Embed Size (px)
Citation preview

DEVELOPMENTAL DYNAMICS 20322S240 (1995)
Pattern of Keratinocyte Growth Factor and Keratinocyte Growth Factor Receptor Expression During Mouse Fetal Development Suggests a Role in Mediating Morphogenetic Mesenchymal-Epithelial Interactions PAUL W. FINCH, GERALD R. CUNHA, JEFFREY S. RUBIN, JANE WONG, AND DINA RON Department of Clinical Neurosciences, Brown University, Rhode Island Hospital, Providence, Rhode Island 02903 (P. W.F.); Department of Anatomy, University of California, San Francisco, California 94143 (G.R.C.); Laboratory of Cellular and Molecular Biology, National Cancer Institute, Bethesda, Maryland 20892 (J.S.R., J . W., D.R.)
ABSTRACT Mesenchymal cells are required for the induction of epithelial development during mammalian organogenesis. Keratinocyte growth factor (KGF) is a mesenchymally derived mitogen with specific activity for epithelial cells, suggest- ing that it may play a role in mediating these in- teractions. To further evaluate this hypothesis, in situ hybridization was used to examine the spatial distribution of KGF and KGF receptor (KGFR) transcripts during organogenesis and limb forma- tion in mouse embryos (days 14.5 through 16.5). To facilitate this aim, mouse KGF cDNA clones were isolated. There was extensive identity between the deduced mouse KGF protein sequence and that of its human and rat cognates, indicating that this gene has been highly conserved during mamma- lian evolution. In addition, mouse KGF protein was purified from fibroblasts and demonstrated to be structurally and functionally similar to hu- man KGF protein. For organs within the integu- mental, respiratory, gastrointestinal, and urogen- ital systems, whose development is dependent upon mesenchymal-epithelial interactions, KGF mRNA was detected in mesenchymal cells, while epithelial cells expressed transcripts for the KGFR. KGF and KGFR mRNA was also expressed in certain other tissues such as perichondrium, cartilage of developing bones, developing skeletal muscle, and visceral smooth muscle whose devel- opment is not regulated by mesenchymal-epithe- lial interactions. KGF expression was also de- tected in tissues isolated from human embryos, suggesting similar functions for KGF in human development. Taken together, our results suggest that KGF plays an important role in mediating mesenchymal-epithelial interactions during orga- nogenesis, but may also have other developmental functions in tissues not governed by such inter- actions. 0 1995 Wiley-Liss, Inc.
INTRODUCTION The normal proliferation of epithelial cells required
for crucial aspects of development is stimulated by in- ductive interactions with the mesenchyme. For exam- ple, organogenesis is initiated when the primitive epi- thelium interacts with specific mesenchymal tissue at the anatomical site of the future organ during defined periods of gestation. The resulting morphogenesis, growth, and functional cytodifferentiation lead to the development of organ-specific types of epithelium. The importance of such interactions has been well estab- lished for the development of many epithelial organ systems (Sengel, 1976; Cunha et al., 1983; Sawyer and Fallon, 1983; Saxen, 1987; Sanders, 1988; Gilbert, 1988; Sakakura, 1991).
Evidence for the involvement of polypeptide growth factors in mediating embryonic induction is accumu- lating. In particular, various members of the fibroblast growth factor (FGF) gene family have been implicated in mesoderm induction in amphibians, and appear to play a role in early pattern formation (Kimelman and Kirschner, 1987; Slack et al., 1987; Paterno et al., 1989). Furthermore, expression of these genes in later stages of mouse embryonic development implies a role in tissue differentiation and organogenesis (Wilkinson et al., 1989; Hebert et al., 1990; Haub and Goldfarb, 1991; Niswander and Martin, 1992; Drucker and Gold- farb, 1993; deLapeyriere et al., 1993). However, since most of the FGF family members have broad target cell specificities, and are expressed by multiple cell types, it is unclear what role they may play in the induction of specific epithelial components by the mesenchyme.
Keratinocyte growth factor (KGF) is a unique mem- ber of the FGF family (FGF-7) (Rubin et al., 1989; Finch et al., 1989). It was originally identified in the conditioned medium of a human embryonic lung fibro- blast cell line as a mitogen that was particularly active
Key words: Gene expression, In situ hybridiza- tion, Keratinocyte growth factor, Mesenchymal-epithelial interaction, Morphogenesis, Organogenesis
0 1995 WILEY-LISS, INC
Received September 30, 1994; accepted February 13, 1995. Paul W. Finch is now a t Ruttenberg Cancer Center, Mount Sinai
School of Medicine, Box 1130, One Gustave L. Levy Place, New York, NY 10029. Address reprint requestskorrespondence there.
Dina Ron is now a t Department of Biology, Technion-Israel Insti- tute of Technology, Technion City, Haifa 32 000, Israel.

224 FINCH ET AL.
on keratinocytes and specific for epithelial cells (Rubin et al., 1989). In contrast to the other FGF members, KGF does not appear to have activity on fibroblasts, endothelial cells, or other non-epithelial targets of FGF action (Rubin et al., 1989). The KGF transcript was detected in several stromal fibroblast cell lines derived from epithelial tissues of embryonic, neonatal, an adult sources, but not in any epithelial cell lines. Demonstra- tion of KGF expression in dermal but not epidermal extracts of whole skin tissue provided further evidence of the stromal pattern of KGF expression in vivo (Finch et al., 1989). The KGF receptor (KGFR) is a transmem- brane tyrosine kinase (Miki et al., 1991a) which binds KGF and aFGF with high affinity and bFGF a t a lower affinity (Bottaro et al., 1990). The KGFR is identical to the FGF receptor 2 (FGFR-2) except for a consecutive stretch of 49 amino acid residues which correspond to the carboxy-terminal half of the third immunoglobu- lin-like loop of the extracellular ligand binding domain and the adjacent stem region (Miki et al., 1992). In contrast, FGFR-2 does no bind KGF but shows high affinity for both aFGF and bFGF. Both receptors are encoded by alternative transcripts of the bek gene with the region of divergence determined by single alterna- tive exons. These exons encode the domains responsible for the different ligand binding properties (Miki et al., 1992). Initial surveys indicated that KGFR transcript expression was limited to epithelial cells, while FGFR-2 mRNA was present in cells of a variety of tis- sue types (Miki et al., 1992). Therefore, KGF repre- sents the only member of the FGF family with an ex- pression pattern and a target cell specificity consistent with a role as a paracrine effector of normal epithelial cell proliferation (Finch et al., 1989).
The aim of the present study was to investigate the role of KGF in mediating mesenchymal-epithelial in- teractions during critical periods of development, by examining KGF/KGFR transcript expression in mouse embryos during advanced organ development and limb formation. To facilitate this aim, we undertook the cloning of the mouse homologue of the human KGF cDNA and characterization of its product. Our results indicate that mouse KGF is analogous to its human counterpart in that i t encodes a mesenchymally de- rived mitogen with activity for epithelial cells. Fur- thermore, the expression patterns of KGF and KGFR in mouse embryos suggests that this factor plays a ma- jor role in mediating mesenchymal-epithelial interac- tions during fetal development.
RESULTS Isolation of Murine KGF cDNA Clone
NIH/3T3 cells have previously been reported to syn- thesize KGF (Miki et al., 1991a). In order to isolate a murine KGF cDNA clone, an NIH/3T3 cDNA library was screened with a human KGF cDNA probe under reduced stringency hybridization and washing condi- tions. Positive plaques were purified and examined by restriction enzyme analysis. Two clones containing
cDNA inserts of 1.9 and 3.0 kb were selected for further study. Sequence analysis of these clones revealed the presence of an open reading frame of 592 bp, encoding a polypeptide of 194 amino acids. Computer analysis of this sequence revealed 95.4 and 92.0% homology at the nucleotide level, and 95.9 and 94.3% identity at the predicted amino acid level with the respective rat and human sequences. The mouse cDNA hybridized to a 2.4 kb transcript present in total cell RNA isolated from NIH/3T3 cells (data not shown), which is comparable in size to the major KGF transcript present in human stromal fibroblasts (Finch et al., 1989).
Partial Purification and Characterization of the Murine KGF Protein
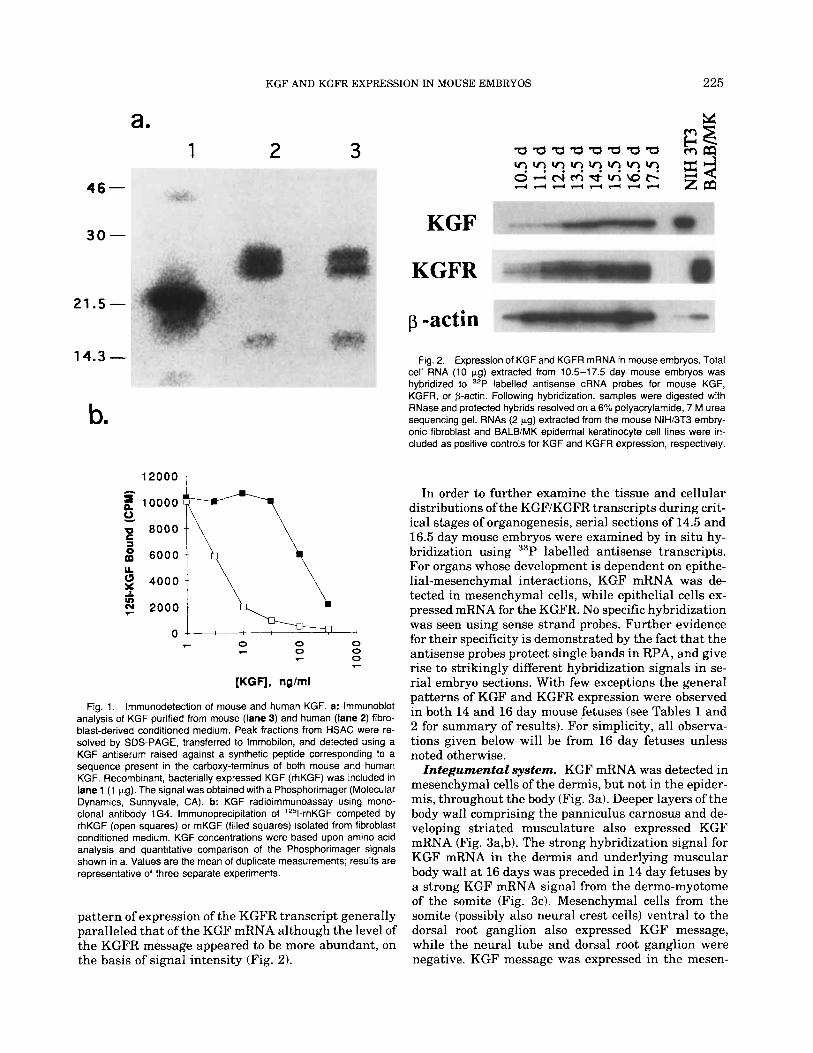
Mouse KGF (mKGF) protein was purified several hundred-fold from NIH/3T3 fibroblast-conditioned me- dium by using a combination of ultrafiltration and hep- arin-Sepharose affinity chromatography (HSAC). Frac- tions containing KGF activity were identified by their ability to stimulate 3H-thymidine incorporation in mouse BALB/MK epidermal keratinocytes. Stimula- tion of 3H-thymidine incorporation in BALB/MK cells was comparable to that elicited by human KGF (hKGF), and was blocked with KGF-specific, neutral- izing monoclonal antibodies raised against rhKGF (Alarid et al., 1994; data not shown). Immunoblotting of mKGF with a carboxy-terminal peptide antiserum revealed a 25-28 kDa doublet band corresponding to hKGF purified from M426 human fibroblast-condi- tioned medium (Fig. la). In addition, a -16 kDa pro- tein was detected in both the human and mouse KGF preparations that probably resulted from proteolysis of the full length molecules (Rubin, unpublished data). The doublet pattern likely reflects differences in gly- cosylation, and the faster mobility of bacterially ex- pressed rhKGF presumably is due to the absence of glycosylation. Using a KGF radioimmunoassay (Bot- taro et al., 1993), we observed that mKGF competed with 1251-rhKGF for binding to 1G4 monoclonal anti- body, although -30-fold higher concentrations were re- quired to achieve the same magnitude of displacement obtained with unlabeled rhKGF (Fig. lb) or fibroblast- derived hKGF (data not shown). Presumably, this re- flects subtle but significant changes in the epitope rec- ognized by 1G4. Taken together, our results indicate that mouse and human KGF proteins are structurally and functionally closely related.
KGF and KGFR mRNA Expression in Fetal Development
To determine the relative levels of expression of the KGF and KGFR transcripts during fetal development, ribonuclease protection assay (RPA) was performed us- ing total cell RNA isolated from mouse embryos at 10.5 through 17.5 day post coitum. As can be seen in Figure 2, the KGF signal was weakest in 10.5 day embryos, reached a peak in intensity at 14.5 days, and then re- mained at a steady state level through day 17.5. The
KGF AND KGFR EXPRESSION IN MOUSE EMBRYOS 225
a.
46 -
30 -
1 2 3
21.5 -
14.3 -
b.
0 C
0 a m LL (3 Y
0000
8000
6000
4000
2000
0 7.- 0 0 0
7 0 0 r 0
7
[KGF], nglml
Fig. 1. lmmunodetection of mouse and human KGF. a: lmmunoblot analysis of KGF purified from mouse (lane 3) and human (lane 2) fibro- blast-derived conditioned medium. Peak fractions from HSAC were re- solved by SDS-PAGE, transferred to Immobilon, and detected using a KGF antiserum raised against a synthetic peptide corresponding to a sequence present in the carboxy-terminus of both mouse and human KGF. Recombinant, bacterially expressed KGF (rhKGF) was included in lane 1 (1 pg). The signal was obtained with a Phosphorimager (Molecular Dynamics, Sunnyvale, CA). b: KGF radioimmunoassay using mono- clonal antibody 1 G4. lmmunoprecipitation of “’I-rhKGF competed by rhKGF (open squares) or mKGF (filled squares) isolated from fibroblast conditioned medium. KGF concentrations were based upon amino acid analysis and quantitative comparison of the Phosphorimager signals shown in a. Values are the mean of duplicate measurements; results are representative of three separate experiments.
pattern of expression of the KGFR transcript generally paralleled that of the KGF mRNA although the level of the KGFR message appeared to be more abundant, on the basis of signal intensity (Fig. 2).
KGF
KGFR
p -actin
Fig. 2. Expression of KGF and KGFR mRNA in mouse embryos. Total cell RNA (10 pg) extracted from 10.5-17.5 day mouse embryos was hybridized to 32P labelled antisense cRNA probes for mouse KGF, KGFR, or p-actin. Following hybridization, samples were digested with RNase and protected hybrids resolved on a 6% polyacrylamide, 7 M urea sequencing gel. RNAs (2 kg) extracted from the mouse NIH/3T3 embry- onic fibroblast and BALBiMK epidermal keratinocyte cell lines were in- cluded as positive controls for KGF and KGFR expression, respectively.
In order to further examine the tissue and cellular distributions of the KGFIKGFR transcripts during crit- ical stages of organogenesis, serial sections of 14.5 and 16.5 day mouse embryos were examined by in situ hy- bridization using 33P labelled antisense transcripts. For organs whose development is dependent on epithe- lial-mesenchymal interactions, KGF mRNA was de- tected in mesenchymal cells, while epithelial cells ex- pressed mRNA for the KGFR. No specific hybridization was seen using sense strand probes. Further evidence for their specificity is demonstrated by the fact that the antisense probes protect single bands in RPA, and give rise to strikingly different hybridization signals in se- rial embryo sections. With few exceptions the general patterns of KGF and KGFR expression were observed in both 14 and 16 day mouse fetuses (see Tables 1 and 2 for summary of results). For simplicity, all observa- tions given below will be from 16 day fetuses unless noted otherwise.
Integumental system. KGF mRNA was detected in mesenchymal cells of the dermis, but not in the epider- mis, throughout the body (Fig. 3a). Deeper layers of the body wall comprising the panniculus carnosus and de- veloping striated musculature also expressed KGF mRNA (Fig. 3a,b). The strong hybridization signal for KGF mRNA in the dermis and underlying muscular body wall at 16 days was preceded in 14 day fetuses by a strong KGF mRNA signal from the dermo-myotome of the somite (Fig. 3c). Mesenchymal cells from the somite (possibly also neural crest cells) ventral to the dorsal root ganglion also expressed KGF message, while the neural tube and dorsal root ganglion were negative. KGF message was expressed in the mesen-

TABLE 1. Expression of KGF mRNA in the 14- to 16-Day Mouse Embryo"
Orpan/tissue KGF KGF (Mason et al.. 1994) Aorta Adrenal Bladder Body wall muscle (ventral) Bone (facial) Braidspinal cord Choroid plexus Diaphragm Duodenum Efferent ducts Esophagus External genitalia Eye
Lens Optic nerve Cornea Eyelid Sclera Eye socket
Hair follicles
Inner ear Intercostal muscles Kidney
Laryngeal cartilage Larynx Limb cartilage Limb muscle Liver Lung Mammary gland Mesenchyme ventral to vertebrae Mullerian duct (male) Nasal cavity Olfactory epithelium Ovary Oviduct Palate Pancreas Pelvic bone Pituitary Rectum Ribs
Salivary gland Seminal vesicle Skin Small intestine
Somite (dermomyotome) 14 d Stomach (pyloris) Stomach (fore-stomach) Testis Thymus Thyroid Tongue Tooth
Trachea Ureter Urogenital sinus Uterus Vertebrae
b - -
Sm. Mus. +; fibroblasts - Mus. and fibroblasts + + + - 16d t + Mesenchyme 2; Sm. Mus. + + Mesenchyme + Mesenchyme + +; Sm. Mus. + t Mesenchyme + +
Mesenchyme + Mesenchyme + + Dermal papillae, mesenchyme + + Mesenchyme 2, + ND Col. duct mesenchyme + + ; Perichondrium + + ; Cart. - Mesenchyme + + Perichondrium + +; Cart. -
-
(-1 16d
nephron and mesenchyme -
+ Mesenchyme + + Mesenchyme + + + + Mesenchyme +
+ , - -
Mesenchyme + + ND -
ND Mesenchyme + Mesenchyme t Perichondrium + + Mesenchyme -, & Sm. Mus. +; fibroblasts - Mesenchyme + + , perichondrium + + , Mesenchyme + + Mesenchyme + + Dermis + + Mesenchyme -, t; Sm. Mus. -, t
cartilage -
Mesenchyme + + 14d Mesenchyme t , + ; Sm. Mus. + Mesenchyme + +; Sm. Mus. + + Mesenchyme 2 Mesenchyme +, Epi and WBC - Mesenchyme + Muscle + + Surrounding mesenchyme + + , Mesenchyme + + , cartilage rings + Mesenchyme + Mesenchyme + + ND Perichondrium + + Dermal papillae and mesenchyme + + Mesenchvme t . +
dental papillae -
NR NR Sm. Mus. +; fibroblasts + Mus. + + NR + 14d NR NR Mesenchyme +; Sm. Mus. ND NR Mesenchyme + + , Sm. Mus. ND NR
NR NR NR NR NR NR Dermal papillae, mesenchyme + t (+ ) 8.5-lld, (-1 after l l d Mesenchyme + + + NR
NR NR Perichondrium + + ; Cart. - + Mesenchyme + + NR NR NR Mesenchyme + + Mesenchyme + + Adult stroma + + Mucosa + NR Mesenchyme + NR NR NR NR
Mesenchyme + + NR Dermis + + Mesenchyme + , Sm. Mus. ND,
submuc + Myotome + + 9- l ld Mesenchyme + Mesenchyme + Mesenchyme - NR NR Muscle + + NR
NR Mesenchyme + NR Mucosa + + Perichondrium + + Dermal papillae and mesenchyme + + NR
Vibrissae Wolffian duct (male)
aComparative analysis of the anatomical sites of KGF expression detected by in situ hybridization with those described by Mason et al. (1994). Cart. = cartilage; Col. duct = collecting ducts; d = day; Epi = epithelium; ND = not determined; NR = not reported; Sm. Mus. = smooth muscle; sub muc = submucosa; WBC = white blood cells. bRelative level of mRNA expression is indicated by - = no expression; 2 = trace expression; +-+ + = relative mRNA expression level.

TABLE 2. Expression of KGFR mRNA in the 14- to 16-Day Mouse Embryo"
KGFR (Orr-Urtreger Orgadtissue KGFR et al., 1993; 14.5 day) Aorta Adrenal Bladder Bone (facial) Brainlspinal cord Choroid plexus Diaphragm Duodenum Efferent ducts Esophagus External genitalia
Lens Optic nerve Cornea Eyelid Sclera Eye socket
Hair follicles Heart Inner ear Kidney Laryngeal cartilage Larynx Limb cartilage Limb muscle Liver Lung Mammary gland Mesenchyme ventral to \ Mullerian duct (male) Nasal cavity Oral epithelium Ovary Palate Pancreas Pelvic bone Pharynx Pituitary Rectum Ribs Salivary gland Seminal vesicle
Eye
Skin Small intestine
Stomach (pyloris) Stomach (fore-stomach) Testis
Thymus Thyroid Tongue Tooth Trachea
Ureter Urogenital sinus Vertebrae Vibrissae
- b
+ Epithelium + +; Sm. Mus. - Periostium + Epithelium + + Epithelium + Epithelium + + Epithelium + + Epithelium + + Epithelium + + + + Epithelium + + Epithelium +
-
-
- -
Epithelium + + Epithelium + + Col. duct + +; nephrons - + + Epithelium + + Perichondrium + + ; cartilage -
t/ + Epithelium + + Epithelium + + Epithelium f Epithelium + + , cartilage + Epithelium + + Epithelium + + Epithelium + + Perichondrium + + , cartilage + Epithelium + + Epithelium + + Sm. Mus. +, Epi t Perichondrium + + , cartilage + Epithelium + + Epithelium + + Epidermis + + Epithelium + , mesenchyme -,
Sm. Mus. + ND Epithelium + Mesenchyme + , 2,
Epithelium + + Epithelium + + Epithelium + + Epithelium + + Epithelium + + , Epithelium + + Epithelium + + Perichondrium + + EDithehm + +
-
-+
rertebrae + +
-
seminiferous tubule -
cartilage rings + +
NR + NR Bone + NR NR NR NR NR NR NR
Epithelium + + NR EDithelium + + NR NR NR NR NR Epithelium + Nephrogenic cords + + NR NR Bone + NR + Epithelium + + Epithelium + + NR NR Epithelium + + Epithelium + + NR Epithelium + + Epithelium + + NR Epithelium + + EDithelium + + H>ndgut Epi + Cartilage + EDithelium + + NR Epidermis + + NR
NR Epithelium + NR
Epithelium + + NR NR Epithelium + + NR
NR NR Perichondrium + + EDithelium + +
Wolffian duct (male) Epithelium + + NR
"Comparative analysis of the anatomical sites of KGF expression detected by in situ hybridization with those described by Orr-Urtreger et al. (1993). Col. duct = collecting ducts; Epi = epithelium; ND = not determined; NR = not reported; Sm. Mus. = smooth muscle. bRelative level of mRNA expression is indicated by - = no expression; t = trace expression; +- + + = relative mRNA expression level.

228 FINCH ET AL.
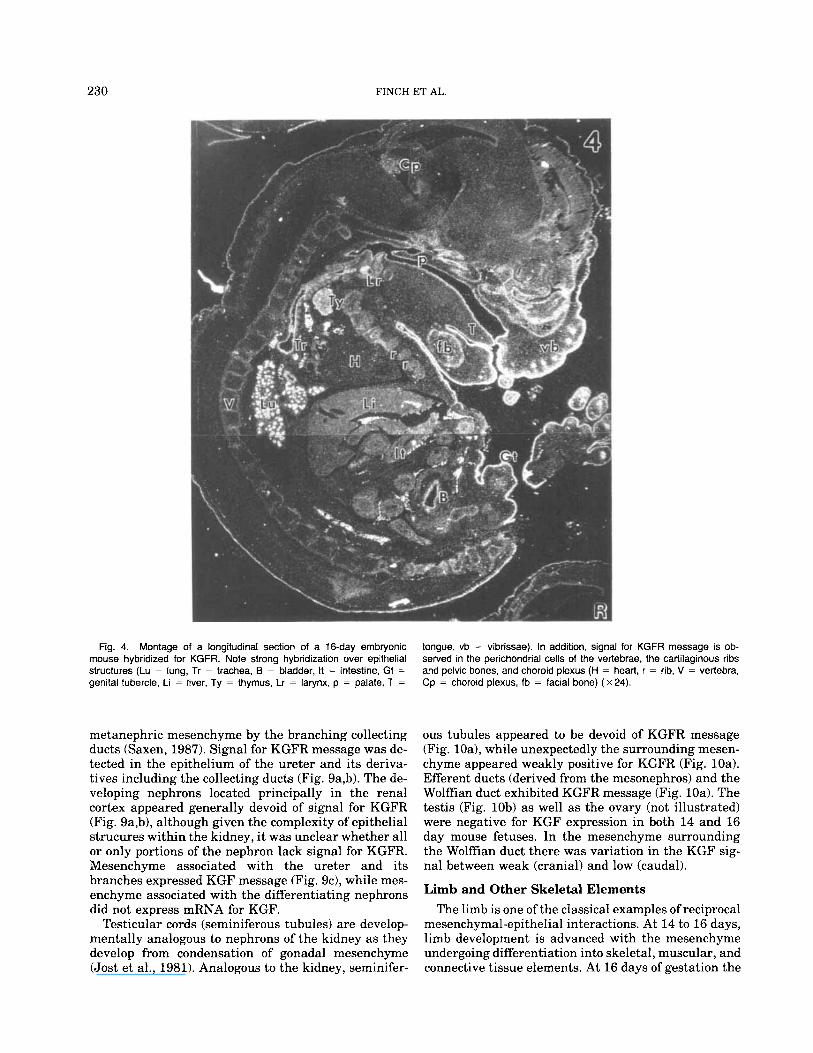
chyme surrounding the large vibrissal (Fig. 3d) and small pelage hair follicles as well as in mesenchyme associated with inter-follicular dermis. Generally, the hybridization signal for KGFR was uniform through- out the epidermis, its appendages, and glandular de- rivatives (Figs. 3-5). Strong hybridization for KGFR was observed in the general body surface epidermis (Fig. 4), epithelium of pelage and vibrissal hair follicles (Fig. 3e), epidermis of the eyelids, corneal epithelium (not illustrated), and epithelium of the external geni- talia (Fig. 4).
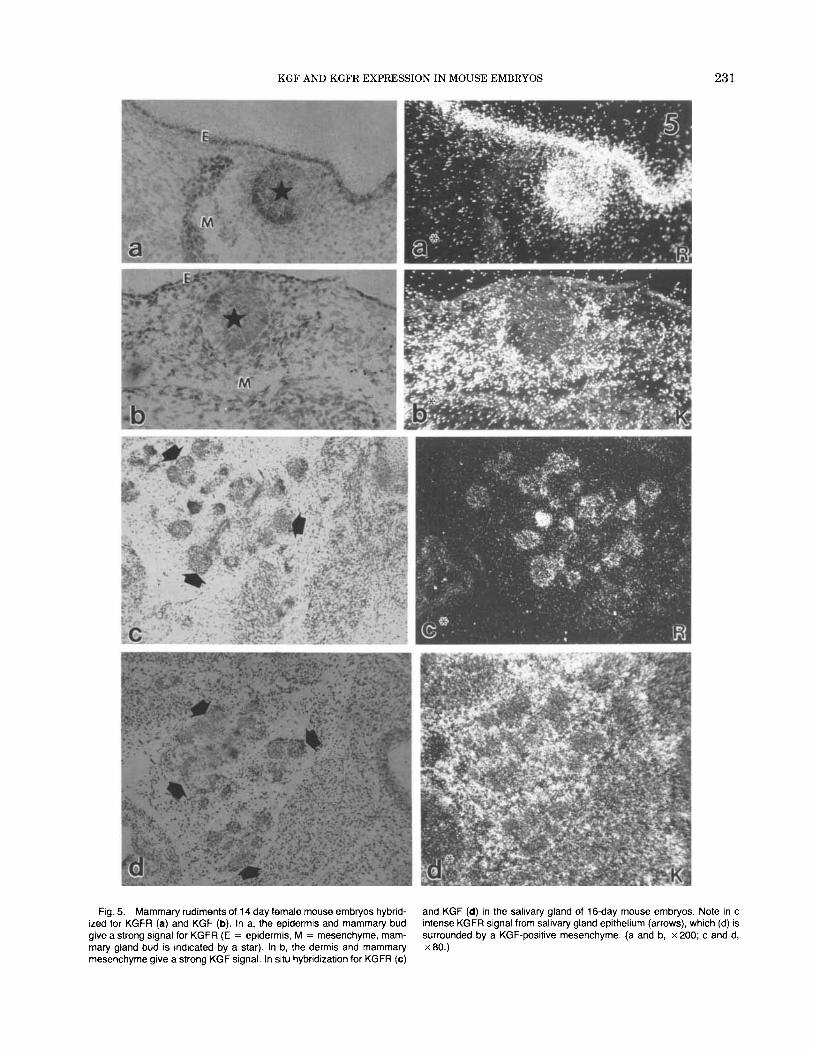
Mammary and submandibular salivary glands are examples of integumental derivatives whose develop- ment and morphogenesis are dependent upon mesen- chymal induction (Grobstein, 1967; Kratochwil, 1987). At 14 days of gestation the mammary anlage of female fetuses are represented as unbranched KGFR-positive epithelial buds (Fig. 5a), which are surrounded by mes- enchyme expressing KGF mRNA (Fig. 5b). Mammary mesenchyme expressing KGF mRNA was in direct con- tinuity with dermal mesenchyme expressing KGF mRNA. The submandibular salivary glands are glan- dular derivatives of the oral cavity. At 16 days of ges- tation the salivary glands were already extensively branched and seen in sections as epithelial cords em- bedded in mesenchyme. The salivary gland epithelium was intensely positive for KGFR message and was sur- rounded by a KGF mRNA-positive mesenchyme (Fig. 5c,d).
The oral and nasal cavities develop in large part from the body surface ectoderm which abuts the endo- derm in the oro-nasal pharynx. All epithelial struc- tures (tooth germs, and palatal, lingual, oral, and nasal epithelia) at 14 to 16 days of gestation strongly ex- pressed message for the KGFR (Fig. 4). Although less uniform, KGF message was detected in the mesen- chyme subadjacent to oral, nasal, lingual, and palatal epithelia (Fig. 6a). The dental papilla, the mesenchy- ma1 inducer of tooth development (Thesleff and Hur- merinta, 1981), was devoid of KGF mRNA, although the surrounding mesenchyme destined to form the fi- brous and skeletal components of the tooth socket ex- pressed KGF message (not illustrated). Unlike the re- mainder of the oral cavity, the epithelium of the tongue is associated with developing striated muscle. This stri- ated muscle was intensely positive for KGF mRNA (Fig. 6d).
Digestive and lower respiratory systems. On the whole, KGFR message was expressed throughout the epithelia of the digestive and lower respiratory systems and their glandular derivatives. The intense signal for KGFR mRNA observed in oral epithelium extended without diminution into the pharynx, larynx, trachea, and esophagus (Figs. 4,6b). Unexpectedly, moderate hybridization signal for KGFR message was also de- tected in the laryngeal and tracheal cartilages (Figs. 6b, 7a). KGF message was detected in the mesenchyme underlying pharyngeal, laryngeal, and esophageal ep- ithelia (Fig. 6a) and in the mesenchymal cells sur-
rounding the laryngeal cartilages. The bronchial tree was highly branched in 14 and 16 day embryos and ductal profiles exhibited an intense signal for KGFR (Figs. 4, 6e). Due to the complex ductal branching of the pulmonary epithelium, signal for KGF mRNA was rather diffuse, but nonetheless distinct hybridization for KGF mRNA was detected in pulmonary mesen- chyme (Fig. 6f).
KGFR expression was observed in epithelial deriva- tives of the foregut, such as the thymus (Figs. 4, 7a). and the thyroid (not illustrated). Epithelia of the thy- mus and thryoid were surrounded by mesenchymal cells that expressed KGF mRNA (not illustrated).
Within the abdominal cavity expression of KGF and KGF message varied considerably on an organ-by-or- gan basis. Signal for KGFR mRNA was intense in ep- ithelium of the abdominal segment of the esophagus (Fig. 7b) and the fore-stomach, while epithelia of the pylorus and small intestine (duodenum, jeunum, and ileum) and colon were at best weakly positive for KGFR mRNA (Fig. 7d). Expression of KGF message in the fibromuscular wall of these organs paralleled that of KGFR (Figs. 6c, 7c). In all of these organs the mes- enchymal wall had differentiated into fibroblastic and smooth muscle layers, and signal for both KGF and KGFR mRNAs of variable intensity was observed over the differentiating smooth muscle cells (Figs. 6c, 7b,d). In contrast, vascular smooth muscle of the aorta was consistently devoid of message for either KGF or KGFR (not illustrated). The liver, which at this stage is primarily a hematopoetic organ, showed diffuse signal for KGF (not illustrated) and KGFR (Fig. 4) barely above background.
Urogenital system. Endodermal epithelium of the urinary bladder and the urogenital sinus, the precursor of the prostate in males and the lower vagina in fe- males, exhibited a strong signal for KGFR (Fig. 8a) while the surrounding fibromuscular wall was nega- tive. In the bladder, KGF hybridization signal was low in fibroblasts underlying the urothelium but moderate in the developing detrusor smooth muscle (Fig. 8b). At 16 days of gestation, differentiation of urogenital sinus mesenchyme is not as advanced as that of the urinary bladder, and thus segregation of the mesenchyme into fibroblastic and smooth muscle layers has not occurred. The homogeneous androgen-receptor-positive mesen- chyme of the urogenital sinus (Cooke et al., 1991; Shannon and Cunha, 1983) expressed KGF message throughout a t moderate levels (Fig. 8b). In males the mesoderm-derived epithelia of the androgen-depen- dent Wolffian duct and its glandular derivative, the seminal vesicle, expressed KGFR and were surrounded by undifferentiated mesenchyme expressing KGF mes- sage at moderate (Wolffian duct) to high (seminal ves- icle) levels (Fig. 8a,b).
The metanephric kidney contains two epithelial lin- eages: (1) the epithelium of the ureter, major and minor calyces, and collecting ducts, all derived from the ure- teric bud, and (2) the nephron which is induced from
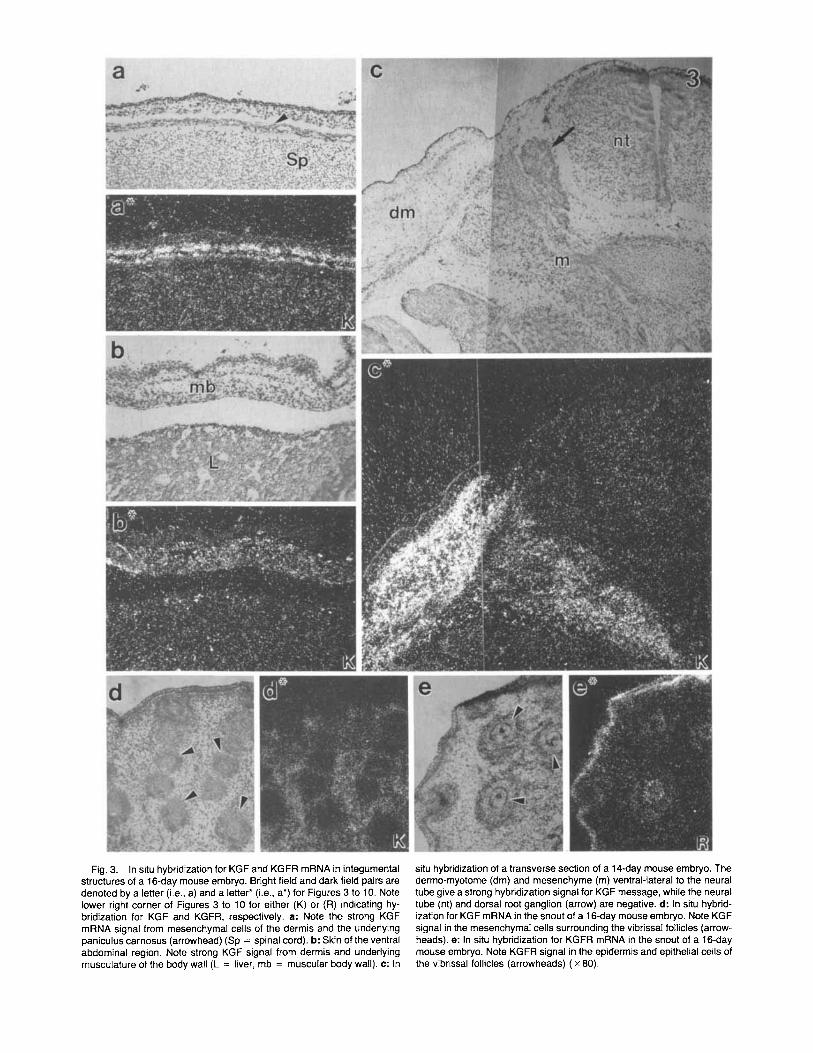
Fig. 3. In situ hybridization for KGF and KGFR mRNA in integumental structures of a 16-day mouse embryo. Bright field and dark field pairs are denoted by a letter (i.e., a) and a letter' (i.e., a*) for Figures 3 to 10. Note lower right corner of Figures 3 to 10 for either (K) or (R) indicating hy- bridization for KGF and KGFR, respectively. a: Note the strong KGF mRNA signal from mesenchymal cells of the dermis and the underlying paniculus carnosus (arrowhead) (Sp = spinal cord). b: Skin of the ventral abdominal region. Note strong KGF signal from dermis and underlying musculature of the body wall (L = liver, mb = muscular body wall). c: In
situ hybridization of a transverse section of a 14-day mouse embryo. The dermo-myotome (dm) and mesenchyme (m) ventral-lateral to the neural tube give a strong hybridization signal for KGF message, while the neural tube (nt) and dorsal root ganglion (arrow) are negative. d: In situ hybrid- ization for KGF mRNA in the snout of a 16-day mouse embryo. Note KGF signal in the mesenchymal cells surrounding the vibrissal follicles (arrow- heads). e: In situ hybridization for KGFR mRNA in the snout of a 16-day mouse embryo. Note KGFR signal in the epidermis and epithelial cells of the vibrissal follicles (arrowheads) ( x 80).
230 FINCH ET AL.
Fig. 4. Montage of a longitudinal section of a 16-day embryonic mouse hybridized for KGFR. Note strong hybridization over epithelial structures (Lu = lung, Tr = trachea, B = bladder, It = intestine, Gt = genital tubercle, Li = liver, Ty = thymus, Lr = larynx, p = palate, T =
tongue, vb = vibrissae). In addition, signal for KGFR message is ob- served in the perichondrial cells of the vertebrae, the cartilaginous ribs and pelvic bones, and choroid plexus (H = heart, r = rib, V = vertebra, Cp = choroid plexus, fb = facial bone) (x24).
metanephric mesenchyme by the branching collecting ducts (Saxen, 1987). Signal for KGFR message was de- tected in the epithelium of the ureter and its deriva- tives including the collecting ducts (Fig. 9a,b). The de- veloping nephrons located principally in the renal cortex appeared generally devoid of signal for KGFR (Fig. 9a,b), although given the complexity of epithelial strucures within the kidney, it was unclear whether all or only portions of the nephron lack signal for KGFR. Mesenchyme associated with the ureter and its branches expressed KGF message (Fig. 9c), while mes- enchyme associated with the differentiating nephrons did not express mRNA for KGF.
Testicular cords (seminiferous tubules) are develop- mentally analogous to nephrons of the kidney as they develop from condensation of gonadal mesenchyme (Jost et al., 1981). Analogous to the kidney, seminifer-
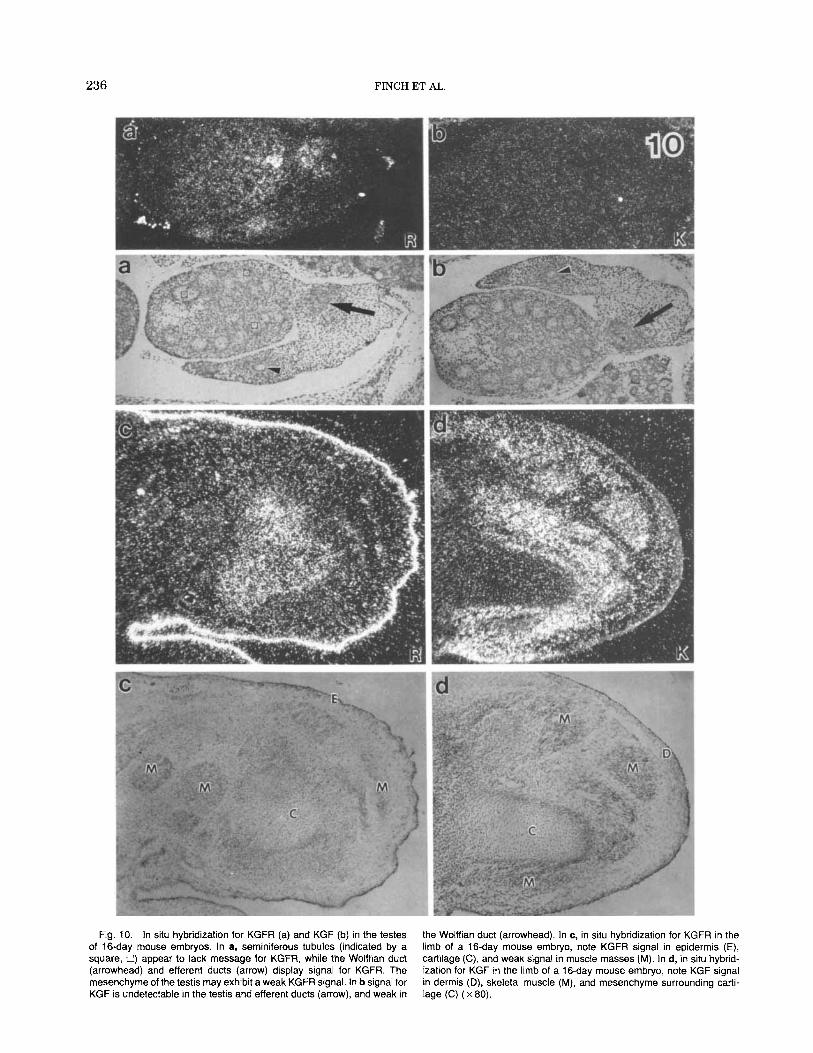
ous tubules appeared to be devoid of KGFR message (Fig. 10a), while unexpectedly the surrounding mesen- chyme appeared weakly positive for KGFR (Fig. 10a). Efferent ducts (derived from the mesonephros) and the WolfEan duct exhibited KGFR message (Fig. 10a). The testis (Fig. lob) as well as the ovary (not illustrated) were negative for KGF expression in both 14 and 16 day mouse fetuses. In the mesenchyme surrounding the Wolffian duct there was variation in the KGF sig- nal between weak (cranial) and low (caudal).
Limb and Other Skeletal Elements The limb is one of the classical examples of reciprocal
mesenchymal-epithelial interactions. At 14 to 16 days, limb development is advanced with the mesenchyme undergoing differentiation into skeletal, muscular, and connective tissue elements. At 16 days of gestation the
KGF AND KGFR EXPRESSION IN MOUSE EMBRYOS 23 1
Fig. 5. Mammary rudiments of 14 day female mouse embryos hybrid- ized for KGFR (a) and KGF (b). In a, the epidermis and mammary bud give a strong signal for KGFR (E = epidermis, M = mesenchyme, mam- mary gland bud is indicated by a star). In b, the dermis and mammary mesenchyme give a strong KGF signal. In situ hybridization for KGFR (c)
and KGF (d) in the salivary gland of 16-day mouse embryos. Note in c intense KGFR signal from salivary gland epithelium (arrows), which (d) is surrounded by a KGF-positive mesenchyme. (a and b, x 200; c and d, x80.)
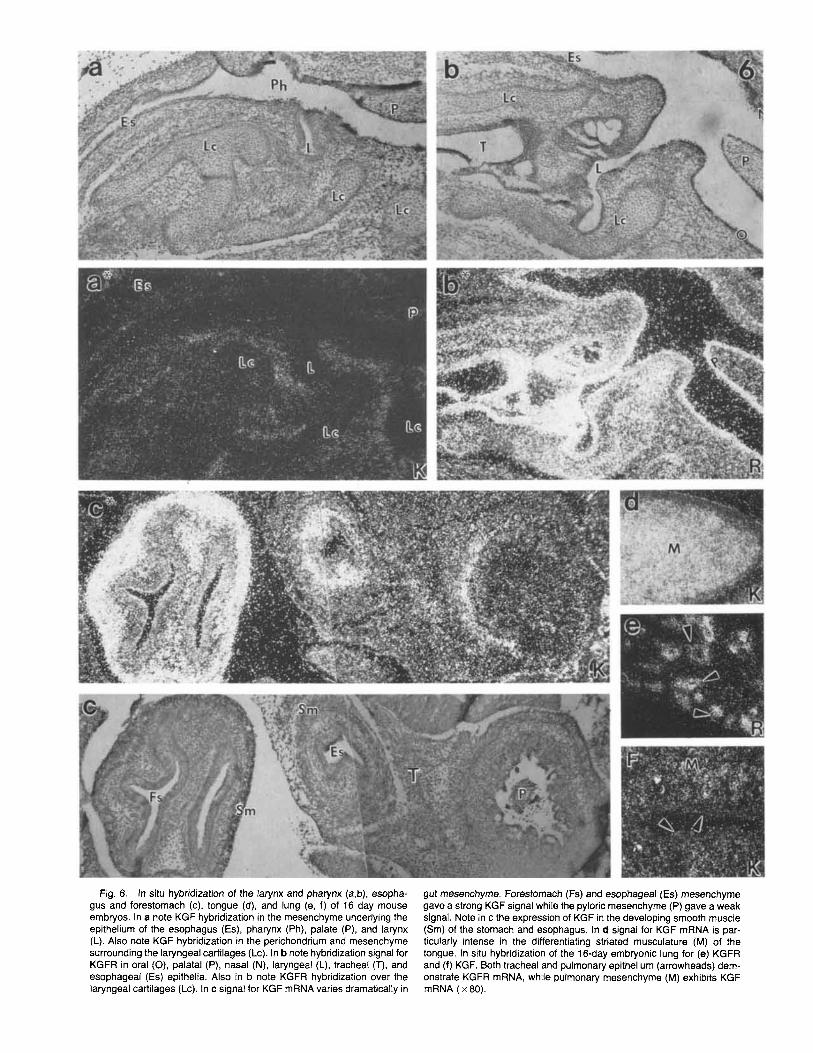
Fig. 6. In situ hybridization of the larynx and pharynx (a,b), esopha- gus and forestomach (c), tongue (d), and lung (e, f) of 16 day mouse embryos. In a note KGF hybridization in the mesenchyme underlying the epithelium of the esophagus (Es), pharynx (Ph), palate (P), and larynx (L). Also note KGF hybridization in the perichondrium and mesenchyme surrounding the laryngeal cartilages (Lc). In b note hybridization signal for KGFR in oral (0), palatal (P), nasal (N), laryngeal (L), tracheal (T), and esophageal (Es) epithelia. Also in b note KGFR hybridization over the laryngeal cartilages (Lc). In c signal for KGF mRNA varies dramatically in
gut mesenchyme. Forestomach (Fs) and esophageal (Es) mesenchyme gave a strong KGF signal while the pyloric mesenchyme (P) gave a weak signal. Note in c the expression of KGF in the developing smooth muscle (Sm) of the stomach and esophagus. In d signal for KGF mRNA is par- ticularly intense in the differentiating striated musculature (M) of the tongue. In situ hybridization of the 16-day embryonic lung for (e) KGFR and (f) KGF. Both tracheal and pulmonary epithelium (arrowheads) dem- onstrate KGFR mRNA, while pulmonary mesenchyme (M) exhibits KGF mRNA ( x 80).
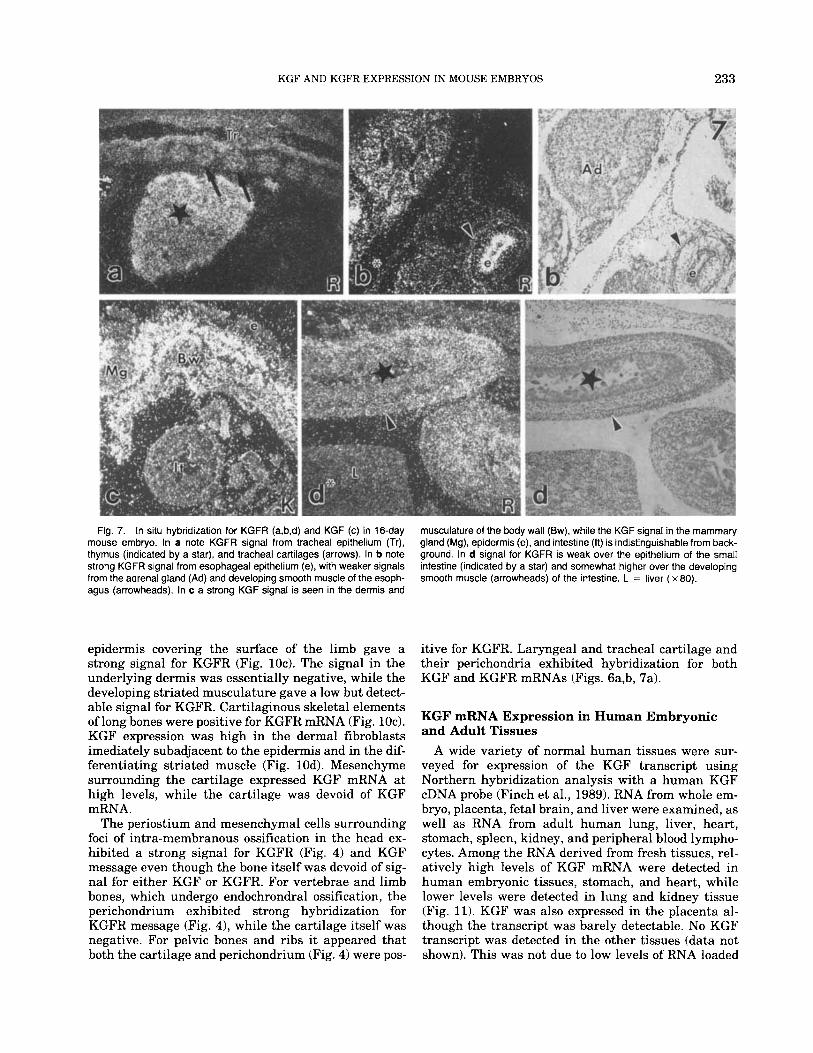
KGF AND KGFR EXPRESSION IN MOUSE EMBRYOS 233
Fig. 7. In situ hybridization for KGFR (a,b,d) and KGF (c) in 16-day mouse embryo. In a note KGFR signal from tracheal epithelium (Tr), thymus (indicated by a star), and tracheal cartilages (arrows). In b note strong KGFR signal from esophageal epithelium (e), with weaker signals from the adrenal gland (Ad) and developing smooth muscle of the esoph- agus (arrowheads). In c a strong KGF signal is seen in the dermis and
musculature of the body wall (Bw), while the KGF signal in the mammary gland (Mg), epidermis (e), and intestine (It) is indistinguishable from back- ground. In d signal for KGFR is weak over the epithelium of the small intestine (indicated by a star) and somewhat higher over the developing smooth muscle (arrowheads) of the intestine. L = liver ( x 80).
epidermis covering the surface of the limb gave a strong signal for KGFR (Fig. 1Oc). The signal in the underlying dermis was essentially negative, while the developing striated musculature gave a low but detect-
itive for KGFR. Laryngeal and tracheal cartilage and their perichondria exhibited hybridization for both KGF and KGFR mRNAs (Figs. 6a,b, 7a).
able signal for KGFR. Cartilaginous skeletal elements of long bones were positive for KGFR mRNA (Fig. 1Oc). KGF expression was high in the dermal fibroblasts
KGF mRNA Expression in Human Embryonic and
imediately subadjacent to the epidermis and in the dif- ferentiating striated muscle (Fig. 10d). Mesenchyme surrounding the cartilage expressed KGF mRNA at high levels, while the cartilage was devoid of KGF mRNA.
The periostium and mesenchymal cells surrounding foci of intra-membranous ossification in the head ex- hibited a strong signal for KGFR (Fig. 4) and KGF message even though the bone itself was devoid of sig- nal for either KGF or KGFR. For vertebrae and limb bones, which undergo endochrondral ossification, the perichondrium exhibited strong hybridization for KGFR message (Fig. 4), while the cartilage itself was negative. For pelvic bones and ribs it appeared that both the cartilage and perichondrium (Fig. 4) were pos-
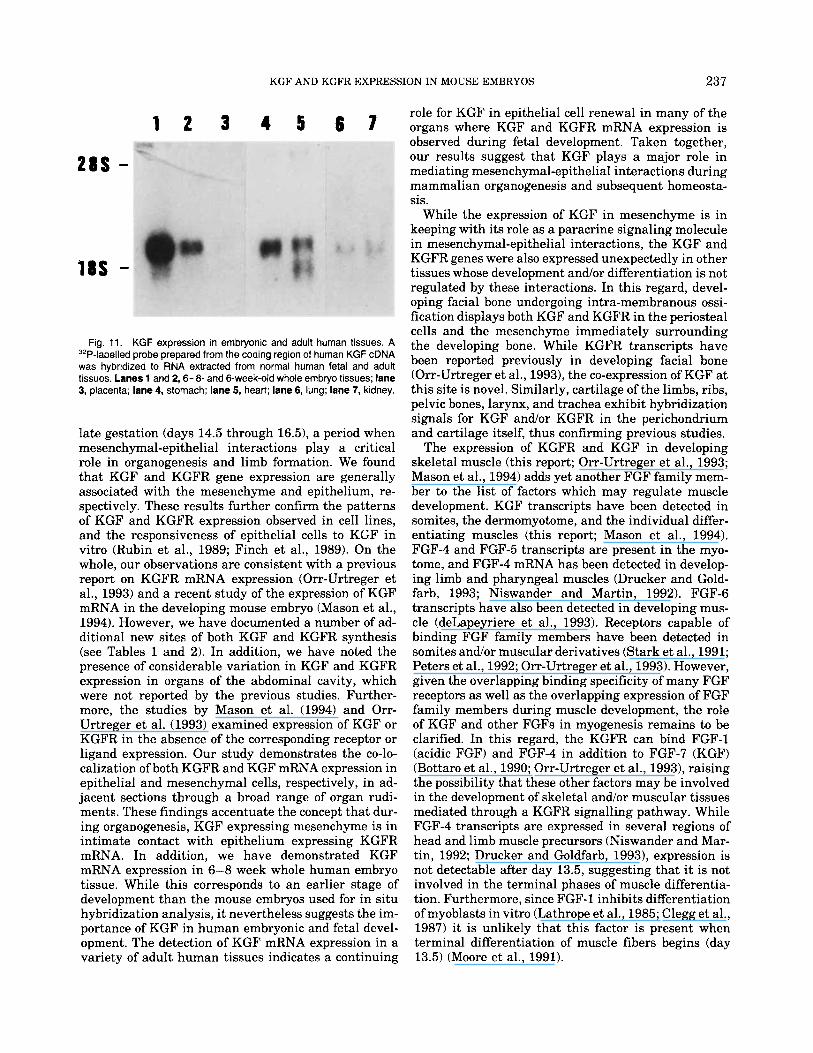
A wide variety of normal human tissues were sur- veyed for expression of the KGF transcript using Northern hybridization analysis with a human KGF cDNA probe (Finch et al., 1989). RNA from whole em- bryo, placenta, fetal brain, and liver were examined, as well as RNA from adult human lung, liver, heart, stomach, spleen, kidney, and peripheral blood lympho- cytes. Among the RNA derived from fresh tissues, rel- atively high levels of KGF mRNA were detected in human embryonic tissues, stomach, and heart, while lower levels were detected in lung and kidney tissue (Fig. 11). KGF was also expressed in the placenta al- though the transcript was barely detectable. No KGF transcript was detected in the other tissues (data not shown). This was not due to low levels of RNA loaded
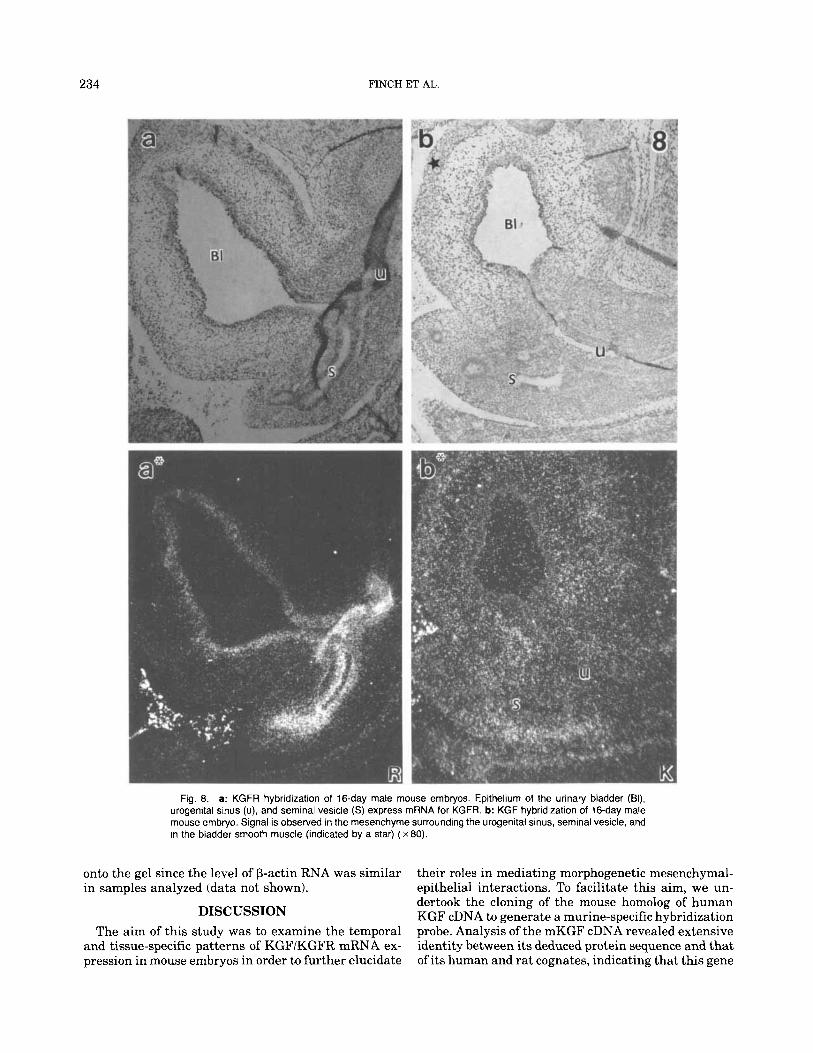
234 FINCH ET AL.
Fig. 8. a: KGFR hybridization of 16-day male mouse embryos. Epithelium of the urinary bladder (BI), urogenital sinus (u), and seminal vesicle (S) express mRNA for KGFR. b: KGF hybridization of 16-day male mouse embryo. Signal is observed in the mesenchyme surrounding the urogenital sinus, seminal vesicle, and in the bladder smooth muscle (indicated by a star) ( x 80).
onto the gel since the level of p-actin RNA was similar in samples analyzed (data not shown).
DISCUSSION The aim of this study was to examine the temporal
and tissue-specific patterns of KGF/KGFR mRNA ex- pression in mouse embryos in order to further elucidate
their roles in mediating morphogenetic mesenchymal- epithelial interactions. To facilitate this aim, we un- dertook the cloning of the mouse homolog of human KGF cDNA to generate a murine-specific hybridization probe. Analysis of the mKGF cDNA revealed extensive identity between its deduced protein sequence and that of its human and rat cognates, indicating that this gene
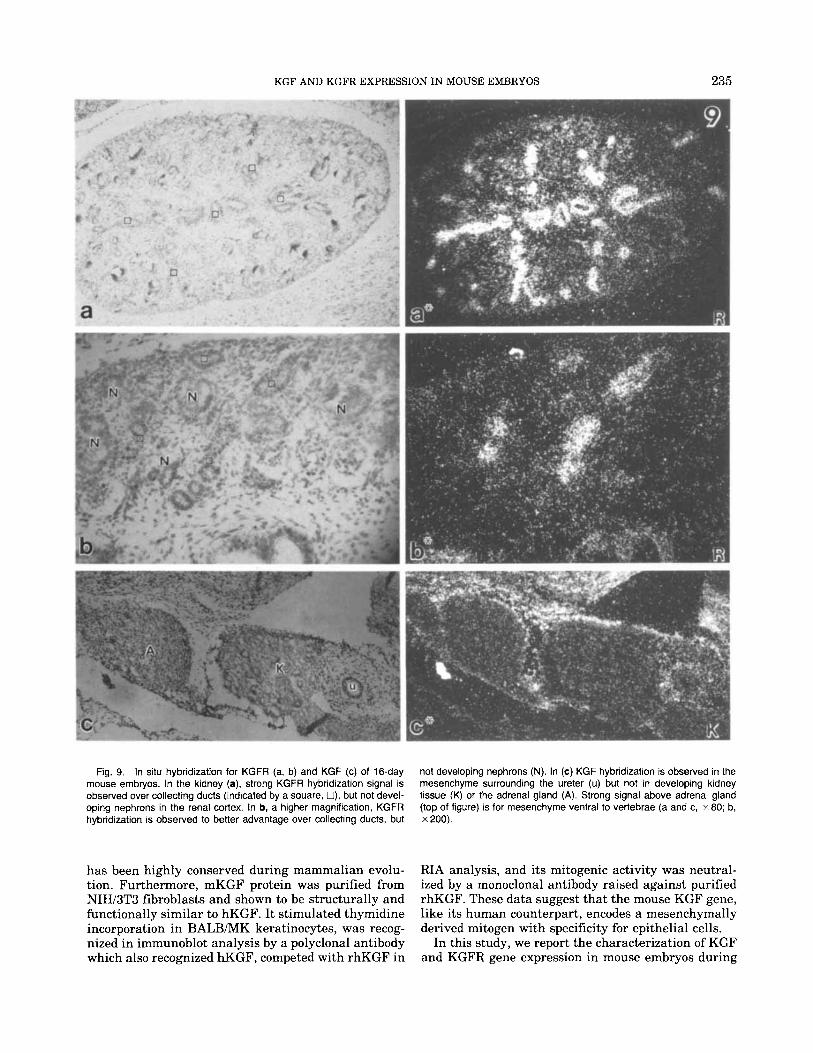
KGF AND KGFR EXPRESSION IN MOUSE EMBRYOS 235
Fig. 9. In situ hybridization for KGFR (a, b) and KGF (c) of 16-day mouse embryos. In the kidney (a), strong KGFR hybridization signal is observed over collecting ducts (indicated by a square, O), but not devel- oping nephrons in the renal cortex. In b, a higher magnification, KGFR hybridization is observed to better advantage over collecting ducts, but
not developing nephrons (N). In (c) KGF hybridization is observed in the mesenchyme surrounding the ureter (u) but not in developing kidney tissue (K) or the adrenal gland (A). Strong signal above adrenal gland (top of figure) is for mesenchyme ventral to vertebrae (a and c, x 80; b, x 200).
has been highly conserved during mammalian evolu- tion. Furthermore, mKGF protein was purified from NIH/3T3 fibroblasts and shown to be structurally and functionally similar to hKGF. It stimulated thymidine incorporation in BALB/MK keratinocytes, was recog- nized in immunoblot analysis by a polyclonal antibody which also recognized hKGF, competed with rhKGF in
RIA analysis, and its mitogenic activity was neutral- ized by a monoclonal antibody raised against purified rhKGF. These data suggest that the mouse KGF gene, like its human counterpart, encodes a mesenchymally derived mitogen with specificity for epithelial cells.
In this study, we report the characterization of KGF and KGFR gene expression in mouse embryos during
236 FINCH ET AL
Fig. 10. In situ hybridization for KGFR (a) and KGF (b) in the testes of 16-day mouse embryos. In a, seminiferous tubules (indicated by a square, U) appear to lack message for KGFR, while the Wolffian duct (arrowhead) and efferent ducts (arrow) display signal for KGFR. The mesenchyme of the testis may exhibit a weak KGFR signal. In b signal for KGF is undetectable in the testis and efferent ducts (arrow), and weak in
the Wolffian duct (arrowhead). In c, in situ hybridization for KGFR in the limb of a 16-day mouse embryo, note KGFR signal in epidermis (E), cartilage (C), and weak signal in muscle masses (M). In d, in situ hybrid- ization for KGF in the limb of a 16-day mouse embryo, note KGF signal in dermis (D), skeletal muscle (M), and mesenchyme surrounding carti- lage (C) ( x 80).
KGF AND KGFR EXPRESSION IN MOUSE EMBRYOS 237
18s -
role for KGF in epithelial cell renewal in many of the 1 2 3 4 5 6 7 organs where KGF and KGFR mRNA expression is
observed during fetal development. Taken together, our results suggest that KGF plays a major role in mediating mesenchymal-epithelial interactions during mammalian organogenesis and subsequent homeosta- sis.
While the expression of KGF in mesenchyme is in keeping with its role as a paracrine signaling molecule in mesenchymal-epithelial interactions, the KGF and KGFR genes were also expressed unexpectedly in other tissues whose development and/or differentiation is not regulated by these interactions. In this regard, devel- oping facial bone undergoing intra-membranous ossi- fication displays both KGF and KGFR in the periosteal cells and the mesenchyme immediately surrounding the developing bone. While KGFR transcripts have been reported previously in bone (Orr-UAreger et al., 1993), the co-expression of KGF at this site is novel. Similarly, cartilage of the limbs, ribs, pelvic bones, larynx, and trachea exhibit hybridization signals for KGF and/or KGFR in the perichondrium
late gestation (days 14.5 through 16.5), a period when and cartilage itself, thus confirming previous studies. mesenchymal-epithelial interactions play a critical The expression of KGFR and KGF in developing role in organogenesis and limb formation. We found skeletal muscle (this report; Orr-Urtreger et al., 1993; that KGF and KGFR gene expression are generally Mason et al., 1994) adds yet another FGF family mem- associated with the mesenchyme and epithelium, re- ber to the list of factors which may regulate muscle spectively. These results further confirm the patterns development. KGF transcripts have been detected in of KGF and KGFR expression observed in cell lines, somites, the dermomyotome, and the individual differ- and the responsiveness of epithelial cells to KGF in entiating muscles (this report; Mason et al., 1994). vitro (Rubin et al., 1989; Finch et al., 1989). On the FGF-4 and FGF-5 transcripts are present in the myo- whole, our observations are consistent with a previous tome, and FGF-4 mRNA has been detected in develop- report on KGFR mRNA expression (Orr-Urtreger et ing limb and pharyngeal muscles (Drucker and Gold- al., 1993) and a recent study of the expression of KGF farb, 1993; Niswander and Martin, 1992). F G F S mRNA in the developing mouse embryo (Mason et al., transcripts have also been detected in developing mus- 1994). However, we have documented a number of ad- cle (deLapeyriere et al., 1993). Receptors capable of ditional new sites of both KGF and KGFR synthesis binding FGF family members have been detected in (see Tables 1 and 2). In addition, we have noted the somites and/or muscular derivatives (Stark et al., 1991; presence of considerable variation in KGF and KGFR Peters et al., 1992; Orr-Urtreger et al., 1993). However, expression in organs of the abdominal cavity, which given the overlapping binding specificity of many FGF were not reported by the previous studies. Further- receptors as well as the overlapping expression of FGF more, the studies by Mason et al. (1994) and Orr- family members during muscle development, the role Urtreger e t al. (1993) examined expression of KGF or of KGF and other FGFs in myogenesis remains to be KGFR in the absence of the corresponding receptor or clarified. In this regard, the KGFR can bind FGF-1 ligand expression. Our study demonstrates the co-lo- (acidic FGF) and FGF-4 in addition to FGF-7 (KGF) calization of both KGFR and KGF mRNA expression in (Bottaro et al., 1990; Orr-Urtreger et al., 1993), raising epithelial and mesenchymal cells, respectively, in ad- the possibility that these other factors may be involved jacent sections through a broad range of organ rudi- in the development of skeletal andlor muscular tissues ments. These findings accentuate the concept that dur- mediated through a KGFR signalling pathway. While ing organogenesis, KGF expressing mesenchyme is in FGF-4 transcripts are expressed in several regions of intimate contact with epithelium expressing KGFR head and limb muscle precursors (Niswander and Mar- mRNA. In addition, we have demonstrated KGF tin, 1992; Drucker and Goldfarb, 1993), expression is mRNA expression in 6-8 week whole human embryo not detectable after day 13.5, suggesting that i t is not tissue. While this corresponds to an earlier stage of involved in the terminal phases of muscle differentia- development than the mouse embryos used for in situ tion. Furthermore, since FGF-1 inhibits differentiation hybridization analysis, it nevertheless suggests the im- of myoblasts in vitro (Lathrope et al., 1985; Clegg et al., portance of KGF in human embryonic and fetal devel- 1987) it is unlikely that this factor is present when opment. The detection of KGF mRNA expression in a terminal differentiation of muscle fibers begins (day variety of adult human tissues indicates a continuing 13.5) (Moore et al., 1991).
28s -
Fig. 11. KGF expression in embryonic and adult human tissues. A 3’P-labelled probe prepared from the coding region of human KGF cDNA was hybridized to RNA extracted from normal human fetal and adult tissues. Lanes 1 and 2’6-8- and 6-week-old whole embryo tissues; lane 3, placenta; lane 4, stomach; lane 5, heart; lane 6, lung; lane 7, kidney.

238 FINCH ET AL.
In addition to expression in skeletal muscle, we have noted signal for both KGF and KGFR in visceral smooth muscle (but not vascular smooth muscle). In organs of the abdominal cavity for example, the signals for KGF and KGFR often co-localized over the differ- entiating smooth muscle cells suggesting an autocrine mechanism of action for this cell type. KGF signal has also been detected in smooth muscle cells of the myo- metrium and walls of the spiral arteries in the adult uterus (Koji et al., 1994). Future studies utilizing func- tion perturbing antibodies and gene knockouts will be required to clarify the exact role of KGF in musculo- skeletal development, as well as in the many other organ systems in which this growth factor is expressed.
While mesenchymal signals are known to be essen- tial for epithelial development, the molecular nature of these signals has remained poorly understood. Consid- ering the complex nature of these signals, i t is likely that many factors are involved, including other growth factor receptor systems, cell adhesion molecules, and proteases (Birchmeier and Birchmeier, 1993). None- theless, KGF fulfills many of the criteria required of a mesenchymal mediator of epithelial development, in- cluding (1) KGF expression by mesenchymal or fibro- blastic cells, (2) epithelial cell expression of the KGFR, (3) inhibition of epithelial growth and development by neutralizing anti-KGF (Alarid et al., 19941, (4) the ability of exogenous KGF to replace androgens in pro- static development (Sugimura et al., 1993), and (5) in- hibition of normal lung development in transgenic mice with targeted expression of a dominant negative KGFR isoform to lung epithelium using the surfactant protein C promoter (Peters et al., 1994). However, other experimental approaches will be required to fully define the precise nature of KGF involvement in epi- thelial development. In this regard, earlier cell and organ culture studies have demonstrated that KGF is a paracrine mitogen for epithelial cells from a variety of different sources (Aaronson et al., 1991; Marchese et al., 1990; Pierce et al., 1994; Rubin et al., 1989; Ulich et al., 1994), as well as a morphogen in the ductal branch- ing morphogenesis of the neonatal mouse seminal ves- icle (Alarid et al., 1994) and the neonatal rat prostate (Sugimura et al., 1993). The extension of similar stud- ies to other organ systems will provide a powerful ap- proach in further elucidating the multiple roles of KGF during development.
EXPERIMENTAL PROCEDURES Collection of Embryos
Timed pregnant Swiss-Webster mice (Taconic, Ger- mantown, NY) were sacrificed at various stages of ges- tation (10.5 through 17.5 days post coitus) and the em- bryos dissected from the uterus. For RNA extraction, the tissues were snap frozen in liquid nitrogen and stored at -70°C. Total cellular RNA was isolated as described by Chirgwin et al. (1979). For in situ hybrid- ization, embryos were fixed overnight in 4% parafor- maldehyde in phosphate-buffered saline at 4°C. Follow-
ing fixation the embryos were dehydrated through graded alcohols, cleared with toluene, and embedded in paraffin. Longitudinal or cross sections were cut at 6 mm and mounted on aminoalkylsilane treated slides.
Generation of Murine KGF and KGFR cDNA Probes
An NIH/3T3 cDNA library was constructed in the pCEV27 phagemid vector (Miki et al., 1991b) using a unidirectional cloning system (Miki et al., 1989). Screening was performed with a SalI-/KpnI fragment consisting of the first 674 nucleotides of human KGF cDNA (Finch et al., 1989) using reduced stringency hybridization conditions. Several positive clones were isolated ranging in size from 0.8-3.0 kb. Two clones of insert size 1.9 and 3.0 kb were taken for sequence anal- ysis. For ribonuclease protection assays (RPA) and in situ hybridization experiments, a 227 bp PstI/SmaI fragment from the 5 ' noncoding region was subcloned into the pGEM3Zf( -) transcription vector (Promega, Madison, WI). After linearization with the appropriate restriction enzyme, antisense transcrips were synthe- sized using T7 RNA polymerase, and sense strand tran- scripts synthesized using SP6 RNA polymerase. A KGFR specific probe was generated by PCR amplifica- tion (Saiki et al., 1985) of a 148 bp fragment encoding exon K of the bek gene (Miki et al., 1992), from mouse chromosomal DNA. The fragment was cloned into the BamHI site of pGEM3Zf( -) such that transcription with T7 RNA polymerase generated antisense tran- scripts and SP6 RNA polymerase generated sense strand transcripts. For RPA, transcription reactions were performed using 32P labelled UTP (800 Ci/mmol, New England Nuclear, Boston, MA) as the labelled nu- cleotide, while for in situ hybridization experiments transcripts were labelled using 33P UTP (1,000-3,000 Ci/mmol, New England Nuclear).
RNase Protection Assay 32P labelled single-stranded antisense RNA probes
were prepared as described (Melton et al., 1984). Ten micrograms total cell RNA were hybridized overnight at 43°C with 1 x lo5 cpm of gel purified probe. Hybrids were digested with 0.1 U RNase A and 20 U RNase T1 (Ambion Inc., Austin, TX) for 60 min at 35°C. Protected fragments were resolved on 6% polyacrylamide, 8 M urea gels, and visualized by autoradiography.
In Situ Hybridization In situ hybridization was performed on deparaf-
finized tissue sections using antisense cRNA probes es- sentially as described by Wilkinson et al. (1987). 33P labelled probes were used at a concentration of 0.2 ng/ ml/kb length of cloned fragment in hybridization buffer (50% deionized formamide; 0.3 M NaCl; 20 mM Tris HCl, pH 8.0; 5 mM EDTA; 10% Dextran sulfate; 1 x Denhardts buffer; 0.5 mg/ml yeast RNA). Tissue sec- tions were hybridized a t 55°C for 16-18 hr and then washed under high stringency conditions (2 x SSC,

KGF AND KGFR EXPRESSION IN MOUSE EMBRYOS 239
50% formamide at 65°C). Non-hybridized probe was di- gested with 20 pg/ml RNase A for 30 min at 37°C. Following further washing under high stringency con- ditions, slides were dehydrated through graded alco- hols containing 0.3 M ammonium acetate. Sections were dipped in NTB-2 emulsion (Eastman Kodak, Rochester, NY) diluted 1:l in H,O, air dried and stored desiccated a t 4°C. After appropriate exposure times, slides were developed in Kodak D-19 developer and counterstained with 0.02% Toluidine blue.
Isolation and Characterization of Murine KGF NIH/3T3 conditioned medium was collected and con-
centrated by ultrafiltration (Rubin et al., 1989). The retentate from ultrafiltration was loaded onto a hepa- rin-Sepharose resin (Pharmacia, Piscataway, NJ), equilibrated in buffer containing 20 mM Tris-HC1, pH 7.5, 0.3 M NaCl. After extensive washing with equili- bration buffer, KGF activity was eluted from the col- umn using 0.6 M NaC1. KGF containing fractions were identified by monitoring 3H-thymidine incorporation into the DNA of BALB/MK mouse keratinocytes (Ru- bin et al., 1989) in the presence or absence of the KGF specific neutralizing monoclonal antibodies 1G4 or 2C4 (Alarid et al., 1994). Fractions containing KGF activity were pooled, concentrated a further 10- to 20-fold with a Centricon-10 microconcentrator (Amicon, Beverly, MA), and stored at -70°C for further analysis. Human KGF (hKGF) was prepared from M426 fibroblasts in a similar fashion.
Immunoblotting and Radioimmunoassay For immunoblotting, 12% polyacrylamide gels were
prepared by the procedure of Laemmli (1970). Samples were boiled for 3 min in the presence of 2.5% (vh) mer- captoethanol. Following electrophoresis, proteins were transferred, to polyvinylidene difluoride membranes (Immobilon, Millipore, Bedford, MA) for immunoblot- ting. A rabbit polyclonal antisera raised against a syn- thetic peptide corresponding to residues 179 to 194 was used for KGF detection (Ron et al., 1993).
For the KGF radioimmunoassay, 1251-labeled recom- binant human KGF (rhKGF) was immunoprecipitated by mouse monoclonal antibody 1G4 (Bottaro et al., 1993), in the presence of unlabeled rhKGF or mouse KGF (mKGF). Antibody-antigen complexes were re- covered and radioactivity measured as previously de- scribed (Bottaro et al., 1993). Bacterially-expressed, rhKGF was prepared as described (Ron et al., 1993).
ACKNOWLEDGMENTS We gratefully acknowledge Dr. Stuart Aaronson for
his advice and encouragement, Dr. Deborah Benzil for dissecting the mouse embryos used in this study, and Robert Interrante for technical assistance in analysis of KGF protein. This work was supported in part by grants from the Rhode Island Foundation (812 and 2010) and Amgen, Inc., to P.W.F., and NIH grants DK
32157, CA 05388, DK 47517, and Human Frontiers of Science Program grant RG-333/93 to G.R.C.
REFERENCES Aaronson, S.A., Bottaro, D.P., Miki, T., Ron, D., Finch, P.W., Fleming,
T.P., Ahn, J., Taylor, W.G., and Rubin, J.S. (1991) Keratinocyte growth factor: A fibroblast growth factor family member with an unusual target cell specificity. Annu. N.Y. Acad. Sci. 638:62-77.
Alarid, E.T., Rubin, J.S., Young, P., Chedid, M., Ron, D., Aaronson, S.A., and Cunha, G.R. (1994) Keratinocyte growth factor functions in epithelial induction during seminal vesicle development. Proc. Natl. Acad. Sci. U.S.A. 91:1074-1078.
Birchmeier, C., and Birchmeier, W. (1993) Molecular aspects of mes- enchymal-epithelial interactions. Annu. Rev. Cell Biol. 9511-540.
Bottaro, D.P., Rubin, J.S., Ron, D., Finch, P.W., Florio, C., and Aaron- son, S.A. (1990) Characterization of the receptor for keratinocyte growth factor. Evidence for multiple fibroblast growth factor recep- tors. J . Biol. Chem. 265:12767-12770.
Bottaro, D.P., Fortney, E., Rubin, J.S., and Aaronson, S.A. (1993) A keratinocyte growth factor receptor-derived peptide antagonist identifies part of the ligand binding site. J. Biol Chem. 268:9180- 9183.
Chirgwin, J., Przybula, A.E., MacDonald, R.J., and Rutter, W.J. (1979) Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 185294-5299.
Clegg, C.H., Linkhart, T.A., Olwin, B.B., and Hauschuka, S.P. (1987) Growth factor control of skeletal muscle differentiation: Commit- ment to terminal differentiation occurs in G1 phase and is repressed by fibroblast growth factor. J. Cell Biol. 105:949-956.
Cooke, P.S., Young, P., and Cunha, G.R. (1991) Androgen receptor expression in developing male reproductive organs. Endocrinology 128:2867-2873.
Cunha, G.R., Chung, L.W.K., Shannon, J.M., Taguchi, O., and Fujii, H. (1983) Hormone-induced morphogenesis and growth: Role of mesenchymal-epithelial interactions. Rec. Prog. Hormone Res. 39: 559-595.
delapeyriere, O., Ollendorff, V., Planche, J., Ott, M.O., Pizette, S., Coulier, F., and Birnbaum, D. (1993) Expression of the Fgfs gene is restricted to developing skeletal muscle in the mouse embryo. De- velopment 118:601-611.
Drucker, B.J., and Goldfarb, M. (1993) Murine FGF-4 expression is spatially restricted within embryonic skeletal muscle and other tis- sues. Mech. Dev. 40:155-163.
Finch, P.W., Rubin, J.S., Miki, T., Ron, D., and Aaronson, S.A. (1989) Human KGF is FGF-related with properties of a paracrine effector of epithelial cell growth. Science 245:752-755.
Gilbert, S.F. (1988) “Developmental Biology,” 3rd ed. New York Sin- auer.
Grobstein, C. (1967) Mechanisms of organogenetic tissue interaction. J. Natl. Cancer Inst. Monogr. 26:279-299.
Haub, O., and Goldfarb, M. (1991) Expression of the fibroblast growth factor-5 gene in the mouse embryo. Development 112:397-406.
Hebert, J.M., Basilico, C., Goldfarb, M., Haub, O., and Martin, G.R. (1990) Isolation of cDNAs encoding four mouse FGF family mem- bers and characterization of their expression patterns during em- bryogenesis. Dev. Biol. 138:454-463.
Jost, A., Magre, S., and Agelopoulou, C. (1981) Early stages of testic- ular differentiation in the rat. Hum. Genet. 58:59-63.
Kimelman, D., and Kirschner, M. (1987) Synergistic induction of me- soderm by FGF and TGFb, and indentification of an mRNA coding for FGF in the early Xenopus embryo. Cell 51:869-877.
Koji, T., Chedid, M., Rubin, J.S., Slayden, O.D., Csaky, K.G., Aaron- son, S.A., and Brenner, R.M. (1994) Progesterone-dependent ex- pression of keratinocyte growth factor mRNA in stromal cells of the primate endometrium: Keratinocyte growth factor as a progestome- din. J. Cell Biol. 125:393-401.
Kratochwil, K. (1987) Tissue combination and organ culture studies in the development of the embryonic mammary gland. In: “Devel- opmental Biology: A Comprehensive Synthesis,” Gwatkin, R.B.L. (ed). New York: Plenum Press, pp 315-334.

240 FINCH ET AL.
Laemmli, U.K. (1970) Cleavage of structural proteins during the as- sembly of the head of bacteriophage T4. Nature 227:680-685.
Lathrope, B., Olson, E., and Glaser, L. (1985) Control by fibroblast growth of differentiation in the BC3Hl muscle cell line. J . Cell Biol. 1001540-1547.
Marchese, C., Rubin, J.S., Ron, D., Faggioni, A,, Torrisi, M.R., Messina, A,, Frati, L., and Aaronson, S.A. (1990) Human kerati- nocyte growth factor activity on proliferation and differentiation of human keratinocytes: Differentiation response distinguishes KGF from EGF family. J. Cell Physiol. 144:326-332.
Mason, I.J., Pace, F.F., Smith, R., and Dickson, C. (1994) FGF-7 (ke- ratinocyte growth factor) expression during mouse development suggests roles in myogenesis, forebrain regionalization and epithe- lial-mesenchymal interactions. Mech. Dev. 15:15-30.
Melton, D.A., Krieg, P.A., Rebagliati, M.R., Maniatis, T., Zinn, K., and Green, M.R. (1984) Efficient in vitro synthesis of biologically active RNA and RNA hybridization probes from plasmids contain- ing a bacteriophage SP6 promoter. Nucleic Acids Res. 12:7035- 7056.
Miki, T., Matsui, T., Heidaran, M.A., and Aaronson, S.A. (1989) Effi- cient directional cloning system to construct cDNA libraries con- taining full-length inserts with high frequency. Gene 83:137-146.
Miki, T., Fleming, T.P., Bottaro, D.P., Rubin, J.S., Ron, D., and Aaron- son, S.A. (1991a) Expression cDNA cloning of the KGF receptor by creation of a transforming autocrine loop. Science 251:72-75.
Miki, T., Fleming, T.P., Crescenzi, M., Molloy, C.J., Blam, S.B, Rey- nolds, S.H., and Aaronson, S.A. (1991b) Development of a highly efficient expression cDNA cloning system: Application to oncogene isolation. Proc. Natl. Acad. Sci. U S A . 88:5167-5171.
Miki, T., Bottaro, D.P., Fleming, T.P., Smith, C.L., Burgess, W.H., Chan, A.M.-L., and Aaronson, S.A (1992) Determination of ligand- binding specificity by alternative splicing: Two distinct growth fac- tor receptors encoded by a single gene. Pror. Natl. Acad. Sci. U.S.A. 89246-250.
Moore, J.W., Dionne, C., Jaye, M., and Swain, J.L. (1991) The mRNAs encoding acidic FGF, basic FGF and FGF receptor are coordinately down regulated during myogenic differentiation. Development 111: 741-748.
Niswander, L., and Martin, G.R. (1992) Fgf-4 expression during gas- trulation, myogenesis, limb and tooth development in the mouse. Development 114:755-768.
Orr-Urtreger, A., Bedford, M.T., Burakova, T., Arman, E., Zimmer, Y., Yayon, A,, Givol, D., and Lonai, P. (1993) Developmental local- ization of the splicing alternatives of fibroblast growth factor re- ceptor-2 (FGFR2). Dev. Biol. 158475-486.
Paterno, G.D., Gillespie, L., Dixon, M., Slack, J.M.W., and Heath, J.K. (1989) Mesoderm-inducing properties of int-2 and kFGF: Two on- cogene encoded growth factors related to FGF. Development 106: 79-84.
Peters, K.G., Werner, S., Chen, G., and Williams, L.T. (1992) Two FGF receptor genes are differentially expressed in epithelial and mesenchymal tissues during limb formation and organogenesis in the mouse. Development 114:233-243.
Peters, K., Werner, S., Liao, X., Wert, S., Whitsett, J., and Williams, L. (1994) Targeted expression of a dominant negative FGF receptor
blocks branching morphogenesis and epithelial differentiation of the mouse lung. EMBO J. 12:973-986.
Pierce, G.F., Yanagihara, D., Klopchin, K., Danilenko, D.M., Hsu, E., Kenney, W.C., and Morris, C.F. (1994) Stimulation of all epithelial elements during skin regeneration by keratinocyte growth factor. J . Exp. Med. 179:831-840.
Ron, D., Bottaro, D.P., Finch, P.W., Morris, D.W., Rubin, J.S., and Aaronson, S.A. (1993) Expression of biologically active recombinant KGF Structure/function analysis of N-terminal truncation mu- tants. J . Biol. Chem. 268:2984-2988.
Rubin, J.S., Osada, H., Finch, P.W., Taylor, W.G., Rudikoff, S., and Aaronson, S.A. (1989) Purification and characterization of a newly identified growth factor specific for epithelial cells. Proc. Natl. Acad. Sci. U.S.A. 86:802-806.
Saiki, R.K., Scharf, S., Faloona, F., Mullis, K.B., Horn, G.T., Erlich, H.A., and Arnheim N. (1985) Enzymatic amplification of the b-globin genomic sequences and restriction site analysis for diag- nosis of sickle cell disease. Science 230:1350-1354.
Sakakura, T. (1991) New aspects of stroma-parenchyma relation in mammary gland differentiation. Int. Rev. Cytol. 125:165-202.
Sanders, E.J. (1988) The roles of epithelial-mesenchymal interactions in developmental processes. Biochem. Cell. Biol. 66530-540.
Sawyer, R.H., and Fallon, J.F. (1983) “Epithelial-Mesenchymal Inter- actions in Development.” New York Praeger.
Saxen, L. (1987) “Organogenesis of the Kidney.” New York: Cam- bridge University Press.
Sengel, P. (1976) “Morphogenesis of Skin.” New York Cambridge University Press.
Shannon, J.M., and Cunha, G.R. (1983) Autoradiographic localization of androgen binding in the developing mouse prostate. Prostate 4367-373.
Slack, J.M.W., Darlington, B.G., Heath, J.K., and Godsave, S.F. (1987) Mesoderm induction in early Xenopus embryos by heparin- binding growth factors. Nature 326:197-200.
Stark, K.L., McMahon, J.A., and McMahon, A.P. (1991) FGFR-4, a new member of the fibroblast growth receptor family, expressed in the definitive endoderm and skeletal muscle lineages of the mouse. Development 113:641-651.
Sugimura, Y., Cunha, G.R., Hayward, S., Hayasahi, N., Arima, K., Kawamura, J., Rubin, J.S., and Aaronson, S.A. (1993) Keratinocyte growth factor (KGF) is a mediator of testosterone-induced prostatic development. Mol. Cell. Differ. 1:423.
Thesleff, I., and Hurmerinta, K. (1981) Tissue interactions in tooth development. Differentiation 18:75-88.
Ulich, T.R., Yi, E.S., Longmuir, K., Yin, S., Blitz, R., Morris, C.F., Housley, R.M., and Pierce, G.F. (1994) Keratinocyte growth factor is a growth factor for type I1 pneumocytes in vivo. J. Clin. Invest. 93: 1298-1306.
Wilkinson, D.G., Bailes, J.A., Champion, J.E., and McMahon, A.P. (1987) A molecular analysis of mouse development from 8 to 10 days post coitum detects changes only in embryonic globin expres- sion. Development 99493-500.
Wilkinson, D.G., Bhatt, S., and McMahon, A.P. (1989) Expression pattern of the FGF-related proto-oncogene int-2 suggests multiple roles in feal development. Development 105: 131-136.





























