Embed Size (px)
Citation preview

Current Biology 17, 1513–1519, September 4, 2007 ª2007 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2007.07.059
ReportHUA2 Caused Natural Variationin Shoot Morphology of A. thaliana
Qing Wang,1,3 Uday Sajja,1 Sarah Rosloski,1
Tania Humphrey,1 Min Chul Kim,2,4 Kirsten Bomblies,2
Detlef Weigel,2 and Vojislava Grbic1,2,3,*1Department of BiologyUniversity of Western OntarioLondon, Ontario N6A 5B8Canada2Department of Molecular BiologyMax Planck Institute for Developmental BiologyD-72076 TubingenGermany
Summary
Differences in life-history strategy are thought to con-tribute to adaptation to specific environmental condi-
tions. Among life-history traits in plants, floweringtime and shoot morphology are particularly important
for reproductive success. Even though flowering timeand shoot morphology are linked, the evolutionary
changes in the genetic circuitry that simultaneouslyaffects both traits remain obscure. Here, we have iden-
tified changes in a putative pre-mRNA processing fac-tor, HUA2, as being responsible for the distinct shoot
morphology and flowering behavior in Sy-0, a natural
strain of Arabidopsis. HUA2 has previously beenshown to positively regulate two MADS box genes af-
fecting flowering time (FLOWERING LOCUS C [FLC])and floral patterning (AGAMOUS [AG]) [1, 2]. We dem-
onstrate that natural changes in HUA2 activity haveopposite effects on its known functions, thus having
implications for the coordinate control of inductionand maintenance of floral fate. The changes in Sy-
0 lead to enhanced FLC expression, resulting in anenlarged basal rosette and aerial rosettes, whereas
suppression of AG function favors a reversion of floralmeristems from determinate to indeterminate develop-
ment. Natural variation in HUA2 activity thus coordi-nates changes in two important life-history traits, flow-
ering time and shoot morphology.
Results and Discussion
Shoot development in plants progresses continuouslyduring postembryonic development through initiationof primordia that can give rise to either vegetative or re-productive structures. Determination of primordium fatedepends on both endogenous and environmental sig-nals, resulting in highly plastic shoot morphology adap-ted to specific environmental conditions. In Arabidopsis
*Correspondence: [email protected] authors contributed equally to this work.4Present address: Department of Agronomy, College of Agriculture
and Life Science, Gyeong Sang National University, 900 Gajwa-
dong, Jinju 660-701, Korea.
thaliana, a complex regulatory network has evolved tointegrate endogenous and environmental signals thatregulate the expression of a set of genes, called the flo-ral pathway integrators, whose activation commits theshoot apical meristem to cease vegetative developmentand to initiate a reproductive program instead [3]. Vari-ability in flowering time influences the shoot morphologyby affecting the developmental fate of shoot meristems,which can give rise to leaves, branches, or flowers. Nu-merous laboratory-induced mutations affect floweringtime and shoot morphology, but it is not known whetherthey contribute in nature to variant morphology andflowering time.
The morphology of the late-flowering A. thalianaaccession Sy-0 is characterized by an enlarged basalrosette, formation of aerial rosettes in the axils of stemleaves, and reversion of early floral meristems to in-determinate growth, a phenomenon known as floral re-version (Figure S1 in the Supplemental Data availableonline). This phenotype, shared by several other acces-sions, arises as a consequence of extended vegetativedevelopment of both shoot apical and axillary meri-stems and indeterminate development of floral meri-stems [4]. Dominant alleles of the floral repressorsFRIGIDA (FRI) and FLC, and a novel locus, AERIALROSETTE1 (ART1), have been identified as factors thatunderlie this morphological divergence [5].
To understand the molecular basis of natural geneticvariation in shoot morphology characteristic for theSy-0 accession, we isolated the ART1 locus by using po-sitional cloning (Figure 1A). ART1 on its own confers lateflowering [5], which was used to map it to a 15 kb geno-mic region. A recombinant line containing this fragmentfrom Sy-0, but not Ler, fully recapitulated the variantmorphology in a FRI-Sf-2 FLC-Sy-0 background, con-firming that this fragment is sufficient for the Sy-0 pheno-type (Figure 1B and Table S1). The 15 kb fragment con-tains three open reading frames (ORFs). Two of theencoded proteins are involved in chloroplast biogenesisand function, whereas the third ORF encodes HUA2,a known regulator of flowering time and floral patterning[1, 2]. HUA2 was therefore an obvious candidate forART1.
ART1 FRI-Sf-2 plants flower late because of synergis-tic activation of a weak FLC-Ler allele by ART1 and FRI[5]. To test whether HUA2-Sy-0 can reconstitute this ef-fect, we introduced a 10.5 kb genomic fragment, cover-ing the HUA2 gene from Sy-0, into plants containingboth a FRI-Sf-2 and a FLC-Ler allele. The transgene con-ditioned late flowering (Figure 1C). Furthermore, overex-pression of a HUA2-Sy-0 minigene in the FRI-Sf-2 hua2-3 background delayed flowering relative to a HUA2-Lerminigene (Figure 1D), further demonstrating that theHUA2-Sy-0 allele has ART1 activity. We thus renamedART1 as HUA2-Sy-0.
The 10.5 kb HUA2-Sy-0 genomic fragment containseight single nucleotide polymorphisms (SNPs) in thenoncoding region, relative to the Col-0 reference allele;
Current Biology Vol 17 No 171514
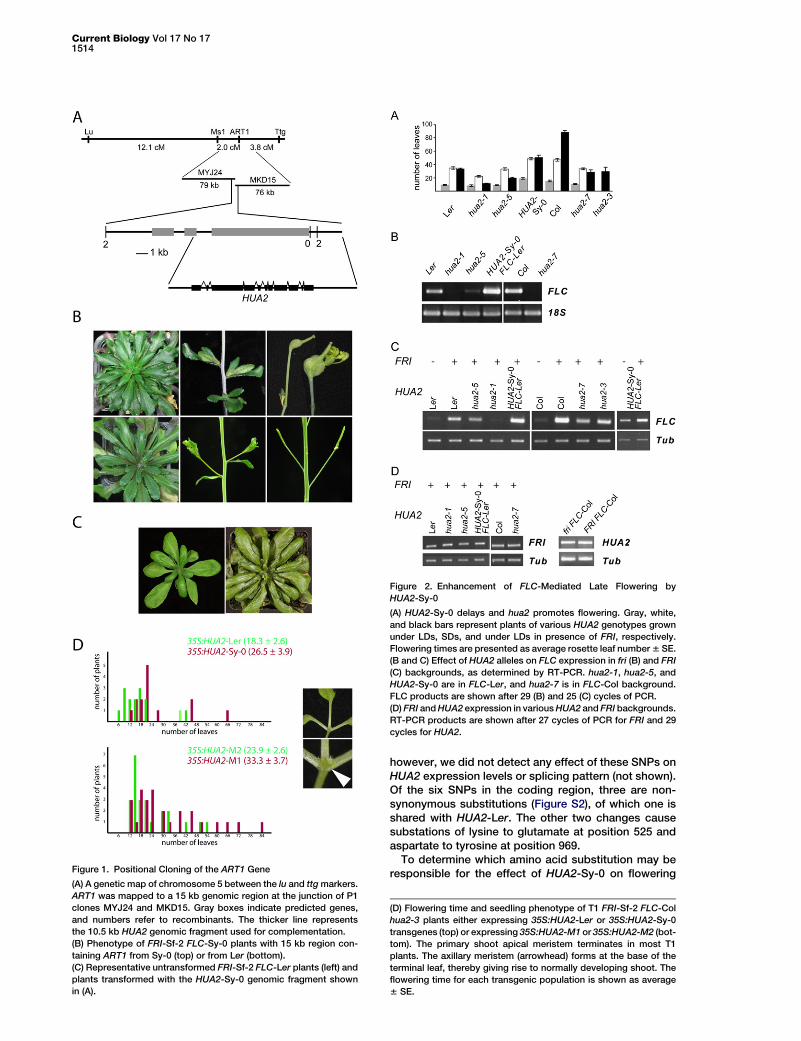
Figure 1. Positional Cloning of the ART1 Gene
(A) A genetic map of chromosome 5 between the lu and ttg markers.
ART1 was mapped to a 15 kb genomic region at the junction of P1
clones MYJ24 and MKD15. Gray boxes indicate predicted genes,
and numbers refer to recombinants. The thicker line represents
the 10.5 kb HUA2 genomic fragment used for complementation.
(B) Phenotype of FRI-Sf-2 FLC-Sy-0 plants with 15 kb region con-
taining ART1 from Sy-0 (top) or from Ler (bottom).
(C) Representative untransformed FRI-Sf-2 FLC-Ler plants (left) and
plants transformed with the HUA2-Sy-0 genomic fragment shown
in (A).
however, we did not detect any effect of these SNPs onHUA2 expression levels or splicing pattern (not shown).Of the six SNPs in the coding region, three are non-synonymous substitutions (Figure S2), of which one isshared with HUA2-Ler. The other two changes causesubstations of lysine to glutamate at position 525 andaspartate to tyrosine at position 969.
To determine which amino acid substitution may beresponsible for the effect of HUA2-Sy-0 on flowering
(D) Flowering time and seedling phenotype of T1 FRI-Sf-2 FLC-Col
hua2-3 plants either expressing 35S:HUA2-Ler or 35S:HUA2-Sy-0
transgenes (top) or expressing 35S:HUA2-M1 or 35S:HUA2-M2 (bot-
tom). The primary shoot apical meristem terminates in most T1
plants. The axillary meristem (arrowhead) forms at the base of the
terminal leaf, thereby giving rise to normally developing shoot. The
flowering time for each transgenic population is shown as average
6 SE.
Figure 2. Enhancement of FLC-Mediated Late Flowering by
HUA2-Sy-0
(A) HUA2-Sy-0 delays and hua2 promotes flowering. Gray, white,
and black bars represent plants of various HUA2 genotypes grown
under LDs, SDs, and under LDs in presence of FRI, respectively.
Flowering times are presented as average rosette leaf number 6 SE.
(B and C) Effect of HUA2 alleles on FLC expression in fri (B) and FRI
(C) backgrounds, as determined by RT-PCR. hua2-1, hua2-5, and
HUA2-Sy-0 are in FLC-Ler, and hua2-7 is in FLC-Col background.
FLC products are shown after 29 (B) and 25 (C) cycles of PCR.
(D) FRI and HUA2 expression in various HUA2 and FRI backgrounds.
RT-PCR products are shown after 27 cycles of PCR for FRI and 29
cycles for HUA2.

Natural Variation in Shoot Morphology1515
Figure 3. FLC Expression Pattern in Vegeta-
tive and Reproductive Apices, Monitored
with a FLC:FLC-GUS Genomic Fusion
(A–N) HUA2-Sy-0 fri (A–D), hua2-3 FRI (E–H),
and HUA2-Sy-0 FRI (I–N).
(M) Whole-mount preparation of primary
stems of HUA2-Sy-0 FRI (main image) and
hua2-3 FRI plants (inset), showing axillary
buds in axils of cauline leaves. GUS staining
is visible in HUA2-Sy-0 FRI but not in hua2-3
FRI axillary buds.
(N) Cross-section of an axillary bud shown in
(M). The primary (1�) and secondary (2�) axil-
ary meristems are visible.
(O and P) The whole-mount GUS stainings of
vegetative and reproductive apices of HUA2-
Col FRI (O) and HUA2-Ler FRI plants. X-gluc
staining (blue under bright field, red under
dark field) marks cells that accumulate FLC-
GUS fusion protein. The following abbrevia-
tions are used: l, leaf; s, stem; and am, axillary
meristem.
time, we expressed two chimeric HUA2 cDNAs, HUA2-M1 and HUA2-M2, in transgenic plants. 35S:HUA2-M1(E525 D969) plants flowered similarly to 35S:HUA2-Sy-0 (E525 Y969) plants, indicating that Glu525 is pre-dominantly responsible for HUA2-Sy-0 activity. In con-trast, 35S:HUA2-M2 (K525 Y969) plants flowered like35S:HUA2-Ler (K525 D969) plants, indicating thatTyr969 is predominantly dispensable for HUA2-Sy-0activity.
To determine the frequency of these substitutions inthe global A. thaliana population, we analyzed additional112 accessions with a world-wide distribution [6]. TheD969Y substitution is common in accessions from theUK and central Europe (Table S2). However, the K525Esubstitution, which is causal of HUA2-Sy-0 activity,was not found outside of Sy-0, indicating that this poly-morphism is rare among natural populations. Sequencecomparison with A. lyrata revealed that both E525 andY969 constitute derived polymorphisms.
To examine the effects of changes in HUA2 activity inSy-0 plants, we assayed a HUA2 allelic series in the Leraccession. HUA2-Sy-0 plants flower late because ofincreased expression levels of the floral repressorFLC, which is further enhanced by the presence ofa functional FRI allele (Figures 2A–2C). In contrast, theloss-of-function alleles hua2-1, hua2-5, and hua2-7(see Figure S3 for information on hua2-7) confer earlyflowering accompanied by lower levels of FLC expres-sion, compared to their corresponding wild-type alleles(Figures 2A–2C). Thus, HUA2-Sy-0 is a gain-of-functionallele regarding its ability to activate FLC expression.The effect of HUA2-Sy-0 appears to be specific to FLCbecause the expression of other FLC-related floral re-pressors, including MAF1, MAF2, and SVP, was similarbetween HUA2-Sy-0 and HUA2-Ler (Figure S4). Our
results for MAF2 and SVP are at variance with previouslypublished work [2], possibly because of differences ingrowth conditions.
The mechanism by which HUA2 and FRI synergisti-cally activate FLC expression is currently not known.One possibility is that HUA2 and FRI regulate eachother’s transcription. However, this is unlikely becausewe found that RNA levels of FRI and HUA2 are indepen-dent of each other (Figure 2D). Alternatively, FRI andHUA2 could interact to affect the spatial pattern ofFLC expression. To investigate this possibility, we intro-duced a reporter that expresses an FLC-GUS fusionsprotein from FLC regulatory sequences into different ge-notypes [7]. In HUA2-Sy-0 fri plants, the fusion proteinaccumulates in the leaf and hypocotyl vasculature dur-ing vegetative development and in mature anthers afterflowering (Figures 3A–3D). In hua2-3 FRI plants, GUS ex-pression is restricted to the vegetative apex, where itis detected in the vasculature and the submeristematicregion. Reporter activity is absent from reproductiveand axillary meristems (Figures 3E–3H and inset inFigure 3M). In contrast, in HUA2-Sy-0 FRI plants, GUSaccumulates throughout the vegetative and reproduc-tive apices (Figures 3I–3N). Thus, HUA2-Sy-0 in combi-nation with FRI can strongly activate FLC expressionin all shoot meristems, including axillary meristems (Fig-ures 3M and 3N). It is particularly interesting that bothHUA2 and FRI are expressed widely throughout theplant, including vegetative and reproductive shoot api-ces, as deduced from microarray profiles and in situhybridization analysis [1, 8], yet confer distinctive andmore restricted patterns of FLC accumulation. Thissuggests that both HUA2 and FRI activities are modifiedby other proteins. HUA2-Sy-0 and FRI together syner-gistically extend the FLC expression pattern to both

Current Biology Vol 17 No 171516
Figure 4. Shoot and Flower Phenotypes of 35S:FLC Plants
(A) An entire shoot of 35S:FLC plants.
(B and C) Secondary inflorescences (arrows) form aerial rosettes in 60% of plants or a branch in 40%.
(D–H) Floral phenotypes. Flowers are subtended by bracts (arrowheads). Early flowers form ectopic shoots (F). Gynoecia are bulged and stig-
matic tissue is reduced (asterisk).
vegetative and reproductive shoot apices. In this con-text, it is important to note that FLC activity in the vascu-lature alone, where FLC is activated independently byboth HUA2-Sy-0 and FRI, has only a limited effect onflowering time. However, extension of FLC activity toshoot meristems, as seen in HUA2-Sy-0 FRI plants,causes extreme delay of flowering, because it allowsFLC to intervene the flowering hierarchy at multiplelevels [9].
In an otherwise isogenic background, HUA2-Col andHUA2-Ler confer the same pattern of FLC expressionas HUA2-Sy-0 (Figures 3O and 3P), suggesting that theSy-0 phenotype arises because of FLC upregulationrather then from a broadening of the FLC expressionpattern. To test this hypothesis, we examined the shootphenotype of plants that express FLC under the controlof the strong and broadly expressed CaMV 35S pro-moter [10]. 35S:FLC plants had extended vegetative de-velopment of both primary and axillary meristems, andsuch development resulted in late flowering frequentlyaccompanied by the formation of aerial rosettes (Fig-ures 4A–4C and Table 1). Although the 35S promotercauses a broader expression pattern than the nativeFLC promoter, the similarity of phenotypes betweenthe Sy-0 and 35S:FLC plants are consistent with thepossibility that high levels of FLC are sufficient for theformation of enlarged basal and aerial rosettes.
In addition to the formation of vegetative structureswithin the inflorescence, Sy-0 plants display floral rever-sion, which is also observed in genotypes with reduced
AG or LFY activity [11, 12]. Because HUA2 was initiallyidentified as a modulator of AG function, on the basisof the ability of hua2-1 to enhance the phenotype ofthe weak ag-4 allele [1], we tested the effect of HUA2-Sy-0 and other hua2 alleles on ag-4 (Figure 5). Interest-ingly, HUA2-Sy-0 had similar effects on the ag-4 pheno-type as several hua2 loss-of-function alleles (Figure 5).In HUA2-Sy-0 ag-4 plants, fertile stamens were replacedby sterile petaloid stamens. Thus, HUA2-Sy-0 is a partialloss-of-function allele regarding its action on AG. Theeffect of HUA2-Sy-0 on AG is enhanced in late-floweringbackgrounds, such that either all stamens (e.g., inHUA2-Sy-0 FLC-Sy-0 ag-4) or some (e.g., in HUA2-Sy-0 FLC-Sy-0 AG) are transformed into petaloid struc-tures (Figures 5M and 5O). Dissected third-whorl organs(Figure 5M) exemplify the spectrum of phenotypes seenin plants heterozygous for HUA2-Sy-0 FLC-Sy-0 alleles.Some of these plants flower early, whereas others flowerlate [5]. The severity of stamen transformation corre-lated with timing of flowering, such that the earliestplants showed the least suppression of stamen identity;the latest plants developed petals in the third whorl,seen also in plants homozygous for HUA2-Sy-0 FLC-Sy-0 alleles. In contrast, plants homozygous for theHUA2-Ler FLC-Ler ag-4 alleles from the same segregat-ing population (see Experimental Procedures) hadflowers indistinguishable from the ag-4 mutant plants.
The observed correlation between the suppression ofstamen identity and flowering time in HUA2-Sy-0 FLC-Sy-0 heterozygous plants raised the possibility that
Table 1. Number of Leaves Formed by Shoot Apical and Axillary Meristems in Ler and 35S:FLC Plants
Genotype n Rosette Leaves Cauline Leaves
Leaves on the First
Branch
Leaves on the Second
Branch
Leaves on the Third
Branch
Ler 10 9.1 6 0.3 3.1 6 0.1 3.1 6 0.2 2.0 6 0.1 1.8 6 0.1
35S:FLC 15 24.2 6 1.2 7.6 6 0.5 8.1 6 0.3 7.0 6 0.3 5.5 6 0.4
Average 6 SE is shown.

Natural Variation in Shoot Morphology1517
Figure 5. Phenotypic Effects of HUA2 Alleles
on ag-4 Flowers
(A) Ler.
(B and C) ag-4 in Ler and Col backgrounds.
Fertile stamens develop in the third whorl in
Ler. In Col, lateral stamens are petaloid and
sterile (shown in the inset in [C]), and medial
stamens are fertile.
(D) ag-1.
(E–N) Flowers and dissected third-whorl or-
gans of ag-4 plants in the presence of various
HUA2 alleles. Some sepals and petals have
been removed from flowers so that the
third-whorl organs (marked with the arrow-
head) are exposed. Second whorl petals (p)
have been included for comparison with pet-
aloid stamens in some genotypes. ‘‘l’’ and
‘‘m’’ stand for lateral and medial third-whorl
organs, respectively. Flowers and dissected
third-whorl organs of HUA2-Sy-0 FLC-Sy-
0 ag-4 plants without vernalization (M) and af-
ter 6 weeks of vernalization (N). Dissected
third-whorl organs in (M) have been collected
from plants heterozygous for HUA2-Sy-
0 FLC-Sy-0. These plants have a wide range
of flowering times. Third-whorl organ on the
left has been dissected from early flowering,
the organ on the middle has been dissected
from the intermediate late flowering, and or-
gan on the right has been dissected from
the late-flowering HUA2-Sy-0 FLC-Sy-0 het-
erozygous plant.
(O) Occasional formation of petal instead of
stamen in the third whorl in HUA2-Sy-
0 FLC-Sy-0 AG flowers.
stamen identity is FLC dependent. Consistent with thishypothesis, flowers of 35S:FLC plants in the Ler back-ground have vegetative characteristics, including sub-tending leaves and enlarged carpels with reduced stig-matic tissue (Figures 4D–4H). If stamen development ismodified by FLC, does HUA2-Sy-0 then directly affectthe ag-4 phenotype, or is this effect due to the HUA2-Sy-0-mediated increased FLC activity in floral meri-stems? To address this question, we vernalized HUA2-Sy-0 FLC-Sy-0 ag-4 plants for 6 weeks. Vernalizationpromotes flowering primarily by repressing the FLC ac-tivity [13–15]. Unvernalized HUA2-Sy-0 FLC-Sy-0 ag-4flowered after producing 42.8 6 3.4 leaves (average 6SE). However, when exposed to 4�C for 6 weeks, theseplants flowered with 11.3 6 0.3 leaves, which is com-parable to vernalized HUA2-Ler FLC-Ler ag-4 plants(11.1 6 0.3 leaves). Vernalized HUA2-Sy-0 FLC-Sy-0ag-4 plants developed flowers in which stamens werereplaced by petaloid stamens (Figure 4N), indicatingthat HUA2-Sy-0 affects the ag-4 phenotype indepen-dently of FLC activity. However, only unvernalizedHUA2-Sy-0 FLC-Sy-0 ag-4 plants (in which FLC is fullyactive) developed petals in the third whorl, indicatingthat full suppression of stamen identity requires bothHUA2-Sy-0 and FLC activities.
In conclusion, ART1 is a naturally occurring allele ofHUA2, which together with active FRI and FLC allelescauses delayed flowering of primary and axillary meri-stems, and this delayed flowering results in the distinc-tive Sy-0 morphology. Allelic variation in FRI and FLC isone of the major determinants of life history in A. thalianabecause active alleles at FRI and FLC loci promoteshoot vegetative development and confer late-floweringwinter annual growth habit, whereas loss-of-function al-leles at these loci lead to early-flowering summer annualhabit [16, 17]. Allelic variation at FRI and FLC influenceshoot morphology by affecting the number of leavesinitiated by the primary shoot apical meristem, but thisvariation usually has little effect on meristems that giverise to branches and flowers.
HUA2-Sy-0 also causes late flowering. Apart from theHUA2-Sy-0 allele, we identified several other polymor-phisms in HUA2 common to a subset of accessions. Al-though the functional significance of these polymor-phisms is not known, additional natural alleles ofHUA2 may be responsible for flowering-time variationbecause several mapping intervals for late-floweringQTL include the HUA2 gene [18, 19]. In contrast to othercharacterized HUA2 alleles, the natural variant HUA2-Sy-0 is unique in that it uncouples the effects on FLC

Current Biology Vol 17 No 171518
and AG observed in laboratory-induced hua2 alleles.HUA2 normally activates FLC expression and enhancesAG function [1, 2]. Consequently, in hua2 mutants, bothof these responses are attenuated. HUA2-Sy-0 en-hances FLC expression in a broad domain that includesall shoot meristems, and this expression leads to late-flowering and aerial-rosette-forming shoot phenotype.On the other hand, it represses AG activity, which corre-lates with floral reversion.
The K525E substitution in HUA2-Sy-0 allele appearsunique to Sy-0. A similar pattern is seen for an FLM de-letion allele and EDI allele of CRY2, other large-effectflowering-time alleles that are found only in accessionsfrom the Niederzenz and the Cape Verde Islands, re-spectively [20, 21]. However, whereas the HUA2-Sy-0 al-lele is rare, the Sy-0 morphology is not. Other Arabidop-sis accessions display similar shoot morphology underlaboratory growth conditions [4]. Furthermore, plantswith aerial rosettes can sometimes be found in naturein individuals with prolonged growth (Figure S5). Thismorphology profoundly changes life history: The vege-tative phase of development is extended beyond the on-set of flowering and is a phenomenon that lengthens thelifespan of the plant and extends the period duringwhich it is capable of seed production. Although thegenetic basis of this morphology across accessions re-mains to be elucidated, its occurrence in natural popula-tions indicates that it is advantageous under some envi-ronments. Theory predicts that pleiotropically actinggenes are normally not the targets of natural variation[22]. The finding that HUA2-Sy-0 affects multiple com-ponents of the plant life-history strategy such as flower-ing time and plant morphology nevertheless highlightsthe potential of multifunctional genes to contribute tophenotypic novelty within a species.
Experimental Procedures
Plant Materials and Growth Conditions
The ART1, ART1 FLC-Sy-0, lu ttg, hua2-5, FRI-Sf-2 in Ler, FRI-Sf-2 in
Col, FRI-Sf-2 hua2-3 strains have been described [2, 5, 23]. The
hua2-1, ag-4, hua2-1 ag-4, and Ler strains were kindly provided by
X. Chen [1], and FRI FLC:FLC-GUS flc3 was provided by R. Amasino
[6]. hua2-7 was isolated from the Syngenta T-DNA collection (refer-
ence number: 314_A08.b.1a.Lb3Fa). The accessions examined for
the Sy-0-specific nucleotide polymorphisms include a set of 96 lines
with extensive characterization of genome-wide sequence variation
[6] and 16 additional accessions obtained from the Arabidopsis
Biological Resource Center. They are listed in Table S2. Plants
were grown in PRO-MIX soil (Plant Products) under 100–150 mmol/
m2/s cool-white fluorescent light at 23�C. Long days (LDs) consisted
of 16 hr light/8 hr darkness, and short day (SD) conditions consisted
of 8 hr light/16 hr darkness. Seven-day-old seedlings for RNA ex-
traction were grown at 21�C on agar with half strength of MS medium
(Sigma) and 1% sucrose under LDs, after stratification at 4�C for
2–3 days. F2 segregating population of a cross between HUA2-
Sy-0 FLC-Sy-0 and ag-4 lines was either grown continuously at
23�C (unvernalized) or first grown at 4�C for 6 weeks before transfer
to 23�C (vernalized plants). Plants were allowed to flower, and ag-4
plants (selected on the basis of the indeterminate floral phenotype)
were genotyped for the HUA2-Sy-0 FLC-Sy-0 alleles with the NIT4
marker located between the two loci. Flowering times are presented
as average rosette leaf number 6 standard error.
Identification of ART1 by Mapping and Complementation
A line carrying recessive mutations in lu ms1 ttg was initially crossed
with a line derived from the Sy-0 accession that carried FLC-
Sy-0 ART1 alleles [5]. Approximately 18,000 F2 plants were screened
for recombination between the Lu and Ttg morphological markers,
thereby resulting in the identification of 250 Lu Ms1 ttg recombinant
lines. The flowering phenotype and the genotype at the ART1 locus
of recombinant lines were scored in F3 progeny. Genomic DNA from
Sy-0 plants was partially digested with Sau3A, and w20 kb frag-
ments were size selected on sucrose gradient and cloned into the
BamHI site of lambdaZAP (Stratagene). One 15 kb-clone, 4B1, start-
ing at nucleotide position 76,808 of BAC MYJ24 and ending at
13,798 of BAC MKD15 was identified. Because it lacked HUA2 50 se-
quences, a full-length genomic fragment was created by joining
2.5 kb upstream HUA2 sequence (isolated by PCR, with primers
HUA2F and HUA2BamH1R; Table S3) and an 8 kb BamHI-ClaI frag-
ment of 4B1. The final 10.5 kb genomic fragment was cloned into the
Bsp120I site of the pPZP111 binary vector [24] and used for com-
plementation analysis. HUA2-Sy-0 and HUA2-Ler cDNAs were
amplified with primers HUA2 cDNA-F and HUA2 cDNA-R from
HUA2-Sy-0 FLC-Sy-0 and Ler strains, respectively. We obtained
HUA2-M1 by replacing T with G (corresponding to Y969D at the
amino acid level) and obtained HUA2-M2 by replacing G with A
(E525K) in HUA2-Sy-0 cDNA. HUA2 cDNAs were cloned into
pGEMT-easy TA cloning vector (Promega). AvrII/BstEII cDNA frag-
ments were inserted into SpeI and BstEII sites of pCAMBIA 1303
binary vector and thus replaced the GUS sequence. The resulting
vectors were transformed into FRI-Sf-2 hua2-3 plants.
Expression Analysis
RNA gel-blot analysis was performed as described [5]. Primers used
for RT-PCR analysis of MAF1, MAF2, or SVP have been described as
well [2]. The remainders of primers used are provided in Table S3.
GUS staining was carried out as described [25]. Whole mounts
were examined under a MZ FLIII (Leica) microscope, and pictures
were taken with an AxioCam HRc digital camera (Zeiss). Thin sec-
tions of tissues stained for GUS activity were prepared from paraf-
fin-embedded tissue, and sections (9–12 mm thick) were prepared
on a EG1160 microtome (Leica).
Sequence Analysis
For A. thaliana accessions, full-length cDNAs were isolated by RT-
PCR, subcloned, and sequenced for strains marked by an asterisk
in Table S2. In the remainder of the accessions, E525K and Y969D
SNPs were detected by direct sequencing of the PCR product
(see Table S3 for primers used). The A. lyrata sequence was assem-
bled from reads in the NCBI trace archive retrieved with discontinu-
ous Mega-BLAST with the A. thaliana HUA2-Col-0 sequence.
Supplemental Data
Five figures and three tables are available at http://www.
current-biology.com/cgi/content/full/17/17/1513/DC1/.
Acknowledgments
We thank Syngenta for the SAIL T-DNA line. We thank Ian Craig for
artwork and Dr. S. Balasubramanian, Dr. Y. Kobayashi, and Dr. R.
Clark for discussions and comments on the manuscript. This work
was supported by the Natural Sciences and Engineering Research
Council of Canada (NSERC) and UWO Academic Development
Funds (ADF) to V.G., by the Korea Science and Engineering Founda-
tion and European Molecular Biology Organisation (EMBO) to
M.C.K., and by the Max Planck Society. D.W. is a director of the
Max Planck Institute.
Received: April 2, 2007
Revised: July 26, 2007
Accepted: July 27, 2007
Published online: August 30, 2007
References
1. Chen, X., and Meyerowitz, E.M. (1999). HUA1 and HUA2 are two
members of the floral homeotic AGAMOUS pathway. Mol. Cell 3,
349–360.
2. Doyle, M.R., Bizzell, C.M., Keller, M.R., Michaels, S.D., Song, J.,
Noh, Y.S., and Amasino, R.M. (2005). HUA2 is required for the

Natural Variation in Shoot Morphology1519
expression of floral repressors in Arabidopsis thaliana. Plant J.
41, 376–385.
3. Baurle, I., and Dean, C. (2006). The timing of developmental tran-
sitions in plants. Cell 125, 655–664.
4. Grbic, V., and Bleecker, A.B. (1996). An altered body plan is con-
ferred on Arabidopsis plants carrying dominant alleles of two
genes. Development 122, 2395–2403.
5. Poduska, B., Humphrey, T., Redweik, A., and Grbic, V. (2003).
The synergistic activation of FLOWERING LOCUS C by FRIGIDA
and a new flowering gene AERIAL ROSETTE 1 underlies a novel
morphology in Arabidopsis. Genetics 163, 1457–1465.
6. Nordborg, M., Hu, T.T., Ishino, Y., Jhaveri, J., Toomajian, C.,
Zheng, H., Bakker, E., Calabrese, P., Gladstone, J., Goyal, R.,
et al. (2005). The pattern of polymorphism in Arabidopsis thali-
ana. PLoS Biol. 3, e196.
7. Michaels, S.D., Himelblau, E., Kim, S.Y., Schomburg, F.M., and
Amasino, R.M. (2005). Integration of flowering signals in win-
ter-annual Arabidopsis. Plant Physiol. 137, 149–156.
8. Schmid, M., Davison, T.S., Henz, S.R., Pape, U.J., Demar, M.,
Vingron, M., Scholkopf, B., Weigel, D., and Lohmann, J.U.
(2005). A gene expression map of Arabidopsis thaliana develop-
ment. Nat. Genet. 37, 501–506.
9. Searle, I., He, Y., Turck, F., Vincent, C., Fornara, F., Krober, S.,
Amasino, R.A., and Coupland, G. (2006). The transcription factor
FLC confers a flowering response to vernalization by repressing
meristem competence and systemic signaling in Arabidopsis.
Genes Dev. 20, 898–912.
10. Benfey, P.N., Ren, L., and Chua, N.H. (1990). Combinatorial and
synergistic properties of CaMV 35S enhancer subdomains.
EMBO J. 9, 1685–1696.
11. Okamuro, J.K., den Boer, B.G., Lotys-Prass, C., Szeto, W., and
Jofuku, K.D. (2006). Flowers into shoots: Photo and hormonal
control of a meristem identity switch in Arabidopsis. Proc.
Natl. Acad. Sci. USA 93, 13831–13836.
12. Parcy, F., Bomblies, K., and Weigel, D. (2002). Interaction of
LEAFY, AGAMOUS and TERMINAL FLOWER1 in maintaining
floral meristem identity in Arabidopsis. Development 129,
2519–2527.
13. Michaels, S.D., and Amasino, R.M. (1999). FLOWERING LOCUS
C encodes a novel MADS domain protein that acts as a repressor
of flowering. Plant Cell 11, 949–956.
14. Sheldon, C.C., Burn, J.E., Perez, P.P., Metzger, J., Edwards,
J.A., Peacock, W.J., and Dennis, E.S. (1999). The FLF MADS
box gene: A repressor of flowering in Arabidopsis regulated by
vernalization and methylation. Plant Cell 11, 445–458.
15. Michaels, S.D., and Amasino, R.M. (2001). Loss of FLOWERING
LOCUS C activity eliminates the late-flowering phenotype of
FRIGIDA and autonomous pathway mutations but not respon-
siveness to vernalization. Plant Cell 13, 935–941.
16. Shindo, C., Aranzana, M.J., Lister, C., Baxter, C., Nicholls, C.,
Nordborg, M., and Dean, C. (2005). Role of FRIGIDA and FLOW-
ERING LOCUS C in determining variation in flowering time of
Arabidopsis. Plant Physiol. 138, 1163–1173.
17. Lempe, J., Balasubramanian, S., Sureshkumar, S., Singh, A.,
Schmid, M., and Weigel, D. (2005). Diversity of flowering re-
sponses in wild Arabidopsis thaliana strains. PLoS Genet. 1,
109–118.
18. Alonso-Blanco, C., El-Assal, S.E.-D., Coupland, G., and Koorn-
neef, M. (1998). Analysis of natural allelic variation at flowering
time loci in the Landsberg erecta and Cape Verde Islands eco-
types of Arabidopsis thaliana. Genetics 149, 749–764.
19. Shindo, C., Lister, C., Crevillen, P., Nordborg, M., and Dean, C.
(2006). Variation in the epigenetic silencing of FLC contributes
to natural variation in Arabidopsis vernalization response.
Genes Dev. 20, 3079–3083.
20. Werner, J.D., Borevitz, J.O., Warthmann, N., Trainer, G.T., Ecker,
J.R., Chory, J., and Weigel, D. (2005). Quantitative trait locus
mapping and DNA array hybridization identify an FLM deletion
as a cause for natural flowering-time variation. Proc. Natl.
Acad. Sci. USA 102, 2460–2465.
21. El-Assal, S.E.-D., Alonso-Blanco, C., Peeters, A.J., Raz, V., and
Koornneef, M. (2001). A QTL for flowering time in Arabidopsis re-
veals a novel allele of CRY2. Nat. Genet. 29, 435–440, Erratum:
(2002). Nat. Genet. 30, 123.
22. Barton, N.H. (1990). Pleiotropic models of quantitative variation.
Genetics 124, 773–782.
23. Lee, I., Aukerman, M.J., Gore, S.L., Lohman, K.N., Michaels,
S.D., Weaver, L.M., John, M.C., Feldmann, K.A., and Amasino,
R.M. (1994). Isolation of LUMINIDEPENDENS: A gene involved
in the control of flowering time in Arabidopsis. Plant Cell 6,
75–83.
24. Hajdukiewicz, P., Svab, Z., and Maliga, P. (1994). The small, ver-
satile pPZP family of Agrobacterium binary vectors for plant
transformation. Plant Mol. Biol. 25, 989–994.
25. Weigel, D., and Glazebrook, J. (2002). Arabidopsis: A Laboratory
Manual (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory
Press).

in Shoot Morphology of A
Supplemental Data S1
HUA2 Caused Natural Variation. thaliana
Qing Wang, Uday Sajja, Sarah Rosloski,Tania Humphrey, Min Chul Kim, Kirsten Bomblies,
Detlef Weigel, and Vojislava Grbic
Figure S1. Morphology of an 80-Day-Old Sy-0 Plant
Sy-0 plants develop an enlarged basal rosette and aerial rosettes
along the inflorescence stem (arrow). They also display floral (inset
and red asterisk) and inflorescence (arrowheads) reversion.
Figure S2. HUA2 Alleles
(A) Diagram of HUA2 genomic structure. Exons are shown as black
boxes, and introns are shown as lines. Vertical lines show the posi-
tion of various mutations within the gene.
(B) Schematic presentation of predicted gene products encoded by
HUA2 alleles used in this study. hua2-7 allele is predicted to lack the
HUA2 protein. HUA2 contains several motifs including a PWWP
domain (white circle), a putative nuclear localization sequences
(NLSs, arrowheads), the RPR domain (gray circle), a putative activa-
tion domain (Ac, white box) and proline-rich domains (P [light-gray
oval] and P/S/V [gray box]). Positions of amino acid substitutions
are shown with a black diamond above each schematic.
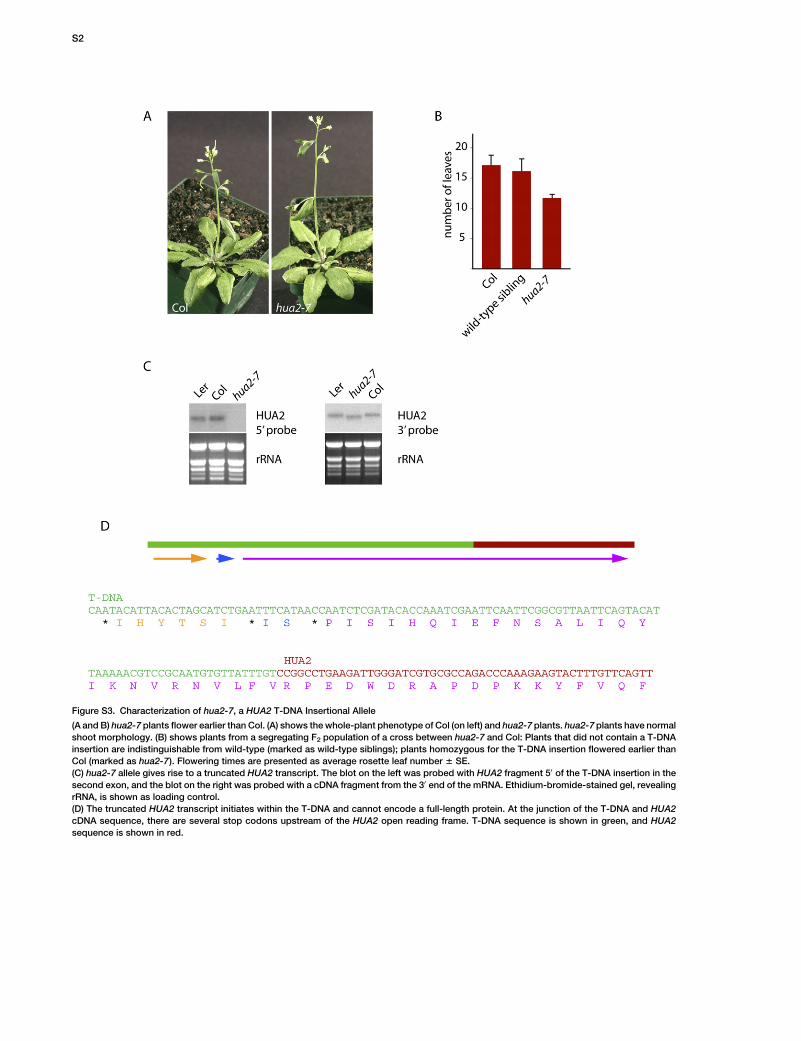
Figure S3. Characterization of hua2-7, a HUA2 T-DNA Insertional Allele
(A and B) hua2-7 plants flower earlier than Col. (A) shows the whole-plant phenotype of Col (on left) and hua2-7 plants. hua2-7 plants have normal
shoot morphology. (B) shows plants from a segregating F2 population of a cross between hua2-7 and Col: Plants that did not contain a T-DNA
insertion are indistinguishable from wild-type (marked as wild-type siblings); plants homozygous for the T-DNA insertion flowered earlier than
Col (marked as hua2-7). Flowering times are presented as average rosette leaf number 6 SE.
(C) hua2-7 allele gives rise to a truncated HUA2 transcript. The blot on the left was probed with HUA2 fragment 50 of the T-DNA insertion in the
second exon, and the blot on the right was probed with a cDNA fragment from the 30 end of the mRNA. Ethidium-bromide-stained gel, revealing
rRNA, is shown as loading control.
(D) The truncated HUA2 transcript initiates within the T-DNA and cannot encode a full-length protein. At the junction of the T-DNA and HUA2
cDNA sequence, there are several stop codons upstream of the HUA2 open reading frame. T-DNA sequence is shown in green, and HUA2
sequence is shown in red.
S2

Figure S4. Expression of MAF1, SVP, and MAF2 in Various HUA2
Backgrounds, as Determined by RT-PCR
Products are shown after 28, 29, and 27 cycles of PCR, respectively.
S3
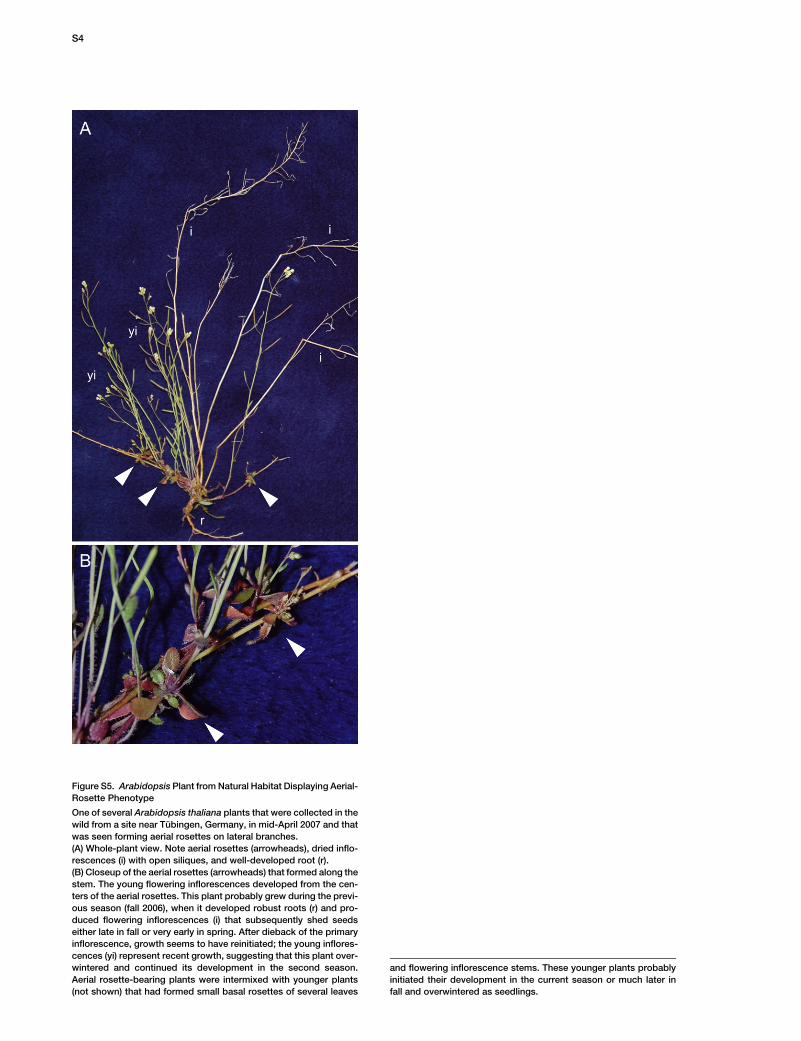
Figure S5. Arabidopsis Plant from Natural Habitat Displaying Aerial-
Rosette Phenotype
One of several Arabidopsis thaliana plants that were collected in the
wild from a site near Tubingen, Germany, in mid-April 2007 and that
was seen forming aerial rosettes on lateral branches.
(A) Whole-plant view. Note aerial rosettes (arrowheads), dried inflo-
rescences (i) with open siliques, and well-developed root (r).
(B) Closeup of the aerial rosettes (arrowheads) that formed along the
stem. The young flowering inflorescences developed from the cen-
ters of the aerial rosettes. This plant probably grew during the previ-
ous season (fall 2006), when it developed robust roots (r) and pro-
duced flowering inflorescences (i) that subsequently shed seeds
either late in fall or very early in spring. After dieback of the primary
inflorescence, growth seems to have reinitiated; the young inflores-
cences (yi) represent recent growth, suggesting that this plant over-
wintered and continued its development in the second season.
Aerial rosette-bearing plants were intermixed with younger plants
(not shown) that had formed small basal rosettes of several leaves
and flowering inflorescence stems. These younger plants probably
initiated their development in the current season or much later in
fall and overwintered as seedlings.
S4
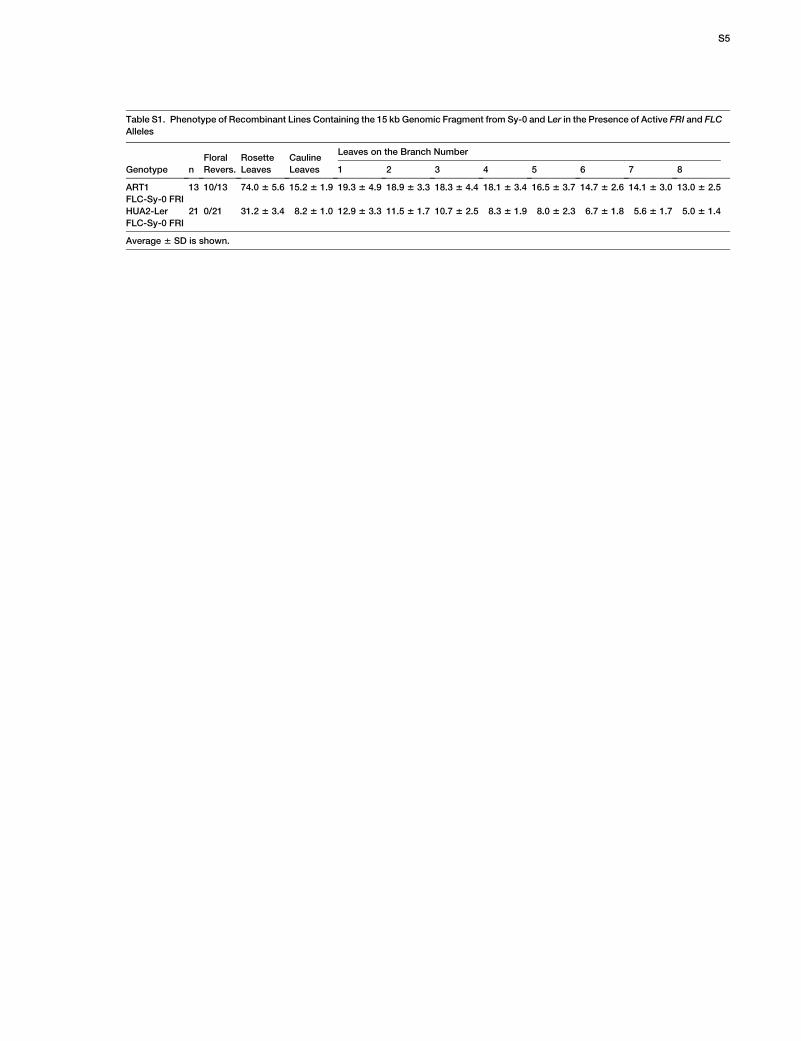
Table S1. Phenotype of Recombinant Lines Containing the 15 kb Genomic Fragment from Sy-0 and Ler in the Presence of Active FRI and FLC
Alleles
Genotype n
Floral
Revers.
Rosette
Leaves
Cauline
Leaves
Leaves on the Branch Number
1 2 3 4 5 6 7 8
ART1
FLC-Sy-0 FRI
13 10/13 74.0 6 5.6 15.2 6 1.9 19.3 6 4.9 18.9 6 3.3 18.3 6 4.4 18.1 6 3.4 16.5 6 3.7 14.7 6 2.6 14.1 6 3.0 13.0 6 2.5
HUA2-Ler
FLC-Sy-0 FRI
21 0/21 31.2 6 3.4 8.2 6 1.0 12.9 6 3.3 11.5 6 1.7 10.7 6 2.5 8.3 6 1.9 8.0 6 2.3 6.7 6 1.8 5.6 6 1.7 5.0 6 1.4
Average 6 SD is shown.
S5
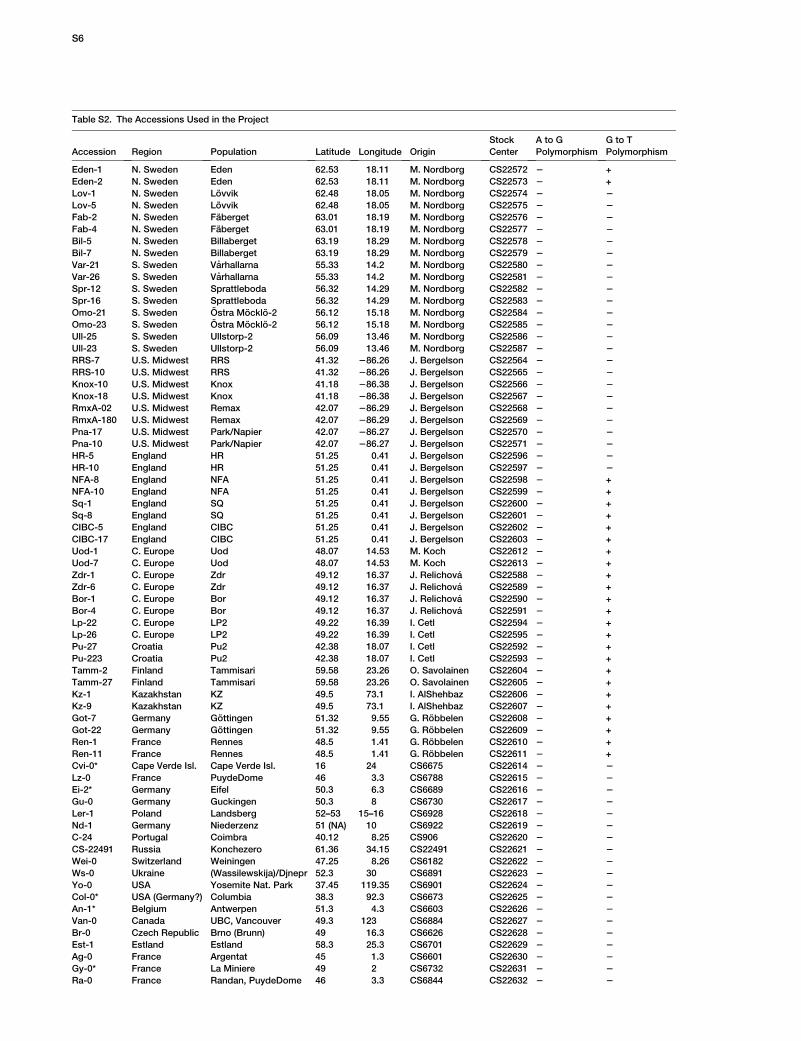
Table S2. The Accessions Used in the Project
Accession Region Population Latitude Longitude Origin
Stock
Center
A to G
Polymorphism
G to T
Polymorphism
Eden-1 N. Sweden Eden 62.53 18.11 M. Nordborg CS22572 2 +
Eden-2 N. Sweden Eden 62.53 18.11 M. Nordborg CS22573 2 +
Lov-1 N. Sweden Lovvik 62.48 18.05 M. Nordborg CS22574 2 2
Lov-5 N. Sweden Lovvik 62.48 18.05 M. Nordborg CS22575 2 2
Fab-2 N. Sweden Faberget 63.01 18.19 M. Nordborg CS22576 2 2
Fab-4 N. Sweden Faberget 63.01 18.19 M. Nordborg CS22577 2 2
Bil-5 N. Sweden Billaberget 63.19 18.29 M. Nordborg CS22578 2 2
Bil-7 N. Sweden Billaberget 63.19 18.29 M. Nordborg CS22579 2 2
Var-21 S. Sweden Varhallarna 55.33 14.2 M. Nordborg CS22580 2 2
Var-26 S. Sweden Varhallarna 55.33 14.2 M. Nordborg CS22581 2 2
Spr-12 S. Sweden Sprattleboda 56.32 14.29 M. Nordborg CS22582 2 2
Spr-16 S. Sweden Sprattleboda 56.32 14.29 M. Nordborg CS22583 2 2
Omo-21 S. Sweden Ostra Mocklo-2 56.12 15.18 M. Nordborg CS22584 2 2
Omo-23 S. Sweden Ostra Mocklo-2 56.12 15.18 M. Nordborg CS22585 2 2
Ull-25 S. Sweden Ullstorp-2 56.09 13.46 M. Nordborg CS22586 2 2
Ull-23 S. Sweden Ullstorp-2 56.09 13.46 M. Nordborg CS22587 2 2
RRS-7 U.S. Midwest RRS 41.32 286.26 J. Bergelson CS22564 2 2
RRS-10 U.S. Midwest RRS 41.32 286.26 J. Bergelson CS22565 2 2
Knox-10 U.S. Midwest Knox 41.18 286.38 J. Bergelson CS22566 2 2
Knox-18 U.S. Midwest Knox 41.18 286.38 J. Bergelson CS22567 2 2
RmxA-02 U.S. Midwest Remax 42.07 286.29 J. Bergelson CS22568 2 2
RmxA-180 U.S. Midwest Remax 42.07 286.29 J. Bergelson CS22569 2 2
Pna-17 U.S. Midwest Park/Napier 42.07 286.27 J. Bergelson CS22570 2 2
Pna-10 U.S. Midwest Park/Napier 42.07 286.27 J. Bergelson CS22571 2 2
HR-5 England HR 51.25 0.41 J. Bergelson CS22596 2 2
HR-10 England HR 51.25 0.41 J. Bergelson CS22597 2 2
NFA-8 England NFA 51.25 0.41 J. Bergelson CS22598 2 +
NFA-10 England NFA 51.25 0.41 J. Bergelson CS22599 2 +
Sq-1 England SQ 51.25 0.41 J. Bergelson CS22600 2 +
Sq-8 England SQ 51.25 0.41 J. Bergelson CS22601 2 +
CIBC-5 England CIBC 51.25 0.41 J. Bergelson CS22602 2 +
CIBC-17 England CIBC 51.25 0.41 J. Bergelson CS22603 2 +
Uod-1 C. Europe Uod 48.07 14.53 M. Koch CS22612 2 +
Uod-7 C. Europe Uod 48.07 14.53 M. Koch CS22613 2 +
Zdr-1 C. Europe Zdr 49.12 16.37 J. Relichova CS22588 2 +
Zdr-6 C. Europe Zdr 49.12 16.37 J. Relichova CS22589 2 +
Bor-1 C. Europe Bor 49.12 16.37 J. Relichova CS22590 2 +
Bor-4 C. Europe Bor 49.12 16.37 J. Relichova CS22591 2 +
Lp-22 C. Europe LP2 49.22 16.39 I. Cetl CS22594 2 +
Lp-26 C. Europe LP2 49.22 16.39 I. Cetl CS22595 2 +
Pu-27 Croatia Pu2 42.38 18.07 I. Cetl CS22592 2 +
Pu-223 Croatia Pu2 42.38 18.07 I. Cetl CS22593 2 +
Tamm-2 Finland Tammisari 59.58 23.26 O. Savolainen CS22604 2 +
Tamm-27 Finland Tammisari 59.58 23.26 O. Savolainen CS22605 2 +
Kz-1 Kazakhstan KZ 49.5 73.1 I. AlShehbaz CS22606 2 +
Kz-9 Kazakhstan KZ 49.5 73.1 I. AlShehbaz CS22607 2 +
Got-7 Germany Gottingen 51.32 9.55 G. Robbelen CS22608 2 +
Got-22 Germany Gottingen 51.32 9.55 G. Robbelen CS22609 2 +
Ren-1 France Rennes 48.5 1.41 G. Robbelen CS22610 2 +
Ren-11 France Rennes 48.5 1.41 G. Robbelen CS22611 2 +
Cvi-0* Cape Verde Isl. Cape Verde Isl. 16 24 CS6675 CS22614 2 2
Lz-0 France PuydeDome 46 3.3 CS6788 CS22615 2 2
Ei-2* Germany Eifel 50.3 6.3 CS6689 CS22616 2 2
Gu-0 Germany Guckingen 50.3 8 CS6730 CS22617 2 2
Ler-1 Poland Landsberg 52–53 15–16 CS6928 CS22618 2 2
Nd-1 Germany Niederzenz 51 (NA) 10 CS6922 CS22619 2 2
C-24 Portugal Coimbra 40.12 8.25 CS906 CS22620 2 2
CS-22491 Russia Konchezero 61.36 34.15 CS22491 CS22621 2 2
Wei-0 Switzerland Weiningen 47.25 8.26 CS6182 CS22622 2 2
Ws-0 Ukraine (Wassilewskija)/Djnepr 52.3 30 CS6891 CS22623 2 2
Yo-0 USA Yosemite Nat. Park 37.45 119.35 CS6901 CS22624 2 2
Col-0* USA (Germany?) Columbia 38.3 92.3 CS6673 CS22625 2 2
An-1* Belgium Antwerpen 51.3 4.3 CS6603 CS22626 2 2
Van-0 Canada UBC, Vancouver 49.3 123 CS6884 CS22627 2 2
Br-0 Czech Republic Brno (Brunn) 49 16.3 CS6626 CS22628 2 2
Est-1 Estland Estland 58.3 25.3 CS6701 CS22629 2 2
Ag-0 France Argentat 45 1.3 CS6601 CS22630 2 2
Gy-0* France La Miniere 49 2 CS6732 CS22631 2 2
Ra-0 France Randan, PuydeDome 46 3.3 CS6844 CS22632 2 2
S6
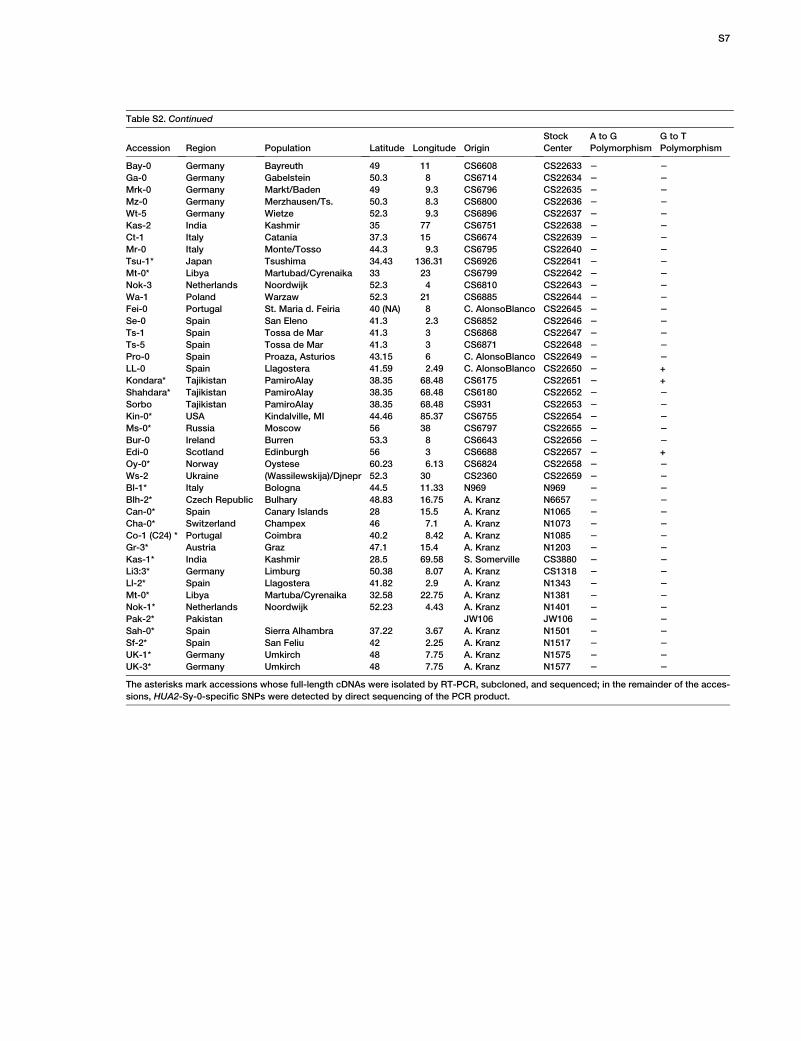
Table S2. Continued
Accession Region Population Latitude Longitude Origin
Stock
Center
A to G
Polymorphism
G to T
Polymorphism
Bay-0 Germany Bayreuth 49 11 CS6608 CS22633 2 2
Ga-0 Germany Gabelstein 50.3 8 CS6714 CS22634 2 2
Mrk-0 Germany Markt/Baden 49 9.3 CS6796 CS22635 2 2
Mz-0 Germany Merzhausen/Ts. 50.3 8.3 CS6800 CS22636 2 2
Wt-5 Germany Wietze 52.3 9.3 CS6896 CS22637 2 2
Kas-2 India Kashmir 35 77 CS6751 CS22638 2 2
Ct-1 Italy Catania 37.3 15 CS6674 CS22639 2 2
Mr-0 Italy Monte/Tosso 44.3 9.3 CS6795 CS22640 2 2
Tsu-1* Japan Tsushima 34.43 136.31 CS6926 CS22641 2 2
Mt-0* Libya Martubad/Cyrenaika 33 23 CS6799 CS22642 2 2
Nok-3 Netherlands Noordwijk 52.3 4 CS6810 CS22643 2 2
Wa-1 Poland Warzaw 52.3 21 CS6885 CS22644 2 2
Fei-0 Portugal St. Maria d. Feiria 40 (NA) 8 C. AlonsoBlanco CS22645 2 2
Se-0 Spain San Eleno 41.3 2.3 CS6852 CS22646 2 2
Ts-1 Spain Tossa de Mar 41.3 3 CS6868 CS22647 2 2
Ts-5 Spain Tossa de Mar 41.3 3 CS6871 CS22648 2 2
Pro-0 Spain Proaza, Asturios 43.15 6 C. AlonsoBlanco CS22649 2 2
LL-0 Spain Llagostera 41.59 2.49 C. AlonsoBlanco CS22650 2 +
Kondara* Tajikistan PamiroAlay 38.35 68.48 CS6175 CS22651 2 +
Shahdara* Tajikistan PamiroAlay 38.35 68.48 CS6180 CS22652 2 2
Sorbo Tajikistan PamiroAlay 38.35 68.48 CS931 CS22653 2 2
Kin-0* USA Kindalville, MI 44.46 85.37 CS6755 CS22654 2 2
Ms-0* Russia Moscow 56 38 CS6797 CS22655 2 2
Bur-0 Ireland Burren 53.3 8 CS6643 CS22656 2 2
Edi-0 Scotland Edinburgh 56 3 CS6688 CS22657 2 +
Oy-0* Norway Oystese 60.23 6.13 CS6824 CS22658 2 2
Ws-2 Ukraine (Wassilewskija)/Djnepr 52.3 30 CS2360 CS22659 2 2
Bl-1* Italy Bologna 44.5 11.33 N969 N969 2 2
Blh-2* Czech Republic Bulhary 48.83 16.75 A. Kranz N6657 2 2
Can-0* Spain Canary Islands 28 15.5 A. Kranz N1065 2 2
Cha-0* Switzerland Champex 46 7.1 A. Kranz N1073 2 2
Co-1 (C24) * Portugal Coimbra 40.2 8.42 A. Kranz N1085 2 2
Gr-3* Austria Graz 47.1 15.4 A. Kranz N1203 2 2
Kas-1* India Kashmir 28.5 69.58 S. Somerville CS3880 2 2
Li3:3* Germany Limburg 50.38 8.07 A. Kranz CS1318 2 2
Ll-2* Spain Llagostera 41.82 2.9 A. Kranz N1343 2 2
Mt-0* Libya Martuba/Cyrenaika 32.58 22.75 A. Kranz N1381 2 2
Nok-1* Netherlands Noordwijk 52.23 4.43 A. Kranz N1401 2 2
Pak-2* Pakistan JW106 JW106 2 2
Sah-0* Spain Sierra Alhambra 37.22 3.67 A. Kranz N1501 2 2
Sf-2* Spain San Feliu 42 2.25 A. Kranz N1517 2 2
UK-1* Germany Umkirch 48 7.75 A. Kranz N1575 2 2
UK-3* Germany Umkirch 48 7.75 A. Kranz N1577 2 2
The asterisks mark accessions whose full-length cDNAs were isolated by RT-PCR, subcloned, and sequenced; in the remainder of the acces-
sions, HUA2-Sy-0-specific SNPs were detected by direct sequencing of the PCR product.
S7

Table S3. Oligonucleotide Primers Used
No. Lab Designation Sequence
1 HUA2F 50-GTTCAAGTTAGGAAGTAACTGG-30
2 HUA2BamHIR 50-GCAAGGATCCACCCTAAAATT-30
3 FLCE12f 50-TTCTCCTCCGGCGATAACCTGGT-30
4 FLCE7r2 50-TCAACCGCCGATTTAAGGTGGCT-30
5 HUA2G 50-CAG TGC TTT AGA CGA CCT GG-30
6 HUA2H 50-GTC CTC ATT TCT GTA TCG CC-30
7 LB3 50-TAG CAT TCT GAA TTT CAT AAC CAA TCT CG-30
8 MKD15p7-3 50-ACCGTCGTAAATGTCTGAAG-30
9 MKD15p7-2 50-GCTCCTTTGTCTTATTCTCA-30
10 G-0632 50-CTT GTG GGA TCA AAT GTC AAA AAT GTG-30
11 G-0633 50-CAT CTC AGC TTC TGC TCC CAC ATG ATG-30
12 HUA2 cDNA-F 50-GCCTCACCTAGGATGGCTCCTGGGCGTAAAAGAGG-30
13 HUA2 cDNA-R 50-GCCCTACAGGTAACCTCAAGCTGGCCTCCAACAATTC-30
14 G-2266 50-TGG AGC ATG CTA CCT CTC CT-30
15 G-2267 50-CCA TGT AAT GGG GTT TTT GG-30
16 G-5431 50-ACTGAAGGAGGATTAGCTGCGGCTGAG-30
17 G-5432 50-GTATAGTTGAATCAAGTATCAAGCGTG-30
18 HUA2-A 50-CTCGTTTCGTGATCCGTAGC-30
19 HUA2-B 50-TTGGGCGATGAGGATTCTC-30
20 N-0078 50-GAG CCT TAC AAC GCT ACT CTG TCT GTC-30
21 N-0079 50-ACA CCA GAC ATA GTA GCA GAA ATC AAG-30
22 G-0640 50-AGA TTA GAG AGA AGT ATT ACC CGA ATC-30
23 G-11926 50-GAT TTC AGA AAA TAA GAG CTG ATT-30
24 MKD15p31 50-CAA GGT CTG TTG CCA TTC TG-30
25 G-2307 50-CCA GGT CGT CTA AAG CAC TGA CAC G-30
26 G-3142 50-GGGCTAATGTGTGTCACAGTAC-30
27 G-2305 50-CCA ACT TTC GTA TGA GAA GTT CCA CC-30
Primer numbers and their uses are as follows: 1/2, amplification of 2.5kb genomic fragment containing an upstream intergenic region and the 50-
end of the HUA2 genomic sequence; 3/4, FLC RT-PCR; 5/6, HUA2-Ler/hua2-5 genotyping (restriction digestion with Tru1I/MseI: HUA2-Ler 434,
413, 166, 42bp; hua2-5 434, 413,208 bp); 1/2/7, Genotyping hua2-6 (T-DNA insertion); 8/9, genotyping hua2-1 (restriction digestion with PstI: WT,
427, 260 bps; hua2-1, 687 bps); 10/11, FLC qRT-PCR; 12/13, amplification of full-length HUA2 cDNA; 14/15, HUA2 RT-PCR; 16/17, FRI RT-PCR;
18/19, genotyping HUA2-Sy-0 (restriction digestion with EcoRI: HUA2-Sy-0, 88, 290, 71, 731 bps; HUA2-Ler 88, 361, 731 bps); 20/21, b-Tubulin2
RT-PCR; 22/23, genotyping ag-4 (restriction digestion with HinfI: WT, 128, 20 bps; and ag-4, 148 bps); 24/25, detection of Y969D HUA2-Sy-0 poly-
morphism by sequencing; and 26/27, detection of E525K HUA2-Sy-0 polymorphism by sequencing.
S8







![[Early liver damage caused by alcohol]](https://img.pdfslide.net/doc/110x75/634ecb9601c04712f40665a3/early-liver-damage-caused-by-alcohol.jpg)





















