Embed Size (px)
Citation preview

Aust. J . Agric. Res., 1977, 28, 449-60
Parthenium Weed (Parthenium hysterophorus L.): Gas Exchange Characteristics as a Basis for Prediction of its Geographical Distribution
D. Doley
Department of Botany, University of Queensland, St. Lucia, Qld. 4067.
Abstract
Gas exchange studies in Parthenium hystevophovus L., a weed recently introduced into central Queensland, indicate that its limits of distribution may be very wide in the humid and subhumid regions of Australia. Under conditions of high leaf water potential, the maximum rate of apparent photosynthesis of cabinet-grown plants was 77 ng cm-2 sec-', with a temperature optimum of 28°C. Gas phase diffusive resistances were very low and insensitive to photosynthetic photon flux density at high water potentials (-5.0 bars), but became greater and quite sensitive to photon flux as the leaf water potential approached -20 bars. At temperatures between 10 and 40°C, transpiration increased slightly, and the dark respiration rate was almost constant, owing to a steady and considerable increase in gas phase diffusive resistance with temperature. The control of gas exchange broke down at about 4 2 T , so that transpiration in the light and dark proceeded at equal rates, and dark respiration rates were very high. Gas exchange in P . hysterophovus appears to be no more sensitive to reduced water potential than it is in several favoured crop and pasture species, but the distribution of this weed may be limited by even brief exposure to very high tempera- tures, or by prolonged drought.
Introduction
Parthenium hysterophorus L. (Asteraceae) is a native of the Americas (Rollins 1950) and had developed as a weed of sugar-cane fields and pastures in India and the islands of the Pacific (Harvey 1976). It has been introduced recently into Australia, and is becoming an important weed of subhumid central Queensland, apparently following a succession of unusually wet years (Everist 1976).
Very little is known of the physiology of P. hysterophorus, and a rapid means of assessing broad environmental limits for its possible distribution may be afforded by the study of gas exchange characteristics. The work reported here was undertaken to test the utility of such gas exchange studies in predicting the likely behaviour of this weed in the field.
Materials and Methods
Seedlings of P. hysterophorus were grown in a potting mixture and maintained in a growth cabinet at daylnight temperatures of 27122°C and relative humidities of 82189%. The photoperiod was 14 hr, with a photosynthetic photon flux density (photon flux) of 13 nE cm-2 sec-'. These conditions induced flowering after a period of about 4 weeks.
Gas exchange characteristics were measured on intact single, young, fully expanded leaves with areas ranging from 14 to 19 cm2. An open gas analysis system was used,

450 D. Doley
similar to that described by Doley and Yates (1976). Illumination of the leaf chamber was provided by a 1500 W tungsten halogen lamp, separated from the leaf chamber by a water bath 7 cm deep and an air gap of 5 cm. Air with known concentrations of carbon dioxide and water vapour was introuduced into the chamber at a flow rate of 3 .5 1. min-' and stirred at a constant rate, so that the boundary layer resistance t o water vapour diffusion (rh) was 0.3 sec cm-l. Photosynthesis was measured by a Grubb Parsons MGA4 infrared gas analyser, and transpiration by differential thermocouple psychrometers.
Leaf temperature was regulated by means of a water jacket in the base of the leaf chamber. The water vapour concentration difference between leaf and air (AX) was regulated by adding appropriate volumes of dried or humidified air to the stream entering the leaf chamber, whilst maintaining a constant leaf temperature. Care was taken to avoid condensation of water in the leaf chamber or in other parts of the apparatus, and this limited the minimum AX. The maximum AX was influenced by both leaf temperature and the capacity of the ice bath air dryer used in the gas supply system.
Mean values of the measured and calculated gas exchange parameters were determined and printed out after 10 scans of each instrument by a computer-controlled scanner. This enabled the monitoring and regulation of all necessary environmental and leaf parameters. A steady rate of gas exchange was assumed to have been reached when five successive printed mean values of apparent photosynthesis varied by no more than k 3 %.
The assumptions made in the calculation of the carbon dioxide diffusive resis- tances were those adopted by Ludlow and Jarvis (1971), namely,
and
where rh and r, are the boundary layer resistances to the diffusion of water vapour and carbon dioxide, respectively, and ri and r, are the stomata1 or gas phase resis- tances to the diffusion of water vapour and carbon dioxide respectively. An estimate of the intercellular carbon dioxide concentration within the leaf (Ci) was obtained by
where C, is the carbon dioxide concentration in the bulk air, and F is the rate of apparent photosynthesis.
All studies except the determination of carbon dioxide compensation concen- tration were carried out on air containing 300 + 2 yl 1- (540 + 4 ng ~ m - ~ ) carbon dioxide. The portion of the plant not enclosed in the leaf chamber was maintained at the laboratory temperature of 24+ O.5"C.
The light responses of carbon dioxide exchange were determined at a leaf tempera- ture of 28 . O f 0 ~ 3 ° C and AX of 11.2 1 0 . 2 yg ~ m - ~ . Reductions in photon flux at the leaf were obtained by interposing sheets of Melinex film and high quality white paper between the light source and leaf chamber. Tests with an Isco spectral radio- meter showed that the spectral quality of the radiation was not appreciably altered by this procedure.
The gas exchange of darkened leaves was determined in two ways. Firstly, measure- ments were made at the end of a light response experiment, following a progressive
Gas Exchange Characteristics of Parthenium Weed
decrease in photon flux from about 160 to 0 pE ~ m - ~ set-l. Secondly, plants were held overnight in darkness, and a leaf was inserted into a darkened gas exchange chamber and allowed to equilibrate for 15 min before the commencement of an experiment. The same methods were used to determine the rates of transpiration and apparent respiration or photosynthesis in darkened as in illuminated leaves.
Temperature response curves were established at a photon flux of 161 +2 nE cmW2 sec-' over periods of 2 days each, firstly with temperatures decreasing from about the optimum, and, after the plant had been returned to the growth cabinet overnight, with temperatures increasing from the optimum on the following day. This procedure was found to be satisfactory by Ludlow and Wilson (1971). Water stress was allowed to develop in plants over a period of 10 days by withholding water from the pots.
Five plants, all raised under similar conditions, were used for the determination of the responses of gas exchange to light and temperature. Three plants were studied in the water stress experiment. Standard deviations of mean values of gas exchange parameters were calculated, but in all cases were too small to represent graphically in the results.
Photon flux ( n ~ cmm2 sec-l)
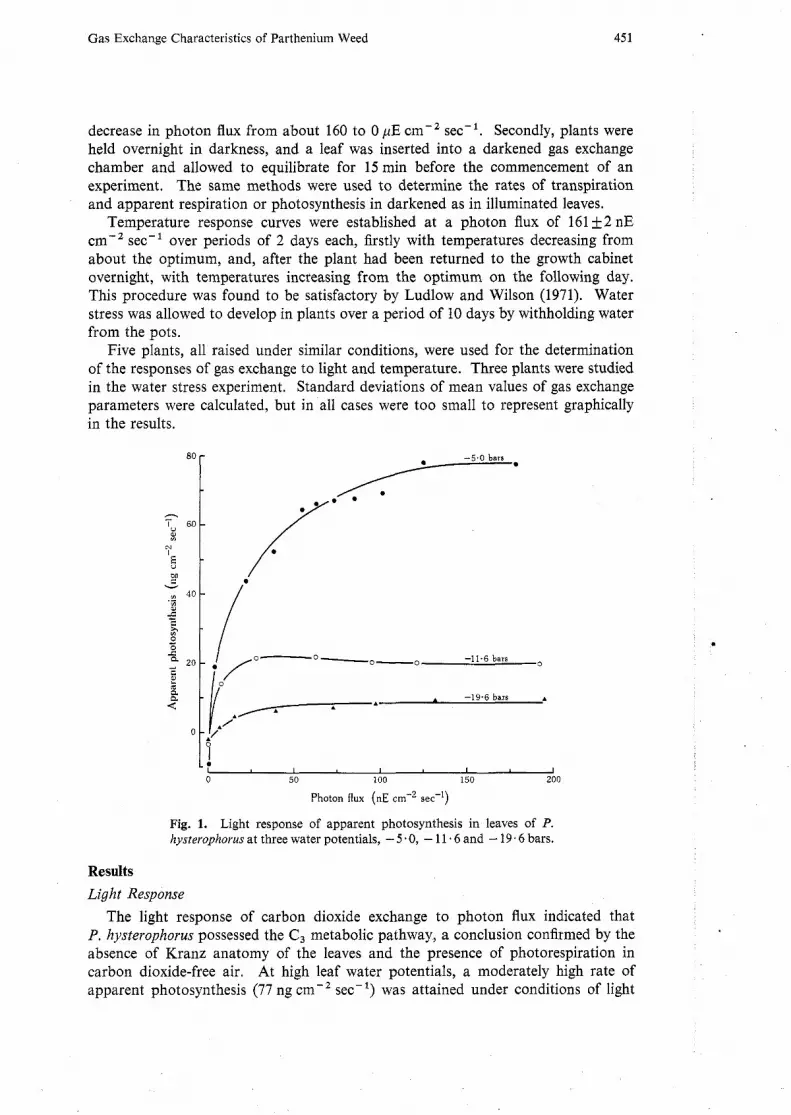
Fig. 1. Light response of apparent photosynthesis in leaves of P. hysterophorus at three water potentials, - 5.0, - 11.6 and - 19.6 bars.
Results Light Response
The light response of carbon dioxide exchange to photon flux indicated that P. hysterophorus possessed the C , metabolic pathway, a conclusion confirmed by the absence of Kranz anatomy of the leaves and the presence of photorespiration in carbon dioxide-free air. At high leaf water potentials, a moderately high rate of apparent photosynthesis (77 ng cm-2 sec-l) was attained under conditions of light

saturation, this occurring at a photon flux of 120 nE cm-2 sec-l, or 50% of noon sunlight (Fig. 1). The photon flux at the light compensation point (less than 4 nE cm-2 sec-l) was low, being about 2 % of full sunlight. It will be noted that these plants were raised in cabinets in which the photon flux was only about 5% of full sunlight.
Water Stress
Fig. 1 also shows that, as the leaf water potential was reduced from -5.0 to - 19.6 bars, apparent photosynthesis declined from 77 to 10 ng cmV2 sec-l. These results were obtained in plants subjected to relatively rapid desiccation over a period of 6-10 days. At the lowest potential studied (- 19.6 bars) the leaves were wilted, but were able to maintain their net uptake of carbon dioxide at photon flux values above about 5 nE cm-2 sec-l. A decrease in dark respiration (from 9.2 to 2.1 ng ~ r n - ~ secL1) occurred as leaf water potential was reduced, most of this decrease taking place between potentials of - 5.0 and - 11.6 bars (Fig. 1). Two days after water potentials were restored from about - 20 to about - 4 bars, apparent photo- synthesis varied from 87 to 104% of the rate recorded before the imposition of water stress.
-5.0 bars Op.- .-.- rn -.-. -.-..-. Pho!on flux ( n ~ cm-2 sec-') Photon flux ( n ~ sec-I)
Fig. 2. Light responses of (a) transpiration and (b) water vapour diffusive resistance of the leaf in P. hysterophorus at three water potentials, - 5.0, - 11.6 and - 19.6 bars.
The responses of transpiration and r; to water stress and photon flux at constant leaf temperature are shown in Fig. 2. Transpiration in the dark of leaves at - 5.0 bars was 4.6 pg cm-2 sec-l, or 40% of the maximum rate in the light, whereas dark transpiration from leaves at lower water potential was negligible. Despite a constant AX, transpiration increased linearly with photon flux between 30 and 190 nE cm-2 sec-I (Fig. 2a). At all water potentials, the leaf was up to 0.5OC cooler than the chamber air at high photon flux, but up to 0.5"C warmer at low photon flux.

Gas Exchange Characteristics of Parthenium Weed 453
As leaf water potential declined from - 5.0 to - 11 -6 bars, the relative changes in transpiration and apparent photosynthesis were similar at a photon flux of 150 nE cm-2 sec-I, but the assimilation ratio (mg carbon dioxide absorbed per gram water transpired) fell from 6.7 at - 11 6 bars to 5 - 7 at - 19.6 bars. These changes were associated with increases in the intracellular resistance to carbon dioxide diffusion (ri) from 4-73seccm-I at -5.0 bars to 1 3 . 5 5 ~ e c c m - ~ at -11.6 bars and 31.8 sec cm-I at - 19.6 bars.
At a leaf water potential of - 5.0 bars, there was little change in r; throughout the entire range of photon flux (Fig. 2b). Lower water potentials induced higher diffusive resistances at the maximum photon flux, and a progressively more sensitive response to light as this flux approached zero.
Leaf temperature (OC)
Fig. 3. Temperature response of apparent photosynthesis (F) and transpiration ( E ) in leaves of P. hysteuophouus.
Temperature Response
Apparent photosynthesis responded to temperature of the leaf in the manner which is characteristic of C, species (Bjorkmann 1975), with a temperature optimum of about 28°C (Fig. 3). The upper temperature compensation point was 50°, and the lower compensation point was estimated to be 7".
Simultaneously with these changes in apparent photosynthesis, transpiration increased in direct proportion to leaf temperature in the range 10-30°C, but increased at a greater rate between 30 and 51" (Fig. 3). The maximum assimilation ratio (7.09 mg carbon dioxide absorbed per gram water transpired) was achieved at a leaf temperature of 14.6". Gas diffusive resistances showed some unusual responses to temperature. Water vapour resistance (ri) increased linearly with leaf temperature between 10 and 42"C, then fell abruptly at higher temperatures (Fig. 4). The change in r i with temperature reflected the variation in apparent photosynthesis between 10 and 40°, r i reaching a minimum of 6.2 sec cm-I at a leaf temperature of 26°C (Fig. 4). At temperatures between 39 and 42" there were marked increases

D. Doley
in ri which were unrelated to any change in water vapour diffusive resistance, although r; did show a significant change when leaf temperature was increased beyond 42".
Because of the need to avoid water vapour condensation in the leaf chamber or in other parts of the apparatus, AX varied from 1.6 pg cm-3 at a leaf temperature of 9.7"C to 84.6 pg cm-3 at 50.9"C. Such a range of AX would obviously affect
1.5
r i 1.0-
0 5 -
0
stomata1 aperture (Lange et al. 1971; Aston 1976), so the relationship between gas exchange and AX was examined (Fig. 5). At values of AX up to 55 pg ~ r n - ~ , attained
10 20 30 40 50
Leaf temperature ("c)
- - 4 -
- /sLO
'O,/O 7' /O'w'o~n-D-o<n
P L o/" -
I
2.0
='.
g p 1.5 - S ,m 'G 2 0 1 0 - .-
G D
0 5
0 -
20 Fig. 4. Temperature response of leaf water vapour diffusive resistance (r;) ( 0 )
,, ,, and intracellular carbon dioxide diffusive resistance (ri) (0) in leaves of P. hysterophorus. ri, ri expressed in sec cm-l.
10
5
0 I
10 20 30 40 50
Leaf-air water vapour concn, difference
- i - i i - . i
-
? * ,/ lA< / A / ./*
i A/A*/*'
- i L' / - p* .*:I d"
A'
40 Fig. 5. Response of transpiration (E) ( 0 ) and leaf
h water vapour diffusive resistance
$ (v;) to difference in water vapour concentration between leaf and
30 ?' air (Ax) at constant and varying 5 2 temperature in leaves of P. - hystevophouus, A r; at varying 8 temperature and - 5 . 5 bars, .- "
-20 ,g Vu;at23.8"Cand-2.0bars , v r; at 23.8"C and - 10.7 bars. B
b
10
0

Gas Exchange Characteristics of Parthenium Weed
at a leaf temperature of 41.6", there was a predictable association between r; and AX, but as the observation continued to higher temperatures there was a change in the leaf such that no difference in the water loss characteristics could be detected between a darkened and an illuminated leaf.
Fig. 5 also shows that when water vapour exchange was examined in relation to AX at a constant temperature of 23 a 8 + O.l°C and at leaf water potentials of - 2.0 or - 10.7 bars, the responses differed quantitatively from those associated with simultaneous changes in both leaf temperature and AX. At constant leaf temperature there was a much greater change in r; with changing AX than occurred when leaf temperature as well as AX changed. This response appeared to be quite independent of leaf water potential.
Fig. 6. Temperature response of respiration rate (RD) and transpiration (E) of a darkened leaf of P. hysterophorus.
, , , , , , , , E 6 1
A A d A n
0
10 20 30 40
1 50
Leaf temperature (OC)
At constant temperature, there was a linear decrease in intercellular carbon dioxide concentration in the leaf from 510 to 396 ng cm-3 as AX increased from 5 to 14 pg ~ r n - ~ . In this experiment, there was no significant change in r i although, as shown in Fig. 4, ri did decrease when increasing AX values were associated with increasing leaf temperature.
Dark respiration responded to leaf temperature in a rather unusual manner (Fig. 6). Between 10 and 35°C there was little change in respiration as temperature increased, and there was even a slight decline between 35 and 42". As leaf temperature rose beyond 42", respiration increased abruptly from 6 .0 to 15.6 ng cm-2 set-I at 49". Dark transpiration followed a very similar course (Fig. 6), except that there was no apparent decrease between 35 and 42". These gas exchange characteristics were reflected by steadily increasing r; from 2.0 to 19.2 sec cm-I between 10 and 42", followed by a decline to 2.6 sec cm-I at a temperature between 46 and 49". These changes in diffusive resistance are similar to though almost twice the magnitude of those observed in the light (cf. Fig. 4). In both light and dark, the abrupt change in diffusive resistance, which may indicate a breakdown in leaf structure, occurred at a leaf temperature of about 42".

D. Doley
Discussion
The gas exchange characteristics of P. hysterophorus reported here conform with the information available on its distribution in other countries, and may be useful in determining its likely range in Australia. It is clear that this species is vigorous when grown under low irradiance, even though the conditions used did not lead to the development of plants of the stature attained in the field (Everist 1976).
Light saturation of apparent photosynthesis is reached at a photon flux equal to about 50 % of noon sunlight. It has been shown that plants grown under high radiant flux have higher rates of photosynthesis than do plants of the same species grown under lower radiant flux (Bjorkmann and Holmgren 1962; Hiesey et al. 1971; Patterson 1975; Gauhl 1976). Therefore, it may be anticipated that the maximum rates of apparent photosynthesis in P. hysterophorus could be higher than the value of 77 ng cm-2 sec-I recorded here, placing it in the same category as some of the more productive C , crop plants (Gifford 1974). The rates of photosynthesis observed at low photon flux in the present study also indicate that P. hysterophorus may compete successfully with many native and exotic pasture species, even when subjected to shading by an overstorey. The fact that P. hysterophorus possesses a rosette form of growth in the early stages would reduce its photosynthetic capacity in an established pasture, but when grown under controlled conditions, up to 12 leaves were produced on an erect stem, and at least some of these would be exposed to full sunlight in the field. The main threat posed by this species is the invasion of disturbed or bare ground (Everist 1976), and the combination of high photosynthetic activity and rosette early growth form would enhance its ability to occupy a cleared site.
The influence of water stress on the shape on the light response curve of apparent photosynthesis is similar to that reported in various species (Doley and Trivett 1974; Pieters and Zima 1975; Ludlow and Ng 1976). Dark respiration is suppressed as leaf water potential is lowered, a response which has been observed in pasture species (Ludlow and Ng 1976), herbaceous crops (Brix 1962; Boyer 1970a) and trees (Brix 1962; Regehr et al. 1975). The initial slopes of the light response curves are similar for plants at - 5.0 and - 11.6 bars, but light saturation is reached at a lower photon flux and at a considerably lower rate of photosynthesis as leaf water potential is reduced. At a water potential of - 11 - 6 bars, there appears to be an optimum photon flux of about 30 nE cm-2 sec-I, above which there is a slight but not statistically significant depression of photosynthesis. When leaf water potential was reduced to - 19.6 bars, there was no evidence of this optimum photon flux, and light saturation was attained only at a flux of 140 nE cm-2 sec-l, a value similar to that recorded for well-watered plants. It is clear that the change in stomatal characteristics between -5.0 and -19.6 bars would influence the appearance or otherwise of an optimum photon flux for photosynthesis.
Given adequate water supply and varying photon flux, it would appear that P. hysterophorus exerts little stomatal control over gas exchange (Fig. 2), which contrasts markedly with the C, grasses Panicum maximum (Ludlow and Ng 1976) and Astrebla lappacea (Doley and Trivett 1974). There is a surprisingly small degree of stomatal closure in the dark when leaf water potential is -5 .0 bars, and this results in a dark transpiration rate which is almost 40% of that at maximum photon flux. After maintenance overnight at a temperature of 2g°C, transpiration was about 23% of the maximum rate in the light, but even this suggests a rather

Gas Exchange Characteristics of Parthenium Weed 457
extravagant consumption of water when it is readily available. Such limited stomatal control over gas exchange is characteristic of some mesophytes, e.g. Nicotiana tabacum (Turner 1974) and Liriodendron tulipifera (Turner 1969), most of which have relatively high rates of dry matter production. However, only in P. hysterophorus does there appear to be such a low diffusive resistance in complete darkness.
There was a greater increase in stomatal diffusive resistance as leaf water potential was lowered from - 5.0 to - 11.6 bars at high irradiance than when plants were transferred from high irradiance to darkness at - 5.0 bars. In addition, there was an increase in the sensitivity of r; to light at - 11 6 bars, which increased further as leaf water potential was lowered to - 19.6 bars. At low leaf water potentials, the response of ri to light resembled more closely that of a variety of species characteris- tic of tropical humid and subhumid regions (Ludlow and Wilson 1971; Van den Driessche et al. 1971), and contrasts markedly with the behaviour of P. hysterophorus at higher leaf water potential. However, it is interesting to note that, at - 19.6 bars, r; in P. hysterophorus was responding to photon flux, whereas Panicum maximum at - 10.5 bars (Ludlow and Ng 1976) showed no response at all. This continued gas exchange at low leaf water potential could confer some competitive advantage on P. hysterophorus, but also could be inimical to its survival under conditions of prolonged drought.
When compared with arid and semiarid zone species such as Eurotia lanata and Atriplex confertifolia (Moore et al. 1972), Acacia harpophylla (Van den Driessche et al. 1971) and Eucalyptus socialis (Collatz et al. 1976), P. hysterophorus shows a more sensitive response of apparent'photosynthesis to water potential at maximum photon flux. For example, the decrease in photosynthesis between - 5 and -20 bars leaf water potential is 23 % in Acacia harpophylla, 55 % in Eucalyptus socialis and 86% in P. hysterophorus. It is also relevant that P. hysterophorus wilts at about -20 bars, whereas Acacia harpophylla phyllodes do not show obvious signs of wilting before death occurs at leaf water potentials below -70 bars.
Some pasture species of the subhumid tropics may show complete suppression of apparent photosynthesis at - 12 to -20 bars when raised in controlled environ- ments, even though photosynthesis in the field may not be so sensitive to water stress (Ludlow and Ng 1976). However, other studies (Boyer 1970a, 1970b) indicate that maize, soybean and sunflower plants grown in controlled environments may sustain apparent photosynthesis rates equal to 20-40 % of the maximum at leaf water poten- tials between - 16 and -20 bars. P. hysterophorus would, therefore, appear to be no more drought-sensitive than several favoured crop and pasture species cultivated in the humid and subhumid areas of Queensland.
Under laboratory conditions, P. hysterophorus leaves recover from severe wilting and regain their former rates of apparent photosynthesis within 2 days of rewatering. This suggests that the species possesses a considerable degree of drought tolerance (Levitt 1972), even though stress becomes evident at a relatively high water potential.
Like Mimulus spp. (Hiesey et al. 1971), and in contrast to Astrebla lappacea (Doley and Yates 1976), P. hysterophorus shows a relatively insensitive response of transpiration to temperature, except at temperatures which are sufficiently high to result in destruction of the leaf surface or of stomatal control. The usual temperature responses of apparent photosynthesis and transpiration are associated with a minimal r', which is reached at a temperature close to the optimum for apparent photosynthesis (Van den Driessche et al. 1971). At high temperatures there may be some restriction

458 D. Doley
of water loss associated with rapidly increasing r ; , particularly in species from relatively dry sites (Wuenscher and Kozlowski 1971; Doley and Yates 1976). In P. hysterophorus, transpiration and r; were both minimal at the lowest temperature achieved (9.7"C), and increased more or less linearly with leaf temperature up to 42°C (Figs 3, 4). Wuenscher and Kozlowski (1971) found similar responses of transpiration and ri to temperature in the leaves of deciduous tree species from relatively mesic sites in Wisconsin. However, these temperature responses must be considered in relation to the associated changes in AX, the importance of which has been emphasized recently by Aston (1976).
In P. hysterophorus there were slight decreases in transpiration and apparent photosynthesis as AX was increased at constant leaf temperature, these changes being mediated by the large increase in ri shown in Fig. 5. Similar observations have been reported for Prunus armeniaca (Schulze et al. 1974), and Sesamum indicum (Hall and Kaufmann 1975), the latter workers also demonstrating the importance of carbon dioxide concentration within the leaf for regulating stomatal resistance. Aston (1976) showed that, when leaf water content was maintained constant, trans- piration in Helianthus annuus increased with increasing AX, which suggests that in the earlier studies there had been a deterioration of leaf water balance as AX increased. Whilst no measurements of leaf water content were made during the experiment shown in Fig. 5, it is likely that there was some deterioration of the leaf water balance. As AX increased from 5 .7 to 13.6 ng ~ m - ~ , carbon dioxide concentration within the leaf fell from 510 to 396 ng ~ m - ~ . This change, at constant leaf water status, would tend to reduce stomatal diffusive resistance and increase transpiration and apparent photosynthesis, rather than lead to the observed responses.
From the foregoing it is clear that the temperature responses of transpiration and apparent photosynthesis shown in Fig. 3 are due not solely to changes in leaf temperature, and that the response of ri to AX at constant temperature (Fig. 5) may be exaggerated, since a deterioration of leaf water balance would reduce AX. Notwithstanding, it is considered that the temperature responses of gas exchange presented here are a close and workable approximation to behaviour in the field, and may be used for predictive purposes.
It is interesting to observe that dark respiration in P. hysterophorus is relatively insensitive to leaf temperature between 10 and 42°C. This behaviour contrasts markedly with that of most species, in which respiration increases at an exponential rate between about 10 and 40". The control of gas exchange in darkened leaves of P, hysterophorus is associated with an unusual degree of stomatal activity, which even suppressed respiration at temperatures between 35 and 42". Above this , temperature, extensive breakdown of the leaf occurred, and respiration was evidently uncontrolled (Fig. 6).
This collapse of leaf functions at high temperature may be critical in determining the distribution of P. hysterophorus in the field, although these extreme temperatures would usually be associated with conditions of aridity too severe for survival of the species (cf. Australian Bureau of Meteorology 1956). It is likely that the long-term temperature tolerance of P. hysterophorus differs somewhat from the responses described here, since many arid and semiarid zone plants can withstand at least brief exposure to temperatures in excess of 50°C (Bjorkmann 1975). However, since it is a short-lived annual with the ability in Queensland to grow and set seed after both summer and winter rains (Everist 1976), P. hysterophorus could well penetrate

Gas Exchange Characteristics of Parthenium Weed 459
to seasonally unfavourable areas provided that there existed at least one period of about 8 weeks' duration which was not subjected to temperature extremes.
Conclusion The pantropic distribution of the species in the humid regions of the Americas,
and its extension into temperate latitudes (Rollins 1950; Harvey 1976) are in conformity with the physiological characteristics described in this paper. Although the present studies were conducted on plants raised under conditions of low photon flux, it is unlikely that plants grown in the field would be more sensitive to temperature or water stress (cf. Ludlow and Ng 1976). It is suggested, therefore, that P. hystero- phorus has the potential to become a weed of great significance throughout the warm and temperate humid and subhumid regions of Australia.
Acknowledgment The advice of Mr G. J. Harvey, Queensland Department of Lands, Sherwood,
is gratefully acknowledged.
References Aston, M. J. (1976). Variation of stomatat diffusion resistance with ambient humidity in sunflower
(Helianthus annuus). Aust. J. Plant Physiol. 3, 489-501. Australia, Bureau of Meteorology (1956). 'Climatic Averages in Australia: Temperature, Relative
Humidity, Rainfall.' (Dep. Supply: Maribyrnong, Vic.). Bjorkmann, 0. (1975). Environmental and biological control of photosynthesis: inaugural
address. In 'Environmental and Biological Control of Photosynthesis', ed. R. Marcelle, pp. 1-16. (W. Junk: The Hague.)
Bjorkmann, O., and Holmgren, P. (1963). Adaptability of the photosynthetic apparatus to light intensity in ecotypes from exposed and shaded habitats. Physiol. Plant. 16, 889-914.
Boyer, J. S. (1970~). Leaf enlargement and metabolic rates in corn, soybean, and sunflower leaves at various leaf water potentials. Plant Physiol. 46, 233-5.
Boyer, J. S. (1970b). Differing sensitivity of photosynthesis to low leaf water potentials in corn and soybean. Plant Physiol. 46, 236-9.
Brix, H. (1962). The effect of water stress on the rates of photosynthesis and respiration in tomato plants and loblolly pine seedlings. Physiol. Plant. 15, 10-20.
Collatz, J., Ferrar, P. J., and Slatyer, R. 0. (1976). Effects of water stress and differential hardening treatments on photosynthetic characteristics of a xeromorphic shrub, Eucalyptus socialis, F. Muell. Oecologia (Berlin) 23, 95-105.
Doley, D., and Trivett, N. B. A. (1974). Effects of low water potentials on transpiration and photosynthesis in Mitchell grass (Astrebla lappacea). Aust. J. Plant Physiol. 1, 539-50.
Doley, D., and Yates, D. J. (1976). Gas exchange of Mitchell grass (Astrebla lappacea (Lindl.) Domin.) in relation to irradiance, carbon dioxide supply, leaf temperature and temperature history. Aust. J. Plant Physiol. 3, 471-87.
Everist, S. L. (1976). Parthenium weed. Qd. Agric. J. 102(2), (cover). Gauhl, E. (1976). Photosynthetic response to varying light intensity in ecotypes of Solanum dulcamara
L. from shaded and exposed habitats. Oecologia (Berlin) 22, 275-86. Gifford, R. M. (1974). A comparison of potential photosynthesis, productivity and yield of plant
species with differing photosynthetic metabolism. Aust. J. Plant Physiol. 1, 107-17. Hall, A. E., and Kaufmann, M. R. (1975). Stomata1 responses to environment with Sesamum
indicum L. Plant Physiol. 55, 455-9. Harvey, G . J. (1976). Pavthenium hysterophorus-literature review. Mimeo. (Queensland Dep.
Lands : Sherwood, Qld.) Hiesey, W. M., Nobs, M. A., and Bjorkmann, 0. (1971). Experimental studies on the nature of
species. V. Biosystematics, genetics and physiological ecology of the Erythranthe section of Mimulus. Publ. Carnegie Inst. Washington No. 628.

D. Doley
Lange, 0. L., Losche, R., Schulze, E.-D., and Kappen, L. (1971). Responses of stomata to changes ill humidity. Planta (Bcrbi?) 100, 76-86.
Levitt, J. (1972). 'Responses of Plants to Environmental Stresses.' (Academic Press: New York.) Ludlow, M. M., and Jarvis, P. G. (1971). Photosynthesis in sitka spruce (Picea sitchensis (Bong.)
Carr.). I. General characteristics. J. Appl. Ecol. 8, 925-53. Ludlow, M. M., and Ng, T. T. (1976). Efiect of water deficit on carbon dioxide exchange and
leaf elongation rate of Panicum n~aximum var. trichoglume. Aust. J. Plant Physiol. 3, 401-13. Ludlow, M. M., and Wilson, G. L. (1971). Photosynthesis of tropical pasture plants. 11. Temperature
and illuminance history. Aust. J. Biol. Sci. 24, 1065-75. Moore, R. T., White, R. S., and Caldwell, M. M. (1972). Transpiration of Atriplex confertifolia
and Eurotia lanata in relation to soil, plant and atmospheric moisture stresses. Can. J. Bot. 50, 2411-18.
Patterson, D. T. (1975). Photosynthetic acclimat~on to irradiance in Celastrus orbiculatus Thunb. Photosynthetica 9, 140-4.
Pieters, G. A., and Zima, M. (1975). Photosynthesis in desiccating leaves of poplar. Physiol. Plant. 34, 56-61.
Regehr, D. L., Bazzaz, F. A., and Boggess, W. R. (1975). Photosynthesis, transpiration and leaf conductance of Populus deltoides in relation to flooding and drought. Photosynthetica 9, 52-61.
Rollins, R. C. (1950). The guayule rubber plant and its relatives. Contrib. Gray Herb. Harv. Univ. No. 172.
Schulze, E.-D., Lange, 0. L., Evanari, M., Kappen, L., and Buschbom, U. (1974). The role of air humidity and leaf temperature in controlling stomatal resistance of Prunus armeniaca L. under desert conditions. I. A simulation of the daily course of stomatal resistance. Oecologia (Berlin) 17, 159-70.
Turner, N. C. (1969). Stomatal resistance to transpiration in three contrasting canopies. Crop Sci. 9, 303-7.
Turner, N. C. (1974). Stomatal behaviour and water status of maize, sorghum, and tobacco under field conditions. 11. At low soil water potential. Plant Physiol. 53, 360-5.
Van den Driessche, R., Connor, D. J., and Tunstall, B. R. (1971). Photosynthetic response of brigalow to irradiance, temperature and water potential. Photosynthetica 5, 210-17.
Wuenscher, J. E., and Kozlowski, T. T. (1971). The response of transpiration resistance to leaf temperature as a desiccation resistance mechanism in tree seedlings. Physiol. Plant. 24, 254-9.
Manuscript received 12 November 1976





























