Embed Size (px)
Citation preview

lable at ScienceDirect
Journal of Human Evolution 61 (2011) 254e269
Contents lists avai
Journal of Human Evolution
journal homepage: www.elsevier .com/locate/ jhevol
People of the ancient rainforest: Late Pleistocene foragers at theBatadomba-lena rockshelter, Sri Lanka
Nimal Perera a,b, Nikos Kourampas c,d,*, Ian A. Simpson c, Siran U. Deraniyagala a, David Bulbeck e,Johan Kamminga e, Jude Perera a, Dorian Q. Fuller f, Katherine Szabó g, Nuno V. Oliveira e
a Sri Lanka Department of Archaeology, Excavation Branch, Sir Marcus Fernando Mawatha, Colombo 7, Sri Lankab School of Archaeology & Anthropology, The Australian National University, Canberra ACT 0200, Australiac School of Natural Sciences, University of Stirling, Stirling FK9 4LA, Scotland, UKdOffice of Lifelong Learning, University of Edinburgh, 11 Buccleuch Place, Edinburgh EH8 9LW, Scotland, UKeDepartment of Archaeology and Natural History, Australian National University, Canberra ACT 0200, Australiaf Institute of Archaeology, University College London, 31-34 Gordon Square, London WC1H 0PY, UKg School of Earth and Environmental Sciences, University of Wollongong, Northfields Avenue, Wollongong, NSW 2522, Australia
a r t i c l e i n f o
Article history:Received 31 March 2010Accepted 2 April 2011
Keywords:Homo sapiensSouth AsiaMicrolithsRainforest foragersEnvironmental archaeologyLate Palaeolithic
* Corresponding author.E-mail addresses: [email protected] (N. Per
ac.uk (N. Kourampas), [email protected] (I.A. S(S.U. Deraniyagala), [email protected] (Danu.edu.au (J. Kamminga), [email protected](D.Q. Fuller), [email protected] (K. Szabó), Nuno.oliveira
0047-2484/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.jhevol.2011.04.001
a b s t r a c t
Batadomba-lena, a rockshelter in the rainforest of southwestern Sri Lanka, has yielded some of theearliest evidence of Homo sapiens in South Asia. H. sapiens foragers were present at Batadomba-lena fromca. 36,000 cal BP to the terminal Pleistocene and Holocene. Human occupation was sporadic before theglobal Last Glacial Maximum (LGM). Batadomba-lena’s Late Pleistocene inhabitants foraged for a broadspectrum of plant and mainly arboreal animal resources (monkeys, squirrels and abundant rainforestsnails), derived from a landscape that retained equatorial rainforest cover through periods of pronouncedregional aridity during the LGM. Juxtaposed hearths, palaeofloors with habitation debris, postholes,excavated pits, and animal and plant remains, including abundant Canarium nutshells, reflect intensivehabitation of the rockshelter in times of monsoon intensification and biome reorganisation after ca.16,000 cal BP. This period corresponds with further broadening of the economic spectrum, evidencedthough increased contribution of squirrels, freshwater snails and Canarium nuts in the diet of therockshelter occupants. Microliths are more abundant and morphologically diverse in the earliest, pre-LGM layer and decline markedly during intensified rockshelter use on the wane of the LGM. Wepropose that changing toolkits and subsistence base reflect changing foraging practices, from shorter-lived visits of highly mobile foraging bands in the period before the LGM, to intensified use ofBatadomba-lena and intense foraging for diverse resources around the site during and, especially,following the LGM. Traces of ochre, marine shell beads and other objects from an 80 km-distant shore,and, possibly burials reflect symbolic practices from the outset of human presence at the rockshelter.Evidence for differentiated use of space (individual hearths, possible habitation structures) is present inLGM and terminal Pleistocene layers. The record of Batadomba-lena demonstrates that Late Pleistocenepathways to (aspects of) behavioural ‘modernity’ (composite tools, practice of symbolism and ritual,broad spectrum economy) were diverse and ecologically contingent.
� 2011 Elsevier Ltd. All rights reserved.
Introduction
The Late Pleistocene, from the warm Last Interglacial (OIS 5e: ca.125,000 BP) to the end of the Pleistocene epoch, was a period critical
era), [email protected]), [email protected]. Bulbeck), johan.kamminga@(J. Perera), [email protected]@anu.edu.au (N.V. Oliveira).
All rights reserved.
for the evolution of our species. During this period, Homo sapiens,which emerged in Africa between 200,000 and 150,000 years ago(Stringer, 2002), expanded its range and became a cosmopolitanspecies. Towards the end of the Pleistocene, by ca. 35e30,000 BP,people of a recognisably modern anatomy were present across theOld World, from the unglaciated parts of Atlantic Europe in thenorthwest to Sahul, then the joined Australia and New Guineacontinent, in the southeast (Bulbeck et al., 2003; Trinkaus, 2005).
In the course of their dispersal, H. sapiens foragers entered andadapted to novel habitats, from the deserts, tropical rainforests,

N. Perera et al. / Journal of Human Evolution 61 (2011) 254e269 255
estuaries and islands of equatorial Asia, to the periglacial tundra ofEurope and Tasmania (Oppenheimer, 2003, 2009; Bulbeck, 2007;Gilligan, 2007; Pope and Terrell, 2008; Petraglia et al., 2010).Many of these landscapes were already parts of a Late Pleistoceneecumene, whose hominin diversity is only hinted at by recentdiscoveries. These include bones and tools of Homo floresiensis,a late member of a clade rooted in early Homo (Morwood et al.,2004; Argue et al., 2009), and Homo erectus (Swisher et al., 1996),whichmay have overlappedwithH. sapiens in Island Southeast Asiafor tens of thousands of years. Outside Africa, H. sapiens pop-ulations, probably diverse at the infraspecific level (c.f. Lahr andFoley, 1998; Gunz et al., 2009), may have interacted with thesehominins in ways possibly more intricate and nuanced than earliermodels of total population replacement (e.g., Stringer and Andrews,1988) allowed. The presence of Neanderthal DNA in the genotypesof living non-Africans (Green et al., 2010) is a testimony to somelevel of interbreeding at the western end of the H. sapiens dispersalarc (Hodgson et al., 2010). The dispersal of H. sapiens out of Africa,with its ecological, demographic and cultural corollaries, is a centraltheme of research in human evolution and Pleistocene archaeology.Matters of contention include the number and timing of LatePleistocene dispersal events, the routes followed by dispersinggroups, the interactions of H. sapiens newcomers with other hom-inins, the degree to which non-sapiens populations were assimi-lated into the modern human gene pool, and the behavioural andtechnological traits that accompanied and, perhaps, facilitatedH. sapiens dispersal and settlement in novel and changing LatePleistocene habitats (e.g., Stringer, 2002; Oppenheimer, 2003, 2009;Cameron and Groves, 2004; Macauley et al., 2005; Trinkaus, 2005;Mellars, 2006a,b; Bulbeck, 2007; Endicott et al., 2007; Atkinsonet al., 2008; Pope and Terrell, 2008; Petraglia et al., 2010).
Homo sapiens in Late Pleistocene South Asia
Evidence from South Asia should be of critical bearing to thesediscussions, owing to South Asia’s location half way between Africaand Sahul (Fig. 1), its inferred high population densities in the later
Figure 1. a) Sri Lankan rockshelters in the Late Pleistocene world (emphasis on early pre-30Qafzeh: Shea, 2001; Darra-i-Kur: Angel, 1972; Sri Lankan rockshelters: Perera, 2010; ZhireBowler et al., 2003). b) Location of archaeological sites mentioned in the text. c) Setting of B
part of the Late Pleistocene (Atkinson et al., 2008), and its diverseLate Pleistocene cultures that differed markedly from their betterknown European and Levantine Upper Palaeolithic contemporaries(James and Petraglia, 2005; James, 2007; Petraglia et al., 2009a,2010).
This critical region, however, has yielded only a scanty andpatchy skeletal record for the Late Pleistocene. With the possibleexception of sporadic finds in its northern periphery (a robustH. sapiens cranial fragment at Darra-i-Kur, Afghanistan, associatedwith a Mousterian industry and dated tentatively to ca. 30,000 BP(Angel, 1972; Dupree and Davis, 1972) (Fig. 1), the earliest reliablydated H. sapiens skeletal remains in South Asia come from itssouthernmost tip: Late Pleistocene rockshelters in Sri Lanka, wherefragmentary remainsmay date back to ca. 38,000 cal BP (Beta 3394:33,070 � 410 BP uncalibrated at Fahien-lena: Kennedy et al., 1987;Kennedy and Deraniyagala, 1989; Deraniyagala, 1992; Wijeyapala,1997; Kennedy, 1999, 2000; Perera, 2010) (Fig. 1). These finds areamong the handful of Late Pleistocene skeletal remains in the vastswath of territory between Arabia and Australia (Kennedy, 2000;Bulbeck et al., 2003). Reconstructions of human dispersal in LatePleistocene South Asia, therefore, draw on the regional archaeo-logical record and genetic evidence from living populations (seeMisra, 2001; James, 2007; Oppenheimer, 2009; and Petraglia et al.,2010 for reviews).
The archaeological record suggests that diverse Middle Palae-olithic industries were present across South Asia duringmost of theLate Pleistocene. The few available absolute dates indicatea temporal range extending from the Last Interglacial (Achyutanet al., 2007) to ca. 38,000 BP or beyond (Petraglia et al., 2007,2009b). Diverse assemblages with a substantial blade and micro-blade component, microliths (small flake- and blade-based stonetools with bipolar blunting along one margin) and bone points,locally accompanied with beads, engraved ostrich shells and otherevidence for symbolic practice, made their appearance from ca.45e40,000 BP onwards (James and Petraglia, 2005; James, 2007;Petraglia et al., 2009a). These industries, variously termed as‘Upper Palaeolithic’ and, when microliths are present, ‘Microlithic’
,000 BP Homo sapiens sites east of Africa). (Pestera cu Oase: Trinkaus et al., 2003; Skhul/ndong: Liu et al., 2010; Niah: Barker et al., 2007; Callao: Mijares et al., 2010; Mungo:atadomba-lena rockshelter. d) Rockshelter entrance, at the base of a steep gneiss cliff.

N. Perera et al. / Journal of Human Evolution 61 (2011) 254e269256
and ‘Mesolithic’, are best understood as local manifestations of thediverse South Asian Late Palaeolithic (James and Petraglia, 2005). InSri Lanka and south India (Jwalapuram Locality 9: Clarkson et al.,2009), where such industries are associated with human remains,the latter are invariably H. sapiens.
When did H. sapiens first enter South Asia and what were theorigins of the region’s Late Palaeolithic? A set of models, which, fora time seemed to tie genetic, archaeological and skeletal evidenceneatly together, infer a single rapid dispersal from Africa sometimebetween ca. 83,000 and 60,000 years ago, during the arid OIS 4.People dispersing during that period of inland desert expansionmay have followed a southern coastal route, hugging coastal Arabiato South Asia (and from there to Southeast Asia and Sahul:Oppenheimer, 2003, 2009; Macauley et al., 2005; Mellars, 2006a;Bulbeck, 2007). The very rapid predicted rates of dispersal, whichbrought humans to Sahul by 50,000 BP if not earlier (Roberts et al.,1994; Balme et al., 2008), are attributed to the inferred maritimeorientation and technological sophistication of coastal foragers(which may have included the use of sea going craft), both tenta-tively traced to developments in Middle Stone Age (MSA) east andsouth Africa (e.g., Walter et al., 2000; Marean et al., 2007). The ca.45e40,000 BP interface between Middle and Late Palaeolithic inSouth Asia has been interpreted as the archaeological expression ofthe H. sapiens dispersal front. The novel technologies and behav-iours of the Late Palaeolithic have thus been viewed as introduc-tions by dispersing ‘modern’ humans (Mellars, 2006a), in obviousanalogy with the dominant interpretation of Europe’s early UpperPalaeolithic (Aurignacian).
Alternatively, and perhaps more realistically, dispersal patternseast of Africa may have beenmuchmore intricate both in space andin time. While including important coastal routes, other modelsinvolve inland pathways along the attractive habitats of South Asia’sbasins and river valleys (e.g., Field et al., 2007; Korrisetar, 2007), andacknowledge that H. sapiens dispersal may have extended overlonger time scales, perhaps stretching back to the clement,monsoonal climates of the Last Interglacial (Petraglia et al., 2010),and/or involved more than one dispersal events out of Africa (Lahrand Foley, 1994, 1998). Recent artefact finds above and belowYounger Toba ash fall at Jwalapuram Locality 3 in southern India,demonstrate that users of aMiddle Palaeolithic industry (with someaffinity with the South African MSA) were present in South Asiafrom as early as ca. 77,000 BP, and were able to endure the severeecological effects of the ca. 74,000 BP Younger Toba eruption, themost powerful volcanic event of the Quaternary (Ambrose, 1998),without any major change in their lithic technology (Petraglia et al.,2007; Haslam et al., 2010). If these resilient hominins wereH. sapiens (the interpretation favoured by the discovering team)they may have had descended from groups dispersing from Africaas early as the earlier part of the Last Interglacial (OIS 5), broadlycontemporaneously with the Skhul and Qafzeh populations in theLevant (Shea, 2001). The dating of skeletal remains with interme-diate H. sapiens and archaic features from Zhirendong cave in southChina to over 100,000 BP (Liu et al., 2010: Fig. 1) renders furthercredit to the suggestion that H. sapiens dispersal from Africa duringthe Last Interglacial may have extended over amuchwider Eurasianrange than hitherto thought.
South Asia may have been a hub of H. sapiens demographicexpansion and further dispersal outside Africa. Genetic evidencefrom living populations shows that, between ca. 60,000 and40,000 BP, the H. sapiens population of South (and neighbouringSoutheast) Asia may have increased five-fold (Atkinson et al., 2008;Fig. 1). This demographic expansion, which possibly renderedSouth Asia the most densely populated region of the Late Pleisto-cene world, may have been a critical driver of the technologicalinnovation and behavioural complexity that characterises the
region’s archaeological record after ca. 45,000 BP (Petraglia et al.,2009a, 2010).
Sri Lankan rockhselters (Fig. 1) have yielded the earliestH. sapiens remains in South Asia (from ca. 38,000 cal BP, at Fahien-lena, through to mid-Holocene times) in firm association with richand diverse Late Palaeolithic assemblages with microliths, boneand antler tools and shell beads (Kennedy et al., 1987; Kennedy andDeraniyagala, 1989; Deraniyagala, 1992; Wijeyapala, 1997; Kennedy,1999, 2000; Perera, 2010). These rockshelters postdate the arrival ofH. sapiens in the region by any reckoning and thus may have littledirect bearing on debates over the timing of dispersal. These sites,however, provide some of the earliest available evidence for thedeparture fromMiddle Palaeolithic industries and the emergence ofaspects of ‘modern’ human behaviour (composite tools, symbolismand ritual, range expansion into novel habitats) in South Asia. Thefirm association of human remains and artefacts with wellpreserved animal and plant remains affords potential proxies forchanging Late Pleistocene environments and allows us to situateLate Pleistocene subsistence, technological innovation and symbolicpractices in their dynamic ecological context (Deraniyagala, 1992;Kourampas et al., 2008, 2009a; Perera, 2010).
Here, we report results from the limited re-excavation ofBatadomba-lena rockshelter, one of these key sites (Fig. 1), in 2005,and from the re-evaluation of unpublished earlier material, exca-vated in 1980e1982. We describe and analyse the site stratigraphyand chronology, and provide assessments of its faunal and floralcontent. Based on these, we offer new insights into the ecologicalcontext of H. sapiens settlement in the rainforest of inland Sri Lankaand discuss some of the implications of the latter for broaderquestions of human behaviour and technological change in LatePleistocene South Asia.
Batadomba-lena: environmental setting and history of research
Batadomba-lena is a small (ca. 10 � 15 m), northeast-facingrockshelter on the steep banks of a stream on the foothills of SriPada (‘Adam’s Peak’), Ratnapura District, approximately 80 km eastof Colombo and the coast (6�46’N, 80�120E; Fig. 1). The rockshelterwas formed in gneiss bedrock, part of the high-grade metamorphicterrain (charnockites, ortho- and paragneisses, metapegmatites) ofthe Precambrian Highland Complex that forms the NNE-SSWtrending mountainous spine of Sri Lanka (Cooray, 1984). Awayfrom sites of agricultural modification, surrounding vegetationmainly comprises dense, three-storey lowland evergreen forest,dominated by species of Doona, Dipterocarpus and Mesua (Gaussenet al., 1968; Perera, 1975).
Batadomba-lena is one of Sri Lanka’s most thoroughlyresearched prehistoric sites. It was first excavated by P.E.P.Deraniyagala in 1938, who reported it as a Late Pleistocene-Holocene site of the Mesolithic ‘Balangoda Culture’ (Deraniyagala,1940, 1943). Post-independence, systematic excavations by the SriLanka Department of Archaeology were directed by Siran Dera-niyagala (1979e1986) and, in 2005, Nimal Perera (Deraniyagala,1992; Perera, 2010).
These excavations unearthed fragmentary and charred skeletalremains of more than 16 robust H. sapiens individuals of both sexesand of ages ranging from childhood to mature adulthood. Humanremains occur in association with abundant material culture andanimal and vegetable remains in largely undisturbed, Late Pleis-tocene habitation layers (Deraniyagala, 1992; Perera, 2010). Thesehuman skeletal elements were interpreted as fractional burials(Kennedy et al., 1987; Kennedy and Deraniyagala, 1989).
A multi-method dating programme (radiocarbon assays oncharcoal, bone and shell, ESR, TL, OSL), linked with the 1980e1982excavations, confirmed the Late to terminal Pleistocene age of the

N. Perera et al. / Journal of Human Evolution 61 (2011) 254e269 257
site, with most reliable dates falling in the period from ca.30e27,000 BP onwards (Agrawal et al., 1985; Deraniyagala, 1992;Abeyratne et al., 1997). The detailed stratigraphy and age of theearliest occupation layers, however, remained poorly documented.Resolving these issues was among the objectives of the 2005excavation.
Materials and methods
Excavation
The 2005 excavation extended over three continuous metre-sized grid squares abutting to the edge of the earlier 1980e1982excavation trench, and reached a depth of ca. 2.8 m from therockshelter floor. Excavation followed the site’s stratigraphicboundaries (Harris, 1989) and resolved the profile into a Harrismatrix of 125 contexts. These contexts were correlated with theseven layers/depositional phases identified in the more extensive1980e1982 excavations (an area of 6 � 5.5 m, with a depth of2.8 m).
Sediment analyses
Excavated sediments were described (lithology, Munsell colour,pH) and logged, sampled for bulk sediment analyses and wet-sieved in the adjacent stream for the collection of smallerfaunal, botanical and lithic finds (one and 2 mm mesh). Laboratoryanalyses at the Australian National University followed standardprocedures of archaeological sediment analysis (Limbrey, 1975;Hughes, 1983) and included grain size distribution (by sievingthrough a set of nested sieves with mesh sizes from 2 to0.0625 mm), moisture content (wet and dry sample weight), grainshape analysis (by low power optical microcopy), organic carbon(loss on ignition) and carbonate content (loss of weight aftertreatment with HCl). Fourteen undisturbed sediment samples werealso collected with Kubiena tins for micromorphological analysis.Thin sections were manufactured at the University of Stirling,following standard procedures of sediment curing, drying and resinimpregnation (http://www.thin.stir.ac.uk), and inspected witha polarising microscope under plane polarised, cross polarised andoblique incident light at magnifications �10e�400, in accordancewith protocols by Bullock et al. (1985), Courty et al. (1989) andStoops (2003).
Radiocarbon chronology
The current chronological framework is based on 14 radio-carbon assays on charcoal collected from wet sieving of the exca-vated sediment. Ten of these dates, from samples collected in thecourse of the 1980e1982 excavation, were published previously byDeraniyagala (1992). The four samples collected in the 2005 exca-vation were dated at the University of Waikato Radiocarbon DatingLaboratory (one conventional and three AMS dates). All 14 radio-carbon dates were calibrated using Oxcal version 4.1.6 (BronkRamsey, 2009: (c) Bronk Ramsey, 2010).
Animal and plant remains
Vertebrate remains were identified with the aid of the referencecollections available at the Sri Lanka Department of Archaeology.Quantitative analyses of the aggregate sample of 2005 and1980e1982 excavations (6677 identified skeletal elements) wereexpressed as number of individual specimens (NISP: Grayson,1984). A small sub-sample of invertebrate remains (ca. 17.4 kg)was also identified and analysed quantitatively at the Australian
National University, within the context of a pilot study. Plantmacroremains were identified as closely as reference collectionsallowed with the aid of low power optical microscopy and SEM (SriLanka Department of Archaeology and Australian NationalUniversity). Phytoliths and other microremains were extractedfrom sediment samples with flotation in sodium polytungstate at2.28 sg and identified with microscopic examination atmagnification �425 and �625 (Doreen Bowdery, pers. comm.,2007). Phytoliths and other plant remains were also identified fromsediment thin sections in the course of micromorphologicalanalysis.
Lithics, bone tools and ornaments
The large assemblage of over 400,000 flaked and/or utilisedlithics, and bone and shell artefacts recovered in the 1980e1982and 2005 excavations were identified and analysed according toDeraniyagala’s (1992) typology for the Sri Lankan prehistory. Deb-itage analysis (19,700 specimens) followed the approach of Sullivanand Rosen (1985). Use-wear/retouch traces and stone tool pathwaymanufacture sequences were identified by examination of 515specimens under a low power microscope, following methods byKamminga (1982) and Holdaway and Stern (2004).
Results
Stratigraphy and radiocarbon chronology
Batadomba-lena contains up to ca. 2.8 m of sediment fill. Thiswas resolved into 125 contexts, reflecting the complex depositionalhistory of the rockshelter (Fig. 2, Table SI). The earliest sediment(layer 8, not shown on Fig. 2) was deposited on vestiges of a redtropical ultisol (layer 9), developed on weathered bedrock (kaoli-nised gneiss). Layer 8 comprises undated loams, interpreted asfloodwater, slackwater and colluvial deposits, and devoid ofarchaeological content (Perera, 2010).
The earliest sediments with cultural content (layer 7, furtherresolved into units 7c, 7b and 7a, from base to top, respectively)consist of reddish gravelly loams with moderate quantities ofcharcoal, bone, snails and débitage, reflecting sporadic humanpresence in the rockshelter. Inferred processes of depositionincluded colluviation, roof disintegration, human-induced deposi-tion of burning residues, food and knapping debris, excavation ofintersecting pits in the rockshelter floor, and possibly shallowburial of disarticulated and fragmented human remains (Table SI).One AMS (30,603 �400 BP, uncalibrated) and one conventional(27,700 þ2090/-1660 BP, uncalibrated) radiocarbon assay on char-coal from the basal unit 7c date the onset of human presence in therockshelter between 36,280 and 31,450 cal BP (95.4% probability:Fig. 3). The earliest of these dates may be as much as 8000 yearsearlier than previously reported (cf. Kennedy et al., 1987; Kennedyand Deraniyagala,1989; Deraniyagala,1992). Four conventional andtwo AMS radiocarbon assays on charcoal from units 7b and 7ahigher up-section suggest that the deposition of layer 7 probablyextended to the onset of the LGM (Fig. 3). The long temporal rangeof layer 7, and clast weathering and palaeofloors manifest in themicromorphological evidence (Table S1), may reflect relatively lowrates of deposition, interspersed with periods of depositional stasis,and, perhaps, erosional truncation.
Human input increased in sediments deposited during andimmediately after the LGM (layer 6). A roof-fall episode separatesthese from juxtaposed hearths (ash and charcoal layers that overliefire-reddened sediment), probable rake-out deposits and palae-ofloors (layer 5) that reflect intensive habitation of Batadomba-lenain the millennia following ca. 16,000 BP (16,474e15,290 cal BP at

Figure 2. a) Schematic section across the Batadomba-lena rockshelter. b) Stratigraphy of the profile excavated in the 2005 season. Frames: samples for micromorphological analysis.Lower parts of the stratigraphy devoid of cultural content (layers 10 to 8: see Table SI), were not sampled by the 2005 trench.
N. Perera et al. / Journal of Human Evolution 61 (2011) 254e269258
95.4% probability) through to the terminal Pleistocene (Fig. 3).Metre-sized roof-fall blocks are incorporatedwithin layer 5 culturaldeposits, indicating that the immediate aftermath of the LGM wasa period of structural instability in the rockshelter (Fig. 2).
Terminal Pleistocene sediments (layers 4 and 3, from ca. 15,000to less than ca. 12,000 BP) comprise charcoal-rich loams andjuxtaposed in situ hearths, with bands of ash and charcoal onreddened loamy substratum (Figs. 2 and 4). Where identifiable,individual hearths are about 1m in diameter. Pits with charcoal-richsediment fill (up to ca. 50 cm deep) are common in post-LGM toterminal Pleistocene layers (Fig. 2). These structures, interpreted asearth ovens (Perera, 2010), are similar to cooking pits for thedetoxification of poisonous tubers reported from over 40,000 cal BPlevels at Niah Cave, Sarawak (Barker et al., 2007). Terminal Pleis-tocene sediments record accelerated deposition under intensehuman use of the rockshelter from ca. 15,000 BP onwards.
These sediments were affected by post-depositional dissolutionof bone and calcareous ash (more advanced in pre-LGM layers:Fig. 4f), neoformation of authigenic calcite and unidentified
Figure 3. Radiocarbon chronology of
(putative) phosphate minerals, and bioturbation by arthropods,which may have included soil-dwelling termites. Arthropod bio-turbationwas particularly intense in organics-rich palaeofloors andhabitation layers (Fig. 4c). Extraction of bat guano and theconstruction of a Buddhist shrine in the early 1940s have disturbedthe Holocene part of the stratigraphy (Fig. 2, Table SI).
Faunal remains
The cultural layers of Batadomba-lena are rich in vertebrate andinvertebrate faunal remains, with ca. 78 kg of total excavated boneand ca. 17.4 kg of shell from the limited 2005 excavation (Table 1and Table SII). Vertebrate remains are highly fragmented andcommonly burned, so that only ca. 20% of the excavated boneweight was identifiable taxonomically. Identified fauna is clearlydominated by small- and medium-bodied taxa (adult body weightgenerally � 20 kg: Fig. 5) of arboreal and mixed, terrestrial andarboreal habitat (over 82% of the total vertebrate assemblage). Thebones of monkeys constitute the majority of the mammalian
the Batdomba-lena rockshelter.

Figure 4. a) Juxtaposed ash laminae in stratigraphic association with postholes in early post-LGM layer 5. b) Detail of ash laminae, with snail shells, other food debris and charrednon-woody plant remains, above fire-reddened sediment (layer 5, context 51, scanned thin section). c) The same laminar deposit of ash and burned organics above intenselybioturbated substratum (note arthropod pellets). Later bioturbation galleries cut across microstratigraphic contacts. d) Articulated palm phytoliths (arrows) embedded ina groundmass of calcitic ash crystals are abundant in these layers, perhaps reflecting the role of palm-based artefacts (mating, thatching or wovenware) in Late Pleistocene materialculture. e) Snail shell (Sn) and non-charred Canarium nutshell (Cn) (layer 5 - context 51). f) Disintegrating bone fragments (arrows) in layer 7 (context 76). g) Palm phytolith fromLayer 6 (Context 81 ¼ 69 e image courtesy of Doreen Bowdery). h) Lithic in layer 7 (context 76). See Fig. 2 for stratigraphic reference
N. Perera et al. / Journal of Human Evolution 61 (2011) 254e269 259
sample, followed by squirrels, civets and mongooses (Fig. 6,Table SII). Ground-dwelling mammals are much more rare, repre-sented mainly by the small-bodied mouse deer and boar. Largeherbivores (e.g., elephants) and predators are very rare. Non-mammalian vertebrate remains mainly comprise jungle fowl,monitor lizards and snakes. Freshwater fish are very rare, althoughthis may reflect preservation bias, in view of the highly fragmentary
and burned character of the assemblage. A diversity of taxa ispresent in every layer: Even the layer 7a assemblage, with only 71identifications, includes giant squirrels, small squirrels, civets,mongoose, mouse deer, porcupine, terrapin, python, rats, bats, toad,fish and monkeys (Fig. 6, Table SII).
Abundant butchery marks on larger bones, widespread calci-nation and blackening from exposure to fire, spatial association of
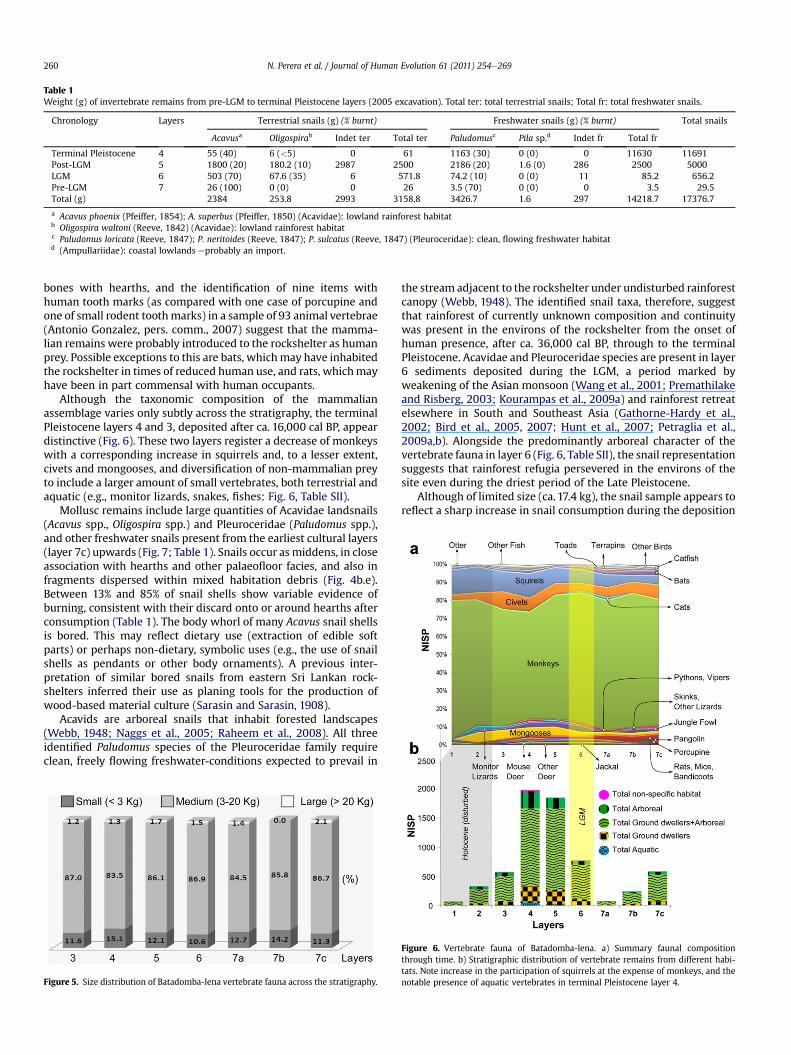
Table 1Weight (g) of invertebrate remains from pre-LGM to terminal Pleistocene layers (2005 excavation). Total ter: total terrestrial snails; Total fr: total freshwater snails.
Chronology Layers Terrestrial snails (g) (% burnt) Freshwater snails (g) (% burnt) Total snails
Acavusa Oligospirab Indet ter Total ter Paludomusc Pila sp.d Indet fr Total fr
Terminal Pleistocene 4 55 (40) 6 (<5) 0 61 1163 (30) 0 (0) 0 11630 11691Post-LGM 5 1800 (20) 180.2 (10) 2987 2500 2186 (20) 1.6 (0) 286 2500 5000LGM 6 503 (70) 67.6 (35) 6 571.8 74.2 (10) 0 (0) 11 85.2 656.2Pre-LGM 7 26 (100) 0 (0) 0 26 3.5 (70) 0 (0) 0 3.5 29.5Total (g) 2384 253.8 2993 3158.8 3426.7 1.6 297 14218.7 17376.7
a Acavus phoenix (Pfeiffer, 1854); A. superbus (Pfeiffer, 1850) (Acavidae): lowland rainforest habitatb Oligospira waltoni (Reeve, 1842) (Acavidae): lowland rainforest habitatc Paludomus loricata (Reeve, 1847); P. neritoides (Reeve, 1847); P. sulcatus (Reeve, 1847) (Pleuroceridae): clean, flowing freshwater habitatd (Ampullariidae): coastal lowlands eprobably an import.
N. Perera et al. / Journal of Human Evolution 61 (2011) 254e269260
bones with hearths, and the identification of nine items withhuman tooth marks (as compared with one case of porcupine andone of small rodent toothmarks) in a sample of 93 animal vertebrae(Antonio Gonzalez, pers. comm., 2007) suggest that the mamma-lian remains were probably introduced to the rockshelter as humanprey. Possible exceptions to this are bats, whichmay have inhabitedthe rockshelter in times of reduced human use, and rats, whichmayhave been in part commensal with human occupants.
Although the taxonomic composition of the mammalianassemblage varies only subtly across the stratigraphy, the terminalPleistocene layers 4 and 3, deposited after ca. 16,000 cal BP, appeardistinctive (Fig. 6). These two layers register a decrease of monkeyswith a corresponding increase in squirrels and, to a lesser extent,civets and mongooses, and diversification of non-mammalian preyto include a larger amount of small vertebrates, both terrestrial andaquatic (e.g., monitor lizards, snakes, fishes: Fig. 6, Table SII).
Mollusc remains include large quantities of Acavidae landsnails(Acavus spp., Oligospira spp.) and Pleuroceridae (Paludomus spp.),and other freshwater snails present from the earliest cultural layers(layer 7c) upwards (Fig. 7; Table 1). Snails occur as middens, in closeassociation with hearths and other palaeofloor facies, and also infragments dispersed within mixed habitation debris (Fig. 4b.e).Between 13% and 85% of snail shells show variable evidence ofburning, consistent with their discard onto or around hearths afterconsumption (Table 1). The body whorl of many Acavus snail shellsis bored. This may reflect dietary use (extraction of edible softparts) or perhaps non-dietary, symbolic uses (e.g., the use of snailshells as pendants or other body ornaments). A previous inter-pretation of similar bored snails from eastern Sri Lankan rock-shelters inferred their use as planing tools for the production ofwood-based material culture (Sarasin and Sarasin, 1908).
Acavids are arboreal snails that inhabit forested landscapes(Webb, 1948; Naggs et al., 2005; Raheem et al., 2008). All threeidentified Paludomus species of the Pleuroceridae family requireclean, freely flowing freshwater-conditions expected to prevail in
Figure 5. Size distribution of Batadomba-lena vertebrate fauna across the stratigraphy.
the stream adjacent to the rockshelter under undisturbed rainforestcanopy (Webb, 1948). The identified snail taxa, therefore, suggestthat rainforest of currently unknown composition and continuitywas present in the environs of the rockshelter from the onset ofhuman presence, after ca. 36,000 cal BP, through to the terminalPleistocene. Acavidae and Pleuroceridae species are present in layer6 sediments deposited during the LGM, a period marked byweakening of the Asian monsoon (Wang et al., 2001; Premathilakeand Risberg, 2003; Kourampas et al., 2009a) and rainforest retreatelsewhere in South and Southeast Asia (Gathorne-Hardy et al.,2002; Bird et al., 2005, 2007; Hunt et al., 2007; Petraglia et al.,2009a,b). Alongside the predominantly arboreal character of thevertebrate fauna in layer 6 (Fig. 6, Table SII), the snail representationsuggests that rainforest refugia persevered in the environs of thesite even during the driest period of the Late Pleistocene.
Although of limited size (ca. 17.4 kg), the snail sample appears toreflect a sharp increase in snail consumption during the deposition
Figure 6. Vertebrate fauna of Batadomba-lena. a) Summary faunal compositionthrough time. b) Stratigraphic distribution of vertebrate remains from different habi-tats. Note increase in the participation of squirrels at the expense of monkeys, and thenotable presence of aquatic vertebrates in terminal Pleistocene layer 4.
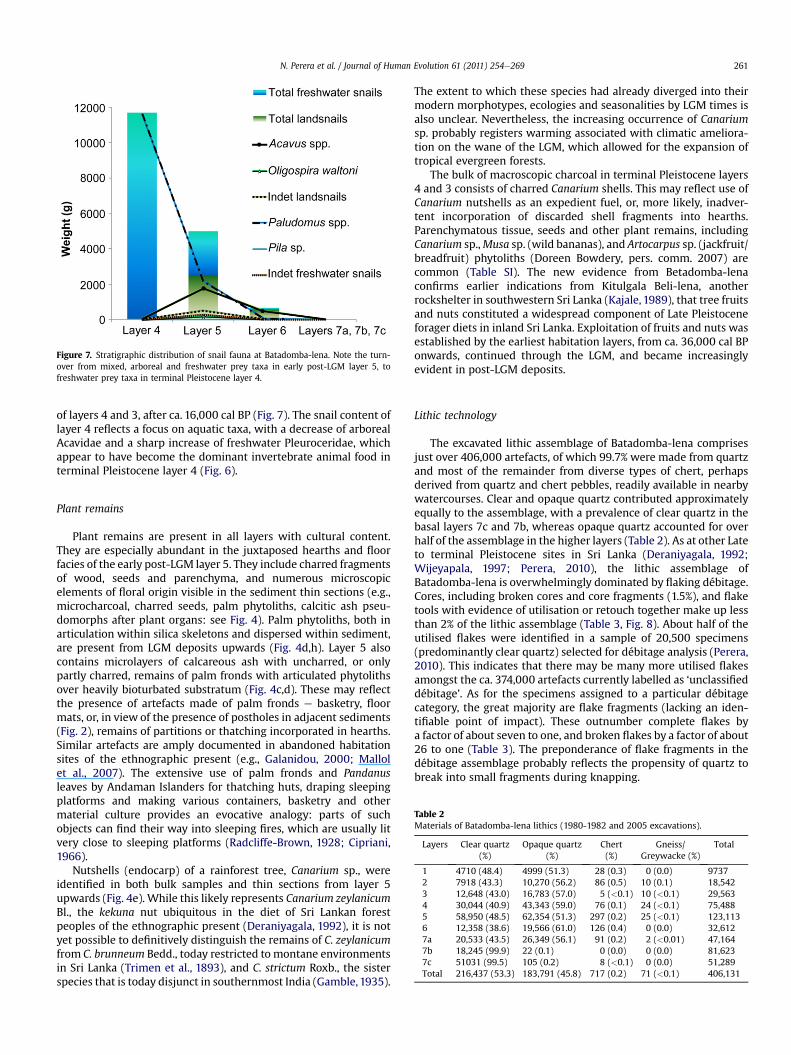
Figure 7. Stratigraphic distribution of snail fauna at Batadomba-lena. Note the turn-over from mixed, arboreal and freshwater prey taxa in early post-LGM layer 5, tofreshwater prey taxa in terminal Pleistocene layer 4.
N. Perera et al. / Journal of Human Evolution 61 (2011) 254e269 261
of layers 4 and 3, after ca. 16,000 cal BP (Fig. 7). The snail content oflayer 4 reflects a focus on aquatic taxa, with a decrease of arborealAcavidae and a sharp increase of freshwater Pleuroceridae, whichappear to have become the dominant invertebrate animal food interminal Pleistocene layer 4 (Fig. 6).
Table 2Materials of Batadomba-lena lithics (1980-1982 and 2005 excavations).
Layers Clear quartz(%)
Opaque quartz(%)
Chert(%)
Gneiss/Greywacke (%)
Total
1 4710 (48.4) 4999 (51.3) 28 (0.3) 0 (0.0) 97372 7918 (43.3) 10,270 (56.2) 86 (0.5) 10 (0.1) 18,5423 12,648 (43.0) 16,783 (57.0) 5 (<0.1) 10 (<0.1) 29,5634 30,044 (40.9) 43,343 (59.0) 76 (0.1) 24 (<0.1) 75,4885 58,950 (48.5) 62,354 (51.3) 297 (0.2) 25 (<0.1) 123,1136 12,358 (38.6) 19,566 (61.0) 126 (0.4) 0 (0.0) 32,6127a 20,533 (43.5) 26,349 (56.1) 91 (0.2) 2 (<0.01) 47,1647b 18,245 (99.9) 22 (0.1) 0 (0.0) 0 (0.0) 81,6237c 51031 (99.5) 105 (0.2) 8 (<0.1) 0 (0.0) 51,289Total 216,437 (53.3) 183,791 (45.8) 717 (0.2) 71 (<0.1) 406,131
Plant remains
Plant remains are present in all layers with cultural content.They are especially abundant in the juxtaposed hearths and floorfacies of the early post-LGM layer 5. They include charred fragmentsof wood, seeds and parenchyma, and numerous microscopicelements of floral origin visible in the sediment thin sections (e.g.,microcharcoal, charred seeds, palm phytoliths, calcitic ash pseu-domorphs after plant organs: see Fig. 4). Palm phytoliths, both inarticulation within silica skeletons and dispersed within sediment,are present from LGM deposits upwards (Fig. 4d,h). Layer 5 alsocontains microlayers of calcareous ash with uncharred, or onlypartly charred, remains of palm fronds with articulated phytolithsover heavily bioturbated substratum (Fig. 4c,d). These may reflectthe presence of artefacts made of palm fronds e basketry, floormats, or, in view of the presence of postholes in adjacent sediments(Fig. 2), remains of partitions or thatching incorporated in hearths.Similar artefacts are amply documented in abandoned habitationsites of the ethnographic present (e.g., Galanidou, 2000; Mallolet al., 2007). The extensive use of palm fronds and Pandanusleaves by Andaman Islanders for thatching huts, draping sleepingplatforms and making various containers, basketry and othermaterial culture provides an evocative analogy: parts of suchobjects can find their way into sleeping fires, which are usually litvery close to sleeping platforms (Radcliffe-Brown, 1928; Cipriani,1966).
Nutshells (endocarp) of a rainforest tree, Canarium sp., wereidentified in both bulk samples and thin sections from layer 5upwards (Fig. 4e). While this likely represents Canarium zeylanicumBl., the kekuna nut ubiquitous in the diet of Sri Lankan forestpeoples of the ethnographic present (Deraniyagala, 1992), it is notyet possible to definitively distinguish the remains of C. zeylanicumfrom C. brunneum Bedd., today restricted to montane environmentsin Sri Lanka (Trimen et al., 1893), and C. strictum Roxb., the sisterspecies that is today disjunct in southernmost India (Gamble,1935).
The extent to which these species had already diverged into theirmodern morphotypes, ecologies and seasonalities by LGM times isalso unclear. Nevertheless, the increasing occurrence of Canariumsp. probably registers warming associated with climatic ameliora-tion on the wane of the LGM, which allowed for the expansion oftropical evergreen forests.
The bulk of macroscopic charcoal in terminal Pleistocene layers4 and 3 consists of charred Canarium shells. This may reflect use ofCanarium nutshells as an expedient fuel, or, more likely, inadver-tent incorporation of discarded shell fragments into hearths.Parenchymatous tissue, seeds and other plant remains, includingCanarium sp.,Musa sp. (wild bananas), and Artocarpus sp. (jackfruit/breadfruit) phytoliths (Doreen Bowdery, pers. comm. 2007) arecommon (Table SI). The new evidence from Betadomba-lenaconfirms earlier indications from Kitulgala Beli-lena, anotherrockshelter in southwestern Sri Lanka (Kajale, 1989), that tree fruitsand nuts constituted a widespread component of Late Pleistoceneforager diets in inland Sri Lanka. Exploitation of fruits and nuts wasestablished by the earliest habitation layers, from ca. 36,000 cal BPonwards, continued through the LGM, and became increasinglyevident in post-LGM deposits.
Lithic technology
The excavated lithic assemblage of Batadomba-lena comprisesjust over 406,000 artefacts, of which 99.7% were made from quartzand most of the remainder from diverse types of chert, perhapsderived from quartz and chert pebbles, readily available in nearbywatercourses. Clear and opaque quartz contributed approximatelyequally to the assemblage, with a prevalence of clear quartz in thebasal layers 7c and 7b, whereas opaque quartz accounted for overhalf of the assemblage in the higher layers (Table 2). As at other Lateto terminal Pleistocene sites in Sri Lanka (Deraniyagala, 1992;Wijeyapala, 1997; Perera, 2010), the lithic assemblage ofBatadomba-lena is overwhelmingly dominated by flaking débitage.Cores, including broken cores and core fragments (1.5%), and flaketools with evidence of utilisation or retouch together make up lessthan 2% of the lithic assemblage (Table 3, Fig. 8). About half of theutilised flakes were identified in a sample of 20,500 specimens(predominantly clear quartz) selected for débitage analysis (Perera,2010). This indicates that there may be many more utilised flakesamongst the ca. 374,000 artefacts currently labelled as ‘unclassifieddébitage’. As for the specimens assigned to a particular débitagecategory, the great majority are flake fragments (lacking an iden-tifiable point of impact). These outnumber complete flakes bya factor of about seven to one, and broken flakes by a factor of about26 to one (Table 3). The preponderance of flake fragments in thedébitage assemblage probably reflects the propensity of quartz tobreak into small fragments during knapping.
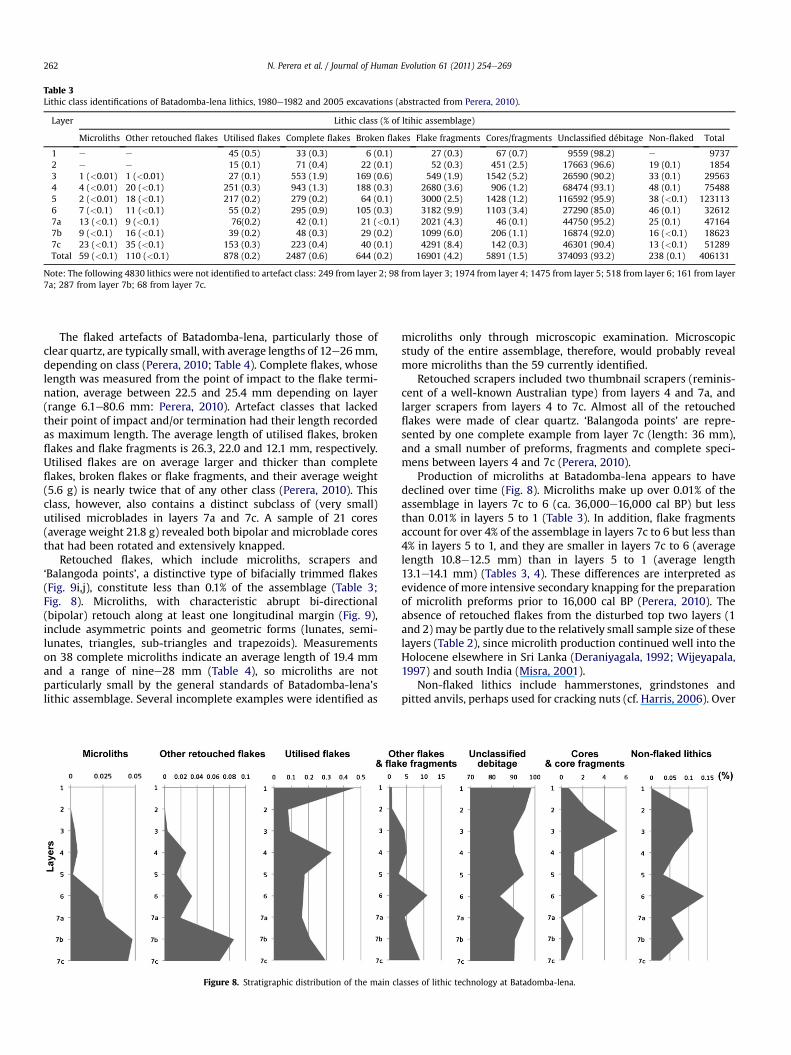
Table 3Lithic class identifications of Batadomba-lena lithics, 1980e1982 and 2005 excavations (abstracted from Perera, 2010).
Layer Lithic class (% of ltihic assemblage)
Microliths Other retouched flakes Utilised flakes Complete flakes Broken flakes Flake fragments Cores/fragments Unclassified débitage Non-flaked Total
1 e e 45 (0.5) 33 (0.3) 6 (0.1) 27 (0.3) 67 (0.7) 9559 (98.2) e 97372 e e 15 (0.1) 71 (0.4) 22 (0.1) 52 (0.3) 451 (2.5) 17663 (96.6) 19 (0.1) 18543 1 (<0.01) 1 (<0.01) 27 (0.1) 553 (1.9) 169 (0.6) 549 (1.9) 1542 (5.2) 26590 (90.2) 33 (0.1) 295634 4 (<0.01) 20 (<0.1) 251 (0.3) 943 (1.3) 188 (0.3) 2680 (3.6) 906 (1.2) 68474 (93.1) 48 (0.1) 754885 2 (<0.01) 18 (<0.1) 217 (0.2) 279 (0.2) 64 (0.1) 3000 (2.5) 1428 (1.2) 116592 (95.9) 38 (<0.1) 1231136 7 (<0.1) 11 (<0.1) 55 (0.2) 295 (0.9) 105 (0.3) 3182 (9.9) 1103 (3.4) 27290 (85.0) 46 (0.1) 326127a 13 (<0.1) 9 (<0.1) 76(0.2) 42 (0.1) 21 (<0.1) 2021 (4.3) 46 (0.1) 44750 (95.2) 25 (0.1) 471647b 9 (<0.1) 16 (<0.1) 39 (0.2) 48 (0.3) 29 (0.2) 1099 (6.0) 206 (1.1) 16874 (92.0) 16 (<0.1) 186237c 23 (<0.1) 35 (<0.1) 153 (0.3) 223 (0.4) 40 (0.1) 4291 (8.4) 142 (0.3) 46301 (90.4) 13 (<0.1) 51289Total 59 (<0.1) 110 (<0.1) 878 (0.2) 2487 (0.6) 644 (0.2) 16901 (4.2) 5891 (1.5) 374093 (93.2) 238 (0.1) 406131
Note: The following 4830 lithics were not identified to artefact class: 249 from layer 2; 98 from layer 3; 1974 from layer 4; 1475 from layer 5; 518 from layer 6; 161 from layer7a; 287 from layer 7b; 68 from layer 7c.
N. Perera et al. / Journal of Human Evolution 61 (2011) 254e269262
The flaked artefacts of Batadomba-lena, particularly those ofclear quartz, are typically small, with average lengths of 12e26mm,depending on class (Perera, 2010; Table 4). Complete flakes, whoselength was measured from the point of impact to the flake termi-nation, average between 22.5 and 25.4 mm depending on layer(range 6.1e80.6 mm: Perera, 2010). Artefact classes that lackedtheir point of impact and/or termination had their length recordedas maximum length. The average length of utilised flakes, brokenflakes and flake fragments is 26.3, 22.0 and 12.1 mm, respectively.Utilised flakes are on average larger and thicker than completeflakes, broken flakes or flake fragments, and their average weight(5.6 g) is nearly twice that of any other class (Perera, 2010). Thisclass, however, also contains a distinct subclass of (very small)utilised microblades in layers 7a and 7c. A sample of 21 cores(average weight 21.8 g) revealed both bipolar and microblade coresthat had been rotated and extensively knapped.
Retouched flakes, which include microliths, scrapers and‘Balangoda points’, a distinctive type of bifacially trimmed flakes(Fig. 9i,j), constitute less than 0.1% of the assemblage (Table 3;Fig. 8). Microliths, with characteristic abrupt bi-directional(bipolar) retouch along at least one longitudinal margin (Fig. 9),include asymmetric points and geometric forms (lunates, semi-lunates, triangles, sub-triangles and trapezoids). Measurementson 38 complete microliths indicate an average length of 19.4 mmand a range of ninee28 mm (Table 4), so microliths are notparticularly small by the general standards of Batadomba-lena’slithic assemblage. Several incomplete examples were identified as
Figure 8. Stratigraphic distribution of the main cl
microliths only through microscopic examination. Microscopicstudy of the entire assemblage, therefore, would probably revealmore microliths than the 59 currently identified.
Retouched scrapers included two thumbnail scrapers (reminis-cent of a well-known Australian type) from layers 4 and 7a, andlarger scrapers from layers 4 to 7c. Almost all of the retouchedflakes were made of clear quartz. ‘Balangoda points’ are repre-sented by one complete example from layer 7c (length: 36 mm),and a small number of preforms, fragments and complete speci-mens between layers 4 and 7c (Perera, 2010).
Production of microliths at Batadomba-lena appears to havedeclined over time (Fig. 8). Microliths make up over 0.01% of theassemblage in layers 7c to 6 (ca. 36,000e16,000 cal BP) but lessthan 0.01% in layers 5 to 1 (Table 3). In addition, flake fragmentsaccount for over 4% of the assemblage in layers 7c to 6 but less than4% in layers 5 to 1, and they are smaller in layers 7c to 6 (averagelength 10.8e12.5 mm) than in layers 5 to 1 (average length13.1e14.1 mm) (Tables 3, 4). These differences are interpreted asevidence of more intensive secondary knapping for the preparationof microlith preforms prior to 16,000 cal BP (Perera, 2010). Theabsence of retouched flakes from the disturbed top two layers (1and 2) may be partly due to the relatively small sample size of theselayers (Table 2), since microlith production continued well into theHolocene elsewhere in Sri Lanka (Deraniyagala, 1992; Wijeyapala,1997) and south India (Misra, 2001).
Non-flaked lithics include hammerstones, grindstones andpitted anvils, perhaps used for cracking nuts (cf. Harris, 2006). Over
asses of lithic technology at Batadomba-lena.
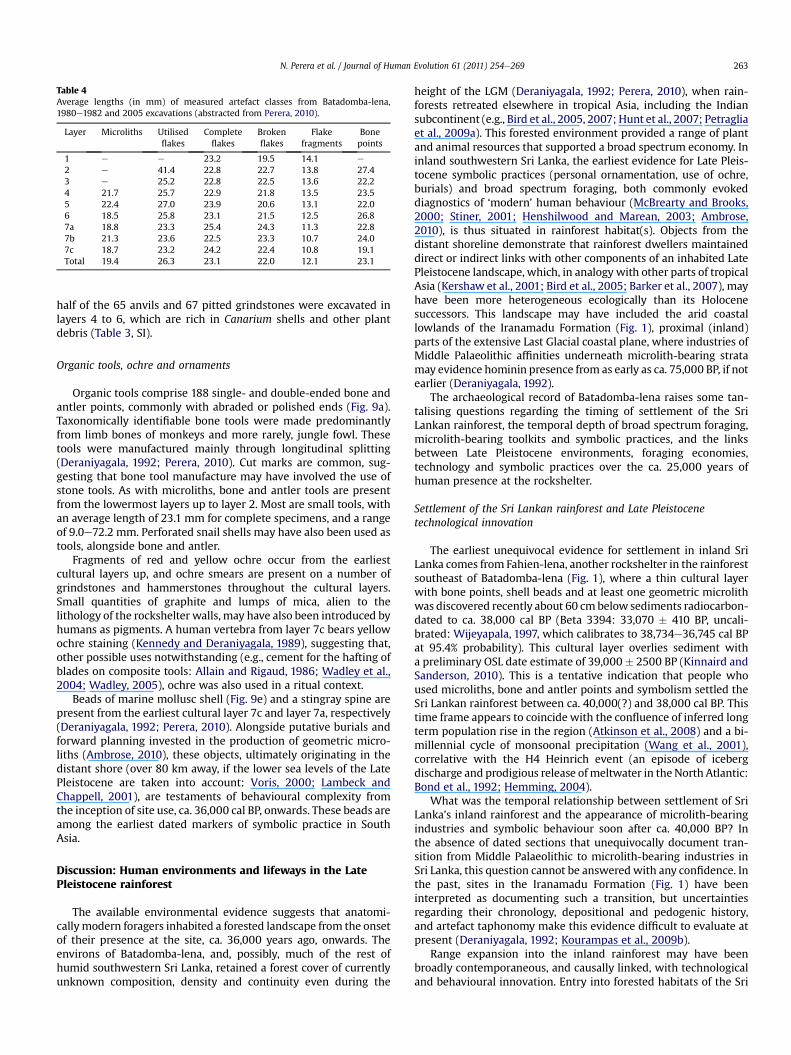
Table 4Average lengths (in mm) of measured artefact classes from Batadomba-lena,1980e1982 and 2005 excavations (abstracted from Perera, 2010).
Layer Microliths Utilisedflakes
Completeflakes
Brokenflakes
Flakefragments
Bonepoints
1 e e 23.2 19.5 14.1 e
2 e 41.4 22.8 22.7 13.8 27.43 e 25.2 22.8 22.5 13.6 22.24 21.7 25.7 22.9 21.8 13.5 23.55 22.4 27.0 23.9 20.6 13.1 22.06 18.5 25.8 23.1 21.5 12.5 26.87a 18.8 23.3 25.4 24.3 11.3 22.87b 21.3 23.6 22.5 23.3 10.7 24.07c 18.7 23.2 24.2 22.4 10.8 19.1Total 19.4 26.3 23.1 22.0 12.1 23.1
N. Perera et al. / Journal of Human Evolution 61 (2011) 254e269 263
half of the 65 anvils and 67 pitted grindstones were excavated inlayers 4 to 6, which are rich in Canarium shells and other plantdebris (Table 3, SI).
Organic tools, ochre and ornaments
Organic tools comprise 188 single- and double-ended bone andantler points, commonly with abraded or polished ends (Fig. 9a).Taxonomically identifiable bone tools were made predominantlyfrom limb bones of monkeys and more rarely, jungle fowl. Thesetools were manufactured mainly through longitudinal splitting(Deraniyagala, 1992; Perera, 2010). Cut marks are common, sug-gesting that bone tool manufacture may have involved the use ofstone tools. As with microliths, bone and antler tools are presentfrom the lowermost layers up to layer 2. Most are small tools, withan average length of 23.1 mm for complete specimens, and a rangeof 9.0e72.2 mm. Perforated snail shells may have also been used astools, alongside bone and antler.
Fragments of red and yellow ochre occur from the earliestcultural layers up, and ochre smears are present on a number ofgrindstones and hammerstones throughout the cultural layers.Small quantities of graphite and lumps of mica, alien to thelithology of the rockshelter walls, may have also been introduced byhumans as pigments. A human vertebra from layer 7c bears yellowochre staining (Kennedy and Deraniyagala, 1989), suggesting that,other possible uses notwithstanding (e.g., cement for the hafting ofblades on composite tools: Allain and Rigaud, 1986; Wadley et al.,2004; Wadley, 2005), ochre was also used in a ritual context.
Beads of marine mollusc shell (Fig. 9e) and a stingray spine arepresent from the earliest cultural layer 7c and layer 7a, respectively(Deraniyagala, 1992; Perera, 2010). Alongside putative burials andforward planning invested in the production of geometric micro-liths (Ambrose, 2010), these objects, ultimately originating in thedistant shore (over 80 km away, if the lower sea levels of the LatePleistocene are taken into account: Voris, 2000; Lambeck andChappell, 2001), are testaments of behavioural complexity fromthe inception of site use, ca. 36,000 cal BP, onwards. These beads areamong the earliest dated markers of symbolic practice in SouthAsia.
Discussion: Human environments and lifeways in the LatePleistocene rainforest
The available environmental evidence suggests that anatomi-callymodern foragers inhabited a forested landscape from the onsetof their presence at the site, ca. 36,000 years ago, onwards. Theenvirons of Batadomba-lena, and, possibly, much of the rest ofhumid southwestern Sri Lanka, retained a forest cover of currentlyunknown composition, density and continuity even during the
height of the LGM (Deraniyagala, 1992; Perera, 2010), when rain-forests retreated elsewhere in tropical Asia, including the Indiansubcontinent (e.g., Bird et al., 2005, 2007; Hunt et al., 2007; Petragliaet al., 2009a). This forested environment provided a range of plantand animal resources that supported a broad spectrum economy. Ininland southwestern Sri Lanka, the earliest evidence for Late Pleis-tocene symbolic practices (personal ornamentation, use of ochre,burials) and broad spectrum foraging, both commonly evokeddiagnostics of ‘modern’ human behaviour (McBrearty and Brooks,2000; Stiner, 2001; Henshilwood and Marean, 2003; Ambrose,2010), is thus situated in rainforest habitat(s). Objects from thedistant shoreline demonstrate that rainforest dwellers maintaineddirect or indirect links with other components of an inhabited LatePleistocene landscape, which, in analogy with other parts of tropicalAsia (Kershaw et al., 2001; Bird et al., 2005; Barker et al., 2007), mayhave been more heterogeneous ecologically than its Holocenesuccessors. This landscape may have included the arid coastallowlands of the Iranamadu Formation (Fig. 1), proximal (inland)parts of the extensive Last Glacial coastal plane, where industries ofMiddle Palaeolithic affinities underneath microlith-bearing stratamay evidence hominin presence from as early as ca. 75,000 BP, if notearlier (Deraniyagala, 1992).
The archaeological record of Batadomba-lena raises some tan-talising questions regarding the timing of settlement of the SriLankan rainforest, the temporal depth of broad spectrum foraging,microlith-bearing toolkits and symbolic practices, and the linksbetween Late Pleistocene environments, foraging economies,technology and symbolic practices over the ca. 25,000 years ofhuman presence at the rockshelter.
Settlement of the Sri Lankan rainforest and Late Pleistocenetechnological innovation
The earliest unequivocal evidence for settlement in inland SriLanka comes from Fahien-lena, another rockshelter in the rainforestsoutheast of Batadomba-lena (Fig. 1), where a thin cultural layerwith bone points, shell beads and at least one geometric microlithwas discovered recently about 60 cm below sediments radiocarbon-dated to ca. 38,000 cal BP (Beta 3394: 33,070 � 410 BP, uncali-brated: Wijeyapala, 1997, which calibrates to 38,734e36,745 cal BPat 95.4% probability). This cultural layer overlies sediment witha preliminary OSL date estimate of 39,000 � 2500 BP (Kinnaird andSanderson, 2010). This is a tentative indication that people whoused microliths, bone and antler points and symbolism settled theSri Lankan rainforest between ca. 40,000(?) and 38,000 cal BP. Thistime frame appears to coincide with the confluence of inferred longterm population rise in the region (Atkinson et al., 2008) and a bi-millennial cycle of monsoonal precipitation (Wang et al., 2001),correlative with the H4 Heinrich event (an episode of icebergdischarge and prodigious release ofmeltwater in the North Atlantic:Bond et al., 1992; Hemming, 2004).
What was the temporal relationship between settlement of SriLanka’s inland rainforest and the appearance of microlith-bearingindustries and symbolic behaviour soon after ca. 40,000 BP? Inthe absence of dated sections that unequivocally document tran-sition from Middle Palaeolithic to microlith-bearing industries inSri Lanka, this question cannot be answered with any confidence. Inthe past, sites in the Iranamadu Formation (Fig. 1) have beeninterpreted as documenting such a transition, but uncertaintiesregarding their chronology, depositional and pedogenic history,and artefact taphonomy make this evidence difficult to evaluate atpresent (Deraniyagala, 1992; Kourampas et al., 2009b).
Range expansion into the inland rainforest may have beenbroadly contemporaneous, and causally linked, with technologicaland behavioural innovation. Entry into forested habitats of the Sri
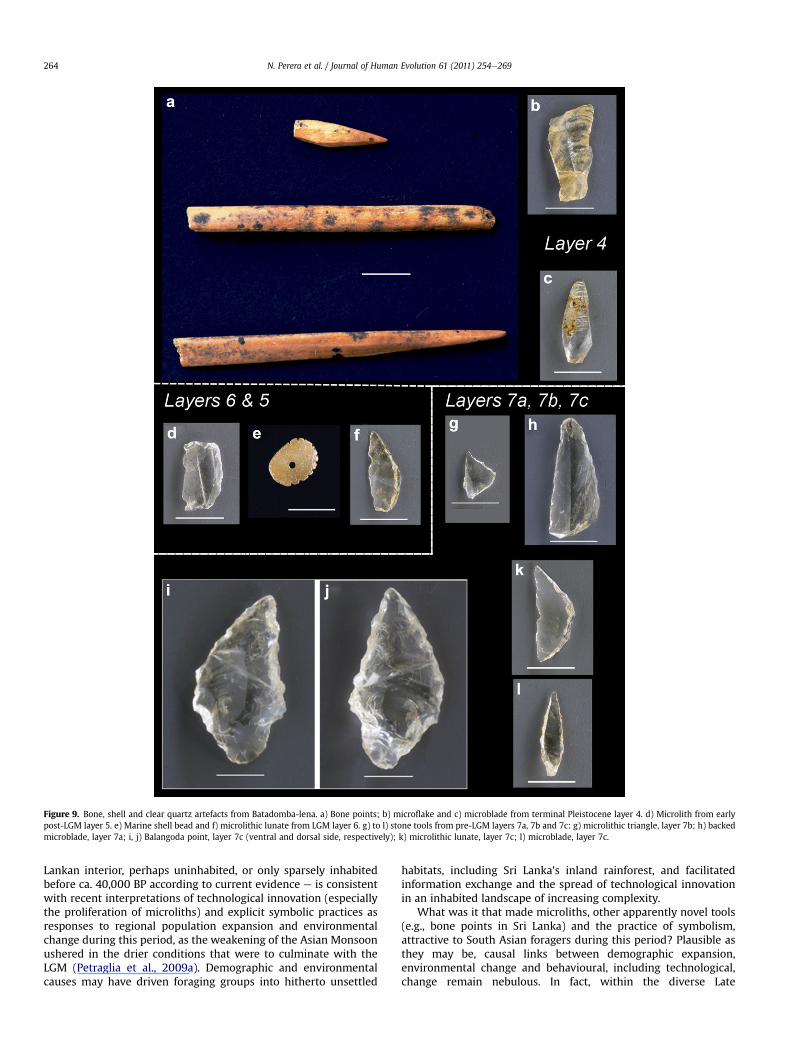
Figure 9. Bone, shell and clear quartz artefacts from Batadomba-lena. a) Bone points; b) microflake and c) microblade from terminal Pleistocene layer 4. d) Microlith from earlypost-LGM layer 5. e) Marine shell bead and f) microlithic lunate from LGM layer 6. g) to l) stone tools from pre-LGM layers 7a, 7b and 7c: g) microlithic triangle, layer 7b; h) backedmicroblade, layer 7a; i, j) Balangoda point, layer 7c (ventral and dorsal side, respectively); k) microlithic lunate, layer 7c; l) microblade, layer 7c.
N. Perera et al. / Journal of Human Evolution 61 (2011) 254e269264
Lankan interior, perhaps uninhabited, or only sparsely inhabitedbefore ca. 40,000 BP according to current evidence e is consistentwith recent interpretations of technological innovation (especiallythe proliferation of microliths) and explicit symbolic practices asresponses to regional population expansion and environmentalchange during this period, as the weakening of the Asian Monsoonushered in the drier conditions that were to culminate with theLGM (Petraglia et al., 2009a). Demographic and environmentalcauses may have driven foraging groups into hitherto unsettled
habitats, including Sri Lanka’s inland rainforest, and facilitatedinformation exchange and the spread of technological innovationin an inhabited landscape of increasing complexity.
What was it that made microliths, other apparently novel tools(e.g., bone points in Sri Lanka) and the practice of symbolism,attractive to South Asian foragers during this period? Plausible asthey may be, causal links between demographic expansion,environmental change and behavioural, including technological,change remain nebulous. In fact, within the diverse Late

N. Perera et al. / Journal of Human Evolution 61 (2011) 254e269 265
Palaeolithic industries of the Indian subcontinent, blades formedonly aminor component well into the end of the Pleistocene (Misra,2001; James and Petraglia, 2005; James, 2007), while the LatePleistocene rainforest of Southeast Asia was settled by people whomade no use of microliths and (recognisable) symbolic objects(Bellwood, 1992, 1997). In other words, small, finely worked toolsand explicitly symbolic material culture do not appear to have beensine qua non in the behavioural repertoire of human societies inLate Pleistocene South and Southeast Asia, perhaps the mostdensely populated part of the world between ca. 45,000 and20,000 BP (Atkinson et al., 2008).
Continuous reliance on less formalised lithic toolkits is alsoevident at Sri Lankan rainforest sites, where microliths constituteonly aminuscule portion of the lithic assemblage (as exemplified byBatadomba-lena). The forested environment of the Sri Lankan sitesand hints for plant-based material culture (e.g., possible palm-derived structures at Batadomba-lena) may indicate that flakes,other lithics and bone tools complemented extensive, but perish-able, plant-derived toolkits (similar to what has been proposed forLate Pleistocene-early Holocene rainforest foragers in SoutheastAsia: cf. Hutterer, 1988).
The available evidence does not permit us to infer whether SriLanka’s microliths and bone points represented technologicaladaptations to the specific conditions of the rainforest (e.g., tips andarmature of projectiles for the hunting of arboreal fauna), orwhether they were introduced to the rainforest from otherecological settings. Suggested pre-40,000 BP sites on the coastallowlands of Sri Lanka are exclusively open-air, situated in the acidicstone lines and sands of the Iranamadu Formation (Deraniyagala,1992). These burial environments are not conducive to the pres-ervation of bone, shell and other organics. Notwithstanding theirarchaeological invisibilibity, therefore, it is possible that aspects ofthe later record, such as organic material culture and symbolicpractices, were deeply rooted in other, longer-inhabited ecozonesof Sri Lanka, from which they were introduced by foragersexpanding into the interior at around 40,000 BP.
Whatever their origins, the functions of microliths in prehistoricSri Lanka, probably hafted on multicomponent tools, as demon-strated for their mainland Indian near-equivalents (Clarkson et al.,2009) and for later, Holocene microliths in Australia (spear barbs:Kamminga, 1980; Attenbrow et al., 2009), remain unknown. Theirminuscule contribution to the lithic assemblage of Batadomba-lenamay indicate specialised uses, or only minimal discard at the site.Integrated use wear and residue analysis in the future may providesome answers. The larger number of microliths in the earlier layers,which reflect less intensive use of the rockshelter (Table 3), mayindicate that microliths featured more prominently in the toolkitsof highly mobile foragers who made shorter-lived visits to therockshelter in the ca. 16 millennia preceding the LGM.
Broad spectrum foraging in the Late Pleistocene rainforest
At Batadomba-lena, as elsewhere in southwestern Sri Lanka (e.g.Kitulgala Beli-lena and Fahien-lena: see faunal evidence inDeraniyagala, 1992; Wijeyapala, 1997; Perera, 2010), microlith- andsymbol-bearing material culture was produced by people whoforaged for a broad spectrum of resources, derived mainly fromevergreen forest traversed by streams. This foraging economy wasbased on a wide variety of mainly small and medium-bodied(�20 kg), agile, largely arboreal animals (monkeys, squirrels andnumerous other mammals), easily collectible land and freshwatersnails and an as yet insufficiently documented assortment of plantfoods, which included tree fruits and nuts. Hunting methodsprobably included the use of projectiles, as evidenced by theabundance of bone points throughout the stratigraphy. Macaque
and langur monkeys, and, perhaps, other arboreal animals, mayhave also been captured by trapping, as observed among contem-porary forest peoples (Hutterer, 1988), and inferred for other LatePleistocene foragers in the Asian rainforests (e.g., the inhabitants ofNiah Cave, Sarawak: Piper et al., 2008; Piper and Rabett, 2009). Thepresence of the predominantly nocturnal civets and mongooses inthe faunal assemblage is a tentative indication for the use of snaresand traps. This hypothesis may be testable through analysis of theage structure of the monkey and other arboreal mammal assem-blage (cf. Piper and Rabett, 2009), but such analysis is not yetavailable. Batadomba-lena also provides some of earliest directevidence for systematic human use of Canarium nuts in Asia, datingfrom ca. 16,000 BP. Canarium became ubiquitous in later terminalPleistocene and Holocene forest-based foraging economies ofSoutheast Asia (e.g., Paz, 2005; Viet, 2008).
This broad spectrum economy was practiced from the outset ofthe rockshelter use from ca. 36,000 cal BP onwards, through to theterminal Pleistocene (and, probably, beyond, as evidenced fromelsewhere in Sri Lanka). Differences in ecological setting, dietcomposition and material culture notwithstanding, Batadomba-lena, thus, adds to the rising number of Late Pleistocene sitesacross the H. sapiens range that demonstrate broad spectrumforaging well before the LGM, (e.g., Munro and Atici, 2009).Comparison of Batadomba-lena with contemporary records illus-trates the diversity and ecological contingency of Late Pleistoceneeconomic practices.
In the circum-Mediterranean, where Pleistocene human ecologyhas been researched more thoroughly than in other bioclimaticregions, broad spectrum foraging is best documented in the animalcomponents of the resource base, especially in the increasedcontribution of agile and elusive small game (e.g., hares, birds) inUpper Palaeolithic, Epi-Palaeolithic and Mesolithic diets (Stiner,2001, 2002, 2004; Stiner et al., 2008). The onset of this broadspectrum trend is viewed as largely coincident with the Middle toUpper Palaeolithic boundary, also marked with technologicaldiversification and evidence for symbolic practices (the ‘UpperPalaeolithic behavioural package’, often taken to reflect a behav-ioural range akin to that of ‘modern’ foragers in similar ecosys-tems). An east-west temporal gradient of dietary diversificationthus interpreted as reflecting the dispersal of H. sapiens foragersacross southern Europe (Stiner, 2001, 2002, 2004). Increased focuson small game of lower preference ranking (due to its low dietaryreturns for a relatively high cost of capture) is interpreted asa response to demographic pressures (Stiner, 2004; Stiner et al.,2008). As increasing populations of human foragers overtaxed thecarrying capacity of their ecosystems, broadening of the dietaryspectrum and technological and social/behavioural innovationwere efforts to buffer or redistribute environmental risk. Sessileand slowmoving small prey (e.g., molluscs, tortoises), however, didplay an important role in Middle as well as in Upper and Epi-Paleolithic circum-Mediterranean diets (Stiner, 2001, 2002) beforeacquiring a marked prominence in many later Mesolithic econo-mies (Lubell, 2004). The vegetable component of broad spectrumdiets in the Late Pleistocene circum-Mediterranean is documentedrather more imperfectly. Direct, albeit sparse, evidence forsystematic use of diverse wild plant foods, including cereals,extends as far back as ca. 23,000 cal BP (Ohallo II Cave in Israel:Weiss et al., 2004). Many Levantine sites register a proliferation ofgrindstones and other plant processing tools immediately after theend of the LGM, indicating intensified harvesting of plant resources(Wright, 1994).
A distinctive feature of Batadomba-lena is that small tomedium-bodied (<20 kg) agile prey species (monkeys, squirrels)formed the cornerstone of its occupants’ diets from the outset, andplayed a significant role in technologies of food procurement (e.g.,

N. Perera et al. / Journal of Human Evolution 61 (2011) 254e269266
bone tips) throughout the occupation of the site. Larger-bodied,ground-living prey animals appear to have made only a minorcontribution to the diet of Batadomba-lena inhabitants. Thiscontrasts strongly with the circum-Mediterranean record, where,increased exploitation of small prey notwithstanding, largerungulates remained central to most Upper Palaeolithic economies(see overview by Munro and Atici, 2009). This disparity does notcome as a surprise, given the marked differences in habitats, foodaffordances and nature of non-human (e.g., climatic) stressesbetween the circum-Mediterranean and tropical South Asia.
What renders Batadomba-lena’s idiosyncrasy particularlyevident is a comparison with the Great Cave of Niah, Sarawak,a complex of Late Pleitsocene sites also situated in Dipterocarpus-dominated lowland rainforest (Barker et al., 2007; Piper et al.,2008; Piper and Rabett, 2009). At Niah’s largest site, the WestMouth, medium to large-bodied ground-living animals, such aspigs, other terrestrial ungulates and monitor lizards, dominated anotherwise diverse diet. The total arboreal and mixed terrestrial/arboreal fauna makes up less than 33% of the assemblage (datafrom Piper et al., 2008), as compared with over 82% of the totalvertebrate assemblage at Batadomba-lena (Figs. 5 and 6; Table SII).Monkeys and squirrels, Batadomba-lena’s vertebrate staples,together make up less than 17% of the recovered faunal assemblage(data from Barker et al., 2007). Even at Lobang Hangus, Niah’s sitewith the highest presence of (mainly) arboreal mammals, such asmonkeys and civets (ca. 56% of the total vertebrate assemblage:Piper et al., 2008; Piper and Rabett, 2009), their representation ismarkedly lower than that at Batadomba-lena.
Comparisonwith the Jwalapuram 9 rockshelter, a microlith-richsite in south India broadly contemporary with Batadomba-lena,further demonstrates the diversity of broad spectrum foragingeconomies in Late Pleistocene South Asia. Jwalapuram 9, asdescribed by Clarkson et al. (2009), registers a very differentsubsistence base, apparently derived from an overall more openlandscape, which ranged from grassland in pre-LGM and LGM timesto openwoodland after ca. 15,000 BP. The occupants of Jwalapuram9 focused their hunting on larger-bodied prey, which mainlycomprised small and medium sized ungulates. Although molluscs(mainly bivalves, but also land snails) were collected systematically,the scarcity of primates and the absence of exclusively arborealanimals contrast this site from Batadomba-lena. In view of theseeconomic contrasts, differences in the relative abundance ofmicroliths between the two sites (microliths are much moreabundant at Jwalapuram 9, making up as much as 12e16% of theassemblage in some layers) may reflect different toolkit adaptationsto local environmental settings and prey ethology. A speculativehypothesis is that multicomponent projectiles (barbed spears orsimilar?) may have had a more prominent role in the open-landscape, larger-prey hunting of the Jwalapuram 9 inhabitants.
How early was broad spectrum foraging in Sri Lanka, and whatwas its temporal relationship with the emergence of diversifiedtoolkits and symbolic practices? Did broad spectrum foraging,microliths and symbolism emerge together, as a South Asianbehavioural equivalent of the ‘Upper Palaeolithic package’, or werethe two records decoupled? These questions are currently unan-swerable, as the subsistence practices of earlier (inferred) MiddlePalaeolithic hominins in Sri Lanka are unknown. Reported MiddlePalaeolithic sites (at Iranamadu Formation) aremade up exclusivelyof lithic assemblages, devoid of any organic remains (Deraniyagala,1992). The composition of earlier Pleistocene faunas (and hence theavailability of potential prey animals) is another unknown. The(rare) reported finds (‘Ratnapura fauna’: Deraniyagala, 1992;Chauhan, 2008) comprise almost exclusively (and perhaps asa result of collection bias) large-bodied mammals (various ungu-lates, elephants, rhinoceroses and carnivores), but these were
derived from undated, and largely undocumented, sedimentarycontexts (possible mass flows on the foothills of eroding slopes:Gunatilaka and Almond, 2001). Until sites with faunal and botanicalpreservation extending to the period before ca. 40,000 cal BP arediscovered, we cannot infer whether broad spectrum foraging(with its technological and behavioural corollaries) was the resultof an abrupt ‘revolution’ (sensu Binford, 1968, and Flannery, 1969),or whether it was assembled over longer time frames, and variousecological zones, in a mosaic process akin to what has beenproposed for the African MSA (McBrearty and Brooks, 2000; Jacobset al., 2008).
Changing Late Pleistocene environments, foraging practices andtoolkits
The ca. 25,000 years of Late Pleistocene human presence inBatadomba-lena bracket episodes of millennial-scale environ-mental change, identified in the pollen and sediment record ofother parts of Sri Lanka (Premathilake and Risberg, 2003;Kourampas et al., 2009a). These events may have affected theenvirons of the rockshelter. Since sediment deposition inBatadomba-lena from ca. 19,000 cal BP to the end of the Pleistocenewas largely human-induced (Table SI), sedimentary indicators ofenvironmental change are not readily identifiable. Remarkably, thetwo millennia following ca. 16,000 cal BP, correlated with abruptintensification of the Asian monsoon (Wang et al., 2001; Thambanet al., 2002) and evidence for landscape reorganisation aroundother rockshelters in southwestern Sri Lanka (e.g. Kitulgala Beli-lena: Kourampas et al., 2009a), were marked by intensification ofhuman occupation in Batadomba-lena and changes in foragingpractices and toolkit. Diversification of the vertebrate prey, withincreased inclusion of other taxa alongside monkeys (Perera, 2010),intensified foraging for snails, Canarium nuts, and possibly otherfruits and tubers, suggest a broadening of the economic spectrumwith the inclusion of a wider array of rainforest resources. Inten-sification of plant resource harvesting at Batadomba-lena on thewake of the LGM presents a remarkable parallel with the circum-Mediterranean of the same period (Wright, 1994).
We do not know whether intensification of occupation on thewane of the LGM (layer 5) reflects more regular use of rockshelterspace in periods of intensifying monsoonal rainfall, increasedpresence of humans in the rainforest (which, by being reshaped byrenewedmonsoonal precipitation, presented hunter-gatherers withnovel and advantageous ecological niches), or a combination of theabove. It is possible that the broadening of human dietary spectrumbetween 16,000 and 14,000 cal BP was a societal response to a localecosystem under stress by intensified resource exploitation andhuman modification of the rainforest, perhaps as a result ofdemographic expansion, and the cascade effects of post-LGMwarming and monsoon intensification. Nonetheless, (apparent)absence of distinctive colluvial fluxes from the sediment record ofthis period, together with the flowing, freshwater conditions ofstreams adjacent to the site (as indicated by Pleuroceridae snails,presumably collected from these streams) suggest that erosion ratesremained moderate in the environs of Batadomba-lena, possiblydue to the retention of forest cover.
Decline of microlith production following the LGM took placeagainst this ecological backdrop. It is unlikely that the decline ofmicroliths signifies reduced use of projectile hunting technology,since bone points are present, and arboreal mammals abundant, interminal Pleistocene layers from which microliths are virtuallyabsent. In view of the faunal evidence from this period, it is possiblethat the post-LGM toolkit reflects reduced mobility of the siteoccupants, with intensified foraging for a broader spectrum ofresources, perhaps in a narrower range around the site. Was this

N. Perera et al. / Journal of Human Evolution 61 (2011) 254e269 267
expansion of the resource base a result of necessity (e.g., the need toderive subsistence from less desirable resources, harvested froma shorter foraging range within a more populous post-LGM land-scape), opportunity (the presence of advantageous habitat mosaics,affording a bounty of easily collectable resources in a period ofenvironmental change), or a combination of the above? Ouremerging working hypothesis is that further broadening of foragingspectrum became feasible in a diversified rainforest ecosystem, withhabitat mosaics fashioned by the combined effects of monsoonintensification and human activity. A plausible alternative scenario,however, is that subsistence practices changed in the face ofmonsoon-driven habitat homogenisation, as habitat mosaics weresuperseded by continuous rainforest. Systematic recovery of faunal,floral and sedimentary proxies from Batadomba-lena and other SriLankan sites, coupled with high-resolution chronologies, has thepotential to refine and ultimately test emerging human-ecologicalhypotheses.
Conclusions
Batadomba-lena preserves evidence for the presence ofH. sapiens foragers in the rainforest of southern Sri Lanka from ca.36,000 cal BP onwards to the end of the Pleistocene. A rich materialculture, marked by microliths, bone points, ornaments, objectsderived from the ca. 80 km-distant shore and possible plant-basedartefacts, is associated with possible human burials, and faunal andfloral remains, providing windows to Late Pleistocene habitats andhuman lifeways.
Batadomba-lena’s Late Pleistocene inhabitants practisedsymbolic behaviour and made a complex toolkit of unretouchedflakes, bone and antler tools, rare blunted-backed microliths and,possibly, plant-based objects and structures from the outset of theirpresence at the site. This toolkit was used to support broad spec-trum foraging, which, although focussing on small to medium sized(adult weight � 20 kg) arboreal mammals (monkeys, squirrels),also included fish, land and freshwater snails and an as yetunidentified range of plant resources. The environs of Batadomba-lena retained forest cover throughout the ca. 25,000 years ofhuman presence recorded at the site, even through periods ofmonsoon weakening and maximum regional aridity during theLGM. The continuity and composition of this rainforest biome,however, remains unknown. Human presence intensified duringthe LGM and especially in its immediate aftermath. Humanforaging practices appear to have changed in this period of inten-sified occupation, with an increased emphasis on small-bodiedprey, especially freshwater snails, and rainforest plant resources,such as Canarium nuts. These shifts may reflect depletion of thesurrounding ecosystem by intensified exploitation by an expandinghuman population, effects of environmental change as a result ofpost-LGM monsoon intensification, or a combination of the above.Production of microliths declined notably from the earliest humanpresence at the site to this period of intensive occupation, invitingthe hypothesis that microliths were mainly associated with highlymobile forager bands that used the rockshelter in earlier times.
The behavioural record of Batadomba-lena is markedly distinctfrom that of the European and Levantine Upper Palaeolithic, whichinform current models of Late Pleistocene ‘cognitive revolution’(Mellars, 1989, 1991; Bar-Yosef, 2002; but see Belfer-Cohen andHovers, 2010 for an alternative reading of the Upper Palaeolithicof the Levant). It is also different from broadly contemporarySoutheast Asian and Australian sites, characterised by the absenceof backed microliths and, in the case of Southeast Asia, lowarchaeological visibility of symbolic practices (Anderson, 1997;Barker et al., 2007; Habgood and Franklin, 2008). Alongsidebroadly contemporary sites in the South Asian mainland (e.g.,
Jwalapuram Locality 9 in Southern India: Clarkson et al., 2009),Batadomba-lena demonstrates that Late Pleistocene pathways totechnological diversification and symbolic practices were diverse,multifaceted and ecologically contingent. Ongoing work aims toplace Sri Lanka’s Late Pleistocene societies in their dynamicecological context, thus contributing to wider debates on thehuman ecology of prehistoric foragers in the tropics, the culturaldiversity of the Late Pleistocene world and the drivers of techno-logical and behavioural change in earlier human societies.
Acknowledgements
We are indebted to the Sri Lanka Department of Archaeology foroutstanding field support, George MacLeod (University of Stirling)for thin-section manufacturing and Doreen Bowdery (AustralianNational University) for making preliminary results of her phytolithanalysis available to us and for commenting on an earlier version ofthis paper. Shanti Pappu (Sharma Centre for Heritage Education),Ana Polo Diaz (University of the Basque Country) and Krista Gilli-land (University of Stirling) provided invaluable insights. Finally,this paper benefited from constructive criticism and suggestions byMary Stiner and two anonymous reviewers. Research funding wasprovided by the Sri Lanka Department of Archaeology, the Austra-lian National University and the British Academy.
Appendix. Supplementary data
Supplementary data related to this article can be found online atdoi:10.1016/j.jhevol.2011.04.001
References
Abeyratne, M., Spooner, N.A., Grün, R., Head, J., 1997. Multidating studies of Bata-domba cave, Sri Lanka. Quatern. Sci. Rev. 16, 243e255.
Achyutan, H., Quade, J., Roe, L., Placzek, C., 2007. Stable isotopic composition ofpedogenic carbonates from the eastern margin of the Thar Desert, Rajasthan,India. Quatern. Int. 162, 50e60.
Agrawal, D.P., Krishnamurthy, R.V., Kusumgar, S., 1985. Physical research laboratoryradiocarbon date list V. Radiocarbon 27, 95e110.
Allain, J., Rigaud, A., 1986. Décor et fonction: quelques exemples tirés du Magda-lénien. L’Anthropologie 90, 713e738.
Ambrose, S.H., 1998. Late Pleistocene human population bottlenecks, volcanicwinter, and differentiation of modern humans. J. Hum. Evol. 34, 623e651.
Ambrose, S.H., 2010. Coeveolution of composite-tool technology, constructivememory, and language: implications for the evolution of modern humanbehaviour. Curr. Anthropol. 51, S135eS147.
Anderson, D.D., 1997. Cave archaeology in southeast Asia. Geoarchaeology 12,607e638.
Angel, J.L., 1972. A Middle Palaeolithic temporal bone from Darra-i-Kur, Afghanistan.Trans. Am. Philos. Soc. 62, 54e56.
Argue, D., Morwood, M.J., Sutikna, T., Jatmiko, Saptomo, E.W., 2009. Homo flor-esiensis: a cladistic analysis. J. Hum. Evol. 57, 623e639.
Atkinson, Q.D., Gray, R.D., Drummond, A.J., 2008. mtDNA variation predicts pop-ulation size in humans and reveals a major Southern Asian chapter in humanprehistory. Mol. Biol. Evol. 25, 468e474.
Attenbrow, V., Robertson, G., Hiscock, P., 2009. The changing abundance of backedartefacts in southeastern Australia: a response to Holocene climate change?J. Archaeol. Sci. 36, 2765e2770.
Balme, J., Davidson, I., McDonald, J., Stern, N., Veth, P., 2008. Symbolic behaviour andthe peopling of the southern arc route to Australia. Quatern. Int. 202, 59e68.
Barker, G., Barton, H., Bird, M., Daly, P., Datan, I., Dykes, A., Farr, L., Gilbertson, D.,Garrisson, B., Hunt, C., Higham, T., Kealhofer, L., Krigbaum, J., Lewis, H.,McLaren, S., Paz, V., Pike, A., Piper, P., Pyatt, B., Rabett, R., Reynolds, T., Rose, J.,Rushworth, G., Stephens, M., Stringer, C., Thompson, J., Turney, C., 2007. The"human revolution" in lowland tropical Southeast Asia: the antiquity andbehavior of anatomically modern humans at Niah Cave (Sarawak, Borneo).J. Hum. Evol. 52, 243e261.
Bar-Yosef, O., 2002. The Upper Paleolithic revolution. Annu. Rev. Anthropol. 31,363e393.
Belfer-Cohen, A., Hovers, E., 2010. Modernity, enhanced working memory, and theMiddle to Upper Paleolithic record of the Levant. Curr. Anthopol. 51, S167eS175.
Bellwood, P., 1992. Southeast Asia before history. In: Tarling, N. (Ed.), The Cam-bridge History of Southeast Asia, vol 1. Cambridge University Press, Cambridge,pp. 55e136.

N. Perera et al. / Journal of Human Evolution 61 (2011) 254e269268
Bellwood, P., 1997. Prehistory of the IndoeMalaysian Archipelago. Revised Edition.University of Hawaii Press, Honolulu.
Binford, R.L., 1968. Post-Pleistocene adaptations. In: Binford, S.R., Binford, L.R. (Eds.),New Perspectives in Archaeology. Aldine, Chicago, pp. 313e341.
Bird, M.I., Boobyer, E.M., Bryant, C., Lewis, H., Paz, V., Stephens, W.E., 2007. A longrecord of environmental change from bat guano deposits in Makangit Cave,Palawan, Philippines. Earth Env. Sci. Trans. R. Soc. Edin. 98, 59e69.
Bird, M.I., Taylor, D., Hunt, C., 2005. Palaeoenvironments of insular Southeast Asiaduring the last glacial period: a savanna corridor in Sundaland? Quatern. Sci.Rev. 24, 2228e2242.
Bond, G., Heinrich, H., Broecker, W., Labeyrie, L., McManus, J., Andrews, J., Huon, S.,Jantschik, R., Clasen, S., Simet, C., Tedesco, K., Klas, M., Bonani, G., Ivy, S., 1992.Evidence for massive discharges of icebergs into the North Atlantic Oceanduring the last glacial period. Nature 360, 245e249.
Bowler, J.M., Johnston, H., Olley, J.M., Prescott, J.R., Roberts, R.G., Shawcross, W.,Spooner, N.A., 2003. New ages for human occupation and climatic change atLake Mungo, Australia. Nature 421, 837e840.
Bronk Ramsey, C., 2009. Bayesian analysis of radiocarbon dates. Radiocarbon 51,337e360.
Bulbeck, D., 2007. Where rivers meet the sea: a parsimonious model for Homosapiens colonisation of the Indian Ocean rim and Sahul. Curr. Anthropol. 48,315e320.
Bulbeck, D., Rayner, D., Groves, C.P., Raghavan, P., 2003. “The contribution of SouthAsia to the Peopling of Australasia" and the relevance of Basel’s NaturhistorischMuseum anthropological collection to the project aims. Bull. Soc. Suisse d’An-thropol. 9, 49e70.
Bullock, P., Fedoroff, N., Jongerius, A., Stoops, G., Tursina, T., Babel, U., 1985. Hand-book for Soil Thin Section Description. Waine Research Publications, Albrington.
Cameron, D.W., Groves, C.P., 2004. Bones, Stones and Molecules. "Out of Africa" andHuman Origins. Elsevier, London.
Chauhan, P.R., 2008. Large mammal fossil occurrences and associated archaeolog-ical evidence in Pleistocene contexts of peninsular India and Sri Lanka. Quatern.Int. 192, 20e42.
Cipriani, L., 1966. The Andaman Islanders. Weidenfield & Nicholson, London.Clarkson, C., Petraglia, M., Korisettar, R., Haslam, M., Boivin, N., Crowther, A.,
Ditchfield, P., Fuller, D., Miracle, P., Harris, C., Connell, K., James, H., Koshy, J.,2009. The oldest and longest enduring microlithic sequence in India: 35,000years of modern human occupation and change at the Jualapuram locality 9rockshelter. Antiquity 83, 326e348.
Cooray, P.G., 1984. The Geology of Sri Lanka. National Museum Publications,Colombo.
Courty, M.A., Goldberg, P., Macphail, R., 1989. Soils and Micromorphology inArchaeology. Cambridge University Press, Cambridge.
Deraniyagala, P.E.P., 1940. The stone age and cave men of Ceylon. Part 1. J. Roy.Asiatic Soc. Ceylon 34, 351e373.
Deraniyagala, P.E.P., 1943. The stone age and cave men of Ceylon. Part 2. J. Roy.Asiatic Soc. Ceylon 35, 159e162.
Deraniyagala, S.U., 1992. The Prehistory of Sri Lanka: An Ecological Perspective,second ed. Department of Archaeological Survey, Colombo.
Dupree, L., Davis, R.S., 1972. The lithic and bone specimens from Aq Kupruk andDara-i-Kur. Trans. Am. Phil. Soc. 62, 14e32.
Endicott, P., Metspalu, M., Kivisild, T., 2007. Genetic evidence on modern humandispersals in South Asia: Y chromosome and mitochondrial DNA perspectives:the world through the eyes of two haploid genomes. In: Petraglia, M.D.,Allchin, B. (Eds.), The Evolution and History of Human Populations in SouthAsia: Interdisciplinary Studies in Archaeology, Biological Anthropology,Linguistics and Genetics. Springer, Dordrecht, pp. 229e244.
Field, J.S., Petraglia, M.D., Lahr, M.M., 2007. The southern dispersal hypothesis andthe South Asian archaeological record: examination of dispersal routes throughGIS analysis. J. Anthropol. Archaeol. 26, 88e108.
Flannery, K.V., 1969. Origins and ecological effects of early domestication in Iran andthe Near East. In: Ucko, P.J., Dimbleby, G.W. (Eds.), The Domestication andExploitation of Plants and Animals. Aldine, Chicago IL, pp. 73e100.
Galanidou, N., 2000. Patterns in caves: foragers, horticulturalists, and the use ofspace. J. Anthropol. Archaeol. 19, 243e275.
Gamble, J.S., 1935. Flora of the Madras Presidency, vol. I. Adlard and Son, London.Gathorne-Hardy, F.J., Syaukani, Davies, R.G., Eggleton, P., Jones, D.T., 2002. Quater-
nary rainforest refugia in SE Asia: using termites (Isoptera) as indicators. Biol. J.Linn. Soc. 75, 453e466.
Gaussen, H., Legris, P., Viart, M., Labroue, L., 1968. Explanatory Notes on the Vege-tation Map of Ceylon. Government Press, Colombo.
Gilligan, I., 2007. Resisting the cold in ice age Tasmania: thermal environment andsettlement strategies. Antiquity 81, 555e568.
Grayson, D.K., 1984. Quantitative Zooarchaeology: Topics in the Analysis ofArchaeological Faunas. Academic Press, London.
Green, R.E., Krause, J., Briggs, A.W., Maricic, T., Stenzel, U., Kircher, M., Patterson, N.,Li, H., Zhai, W., Fritz, MH.-Y., Hansen, N.F., Durand, E.Y., Malaspinas, A.S.,Jensen, J.D., Marques-Bonet, T., Alkan, C., Prüfer, K., Meyer, M., Burbano, H.A.,Good, J.M., Schultz, R., Aximu-Petri, A., Butthof, A., Höber, B., Höffner, B.,Siegemund, M., Weihmann, A., Nusbaum, C., Lander, E.S., Russ, C., Novod, N.,Affourtit, J., Egholm, M., Verna, C., Rudan, P., Brajkovic, D., Kucan, �Z, Gu�sic, I.,Doronichev, V.B., Golovanova, L.V., Lalueza-Fox, C., de la Rasilla, M., Fortea, J.,Rosas, A., Schmitz, R.W., Johnson, P.L.F., Eichler, E.E., Falush, D., Birney, E.,Mullikin, J.C., Slatkin, M., Nielsen, R., Kelso, J., Lachmann, M., Reich, D., Pääbo, S.,2010. A draft sequence of the Neandertal genome. Science 328, 710e722.
Gunatilaka, A., Almond, D.C., 2001. Serendipity and the jewels of Serendib: Part 2.External processes and climatic framework that influenced the genesis of themulti-species gemstone province in Sri Lanka. J. Geol. Soc. Sri Lanka 10, 49e67.
Gunz, P., Bookstein, F.L., Mitteroeker, P., Stadlmayr, A., Seidler, H., Weber, G.W., 2009.Early modern human diversity suggests subdivided population structure anda complex out-of-Africa scenario. Proc. Natl. Acad. Sci. 106, 6094e6098.
Habgood, P.J., Franklin, N.R., 2008. The revolution that didn’t arrive: a review ofPleistocene Sahul. J. Hum. Evol. 55, 187e222.
Harris, D.R., 2006. The interplay of ethnographic and archaeological knowledge in thestudy of past human subsistence in the tropics. J. Roy. Anthropol. Inst., S63eS78.
Harris, E.C., 1989. Principles of Archaeological Stratigraphy. Academic Press, London.Haslam, M., Clarkson, C., Petraglia, M., Korisettar, R., Jones, S., Shipton, C.,
Ditchfield, P., Ambrose, S.H., 2010. The 74 ka Toba super-eruption and southernIndian hominins: archaeology, lithic technology and environments at Jwala-puram Locality 3. J. Archaeol. Sci. 37, 3370e3384.
Hemming, S.R., 2004. Heinrich Events: massive late Pleistocene detritus layers ofthe North Atlantic and their global climate imprint. Rev. Geophys. 42, RG1005.
Henshilwood, C.S., Marean, C.W., 2003. The origin of modern human behavior.Critique of the models and their test implications. Curr. Anthropol. 44, 627e651.
Hodgson, J.A., Bergey, C.M., Disotell, T.R., 2010. Neandertal genome: the ins and outsof African genetic diversity. Curr. Biol. 20, R517eR519.
Holdaway, S., Stern, N., 2004. A Record in Stone: The Study of Australia’s FlakedStone Artefacts. Museum Victoria/Canberra: The Australian Institute ofAboriginal and Torres Strait Islander Studies, Melbourne.
Hughes, P., 1983. Geoarchaeology in Australia. In: Beaton, J., Connah, G. (Eds.),Australian Field Archaeology: A Guide to Techniques. Australian Institute ofAboriginal Studies, Canberra, pp. 109e117.
Hunt, C.O., Gilbertson, D.D., Rushworth, G., 2007. Modern humans in Sarawak,Malaysian Borneo, during oxygen isotope stage 3: palaeonvironmentalevidence from the great cave of Niah. J. Archaeol. Sci. 34, 1953e1969.
Hutterer, K.L., 1988. The prehistory of the Asian rainforests. In: Denslow, J.S.,Padoch, C. (Eds.), People of the Tropical Rainforest. University of CaliforniaPress, Berkley and Loss Angeles, pp. 63e72.
Jacobs, Z., Roberts, R.G., Galbraith, R.F., Deacon, H.J., Grün, R., Mackay, A., Mitchell, P.,Vogelsang, R., Wadley, L., 2008. Ages for the Middle stone age of southernAfrica: implications for human behavior and dispersal. Science 322, 733e735.
James, H.V.A., 2007. The emergence of modern human behaviour in South Asia:a review of the current evidence and discussion of the possible implications. In:Petraglia, M.D., Allchin, B. (Eds.), The Evolution and History of Human Pop-ulations in South Asia: Interdisciplinary Studies in Archaeology, BiologicalAnthropology, Linguistics and Genetics. Springer, Dordrecht, pp. 268e281.
James, H.V.A., Petraglia, M.D., 2005. Modern human origins and the evolution ofbehavior in the Later Pleistocene record of South Asia. Curr. Anthropol. 46, S3eS26.
Kajale, M.D., 1989. Mesolithic exploitation of wild plants in Sri Lanka: archae-obotanical study at the cave of Beli-Lena. In: Harris, D.R., Hillman, G.C. (Eds.),Foraging and Farming. The Evolution of Plant Exploitation. Unwin Hyman,London, pp. 269e281.
Kamminga, J., 1980. A functional investigation of Australian microliths. The Artefact5, 1e18.
Kamminga, J., 1982. Over the Edge: Functional Analysis of Australian Stone Tools.Occasional Papers in Anthropology 12. Queensland University AnthropologyMuseum, Brisbane.
Kennedy, K.A.R., 1999. Paleoanthropology of south Asia. Evol. Anthropol. 8,165e185.
Kennedy, K.A.R., 2000. God-apes and Fossil Men: Paleoanthropology of South Asia.University of Michigan Press, Ann Arbor.
Kennedy, K.A.R., Deraniyagala, S.U., 1989. Fossil remains of 28,000-year-old homi-nids from Sri Lanka. Curr. Anthropol. 30, 394e399.
Kennedy, K.A.R., Deraniyagala, S.U., Roertgen, W.J., Chiment, J., Disotell, T., 1987.Upper Pleistocene fossil hominids from Sri Lanka. Am. J. Phys. Anthropol. 72,441e461.
Kershaw, P.A., Penny, D., van der Kaars, S., Anshari, D., Thamotherampillai, A., 2001.Vegetation and climate in lowland Southeast Asia at the last glacial maximum.In: Metcalfe, I., Smith, J.M.B., Morwood, M., Davidson, I. (Eds.), Faunal and FloralMigrations and Evolution in SE Asia-Australasia. Balkema, Lisse, pp. 227e236.
Kinnaird, T.C., Sanderson, D.C.W., 2010. Luminescence Dating of Sediments from theFahien-lena Rockshelter, Southern Sri Lanka. Scottish Universities Environ-mental Research Centre, East Kilbride.
Korrisetar, R., 2007. Towards developing a basin model for the Paleolithic settle-ment of the Indian Subcontinent: geodynamics, monsoon dynamics anddispersal routes. In: Petraglia, M.D., Allchin, B. (Eds.), The Evolution and Historyof Human Populations in South Asia: Interdisciplinary Studies in Archaeology,Biological Anthropology, Linguistics and Genetics. Springer, Dordrecht, pp.69e96.
Kourampas, N., Simpson, I.A., Perera, N., Deraniyagala, S.U., 2008. Late Pleistocenehunter-gatherers in the South Asian rainforest: geoarchaeology of inhabitedrockshelters in south-western Sri Lanka. Antiquity 82.
Kourampas, N., Simpson, I.A., Perera, N., Deraniyagala, S.U., Wijeyapala, W.H., 2009a.Rockshelter sedimentation in a dynamic tropical landscape: late Pleistocene-eEarly Holocene archaeological deposits in Kutulgala Beli-lena, southwesternSri Lanka. Geoarchaeology. 24, 677e714.
Kourampas, N., Simpson, I.A., Polo Diaz, A., Perera, N., Deraniyagala, S.U., 2009b.Geoarchaeology of Pleistocene Sites in Southern Sri Lanka: Fahien rockshelterand Tool-bearing Sediments of the Iranamadu Formation. Report to the SriLanka Department of Archaeology. University of Stirling, Stirling.

N. Perera et al. / Journal of Human Evolution 61 (2011) 254e269 269
Lahr, M., Foley, R., 1994. Multiple dispersals and modern human origins. Evol.Anthropol. 3, 48e60.
Lahr, M.M., Foley, R.A., 1998. Towards a theory of modern human origins: geog-raphy, demography, and diversity in recent human evolution. Yearb. Phys.Anthropol. 41, 137e176.
Lambeck, K., Chappell, J., 2001. Sea level change through the last glacial cycle.Science 292, 679e686.
Limbrey, S., 1975. Soil Science and Archaeology. Academic Press, London.Liu, W., Jin, C.-J., Zhang, Y.-Q., Cai, Y.-J., Xing, S., Wu, X.-J., Cheng, H., Edwards, R.L.,
Pan, W.-S., Qin, D.-G., An, Z.-S., Trinkaus, E., Wu, X.-Z., 2010. Human remainsfrom Zhirendong, south China, and modern human emergence in East Asia.Proc. Natl. Acad. Sci. 108, 1267e1271.
Lubell, D., 2004. Prehistoric edible land snails in the circum-Mediterranean: thearchaeological evidence. XXIVe Rencontres Internationales d’ Archéologie et d’Histoire d’ Antibes. In: Brugal, J.-P., Desse, J. (Eds.), Petits Animaux et SociétésHumaines. Du Complément Alimentaire des Ressources Utilitaires. APDCA,Antibes, pp. 77e98.
Macauley, V., Hill, C., Achilli, A., Rengo, C., Clarke, D., Meehan, W., Blackburn, J.,Semino, O., Scozzari, R., Cruciani, F., Taha, A., Shaari, N.K., Raja, J.M., Ismail, P.,Zainuddin, Z., Goodwin, W., Bulbeck, D., Bandelt, H.-J., Oppenheimer, S.,Torroni, A., Richards, M., 2005. Single, rapid coastal settlement of Asia revealedby analysis of complete mitochondrial genomes. Science 308, 1034e1036.
Mallol, C., Marlowe, F.W., Wood, B., Porter, C., 2007. Earth, wind, and fire: ethno-archaeological signals of Hadza fires. J. Archaeol. Sci. 34, 2035e2052.
Marean, C.W., Bar-Matthews, M., Bernatchez, J., Fisher, E., Goldberg, P., Herries, A.I.R.,Jacobs, Z., Jerardino, A., Karkanas, P., Minichillo, T., Nilssen, P.J., Thompson, E.,Watts, I.,Williams,H.M., 2007. Early humanuse ofmarine resources andpigment inSouth Africa during the Middle Pleistocene. Nature 449, 905e908.
McBrearty, A., Brooks, A., 2000. The revolution that wasn’t: a new interpretation ofthe origin of modern human behavior. J. Hum. Evol. 39, 453e563.
Mellars, P., 1989. Technological changes at the Middle-Upper Palaeolithic transition:economic, social and cognitive perspectives. In: Mellars, P., Stringer, C. (Eds.),The Human Revolution: Behavioural and Biological Perspectives on the Originsof Modern Humans. Edinburgh University Press, Edinburgh, pp. 338e365.
Mellars, P., 1991. Cognitive changes and the emergence of modern humans inEurope. Cambridge Archaeol. J. 1, 63e76.
Mellars, P., 2006a. Going east: new genetic and archaeological perspectives on themodern human colonization of Eurasia. Science 313, 796e800.
Mellars, P., 2006b. Why did modern human populations disperse from Africa ca.60,000 years ago? A new model. Proc. Natl. Acad. Sci. 103, 9381e9386.
Mijares, A.S., Détroit, F., Piper, P., Grün, R., Bellwood, P., Aubert, M., Champion, G.,Cuevas, N., de Leon, A., Dizon, E., 2010. New evidence for a 67,000-year-oldhuman presence at Callao Cave, Luzon, Philippines. J. Hum. Evol. 59, 123e132.
Misra, V.N., 2001. Prehistoric human colonisation of India. J. Biosci. 26, 491e531.Morwood, M.J., Soejono, R.P., Roberts, R.G., Sutikna, T., Turney, C.S.M.,
Westaway, K.E., Rink, W.J., Zhao, J.-X., van den Bergh, G.D., Awe Due, Rokus,Hobbs, D.R., Moore, M.W., Bird, M.I., Fifield, L.K., 2004. Archaeology and age ofa new hominin from Flores in eastern Indonesia. Nature 431, 1087e1091.
Munro, N.D., Atici, L., 2009. Human subsistence change in the Late PleistoceneMediterranean Basin: the status of research on faunal intensification, diversi-fication & specialization. Before Farming 2009 (1), 1e6.
Naggs, F., Raheem, D., Ranawana, K., Mapatuna, Y., 2005. The Darwin Initiativeproject on Sri Lankan land snails: patterns of diversity in Sri Lankan forests.Raffles B. Zool. Supp. 12, 23e29.
Oppenheimer, S., 2003. Out of Eden: The Peopling of the World. Robinson, London.Oppenheimer, S., 2009. The great arc of dispersal of modern humans: Africa to
Australia. Quatern. Int. 202, 2e13.Paz, V., 2005. Rock shelters, caves, and archaeobotany in Island Southeast Asia.
Asian. Perspect. 44, 107e118.Perera, H.N., 2010. Prehistoric Sri Lanka: Late Pleistocene Rockshelters and an Open
Air Site. BAR International Series. Archaeopress, Oxford.Perera, N.P., 1975. A physiognomic vegetation map of Sri Lanka (Ceylon). J. Biogeogr.
2, 185e203.Petraglia, M., Korisettar, R., Boivin, N., Clarkson, C., Ditchfield, P., Jones, S., Koshy, J.,
Lahr, M.M., Oppenheimer, C., Pyle, D., Roberts, R., Schwenninger, J.-L., Arnold, L.,White, K., 2007. Middle Paleolithic assemblages from the Indian subcontinentbefore and after the Toba super-eruption. Science 317, 114e116.
Petraglia, M., Korisettar, R., Kasturi Bai, M., Boivin, N.B.J., Clarkson, C.,Cunningham, K., Ditchfield, P., Fuller, D., Hampson, J., Haslam, M., Jones, S.,Koshy, J., Miracle, P., Oppenheimer, C., Roberts, P., White, K., 2009b. Humanoccupation, adaptation and behavioural change in the Pleistocene and Holoceneof south India: recent investigations in the Kurnool District, Andhra Pradesh.Eurasian Prehist. 6, 119e166.
Petraglia, M., Clarkson, C., Boivin, N., Haslam, M., Korisettar, R., Chaubey, G.,Ditchfield, P., Fuller, D., James, H., Jones, S., Kivisild, T., Koshy, J., Lahr, M.M.,Metspalu, M., Roberts, R., Arnold, L., 2009a. Population increase and environ-mental deterioration correspond with microlithic innovations in South Asia ca.35,000 years ago. Proc. Natl. Acad. Sci. 106, 12261e12266.
Petraglia, M.D., Haslam, M., Fuller, D., Boivin, N., Clarkson, C., 2010. Out of Africa:new hypotheses and evidence for the dispersal of Homo sapiens along theIndian Ocean rim. Annls. Hum. Biol. 37, 288e311.
Piper, P.J., Rabett, R.J., 2009. Hunting in a tropical rainforest; evidence from theterminal Pleistocene at Lobang Hangus, Niah caves, Sarawak. Int. J. Osteo-archaeol. 19, 551e565.
Piper, P.J., Rabett, R.J., Bin Kurui, E., 2008. Using community, composition and struc-tural variation in terminal Pleistocene vertebrate assemblages to identify humanhunting behaviour at the Niah Caves, Borneo. Bull. Indo. Pac. Prehist. 28, 88e98.
Pope, K.O., Terrell, J.E., 2008. Environmental setting of human migrations in thecircum-Pacific region. J. Biogeogr. 35, 1e21.
Premathilake, R., Risberg, J., 2003. Late Quaternary history of the Horton Plains,central Sri Lanka. Quatern. Sci. Rev. 22, 1525e1541.
Radcliffe-Brown, A.R., 1928. The Andaman Islanders. The Free Press of Glencoe, NewYork.
Raheem, D.C., Naggs, F., Preece, R.C., Mapatuna, Y., Kariyawasam, L., Eggleton, P., 2008.Structure and conservation of Sri Lankan landsnail assemblages in fragmentedlowland rainforest and village home gardens. J. Appl. Ecol. 45, 1019e1028.
Reimer, P.J., Baillie, M.G.L., Bard, E., Bayliss, A., Beck, J.W., Blackwell, P.G., BronkRamsey, C., Buck, C.E., Burr, G.S., Edwards, R.L., Friedrich, M., Grootes, P.M.,Guilderson, T.P., Hajdas, I., Heaton, T.J., Hogg, A.G., Hughen, K.A., Kaiser, K.F.,Kromer, B., McCormac, F.G., Manning, S.W., Reimer, R.W., Richards, D.A.,Southon, J.R., Talamo, S., Turney, C.S.M., van der Plicht, J., Weyhenmeyer, C.E.,2009. IntCal09 and Marine09 radiocarbon age calibration curves, 0-50,000years cal BP. Radiocarbon 51, 1111e1150.
Roberts, R.G., Jones, R., Spooner, N.A., Head, M.J., Murray, A.S., Smith, M.A., 1994. Thehuman colonisation of Australia: optical dates of 53,000 and 60,000 yearsbracket human arrival at Deaf Adder Gorge, Northern Territory. Quatern. Sci.Rev. 13, 575e583.
Sarasin, P., Sarasin, F., 1908. Die Steinzeit auf Ceylon. Ergebnisse natur-wissenschaftlicher Forschungen auf Ceylon, IV. C.W. Kreidel’s Verlag, Wiesbaden.
Shea, J.J., 2001. The Middle Paleolithic: early modern humans and Neandertals inthe Levant. Near East. Archaeol. 64, 38e64.
Stiner, M.C., 2001. Thirty years on the ‘’Broad Spectrum Revolution’’ and paleolithicdemography. Proc. Natl. Acad. Sci. 98, 6993e6996.
Stiner, M.C., 2002. Carnivory, coevolution, and the geographic spread of the genusHomo. J. Archaeol. Res. 10, 1e63.
Stiner, M.C., 2004. Small game use and expanding diet breadth in the EasternMediterranean Basin during the Palaeolithic. XXIVe Rencontres Internationalesd’ Archéologie et d’ Histoire d’ Antibes. In: Brugal, J.-P., Desse, J. (Eds.), PetitsAnimaux et Sociétés Humaines. Du Complément Alimentaire des RessourcesUtilitaires. Éditions APDCA, Antibes, pp. 499e513.
Stiner, M.C., Beaver, J.E., Munro, N.D., Surovell, T.A., 2008. Modelling Paleolithicpredator-prey dynamics and the effects of hunting pressure on prey ‘choice’. In:Bocquet-Appel, J.-P. (Ed.), Recent Advances in Palaeodemography. Springer,Dordrecht, pp. 143e178.
Stoops, G., 2003. Guidelines for Analysis and Description of Soil and Regolith ThinSections. Soil Science Society of America, Madison.
Stringer, C., 2002. Modern human origins: progress and prospects. Phi. Trans. R. Soc.Lond. B 357, 563e579.
Stringer, C.B., Andrews, P., 1988. Modern human origins. Science 241, 773e774.Sullivan III, A.P., Rosen, K.C., 1985. Debitage analysis and archaeological interpre-
tation. Am. Antiq. 50, 755e779.Swisher III, C.C., Rink, W.J., Antón, S.C., Schwarcz, H.P., Curtis, G.H., Suprijo
Widiasmoro, A., 1996. Latest Homo erectus of Java: potential contemporaneitywith Homo sapiens in southeast Asia. Science 274, 1870e1874.
Thamban, M., Purnachandra Rao, V., Schneider, R.R., 2002. Reconstruction of lateQuaternary monsoon oscillations based on clay mineral proxies using sedimentcores from the western margin of India. Mar. Geol. 186, 527e539.
Trimen, H., Alston, A.H.G., Hooker, J.D., 1893. A Handbook to the Flora of Ceylon.Dulau and Co., London.
Trinkaus, E., 2005. Early modern humans. Annu. Rev. Anthropol. 34, 207e230.Trinkaus, E., Moldovan, O., Milota, S, Bîlg�ar, A., Sarcina, L., Athreya, S., Bailey, S.E.,
Rodrigo, R., Mircea, G., Higham, T., Bronk Ramsey, C., van der Plicht, J., 2003. Anearly modern human from the Pestera cu Oase, Romania. Proc. Natl. Acad. Sci.100, 11231e11236.
Viet, N., 2008. Hoabinhian macrobotanical remains from archaeological sites inVietnam: indicators of climate changes from the late Pleistocene to the earlyHolocene. Bull. Indo. Pac. Prehist. 28, 80e83.
Voris, H.K., 2000. Maps of Pleistocene sea levels in Southeast Asia: shorelines, riversystems and time durations. J. Biogeogr. 27, 1153e1167.
Wadley, L., 2005. Putting ochre to the test: replication studies of adhesives that mayhave been used for hafting tools in the Middle Stone Age. J. Hum. Evol. 49,587e601.
Wadley, L., Williamson, B., Lombard, M., 2004. Ochre in hafting in Middle Stone Agesouthern Africa: a practical role. Antiquity 78, 661e675.
Walter, R.C., Buffler, R.T., Bruggemann, J.H., Gulliaume, M.M.M., Berhe, S.M.,Negassi, B., Libsekal, Y., Cheng, H., Edwards, R.L., von Cosel, R., Néraudeau, D.,Gagnon, M., 2000. Early human occupation of the Red Sea coast of Eritreaduring the last Interglacial. Nature 405, 65e69.
Wang, Y.J., Cheng, H., Edwards, R.L., An, Z.S., Wu., J.Y., Shen, C.-C., Dorale, J.A., 2001.A high-resolution absolute-dated late Pleistocene monsoon record from HuluCave, China. Science 294, 2345e2348.
Webb, W.F., 1948. Foreign Land and FreshWater Shells. W.F. Webb, St. Petersberg, FL.Weiss, E., Wetterstrom, W., Nadel, D., Bar-Yosef, O., 2004. The broad spectrum
revisited: evidence from plant remains. Proc. Natl. Acad. Sci. 101, 9551e9555.Wijeyapala, W.H., 1997. New Light on the Prehistory of Sri Lanka in the Context of
Recent Investigations of Cave Sites. Ph.D. Dissertation, University of Peradeniya.Wright, K.I., 1994. Ground-stone tools and hunter-gatherer subsistence in South-
west Asia: implications for the transition to farming. Am. Antiq. 59, 238e263.





























