Embed Size (px)
Citation preview

Arch Virol (1989) 104:287-298 Archives
Vifrology © by Springer-Verlag 1989
Phagocytosis of herpes simplex virus by human granuloeytes and monocytes
J. A. G. Van Strijp, K. P. M. Van Kessel, Marijke E. van der Tol, A. C. Fluit, H. Snippe, and J. Verhoef
Laboratory for Microbiology, University of Utrecht, Utrecht, The Netherlands
Accepted October 25, 1988
Summary. Polymorphonuclear leukocytes (PMN) can mediate cytotoxic reac- tions against virus infected targets cells. We observed very efficient binding of PMN to HSV-infected fibroblasts when loaded with HSV-specific antibodies. Using electron microscopy, infected fibroblasts were found to be totally sur- rounded by PMN and the phagocytosis of virions and fragments of infected cells was demonstrated. To quantify and study this phenomenon, and to com- pare PMN with monocytes, we developed radiometric and fluorometric phago- cytosis assays. Leukocytes were mixed with [3H]glucosamine- or FITC-labeled virus and incubated at 37 °C. PMN associated radioactivity or fluorescence per cell as measured by flow cytometry was determined. PMN phagocytosis was dependent on the presence of specific anti-HSV antibodies and could be en- hanced by addition of complement. Monocytes were also able to phagocytize virions; however, the rate of uptake was less than that for PMN. Under optimal conditions the total amount of herpes simplex particles that could be associated with one PMN or monocyte was about 10,000.
PMN and monocytes are capable of phagocytosis of HSV. This may be an important factor in preventing the spread of infection in vivo.
Introduction
Granulocytes (polymorphonuclear leukocytes, PMN) are the first leukocytes to arrive in herpes simplex lesions [11, 13].
Several researchers have shown that PMN are able to bind to virus infected cells [9, 16, 27] and to internalize virus particles [23]. Although granulocytes are able to lyse HSV-infected cells in vitro, the high effector to target ratios and the long incubation periods required cast some doubt on the role of this process in vivo [14, 18, 21]. Furthermore, the observation that PMN phago- cytize virions or fragments of infected cells has not been well established in vivo nor has the efficacy of this process to halt viral spread been compared to

288 J . A . G . Van Strijp et al.
classical neutral izat ion by antibodies and complement [ 12, 21, 31]. We therefore studied in greater detail the phagocytosis of HSV by P M N and monocytes .
Elect ron microscopy o f P M N and infected fibroblasts in the presence o f antibodies and complement was used to conf i rm that phagocytosis occurs. A quant i ta t ive radiometr ic phagocytosis assay for HSV was developed as a mod- ification o f our s tandard assay [30]. To quant i fy phagocytosis on a per cell basis we developed a f luorometr ic assay based on analysis by a flow cy tomete r [-1, 24]. The latter technique provided an efficient means to s tudy viral c learance by P M N and monocytes .
Materials and methods
Chemicals"
RPMI 1640, Medium 199, fetal calf serum (FCS) and gentamicin were obtained from Gibco (Paisley, Renfrewshire, U.K.), Hepes (N-2-hydroxyethylpiperazine-N'-2-ethane-sulphonic acid) was purchased from BDH Chemicals Ltd (Poole, Dorset, U.K.), [3H]glucosamine (specific activity 138 mCi/mg) from Amersham International (Bucks, U.K.), and dextran T-70, T-10, and Ficoll-Paque from Pharmacia (Pharmacia Fine Chemicals AB, Uppsala, Sweden). Fluorescein-isothyocyanate (FITC), Isomer I from Sigma (St. Louis, MO, U.S.A.). Anti-Leu-M 3 and anti-mouse kappa, phycoerythrin conjugated were purchased from Bec- ton Dickinson (Mountain View, CA, U.S.A.).
Cells and viruses
Human embryonic fibroblasts were established in our own laboratory and HEp-2 cells obtained from Flow (Irvine, U.K.). The cells were cultured in RPMI 1640/Medium 199 (1 : 1, v/v), supplemented with 10% FCS, 5 mM Hepes, 2 g/1 sodium bicarbonate, and I0 gg/ mt gentamicin. Herpes simplex virus type 1 (HSV-1), strain F, was a gift from Dr. B. Roizman (University of Chicago, Chicago, IL).
For determination of virus titer, serial tenfold dilutions were made and each dilution was mixed with 10 4 fibroblasts per well of a microtiter plate. After six days each well was scored for the presence of cytopathic effect and the 50% tissue infective dose (TCIDs0) was calculated.
Sera
Sera from healthy volunteers were screened for HSV specific antibodies in an ELISA and in an antibody dependent cell mediated cytotoxicity assay (ADCC) with killer cells, both as described previously [29]. Sera with high levels of specific anti HSV-antibodies were pooled and stored at --70 °C. All sera were tested in a 50% hemolytic complement activity assay and only those with normal complement levels were used. To deplete complement activity, sera were heated for 30 minutes at 56 °C.
IgG was purified from these sera by using high performance liquid chromatography (HPLC) on a ABx-column (J. T. Baker Chemical company, Phillipsburg, NJ, U.S.A.). Fab fragments were prepared by papain digestion, and non-digested IgG and Fc fragments were removed by protein A affinity-chromatography.
Isolation of human PMN and monocytes
Polymorphonuclear leukocytes (PMN) and monocytes were isolated according to a modified method of Boyum as described earlier [27, 28]. Briefly, heparinized human blood was mixed with dextran T-70 to a final concentration of 1.2% and sedimented for 60 minutes.

Phagocytosis of HSV 289
Leukocyte-rich plasma was withdrawn and leukocytes were separated on Ficoll-Paque. Residual erythrocytes in the PMN fraction were lysed by hpotonic shock. The interface layer of mononuclear cells was washed and incubated for 60 minutes on autologous plasma coated Petri dishes. Nonadherent cells were removed and adherent cells, in which monocytes were highly enriched (80-90% purity) were harvested using EDTA.
Virus purification and labeling
Radiolabeled virus
HEp-2 cells were grown to confluence on 15 cm culture dishes (Greiner, Nurtingen, Federal Republic of Germany) and infected with HSV-1 (multiplicity of infection was 20). After one hour medium with residual virus was removed and fresh medium was added. After 3 to 4 hours the medium was replaced by medium containing 80 gCi [3H]glucosamine per milliliter and incubated for an additional 20 hrs. Cells were harvested by scraping, washed and then lysed in a Dounce homogenizer (Wheaton Scientific, Milville, NJ, U.S.A.) by 20 strokes with each pestle. Cell fragments and nuclei were pelleted at 500 x g and the cytosol was dialyzed to remove excess non-incorporated radioactivity. One ml was layered on top of a block-gradient consisting of 5 ml 2.5% and 5 ml 25% dextran-10, a modification of the technique described by Cassai [5]. Gradients were centrifuged at 100,000 x g for two hours at 4 °C. Virus bands were aspirated and dextran was removed by dialysis against PBS in Spectrum/Por 6 dialysis membranes with a molecular weight cutoff of 50,000 (Spec- trum Medical Industries Inc., Los Angeles, CA, U.S.A.). Virus was stored at --70 °C in small aliquots. Purity was checked by electron microscopy and it was shown that over 95% of the material was enveloped virus.
FITC labeled virus
Virus was purified as described above without the addition of [3H]glucosamine. Purified virus was mixed with FITC (1 mg/ml in 1 M sodium carbonate buffer pH 9.6), and incubated at room temperature for one hour under constant rotation. After dialysis against PBS for 18 hours at 4 °C, fluorescein labeled virus was stored at --70 °C in small aliquots until use.
Electron microscopy
Virus infected fibroblasts were washed and incubated with PMN in the presence of anti- bodies. Cells were fixed with 3% glutaraldehyde, suspended in agarose, fixed with 1% OsO4, dehydrated and embedded in EPON. Specimens were cut using a diamond knife and contrasted using lead-citrate according to Reynolds and uracil-magnesium-acetate. Coupes were examined and photographed in a Zeiss EM 109 electron microscope. Photos examined contained more then 200 PMN and 40 infected fibroblasts.
Radiometric phagocytosis assay
100 ~tl PMN [3 x 107/ml Hanks' balanced salt solution supplemented with 5% FCS (HBSSf)] were mixed with 50 ~tl purified radiolabeled virus and 50 ~tl human serum (diluted in HBSSf). The cells were incubated for various time periods in a shaking water bath (150rpm), the reaction was stopped by adding ice-cold PBS. PMN were separated from free virus by centrifugation at 160 x g and washed thrice. Samples were solubilized in scintillation cocktail (aqualuma plus, Lumac, Landgraaf, the Netherlands) and counted in a liquid scintillation counter (PW 4800, Philips, Almelo, The Netherlands). Percentage phagocytosis was ex- pressed as:
measured radioactivity x 100%
total added radioactivity

290 J .A.G. Van Strijp et al.: Phagocytosis of HSV
Fluorometric phagocytosis assay
60 gl PMN (3 × 107/ml) in HBSSf were mixed with 30 ~tl purified FITC-labeled virus and 30 gl human serum (diluted in HBBSf). After incubation for various time periods in a shaking water bath (150rpm), the reaction was stopped by adding ice-cold PBS. PMN were separated from free virus by centrifugation at 160 × g, washed twice and the cells were suspended in 0.5ml PBS and fixed by adding 0.5ml 1% paraformaldehyde. Cells were analyzed in a FACStar flow cytometer (Becton Dickinson, Mountain View, CA, U.S.A.) with an excitation wavelength of 488 nm. Phagocytosis was expressed as percentage relative to the maximum observed phagocytosis using 20% human anti HSV-serum. To study phagocytosis by monocytes, lymphocyte contamination of adherent cells was inevitable. To discriminate between non phagocytic lymphocytes and monocytes, cells were fixed by adding 0.5% paraformaldehyde after the phagocytosis assay. The cells were subsequently incubated with a mouse anti LeuM 3 monoclonal antibody for 30 minutes.
After washing cells were incubated with an anti-mouse Kappa antibody conjugated with phycoerythrin, washed and measured. Only red fluorescent cells were analyzed and phagocytosis was measured in the same way as with granutocytes.
Statistical analysis
Results are expressed as the mean of three or more independent observations ± standard error of the mean.
Results
Electron microscopy
The interaction between granulocytes and HSV infected fibroblasts was studied by electron microscopy. We focused on the phagocytosis of HSV by P M N in the presence of antibodies. Figure 1 A shows a human fibroblast infected with HSV-1 for 20 hours then coated with antibodies and incubated with human PMN. Several P M N adhere to the infected fibroblast. Free virions are located mainly in the space between P M N and infected fibroblast. The infected fibro- blast is still intact and has the same morphology as fibroblasts infected with HSV and incubated for the same time period in the absence of PMN. Phag- ocytosis of virions is more evident in Fig. 1 B, C, where the ingestion stage of free virions and large immune complexes containing many virions is shown. When serum was present more then eight3, percent of the adhering P M N con- tained virions in phagosomes, and in 20% of the P M N more then 40 virions could be observed in one coupe. Membrane vesicles, probably from the plasma membrane of infected cells, which possess the same antigens as the virions are also invaginated. No discrimination can be made between degraded HSV par-
Fig. 1A-D. HSV infected fibroblasts were incubated with PMN in the presence of both HSV-specific antibodies and complement for 30 minutes at 37 °C. A An infected fibroblast is surrounded by several PMN. Good adherence is obvious and there are phagocytic vacuoles within the PMN. Internalization occurs of single virions (B) and of larger immune complexes (C). D In the phagosome there are three intact herpes virions and several membrane fragments. Also phagosome-lysosome fusion is evident. The bar represents 0.5 lam, respec-
tively
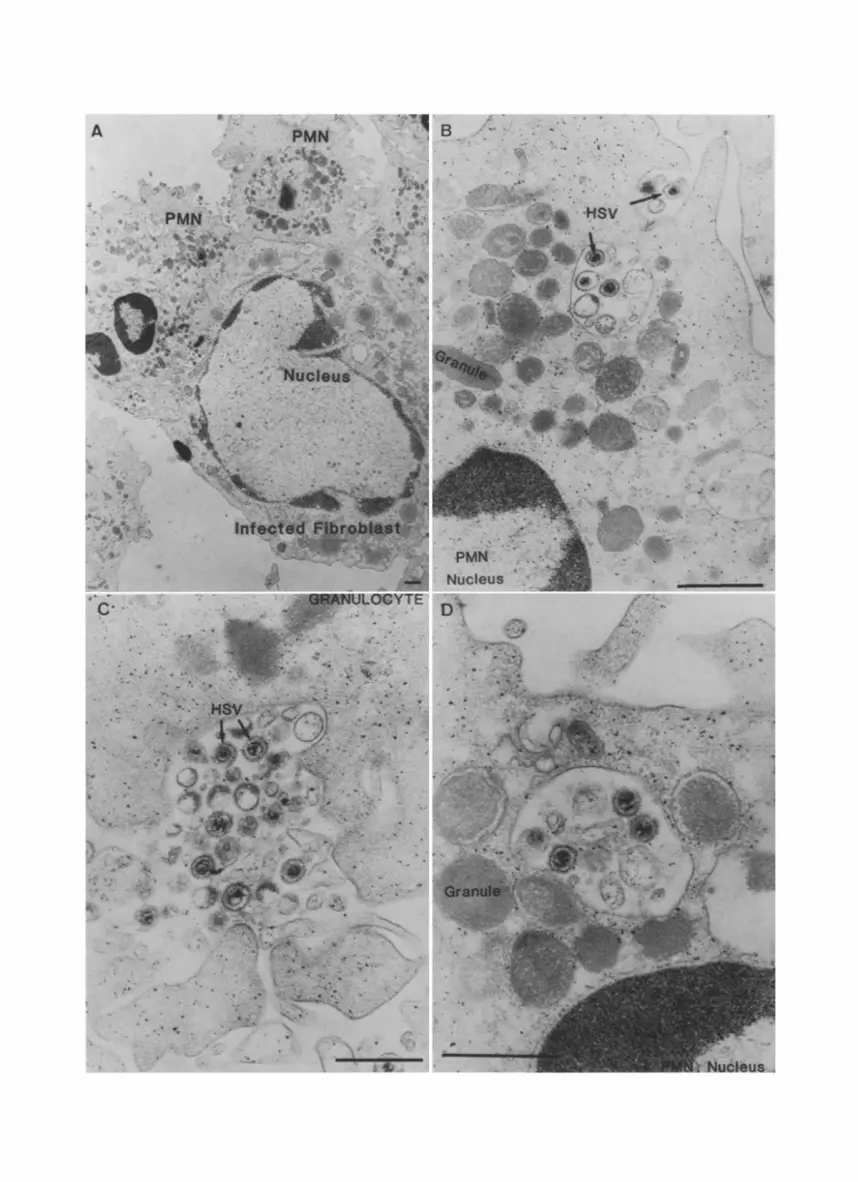

292 J . A . G . Van Strijp et al.
ticles and the smaller damaged membrane vesicles of about the same size as the virions. Although phagosome-lysosome fusion can be observed in Fig. t D, and an overall breakdown of phagocytized material after longer incubations is obvious, it is hard to determine whether the virions are degraded or not. Virions inside the PMN are only observed in the vacuoles, and never free in the cy- toplasm or in the PMN-nucleus.
Phagocytosis using FH]glucosamine labeled virions
Figure 2 shows the kinetics of uptake of herpes simplex virions in the presence of both HSV-specific antibodies and human complement (both 20%). Within 15 minutes a maximum uptake of over 70% was reached. When no antibodies or complement was present, uptake was always less then 10%. Figure3 shows
80
60
U P 4o T A K E 20
~ I . . . . .~ ANTISERUM
~ HBSSf
7.5 15 30 60 120 TIME (rain)
Fig. 2. PMN were incubated with radiolabeted herpes virions in the presence of 20% human anti-HSV serum or the buffer control (HBSSJ), for various time periods. Uptake by PMN
was expressed as a percentage of total added radioactivity
80
60
U P 40 T A K E ZO
o o
~_._]~ WITH COMPLEMENT
HEATED
o'., ; 2'.5 ; /o 2'0 PERCENTAGE SERUM
Fig. 3. PMN were incubated with radiolabeled herpes virions and different concentrations of human anti-HSV serum containing intact human complement (WITH COMPLEMENT) or the same serum which was incubated for 30 minutes at 56°C (HEATED). After 30 minutes the reaction was stopped and leukocyte associated radioactivity was determined

Phagocytosis of HSV 293
that phagocytosis depends on antibody concentration. The presence of an intact human complement system enhances the uptake of virions markedly (over 70% uptake).
Phagocytosis using FITC labeled virions
To study the efficiency ofmonocytes and granulocytes in the clearance of virions, a fluorometric method was developed. Figure 4 shows that the time curves for human PMN phagocytosis in the presence of specific antibodies with or without complement were comparable with those measured with the radiometric assay. Without antibodies or complement no uptake was observed. For monocytes the uptake process proceeds over a longer period of time. Spontaneous uptake of virions by monocytes was much higher than that by granulocytes.
Figure 5 shows that the use of heated serum gives the same results as an equal amount of purified IgG. Fab-fragments of this IgG, in comparable molar concentrations did not promote phagocytosis. When the experiments were per- formed at 4 °C no uptake was observed.
The maximum amount of virions that can be taken up by both types of phagocytes was evaluated. Different amounts of virions were incubated with either monocytes or PMN. In Fig. 6 the results are expressed as mean fluores- cence channel of the histogram obtained when fluorescence is plotted against number of cells. A representative experiment is shown with PMN and monocytes from the same donor, performed on the same day. Above 200 pl of added virus suspension a saturation of both PMN and monocytes was reached. Equal amounts of fluorescent virions could be taken up by granulocytes and by monocytes when both complement and antibodies were present.
R E L A T I V E
g
U P T A K E
125 I GRANULOCYTES 100 ~.
7.5 15 3o 60
1251~ tl MONOCYTES [ ~
7.5 15 30 60 120
[ ] ANTISERUM WITH COMPLEMENT
[] HEATED ANTISERUM
• HBSS
TIME (rain)
Fig. 4. Granulocytes and monocytes were compared in their ability to phagocytize FITC- labeled herpes virions, in the presence of 20% human anti HSV serum with or without complement. At various time intervals leukocytes were analyzed in a FACS. The mean fluorescence channel was calculated and the percentage of uptake was expressed relative to the uptake by PMN after 30 minutes in the presence of 20% antiserum with complement
(lOO%)

100.
20
R E L A 75 T 1 V E
50 %
U P 25. T A K E o
MEAN FLUORESCENCE CHANNEL
1000
BOO
600
400
200
O--
20 % IgG IgG IgG IgG heated 2 1 0 .5 0 .25
mg/r.1
Fob hbee 20 1.4 4°C
Fig. 5. Phagocytosis of FITC labeled virus by human PMN. Purified IgG was used in concentrations comparable to that in 20% human serum (2 mg/ml) and two step dilutions of this concentration. Fab fragment of the same IgG were also present in the same molar concentration (1.4 mg/ml). 20% 20% human anti-HSV serum. 4 °C The phagocytosis assay
was performed under the same conditions but at 4 °C
294 J .A.G. Van Strijp et at.
PkiN + SERUM
MN + SERUki
kiN + HBSS
PkiN + HBSS
0 50 I O0 150 2oo 250 300 ul PURIFIED VIRUS
Fig. 6. Different amounts of virus were incubated with PMN or monocytes with serum containing both HSV-specific antibodies and complement. The mean fluorescence channel
was calculated after FACS analysis
Estimation of number of viruses per PMN
When antibodies and complement are present in concentrations higher then in 5% serum, 80% of the radiolabeled viruses were taken up by P M N after 45 minutes of phagocytosis (Figs. 2 and 3). The titer of the purified virus was 2 x 109 plaque forming units per ml; less than 1% of these virions were infectious. In Fig. 6 we observed that with 200 ~tt of this virus suspension, saturation of the P M N is still not reached. This means that of 4 x 10 l° virus particles added, at least 3 x 10 l° can be taken up by 3 x 106 P M N or monocytes. Thus there must be at least 10,000 viruses present in each cell. This is in agreement with estimations based on our electron microscopic study.

Phagocytosis of HSV 295
Discussion
As Kirchner stated in 1982: "Granulocytes certainly are the stepchildren among the cells of the lymphoreticular system in regard to that is known about their role in the defense against viruses" [11]. The role of phagocytosis has long been underestimated in antiviral defense. Only recently some publications have appeared concerning this phenomenon. For influenza virus it was shown by means of chemiluminescence that PMN are capable of phagocytosis of virions [8, 26]. West, using radiolabeled virions, showed that vaccinia virus can be taken up by PMN [33] and Smith showed that radiolabeled immune complexes containing Herpes Simplex virions were bound to granulocytes that adhered to coverslip [23].
Several factors point to a role for PMN in the defense against viruses. First, PMN are abundantly present in herpes lesions [1 l, 13] and bind effectively to infected cells. They are therefore, the first to encounter the newly freed virions. Secondly, most viruses are not able to replicate inside PMN [8, 11], probably because of the very low level of protein and DNA synthesis in PMN [10]. Furthermore, when virus infected cells are lysed in a late stage of infection by complement or killer cells, large amounts of infective virus can be liberated into the host [4]. We were interested in the capacity of PMN to phagocytize and neutralize viruses. In earlier studies we showed that human granulocytes adhere to infected fibroblasts [27, 29]. In our observations and those in the literature, the in vivo relevance of the killing of HSV-infected cells by PMN is not yet clear [t4, 18, 21]. In the present study we extended this work by looking at the process of phagocytosis using electron microscopy, radiometry, and flow cytometry.
Electron microscopy showed that PMN adhered efficiently to infected fi- broblasts. The fibroblasts were totally surrounded by PMN, with a large area of contact. No events that were related to cytotoxic processes were observed: no swelling of the periplasmatic space or endoplasmic reticulum and no visible damage to the fibroblast membrane. Nor was fusion of membranes between PMN and fibroblast, degranulation or polarization of the PMN [6, 7, 15] observed. However, the PMN surrounding the HSV infected cells contained vacuoles loaded with virions. Close examination using larger magnifications revealed adherence and ingestion as well as phagosome-lysosome fusion. Once phagocytosis has occurred and phagosome-lysosome fusion takes place it is likely that virus particles are degraded due to the antiviral effects of lysosome constituents [ 19, 20, 25, 36]. Also, PMN elicit a respiratory burst when exposed to HSV and antibodies [3] whereby reactive oxygen species liberated by the PMN must be able to degrade virions [2].
PMN, therefore, are good candidates for viral clearance by means of phag- ocytosis. To quantify this, we developed a radiometric assay. With this assay we were able to show that uptake of virus by PMN depended on antibody concentration and incubation time. There was a drastic enhancement of uptake when complement was added. This can be explained by both the formation of

296 J .A.G. Van Strijp et al.
larger immune complexes, as measured by Smith [23] and by better adhesion as caused by complement receptor binding [35].
The fluorometric assay, although easier to perform, did not offer the pos- sibility to quantify the uptake as a percentage of the added material. It was, however, a very useful technique for comparing different cells, different sera or different time points. This is demonstrated by the kinetic studies done with monocytes versus PMN. Here we showed that monocytes are slower in their capacity of virus uptake. The amount of virions that can associate with mon- ocytes or PMN is between 5000 and 10,000. Thus it appears that monocytes, the classic mediators of immune complex removal, can not remove more virus than PMN. Monocytes can, however, bind virions in the absence of antibodies and/or complement. This may be caused by a stimulating effect of contaminating lymphocytes or NK-cells or by binding of HSV to a receptor present on mon- ocytes and not on granulocytes. We found that decreasing the number of non adherent cells did not diminish this spontaneous binding. However, we were not able to remove all the contaminating cells. In the electron micrographs the virions that were ingested could be emanating from fibroblasts lysed either by the virus itself or by the PMN. We cannot, however, rule out the possibility that the virions emerge by budding through the plasma membrane from the infected fibroblast [34]. This is underscored by the observation that the side of the PMN that contained most, if not all virions was facing and partially adhering to the fibroblast, so that probably all virions originated from the fibroblast.
We conclude that PMN are effective mediators of viral clearance in vitro and that it will be of importance to continue this study in vivo. Especially since no correlation was found between the in vivo protection of antibodies and the lack in vitro of neutralization and Fc-receptor mediated processses other than phagocytosis [17-1.
Acknowledgements
We thank Ingrid Voss and Edith Peters for the excellent care of the cell lines, Prof. H. Geuze for the possibility of performing the electron microscopic studies, and John Kovarik for reviewing the manuscript.
References
I. Bassoe CF, Laerum OD, Solberg CO, Haneberg B (1983) Phagocytosis of bacteria by human leukocytes measured by flow cytometry. Proc Soc Exp Biol Med 174:182-t86
2. Belding ME, Klebanoff SJ, Ray CG (1970) Peroxidase mediated virucidal systems. Science 167:195-196
3. Bingham EL, Fenger TW, Sugar A, Smith JW (t985) Dependence on antibody for induction of chemiluminescence in polymorphonuclear leukocytes by herpes simplex virus. Invest Ophtamol Vis Sci 26:1236-1243
4. Brier AM, Wholenberg C, Rosenthal J, Mage M, Notkins L (1971) Inhibition or enhancement of immunological injury of virus infected cells. Proc Natl Acad Sci USA 68 : 3073-3077
5. Cassai EN, Sarmiento M, Spear P (1975) Comparison of the virion proteins specified by herpes simplex virus types 1 and 2. J Virol 16:1327---1331

Phagocytosis of HSV 297
6. Daniels CA, Bodner S, Trofatter KF (1980) Scanning and transmission electron mi- croscopic studies of complement-mediated lysis and antibody-dependent cell-mediated cytolysis of herpes simplex virus-infected human flbroblasts. Am J Pathol 100: 663- 682
7. Frey T, Petty HR, McConnel HM (1982) Electron microscopic study of natural killer cell-tumor cell conjugates. Proc Natl Acad Sci USA 79:5317-5321
8. Fujisawa H, Tsuru S, Taniguchi M, Zinnaka Y, Nomoto K (1987) Protective mech- anisms against pulmonary infection with influenza virus. I. Relative contribution of polymorphonuclear leukocytes and of alveolar macrophages to protection during the early phase of intranasal infection. J Gen Virol 88: 425~432
9. Grewal AS, Carpio M, Babuik LA (1980) Polymorphonuclear neutrophil-mediated antibody dependent cell cytotoxicity of herpesvirus-infected cells: ultrastructurat stud- ies. Can J Microbiol 26:ig: 427435
10. Jack RM, Fearon DT (1988) Selective synthesis of mRNA and proteins by human peripheral blood neutrophils. J Immunology 140:4288-4293
11. Kirchner H (1982) Immunobiology of infection with herpes simplex virus. In: Melnick JL (ed) Monographs in virology, vo113. S Karger, Basel, p 39
12. McKendall RR (1985) IgG-mediated viral clearance in experimental infection with Herpes Simplex virus type 1 : role for neutralization and Fc-dependent functions but not C' cytolysis and C 5 chemotaxis. J Infect Dis 151 : 464-470
13. Meyers-Elliott RH, Chitjian PA (1981) Immunopathogenesis of corneal inflammation in herpes simplex virus stromal keratitis: role of the polymorphonuclear leukocyte. Invest Ophtamol Vis Sci 20:784-798
14. Oleske JM, Ashman RB, Kohl S, Starr SE, Wood Ph, Nahmias AJ (1977) Human polymorphonuclear leukocytes as mediators of antibody-dependent cellular cytotoxicity to herpes simplex virus-infected cells. Clin Exp Immunol 27:446-453
15. Patterson S, Byrne JA, Lampert PW, Oldstone MBA (t987) Morphologic analysis of the interactions between lymphocytic choriomeningitis virus-specific cloned cytotoxic T cells and virus infected targets. Lab Invest 57:29-36
16. Penfold PL, Greenberg AH, Roitt IM (1976) Characteristics of the effector cells me- diating cytotoxicity against antibody-coated targets. III. Ultrastructural studies. Clin Exp Immunol 23:91-97
17. Rector JT, Laush RN, Oakes JE (1982) Use of monoclonal antibodies for analysis of antibody-dependent immunity of ocular Herpes Simplex virus type 1 infection. Infect Immun 38:168-174
18. Rouse BT, Wardly RC, Babuik LA (1976) The role of antibody-dependent cytotoxicity in recovery from herpesvirus infections. Cell Immunol 22:182-186
19. Rouse BT, Babuik LA, Henson PM (1980) Neutrophils in antiviral immunity: inhibition of virus replication by a mediator produced by bovine neutrophils. J Infect Dis 141 : 223-232
20. Selsted ME, Harwig SSL (1987) Purification, primary structure, and antimicrobial activities of a guinea pig neutrophil defensin. Infect Immun 55:2281-2286
21. Siebens H, Tevethia SS, Babior BM (1979) Neutrophil-mediated antibody-dependent killing of herpes simplex virus-infected cells. Blood 54:88--94
22. Simmons A, Nash A (1985) Role of antibody in primary and recurrent herpes simplex infection. J Virol 53:944-948
23. Smith JW, Jachimowicz JR, Bingham EL (1986) Binding and internalization of Herpes Simplex Virus-antibody complexes by polymorphonuclear leukocytes. J Med Vir 20: 151-163
24. Steinkamp JA, Wilson JS, Saunders GC, Stewart CC (1982) Phagocytosis: flow cy- tometric quantitation with fluorescent microspheres. Science 215:64-66
25. Thorne KJI, Norman JM, Haydock SF, Laminas DA, Duffus PH (1984) Antibody-

298 J .A .G. Van Strijp et al. : Phagocytosis of HSV
dependent celt-mediated cytotoxicity against IBR-infected bovine kidney cells by ru- minant neutrophils: the role of lysosomal cationic protein. Immunology 53:275-282
26. Tsuru S, Fujisawa H, Taniguchi M, Zinnaka Y, Nomoto K (1987) Mechanism of protection during the early phase of a generalized viral infection. II. Contribution of polymorphonuclear leukocytes to protection against intravenous infection with influ- enza virus. J Gen Virol 68:419-424 Van Kessel KPM, Van Kats-Renaud H J, Van Strijp JAG, Visser MR, Verhoef J (1986) Measurement of antibody-mediated binding of human polymorphonuclear leukocytes to HSV-1 infected anchorage fibroblasts. J Immunol Methods 88:191-107
28. Van Kessel KPM, Visser MR, Van Strijp JAG, Van Kats-Renaud H J, Verhoef J (1986) Cytotoxicity by human adherent Cells: oxygen-dependent and -independent cytotoxic reactions by different cell populations. Immunology 58:291-296
29. Van Strijp JAG, Van Kessel KPM, Miltenburg LAM, Fluit AC, Verhoef J (1988) Attachment of human potymorphonuclear leukocytes to herpes simplex virus-infected fibroblasts mediated by antibody-independent complement activation. J Virol 62: 847- 850
30. Verhoef J, Peterson PK, Quie PG (1977) Kinetics of staphylococcal opsonization, attachment, ingestion and killing by human polymorphonuclear leukocytes: a quan- titative assay using 3H-Thymidine labeled bacteria. J Immunol Methods 14:303 313
31. Wallis C, Melnick JL (1971) Herpesvirus neutralization: The role of complement. J Immunol 107:1235-1242
32. Wardly RC, Rouse BT, Babuik LA (1976) Antibody-dependent cytotoxicity mediated by neutrophils: a possible mechanism of antiviral defense. J Reticuloendoth Soc 19: 323-332
33. West BC, Eschete ML, Cox ME, King JW (1987) Neutrophil uptake of vaccinia virus in vitro. J Infect Dis 156:597-606
34. Witdy P (1986) Herpesvirus. Intervirotogy 25: 117-I 40 35. Wright SD, Griffin FM Jr (1985) Activation of phagocytic cells' C3 receptors for
phagocytosis. J Leukocyte Biol 38:327-339 36. Zerial A, Skerlavaj B, Gennaro R, Romeo D (1987) Inactivation of Herpes Simplex
Virus by protein components of bovine neutrophil granules. Antiviral Res 7: 341 352
27.
Authors' address: J. A. G. Van Strijp, Laboratory for Microbiology, State University of Utrecht, Catharynesingel 59, NL-3511 GG Utrecht, The Netherlands.
Received July 27, 1988





























