Embed Size (px)
Citation preview

J Physiol 588.23 (2010) pp 4769–4783 4769
Phenotypic distinctions between neural crest and placodalderived vagal C-fibres in mouse lungs
Christina Nassenstein1,2, Thomas E. Taylor-Clark1, Allen C. Myers1, Fei Ru1, Rajender Nandigama2,Weston Bettner1 and Bradley J. Undem1
1The Johns Hopkins School of Medicine, 5501 Hopkins Bayview Circle, Baltimore, MD 21224, USA2Institute of Anatomy and Cell Biology, Justus-Liebig-University, Aulweg 123, 35835 Giessen, Germany
Two major types of nociceptors have been described in dorsal root ganglia (DRGs). Incomparison, little is known about the vagal nociceptor subtypes. The vagus nerves providemuch of the capsaicin-sensitive nociceptive innervation to visceral tissues, and are likely tocontribute to the overall pathophysiology of visceral inflammatory diseases. The cell bodies ofthese afferent nerves are located in the vagal sensory ganglia referred to as nodose and jugularganglia. Neurons of the nodose ganglion are derived from the epibranchial placodes, whereasjugular ganglion neurons are derived from the neural crest. In the adult mouse, however, thereis often only a single ganglionic structure situated alone in the vagus nerve. By employingWnt1Cre/R26R mice, which express β-galactosidase only in neural crest derived neurons, wefound that this single vagal sensory ganglion is a fused ganglion consisting of both neuralcrest neurons in the rostral portion and non-neural crest (nodose) neurons in the more centraland caudal portions of the structure. Based on their activation and gene expression profiles,we identified two major vagal capsaicin-sensitive nociceptor phenotypes, which innervated adefined target, namely the lung in adult mice. One subtype is non-peptidergic, placodal inorigin, expresses P2X2 and P2X3 receptors, responds to α,β-methylene ATP, and expressesTRKB, GFRα1 and RET. The other phenotype is derived from the cranial neural crest and doesnot express P2X2 receptors and fails to respond to α,β-methylene ATP. This population can befurther subdivided into two phenotypes, a peptidergic TRKA+ and GFRα3+ subpopulation, anda non-peptidergic TRKB+ and GFRα1+ subpopulation. Consistent with their similar embryonicorigin, the TRPV1 expressing neurons in the rostral dorsal root ganglia were more similar tojugular than nodose vagal neurons. The data support the hypothesis that vagal nociceptorsinnervating visceral tissues comprise at least two major subtypes. Due to distinctions in theirgene expression profile, each type will respond to noxious or inflammatory conditions in theirown unique manner.
(Received 28 June 2010; accepted after revision 6 October 2010; first published online 11 October 2010)Corresponding author B. J. Undem: Johns Hopkins Allergy and Asthma Center, 5501 Hopkins Bayview Circle, Baltimore,MD 21224, USA. Email: [email protected]
Abbreviations DRG, dorsal root ganglion; GFR, GDNF family ligand receptor; NGF, nerve growth factor; PPT-A,preprotachykinin-A; RET, ret proto-oncogene; TRK, tropomyosin-related kinase.
Introduction
Nociceptors derived from dorsal root ganglia (DRGs) arebroadly subcategorized into two general subtypes, referredto as peptidergic and non-peptidergic based on expressionof tachykinins and calcitonin gene-related peptide (CGRP)in the former type (Woolf & Ma, 2007). Isolectin B4 hasproven useful in defining these nociceptive subtypes inthat it stains somewhat selectively the non-peptidergicphenotype. These two subtypes innervate different regions
of the CNS, and thus clearly subserve distinct sensationsand functions in the organism (Molliver et al. 1995; Woolf& Ma, 2007).
At a more molecular level, recent studies are refiningthese two major nociceptive subtypes in dorsal root gangliaby evaluating the expression of transcription factorssuch as runt-related transcription factor 1 (RUNX1)and receptors for neurotrophic factors. The data fromstudies with mice support the hypothesis that RUNX1 isexpressed embryologically in virtually all nociceptors (but
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society DOI: 10.1113/jphysiol.2010.195339

4770 C. Nassenstein and others J Physiol 588.23
seldom in non-nociceptors) (Chen et al. 2006; Krameret al. 2006). Perinatally, however, RUNX1 expression isextinguished in a subset of nociceptors (Chen et al. 2006)and this coordinates a series of events that define thisRUNX1 negative phenotype as peptidergic nociceptors.The RUNX1 expressing nociceptors not only fail to expressneuropeptides but also differ from the peptidergic sub-type in that they express ret proto-oncogene (RET), thekey transmembrane signalling component of the glialcell derived neurotrophic factor (GDNF) family ligandreceptors (GFRs). By contrast, rather than depending onRET/GFRs, the small peptidergic nociceptors in the DRGexpress tropomyosin-related kinase (TRK)A and dependon nerve growth factor (NGF) for survival (Yoshikawaet al. 2007).
There is little information on nociceptor subtypes in thevagal afferent system. Vagal afferent C-fibres innervatingthe respiratory tract are generally quiet during healthyrespiration, but are activated by noxious stimuli andinflammatory mediators (Taylor-Clark & Undem, 2006).Inasmuch as they serve to provide the organ with a‘sense of its own potential injury’, they fit Sherrington’sdefinition of a nociceptor. These nociceptors do nottypically evoke pain sensations; rather their activation canlead to coughing, sneezing, dyspnoeic sensations, as wellas reflex secretion and bronchoconstriction (Coleridge &Coleridge, 1977; Coleridge et al. 1993). In most mammalsstudied thus far, C-fibres are categorically activated bycapsaicin. Thus, transient receptor potential vanilloid 1(TRPV1) expression is a useful molecular marker of vagalrespiratory nociceptors.
An immediate distinction between vagal and spinalnociceptors in larger mammals, including guinea pigs,is that vagal neurons are situated in only two ganglia,termed nodose and jugular (supranodose). The jugularganglion has a neural crest origin similar to the DRG,whereas the neurons comprising the nodose ganglion arederived from the epibranchial placodes (Baker & Schlosser,2005). Based on electrophysiological, histological, andgene expression analysis we have previously demonstratedthat the vagal nociceptors innervating the respiratorytract and oesophagus of guinea pigs can be broadlysubdivided into two subtypes (Kwong et al. 2008). Similarto spinal nociceptors one subtype is peptidergic and theother largely non-peptidergic (Undem et al. 2004). Thepeptidergic neurons are derived from the jugular (neuralcrest) ganglia, whereas the non-peptidergic neurons werederived from the placodal ganglion.
The extent to which knowledge about vagal nociceptorsubtypes can be inferred from information regardingsubtypes in the mouse DRG is unknown as there have beenno studies on vagal nociceptor subtypes in the mouse.In the present study we addressed the hypothesis thatthere are two general nociceptor phenotypes in the mousevagus, and these subtypes can be differentiated based on
expression of the P2X2 receptor, certain neurotrophicfactor receptors and the sensory neuropeptide, substanceP. The mouse offers the advantage of defining bona fideneural crest lineage neurons by using Wnt1Cre/R26Rmice. We therefore also directly address the hypothesis thatwhether the neurons are placodal or neural crest in originlargely predicts the nociceptive phenotype in the adultanimal. To keep other variables at a minimum, we focusedon nociceptor subtypes innervating a single defined peri-pheral compartment, namely the mouse lung.
Methods
Animal experiments
All experiments were performed with approval from theJohns Hopkins Animal Use and Care Committee.
Retrograde labelling and cell dissociation
Bronchopulmonary afferent neurons of C57/BL6mice (male, 6–8 weeks) were retrogradelylabelled using DiI (1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine perchlorate; DiC18(3);Molecular Probes/Invitrogen, Eugene, OR, USA) solution(0.1%, 50 μl; dissolved in 10% DMSO and 90%normal saline) or aminostilbamidine methanesulfonate(Molecular Probes/Invitrogen; 0.05%, 50 μl, dissolvedin normal saline). In comparison to the lipophilicDiI (absorption maximum: 549 ± 3 nm), the polaraminostilbamidine (absorption maximum: 386 ± 5 nm)was compatible with the detergent-containing washingbuffers used for immunohistochemistry. For all otherexperiments (single-cell RT-PCR, calcium imaging), weused DiI as a neuronal tracer.
Under anaesthesia (2 mg ketamine and 0.2 mg xylazineI.P. per mouse), mice were orotracheally intubated, andDiI or aminostilbamidine was instilled into the tracheallumen 5–9 days before an experiment.
After the animals were killed by CO2 asphyxiation,the jugular/nodose ganglia or DRGs (C1, T1–6) weredissected and cleared of adhering connective tissue.Isolated ganglia were incubated in an enzyme buffer(2 mg ml−1 collagenase type 1A and 2 mg ml−1 dispaseII in Ca2+-, Mg2+-free Hanks’ balanced salt solution) for30 min at 37◦C. Neurons were dissociated by triturationwith three glass Pasteur pipettes of decreasing tip pore size,then washed by centrifugation (three times at 1000 g for2 min) and suspended in L-15 medium containing 10%fetal bovine serum (FBS). The cell suspension was trans-ferred onto poly-D-lysine/laminin-coated coverslips. Afterthe suspended neurons had adhered to the coverslips for2 h, the neuron-attached coverslips were flooded with theL-15 medium (10% FBS) and used within 8 h.
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

J Physiol 588.23 Vagal C-fibre subtypes innervating the lung 4771
Single-cell RT-PCR
First strand cDNA was synthesized from singlelung-labelled jugular/nodose cells by using the Super-Script III CellsDirect cDNA Synthesis System (Invitrogen,Carlsbad, CA, USA) according to the manufacturer’srecommendations.
Cell picking. Coverslips of retrogradelly labelled,dissociated neurons were constantly perfused by Locke’ssolution and identified by using fluorescence micro-scopy. Single cells were harvested into a glass-pipette (tipdiameter 50–150 μm) pulled with a micropipette puller(Model P-87, Sutter Instruments Co., Novato, CA, USA)by applying negative pressure. The pipette tip was thenbroken in a PCR tube containing 1 μl RNAse Inhibitor(RNAseOUT, 2 U μl−1), immediately snap frozen andstored on dry ice. From one coverslip, one to four cells werecollected. A sample of the bath solution from the vicinityof a labelled neuron was collected from each coverslip forno-template experiments (bath control).
RT-PCR. Samples were defrosted, lysed (10 min at 75◦C)and treated with DNAse I. Then, poly(dT) and randomhexamer primers (Roche Applied Bioscience) were added.Twenty-five microlitres of the volume was reverse trans-cribed by adding SuperscriptIII RT for cDNA synthesis,whereas water was added to the remaining sample, whichwas used in the following as –RT control.
PCR. Of each sample, 1.2 μl (cDNA, RNA control or bathcontrol) was used for PCR amplification of mouse β-actin,TRPV1, P2X2, preprotachykinin A (PPT-A), RUNX1 andthe diverse nerve factor receptors by the HotStar TaqPoymerase Kit (Qiagen) according to the manufacturer’srecommendations in a final volume of 20 μl. After aninitial activation step at 95◦C for 15 min, cDNAs wereamplified with custom-synthesized primers (Invitrogen)(Table 1) by 45 cycles (b-Actin and TRPV1) or 50 cycles (allother genes) of denaturation at 94◦C for 30 s, annealingat 60◦C for 30 s and extension at 72◦C for 1 min followedby a final extension at 72◦C for 10 min. Products werethen visualized in ethidum bromide-stained 1.5% agarosegels.
Immunohistochemistry
The ganglia were fixed with 1% formalin in phosphatebuffered saline (PBS, 4 h, 4◦C), rinsed with PBS,cryoprotected overnight in PBS containing 18% sucrose,frozen and sectioned (10–12 μm). For GFRα3 staining, thesections were blocked with goat serum (10% in PBS with1% BSA, 0.5% Tween 20, 2 h), and incubated overnight(4◦C) with rabbit pAbs to GFRα3 (M-210, Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA; 7.5 μg ml−1),or normal rabbit IgG (R&D Systems, Minneapolis, MN,USA) as isotype control antibodies. For RET staining,sections were blocked with donkey serum and incubatedovernight with goat pAbs to RET (AF482, R&D Systems;2.5 μg ml−1) or normal goat IgG (R&D Systems) as isotypecontrol antibodies.
Sections were then rinsed with PBS containing 0.3%Triton X-100 and 1% BSA and incubated with AlexaFluor 647 goat anti-rabbit IgG (for GFRα3 staining),or Alexa Fluor Cy3 donkey anti-goat (for RET staining;both Molecular Probes/Invitrogen) for 2 h at roomtemperature. Washed slides were coverslipped with PBS(pH 8.6). The tissues were photographed (Q-ImagingRetiga EXi camera, BioVision, Exton, PA, USA), withan epifluorescence microscope (Olympus BX60, OlympusAmerica, Inc., Melville, NY, USA) equipped withappropriate filter sets to allow separate visualization ofAlexa 647, Alexa Cy3, and aminostilbamidine labellingusing IPLab software (BioVision).
X-Gal staining in Wnt1Cre/R26R mice
The Wnt1 gene is uniformly, albeit briefly, expressedin early migratory neural crest cells at all axial levels.The expression is extinguished as the cells migrateaway from the neural tube. The Wnt1Cre/R26R micehave a transgene expressing Cre recombinase under theWnt1 promoter and enhancer. The second componentis a reporter gene referred to as R26R that expressesβ-galactosidase. In these mice the neural crest lineageneurons (but not the placodal lineage) can be observedby their expression of β-galactosidase; for more details seeJiang et al. (2000). The enzyme activity of β-galactosidasein vagal ganglia of Wnt1Cre/R26R mice was visualizedby using the Marker Gene β-galactosidase staining kitaccording the manufacturer’s recommendations (MarkerGene Technologies Inc., Eugene, OR, USA).
Ca2+ imaging
The intracellular [Ca2+]free measurements were performedin dissociated jugular/nodose neurons fromWnt1/R26Rmice (n = 3). The coverslips were loaded with Fura-2AM (8 μM) in L-15 medium containing 20% FBS andincubated for 40 min at 37◦C. The coverslip was placedin a custom-built chamber (600 μl bath volume) thatwas superfused with Locke’s solution (at 35◦C) for20 min before the experiment by an infusion pump(8 ml min−1). Changes in intracellular [Ca2+]free weremeasured by digital microscopy (Universal; Carl Zeiss,Inc., Thornwood, NY, USA) equipped with in-houseequipment for ratiometric recording of single cells. A fieldof cells was monitored by sequential dual excitation, 352
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

4772 C. Nassenstein and others J Physiol 588.23
Table 1. Primer sequences for single cell RT-PCR
Gene Primer Sequence (5′ to 3′) GenBank Product length
β-Actin Forward CTG GTC GTC GAC AAC GGC TCC NM_007393 302 bpReverse GCC AGA TCT TCT CCA TG
TRPV1 Forward TCA CCG TCA GCT CTG TTG TC NM_001001445 285 bpReverse GGG TCT TTG AAC TCG CTG TC
P2X2 Forward GGG GCA GTG TAG TCA GCA TC NM_153400 241 bpReverse TCA GAA GTC CCA TCC TCC A
PPT-A Forward AGA CCC AAG CCT CAG CAG TT D 17584 215/171/162/118 bpReverse CGT CTT CTT TCG TAG TTC TGC ATT
RET Forward CCT GCT GTC CAT CTT CTG TGT NM_009050 260 bpReverse TTC CAA ACT CGC CTT CTC C
RET9 Forward GGA AGC AGG AGC CAG ACA A NM_001080780 357 bpReverse AAA GAA AAG GGT TCG GAG GA
RET51 Forward AAG CAG GAG CCA GAC AAG AG NM_001080780 344 bpReverse GGT GAA ACC ATC CAG TTA GCA
GFRα1 Forward CAC GAC TAC CAC TGC CTT CC NM_010279 185 bpReverse CAG CGA GAC CAT CCT TTC C
GFRα2 Forward TGA CGG AGG GTG AGG AGT T NM_008115 209 bpReverse GCA GAT GGA GAT GTA GGA GGAG
GFRα3 Forward CTG CTA CTG GTG CTG TCG TT NM_010280 230 bpReverse CTG AGT TGT TCT GCT GCC TCT
GFRα4 Forward AAC CCC TGC TTG GAT GGT NM_020014 232 bpReverse GTC GTC CAC GGT TCA TGT T
TRKA Forward GGT GGC TGC TGG TAT GGT AK081588 216 bpReverse CTG AAC TTG CGG TAG AGG ATG
TRKB Forward CTA AAT CCA GCC CCG ACA C NM_008745 151 bpReverse GTC ACA GAC TTT CCT TCC TCC A
TRKC Forward TCA ACA AGC CCA CCC ACT AC NM_008746 213 bpReverse CTG CTA TGG ACA CCC CAA AA
RUNX1 Forward CTC CGT GCT ACC CAC TCA CT NM_009821 184 bpReverse GTC GTT GAA TCT CGC TAC CTG
lacZ Forward GAT GTG GAT TGG CGA TAA AAA J01636 433 bpReverse AGT AAG GCG GTC GGG ATA GT
VGluT1 Forward CAT CTC TCA GCC CGC CTA C NM_182993 230 bpReverse GCG AGT ATC CGA CCA CCA
VGluT2 Forward CTG CGA TAC TGC TCA CCT CTA C AF324864 175 bpReverse GCC AAC CTA CTC CTC TCC AA
and 380 nm, and the analysis of the image ratios usedmethods previously described (MacGlashan, 1989). Theratio images were acquired every 6 s. Superfused bufferwas stopped 20 s prior to drug applications. In eachexperiment, the cells on the coverslip were exposed toα,β-methylene ATP (10 μM), capsaicin (1 μM) and KCl(75 mM) for 1 min each. The KCl was used as an indicatorof voltage-sensitive cells. Between each stimulus, the cellswere washed with fresh buffer for at least 3 min forthe cells to recover prior to the addition of the secondstimulus. After the last stimulation, cells were fixed inglutaraldehyde and were stained in situ with the MarkerGene β-galactosidase staining kit as described above. Eachset of images for the Ca2+ measurements also included abrightfield image of the field of cells under study beforeand after the X-Gal staining. Neural crest derived cellswere identified based on their dark appearance. Only
cells that had an average diameter (long and short axis)of over 15 μm were analysed. Those cells that failed torespond to capsaicin with a rapid rise in Ca2+ wereconsidered non-nociceptive and thus were not consideredfor statistical analysis.
Extracellular recording
The animals were killed by CO2 asphyxiation followedby exsanguination. The innervated isolated trachea-lungpreparation was prepared as previously described(Kollarik et al. 2003). Briefly, the airways and lungswith their intact right-side extrinsic innervation (vagusnerve including jugular/nodose ganglia) were taken andplaced in a dissecting dish containing Krebs bicarbonatebuffer solution composed of (mM): 118 NaCl, 5.4 KCl,
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

J Physiol 588.23 Vagal C-fibre subtypes innervating the lung 4773
1.0 NaH2PO4, 1.2 MgSO4, 1.9 CaCl2, 25.0 NaHCO3 and11.1 dextrose, and equilibrated with 95% O2 and 5%CO2 (pH 7.2–7.4). Connective tissue was trimmed awayleaving the larynx, trachea, main bronchi and lungs withtheir intact nerves. The airways were then pinned to thelarger compartment of a custom-built two-compartmentrecording chamber which was lined with siliconeelastomer (Sylgard 184 Silicone Elastomer Kit, DowCorning, Midland, MI, USA). The left jugular/nodoseganglion was gently pulled into the adjacent compartmentof the chamber through a small hole and pinned.Both compartments were separately superfused withKrebs solution, which was warmed by a warming jacket(39–42◦C) to keep airway tissues and ganglia at 37◦C.A sharp glass electrode was pulled by a Flaming Brownmicropipette puller (P-87; Sutter Instrument Co.) andfilled with 3 M NaCl solution. The electrode was gentlyinserted into the jugular/nodose ganglion so as to beplaced near the cell bodies. The recorded action potentialswere amplified (Microelectrode AC amplifier 1800; A-MSystems, Everett, WA, USA), filtered (0.3 kHz of lowcut-off and 1 kHz of high cut-off), and monitored on anoscilloscope (TDS340; Tektronix, Beaverton, OR, USA)and a chart recorder (TA240; Gould, Valley View, OH,USA). The scaled output from the amplifier was capturedand analysed by an Apple Macintosh computer usingNerveOfIt software (Phocis, Baltimore, MD, USA). Formeasuring conduction velocity, an electrical stimulation(S44; Grass Instruments, Quincy, MA,USA) was appliedon the core of the receptive field. The conduction velocitywas calculated by dividing the distance along the nervepathway by the time delay between the shock artifact andthe action potential evoked by electrical stimulation. If aC-fibre (<1 m s−1) was found, the recording was started.One millilitre of vehicle, α,β-methylene ATP (10 μM) orcapsaicin (1 μM) was intratracheally applied for 10 s. Wehave found that this technique does not record actionpotentials in ‘through fibres’. For example, positioning theelectrode away from cell bodies on the vagus itself fails torecord action potential stimulated by receptive fields, orby electrical nerve stimulation.
Statistical analysis
There was little (<1 Hz) or no background activity inthe extracellular recordings. In all experiments, a singleunit was recorded. The action potential discharge evokedby vehicle, α,β-methylene ATP and capsaicin stimulationwas quantified off-line and segregated into consecutive 1 sbins. The response was considered to be terminated whenthe number of spikes in the bins declined to <2 × baseline.The total number of action potentials recorded followingvehicle, α,β-methylene ATP and capsaicin application wascounted. The peak frequency evoked by a stimulus was
quantified as the maximum number of action potentialsthat occurred within any 1 s bin.
The intracellular [Ca2+]free of vagal mouse neuronswas calculated as previously described using the Tsiencalculation (Taylor-Clark et al. 2008). If a cell lacked arobust response to capsaicin (1 μM) or KCl (75 mM), orhad an averaged diameter (long and short axis) of lessthan 15 μm, it was not included in the analysis. A cell wasconsidered as α,β-methylene, capsaicin or KCl positiveif the drug-induced peak increase was greater than twostandard deviations above the mean baseline intracellular[Ca2+]free.
Student’s paired or unpaired t test or ANOVA was usedwhen appropriate. P < 0.05 was considered statisticallysignificant. All data are expressed as means ± S.E.M.
Drug preparations and applications
Capsaicin was dissolved in ethanol, and α,β-methyleneATP and KCl were dissolved in H2O, respectively,and diluted with the appropriate buffer to the finalconcentrations. The final ethanol concentration was 0.1%for calcium measurements and 0.01% for extracellularrecordings. Capsaicin was purchased from Sigma-Aldrich(St Louis, MO, USA). Fura-2 AM was purchasedfrom Molecular Probes/Invitrogen. L-15 and Hanks’balanced salt solution (HBSS) were purchased fromGibco/Invitrogen (Carlsbad, CA, USA).
Results
Anatomy of the mouse jugular nodose complex
Vagal sensory neurons are situated in the nodose (placodederived) and jugular ganglion (neural crest derived). Thesedisparate ganglia are very evident in guinea pigs andlarger mammals, but there has been some issue of theirlocation in mice. We carried out dissections of the vagusand accompanying ganglia in adult mice, and foundthat indeed, compared to the guinea pig, there is oftenno discrete nodose and jugular ganglion on the vagusnerve of the mouse, and only occasionally a minimalganglionic structure is observed rostral to the nodoseganglion (Fig. 1).
The nodose ganglion in the mouse often appearssomewhat elongated, so we hypothesized it may representa fused nodose–jugular complex with the rostral pole morejugular-like, and the body and caudal pole, nodose-like. Toaddress this we obtained mice in which the premigratoryneural crest neurons expressed β-galactosidase encodedby the E.coli lacZ gene (Wnt1Cre/R26R mice) (Jianget al. 2000; Makita et al. 2008). When the vagal gangliaof these mice (n = 16) were evaluated it was clear thatneural crest derived cells formed the more rostral aspect
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

4774 C. Nassenstein and others J Physiol 588.23
Figure 1. Dissection of the vagal ganglia in guinea pigs and miceThe right vagus and accompanying ganglia were dissected in guinea pigs (left photograph) and mice (rightphotograph). Compared to the guinea pig in which the nodose and the jugular ganglion are clearly separated,mice possess a fused jugular nodose complex.
of the ganglion whereas the caudal pole of the ganglioncomprised non-neural crest (placodal) neurons. In severalganglia, a significant number of neural crest neurons wereobserved down to the central body of the ganglion (Fig. 2).In about 1 in 3 mice a jugular ganglion, entirely consistingof neural crest derived cells, can be recognized as the tophalf of an hourglass like structure on the vagus.
Calcium imaging
In a previous study with guinea pigs we noted thatthe nodose C-fibre terminals in the respiratory tractresponded strongly with action potential discharge tothe P2X-2/3 selective agonist α,β-methylene ATP, whereas
the jugular neurons were categorically unresponsive. Wealso noted at the level of the cell body that the nodoseneurons responded to ATP or α,β-methylene ATP witha large persistent inward current that was explained bythe presence of both P2X2 and P2X3 receptors. Jugularneurons expressed P2X3 but not P2X2 receptors andresponded to ATP with a very transient inward current,which is consistent with the current signature of P2X3homomeric receptors (Kwong et al. 2008).
We evaluated the α,β-methylene ATP responsiveness inmouse vagal ganglion neurons. Using the Wnt1Cre/R26Rmice, we dissociated the vagal ganglion complex (8ganglia from 4 mice) and evaluated the calciumrise in β-galactosidase+ neural crest neurons andin β-galactosidase− placodal neurons. About 58%
Figure 2. X-Gal staining of the jugular nodosecomplex in miceIn order to evaluate the localization of neural crestderived and placodal neurons within the jugular–nodosecomplex, X-Gal staining was performed inWnt1Cre/R26R mice. Neural crest derived cells formedthe rostral pole of the ganglion whereas the body andcaudal pole of the ganglion comprised the placodal cells(A–D). A jugular ganglion, entirely consisting of neuralcrest derived cells, can be recognized as the top half ofan hourglass like structure on the vagus in some ganglia(C and D).
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society
J Physiol 588.23 Vagal C-fibre subtypes innervating the lung 4775
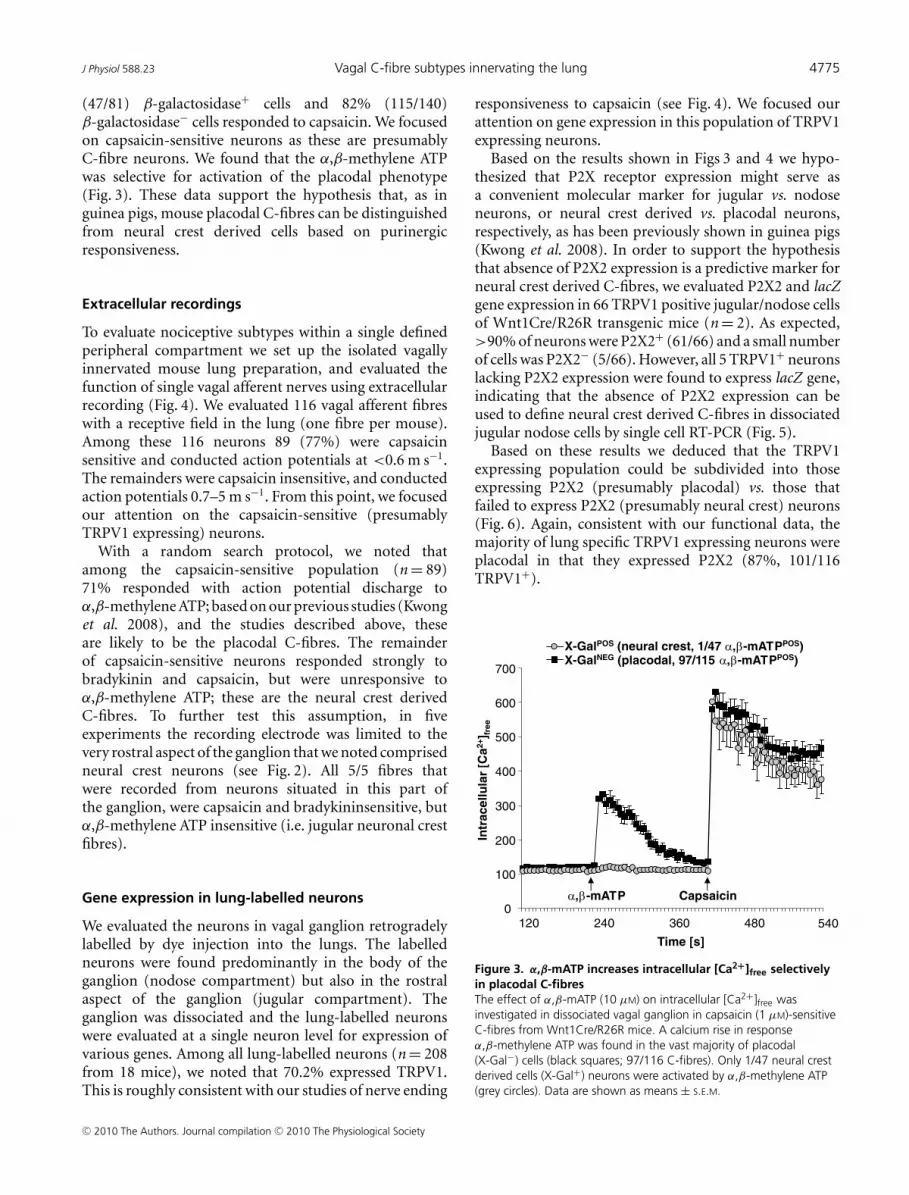
(47/81) β-galactosidase+ cells and 82% (115/140)β-galactosidase− cells responded to capsaicin. We focusedon capsaicin-sensitive neurons as these are presumablyC-fibre neurons. We found that the α,β-methylene ATPwas selective for activation of the placodal phenotype(Fig. 3). These data support the hypothesis that, as inguinea pigs, mouse placodal C-fibres can be distinguishedfrom neural crest derived cells based on purinergicresponsiveness.
Extracellular recordings
To evaluate nociceptive subtypes within a single definedperipheral compartment we set up the isolated vagallyinnervated mouse lung preparation, and evaluated thefunction of single vagal afferent nerves using extracellularrecording (Fig. 4). We evaluated 116 vagal afferent fibreswith a receptive field in the lung (one fibre per mouse).Among these 116 neurons 89 (77%) were capsaicinsensitive and conducted action potentials at <0.6 m s−1.The remainders were capsaicin insensitive, and conductedaction potentials 0.7–5 m s−1. From this point, we focusedour attention on the capsaicin-sensitive (presumablyTRPV1 expressing) neurons.
With a random search protocol, we noted thatamong the capsaicin-sensitive population (n = 89)71% responded with action potential discharge toα,β-methylene ATP; based on our previous studies (Kwonget al. 2008), and the studies described above, theseare likely to be the placodal C-fibres. The remainderof capsaicin-sensitive neurons responded strongly tobradykinin and capsaicin, but were unresponsive toα,β-methylene ATP; these are the neural crest derivedC-fibres. To further test this assumption, in fiveexperiments the recording electrode was limited to thevery rostral aspect of the ganglion that we noted comprisedneural crest neurons (see Fig. 2). All 5/5 fibres thatwere recorded from neurons situated in this part ofthe ganglion, were capsaicin and bradykininsensitive, butα,β-methylene ATP insensitive (i.e. jugular neuronal crestfibres).
Gene expression in lung-labelled neurons
We evaluated the neurons in vagal ganglion retrogradelylabelled by dye injection into the lungs. The labelledneurons were found predominantly in the body of theganglion (nodose compartment) but also in the rostralaspect of the ganglion (jugular compartment). Theganglion was dissociated and the lung-labelled neuronswere evaluated at a single neuron level for expression ofvarious genes. Among all lung-labelled neurons (n = 208from 18 mice), we noted that 70.2% expressed TRPV1.This is roughly consistent with our studies of nerve ending
responsiveness to capsaicin (see Fig. 4). We focused ourattention on gene expression in this population of TRPV1expressing neurons.
Based on the results shown in Figs 3 and 4 we hypo-thesized that P2X receptor expression might serve asa convenient molecular marker for jugular vs. nodoseneurons, or neural crest derived vs. placodal neurons,respectively, as has been previously shown in guinea pigs(Kwong et al. 2008). In order to support the hypothesisthat absence of P2X2 expression is a predictive marker forneural crest derived C-fibres, we evaluated P2X2 and lacZgene expression in 66 TRPV1 positive jugular/nodose cellsof Wnt1Cre/R26R transgenic mice (n = 2). As expected,>90% of neurons were P2X2+ (61/66) and a small numberof cells was P2X2− (5/66). However, all 5 TRPV1+ neuronslacking P2X2 expression were found to express lacZ gene,indicating that the absence of P2X2 expression can beused to define neural crest derived C-fibres in dissociatedjugular nodose cells by single cell RT-PCR (Fig. 5).
Based on these results we deduced that the TRPV1expressing population could be subdivided into thoseexpressing P2X2 (presumably placodal) vs. those thatfailed to express P2X2 (presumably neural crest) neurons(Fig. 6). Again, consistent with our functional data, themajority of lung specific TRPV1 expressing neurons wereplacodal in that they expressed P2X2 (87%, 101/116TRPV1+).
X-GalNEG (placodal, 97/115 , -mATPPOS)X-GalPOS (neural crest, 1/47 , -mATPPOS)
120 240 360 480 540
Intr
ac
ell
ula
r[C
a2+] f
ree
Time [s]
0
100
200
300
400
500
600
700
, -mATP Capsaicin
Figure 3. α,β-mATP increases intracellular [Ca2+]free selectivelyin placodal C-fibresThe effect of α,β-mATP (10 μM) on intracellular [Ca2+]free wasinvestigated in dissociated vagal ganglion in capsaicin (1 μM)-sensitiveC-fibres from Wnt1Cre/R26R mice. A calcium rise in responseα,β-methylene ATP was found in the vast majority of placodal(X-Gal−) cells (black squares; 97/116 C-fibres). Only 1/47 neural crestderived cells (X-Gal+) neurons were activated by α,β-methylene ATP(grey circles). Data are shown as means ± S.E.M.
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

4776 C. Nassenstein and others J Physiol 588.23
Figure 4. α,β-mATP stimulation of vagal sensory nerves elicits action potential discharge in a sub-population of capsaicin-sensitive bronchopulmonary C-fibresThe action potential discharge in response to α,β-mATP and capsaicin stimulation was evaluated in broncho-pulmonary vagal afferent nerves by extracellular recording. 89/116 neurons (77%) were capsaicin sensitive andconducted action potentials at <0.6 m s−1. We noted that among the capsaicin-sensitive population (n = 89), 71%responded with action potential discharge to α,β-methylene ATP; these are likely to be the placodal C-fibres. Theremainder of capsaicin-sensitive neurons responded strongly to bradykinin and capsaicin, but were unresponsiveto α,β-methylene ATP; these are the neural crest derived C-fibres
Gene expression in P2X2-negative ‘jugular’ neurons.Previous studies have indicated that spinal (neural crest)nociceptors can be subdivided into a peptidergic TRKA+
group and a non-peptidergic TRKA− subpopulation(Woolf & Ma, 2007). The occurrence of the two differentneural crest C-fibre subpopulations was confirmed invagal TRPV1+lacZ+P2X2− cells. Three of five cells
Figure 5. Neural crest derived C-fibres lack P2X2 mRNAexpressionSingle cell RT-PCR was performed in vagal C-fibres fromWnt1Cre/R26R mice. Five out of 66 TRPV1+ cells did not expressP2X2. All 5 cells were lacZ+ and therefore considered to be of neuralcrest origin. Further analysis revealed that this cell population consistsof two different phenotypes as revealed by TRK receptor mRNAexpression.
obtained from two mice expressed TRKA but not TRKB,and 2/5 cells were TRKB+ but did not express TRKA(Fig. 5).
Next, we evaluated gene expression in target-specificcranial neural crest cells, namely the lung-specificnociceptors, which were stained by intratrachealapplication of DiI. As mentioned above, the neuralcrest derived cells represent a relatively small populationof capsaicin-sensitive (or TRPV1 expressing) neuronsinnervating the lungs. Nevertheless we were able toevaluate gene expression in 15 TRPV1+P2X2− neuronsobtained from eight animals (Fig. 6), and noted thatboth TRKA+ (6/15) and TRKA− neurons were present(9/15). TRKB was expressed in 5 of the 9 TRKA− neuronsleaving only four neurons that failed to express eitherTRKA or TRKB. None of the neurons expressed TRKC(data not shown). We next evaluated whether expressionof preprotachykinin-A (PPT-A) mRNA correlated withTRKA (as it does in DRG neurons). Indeed 5/6 TRKA+ cellwere PPT-A+, whereas only 3 of 8 TRKA− cells expressedPPT-A (Fig. 6).
All 15 of the lung-specific neural crest neuronsexpressed RET, but the expression of GDNF familyreceptor subtypes was selective for TRKA+ vs. TRKA−
neurons. The GDNF receptor, GFRα1, was selectivelyexpressed in the non-peptidergic TRKA− neurons (8/9neurons were GFRα1+), and only rarely expressed inTRKA+ neurons (1/6). By contrast, GFRα3, the receptorfor artemin, was expressed selectively in the peptidergicTRKA+ subtype. All of the TRKA+ neurons expressedGFRα3, whereas only 1 of 9 TRKA− neurons expressedthis receptor. GFRα2 (2 of 15) and GFRα4 (2of 15) were
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

J Physiol 588.23 Vagal C-fibre subtypes innervating the lung 4777
rarely expressed in these neural crest nociceptive neurons(data not shown).
Gene expression in placodal P2X2-positive ‘nodose’neurons. As mentioned the majority of vagal sensoryneurons innervating the mouse lungs are placodal innature, and thus we were able to evaluate gene expressionin a larger number (up to n = 82) of lung-specific TRPV1+
placodal neurons than neural crest neurons obtained from18 mice.
By contrast to the neural crest neurons, nearly allof the TRPV1+ placodal ‘nodose’ neurons expressedTRKB (72/82, 87.2%), whereas TRKA was relatively rarelyexpressed (17/82, 20.7%) (Fig. 6). Comparable with theneural crest neurons, TRKC was rarely expressed in
the TRPV1+ placodal neurons (7/73, 9.6%) (data notshown).
PPT-A was expressed in <10% (5/51) of lung-specificC-fibre placodal neurons. These data show that in the adultmouse the majority of tachykinergic C-fibres innervatingthe lungs are by and large neural crest and not placodalin nature, consistent with reports in other species (Katz &Karten, 1980; Lundberg et al. 1983; Springall et al. 1987;Kummer et al. 1992; Undem et al. 2004).
As with the neural crest population of neurons, wefound that that RET mRNA was uniformly expressed byTRPV1 expressing lung labelled placodal neurons (80/80neurons, 100%). Single cell RT-PCR with primers thatwere able to discriminate between RET9 and RET51isoforms showed that most of the cells co-expressed bothvariants (data not shown; RET9 was expressed in 20/21
Figure 6. Characterization of distinct bronchopulmonary C-fibre subsets by single cell RT-PCRSince the lack of P2X2 receptors was specific for neural crest derived neurons, lung-labelled TRPV1+ vagal neuronswere divided in two groups according to their P2X2 expression. In each individual cell, the expression of PPT-A gene(preprotachykinin A), glial cell derived neurotrophic family ligand receptors (GFLRs) and neurotrophin receptors(NTRs) were analysed. Up to 82 placodal cells and 15 neural crest derived cells were analysed per individual target.The figure shows representative results from 15 individual placodal cells and all neural crest neurons. In order tofacilitate the understanding of the figure, the neural crest cells were sorted according to their TRKA expression. P,positive control (cDNA from whole JNG); N, –RT negative control; B, bath control; ∗not done
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

4778 C. Nassenstein and others J Physiol 588.23
cells (95%); RET51 was expressed in 18/21 cells (85.7%)).Thus, RET is expressed indiscriminately in vagal sensoryneurons innervating adult mouse lungs.
This conclusion based on gene expression wassupported by immunohistochemical data revealing thatindeed virtually all vagal ganglion neurons were RET+
(Fig. 7).The GDNF receptor GFRα1 was expressed in nearly all
placodal nociceptors innervating the lungs (53/60, 88%).The artemin receptor GFRα3 was not expressed in placodalC-fibre neurons (0/51). Thus, this receptor is expressedselectively in neural crest C-fibre neurons (Fig. 6). Tosupport the conclusion that GFRα3 is selectively expressedonly in vagal ganglion neurons of neural crest origin,we evaluated GFRα3 immunohistologically in gangliaisolated from the Wnt1Cre/R266R mice. As predictedfrom our gene expression studies, neurons stained withanti-GFRα3 antibodies were limited to the small minorityof neural crest neurons with the majority of lung labelledneurons being placodal neurons that are negative forGFRα3 (Fig. 8). GFRα2 mRNA was expressed in 15/56(27%) and GFRα4 mRNA was expressed in 28/56 (50%)of the neurons (data not shown).
RUNX1 expression. In DRG neurons, RUNX1 expression,as with RET expression, was found to be selectivelyexpressed in the non-peptidergic subtype of nociceptor,at least in early post-natal life (Chen et al. 2006; Woolf& Ma, 2007; Yoshikawa et al. 2007). We addressed thehypothesis that the placodal subtype of vagal nociceptorwould preferentially express RUNX1. RUNX1 mRNA
was expressed in the majority of TRPV1 expressinglung-labelled neurons of both placodal (24/37) and neuralcrest (9/15) origin. RUNX1 expression was not selectivelyassociated with other genes we evaluated (PPT-A, TRKreceptors, or GFLR receptors) (Fig. 6).
Glutamate tansporter expression. Different sensorynerve phenotypes have been shown to express differentglutamate transporters (Oliveira et al. 2003; Morris et al.2005; Brumovsky et al. 2007; Mazzone & McGovern,2008). We evaluated 28 lung specific vagal TRPV1+
neurons for expression of VGluT1 and VGluT2. BothVGluT1 and VGluT2 were expressed in the total vagalganglion. However, none of the 28 TRPV1+ neuronsexpressed VGluT1 (data not shown), whereas each of the28 neurons expressed VGluT2 mRNA. The 28 neuronswere of both placodal and neural crest origin (Fig. 6).VGluT1 was expressed in 2 of 7 lung-specific neurons thatwere TRPV1-negative (data not shown).
Dorsal root ganglion neurons. Dinh et al. (2004) notedthat a portion of DRG neurons provide TRPV1+ axons tothe mouse lungs. These DRG neurons arise from the sameregion of postotic hindbrain that also gives rise to vagaljugular neurons (Baker & Schlosser, 2005). Therefore, wepredicted that the gene expression in the TRPV1+ DRGneurons should be similar to the jugular neurons.
In comparison to the percentage of TRPV1+ cellsin vagal ganglia (70.2% from 208 neurons), TRPV1expression was lower in T1–T6 DRG (51.1% from 45neurons obtained from 6 mice) and in C1 DRG (25.5%
Figure 7. RET expression in mouse jugular nodosecomplexRET expression was tested in lung-labelled jugularnodose ganglia sections of male C57BL/6 mice. Almostall neurons expressed RET. A, the left section wasstained with a RET mAbs (red fluorescence) and theright section was incubated with the correspondingisotype control. Vagal nerve cells projecting to theairways were retrogradely stained by intratrachealapplication of aminostilbamidine (green fluorescentcells). B, higher magnification reveals the presence ofRET immunoreactivity in lung-labelled neurons.
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

J Physiol 588.23 Vagal C-fibre subtypes innervating the lung 4779
from 98 neurons obtained from 5 mice). Since no obviousdifferences in the gene expression profile from TRPV1+
neurons could be observed in T1–T6 (n = 25) and C1DRG (n = 23), the data were pooled.
Consistent with our prediction, the vast majority(87.5%) of these neurons did not express P2X2. Alsoconsistent with what we observed in the jugular ganglionneurons, the TRPV1+ neurons expressed TRKA (35/48neurons). The vast majority (92%) of these neurons alsoexpressed RET, and at least one GFR receptor (Table 2).
Discussion
This study demonstrates that in the adult mouse,capsaicin-sensitive vagal C-fibres innervating a singleorgan (lungs) comprise two major subtypes, based onembryological origin. One subtype is neural crest in origin,the other placodal. The data also support our preliminaryhypothesis that the capsaicin-sensitive neural crest vagalC-fibres can be further subdivided into two phenotypesbased on expression of TRKA receptors. The main threenociceptive C-fibre subtypes differ in activation profileand expression of neuropeptide and growth factor receptorgenes. They therefore likely subserve disparate sensationsand reflexes, and will respond to noxious or inflammatory
Table 2. Gene expression in TRPV1+ neurons in neurons isolatedfrom the cervical and T1-T6 DRGS of the adult mouse
Gene Neurons Percentage of positve neurons
P2X2 8/48 17%PPTA 39/48 81%TRKA 35/48 73%RET 44/48 92%TRKA and RET 35/48 73%GFRα1 35/48 73%GFRα2 20/48 42%GFRα3 14/48 29%GFRα4 20/48 42%
Neurons were isolated from C1 and from T1–T6 DRG of adultmice. The neurons were first screened for expression of TRPV1. Atotal of 48 TRPV1 expressing neurons were identified obtainedfrom 8 mice.
conditions in their own unique manner. The recognitionof distinct vagal nociceptive phenotypes is essential in thedevelopment of our understanding of the role these nervesplay in respiratory physiology and in inflammatory airwaydiseases such as asthma, chronic obstructive pulmonarydisease, and chronic bronchitis.
In guinea pigs and larger mammals, the cell bodies ofvagal sensory neurons form two distinct ganglia referred
Figure 8. GFRα3 expression in lung-labelledneuronsGFRα3 expression was evaluated byimmunohostistochemistry in lung-labelled vagal gangliaof Wnt1Cre/R26R mice. GFRα3 expression wasselectively expressed in neural crest derived (X-Gal+)neurons. B, arrowheads indicate lung-labelled GFRα3+neural crest derived neurons.
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

4780 C. Nassenstein and others J Physiol 588.23
to as the nodose and jugular (or supranodose) ganglion.The neural crest derived cells form a jugular ganglion,which is clearly separated from the nodose ganglionthat comprises neurons from the epibranchial placode(Yntema, 1942; D’Amico-Martel & Noden, 1983). Wepreviously noted in guinea pig airways and oesophagusthat P2X2 expression and responsiveness to α,β-mATPis a useful molecular marker of nodose (placodal)vagal sensory neurons (Kwong et al. 2008). In themouse, the vagal sensory neurons most often formed asingle elongated ganglion (see Fig. 1). In our studies ofaction potential discharge of capsaicin-sensitive C-fibresin the lungs, recorded with an extracellular electrodepositioned in this vagal sensory ganglion, we notedthat C-fibres could be subdivided into ATP sensitive(nodose-like) and ATP insensitive (jugular-like) cells. Thispredicts that the vagal ‘nodose’ ganglion in the mouseis actually a combination of neural crest and placodalneurons.
The hypothesis that the single ganglion structure in themouse vagus is a combination of nodose (placodal) andjugular (neural crest) neurons is directly supported byour data using Wnt1Cre/R26R mice. These mice expressβ-galactosidase only in premigratory neural crest derivedneurons. The neural crest neurons were consistently foundwithin the single vagal ganglion structure; moreover,consistent with the predictions from the nerve endingsstudies, only the non-neural crest neurons (placodalneurons) responded to α,β-mATP with large increasesin intracellular calcium. In a minority of mice we notedthat the nodose and jugular neurons were separated withthe later forming a very small swelling distinct from thenodose ganglion. Most often, however, the neural crestneurons formed the rostral aspect of a single ganglionstructure with a significant minority of these neurons asfar caudally as the ganglion centre. Thus, one must becautious when evaluating neurons from these ganglia andassuming that they are all ‘nodose’ neurons. Consistentwith the hypothesis that the ATP-unresponsive C-fibresterminating in the mouse lungs are jugular in nature wasthe observation that when the electrode was positionedin the rostral aspect of the vagal ganglion complex, thecapsaicin-sensitive C-fibre under study categorically failedto respond with action potential discharge to α,β-mATP.In guinea pigs and rats, the number of C-fibre neuronsinnervating the respiratory tract is similar to the number ofnodose neurons (Springall et al. 1987; Ricco et al. 1996; Yuet al. 2005). In the present study we found that in the mousethe nodose C-fibre neurons far outnumbered the jugularneurons. This may represent a species difference. It shouldbe kept in mind that the neural crest C-fibres in the lungcomprise not only jugular neurons, but also DRG neurons.Thus the neural crest derived C-fibres innervating therespiratory tract and oesophagus may actually outnumberthe nodose C-fibres.
In guinea pigs and rats the sensory neurokinins in thelungs are preferentially localized to jugular (neural crest)ganglion C-fibres. Consistent with this, we found that thepreprotachykinin gene, PPTA, was selectively expressedin the neural crest subpopulation of lung-labelledneurons. However, not all neural crest C-fibres wereneurokinin-positive. The PPT-A gene was predominantlyexpressed in the TRKA+ subset of jugular neurons. Thisis consistent with findings in the somatosensory systemwhere PPT-A is also expressed in the majority of TRKA+
nociceptors (Kashiba et al. 1996).Vagal and spinal nociceptors are neurons that exhibit
plasticity. They respond to local insults and inflammationwith respect to both excitability changes and actionpotential discharge, but also via alterations in geneexpression. For example at sites of airway inflammationinduced by either allergen or viral exposure, there arephenotypic switches in the neuropeptide expression suchthat even capsaicin-insensitive placodal A-fibre neuronsbecome peptidergic (Hunter et al. 2000; Carr et al. 2002;Chuaychoo et al. 2005; Dinh et al. 2005). Neurotrophicfactor molecules produced at sites of inflammation arelikely to be responsible for these types of phenotypicchanges. It is therefore important to understand the natureof neurotrophic receptor expression in the subsets ofadult vagal neurons. With respect to the neurotrophinreceptors we noted that the placodal C-fibres are TRKBexpressing neurons, whereas about 50% of the neural crestC-fibres were TRKA expressing neurons. Relatively fewC-fibre placodal neurons expressed TRKC. This is basicallyconsistent with pre- and perinatal studies of nodose andjugular neurons, and indicates that in the adult animalsBDNF will be more apt to modulate the function ofplacodal neurons, whereas many neural crest neurons aremore apt to be under the control of NGF.
The GDNF family ligands include GDNF, neurturin,artemin, and persephin. These growth factors activateGDNF family ligand receptors (GFRs) GFRα1, GFRα2,GFRα3 and GFRα4, respectively (Bespalov & Saarma,2007). Our findings indicate that GDNF will most likelymodulate the function of placodal C-fibre neurons in thelungs, as well as the TRKA-negative neural crest C-fibres,in that these neurons express GFRα1 receptor. The GFRα3receptor agonist artemin, on the other hand, is unlikelyto influence placodal C-fibres but may modulate thepeptidergic TRKA-positive neural crest C-fibre populationin the lungs. Although our studies focused mainly onlung-specific neurons, this idea may be relevant toother visceral organs. Nodose neurons labelled from themouse pancreas do not express GFRα3, whereas thisreceptor is expressed in the majority of pancreas specificDRG neurons (neural crest neurons) (Fasanella et al.2008).
RET functions as an important co-receptor for theGFRs. Both in our gene expression analysis and in
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

J Physiol 588.23 Vagal C-fibre subtypes innervating the lung 4781
our immunohistochemical analysis we found RET to beuniformally expressed in the adult vagal sensory neurons.This is different from observations in the perinatal DRGneurons, where RET was found to be selectively expressedin the non-peptidergic, non-TRKA subtype of nociceptors(Woolf & Ma, 2007). The expression of the transcriptionfactor RUNX1 is also a marker of the non-peptidergicsubtype of nociceptor in DRGs (Chen et al. 2006). Wefound, however, that RUNX1 was expressed in about50% of the lung-specific vagal noceptors, but it wasnot apparently coordinated with RET, TRK, or PPT-Aexpression.
The vagal jugular ganglion neurons emigrate from thepostotic hindbrain at the level of the first seven somites.This region also gives rise to the neuron in the morerostral DRG (Baker & Schlosser, 2005). If the nociceptorphenotype is explained largely by embryonic origin, wewould predict that the capsaicin-sensitive neurons in theDRGs innervating the lungs would be similar to the jugularneurons. Although we did not investigate lung-specificDRG neurons, the TRPV1 expressing neurons in therostral DRGs are similar to jugular neurons in their lack ofP2X2 expression, and in their expression of TRKA alongwith RET and in some neurons GFRα3. As we observedin the vagal ganglia, nearly all the TRPV1-expressingneurons comprising the rostral DRGs in the adult mice,including those that express PPT-A, express both TRKreceptors and RET/GFRα receptors. This is similar to adulthuman DRG where a substantial percentage of TRKAexpressing neurons co-express RET (Josephson et al.2001). Therefore the idea, based largely on elegant peri-natal studies of lumbar DRG neurons, that a nociceptorcan be categorically segregated into TRK expressing orRET/GFRα expressing subtypes should be cautiouslyextrapolated to DRG neurons of the adult animal ingeneral.
Glutamate transporters have been shown to bedifferentially expressed in sensory neurons of differentphenotypes (Oliveira et al. 2003; Morris et al. 2005;Brumovsky et al. 2007; Mazzone & McGovern, 2008).There is little information on differential VGluTexpression in mouse vagal sensory ganglia. Generally, inthe DRG, VGluT1 has been shown to be expressed inlarge diameter non-nociceptive neurons, whereas VGluT2has been found to be expressed in smaller diameter,presumed nociceptive neurons. There is some controversyabout whether VGluTs may be absent from most smalldiameter peptidergic neurons in the DRG. In guineapigs and mice, Morris et al. (2005) concluded thatmost peptidergic DRG neurons lack VGluT expression.Others have noted that nearly all CGRP containing andIB4-binding mouse DRG neurons are immunoreactive forVGluT2 (Brumovsky et al. 2007). We show that VGluT2,but not VGluT1 mRNA was expressed in virtually allTRPV1-expressing lung-specific neurons irrespective of
their placodal vs. neural crest origin, including neuronsexpressing PPT-A. VGluT1 was found to be expressedonly in those neurons not expressing TRPV1 (presumablynon-nociceptor neurons). Our findings are consistent withhistological studies of guinea pig airway-specific vagalneurons that show that VGluT1 identifies a large diameterpopulation of neurons, whereas VGluT2 identifies smallerdiameter neurons (Mazzone & McGovern, 2008). Tonget al. (2001) likewise noted that the majority of nodoseand DRG neurons labelled from the rat stomach have alsobeen found to express VGluT2. Therefore, VGluT1 andVGluT2 expression may differentially be expressed in vagalnociceptive vs. non-nociceptive neurons, but they are notdifferentially expressed in subsets of TRPV1 expressingvisceral nociceptors.
The presence of different C-fibre subpopulationsinnervating the lung was first hypothesized by theColeridges. In dogs, the capsaicin-sensitive vagal C-fibrescould be subdivided into ‘pulmonary’ and a ‘bronchial’subtypes based on different latencies of responses tocapsaicin injection to the pulmonary or the bronchialcirculation, and different sensitivities to phenyl diguanide,bradykinin and ozone (Coleridge & Coleridge, 1977;Kaufman et al. 1980; Coleridge et al. 1993). In guineapigs we had noted that the neural crest, but not placodal,C-fibres innervate large extrapulmonary airways whichwould be consistent with the Colerdidges’ ‘bronchial’C-fibre subtype. The nodose C-fibres deeper in the lungtissue may comprise ‘pulmonary C-fibres’. It is tempting tospeculate, therefore, that the respiratory C-fibres accessibleto stimuli in the external environment (‘extero-receptors’that terminate in the epithelium of the larger airways) aremainly neural crest in nature, whereas those that terminatedeeper within the lung tissue (‘intero-receptors’) are moreplacodal in nature. In any event, the sensations andreflexes that are evoked upon noxious stimuli will likelydepend on the nature of the C-fibre subtype activated.Our data revealing the selective expression of variousneurotrophic factor receptors indicate that the type ofphenotypic neuromodulation that occurs in inflammatoryairway diseases will depend on the subtype and locationof the C-fibre terminals.
References
Baker CV & Schlosser G (2005). The evolutionary origin ofneural crest and placodes. J Exp Zoolog B Mol Dev Evol 304,269–273.
Bespalov MM & Saarma M (2007). GDNF family receptorcomplexes are emerging drug targets. Trends Pharmacol Sci28, 68–74.
Brumovsky P, Watanabe M & Hokfelt T (2007). Expression ofthe vesicular glutamate transporters-1 and -2 in adult mousedorsal root ganglia and spinal cord and their regulation bynerve injury. Neuroscience 147, 469–490.
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

4782 C. Nassenstein and others J Physiol 588.23
Carr MJ, Hunter DD, Jacoby DB & Undem BJ (2002).Expression of tachykinins in nonnociceptive vagal afferentneurons during respiratory viral infection in guinea pigs. AmJ Respir Crit Care Med 165, 1071–1075.
Chen CL, Broom DC, Liu Y, de Nooij JC, Li Z, Cen C, SamadOA, Jessell TM, Woolf CJ & Ma Q (2006). Runx1 determinesnociceptive sensory neuron phenotype and is required forthermal and neuropathic pain. Neuron 49, 365–377.
Chuaychoo B, Hunter DD, Myers AC, Kollarik M & Undem BJ(2005). Allergen-induced substance P synthesis inlarge-diameter sensory neurons innervating the lungs. JAllergy Clin Immunol 116, 325–331.
Coleridge HM & Coleridge JC (1977). Impulse activity inafferent vagal C-fibres with endings in the intrapulmonaryairways of dogs. Respir Physiol 29, 125–142.
Coleridge JC, Coleridge HM, Schelegle ES & Green JF (1993).Acute inhalation of ozone stimulates bronchial C-fibers andrapidly adapting receptors in dogs. J Appl Physiol 74,2345–2352.
D’Amico-Martel A & Noden DM (1983). Contributions ofplacodal and neural crest cells to avian cranial peripheralganglia. Am J Anat 166, 445–468.
Dinh QT, Groneberg DA, Peiser C, Springer J, Joachim RA,Arck PC, Klapp BF & Fischer A (2004). Nerve growthfactor-induced substance P in capsaicin-insensitive vagalneurons innervating the lower mouse airway. Clin ExpAllergy 34, 1474–1479.
Dinh QT, Mingomataj E, Quarcoo D, Groneberg DA, Witt C,Klapp BF, Braun A & Fischer A (2005). Allergic airwayinflammation induces tachykinin peptides expression invagal sensory neurons innervating mouse airways. Clin ExpAllergy 35, 820–825.
Fasanella KE, Christianson JA, Chanthaphavong RS & DavisBM (2008). Distribution and neurochemical identification ofpancreatic afferents in the mouse. J Comp Neurol 509, 42–52.
Hunter DD, Myers AC & Undem BJ (2000). Nerve growthfactor-induced phenotypic switch in guinea pig airwaysensory neurons. Am J Respir Crit Care Med 161, 1985–1990.
Jiang X, Rowitch DH, Soriano P, McMahon AP & Sucov HM(2000). Fate of the mammalian cardiac neural crest.Development 127, 1607–1616.
Josephson A, Widenfalk J, Trifunovski A, Widmer HR, Olson L& Spenger C (2001). GDNF and NGF family members andreceptors in human fetal and adult spinal cord and dorsalroot ganglia. J Comp Neurol 440, 204–217.
Kashiba H, Ueda Y & Senba E (1996). Coexpression ofpreprotachykinin-A, α-calcitonin gene-related peptide,somatostatin, and neurotrophin receptor family messengerRNAs in rat dorsal root ganglion neurons. Neuroscience 70,179–189.
Katz DM & Karten HJ (1980). Substance P in the vagal sensoryganglia: localization in cell bodies and pericellulararborizations. J Comp Neurol 193, 549–564.
Kaufman MP, Coleridge HM, Coleridge JC & Baker DG(1980). Bradykinin stimulates afferent vagal C-fibers inintrapulmonary airways of dogs. J Appl Physiol 48, 511–517.
Kollarik M, Dinh QT, Fischer A & Undem BJ (2003).Capsaicin-sensitive and -insensitive vagalbronchopulmonary C-fibres in the mouse. J Physiol 551,869–879.
Kramer I, Sigrist M, de Nooij JC, Taniuchi I, Jessell TM & ArberS (2006). A role for Runx transcription factor signaling indorsal root ganglion sensory neuron diversification. Neuron49, 379–393.
Kummer W, Fischer A, Kurkowski R & Heym C (1992). Thesensory and sympathetic innervation of guinea-pig lung andtrachea as studied by retrograde neuronal tracing anddouble-labelling immunohistochemistry. Neuroscience 49,715–737.
Kwong K, Kollarik M, Nassenstein C, Ru F & Undem BJ (2008).P2X2 receptors differentiate placodal vs. neural crest C-fiberphenotypes innervating guinea pig lungs and esophagus. AmJ Physiol Lung Cell Mol Physiol 295, L858–L865.
Lundberg JM, Brodin E & Saria A (1983). Effects anddistribution of vagal capsaicin-sensitive substance P neuronswith special reference to the trachea and lungs. Acta PhysiolScand 119, 243–252.
MacGlashan D Jr (1989). Single-cell analysis of Ca++ changesin human lung mast cells: graded vs. all-or-nothingelevations after IgE-mediated stimulation. J Cell Biol 109,123–134.
Makita T, Sucov HM, Gariepy CE, Yanagisawa M & Ginty DD(2008). Endothelins are vascular-derived axonal guidancecues for developing sympathetic neurons. Nature 452,759–763.
Mazzone SB & McGovern AE (2008). Immunohistochemicalcharacterization of nodose cough receptor neuronsprojecting to the trachea of guinea pigs. Cough 4, 9.
Molliver DC, Radeke MJ, Feinstein SC & Snider WD (1995).Presence or absence of TrkA protein distinguishes subsets ofsmall sensory neurons with unique cytochemicalcharacteristics and dorsal horn projections. J Comp Neurol361, 404–416.
Morris JL, Konig P, Shimizu T, Jobling P & Gibbins IL (2005).Most peptide-containing sensory neurons lack proteins forexocytotic release and vesicular transport of glutamate. JComp Neurol 483, 1–16.
Oliveira AL, Hydling F, Olsson E, Shi T, Edwards RH, FujiyamaF, Kaneko T, Hokfelt T, Cullheim S & Meister B (2003).Cellular localization of three vesicular glutamate transportermRNAs and proteins in rat spinal cord and dorsal rootganglia. Synapse 50, 117–129.
Ricco MM, Kummer W, Biglari B, Myers AC & Undem BJ(1996). Interganglionic segregation of distinct vagal afferentfibre phenotypes in guinea-pig airways. J Physiol 496,521–530.
Springall DR, Cadieux A, Oliveira H, Su H, Royston D & PolakJM (1987). Retrograde tracing shows that CGRP-immunoreactive nerves of rat trachea and lung originatefrom vagal and dorsal root ganglia. J Auton Nerv Syst 20,155–166.
Taylor-Clark T & Undem BJ (2006). Transductionmechanisms in airway sensory nerves. J Appl Physiol 101,950–959.
Taylor-Clark TE, McAlexander MA, Nassenstein C, SheardownSA, Wilson S, Thornton J, Carr MJ & Undem BJ (2008).Relative contributions of TRPA1 and TRPV1 channels in theactivation of vagal bronchopulmonary C-fibres by theendogenous autacoid 4-oxononenal. J Physiol 586,3447–3459.
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

J Physiol 588.23 Vagal C-fibre subtypes innervating the lung 4783
Tong Q, Ma J & Kirchgessner AL (2001). Vesicular glutamatetransporter 2 in the brain-gut axis. Neuroreport 12,3929–3934.
Undem BJ, Chuaychoo B, Lee MG, Weinreich D, Myers AC &Kollarik M (2004). Subtypes of vagal afferent C-fibres inguinea-pig lungs. J Physiol 556, 905–917.
Woolf CJ & Ma Q (2007). Nociceptors: noxious stimulusdetectors. Neuron 55, 353–364.
Yntema CL (1942). Experiments on the origin of some of someof the cranial sensory ganglia in the chick. Anat Rec 82, 455.
Yoshikawa M, Senzaki K, Yokomizo T, Takahashi S, Ozaki S &Shiga T (2007). Runx1 selectively regulates cell fatespecification and axonal projections of dorsal root ganglionneurons. Dev Biol 303, 663–674.
Yu S, Undem BJ & Kollarik M (2005). Vagal afferent nerveswith nociceptive properties in guinea-pig oesophagus.J Physiol 563, 831–842.
Author contributions
C.N. and B.J.U. were responsible for the conception and designof the experiments, and the drafting of the article; C.N., T.E.T.C.,A.M., F.R., R.N., W.B. and B.J.U. collected, analysed and inter-preted the data. All authors approved the final version of themanuscript.
Acknowledgements
This work was funded by The National Institutes of Health (NIH)and the German Research Foundation (DFG NA 836/2-1). Theauthors thank Mrs Sonya Meeker and Ms Silke Wiegand forexcellent technical assistance and D. D. Ginty and T. Makita forproviding the Wnt1Cre/R26R mice.
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society





























