Embed Size (px)
Citation preview

Peptide display on Potato virus X: molecularfeatures of the coat protein-fused peptide affectingcell-to-cell and phloem movement of chimeric virusparticles
Chiara Lico,1 Floriana Capuano,1 Giovanni Renzone,2 Marcello Donini,1
Carla Marusic,1 Andrea Scaloni,2 Eugenio Benvenuto1
and Selene Baschieri1
Correspondence
Selene Baschieri
1Sezione di Genetica e Genomica Vegetale, ENEA CR Casaccia, Via Anguillarese 301,00060 Rome, Italy
2Proteomics and Mass Spectrometry Laboratory, ISPAAM, National Research Council,80147 Naples, Italy
Received 4 April 2006
Accepted 26 June 2006
The potexvirus Potato virus X (PVX) can be modified genetically to generate chimeric virus particles
(CVPs) carrying heterologous peptides fused to coat protein (CP) subunits. A spontaneous PVX
mutant expressing a truncated, but functional, form of the CP has been isolated. With the aim
of exploiting this virus to display peptides useful for vaccine formulations, two novel viral expression
vectors based on pPVX201 (bearing the wild-type PVX genome) were constructed encoding the
truncated CP. Both vectors were able to produce infectious virus particles in planta and were used
to insert a panel of sequences encoding peptides of biopharmaceutical interest as N-terminal
fusions to the truncated cp gene. The analysis of infection progression induced by the different
constructs enabled identification of two important structural features of the fused peptide, namely
tryptophan content and isoelectric point, critically affecting the formation of PVX CVPs and virus
movement through the plant. These results are discussed in view of the rising interest in engineered
plant viruses for development of peptide-based epitope vaccines.
INTRODUCTION
Genetically modified plant viruses can be used to transientlyexpress heterologous sequences in susceptible plant hosts(Pogue et al., 2002). A promising application of engineeredplant viruses is the development of epitope vaccines by thegeneration of chimeric virus particles (CVPs) that displaypeptides of interest on their surface as fusions to the coatprotein (CP) (Scholthof et al., 1996). The potexvirus Potatovirus X (PVX) has been shown to be an ideal, highly ordered,multivalent scaffold to be used to this purpose (Brennanet al., 1999; Marusic et al., 2001). PVX could also be used as acarrier of whole proteins, although this approach presentssome disadvantages in vaccine applications due to theunpredictability of the number of chimeric CP units pervirion, precluding an exact determination of the vaccinedose (Santa Cruz et al., 1996).
PVX filamentous particles consist of a single-stranded,positive-sense RNA molecule embedded in a capsid made ofapproximately 1300 units of the same protein. Although no
atomic-resolution data are available defining the organiza-tion of the virus particle and CP folding, it is assumed thatPVX has a helical structure, with the CP N terminus exposedon the viral surface (Baratova et al., 1992a, b; Parker et al.,2002). For this reason, the construction of PVX CVPsrequires the fusion of exogenous sequences at the 59
terminus of the cp gene. The genetic manipulation of PVXhas been greatly improved by the development of expressionvectors, such as pPVX201, that contain the cDNA encodingthe complete viral genome (Baulcombe et al., 1995;Chapman et al., 1992a).
In this work, we have isolated a spontaneous PVX mutantcharacterized by a deletion at the 59 terminus of the cp gene.Two pPVX201-derived vectors encoding the cp mutant genehave been constructed and were used to fuse, to the 59 end ofthe cp gene, sequences encoding a panel of peptides varyingin both length and amino acid composition. From theobservation of the infectious phenotypes induced in plantaby the different constructs, we concluded that cell-to-celland phloem movement of PVX CVPs is critically affected bythe occurrence of tryptophan (Trp), as well as the isoelectricpoint (pI) of the peptide.Supplementary methods are available in JGV Online.
0008-2097 G 2006 SGM Printed in Great Britain 3103
Journal of General Virology (2006), 87, 3103–3112 DOI 10.1099/vir.0.82097-0

METHODS
DNA constructs. Construction of the pPVX201 viral expressionvector (kindly provided by D. Baulcombe, The Sainsbury Laboratory,Norwich, UK) (Baulcombe et al., 1995) encoding the peptide KSS(amino acid sequence CKSSGKLISLC) (Scala et al., 1999) as an N-terminal fusion with the CP was performed as described previously(Marusic et al., 2001), adopting a strategy that removes the dupli-cated cp subgenomic promoter.
To construct the viral expression vector pPVXCC, the 59-deletedportion of the mutant cp gene was substituted to the wild-type cp inpPVX201. Briefly, an RT-PCR fragment was obtained by using totalRNA extracted from the systemically infected leaves of pPVX201-KSS-inoculated plants as a template and primers PVXBack(59-CTGGGGAATCAATCACAGTGTTG-39) and PVXXho (59-GAC-GTAGTTATGGTGGTGGTAG-39). The PCR product was digestedwith the restriction enzymes NheI and XhoI and ligated into similarlydigested pPVX201. To construct the viral expression vector pPVXSma,in which sequences recognized by the restriction enzyme SmaI areinserted immediately downstream of the start codon of the mutant cpgene, sense (59-TCGACATGCCCGGGACTCCTGCCACAGCTTCA-GG-39) and antisense (59-CCTGAAGCTGTGGCAGGAGTCCCG-GGCATG-39) primers were designed and annealed in vitro to generatea fragment with NheI-compatible 59 and StuI-compatible 39 ends. Thisfragment was ligated into NheI–StuI-digested pPVXCC.
To insert sequences encoding the heterologous peptides as 59-endfusions with the cp gene in pPVXCC, primer pairs were designed(following the PVX cp codon usage) and annealed in vitro to obtainfragments with NheI-compatible 59 and StuI-compatible 39 ends to beligated into the NheI–StuI-digested vector. All of the sequencesencoding the peptides were preceded by an ATG codon and followed bythe cp gene portion that is lost by StuI digestion.
To insert the sequences encoding the heterologous peptides as fusionswith the cp gene into pPVXSma, oligonucleotide pairs were designed toobtain NheI-compatible 59- and SmaI-compatible 39-end DNAfragments that were ligated into the NheI–SmaI-digested vector. Asreported previously, sequences encoding the peptides were preceded byan ATG codon.
The biochemical features of the N-terminal CP peptides encoded bypPVXCC- and pPVXSma-derived constructs were calculated by usingthe ProtParam tool (http://www.expasy.org/tools/protparam.html).
Plant infection. Nicotiana benthamiana plants were inoculatedwith the different constructs or with pPVX201 (as control) asdescribed previously (Marusic et al., 2001). To verify the genomicstability and infectivity of virus particles that cause systemic infec-tion, repeated cycles of reinfection were performed using leaf sap,prepared by homogenizing infected tissue in 16 PBS and centrifu-ging the sample for 3 min at 20 000 g and 4 uC. The supernatant wasused directly to infect the plants. Briefly, sap obtained from sympto-matic systemic leaves of the plants inoculated with the construct ofinterest (#1) was used to inoculate a second group of plants (#2).Afterwards, a third group of plants (#3) was infected by using thesap of the symptomatic systemic leaves from #2 plants. The presenceof the expected cp gene was assessed by RT-PCR on total RNAextracted from systemic leaves of plants #1, #2 and #3, as describedbelow.
RNA extraction and RT-PCR. Ten to twelve days post-infection(p.i.), expression of the chimeric cp genes was verified by RT-PCR.Briefly, total RNA from systemically infected leaves was extracted byusing an RNeasy plant mini kit (Qiagen) and RT-PCRs were per-formed by using a GeneAmp RNA PCR Kit (Perkin Elmer). cDNAwas synthesized by using oligo d(T)16 and PCR was carried out
with PVXBack and PVXNew (59-CAGTCTAGCTCTGCTGATGC-CGTTG-39) primers. PCR fragments were purified and verified bysequencing.
Western blot analysis and ELISA. Protein extracts were obtainedfrom inoculated and systemic leaves as described previously (Marusicet al., 2001). Aliquots (5 mg for extracts of symptomatic leaves or20 mg for extracts of asymptomatic leaves) were separated on a12?5 % (w/v) SDS-PAGE gel before transferring onto a PVDF mem-brane (Millipore). Immunodetection was performed as described pre-viously (Donini et al., 2005).
Alternatively, the presence of PVX CP was revealed by ELISA using anAgdia Inc. kit (catalogue no. SRP10 000/0500) following themanufacturer’s instructions.
Site-directed mutagenesis. The mutagenesis of pPVXSma-NYESOand pPVXSma-NYESOSh constructs was performed by using aQuikChange Multi site-directed mutagenesis kit following the manu-facturer’s instructions (Stratagene). The same oligonucleotide(59-Pho-AGCCTACTAATGGGGATTACACAATGT-39) allowed theTGG(Trp)RGGG(glycine, Gly) codon substitution in both constructs.
Electron microscopy. Small pieces of the inoculated leaves, includ-ing necrotic ringspots, were fixed in 2?5 % (v/v) glutaraldehyde solu-tion in 0?08 M phosphate buffer (pH 7?0) and then transferred into1 % (w/v) osmium tetroxide. After dehydration in ethanol solutions,the samples were embedded in araldite resin. Ultrathin sections pre-pared by using an Ultracut-E ultramicrotome (Reichert–Jung) werecollected on copper grids and stained with 1 % (w/v) uranyl acetatesolution. Sections were then analysed by using a transmission elec-tron microscope EM208 (Philips).
RESULTS
Isolation of a PVX CP mutant
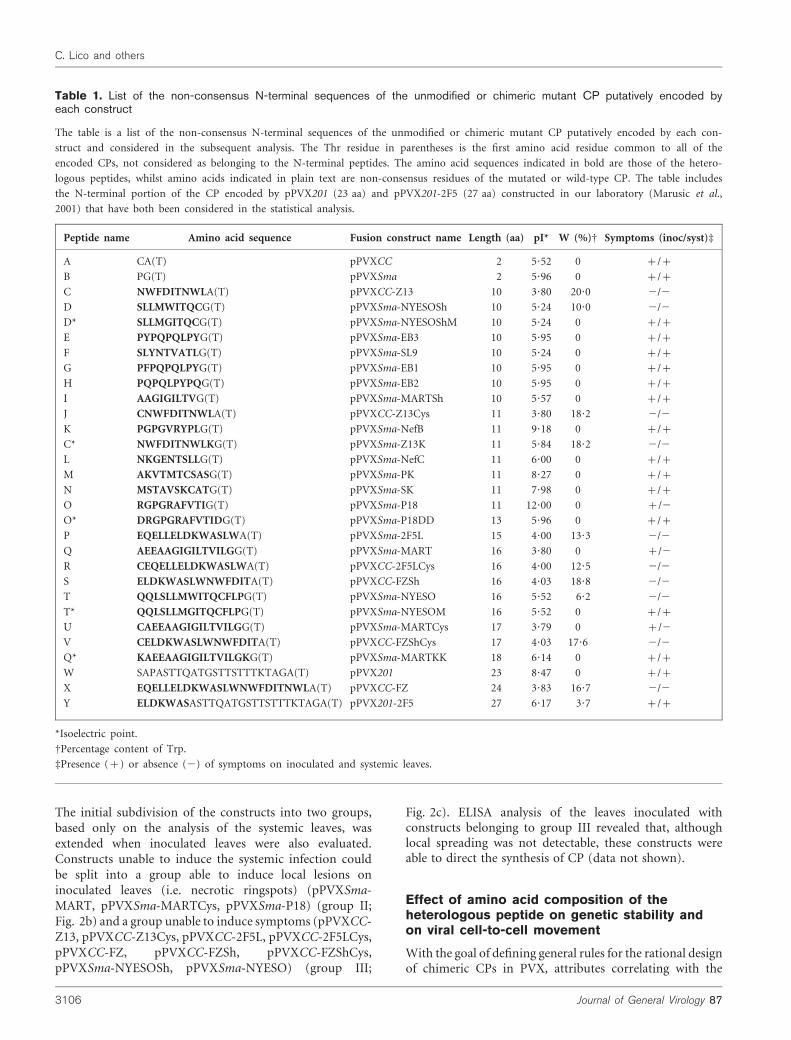
The PVX viral expression vector pPVX201 (strain X3)(Fig. 1a, b) was modified by inserting at the 59 end of the cpgene the sequence encoding a peptide of interest (KSS) forvaccine formulations. The resulting construct, pPVX201-KSS, was used to inoculate N. benthamiana plants with theaim of obtaining CVPs displaying the KSS peptide on theirsurface. Ten days p.i., the systemic leaves showing veinalchlorosis (a typical symptom of PVX infection) weresampled. To verify the presence of the expected modifiedviral genome within the leaves, total RNA was extracted,reverse-transcribed and amplified in the region includingthe 59 end of the cp gene. Surprisingly, we found arearrangement of the cp gene, causing the loss of nt 4–67from the 59 end. Analysis of the deduced amino acidsequence revealed that residues 2–22 at the N terminus werelost and substituted by a cysteine (Cys) residue in the correctreading frame.
Construction of pPVXCC and pPVXSMA viralexpression vectors
The rearranged cp gene was substituted to the wild-type cp inpPVX201, generating the pPVXCC viral expression vector(Fig. 1d). To verify the ability of pPVXCC to produceinfectious virus particles, the vector was used to inoculate N.benthamiana plants. Ten days p.i., systemic leaves of theplants displayed the same symptoms induced by pPVX201
3104 Journal of General Virology 87
C. Lico and others

(Fig. 1c, e) and contained the expected viral RNA, asassessed by RT-PCR and sequence analysis (not shown).Moreover, after repeated infection cycles with sap fromsystemic leaves of pPVXCC-inoculated plants, the sequenceof the viral genome did not change, demonstrating viralgenome stability.
To facilitate the insertion of sequences encoding hetero-logous peptides into pPVXCC, the vector was engineered by
adding at the 59 end of the deleted cp gene a uniquerestriction site (SmaI) (Fig. 1f). This modification producedthe substitution of Cys (CP position 2) and alanine (Ala)(CP position 3) with proline (Pro) and Gly, respectively(Table 1). The resulting pPVXSma vector induced symp-toms on systemic leaves that were identical to those of thecontrols (Fig. 1g). The stability of the genome of these virusparticles was verified as described previously (data notshown).
Heterologous peptides
To determine peptide traits influencing structural stabilityand infectivity of the chimeric viruses, peptides of differentlength and amino acid composition were chosen to bedisplayed as CP fusions (Table 1). The panel of peptidesincluded epitopes selected from melanoma-associated(MART, MARTSh, NYESO, NYESOSh) (Kirkin et al.,1998) or human immunodeficiency virus type 1 (Z13,Z13Cys, 2F5L, 2F5LCys, FZ, FZSh, FZShCys, SL9, P18,NefB, NefC) (Hurwitz et al., 2005; Shiver & Emini, 2004)proteins. Other peptides, associated with coeliac disease(EB1, EB2, EB3) (Shan et al., 2002) or endowed withantimicrobial activity (PK, SK) (Donini et al., 2005;Polonelli et al., 2003), were also included. To elucidate apossible role of the Cys residue in position 2 of pPVXCC CPon viral stability, some of the peptide-encoding sequenceswere designed both with and without this amino acid in thisposition.
Symptoms and molecular analysis of plantsinoculated with the chimeric CP-encodingconstructs
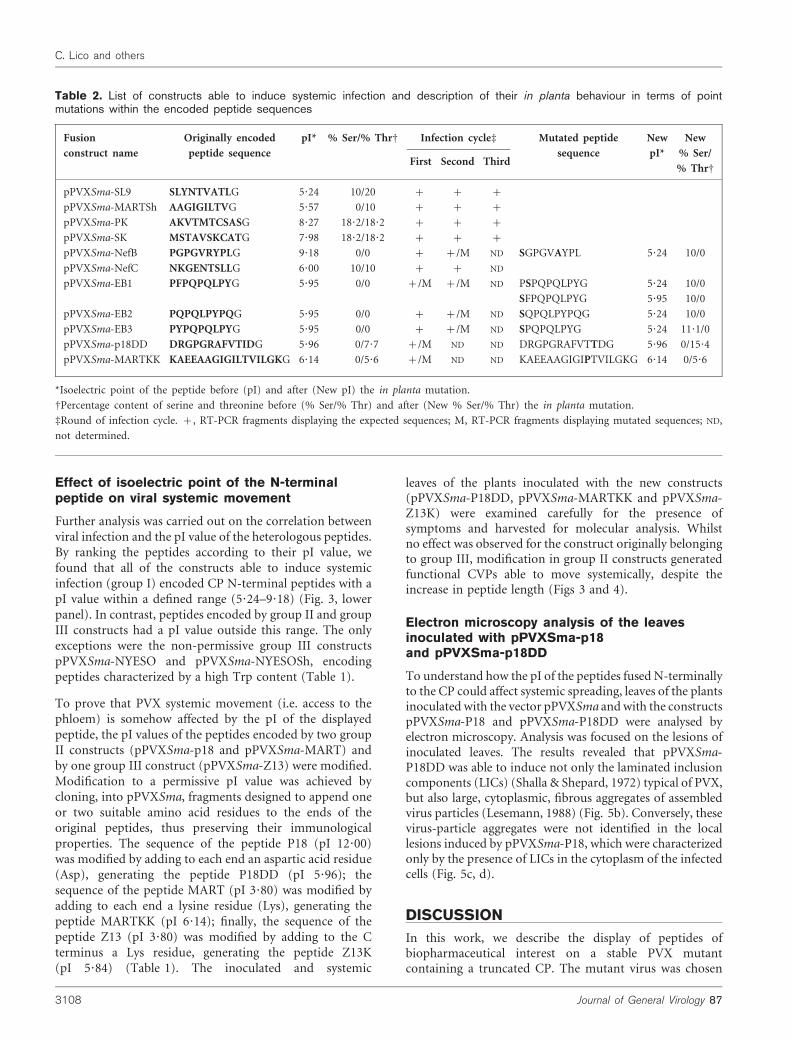
Ten to twelve days p.i., systemic leaves were examined toevaluate the presence of virus symptoms. Subsequently, planttissues were analysed for the presence of the expected viralRNAs and CPs by sequencing and Western blotting. Theconstructs were divided easily into two main groups: thoseable to induce systemic infection (pPVXSma-SL9, pPVXSma-EB1, pPVXSma-EB2, pPVXSma-EB3, pPVXSma-MARTSh,pPVXSma-KS, pPVXSma-SK, pPVXSma-NefB, pPVXSma-NefC) and those that were not. Viral RNA was present insystemic leaves of plants inoculated with constructs able toinduce systemic infection (group I; Fig. 2a). Although majorsequence rearrangements did not occur, occasional pointmutations affected the heterologous nucleotide sequenceselectively, resulting in amino acid substitutions in thedisplayed peptide. This phenomenon was mostly evidentduring reinfections, but in one case (pPVXSma-EB1), thesubstitution occurred during the infection cycle produced byplasmid inoculation (Table 2). In view of the possibleapplication of the systemically moving and geneticallystable CVPs to biopharmaceutical research, some of themwere produced on a large scale in plants and, afterpurification, the nature of CP–peptide fusion proteins wasascertained by extensive mass mapping experiments (seesupplementary material, available in JGV Online).
Fig. 1. (a) Schematic representation of pPVX201 viral expres-sion vector. Hatched box, multicloning site; 35S, 35S promoterof Cauliflower mosaic virus; filled box, Agrobacterium tume-
faciens nopaline synthase gene transcription terminator; RdRP,RNA-dependent RNA polymerase; M, movement proteins; CP,coat protein. (b) Detail of the cp gene region in pPVX201. (c)Systemic leaves of N. benthamiana plants 10 days p.i. withpPVX201. (d) Detail of the deleted cp gene region inpPVXCC. (e) Systemic leaves of N. benthamiana plants10 days p.i. with pPVXCC. (f) Detail of the deleted and engi-neered cp gene region in pPVXSma. (g) Systemic leaves of N.
benthamiana plants 10 days p.i. with pPVXSma. Filled arrows,cp gene original and duplicated subgenomic promoter; hatchedhorizontal arrows, PVXBack oligonucleotide; empty arrow,PVXNew oligonucleotide; shaded arrow, PVXXho oligonucleo-tide. Major restriction sites are indicated.
http://vir.sgmjournals.org 3105
Peptide features affecting mutant PVX CVP fitness
The initial subdivision of the constructs into two groups,based only on the analysis of the systemic leaves, wasextended when inoculated leaves were also evaluated.Constructs unable to induce the systemic infection couldbe split into a group able to induce local lesions oninoculated leaves (i.e. necrotic ringspots) (pPVXSma-MART, pPVXSma-MARTCys, pPVXSma-P18) (group II;Fig. 2b) and a group unable to induce symptoms (pPVXCC-Z13, pPVXCC-Z13Cys, pPVXCC-2F5L, pPVXCC-2F5LCys,pPVXCC-FZ, pPVXCC-FZSh, pPVXCC-FZShCys,pPVXSma-NYESOSh, pPVXSma-NYESO) (group III;
Fig. 2c). ELISA analysis of the leaves inoculated withconstructs belonging to group III revealed that, althoughlocal spreading was not detectable, these constructs wereable to direct the synthesis of CP (data not shown).
Effect of amino acid composition of theheterologous peptide on genetic stability andon viral cell-to-cell movement
With the goal of defining general rules for the rational designof chimeric CPs in PVX, attributes correlating with the
Table 1. List of the non-consensus N-terminal sequences of the unmodified or chimeric mutant CP putatively encoded byeach construct
The table is a list of the non-consensus N-terminal sequences of the unmodified or chimeric mutant CP putatively encoded by each con-
struct and considered in the subsequent analysis. The Thr residue in parentheses is the first amino acid residue common to all of the
encoded CPs, not considered as belonging to the N-terminal peptides. The amino acid sequences indicated in bold are those of the hetero-
logous peptides, whilst amino acids indicated in plain text are non-consensus residues of the mutated or wild-type CP. The table includes
the N-terminal portion of the CP encoded by pPVX201 (23 aa) and pPVX201-2F5 (27 aa) constructed in our laboratory (Marusic et al.,
2001) that have both been considered in the statistical analysis.
Peptide name Amino acid sequence Fusion construct name Length (aa) pI* W (%)D Symptoms (inoc/syst)d
A CA(T) pPVXCC 2 5?52 0 +/+
B PG(T) pPVXSma 2 5?96 0 +/+
C NWFDITNWLA(T) pPVXCC-Z13 10 3?80 20?0 2/2
D SLLMWITQCG(T) pPVXSma-NYESOSh 10 5?24 10?0 2/2
D* SLLMGITQCG(T) pPVXSma-NYESOShM 10 5?24 0 +/+
E PYPQPQLPYG(T) pPVXSma-EB3 10 5?95 0 +/+
F SLYNTVATLG(T) pPVXSma-SL9 10 5?24 0 +/+
G PFPQPQLPYG(T) pPVXSma-EB1 10 5?95 0 +/+
H PQPQLPYPQG(T) pPVXSma-EB2 10 5?95 0 +/+
I AAGIGILTVG(T) pPVXSma-MARTSh 10 5?57 0 +/+
J CNWFDITNWLA(T) pPVXCC-Z13Cys 11 3?80 18?2 2/2
K PGPGVRYPLG(T) pPVXSma-NefB 11 9?18 0 +/+
C* NWFDITNWLKG(T) pPVXSma-Z13K 11 5?84 18?2 2/2
L NKGENTSLLG(T) pPVXSma-NefC 11 6?00 0 +/+
M AKVTMTCSASG(T) pPVXSma-PK 11 8?27 0 +/+
N MSTAVSKCATG(T) pPVXSma-SK 11 7?98 0 +/+
O RGPGRAFVTIG(T) pPVXSma-P18 11 12?00 0 +/2
O* DRGPGRAFVTIDG(T) pPVXSma-P18DD 13 5?96 0 +/+
P EQELLELDKWASLWA(T) pPVXSma-2F5L 15 4?00 13?3 2/2
Q AEEAAGIGILTVILGG(T) pPVXSma-MART 16 3?80 0 +/2
R CEQELLELDKWASLWA(T) pPVXCC-2F5LCys 16 4?00 12?5 2/2
S ELDKWASLWNWFDITA(T) pPVXCC-FZSh 16 4?03 18?8 2/2
T QQLSLLMWITQCFLPG(T) pPVXSma-NYESO 16 5?52 6?2 2/2
T* QQLSLLMGITQCFLPG(T) pPVXSma-NYESOM 16 5?52 0 +/+
U CAEEAAGIGILTVILGG(T) pPVXSma-MARTCys 17 3?79 0 +/2
V CELDKWASLWNWFDITA(T) pPVXCC-FZShCys 17 4?03 17?6 2/2
Q* KAEEAAGIGILTVILGKG(T) pPVXSma-MARTKK 18 6?14 0 +/+
W SAPASTTQATGSTTSTTTKTAGA(T) pPVX201 23 8?47 0 +/+
X EQELLELDKWASLWNWFDITNWLA(T) pPVXCC-FZ 24 3?83 16?7 2/2
Y ELDKWASASTTQATGSTTSTTTKTAGA(T) pPVX201-2F5 27 6?17 3?7 +/+
*Isoelectric point.
DPercentage content of Trp.
dPresence (+) or absence (2) of symptoms on inoculated and systemic leaves.
3106 Journal of General Virology 87
C. Lico and others
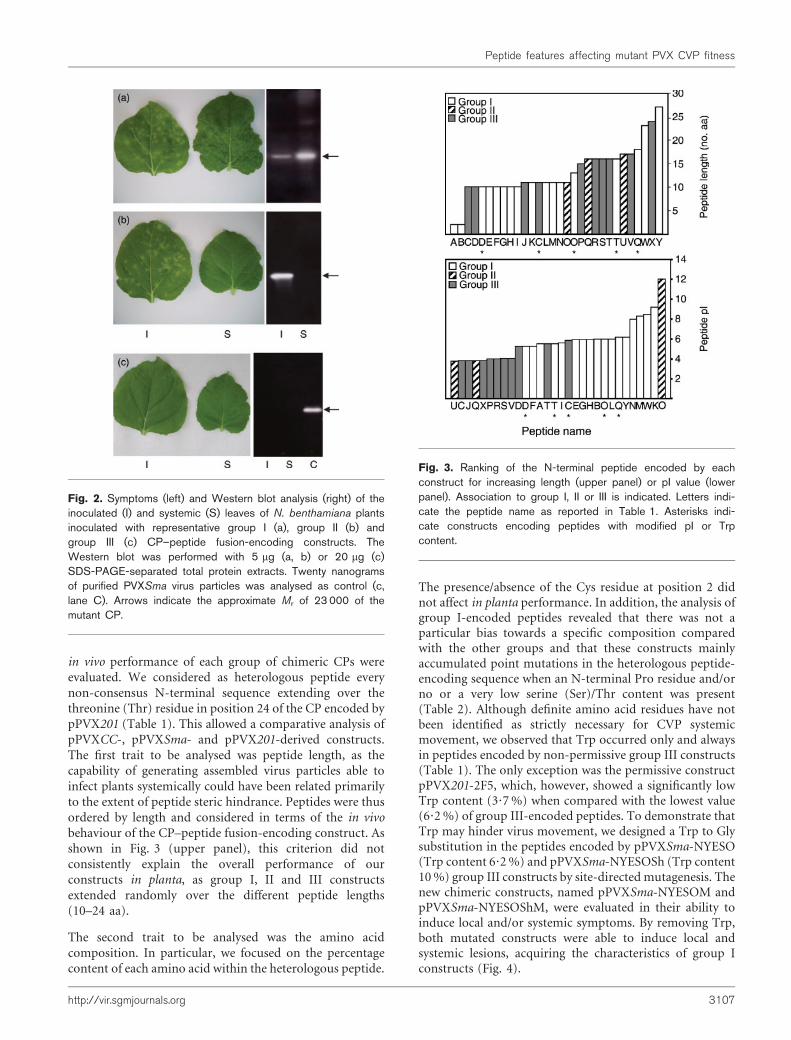
in vivo performance of each group of chimeric CPs wereevaluated. We considered as heterologous peptide everynon-consensus N-terminal sequence extending over thethreonine (Thr) residue in position 24 of the CP encoded bypPVX201 (Table 1). This allowed a comparative analysis ofpPVXCC-, pPVXSma- and pPVX201-derived constructs.The first trait to be analysed was peptide length, as thecapability of generating assembled virus particles able toinfect plants systemically could have been related primarilyto the extent of peptide steric hindrance. Peptides were thusordered by length and considered in terms of the in vivobehaviour of the CP–peptide fusion-encoding construct. Asshown in Fig. 3 (upper panel), this criterion did notconsistently explain the overall performance of ourconstructs in planta, as group I, II and III constructsextended randomly over the different peptide lengths(10–24 aa).
The second trait to be analysed was the amino acidcomposition. In particular, we focused on the percentagecontent of each amino acid within the heterologous peptide.
The presence/absence of the Cys residue at position 2 didnot affect in planta performance. In addition, the analysis ofgroup I-encoded peptides revealed that there was not aparticular bias towards a specific composition comparedwith the other groups and that these constructs mainlyaccumulated point mutations in the heterologous peptide-encoding sequence when an N-terminal Pro residue and/orno or a very low serine (Ser)/Thr content was present(Table 2). Although definite amino acid residues have notbeen identified as strictly necessary for CVP systemicmovement, we observed that Trp occurred only and alwaysin peptides encoded by non-permissive group III constructs(Table 1). The only exception was the permissive constructpPVX201-2F5, which, however, showed a significantly lowTrp content (3?7 %) when compared with the lowest value(6?2 %) of group III-encoded peptides. To demonstrate thatTrp may hinder virus movement, we designed a Trp to Glysubstitution in the peptides encoded by pPVXSma-NYESO(Trp content 6?2 %) and pPVXSma-NYESOSh (Trp content10 %) group III constructs by site-directed mutagenesis. Thenew chimeric constructs, named pPVXSma-NYESOM andpPVXSma-NYESOShM, were evaluated in their ability toinduce local and/or systemic symptoms. By removing Trp,both mutated constructs were able to induce local andsystemic lesions, acquiring the characteristics of group Iconstructs (Fig. 4).
Fig. 2. Symptoms (left) and Western blot analysis (right) of theinoculated (I) and systemic (S) leaves of N. benthamiana plantsinoculated with representative group I (a), group II (b) andgroup III (c) CP–peptide fusion-encoding constructs. TheWestern blot was performed with 5 mg (a, b) or 20 mg (c)SDS-PAGE-separated total protein extracts. Twenty nanogramsof purified PVXSma virus particles was analysed as control (c,lane C). Arrows indicate the approximate Mr of 23 000 of themutant CP.
Fig. 3. Ranking of the N-terminal peptide encoded by eachconstruct for increasing length (upper panel) or pI value (lowerpanel). Association to group I, II or III is indicated. Letters indi-cate the peptide name as reported in Table 1. Asterisks indi-cate constructs encoding peptides with modified pI or Trpcontent.
http://vir.sgmjournals.org 3107
Peptide features affecting mutant PVX CVP fitness
Effect of isoelectric point of the N-terminalpeptide on viral systemic movement
Further analysis was carried out on the correlation betweenviral infection and the pI value of the heterologous peptides.By ranking the peptides according to their pI value, wefound that all of the constructs able to induce systemicinfection (group I) encoded CP N-terminal peptides with apI value within a defined range (5?24–9?18) (Fig. 3, lowerpanel). In contrast, peptides encoded by group II and groupIII constructs had a pI value outside this range. The onlyexceptions were the non-permissive group III constructspPVXSma-NYESO and pPVXSma-NYESOSh, encodingpeptides characterized by a high Trp content (Table 1).
To prove that PVX systemic movement (i.e. access to thephloem) is somehow affected by the pI of the displayedpeptide, the pI values of the peptides encoded by two groupII constructs (pPVXSma-p18 and pPVXSma-MART) andby one group III construct (pPVXSma-Z13) were modified.Modification to a permissive pI value was achieved bycloning, into pPVXSma, fragments designed to append oneor two suitable amino acid residues to the ends of theoriginal peptides, thus preserving their immunologicalproperties. The sequence of the peptide P18 (pI 12?00)was modified by adding to each end an aspartic acid residue(Asp), generating the peptide P18DD (pI 5?96); thesequence of the peptide MART (pI 3?80) was modified byadding to each end a lysine residue (Lys), generating thepeptide MARTKK (pI 6?14); finally, the sequence of thepeptide Z13 (pI 3?80) was modified by adding to the Cterminus a Lys residue, generating the peptide Z13K(pI 5?84) (Table 1). The inoculated and systemic
leaves of the plants inoculated with the new constructs(pPVXSma-P18DD, pPVXSma-MARTKK and pPVXSma-Z13K) were examined carefully for the presence ofsymptoms and harvested for molecular analysis. Whilstno effect was observed for the construct originally belongingto group III, modification in group II constructs generatedfunctional CVPs able to move systemically, despite theincrease in peptide length (Figs 3 and 4).
Electron microscopy analysis of the leavesinoculated with pPVXSma-p18and pPVXSma-p18DD
To understand how the pI of the peptides fused N-terminallyto the CP could affect systemic spreading, leaves of the plantsinoculated with the vector pPVXSma and with the constructspPVXSma-P18 and pPVXSma-P18DD were analysed byelectron microscopy. Analysis was focused on the lesions ofinoculated leaves. The results revealed that pPVXSma-P18DD was able to induce not only the laminated inclusioncomponents (LICs) (Shalla & Shepard, 1972) typical of PVX,but also large, cytoplasmic, fibrous aggregates of assembledvirus particles (Lesemann, 1988) (Fig. 5b). Conversely, thesevirus-particle aggregates were not identified in the locallesions induced by pPVXSma-P18, which were characterizedonly by the presence of LICs in the cytoplasm of the infectedcells (Fig. 5c, d).
DISCUSSION
In this work, we describe the display of peptides ofbiopharmaceutical interest on a stable PVX mutantcontaining a truncated CP. The mutant virus was chosen
Table 2. List of constructs able to induce systemic infection and description of their in planta behaviour in terms of pointmutations within the encoded peptide sequences
Fusion
construct name
Originally encoded
peptide sequence
pI* % Ser/% ThrD Infection cycled Mutated peptide
sequence
New
pI*
New
% Ser/
% ThrDFirst Second Third
pPVXSma-SL9 SLYNTVATLG 5?24 10/20 + + +
pPVXSma-MARTSh AAGIGILTVG 5?57 0/10 + + +
pPVXSma-PK AKVTMTCSASG 8?27 18?2/18?2 + + +
pPVXSma-SK MSTAVSKCATG 7?98 18?2/18?2 + + +
pPVXSma-NefB PGPGVRYPLG 9?18 0/0 + +/M ND SGPGVAYPL 5?24 10/0
pPVXSma-NefC NKGENTSLLG 6?00 10/10 + + ND
pPVXSma-EB1 PFPQPQLPYG 5?95 0/0 +/M +/M ND PSPQPQLPYG 5?24 10/0
SFPQPQLPYG 5?95 10/0
pPVXSma-EB2 PQPQLPYPQG 5?95 0/0 + +/M ND SQPQLPYPQG 5?24 10/0
pPVXSma-EB3 PYPQPQLPYG 5?95 0/0 + +/M ND SPQPQLPYG 5?24 11?1/0
pPVXSma-p18DD DRGPGRAFVTIDG 5?96 0/7?7 +/M ND ND DRGPGRAFVTTDG 5?96 0/15?4
pPVXSma-MARTKK KAEEAAGIGILTVILGKG 6?14 0/5?6 +/M ND ND KAEEAAGIGIPTVILGKG 6?14 0/5?6
*Isoelectric point of the peptide before (pI) and after (New pI) the in planta mutation.
DPercentage content of serine and threonine before (% Ser/% Thr) and after (New % Ser/% Thr) the in planta mutation.
dRound of infection cycle. +, RT-PCR fragments displaying the expected sequences; M, RT-PCR fragments displaying mutated sequences; ND,
not determined.
3108 Journal of General Virology 87
C. Lico and others

mainly because it is hypothetically more suitable for thedisplay of longer peptides than the wild type, but also as it isputatively less immunogenic, due to reduced CP dimensions(Koenig & Torrance, 1986). This could be an importantfeature when chimeric PVX is used as carrier of vaccinepeptides, an application that requires repeated administra-tion (Borisova et al., 1999; Tangri et al., 2005). pPVXCC andits derivative pPVXSma vectors, harbouring the mutated cp
gene, were able to induce plant infections indistinguishablefrom those induced by the wild-type pPVX201 vector, withthe formation of virus particles carrying genetically stablegenomes. These data, together with a recent study aimed atdefining PVX structure (Shanmugam et al., 2005), indicatethat the formation of functional virus particles is notaffected significantly by the removal of CP N-terminalamino acids. However, our observations also demonstratedthat the fusion of heterologous peptides of varying lengthand composition to the truncated N terminus does notalways result in systemic infection. This could be expected,as CP is essential for PVX movement (Baulcombe et al.,1995; Chapman et al., 1992a, b) through both plasmodes-mata and phloem (Fedorkin et al., 2001; Santa Cruz et al.,1998). By the visual inspection of inoculated plants, thechimeric constructs were classified into three main groups.In the case of groups I (symptoms on both inoculated andsystemic leaves) and II (symptoms only on inoculatedleaves), the infection phenotype matched perfectly with themolecular analysis performed. On the contrary, group IIIconstructs, despite being unable to induce detectablesymptoms, were able to direct the synthesis of the CP inthe inoculated leaves, although at low levels. This resultdemonstrated that the performance of these constructscould not simply be ascribed to transcriptional problems,but rather to a functional failure of the chimeric CP thatprevents the transport of the viral genome from initiallyinfected cells to the neighbours.
Because group III peptides limited viral local movement,whereas group II peptides were able to hinder CP functionselectively in phloem transport, our data indicate that theCP exerts distinct functions in cell-to-cell and systemicmovement. To shed light on how peptide attributes affecteach function selectively, we tried to identify commonphysicochemical parameters that characterized each groupof peptides unambiguously. Ranking peptides according totheir length resulted in a random distribution of the threegroups, indicating that this parameter was unrelated toability (group I) or not (group II and group III) to generateviral forms moving systemically. Peptide groupings werefurther analysed in terms of amino acid composition. Thisapproach revealed that, although systemic movement wasnot related to a specific amino acid composition, thepresence of Trp residues was peculiar to group III peptides.One exception to this rule was pPVX201-2F5 (group I), yetencoding the peptide with the lowest Trp content. Trpresidues are often found associated with the end ofhydrophobic regions in transmembrane proteins and aresupposed to act as ‘floats’ at the membrane boundary, byfixing the protein to the lipid bilayer (Lee, 2003). On thisbasis, it could be hypothesized that the inability of group IIIconstructs to promote cell-to-cell spreading might beascribed to peptide-mediated membrane anchoring ofchimeric CP, inhibiting cell-to-cell transfer. This hypothesiswas supported by the fact that group III peptides weremainly putative transmembrane or membrane-proximalpeptides (Chen et al., 1997; Salzwedel et al., 1999) and was
Fig. 4. Effect of TrpRGly substitution and of pI value modifica-tion on viral cell-to-cell and systemic movement, respectively.Left side, inoculated (I) and systemic (S) leaves of plants inocu-lated with the original constructs; right side, inoculated (I) andsystemic (S) leaves of plants inoculated with the constructsencoding peptides bearing the TrpRGly substitution or the pImodification.
http://vir.sgmjournals.org 3109
Peptide features affecting mutant PVX CVP fitness

finally confirmed by site-directed mutagenesis experimentsshowing that Trp to Gly substitution completely restored theability of the CP to induce systemic infection.
Another correlation found to be highly significant wasbetween the pI range of group I peptides and systemicmovement. The robustness of this correlation was tested bymodifying the pI value of peptides encoded by group II andgroup III constructs (i.e. MART, P18 and Z13). The resultsof these experiments showed that, whilst pI refinement ofgroup II peptides resulted in the generation of viral formsable to move systemically (hence acting as group I peptides),pI adjustment of group III peptides had no effect. These datasupport the idea that the influence of pI on systemicinfection may be hindered by a particularly high Trpcontent, interfering with earlier stages of viral movement.Thus, we can conclude that the effect of pI is selective onsystemic movement, whilst Trp affects cell-to-cell move-ment independently of the pI value.
Despite the fact that all constructs belonging to group Iconformed to the Trp and pI ‘rules’, some of them weresubjected to point mutations in the sequences encoding theheterologous peptides. These mutations occurred mainlyduring reinfection cycles and were mainly biased towardsthe increase of Ser/Thr content, supporting findings thatidentify in these residues the phosphorylation sites involvedin virus unpackaging (Baratova et al., 2004; Kozlovsky et al.,2003).
Electron microscopy studies, aimed to define differencesbetween group II and pI-remodelled group I constructs,showed that whilst LICs were present in both samples,formation of virus-particle aggregates in infected cells wasevident only after pI value adjustment. These results indicatethat a non-permissive pI value (i.e. a charged N terminus)could interfere with the correct assembly of chimeric CPsubunits to form complete virions. It is indeed a commonbelief that the introduction of repulsive charges in the N-terminal domain through phosphorylation is associatedwith PVX disassembly (Atabekov et al., 2001; Kozlovskyet al., 2003).
The major drawback to the production of CVPs harbouringpeptides of interest in plants has been ascribed primarily tothe steric interference of the foreign peptide with the correctassembly of virus particles (Scholthof et al., 1996), butpeptide pI/charge has also been shown to affect viral fitness.In the case of rod-shaped Tobacco mosaic virus, thisparameter affects cell-to-cell movement and this phenom-enon has been ascribed to a lethal effect of the CP–peptidefusion on host cells (Bendahmane et al., 1999). Conversely,in both icosahedral Cowpea mosaic virus and filamentousZucchini yellow mosaic virus, the effect is on systemicmovement and is explained in terms of interference in theinteraction between the CP and undefined host components(Kimalov et al., 2004; Porta et al., 2003). From these data, itis evident that common physicochemical features of adisplayed peptide can influence the fitness of diverse viral
(a) (b)
(c)
L
(d)
L
Fig. 5. Electron micrographs of N.
benthamiana leaves 10 days p.i. with: (a)pPVXSma (32 0006), showing the presenceof assembled virus particles (white arrows);(b) pPVXSma-P18DD (16 0006), showingthe presence of both assembled virus parti-cles (white circles) and LICs (L); (c, d)pPVXSma-P18 (60006 and 25 0006,respectively), showing the presence of LICs.Bars, 500 nm.
3110 Journal of General Virology 87
C. Lico and others

genera (i.e. Tobamovirus, Comovirus and Potyvirus), but thatthe effect is different depending on the different role playedby virus particles and coat proteins in the virus life cycle.This is further confirmed by our work that has extended theknowledge base of the effects of peptide display to thefilamentous potexvirus PVX. Our findings, by providingnew insights into the prediction of peptide sequencescompatible with the production of infectious CVPs forcandidate vaccines, reinforce the notion that PVX movesfrom cell to cell and through the phloem in two structurallydifferent forms (i.e. cell-to-cell movement as a ribonucleo-proteic complex and systemic movement as assembled virusparticles) (Lough et al., 1998, 2000).
From the current and prior studies, we can conclude thatinformation derived from peptide-display technologyapplied to viruses of different phylogenetic origins hasimportant implications both to produce candidate vaccinesand to deepen knowledge of virus biology.
ACKNOWLEDGEMENTS
This work was partially supported by grant FIRB-MIURRBNE01N9EE_003, project ‘Development of novel strategies forcancer vaccines’. We thank A. Giovanetti, M. T. Mancuso and A.Polverari for their support in electron microscopy experiments andM. E. Bitti for helpful technical assistance.
REFERENCES
Atabekov, J. G., Rodionova, N. P., Karpova, O. V., Kozlovsky, S. V.,Novikov, V. K. & Arkhipenko, V. (2001). Translational activation ofencapsidated potato virus X RNA by coat protein phosphorylation.Virology 286, 466–474.
Baratova, L. A., Grebenshchikov, N. I., Dobrov, E. N. & 7 otherauthors (1992a). The organization of potato virus X coat proteins invirus particles studied by tritium planigraphy and model building.Virology 188, 175–180.
Baratova, L. A., Grebenshchikov, N. I., Shishkov, A. V., Kashirin, I. A.,Radavsky, Y. L., Jarvekulg, L. & Saarma, M. (1992b). Thetopography of the surface of potato virus X: tritium planigraphyand immunological analysis. J Gen Virol 73, 229–235.
Baratova, L. A., Fedorova, N. V., Dobrov, E. N. & 7 other authors(2004). N-terminal segment of potato virus X coat protein subunitsis glycosylated and mediates formation of a bound water shell on thevirion surface. Eur J Biochem 271, 3136–3145.
Baulcombe, D. C., Chapman, S. & Santa Cruz, S. (1995). Jellyfishgreen fluorescent protein as a reporter for virus infections. Plant J 7,1045–1053.
Bendahmane, M., Koo, M., Karrer, E. & Beachy, R. N. (1999).Display of epitopes on the surface of tobacco mosaic virus: impact ofcharge and isoelectric point of the epitope on virus-host interactions.J Mol Biol 290, 9–20.
Borisova, G., Borschukova, O., Skrastina, D., Dislers, A., Ose, V.,Pumpens, P. & Grens, E. (1999). Behavior of a short preS1 epitopeon the surface of hepatitis B core particles. Biol Chem 380, 315–324.
Brennan, F. R., Jones, T. D., Longstaff, M., Chapman, S., Bellaby, T.,Smith, H., Xu, F., Hamilton, W. D. & Flock, J. I. (1999).Immunogenicity of peptides derived from a fibronectin-binding
protein of S. aureus expressed on two different plant viruses. Vaccine
17, 1846–1857.
Chapman, S., Kavanagh, T. & Baulcombe, D. (1992a). Potato virus
X as a vector for gene expression in plants. Plant J 2, 549–557.
Chapman, S., Hills, G., Watts, J. & Baulcombe, D. (1992b). Mutational
analysis of the coat protein gene of potato virus X: effects on virion
morphology and viral pathogenicity. Virology 191, 223–230.
Chen, Y.-T., Scanlan, M. J., Sahin, U. & 7 other authors (1997). A
testicular antigen aberrantly expressed in human cancers detected
by autologous antibody screening. Proc Natl Acad Sci U S A 94,
1914–1918.
Donini, M., Lico, C., Baschieri, S., Conti, S., Magliani, W., Polonelli, L.
& Benvenuto, E. (2005). Production of an engineered killer peptide
in Nicotiana benthamiana by using a Potato virus X expression
system. Appl Environ Microbiol 71, 6360–6367.
Fedorkin, O. N., Solovyev, A. G., Yelina, N. E., Zamyatnin, A. A., Jr,
Zinovkin, R. A., Makinen, K., Schiemann, J. & Morozov, S. Yu.
(2001). Cell-to-cell movement of potato virus X involves distinct
functions of the coat protein. J Gen Virol 82, 449–458.
Hurwitz, J. L., Slobod, K. S., Lockey, T. D., Wang, S., Chou, T.-H. W.& Lu, S. (2005). Application of the polyvalent approach to HIV-1
vaccine development. Curr Drug Targets Infect Disord 5, 143–156.
Kimalov, B., Gal-On, A., Stav, R., Belausov, E. & Arazi, T. (2004).Maintenance of coat protein N-terminal net charge and not primary
sequence is essential for zucchini yellow mosaic virus systemic
infectivity. J Gen Virol 85, 3421–3430.
Kirkin, A. L., Dzhandzhugazyan, K. & Zeuthen, J. (1998). Melanoma-
associated antigens recognized by cytotoxic T lymphocytes. APMIS
106, 665–679.
Koenig, R. & Torrance, L. (1986). Antigenic analysis of potato virus
X by means of monoclonal antibodies. J Gen Virol 67, 2145–2151.
Kozlovsky, S. V., Karpova, O. V., Arkhipenko, M. V., Zayakina, O. V.,
Rodionova, N. P. & Atabekov, I. G. (2003). Effect of the N-terminal
domain of the coat protein of Potato virus X on the structure of viral
particles. Dokl Biochem Biophys 391, 189–191.
Lee, A. G. (2003). Lipid–protein interactions in biological mem-
branes: a structural perspective. Biochim Biophys Acta 1612, 1–40.
Lesemann, D. E. (1988). Cytopathology. In The Plant Viruses: The
Filamentous Plant Viruses, pp. 179–235. Edited by R. G. Milne. New
York: Plenum.
Lough, T. J., Shash, K., Xoconostle-Cazares, B., Hofstra, K. R.,
Beck, D. L., Balmori, E., Forster, R. L. S. & Lucas, W. J. (1998).
Molecular dissection of the mechanism by which potexvirus triple
gene block proteins mediate cell-to-cell transport of infectious RNA.
Mol Plant Microbe Interact 11, 801–814.
Lough, T. J., Netzler, N. E., Emerson, S. J., Sutherland, P., Carr, F.,
Beck, D. L., Lucas, W. J. & Forster, R. L. S. (2000). Cell-to-cell
movement of potexviruses: evidence for a ribonucleoprotein
complex involving the coat protein and the first triple gene block
protein. Mol Plant Microbe Interact 13, 962–974.
Marusic, C., Rizza, P., Lattanzi, L., Mancini, C., Spada, M., Belardelli, F.,
Benvenuto, E. & Capone, I. (2001). Chimeric plant virus particles as
immunogens for inducing murine and human immune responses
against human immunodeficiency virus type 1. J Virol 75, 8434–8439.
Parker, L., Kendall, A. & Stubbs, G. (2002). Surface features of
potato virus X from fiber diffraction. Virology 300, 291–295.
Pogue, G. B., Lindbo, J. A., Garger, S. J. & Fitzmaurice, W. P. (2002).Making an ally from an enemy: plant virology and the new
agriculture. Annu Rev Phytopathol 40, 45–74.
Polonelli, L., Magliani, W., Conti, S., Bracci, L., Lozzi, L., Neri, P.,Adriani, D., De Bernardis, F. & Cassone, A. (2003). Therapeutic
http://vir.sgmjournals.org 3111
Peptide features affecting mutant PVX CVP fitness

activity of an engineered synthetic killer antiidiotypic antibody
fragment against experimental mucosal and systemic candidiasis.
Infect Immun 71, 6205–6212.
Porta, C., Spall, V. E., Findlay, K. C., Gergerich, R. C., Farrance, C. E.& Lomonossoff, G. P. (2003). Cowpea mosaic virus-based chimaeras.
Effects of inserted peptides on the phenotype, host range, and
transmissibility of the modified viruses. Virology 310, 50–63.
Salzwedel, K., West, J. T. & Hunter, E. (1999). A conserved tryptophan-
rich motif in the membrane-proximal region of the human
immunodeficiency virus type 1 gp41 ectodomain is important for
Env-mediated fusion and virus infectivity. J Virol 73, 2469–2480.
Santa Cruz, S., Chapman, S., Roberts, A. G., Roberts, I. M., Prior,D. A. M. & Oparka, K. J. (1996). Assembly and movement of a plant
virus carrying a green fluorescent protein overcoat. Proc Natl Acad
Sci U S A 93, 6286–6290.
Santa Cruz, S., Roberts, A. G., Prior, D. A. M., Chapman, S. &Oparka, K. J. (1998). Cell-to-cell and phloem-mediated transport of
potato virus X: the role of virions. Plant Cell 10, 495–510.
Scala, G., Chen, X., Liu, W., Telles, J. N., Cohen, O. J., Vaccarezza, M.,Igarashi, T. & Fauci, A. S. (1999). Selection of HIV-specific
immunogenic epitopes by screening random peptide libraries withHIV-1-positive sera. J Immunol 162, 6155–6161.
Scholthof, H. B., Scholthof, K.-B. G. & Jackson, A. O. (1996). Plantgene virus vectors for transient expression of foreign proteins inplants. Annu Rev Phytopathol 34, 299–323.
Shalla, T. A. & Shepard, J. F. (1972). The structure and antigenicanalysis of amorphous inclusion bodies induced by potato virus X.Virology 49, 654–667.
Shan, L., Molberg, Ø., Parrot, I., Hausch, F., Filiz, F., Gray, G. M.,Sollid, L. M. & Khosla, C. (2002). Structural basis for glutenintolerance in celiac sprue. Science 297, 2275–2279.
Shanmugam, G., Polavarapu, P. L., Kendall, A. & Stubbs, G. (2005).Structures of plant viruses from vibrational circular dichroism. J GenVirol 86, 2371–2377.
Shiver, J. W. & Emini, E. A. (2004). Recent advances in thedevelopment of HIV-1 vaccines using replication-incompetentadenovirus vectors. Annu Rev Med 55, 355–372.
Tangri, S., Mothe, B. R., Eisenbraun, J. & 9 other authors (2005).Rationally engineered therapeutic proteins with reduced immuno-genicity. J Immunol 174, 3187–3196.
3112 Journal of General Virology 87
C. Lico and others





























