suitable habitats during settlement can have significant advantages in terms of
enhancing growth and/or increasing survival of young reef fish (Bonin et al. 2011; Vail and
McCormick 2011; Rankin and Sponaugle 2014). Young reef fish have highly developed visual
(Wenger et al. 2011; Lecchini et al. 2014), olfactory (McCormick et al. 2010; Paris et al. 2013) and
auditory senses (Radford et al. 2011; Huijbers et al. 2012), which are known to play a critical role
when selecting suitable habitats. The humbug damselfish Dascyllus aruanus has been demonstrated
to use olfactory cues released by conspecifics in selecting reefs to settle on (Sweatman 1988). Vail
and McCormick (2011) demonstrated that settling damselfishes use olfaction to recognise predators
and thus avoid settling onto habitats where predators are present.
At present, it is not known whether young reef fish that are found near a cleaning station
have selected these reefs by using visual or other cues. It is known that naïve larvae of Hawaiian
humbug Dascyllus albisella, raised in captivity innately recognise the Hawaiian cleaner wrasse,
Labroides phthirophagus (Losey et al. 1995). That study demonstrated that experience with a

11
cleaner fish is not required for D. albisella to recognise a cleaner fish and to adopt the client ‘pose’.
Although this study showed that young reef fish recognise cleaner fish, no studies have tested
whether cleaner fish presence plays a role in larval habitat choice. A similar mechanism may occur
with young reef fish that are approaching or are making the transition to the benthos, in that they
may partly select reefs based on cues related, directly or indirectly, to cleaner fish presence. As
most coral reef fish (especially damselfishes) are relatively site attached and will not leave their
territory to seek a cleaner fish (Randall et al. 1997), any selection would have to occur while they
are in the process of settling onto the reef or soon after as post-settlement movement (Lewis 1997;
Simpson et al. 2008).
There are other potential benefits besides a reduction in parasite loads for fish that settle in
the vicinity of a cleaner wrasse. Cheney et al. (2008) showed that the presence of a cleaner wrasse
reduces the frequency of aggressive chases by a piscivore towards potential prey species. This
reduction in aggression by predators at cleaning stations may indirectly benefit prey species, by
reducing the risk of being consumed and influencing how long they may remain at a cleaning
station when a predator is present. No doubt such an indirect effect of cleaner fish presence would
benefit such vulnerable young reef fish during settlement, as predation is the major cause of
mortality at that period.
1.7 Aims and thesis structure
The broad aim of this study was to improve understanding of a range of parasite-related issues from
the richness of digenean trematodes to the importance of cleaning behaviour in damselfishes. The
thesis thus explores a series of interrelated topics relating to the parasites of damselfishes and their
ecological implications: (1) the richness and prevalence of digenean trematodes in damselfishes
from the northern GBR, Lizard Island; (2) the effects of cleaner wrasse presence on the abundance
of recruiting damselfishes; (3) the effect of cleaner wrasse presence in a microhabitat choice of a
damselfish; and (4) the cumulative number of gnathiid isopods infecting a species of damselfish
relative to cleaner wrasse presence. These topics are explored through both field and laboratory-
based experiments. The thesis is presented as a collection of chapters written in a style that is
suitable for publication. The thesis consists of six chapters of which four (2 ̶ 5) are in a manuscript
format.
Chapter 2. Trematodes of northern Great Barrier Reef Pomacentridae: richness, host-
specificity and beta diversity

12
There are 326 species of trematodes reported so far for the GBR (Cribb et al. 2014b) including 18
species known from damselfishes. Although damselfishes (Pomacentridae) are a major component
of the coral reef fish assemblage, and much is known in regards to their ecology and biology, the
trematode fauna of damselfishes on the northern GBR, has not been examined systematically.
Current knowledge regarding the trematode fauna of damselfishes is mainly from systematic
surveys conducted on Heron Island, southern GBR (Cribb et al. 1992; Bray et al. 1993a; Bray et al.
1993b; Barker et al. 1994). In contrast to damselfishes, other families of fishes have been
extensively surveyed for trematodes at both Heron and Lizard Island - chaetodontids (Bray et al.
1994; McNamara et al. 2012; Diaz et al. 2013), lutjanids (Miller and Cribb 2007; Miller and Cribb
2008) and serranids (Bott and Cribb 2009; Bott et al. 2013). An extensive survey of damselfish
trematodes from Lizard Island, far north GBR formed the first component of my PhD research. This
study led to the description of one new species, Pseudobacciger cheneyae, published as separate
standalone paper (incorporated in chapter 2).
Chapter 3. Presence of cleaner wrasse increases the recruitment of fishes to coral reefs. Although the direct and indirect effects of cleaner fish have been demonstrated for adult visitor reef
fish, they have yet to be examined for resident larval reef fishes. Clague et al. (2011) showed that,
on reefs with cleaner fish, older Pomacentrus moluccensis were larger at a certain age than those
from reefs without cleaner fish. Gnathiids have been demonstrated to be important during
settlement, in that a single gnathiid can decrease the success of settlement of P. amboinensis larvae
significantly (Grutter et al. 2011). Therefore, if gnathiids are not removed from fish by cleaner fish
(direct affect) or gnathiid population densities are higher on reefs without cleaner fish (indirect
affect), there could be a reduction in juvenile fish abundance. Cleaner presence could also affect
larval fish growth or survival through modification of the behaviour of predatory fish and thus the
risk of predation, as there is evidence that predators reduce aggression towards bystanders in the
presence of a cleaner wrasse (Cheney et al. 2008). I examined the effects of cleaner fish presence on
juvenile density using patch reefs that have been experimentally manipulated for 12 years by being
kept free of cleaner wrasses (Clague et al. 2011; Waldie et al. 2011).
Chapter 4. Cleaner wrasse influence habitat selection of young damselfish.
The ability to locate suitable habitats during settlement can confer significant advantages on young
fish by enhancing growth and/or increasing survival (McCormick and Hoey 2004). Young reef fish
have highly developed visual and olfactory senses (Lecchini et al. 2007b) and these play a critical
role in selecting suitable habitats. Naïve captive-raised larval Hawaiian humbug, Dascyllus
albisella, innately recognise Hawaiian cleaner wrasse, Labroides phthirophagus (Losey et al. 1995).

13
Although that study showed that reef fish larvae recognise cleaner fish, no studies have
demonstrated a link between cleaner fish presence and larval microhabitat choice. Parasites such as
gnathiids have already been shown to decrease settlement success of a fish, thus I hypothesise that it
would be an advantage for young reef fish to settle near cleaning stations. I tested whether young
reef fish preferentially choose a microhabitat that is adjacent to a cleaner fish and whether young
fish found at cleaning stations receive direct benefits in the form of cleaning services. If young fish
actively select a microhabitat that is near a cleaner fish, this selection may help increase their
survival after settlement.
Chapter 5. Diel patterns of infection by gnathiid isopods on the ambon damsel, Pomacentrus
amboinensis, and the influence of the presence of cleaner wrasse.
Gnathiid isopods are the most common temporary ectoparasites of coral reef fishes (Grutter and
Poulin 1998), and are the key food of cleaner wrasse (Grutter 1997). The presence of cleaner fish is
known to reduce the number of gnathiids on reef fish (Grutter 1999a; Cheney and Côté 2001) which
could explain the higher abundances of fishes on reefs where cleaner wrasse are present (Grutter et
al. 2003; Waldie et al. 2011) and also the abundance of recruiting fishes. I hypothesise that
predation of gnathiids by cleaner wrasse leads to a reduction in the overall population of gnathiids
on reefs with cleaners relative to those without. Although previous studies have examined the
number of gnathiids on reef fishes (Grutter 1994; Grutter 1999a; Cheney and Côté 2001; Grutter et
al. 2011), these studies only characterise the number of gnathiids on an individual at a specific time
point. As a result, the rate at which gnathiids infect reef fish, and whether that rate changes over the
course of a day, is not understood. Fish might be under surveyed due to gnathiids either dropping
off due to being fully fed or having been eaten by a cleaner fish. In this study I used sentinel traps
that retain gnathiids once they drop off and prevent physical interaction between cleaner wrasse and
client. It is possible that individuals that are vulnerable to gnathiid infection such as young fish are
more commonly infected with gnathiids than has been shown previously. By collecting samples
throughout the day and night (24 hours), I explored the overall cumulative number of gnathiids and
also examined of how it varies with respect to cleaner wrasse presence.
Chapter 6. General Discussion
This chapter integrates the major findings of each study and makes recommendations for future
research.

14
Chapter 2: Trematodes of northern Great Barrier Reef
Pomacentridae: richness, host-specificity and beta
diversity

15
Chapter 2: Trematodes of northern Great Barrier Reef Pomacentridae:
richness, host-specificity and beta diversity
Derek Sun, Rodney A. Bray, Russell Q-Y. Yong, Scott C. Cutmore & Thomas H. Cribb
Abstract
Species richness and host specificity of the digenean fauna of the family Pomacentridae was studied
by examining 358 individuals of 32 species of pomacentrids from Lizard Island, northern Great
Barrier Reef (GBR). In total, 19 clear morphotype species of digeneans, 54 host/parasite
combinations, 19 new host distributions, 18 new host records and three new species,
Fellodistomidae n. sp., Gyliauchenidae n. sp., and Pseudobacciger cheneyae Sun, Bray, Yong,
Cutmore & Cribb, 2014, were recorded from this large and important group of coral reef fishes.
Combined morphological and molecular analysis of specimens initially identified as Hysterolecitha
nahaensis suggested that on the basis of morphology it is possible to recognise three species and
from molecular analysis as many as seven. The two sources of data could not be completely
reconciled and the system requires further work. For the purpose of this analysis all such forms are
identified as “H. nahaensis complex”. Further examination of the host-parasite relationship showed
that host specificity is relatively low; only three species are apparently oioxenic whereas the
majority possessed either stenoxenic or euryxenic host specificity. Overall, a total of 23 species of
digenean trematodes have now been reported in pomacentrids from the GBR of which 13 are
present at both Lizard and Heron Island. Five and four species have been reported only from Lizard
and Heron Island, respectively. Lastly, 14 of the 23 species have been reported from only
pomacentrids. The richness that was recorded is similar to that reported previously at Heron Island.
This study suggests that the diversity of pomacentrids species far exceeds that of their digenean
parasites at Lizard Island.
Introduction
Damselfishes (Pomacentridae) are one of the most diverse families of marine fishes occurring on
coral reefs (Randall et al. 1997). Currently, there are 387 species of pomacentrids recognised
globally in both marine and brackish systems (Froese and Pauly 2012). The family is divided into

16
five subfamilies: Abudefdufinae, Chrominae, Lepidozyginae (monospecific), Pomacentrinae and
Stegastinae (Cooper et al. 2009); all five subfamilies are represented on the GBR and more than 80
species are known in the region (Randall et al. 1997).
Habitat selection and a wide diversity of diet preferences of damselfishes may have aided
diversification of this family on coral reefs. The large diversity within this group has allowed
damselfishes to occupy and exploit a range of microhabitats such as living corals (Wilson et al.
2008), rubble/dead coral (Bay et al. 2001), and algae-dominated areas (Medeiros et al. 2010), as
well as exploiting both benthic and pelagic niches (Randall et al. 1997). Damselfishes can be
divided into three major trophic guilds: planktivores, herbivores and omnivores (Cooper et al.
2009); the scope for exploitation by trophically transmitted parasites, including trematodes, in this
family is thus substantial.
Host specificity is an important aspect of the biology of parasites (Adamson and Caira
1994). Host specificity of parasites can be classified into three broad categories: oioxenicity, where
an individual parasite species is known to only infect a single host species; stenoxenicity, where the
parasite infects multiple phylogenetically related species; and euryxenicity, where a species infects
a diverse range of hosts united by ecology and physiology rather than taxonomy (Miller et al.
2011b). Miller et al. (2011b) showed that 60 % of the trematodes then known from the GBR were
oioxenic, 32 % were stenoxenic and just 8 % were euryxenic. For the Pomacentridae specifically,
Barker et al. (1994) found that, of the 18 species of digeneans known from pomacentrids at Heron
Island, six [Bivesicula unexpecta Cribb, Bray & Barker, 1994, Haplosplanchnidae sp., Lepotrema
adlardi (Bray, Cribb & Barker, 1993), Lepotrema sp. 1, Lepotrema sp. 2 and Steganoderma gibsoni
Cribb, Bray &Barker, 1992] were oioxenic, three (Derogenes pearsoni Bray, Cribb & Barker, 1993,
D. pharyngicola Bray, Cribb & Barker, 1993 and Hysterolecitha heronensis Bray, Cribb & Barker,
1993) were strictly stenoxenic, another two [Hysterolecitha nahaensis Yamaguti, 1942 and
Schikhobalotrema pomacentri (Manter, 1937)] were predominantly stenoxenic (i.e. primarily found
in one group of closely-related hosts, but occasionally found in hosts from unrelated families) and
seven [Aponurus laguncula Loos, 1907, Lepotrema clavatum (Ozaki, 1932), Lecithaster stellatus
Looss, 1907, Macvicaria heronensis Bray & Cribb, 1989, Preptetos cannoni Barker, Bray & Cribb,
1993, P. xesuri (Yamaguti, 1940) Pritchard, 1960 and Thulinia microrchis (Yamaguti, 1934) Bray,
Cribb & Barker, 1993] were euryxenic. The host range thus varied greatly between species, with
one, Hysterolecitha nahaensis, found in 24 pomacentrid species.
Although damselfishes have been the focus of many ecological studies in reef fishes their
parasitic fauna remains incompletely known. The first dedicated investigation into damselfish
digenean fauna was conducted at Heron Island in the early 1990s. A total of 39 pomacentrid species
was examined, resulting in the discovery of 18 species of trematodes and the description of 8 new

17
species (Cribb et al. 1992; Bray et al. 1993b; Bray et al. 1993a; Bray and Cribb 1998). Relatively
few studies have been conducted elsewhere on the GBR; of these, work was primarily conducted at
Lizard Island. However, unlike at Heron Island, these studies focused on individual species of
damselfishes or a particular family of digenean trematode (Grutter et al. 2010; Hunter and Cribb
2010; Sun et al. 2012; Sun et al. 2014). Thus, the trematode fauna of damselfishes at other locations
on the GBR, including Lizard Island, can be considered to be under-studied. Studies conducted
elsewhere in the world (e.g. Yeo and Spieler 1980; Dyer et al. 1985) have demonstrated a relatively
depauperate digenean fauna in damselfishes.
This is the first study to have systematically examined the diversity and host specificity of
digenean trematodes in the family Pomacentridae from Lizard Island in the northern Great Barrier
Reef and compared its nature relative to the existing known fauna at Heron Island reef 1,184 km to
the south.
Materials and methods
Specimen collection
Between 2011 and 2012, pomacentrids were collected from Lizard Island (1440S, 14528E),
Great Barrier Reef, Australia. These fish were collected using a barrier net, spear gun or clove oil,
and were returned to the research station and euthanized prior to dissection via cranial pithing. As
transversotrematids live beneath the scales of fish the body was soaked in vertebrate saline for a
minimum of 10 minutes. This allowed any transversotrematids to fall to the bottom of the container.
The heart was removed from the body and examined carefully for aporocotylids. The contents of
the gut cavity were removed and examined under a dissecting microscope using fine scissors and
forceps. A “gut wash” was conducted in an attempt to dislodge parasites attached to the gut wall
(Cribb and Bray 2010). Specimens were killed and heat-fixed by pipetting the worms into 0.85 %
saline just off the boil and transferred to Eppendorf tubes containing 70 % absolute ethanol (Cribb
and Bray 2010).
Morphological analysis
Preserved worms were stained using Mayer’s haematoxylin, destained in 1 % hydrochloric acid,
neutralised with 1 % ammonia solution, dehydrated through a graded series of ethanols (50, 70, 80,
90 and 100 %), cleared in methyl salicylate and then mounted on slides in Canada balsam. All
measurements were made using a digital camera and the programme SPOT Advanced (version 4.6).
Voucher specimens are lodged within the Queensland Museum.

18
Molecular analysis
All voucher specimens are paragenophores (sensu Pleijel et al. 2008). All vouchers are
lodged in the Queensland Museum, Brisbane, Australia. Total genomic DNA was extracted using
standard phenol/chloroform extraction techniques (Sambrook and Russell 2001). The D1-D3
regions of 28S nuclear ribosomal DNA was amplified using the primers LSU5 (5'-TAG GTC GAC
CCG CTG AAY TTA AGC-3') (Littlewood et al. 2000) and ECD2 (5'-CTT GGT CCG TGT TTC
AAG ACG GG-3') (Littlewood et al. 2000) and the ITS2 region using the primers 3S (5'-GGT ACC
GGT GGA TCA CGT GGC TAG TG-3') (Bowles et al. 1993) and ITS2.2 (5'-CCT GGT TAG TTT
CTT TTC CTC CGC-3' ) (Cribb et al. 1998) PCR for both the 28S and ITS2 regions was performed
with a total volume of 20 μl consisting of approximately 10 ng of DNA, 5μl of 5x MyTaq Reaction
Buffer (Bioline), 0.75 μl of each primer (10 pmols) and 0.25 μl of Taq DNA polymerase
(BiolineMyTaq™ DNA Polymerase), made up to 20 μl with Invitrogen™ ultraPURE™ distilled
water. Amplification was carried out on a MJ Research PTC-150 thermocycler. The following
profile was used to amplify the 28S region: an initial 95°C denaturation for 4 min, followed by 30
cycles of 95°C denaturation for 1 min, 56°C annealing for 1 min, 72°C extension for 2 min,
followed by a single cycle of 95°C denaturation for 1 min, 55°C annealing for 45 sec and a final
72°C extension for 4 min. The following profile was used to amplify the ITS2 region: an initial
single cycle of 95°C denaturation for 3 min, 45°C annealing for 2 min, 72°C extension for 90 sec,
followed by 4 cycles of 95°C denaturation for 45 sec, 50°C annealing for 45 sec, 72°C extension for
90 sec, followed by 30 cycles of 95°C denaturation for 20 sec, 52°C annealing for 20 sec, 72°C
extension for 90 sec, followed by a final 72°C extension for 5 min. Amplified DNA was purified
using a BiolineISOLATE II PCR and Gel Kit, according to the manufacturer’s protocol. Cycle
sequencing of purified DNA was carried out for the ITS2 rDNA region using the forward primer
GA1 (5'-AGA ACA TCG ACA TCT TGA AC-3') (Anderson and Barker 1998) and the reverse
primer ITS2.2 and for the 28S rDNA region using the same primers used for PCR amplification as
well as the additional primer 300F (5'-CAA GTA CCG TGA GGG AAA GTT-3')(Littlewood et al.
2000). Cycle sequencing was carried out at the Australian Genome Research Facility using an
AB3730xl capillary sequencer. Sequencher™ version 4.5 (GeneCodes Corp.) was used to assemble
and edit contiguous sequences. Sequences were aligned using Muscle implemented with MEGA
5.2. Differences between sequences were explored by creating distance matrices and neighbour
joining (NJ) trees.

19
Results
A total of 358 individual fish were examined from 32 species of pomacentrids from Lizard Island.
In total, 19 clear morphotype species of digeneans, 54 host/parasite combinations, 18 new host
records, 19 new host parasite/locality records and three new species were recorded (see Table 1).
Below, the details of observations relating to the 19 species are summarised.
Bivesiculidae Yamaguti, 1934
Bivesicula unexpecta Cribb, Bray & Barker, 1994
Host: Acanthochromis polyacanthus (Bleeker, 1855).
Locality: Lizard Island
Voucher specimens: QM XXXXX
Remarks: This species was described from Acanthochromis polyacanthus by Cribb et al. (1994a) at
Heron Island. Sequence data for this species from the same host species was subsequently reported
for both Heron and Lizard Islands (Cribb et al. 1998). The present specimens are entirely consistent
with the original description of this species. Bivesicula unexpecta remains the only bivesiculid
species to have been reported as an adult from a pomacentrid and A. polyacanthus remains the only
known host species. Immature bivesiculids were found in several other pomacentrids in this study,
but these were all consistent with Bivesicula claviformis Yamaguti, 1934, a species that matures in
serranid fishes and has immature stages in the intestine of a range of fishes preyed on by serranids
(Cribb et al. 1998).
Derogenidae Nicoll, 1910
Derogenes pearsoni Bray, Cribb & Barker, 1993
Hosts: Chrysiptera cyanea (Quoy & Gaimard, 1825), Pomacentrus adelus Allen, 1991, Stegastes
apicalis (De Vis, 1885).
Locality: Lizard Island
Voucher specimens: QM XXXXX
Remarks: Our specimens of this species match those reported by Bray et al. (1993a) from the
damselfishes Amblyglyphidodon curacao (Bloch, 1787), Amphiprion akindynos Allen, 1972,
Plectroglyphidodon lacrymatus (Quoy & Gaimard, 1985), Pomacentrus chrysurus Cuvier, 1830, P.
moluccensis Bleeker, 1853, P. tripunctatus Cuvier, 1830, and an unidentified species of
Pomacentrus sp. from Heron Island, in the southern GBR. Sun et al. (2012) reported D. pearsoni
from Pomacentrus amboinensis Bleeker, 1868 from Lizard Island. The three hosts reported here
represent new host records. Derogenes pearsoni differs from the other species of Derogenes found
in GBR pomacentrids, D. pharyngicola Bray, Cribb & Barker, 1993, in having shorter caeca, the

20
structure of the sinus-sac, the location of the posterior genital pore and the development of the
uterus in the forebody (Bray et al. 1993a). Derogenes pharyngicola was not detected in the present
study.
Faustulidae Poche, 1926
Pseudobacciger cheneyae Sun, Bray, Yong, Cutmore & Cribb, 2014
Host: Chromis weberi Fowler & Bean, 1928.
Locality: Lizard Island
Voucher specimens: QM G 234368G 234387
Remarks: This species was described from Chromis weberi by Sun et al. (2014) (as part of this
study, published paper appended to this chapter) from Lizard Island. Pseudobacciger cheneyae is
the first faustulid to have been reported from a pomacentrid, with other members of the genus
known primarily from clupeids, but also from pomacanthids and sparids (Parukhin 1976b; Machida
1984; Dimitrov et al. 1999).
The systematics of this species, its family and higher classification are problematic. Although the
genus Pseudobacciger is classified within the family Faustulidae, phylogenetic analyses show that
P. cheneyae is more closely related to members of the superfamily Gymnophalloidea, which
includes, amongst others, the Fellodistomidae and Tandanicolidae, to which P. cheneyae is sister
(Sun et al. 2014). Currently, P. cheneyae displays oioxenic host-specificity in that it has only been
recorded from one host species, despite the ecology and behaviour of C. weberi resembling that of
other planktivorous damselfishes.
Fellodistomidae Nicoll, 1909
Fellodistomidae n. sp.
Host: Chrysiptera rollandi (Whitley, 1961).
Locality: Lizard Island
Voucher specimens: QM XXXXX
Remarks: This species was represented by just a single specimen from 1 of 9 Chrysiptera rollandi
examined. The specimen agrees with the family Fellodistomidae in its smooth tegument, restricted
vitellarian and Y-shaped excretory vesicle but not with any presently recognised genus. The fact
that only a single specimen was found casts doubt on its status as a genuine parasite of
pomacentrids, but no such form has been seen in any of thousands of other fishes examined at
Lizard Island. The status of this form remains unclear.

21
Gyliauchenidae Fukui, 1929
Gyliauchenidae n. sp.
Host: Stegastes apicalis (De Vis, 1885).
Locality: Lizard Island
Deposited specimens: QM XXXXX
Remarks: The Gyliauchenidae is a small family of trematodes in the superfamily Lepocreadioidea,
whose members primarily infect herbivorous reef fishes (Bray et al. 2009). Only one species,
Gyliauchen pomacentri Nahhas & Wetzel, 1995, has been previously reported from a pomacentrid
fish, Pomacentrus philippinus from off Suva, Fiji (Nahhas and Wetzel 1995). A description of this
new species is being prepared in collaboration with Dr Kathryn Hall of the Queensland Museum.
Haplosplanchnidae Poche, 1926
Schikhobalotrema pomacentri Skrjabin & Guschanskaja, 1955
Hosts: Amblyglyphidodon curacao (Bloch, 1787), Chromis viridis (Cuvier, 1830), Pomacentrus
moluccensis Bleeker, 1853.
Voucher specimen: QM XXXXX
Remarks: The family Haplosplanchnidae is known from a wide range of marine teleosts, with three
species from the genus Schikhobalotrema Skrjabin & Guschanskaja, 1955, reported from
pomacentrids. Schikhobalotrema crassum Pritchard & Manter, 1961, was described from Stegastes
fasciolatus (Ogilby, 1889) from Hawaii, S. robustum Pritchard & Manter, 1961 was described from
S. fasciolatus as well as two acanthurid and one chaetodontid species, also from Hawaii (Pritchard
and Manter 1961), and S. pomacentri (Manter, 1937) Skrjabin & Guschanskaja, 1955, has been
reported from 17 species of pomacentrid (plus one labrid species), from both Pacific and Atlantic
coasts of North, Central and South America, as well as Heron Island (Cribb et al. 1994). The
present specimens were consistent with S. pomacentri. The taxonomy of this genus has not been
considered in any detail for specimens from the GBR and we think it is likely that parallel
molecular studies will be needed to resolve it. The identification of S. pomacentri should be
considered provisional.
Lecithasteridae Odhner, 1905
Aponurus laguncula Looss, 1907
Host: Abudefduf whitleyi Allen & Robertson, 1974
Locality: Lizard Island
Voucher specimens: QM XXXXX

22
Remarks: This species is reported as being cosmopolitan, being found in various locations
worldwide and infecting over 60 different host species from 28 families (Bray & Mackenzie, 1990).
Bray et al. (1993a) first reported and described this species from the Pomacentridae, having
discovered it in Pomacentrus moluccensis from Heron Island, while Grutter et al. (2010) and Sun et
al. (2012) reported it from P. moluccensis and P. amboinensis from Lizard Island, respectively. Our
discovery of A. laguncula from the Whitley’s sergeant, Abudefduf whitleyi, represents a new host
record for this species. This species differs from the closely-related A. rhinoplagusiae Yamaguti,
1934, which also infects pomacentrids, in having a smaller seminal receptacle (Bray et al. 1993a).
Hysterolecitha heronensis Bray, Cribb & Barker, 1993
Host: Pomacentrus amboinensis Bleeker, 1868
Locality: Lizard Island
Voucher specimens: QM XXXXX
Remarks: Our specimens of this species match the original description by Bray et al. (1993a) from
five species of Pomacentridae from Heron Island, Pomacentrus amboinensis, P. moluccensis, P.
nigromarginatus Allen, 1973, P. philippinus Evermann & Searle, 1907, and an unidentified
Pomacentrus sp. It was later reported from P. amboinensis from Lizard Island (Sun et al. 2012).
Hysterolecitha heronensis is similar to H. nahaensis Yamaguti, 1942, with which it may have been
confused historically (Bray et al. 1993a). It is distinguished from H. nahaensis by possessing
notably digitiform vitelline lobes, a larger sinus-sac, a greater average sucker ratio and more distinct
genital atrium. It is noticeable that at Lizard Island this species has been found only in species of
one pomacentrid genus, Pomacentrus.
Hysterolecitha nahaensis Yamaguti, 1942 complex
Host: Abudefduf bengalensis (Bloch, 1787), Abudefduf sexfasciatus (Lacépède, 1801),
Acanthochromis polyacanthus (Bleeker, 1855), Amblyglyphidodon curacao (Bloch, 1787), Chromis
atripectoralis Welander & Schultz, 1951, Dascyllus reticulatus (Linnaeus, 1758), Dischistodus
melanotus (Bleeker, 1858), Neopomacentrus melas (Cuvier, 1830), Pomacentrus adelus Allen,
1991, Pomacentrus amboinensis Bleeker 1868, Pomacentrus chrysurus Cuvier, 1830, Pomacentrus
moluccensis Bleeker, 1853, Pomacentrus nagasakiensis Tanaka, 1917, Pomacentrus pavo (Bloch,
1787), Pomacentrus wardi Whitley, 1927.
Locality: Lizard Island
Voucher specimens: QM XXXXX
Remarks: Specimens recorded here as “Hysterolecitha nahaensis complex” created a special
identification problem. This species was described by Yamaguti (1942) from two species of

23
Pomacentridae and one of Scaridae with Dascyllus aruanus (Pomacentridae) as the type-host. From
the same general region it was subsequently re-reported from Japanese waters (Ichihara 1974; Dyer
et al. 1988), south Vietnam (King 1964), Indonesia (Sulawesi) (Yamaguti 1953), and the Great
Barrier Reef (Lester and Sewell 1990; Bray et al. 1993a; Barker et al. 1994; Sun et al. 2012). It has
also been reported from “Southern Seas” by Parukhin (1989) and from the Mediterranean by Rizk
et al. (1996). In Asia and waters from the Great Barrier Reef, records of this species have been
overwhelmingly from pomacentrids although there are single records from an acanthurid, a lobotid
and a scarid. Bray et al. (1993a) reported 23 species of Pomacentridae (and no other families of
fishes) as hosts for this species on the GBR. In that study a multivariate analysis of 62 specimens
identified some variation in which specimens from Dascyllus reticulatus showed the greatest
tendency to clumping, but the variation was considered inconclusive and all the specimens were
identified as a single species. Bray et al. (1993a) considered the record of Parukhin (1989) from
“Southern Seas” to be unconvincing.
In the present study H. nahaensis was by far the most common infection. However,
morphological examination of 105 specimens initially identified as H. nahaensis showed evidence
of representing three clearly distinct species as follows:
Type 1. Multiple specimens from Dascyllus reticulatus, two from Pomacentrus moluccensis, and
single specimens from Acanthochromis polyacanthus and Stegastes apicalis were distinguishable
from all other samples on the basis of their ventral suckers being proportionally distinctly smaller
than those of comparable-sized individuals from all other fishes (Figure 1).
Type 2. Eight morphological specimens (1 ex N. melas, 3 ex P. amboinensis, 2 ex P. moluccensis
and 2 ex P. wardi) differed from all other specimens in the sample in the ventral sucker being
proportionally far larger than those of comparable-sized specimens (Figure 1). The sucker length
ratio in these specimens ranged from 4.31-5.90. In addition, in these specimens the testes are
unusually close to the ventral sucker, the vitellarium is proportionally large, and the oral sucker is
proportionally small.
Type 3. This was by far the most common type (71 measured specimens) and it was found in 14
pomacentrid species Abudefduf sexfasciatus, Acanthochromis polyacanthus, Amblyglyphidodon
curacao, Dascyllus reticulatus, Dischistodus melanotus, Neoglyphidodon melas, Pomacentrus
adelus, P. amboinensis, P. chrysurus, P. moluccensis, P. nagasakiensis, P. pavo and P. wardi. The
size of the ventral sucker of these specimens was clearly intermediate between those of
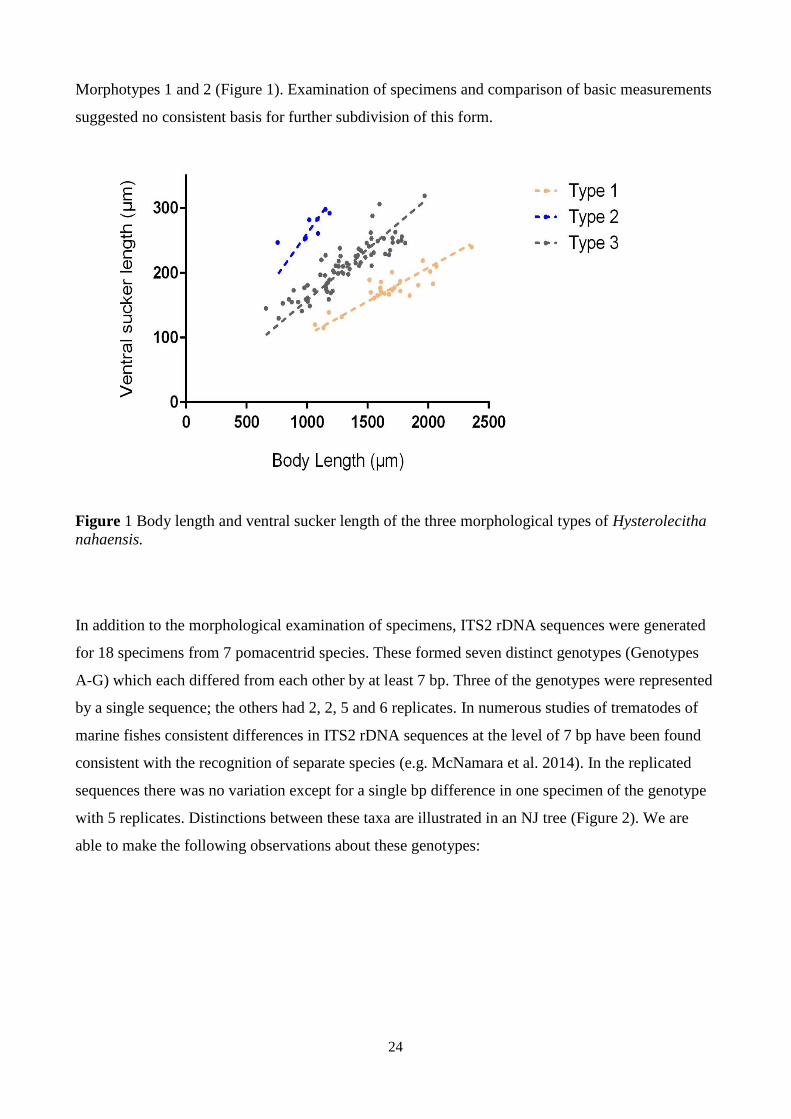
24
Morphotypes 1 and 2 (Figure 1). Examination of specimens and comparison of basic measurements
suggested no consistent basis for further subdivision of this form.
Figure 1 Body length and ventral sucker length of the three morphological types of Hysterolecitha
nahaensis.
In addition to the morphological examination of specimens, ITS2 rDNA sequences were generated
for 18 specimens from 7 pomacentrid species. These formed seven distinct genotypes (Genotypes
A-G) which each differed from each other by at least 7 bp. Three of the genotypes were represented
by a single sequence; the others had 2, 2, 5 and 6 replicates. In numerous studies of trematodes of
marine fishes consistent differences in ITS2 rDNA sequences at the level of 7 bp have been found
consistent with the recognition of separate species (e.g. McNamara et al. 2014). In the replicated
sequences there was no variation except for a single bp difference in one specimen of the genotype
with 5 replicates. Distinctions between these taxa are illustrated in an NJ tree (Figure 2). We are
able to make the following observations about these genotypes:

25
Figure 2 Neighbour joining tree based on ITS2 rDNA sequences of Hysterolecitha nahaensis
complex from Lizard Island. Samples are identified by host and dissection numbers.
Genotype A. 5 sequences ex Dascyllus reticulatus and 1 ex Stegastes apicalis. This genotype
corresponds with Morphotype 1 in that all sequences from D. reticulatus were of this genotype and
the great majority of morphological specimens were of Morphotype 1. The single sequence and
morphological specimen from S. apicalis corresponded in the same way.
Genotype B. Two identical sequences ex Chromis atripectoralis. No complete morphological
specimens corresponding to this genotype are available; partial paragenophore specimens are
consistent with H. nahaensis but are unusually large. One paragenophore specimen with part of the
posterior body removed measures 1,816 µm to the vitellarium, far longer than any other specimen
in the complete collection for which the maximum measurement is 1,505 µm. The combination of
genetic and morphological distinction are presumably consistent with the presence of a distinct
species.
Genotype C. A single completely distinct sequence ex Pomacentrus moluccensis. This is one of
three genotypes reported from this species; it is the repeated sharing of hosts by multiple genotypes

26
that makes delineation in this group especially problematic. The single morphological specimen
from the dissection corresponding to the sequenced specimen is a standard and unremarkable
Morphotype 3. It is not presently possible to relate this genotype to a distinct morphotype.
Genotype D. A single completely distinct sequence ex Pomacentrus adelus. This host is also
represented in Genotype E. There is no corresponding morphological specimen from the fish from
which the sequenced specimen was obtained. It is not presently possible to relate this genotype to a
distinct morphotype.
Genotype E. Five sequences from Chromis viridis, Amblyglyphidodon curacao, Pomacentrus
adelus and P. moluccensis (4 x identical, the C. viridis sample differs at 1 bp). The great bulk of the
specimens from these hosts relate to Morphotype 3 including two sequenced from the same
dissections as are represented among the morphological material. This combination presumably
reflects a distinct species and Genotype C and Morphotype 3 are here provisionally identified as the
same species. However, this identification is compromised by the fact that other genotypes have
been found in two of the fish species (P. adelus and P. moluccensis).
Genotype F. A single completely distinct sequence ex Abudefduf whitleyi. No morphological
specimens at all are available from this host. The few specimens from Abudefduf sexfasciatus (for
which there are no sequences) are slightly distinctive in having unusually large ventral suckers for
the specimens in Morphotype 3. It is not presently possible to relate this genotype to a distinct
morphotype, but it seems possible that it represents a distinct “Abudefduf” species.
Genotype G. Two identical sequences ex P. moluccensis from two separate individual fish. One of
those fish had infections of both Morphotype 2 and 3, the other only Morphotype 3 among the
examined morphological specimens. It is not presently possible to relate this genotype to a distinct
morphotype.
There are no ITS2 rDNA sequences for specimens of Hysterolecitha nahaensis available for
comparison from any other locality; indeed, there are none for the genus except for an 18S rDNA
sequence of H. nahaensis (see Blair et al. 1998). Perusal of specimens identified as H. nahaensis by
Bray et al. (1993a) from pomacentrids from Heron Island suggests that there is almost certainly
more than one species represented.
We conclude that it is impossible to be definitive about the richness of Hysterolecitha in
GBR pomacentrids. The subject requires further intensive study, especially the sequencing of more
paragenophore specimens. It is noteworthy that the type-host of H. nahaensis, Dascyllus aruanus,

27
was surveyed thoroughly (n = 20) but had no infections of any sort at Lizard Island. For the
purposes of the present analysis we have chosen to record all the forms discussed above as “H.
nahaensis complex”. This is for two reasons. First, although some of the putative taxa within the
complex appear to be clearly distinct, there are many grey areas in which the differences between
putative taxa are completely unresolved; we are unable to propose a convincing hypothesis for the
number of species present although we conclude that there are at least four, but potentially as many
as seven. Given that three genotypes were represented by only single sequences and that specimens
from many pomacentrid species have not been sequenced at all, it seems possible that even more
diversity remains undetected in this system. Second, a key aim of this study was to compare the
trematode fauna of pomacentrids of the northern GBR with that reported previously for the southern
GBR. We conclude that the identifications of the H. nahaensis complex for the southern GBR are
also confounded by the presence of a complex of species and reiterate that no molecular data at all
are available from there. We thus propose to identify all these forms as “H. nahaensis complex”
with a view to future resolution; determining the extent to which the fauna of the northern and
southern GBR is comparable for this complex is thus beyond the scope of the present study.
Lecithaster stellatus Looss, 1906
Hosts: Chrysiptera rollandi (Whitley, 1961), Pomacentrus amboinensis Bleeker, 1868,
Pomacentrus moluccensis Bleeker, 1853.
Locality: Lizard Island
Voucher specimens: QM XXXXX
Remarks: Our specimens of this species match the description by Bray et al. (1993a). This species
has a large distribution range, having been recorded throughout the Indo-west and west-central
Pacific, as well as from the Mediterranean Basin, in both tropical and temperate waters (Bray et al.
1993a). It has been recorded from 17 fish families so far (reviewed by Bray et al. 1993a), including
at least one record from fresh water in anadromous fishes (Salmonidae) (Bykhovskaya-Pavlovskaya
et al. 1962). Bray et al. (1993a) reported L. stellatus from 11 species of Pomacentridae from Heron
Island, while Sun et al. (2012) reported it from Pomacentrus amboinensis from Lizard Island. Our
discovery of this species in Chrysiptera rollandi and Pomacentrus moluccensis represent new host
records. The species has also been reported from Lizard Island, from the labrid Coris batuensis
(Bleeker, 1856) by Muñoz and Cribb (2006).
Thulinia microrchis Yamaguti, 1934
Hosts: Abudefduf septemfasciatus (Cuvier, 1830), Acanthochromis polyacanthus (Bleeker, 1855),
Dischistodus melanotus (Bleeker, 1858), Pomacentrus amboinensis Bleeker, 1868.

28
Locality: Lizard Island
Voucher specimens: QM XXXXX
Remarks: Our specimens match the description of Bray et al. (1993a). This species has a
widespread distribution and is reported to be highly euryxenic (Bray et al. 1993a; Miller et al.
2011b). It has been reported from Heron Island in 15 species of Pomacentridae as well as in species
of Acanthuridae, Chaetodontidae, Kyphosidae, Lethrinidae, Pomacanthidae, Serranidae and
Siganidae (Yamaguti 1938; Manter 1969a; Ichihara 1986; Lester and Sewell 1990; Bray et al.
1993a; Barker et al. 1994). Despite the disparity in dietary preferences of these fishes, unpublished
molecular evidence indicates the conspecificity of this species, at least across several host families
(Miller et al. 2011b). Our finding of T. microrchis in Abudefduf septemfasciatus represents a new
host record; T. microrchis was previously reported from three other species of Abudefduf, A.
bengalensis, A. sexfasciatus and A. whitleyi (see Bray et al. 1993a).
Lepocreadiidae Odhner, 1905
Lepocreadium oyabitcha Machida, 1984
Hosts: Abudefduf bengalensis (Bloch, 1787), Abudefduf sexfasciatus (Lacépède, 1801), Abudefduf
whitleyi Allen & Robertson, 1974.
Locality: Lizard Island
Voucher specimens: QM XXXXX
Remarks: This species was described by Machida (1984) from Abudefduf vaigiensis (Quoy &
Gaimard, 1825) from Okinawa, Japan. A single specimen was reported from A. whitleyi from
Lizard Island by Bray and Cribb (1998), who noted several differences between their specimens and
those of Machida (1984), including in body shape, having a tri-lobed (instead of ovoid) ovary, and
in infection site (the gut, as opposed to the gall-bladder). Despite this, Bray and Cribb (1998) regard
the two as being conspecific, pointing out the distinctive “ramifying moniliform-vitellarium” as an
important unifying feature. Our findings agree with those of Bray and Cribb (1998), and expand the
host range of this species by two (Abudefduf bengalensis and A. sexfasciatus). It seems likely that
this species is restricted to fishes of the genus Abudefduf.
Lepotrema adlardi (Bray, Cribb & Barker, 1993)
Host: Abudefduf bengalensis (Bloch, 1787).
Locality: Lizard Island
Voucher specimens: QM XXXXX
Remarks: This species was described from Abudefduf bengalensis from Heron Island by Bray et al.
(1993b) (as Lepocreadium adlardi). Revision of the Lepocreadiidae by Bray and Cribb (1996) led

29
to the transfer of this species to Lepotrema. All observed features in our material are consistent with
those of L. adlardi. The species differs from other species of Lepotrema in the length of its
forebody, the extent of its vitelline follicles and having a relatively long prepharynx. Abudefduf
bengalensis remains the only known host of L. adlardi, despite the examination of several other
species of Abudefduf at both Heron and Lizard Island.
Lepotrema clavatum Ozaki, 1932
Hosts: Acanthochromis polyacanthus (Bleeker, 1855), Pomacentrus amboinensis Bleeker, 1868.
Locality: Lizard Island
Voucher specimens: QM XXXXX
Remarks: This species was described by Ozaki (1932) from the monocanthid Stephanolepis
cirrhifer (Temminck & Schlegel, 1850) from Japan. It has since been reported in four fish families
from across the Indo-Pacific (Balistidae, Bothidae, Chaetodontidae and Pomacentridae) (see Bray
and Cribb 1996 for a full list of infected fish species). This species has been recorded in two species
of pomacentrids (Acanthochromis polyacanthus and Parma polylepis Günther, 1862) at Heron
Island on the GBR. It has also been recorded from another pomacentrid species in Hawaii,
Dascyllus albisella Gill, 1862, (Pritchard 1963). The specimens we collected are consistent with
those reported by Bray et al. (1993b) from Heron Island (as Lepocreadium clavatum), with
Pomacentrus amboinensis representing a new host record for this species. Lepotrema clavatum is
easily distinguished from the other two species of Lepotrema which infect pomacentrids. From L.
adlardi it differs in the extent of the vitelline follicles, the length of the prepharynx and length of
the forebody, and from L. monile Bray & Cribb, 1998 it differs in having a tri-lobed ovary and
lacking a sphincter around the distal metraterm. The host-specificity of L. clavatum is unusual for
this genus, being present in several fish families. Research presently underway suggests that L.
clavatum will prove to be a species complex comprising species with narrower specificity than
currently reported. This form from pomacentrids may ultimately became known under a separate
name. Records for the Great Barrier Reef show that this species is abundant in A. polyacanthus
(This study; Bray et al. 1993b) and at best uncommon in all other pomacentrids.
Lepotrema monile Bray & Cribb, 1998
Hosts: Abudefduf septemfasciatus (Cuvier, 1830), Pomacentrus amboinensis Bleeker, 1868,
Pomacentrus chrysurus Cuvier, 1830, Pomacentrus wardi Whitley, 1927.
Locality: Lizard Island
Voucher specimens: QM XXXXX

30
Remarks: This species was first reported as Lepocreadium sp. from Pomacentrus wardi from Heron
Island (Bray et al. 1993b). Bray and Cribb (1996) revived the genus Lepotrema and included five
species within it, defined by having a combination of a dorsally-positioned excretory pore, a tri-
lobed ovary and a “folded muscular pad” on the distal surface of the metraterm. Bray and Cribb
(1998) proposed L. monile for this form, noting that while it lacks a tri-lobed ovary and the folded
muscular pad, it does possess a dorsal excretory pore. This species was subsequently reported from
Pomacentrus amboinensis from Lizard Island by Sun et al. (2012). Our specimens agree closely
with the original description of this species. Abudefduf septemfasciatus and P. chrysurus represent
new hosts for this species.
Preptetos cannoni Barker, Bray and Cribb, 1993
Hosts: Abudefduf septemfasciatus (Cuvier, 1830).
Locality: Lizard Island
Deposited specimen: QM XXXXX
Remarks: This species was described by Barker et al. (1993) from three species of Siganidae from
Heron Island. It has also been reported from Pomacentrus bankanensis (see Bray et al. 1993b).
Species of the genus Preptetos resemble Lepocreadium Stossich, 1903, but can be distinguished by
the excretory vesicle passing between the testes. The specimen from A. septemfasciatus agrees with
the original descriptions by Barker et al. (1993) and represents a new host record for this species. In
this study, only a single individual of P. cannoni was found. The low prevalence and intensity
detected is consistent with the findings of Bray et al. (1993b) who also reported just a single
individual pomacentrid (Pomacentrus bankanensis) being infected. It seems that the species
overwhelmingly infects siganids, and occurs in pomacentrids only incidentally.
Monorchiidae Odhner, 1911
Hurleytrematoides morandi McNamara & Cribb, 2011
Host: Pomacentrus wardi Whitley, 1927.
Locality: Lizard Island
Voucher specimens: QM XXXXX
Remarks: Species of Hurleytrematoides have been reported from several locations including Florida
(Manter 1942), China (Wang and Wang 1993), Indonesia (Machida 2005) and Australia
(McNamara et al. 2012). They overwhelmingly infect fishes from the family Chaetodontidae, with
some records from Acanthuridae, Pomacanthidae and Tetraodontidae (McNamara and Cribb 2011).
This species can be distinguished from other species of Hurleytrematoides by its distinctive body
shape and by its distinctively spined cirrus-sac (McNamara and Cribb 2011). This record, based on

31
the discovery of a single specimen in Pomacentrus wardi, is the first of this family from the
Pomacentridae. This species has previously been found only (and repeatedly) in species of
Chaetodon. This individual infection is best considered a straggler or ‘accidental’.
Transversotrematidae Witenberg, 1944
Transversotrema damsella Hunter & Cribb, 2012
Hosts: Abudefduf septemfasciatus (Cuvier, 1830), Abudefduf sexfasciatus (Lacépède, 1801).
Locality: Lizard Island
Deposited specimens: QM XXXXX
Remarks: This species was described from Abudefduf bengalensis from Lizard Island by Hunter and
Cribb (2012) and has also been recorded from several other pomacentrid species: Abudefduf
septemfasciatus, A. sexfasciatus, Acanthochromis polyacanthus and Amblyglyphidodon curacao, all
from Lizard Island. Further examination of the host records of T. damsella suggests that this species
predominantly infects damselfishes of the genus Abudefduf (see Hunter and Cribb 2012).
Interestingly, T. damsella has not been found at Heron Island despite numerous damselfish species
having been examined there (Barker et al. 1994). Transversotrema damsella is distinguished from
other Transversotrema species by the ovary being larger than the testes and also has a relatively
distinctive overall shape.
Zoogonidae Odhner, 1902
Steganoderma gibsoni Cribb, Bray & Barker, 1992
Hosts: Abudefduf septemfasciatus (Cuvier, 1830), Abudefduf sexfasciatus (Lacépède, 1801),
Abudefduf whitleyi Allen & Robertson, 1974.
Locality: Lizard Island
Voucher specimens: QM XXXXX
Remarks: The present specimens agree well with S. gibsoni as described originally from A. whitleyi
from the southern GBR (Cribb et al. 1992) and it was found in two further Abudefduf species. This
is the only zoogonid known from a GBR pomacentrid. The records reported here suggest that this
species is probably restricted to species of Abudefduf. There have been several other records of
zoogonids from pomacentrids; Deretrema fusillus Linton, 1910 from Abudefduf saxatilis (Linnaeus,
1758) from Florida (Linton 1910; Manter 1934,1947), Deretrema sp. from A. sordidus (Forsskål,
1775) from Saipan in Micronesia (Toman and Kamegai 1974) and Diphterostomum americanum
Manter, 1947 from Stegastes leucostictus (Müller & Troschel, 1848) from Florida (Overstreet
1969).
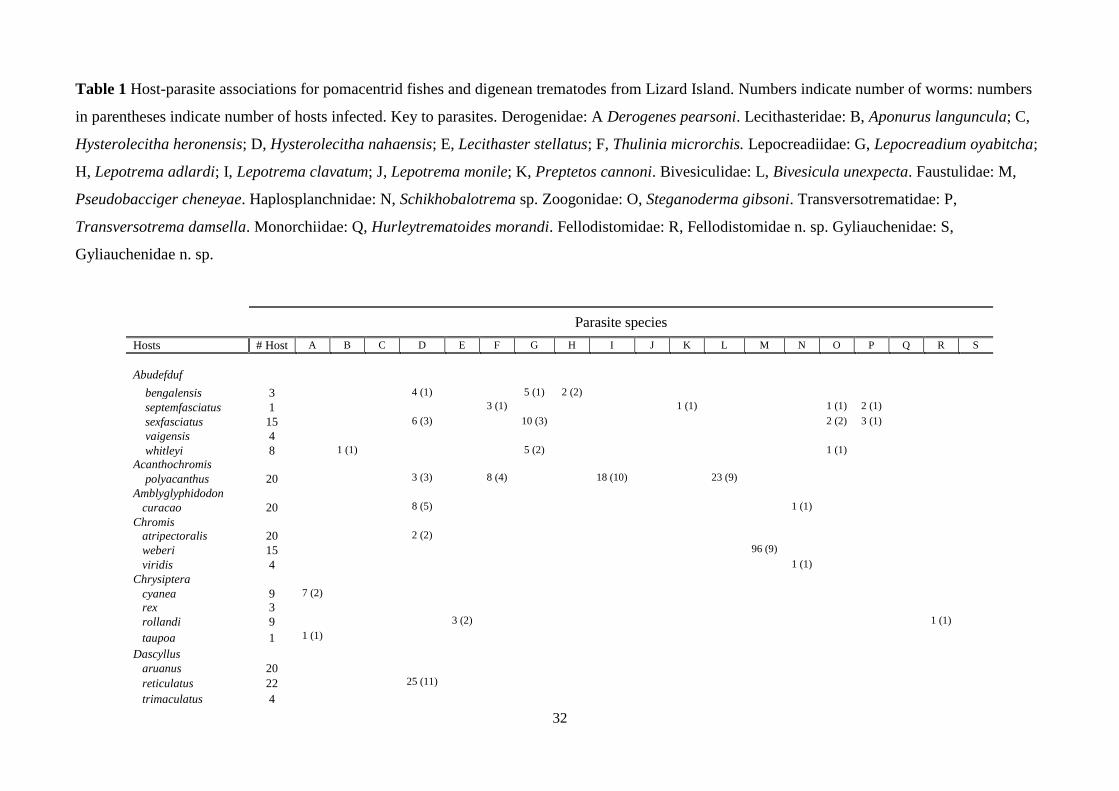
32
Table 1 Host-parasite associations for pomacentrid fishes and digenean trematodes from Lizard Island. Numbers indicate number of worms: numbers
in parentheses indicate number of hosts infected. Key to parasites. Derogenidae: A Derogenes pearsoni. Lecithasteridae: B, Aponurus languncula; C,
Hysterolecitha heronensis; D, Hysterolecitha nahaensis; E, Lecithaster stellatus; F, Thulinia microrchis. Lepocreadiidae: G, Lepocreadium oyabitcha;
H, Lepotrema adlardi; I, Lepotrema clavatum; J, Lepotrema monile; K, Preptetos cannoni. Bivesiculidae: L, Bivesicula unexpecta. Faustulidae: M,
Pseudobacciger cheneyae. Haplosplanchnidae: N, Schikhobalotrema sp. Zoogonidae: O, Steganoderma gibsoni. Transversotrematidae: P,
Transversotrema damsella. Monorchiidae: Q, Hurleytrematoides morandi. Fellodistomidae: R, Fellodistomidae n. sp. Gyliauchenidae: S,
Gyliauchenidae n. sp.
Parasite species
Hosts # Host A B C D E F G H I J K L M N O P Q R S
Abudefduf
bengalensis 3 4 (1) 5 (1) 2 (2)
septemfasciatus 1 3 (1) 1 (1) 1 (1) 2 (1)
sexfasciatus 15 6 (3) 10 (3) 2 (2) 3 (1)
vaigensis 4
whitleyi 8 1 (1) 5 (2) 1 (1)
Acanthochromis
polyacanthus 20 3 (3) 8 (4) 18 (10) 23 (9)
Amblyglyphidodon
curacao 20 8 (5) 1 (1)
Chromis
atripectoralis 20 2 (2)
weberi 15 96 (9)
viridis 4 1 (1)
Chrysiptera
cyanea 9 7 (2)
rex 3
rollandi 9 3 (2) 1 (1)
taupoa 1 1 (1)
Dascyllus
aruanus 20
reticulatus 22 25 (11)
trimaculatus 4

33
Dischistodus
melanotus 4 3 (1) 2 (2)
pseudochrysopecilus 4
Neoglyphidodon
melas 20 48 (6)
Neopomacentrus
azysron 21 1 (1)
Plectroglyphidodon
lacrymatus 2
Pomacentrus
adelus 17 7 (1) 2 (1) 10 (5)
amboinensis 23 1 (1) 10 (6) 3 (3) 1 (1)
chrysurus 22 7 (4) 1 (1) 1 (1)
coelestis 12 1 (1)
moluccensis 27 18 (6) 2 (2) 1 (1) 2 (1)
nagasakiensis 7 3 (2)
pavo 5 1 (1)
wardi 8 4 (3) 1 (1)
Stegastes
apicalis 7 8 (2) 1 (1) 7 (3)
fasciolatus 1

34
Discussion
Identification of the fauna
From a total of 358 individuals of 32 species of pomacentrids examined we found 19 species of
digenean trematodes and 54 host/parasite combinations. A total of 19 host distributions were new,
18 were new host records, and three were new species. The characterisation of this fauna proved in
part straightforward and in part complex. It was straightforward in that the great majority of the
species were easily identified as forms previously reported from pomacentrids or, in a few cases,
other fishes. The three new species, a faustulid, a fellodistomid and a gyliauchenid, were all readily
recognisable as distinct and were the sole representatives of their respective families encountered.
The complexities of identification relate almost entirely to the delineation of the Hysterolecitha
nahaensis complex. As outlined above, this species was by far the most common infection
encountered but combined molecular and morphological evidence suggests that it comprises at least
four species, there is limited evidence for as many as seven species, and it seems entirely possible
that there could be more than that. On the basis of the present evidence it is possible to distinguish
only three taxa by morphology of which the most common, Morphotype 3, almost certainly is a
complex of multiple species. In addition to the problem relative to H. nahaensis, it seems likely that
some of the other species identified (especially Aponurus languncula, Lecithaster stellatus and
Lepotrema clavatum) will ultimately become recognised under different names, but their distinction
as single taxa in pomacentrids does not appear to be in doubt.
The second complexity in the characterisation of this fauna is in the size of the task. The
study incorporated the examination of 372 individuals of 32 pomacentrid species. Despite the fact
that these numbers represent a substantial number of individual fishes for field and processing
effort, the incompleteness of the survey of the fauna is demonstrated by two features of the data.
First, there remain a further 58 pomacentrid species known from Lizard Island that have never been
examined for parasites; undoubtedly these will harbour many new host-parasite combinations. The
second feature of the fauna is the rarity of many infections. Of 54 host/parasite combinations, 27
(exactly half) were detected in only a single individual fish. Such a level of rarity implies that a
great many more host/parasite combinations remain undetected for the hosts that have been
examined. Some of the infections found only once are evidently stragglers (e.g. Preptetos cannoni
and Hurleytrematoides morandi) and are thus insignificant in the understanding of the fauna. In
contrast, the distribution of oioxenic (single host) species is difficult to predict and there are likely
to be a number of those that remain completely undetected so far.
Below we consider the nature of host-specificity, overall richness patterns and beta diversity
for this fauna with the understanding that the data set remains incomplete.

35
Richness
Despite the Pomacentridae being a large and diverse group of coral reef fishes, the mean
species richness of digeneans in species within this family was low at 1.69. Results show that 20 of
32 pomacentrid species examined from Lizard Island were infected with only 1 ̶ 3 species of
digeneans; just five species harboured four species of digenean; seven of the 32 species of
pomacentrids had no digeneans at all. For many pomacentrid species the samples were low, but it is
striking that the 20 Dascyllus aruanus produced no infections at all.
It is well-established that parasite richness within host species is not random, but is broadly
related to host species traits, especially size, density and distribution (Guégan et al. 1992; Kamiya et
al. 2014). Host size probably explains the relatively low richness of adult digeneans among
pomacentrid species relative to larger coral reef fishes such as chaetodontids, lutjanids and serranids
as well as among the pomacentrid species. Notably the larger species in the study, species of
Abudefduf, were infected with most digenean species. Generally, host size is a strong predictor for
parasite richness because larger-bodied individuals provide greater space and resources for parasites
than smaller-bodied individuals (Sasal et al. 1997; Kamiya et al. 2014). However, at the
infracommunity level, parasite richness is more variable when examining the effects of increasing
host size. Two medium-sized pomacentrid species, Pomacentrus amboinensis and P. moluccensis,
were also infected with 4 species of digeneans; notably, however these two species had the two
largest samples examined in this study indicating the importance of sample size. Overall, although
host size is a good predictor of parasite richness it will not necessarily over-rule the effects of
different habitats, densities and diet (Lo et al. 1998; Torres et al. 2006; Morand 2015).
The reported digenean fauna of pomacentrids from both the northern and southern GBR
suggests that there has been little radiation of families of trematodes species within this group; the
total of 23 species are drawn from 11 trematode families. Apart from the presence of three species
of Lepotrema the clear exception is the H. nahaensis complex; however, interpretation of the nature
of the H. nahaensis complex must await its better characterisation. The limited radiation of
individual genera is in contrast to many other GBR fish family/trematode family complexes
Bucephalidae Poche, 1907 in serranids (Bott and Cribb 2009), Paradiscogaster Yamaguti, 1934
(Diaz et al. 2013) and Hurleytrematoides Yamaguti,1953 (McNamara and Cribb 2011) in
chaetodontids, Opecoelidae Ozaki, 1925 in mullids (Rohner and Cribb 2013) and Cryptogonimidae
Ward, 1917 in lutjanids (Miller and Cribb 2007). In all these combinations there is much greater
parasite richness than seen for any of the families represented in pomacentrids.
As found by Barker et al. (1994) at Heron Island, the diversity of pomacentrid species is far
greater than the diversity of digenean trematodes at Lizard Island. Other studies from other
locations have also shown that the diversity of digeneans in pomacentrids is relatively low (Yeo and

36
Spieler 1980; Dyer et al. 1985). However, it should be noted that this study only examined 32
species of pomacentrids from a possible 89 species that have been recorded at Lizard Island
(Grutter et al. in review). Thus, there is the likelihood that other species of digenean worms will be
found, as well as extending the host records for already known species.
Host-specificity
The host specificity of the digeneans of pomacentrids varies, with a tendency towards being
relatively low. We found just three1 species to be convincingly oioxenic (i.e found repeatedly in
only one host species) at Lizard Island: Bivesicula unexpecta was only found in Acanthochromis
polyacanthus, Lepotrema adlardi only in Abudefduf bengalensis and Pseudobacciger cheneyae only
in Chromis weberi. Due to only a single specimen being recorded, further examination is required
to determine whether Fellodistomidae sp. from Chrysiptera rollandi is indeed oioxenic or in fact
occurs in other pomacentrids. Our findings are in support of earlier studies regarding Bivesicula
unexpecta and Lepotrema adlardi (Bray et al. 1993b; Cribb et al. 1994a). The majority of the
digenean species reported in this study are apparently stenoxenic, i.e. infecting multiple species
within the same family; Derogenes pearsoni, Hysterolecitha heronensis, H. nahaensis complex,
Lepocreadium oyabitcha, Lepotrema monile, Steganoderma gibsoni (see Cribb et al. 1992; Bray et
al. 1993a; Bray and Cribb 1998) and Transversotrema damsella (Hunter and Cribb 2012) are all in
this category. Two of these stenoxenic species, H. heronensis and L. oyabitcha, can be further
categorised as being restricted to multiple congeners of Pomacentrus and Abudefduf, respectively.
The remaining species are classified as euryxenic, infecting a wide range of hosts from multiple
families; Aponurus laguncula, Lecithaster stellatus, Lepotrema clavatum, Hurleytrematoides
morandi, Preptetos cannoni and Thulinia microrchis. However, it should be noted that H. morandi
and P. cannoni, which are common in chaetodontids and siganids respectively are technically
euryxenic, but are effectively stenoxenic to the Chaetodontidae and Siganidae, infecting
pomacentrids only incidentally. This could also be the case for A. languncula which has not been
reported other than from three species of pomacentrids. Infections of these pomacentrid species are
rare, suggesting that the infections may also be incidental.
A likely explanation for the generally broad host range of many species are the eco-
physiological similarities between pomacentrid species, especially their broad and overlapping
diets, which may facilitate host-sharing in digeneans (Marcogliese 2002). Use of intermediate hosts
by metacercariae facilitates transmission into the definitive host (Poulin and Cribb 2002). Broadly
overlapping diets increase the possibility that multiple species of fish will encounter the same suite
1 In this study the gyliauchenid was oioxenic, but subsequent collections by colleagues (not reported
here) has shown that it infects at least two species in addition to Stegastes apicalis.

37
of metacercariae. The fact that hemiuroid metacercariae are known to infect a range of zooplankton
such as calanoid copepods and polychaetes (Marcogliese 1995) which in turn are consumed by
many pomacentrids, may explain the high prevalence of digeneans from this superfamily infecting
multiple pomacentrid species. Although diet may explain some of the variation in the infection of
pomacentrids by certain digeneans species (e.g. Hysterolecitha nahaensis complex), it does not
explain the high specificity observed in other species (e.g. Pseudobacciger cheneyae to just one
species of Chromis, Lepotrema adlardi to just one species of Abudefduf). In such cases the
explanation may be physiological compatibility (Adamson and Caira 1994). The physiological
compatibility between a parasite and its potential hosts may play an important role in determining
the suitability of those hosts, thus driving host specificity. However, the extent to which physiology
influences host specificity has not been explored for most fish-infecting digeneans and is poorly
understood (Miller et al. 2011b).
It is notable that some of the digeneans we found were species which are interpreted as
having cosmopolitan distributions and exhibiting euryxenicity, i.e. extremely wide host ranges. We
encountered five species which could be classed as being euryxenic, including two (Aponurus
languncula and Lecithaster stellatus) that are reported to be distributed circumglobally.
Euryxenicity is observed in a small number of marine digenean species worldwide (Bartoli et al.
2005) and has been reported for only 23 of the 290 species of digeneans on the GBR (Miller et al.
2011b). It is possible, perhaps probable, that some of these euryxenic species may comprise
complexes of cryptic species that have distinct geographical distributions. The two cosmopolitan
species reported in this study are from the superfamily Hemiuroidea, which are characterised by
many of its member taxa exhibiting low host specificity (Gibson and Bray 1979).
Although A. laguncula has not been reported from any fish family on the GBR other than
pomacentrids (reported from 3 species only), it has been recorded in 67 teleost species from 29
families, making it putatively one of the most widespread digeneans with respect to both
geographical distribution and host range (Bray and MacKenzie 1990; Bray et al. 1993a). It is
currently unclear whether all records of A. laguncula comprise one species or form a species
complex. Recent molecular data have indicated the presence of a cryptic species of Aponurus within
the A. laguncula complex, A. mulli Carreras-Aubets, Repulles-Albelda, Kostadinova & Carrasson,
2011, which infects red mullet, Mullus barbatus Linnaeus, 1758, from the Mediterranean (Carreras-
Aubets et al. 2011). It is possible that further exploration will reveal that Aponurus laguncula is
actually comprised of multiple cryptic species, and that A. laguncula sensu stricto may actually not
be found in Australian waters. Conversely, it is also possible that molecular work may reveal that A.
laguncula is indeed one species, despite its apparent cosmopolitan distribution, as has been
previously demonstrated for the tuna-infecting aporocotylid fluke Cardicola forsteri Cribb, Daintith

38
& Munday, 2000 (Aiken et al. 2007). The same principles may also apply to the other reportedly
cosmopolitan species, Lecithaster stellatus.
Cryptic species may also occur within some of the stenoxenic species found on the GBR, in
particular Hysterolecitha nahaensis. Currently, H. nahaensis has been recorded in a total of 26
pomacentrid species from Lizard and Heron Islands, making it the digenean with the widest host
range. This species closely resembles H. heronensis; the two species may have been historically
confused with each other due to their morphological similarities (Bray et al. 1993a). These species
require comprehensive reconsideration with combined morphological or molecular analysis. It is
apparent that a number of cryptic species are present within these species, as has been demonstrated
for other digeneans in other hosts (Jousson et al. 2000; Nolan and Cribb 2005; Diaz et al. 2015).
Beta diversity
These data create a rare opportunity to consider beta diversity in parasite faunal composition on the
GBR. Both the depth of study and richness of digenean trematodes encountered in this study is
comparable to that of Barker et al. (1994), who examined 312 individuals from 39 species of
pomacentrids from Heron Island, on the southern GBR (see Table 2). Twenty-one pomacentrid
species were examined at both sites. Of these, six had robust samples of at least 10 individuals at
both sites. Although the overall trematode richness is almost identical for the two sites, it is striking
that mean richness and prevalence were reported as much higher at Heron than at Lizard Island.
Table 2 Comparison between data collected from Heron Island (Barker et al. 1994) and Lizard
Island (present study) for digeneans from pomacentrid fishes.
HI LI
Fish species 39 32
Fish individuals 312 358
Parasite species 18 19
Host/parasite combinations 86 54
Mean richness 2.205 1.688
Total infections 209 140
Mean overall prevalence 0.67 0.39

39
The total digenean fauna for GBR pomacentrids now stands at 23 species in 50 pomacentrid
species of which 14 are now known from both locations, five have been only recorded from Lizard
Island (Lepocreadium oyabitcha, Pseudobacciger cheneyae, Transversotrema damsella,
Fellodistomid sp and Gyliauchenid sp.) and four only from Heron Island (Derogenes pharyngicola,
Lepocreadium sp., Preptetos xesuri and Macvicaria heronensis). Of the five species known only
from Lizard Island, only three can be inferred to be absent from Heron Island. As only a single
specimen of the fellodistomid digenean was recorded and given that no Chromis weberi have been
examined at Heron Island for P. cheneyae, despite records indicating that C. weberi does occur
there (Russell 1983), we cannot conclude that these species do not occur at Lizard Island; it is
possible that with further examination, these species may prove to be present at Heron Island. In
contrast, the evidence is good that three species (L. oyabitcha, T. damsella and Gyliauchenid n. sp.)
do not occur at Heron Island. Of the four species known only from Heron Island, only D.
pharyngicola is a regularly occurring parasite restricted to pomacentrids; Preptetos xesuri and
Macvicaria heronensis are mainly parasites of other fishes and the Lepocreadium sp. from
Amblyglyphidodon curacao has been collected only twice. We conclude that it is possible to
identify a “core pomacentrid fauna” of species that are regular in and dependent on pomacentrids
(Table 3). From this core fauna are excluded species that are rare to the point that their status is
uncertain (e.g. Aponurus laguncula, Fellodistomid sp.) and those that are clearly principally
parasites of other fishes (e.g. Preptetos cannoni and Hurleytrematoides morandi). With such
exclusions we recognise 14 morphospecies in this category with the understanding that “H.
nahaensis complex” certainly represents multiple species.

40
Table 3 Core pomacentrid specific digeneans from Heron and Lizard Island.
The only previous detailed comparison of the parasite fauna of comparable fish from the
southern and northern GBR was that of monorchiid trematodes of chaetodontid fishes (McNamara
and Cribb 2011; McNamara et al. 2012). A total of 10 species in total was reported, of which all
occurred at Heron Island but only seven were found at Lizard Island. This finding ran counter to the
general pattern of greater biological richness on the northern than on the southern GBR (Russell
1983). Here the comparison is less straightforward because of the complex (and partly uncertain)
host-specificity of the pomacentrid parasites (whereas the monorchiids of chaetodontids show
reliably high specificity) and because a smaller number of the host species sampled at the two sites
were in common. Of the 14 core pomacentrid species, 10 (71 %) have been found at Heron Island
and 13 (93 %) at Lizard Island (Table 3). These numbers are consistent with the interpretation that
there are significant differences in the parasite fauna at the two sites and that overall richness is
slightly higher in the north. That both mean species richness and mean overall prevalence were
reported as much higher at Heron Island by Barker et al. (1994) is puzzling. There are so many
areas of uncertainty in these comparisons (from the true identity of several of the species to the
robustness of the sample sizes) that these findings can only be a basis for future analyses.
Family Genus Species HI LI
Bivesiculidae Bivesicula unexpecta ● ●
Derogenidae Derogenes pearsoni ● ●
Derogenes pharyngicola ●
Faustulidae Pseudobacciger cheneyae ●
Gyliauchenidae Gyliauchen sp. ●
Haplosplanchnidae Schickhobalotrema pomacentri ● ●
Lecithasteridae Hysterolecitha heronensis ● ●
Hysterolecitha nahaensis
(complex) ● ●
Lepocreadiidae Lepocreadium oyabitcha ●
Lepotrema adlardi ● ●
Lepotrema clavatum ● ●
Lepotrema monile ● ●
Transversotrematidae Transversotrema damsella ●
Zoogonidae Steganoderma gibsoni ● ●
Count 10 13

41
Acknowledgments
We thank E. C. McClure, F. Cortesi, K. L. Cheney, S. Martin for their help collecting specimens
and the staff of the Lizard Island Research Station for their support. This work was funded by a Sea
World Research and Rescue Foundation, Australia (SWRRFI) awarded to D. Sun and T. H. Cribb
and an Australian Research Council Discovery Grant awarded to T. H. Cribb.

42
Pseudobacciger cheneyae n. sp. (Digenea:
Gymnophalloidea) from Weber’s chromis (Chromis
weberi Fowler & Bean) (Perciformes: Pomacentridae) at
Lizard Island, Great Barrier Reef, Australia

43
Pseudobacciger cheneyae n. sp. (Digenea: Gymnophalloidea) from Weber’s
chromis (Chromis weberi Fowler & Bean) (Perciformes: Pomacentridae) at
Lizard Island, Great Barrier Reef, Australia
Derek Sun, Rodney A. Bray, Russell Q-Y. Yong, Scott C. Cutmore & Thomas H. Cribb
This section is published as an original article in Systematic Parasitology (2014) 8:141-152
Abstract
A new species of digenean, Pseudobacciger cheneyae n. sp., is described from the intestines of
Weber’s chromis (Chromis weberi Fowler & Bean) from off Lizard Island, Great Barrier Reef,
Australia. This species differs from the three described species of Pseudobacciger Nahhas & Cable,
1964 [P. cablei Madhavi, 1975, P. harengulae (Yamaguti, 1938) and P. manteri Nahhas & Cable,
1964], in combinations of the size of the suckers and the length of the caeca. The host of the present
species is a perciform (Family Pomacentridae) which contrasts with previous records of the genus
which are almost exclusively from clupeiform fishes. The genus Pseudobacciger is presently
recognised within the family Faustulidae (Poche, 1926) but phylogenetic analyses of 28S and ITS2
rDNA show that the new species bears no relationship to species of four other faustulid genera
(Antorchis Linton, 1911, Bacciger Nicoll, 1924, Paradiscogaster Yamaguti, 1934 and
Trigonocryptus Martin, 1958) but that instead it is nested within the Gymnophalloidea (Odhner,
1905) as sister to the Tandanicolidae (Johnston, 1927). This result suggests that the Faustulidae is
polyphyletic.
Introduction
The Faustulidae Poche, 1926 is a small family of digenean trematodes found in a wide range of
mainly marine teleost families (Bray 2008). There are 11 genera, most of which have few species.
Only four genera, Antorchis Linton, 1911, Bacciger Nicoll, 1924, Faustula Poche, 1926 and
Paradiscogaster Yamaguti, 1934, have five or more described species. In Australia, faustulids have
been reported from several fish families in tropical, sub-tropical and temperate waters (Korotaeva
1969; Kurochkin 1970; Bray 1982; Bray et al. 1994; Cribb et al. 1999; Diaz and Cribb 2013; Diaz
et al. 2013).

44
Pseudobacciger Nahhas & Cable, 1964, as reviewed by Dimitrov et al. (1999), is
represented by three species, all infecting marine teleosts. The hosts are overwhelmingly
clupeiforms (Clupeidae and Engraulidae), with single records from a sparid and a pomacanthid
(Madhavi 1975; Parukhin 1976a; Gaevskaja and Naidenova 1996; Dimitrov et al. 1999; Machida
and Uchida 2001). Species of Pseudobacciger primarily infect the intestine, but also occur
occasionally in the pyloric caeca (Dimitrov et al. 1999). There are reports of species of
Pseudobacciger from the temperate to subtropical Atlantic, Pacific and Indian Oceans (Bray 2008)
but to date there are no records from Australian waters. Sporocysts have been reported in bivalves
and metacercariae in shrimps from Japan and Korea (Chun et al. 1981; Kim and Chun 1984).
Pseudobacciger was previously included in the subfamily Baccigerinae Yamaguti, 1958 (family
Fellodistomidae Nicoll, 1909) but molecular analysis by Hall et al. (1999) revealed the
Fellodistomidae to be polyphyletic, and the Baccigerinae was raised to family level as its senior
synonym, Faustulidae.
Pomacentrids have been extensively examined for trematodes in Australia (Manter 1969b;
Cribb et al. 1992; Bray et al. 1993a; Bray et al. 1993b; Bray and Cribb 1998; Bray and Cribb 2004;
Hunter and Cribb 2012). To date 20 fully identified species from eight families of trematodes have
been reported, but no faustulids have been recorded from pomacentrids from Australia or
elsewhere. Between July 2011 and August 2012, several specimens of a planktivorous damselfish,
Weber’s chromis (Chromis weberi Fowler & Bean), were caught from off Lizard Island, on the
northern GBR and examined for trematodes. In these we discovered a previously unreported
trematode, of which morphological analysis suggests is a new species of Pseudobacciger.
Molecular phylogenetic analysis suggests that the family-level affiliation of this new species is not
clear.
Materials and methods
Specimen collection
Fifteen Chromis weberi were collected from off Lizard Island, Great Barrier Reef, Australia in 2011
and 2012. Fish were collected using a barrier net and were killed prior to dissection by cranial
pithing. The contents of the body cavity were removed and examined under a dissecting microscope
using fine scissors and forceps. The gastrointestinal tract was examined for parasites using the gut-
wash approach as described by Cribb and Bray (2010). Specimens were fixed by pipetting them into
near boiling saline followed by immediate preservation in 70 % ethanol for both morphological and
molecular analysis. Worms for morphological analysis were stained using Mayer’s haematoxylin,
destained in 1 % hydrochloric acid, neutralized with 0.5 % ammonia solution, dehydrated through a

45
graded series of ethanol, cleared in methyl salicylate, and mounted on slides in Canada balsam.
Drawings were made with the aid of a camera lucida. Measurements were made using a SPOT
Insight™ digital camera (Diagnostic Instruments, Inc.) mounted on an Olympus BH-2 compound
microscope using SPOT™ imaging software. Measurements are in micrometres (µm) and are
presented as the range, followed by the mean in parentheses. Type-specimens are lodged in the
Queensland Museum (QM), Brisbane, Australia.
Molecular analysis
All voucher specimens are lodged in the Queensland Museum, Brisbane, Australia and are
paragenophores (sensu Pleijel et al. 2008). Total genomic DNA was extracted using standard
phenol/chloroform extraction techniques (Sambrook and Russell 2001). Partial 28S nuclear
ribosomal DNA was amplified using the primers LSU5 (5'-TAG GTC GAC CCG CTG AAY TTA
AGC-3') (Littlewood et al. 2000) and ECD2 (5'-CTT GGT CCG TGT TTC AAG ACG GG-3')
(Littlewood et al. 2000) and the ITS2 region using the primers 3S (5'-GGT ACC GGT GGA TCA
CGT GGC TAG TG-3') (Bowles et al. 1993) and ITS2.2 (5'-CCT GGT TAG TTT CTT TTC CTC
CGC-3' ) (Cribb et al. 1998). PCR for both the 28S and ITS2 regions was performed with a total
volume of 20 μl consisting of approximately 10 ng of DNA, 5 μl of 5 MyTaq Reaction Buffer
(Bioline), 0.75 μl of each primer (10 pmols) and 0.25 μl of Taq DNA polymerase (Bioline
MyTaq™ DNA Polymerase), made up to 20 μl with Invitrogen™ ultraPURE™ distilled water.
Amplification was carried out on a MJ Research PTC-150 thermocycler. The following profile was
used to amplify the 28S region: an initial 95°C denaturation for 4 min, followed by 30 cycles of
95°C denaturation for 1 min, 56°C annealing for 1 min, 72°C extension for 2 min, followed by a
single cycle of 95°C denaturation for 1 min, 55°C annealing for 45 sec and a final 72°C extension
for 4 min. The following profile was used to amplify the ITS2 region: an initial single cycle of 95°C
denaturation for 3 min, 45°C annealing for 2 min, 72°C extension for 90 sec, followed by 4 cycles
of 95°C denaturation for 45 sec, 50°C annealing for 45 sec, 72°C extension for 90 sec, followed by
30 cycles of 95°C denaturation for 20 sec, 52°C annealing for 20 sec, 72°C extension for 90 sec,
followed by a final 72°C extension for 5 min. Amplified DNA was purified using a Bioline
ISOLATE II PCR and Gel Kit, according to the manufacturer’s protocol. Cycle sequencing of
purified DNA was carried out for the 28S rDNA region using the same primers used for PCR
amplification as well as the additional primer 300F (5'-CAA GTA CCG TGA GGG AAA GTT-3')
(Littlewood et al. 2000) and for the ITS2 rDNA region using the forward primer GA1 (5'-AGA
ACA TCG ACA TCT TGA AC-3') (Anderson and Barker 1998) and the reverse primer ITS2.2.
Cycle sequencing was carried out at the Australian Genome Research Facility using an AB3730xl
capillary sequencer. Sequencher™ version 4.5 (GeneCodes Corp.) was used to assemble and edit
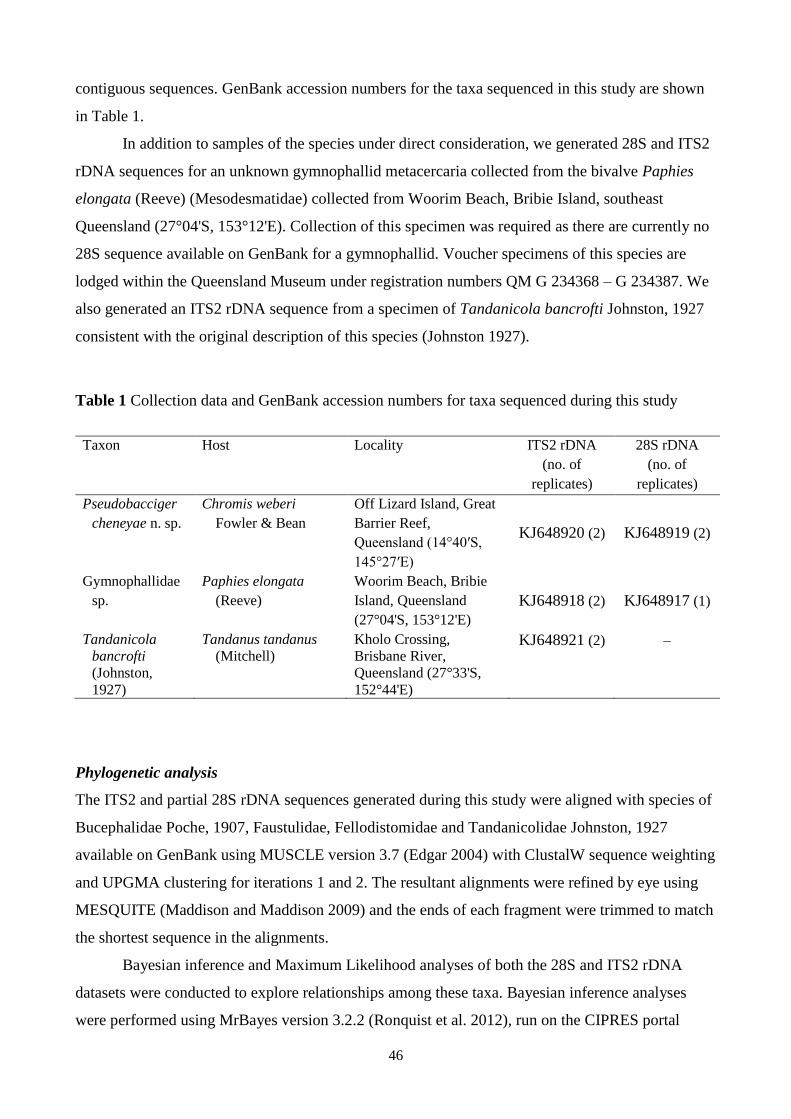
46
contiguous sequences. GenBank accession numbers for the taxa sequenced in this study are shown
in Table 1.
In addition to samples of the species under direct consideration, we generated 28S and ITS2
rDNA sequences for an unknown gymnophallid metacercaria collected from the bivalve Paphies
elongata (Reeve) (Mesodesmatidae) collected from Woorim Beach, Bribie Island, southeast
Queensland (27°04'S, 153°12'E). Collection of this specimen was required as there are currently no
28S sequence available on GenBank for a gymnophallid. Voucher specimens of this species are
lodged within the Queensland Museum under registration numbers QM G 234368 – G 234387. We
also generated an ITS2 rDNA sequence from a specimen of Tandanicola bancrofti Johnston, 1927
consistent with the original description of this species (Johnston 1927).
Table 1 Collection data and GenBank accession numbers for taxa sequenced during this study
Taxon Host Locality ITS2 rDNA
(no. of
replicates)
28S rDNA
(no. of
replicates)
Pseudobacciger
cheneyae n. sp.
Chromis weberi
Fowler & Bean
Off Lizard Island, Great
Barrier Reef,
Queensland (14°40′S,
145°27′E)
KJ648920 (2) KJ648919 (2)
Gymnophallidae
sp.
Paphies elongata
(Reeve)
Woorim Beach, Bribie
Island, Queensland
(27°04'S, 153°12'E)
KJ648918 (2) KJ648917 (1)
Tandanicola
bancrofti
(Johnston,
1927)
Tandanus tandanus
(Mitchell)
Kholo Crossing,
Brisbane River,
Queensland (27°33'S,
152°44'E)
KJ648921 (2)
Phylogenetic analysis
The ITS2 and partial 28S rDNA sequences generated during this study were aligned with species of
Bucephalidae Poche, 1907, Faustulidae, Fellodistomidae and Tandanicolidae Johnston, 1927
available on GenBank using MUSCLE version 3.7 (Edgar 2004) with ClustalW sequence weighting
and UPGMA clustering for iterations 1 and 2. The resultant alignments were refined by eye using
MESQUITE (Maddison and Maddison 2009) and the ends of each fragment were trimmed to match
the shortest sequence in the alignments.
Bayesian inference and Maximum Likelihood analyses of both the 28S and ITS2 rDNA
datasets were conducted to explore relationships among these taxa. Bayesian inference analyses
were performed using MrBayes version 3.2.2 (Ronquist et al. 2012), run on the CIPRES portal

47
(Miller et al. 2010). Maximum Likelihood analyses were performed using RAxML version 7.2.8
(Stamatakis et al. 2008). The software jModelTest version 0.1.1 (Posada 2008) was used to estimate
the best nucleotide substitution model for the datasets. Bayesian inference analyses were conducted
on the 28S dataset using the GTR+G model and on the ITS2 dataset using the TVM+G model
predicted as the best estimators by the Akaike Information Criterion (AIC) in jModelTest. The
closest approximation of these models was implemented in the subsequent Maximum Likelihood
analyses. Bayesian inference analyses were run over 10,000,000 generations (ngen = 10,000,000)
with two runs each containing four simultaneous Markov Chain Monte Carlo (MCMC) chains
(nchains = 4) and every 1,000th tree saved (samplefreq = 1,000). Bayesian analyses used the
following parameters: nst = 6, rates = gamma, ngammacat = 4, and the priors parameters of the
combined dataset were set to ratepr = variable. Samples of substitution model parameters, and tree
and branch lengths were summarised using the parameters ‘sump burnin = 3000’ and ‘sumt burnin
= 3,000’. These ‘burnin’ parameters were chosen because the log likelihood scores ‘stabilised’ well
before 300,000 replicates in the Bayesian inference analyses. Nodal support in the Maximum
Likelihood analyses was estimated by performing 100 bootstrap pseudoreplicates. Species of the
Hemiuridae (Looss, 1899) were designated as functional outgroups in all analyses.
Gymnophalloidea Odhner, 1905 incertae familiae
Pseudobacciger Nahhas & Cable, 1964
Pseudobacciger cheneyae n. sp.
Type-host: Chromis weberi Fowler & Bean (Perciformes: Pomacentridae), Weber’s chromis.
Type-locality: Off Lizard Island, northern Great Barrier Reef, Queensland, Australia (14°40'S,
145°27'E).
Site in host: Intestine.
Material examined: A total of 100 specimens collected from C. weberi; 2 for molecular analysis,
the remainder for morphological analysis.
Prevalence: 73% (11 of 15 fish infected).
Intensity: 1−44 individuals per fish with a mean ( SE) intensity (10 4.128)
Type-material: Holotype QM G 234368 and 19 paratypes QM G 234369–G 234387 deposited in
the Queensland Museum, Brisbane, Australia.
urn:lsid:zoobank.org:act: 369B1E27-B899-48CD-ACD6-0893B314C1C4 urn:lsid:zoobank.org:act: AB0A60B3-E5AC-4EE4-8990-8664A738A1E9

48
Molecular sequence data: 28S rDNA, 2 replicates (1 submitted; GenBank KJ648919); ITS2 rDNA,
2 replicates (1 submitted; GenBank KJ648920).
Etymology: This species is named in honour of Dr Karen Cheney, The University of Queensland, in
recognition of her contribution to marine parasitology.
Description (Figure 1, 2)
[Measurements taken from 20 specimens.] Body pyriform, posterior half somewhat broader than
anterior, 371453 (407) 230327 (281). Forebody, 98138 (119) long, occupies 2334 (28) %
body length. Tegumental spines minute, <1 long, cover entire body evenly, including surfaces of
oral and ventral suckers. Oral sucker well delineated, opening ventro-subterminally, muscular,
6891 4663 (79 57). Ventral sucker well developed, in anterior half of body, 80109
69101 (94 87). Oral sucker length to ventral sucker length ratio 1:1.211.67 (1.47). Oral sucker
width to ventral sucker width ratio 1: 1.031.24 (1.13). Prepharynx absent. Pharynx muscular,
immediately posterior to oral sucker, 3647 2534 (40 29). Oesophagus distinct, often sinuous
or coiling once before intestinal bifurcation, 3355 (43) long. Intestine bifurcates dorsal to ventral
sucker; caeca usually hidden by uterine coils, extend to anterior hindbody, close to posterior
margins of testes, 115166 (133) long (Figure 2). Testes 2, entire, nearly round or somewhat
irregular in outline, opposite in anterior hindbody, intercaecal, well-separated from lateral margins
of body, 4781 5181 (65 70). Sperm ducts inconspicuous. Cirrus-sac absent. Seminal vesicle
bipartite, median, posterior to caecal bifurcation, reaching anteriorly from level of posterior margin
of ventral sucker, 2671 3270 (43 47). Pars prostatica and ejaculatory ducts not detected.
Genital pore median, immediately anterior to margin of ventral sucker. Ovary trilobed, median,
between testes, posterior margin usually hidden by uterine coils, extends to anterior hindbody,
4473 5596 (54 75). Oviduct and oötype inconspicuous. Laurer’s canal opening dorsally
towards posterior end of body (usually obscured by eggs, seen clearly in only a single laterally
mounted specimen). Seminal receptacle not detected. Uterine coils extensive in posterior half of
body from posterior margin of vitelline follicles and vitelline duct, and overlapping testes, ovary
and seminal vesicle. Vitelline follicles in 2 small lateral fields at level of ventral sucker, 64100
3373 (84 47). Vitelline ducts undulate, unite medially anterior to ovary and posterior to seminal
vesicle, straight length 82115 (96), total length 89143 (115). Eggs elliptical, 1821 1215 (20
13.5). Excretory vesicle Y-shaped; arms extend to level of anterior margin of ventral sucker,
243326 (286) long. Excretory pore terminal.
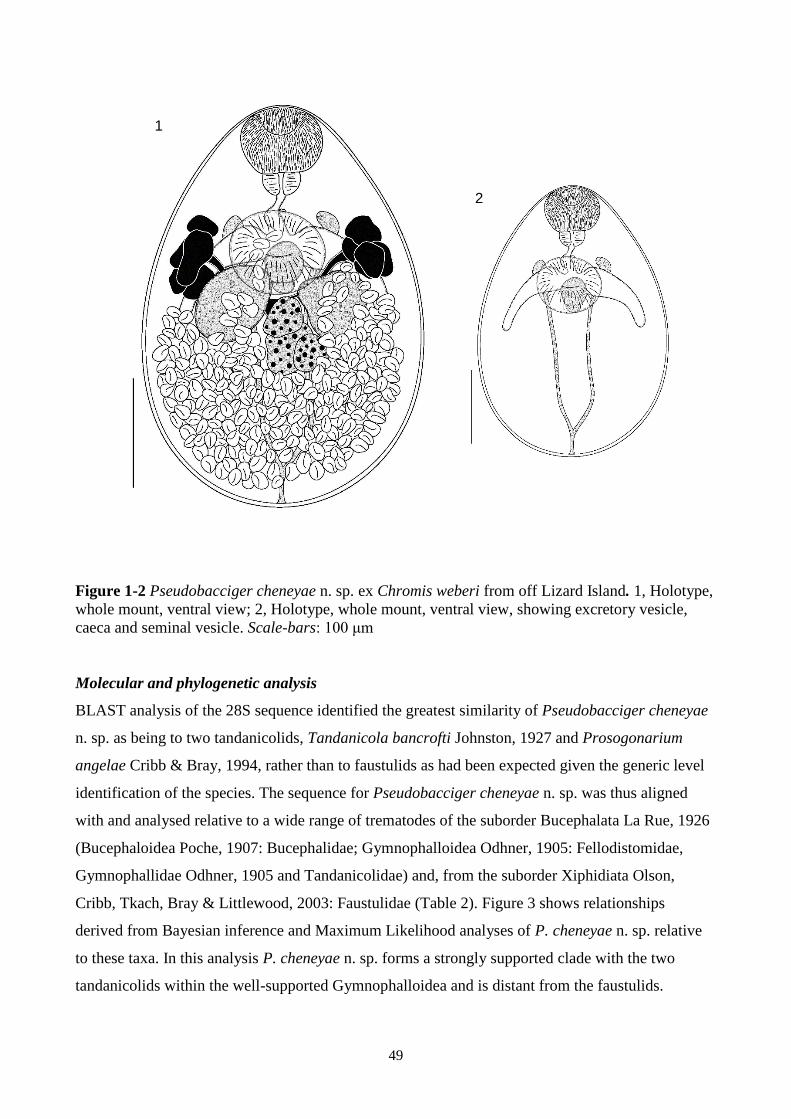
49
2
Figure 1-2 Pseudobacciger cheneyae n. sp. ex Chromis weberi from off Lizard Island. 1, Holotype,
whole mount, ventral view; 2, Holotype, whole mount, ventral view, showing excretory vesicle,
caeca and seminal vesicle. Scale-bars: 100 μm
Molecular and phylogenetic analysis
BLAST analysis of the 28S sequence identified the greatest similarity of Pseudobacciger cheneyae
n. sp. as being to two tandanicolids, Tandanicola bancrofti Johnston, 1927 and Prosogonarium
angelae Cribb & Bray, 1994, rather than to faustulids as had been expected given the generic level
identification of the species. The sequence for Pseudobacciger cheneyae n. sp. was thus aligned
with and analysed relative to a wide range of trematodes of the suborder Bucephalata La Rue, 1926
(Bucephaloidea Poche, 1907: Bucephalidae; Gymnophalloidea Odhner, 1905: Fellodistomidae,
Gymnophallidae Odhner, 1905 and Tandanicolidae) and, from the suborder Xiphidiata Olson,
Cribb, Tkach, Bray & Littlewood, 2003: Faustulidae (Table 2). Figure 3 shows relationships
derived from Bayesian inference and Maximum Likelihood analyses of P. cheneyae n. sp. relative
to these taxa. In this analysis P. cheneyae n. sp. forms a strongly supported clade with the two
tandanicolids within the well-supported Gymnophalloidea and is distant from the faustulids.
1
50
BLAST analysis of the ITS2 sequence identified the greatest similarity of P. cheneyae n. sp.
as being to a range of bucephalids. Given this, and the results of the 28S rDNA analyses, the
sequence was aligned with and analysed relative to a wide range of trematodes of the suborder
Bucephalata (Bucephaloidea: Bucephalidae; Gymnophalloidea: Fellodistomidae, Gymnophallidae
and Tandanicolidae) and, as above, from the suborder Xiphidiata, representatives of Faustulidae. In
all analyses P. cheneyae n. sp. formed clades with a range of taxa from the Bucephalata; it was
never associated with the clade comprising faustulids. However, posterior probability and bootstrap
support were typically low and these analyses are not therefore shown.
Table 2 28S rDNA sequence data from GenBank included in phylogenetic analyses
Species GenBank
accession
number
Reference
Family Fellodistomidae
Coomera brayi Dove & Cribb, 1995 KJ425462 Cribb et al. (2014a)
Fellodistomum agnotum Nicoll, 1909 AJ405289 Bray et al. (1999)
Fellodistomum fellis (Olsson, 1868) AJ405290 Bray et al. (1999)
Oceroma praecox Cribb, Miller, Bray & Cutmore, 2014 KJ425464 Cribb et al. (2014a)
Olssonium turneri Bray & Gibson, 1980 AY222283 Olson et al. (2003)
Steringophorus blackeri Bray, 1973 AJ405296 Bray et al. (1999)
Steringophorus dorsolineatus (Reimer, 1985) AJ405291 Bray et al. (1999)
Steringophorus furciger (Olsson, 1867) AJ405292 Bray et al. (1999)
Steringophorus margolisi Bray, 1995 AY222281 Olson et al. (2003)
Steringophorus pritchardae (Campbell, 1975) AJ405295 Bray et al. (1999)
Steringophorus thulini Bray & Gibson, 1980 AJ405298 Bray et al. (1999)
Tergestia sp. KJ425467 Cribb et al. (2014a)
Family Tandanicolidae
Prosogonarium angelae Cribb & Bray, 1994 AY222285 Olson et al. (2003)
Tandanicola bancrofti Johnston, 1927 KJ425466 Cribb et al. (2014a)
Family Bucephalidae
Rhipidocotyle minima (Wagener, 1852)* AY222225 Olson et al. (2003)
Family Faustulidae
Antorchis pomacanthi (Hafeezullah & Siddiqi, 1970) AY222268 Olson et al. (2003)
Bacciger lesteri Bray, 1982 AY222269 Olson et al. (2003)
Trigonocryptus conus Martin, 1958 AY222270 Olson et al. (2003)
Outgroup Taxa
Superfamily Hemiuroidea
Aponurus sp. DQ354368 Pankov et al. (2006)
Bunocotyle progenetica Chabaud & Buttner, 1959 DQ354365 Pankov et al. (2006)
Robinia aurata Pankov, Webster, Blasco-Costa, Gibson,
Littlewood & Kostadinova, 2006
DQ354367 Pankov et al. (2006)
*Bartoli et al. (2006) re-identified this material as R. minima (Wagener, 1852) not R. galeata (Rudolphi,
1819) as originally identified. R. galeata, the type-species of the genus, was poorly known until redescribed
by Derbel et al. (2011), who showed that it is clearly distinct from R. minima.

51
Figure 3 Relationships between Pseudobacciger cheneyae n. sp. and other Gymnophalloidea,
Bucephalidae and Faustulidae taxa based on Bayesian inference and Maximum Likelihood analyses
of the 28S rDNA dataset. Bayesian inference posterior probabilities are shown above the nodes and
Maximum Likelihood bootstrap support values below the nodes; values < 50 not shown.
Abbreviations: Bu, Bucephalidae; Faustul, Faustulidae; Gy, Gymnophallidae; Tand,
Tandanicolidae.
Discussion
Taxonomy
The present species possesses features that are fully consistent with species of the faustulid genus
Pseudobacciger as recognised by Bray (2008). The present species has the same general
organization and, in particular, possesses the naked two-chambered seminal vesicle reported for that
genus. As reviewed by Dimitrov et al. (1999), Pseudobacciger presently is represented by just three
species: Pseudobacciger cablei Madhavi, 1975, P. manteri Nahhas & Cable, 1964 and P.
harengulae (Yamaguti, 1938). The present form is distinguished from two of these species as
follows. Pseudobacciger cablei is reported as a distinctly larger worm (528704 μm long vs
371453 μm for the present material) in which the suckers (especially the oral sucker) are actually

52
smaller than in the present form (Madhavi 1975). Pseudobacciger cablei differs further from the
new species in having intestinal caeca that terminate close to the anterior margins of the testes
rather than towards the posterior margin, and in having vitelline follicles that reach relatively close
to the anterior end of the body. Finally, P. cablei differs from the present form in being reported as
having the genital pore well anterior to the ventral sucker as opposed to immediately anterior to it.
Pseudobacciger manteri is reported at about the same overall size as the present material (373747
μmlong vs 371453 μm for the present material), but possesses a distinctly smaller oral sucker and
caeca that terminate well posterior to the testes (Nahhas and Cable 1964).
The present material is most similar to the type-species of Pseudobacciger, P. harengulae,
as originally described by Yamaguti (1938) from the clupeid Sardinella zunasi (Bleeker) (as
Harengula zunasi), and reported several times subsequently as reviewed by Dimitrov et al. (1999)
[We are doubtful that the form reported from a pomacanthid by Machida and Uchida (2001) as P.
harengulae can be conspecific with the species reported originally by Yamaguti (1938); see further
comments below]. The overall size and shape of P. harengulae as reported in those studies and the
present form are comparable as are the relative arrangements of their digestive systems and
reproductive organs, but we detect a difference, however, in the larger size of the suckers of the
present form: both oral and ventral suckers have size ranges either completely non-overlapping with
those reported for P. harengulae (see Gaevskaja 1996; Dimitrov et al. 1999) or overlapping with a
higher range (Yamaguti 1938). In particular, the ventral sucker of the present form has a length of
80109 μm, in contrast to 5885 μm in diameter as reported by Yamaguti (1938). Yamaguti’s
specimens appear to have been slightly flattened and it might thus be expected that the size of the
suckers would be inflated whereas they are distinctly smaller than reported here. Machida and
Uchida (2001) identified specimens from the pomacanthid Genicanthus lamarck Lacépède as P.
harengulae. These are much larger (9201,270 670890 μm) than previously described for this
species (and indeed for the genus), with proportionally larger dimensions for most characters,
including suckers, pharynx, oesophagus, gonads and eggs. In addition the forebody is relatively
longer than that of the new species (3545 vs 2334% of body length). Machida and Uchida (2001)
noted that the ‘variation with P. harengulae [as extended by their description] seems to make the
status of each of the three species rather obscure’. We think it is unlikely that Machida and Uchida’s
(2001) worms are conspecific with the tiny worm reported earlier from a clupeiform by Yamaguti
(1938)
Our conclusions regarding the identity of the present form are influenced by our
understanding of host-specificity of these parasites. Miller et al. (2011a) analysed the reported host-
specificity of 290 trematodes known from fishes of the Great Barrier Reef. Of these, just four had
been reported from more than one order of fishes and for none of the four had the wide specificity

53
been tested by molecular data. The high prevalence and intensity reported in the present study
suggest strongly that the infection of Chromis weberi is in no way accidental yet P. cheneyae n. sp.
has not been seen in a wide range of other pomacentrid species examined on the GBR. The three
presently-recognised species of Pseudobacciger are reported almost entirely from clupeiform
fishes; the exceptions are reports of P. harengulae from a herbivorous sparid (Perciformes, the
Bogue Boops boops Linnaeus) by Parukhin (1976a) and from a planktivorous pomacanthid
(Perciformes, Lamarck’s Angelfish Genicanthus lamarck) by Machida and Uchida (2001). For the
present form to occur widely in clupeiforms but in just one of many examined species of
Pomacentridae (Perciformes) seems highly anomalous. We thus think that the apparent distinctions
between the host of the present species and those reported previously for species of Pseudobacciger
are a likely indication that the present form is specifically distinct.
We consider taxonomic decision making in this genus to be unusually difficult. Species of
Pseudobacciger are usually exceptionally small (of the more than 300 species of trematodes
reported from fishes of the GBR, only the bivesiculid Paucivitellosus fragilis Coil, Reid & Kuntz,
1965 is as small as the new species (Pearson 1968), and important morphological features tend to be
obscured by eggs. We note that the analysis of Dimitrov et al. (1999) implied that P. harengulae is
an essentially cosmopolitan species given that there are records from Japan, Korea, the Atlantic and
Indian Oceans and the Black Sea. Although this is certainly possible, as demonstrated by the
molecular evidence that the aporocotylid Cardicola forsteri Cribb, Daintith & Munday, 2000 is
cosmopolitan (Aiken et al. 2007), such widespread species appear to be exceptional. We think it
possible, perhaps likely, that Pseudobacciger will prove to comprise a complex of significantly
more species than are presently recognised. Such a conclusion will almost certainly depend on the
generation of molecular data which, unfortunately, are available only for the present species so far.
In the light of analysis of the morphology and host-specificity of the present form we conclude that
it is best treated as a new species. However, the new name is proposed with a heightened sense that
all such proposals are hypotheses open to testing.
Systematics
The classification of the present form raises a systematic problem in that although its morphology
makes it convincing as a species of Pseudobacciger, it has been shown to be phylogenetically
remote from the representatives of four faustulid genera for which 28S or ITS2 rDNA sequence
data are available. Instead, its position, as can best be judged on the basis of molecular data, is
robustly within the Gymnophalloidea and as sister taxon to species of the Tandanicolidae. The
Tandanicolidae is a small family with species distinctive especially in the form of the terminal
genitalia and in usually infecting both marine and freshwater siluriforms (catfish) (Cribb and Bray

54
1994; Bray 2001). The present form certainly cannot be readily recognised in that family. The
gymnophallid metacercaria sequenced for this study formed the sister taxon to Pseudobacciger +
Tandanicolidae [We note that its identity as a gymnophallid was confirmed by its nesting among
several other identified gymnophallids in the analyses of ITS2 rDNA sequence data].
Gymnophallids are essentially parasites of birds (Scholz 2002) so that, although the present form is
very small, like most gymnophallids, it would have been a surprise if, of all the gymnophalloids, P.
cheneyae had proven most closely related to that family. Our analyses incorporated six genera of
Fellodistomidae but these form a well-supported clade sister to the Gymnophallidae +
(Pseudobacciger + Tandanicolidae). In addition to the Fellodistomidae, Gymnophallidae and
Tandanicolidae, Bray (2002) included the Botulisaccidae Yamaguti, 1971 and the Callodistomidae
Odhner, 1910 in the Gymnophalloidea; the latter two were included “for convenience of
identification”. No molecular data are available for the former family, but the sole member of this
monotypical family does not resemble Pseudobacciger cheneyae n. sp. The Callodistomidae, as
represented by the species Prosthenhystera obesa (Diesing, 1850) was found, utilising 28S rDNA,
to be the sister group to the Allocreadiidae Looss, 1902 within the superfamily Gorgoderoidea
Looss, 1901 by Curran et al. (2006). Phylogenetically, therefore, P. cheneyae n. sp. can thus be best
considered as Gymnophalloidea incertae sedis with the possibility that it may ultimately require a
family separate from those presently recognised. We speculate, thus, that the Faustulidae as
presently conceived may require division into two families, one related to the Zoogonidae as
presently recognised from the analysis of Olson et al. (2003) and a separate family within the
Gymnophalloidea.
Potential division of the Faustulidae has implications for the understanding of morphology
in the group. We observe that there is a potentially relevant dichotomy in the anatomy of the genera
presently recognised in the Faustulidae. Species of Allofellodistomum Yamaguti, 1971,
Pronoprymna Poche, 1926, Pseudobacciger and Pseudosellacotyle Yamaguti, 1953 have cirrus-
sacs or naked terminal genitalia with a very small pars prostatica whereas species of Antorchis,
Bacciger (but we note some variation within this genus), Echinobreviceca Dronen, Blend &
McEachran, 1994, Faustula, Paradiscogaster, Parayamagutia Machida, 1971, Trigonocryptus
Martin, 1958 and Yamagutia Srivastava, 1937 all have a cirrus-sac enclosing a large, prominent
pars prostatica. The three faustulid genera for which 28S rDNA sequences were previously
available (Antorchis, Bacciger and Trigonocryptus) and the single genus for which ITS2 rDNA
sequences were available (Paradiscogaster) are all of the latter type whereas P. cheneyae n. sp. is
of the former type. This difference may form a morphological basis for the recognition of separate
families. Notably, there are already two family group names, Pseudosellacotylinae Yamaguti, 1958
and Pentagramminae Yamaguti, 1958, which might be raised and applied to a distinct family within

55
the Gymnophalloidea. Although we consider the evidence too preliminary to justify a formal
proposal relating to family names at this stage, we feel unable to continue to recognise
Pseudobacciger in the Faustulidae. We recognise that the morphological similarity between P.
cheneyae n. sp. and other Pseudobacciger spp. may be convergent, but the similarity is so striking
that this seems unlikely.
Potential division of the Faustulidae also has implications for the understanding of life-
cycles in the group. A close relationship of the Faustulidae with the Zoogonidae has been
previously intriguing in that all the many known zoogonid life-cycles involve gastropods as first
intermediate hosts as recently reviewed by Barnett et al. (2014) whereas the handful of known
faustulid cycles (species of Bacciger and Pseudobacciger) involve bivalves as first intermediate
hosts (see Kim and Chun 1984; Bartoli and Gibson 2007). Should these species of Bacciger and
Pseudobacciger prove to form a clade within the Gymnophalloidea then that relationship would be
consistent with the use of bivalves by all known members of that clade, as well as the sister taxon
Bucephalidae. Such an outcome would also lead to the prediction that the Faustulidae that are
related to the Zoogonidae may prove to have gastropod rather than bivalve first intermediate hosts.
Ecology of damselfish and their parasites
The Pomacentridae is one of the most prominent families of fishes on the GBR with 79 species
known from the region (Randall et al. 1997). Pomacentrids are found mainly on shallow coral reefs,
but a few species have been recorded at depths of 80 metres or more. Despite this family being
diverse, occupying a range of habitats (live and dead coral, sand, coral rubble and the water-
column) (Wilson et al. 2008), having varying dietary preferences (herbivorous, omnivorous, and
planktivorous) (Randall et al. 1997), and a variety of social habits (social or solitary) (Fishelson
1998), the trematode fauna of this family is considered depauperate relative to some other fish
families (Barker et al. 1994). Although, pomacentrids have been extensively examined for
trematode parasites on the Great Barrier Reef (Manter 1969b; Cribb et al. 1992; Bray et al. 1993a;
Bray et al. 1993b; Bray and Cribb 1998; Bray and Cribb 2004; Hunter and Cribb 2012), no
gymnophalloid taxa have been recorded previously.
Trematodes of pomacentrids in Australian waters demonstrate a wide spectrum of patterns
of host specificity. Several species are evidently euryxenic (infecting taxa related only by ecology).
In this category are Hysterolecitha nahaensis Yamaguti, 1942, Lecithocladium moretonense Bray &
Cribb, 2004, Lepotrema clavatum Ozaki, 1932 and Thulinia microrchis (Yamaguti 1934) which
have all been reported from non-pomacentrids in addition to pomacentrids (Bray et al. 1993a;
Machida and Uchida 2001; Bray and Cribb 2002,2004) although the host range of these species
have not been tested by molecular approaches. Several species are stenoxenic (infecting

56
phylogenetically related host species); the haplosplanchnid Schickhobalotrema pomacentri (Manter,
1937) and the transversotrematid Transversotrema damsella Hunter & Cribb, 2012 have been
reported from multiple pomacentrid species (Cribb et al., 1994; Hunter & Cribb, 2012). Finally, two
species of Lepotrema Ozaki, 1932 (Lepocreadiidae), one species of zoogonid, and one species of
bivesiculid have each been reported from only a single pomacentrid species (Barker et al. 1994;
Cribb et al. 1994a; Bray and Cribb 1998). The specificity of P. cheneyae n. sp. is consistent with
this latter pattern of strict specificity. The ecology and behaviour of C. weberi is similar to that of
other planktivorous damselfish species, such as Abudefduf whitleyi Allen & Robertson, Chromis
ternatensis Bleeker and C. viridis Cuvier, all of which share the habitat of infected C. weberi. It is
thus unclear why P. cheneyae n. sp. should infect only C. weberi. As C. weberi is classified as a
non-territorial fish species (Ormond et al. 1996), it is unlikely that infection is the direct result of a
narrow habitat choice. Host-specificity is often thought of as being generated by encounter and
compatibility filters (Euzet and Combes 1980). In the present case it is not yet possible to determine
which filter might be driving the apparently strict specificity seen here.
Acknowledgements
We thank K. L. Cheney and E. C. McClure for their help collecting specimens and the staff of the
Lizard Island Research Station for their support. This work was funded by an Australian Research
Council Discovery grant awarded to T. H. Cribb and a Sea World Research and Rescue Foundation,
Australia (SWRRFI) awarded to D. Sun and T. H. Cribb.

57
Chapter 3: Presence of cleaner wrasse increases the
recruitment of fishes to coral reefs

58
Chapter 3: Presence of cleaner wrasse increases the recruitment of fishes to
coral reefs
Derek Sun, Karen L. Cheney, Johanna Werminghausen, Mark G. Meekan, Mark I. McCormick,
Thomas H. Cribb & Alexandra S. Grutter
Submitted to a journal for review
Abstract
Mutualisms affect the biodiversity, distribution and abundance of biological communities.
However, ecological processes that drive mutualism-related shifts in population structure are often
unclear and must be examined to elucidate how complex, multi-species mutualistic networks are
formed and structured. In this study, we investigated how the presence of key marine mutualistic
partners can drive the organisation of local communities on coral reefs. The cleaner wrasse,
Labroides dimidiatus, removes ectoparasites and reduces stress hormones for multiple reef fish
species, and their presence on coral reefs increases fish abundance and diversity. Such changes in
population structure could be driven by increased recruitment of larval fish at settlement, or by post-
settlement processes such as modified levels of migration or predation. We conducted a controlled
field experiment to examine the effect of cleaners on recruitment processes of a common group of
reef fishes, and showed that small patch reefs (61–285 m2) with cleaner wrasse had higher
abundances of damselfish recruits, than reefs from which cleaner wrasse had been removed over a
12-year period. However, the presence of cleaner wrasse did not affect species diversity of
damselfish recruits. Our study provides evidence of the ecological processes that underpin changes
in local population structure in the presence of a key mutualistic partner.
Introduction
On coral reefs, cleaning symbioses are key mutualistic networks that increase abundance and
promote species diversity in local fish communities (Grutter et al. 2003; Waldie et al. 2011).
Cleaner organisms, such as the bluestreak cleaner wrasse, Labroides dimidiatus, interact with
multiple species of fish, and provide fitness benefits to their clients by reducing stress hormones

59
(Soares et al. 2011) and ectoparasite loads such as gnathiid isopods (Grutter 1999a). The presence
of cleaner wrasse also affects fish population structure, size-frequency distribution and growth
(Grutter 1999a; Clague et al. 2011; Waldie et al. 2011). Abundance and diversity of fishes, both
adult damselfishes (site-attached) (Bshary 2003; Waldie et al. 2011) and visitors (moving between
patch reefs) (Grutter et al. 2003) increase, as does the abundance of juvenile visitor fishes (Waldie
et al. 2011) on reefs with cleaner wrasse.
Damselfishes (Pomacentridae) are one of the most abundant groups of fishes found on coral
reefs. Damselfishes play a critical role in structuring coral reef benthic communities (Ceccarelli et
al. 2005), with their presence indirectly increasing the survival of juvenile fishes (McCormick and
Meekan 2007). Although the abundance and diversity of adult damselfishes are higher on reefs
where cleaner wrasse are present (Bshary 2003; Waldie et al. 2011), it is not known if this
difference occurs during recruitment, as damselfishes are relatively sedentary after settlement (when
fish first join the reef assemblage after a pelagic larval stage). Whether differences in reef fish
population structure in the presence of cleaning organisms involve regulatory processes such as
migration or predation during settlement, or post-settlement during the juvenile and adult life
stages, has not been investigated. Understanding recruitment, a critical demographic process, is
important as it determines the structure of many marine and terrestrial communities (Caley et al.
1996). In this study, we examined whether the presence of cleaner wrasses influences recruitment
patterns of larval coral reef fishes on patch reefs, which could explain differences in the abundance
and diversity of local assemblages of adult fishes. As previous studies have shown a decrease in
abundance and diversity of reef fishes such as damselfishes and juvenile visitor fishes on reefs
where cleaner wrasse are absent (Bshary 2003; Grutter et al. 2003; Waldie et al. 2011), we predict a
similar trend occurring for damselfish recruits.
Material and methods
Our study was conducted on 16 small patch reefs in shallow (3−7 m depth) areas in lagoon and
back-reef habitats near Lizard Island, GBR, Australia. Since 2000, ‘removal’ reefs (n = 7) were
inspected at 3-month intervals and cleaner wrasse removed with hand and barrier nets; ‘control’
reefs (n = 9) were left undisturbed (see Clague et al. 2011 for a description of reefs; Waldie et al.
2011). The number of cleaner wrasse present on control reefs were recorded.
To investigate whether the abundance and species richness of newly-settled damselfishes
differed between removal and control reefs, we counted young recruits (< 20 mm TL) for five days
in each of three lunar cycles, beginning four days after the new moon (18−22 November 2012,
17−21 December 2012 and 16−20 January 2013). These times were selected because settlement of

60
fishes to coral reefs peak around the time of the new moon during these summer months (Milicich
and Doherty 1994). Counts alternated between randomly-selected control and removal reefs and
fish were identified to species where possible. Fish were counted once per reef by the same
observer (D.S.) on SCUBA for 60−120 min, depending on reef size. Fish were counted by the same
observer to reduce variability. As damselfishes are most active from morning to midday (Bosiger
and McCormick 2014), reefs were surveyed between 0900−1200 hr. The observer circled ~ 1 m
above the reef counting an abundant, or several less-abundant, species (in the same order for each
reef) on each pass. For analysis, fish were categorised either as common, wherein recruits of these
species were found on 90% of reefs across all three months (of which there were five species:
Pomacentrus adelus, P. amboinensis, P. moluccensis, P. nagasakiensis and Chrysiptera rollandi),
or uncommon (all other damselfishes). We compared the abundance and three measures of diversity
of damselfish recruits on control and removal reefs.
We conducted generalized linear mixed effect models (GLMER) with a Poisson distribution
error, to examine whether the total abundance of all the five common damselfishes combined (and
also separately by species) and the total abundance of uncommon damselfishes differed relative to
the presence of cleaner wrasse. The terms treatment (cleaner or no cleaner presence), site (Lagoon
or Casuarina Beach) and time (November 2012, December 2012 and January 2013) were fixed
factors; reef identity was a random factor and reef area was a covariate. A posteriori Tukey-Kramer
HSD post-hoc test was used to interpret significant interactions. We also used full linear mixed-
effect models (LMER) with a REML procedure to analyse species richness and Simpson’s indices
of diversity (Krebs 1999) and used a permutational multivariate analysis of variance
(PERMANOVA) and principle coordinates analysis (PCO) for the species composition. To
determine the best model in all analyses, we compared the full model with models in which one of
the explanatory terms was dropped using the “drop1” function (Chambers 1992). The term was
dropped if the analysis of variance found that a dropped term had no significant effect on the model
using the Chi-square distribution. A Bonferroni correction (α = 0.01) was applied to account for
multiple comparisons. All statistical analyses were conducted using R version 3.0.1 (R
Development Core Team 2013), except for the species composition which was analysed using
PRIMER 6.0 (Anderson et al. 2008).
Results
Thirty-one species of damselfish recruits were identified (see Appendix I). The total abundance of
common damselfishes was lower on removal than on control reefs (p = 0.006, Figure 1). At the
species level, the abundances of C. rollandi (p = 0.0001, December only) and P. amboinensis (p <
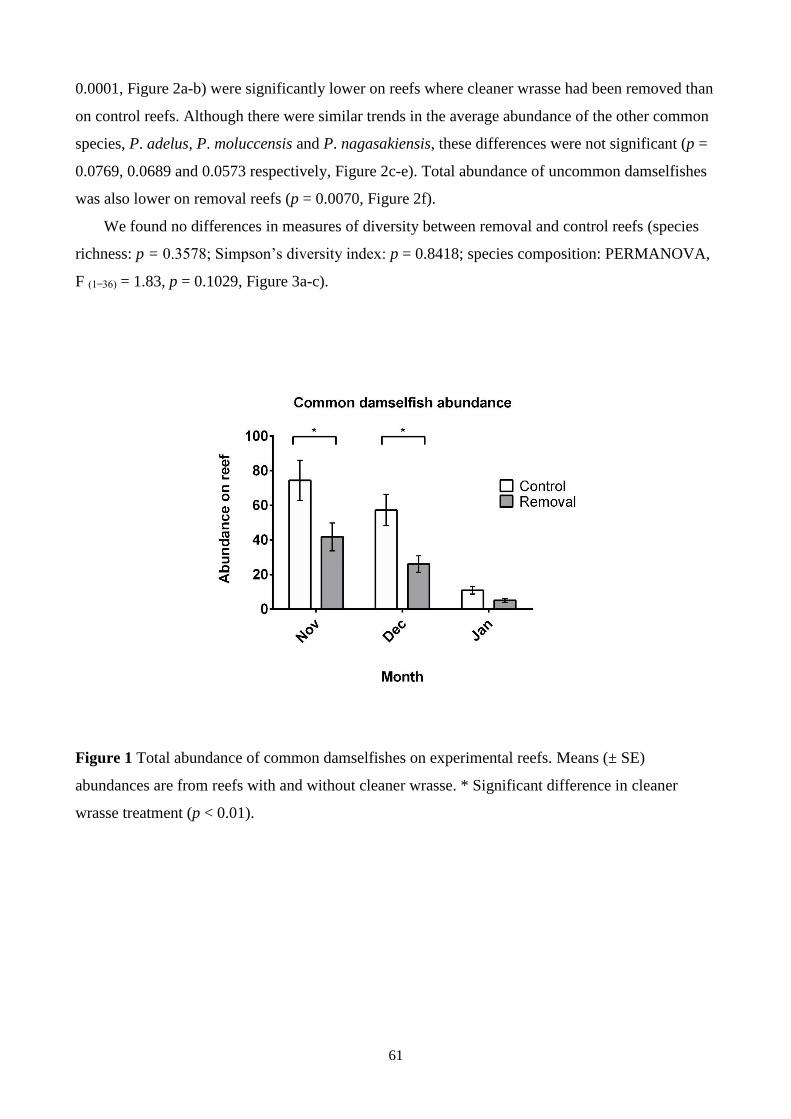
61
0.0001, Figure 2a-b) were significantly lower on reefs where cleaner wrasse had been removed than
on control reefs. Although there were similar trends in the average abundance of the other common
species, P. adelus, P. moluccensis and P. nagasakiensis, these differences were not significant (p =
0.0769, 0.0689 and 0.0573 respectively, Figure 2c-e). Total abundance of uncommon damselfishes
was also lower on removal reefs (p = 0.0070, Figure 2f).
We found no differences in measures of diversity between removal and control reefs (species
richness: p = 0.3578; Simpson’s diversity index: p = 0.8418; species composition: PERMANOVA,
F (1 ̶36) = 1.83, p = 0.1029, Figure 3a-c).
Figure 1 Total abundance of common damselfishes on experimental reefs. Means (± SE)
abundances are from reefs with and without cleaner wrasse. * Significant difference in cleaner
wrasse treatment (p < 0.01).
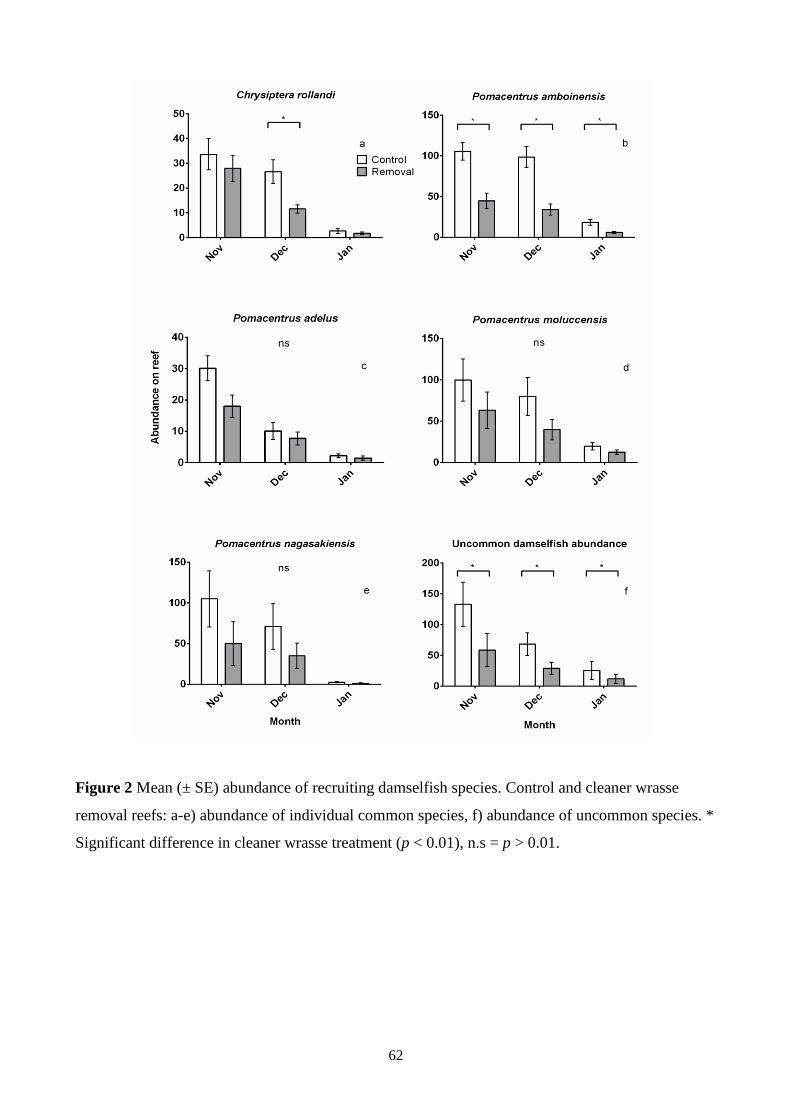
62
Figure 2 Mean (± SE) abundance of recruiting damselfish species. Control and cleaner wrasse
removal reefs: a-e) abundance of individual common species, f) abundance of uncommon species. *
Significant difference in cleaner wrasse treatment (p < 0.01), n.s = p > 0.01.
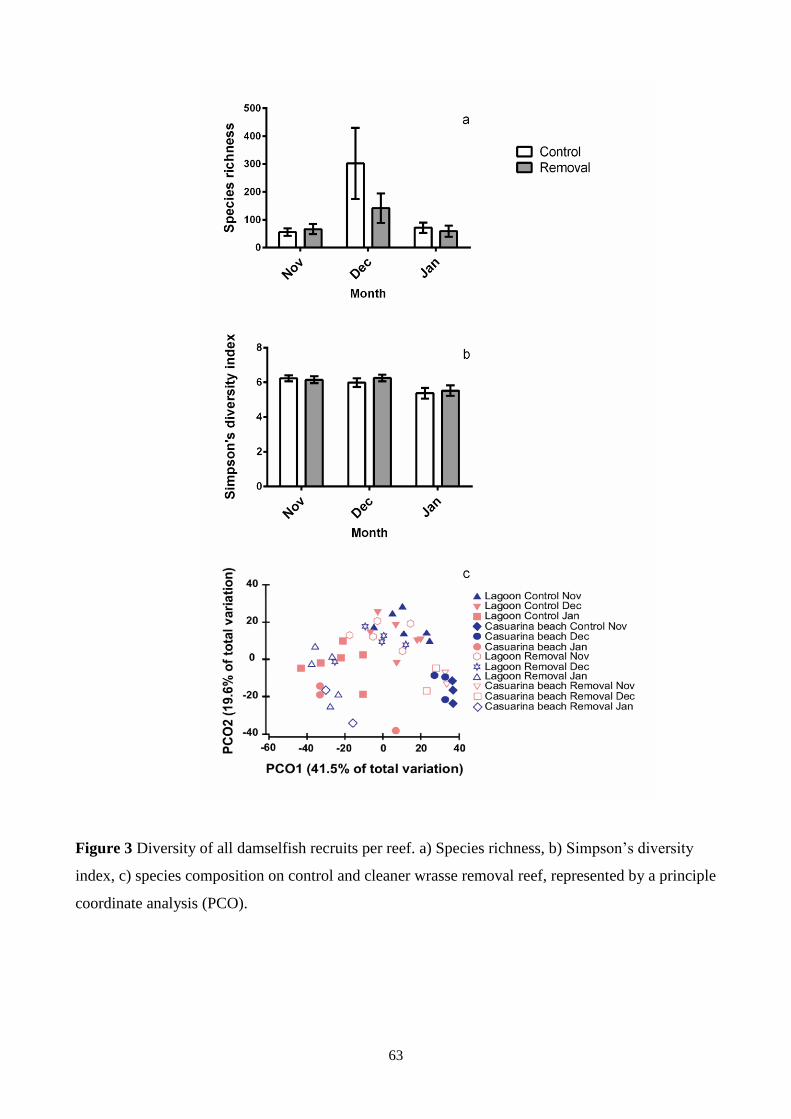
63
Figure 3 Diversity of all damselfish recruits per reef. a) Species richness, b) Simpson’s diversity
index, c) species composition on control and cleaner wrasse removal reef, represented by a principle
coordinate analysis (PCO).

64
Discussion
The combined abundances of the five common species, the individual abundances of two common
species (C. rollandi and P. amboinensis), and the total abundance of uncommon recruited
damselfishes were higher on reefs with cleaner wrasse present than on those without. There was
also a strong (but not statistically significant) trend for abundances of the other common
damselfishes (P. adelus, P. moluccensis and P. nagasakiensis) to be higher on reefs with cleaners.
These results suggest that the presence of a key mutualistic species, creates changes in population
structure around the time of settlement and recruitment. This in turn could explain the observed
changes in abundance and distribution of coral reef fish populations in relation to cleaner wrasse
presence (Waldie et al. 2011).
The presence of cleaner wrasse could influence recruitment processes by a number of direct
or indirect mechanisms. If young fish use visual or olfactory cues to locate potential microhabitats,
then sensory cues may enable them to locate cleaners directly. Indeed, a recent study suggests
recently-settled fish can recognise and preferentially choose a microhabitat near a cleaner (Sun et al.
in review, Chapter 4). Alternatively, cleaner wrasse could influence recruitment processes indirectly
by reducing the abundance of ectoparasites in the local environment. Higher abundances of
parasites in a system have the potential to detrimentally affect the recruitment of juveniles, as
demonstrated in lower recruitment success of juvenile cricket frogs (Acris crepitans) in ponds with
high rates of parasitic infection (Beasley et al. 2005). Although both adults and recruit damselfishes
are known to be infected with gnathiids, cleaner wrasse rarely clean recruits and tend to clean larger
individuals (Sun et al. in review, Chapter 4). However, with gnathiids decreasing swimming
performance and increasing mortality of settling fish (Grutter et al. 2011) the removal of even a
single gnathiid can increase the survival of an individual. Cleaner wrasse may also indirectly
promote fish recruitment by influencing the abundance of conspecifics, which have been shown to
influence microhabitat selection by settling fish, as settling fish may use the presence of
conspecifics as a measure of habitat quality (Coppock et al. 2013) or the behaviour of predators
(Cheney et al. 2008). The abundance of all (adult and juvenile) conspecifics for P. amboinensis, P.
moluccensis and P. nagasakiensis are also higher on reefs with cleaner wrasse than those without
(D. Sun, unpublished data). Further investigation is required to disentangle the relative importance
of these direct and indirect mechanisms. It should be noted that the gradual decrease in the
abundance of damselfish recruits from November to January is due to the larval fish season ending.
Interestingly, there was no difference in species richness, Simpson’s diversity index, or
composition of damselfish recruits between control and removal reefs, suggesting that the presence
of cleaner wrasse affects the abundance of individual species but not the overall community
diversity of recruit damselfishes. Previous studies have shown an increase in the abundance and

65
species richness of resident adult fishes in the presence of cleaner wrasse (Bshary 2003; Waldie et
al. 2011). Our results suggest that such patterns in diversity are not likely due to differential
recruitment, but due to post-settlement events, including migration in response to cleaner wrasse
presence or the effects of parasitism (Waldie et al. 2011). Parasitic infection by gnathiid isopods can
make recruits more susceptible to the high rates of predation that occur immediately after settlement
(typically 56 % within two days of settlement) (Almany and Webster 2006). Ultimately, this
process could affect the diversity of fishes in the presence of cleaner wrasse.
In summary, we have demonstrated for the first time that the presence of cleaner wrasse has
the potential to influence the structure and demography of reef fish populations via effects on the
process of recruitment. Thus, mutualism is an important mechanism that contributes to the already-
complex recruitment processes of reef fishes.
Acknowledgements
We thank Lizard Island Research Station staff. We also like to thank E. C. McClure, M. De
Brauwer for field assistance. S.P. Blomberg for statistical advice and R. Q-Y. Yong for reading
drafts of this manuscript. Funding was provided by Sea World Research and Rescue Foundation,
Australia (SWRRFI) to D.S, T.H.C and A.S.G and an Australian Research Council Discovery Grant
to A.S.G and M.G.M.

66
Chapter 4: Cleaner wrasse influence habitat selection of
young damselfish

67
Chapter 4: Cleaner wrasse influence habitat selection of young damselfish
Derek Sun, Kare L. Cheney, Johanna Werminghausen, Eva C. McClure, Mark G. Meekan, Mark I.
McCormick, Thomas H. Cribb & Alexandra S. Grutter
Submitted to a journal for review
Abstract
The presence of bluestreak cleaner wrasse, Labroides dimidiatus, on coral reefs increases total
abundance and biodiversity of reef fishes. The mechanism(s) that cause such shifts in population
structure are unclear, but it is possible that young fish preferentially settle into microhabitats where
cleaner wrasse are present. To investigate this possibility, we conducted aquarium experiments to
examine whether settlement-stage and young juveniles of ambon damselfish, Pomacentrus
amboinensis, selected a microhabitat near a cleaner wrasse (adult or juvenile). Both settlement-stage
(0 d post-settlement) and juvenile (~ 5 weeks post-settlement) fish spent a greater proportion of
time in a microhabitat adjacent to L. dimidiatus than in one next to a control fish (a non-cleaner
wrasse, Halichoeres melanurus) or one where no fish was present. This suggests that cleaner wrasse
may serve as a positive cue during microhabitat selection. We also conducted focal observations of
cleaner wrasse and counts of nearby damselfishes (1 m radius) to examine whether newly-settled
fish obtained direct benefits, in the form of cleaning services, from being near a cleaner wrasse in
the field. Although abundant, newly-settled recruits (< 20 mm TL) were rarely (2 %) observed
being cleaned per 20 min observations compared with larger damselfishes (58 %). Individual
damselfish that were cleaned were significantly larger than the median size of the surrounding
nearby non-cleaned conspecifics; this was consistent across four species. The selection by
settlement-stage fish of a microhabitat adjacent to cleaner wrasse in the laboratory, despite only
being rarely cleaned, suggests that even rare cleaning events and/or indirect benefits may drive their
settlement choices. This behaviour may also explain the decreased abundance of young fishes on
reefs from where cleaner wrasse had been experimentally removed. This study reinforces the
potentially important role of mutualism during the processes of settlement and recruitment of young
reef fishes.

68
Introduction
On coral reefs there are many mutualistic cleaners that remove parasites from other organisms,
including coral reef fishes. One of the most obvious of these is the bluestreak cleaner wrasse,
Labroides dimidiatus (Labridae), a cleaner wrasse common throughout the Indo-Pacific (Randall et
al. 1997). Labroides dimidiatus has been shown to reduce the ectoparasite load of individual client
fishes (Gorlick et al. 1987; Grutter 1999a; Clague et al. 2011), aggression by predators (Cheney et
al. 2008), stress (measured in cortisol levels) via physical contact (Bshary et al. 2007; Soares et al.
2011) and increase growth and size of client fishes (Clague et al. 2011; Waldie et al. 2011). In
addition, these cleaner wrasse affect local populations of reef fishes, so that in their absence there
are reductions in the diversity and abundance of site-attached adult residents, visitors (fishes that
move between patch reefs) and juvenile visitor fishes (Bshary 2003; Grutter et al. 2003; Waldie et
al. 2011). Although the positive effects of the presence of L. dimidiatus on individuals and
populations have been demonstrated in experimental manipulations of patch reefs ranging in size
from ~61 to 285 m2, the mechanisms involved, particularly at small spatial scales (1 m–10s of m),
remain largely unstudied (Waldie et al. 2011).
One of the most critical periods that effects the population dynamics of coral reef fishes
occurs during the settlement of young fish from the plankton to the reef and the few days or weeks
of life in benthic habitats that immediately follow this event (post-settlement). Processes at this time
ultimately determine which individuals are recruited to the population of juveniles. Young fish can
actively choose the habitat into which they will settle and the processes that underlie these choices
are complex (Hoey and McCormick 2004; Heinlein et al. 2010). Additionally, the mortality rate at
this time is high; indeed, it has been estimated that 57 % of individuals die within the first 1−2 days
of settlement (Almany and Webster 2006), which could be due to microhabitat selection. Thus, the
choice of a suitable microhabitat at settlement is critical for successful recruitment. Selecting the
optimum microhabitat depends on numerous factors, which may include the structure of the
microhabitat (Tolimieri 1995), and the presence of potential predators (Vail and McCormick 2011)
and conspecifics (Sweatman 1985). It was recently shown that the experimental removal of L.
dimidiatus was associated with a decrease in the abundance of new recruits on patch reefs, relative
to control reefs where cleaner wrasse were present (Sun et al., in review, Chapter 3). However, it is
unknown whether the presence of L. dimidiatus can serve as a positive cue for microhabitat choice
at settlement.
Our study used experimental and observational approaches to determine whether the
presence of juvenile or adult cleaner wrasse influenced the choice of microhabitat by settling
juveniles of the ambon damselfish, Pomacentrus amboinensis. In the laboratory, we examined
whether the presence of cleaner wrasse influences habitat choice of P. amboinensis. To determine

69
the extent to which recently-settled damselfishes were cleaned by L. dimidiatus juveniles, and thus
whether the benefits of settling near a cleaner wrasse could be related to the cleaning services they
provide or to other indirect benefits, focal observations of the cleaning interactions between L.
dimidiatus and damselfishes were conducted in the field. Lastly, to determine whether juvenile
cleaner wrasse are size selective in their choice of clients, we compared the size of individuals of
the four most abundant damselfishes that were cleaned by the wrasse with the median size of non-
cleaned conspecifics.
Materials and methods
Laboratory experiment: Does the presence of L. dimidiatus affect microhabitat choice in young
damselfish?
We conducted an aquarium experiment at Lizard Island Research Station, GBR, Australia, to
investigate whether the presence of L. dimidiatus influenced microhabitat choice of P. amboinensis,
a species chosen because of its high abundance at the study site (Sun et al. in review, Chapter 3).
Pomacentrus amboinensis were collected between November 2011 and January 2012 using light-
traps (see Meekan et al. 2001 for design) moored overnight approximately 100 m from the reef.
These fish were juveniles caught just prior to settlement from the plankton and thus had no previous
exposure to L. dimidiatus. Fish were held in groups of ~ 20 in clear plastic holding aquaria (29 × 17
× 12 cm) with a constant flow of seawater. Fish collected in November were held for around 5
weeks (at which point they were classified as juveniles, i.e. ~ 5 weeks post-settlement), whereas
fish collected in January were tested within a day of capture (classified as settlement-stage, i.e. 0
days post-settlement). Fish were fed ad libitum twice a day with live nauplii of brine shrimp
(Artemia sp.).
Both adult and juvenile cleaner wrasse (L. dimidiatus) and a non-cleaner control fish
(pinstripe wrasse, Halichoeres melanurus), were collected from the Lizard Island lagoon using
hand-nets, barrier nets and anaesthetic clove oil mixed with alcohol and seawater (10 % clove oil;
40 % ethanol; 50 % seawater). Halichoeres melanurus was chosen as a control due to similarity in
body shape to L. dimidiatus (Grutter 2004a; Cheney et al. 2008; Cheney et al. 2009). All wrasses
were maintained in aquaria (43 × 32 × 30 cm) with running unfiltered seawater.
Clear Perspex sheets were placed at either end of a glass experimental aquaria (35 × 36 × 65
cm), with flow-through seawater, to create two end compartments (10 cm away from the end of the
aquarium). The middle section of the aquarium was divided into three 15 cm subsections, using
vertical black lines drawn on the front and back of the aquarium. Three polyvinyl chloride (PVC)
tube shelters (6 × 5 cm diameter) were placed in the centre of each subsection (Fig. 1). Aquaria

70
were covered with black plastic on three sides to isolate the fish from external activity, and a green
plastic shade cloth hung in front of the tanks to create an observation blind.
In each end compartment a L. dimidiatus, a control fish (H. melanurus) or no fish was
placed, depending on the treatment: 1) L. dimidiatus and H. melanurus; 2) L. dimidiatus and no
fish; or 3) H. melanurus and no fish. These combinations were presented to a randomly-selected P.
amboinensis ‘client’ and the side in which a cleaner or control (or no fish) was placed was
randomised, but balanced. The ‘no fish’ treatment at one end compartment of the aquarium was
used to control for the ‘client’ potentially avoiding any heterospecifics, irrespective of identity.
A P. amboinensis was placed in the middle section of the aquarium in a bottomless clear
plastic container (8 × 8 × 8 cm) with numerous small (5 mm) ventilation holes for 20 min prior to
commencing the trial, allowing the fish to acclimate to the new surroundings. The bottomless
container was removed using an attached monofilament string by the observer, who was positioned
behind the observation blind. Each trial was conducted for 20 min, and the position of P.
amboinensis within the aquarium (left, middle, or right section) was recorded every 15 s. It is
inferred that the degree of proximity to the stimulus represented its choice of microhabitat.
Experiment 1 was divided into three parts (A, B, C) with different ontogenetic stages of
cleaner, control and P. amboinensis per treatment (see Table 1 for a summary of the fish stage and
species). For each part, we used 8 cleaner and 8 control fish that were randomly selected, and 75 P.
amboinensis (n = 25 individuals per treatment). At the end of each trial, cleaner and control fish
were replaced. Individual P. amboinensis were only used once, so that their behaviour was not
affected by previous exposure.
Part A used adult L. dimidiatus and H. melanurus, and Part B used juveniles of both species.
These experiments tested whether the ontogenetic stage/size of L. dimidiatus affected habitat choice
of juvenile P. amboinensis. Part C was conducted when settlement-stage fish became available in
the light-traps; juvenile L. dimidiatus and H. melanurus were used for this component. The aim of
this experiment, relative to Part B, was to determine whether P. amboinensis’ ontogenetic stage/size
affected its habitat choice.

71
Figure 1 Front view of the laboratory experiment tank design. A: clear Perspex sheet placed on
either side to create compartments for stimuli, B: vertical dashed lines on front and back of
aquarium to create subsections, and C: PVC tube shelters placed in each subsection. Not drawn to
scale.
Table 1 Ontogenetic stage and size of fish used in each part for laboratory experiment. Size is mean
± SE, TL.
Part Date Labroides
dimidiatus
Halichoeres
melanurus
Pomacentrus
amboinensis
(cleaner) (control) (client)
A December 2011 Adult
(79.1 ± 4.0 mm)
Adult
(86.1 ± 0.8 mm)
Juvenile
(19.3± 0.3 mm)
B December 2011 Juvenile
(24.2 ± 0.5 mm)
Juvenile
(24.8 ± 0.4 mm)
Juvenile
(18.0 ± 0.3 mm)
C January 2012 Juvenile
(23.1 ± 0.7 mm)
Juvenile
(24.3 ± 0.8 mm)
Settlement-stage
(15.2 ± 0.1 mm)

72
Field observations: Do juvenile L. dimidiatus clean recently-settled damselfish recruits and are
they selective in their choice of clients?
To assess whether recently-settled damselfish recruits are cleaned by L. dimidiatus, and thus gain
direct fitness benefits from settling near them, we conducted 20 min behavioural observations on
juvenile L. dimidiatus (n = 79) on a section of continuous reef (~ 200 m) at Coconut Beach
(1440S, 14528E), Lizard Island. Although cleaner wrasse of all sizes mainly consume
ectoparasites (Grutter 2000), we observed juvenile L. dimidiatus, rather than adults, due to the
higher frequency of juvenile L. dimidiatus interacting with young reef fish (Robertson 1974, D.
Sun, unpublished data). Before observations commenced, observers learned to accurately estimate
fish size (TL) underwater by using printed and laminated outlines of model fish (12−74 mm TL).
The fish models were placed on the reef at varying distances from the observer, and sizes were
estimated and compared with the actual length of the model until observers were more than 80 %
accurate at estimating length. Rulers printed onto dive slates were also used as a guide for
estimating fish length.
For each observation, a juvenile L. dimidiatus was located and its estimated TL was
recorded. A 1 m radius was measured around the area where the L. dimidiatus was first sighted
using a 1 m string and marked using five lead sinkers (7 mm) attached to floats (25 mm) and all fish
within this area were recorded. All fish present were recorded to account for the availability of other
sizes of fish as potential clients that could influence the cleaners’ choice of client. Surrounding fish
were allowed to acclimate (~ 5 min) to the presence of the markers and observer, before the
commencement of observations. Cleaning interactions were recorded as any physical contact
between cleaner and client, or significant inspection by the cleaner (> 1 s). For each interaction, we
recorded the client species (where possible), cleaning duration, and estimated client size (TL, mm).
To compare client size and the median size of nearby (non-cleaned) conspecifics, the size of all
other fish present within the marked observation site (TL = > 10 to < 80 mm), but not involved in
any cleaning interaction, was also estimated. All other fish were identified to species where
possible. To ensure that each L. dimidiatus was observed only once, observers began at one end of
the reef and continued without backtracking. In contrast to adults, juvenile L. dimidiatus stay within
a highly restricted home range of around 4 m2 (Robertson 1974), further reducing the possibility of
observing an individual twice. Although every individual within the observation area was identified
to species, ultimately, in order to adequately perform statistical analyses, only the most abundant
damselfish species (both cleaned and non-cleaned conspecifics) from the observations were used.
These four common species were Pomacentrus amboinensis, P. moluccensis, P. nagasakiensis and
P. wardi.

73
Statistical analyses
Laboratory experiment
A full linear mixed-effects (LMER) model with a restricted maximum likelihood (REML)
procedure was used to examine whether P. amboinensis spent more time near a L. dimidiatus than
near a control fish or no fish compartment. Two analyses were conducted; the first model used data
from parts A and B (Table 1) to determine whether ontogenetic stage of the cleaner had an effect on
juvenile P. amboinensis’ habitat choice, while the second model used data from part B and C, and
included ontogenetic stage (settlement-stage and juvenile P. amboinensis) of the client as a factor.
In the first model, cleaner ontogenetic stage (adult or juvenile), settlement stimulus type to which
the microhabitat was adjacent (cleaner, control, no fish), aquarium side the cleaner was placed (left
or right) and fish stimulus treatment combination (cleaner/control, cleaner/no fish or control/no
fish) were used as fixed factors. Replicate trial identity was added as a random factor, to account for
an order effect. In both models, proportion of time spent in each section of the aquarium (right, left
and centre) was transformed by taking the arcsine of the square root of the proportion to meet the
assumptions of normality.
Field observations
A LMER model with a REML procedure was used to test whether there was a significant difference
between client fish and the median size of the nearby conspecifics. The terms treatment (client or
nearby conspecifics), nearby conspecific size and species (four damselfish species: P. amboinensis,
P. moluccensis, P. nagasakiensis and P. wardi) were fixed factors, and the identity of the individual
cleaner fish and the cleaning event identity (specifying which client and nearby conspecifics
corresponded with one another) were random factors. To determine whether client size varied with
cleaner size or the size of all other nearby species, separate (LMER) analyses were run with cleaner
size and other nearby fish size as covariates and cleaner identity as a random factor. A general
linear model (GLM) with a binomial distribution was used to determine whether there was a
significant difference between recruits that were cleaned and the abundance of other fish species
and recruits within the 1 m radius observation area.
To determine the best model in all analyses, we compared the full model with models in
which one of the explanatory terms was dropped using the ‘drop1’ function (Chambers 1992). The
term was dropped if the analysis of variance found that a dropped term had no significant effect on
the model using the Chi-square distribution. A Tukey-Kramer HSD test (TK-HSD) using the glht
function in the ‘multcomp’ package (Horthorn et al. 2013) identified where differences occurred
and a summary output table was reconstructed using the results from the ‘drop1’ function. Prior to
all analyses, the assumptions of normality and homogeneity of variance were assessed using

74
histograms, residuals and quantile-quantile plots. All statistical analyses used R version 3.0.1 (R
Development Core Team 2013).
Results
Laboratory experiment: Does the presence of L. dimidiatus affect microhabitat choice in young
damselfish?
In all three parts (A-C) of the experiment, P. amboinensis spent significantly more time in the
subsection or PVC tube that is adjacent to a L. dimidiatus than next to a control fish or no fish
compartment (Fig. 2a-c). The first statistical analysis, which used data from parts A and B to
examine whether cleaner ontogenetic stage had an effect, showed that the proportion of time that
juvenile P. amboinensis spent next to the stimulus fish (cleaner, control or no fish) differed
according to an interaction between fish stimulus and cleaner stage (LRT = 19.98, df = 2, P <
0.0001; Fig. 2a, b); a TK-HSD test showed that juvenile P. amboinensis spent significantly more
time near a juvenile than an adult L. dimidiatus (P < 0.05). There was no significant effect of
treatment combination (LRT = 0.474, df = 2, P = 0.788) or aquarium side (LRT = - 4.18, df = 1, P =
0.772).
For the second statistical analysis, which used data from parts B and C to examine whether
ontogenetic stage of the client affected habitat choice, the proportion of time that P. amboinensis
spent next to a habitat did not differ with stage (LRT = - 4.40, df = 1, P = 0.749) but did differ
according to fish stimulus (LRT = 144.22, df = 2, P < 0.0001; Fig. 2a, c); a TK-HSD test showed
that P. amboinensis spent significantly more time near L. dimidiatus (P < 0.05). The proportion of
time spent in a habitat differed according to treatment combination (LRT = 37.11, df = 2, P <
0.0001). A TK-HSD test showed that P. amboinensis spent more time in both chosen microhabitats
(i.e. spent less time in the middle section) in the cleaner/no fish and control/no fish stimulus
treatment combinations than in the cleaner/control treatment. The effect of aquarium side was not
significant (LRT = -1.44, df = 1, P = 0.182). There was no significant effect of size between cleaner
and control wrasse (> 0.05).
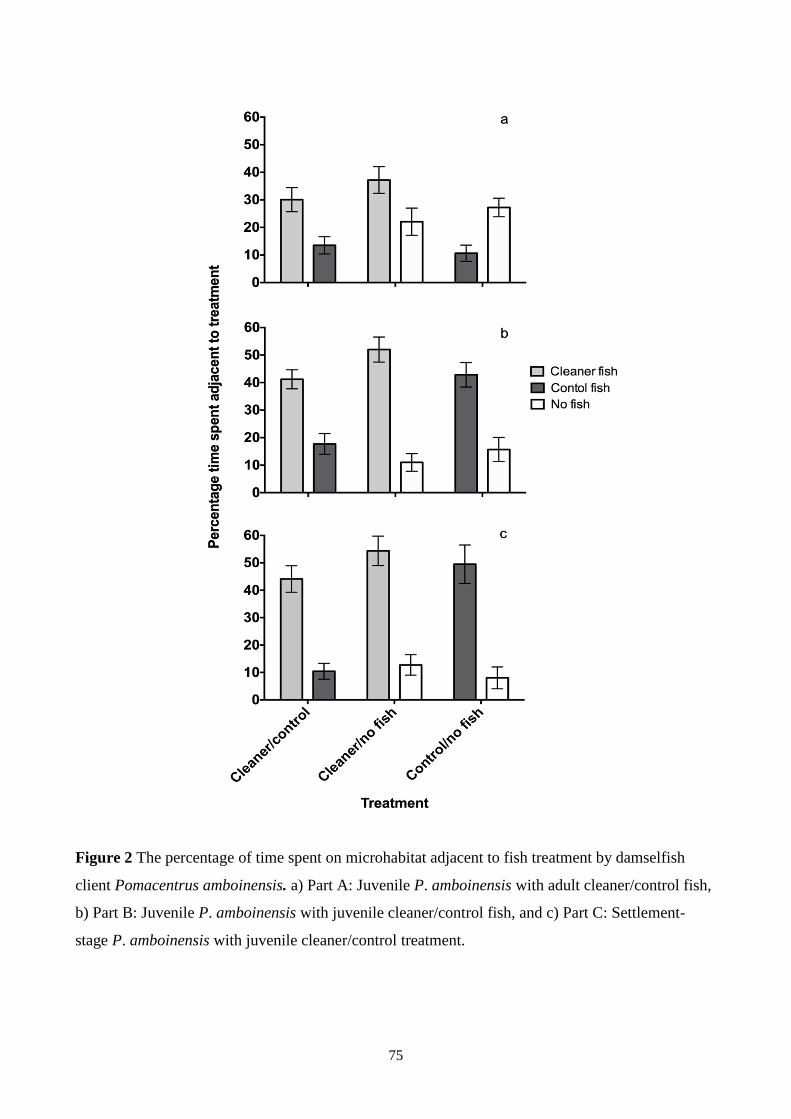
75
Figure 2 The percentage of time spent on microhabitat adjacent to fish treatment by damselfish
client Pomacentrus amboinensis. a) Part A: Juvenile P. amboinensis with adult cleaner/control fish,
b) Part B: Juvenile P. amboinensis with juvenile cleaner/control fish, and c) Part C: Settlement-
stage P. amboinensis with juvenile cleaner/control treatment.

76
Field observation: Do juvenile L. dimidiatus clean recently-settled damselfish recruits and are
they selective in their choice of clients?
A total of 1107 cleaning interactions were recorded during 79 observations of individual L.
dimidiatus juveniles over a period of 1580 min. Cleaner wrasse cleaned 104 species of fish from 19
families, including 32 species of damselfishes of which the following were cleaned most often
(listed from most to fewest): Acanthochromis polyacanthus, Stegastes apicalis, Amblyglyphidodon
curacao, P. moluccensis, Plectroglyphidodon lacrymatus, P. amboinensis, P. nagasakiensis, P.
wardi and Chromis lepidolepis (see Appendix 2 for a full list of client species). These interaction
frequencies were not adjusted for relative abundance of each client species. The mean number (±
SE) of clients cleaned and the duration spent cleaning per 20 min observation was 14 ± 1 fish and
83.8 ± 7.3 s, respectively. Overall, only 14 % of cleaning interactions were between juvenile L.
dimidiatus and the four common damselfishes (P. amboinensis, P. moluccensis, P. nagasakiensis
and P. wardi) that had the greatest abundances within the 1 m radius of a cleaner wrasse. A total of
46 % of interactions occurred with other damselfishes (excluding the latter common species listed),
while the remaining 40 % of interactions occurred between non-damselfishes and juvenile L.
dimidiatus. Overall, 60 % of the clients of juvenile L. dimidiatus were damselfishes. The range and
median size (mm TL) of the common damselfishes that were cleaned were: P. amboinensis (20–80
and 50 mm), P. moluccensis (25–80 and 50 mm), P. nagasakiensis (15–90 and 40 mm), and P.
wardi (50–120 and 90 mm); the duration (seconds, mean ± SE) of time spent cleaning per 20 min
observation for the common species was 14 ± 2.5 s. Client size differed among the four species
(LRT = 68.01, df = 3, P < 0.0001; Fig. 3a); a TK-HSD test showed that P. wardi clients were
significantly larger than the other common client damselfishes. Damselfishes that were cleaned
were significantly larger than nearby non-cleaned conspecifics occurring within a 1 m radius; this
effect was consistent for all four species (LRT = 173.72, df = 1, P < 0.0001; Fig. 3a). Separate
analyses showed that the size of the damselfish client was not correlated with the size of the cleaner
wrasse (LRT = 0.17, df = 1, P = 0.675) nor was it correlated with the median size of all other fish
species combined (excluding common damselfishes) within the 1 m radius of the cleaner (LRT =
2.55, df = 1, P = 0.110).
Despite the presence of numerous recently-settled damselfishes (size range 15–20 mm)
within the 1 m radius observation areas (Fig. 3b), only 2 % (22 out of 1107 cleaning interactions) of
fishes that were cleaned were < 20 mm TL. There was a significant negative relationship between
the number of recruits that were cleaned and the abundance of both other fish species (> 20 mm,
TL) (LRT = 7.97, df = 1, P = 0.004, Fig. 4a) and recruits (LRT = 16.95, df = 1, P = < 0.0001, Fig,
4b) within the 1 m radius observation area. The probability of a recruit (< 20 mm) being cleaned by
a juvenile cleaner wrasse in a 20 min period is 0.002 (95 % CI = 3.22 × 10-6 ̶ 0.22) when there are
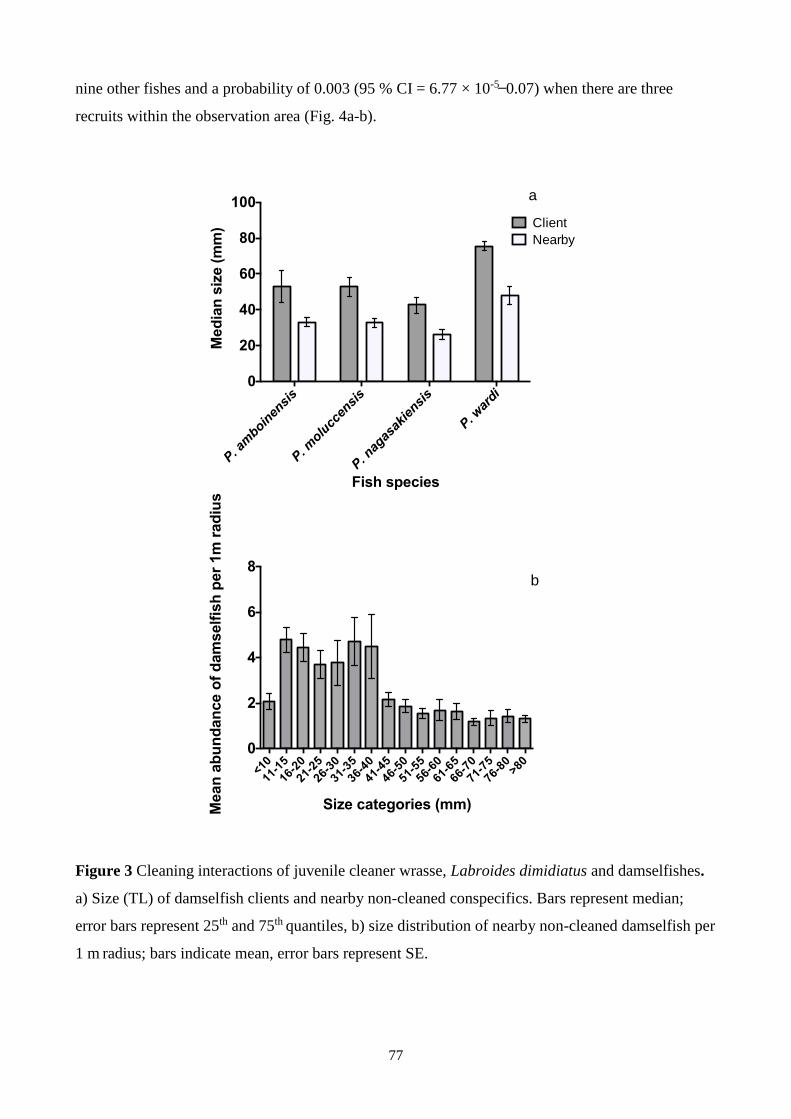
77
nine other fishes and a probability of 0.003 (95 % CI = 6.77 × 10-5 ̶ 0.07) when there are three
recruits within the observation area (Fig. 4a-b).
Figure 3 Cleaning interactions of juvenile cleaner wrasse, Labroides dimidiatus and damselfishes.
a) Size (TL) of damselfish clients and nearby non-cleaned conspecifics. Bars represent median;
error bars represent 25th and 75th quantiles, b) size distribution of nearby non-cleaned damselfish per
1 m radius; bars indicate mean, error bars represent SE.
<10
11-1
5
16-2
0
21-2
5
26-3
0
31-3
5
36-4
0
41-4
5
46-5
0
51-5
5
56-6
0
61-6
5
66-7
0
71-7
5
76-8
0>8
00
2
4
6
8
Size categories (mm)Mean
ab
un
dan
ce o
f d
am
self
ish
per
1m
rad
ius
b
P. a
mboin
ensi
s
P. m
olucc
ensi
s
P. n
agas
akie
nsis
P. war
di0
20
40
60
80
100
Fish species
Med
ian
siz
e (
mm
) Client
Nearby
a
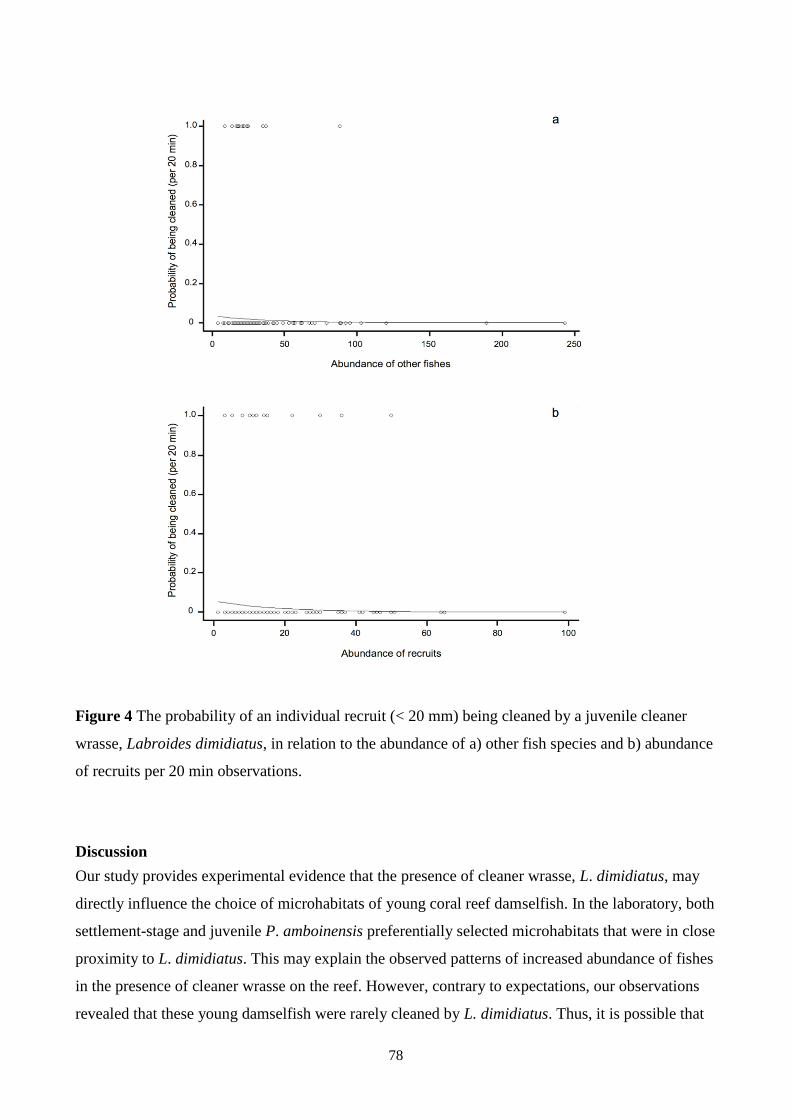
78
Figure 4 The probability of an individual recruit (< 20 mm) being cleaned by a juvenile cleaner
wrasse, Labroides dimidiatus, in relation to the abundance of a) other fish species and b) abundance
of recruits per 20 min observations.
Discussion
Our study provides experimental evidence that the presence of cleaner wrasse, L. dimidiatus, may
directly influence the choice of microhabitats of young coral reef damselfish. In the laboratory, both
settlement-stage and juvenile P. amboinensis preferentially selected microhabitats that were in close
proximity to L. dimidiatus. This may explain the observed patterns of increased abundance of fishes
in the presence of cleaner wrasse on the reef. However, contrary to expectations, our observations
revealed that these young damselfish were rarely cleaned by L. dimidiatus. Thus, it is possible that
Contr
ol
Rem
oval
0
20
40
60
80
100
P. amboinensis
Me
an
ab
un
da
nc
e p
er
ree
f
1a*
Contr
ol
Rem
oval
0
20
40
60
80
100
P. moluccensis
Me
an
ab
un
da
nc
e p
er
ree
f
1cn.s.
Contr
ol
Rem
oval
0
5
10
15
20
P. adelus
Me
an
ab
un
da
nc
e p
er
ree
f
1en.s.
Contr
ol
Rem
oval
0
10
20
30
C. rollandi
Me
an
ab
un
da
nc
e p
er
ree
f
1b*
Contr
ol
Rem
oval
0
20
40
60
80
100
P. nagasakiensis
Me
an
ab
un
da
nc
e p
er
ree
f
1d*
Contr
ol
Rem
oval
0
20
40
60
80
100
Total damselfish abundance
Me
an
ab
un
da
nc
e p
er
ree
f
1f*

79
even these rare cleaning events and/or indirect benefits resulting from the presence cleaner wrasse
may drive the selective behaviour of settling damselfish.
We assessed microhabitat choice by settlement-stage (0 days post-settlement) and slightly
older juvenile (~5 weeks post-settlement) P. amboinensis and found that individuals preferentially
selected a microhabitat adjacent to L. dimidiatus. This indicates that the ‘attraction’ towards cleaner
wrasse occurs in both stages. Habitat selectivity at settlement appears common in young
damselfishes, with some species attracted to particular microhabitats such as coral heads that
harbour conspecifics (Lecchini et al. 2007a; Coker et al. 2012a), while others are attracted to coral
heads associated with heterospecifics (Green 1998; Almany 2003). Our study adds cleaner wrasse
to the list of organisms that settlers are attracted to, so that the presence of cleaner wrasse could act
as an indicator of microhabitat quality to settlement-stage fish. Furthermore, these results suggest
that if juveniles move between microhabitats, that their site selection may also be influenced by the
presence of L. dimidiatus.
The apparent attraction of L. dimidiatus may explain the greater number of recently-settled
recruits (Sun et al. in review, Chapter 3), juvenile visiting fishes and even adult damselfishes, found
on patch reefs where L. dimidiatus are present relative to reefs without cleaner wrasse (Bshary
2003; Waldie et al. 2011). There are two ways in which these patterns may arise, using the presence
of L. dimidiatus as a cue. Settlement-stage fish may either choose to settle on certain patch reefs, or
engage in post-settlement movement between patch reefs as juveniles (Simpson et al. 2008). Both
of these processes may additively result in damselfishes being more abundant on reefs with cleaner
wrasse than on those without.
The fish used in the laboratory experiment were naïve with respect to the shape or odour of
L. dimidiatus, since they were collected by light-traps while still in their planktonic phase. Despite
this naïveté, young fish were able to differentiate between L. dimidiatus and another species that
was similar in size and shape but differed in colour, pattern and behaviour. The attraction to cleaner
wrasse thus appears to be an innate behaviour. Slightly older recruits displayed the same abilities.
However, we did not test whether P. amboinensis recognised L. dimidiatus as a cleaner per se. On
some occasions, the fish did behave like a client by remaining in the same position and adopting a
pose with spread fins when next to the compartment with a cleaner wrasse. This behaviour did not
occur when they were adjacent to the control fish or an empty compartment. Prospective clients of
cleaner fish often assume this posture when they attempt to attract a cleaner to inspect them (Côté et
al. 1998). Because the cleaner wrasse was separated by Perspex, both the cleaner and client were
unable to physically interact; such physical contact likely acts as a positive feedback (Bshary and
Würth 2001). Losey et al. (1995) previously showed that newly-metamorphosed laboratory-reared,
and hence ‘cleaner-naïve’ Hawaiian humbug damselfish, Dascyllus albisella, recognised the

80
Hawaiian cleaner wrasse, Labroides phthirophagus by adopting the cleaning ‘pose’. Our study thus
confirms the hypothesis that client fish have an innate ability to recognise cleaners.
It seems likely that P. amboinensis used visual cues to differentiate between the two
stimulus fish that were presented. A combination of black and blue and/or yellow body patterns
displayed by cleaner fish, including L. dimidiatus, allows clients to visually recognise cleaners
(Cheney et al. 2009). Indeed, the particular colouration and patterns displayed by cleaner fish attract
the higher numbers of client fish than other colour and pattern combinations. Client fish are also
known to recognise cleaner fish based on their small body size and the presence of lateral stripes
(Stummer et al. 2004).
Pomacentrus amboinensis both chose adult and juvenile L. dimidiatus over the other stimuli.
This occurred despite the two cleaner stages being differently coloured, with both having a black
stripe contrasted by blue, but the adults also having the colours yellow and white. However,
juvenile P. amboinensis spent more time near juvenile L. dimidiatus than adults implying the
possibility of ontogenetic variation in choice of cleaner fish.
Alternatively, this could simply be due to the difference in relative size between the client
and cleaner at different ontogenetic stages. The same choice test was not performed using
settlement-stage fish due to their limited numbers in the light traps. Previous studies have shown
that fish larvae have good visual senses, which enables them to detect and recognise conspecifics,
heterospecifics and predator fishes (Lecchini et al. 2005; Lecchini et al. 2014).
Focal observations of juvenile L. dimidiatus on reefs revealed that only 2 % of their clients
were < 20 mm TL. This suggests that small individuals are rarely cleaned by L. dimidiatus and so
the interaction may contribute little to the cleaner wrasse’s diet. Small-bodied individuals, such as
recently-settled damselfishes, are infected with very few ectoparasites (Grutter et al. 2010; Sun et
al. 2012) and the presence of gnathiid isopods (the main food source of L. dimidiatus) is relatively
low in juvenile fishes (only 3.5 % of recruit P. amboinensis are infected) (Grutter et al. 2011). From
the cleaner wrasse’s perspective, these factors likely make settlers an unsuitable source of
ectoparasites. It was therefore unsurprising that we found that the damselfish that were cleaned by
juvenile L. dimidiatus were larger than the median size of non-cleaned conspecifics within a 1 m
radius. This result was consistent with previous studies that show that L. dimidiatus selectively
clean larger fish (Grutter et al. 2005; Clague et al. 2011), probably due to client size and parasite
load being closely correlated (Grutter 1994,1995a; Grutter and Poulin 1998); L. dimidiatus may
thus use size as an indicator of food availability.
From the client’s perspective, the odds of an individual new recruit (< 20 mm, TL) being
cleaned per 20 min, was low, with only two or three out of 1000 fish predicted to be cleaned when
there were no other fishes or recruits (< 20 mm) within a 1 m radius, and this rate rapidly decreased

81
towards zero with increasing abundance of nearby other fish species and recruits. Over a day,
however, the cumulative odds would be much higher. Such rare events could have
disproportionately significant benefits as some parasites can have especially harmful effects on
small individuals. While gnathiid isopods are generally considered micropredators, taking small
amounts of blood from their significantly larger host (Lafferty and Kuris 2002), they are capable of
consuming up to 85 % of the total blood volume of a settlement-stage P. amboinensis (Grutter et al.
2011). Indeed, infection by a single gnathiid isopod decreases the oxygen consumption and
swimming performance of a recruit damselfish and reduces the likelihood of successful settlement
on the reef (Grutter et al. 2011). If an infected fish has a gnathiid removed before it feeds on its
blood, this could provide a major benefit for survival. Since observed cleaning interactions were
brief (~ 1 s), it is unlikely that they obtained other direct benefit of cleaning, such as tactile
stimulation which leads to a reduction in client stress (cortisol) levels (Soares et al. 2011).
As direct cleaning interactions between cleaners and recently-settled recruits were relatively
uncommon, it is possible that the benefits of preferentially selecting a microhabitat near cleaners
may be at least partly indirect. For example, cleaner wrasse may reduce infection rate of parasites in
the vicinity of a cleaning station. As each L. dimidiatus eats around 1200 gnathiids per day (Grutter
1996a), this could significantly reduce the number of ectoparasites in the immediate vicinity of a L.
dimidiatus and hence lower the infection rate onto nearby settlers (Gorlick et al. 1987; Grutter
1999a; Clague et al. 2011). Other indirect benefits of the presence of cleaner wrasses may also exist
for settling fish. ‘Safe havens’ from predators may be created by L. dimidiatus (Cheney et al. 2008)
in that, in the presence of cleaners, some predators behave with notably reduced aggression. Settling
reef fish may therefore prefer to settle in a habitat with L. dimidiatus present, due to reduced risk of
predator aggression. Alternatively, or in addition, selection by young damselfish of microhabitats
near cleaners may simply be an investment where the return occurs only when recruits have grown
significantly.
On reefs lacking cleaner wrasse, the size at age of the lemon damselfish, P. moluccensis, is
smaller in older individuals (> 1 year old) and older fish have more parasitic copepods than fish on
reefs where cleaners are present (Clague et al. 2011). Additionally, size frequency distributions of
P. amboinensis and P. moluccensis are skewed towards smaller individuals on reefs without cleaner
wrasse (Waldie et al. 2011). Ultimately, this might affect fecundity, and hence the supply of future
settlers, since larger body sizes are correlated with egg production (Green 2008).
In summary, our study provides evidence that settlement-stage reef fish may use the
presence of cleaner wrasse as a cue when deciding what microhabitat to settle on. As the presence
of this mutualistic interaction creates favourable environments, this may ultimately enhance the

82
survival and abundance of these species. This study demonstrates that mutualism contributes to the
already-complex settlement processes of reef fishes.
Acknowledgements
We thank F. Cortesi, M. De Brauwer, F. R. A. Jaine, J. E. Lange, A. Marshell, G. A. C. Phillips,
and M. D. Mitchell for their help in the field; J. O. Hanson for statistical advice and R. Q-Y. Yong
and S. C. Cutmore for comments on earlier drafts of the manuscript. We are particularly grateful for
the support of the staff of the Lizard Island Research Station. This work was funded by a Sea World
Research and Rescue Foundation, Australia (SWRRFI) awarded to D. Sun, A. S. Grutter and T. H.
Cribb and funds from the Australian Research Council awarded to A. S. Grutter and M. G. Meekan
and from The University of Queensland awarded to A. S. Grutter.

83
Chapter 5: Diel patterns of infection by gnathiid isopods
on the ambon damsel, Pomacentrus amboinensis, and
the influence of the presence of cleaner wrasse

84
Chapter 5: Diel patterns of infection by gnathiid isopods on the ambon damsel,
Pomacentrus amboinensis, and the influence of the presence of cleaner wrasse
Derek Sun, Karen L. Cheney, Johanna Werminghausen, Thomas H. Cribb & Alexandra S. Grutter
Abstract
That the presence of bluestreak cleaner wrasse, Labroides dimidiatus, affects fish abundance and
fish traits is well known, yet little is known of the causal mechanism (s) involved. Since juvenile
gnathiid isopods are the main ectoparasite eaten by cleaner wrasse, a difference in their infection
number as a consequence of cleaner wrasse presence could be one such causal mechanism. To test
this hypothesis we used ambon damselfish, Pomacentrus amboinensis as a model host to estimate
the cumulative number of gnathiid isopods on patch reefs where the presence and absence of
cleaner wrasse had been experimentally manipulated for 12 years. Gnathiids were collected using
sentinel traps containing three live juvenile P. amboinensis as hosts; traps were sampled after 12 h
at dawn and dusk. There was no significant difference in the estimated cumulative number of
gnathiids per trap between reefs with and without L. dimidiatus. The probability of capturing one or
more gnathiid per trap on reefs without L. dimidiatus was 0.167 (95% CI = 0.112–0.240) compared
with 0.148 (95% CI = 0.096–0.221) on control reefs. However, the probability of capturing
gnathiids in traps was higher during the night [0.222 (95% CI = 0.144–0.327)], than during the day
[0.088 (95% CI = 0.061 – 0.126)]; or a 2.934 (95% CI = 1.725–4.985) times higher probability
during the night than during the day. We conclude that the rate of gnathiid isopod
attraction/detection measured here does not explain the patterns observed involving the effect of the
presence of L. dimidiatus on this fish reported in other studies.
Introduction
Parasites are often difficult to observe, yet many studies have shown that they may play a vital role
in influencing ecosystem functions (Hudson et al. 2006), regulate host populations (Scott and
Dobson 1989), and influence overall biodiversity (Hudson et al. 2006). On coral reefs, fish parasites
can have negative effects on their hosts (Ferguson et al. 2011; Binning et al. 2013; Krkošek et al.

85
2013). Understanding how fish manage parasite infections is thus critical to understanding their
ecology.
Cleaning behaviour, which involves organisms that remove ectoparasites from cooperating
clients, may be one way that reduces parasitic loads. Indeed, the frequent nature of cleaning
undergone by individual client fish, as often as every ~ 5 min for some species (Grutter 1995a),
suggests that the removal of ectoparasites is important to them. On Indo-Pacific coral reefs, L.
dimidiatus is the most common of the five Labroides species (Randall et al. 1997), all of which are
obligate cleaner fish that remove ectoparasites and sometimes tissue from the surfaces of other
fishes, known as clients.
The presence of the cleaner wrasse L. dimidiatus greatly influences the richness and
abundance of adult (Bshary 2003; Grutter et al. 2003), juvenile (Waldie et al. 2011) and recruiting
(Sun et al. in review, Chapter 3) reef fish. After 4 ̶ 20 months, in the Red Sea, the diversity of reef
fish on patch reefs was reduced when cleaner wrasse disappeared or were removed (Bshary 2003).
At Lizard Island, Great Barrier Reef, the species richness and abundance of visitors (fish species
that move between patch reefs) were reduced 18 months after cleaner wrasse had been removed
(Grutter et al. 2003); after 8.5 years, there was a reduction in the abundance and diversity of both
resident and visitor as well as adult herbivorous (Acanthuridae) fishes (Waldie et al. 2011). Only
recently, it was found that the abundance of damselfish recruits (species that are site-attached),
specifically Chrysiptera rollandi and Pomacentrus amboinensis, were reduced on these same patch
reefs after 12 years of regular removal of cleaner wrasse (Sun et al. in review, Chapter 3). These
studies highlight the importance of understanding the mechanism (s) via which a relatively
uncommon and small fish maintains the diversity and abundance of so many coral reef fishes.
An obvious way by which fish may benefit from the presence of cleaner fish is by obtaining
health benefits from a reduction in ectoparasites, which ultimately leads to changes in individuals or
communities. How ectoparasites can be influenced by cleaner fish will be determined by the
ecology and behaviour of the parasite and particularly the level of association between the parasite
and the host. There are two main types of ectoparasites, those that remain on the host for almost
their entire lives, i.e. ‘permanent’ parasites such as copepods (Finley and Forrester 2003) and
cymothoid isopods (Adlard and Lester 1994; Roche et al. 2013a), and those that remain on the host
only briefly while feeding, i.e. ‘temporary’ parasites such as gnathiid (Smit et al. 2003; Ota et al.
2012) and corallanid isopods (Grutter and Lester 2002), which are often classified as mobile
micropredators (Lafferty and Kuris 2002).
Surprisingly, only a few studies have tested the effect of L. dimidiatus on parasite loads.
The removal of L. dimidiatus did not have an effect on number of copepods per fish after 6 months
on a damselfish P. moluccensis at Lizard Island (Grutter 1996b). After 2 y the number of copepods

86
on P. vaiuli in the Marshall Islands was also not affected but the number of larger copepods (and
hence overall biomass) was higher (Gorlick et al. 1987). At Lizard Island, Clague et al. (2011)
showed that copepod number, summed across 10 fish sampled per reef, was higher on P.
moluccensis on reefs without a cleaner wrasse for 8.5 y, but only on larger P. moluccensis. While
copepods are not often consumed by cleaner wrasse (Grutter 1997) and individual damselfish are
cleaned relatively infrequently (Grutter 1995a), that larger P. moluccensis individuals are
preferentially cleaned (Clague et al. 2011; Sun et al. in review, Chapter 4) could explain why an
effect on copepod number was only apparent in larger individuals.
The presence of cleaners also has an effect on parasitic isopods. For caged thicklip wrasse,
Hemigymnus melapterus, gnathiid number per fish was higher on reefs without L. dimidiatus after
24 h and 12 d (Grutter 1999a), while on the same removal reefs the prevalence and number of
corallanid isopods was higher after 24 h, while a noticeable shift towards smaller isopods was
detected after 12 d (Grutter and Lester 2002). In a very different system involving cleaner gobies
(Elacatinus spp.) in the Caribbean, gnathiid number for Stegastes diencaeus damselfish captured
using handnets was lower on individuals that had cleaning stations within their territory than on
individuals that lived further away from a cleaning station (Cheney and Côté 2001).
Gnathiids can inhibit the growth (Jones and Grutter 2008), swimming performance and
oxygen consumption (Grutter et al. 2011) and decrease the survival rate (Grutter et al. 2008; Grutter
et al. 2011) of damselfish juveniles. High numbers of gnathiids can cause anaemia (Paperna 1977;
Grutter 2008) and mortality in adult fish (Mugridge and Stallybrass 1983; Marino et al. 2004).
Thus, a reduction in gnathiids could ultimately lead to the higher abundance of damselfishes
observed on reefs where cleaner wrasse are present (Bshary 2003; Waldie et al. 2011; Sun et al. in
review, Chapter 3).
The size frequency distribution of both P. amboinensis and P. moluccensis was shifted
towards smaller individuals on reefs where cleaner wrasse have been removed for 8.5 years (Waldie
et al. 2011). The shift in size distribution could be due to decreased rate of growth in individuals on
reefs without a cleaner wrasse. Although P. moluccensis had a lower growth rate and higher
copepod numbers on reefs without cleaners, this was only apparent in larger individuals (Clague et
al. 2011). Thus, it is possible that other ectoparasites such as gnathiid isopods may explain the
overall shift in size distribution seen between reefs with and without cleaner wrasse.
As L. dimidiatus can consume an average of 1200 ectoparasites a day with the majority of
their diet being made up of gnathiid isopods (99.7 %) (Grutter 1996a), it is possible that their effect
on parasites is likely to be the greatest on this group. Gnathiid isopods are one of the main
ectoparasites that infect coral reef fishes (Grutter et al. 2000). Unlike copepods, gnathiids are
mobile temporary parasites, emerging either during the day or at night to locate a host to feed

87
(Sikkel et al. 2006). Gnathiids usually detach from a host once they are fully fed (Grutter 2003) or
are disturbed (Grutter 1995b). Due to this unique life cycle and behaviour, several measures have
been used to quantify the ‘abundance’ of gnathiids. These terms are defined as density of
cumulative number of emerging gnathiids (i.e. number per reef area/per time), number (number per
fish) and infestation (number present per unit time). Therefore, the sampling method used to
quantify gnathiid isopod ‘abundance’ on fish could also affect the estimate; for example due to
differences in their emergence from the benthos (Grutter 1999b; Sikkel et al. 2006), the time of day
that a fish are sampled will affect how many parasites are counted. Damselfishes also tend to have
few gnathiids (Grutter 1994; Sun et al. 2012) and so the odds of sampling them on fish are low. A
method that increases the time they are sampled on fish would increase these odds.
The use of caged fish in previous studies to quantify the number or infestation of gnathiids
(Grutter 1999a; Sikkel et al. 2006; Coile and Sikkel 2013) does not exclude the potential direct
interaction between cleaner and the caged fish and hence the removal of gnathiids. The use of cages
also does not prevent the potential loss of gnathiids once they are fed and have dropped off.
Therefore, it is possible that the initial number of gnathiids in previous studies was similar on caged
fish placed on reefs either with or without cleaner wrasse and that a lower infection was recorded
simply due to the direct cleaning interaction between cleaners and the caged fish. As gnathiids can
easily detach themselves from a host, conventional methods of capturing fish (handnets) to examine
the number of gnathiids are not ideal (Grutter 1995b). Such detachment could have contributed to
the low prevalence rate and number of gnathiids that has been reported on damselfishes (Grutter
and Poulin 1998; Cheney and Côté 2001; Grutter et al. 2011; Sun et al. 2012).
The high emergence rates of gnathiids (41.7 ± 6.9 m-2 day-1) (Grutter 2008) means it is
likely that hosts are put under constant physical stress, as well-fed gnathiids are constantly replaced
by unfed individuals. Although previous studies have shown an effect of cleaner fish presence on
the number of gnathiids on a fish (Grutter 1999a; Cheney and Côté 2001), the results are limited in
that they only provide information of the number of gnathiids on a host at a given time, when the
host was captured. As a result, these studies may not provide a true representation of overall number
in relation to cleaner fish presence. Therefore, an alternative sampling technique is required: one
which prevents cleaner wrasse from interacting with the host fish, retains gnathiids once they have
dropped off, and provides as estimate of the cumulative number of gnathiids on fish over time. One
such method involves the use of ‘sentinel traps’ which hold ‘bait’ fish and allow gnathiids, but not
cleaner fish, to enter and feed off hosts, but which do not allow gnathiids to escape (Sikkel et al.
2011).
Despite damselfishes being relatively sedentary on coral reefs and unable to move between
reefs to seek a cleaner wrasse, their abundance is higher in the presence of a cleaner wrasse (Sun et

88
al. in review, Chapter 3). Yet, the mechanism(s) remains unclear. Currently, it is not known if the
cumulative number of gnathiids on damselfishes differ according to cleaner wrasse presence. As
cleaner wrasse can consume large quantities of gnathiid isopods, we therefore predict that the
cumulative number of gnathiids will be lower on reefs where cleaner wrasse are present. In this
study, we examined whether the presence of cleaner wrasse has an effect on the cumulative number
of gnathiid isopods per trap using relatively large juvenile ambon damsel, P. amboinensis.
Pomacentrus amboinensis were placed in sentinel traps and placed on experimental patch reefs with
or without cleaner wrasse. Because cleaner fish are only active during the day (Grutter 1996a) and
thus could potentially affect only ‘diurnal gnathiids’, sampling was divided between the day and the
night, over a 24 h time period.
Materials and methods
Study site
This study was conducted at Lizard Island (1440S, 14528E) in the northern GBR, Australia during
late summer (January and February) of 2013. We used 16 isolated patch reefs (3−7 m depth) located
at two sites: the Lagoon (11 reefs) and at Casuarina Beach (5 reefs) (Grutter et al. 2003; Clague et al.
2011). Reefs were randomly allocated as either removal or control treatment reefs in September 2000.
Subsequently, reefs were checked at several-month intervals and were inspected for cleaner wrasse
L. dimidiatus; any new recruits on removal reefs were removed using barrier and handnets. Control
reefs were surveyed periodically for L. dimidiatus by swimming around the reef several times and the
abundances of cleaners recorded.
Study species
The ambon damselfish, Pomacentrus amboinensis, is a common coral reef fish species on the GBR.
As much is known about its ecology, including habitat preference, recruitment, and feeding habits
(Kerrigan 1996; McCormick 2006; McCormick et al. 2010), this species has been extensively used
as a model species in both field and laboratory experiments and it is abundant on all of the
experimental reefs.
Sampling design
A total of 72 large juvenile P. amboinensis (mean TL per fish ± SE = 50.01 ± 0.38 mm) were
collected at Lizard Island in January 2013 using barrier and handnets. Fish were initially maintained
in aerated 40 L tanks, then, due to their known tendency for aggressive behaviour towards each
other, were transferred to individual clear plastic holding aquaria (29 × 17 × 12 cm) with a constant

89
flow of unfiltered seawater and maintained on a diet of commercial fish flakes. Fish were then
randomly divided into four groups (A, B, C, and D), each group consisting of 18 fish. Within each
group, the fish were then further divided into 6 labelled subgroups (numbered from 1–24) with 3
fish in each subgroup. These three fish were used as the sampling unit per fish trap. Gnathiids
mainly locate their hosts using olfactory cues (Sikkel et al. 2011) so three fish per trap were used in
order to maximise the amount of chemical cue emitted from the trap.
A total of 14 individually labelled sentinel traps were constructed using a T-junction PVC
pipe (370 mm long × 150 mm diameter) with a threaded lid attached to each of the three ends
(Figure 1) (see Sikkel et al. 2011 for a base design). A 100 mm diameter hole was cut out of each
lid and a clear plastic funnel (10 mm diameter opening) attached to the inside surface of the lid,
using plastic cable ties and aquarium grade silicone glue. To facilitate filling and emptying the
water in the tubes, three 32 mm diameter holes were drilled into the sides of the tube. The drainage
holes were covered with 100 μm plankton mesh glued inside the tube; this allowed only water (no
gnathiids) to circulate in and out of the trap. The six openings allowed sufficient water circulation to
maintain adequate oxygen levels.
Twelve traps in a paired design, with 6 traps placed each time on a removal and control
reef, were used per day. This was repeated on all 16 experimental reefs; due to the odd number of
reefs at each of the two sites sampled, this meant only one reef was sampled on one day at each of
the sites, resulting in 9-day sampling periods. For each sampling day, two fish groups (A and B, or
C and D) were used. The traps were placed on the reef at dusk, sampled (traps emptied and
redeployed with same fish) at dawn the following day (~ 12 h after initial placement) and the
second and final sample collected at dusk later that day (~ 12 h after first sampling). The sampling
design allowed each group to be used a total of 4 different times throughout the experiment, with
each group being re-used every 48 h. This provided each group with a recovery period of at least 24
h between deployments.
To determine whether gnathiids were attracted to traps and not the fish in them, we
deployed empty traps as a control. Empty trap sampling was conducted with 6 traps placed on a
control and 6 on a removal reef, one day before the commencement (day 0) and the day after the
conclusion (day 10) of the 9-day experiment. As an additional control, every second day throughout
the experiment, an extra empty trap was placed alongside the six traps containing fish. The entire
experiment was replicated after a one-week recovery period, with the same groups of fish, in the
same order, for the same duration (Table 1).
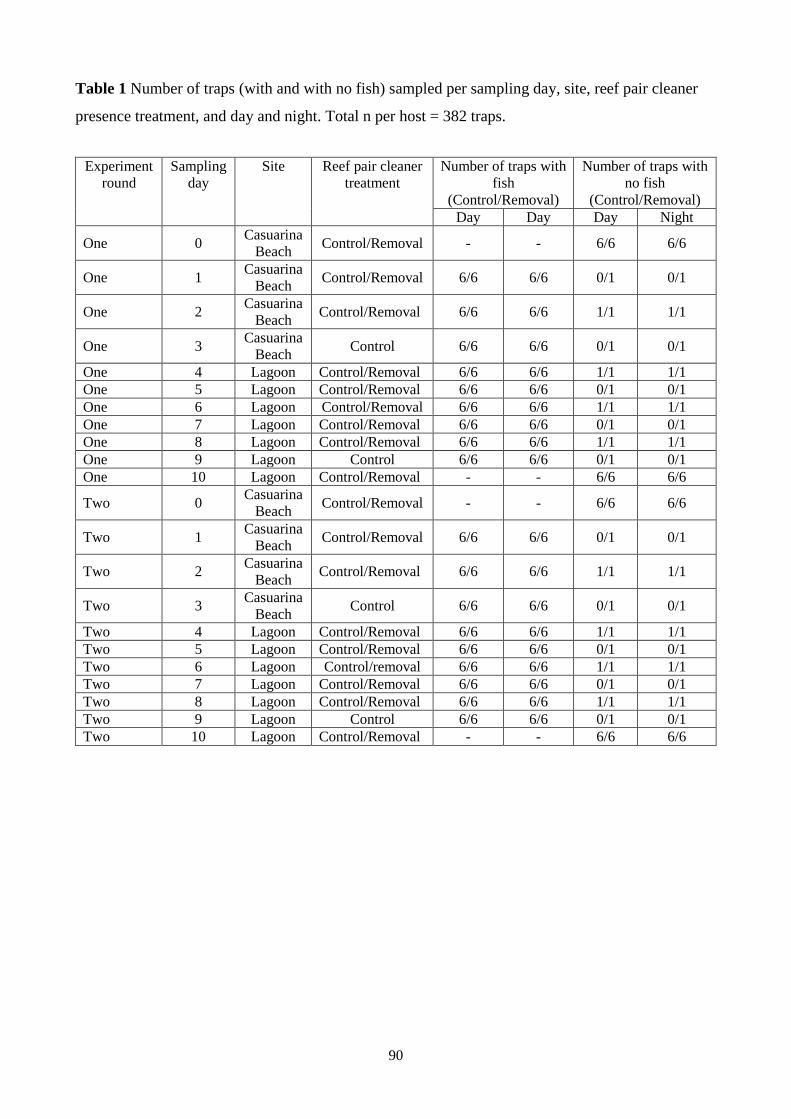
90
Table 1 Number of traps (with and with no fish) sampled per sampling day, site, reef pair cleaner
presence treatment, and day and night. Total n per host = 382 traps.
Experiment
round
Sampling
day
Site Reef pair cleaner
treatment
Number of traps with
fish
(Control/Removal)
Number of traps with
no fish
(Control/Removal)
Day Day Day Night
One 0 Casuarina
Beach Control/Removal - - 6/6 6/6
One 1 Casuarina
Beach Control/Removal 6/6 6/6 0/1 0/1
One 2 Casuarina
Beach Control/Removal 6/6 6/6 1/1 1/1
One 3 Casuarina
Beach Control 6/6 6/6 0/1 0/1
One 4 Lagoon Control/Removal 6/6 6/6 1/1 1/1
One 5 Lagoon Control/Removal 6/6 6/6 0/1 0/1
One 6 Lagoon Control/Removal 6/6 6/6 1/1 1/1
One 7 Lagoon Control/Removal 6/6 6/6 0/1 0/1
One 8 Lagoon Control/Removal 6/6 6/6 1/1 1/1
One 9 Lagoon Control 6/6 6/6 0/1 0/1
One 10 Lagoon Control/Removal - - 6/6 6/6
Two 0 Casuarina
Beach Control/Removal - - 6/6 6/6
Two 1 Casuarina
Beach Control/Removal 6/6 6/6 0/1 0/1
Two 2 Casuarina
Beach Control/Removal 6/6 6/6 1/1 1/1
Two 3 Casuarina
Beach Control 6/6 6/6 0/1 0/1
Two 4 Lagoon Control/Removal 6/6 6/6 1/1 1/1
Two 5 Lagoon Control/Removal 6/6 6/6 0/1 0/1
Two 6 Lagoon Control/removal 6/6 6/6 1/1 1/1
Two 7 Lagoon Control/Removal 6/6 6/6 0/1 0/1
Two 8 Lagoon Control/Removal 6/6 6/6 1/1 1/1
Two 9 Lagoon Control 6/6 6/6 0/1 0/1
Two 10 Lagoon Control/Removal - - 6/6 6/6

91
Figure 1 Photograph of the sentinel trap used in the experiment. Traps were made from plastic PVC
material. A) Each lid is fitted with a transparent plastic funnel, allowing gnathiid isopods to enter
the trap while providing fish with adequate water circulation; B) Each trap is fitted with a 1.4 kg
dive weight, which is secured with a dive belt at the front opening; C) Three drainage holes (3.2 cm
in diameter) are drilled at the back of the traps. Photo: Maarten De Brauwer.
Sampling protocol
The three fish were held individually in subcages within the trap. Subcages reduced any
potential negative interaction amongst the fish within the trap and were constructed from flexible
corrugated plastic pipe (100 diameter × 50 length mm) with mesh (15 mm) covering each circular
end (Figure 2). Although mesh size was large enough to allow gnathiids to enter the subcage, ~10
larger holes (3 mm) were also drilled in the mesh to increase the opportunity for gnathiids to enter
the subcage. The subcages had small holes bored along its length in order to increase the entry
points for gnathiid isopods. Prior to transporting fish to the field site, each fish was placed
individually into subcages labelled with the corresponding subgroup identification.
Fish were transported to the field in a 60 L container, which was aerated with regular water
changes. At the field site, each subgroup was placed in a trap. Traps were then passed to a snorkeler
and were placed carefully on the reef at dusk. Traps were secured with a 1.4 kg dive weight
attached to the outside of each trap using a dive belt and buckle. Traps were subsampled at dawn
the following day. To ensure no trapped gnathiids were lost while moving traps off the reef to the
boat, all funnel holes were blocked using a 1 cm diameter ball of household gum (Blu-tack,
Thomastown, Australia). The contents of each trap were emptied and well-rinsed into
correspondingly-numbered 10 L buckets. Subcages, still containing a fish, from each trap were
given a 1-minute freshwater bath (3 L) in a separate bucket, and the contents then correspondingly
B
C
A

92
placed into the aforementioned 10 L buckets. Freshwater baths are a standard method used to
remove gnathiids that may be attached on a fish (Soares et al. 2008). Subcages and fish were then
returned to the traps and placed on their original locations on the reef. This procedure was repeated
the next dusk but using the next pair of reefs and the next two fish groups.
Twelve buckets holding the trap, rinse and bath water samples were brought back to the
research station, along with (after the dusk sampling) the recently-used fish in their subcages. For
the dusk samples, an airstone was placed into each bucket for 3 hours to keep the fish alive while
allowing any remaining gnathiids to drop off naturally. Each fish subgroup was also given a final 1-
minute freshwater bath, during which their body was gently rubbed before being placed back into
their allocated aquaria. The combined water samples were filtered through a 100 μm plankton sieve
into vials to remove any gnathiids, and then preserved with a 10% formaldehyde solution in
seawater. During the experiment a total of six fish died (two traps, each on a control and removal
reef) and were replaced with new ones.
Figure 2 Individual subcages used to hold Pomacentrus amboinensis in the sentinel traps
Statistical analysis
Since 53 of the 62 samples with gnathiids present contained only one individual, we used the
presence/absence of gnathiids as the response variable. A generalized linear mixed effect model
(GLMER) with a binomial distribution error was used to examine whether gnathiid presence
differed with cleaner wrasse presence. The terms treatment (cleaner or no cleaner), site (Lagoon or
Casuarina Beach), time (day or night) and experiment replicate (Experiment one or two) were fixed
factors, and reef ID, sampling day, and fish group as random factors. Fish subgroup identity was
nested with fish group. The sum of the fish weight of the three fish per subgroup was included as a
covariate, as gnathiids are known to use fish scent to locate a potential host (Sikkel et al. 2011) and
we expected that the scent of fish would be correlated with the mass of fish.

93
In a separate analysis, the same model was used to estimate the rate at which gnathiids
entered empty traps (no fish), but without fish weight, fish group, and fish subgroup identity as
factors.
To determine the best model for the data, we compared the full model with models in
which one of the explanatory terms was dropped using the “drop1” function (Chambers 1992). If
the maximum likelihood ratio test (LRT) found that a dropped term had no significant effect on the
model by using the Chi-square distribution, then the term was dropped. All statistical analyses were
performed using R version 3.0.1 (R Development Core Team 2013). As information regarding
species-level taxonomy of gnathiid isopods is scarce, gnathiids encountered in this study were not
identified further.
Results
Diel patterns
Of the 382 trap samples containing fish, 62 contained a total of 72 gnathiids, with a maximum of
three gnathiids in a single trap (Table 2).
The presence of gnathiid isopods differed according to time of day (LRT= 13.61, df = 1, P
= 0.0002), with more gnathiids being caught at night than in the day. Traps at night had a higher
probability of capturing a gnathiid 0.222 (95% CI = 0.144–0.327), than those set during the day
0.088 (95% CI = 0.061–0.126); or a 2.934 (95% CI = 1.725–4.985) times higher probability during
the night than during the day.
Cleaner wrasse presence
The simplified model showed that the presence/absence of gnathiid isopods did not differ according
to cleaner wrasse presence (LRT = 0.216, df = 1, P = 0.642), site (LRT =0.050, df = 1, P = 0.822),
fish weight (LRT = 1.185, df = 1, P = 0.276) or experiment replicate (LRT = 0.299, df = 1, P =
0.583). Traps had a 0.167 (95% CI = 0.112–0.240) probability of capturing a gnathiid on removal
reefs, compared with 0.148 probability (95% CI = 0.096–0.221) on control reefs, or a 1.153 (95%
CI = 0.731–1.818) times higher probability on removal than control reefs.
Of the 128 trap samples containing no fish, 11 contained a sum of 12 gnathiids, with a
maximum of two gnathiids per trap (Table 2). The presence/absence of gnathiids did not differ
according to cleaner wrasse presence (LRT = 0.136, df = 1, P = 0.711), time of day (LRT = 0.239,
df = 1, P = 0.624), site (LRT = 2.43, df = 1, P = 0.118) and experiment replicate (LRT = 1.22, df =
1, P = 0.268).
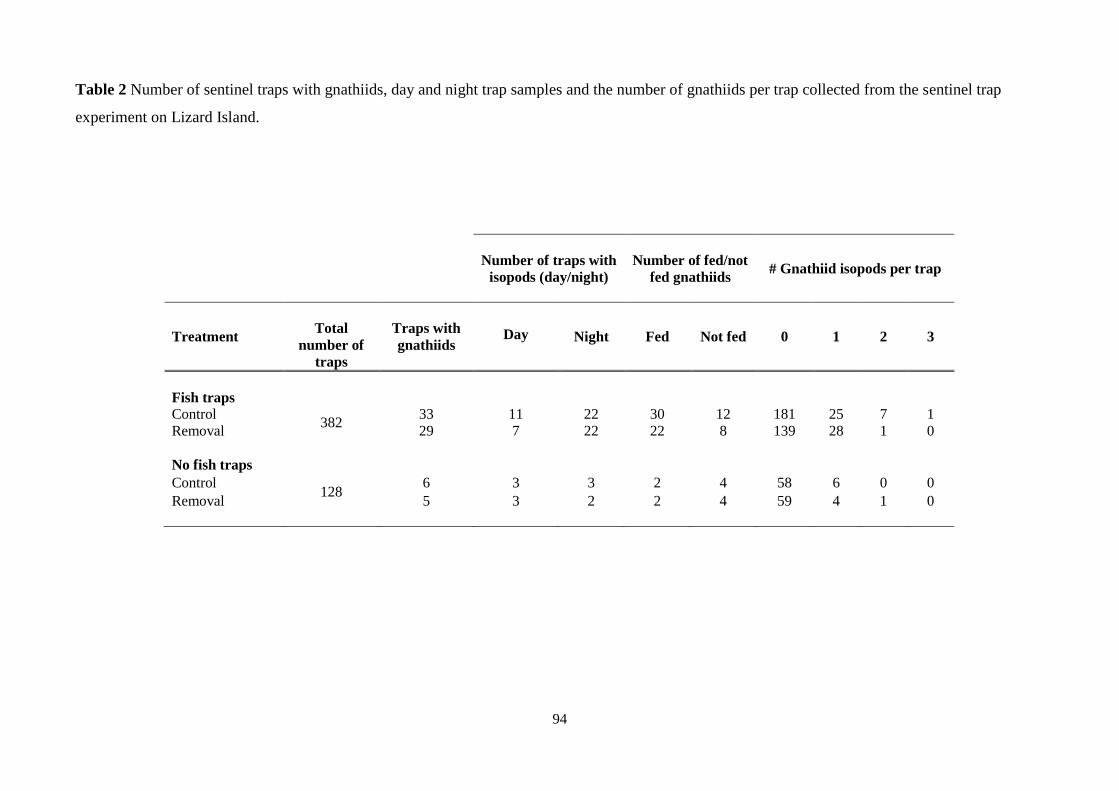
94
Table 2 Number of sentinel traps with gnathiids, day and night trap samples and the number of gnathiids per trap collected from the sentinel trap
experiment on Lizard Island.
Number of traps with
isopods (day/night)
Number of fed/not
fed gnathiids # Gnathiid isopods per trap
Treatment
Total
number of
traps
Traps with
gnathiids
Day Night Fed Not fed 0 1 2 3
Fish traps
Control
382
33
11
22
30
12
181
25
7
1
Removal 29 7 22 22 8 139 28 1 0
No fish traps
Control 128
6 3 3 2 4 58 6 0 0
Removal 5 3 2 2 4 59 4 1 0

95
Discussion
Our study found that the cumulative number of gnathiid isopods per trap using the three ambon
damsel did not differ between patch reefs that have cleaner wrasse present and those from which
cleaners had been experimentally removed over a 12-year period. There was a clear difference in
the number of gnathiids according to day and night. The results from this study suggest that factors
related to cleaner wrasse presence other than gnathiids alone in this system may explain the benefits
that occur to damselfishes.
Diel patterns
Although the number of gnathiids did not differ with cleaner wrasse presence or absence, it
did differ according to day or night. There was a 2.9 times higher probability that traps sampled at
night would capture a gnathiid than traps that where sampled during the day. The diel variation seen
in this study is consistent with findings using other sampling methods, which have also reported
that the number of gnathiids on caged fish is higher at dawn than at dusk (Grutter and Hendrikz
1999; Sikkel et al. 2006). Although, gnathiids clearly can emerge and infect fish during the day, this
is usually at a much lower rate than during the night, with peak activity being recorded at night and
at pre-dawn periods (Grutter and Hendrikz 1999; Sikkel et al. 2006).
The higher abundance of gnathiid isopods active at night may have arisen as a result of
selective pressures from diurnal parasite predators, principally cleaner fish, that feed on them
during the day (Grutter 2002). As different ontogenetic stages (Grutter et al. 2000; Sikkel et al.
2006) and species (Jones and Grutter 2007) of gnathiids are known to emerge differentially during
either the day or at night, it is possible that some species have evolved to emerge at night when
there are fewer diurnal planktivores, cleaners and microinvertebrate feeders (Helfman 1986; Grutter
1996a), therefore, reducing the risk of predation. Although predation of gnathiid isopods can still
occur from nocturnal cleaning activities as observed in the cleaner shrimp, Urocaridella sp. on coral
reefs (Bonaldo et al. 2014) and on monogeneans by Lysmata amboinensis in tanks (Militz and
Hutson 2015), gut contents analyses reveal that cleaner shrimps consume few gnathiids (Becker and
Grutter 2004). Therefore, it is unlikely that cleaning activities by shrimps will have a major impact
on the abundance of gnathiids, compared with the cleaning activities of cleaner fish.
Cleaner wrasse presence
We found that there was no difference in the probability that a gnathiid was present in a trap
on reefs with (0.148) and without cleaner wrasse (0.167). As gnathiid presence did not differ
according to cleaner wrasse presence, it may therefore not explain some of the positive effects
observed on fish from reefs with cleaner wrasse present. For example, the smaller size distribution

96
(Waldie et al. 2011) of P. amboinensis and P. moluccensis and reduced growth rate (Clague et al.
2011) reported in earlier studies which used the same experimental reefs may not be due to
differing levels of gnathiids on these fishes. Indeed Clague et al. (2011) showed that the number of
copepods was higher on larger P. moluccensis from reefs that have had cleaners removed than on
reefs with cleaners and suggested this could explain their reduced growth rate. There is evidence
that infection by copepods can affect the growth, gonad mass and survival of fish (Finley and
Forrester 2003; Palacios-Fuentes et al. 2012). Other ectoparasites such as cymothoid isopods have
also been shown to affect growth and survival in reef fishes (Adlard and Lester 1994; Fogelman and
Grutter 2008). It is thus possible that other ectoparasites such as copepods and cymothoid isopods,
in isolation or even in synergy with gnathiids, may cause this downward shift in size. However, we
did not find any cymothoids in the traps. Similarly, the recent study that showed that the abundance
of recruiting damselfishes was higher on reefs where cleaner wrasse are present (Sun et al. in
review, Chapter 3) is thus unlikely due to higher gnathiid numbers.
Both the prevalence and number of gnathiid isopods was low, with only 16 % of traps over a
24 h time period having at least one gnathiid, with the number of gnathiids ranging only from one to
three gnathiids per trap. This low prevalence and number of gnathiids is consistent with previous
studies that demonstrated that small juvenile P. amboinensis (mean TL ± SE = 14.28 ± 0.26) are
rarely infected with gnathiids, with instantaneous prevalence being 3 and 3.5 % and never more
than one gnathiid per infected fish (Grutter et al. 2011; Sun et al. 2012). The feeding success of a
micropredator should increase as host mass increases, as the likelihood of being detected and thus
potentially being preyed upon decreases (Kuris and Lafferty 2000). This could explain the lower
number of gnathiids we recorded infecting the relatively small-sized P. amboinensis.
Due to the low prevalence of gnathiids infecting these fish, it is unlikely that gnathiids are
the likely mechanisms that have caused the higher abundance of damselfish recruits on reefs with
cleaner wrasse than on reefs without. Therefore, it is likely that other factors related to cleaner
wrasse presence could explain those results. For example, decreased aggression from predators
when in the presence of cleaner wrasse (Cheney et al. 2008) or reduced stress via tactile stimulation
provided by cleaner wrasse (Soares et al. 2011).
It is also possible that we could not detect a difference due to cleaner wrasse presence with
the sentinel traps due to the sampling method. The amount of sampling time (24 h) may have been
too limited to detect slight differences in gnathiid abundance. Convincingly, further sampling would
detect a small yet significant difference in gnathiid abundance between cleaned and non-cleaned
reefs, such a small difference might still be important. We emphasize that although small fish may
only be attacked by a few gnathiids over its life time this might be enough to reduce their growth
and potentially affect their survival.

97
A speculative scenario that agrees with our results as well as the observed beneficial effects
on fishes, is that the advantage to fish for being on reefs with L. dimidiatus could be due to the
length of infection (feeding period) being reduced, rather than the number of gnathiids attacking a
fish. This could occur if cleaners disturb gnathiids and reduced the time they spend (and the blood
they remove) on fish. Previous studies have shown that the feeding rate (time spent ingesting blood)
of gnathiid isopods can vary among individuals, with gnathiids remaining on a host for as little as
30 min or up to 10 h in order to feed (Grutter 2003; Smit et al. 2003; Grutter et al. 2011).
The efficiency of L. dimidiatus in seeking and consuming gnathiids, coupled with the
frequency at which clients are cleaned (e.g. barred rabbit fish, Siganus doliatus, are cleaned on
average every 5 min in a day), means that the risk of predation can be exceedingly high for some
gnathiids (Grutter 1995a). Grutter (2003) showed that for third-stage larval gnathiids, the likelihood
of red blood being present in the gut increased over time, with 20 % of gnathiids being engorged at
15 minutes and 46 % engorged at 30 min. Given that clients such as S. doliatus are cleaned
frequently, the amount of red blood that they may lose from gnathiids should be significantly
reduced on reefs with cleaner wrasse. Gnathiids may not be able to become fully engorged on red
blood within the shortened time frame, due to the client being cleaned (gnathiid predation). This
scenario fits the paradigm of cleaner wrasse driving changes in gnathiid feeding behaviour,
ultimately limiting their ability to adversely affect their host and thus conferring benefits upon fish
clients.
Although studies have shown a clear size preference by L. dimidiatus for larger host
individuals (Grutter 1995a; Grutter et al. 2005; Clague et al. 2011), it should be emphasised that a
reduction in parasitic numbers would be beneficial to any host, regardless of how frequently it may
be cleaned. A single gnathiid can consume up to 85 % of the total blood volume of a settlement-
stage damselfish (Grutter et al. 2011) and might transmit blood parasites and other infectious agents
(Davies et al. 2009; Curtis et al. 2013). Therefore, fish species that are cleaned less regularly (e.g. P.
moluccensis, which are only cleaned on average once per 30 min) (Grutter 1995a) would still
benefit from a reduction in gnathiid feeding time due to a reduced blood loss and exposure to
parasite transmission.
Previous studies have clearly demonstrated that there is an advantage for fish being on reefs
with cleaners present (e.g. Grutter 1999a; Soares et al. 2011; Waldie et al. 2011). In this light the
results of this study showing no differences in relation to cleaner wrasse presence are surprising. In
consideration, these results suggest that the benefits gained from being on reefs with cleaner wrasse
(such as increased recruitment), could relate to the minimisation of the infection that do occur. In
summary, this study demonstrates that the relationship between cleaner wrasse and their clients is

98
complex and that factors other than the abundance of gnathiid isopods in the environment may
explain some of the advantages to fish of being on reefs where cleaner wrasse are present.
Acknowledgements
We thank E. C. McClure, M. De Brauwer, F. Cortesi, G. A. C. Phillips and A. E. Winters for their
help in the field; S. P. Blomberg for statistical advice, P. C. Sikkel for advice on sentinel trap and
experimental design and R. Q-Y. Yong for comments on earlier drafts of the manuscript. We are
particularly grateful for the support of the staff of the Lizard Island Research Station. This work was
funded by a Sea World Research and Rescue Foundation, Australia (SWRRFI) awarded to D. Sun,
A. S. Grutter and T. H. Cribb and funds from the Australian Research Council awarded to A. S.
Grutter.

99
Chapter 6: General Discussion

100
Chapter 6: General Discussion
The Preamble to this thesis referred to the complexity of interactions between three groups of
organisms – hosts, parasites and cleaners. Although a lot of work has been done on the various
elements of this system prior to my study, and although substantial progress has been made here,
the questions arising from the study appear to leave at least as much to be done as when I started.
From the study of the diversity of digenean trematodes of adult pomacentrid fishes at Lizard
Island it was established that digenean richness is low relative to that of other coral reef fish groups.
Host-specificity ranges dramatically from a few taxa that are strongly oioxenic through to several
that are clearly stenoxenic and others that are seemingly euryxenic. In comparison with the known
fauna of Heron Island, the fauna at Lizard Island was generally comparable but with several key
distinctions. Questions arising from these findings start with the problems of identification. It was a
surprise to find evidence of so much cryptic richness among specimens of just one of the 19
morphospecies in the study. Hysterolecitha nahaensis clearly is deserving of and requires dedicated
independent study. There is no a posteriori explanation for why this one species has radiated so
dramatically. In terms of the discovery of the total richness of the system, the distribution of
oioxenic species presents a challenge. Not only do we not understand why only one faustulid is
present in one species of Chromis, one bivesiculid in Acanthochromis polyacanthus and Lepotrema
adlardi in just one species of Abudefduf – we are left uncertain as to what richness may remain
undetected in the pomacentrid species that have never been examined. In terms of beta diversity, it
might have been predicted that, as found, there would be some relatively low-level differences
between the fauna found at Heron and Lizard Islands. What remains completely unexplained is why
the species that have limited distributions are so restricted thus. Further, the striking differences in
mean richness and mean prevalence between Heron and Lizard Islands remain unexplained. For
convincing explanations to emerge on such topics we will need at least to understand the life-cycles
of these taxa. Presently not a single life-cycle of a pomacentrid-infecting digenean is known.
This study has also highlighted the importance of the presence of cleaner wrasse in the
ecology of young reef fishes. I demonstrated that the positive effects of cleaner wrasse presence on
adult fish population size reported in other studies (Bshary 2003; Grutter et al. 2003; Waldie et al.
2011), begins during the recruitment period by enhancing the abundance of damselfish recruits.
Thus, we now know that cleaner wrasse can influence the population dynamics from the start of the
recruitment period. Although cleaner wrasse species are important key stone mutualist on coral
reefs, they are collected in large numbers for the aquarium trade; 20,000 individuals have been
caught in Sri Lanka in one year alone. The removal of such a key organism in large numbers should

101
have a detrimental effect on the biodiversity of reef fishes. Therefore, studies on reef fish recovery
on coral reefs should consider the beneficial role of cleaner wrasse alongside that of other key
organisms. However, it remains unclear why the abundance of only two species of damselfishes
(Chrysiptera rollandi and Pomacentrus amboinensis) which differ from each other in regards to
ecology (e.g. habitat and social structure), were affected by the presence of a cleaner wrasse, when
similar species showed no differences. It is possible that factors other than the presence of cleaner
wrasse dominate when it comes to microhabitat selection during their recruitment. Despite the
overall higher abundance of recruiting damselfishes on reefs where cleaner wrasse are present, it is
not understood how these fish are selecting these particular reefs. As cleaner wrasse are inactive and
shelter within the reef matrix at night when the majority of settlement-stage fish settle, it is possible
that they are selecting reefs based on olfactory in addition to visual cues. This hypothesis is yet to
be tested, however, doing so will provide greater insight into the dynamics that occur during the
recruitment period.
The results from chapter 4 showed that when given the option, a young damselfish would
preferentially select a microhabitat that is adjacent to a cleaner wrasse. This finding indicates that
cleaner wrasse may influence small-scale post-settlement migration. Field observations indicated
that small recruits are rarely cleaned by juvenile cleaner wrasse. It was therefore unsurprising that
the damselfish that were preferentially cleaned by juvenile L. dimidiatus were larger. It is thus
likely that indirect rather than direct benefits are the driver for such small recruits seeking proximity
to a cleaner. As cleaner wrasse are known to consume large quantities of parasites per day such as
gnathiids, a reduction in gnathiid abundance within cleaning stations of juvenile cleaner wrasses
could be one possible benefit. However, it is not yet known if the abundance of gnathiids are
actually reduced within cleaning stations. Thus, further studies are required to explore this scenario.
It is also not clear what other indirect benefits a recruit may obtain from being in a microhabitat that
is near a cleaner wrasse. If gnathiid infestation is lower in the vicinity of a cleaner wrasse, we
should expect both growth and survival to increase, thus possibly explaining the results found in
this study. However, alternatively, or in addition, selection by young damselfish of microhabitats
near cleaners may simply be an investment where the return only occurs when recruits have grown
significantly.
Given that past studies have shown reduction in gnathiid infestation on hosts and that
cleaner wrasse are known to consume enormous numbers of gnathiids when cleaner wrasse are
present, it was somewhat surprising to find no effect of cleaner wrasse presence in relation to the
abundance of gnathiids caught in sentinel traps. As this study only examined the infection of one
species of damselfishes, future studies should examine whether the infection of gnathiids is
consistently the same or different for other species of damselfishes. It is possible that different

102
species of damselfishes may be infected with more gnathiids. However, the results shown in this
study may suggest that the benefits of fish being on reefs with cleaner wrasse are not significantly
from a reduction in gnathiid isopod load but due to exposure of gnathiid infection being minimised
by cleaner wrasse presence. Further examination is required to test this idea. If supported, the
implications could be an intriguing “minimal effect/win/win” outcome whereby the population of
gnathiids is not materially affected by the presence of cleaners, the cleaners benefit from their
presence by the provision of most of their food, and client fish benefit from the presence of the
cleaners by the reduction of the impact of gnathiid parasitism. Could there be further complexities
such as cleaners preferentially eating engorged gnathiids? Only further studies will tell.

103
References
Adam TC (2010) High-quality habitat and facilitation ameliorate competitive effects of prior residents on new settlers.
Oecologia 166:121-130
Adamson ML, Caira JN (1994) Evolutionary factors influencing the nature of parasite specificity. Parasitology
109:S85-S95
Adlard RD, Lester RJG (1994) Dynamics of the interaction between the parasitic isopod, Anilocra pomacentri, and the
coral reef fish, Chromis nitida. Parasitology 109:311-324
Aiken HM, Bott NJ, Mladineo I, Montero FE, Nowak BF, Hayward CJ (2007) Molecular evidence for cosmopolitan
distribution of platyhelminth parasites of tunas (Thunnus spp.). Fish Fish 8:167-180
Allen GR (2008) Conservation hotspots of biodiversity and endemism for Indo-Pacific coral reef fishes. Aquat Conserv
18:541-556
Almany GR (2003) Priority effects in coral reef fish communities. Ecology 84:1920-1935
Almany GR, Webster MS (2006) The predation gauntlet: early post-settlement mortality in reef fishes. Coral Reefs
25:19-22
Anderson GR, Barker SC (1998) Inference of phylogeny and taxonomy within the Didymozoidae (Digenea) from the
second internal transcribed spacer (ITS2) of ribosomal DNA. Syst Parasitol 41:87-94
Anderson MJ, Gorley RN, Clarke KR (2008) PERMANOVA+ for Primer: Guide to software and statistical methods,
Plymouth
Arnal C, Côté IM (2000) Diet of broadstrip cleaning gobies on a Barbadian reef. J Fish Biol 57:1075-1082
Arnal C, Morand S (2001) Importance of ectoparasites and mucus in cleaning interactions in the Mediterranean cleaner
wrasse Symphodus melanocercus. Mar Biol 138:777-784
Barber I, Hoare D, Krause J (2000) Effects of parasites on fish behaviour: a review and evolutionary perspective. Rev
Fish Biol Fish 10:131-165
Barker SC, Bray RA, Cribb TH (1993) Preptetos cannoni n. sp. (Digenea: Lepocreadiidae) from Siganus lineauts
(Teleostei: Siganidae) from the southern Great Barrier Reef, Australia. Syst Parasitol 26:151-155
Barker SC, Cribb TH, Bray RA, Adlard RD (1994) Host-parasite associations on a coral reef: Pomacentrid fishes and
digenean trematodes. Int J Parasitol 24:643-647
Barnett LJ, Miller TL, Cribb TH (2014) A review of the currently recognised zoogonid cercariae, including the
identification and emergence ecology of Cercaria capricornia XI (Digenea: Zoogonidae) from Nassarius
olivaceus (Gastropoda: Nassariidae) in Central Queensland, Australia. Folia Parasitol (Praha) 61:322-330
Bartoli P, Gibson DI (2007) Synopsis of the life cycles of Digenea (Platyhelminthes) from lagoons of the northern coast
of the western Mediterranean. J Nat Hist 41:1553-1570
Bartoli P, Gibson DI, Bray RA (2005) Digenean species diversity in teleost fish from a nature reserve off Corsica,
France (Western Medterranean), and a comparison with other Mediterranean regions. J Nat Hist 39:47-70
Bartoli P, Bray RA, Gibson DI (2006) Four closely related but forgotten species of Rhipidocotyle Diesing, 1858
(Digenea : Bucephalidae) in fishes from European seas. Syst Parasitol 65:129-149
Bascompte J, Jordano P (2007) Plant-animal mutualistic networks: The architecture of biodiversity. Annu Rev Ecol
Evol Syst 38:567-593
Bay LK, Jones GP, McCormick MI (2001) Habitat selection and aggression as determinants of spatial segregation
among damselfish on a coral reef. Coral Reefs 20:289-298
Beasley VR, Faeh SA, Wikoff B, Eisold J, Nichols D, Cole R, Schotthoefer AM, Staehle C, Greenwell M, Brown LE
(2005) Risk factors and the decline of the northern cricket frog, Acris crepitans: evidence for involvement of
herbicides, parasitism, and habitat modifcations. In: Lanoo MJ (ed) Amphibian declines: the conservation
status of United States species. University of California Press, Berkeley, CA,
Becker JHA, Grutter AS (2004) Cleaner shrimp do clean. Coral Reefs 23:515-520
Becker JHA, Curtis LM, Grutter AS (2005) Cleaner shrimp use a rocking dance to advertise cleaning service to clients.
Curr Biol 15:760-764
Biani NB, Mueller UG, Wcisio WT (2009) Cleaner mites: Sanitary mutualism in the miniature ecosystem of neotropical
bee nests. The American Naturalist 173:841-847
Binning SA, Roche DG, Layton C (2013) Ectoparasites increase swimming costs in a coral reef fish. Biol Lett
9:20120927
Blair D, Bray RA, Barker SC (1998) Molecules and morphology in phylogenetic studies of the Hemiuroidea (Digenea:
Trematoda: Platyhelminthes). Mol Phylogenet Evol 9:15-25
Bonaldo RM, Grutter AS, Sazima I, Krajewski JP (2014) 24/7 service: nocturnal cleaning in a tropical Indo-Pacific reef.
Mar Biodivers:1-2
Bond WJ (1994) Keystone species. Springer Berlin Heidelberg
Bonin MC, Almany GR, Jones GP (2011) Contrasting effects of habitat loss and fragmentation on coral-associated reef
fishes. Ecology 92:1503-1512

104
Bosiger YJ, McCormick MI (2014) Temporal links in daily activity patterns between coral reef predators and their prey.
PLoS ONE 9:e111723
Bott NJ, Cribb TH (2009) Prosorhynchine trematodes (Digenea: Bucephalidae) from epinephelines (Perciformes:
Serranidae) on the Great Barrier Reef, Australia. Syst Parasitol 72:57-69
Bott NJ, Miller TL, Cribb TH (2013) Bucephalidae (Platyhelminthes: Digenea) of Plectropomus (Serranidae:
Epinephelinae) in the tropical Pacific. Parasitol Res 112:2561-2584
Bowles J, Hope M, Tiu WU, Liu X, McManus DP (1993) Nuclear and mitochondrial genetic markers highly conserved
between Chinese and Philippines Schistosoma japonicum. Acta Trop 55:217-229
Bray R, Cribb T (2002) Lepocreadiinae lepocreadiids (Digenea) from tetraodontiform fishes in the waters off Tasmania,
Queensland and Moorea, French Polynesia. Acta Parasitol 47:6-13
Bray R, Cribb T (2004) Species of Lecithocladium Luhe, 1901 (Digenea, Hemiuridae) from Australian marine fishes,
with a description of a new species from various hosts off eastern Australia. Acta Parasitol 49:3-11
Bray RA (1982) Two new species of Bacciger Nicoll, 1914 (Digenea: Fellodistomidae) from mullet in Australia. J Nat
Hist 16:23-29
Bray RA (2001) Family Tandanicolidae Johnston, 1927. In: Gibson DI, Jones A, Bray RA (eds) Keys to the Trematoda,
Volume 1. CABI Publishing and the Natural History Museum, Wallingford,
Bray RA (2002) Superfamily Gymnophalloidea Odhner, 1905. In: Gibson DI, Jones A, Bray RA (eds) Keys to the
Trematoda Vol 1. CAB International, Wallingford, pp243-244
Bray RA (2008) Family Faustulidae Poche, 1926. In: Bray RA, Gibson DI, Jones A (eds) Keys to the Trematoda Vol 3.
CABI Publishing and the Natural History Museum, Wallingford, pp509-522
Bray RA, MacKenzie K (1990) Aponurus laguncula Looss, 1907 (Digenea: Lecithasteridae): a report from herring,
Clupea harengus L., in the easter English Channel and a review of its biology. Syst Parasitol 17:115-124
Bray RA, Cribb TH (1996) Two Lepotrema Ozaki, 1932 species (Digenea: Lepocreadiidae) from marine fishes from
the southern Great Barrier Reef, Queensland, Australia. Syst Parasitol 35:111-117
Bray RA, Cribb TH (1998) Lepocreadiidae (Digenea) of Australian coastal fishes: new species of Opechona Loos,
1907, Lepotrema Ozaki, 1932 and Bianium Stunkard, 1930 and comments on other species reported for the
first time or poorly known in Australian waters. Syst Parasitol 41:123-148
Bray RA, Cribb TH, Barker SC (1993a) The Hemiuroidea (Digenea) of pomacentrid fishes (Perciformes) from Heron
Island, Queensland, Australia. Syst Parasitol 24:159-184
Bray RA, Cribb TH, Barker SC (1993b) The Lepocreadiidae (Digenea) of pomacentrid fishes (Perciformes) from Heron
Island, Queensland, Australia. Systematic Parasitology 26:189-200
Bray RA, Cribb TH, Barker SC (1994) Fellodistomidae and Lepocreadiidae (Platyhelminthes: Digenea) from
chaetodontid fishes (Perciformes) from Heron Island, southern Great Barrier Reef, Queensland, Australia.
Invertebr Taxon 8:545-581
Bray RA, Littlewood DTJ, Herniou EA, Williams B, Henderson RE (1999) Digenean parasites of deep-sea teleosts: a
review and case studies of intrageneric phylogenies. Parasitology 119:S125-S144
Bray RA, Waeschenbach A, Cribb TH, Weedall GD, Dyal P, Littlewood DTJ (2009) The phylogeny of the
Lepocreadioidea (Platyhelminthes, Digenea) inferred from nuclear and mitochondrial genes: Implications for
their systematics and evolution. Acta Parasitol 54:310-329
Bronstein JL (1994) Conditional outcomes in mutualistic interactions. Trends Ecol Evol 9:214-217
Bronstein JL, Dieckmann U, Ferrière R (2004) Evolutionary Conservation Biology. Cambridge University Press,
Cambridge
Brown BL, Creed RP, Skelton J, Rollins MA, Farrell KJ (2012) The fine line between mutualism and parasitism:
complex effects in a cleaning symbiosis demonstrated by mutliple field experiments. Oecologia 170:199-207
Brown SP, Renaud F, Guegan JF, Thomas F (2001) Evolution of trophic transmission in parasites: the need to reach a
mating place? J Evol Biol 14:815-820
Bshary R (2003) The cleaner wrasse, Labroides dimidiatus, is a key organism for reef fish diversity at Ras Mohammed
National Park, Egypt. J Anim Ecol 72:169-176
Bshary R, Würth M (2001) Cleaner fish Labroides dimidiatus manipulate client reef fish by providing tactile
stimulation. Proc R Soc Biol Sci Ser B 268:1495-1501
Bshary R, Oliveira RF, Oliveira TS, Canário AV (2007) Do cleaning organisms reduce the stress response of client reef
fish? Front Zool 4:21
Bykhovskaya-Pavlovskaya I, Gusev A, Dubinia M, Izyumova N, Smirnova T, Sokolovskaya I, Shtein G, Shul'man S,
Epshtein V (1962) Key to parasites of freshwater fish of the USSR Akad. Sci USSR Zool Inst Keys to fauna of
the USSR 80:180-223
Caley MJ, Carr MH, Hixon MA, Hughes TP, Jones GP, Menge BA (1996) Recruitment and the local dynamics of open
marine populations. Annu Rev Ecol Syst 27:477-500
Candolin U, Voigt HR (2001) No effect of a parasite on reproduction in stickleback males: a laboratory artefact?
Parasitology 122:457-464
Carrassón M, Cribb TH (2014) Benign effect of the fish parasitic isopod Ceratothoa cf. imbricata on Selenotoca
multifasciata (Scatophagidae) from Australia. Dis Aquat Organ 110:173-180

105
Carreras-Aubets M, Repullés-Albelda A, Kostadinova A, Carrassón M (2011) A new cryptic species of Aponurus
Looss, 1907 (Digenea: Lecithasteridae) from Mediterranean goatfish (Teleostei: Mullidae). Syst Parasitol
79:145-159
Ceccarelli D (2007) Modification of benthic communities by territorial damselfish: a multi-species comparison. Coral
Reefs 26:853-866
Ceccarelli D, Jones G, McCook L (2005) Effects of territorial damselfish on an algal-dominated coastal coral reef.
Coral Reefs 24:606-620
Chambers JM (1992) Linear models. In: Chambers JM, Hastie TJ (eds) Statistical models in S. Wadsworth &
Brooks/Cole, Pacific Grove, California, pp95-144
Cheney KL, Côté IM (2001) Are Caribbean cleaning symbioses mutualistic? Costs and benefits of visiting cleaning
stations to longfin damselfish. Anim Behav 62:927-933
Cheney KL, Côté IM (2003) The ultimate effect of being cleaned: does ectoparasite removal have reproductive
consequences for damselfish clients? Behav Ecol 14:892-896
Cheney KL, Bshary R, Grutter AS (2008) Cleaner fish cause predators to reduce aggression toward bystanders at
cleaning stations. Behav Ecol 19:1063-1067
Cheney KL, Grutter AS, Blomberg SP, Marshall NJ (2009) Blue and yellow signal cleaning behavior in coral reef
fishes. Curr Biol 19:1283-1287
Chun SK, Chang DS, Park CK, Kim YG, Rho YG (1981) Basic studies for the production of the hard clam Meretrix
lusoria (Roding) in Jeonnbug farming area. Bulletin of Fisheries Research and Development Agency 26:7-36
Clague GE, Cheney KL, Goldizen AW, McCormick MI, Waldie PA, Grutter AS (2011) Long-term cleaner fish
presence affects growth of a coral reef fish. Biol Lett 6:863-865
Coile AM, Sikkel PC (2013) An experimental field test of susceptibility to ectoparasitic gnathiid isopods among
Caribbean reef fishes. Parasitology 140:888-896
Coker DJ, Pratchett MS, Munday PL (2012a) Influence of coral bleaching, coral mortality and conspecific aggression
on movement and distribution of coral-dwelling fish. J Exp Mar Biol Ecol 414-415:62-68
Coker DJ, Graham NAJ, Pratchett MS (2012b) Interactive effects of live coral and structural complexity on the
recruitment of reef fishes. Coral Reefs 31:919-927
Cooper WJ, Smith JJ, Westneat MW (2009) Exploring the radiation of a diverse reef fish family: phylogenetics of the
damselfishes (Pomacentridae), with new classifications based on molecular analyses of all genera. Mol
Phylogenet Evol 52:1-16
Coppock AG, Gardiner NM, Jones GP (2013) Olfactory discrimination in juvenile coral reef fishes: Responses to
conspecifics and corals. J Exp Mar Biol Ecol 443:21-26
Côté IM (2000) Evolution and ecology of cleaning symbioses in the sea. Oceanography and Marine Biology 38:311-
355
Côté IM, Arnal C, Reynolds JD (1998) Variation in posing behaviour among fish species visiting cleaning stations. J
Fish Biol 53 (sA):256-266
Cribb T, Bray R (1994) A review of the Tandanicolinae (= Monodhelminthinae)(Digenea: Fellodistomidae), with a
description of Prosogonarium angelae n. sp. Syst Parasitol 27:187-205
Cribb TH, Bray RA (2010) Gut wash, body soak, blender and heat-fixation: approaches to the effective collection,
fixation and preservation of trematodes of fishes. Syst Parasitol 76:1-7
Cribb TH, Bray RA, Barker SC (1992) Zoogonidae (Digenea) from southern Great Barrier Reef fishes with a
description of Steganoderma (Lecithostaphylus) gibsoni n. sp. Syst Parasitol 23:7-12
Cribb TH, Bray RA, Barker SC (1994a) Bivesiculidae and Haplosplanchnidae (Digenea) from fishes of the southern
Great Barrier Reef, Australia. Syst Parasitol 28:81-97
Cribb TH, Anderson GR, Bray RA (1999) Faustulid trematodes (Digenea) from marine fishes of Australia. Syst
Parasitol 44:119-138
Cribb TH, Anderson GR, Adlard RD, Bray RA (1998) A DNA-based demonstration of a three-host life cycle for the
Bivesiculidae (Platyhelminthes: Digenea). Int J Parasitol 28:1791-1795
Cribb TH, Miller TL, Bray RA, Cutmore S, C. (2014a) The sexual adult of Cercaria praecox Walker, 1971 (Digenea:
Fellodistomidae), with the proposal of Oceroma n. g. Syst Parasitol 88:1-10
Cribb TH, Miller TL, Bray RA, Cutmore SC (in press) The sexual adult of Cercaria praecox Walker, 1971 (Digenea:
Fellodistomidae), with the proposal of Oceroma n. g. Syst Parasitol
Cribb TH, Bray RA, Barker SC, Adlard RD, Anderson GR (1994b) Ecology and diversity of digenean trematodes of
reef and inshore fishes of Queensland. Int J Parasitol 24:851- 860
Cribb TH, Bray RA, Littlewood DTJ, Pichelin SP, Herniou EA (2001) The Digenea. In: Littlewood DTJ, Bray RA (eds)
Interrelationships of the Platyhelminthes. Taylor and Francis, London, pp168-185
Cribb TH, Bott NJ, Bray RA, McNamara MKA, Miller TL, Nolan MJ, Cutmore SC (2014b) Trematodes of the Great
Barrier Reef, Australia: emerging patterns of diversity and richness in coral reef fishes. Int J Parasitol 44:929-
939
Curran SS, Tkach VV, Overstreet RM (2006) A review of Polylekithum Arnold, 1934 and its familial affinities using
morphological and molecular data, with description of Polylekithum catahoulensis sp. nov. Acta Parasitol
51:238-248

106
Curtis LM, Grutter AS, Smit NJ, Davies AJ (2013) Gnathia aureamaculosa, a likely definitive host of Haemogregarina
balistapi and potential vector for Haemogregarina bigemina between fishes of the Great Barrer Reef,
Australia. Int J Parasitol 43:361-370
Danilowicz BS (1996) Choice of coral species by naive and field caught damselfishes. Copeia 3:735-739
Davies AJ, Smit NJ (2001) The life cycle of Haemogregarina bigemina (Adeleina: Haemogregarinidae) in South
African hosts. Folia Parasitol (Praha) 48:169-177
Davies AJ, Curtis L, Grutter AS, Smit NJ (2009) Suspected viral erythrocytic necrosis (VEN) in a juvenile blackbar
triggerfish, Rhinecanthus aculeatus, from Lizard Island, Great Barrier Reef, Australia. Marine Biodiversity
Records 2:e149
Davies AJ, Smit NJ, Hayes PM, Seddon AM, Wertheim D (2004) Haemogregarina bigemina (Protozoa: Apicomplexa:
Adeleorina) - Past, present and future. Folia Parasitol (Praha) 51:99-108
Derbel H, Chaari M, Neifar L (2011) Redescription of Rhipidocotyle galeata (Rudolphi, 1819) (Digenea,
Bucephalidae), the type species of Rhipidocotyle Diesing, 1907. Zoosystema 33:133-139
Diaz EP, Bray RA, Cribb TH (2013) Paradiscogaster flindersi and P. oxleyi n. sp. (Digenea: Faustulidae): overlapping
host and geographical distributions in corallivore chaetodontid fishes in the tropical Indo-west Pacific. Syst
Parasitol 86:87-99
Diaz PE, Cribb TH (2013) Paradiscogaster leichhardti sp. nov. (Digenea: Faustulidae) in Chaetodontoplus meredithi
(Perciformes: Pomacanthidae) from Heron Island, Great Barrier Reef. Memoirs of the Queensland Museum
58:49-53
Diaz PE, Bray RA, Cutmore SC, Ward S, Cribb TH (2015) A complex of species related to Paradiscogaster glebulae
(Digenea: Faustulidae) in chaetodontid fishes (Teleostei: Perciformes) of the Great Barrier Reef. Parasitol Int
Dimitrov GI, Bray RA, Gibson DI (1999) A redescription of Pseudobacciger harengulae (Yamaguti, 1938) (Digenea:
Faustulidae) from Sprattus sprattus phalericus (Risso) and Engraulis encrasicholus ponticus Alexandrov off
the Bulgarian Black Sea coast, with a review of the genus Pseudobacciger Nahhas & Cable, 1964. Syst
Parasitol 43:133-146
Dyer WG, Williams EH, Williams LB (1985) Digenetic trematodes of marine fishes of the western and southern coasts
of Puerto Rico. Proc Helminthol Soc Wash 52:85-94
Dyer WG, Williams EH, Bunkley-Williams L (1988) Digenetic trematodes of marine fishes of Okinawa, Japan. J
Parasitol 74:638-645
Edgar RC (2004) MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res
32:1792-1797
Euzet L, Combes Cle (1980) Les problèmes de l'espèce chez les animaux parasites. Bull Soc Zool Fr 40:239-285
Feary DA, Almany GR, McCormick MI, Jones GP (2007) Habitat choice, recruitment and the response of coral reef
fishes to coral degradation. Oecologia 153:727-737
Ferguson JA, Koketsu W, Ninomiya I, Rossignol PA, Jacobson KC, Kent ML (2011) Mortality of coho salmon
(Oncorhynchus kisutch) associated with burdens of multiple parasite species. Int J Parasitol 41:1197-1205
Finley RJ, Forrester GE (2003) Impact of ectoparasites on the demography of a small reef fish. Mar Ecol Prog Ser
248:305-309
Fishelson L (1998) Behaviour, socio-ecology and sexuality in damselfishes (Pomacentridae). Ital J Zool 65:387-398
Fogelman RM, Grutter AS (2008) Mancae of the parasitic cymothoid isopod, Anilocra apogonae: early life history,
host-specificty, and effect on growth and survival of preferred young cardinal fishes. Coral Reefs 27:685-693
Fogelman RM, Kuris AM, Grutter AS (2009) Parasitic castration of a vertebrate: effect of the cymothoid isopod,
Anilocra apogonae, on the five-lined cardinalfish, Cheilodipterus quinquelineatus. Int J Parasitol 39:577-583
Froese R, Pauly D (2012) FishBase (version 02/2014), www.fishbase.org
Gaevskaja AV (1996) [New records of trematodes from eastern Atlantic fishes] (In Russian). Parasitlogiya 30:504-509
Gaevskaja AV, Naidenova NN (1996) [Revision of Bacciger bacciger (Trematoda: Fellodistomidae) from fishes of the
Black and Mediterranean Seas] (In Russian). Parazitologiya (St Petersb) 30:39-44
Gibson DI, Bray RA (1979) The Hemiuroidea: terminology, systematics and evolution. Bulletin of the British Museum
Natural History (Zoology) 36:35-146
Gorlick DL, Atkins PD, Losey GS (1987) Effect of cleaning by Labroides dimidiatus (Labridae) on an ectoparasite
population infecting Pomacentrus vaiuli (Pomacentridae) at Enewetak Atoll. Copeia 1:41-45
Green AL (1994) The early life history of labroid fishes at Lizard Island, Northern Great Barrier Reef. James Cook
University,
Green AL (1998) Spatio-temporal patterns of recruitment of labroid fishes (Pisces: Labridae and Scaridae) to
damselfish territories. Environ Biol Fishes 51:235-244
Green BS (2008) Maternal effects in fish populations. Adv Mar Biol 54:1-105
Grorud-Colvert K, Sponaugle S (2011) Variability in water temperature affects trait-mediated survival of a newly
settled coral reef fish. Oecologia 165:675-686
Grutter AS (1994) Spatial and temporal variations of the ectoparasites of seven reef fish species from Lizard Island and
Heron Island, Australia. Mar Ecol Prog Ser 115:21-30
Grutter AS (1995a) Relationship between cleaning rates and ectoparasite loads in coral reef fishes. Mar Ecol Prog Ser
118:51-58

107
Grutter AS (1995b) A comparison of methods for sampling ectoparasites from coral reef fishes. Mar Freshw Res
46:897-903
Grutter AS (1996a) Parasite removal rates by the cleaner wrasse, Labroides dimidiatus. Mar Ecol Prog Ser 130:61-70
Grutter AS (1996b) Experimental demonstration of no effect by the cleaner Labroides dimidiatus (Cuvier and
Valenciennes) on the host fish Pomacentrus moluccensis (Bleeker). J Exp Mar Biol Ecol 196:285-298
Grutter AS (1997) Spatiotemporal variation and feeding selectivity in the diet of the cleaner fish Labroides dimidiatus.
Copeia 1997:346-355
Grutter AS (1999a) Cleaner fish really do clean. Nature 398:672-673
Grutter AS (1999b) Infestation dynamics of gnathiid isopod juveniles parasitic on the coral-reef fish Hemigymnus
melapterus (Labridae). Mar Biol 135:545-552
Grutter AS (2000) Ontogenetic variation in the diet of the cleaner fish Labroides dimidiatus and its ecological
consequences Mar Ecol Prog Ser 197:241-246
Grutter AS (2001) Parasite infection rather than tactile stimulation is the proximate cause of cleaning behaviour in reef
fish. Proc R Soc Biol Sci Ser B 268:1361-1365
Grutter AS (2002) Cleaning symbioses from the parasites' perspective. Parasitology 124:65-81
Grutter AS (2003) Feeding ecology of the fish ectoparasite Gnathia sp. (Crustacea: Isopoda) from the Great Barrier
Reef, and its implications for cleaning behaviour. Mar Ecol Prog Ser 2559:295-302
Grutter AS (2004a) Cleaner fish use tactile dancing behaviour as a preconflict management strategy. Curr Biol 14:1080-
1083
Grutter AS (2004b) Cleaner fish use tactile dancing behaviour as a preconflict management strategy. Current biology
14:1080-1083
Grutter AS (2008) Interactions between gnathiid isopods, cleaner fish and other fishes on Lizard Island, Great Barrier
Reef. J Fish Biol 73:2094-2109
Grutter AS, Poulin R (1998) Intraspecific and interspecific relationships between host size and the abundance of
parasitic larval gnathiid isopods on coral reef fishes. Mar Ecol Prog Ser 164:263-271
Grutter AS, Hendrikz J (1999) Diurnal variation in the abundance of juvenile parasitic gnathiid isopods on coral reef
fish- implications for parasite-cleaner fish interactions. Coral Reefs 18:187-191
Grutter AS, Lester RJG (2002) Cleaner fish Labroides dimidiatus reduce 'temporary' parasitic corallanid isopods on the
coral reef fish Hemigymnus melapterus. Mar Ecol Prog Ser 234:247-255
Grutter AS, Lester RJG, Greenwood JG (2000) Emergence rates from the benthos of the parasitic juveniles of gnathiid
isopods. Mar Ecol Prog Ser 207:123-127
Grutter AS, Murphy J, Choat H (2003) Cleaner fish drives local fish diversity on coral reefs. Curr Biol 13:64-67
Grutter AS, Glover S, Bshary R (2005) Does client size affect cleaner fish choice of client? An empirical test using
client fish models. J Fish Biol 66:1748-1752
Grutter AS, Pickering JL, McCallum H, McCormick MI (2008) Impact of micropredatory gnathiid isopods on young
coral reef fishes. Coral Reefs 27:655-661
Grutter AS, Cribb TH, McCallum H, Pickering JL, McCormick MI (2010) Effects of parasites on larval and juvenile
stages of the coral reef fish Pomacentrus moluccensis. Coral Reefs 29:31-40
Grutter AS, Crean AJ, Curtis LM, Kuris AM, Warner RR, McCormick MI (2011) Indirect effect of an ectoparasite
reduce successful establishment of a damselfish at settlement. Funct Ecol 25:586-594
Grutter AS, Blomberg SP, Fargher B, Kuris AM, McCormick MI, Warner RR (in review) A reason for pelagic larval
stages: Small size of coral reef fish at settlement and high mortality risk from gnathiid isopod micropredators
Guégan JF, Lambert A, Lévêque C, Combes C, Euzet L (1992) Can host body size explain the parasite species richness
in tropical freshwater fishes? Oecologia 90
Hall KA, Cribb TH, Barker SC (1999) V4 region of small subunit rDNA inidicates polyphyly of the Fellodistomidae
(Digenea) which is supported by morphology and life-cycle data. Syst Parasitol 43:81-92
Harms KE, Wright SJ, Calderón O, Hernández A, Herre EA (2000) Pervasive density-dependent recruitment enhances
seedling diveristy in a tropical forest. Nature 404:493-495
Hayes PM, Smit NJ, Grutter AS, Davies AJ (2011) Unexpected response of a captive blackeye thicklip, Hemigymnus
melapterus (Bloch), from Lizard Island, Australia, exposed to juvenile isopods Gnathia aureamaculosa
Ferreira & Smit. J Fish Dis 34:563-566
Heinlein J, M., Stier AC, Steele MA (2010) Predators reduce abundance and species richness of coral reef fish recruits
via non-selective predation. Coral Reefs 29:527-532
Helfman GS (1986) Fish behaviour by day, night and twilight The behaviour of teleost fishes. Springer, pp366-387
Heupel MR, Bennett MB (1999) The occurrence, distribution and pathology associated with gnathiid isopod larvae
infecting the epaulette shark, Hemiscyllium ocellatum. International Journal for Parasitology 29:321-330
Hixon MA, Brostoff WN (1983) Damselfish as keystone species in reverse: intermediate disturbance and diversity of
reef algae. Science 220:511-513
Ho HW, Bray RA, Cutmore SC, Ward S, Cribb TH (2014) Two new species of Phyllodistomum Braun, 1899
(Trematoda: Gorgoderidae Looss, 1899) from Great Barrier Reef fishes. Zootaxa 3779:551-562
Hoey AS, McCormick MI (2004) Selective predation for low body condition at the larval-juvenile transition of a coral
reef fish. Oecologia 139:23-29

108
Horthorn T, Bretz F, Westfall P, Hothorn MT (2013) Package 'multcomp' (ed. CRAN) See http://cran.r-
project.org/web/packages/multcomp.
Hudson PJ, Dobson AP, Lafferty KD (2006) Is a healthy ecosystem one that is rich in parasites? Trends Ecol Evol
21:381-385
Huijbers CM, Nagelkerken I, Lössbroek PA, Schulten IE, Siegenthaler A, Holderied MW, Simpson SD (2012) A test of
the senses: Fish select novel habitats by responding to multiple cues. Ecology 93:46-55
Hunter JA, Cribb TH (2010) Transversotrematidae (Plathelminthes: Trematoda) are rich and abundant on Indo-Pacific
fishes. Zootaxa 2442:25-38
Hunter JA, Cribb TH (2012) A cryptic complex of species related to Transversotrema licinum Manter, 1970 from fishes
of the Indo-West Pacific, including descriptions of ten new species of Transversotrema Witenberg, 1944
(Digenea: Transversotrematidae). Zootaxa 3176:1-44
Ichihara A (1974) Hemiurid trematodes from marine fishes near the Tsushima Islands in the Sea of Japan. Third
International Congress of Parasitology
Ichihara A (1986) On the helminths of marine fishes in Sagami Bay (II) Parasitology - quo vadit? Sixth International
Congress of Parasitology
Jacoby CA, Greenwood JG (1988) Spatial, temporal, and behavioural patterns in emergence of zooplankton in the
lagoon of Heron Reef, Great Barrier Reef, Australia. Mar Biol 97:309-328
Johnston SJ (1913) On some Queensland trematodes, with anatomical observations and descriptions of new species and
genera. Q J Microsc Sci 59:361-400
Johnston TH (1927) New trematodes from an Australian siluroid. Trans R Soc S Aust 51:129-136
Jones CM, Grutter AS (2005) Parasitic isopods (Gnathia sp.) reduce haematocrit in captive blackeye thicklip (Labridae)
on the Great Barrier Reef. Journal of Fish Biology 66:860-864
Jones CM, Grutter AS (2007) Variation in emergence of parasitic and predatory isopods among habitats at Lizard
Island, Great Barrier Reef. Mar Biol 150:919-927
Jones CM, Grutter AS (2008) Reef-based micropredators reduce the growth of post-settlement damselfish in captivity.
Coral Reefs 27:677-684
Jones GP (1988) Experimental evaluation of the effects of habitat structure and competitive interactions on the juveniles
of two coral reef fishes. J Exp Mar Biol Ecol 123:115-126
Jordan LKB, Lindeman KC, Spieler RE (2012) Depth-variable settlement patterns and predation influence on newly
settled reef fish (Haemulon spp., Haemulidae). PLoS ONE 7:e50897
Jordano P (2000) Fruits and frugivory. In: Fenner M (ed) Seeds: The ecology of regeneration in natural plant
communities. CABI, Wallingford, UK, pp125-166
Jousson O, Bartoli P, Pawlowski J (2000) Cryptic speciation among intestinal parasites (Trematoda: Digenea) infecting
sympatric host fishes (Sparidae). J Evol Biol 13:778-785
Justine JL (2010) Parasites of coral reef fish: how much do we know? With a bibliography of fish parasites in New
Caledonia. Belg J Zool 140:155-190
Justine JL, Beveridge I, Boxshall GA, Bray RA, Moravec F, Trilles JP, Whittington ID (2010) An annotated list of
parasites (Isopoda, Copepoda, Monogenea, Digenea, Cestoda and Nematoda) collected in groupers
(Serranidae, Epinephelinae) in New Caledonia emphasizes parasite biodiversity in coral reef fish. Folia
Parasitol (Praha) 57:237-262
Kamiya T, O'Dwyer K, Nakagawa S, Poulin R (2014) What determines species richness of parasitic organisms? A
meta‐analysis across animal, plant and fungal hosts. Biological Reviews 89:123-134
Kerrigan BA (1996) Temporal patterns in size and condition at settlement in two tropical reef fishes (Pomacentridae:
Pomacentrus amboinensis and P. nagasakiensis). Mar Ecol Prog Ser 135:27-41
Kim YG, Chun SK (1984) Studies on the life history of Bacciger harengulae. J Korean Fish Soc 17:449-470
King RE (1964) Three hemiurid trematodes from South Vietnam. Trans Am Microsc Soc 83:435-439
Knowlton N, Rohwer F (2003) Multispecies microbial mutualisms on coral reefs: The host as a habitat. The American
Naturalist 162:51-62
Korotaeva VD (1969) [Helminths of some food-fishes of the order Clupeiformes in the sea near Australia and New
Zealand] (In Russian). In: Markevich AP (ed) Problemy Parazitologii Trudy Nauchno Konferentsii
Parazitologov USSR (6th) 237-238
Krebs CJ (1999) Ecological methodology. Benjamin/Cummings, Menlo Park, California
Krkošek M, Revie CW, Gargan PG, Skilbrei OT, Finstad B, Todd CD (2013) Impact of parasites on salmon recruitment
in the Northeast Atlantic Ocean. Proc R Soc Biol Sci Ser B 280:doi: 10.1098/rspb.2012.2359
Kuris A, Lafferty K (2000) Parasite-host modeling meets reality: adaptive peaks and their ecological attributes.
Evolutionary biology of host–parasite relationships: theory meets reality:9-26
Kurochkin YV (1970) [New trematode species Discogasteroidinae (Trematoda, Fellodistomatidae) from marine fishes
of Australia] (In Russian). Parazitologiya 4:111-115
Lafferty KD, Kuris AM (2002) Trophic strategies, animal diversity and body size. Trends Ecol Evol 17:507-513
Lecchini D, Planes S, Galzin R (2005) Experimental assessment of sensory modalities of coral-reef fish larvae in the
recognition of their settlement habitat. Behav Ecol Sociobiol 58:18-26
Lecchini D, Planes S, Galzin R (2007a) The influence of habitat characteristics and conspecifics on attration and
survival of coral reef fish juveniles. J Exp Mar Biol Ecol 341:85-90

109
Lecchini D, Peyrusse K, Lanyon RG, Lecellier G (2014) Importance of visual cues of conspecifics and predators during
the habitat selection of coral reef fish larvae. Ethology 337:345-351
Lecchini D, Osenberg CW, Shima JS, St Mary CM, Galzin R (2007b) Ontogenetic changes in habitat selection during
settlement in a coral reef fish: ecological determinants and sensory mechanisms. Coral Reefs 26:423-432
Lemly AD, Esch GW (1984) Effects of the trematode Uvulifer ambloplitis on juvenile Bluegill Sunfish, Lepomis
macrochirus: ecological implications. J Parasitol 70:475-492
Lester RJG, Sewell KB (1990) Checklist of parasites from Heron Island, Great Barrier Reef. Aust J Zool 37:101-128
Lewis AR (1997) Recruitment and post-recruit immigration affect the local population size of coral reef fishes. Coral
Reefs 16:139-149
Lienart GDH, Mitchell MD, Ferrari MC, McCormick MI (2014) Temperature and food availability affect risk
assessment in an ectotherm. Anim Behav 89:199-204
Linton E (1910) Helminth fauna of the Dry Tortugas. II. Trematodes. Papers of the Tortugas Laboratory of the Carnegie
Institute of Washington Publication 4:11-98
Littlewood DTJ, Curini-Galletti M, Herniou EA (2000) The interrelationships of Proseriata (Platyhelminthes: Seriata)
tested with molecules and morphology. Mol Phylogenet Evol 16:449-466
Lo CM, Morand S, Galzin R (1998) Parasite diversity/host age and size relationship in three coral-reef fishes from
French Polynesia. Int J Parasitol 28:1695-1708
Losey GS (1972) The ecological importance of cleaning symbiosis. Copeia 4:820-833
Losey GS (1979) Fish cleaning symbiosis: proximate cauases of host behaviour. Anim Behav 27:669-685
Losey GS, Mahon JL, Danilowicz BS (1995) Innate recognition by host fish of their cleaning symbiont. Ethology
100:277-283
Machida M (1984) Two new trematodes from gallbladder of tropical marine fishes, Myripristis and Abudefduf. Bulletin
of the Natural Museum, Tokyo 10:1-5
Machida M (2005) Monorchiidae (Trematoda, Digenea) from fishes of Japanese and adjacent waters. Bulletin of the
National Science Museum, Tokyo Series A Zoology 31:123-136
Machida M, Uchida K (2001) Digenean trematodes from pomacenthid fishes of Japan and Palau. Bulletin of the
National Science Museum, Tokyo, Series A, Zoology 27:217-227
Maddison WP, Maddison DR (2009) Mesquite: a modular system for evolutionary analysis. Version 2.72
http://mesquiteproject.org
Madhavi R (1975) Digenetic trematodes from marine fishes of Waltair coast, Bay of Bengal: Family Fellodistomidae.
Revista di Parasitologia 36:115-128
Manter HW (1934) Some digenetic trematodes from deep-water fish of Tortugas, Florida. Papers from Tortugas
Laboratory 28:257-345
Manter HW (1942) Monorchiidae (Trematoda) from fishes of Tortugas, Florida. Trans Am Microsc Soc 61:349-360
Manter HW (1947) The digenetic trematodes of marine fishes of Tortugas, Florida. The American Midland Naturalist
38:257-416
Manter HW (1969a) Three new genera hemiurid trematodes from Australian fishes. Journal of the Fisheries Board of
Canada 26:787-792
Manter HW (1969b) Some digenetic trematodes of marine fishes of New Caledonia. Part IV. Hemiuridae and summary.
Proc Helminthol Soc Wash 36:194-204
Marcogliese DJ (1995) The role of zooplankton in the transmission of helminth parasites to fish. Rev Fish Biol Fish
5:336-371
Marcogliese DJ (2002) Food webs and the transmission of parasites to marine fish. Parasitology 124:83-99
Marino F, Giannetto S, Paradiso ML, Bottari T, De Vico G, Macri B (2004) Tissue damage and haematophagia due to
praniza larvae (Isopoda: Gnathiidae) in some aquarium seawater teleosts. Dis Aquat Organ 59:43-47
McCormick MI (2006) Mothers matter: crowding leads to stressed mothers and smaller offspring in marine fish.
Ecology 87:1104-1109
McCormick MI, Hoey AS (2004) Larval growth history determines juvenile growth and survival in a tropical marine
fish. Oikos 106:225-242
McCormick MI, Meekan MG (2007) Social facilitation of selective mortality Ecology 88:1562-1570
McCormick MI, Moore JAY, Munday PL (2010) Influence of habitat degradation on fish replenishment. Coral Reefs
29:537-546
McNamara M, Cribb T (2011) Taxonomy, host specificity and dietary implications of Hurleytrematoides (Digenea:
Monorchiidae) from chaetodontid fishes on the Great Barrier Reef. Parasitol Int 60:255-269
McNamara M, Miller T, Cribb T (2014) Evidence for extensive cryptic speciation in trematodes of butterflyfishes
(Chaetodontidae) of the tropical Indo-West Pacific. Int J Parasitol 44:37-48
McNamara M, Adlard R, Bray R, Sasal P, Cribb T (2012) Monorchiids (Platyhelminthes: Digenea) of chaetodontid
fishes (Perciformes): biogeographical patterns in the tropical Indo-West Pacific. Parasitol Int 61:288-306
Medeiros PR, Souza AT, Ilarri MI (2010) Habitat use and behavioural ecology of the juveniles of two sympatric
damselfishes (Actinopterygii: Pomacentridae) in the south-western Atlantic Ocean. J Fish Biol 77:1599-1615
Meekan M, Steven A, Fortin M (1995) Spatial patterns in the distribution of damselfishes on a fringing coral reef. Coral
Reefs 14:151-161

110
Meekan MG, Wilson SG, Halford A, Retzel A (2001) A comparison of catches of fishes and invertebrates by two light
trap designs, in tropical NW Australia. Mar Biol 139:373-381
Mensink PJ, Geange SW, Shima JS (2014) Reproductive success of parasitized males in a marine reef fish. Mar Biol
161:2689-2696
Milicich MJ, Doherty PJ (1994) Larval supply of coral reef fish populations - magnitude and synchrony of
replenishment to Lizard Island, Great Barrier Reef. Mar Ecol Prog Ser 110:121-134
Militz TA, Hutson KS (2015) Beyond symbiosis: cleaner shrimp clean up in culture. PLoS ONE 10:e0117723
Miller MA, Pfeiler E, Schwartz T (2010) Creating the CIPRES Science Gateway for inference of large phylogenetic
trees. In: Proceedings of the Gateway Computing Environments Workshop (GCE), 14 Nov. 2010, New
Orleans, LA, pp. 1-8
Miller TL, Cribb TH (2007) Coevolution of Retrovarium n. gen. (Digenea: Cryptogonimidae) in Lutjanidae and
Haemulidae (Perciformes) in the Indo-West Pacific. Int J Parasitol 37:1023-1045
Miller TL, Cribb TH (2008) Eight new species of Siphoderina Manter, 1934 (Digenea, Cryptogonimidae) infecting
Lutjanidae and Haemulidae (Perciformes) off Australia. Acta Parasitol 53:344-364
Miller TL, Bray RA, Cribb TH (2011a) Taxonomic approaches to and interpretation of host-specificity of trematodes of
fishes: lessons from the Great Barrier Reef. Parasitology 138:1710-1722
Miller TL, Bray RA, Cribb TH (2011b) Taxonomic approaches to and interpretation of host specificity of trematodes of
fishes: lessons from the Great Barrier Reef. Parasitology
Monod T (1926) Les Gnathiidae. Essai monographique (morphologie, biologie, systématique). Mémoires de la Société
des Sciences Naturelles du Maroc 13:1-668
Morand S (2015) (macro-) Evolutionary ecology of parasite diversity: From determinants of parasite species richness to
host diversification. International Journal for Parasitology: Parasites and Wildlife 4:80-87
Morand S, Cribb TH, Kulbicki M, Rigby MC, Chauvet C, Dufour V, Faliex E, Galzin R, Lo CM, Lo-Yat A, Pichelin S,
Sasal P (2000) Endoparasite species richness of New Caledonian butterfly fishes: host density and diet matter.
Parasitology 121:65-73
Mugridge RER, Stallybrass HG (1983) A mortality of eels, Anguilla anguilla L., attributed to Gnathiidae. J Fish Dis
6:81-82
Muñoz G, Cribb TH (2006) Parasite communities and diet of Coris batuensis (Pisces: Labridae) from Lizard Island,
Great Barrier Reef. Mem Queensl Mus 52:191-198
Muñoz G, Grutter A, Cribb T (2006) Endoparasite communities of five fish species (Labridae: Cheilininae) from Lizard
Island: how important is the ecology and phylogeny of the hosts? Parasitology 132:363-374
Muñoz G, Grutter AS, Cribb TH (2007) Structure of the parasite communities of a coral reef fish assemblage
(Labridae): testing ecological and phylogenetic host factors. J Parasitol 93:17-30
Nahhas FM, Cable RM (1964) Digenetic and aspidogasterid trematodes from marine fishes of Curacao and Jamaica.
Tulane Studies of Zoology 11:167-228
Nahhas FM, Wetzel JA (1995) Digenetic trematodes of marine fishes from Suva, Fiji: the family Gyliauchenidae Ozaki,
1933. Journal of Helminthological Society Washington 62:117-130
Nolan MJ, Cribb TH (2005) The use and implications of ribosomal DNA sequencing for the discrimination of digenean
species. Adv Parasitol 60:101-163
Ohman MC, Munday PL, Jones GP, Caley MJ (1998) Settlement strategies and distribution patterns of coral-reef fishes.
J Exp Mar Biol Ecol 225:219-238
Olson PD, Cribb TH, Tkach VV, Bray RA, Littlewood DTJ (2003) Phylogeny and classification of the Digenea
(Platyhelminthes: Trematoda). Int J Parasitol 33:733-755
Ormond RFG, Roberts JM, Jan R-Q (1996) Behavioural differences in microhabitat use by damselfishes
(Pomacentridae): implications for reef fish biodiversity. J Exp Mar Biol Ecol 202:85-95
Ostlund-Nilsson S, Curtis L, Nilsson G, Grutter AS (2005) Parasitic isopod Anilocra apogonae, a drag for the cardinal
fish Cheilodipterus quinquelineatus. Mar Ecol Prog Ser 287:209-216
Ota Y, Hoshino O, Hirose M, Tanaka K, Hirose E (2012) Third-stage larva shifts host fish from teleost to elasmobranch
in the temporary parasitic isopod, Gnathia trimaculata (Crustacea: Gnathiidae). Mar Biol 159:2333-2347
Overstreet RM (1969) Digenetic trematodes from Biscayne Bay, Florida. Tulane Stud Zool Bot 15:119-176
Ozaki Y (1932) A new trematode from File-fish: Lepotrema clavatum ngn sp. Proceedings of the Imperial Academy
8:44-47
Palacios-Fuentes P, Landaeta MF, Munoz G, Plaza G, Ojeda FP (2012) The effects of a parasitic copepod on the recent
larval growth of a fish inhabiting rocky coasts. Parasitol Res 111:1661-1671
Pankov P, Webster BL, Blasco-Costa I, Gibson DI, Littlewood DTJ, Balbuena JA, Kostadinova A (2006) Robinia
aurata n. g., n. sp. (Digenea: Hemiuridae) from the mugilid Liza aurata with a molecular confirmation of its
position within the Hemiuroidea. Parasitology 133:217-227
Paperna I (1977) Preliminary data on the Gnathiidae (Isopoda) of the northern Red Sea, the Bitter Lakes and the Eastern
Mediterranean and the biology of Gnathia piscivora n. sp. Rapports et Proces-Verbaux des Reunions-
Commission Internationale pour l'Exploration Scientifique de la Mer Mediterranee (CIESM)
Paris CB, Atema J, Irisson J-O, Kingsford M, Gerlach G, Guigand CM (2013) Reef odor: a wake up call for navigation
in reef fish larvae. PLoS ONE 8:e72808
Parukhin AM (1976a) [Parasitic worms of benthic fishes of the southern seas] (In Russian). Naukova Dumka, Kiev

111
Parukhin AM (1976b) [Parasitic worms of commercially important fishes of the southern Seas] (In Russian), Kiev:
Naukova Dumka
Parukhin AM (1989) [Parasitic worms of bottom fishes of the southern seas] (In Russian). Naukova Dumka, Kiev 153
Pearson JC (1968) Observations on the morphology and life-cycle of Paucivitellosus fragilis Coil, Reid & Kuntz, 1965
(Trematoda: Bivesiculidae). Parasitology 58:769-788
Penfold R, Grutter AS, Kuris AM, McCormick MI, Jones CM (2008) Interactions between juvenile marine fish and
gnathiid isopods: predation versus micropredation. Mar Ecol Prog Ser 357:111-119
Pleijel F, Jondelius U, Norlinder E, Nygren A, Oxelman B, Schander C, Sundberg P, Thollesson M (2008) Phylogenies
without roots? A plea for the use of vouchers in molecular phylogenetic studies. Mol Phylogenet Evol 48:369-
371
Posada D (2008) jModelTest: Phylogenetic model averaging. Mol Biol Evol 25:1253-1256
Poulin R, Grutter AS (1996) Cleaning symbioses: Proximate and adaptive explanations. Bioscience 46:512-517
Poulin R, Morand S (2000) The diversity of parasites. Q Rev Biol 75:277-293
Poulin R, Cribb TH (2002) Trematode life cycles: short is sweet? Trends Parasitol 18:176-183
Pritchard MH (1963) Studies on digenetic trematodes of Hawaiian fishes, primarily families Lepocreadiidae and
Zoogonidae. The Journal of Parasitology 49:578-587
Pritchard MH, Manter HW (1961) Studies of digenetic trematodes of Hawaiian fishes-family Haplosplanchnidae. Proc
Helminthol Soc Wash 28:191
R Development Core Team (2013) R: A language and environment for statistical computing , R Foundation for
Statistical Computing. http://www.R-project.org), Vienna
Radford C, Stanley J, Simpson S, Jeffs A (2011) Juvenile coral reef fish use sound to locate habitats. Coral Reefs
30:295-305
Randall JE, Allen GR, Steene RC (1997) Fishes of the Great Barrier Reef and Coral Sea. Crawford House Publishing,
Bathurst, N.S.W
Rankin TL, Sponaugle S (2014) Characteristics of settling coral Rref fish are related to recruitment timing and success.
PLoS ONE 9:e108871
Rizk MH, Taha SA, Ab El-Rahman A (1996) Helminth parasites of some fishes from Mediterranean Sea and their
effect on the coefficient of condition. J Egypt Ger Soc Zool 19:129-144
Robertson DR (1974) A study of the biology and reproductive biology of the labrid fish, Labroides dimidiatus, at Heron
Island, Great Barrier Reef. Ph.D, thesis. The University of Queensland, pp 174
Roche DG, Strong LE, Binning SA (2013a) Prevalence of the parasitic cymothoid isopod Anilocra nemipteri on its fish
host at Lizard Island, Great Barrier Reef. Aust J Zool 60:330-333
Roche DG, Binning SA, Strong LE, Davies JN, Jennions MD (2013b) Increased behavioural lateralization in parasitized
coral reef fish. Behav Ecol Sociobiol 67:1339-1344
Rohde K (2002) Ecology and biogeography of marine parasites. Adv Mar Biol 43:1-86
Rohner CA, Cribb TH (2013) Opecoelidae (Digenea) in northern Great Barrier Reef goatfishes (Perciformes: Mullidae).
Syst Parasitol 84:237-253
Ronquist F, Teslenko M, van der Mark P, Ayres DL, Darling A, Hohna S, Larget B, Liu L, Suchard MA, Huelsenbeck
JP (2012) MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model
space. Syst Biol 61:539-542
Ros AF, Lusa J, Meyer M, Soares MC, Oliveira RF, Brossard M, Bshary R (2011) Does access to the bluestreak cleaner
wrasse Labroides dimidatus affect indicators of stress and health in resident reef fishes in the Red Sea? Horm
Behav 59:151-158
Russell BC (1983) Annotated checklist of the coral reef fishes in the Capricorn-Bunker group Great Barrier Reef,
Australia. Great Barrier Reef Marine Park Authority, Mackay
Sambrook J, Russell DW (2001) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold
Spring, New York
Sasal P, Morand S, Guégan JF (1997) Determinants of parasite species richness in Meditteranean marine fishes. Mar
Ecol Prog Ser 149:61-71
Scholz T (2002) Family Gymnophallidae Odhner, 1905. In: Gibson DI, Jones A, Bray RA (eds) Keys to the Trematoda,
Volume 1. CABI Publishing and the Natural History Museum, Wallingford,
Scott ME, Dobson A (1989) The role of parasites in regulating host abundance. Parasitol Today 5:176-183
Sikkel PC, Cheney KL, Côté IM (2004) In situ evidence for ectoparasites as a proximate cause of cleaning interactions
in reef fish. Anim Behav 68:241-247
Sikkel PC, Schaumburg CS, Mathenia JK (2006) Diel infestation dynamics of gnathiid isopod larvae parasitic on
Caribbean reef fish. Coral Reefs 25:683-689
Sikkel PC, Ziemba RE, Sears WT, Wheeler JC (2009) Diel ontogenetic shift in parasitic activity in a gnathiid isopod on
Caribbean coral reefs. Coral Reefs 28:489-495
Sikkel PC, Sears WT, Weldon B, Tuttle BC (2011) An experimental field test of host-finding mechanisms in a
Caribbean gnathiid isopod. Mar Biol 158:1075-1083
Simpson SD, Jeffs A, Montgomery JC, McCauley R, Meekan MG (2008) Nocturnal relocation of adult and juvenile
coral reef fishes in response to reef noise. Coral Reefs 27:97-104
Smit NJ, Davies AJ (2004) The curious life-style of the parasitic stages of gnathiid isopods. Adv Parasitol 58:289-391

112
Smit NJ, Basson L, Van As JG (2003) Life cycle of the temporary fish parasite, Gnathia africana (Crustacea: Isopoda:
Gnathiidae). Folia Parasitol (Praha) 50:135-142
Smith JW (2002) Family Sanguinicolidae Graff, 1907. In: Gibson DI, Jones A, Bray RA (eds) Keys to the digenean
parasites of vertebrates. CAB International, Cambridge,
Soares MC, Bshary R, Côté IM (2008) Does cleanerfish service quality depend on client value or choice options? Anim
Behav 76:123-130
Soares MC, Oliveira R, Ros AFH, Grutter AS, Bshary R (2011) Tactile stimulation lowers stress in fish. Nat Commun
2:534
Srinivasan M (2003) Depth distibutions of coral reef fishes: the influence of microhabitat structure, settlement, and
post-settlement processes. Oecologia 137:76-84
Stachowicz JJ (2001) Mutualism, facilitation, and the structure of ecological communities. BioScience 51:235-246
Stamatakis A, Hoover P, Rougemont J (2008) A rapid bootstrap algorithm for the RAxML web servers. Syst Biol
57:758-771
Stummer LE, Weller JA, Johnson ML, Côté IM (2004) Size and stripes: how fish clients recognize cleaners. Anim
Behav 68:145-150
Sun D, Blomberg SP, Cribb TH, McCormick MI, Grutter AS (2012) The effects of parasites on the early life stages of a
damselfish. Coral Reefs 31:1065-1075
Sun D, Bray RA, Yong R, Q-Y, Cutmore SC, Cribb TH (2014) Pseudobacciger cheneyae n. sp. (Digenea:
Gymnophalloidea) from Weber’s chromis (Chromis weberi Fowler & Bean) (Perciformes: Pomacentridae) at
Lizard Island, Great Barrier Reef, Australia. Syst Parasitol 88:141-152
Sweatman H (1985) The influence of adults of some coral reef fishes on larval recruitment. Ecol Monogr 55:469-485
Sweatman H (1988) Field evidence that settling coral reef fish larvae detect resident fishes using dissolved chemical
cues. J Exp Mar Biol Ecol 124:163-174
Tolimieri N (1995) Effects of microhabitat characteristics on the settlement and recruitment of a coral reef fish at two
spatial scales. Oecologia 102:52-63
Toman G, Kamegai S (1974) Some digenetic trematodes of marine fishes from Saipan, Micronesia Proceedings of the
Third International Congress of Parasitology, Munich 1617-1618
Torres J, Miquel J, Casanova J-C, Ribas A, Feliu C, Morand S (2006) Endoparasite species richness of Iberian
carnivores: influences of host density and range distribution. Biodivers Conserv 15:4619-4632
Vail AL, McCormick MI (2011) Metamorphosing reef fishes avoid predator scent when choosing a home. Biol Lett
7:921-924
Waldie PA, Blomberg SP, Cheney KL, Goldizen AW, Grutter AS (2011) Long-term effects of the cleaner fish
Labroides dimidiatus on coral reef fish communities. PLoS ONE 6:e21201-21201-e21201-21207
Wang P, Wang Y (1993) Some digenetic trematodes from marine fishes from Fujian Province. Journal of Fujian
Teachers University Natural Science Edition 9:66-73
Weeks P (2000) Red-billed oxpeckers: vampires or tickbirds? Behav Ecol 11:154-160
Wenger AS, Johansen J, Jones G (2011) Suspended sediment impairs habitat choice and chemosensory discrimination
in two coral reef fishes. Coral Reefs 30:879-887
Whittington ID (1998) Diversity "down under": monogeneans in the Antipodes (Australia) with a prediction of
monogenean biodiversity worldwide. Int J Parasitol 28:1481-1493
Wilson SK, Burgess SC, Cheal AJ, Emslie M, Fisher R, Miller I, Polunin NVC, Sweatman HPA (2008) Habitat
utilization by coral reef fish: implications for specialists vs. generalists in a changing environment. J Anim
Ecol 77:220-228
Yamaguti S (1934) Studies on the helminth fauna of Japan. Part 2. Trematodes of fishes. Japanese Journal of Zoology
5:249-541
Yamaguti S (1938) Studies on the Helminth Fauna of Japan. Part 21. Trematodes of Fishes, IV. Yamaguti, S., Kyöto
Yamaguti S (1942) Studies on the helminth fauna of Japan. Part 39. Trematodes of fishes mainly from Naha.
Transactions of the Biogeographical Society of Japan 3:329-398
Yamaguti S (1953) Parasitic worms mainly from Celebes. Part 3. Digenetic trematodes of fishes II. Acta Med
Okayama 8:257-295
Yeo SE, Spieler RE (1980) Habitat effects on the occurence of parasites inhabiting the Sergeant Major, Abudefduf
saxatilis (Linnealus), with a list of parasites of Caribbean damselfish. Bull Mar Sci 30:313-324

113
Appendices
Appendix I
All damselfish recruits observed on experimental patch reefs (Chapter 3).
Fish Genus Species
Acanthochromis polyacanthus
Amblyglyphidodon curacao
Amblyglyphidodon leucogaster
Chromis ternatensis
Chromis viridis
Chrysiptera cyanea †
Chrysiptera flavipinnis
Chrysiptera rex *
Chrysiptera rollandi
Chrysiptera talboti
Dascyllus aruanus
Dascyllus reticulatus
Dischistodus perspicillatus
Dischistodus prospotaenia
Dischistodus spp.
Neoglyphidodon melas
Neoglyphidodon nigroris
Neopomacentrus bankieri
Neopomacentrus cyanomos
Pomacentrus adelus
Pomacentrus amboinensis
Pomacentrus bankanensis †
Pomacentrus branchialis
Pomacentrus chrysurus
Pomacentrus coelestis
Pomacentrus
Pomacentrus
grammorhynchus
lepidogenys
Pomacentrus moluccensis
Pomacentrus nagasakiensis
Pomacentrus pavo
Pomacentrus simsiang
* Species only recorded at reefs with L. dimidiatus present.
† Species only recorded at reefs without L. dimidiatus present.

114
Appendix II
Species of fish cleaned by juvenile Labroides dimidiatus (Chapter 4).
Species Family
Ctenochaetus binotatus
Acanthuridae
Ctenochaetus striatus Acanthuridae
Zebrasoma scopas Acanthuridae
Zebrasoma veliferum Acanthuridae
Apogon doederleini Apogonidae
Apogon compressus Apogonidae
Apogon cookii Apogonidae
Apogon properupta Apogonidae
Apogon trimaculatus Apogonidaae
Cheilodipterus intermedius Apogonidae
Cirripectes chelomatus Blennidae
Cirripectes stigmaticus Blennidae
Ecsenius aequalis Blennidae
Ecsenius bicolor Blennidae
Ecsenius mandibularis Blennidae
Ecsenius stictus Blennidae
Exallias brevis Blennidae
Meiacanthus atrodorsalis Blennidae
Plagiotremus rhinorhynchos Blennidae
Plagiotremus tapeinosoma Blennidae
Caesio caerulaurea Caesionidae
Caesio teres Caesionidae
Chaetodon aureofasciatus Chaetodontidae
Chaetodon baronessa Chaetodontidae
Chaetodon citrinellus Chaetodontidae
Chaetodon klienii Chaetodontidae
Chaetodon lunulatus Chaetodontidae
Chaetodon plebius Chaetodontidae
Chaetodon rainfordi Chaetodontidae
Chaetodon ulietensis Chaetodontidae
Amblygobius rainfordi Gobidae
Valenciennea strigata Gobidae
Neoniphon opercularis Holocentridae
Neoniphon sammara Holocentridae
Sarocentron cornutum Holocentridae
Sargocentron diadema Holocentridae
Sargocentron spiniferum Holocentridae
Bodianus axillaris Labridae
Cheilinus chlorourus Labridae
Chlorurus sordidus Labridae
Choerodon fasciatus Labridae
Coris batuensis Labridae

115
Epibulis insidiator Labridae
Gomphosus varius Labridae
Halichoeres hortulanus Labridae
Halichoeres malanurus Labridae
Halichoeres melanurus Labridae
Hemigymnus melapterus Labridae
Labrichtys unilineatus Labridae
Labroides dimidiatus Labridae
Pseudocheilinus hexataenia Labridae
Thalassoma lunare Labridae
Scarus frenatus Labridae
Scarus ghobban Labridae
Scarus niger Labridae
Stethojulis bandanensis Labridae
Lutjanus carponotatus Lutjanidae
Pervagor janthinosoma Monacanthidae
Parupeneus multifasciatus Mullidae
Scolopsis bilineatus Nemipteridae
Centropyge bicolor Pomacanthidae
Pomacanthus sexstriatus Pomacanthidae
Abudefduf sexfaciatus Pomacentridae
Abudefduf whitleyi Pomacentridae
Acanthochromis polyacanthus Pomacentridae
Amblyglyphididon curacao Pomacentridae
Amblyglyphididon leucogaster Pomacentridae
Chromis actripectoralis Pomacentridae
Chromis lepidolepis * Pomacentridae
Chromis viridis Pomacentridae
Chrysiptera rex Pomacentridae
Dischistodus melanotus Pomacentridae
Dischistodus prosopotaenia Pomacentridae
Dischistodus pseudochrysopoecilus Pomacentridae
Hemiglyphidodon plagiometopon Pomacentridae
Neoglyphidodon melas Pomacentridae
Neopomacentrus azysron Pomacentridae
Plectroglyphidodon lacrymatus Pomacentridae
Pomacentrus adelus * Pomacentridae
Pomacentrus amboinensis * Pomacentridae
Pomacentrus bankanensis Pomacentridae
Pomacentrus branchialis * Pomacentridae
Pomacentrus coelestis Pomacentridae
Pomacentrus chrysurus Pomacentridae
Pomacentrus grammorhynchus Pomacentridae
Pomacentrus imitator Pomacentridae
Pomacentrus moluccensis Pomacentridae
Pomacentrus nagasakiensis * Pomacentridae
Pomacentrus simsiang Pomacentridae
Pomacentrus wardi Pomacentridae
Premnas biaculeatus Pomacentridae
Stegastes apicalis Pomacentridae

116
Stegastes fasciolatus Pomacentridae
Stegastes nigricans Pomacentridae
Ogilbyina queenslandia Pseudochromidae
Pseudochromis fuscus Pseudochromidae
Cephalopholis boenak Epinephelidae
Cephalopholis cyanostigma Epinephelidae
Cephalopholis microprion Epinephelidae
Siganus corallinus Siganidae
Synodus variegatus Synodontidae
Canthigaster papua Tetraodontidae
Canthigaster valentini Tetraodontidae
* Recently-settled individuals (< 2 cm) of these species were cleaned by L. dimidiatus.





























