Embed Size (px)
Citation preview

Plant Cell, Tissue and Organ Culture60: 139–149, 2000.© 2000Kluwer Academic Publishers. Printed in the Netherlands.
139
Phloroglucinol enhances growth and rate of axillary shoot proliferation inpotato shoot tip cultures in vitro
Debabrata Sarkar∗ & Prakash S. NaikDivision of Crop Improvement, Central Potato Research Institute, Shimla-171 001, Himachal Pradesh, India(∗requests for offprints; Fax: +91-177 224460; E-mail: [email protected])
Received 12 November 1999; accepted in revised form 17 April 2000
Key words:1,3,5-benzenetriol, Dilospan S, meristem culture, micropropagation, phenolic compound, phloroglu-cinol, Solanum tuberosumL., 1,3,5-trihydroxybenzene
Abstract
Efficacy of phloroglucinol in promoting growth and development ofin vitro-derived shoot tips was studied in sixpotato (Solanum tuberosumL.) genotypes. Different concentrations of phloroglucinol (0, 0.08, 0.4, 0.8, 1.2 and1.6 mM) were tested in combination with either 0.1 or 0.2 M sucrose in shoot tip proliferation medium based onMS (Murashige and Skoog, 1962) medium supplemented with 5.8µM GA3 (gibberellic acid), 1.1µM BA (N6-benzyladenine) and 8.39µM D-calcium pantothenate. Phloroglucinol fostered multiple shoot formation, promotedaxillary shoot proliferation in terms of shoot tip fresh weight and shoot length, and stimulated root formation on theshoot tips. There was significant phloroglucinol× sucrose interaction for number of shoots developed per shoot tip,shoot tip fresh weight and number of roots induced per shoot tip. The beneficial effect of phloroglucinol on shoottip survival was conspicuous only in genotypes that showed poor survival in the control proliferation medium.There were significant differences in response between the two sucrose levels with regard to shoot tip fresh weightand number of roots per shoot tip. Phloroglucinol in combination with 0.2 M sucrose induced maximum numberof roots per shoot tip. Optimum shoot tip growth was fostered in medium containing 0.8 mM phloroglucinol and0.2 M sucrose. High frequency multiple shoot formation in this medium ensures a faster rate of potato shoot tipmultiplication within a limited time and space.
Abbreviations: BA – N6-benzyladenine; GA3 – gibberellic acid; IAA – indole-3-acetic acid; MS medium –Murashige and Skoog’s medium (1962); PG – phloroglucinol
Introduction
In potato (Solanum tuberosumL.), shoot tip cultureis used in combination with thermotherapy and/orchemotherapy for obtaining virus-free plants. Themethod basically involves removal of very small tipsof the growing shoots, usually 300–500µm in size,and growing these on suitable nutrient medium underdefined cultural conditions. Although the cultural re-quirements for potato shoot tips are not very strict,they grow very slowly in cultures (Tao et al., 1987).In addition, the shoot tips do not readily root in theproliferation/differentiation medium, thus requiring aseparate rooting stage (Lozoya-Saldana and Madrigal-
Vargas, 1985). Therefore, over the years, differentmedia and culture schedules have been developed forgrowing potato shoot tipsin vitro (Cassells, 1987;Mellor and Stace-Smith, 1987). However, these elab-orate culture schedules limit the effectiveness of thistechnique for the production of disease-free plants,since it often takes from four to eight months, withthe number of produced individuals being very small(Sanchez et al., 1991).
The growth-regulating properties of the phenoliccompounds and their glycosides are well documentedin plant tissue cultures (Murashige, 1974; James andThurbon, 1981a; George and Sherrington, 1984).Phloroglucinol (syn. 1,3,5-trihydroxybenzene), one of

140
the degradation products of phloridzin, is known forits growth-regulating property. Workers at the EastMailing Research Station in England have reportedthat phloroglucinol enhances growth and rate of ax-illary shoot proliferation from shoot tip cultures ofseveral woody plants (Jones, 1976) or acts as an auxinsynergist during the auxin-sensitive phase of root ini-tiation (James and Thurbon, 1981b). Preconditioningof shoot cultures by phloroglucinol has been reportedto initiate adventitious roots in different woody spe-cies in vitro (James, 1979, 1983). Subsequently, theeffectiveness of this phenolic compound as a growthregulator has been reported in other studies also (Ham-matt and Grant, 1997; Ramirez-Malagon et al., 1997;Al-Wasel, 1998; Zanol et al., 1998). However, themost interesting aspect of this phenolic compound isits reported ability to enhance survival of, and shootformation from small tissue explants like meristemsand/or shoot tipsin vitro (Goudarzi et al., 1997; De-miralay et al., 1998). There are no reports on the effectof phloroglucinol on potato shoot tip culturevis-à-vis micropropagation. For its beneficial role in shoottip proliferation and promotive effect on root formingactivity in different plant species, we decided to exam-ine what influence PG might have on potato shoot tipculturesin vitro.
Materials and methods
Plant materials
Six potato (Solanum tuberosumL.) genotypes wereused in the present study: Kufri Chipsona-1, KufriChipsona-2 and Kufri Jyoti belong toS. tuberosumssp. tuberosum; CUP-199 toS. tuberosumssp. an-digena; Kufri Lalima and Kufri Sindhuri are hybridsof andigenaand tuberosum. They represent a widegenetic base, and were selected because of their con-trasting response to shoot tip culture as observedin our laboratory. These genotypes were maintainedand multiplied through shoot cuttings following themethod described earlier by Sarkar et al. (1997).
Shoot tip culture
Apical shoot tips about 300–500µm long were dis-sected from 30-day-old aseptically grownin vitroplantlets following the procedure previously described(Sarkar and Naik, 1998). Ten shoot tips were cul-tured per Petri dish (sterile polystyrene; ø 90 mm)containing 30 ml of MS (Murashige and Skoog, 1962)
medium supplemented with 5.8µM GA3, 1.1µM BA,8.39µM D-calcium pantothenate and either 0.1 or 0.2M sucrose. The medium was solidified with 7 g l−1
Nobel agar (HiMedia, India). Petri dishes were sealedwith Parafilm M (™American National Can, USA),and were incubated under a 16-h photoperiod (approx.50µmol m−2 s−1 light intensity) at 24±1 ◦C.
PG-sucrose treatments
Freshly prepared PG (P 1178, Sigma, USA; anhyd-rous; MW 126.1) stock solutions were filter-sterilizedand diluted as required. Altogether, five concentra-tions of PG (0.08, 0.4, 0.8, 1.2 and 1.6 mM) weretested in combination with either 0.1 or 0.2 M sucrose.Shoot tip culture medium without PG was used as thecontrol.
Data observations and analyses
After six weeks’ incubation, observations were re-corded on the number of shoots developed per shoottip, shoot length (mm), shoot tip fresh weight (mg),number of roots developed per shoot tip and per-centage shoot tip survival. As there were ten shoottips per Petri dish, data were recorded and averaged.The experiment was conducted in a factorial (6× 2× 6) completely randomised design with six geno-types, two concentrations of sucrose (0.1 and 0.2 M)and six concentrations of PG (0, 0.08, 0.4, 0.8, 1.2and 1.6 mM). Each treatment comprised three rep-licate Petri dishes, and the experiment was repeatedonce. Before statistical analyses, data on percentageshoot tip survival and number of roots per shoot tipwere transformed into arc sine square root and squareroot (
√x+0.5), respectively. As the experiment was
conducted twice, data were pooled over individualexperiments, and the three-way analyses of variance(six genotypes× two levels of sucrose× six levelsof PG) were computed using the standard procedure(Steel and Torrie, 1980). Mean separations were car-ried out by LSD test, and associations between shootand root parameters were estimated by nonparametricmethod of Kendall’s coefficient (tau-b) of rank correl-ation (Sokal and Rohlf, 1996) using statistical packageSPSS, Release 6.0.
Results
The analyses of variance showed that PG had a majoreffect on all the characters studied. The variation due
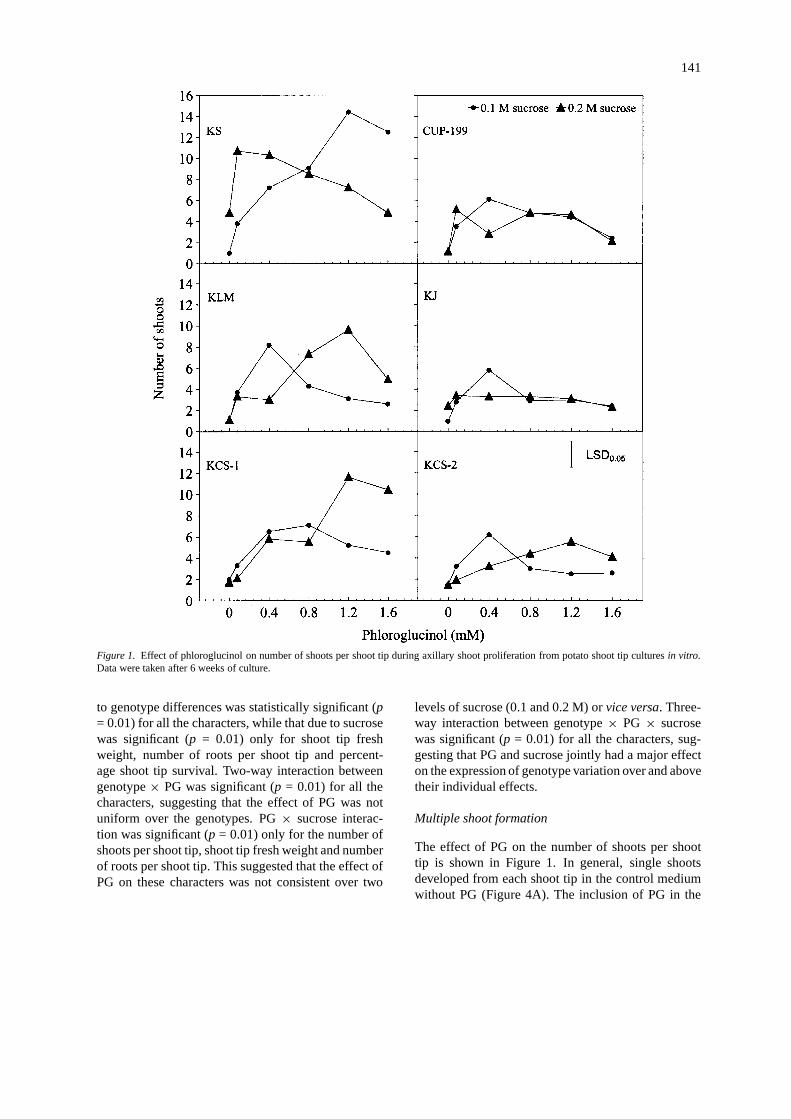
141
Figure 1. Effect of phloroglucinol on number of shoots per shoot tip during axillary shoot proliferation from potato shoot tip culturesin vitro.Data were taken after 6 weeks of culture.
to genotype differences was statistically significant (p= 0.01) for all the characters, while that due to sucrosewas significant (p = 0.01) only for shoot tip freshweight, number of roots per shoot tip and percent-age shoot tip survival. Two-way interaction betweengenotype× PG was significant (p = 0.01) for all thecharacters, suggesting that the effect of PG was notuniform over the genotypes. PG× sucrose interac-tion was significant (p = 0.01) only for the number ofshoots per shoot tip, shoot tip fresh weight and numberof roots per shoot tip. This suggested that the effect ofPG on these characters was not consistent over two
levels of sucrose (0.1 and 0.2 M) orvice versa. Three-way interaction between genotype× PG × sucrosewas significant (p = 0.01) for all the characters, sug-gesting that PG and sucrose jointly had a major effecton the expression of genotype variation over and abovetheir individual effects.
Multiple shoot formation
The effect of PG on the number of shoots per shoottip is shown in Figure 1. In general, single shootsdeveloped from each shoot tip in the control mediumwithout PG (Figure 4A). The inclusion of PG in the
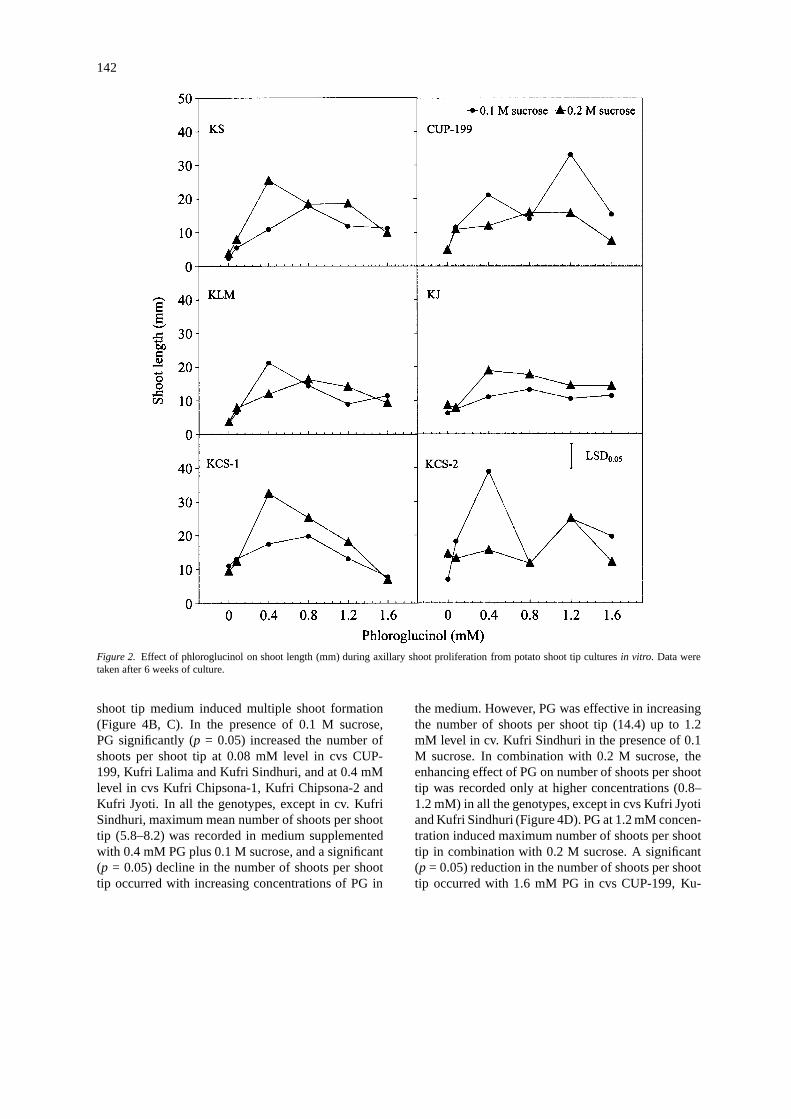
142
Figure 2. Effect of phloroglucinol on shoot length (mm) during axillary shoot proliferation from potato shoot tip culturesin vitro. Data weretaken after 6 weeks of culture.
shoot tip medium induced multiple shoot formation(Figure 4B, C). In the presence of 0.1 M sucrose,PG significantly (p = 0.05) increased the number ofshoots per shoot tip at 0.08 mM level in cvs CUP-199, Kufri Lalima and Kufri Sindhuri, and at 0.4 mMlevel in cvs Kufri Chipsona-1, Kufri Chipsona-2 andKufri Jyoti. In all the genotypes, except in cv. KufriSindhuri, maximum mean number of shoots per shoottip (5.8–8.2) was recorded in medium supplementedwith 0.4 mM PG plus 0.1 M sucrose, and a significant(p = 0.05) decline in the number of shoots per shoottip occurred with increasing concentrations of PG in
the medium. However, PG was effective in increasingthe number of shoots per shoot tip (14.4) up to 1.2mM level in cv. Kufri Sindhuri in the presence of 0.1M sucrose. In combination with 0.2 M sucrose, theenhancing effect of PG on number of shoots per shoottip was recorded only at higher concentrations (0.8–1.2 mM) in all the genotypes, except in cvs Kufri Jyotiand Kufri Sindhuri (Figure 4D). PG at 1.2 mM concen-tration induced maximum number of shoots per shoottip in combination with 0.2 M sucrose. A significant(p = 0.05) reduction in the number of shoots per shoottip occurred with 1.6 mM PG in cvs CUP-199, Ku-
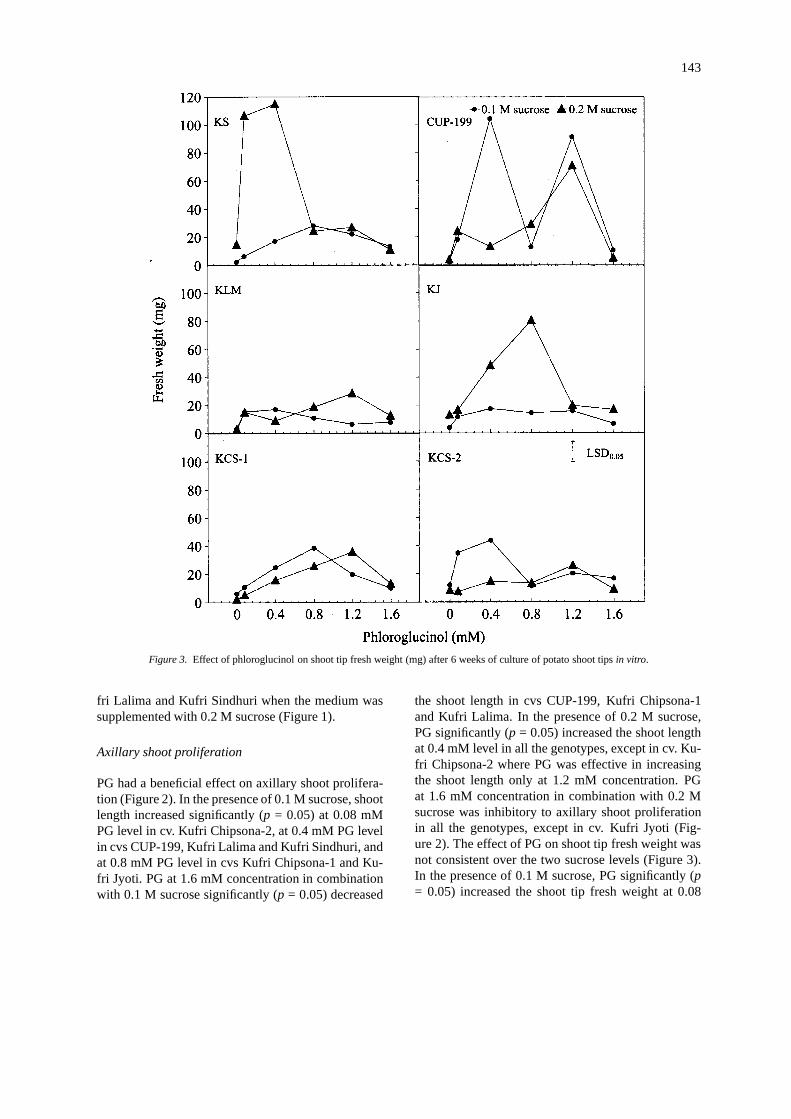
143
Figure 3. Effect of phloroglucinol on shoot tip fresh weight (mg) after 6 weeks of culture of potato shoot tipsin vitro.
fri Lalima and Kufri Sindhuri when the medium wassupplemented with 0.2 M sucrose (Figure 1).
Axillary shoot proliferation
PG had a beneficial effect on axillary shoot prolifera-tion (Figure 2). In the presence of 0.1 M sucrose, shootlength increased significantly (p = 0.05) at 0.08 mMPG level in cv. Kufri Chipsona-2, at 0.4 mM PG levelin cvs CUP-199, Kufri Lalima and Kufri Sindhuri, andat 0.8 mM PG level in cvs Kufri Chipsona-1 and Ku-fri Jyoti. PG at 1.6 mM concentration in combinationwith 0.1 M sucrose significantly (p = 0.05) decreased
the shoot length in cvs CUP-199, Kufri Chipsona-1and Kufri Lalima. In the presence of 0.2 M sucrose,PG significantly (p = 0.05) increased the shoot lengthat 0.4 mM level in all the genotypes, except in cv. Ku-fri Chipsona-2 where PG was effective in increasingthe shoot length only at 1.2 mM concentration. PGat 1.6 mM concentration in combination with 0.2 Msucrose was inhibitory to axillary shoot proliferationin all the genotypes, except in cv. Kufri Jyoti (Fig-ure 2). The effect of PG on shoot tip fresh weight wasnot consistent over the two sucrose levels (Figure 3).In the presence of 0.1 M sucrose, PG significantly (p= 0.05) increased the shoot tip fresh weight at 0.08
144
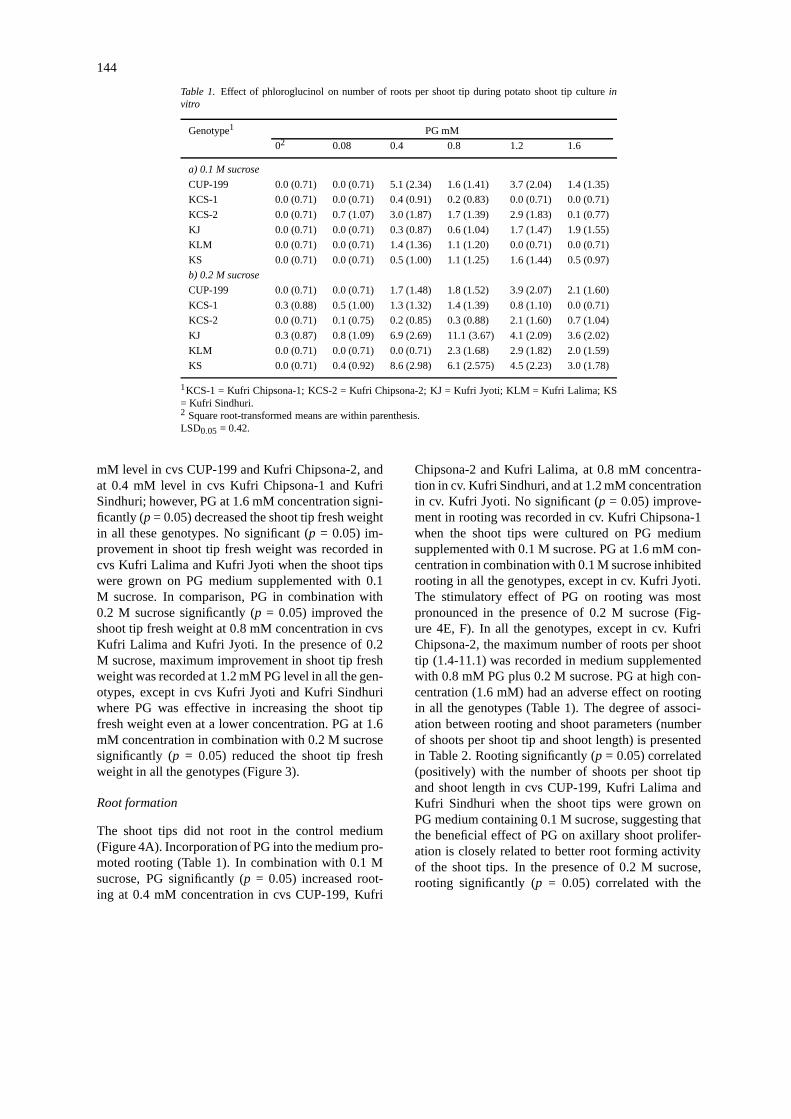
Table 1. Effect of phloroglucinol on number of roots per shoot tip during potato shoot tip cultureinvitro
Genotype1 PG mM
02 0.08 0.4 0.8 1.2 1.6
a) 0.1 M sucrose
CUP-199 0.0 (0.71) 0.0 (0.71) 5.1 (2.34) 1.6 (1.41) 3.7 (2.04) 1.4 (1.35)
KCS-1 0.0 (0.71) 0.0 (0.71) 0.4 (0.91) 0.2 (0.83) 0.0 (0.71) 0.0 (0.71)
KCS-2 0.0 (0.71) 0.7 (1.07) 3.0 (1.87) 1.7 (1.39) 2.9 (1.83) 0.1 (0.77)
KJ 0.0 (0.71) 0.0 (0.71) 0.3 (0.87) 0.6 (1.04) 1.7 (1.47) 1.9 (1.55)
KLM 0.0 (0.71) 0.0 (0.71) 1.4 (1.36) 1.1 (1.20) 0.0 (0.71) 0.0 (0.71)
KS 0.0 (0.71) 0.0 (0.71) 0.5 (1.00) 1.1 (1.25) 1.6 (1.44) 0.5 (0.97)
b) 0.2 M sucrose
CUP-199 0.0 (0.71) 0.0 (0.71) 1.7 (1.48) 1.8 (1.52) 3.9 (2.07) 2.1 (1.60)
KCS-1 0.3 (0.88) 0.5 (1.00) 1.3 (1.32) 1.4 (1.39) 0.8 (1.10) 0.0 (0.71)
KCS-2 0.0 (0.71) 0.1 (0.75) 0.2 (0.85) 0.3 (0.88) 2.1 (1.60) 0.7 (1.04)
KJ 0.3 (0.87) 0.8 (1.09) 6.9 (2.69) 11.1 (3.67) 4.1 (2.09) 3.6 (2.02)
KLM 0.0 (0.71) 0.0 (0.71) 0.0 (0.71) 2.3 (1.68) 2.9 (1.82) 2.0 (1.59)
KS 0.0 (0.71) 0.4 (0.92) 8.6 (2.98) 6.1 (2.575) 4.5 (2.23) 3.0 (1.78)
1KCS-1 = Kufri Chipsona-1; KCS-2 = Kufri Chipsona-2; KJ = Kufri Jyoti; KLM = Kufri Lalima; KS= Kufri Sindhuri.2 Square root-transformed means are within parenthesis.LSD0.05 = 0.42.
mM level in cvs CUP-199 and Kufri Chipsona-2, andat 0.4 mM level in cvs Kufri Chipsona-1 and KufriSindhuri; however, PG at 1.6 mM concentration signi-ficantly (p = 0.05) decreased the shoot tip fresh weightin all these genotypes. No significant (p = 0.05) im-provement in shoot tip fresh weight was recorded incvs Kufri Lalima and Kufri Jyoti when the shoot tipswere grown on PG medium supplemented with 0.1M sucrose. In comparison, PG in combination with0.2 M sucrose significantly (p = 0.05) improved theshoot tip fresh weight at 0.8 mM concentration in cvsKufri Lalima and Kufri Jyoti. In the presence of 0.2M sucrose, maximum improvement in shoot tip freshweight was recorded at 1.2 mM PG level in all the gen-otypes, except in cvs Kufri Jyoti and Kufri Sindhuriwhere PG was effective in increasing the shoot tipfresh weight even at a lower concentration. PG at 1.6mM concentration in combination with 0.2 M sucrosesignificantly (p = 0.05) reduced the shoot tip freshweight in all the genotypes (Figure 3).
Root formation
The shoot tips did not root in the control medium(Figure 4A). Incorporation of PG into the medium pro-moted rooting (Table 1). In combination with 0.1 Msucrose, PG significantly (p = 0.05) increased root-ing at 0.4 mM concentration in cvs CUP-199, Kufri
Chipsona-2 and Kufri Lalima, at 0.8 mM concentra-tion in cv. Kufri Sindhuri, and at 1.2 mM concentrationin cv. Kufri Jyoti. No significant (p = 0.05) improve-ment in rooting was recorded in cv. Kufri Chipsona-1when the shoot tips were cultured on PG mediumsupplemented with 0.1 M sucrose. PG at 1.6 mM con-centration in combination with 0.1 M sucrose inhibitedrooting in all the genotypes, except in cv. Kufri Jyoti.The stimulatory effect of PG on rooting was mostpronounced in the presence of 0.2 M sucrose (Fig-ure 4E, F). In all the genotypes, except in cv. KufriChipsona-2, the maximum number of roots per shoottip (1.4-11.1) was recorded in medium supplementedwith 0.8 mM PG plus 0.2 M sucrose. PG at high con-centration (1.6 mM) had an adverse effect on rootingin all the genotypes (Table 1). The degree of associ-ation between rooting and shoot parameters (numberof shoots per shoot tip and shoot length) is presentedin Table 2. Rooting significantly (p = 0.05) correlated(positively) with the number of shoots per shoot tipand shoot length in cvs CUP-199, Kufri Lalima andKufri Sindhuri when the shoot tips were grown onPG medium containing 0.1 M sucrose, suggesting thatthe beneficial effect of PG on axillary shoot prolifer-ation is closely related to better root forming activityof the shoot tips. In the presence of 0.2 M sucrose,rooting significantly (p = 0.05) correlated with the

145
Table 2. Kendall’s coefficient (τ )1 of rank correlation between number of roots pershoot tip and shoot parameters during potato shoot tip culture in phloroglucinol media.
Genotype2 Number of shoots Shoot length
0.1 M sucrose 0.2 M sucrose 0.1 M sucrose 0.2 M sucrose
CUP-199 0.690∗ 0.000ns 0.690∗ 0.414ns
KCS-1 0.602ns 0.067ns 0.624ns 0.867∗∗KCS-2 0.467ns 0.867∗∗ 0.600ns 0.067ns
KJ 0.138ns 0.276ns 0.552ns 0.733∗KLM 0.775∗∗ 0.894∗∗ 0.775∗∗ 0.596ns
KS 0.788∗ 0.414ns 0.788∗ 0.867∗∗
1 τ = N/n (n-1), wheren is the conventional sample size andN is a count of ranks;n = 6.2 Same as in Table 1.∗, ∗∗ Significant atp≤ 0.05 andp≤ 0.01, respectively; ns = non-significant.
number of shoots per shoot tip in cvs Kufri Chipsona-2 and Kufri Lalima, and with shoot length in cvs KufriChipsona-1, Kufri Jyoti and Kufri Sindhuri (Table 2).
Percentage shoot tip survival
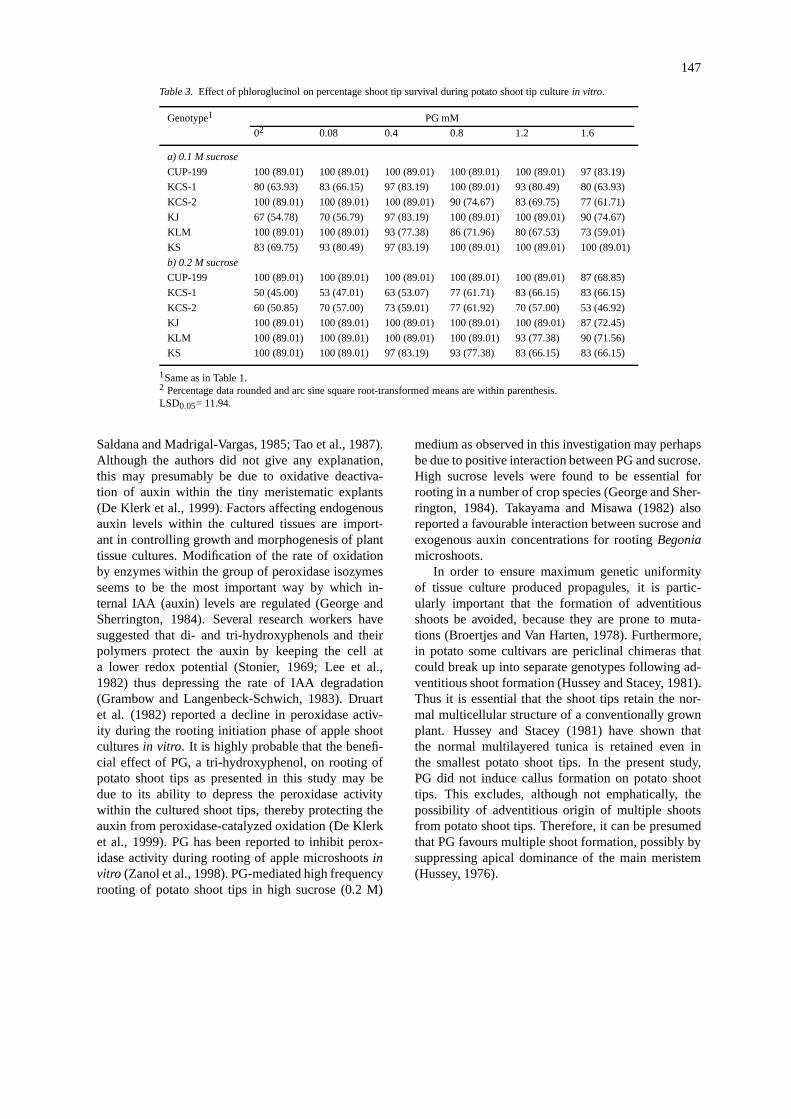
The effect of PG on percentage shoot tip survival isshown in Table 3. In cvs Kufri Chipsona-1, KufriJyoti and Kufri Sindhuri, PG at 0.4 mM concentra-tion in combination with 0.1 M sucrose significantly(p = 0.05) increased the percentage shoot tip survival.However, except in cv. Kufri Chipsona-1, PG didnot have any beneficial effect on shoot tip survivalin cvs Kufri Jyoti and Kufri Sindhuri when the me-dium was supplemented with 0.2 M sucrose; rather PGat a higher concentration in combination with 0.2 Msucrose was detrimental to shoot tip survival. In con-trast, optimum shoot tip survival was recorded even inthe control medium in cvs CUP-199, Kufri Chipsona-2 and Kufri Lalima when the shoot tips were culturedon medium containing 0.1 M sucrose. PG at 0.8 mMlevel in combination with 0.1 M sucrose decreased thepercentage shoot tip survival in cvs Kufri Chipsona-2and Kufri Lalima. However, in the presence of 0.2 Msucrose, PG was inhibitory to shoot tip survival only at1.6 mM level in cv. Kufri Lalima, and did not have anypromoting effect on survival in cv. Kufri Chipsona-2as compared to the control (Table 3).
Discussion
The present study showed that PG had a beneficialeffect on potato shoot tip survival and growth. Thebeneficial effect of PG on shoot tip survival was con-spicuous only in genotypes that showed poor survival
in the control shoot tip medium. However, PG signi-ficantly improved shoot tip growth and developmentin all the genotypes. It not only induced multiple shootformation, but also enhanced axillary shoot prolifera-tion. The effectiveness of certain phenolic substancesin enhancing shoot formation is well established inplant tissue cultures (Murashige, 1974; George andSherrington, 1984). As early as in 1976, Jones (1976)reported the positive influence of PG, a phenolic com-pound, on apple shoot tip growth. The effectivenessof this phenolic substance in enhancing shoot pro-liferation vis-à-visshoot tip growth was reported inother crop species also (Hunter, 1979; Pontikis andSapoutzaki, 1984; Mosella and Fernandez, 1985;Pontikis and Melas, 1986; Aklan et al., 1997; Jordanand Piwanski, 1997; Demiralay et al., 1998). In allthese experiments, the results were based on testing ofonly single concentration (0.5–1.0 mM) of PG. Andfurthermore, the interactive influence of PG in associ-ation with sucrose was never studied. In comparison, awide range of PG was tested in the present experimentin combination with 0.1 or 0.2 M sucrose. The studyclearly demonstrated that PG significantly interactedwith sucrose for multiple shoot formation and axillaryshoot proliferation during potato shoot tip culturesinvitro. We observed close relationship between rootingand shoot parameters in some of the genotypes duringpotato shoo tip cultures under the promotive influenceof PG. PG-mediated better root forming activity whichwill be discussed in the following section may pre-sumably be responsible for enhanced growth of potatoshoot tip culturesin vitro.
The most prominent interactive effect between PGand sucrose was evident in the rooting behaviour ofthe shoot tips. The promotive effect of PG on rooting
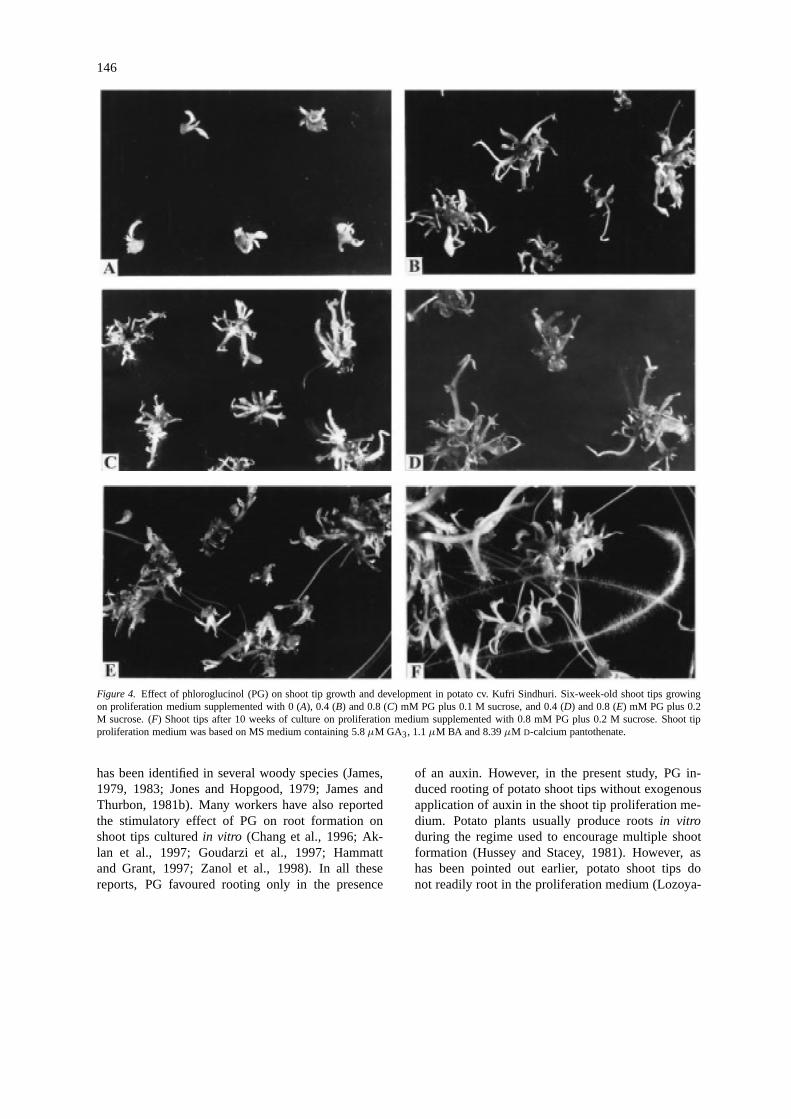
146
Figure 4. Effect of phloroglucinol (PG) on shoot tip growth and development in potato cv. Kufri Sindhuri. Six-week-old shoot tips growingon proliferation medium supplemented with 0 (A), 0.4 (B) and 0.8 (C) mM PG plus 0.1 M sucrose, and 0.4 (D) and 0.8 (E) mM PG plus 0.2M sucrose. (F) Shoot tips after 10 weeks of culture on proliferation medium supplemented with 0.8 mM PG plus 0.2 M sucrose. Shoot tipproliferation medium was based on MS medium containing 5.8µM GA3, 1.1µM BA and 8.39µM D-calcium pantothenate.
has been identified in several woody species (James,1979, 1983; Jones and Hopgood, 1979; James andThurbon, 1981b). Many workers have also reportedthe stimulatory effect of PG on root formation onshoot tips culturedin vitro (Chang et al., 1996; Ak-lan et al., 1997; Goudarzi et al., 1997; Hammattand Grant, 1997; Zanol et al., 1998). In all thesereports, PG favoured rooting only in the presence
of an auxin. However, in the present study, PG in-duced rooting of potato shoot tips without exogenousapplication of auxin in the shoot tip proliferation me-dium. Potato plants usually produce rootsin vitroduring the regime used to encourage multiple shootformation (Hussey and Stacey, 1981). However, ashas been pointed out earlier, potato shoot tips donot readily root in the proliferation medium (Lozoya-
147
Table 3. Effect of phloroglucinol on percentage shoot tip survival during potato shoot tip culturein vitro.
Genotype1 PG mM
02 0.08 0.4 0.8 1.2 1.6
a) 0.1 M sucrose
CUP-199 100 (89.01) 100 (89.01) 100 (89.01) 100 (89.01) 100 (89.01) 97 (83.19)
KCS-1 80 (63.93) 83 (66.15) 97 (83.19) 100 (89.01) 93 (80.49) 80 (63.93)
KCS-2 100 (89.01) 100 (89.01) 100 (89.01) 90 (74.67) 83 (69.75) 77 (61.71)
KJ 67 (54.78) 70 (56.79) 97 (83.19) 100 (89.01) 100 (89.01) 90 (74.67)
KLM 100 (89.01) 100 (89.01) 93 (77.38) 86 (71.96) 80 (67.53) 73 (59.01)
KS 83 (69.75) 93 (80.49) 97 (83.19) 100 (89.01) 100 (89.01) 100 (89.01)
b) 0.2 M sucrose
CUP-199 100 (89.01) 100 (89.01) 100 (89.01) 100 (89.01) 100 (89.01) 87 (68.85)
KCS-1 50 (45.00) 53 (47.01) 63 (53.07) 77 (61.71) 83 (66.15) 83 (66.15)
KCS-2 60 (50.85) 70 (57.00) 73 (59.01) 77 (61.92) 70 (57.00) 53 (46.92)
KJ 100 (89.01) 100 (89.01) 100 (89.01) 100 (89.01) 100 (89.01) 87 (72.45)
KLM 100 (89.01) 100 (89.01) 100 (89.01) 100 (89.01) 93 (77.38) 90 (71.56)
KS 100 (89.01) 100 (89.01) 97 (83.19) 93 (77.38) 83 (66.15) 83 (66.15)
1Same as in Table 1.2 Percentage data rounded and arc sine square root-transformed means are within parenthesis.LSD0.05= 11.94.
Saldana and Madrigal-Vargas, 1985; Tao et al., 1987).Although the authors did not give any explanation,this may presumably be due to oxidative deactiva-tion of auxin within the tiny meristematic explants(De Klerk et al., 1999). Factors affecting endogenousauxin levels within the cultured tissues are import-ant in controlling growth and morphogenesis of planttissue cultures. Modification of the rate of oxidationby enzymes within the group of peroxidase isozymesseems to be the most important way by which in-ternal IAA (auxin) levels are regulated (George andSherrington, 1984). Several research workers havesuggested that di- and tri-hydroxyphenols and theirpolymers protect the auxin by keeping the cell ata lower redox potential (Stonier, 1969; Lee et al.,1982) thus depressing the rate of IAA degradation(Grambow and Langenbeck-Schwich, 1983). Druartet al. (1982) reported a decline in peroxidase activ-ity during the rooting initiation phase of apple shootculturesin vitro. It is highly probable that the benefi-cial effect of PG, a tri-hydroxyphenol, on rooting ofpotato shoot tips as presented in this study may bedue to its ability to depress the peroxidase activitywithin the cultured shoot tips, thereby protecting theauxin from peroxidase-catalyzed oxidation (De Klerket al., 1999). PG has been reported to inhibit perox-idase activity during rooting of apple microshootsinvitro (Zanol et al., 1998). PG-mediated high frequencyrooting of potato shoot tips in high sucrose (0.2 M)
medium as observed in this investigation may perhapsbe due to positive interaction between PG and sucrose.High sucrose levels were found to be essential forrooting in a number of crop species (George and Sher-rington, 1984). Takayama and Misawa (1982) alsoreported a favourable interaction between sucrose andexogenous auxin concentrations for rootingBegoniamicroshoots.
In order to ensure maximum genetic uniformityof tissue culture produced propagules, it is partic-ularly important that the formation of adventitiousshoots be avoided, because they are prone to muta-tions (Broertjes and Van Harten, 1978). Furthermore,in potato some cultivars are periclinal chimeras thatcould break up into separate genotypes following ad-ventitious shoot formation (Hussey and Stacey, 1981).Thus it is essential that the shoot tips retain the nor-mal multicellular structure of a conventionally grownplant. Hussey and Stacey (1981) have shown thatthe normal multilayered tunica is retained even inthe smallest potato shoot tips. In the present study,PG did not induce callus formation on potato shoottips. This excludes, although not emphatically, thepossibility of adventitious origin of multiple shootsfrom potato shoot tips. Therefore, it can be presumedthat PG favours multiple shoot formation, possibly bysuppressing apical dominance of the main meristem(Hussey, 1976).

148
Slow growth of potato shoot tips in culture is dueto reduced rate of axillary shoot proliferation and inab-ility of shoot tips to root in the proliferation medium.From the present experiment, it is clear that PG canfavourably be used to promote potato shoot tip growthin culture. To foster optimum growth duringin vitroculture of potato shoot tips, 0.8 mM PG is recom-mended for use in combination with 0.2 M sucrosein the shoot tip proliferation medium. High frequencymultiple shoot formation in this medium will ensurea faster rate of shoot tip multiplication within a lim-ited time. And, most importantly, the amenability ofthe shoot tips to rooting in this proliferation mediumwill eliminate the requirement for a separate rootingstage. This will greatly save on time and labour forthe production of virus-free plant materialsvis-à-vismicropropagation in potato.
Acknowledgements
We thank Dr GS Shekhawat, Director, Central PotatoResearch Institute, Shimla and Dr PC Gaur, Head,Division of Crop Improvement for providing neces-sary facilities. Photographic assistance from Mr SKDey is much appreciated. The comments and sugges-tions on the manuscript from Dr GJ de Klerk and twoanonymous reviewers are gratefully acknowledged.
References
Aklan K, Cetiner S, Aka-Kacar Y, Yalcin-Mendi Y, Kuden AB &Dennis FG Jr (1997)In vitro multiplication of clonal apple root-stocks M. 9, M. 26 and MM. 106 by meristem culture. Acta Hort.441: 325–327
Al-Wasel AS (1998)In vitro propagation ofAtriplex nummulariaL.Arab Univ. J. Agric. Sci. 6: 373–383
Broertjes C & Van Harten AM (1978) Application of MutationBreeding Methods in the Improvement of Vegetatively Propag-ated Crops. Elsevier Scientific Publishing Company, Amsterdam
Cassells AC (1987)In vitro induction of virus-free potatoes bychemotherapy. In: Bajaj YPS (ed) Biotechnology in Agricultureand Forestry, Vol 3, Potato (pp 40–50). Springer-Verlag, Berlin,Heidelberg, New York, Tokyo
Chang DCN, Peng KH, Nichols M & Swain D (1996) Phloroglu-cinol and tryptone enhancein vitro rooting and survival rate ofasparagus nodal sections. Acta Hortic. 415: 411–416
De Klerk GJ, vander Krieken W & de Jong JC (1999) The formationof adventitious roots: new concepts, new possibilities.In VitroCell. Dev. Biol. Plant 35: 189–199
Demiralay A, Yalcin-Mendi Y, Aka-Kacar Y, Cetiner S, Aksoy U,Ferguson L & Hepksoy S (1998)In vitro propagation ofFicus ca-rica L. var. Bursa Siyahi through meristem culture. Acta Hortic.480: 165–167
Druart Ph, Kevers C, Boxus Ph & Gaspar Th (1982)In vitro promo-tion of root formation by apple shoots through darkness effect onendogenous phenols and peroxidases. Z. Pflanzenphysiol. 108:429–436
George EF & Sherrington PD (1984) Plant Propagation by TissueCulture: Handbook and Directory of Commercial Laboratories.Exegetics Ltd., London
Goudarzi R, Majedi A, Talaie AR & Mostafavi M (1997) Microp-ropagation of cherry rootstock (Prunus aviumcv. F12/1) by shoottip culture. Iranian J. Agric. Sci. 28: 133–143
Grambow HJ & Langenbeck-Schwich B (1983) The relationshipbetween oxidase activity, peroxidase activity, hydrogen peroxide,and phenolic compounds in the degradation of indole-3-aceticacid in vitro. Planta 157: 131–137
Hammatt N & Grant NJ (1997) Micropropagation of mature Britishwild cherry. Plant Cell Tiss. Org. Cult. 47: 103–110
Hunter CS (1979)In vitro culture of Cinchona ledgerianaL. J.Hortic. Sci. 54: 111–114
Hussey G (1976)In vitro release of axillary shoots from ap-ical dominance in monocotyledonous plantlets. Ann. Bot. 40:1323–1325
Hussey G & Stacey NJ (1981)In vitro propagation of potato(Solanum tuberosumL.). Ann. Bot. 48: 787–796
James DJ (1979) The role of auxins and phloroglucinol in adven-titious root formation inRubusandFragaria grown in vitro. J.Hortic. Sci. 54: 273–277
James DJ (1983) Adventitious root formationin vitro in applerootstocks (Malus pumila) I. Factors affecting the length of theauxin-sensitive phase in M.9. Physiol. Plant. 57: 149–153
James DJ & Thurbon IJ (1981a) Phenolic compounds and otherfactors controlling rhizogenesisin vitro in the apple rootstocksM. 9 and M. 26. Z. Pflanzenphysiol. 105: 11–20
James DJ & Thurbon IJ (1981b) Shoot and root initiationinvitro in the apple rootstock M. 9 and the promotive effects ofphloroglucinol. J. Hortic. Sci. 56: 15–20
Jones OP (1976) Effect of phloridzin and phloroglucinol on appleshoots. Nature 262: 392–393
Jones OP & Hopgood ME (1979) The successful propagationinvitro of two rootstocks ofPrunus: the plum rootstock Pixy (P.insititia) and the cherry rootstock F 12/1 (P. avium). J. Hortic.Sci. 54: 63–66
Jordan M & Piwanski D (1997) Regeneration of babaco (Caricapentagona(Heilborn) Badillo) using leaf explants and shoot-tipculture. Phyton. Buenos Aires 61: 109–115
Lee TT, Starratt AN & Jevnikar JJ (1982) Regulation of enzymeoxidation of indole-3-acetic acid by phenols: structure-activityrelationships. Phytochemistry 21: 517–523
Lozoya-Saldana H & Madrigal-Vargas A (1985) Kinetin, thermo-therapy, and tissue culture to eliminate potato virus (PVX) inpotato. Amer. Potato. J. 62: 339–345
Mellor FC & Stace-Smith R (1987) Virus-free potatoes throughmeristem culture. In: Bajaj YPS (ed) Biotechnology in Agri-culture and Forestry, Vol 3, Potato (pp 30–39). Springer-Verlag,Berlin, Heidelberg, New York, Tokyo
Mosella Ch-L & Fernandez MR (1985)In vitro tissue culture as atool for plant research and propagation. II.In vitro culture of pinkgarlic (Allium sativumL.). Simiente 55: 60–63
Murashige T (1974) Plant propagation through tissue cultures. Ann.Rev. Plant Physiol. 25: 135–166
Murashige T & Skoog F (1962) A revised medium for rapid growthand bioassays with tobacco tissue cultures. Physiol. Plant. 15:473–497
Pontikis C & Melas P (1986) Micropropagation ofFicus caricaL.HortScience 21: 153

149
Pontikis C & Sapoutzaki E (1984) Effect of phloroglucinol on suc-cessful propagationin vitro of Troyer citrange. Plant Propag. 30:3–5
Ramirez-Malagon R, Borodanenko A, Barrera-Guerra JL & Ochoa-Alejo N (1997) Micropropagation forFraser photinia(Photinia× fraseri). Plant Cell Tiss. Org. Cult. 48: 219–222
Sanchez GE, Slack SA & Dodds JH (1991) Response of selectedSolanumspecies to virus eradication therapy. Amer. Potato J. 68:299–315
Sarkar D & Naik PS (1998) Cryopreservation of shoot tips of tet-raploid potato (Solanum tuberosumL.) clones by vitrification.Ann. Bot. 82: 455–461
Sarkar D, Naik PS & Chandra R (1997) Effect of inoculation densityon potato micropropagation. Plant Cell Tiss. Org. Cult. 48: 63–66
Sokal RR & Rohlf FJ (1996) Introduction to Biostatistics. WHFreeman and Company, New York
Steel RGD & Torrie JH (1980) Principles and Procedures of Statist-ics: A Biometrical Approach. McGraw Hill, New York
Stonier T (1969) Studies on auxin protectors. VII. Associationof auxin protectors with crown gall development in sunflowerstems. Physiol. Plant. 44: 1169–1174
Takayama S & Miswa M (1982) Factors affecting differentiationand growthin vitro, and a mass propagation scheme forBegonia× hiemalis. Scientia Hortic. 16: 65–75
Tao GQ, Yin WY, Gong GP & Cui C (1987)In vitro production andrelease of potato varieties in China. In: Bajaj YPS (ed) Biotech-nology in Agriculture and Forestry, Vol 3, Potato (pp 62–79).Springer-Verlag, Berlin, Heidelberg, New York, Tokyo
Zanol GC, Fortes GR de L, Campos AD, da Silva JB , Centellas AQ& da Silva JB (1998)In vitro rooting and peroxidase activity ofapple rootstock cv. ‘Marubakaido’ treated with indolbutyric acidand phloroglucinol. Revista Brasil. Fisiol. Vegetal. 10: 65–68





























