Embed Size (px)
Citation preview

Induced Variations in Brassinosteroid Genes DefineBarley Height and Sturdiness, and Expand the GreenRevolution Genetic Toolkit1[C][W][OPEN]
Christoph Dockter2, Damian Gruszka2, Ilka Braumann, Arnis Druka, Ilze Druka, Jerome Franckowiak,Simon P. Gough, Anna Janeczko, Marzena Kurowska, Joakim Lundqvist, Udda Lundqvist,Marek Marzec, Izabela Matyszczak, André H. Müller3, Jana Oklestkova, Burkhard Schulz,Shakhira Zakhrabekova, and Mats Hansson4*
Carlsberg Laboratory, DK–1799 Copenhagen V, Denmark (C.D., I.B., S.P.G., J.L., I.M., A.H.M., S.Z., M.H.); Departmentof Genetics, Faculty of Biology and Environment Protection, University of Silesia, PL–40–032 Katowice, Poland(D.G., M.K., M.M.); James Hutton Institute, Invergowrie, Dundee DD2 5DA, United Kingdom (A.D., I.D.);Department of Agriculture, Fishery, and Forestry, Agri-Science Queensland, Hermitage Research Facility, Warwick,Queensland 4370, Australia (J.F.); Institute of Plant Physiology, Polish Academy of Sciences, 30–239 Krakow, Poland(A.J.); Nordic Genetic Resource Center, SE–230 53 Alnarp, Sweden (U.L.); Laboratory of Growth Regulators, Centreof the Region Haná for Biotechnological and Agricultural Research, Palacký University, and Institute of ExperimentalBotany, Academy of Sciences of the Czech Republic, CZ–783 71 Olomouc, Czech Republic (J.O.); and Department ofPlant Science and Landscape Architecture, University of Maryland, College Park, Maryland 20742 (B.S.)
Reduced plant height and culm robustness are quantitative characteristics important for assuring cereal crop yield and quality underadverse weather conditions. A very limited number of short-culm mutant alleles were introduced into commercial crop cultivars duringthe Green Revolution. We identified phenotypic traits, including sturdy culm, specific for deficiencies in brassinosteroid biosynthesis andsignaling in semidwarf mutants of barley (Hordeum vulgare). This set of characteristic traits was explored to perform a phenotypic screen ofnear-isogenic short-culm mutant lines from the brachytic, breviaristatum, dense spike, erectoides, semibrachytic, semidwarf, and slender dwarfmutant groups. In silico mapping of brassinosteroid-related genes in the barley genome in combination with sequencing of barley mutantlines assigned more than 20 historic mutants to three brassinosteroid-biosynthesis genes (BRASSINOSTEROID-6-OXIDASE,CONSTITUTIVE PHOTOMORPHOGENIC DWARF, and DIMINUTO) and one brassinosteroid-signaling gene (BRASSINOSTEROID-INSENSITIVE1 [HvBRI1]). Analyses of F2 and M2 populations, allelic crosses, and modeling of nonsynonymous amino acid exchangesin protein crystal structures gave a further understanding of the control of barley plant architecture and sturdiness by brassinosteroid-related genes. Alternatives to the widely used but highly temperature-sensitive uzu1.a allele ofHvBRI1 represent potential genetic buildingblocks for breeding strategies with sturdy and climate-tolerant barley cultivars.
The introduction of dwarfing genes to increase culmsturdiness of cereal crops was crucial for the first GreenRevolution (Hedden, 2003). The culms of tall cereal cropswere not strong enough to support the heavy spikes ofhigh-yielding cultivars, especially under high-nitrogenconditions. As a result, plants fell over, a process knownas lodging. This caused losses in yield and grain-qualityissues attributable to fungal infections, mycotoxin con-tamination, and preharvest germination (Rajkumara,2008). Today, a second Green Revolution is on its way, torevolutionize the agricultural sector and to ensure foodproduction for a growing world population. Concur-rently, global climate change is expected to cause morefrequent occurrences of extreme weather conditions, in-cluding thunderstorms with torrential rain and strongwinds, thus promoting cereal culm breakage (Porter andSemenov, 2005; National Climate Assessment Develop-ment Advisory Committee, 2013). Accordingly, plantarchitectures that resist lodging remain a major crop-improvement goal and identification of genes that reg-ulate culm length is required to enhance the genetictoolbox in order to facilitate efficient marker-assistedbreeding. The mutations and the corresponding genes
1 This work was supported by the Carlsberg Foundation, GrøntUdviklings- og Demonstrations Program (grant no. 34009–12–0522), theDeutche Forschungsgemeinschaft (grant no. DO1482/1–1), the Minister-stwoNauki i SzkolnictwaWyższego (grant nos. PBZ–MNiSW–2/3/2006/8 and IP2011–016471), the Ministerstwo Rolnictwa i Rozwoju Wsi (grantno. HOR hn–801/11/14–PO–0114–002), the National Science Foundation(grant no. CAREER-IOS 1054918), Czech Republic Ministry of Education,Youth and Sports (grant no. LK 21306), and the European Commission(funding from the CropLife Marie Curie Initial Training Network and theEuropean Research Area in Plant Genomics BARCODE project).
2 These authors contributed equally to the article.3 Present address: Department of Structural Biology, StanfordUniversity
School of Medicine, Fairchild Science Building, D143, Stanford, CA 94305.4 Present address: Copenhagen Plant Science Center, University of
Copenhagen, DK–1871 Frederiksberg C, Copenhagen, Denmark.* Address correspondence to [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policy de-scribed in the Instructions for Authors (www.plantphysiol.org) is:Mats Hansson ([email protected]).
[C] Some figures in this article are displayed in color online but inblack and white in the print edition.
[W] The online version of this article contains Web-only data.[OPEN] Articles can be viewed online without a subscription.www.plantphysiol.org/cgi/doi/10.1104/pp.114.250738
1912 Plant Physiology�, December 2014, Vol. 166, pp. 1912–1927, www.plantphysiol.org � 2014 American Society of Plant Biologists. All Rights Reserved. www.plant.org on December 22, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2014 American Society of Plant Biologists. All rights reserved.

that enabled the Green Revolution in wheat (Triticumaestivum) and rice (Oryza sativa) have been identified(Hedden, 2003). They all relate to gibberellin metabolismand signal transduction. It is now known that other planthormones such as brassinosteroids are also involved inthe regulation of plant height. Knowledge of the molec-ular mechanisms underlying the effects of the two hor-mones on cell elongation and division has mainly comefrom studies in Arabidopsis (Arabidopsis thaliana; Baiet al., 2012). Mutant-based breeding strategies to fine-tune brassinosteroid metabolism and signaling path-ways could improve lodging behavior in modern crops(Vriet et al., 2012) such as barley (Hordeum vulgare),which is the fourth most abundant cereal in both areaand tonnage harvested (http://faostat.fao.org).A short-culm phenotype in crops is often accompanied
by other phenotypic changes. Depending on the pene-trance of such pleiotropic characters, but also the pa-rental background and different scientific traditions andexpertise, short-culmed barley mutants were historicallydivided into groups, such as brachytic (brh), breviaristatum(ari), dense spike (dsp), erectoides (ert), semibrachytic (uzu),semidwarf (sdw), or slender dwarf (sld; Franckowiak andLundqvist, 2012). Subsequent mutant characterizationwas limited to intragroup screens and very few allelismtests between mutants from different groups have beenreported (Franckowiak and Lundqvist, 2012). Althoughthe total number of short-culm barley mutants exceeds500 (Franckowiak and Lundqvist, 2012), very few havebeen characterized at the DNA level (Helliwell et al.,2001; Jia et al., 2009; Chandler and Harding, 2013;Houston et al., 2013). One of the first identified haplo-types was uzu barley (Chono et al., 2003). TheUzu1 geneencodes the brassinosteroid hormone receptor and isorthologous to the BRASSINOSTEROID-INSENSITIVE1(BRI1) gene of Arabidopsis, a crucial promoter of plantgrowth (Li and Chory, 1997). The uzu1.a allele has beenused in East Asia for over a century and is presentlydistributed in winter barley cultivars in Japan, the Ko-rean peninsula, and China (Saisho et al., 2004). Its agro-nomic importance comes from the short and sturdy culmthat provides lodging resistance, and an upright plantarchitecture that tolerates dense planting.Today, more than 50 different brassinosteroids have
been identified in plants (Bajguz and Tretyn, 2003). Mostare intermediates of the complex biosynthetic pathway(Shimada et al., 2001). Approximately nine genes codefor the enzymes that participate in the biosyntheticpathway from episterol to brassinolide (SupplementalFig. S1). Brassinosteroid deficiency is caused by down-regulation of these genes, but it can also be associatedwith brassinosteroid signaling. The first protein in thesignaling network is the brassinosteroid receptor enco-ded by BRI1 (Li and Chory, 1997; Kim and Wang,2010). In this work, we show how to visually identifybrassinosteroid-mutant barley plants and we describemore than 20 relevant mutations in four genes of thebrassinosteroid biosynthesis and signaling pathways thatcan be used in marker-assisted breeding strategies.
RESULTS
Phenotypic Identification of Brassinosteroid Mutantsin Barley
A large number of brh, ari, dsp, ert, uzu, sdw, and sldshort-culm mutants have been induced in various barleycultivars, and many mutants within each group werefound to be allelic (Franckowiak and Lundqvist, 2012).We hypothesized that mutants from different groupsmight be allelic too; however, because of their differentgenetic backgrounds, direct comparison of their mutantphenotypes was impossible. Representative lines fromthese groups, along with many other mutants, were re-cently crossed into the genetic background of barley‘Bowman’ and genotyped with up to 3,000 markers(Druka et al., 2011). By growing such near-isogenic linesin parallel in the same environment, we were able toperform a comprehensive comparison of their mu-tant phenotypes independently of their original geneticbackground or preconceptions derived from previoushistoric classification (Fig. 1).
The brassinosteroid receptor, encoded by Uzu1(HvBRI1), is known to regulate plant height in barley(Chono et al., 2003). We used BW885, the near-isogenicline carrying the uzu1.a allele (Druka et al., 2011), as anideotype for a barley brassinosteroid mutant. Whengrown at low temperatures, the uzu1.a mutant is a semi-dwarf with 80% of wild-type culm length (Fig. 2). Theelongation of upper-stem internodes is particularly re-duced while the stem diameter remains unaltered. Theoverall plant architecture is more erect, with acute leaf-blade angles. Short-awned spikes are compact with densebasal spikelets, and frequently with opposite spikelets inthe tip caused by irregular elongation of rachis internodes.Leaf margins and auricles of uzu1.a have a slightly un-dulating appearance, similar to the wavy leaf phenotypefound in maize (Zea mays) Wavy auricle in blade1 mutants(Hay and Hake, 2004). The same phenotypic detail, but toa more extreme extent, was found in cv Bowman wild-type plants treated with propiconazole (Fig. 2), a potentinhibitor of brassinosteroid biosynthesis (Hartwig et al.,2012). In anatomical and microscopic analyses, we foundother phenotypic details (Supplemental Fig. S2).
We used the unique combination of visiblebrassinosteroid-mutant characters once they wereestablished to perform a phenotypic screen of 160 cvBowman near-isogenic lines with reduced culm length.Sixteen lines fulfilled the brassinosteroid phenotypecriteria. These mutants were found in the brh, uzu, ari,and ert groups (Fig. 3). We also tested the response ofdark-grown seedling leaves to exogenously appliedbrassinolide (Honda et al., 2003), in order to differentiatepotential brassinosteroid-signaling mutants from thosewith impaired brassinosteroid biosynthesis. In this so-called leaf-unrolling test, the mutants were grown indarkness for 6 d at 26°C and tightly rolled leaf segmentswere immersed in water or in a solution containing themost active brassinosteroid, brassinolide, for 72 h. Mu-tants ari.256, ert-ii.79, and uzu1.a showed greatly reduced
Plant Physiol. Vol. 166, 2014 1913
Variation in Barley Brassinosteroid Genes
www.plant.org on December 22, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

leaf-unrolling response to brassinolide (Fig. 3D), sug-gesting defective brassinosteroid signaling. In rice, a leaflamina inclination assay has been successfully used toidentify brassinosteroid signaling mutants (Fujioka et al.,1998; Hong et al., 2003). We adopted this method tobarley. Addition of brassinolide to the tip of barleyseedlings resulted in an increased angle between the firstleaf blade and culm in wild-type plants and those mu-tants that responded to exogenously applied brassino-lide. Mutants insensitive in the leaf-unrolling assay alsomissed increased leaf inclination, an indication for a de-fect in brassinosteroid signaling (Supplemental Fig. S3).
In order to test the robustness of the brassinosteroid-deficient characters for direct phenotypic screening, westudied 950 M2 plants of a chemically mutagenizeddoubled-haploid population of the barley line H930-36.Of 16 candidate plants with short-culm phenotypes, oneshowed the specific brassinosteroid-deficient characters.In the leaf-unrolling test, the mutant leaf segmentsshowed reduced response to added brassinolide (Fig. 4,A–F). Thus, the mutant was grouped with ari.256, ert-ii.79,and uzu1.a, and selected for further analysis of thebrassinosteroid-signaling gene, HvBRI1.
Connecting Brassinosteroid Biosynthesis Genes withBarley Mutant Loci
A rich variety of brassinosteroids are intermediatesof the complex biosynthetic pathway (SupplementalFig. S1). After initial steps, the pathway branches intoan early or late C-6 oxidation route. Several enzymeshave broad substrate specificity. Therefore, they cata-lyze multiple steps of the pathway. The final product isbrassinolide, which is generated by conversion fromcastasterone. The oxidase catalyzing the final step hasnot been found in cereal grasses, like barley. In theseplants, castasterone was suggested to be the bioactiveend product of the brassinosteroid biosynthetic path-way (Kim et al., 2008). We identified nine genes en-coding enzymes of the pathway in the barley genomeby homology searches between deduced polypeptidesequences of barley and Arabidopsis (SupplementalFig. S4). These were mapped on the barley genome insilico using the barley genome zipper (Mayer et al.,2011) and the barley physical map (The InternationalBarley Genome Sequencing Consortium, 2012). Thegenomic locations were then compared with the mutant
Figure 2. The brassinosteroid pheno-type of the cv Bowman near-isogenicline BW885 (uzu1.a) compared withwild-type cv Bowman (Bow). A, Grossmorphology and overall plant archi-tecture. B and C, Awns and spikes areshort with reduced rachis-internodelength at the base and tip. D and E,Leaves are erect with undulating leafmargins. The latter phenotype was alsofound in wild-type cv Bowman grownin the presence of the brassinosteroid-biosynthetic inhibitor propiconazole(PCZ). Scale bars = 10 cm. [See onlinearticle for color version of this figure.]
Figure 1. Main barley complementationgroups in the cv Bowman near-isogenicline collection, which contain mutantswith a short-culm phenotype. Gross mor-phology of representatives of the barleymutant groups brh, ari, dsp, ert, uzu, sdw,and sld comparedwith cv Bowman. Scalebar = 10 cm. [See online article for colorversion of this figure.]
1914 Plant Physiol. Vol. 166, 2014
Dockter et al.
www.plant.org on December 22, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

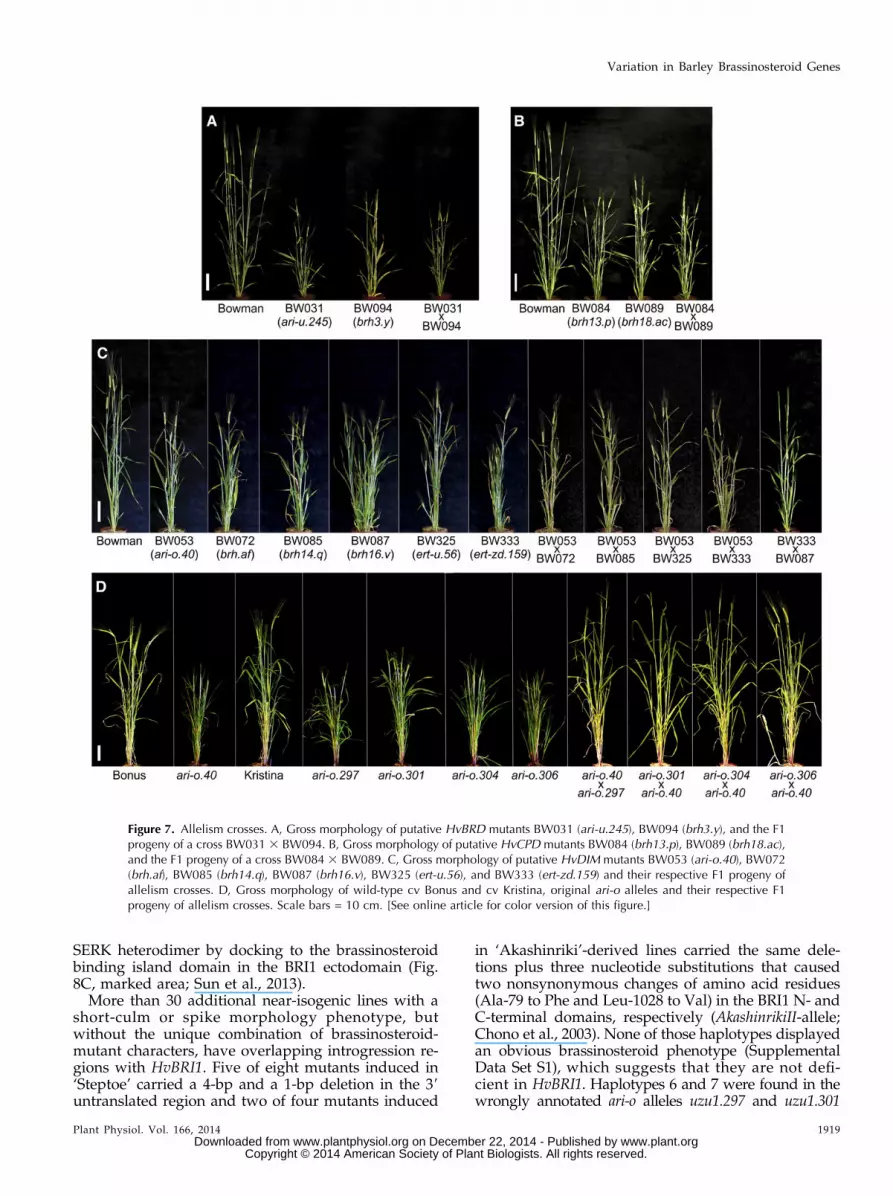
donor introgressions of the selected near-isogenic lines,which suggest that the mutants are alleles of three bio-synthetic genes, (BRASSINOSTEROID-6-OXIDASE[HvBRD], CONSTITUTIVE PHOTOMORPHOGENICDWARF [HvCPD], and DIMINUTO [HvDIM]), or ofthe brassinosteroid receptor gene HvBRI1 (Fig. 5). Thiswas tested by sequencing of the genes from availablenear-isogenic lines and historic barley mutants, and byperforming allelic crosses.
Genetic Characterization of HvBRD Mutants
HvBRD, encoding a brassinosteroid-6-oxidase, is lo-cated in the telomeric region of the short arm of barleychromosome 2H (Fig. 5; Supplemental Fig. S5A). Thelocation of HvBRD is within the mutant donor intro-gressions of the near-isogenic lines ari-u.245, brh3.g, brh3.h,brh3.y, and ert-t.55. In the barley genome zipper, thecommon genetic donor interval of those lines contained21 gene models but did not include the HvBRD orthologof Brachypodium distachyon, rice, and sorghum (Sorghumbicolor). In EnsemblPlant (http://plants.ensembl.org),the same interval contained 68 gene models includingHvBRD. Sequencing of HvBRD identified nonsense
mutations in ari-u.245 (T1676A, Leu-257 replaced bystop), brh3.g, and brh3.h (G2723A, Trp-444 replaced bystop), and a point mutation in brh3.y (G2183A) causing aGly-353 to Asp modification in the dioxygen-binding siteof HvBRD (Fig. 6; Supplemental Fig. S5B; SupplementalData Set S1). Allelism between available brh3 and ert-t lociwas recently proven (Dahleen et al., 2005; Franckowiakand Lundqvist, 2012). In this study, we additionallyverified the allelic nature of ari-u.245 to the brh3 and ert-tgroups by a cross between ari-u.245 and brh3.y. The F1progeny, heterozygous for the parental mutations inHvBRD, clearly resembled the brassinosteroid-deficientphenotype (Fig. 7A). We also identified mutations inbrh3.i and ert-t.437, which have been reported as allelicmutants (Dahleen et al., 2005) but are not representedin the collection of near-isogenic lines. The nonsensemutation in brh3.i was identical to brh3.g and brh3.h,and the point mutation in ert-t.437 was identical tobrh3.y. Other genetic differences in HvBRD clearlyindicate an independent history of ert-t.437 and brh3.y(Supplemental Data Set S1). No mutation within thecoding region of HvBRD could be found in BW324(ert-t.55) or the original ert-t.55 mutant, even though themutant was reported allelic to brh3 mutants (Dahleenet al., 2005) and to ert-t.437 (Franckowiak and Lundqvist,
Figure 3. Phenotypic details of mutants in thecollection of cv Bowman near-isogenic lines with anuzu1.a-like plant architecture. A, Reduced culm lengthdue to short upper internodes. B, Irregular rachis-internode length and short awns. C, Acute leaf anglesand undulating leaf margins. D, Insensitivity to exoge-nously applied brassinolide (uzu1.a, uzu1.256, andert-ii.79) in a leaf-unrolling bioassay is indicative ofdefects in brassinosteroid signaling. Mutant uzu1.256was renamed from its previous name ari.256. Scalebar = 10 cm. [See online article for color version ofthis figure.]
Plant Physiol. Vol. 166, 2014 1915
Variation in Barley Brassinosteroid Genes
www.plant.org on December 22, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

2012). We therefore suggest that the ert-t.55 mutation islocated outside the sequenced region ofHvBRD, possiblyin a regulatory element, or that the mutation has causeda rearrangement in the telomeric region of chromosome 2H,not detectable in our experimental setup. Mutant ert-t.55was induced by x-ray treatment (Supplemental Data Set
S1). BW093, the presumed near-isogenic line of brh3.i,neither had an introgression on chromosome 2H (Drukaet al., 2011) nor harbored the brh3.i haplotype although itshowed a short-culm phenotype (Supplemental Data SetS1). In contrast with the original line brh3.i, BW093 wasalso missing the combination of brassinosteroid-mutantcharacters, indicating a diverse ancestry of the geno-types. The connection between mutations in HvBRD andthe brassinosteroid-deficient phenotype, was furtherstrengthened by an exact cosegregation of the phenotypewith the brh3.g allele in an F2-mapping population. Thatis, 103 plants genotyped as homozygous brh3.g mutantsall had a short awn length (range, 4.5–8.1 cm; average 6SD, 6.86 0.81 cm) in contrast with the long awn lengths of190 heterozygous mutants (10.5–14.6 cm; 12.6 6 0.84 cm)and 90 wild-type plants (11.4–15.0 cm; 12.8 6 0.90 cm).
Genetic Characterization of HvCPD Mutants
HvCPD, encoding the barley C-23a-hydroxylase cy-tochrome P450 90A1 (CYP90A1), could not be localizedon the physical map of the barley genome. Instead, thesynteny to B. distachyon, rice, and sorghum genomes(Mayer et al., 2011) was used to identify likely markersfor HvCPD. Among mutants with a brassinosteroid-deficient phenotype, only brh13.p has an introgressionregion (Druka et al., 2011) colocalizing with the sug-gested position of HvCPD at 44.24 centimorgans (cM) onchromosome 5H (Fig. 5; Supplemental Fig. S6A). How-ever, the introgression region of brh13.p is large andsupposed to contain 1,380 gene models according to thebarley genome zipper (Mayer et al., 2011) and 808 genemodels according to EnsemblPlant (http://plants.ensembl.org) but only one brassinosteroid-biosynthesisgene, HvCPD. The near-isogenic line of brh18.acwas notincluded in the marker analysis by Druka et al. (2011),but brh18.ac has previously been linked to brh13.p viathe Simple Sequence Repeat markers Bmag0387 andHvLEU (Dahleen et al., 2005). The latter is anchoredapproximately 6 cM from HvCPD. We performed across between mutant lines brh13.p and brh18.ac. TheF1 progeny, heterozygous for the parental mutationsin HvCPD, clearly displayed a brassinosteroid-mutantphenotype, verifying that brh13.p and brh18.ac are allelic(Fig. 7B). Sequencing of HvCPD showed brh13.p andbrh18.ac to carry point mutations (C2562T and C2807T,respectively) causing substitutions of highly conservedamino acid residues (Pro-445 to Leu in brh13.p and Pro-479 to Ser in in brh18.ac). Pro-445 and Pro-479 are lo-cated within and close to the highly conserved hemebinding site, respectively, in the C-terminal part ofHvCPD (Fig. 6; Supplemental Fig. S6B; SupplementalData Set S1). It was noted that 11 additional near-isogenic lines with a short-culm phenotype, but with-out other distinct brassinosteroid-mutant characters,have overlapping introgression regions with HvCPD(Supplemental Data Set S1). Sequencing of HvCPD fromthese 11 lines revealed no mutations, which suggeststhat they are not deficient in HvCPD.
Figure 4. Phenotypic characterization of the HvBRI1 allele uzu1.ccompared with the double-haploid line H930-36. A, Gross morphol-ogy and overall plant architecture with reduced culm length. B, Acuteleaf angles. C, Awns and spikes are short with reduced rachis-internodelength. D, Undulated leaf margins. E, Reduced leaf elongation underdark growing conditions. F, Reduced sensitivity to exogenously appliedbrassinolide (BL) in a leaf-unrolling bioassay. G and H, Gross mor-phology ofHvBRI1 alleles uzu1.a, uzu1.b, uzu1.c, and their respectiveF1 progeny of allelism crosses. Scale bars = 10 cm. [See online articlefor color version of this figure.]
1916 Plant Physiol. Vol. 166, 2014
Dockter et al.
www.plant.org on December 22, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

Genetic Characterization of HvDIM Mutants
The sequence of HvDIM, encoding the barley D5-sterol-D24-reductase DIMINUTO, corresponds directly tosingle-nucleotide polymorphism (SNP) marker 1_0547in the telomere on the long arm of chromosome 7H. Thesix near-isogenic lines of ari-o.40, brh.af, brh14.q, brh16.v,ert-u.56, and ert-zd.159 have a small, common geneticdonor interval containing the HvDIM locus (Fig. 5;Supplemental Fig. S7A). In addition, the six mutantsshow a brassinosteroid-deficient phenotype including ashort culm mainly caused by a specific and extremeshortening of the second culm internode (Fig. 3A). Inthe barley genome zipper (Mayer et al., 2011), the in-terval contained three gene models including theHvDIM ortholog of B. distachyon, rice, and sorghum.In EnsemblPlant (http://plants.ensembl.org), the sameinterval contained 27 gene models. Severe DNA changeswere identified in the six mutants (Supplemental DataSet S1). In brh14.q, induced by ethyl methanesulfonatetreatment, a C628T substitution was detected resultingin the exchange of Leu-210 to Phe in the highly con-served FAD-binding site. Mutation ert-zd.159 was foundin the same domain (A530T, Glu-177 to Val), whereasthe mutant ari-o.40 had a T1171A substitution re-sulting in a Trp-391 to Arg exchange. Although Trp-391is located in a domain of unknown function, Trp-391 ishighly conserved among plant DIM polypeptides and
also in comparison to the human ortholog of DIM,SELADIN-1 (Supplemental Fig. S7B; Greeve et al.,2000). The deletions found in brh16.v (C3937 deleted,causing a frame shift followed by a premature stopcodon), ert-u.56 (exon 1 deleted, induced by x-rays), andbrh.af (deletion of .12 kb, induced by neutrons) furtherverified HvDIM as the candidate gene. Crosses gener-ated F1 progenies with identical phenotypes to theirparental lines, verifying the allelic character of thegene loci (Fig. 7C). In addition to ari-o.40, six otheralleles of ari-o are available. HvDIMmutations in ari-o.43and ari-o.143 were identical to ari-o.40, whereas nomutations in HvDIM were found in ari-o.297, ari-o.301,ari-o.304, and ari-o.306 even though they showed abrassinosteroid-deficient phenotype. Crosses of ari-o.297,ari-o.301, ari-o.304, and ari-o.306 to ari-o.40 resulted inF1 progenies with a tall wild-type phenotype withoutbrassinosteroid-mutant characters (Fig. 7D). This con-firmed that ari-o.297, ari-o.301, ari-o.304, and ari-o.306 arewrongly annotated alleles of HvDIM. Sequencing thelatter four lines for HvCPD, HvBRD, and HvBRI1identified three mutants: nonsynonymous point muta-tions in HvBRI1 in ari-o.297 (T158C, Phe-53 to Ser;renamed uzu1.297) and ari-o.301 (T845A, Val-282 toAsp; renamed uzu1.301), and a nonsense mutation inHvBRD in ari-o.304 (A2736T; Lys-449 replaced by stop;renamed ari-u.304; Fig. 6). No mutation was found inari-o.306 (renamed ari.306).
Figure 5. In silico mapping of brassinosteroid bio-synthesis genes and receptor gene HvBRI1 revealedmutant candidates in the collection of cv Bowmannear-isogenic lines. MLOC-gene positions on barleychromosomes (gray) according to the barley ge-nome map (The International Barley GenomeSequencing Consortium, 2012); mutant donorintrogressions (black) according to Druka et al.(2011). Loci HvDET2 and HvCPD are not anchoredin the barley genome map, but were localized in thebarley genome zipper (Mayer et al., 2011). [Seeonline article for color version of this figure.]
Plant Physiol. Vol. 166, 2014 1917
Variation in Barley Brassinosteroid Genes
www.plant.org on December 22, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

Genetic Characterization and Novel Alleles of HvBRI1
In addition to the uzu1 standard line BW885, thereported uzu1.a mutation (Chono et al., 2003) is alsopresent in BW860 (sld1.a + uzu1.a) and BW912 (wst1.c +uzu1.a). These three near-isogenic lines delimit theHvBRI1 position to a shared interval covered by nineSNPs on chromosome 3H (Fig. 5; Supplemental Fig. S8A).This interval contained 150 gene models according toEnsemblPlant (http://plants.ensembl.org) and 217 genemodels according to the barley genome zipper, includingthe BRI1 ortholog of B. distachyon, rice, and sorghum.Sequencing of HvBRI1 from those lines revealed the ex-pected single substitution A2612G (His-857 to Arg),known as allele uzu1.a, which is found in Northeast Asianshort-culm cultivars and landraces such as ‘Akashinriki’(Chono et al., 2003; Supplemental Data Set S1).
HvBRI1 is also located within the introgressionregions of near-isogenic lines ari.256 and ert-ii.79(Supplemental Fig. S8A), which, like BW885 (uzu1.a),showed a brassinosteroid signaling-deficient pheno-type in a leaf-unrolling test (Fig. 3D). Sequencing ofHvBRI1 revealed mutations in both ari.256 (A1733T;renamed uzu1.256) and ert-ii.79 (C1760A and C1761A;Fig. 6; Supplemental Fig. S8B; Supplemental Data SetS1). The amino acid residues affected by the muta-tions in uzu1.256 (Arg-564 to Trp) and ert-ii.79 (Thr-573 to
Lys) are located in the region encoding the steroid-binding island domain (Fig. 8, A and B). The introduc-tion of the charged Lys-573 to the hydrophobic active sitesurrounding residues, destroying charge neutrality, isexpected to prevent brassinosteroid binding. In bothcases, steric clashes with neighboring residues arelikely to occur (Fig. 8B, inserts); as a consequence, thesteroid binding and the general positioning of thebrassinosteroid-binding site may be affected. Alter-natively, the change in the surface polarity (Fig. 8,C and D) in the putative docking site of BRASSINOS-TEROID INSENSITIVE1-ASSOCIATED RECEPTORKINASE1 (BAK1) and other SOMATIC EMBRYO-GENESIS RECEPTOR KINASE (SERK)-like coreceptors(Fig. 8C, yellow circle) may simply hinder the BRI1-SERK interaction, although less severe than in Arabi-dopsis bri1-102 (AtThr750 to Ile), which is a strongdwarf mutant (Supplemental Fig. S8B; Hothorn et al.,2011; She et al., 2011; Sun et al., 2013). By using theArabidopsis BAK1/SERK3 (AT4G33430) sequence asprobe, we identified at least five SERK-like barley genes(MLOC_19580, AK252995 [identical to MLOC_80630and MLOC_59982], MLOC_8229, MLOC_6922, andMLOC_79186) in available barley sequence databases(IPK Barley BLAST Server, http://webblast.ipk-gatersleben.de/). Their specific functions in barleyare not yet known, but they are likely to form a BRI1/
Figure 6. Structures of analyzed brassinosteroidgenes. A,HvBRD. B, HvCPD. C,HvDIM. D, HvBRI1.The location and number of exons (E) and the identityof mutations at the gene and peptide levels (threeletter code, italic) are indicated. Dotted lines in Crepresent deleted gene fragments in the respectivemutant accessions. Asterisks indicate nonsense mu-tations where amino-acid residues have been ex-changed by stop.
1918 Plant Physiol. Vol. 166, 2014
Dockter et al.
www.plant.org on December 22, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.
SERK heterodimer by docking to the brassinosteroidbinding island domain in the BRI1 ectodomain (Fig.8C, marked area; Sun et al., 2013).More than 30 additional near-isogenic lines with a
short-culm or spike morphology phenotype, butwithout the unique combination of brassinosteroid-mutant characters, have overlapping introgression re-gions with HvBRI1. Five of eight mutants induced in‘Steptoe’ carried a 4-bp and a 1-bp deletion in the 39untranslated region and two of four mutants induced
in ‘Akashinriki’-derived lines carried the same dele-tions plus three nucleotide substitutions that causedtwo nonsynonymous changes of amino acid residues(Ala-79 to Phe and Leu-1028 to Val) in the BRI1 N- andC-terminal domains, respectively (AkashinrikiII-allele;Chono et al., 2003). None of those haplotypes displayedan obvious brassinosteroid phenotype (SupplementalData Set S1), which suggests that they are not defi-cient in HvBRI1. Haplotypes 6 and 7 were found in thewrongly annotated ari-o alleles uzu1.297 and uzu1.301
Figure 7. Allelism crosses. A, Gross morphology of putative HvBRD mutants BW031 (ari-u.245), BW094 (brh3.y), and the F1progeny of a cross BW031 3 BW094. B, Gross morphology of putative HvCPD mutants BW084 (brh13.p), BW089 (brh18.ac),and the F1 progeny of a cross BW084 3 BW089. C, Gross morphology of putative HvDIM mutants BW053 (ari-o.40), BW072(brh.af), BW085 (brh14.q), BW087 (brh16.v), BW325 (ert-u.56), and BW333 (ert-zd.159) and their respective F1 progeny ofallelism crosses. D, Gross morphology of wild-type cv Bonus and cv Kristina, original ari-o alleles and their respective F1progeny of allelism crosses. Scale bars = 10 cm. [See online article for color version of this figure.]
Plant Physiol. Vol. 166, 2014 1919
Variation in Barley Brassinosteroid Genes
www.plant.org on December 22, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

(Fig. 7D). The amino acid exchange found in uzu1.297(Phe-53 to Ser; Fig. 8A) is located within the C lobe of theN-terminal cap (Hothorn et al., 2011). The exchange toSer-53 may destabilize the structure of this domaincausing endoplasmic reticulum retention of BRI1 (Honget al., 2008). The exchange of the nonpolar Val-282 resi-due to the negatively charged Asp in uzu1.301will resultin the formation of a salt bridge between Asp-282 andLys-302 (a-C distance 4.46Å), which may destabilize thestructure of the folded protein due to a loss of solvationenergy of Lys-302, which is solvent exposed (Bosshardet al., 2004). Alternatively, the exchange of the nonpolar
Val-282 to the negatively charged Asp may inhibit thesuperhelical arrangement (Ile-Pro spine) in leucine-richrepeat (LRR)-12 and thus cause a repositioning of theupstream LRRs and the N-terminal cap (Hothorn et al.,2011; Santiago et al., 2013).
Interestingly, the ert-ii.79 mutation was identifiedearlier in the uzu1 mutant 093AR (uzu1.b; Gruszkaet al., 2011b). To exclude a common ancestry between093AR and ert-ii.79, we genotyped the original acces-sions using 383 Amplified Fragment Length Poly-morphism (AFLP) markers. We found a high levelof polymorphism (7.6%) throughout the genomes,
Figure 8. Structural basis of brassinosteroid per-ception and signaling in barley HvBRI1 mutants.A to D, X-ray structure of the Arabidopsis BRI1brassinosteroid-binding domain (Protein DataBank accession no. 3RGZ; Hothorn et al., 2011;She et al., 2011) displayed as ribbon (A and B)and hydrophobicity (C and D) models, respec-tively. LRRs missing in monocots are shown inwhite, the brassinosteroid-binding site (islanddomain) in yellow, and the bound steroid hor-mone brassinolide in red. E and F, X-ray struc-ture of the Arabidopsis BRI1 kinase domain(Protein Data Bank accession no. 4OAB). Thetwo views are rotated 90˚ relative to each other.BRI1 in complex with ATP (red), the BRI1 acti-vation loop with phosphorylation sites (in yel-low), and the proposed protein-docking platformfor the BRI1 inhibitor BKI or the SERK cor-eceptors (circled) are highlighted (Bojar et al.,2014). The numbering of amino acid residuesrefers to the barley peptide sequence shown inSupplemental Figure S8B online in whichcorresponding residue positions in ArabidopsisAtBRI1 or rice OSBRI1 are indicated. Proteinstructures were visualized using Chimera soft-ware (Pettersen et al., 2004).
1920 Plant Physiol. Vol. 166, 2014
Dockter et al.
www.plant.org on December 22, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

indicating independent mutagenic events. The phe-notypic characteristics of uzu1.b could be seen in allthree genetic backgrounds (Fig. 9), supporting therobustness of the brassinosteroid-deficient charac-ters for direct phenotypic screening of barley mutantcollections.Sequencing of HvBRI1 from the brassinosteroid-
insensitive mutant isolated among the 950 M2 plants ofa chemically mutagenized population derived from thedoubled-haploid line H930-36, revealed a novel HvBRI1allele (a single-nucleotide G2171A substitution), whichwe named uzu1.c (Fig. 6; Supplemental Fig. S8B;Supplemental Data Set S1). Cosegregation of genotypeand phenotype in M3 plants and allelism tests with bothuzu1.a and uzu1.b verified the new uzu1 haplotype (Fig.4, G and H). Substitution of the semiconserved Arg-710
to Lys in close proximity to the BRI1 transmembrane-binding domain causes a mild brassinosteroid phenotype.No other mutations were found in HvBRI1 amongthe remaining 949 M2 plants. The successful iden-tification of the HvBRI1 mutant in a chemically in-duced mutant population and the clear brassinosteroidphenotypes seen in crosses and in different geneticbackgrounds (Figs. 4, 7, and 9) strongly suggest thatbrassinosteroid phenotypes are identifiable in mutantpopulations of most barley cultivars.
Temperature Sensitivity of uzu1.a
The uzu1.a allele, widely distributed in NortheastAsian winter barley cultivars, reduces lodging (Chono
Figure 9. Phenotypic characterization of uzu1.b lines ert-ii.79 (uzu1.b in cv Bonus), 093AR (uzu1.b in cv Aramir), and BW312(uzu1.b in cv Bowman). A, Gross morphology and general semidwarf phenotype. B, General spike phenotype with short awns,irregular rachis internodes, and elongated basal rachis internodes (a character not so pronounced in a cv Bowman geneticbackground). C, Erect leaf architecture. D, Undulating leaf margins of mutants compared with their respective parental cul-tivars. Scale bars = 10 cm in A and 5 cm in B.
Plant Physiol. Vol. 166, 2014 1921
Variation in Barley Brassinosteroid Genes
www.plant.org on December 22, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

et al., 2003; Saisho et al., 2004) but shows sensitivity toelevated temperatures. We compared the near-isogenicline of uzu1.a with other brassinosteroid signaling andbiosynthesis mutants grown at 14°C and 26°C. At 14°C,all mutants had an erect phenotype with height reduc-tion ranging from strong (brh3.h, 61% of wild-typelength) to weak semidwarfism (uzu1.256, 85%; Fig. 10).The average culm length of uzu1.a was 72% of the wild-type length. At 26°C, uzu1.a showed extreme dwarfing(29% of wild-type length) in contrast with the othermutants (65%–91%). When moved from 26°C to 14°C,already developed tillers remained short but new tillersgrew taller (Fig. 10). We conclude that the temperature-sensitive phenotype is not associated with brassinoste-roid or HvBRI1 mutations in general, but is specific forthe uzu1.a allele. We also measured the level of castas-terone, the suggested end product of the brassinosteroidbiosynthetic pathway in monocots (Kim et al., 2008). Thehighest level was found in uzu1.a grown at 26°C andin ert-ii.79, which are the signaling mutants with thestrongest phenotype. This finding correlates well withthe high levels of castasterone and brassinolide found inthe Arabidopsis mutant bri1-4 (Noguchi et al., 1999).Barley lines mutated in HvBRD, HvCPD, or HvDIM aredeficient in brassinosteroid biosynthesis prior the for-mation of castasterone and thus had the lowest castas-terone levels, as expected (Table I).
DISCUSSION
The Group of Brassinosteroid-Deficient Barley
The use of nitrogen fertilizers has an enormous impacton global cereal production and was a key componentof the Green Revolution. Initially, the culms were notstrong enough to support the heavy spikes of fertilizedplants, which resulted in lodging and overall loss inyield. It was therefore necessary to combine the use offertilizers with breeding of short-culm cultivars, whichare less prone to lodging due to their compact and sturdyculms (Hedden, 2003). Thus far, only a few genes havebeen explored in short-culm cultivars, such as Reducedheight1 (Rht1), Rht2, and Rht8 in wheat (Daoura et al.,2014), semidwarf1 (sd1) in rice (Nagano et al., 2005), andsdw1/denso in barley (Hellewell et al., 2000). The knownRht gene products belong to the DELLA subgroup of theGRAS protein family, which are thought to function astranscriptional regulators and interfere with the signaltransduction pathway of the gibberellin growth hormone(Pysh et al., 1999). By contrast, sd1 and sdw1/denso areinvolved in the production of gibberellin because theyencode gibberellin 20-oxidases of the biosynthetic path-way (Monna et al., 2002; Sasaki et al., 2002; Spielmeyeret al., 2002; Jia et al., 2009).
Plant height and cell elongation are also regulated bybrassinosteroids (Vriet et al., 2012). The access to a collec-tion of 160 short-culm mutants in a near-isogenic back-ground and with roughly mapped mutant gene locations(Druka et al., 2011), in combination with in silico genemapping using the latest barley genome resources (Mayeret al., 2011; The International Barley Genome SequencingConsortium, 2012), enabled us to effectively discoverbrassinosteroid mutants by a reverse approach of pheno-typing followed by genotyping. Unexpectedly, the identi-fied semidwarf gene loci do not belong to the same barleycomplementation group but were described as brh, uzu, ari,and ert barley. The history of those mutants might explainthis misleading pregrouping. The original mutations wereinduced and isolated in independent programs all over theworld over a time period of several decades. Thus, mu-tagenic events were described and grouped by differentbarley researchers. An aggravating factor was that theplant material used for mutagenesis was dependent on
Table I. Level of castasterone in barley near-isogenic lines carryingmutations in the brassinosteroid biosynthesis genes HvCPD, HvBRD,and HvDIM or the HvBRI1 gene encoding the brassinosteroid receptor
Data are presented as the average 6 SD unless otherwise indicated.
Near-Isogenic Line Genotype Castasterone
pmol g leaf 21
‘Bowman’ Wild type 2.68 6 0.40BW084 brh13.p (HvCPD) 0.36 6 0.06BW091 brh3.g (HvBRD) 0.50 6 0.06BW333 ert-zd.159 (HvDIM) 0.84 6 0.29BW033 uzu1.256 (HvBRI1) 4.52 6 0.53BW312 ert-ii.79 (HvBRI1) 9.39 6 1.76BW885 uzu1.a (HvBRI1) 7.43 6 0.21BW885 (26˚C) uzu1.a (HvBRI1) 10.31 6 0.18
Figure 10. Temperature-responsive phenotypes of brassinosteroidmutants in the collection of cv Bowman near-isogenic lines comparedwith wild-type cv Bowman. A, Culm length of plants grown at 14˚C(dark-colored bars) or 26˚C (light-colored bars). B, HvBRI1 mutantsgrown at 26˚C. At high temperatures, elongation of uzu1.a culms isstrongly reduced compared with cv Bowman and other brassinosteroidmutants. C and D, At 26˚C, uzu1.a leaf margins are strongly undulatingand internodes fail to elongate. E, Height reduction is partly neutral-ized by moving plants from warm temperature (26˚C) to cool tem-perature (14˚C). Squares and stars mark tillers developed at 26˚C and14˚C, respectively. Scale bars = 10 cm. [See online article for colorversion of this figure.]
1922 Plant Physiol. Vol. 166, 2014
Dockter et al.
www.plant.org on December 22, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

national cultivar preferences and, over time, was oftenchanged. Thus, even though the isolated brassinosteroidbarley mutants share the same characteristics, the con-nection between those original mutations was less obviousbecause the different parental backgrounds contributeother, distracting characters such as two- versus six-rowspikes. Only the unification of the genetic background inthe near-isogenic cv Bowman collection (Druka et al., 2011)helped to overcome this problem.The short culm of brassinosteroid and gibberellin
mutants is a trait of agronomic interest because itprovides resistance to lodging. The erect and uprightgrowth is unique to brassinosteroid mutants only anddistinguishes them from gibberellin mutants. The erectleaves enhance light capture for photosynthesis, serveas a nitrogen reservoir for grain filling, and enabledenser planting with a higher leaf surface per area ofland (Sakamoto et al., 2006). It was shown that ricemutants deficient in brassinosteroid biosynthesis en-hanced grain yield under conditions of dense planting(Sakamoto et al., 2006). Thus, brassinosteroid mutantsprovide additional traits for plant breeding that are notfound in gibberellin mutants.Interestingly, we only found mutations in three of
approximately nine genes involved in brassinosteroidbiosynthesis starting from episterol (Supplemental Fig.S1). The genes HvDIM, HvCPD, and HvBRD encodeenzymes from different parts of the pathway. Mutationsin the three genes show almost identical visible pheno-types, which suggests that loss of function of any otherbrassinosteroid biosynthesis gene should result in asimilar plant architecture. However, it was found that aknockout of DWARF4 (DWF4) in rice causes mild sem-idwarfism only (Sakamoto et al., 2006). Thus, it is pos-sible that barley HvDWF4 mutants were not isolated asshort-culm mutants and therefore were not included inthe mutant collections explored in this study. Similar,loss of function of a CYP90-like gene in rice resulted insevere dwarfism (Hong et al., 2003). Because severebarley dwarfs are not included in the collection of near-isogenic lines, it is possible that no suitable candidateplant for HvCYP90D1 was present in our screen.In addition, the existence of gene duplications in the
barley genome or gene families with overlapping func-tions might explain why brassinosteroid biosynthesismutants were only found in three genes. For example,DWF4 appears to be represented by two very similarDWF4-like genes at almost identical positions on chro-mosome 4H at 60 cM in the physical map of barley (TheInternational Barley Genome Sequencing Consortium,2012). It is possible that only a double mutant mightcause a brassinosteroid phenotype. Both HvDWF4 genesoverlap with the introgression regions of five near-isogenic lines with short-culm phenotypes, but withoutthe unique combination of brassinosteroid features. Se-quencing of HvDWF4 from these lines did not reveal anymutations. Thus, it is likely that these lines are deficientin one or several genes linked to HvDWF4, but not in-volved in brassinosteroid biosynthesis or signaling.
Future studies on barley mutants deficient in otherbrassinosteroid-related genes will show whether weshould regroup barley semidwarfs and introduce thegroup of brassinosteroid-deficient barley defined bythe unique combination of phenotypic characters andgenotypes described in this study. For example, screen-ing for genetic polymorphism in known brassinosteroid-related genes in Targeting Induced Local Lesions inGenomes (TILLING) populations would reveal thephenotypic variation, which is not covered in this studydue to the preset selection for short-culm mutants.
Induced Mutations in Barley
In our extensive screen for brassinosteroid mutants,we noted four interesting cases of identical mutations.In the first case, the mutants were induced by identicalchemical treatment (brh3.g, brh3.h, and brh3.i), in thesecond case by different treatments (ari-o.40, ari-o.43,and ari-o.143). In each case, the mutants were inducedin the same mutagenesis program, and thus we cannotexclude a mix-up of mutant accessions after the mu-tagenic treatment as it was suggested earlier for allelesof barley loci praematurum-a and third outer glume1(Houston et al., 2012; Zakhrabekova et al., 2012). Twoother cases, however, strongly point toward an inde-pendent ancestry of identical mutations. Mutants uzu1.b (093AR) and ert-ii.79 were induced in independentprograms in different decades and countries. Here, wetested the genetic background and found a high levelof polymorphism throughout the mutant genomes,strongly supporting their independent ancestry. In theother case, brh3.y and ert-t.437, silent mutations withinHvBRD, in addition to the loss-of-function mutation, areclear evidence of an independent ancestry (SupplementalData Set S1). Both cases invite speculation concerningmutation hotspots in barley, preferentially selected dueto the strong brassinosteroid-mutant phenotype. Mod-ern analytical techniques such as exome capture andgenotyping-by-sequencing will help to clarify this find-ing and other cases in the near future.
Lodging-Resistant Barley in a Changing Climate
The barley uzu1.a mutant is known to have al-terations in brassinosteroid signaling due to a non-synonymous amino acid exchange in the BRI1 kinasedomain (Chono et al., 2003). Although the uzu1.a genemodification has a long history in barley breeding, itremained exclusively in barley winter cultivars inNortheast Asia (Saisho et al., 2004). As shown in thisstudy, the uzu1.a heat-sensitive reduction in plantheight, severe in two-row spring barley grown inwarmer climates, might be a reason why this haplo-type did not spread globally during the Green Revo-lution. It is obvious that other alleles of HvBRI1,especially uzu1.256 with its milder phenotype andwithout the pronounced temperature sensitivity of
Plant Physiol. Vol. 166, 2014 1923
Variation in Barley Brassinosteroid Genes
www.plant.org on December 22, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

uzu1.a, should be considered as more reliable alter-natives for culm-length control in barley breeding.Interestingly, the uzu1.a mutation causes a change(His-857 to Arg) in a kinase surface domain (Bojaret al., 2014) that is not involved in ATP binding, nordoes it interfere with the phosphorylation of the acti-vation loop or with the position of the putative in-hibitor BKI1 and SERK coreceptor binding site (Fig. 8,E and F). This is in agreement with the mild brassi-nosteroid phenotype we found in uzu1.a grown inmoderate climate regimes. Mutations interfering withATP binding, phosphorylation, or SERK interactionare known to cause drastic phenotypes in Arabidopsisand rice (Supplemental Fig. S8B), whereas mutationsuzu1.a (at 14°C), d61-10, and bri1-301, all located withinthe same peripheral domain, result in mild phenotypesonly (Fig. 8, E and F; Supplemental Fig. S8B). Here,exchanges of amino acid residues might not disturbthe general structure of the kinase domain but in-stead interactions with other proteins of the signalingcascade, such as CONSTITUTIVE DIFFERENTIALGROWTH1 or BRASSINOSTEROID-SIGNALING-KINASEs. However, because the temperature sensitivityis specific for uzu1.a only, it is very likely that thedynamic of BRI1 is affected by the modification ofHis-857. Such dynamic conformational changes mightinvolve temporal stabilization by the cation-p interac-tion between His-857 and Phe-917 in the underlyinga-helix (Fig. 8, E and F). A highly charged Arg-857 mightpush the equilibrium to the conformational state wherethe protein is stabilized in its inactive state via thecation-p interaction. Interestingly, it has been shownthat elevated temperatures stabilize cation-p interactions(Prajapati et al., 2006). A temperature-dependent inacti-vation of BRI1 would be consistent with the temperature-sensitive dwarfism found in uzu1.a. We note that His-Pheor His-Tyr pairs are often arranged in very similarpositions (His-[X57–59]-Phe/Tyr) in many kinases, in-cluding plant receptor-like kinases (SupplementalData Set S2).
The global temperature is currently rising and it hasalso been postulated that the climate may become moreextreme with weather conditions that promote cereallodging (Porter and Semenov, 2005; National ClimateAssessment Development Advisory Committee, 2013).Accordingly, it would be desirable to have access to anincreased repertoire of lodging-tolerance genes that alsowork under higher temperatures. The more than 20temperature-insensitive mutations identified in thisstudy can be transferred to elite cultivars by recurrentbackcrosses. However, this is time-consuming and maybring unwanted variants through linked genes. Analternative approach is to screen for novel mutationsdirectly induced in elite cultivars that already havefavorable agronomic and grain-quality attributes. Thiscan be done when the gene of interest is known. Thevisual phenotyping provides a fast and inexpensivemethod of identifying brassinosteroid mutants, therebyefficiently reducing the number of candidate genes to beanalyzed in order to reveal the molecular cause of a short
and sturdy culm. An alternative approach is to createmutations through targeted mutagenesis and genomeediting. However, these techniques are still in their in-fancy in barley (Wendt et al., 2013) and although itwould be possible to create mutations without leavingany traces of foreign DNA in the resulting breeding lines,most countries regard such plants as genetically modi-fied or legal decisions are pending. Regardless of theoutcome of such decisions, public opinion has to show apositive acceptance to gene editing before it will be ofeconomic importance. In view of these obstacles, wepostulate that the more than 20 identified mutationsand the visual identification of brassinosteroid-deficientshort-culm phenotypes in induced mapping populationswill remain attractive for many years. We propose thatthe phenotypic screening can be performed in any ge-netic background. We are currently screening a largenumber of uncharacterized short-culm mutants (includ-ing severe dwarfs) in order to identify mutations in othergenes involved in brassinosteroid signaling and biosyn-thesis that were not revealed in this study.
MATERIALS AND METHODS
Plant Material and Growth Conditions
Barley (Hordeum vulgare ‘Bowman’) and other parental cultivars, cv Bowmannear-isogenic mutant lines (Druka et al., 2011) and original mutant lines(Supplemental Data Set S1), F1 plants derived from allelism crosses, and anF2-mapping population from a cross between BW091 (brh3.g) 3 ‘Bowman’were grown in a greenhouse at 18°C under 16-h-light/8-h-dark cycles.Temperature-dependent experiments were performed in a growth chamber ateither 14°C or 26°C under 16-h-light/8-h-dark cycles. Light intensity was set toa photon flux of 300 mmol m22 s21.
For propiconazole treatments, barley seeds of cv Bowman’ were imbibedand germinated on filter paper drenched with the respective propiconazoledilutions in water. Seeds were transplanted in soil after radicle emergence. Forpropiconazole treatments, barley plants were grown under greenhouse con-ditions at 26°C (day) and 18°C (night). Single plants were grown in 20-cm-wide pots in 1:1 (v/v) mixtures of Turface Athletics MVP (Profile Products)and Pro-Mix-BX with microrise (Premier Horticulture). Plants were fertilizedwith 0.02% (v/v) Miracle-Gro Excel (Scotts) adjusted to pH 6.0 following themanufacturer’s recommendations. The watering solution was supplementedwith 500 mM propiconazole solution using the commercial fungicide formu-lation Banner Maxx (Syngenta). Propiconazole treatments were applied dur-ing each watering cycle via soil drench. At full development of spikes,morphometric data were gathered and photographs were taken.
Plant seeds are either available from the Nordic Genetic Resource Center(http://www.nordgen.org) or from the National Small Grains Collection(http://www.ars.usda.gov). Seeds of mutant brh.af were kindly provided byAndy Kleinhofs.
Allelism Crosses
Allelism testswereperformed through crosses betweenbarley accessions carryingmutations in identical brassinosteroid-related genes. Plants were grown in agreenhouse as described above, emasculated, and pollinated within 3 d. The fol-lowing cv Bowman near-isogenic lines (Druka et al., 2011) were used for allelismtests concerning HvBRD, HvDIM and HvCPD: BW031 (ari-u.245) 3 BW094 (brh3.y),BW053 (ari-o.40) 3 BW072 (brh.af), BW053 (ari-o.40) 3 BW085 (brh14.q), BW053(ari-o.40) 3 BW325 (ert-u.56), BW053 (ari-o.40) 3 BW333 (ert-zd.159),BW333 (ert-zd.159)3 BW087 (brh16.v), and BW084 (brh13.p)3 BW089 (brh18.ac). Inaddition, we crossed the original mutant accession ari-o.40 with ari-o.297, ari-o.301,ari-o.304, and ari-o.306. In the case of HvBRI1, mutant uzu1.c (R710K) was crossed toboth BW885 (uzu1.a) and 093AR (uzu1.b). F1 progenies of all crosses were geno-typed to confirm successful crossings, and were grown to maturity for phenotypic
1924 Plant Physiol. Vol. 166, 2014
Dockter et al.
www.plant.org on December 22, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

analysis. Allelism tests between other mutants identified in this study are describedelsewhere (Dahleen et al., 2005; Franckowiak and Lundqvist, 2012).
Leaf-Segment Unrolling and Leaf LaminaInclination Assay
Leaf-segment unrolling tests were performed as described in Chono et al.(2003) and Gruszka et al. (2011a). The rate of leaf-segment unrolling wascalculated according to Wada et al. (1985). For leaf lamina inclination tests,plants were grown in vermiculite until the first leaf protruded 2 cm above thecoleoptile. One microliter of 2 mg mL21 brassinolide (24-epi-brassinolide;Sigma-Aldrich Chemie) solved in 96% (v/v) ethanol was added to the tip ofthe barley seedlings. The angle between the first leaf and the second leaf wasmeasured when the second leaf emerged.
Quantification of Castasterone Levels
Isolation and identification of the brassinosteroid biosynthetic intermediatecastasterone was performed according to a modified protocol described inSwaczynová et al. (2007) and Janeczko et al. (2010). All plants were grownfor 14 d at 17°C, except BW885, which was also grown at 26°C. For each genotype,measurements were performed in three replicates. The leaf material (2 g freshweight) was homogenized in 20 mL of 80% (v/v) cold methanol. After centrifu-gation, the supernatant was enriched in deuterium-labeled brassinosteroids (in-ternal standards) and passed through a Strata X column (Phenomenex). The boundfraction was eluted with acetonitrile and evaporated to dryness. Brassinosteroidswere determined by ultra-HPLC with tandem mass spectrometry.
In Silico Gene Mapping
Coding DNA sequences and protein sequences of Arabidopsis (Arabidopsisthaliana) genes STE1 (AT3G02580), DWF5 (AT1G50430), DIM/DWF1 (AT3G19820),DET2 (AT2G38050), CYP90B1/DWF4 (AT3G50660), CYP90C1/ROT3 (AT4G36380),CYP90D1 (AT3G13730), CYP90A1/CPD (AT5G05690), CYP85A/BR6ox/BRD1(AT5G38970), or BRD2 (AT3G30180) and rice (Oryza sativa) CYP724B1/DWF11(LOC_Os04g39430) were used as probes to identify full-length barley gene andpeptide sequences via the IPK Barley BLAST Server of the Leibniz Institute ofPlant Genetics and Crop Plant Research (http://webblast.ipk-gatersleben.de/).Inferred and anchored positions of barley high-confidence genes were extractedfrom the barley genome zipper (Mayer et al., 2011) and the barley genome map(The International Barley Genome Sequencing Consortium, 2012), respectively.Barley and Arabidopsis peptide sequences were aligned using Clustal Omegasoftware (http://www.ebi.ac.uk/Tools/msa/clustalo/).
Resequencing of HvBRD, HvCPD, HvDIM, HvDWF4,and HvBRI1
Genomic DNA was extracted from green leaf material using theREDExtract-N-Amp Plant PCR Kit (Sigma-Aldrich) according to the manu-facturer’s instructions. Green leaves of an F2-mapping population were sam-pled in 96-well format and genomic DNA was isolated using a Freedom EVO200 robot (Tecan Group). PCR amplifications of AK372445 (HvBRD),MLOC_10658 (HvCPD), MLOC_52405 (HvDIM), AK355174 (HvDWF4), andMLOC_5176 (HvBRI1) were performed according to the manufacturer’s pro-tocol (REDExtract-N-Amp PCR ReadyMix; Sigma-Aldrich).
PCRs for HvBRD fragments 1, 2, and 5 were performed for 42 cycles (initialdenaturation at 94°C/3 min followed by three cycles of 94°C/45 s, 61.5°C/45 s,and 72°C/90 s for extension; followed by three cycles of 94°C/45 s, 59.5°C/45 s,and 72°C/90 s for extension; and followed by 36 cycles of 94°C/45 s, primerpair-dependent annealing temperature/45 s, and 72°C/90 s for extension,with a final extension step of 72°C/10 min). PCRs for HvBRD fragments 3 and4 were performed for 30 cycles (initial denaturation at 94°C/3 min followed by30 cycles of 94°C/45 s, 54.5°C/45 s, and 72°C/105 s for extension, with a finalextension step of 72°C/10 min). PCRs for HvBRI1 were performed using theprotocol published in Gruszka et al. (2011b). PCRs for HvCPDwere performedfor 37 cycles (initial denaturation at 94°C/3 min followed by 37 cycles of94°C/45 s, 54.5°C/45 s, and 72°C/90 s for extension, with a final extension stepof 72°C/10 min). PCRs for HvDIM were performed for 40 cycles (initial dena-turation at 94°C/3 min followed by 40 cycles of 94°C/30 s, 59°C/30 s, and72°C/60–90 s for extension, with a final extension step of 72°C/10 min). PCRsfor HvDWF4 were performed for 42 cycles (initial denaturation at 94°C/3 min
followed by three cycles of 94°C/45 s, 58°C/45 s, and 72°C/90 s for extension;followed by three cycles of 94°C/45 s, 56°C/45 s, and 72°C/90 s for ex-tension; and followed by 36 cycles of 94°C/45 s, 54.5°C/45 s, and 72°C/90 sfor extension, with a final extension step of 72°C/10 min). Primer sequencesincluding specific annealing temperatures used for PCR amplifications arelisted and described in Supplemental Data Set S3.
PCR productswere resolved by agarose gel electrophoresis (GenAgaroseLE;GENAXXON Bioscience) and visualized after ethidium bromide staining. PCRproducts were purified using the NucleoSpin Gel and PCR Clean-Up Kit(Macherey-Nagel) according to the manufacturer’s instructions. Purified PCRproducts were sequenced by StarSEQ.
Molecular Analysis of uzu1.b Mutants ert-ii.79 and 093AR
Genomic DNA of two uzu1.b alleles, ert-ii.79 and 093AR, and of their re-spective parents, ‘Bonus’ and ‘Aramir,’ was isolated from green leaf materialusing the micro cetyl trimethylammonium bromide protocol described inGruszka et al. (2011b). For this analysis, the AFLP marker technique wasapplied. Two parental varieties were compared with each other. Similarly,AFLP band patterns were compared between the mutants and mutants alsowere reciprocally compared with the parental varieties. During the analysis,on average, more than 380 AFLP bands were scored for each comparison.
Mutagenesis and TILLING Screen of a BarleyM2 Generation
The doubled-haploid line H930-36 was developed from the F1 generation of across between barley ‘Klages’ and variety Mata. H930-36 was mutagenized bychemical treatment with N-methyl-N-nitrosourea at a dose of 1.5 mM/3 h. Then, 950individual plants of the M2 generation were grown in the field and phenotyped atmaturity. Leaf material of individual plants was harvested, dried in Silica Gel TypeIII (Sigma-Aldrich), and groundwith an electric grinder (Retsch). DNAwas extractedusing the micro cetyl trimethylammonium bromide protocol (Gruszka et al., 2011b)and genotypes were analyzed using the TILLING approach described in Kurowskaet al. (2012). Two fragments of the HvBRI1 spanning the gene region from 1,639 to2,994 bp (Supplemental Fig. S8B), were analyzed for sequence alterations.
Anatomical and Microscopic Analysis
For histological assays of 2-mm root segments of 5-d-old barley seedlings,conventional fixation, substitution, and embedding procedures were per-formed as described elsewhere (Marzec et al., 2013). Semithin sections(approximately 2 mm thick) were stained for 2 min with 1% (w/v) methyleneblue/1% (w/v) Azur II in 1% (w/v) aqueous borax at 60°C before light mi-croscopic examination with a Zeiss Axiovert 135 microscope. In order to an-alyze the root architecture, one seed per plastic tube (50-cm length and 40-mmdiameter) was sown in vermiculite. Roots of 18-d-old plants were analyzedusing a standard Epson scanner and WinRhizo software (Regent Instruments).
Sequence data from this article can be found in the GenBank/EMBL datalibraries under accession numbers KF318307 (HvDIM), KF318308 (HvBRD),and KF360233 (HvCPD).
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure S1. The brassinosteroid biosynthetic pathway asfound in Arabidopsis.
Supplemental Figure S2. Anatomical and microscopic analysis of seed-lings of cv Bowman near-isogenic lines BW031 (ari-u.245), BW084(brh13.p), BW333 (ert-zd.159), and BW312 (ert-ii.79).
Supplemental Figure S3. Leaf-inclination assay of seedlings of cv Bowmannear-isogenic lines BW031 (ari-u.245), BW084 (brh13.p), BW312 (ert-ii.79),and BW333 (ert-zd.159).
Supplemental Figure S4. Alignments of brassinosteroid-biosynthesis en-zymes from Arabidopsis and barley.
Supplemental Figure S5. In silico mapping and detailed analysis of con-served amino acid residues of HvBRD.
Plant Physiol. Vol. 166, 2014 1925
Variation in Barley Brassinosteroid Genes
www.plant.org on December 22, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

Supplemental Figure S6. In silico mapping and detailed analysis of con-served amino acid residues of HvCPD.
Supplemental Figure S7. In silico mapping and detailed analysis of con-served amino acid residues of HvDIM.
Supplemental Figure S8. In silico mapping and detailed analysis of con-served amino acid residues of HvBRI1.
Supplemental Data Set S1. Overview and description of analyzed barleymutant accessions and parental cultivars.
Supplemental Data Set S2. Examples of kinases with a cation-Pi His-[X57–59]-Phe/Tyr motif.
Supplemental Data Set S3. Primers used for amplification of the HvBRD,HvBRI1, HvCPD, HvDIM, and HvDWF4 genomic DNA sequences.
ACKNOWLEDGMENTS
We thank Lisbeth Faldborg, Christa Augsburg, Charlotte Riis, Ond�rejNovák, Ewelina Glodowska, and Ewa Wierus for technical assistance, aswell as Andy Kleinhofs, the Nordic Genetic Resource Center, and the NationalSmall Grains Collection for providing seeds.
Received September 19, 2014; accepted October 16, 2014; published October20, 2014.
LITERATURE CITED
Bai MY, Shang JX, Oh E, Fan M, Bai Y, Zentella R, Sun TP, Wang ZY(2012) Brassinosteroid, gibberellin and phytochrome impinge on acommon transcription module in Arabidopsis. Nat Cell Biol 14: 810–817
Bajguz A, Tretyn A (2003) The chemical characteristic and distribution ofbrassinosteroids in plants. Phytochemistry 62: 1027–1046
Bojar D, Martinez J, Santiago J, Rybin V, Bayliss R, Hothorn M (2014)Crystal structures of the phosphorylated BRI1 kinase domain and im-plications for brassinosteroid signal initiation. Plant J 78: 31–43
Bosshard HR, Marti DN, Jelesarov I (2004) Protein stabilization by saltbridges: concepts, experimental approaches and clarification of somemisunderstandings. J Mol Recognit 17: 1–16
Chandler PM, Harding CA (2013) ‘Overgrowth’ mutants in barley andwheat: new alleles and phenotypes of the ‘Green Revolution’ DELLAgene. J Exp Bot 64: 1603–1613
Chono M, Honda I, Zeniya H, Yoneyama K, Saisho D, Takeda K,Takatsuto S, Hoshino T, Watanabe Y (2003) A semidwarf phenotype ofbarley uzu results from a nucleotide substitution in the gene encoding aputative brassinosteroid receptor. Plant Physiol 133: 1209–1219
Dahleen LS, Vander Wal LJ, Franckowiak JD (2005) Characterization andmolecular mapping of genes determining semidwarfism in barley. JHered 96: 654–662
Daoura BG, Chen L, Du Y, Hu YG (2014) Genetic effects of dwarfing geneRht-5 on agronomic traits in common wheat (Triticum aestivum L.) andQTL analysis on its linked traits. Field Crops Res 156: 22–29
Druka A, Franckowiak J, Lundqvist U, Bonar N, Alexander J, Houston K,Radovic S, Shahinnia F, Vendramin V, Morgante M, et al (2011) Geneticdissection of barley morphology and development. Plant Physiol 155: 617–627
Franckowiak JD, Lundqvist U (2012) Descriptions of barley genetic stocksfor 2012. Barley Genet Newsl 42: 36–792
Fujioka S, Noguchi T, Takatsuto S, Yoshida S (1998) Activity of brassi-nosteroids in the dwarf rice lamina inclination bioassay. Phytochemistry49: 1841–1848
Greeve I, Hermans-Borgmeyer I, Brellinger C, Kasper D, Gomez-Isla T, BehlC, Levkau B, Nitsch RM (2000) The human DIMINUTO/DWARF1 homologseladin-1 confers resistance to Alzheimer’s disease-associated neuro-degeneration and oxidative stress. J Neurosci 20: 7345–7352
Gruszka D, Szarejko I, Maluszynski M (2011a) Identification of barley DWARFgene involved in brassinosteroid synthesis. Plant Growth Regul 65: 343–358
Gruszka D, Szarejko I, Maluszynski M (2011b) New allele of HvBRI1 geneencoding brassinosteroid receptor in barley. J Appl Genet 52: 257–268
Hartwig T, Corvalan C, Best NB, Budka JS, Zhu JY, Choe S, Schulz B(2012) Propiconazole is a specific and accessible brassinosteroid (BR)biosynthesis inhibitor for Arabidopsis and maize. PLoS ONE 7: e36625
Hay A, Hake S (2004) The dominant mutant Wavy auricle in blade1 disruptspatterning in a lateral domain of the maize leaf. Plant Physiol 135: 300–308
Hedden P (2003) The genes of the Green Revolution. Trends Genet 19: 5–9Hellewell KB, Rasmusson DC, Gallo-Meagher G (2000) Enhancing yield
in semidwarf barley. Crop Sci 40: 352–358Helliwell CA, Chandler PM, Poole A, Dennis ES, Peacock WJ (2001) The
CYP88A cytochrome P450, ent-kaurenoic acid oxidase, catalyzes three steps ofthe gibberellin biosynthesis pathway. Proc Natl Acad Sci USA 98: 2065–2070
Honda I, Zeniya H, Yoneyama K, Chono M, Kaneko S, Watanabe Y (2003)Uzu mutation in barley (Hordeum vulgare L.) reduces the leaf unrollingresponse to brassinolide. Biosci Biotechnol Biochem 67: 1194–1197
Hong Z, Jin H, Tzfira T, Li J (2008) Multiple mechanism-mediated reten-tion of a defective brassinosteroid receptor in the endoplasmic reticulumof Arabidopsis. Plant Cell 20: 3418–3429
Hong Z, Ueguchi-Tanaka M, Umemura K, Uozu S, Fujioka S, TakatsutoS, Yoshida S, Ashikari M, Kitano H, Matsuoka M (2003) A ricebrassinosteroid-deficient mutant, ebisu dwarf (d2), is caused by a loss offunction of a new member of cytochrome P450. Plant Cell 15: 2900–2910
Hothorn M, Belkhadir Y, Dreux M, Dabi T, Noel JP, Wilson IA, Chory J(2011) Structural basis of steroid hormone perception by the receptorkinase BRI1. Nature 474: 467–471
Houston K, Druka A, Bonar N, Macaulay M, Lundqvist U, Franckowiak J,Morgante M, Stein N, Waugh R (2012) Analysis of the barley bractsuppression gene Trd1. Theor Appl Genet 125: 33–45
Houston K, McKim SM, Comadran J, Bonar N, Druka I, Uzrek N, CirilloE, Guzy-Wrobelska J, Collins NC, Halpin C, et al (2013) Variation inthe interaction between alleles of HvAPETALA2 and microRNA172 de-termines the density of grains on the barley inflorescence. Proc NatlAcad Sci USA 110: 16675–16680
Janeczko A, Biesaga–Ko�scielniak J, Oklešťková J, Filek M, Dziurka M,Szarek-Łukaszewska G, Ko�scielniak J (2010) Role of 24-epibrassinolidein wheat production: physiological effects and uptake. J Agron Crop Sci196: 311–321
Jia Q, Zhang J, Westcott S, Zhang XQ, Bellgard M, Lance R, Li C (2009)GA-20 oxidase as a candidate for the semidwarf gene sdw1/denso inbarley. Funct Integr Genomics 9: 255–262
Kim BK, Fujioka S, Takatsuto S, Tsujimoto M, Choe S (2008) Castasteroneis a likely end product of brassinosteroid biosynthetic pathway in rice.Biochem Biophys Res Commun 374: 614–619
Kim TW, Wang ZY (2010) Brassinosteroid signal transduction from re-ceptor kinases to transcription factors. Annu Rev Plant Biol 61: 681–704
Kurowska M, Labocha-Pawłowska A, Gnizda D, Maluszynski M, Szarejko I(2012) Molecular analysis of point mutations in a barley genome exposed toMNU and gamma rays. Mutat Res 738-739: 52–70
Li J, Chory J (1997) A putative leucine-rich repeat receptor kinase involvedin brassinosteroid signal transduction. Cell 90: 929–938
Marzec M, Melzer M, Szarejko I (2013) Asymmetric growth of root epi-dermal cells is related to the differentiation of root hair cells in Hordeumvulgare (L.). J Exp Bot 64: 5145–5155
Mayer KF, Martis M, Hedley PE, Simková H, Liu H,Morris JA, Steuernagel B,Taudien S, Roessner S, Gundlach H, et al (2011) Unlocking the barleygenome by chromosomal and comparative genomics. Plant Cell 23: 1249–1263
Monna L, Kitazawa N, Yoshino R, Suzuki J, Masuda H, Maehara Y, Tanji M,Sato M, Nasu S, Minobe Y (2002) Positional cloning of rice semidwarfinggene, sd-1: rice “green revolution gene” encodes a mutant enzyme involvedin gibberellin synthesis. DNA Res 9: 11–17
Nagano H, Onishi K, Ogasawara M, Horiuchi Y, Sano Y (2005) Genealogyof the “Green Revolution” gene in rice. Genes Genet Syst 80: 351–356
National Climate Assessment Development Advisory Committee (2013)The U.S. Global Change Research Program http://ncadac.globalchange.gov/
Noguchi T, Fujioka S, Choe S, Takatsuto S, Yoshida S, Yuan H, FeldmannKA, Tax FE (1999) Brassinosteroid-insensitive dwarf mutants of Arabi-dopsis accumulate brassinosteroids. Plant Physiol 121: 743–752
Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, MengEC, Ferrin TE (2004) UCSF Chimera—a visualization system for ex-ploratory research and analysis. J Comput Chem 25: 1605–1612
Porter JR, Semenov MA (2005) Crop responses to climatic variation. PhilosTrans R Soc Lond B Biol Sci 360: 2021–2035
Prajapati RS, Sirajuddin M, Durani V, Sreeramulu S, Varadarajan R(2006) Contribution of cation-pi interactions to protein stability. Bio-chemistry 45: 15000–15010
1926 Plant Physiol. Vol. 166, 2014
Dockter et al.
www.plant.org on December 22, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

Pysh LD, Wysocka-Diller JW, Camilleri C, Bouchez D, Benfey PN (1999) TheGRAS gene family in Arabidopsis: sequence characterization and basic expres-sion analysis of the SCARECROW-LIKE genes. Plant J 18: 111–119
Rajkumara S (2008) Lodging in cereals. Agric Rev 29: 55–60Saisho D, Tanno K, Chono M, Honda I, Kitano H, Takeda K (2004)
Spontaneous brassinolide-insensitive barley mutants “uzu” adapted toEast Asia. Breed Sci 54: 409–416
Sakamoto T, Morinaka Y, Ohnishi T, Sunohara H, Fujioka S, Ueguchi-Tanaka M, Mizutani M, Sakata K, Takatsuto S, Yoshida S, et al (2006)Erect leaves caused by brassinosteroid deficiency increase biomassproduction and grain yield in rice. Nat Biotechnol 24: 105–109
Santiago J, Henzler C, Hothorn M (2013) Molecular mechanism for plantsteroid receptor activation by somatic embryogenesis co-receptor ki-nases. Science 341: 889–892
Sasaki A, Ashikari M, Ueguchi-Tanaka M, Itoh H, Nishimura A, Swapan D,Ishiyama K, Saito T, KobayashiM, Khush GS, et al (2002) Green revolution: amutant gibberellin-synthesis gene in rice. Nature 416: 701–702
She J, Han Z, Kim TW, Wang J, Cheng W, Chang J, Shi S, Wang J, YangM, Wang ZY, et al (2011) Structural insight into brassinosteroid per-ception by BRI1. Nature 474: 472–476
Shimada Y, Fujioka S, Miyauchi N, Kushiro M, Takatsuto S, Nomura T,Yokota T, Kamiya Y, Bishop GJ, Yoshida S (2001) Brassinosteroid-6-oxidases from Arabidopsis and tomato catalyze multiple C-6 oxidationsin brassinosteroid biosynthesis. Plant Physiol 126: 770–779
Spielmeyer W, Ellis MH, Chandler PM (2002) Semidwarf (sd-1), “greenrevolution” rice, contains a defective gibberellin 20-oxidase gene. ProcNatl Acad Sci USA 99: 9043–9048
Sun Y, Han Z, Tang J, Hu Z, Chai C, Zhou B, Chai J (2013) Structure re-veals that BAK1 as a co-receptor recognizes the BRI1-bound brassino-lide. Cell Res 23: 1326–1329
Swaczynová J, Novák O, Hauserová E, Funksová K, Šíša M, Strnad M(2007) New techniques for estimation of naturally occurring brassinos-teroids. J Plant Growth Regul 26: 1–14
The International Barley Genome Sequencing Consortium (2012) Aphysical, genetic and functional sequence assembly of the barley ge-nome. Nature 491: 711–716
Wada K, Kondo H, Marumo S (1985) A simple bioassay for brassinoste-roids: A wheat leaf-unrolling test. Agric Biol Chem 49: 2249–2251
Wendt T, Holm PB, Starker CG, Christian M, Voytas DF, Brinch-Pedersen H,Holme IB (2013) TAL effector nucleases induce mutations at a pre-selected lo-cation in the genome of primary barley transformants. PlantMol Biol 83: 279–285
Vriet C, Russinova E, Reuzeau C (2012) Boosting crop yields with plantsteroids. Plant Cell 24: 842–857
Zakhrabekova S, Gough SP, Braumann I, Müller AH, Lundqvist J, AhmannK, Dockter C, Matyszczak I, Kurowska M, Druka A, et al (2012) Inducedmutations in circadian clock regulator Mat-a facilitated short-season ad-aptation and range extension in cultivated barley. Proc Natl Acad Sci USA109: 4326–4331
Plant Physiol. Vol. 166, 2014 1927
Variation in Barley Brassinosteroid Genes
www.plant.org on December 22, 2014 - Published by www.plantphysiol.orgDownloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

1
Supplemental Data Induced Variations in Brassinosteroid Genes Define Barley Height and Sturdiness, and Expand the “Green Revolution” Genetic Toolkit Christoph Dockter, Damian Gruszka, Ilka Braumann, Arnis Druka, Ilze Druka, Jerome Franckowiak, Simon P. Gough, Anna Janeczko, Marzena Kurowska, Joakim Lundqvist, Udda Lundqvist, Marek Marzec, Izabela Matyszczak, André H. Müller, Jana Oklestkova, Burkhard Schulz, Shakhira Zakhrabekova, and Mats Hansson
The following materials are available in the online version of this article.
Supplemental Figure 1. The Brassinosteroid Biosynthetic Pathway as Found in Arabidopsis. Supplemental Figure 2. Anatomical and Microscopic Analysis of Seedlings of Bowman Near-Isogenic Lines BW031 (ari-u.245), BW084 (brh13.p), BW333 (ert-zd.159), and BW312 (ert-ii.79). Supplemental Figure 3. Leaf-Inclination Assay of Seedlings of Bowman Near-Isogenic Lines BW031 (ari-u.245), BW084 (brh13.p), BW312 (ert-ii.79), and BW333 (ert-zd.159). Supplemental Figure 4. Alignments of Brassinosteroid-Biosynthesis Enzymes from Arabidopsis and Barley. Supplemental Figure 5. In Silico Mapping and Detailed Analysis of Conserved Amino Acid Residues of Hv-BRD (Hordeum vulgare BRASSINOSTEROID-6-OXIDASE). Supplemental Figure 6. In Silico Mapping and Detailed Analysis of Conserved Amino Acid Residues of Hv-CPD (Hordeum vulgare CONSTITUTIVE PHOTOMORPHOGENIC DWARF). Supplemental Figure 7. In Silico Mapping and Detailed Analysis of Conserved Amino Acid Residues of Hv-DIM (Hordeum vulgare DIMINUTO). Supplemental Figure 8. In Silico Mapping and Detailed Analysis of Conserved Amino Acid Residues of Hv-BRI1 (Hordeum vulgare BRASSINOSTEROID-INSENSITIVE 1).
Supplemental Dataset 1. Dataset 1.1. Overview and Description of Analyzed Barley Mutant Accessions and Parental Cultivars. Dataset 1.2. Examples of Kinases with a Cation-Pi His-[X57-59]-Phe/Tyr Motif. Dataset 1.3. Primers Used for Amplification of the Hv-BRD, Hv-BRI1, Hv-CPD, Hv-DIM and Hv-DWF4 Genomic DNA Sequences.

2
SI Figures

3
Supplemental Figure 1. The Brassinosteroid Biosynthetic Pathway as Found in Arabidopsis
(Bishop and Yokota, 2001; Ohnishi et al., 2012).
The green section marks the general phytosterol pathway preceding the branch of brassinosteroid
biosynthesis. Eight enzymatic steps between squalene-2,3-epioxide and episterol are not shown
in detail. The C-22 oxidation pathway is highlighted in pink, the late C-6 oxidation pathway is
marked in yellow, and the early C-6 oxidation pathway is marked in blue. The symbols of genes
encoding enzymes catalyzing the different reaction steps are indicated next to the arrows. Several
enzymes have broad substrate specificity. Therefore, they occur at multiple steps in the pathway.
Dotted arrows signify more than one enzymatic reaction catalyzed by a multifunctional enzyme.
Arrows without gene symbol assigned to it signify unidentified enzymatic steps. The final
product of the pathway is brassinolide, which is generated by conversion from castasterone. The
enzyme catalyzing this reaction and the corresponding gene has not been found in cereal grasses,
like barley. In these plants castasterone seems to be the final product of the brassinosteroid
synthesis (Kim et al., 2008). STE1, Δ7-sterol-C5-desaturase; DWF5, Δ5,7-sterol-Δ7-reductase;
DIM/DWF1, Δ5-sterol-Δ24-reductase; DWF4, C-22 hydroxylase; DET2, 5α reductase; CPD, C-
23α-hydroxylase/C-3 dehydrogenase; ROT3 and CYP90D1, C-23 hydroxylases; BR6ox1,
brassinosteroid-6-oxidase 1; BR6ox2, brassinosteroid-6-oxidase 2.

4

5

6
Supplemental Figure 2. Anatomical and Microscopic Analysis of Seedlings of Bowman Near-
Isogenic Lines BW031 (ari-u.245), BW084 (brh13.p), BW333 (ert-zd.159), and BW312 (ert-
ii.79).
(A) Phenotypes of 5-day-old seedlings grown in light. All mutant lines show significant
shortening of the leaf length but no difference in the length of seminal roots when compared with
Bowman. Scale bar: 2 cm.
(B) Brassinosteroid deficiency-dependent length reduction is observed only in leaves, whereas
no significant differences in the length of the longest seminal root, total length of seminal roots,
or the number of seminal roots were identified between the analyzed genotypes. Slightly reduced
total length of seminal roots in brh13.p is caused by reduced number of seminal roots.
(C) Root stele of the longest seminal root on the cross sections through the mature root-hair
zone. Number of cell layers and number of cells in each layer were very similar between the

7
analyzed genotypes. However, increased size of the outer metaxylem vessels (red arrows) was
observed in the brassinosteroid-mutant lines compared to Bowman. Scale bar: 50 µm.
(D) Longitudinal sections through the mature root-hair zone of the longest seminal roots. No
significant difference was observed in the average length of parenchyma cells between the
analyzed genotypes (250 cells from 5 roots were measured for each genotype). Scale bar: 100
µm.
(E) Density of lateral roots developed on the longest seminal root of 18-day-old seedlings. A
lower density of lateral roots was observed in all mutant lines in comparison with Bowman.
Scale bar: 5 cm.

8
Supplemental Figure 3. Leaf Lamina Inclination Assay of Barley Wild Type and Mutant Plants.
One µl of 2 mg/ml brassinolide (24-epi-brassinolide) solved in 96% ethanol was added to the tip
of barley seedlings. Control plants were treated with one µl 96% ethanol. The photos were taken
4 days later when the second leaf emerged. Plants sensitive to brassinolide show a pronounced
inclination between the lamina and leaf sheath. The method was developed from (Fujioka et al.,
1998; Hong et al., 2003). (A) Barley cultivar Bowman, (B) near-isogenic line BW031 (ari-
u.245), (C) BW084 (brh13.p), (D) BW312 (ert-ii.79), and (E) BW333 (ert-zd.159). The left
plant in each photo was treated with brassinosteroid, whereas the right plant was only exposed to
ethanol. BW312 (ert-ii.79) is insensitive to brassinolide due to its mutation in the brassinosteroid
receptor gene Hv-BRI1.

9
A AT3G02580 At-STE1 1 -------MAADNAYLMQFVDETSFYNRIVLSHLLPA-—NLWEPLPHFLQTWLRNYLAGTLLYFISGFLWCFYIYYLKINVYLPKDAIPTI 81 AK372389 Hv-STE1 1 MAAAAAAAAAGEDYWGLFREETDWYNEIFLSAVVPGGGGWWRALPHPLRSWLRNCIGGYLLYFATGFLWCFVIYYWKRNAYIPKDAVPTV 90 ** : * * :**.:**.*.** ::*. *. *** *::**** :.* **** :****** *** * *.*:****:**: AT3G02580 At-STE1 82 KAMRLQMFVAMKAMPWYTLLPTVSESMIERGWTKCFASIGEFGWILYFVYIAIYLVFVEFGIYWMHRELHDIKPLYKYLHATHHIYNKQN 171 AK372389 Hv-STE1 91 EAMKKQIIVASKAMPFYCALPSVSEHMIESGWTRCFFHISEVGWPMYFVYVALYLTFVEFGIYWMHRELHDIKPLYKHLHATHHIYNKEN 180 :**: *::** ****:* **:*** *** ***:** *.*.** :****:*:**.*********************:**********:*
AT3G02580 At-STE1 172 TLSPFAGLAFHPVDGILQAVPHVIALFIVPIHFTTHIGLLFMEAIWTANIHDCIHGNIWPVMGAGYHTIHHTTYKHNYGHYTIWMDWMFG 261 AK372389 Hv-STE1 181 TLSPFAGLAFHPLDGILQAISHVIALFILPVHFRTHVALIFIEAVWTANIHDCIHGKVWPVMGAGYHTIHHTTYRHNYGHYTVWMDWLFG 270 ************:******: *******:*:** **:.*:*:**:***********::****************:*******:****:** AT3G02580 At-STE1 262 SLRDPLLEEDDNKDSFKKAE 281 AK372389 Hv-STE1 271 TLREPEDLLKKD-------- 282 :**:* ..:
B
AT1G50430 At-DWF5 1 ------------------MAETVHSPIVTYASMLSLLAFCPPFVILLWYTMVHQDGSVTQTFGFFWENGV-QGLINIWPRPTLIAWKIIF 71 MLOC_69179 Hv-DWF5 1 MGKQQQQPKPAPAAPPPQPSKTAHSPLLTYASMVTLLSLCPPFVILLWYTMVHADGSVLQIYEHLRDHGILEGLKAIWPAPTLVAGKIIL 90 ::*.***::*****::**::************** **** * : .: :.*: :** *** ***:* ***: AT1G50430 At-DWF5 72 CYGAFEAILQLLLPGKRVEGPISPAGNRPVYKANGLAAYFVTLATYLGLWWFGIFNPAIVYDHLGEIFSALIFGSFIFCVLLYIKGHVAP 161 MLOC_69179 Hv-DWF5 91 GFALFEAALQLLLPGKRFEGPISPSGNVPVYKANGLLAYAVTLLTYLSLWWFGIFNPAIVYDHLGEIYSALVFGSLVFCLLLYIKGHVAP 180 :. *** *********.******:** ******** ** *** ***.*******************:***:***::**:********** AT1G50430 At-DWF5 162 SSSDSGSCGNLIIDFYWGMELYPRIGKSFDIKVFTNCRFGMMSWAVLAVTYCIKQYEINGKVSDSMLVNTILMLVYVTKFFWWEAGYWNT 251 MLOC_69179 Hv-DWF5 181 SSSDSGSAGNVIIDFYWGMELYPRIGKHFDIKVFTNCRFGMMSWAVLAVTYCIKQYEMNGRVADSMLVNTALMLIYITKFFWWESGYWCT 270 *******.**:**************** *****************************:**:*:******* ***:*:*******:*** * AT1G50430 At-DWF5 252 MDIAHDRAGFYICWGCLVWVPSVYTSPGMYLVNHPVELGTQLAIYILVAGILCIYINYDCDRQRQEFRRTNGKCLVWGRAPSKIVASYTT 341 MLOC_69179 Hv-DWF5 271 MDIAHDRAGFYICWGCLVWVPSIYTSPGMYLVNHPVNLGPQLALSILLAGILCIYINYDCDRQRQEFRRTNGKATVWGKAPSKIIASYQT 360 **********************:*************:** ***: **:*************************. ***:*****:*** * AT1G50430 At-DWF5 342 TSGETKTSLLLTSGWWGLARHFHYVPEILSAFFWTVPALFDNFLAYFYVIFLTLLLFDRAKRDDDRCRSKYGKYWKLYCEKVKYRIIPGIY 432 MLOC_69179 Hv-DWF5 361 TKGETKTSLLLTSGWWGFSRHFHYAPEILAAFFWTVPALFTHFMPYFYVIFLTILLLDRAKRDDDRCSTKYGKYWKMYCNRVPYRVVPGIY 451 *.***************::*****.****:********** .*: ********:**:********** :*******:**::* **::****
C
AT3G19820 At-DIM 1 MSDLQTPLVRPKRKKTWVDYFVKFRWIIVIFIVLPFSATFYFLIYLGDMWSESKSFEKRQKEHDENVKKVIKRLKGRDASKDGLVCTARK 90 MLOC_52405 Hv-DIM 1 MADLQTPLVRPKRKKVLVDYLVQFRWILVIFVVLPASALIYFNIYLGDMWSAMKSEKKRQKEHDDNVQKVVKRLKQRNPKKDGLVCTARK 90 *:*************. ***:*:****:***:*** ** :** ******** ** :*******:**:**:**** *: .********** AT3G19820 At-DIM 91 PWIAVGMRNVDYKRARHFEVDLGEFRNILEINKEKMTARVEPLVNMGQISRATVPMNLSLAVVAELDDLTVGGLINGYGIEGSSHIYGLF 180 MLOC_52405 Hv-DIM 91 PWIAVGMRNVDYKRVRHFEVDLSAFRNILEIDAERMVAKVEPLVNMGQISRATCPMNLSLAVVAELDDLTVGGLINGYGIEGSSHIYGLF 180 **************.*******. *******: *:*.*:************** ************************************ AT3G19820 At-DIM 181 ADTVEAYEIVLAGGELVRATRDNEYSDLYYAIPWSQGTLGLLVAAEIRLIKVKEYMRLTYIPVKGDLQALAQGYIDSFAPKDGDKSKIPD 270 MLOC_52405 Hv-DIM 181 SDTVVALEIVLADGRIVRATKDNEYSDLFYGVPWSQGTLGFLVSAEIKLIPIKEYMRLTYTPVKGPLKELAQAYADAVAPRDGDPAKVPD 270 :*** * ***** *.:****:*******:*.:********:**:***:** :******** **** *: ***.* *:.**:*** :*:** AT3G19820 At-DIM 271 FVEGMVYNPTEGVMMVGTYASKEEAKKKGNKINNVGWWFKPWFYQHAQTALKKGQFVEYIPTREYYHRHTRCLYWEGKLILPFGDQFWFR 360 MLOC_52405 Hv-DIM 271 FVEGMVYNATEGVMMTGVYASKEEAKKKGNKINSVGWWFKPWFYQHAQTALKKGEFVEYIPTREYYHRHTRCLYWEGKLILPFGDQFWFR 360 ******** ******.*.***************.********************:*********************************** AT3G19820 At-DIM 361 YLLGWLMPPKVSLLKATQGEAIRNYYHDMHVIQDMLVPLYKVGDALEWVHREMEVYPIWLCPHKLFKQPIKGQIYPEPGFEYENRQGDTE 450 MLOC_52405 Hv-DIM 361 FLFGWLMPPKVSLLKATQGDAIRNYYHDNHVIQDMLVPLYKVGDALEFVHQEMEVYPLWLCPHRLFKLPVKTMIYPEPGFEHHQRQGDTS 450 :*:****************:******** ******************:**:******:*****:*** *:* ********:.:*****. AT3G19820 At-DIM 451 DAQMYTDVGVYYAPGCVLRGEEFDGSEAVRRMEKWLIENHGFQPQYAVSELDEKSFWRMFNGELYEECRKKYRAIGTFMSVYYKSKKGRK 540 MLOC_52405 Hv-DIM 451 YAQMFTDVGVYYTPAFIFRGEQFDGVEAVKRLEQWLIENHSYQPQYAVTELNEKDFWRMFDASHYEHCRQKYGAVGTFMSVYYKSKKGRK 540 ***:*******:*. ::***:*** ***:*:*:******.:******:**:**.*****:.. **.**:** *:*************** AT3G19820 At-DIM 541 TEKEVREAEQAHLETAYAEAD 561 MLOC_52405 Hv-DIM 541 SEKEVQEAEAAILEPAYADEA 561 :****:*** * ** ***:
D
AT2G38050 At-DET2 1 --------MEEIADKTFFRYCLLTLIFAGPPTAVLLKFLQAPYGKHNRTGWGPTVSPPIAWFVMESPTLWLTLLLFPFGRHA---LNPKSL 80 AK357682 Hv-DET2 1 MPDQTAAAAGAGGDGALFSRCLLALYIISPVTVFLLRYVSAPYGKLSRPGWGPAIPAALAWCAMESPTLWLPPLVAPLPLLLAAAASPLAA 91 .* ::* ***:* : .* *..**:::.***** .* ****:: :** .******** *: *: .* : AT2G38050 At-DET2 81 LLFSPYLIHYFHRTIIYPLRLFRSSFPAGKNGFPITIAALAFTFNLLNGYIQAR-WVSHYKDDYEDGNWFWWRFVIGMVVFITGMYINITS 170 AK357682 Hv-DET2 92 LPPALYALHYVNRTVLHPLRILRLRR--APAPVPVLVAACAFGFNLLNAYVQARSWALHAARPAPAPALALARCLAGLALFAWGMRVNVAA 180 * : * :**..**:::***::* . .*: :** ** *****.*:*** *. * * : *:.:* ** :*::: AT2G38050 At-DET2 171 DRTLVRLKKENRGGYVIPRGGWFELVSCPNYFGEAIEWLGWAVMTWSWAGIGFFLYTCSNLFPRARASHKWYIAKFKEEYPKTRKAVIPFVY 262 AK357682 Hv-DET2 181 DRELLRLK-EAGGGYKIPRGGWFDAVTCPNYFGEIVEWLGYCLVAWSPAAWAFFLYTCANLMPRARDHRLWYLNKFGGEYPASRRAVIPYIY 271 ** *:*** * *** *******: *:******* :****:.:::** *. .******:**:**** : **: ** *** :*:****::*

10
E AT3G50660 At-DWF4 1 ---MF---ETEHHTLLPLLLLPSLL------SLLLFLILLKRRNRKTRFNLPPGKSGWPFLGETIGYLKPYTATTLGDFMQQHVSKYGKI 78 AK355174 Hv-DWF4 1 MAAMMASITSELLFFLPFILLALLTFYTSSVAKCHGLHRWSGRTKKKRPNLPPGAAGWPFVGETFGYLRAHPATSIGQFMNQHIARYGKI 90 *: :* :**::** * : * . *.:*.* ***** :****:***:***: : **::*:**:**:::**** AT3G50660 At-DWF4 79 YRSNLFGEPTIVSADAGLNRFILQNEGRLFECSYPRSIGGILGKWSMLVLVGDMHRDMRSISLNFLSHARLRTILLKDVERHTLFVLDSW 168 AK355174 Hv-DWF4 91 YRSSLFGERTVVSADAGLNRYILQNEGRLFECSYPRSIGGILGKWSMLVLVGDPHREMRSISLNFLSSLRLRAVLLPEVERHTLLVLRGW 180 ***.**** *:*********:******************************** **:********** ***::** :******:** .* AT3G50660 At-DWF4 169 QQN---SIFSAQDEAKKFTFNLMAKHIMSMDPGEEETEQLKKEYVTFMKGVVSAPLNLPGTAYHKALQSRATILKFIERKMEERKLDIKE 255 AK355174 Hv-DWF4 181 LPSSSSAVFSAQHEAKKFTFNLMAKNIMSMDPGEEETERLRLEYITFMKGVVSAPLNFPGTAYWKALKSRATILGVIERKMEDRLEKMNK 270 . ::****.************.************:*: **:************:***** ***:****** .******:* .::: AT3G50660 At-DWF4 256 EDQEEEEVKTEDEAEMSKSDHVRKQRTDDDLLGWVLKHSNLSTEQILDLILSLLFAGHETSSVAIALAIFFLQACPKAVEELREEHLEIA 345 AK355174 Hv-DWF4 270 E---------------------ASSMEEDDLLGWAMKQSNLSKEQILDLLLSLLFAGHETSSMALALAIFFLEGCPKAVEELREEHLEIA 339 * .. :******.:*:****.******:************:*:*******:.**************** AT3G50660 At-DWF4 346 RAKKELGESELNWDDYKKMDFTQCVINETLRLGNVVRFLHRKALKDVRYKGYDIPSGWKVLPVISAVHLDNSRYDQPNLFNPWRWQQQNN 435 AK355174 Hv-DWF4 340 RRQKLRGECKLSWEDYKEMVFTQCVINETLRLGNVVRFLHRKVIRDVHYNGYDIPSGWKILPVLAAVHLDSSLYEDPNSFNPWRWKGNAS 429 * :* **.:*.*:***:* **********************.::**:*:*********:***::*****.* *::** ******: : . AT3G50660 At-DWF4 436 GASSSGSGSFSTWGNNYMPFGGGPRLCAGSELAKLEMAVFIHHLVLKFNWELAEDDKPFAFPFVDFPNGLPIRVSRIL------- 513 AK355174 Hv-DWF4 430 GVA---------QNSNFMPYGGGTRLCAGSELAKLEMAIFLHHLVLNFRWELAEPDQAFVYPFVDFPKGLPIRVHRIAQEKGGDE 505 *.: .*:**:*** **************:*:*****:*.***** *: *.:******:****** **
F
LOC_Os04g39430 Os-DWF11 1 --MVGGELVLAALVILLALLLTLVLSHFLPLLLNPKAPKGSFGWPLLGETLRFLSPHASNTLGSFLEDHCSRYGRVFKSHLFCTPTIVSC 88 AK371371 Hv-DWF11 1 MSMTVGDLVLAAPAILLALLLALVLSHFLPLLLNPKAPRGSFGWPLVGETLRFLTPHASNTLGSFLEDHCSRYGRVFKSHLFCTPTVVSC 90 *. *:***** .*******:****************:*******:*******:*******************************:*** LOC_Os04g39430 Os-DWF11 89 DQELNHFILQNEERLFQCSYPRPIHGILGKSSMLVVLGEDHKRLRNLALALVTSTKLKPSYLGDIEKIALHIVGSWHGKSKDKGMVNVIA 178 AK371371 Hv-DWF11 91 DQELNHFILQNEERLFQCSYPRPIHGILGKSSMLVVLGEDHKRLRNLALALVTSTKLKPSYLGDIERIALHIVGSWHGKGS------NIT 174 ******************************************************************:************.. *: LOC_Os04g39430 Os-DWF11 179 FCEEARKFAFSVIVKQVLGLSPEEPVTAMILEDFLAFMKGLISFPLYIPGTPYAKAVQARARISSTVKGIIEERRNAGSSNKGDFLDVLL 268 AK371371 Hv-DWF11 175 FCEEARKFAFSVIVKQVLGLSPEEPVTAMILEDFLTFMKGLISFPLRIPGTPYAKAVQARERISSTVKGIIEERRKADCCKRDDFLNVLL 264 ***********************************:********** ************* **************:* ..:: ***:*** LOC_Os04g39430 Os-DWF11 269 SSNELSDEEKVSFVLDSLLGGYETTSLLISMVVYFLGQSAQDLELVKREHEGIRSKKEKDEFLSSEDYKKMEYTQHVINEALRCGNIVKF 358 AK371371 Hv-DWF11 265 STDELSDEEKVSFVLDSLLGGYETTSLMISMVVYFLGQSAQDLDLVKREHQGIRSTKAKEECLSSEDYKKMEYTQHVINEALRCGNIVKF 354 *::************************:***************:******:****.* *:* **************************** LOC_Os04g39430 Os-DWF11 359 VHRKALKDVRYKEYLIPSGWKVLPVFSAVHLNPLLHGNAQQFQPCRWEGASQGTSKKFTPFGGGPRLCPGSELAKVEAAFFLHHLVLNYR 448 AK371371 Hv-DWF11 355 VHRKALKDVRYKEYLIPSGWKVLPVFSAVHLNPSLHGNAQQFQPCRWEGPSQGTSKKFTPFGGGTRLCPGSELAKVEAAFFLHHLVLNFR 444 ********************************* *************** ************** ***********************:* LOC_Os04g39430 Os-DWF11 449 WRIDGDDIPMAYPYVEFQRGLPIEIEPLCSES 480 AK371371 Hv-DWF11 445 WKIDGDDIPMAYPYVEFPRGLPIEIEPICSES 476 *:*************** *********:****
G
At4G36380 At-CYP90C1/ROT3 1 MQPPASAGLFRSPENLPWPYNYMDYLVAGFLVLTAGILLRPWLWLRLRNSKTKDGDEEEDNEEKKKGMIPNGSLGWPVIGETLNFIACGY 90 MLOC_62829 1 -------------MSVSWPAPSTWA-AAG------AFLAAAWLLC--FRLLPAAARPRRRLATKTGARLPPGSFGWPVVGETLDFVSCAY 68 .: ** .** .:* ** . . .. *. . :* **:****:****:*::*.* At4G36380 At-CYP90C1/ROT3 91 SSRPVTFMDKRKSLYGK-VFKTNIIGTPIIISTDAEVNKVVLQNHGNTFVPAYPKSITELLGENSILSINGPHQKRLHTLIGAFLRSPHL 179 MLOC_62829 67 SPRPEAFVEKRRLRYGSAVFRSHLFGSATVVTADAEVSRCVLQSDARSFVPWYPRSLTELMGKSSILLINGSLQRRVHGLVGAFFKSPQL 158 * ** :*::**: **. **::.::*: ::::****.: ***....:*** **:*:***:*:.*** *** *:*:* *:***::**:* At4G36380 At-CYP90C1/ROT3 180 KDRITRDIEASVVLTLASWAQLP---LVHVQDEIKKMTFEILVKVLMSTSPGEDMNILKLEFEEFIKGLICIPIKFPGTRLYKSLKAKER 266 MLOC_62829 159 KAQVTADMQRRLAPALAAWREQGPGARLRIQDHAKTIVFQILVRGLIGLEAGPEMQQLKQQFQEFIVGLMSLPIKLPGTRLYRSLQAKKR 248 * ::* *:: :. :**:* : :::**. *.:.*:***: *:. . * :*: ** :*:*** **:.:***:******:**:**:* At4G36380 At-CYP90C1/ROT3 267 LIKMVKKVVEER---QVAMTTTSPANDVVDVLLRDGGDSEKQSQPSDFVSGKIVEMMIPGEETMPTAMTLAVKFLSDNPVALAKLVEENM 353 MLOC_62829 249 MARVIQRIIQDKRRRRRALDEAPPARDAIDVLMGDGSEEL----TDELISDNMIDLMIPAEDSVPVLITLAVKFLSECPLALQQLEEENM 334 : :::::::::: : *: : **.*.:***: **.:. .:::* :::::***.*:::*. :********: *:** :* **** At4G36380 At-CYP90C1/ROT3 354 EMKRRKLELGEEYKWTDYMSLSFTQNVINETLRMANIINGVWRKALKDVEIKGYLIPKGWCVLASFISVHMDEDIYDNPYQFDPWRWDRI 443 MLOC_62829 335 QLKMRKAGLGETLQWTDYMSLSFTQHVITETLRMGNIISGIMRKAVRDVEVKGHLIPKGWCVFMYFRSVHLDDMLYEDPCKFNPWRWKEK 424 ::* ** *** :***********.**.*****.***.*: ***::***:**:********: * ***:*: :*::* :*:****.. At4G36380 At-CYP90C1/ROT3 444 NGSANSSICFTPFGGGQRLCPGLELSKLEISIFLHHLVTRYSWTAEEDEIVSFPTVKMKRRLPIRVATVDDSASPISLEDH 524 MLOC_62829 425 DVS---ASSFTPFGGGQRLCPGLDLARLEASIFLHHLVTSFRWVAEEDHIVNFPTVRLKGGMPIRVTARD----------- 491 : * : .**************:*::** ********* : *.****.**.****::* :****:: *

11
H AT3G13730 At-CYP90D1 1 ----------MDTSSSLLFFSFFFFIIIVIFNKINGLRSSPASKKKLNDHHVTSQSHGPKFPHGSLGWPVIGETIEFVSSAYSDRPESFM 80 MLOC_62829 Hv-CYP90D1 1 MSVSWPAPSTWAAAGAFLAAA--WLL---CF------RLLPAAA---RPRRRLATKTGARLPPGSFGWPVVGETLDFVSCAYSPRPEAFV 76 ::.::* : ::: * * **: . :: : . * ::* **:****:***::***.*** ***:*: AT3G13730 At-CYP90D1 81 DKRRLMYGR-VFKSHIFGTATIVSTDAEVNRAVLQSDSTAFVPFYPKTVRELMGKSSILLINGSLHRRFHGLVGSFLKSPLLKAQIVRDM 169 MLOC_62829 Hv-CYP90D1 77 EKRRLRYGSAVFRSHLFGSATVVTADAEVSRCVLQSDARSFVPWYPRSLTELMGKSSILLINGSLQRRVHGLVGAFFKSPQLKAQVTADM 166 :**** ** **:**:**:**:*::****.*.*****: :***:**::: ***************:**.*****:*:*** ****:. ** AT3G13730 At-CYP90D1 170 HKFLSESMDLWSEDQPV---LLQDVSKTVAFKVLAKALISVEKGEDLEELKREFENFISGLMSLPINFPGTQLHRSLQAKKNMVKQVERI 256 MLOC_62829 Hv-CYP90D1 167 QRRLAPALAAWREQGPGARLRIQDHAKTIVFQILVRGLIGLEAGPEMQQLKQQFQEFIVGLMSLPIKLPGTRLYRSLQAKKRMARVIQRI 256 :: *: :: * *: * :** :**:.*::*.:.**.:* * ::::**::*::** *******::***:*:*******.*.: ::** AT3G13730 At-CYP90D1 257 IEGKIRKTKNKEEDDVIAKDVVDVLLKDSSEHLTHNLIANNMIDMMIPGHDSVPVLITLAVKFLSDSPAALNLLTEENMKLKSLKELTGE 346 MLOC_62829 Hv-CYP90D1 257 IQDKRRRRRAL-DEAPPARDAIDVLMGDGSEELTDELISDNMIDLMIPAEDSVPVLITLAVKFLSECPLALQQLEEENMQLKMRKAGLGE 345 *: * *: : :: *:*.:***: *.**.**.:**::****:***..***************:.* **: * ****:** * ** AT3G13730 At-CYP90D1 347 PLYWNDYLSLPFTQKVITETLRMGNVIIGVMRKAMKDVEIKGYVIPKGWCFLAYLRSVHLDKLYYESPYKFNPWRWQERDMNTSSFSPFG 436 MLOC_62829 Hv-CYP90D1 346 TLQWTDYMSLSFTQHVITETLRMGNIISGIMRKAVRDVEVKGHLIPKGWCVFMYFRSVHLDDMLYEDPCKFNPWRWKEKDVSASSFTPFG 435 * *.**:** ***:**********:* *:****::***:**::******.: *:******.: **.* *******:*:*:.:***:*** AT3G13730 At-CYP90D1 437 GGQRLCPGLDLARLETSVFLHHLVTRFRWIAEEDTIINFPTVHMKNKLPIWIKRI- 491 MLOC_62829 Hv-CYP90D1 436 GGQRLCPGLDLARLEASIFLHHLVTSFRWVAEEDHIVNFPTVRLKGGMPIRVTARD 491 ***************:*:******* ***:**** *:*****::* :** :.
I
AT5G05690 At-CYP90A1/CPD 1 -------MAFTAFLLLLSSIAAGFLL--LLRRTRYRRMGLPPGSLGLPLIGETFQLIGAYKTENPEPFIDERVARYGSVFMTHLFGEPTI 81 MLOC_10658 Hv-CPD 1 MDAGALQLAATAVAIVAVALVCRWFLVACAARGRKQRPRLPPGSTGLPLIGETLRLISAYKTPDPEPFIDERVARHGGVFTTHIFGERTV 90 :* **. :: ::.. ::* * * :* ***** ********::**.**** :***********:*.** **:*** *: AT5G05690 At-CYP90A1/CPD 82 FSADPETNRFVLQNEGKLFECSYPASICNLLGKHSLLLMKGSLHKRMHSLTMSFANSSIIKDHLMLDIDRLVRFNLDSWSS--RVLLMEE 169 MLOC_10658 Hv-CPD 91 FSADPAFNRLLLAAEGRAVSCSYPSSITTLLGARSLLLTRGTAHKRLHSLTLTRLGRP-ASQPLLAHIDRLVLATMRQWEPTATVRLLDE 179 ***** **::* **: ..****:** .*** :**** :*: ***:****:: .: *: .***** .: .*. * *::* AT5G05690 At-CYP90A1/CPD 170 AKKITFELTVKQLMSFDPGEWSESLRKEYLLVIEGFFSLPLPLFS----TTYRKAIQARRKVAEALTVVVMKRREE---------EEEGA 246 MLOC_10658 Hv-CPD 180 AKKITFNLTVKQLVSIEPGPWTESLRCEYVKLIDGFFSIPFPFASFLPFTTYGQALKSRKKVAGALREVIRRRMEERGEENGVKEEMDGK 269 ******:******:*::** *:**** **: :*:****:*:*: * *** :*:::*:*** ** *: :* ** * :* AT5G05690 At-CYP90A1/CPD 247 ERKKDMLAALLAADD-GFSDEEIVDFLVALLVAGYETTSTIMTLAVKFLTETPLALAQLKEEHEKIRAMKSDSYSLEWSDYKSMPFTQCV 335 MLOC_10658 Hv-CPD 270 REKKDMVEELLEADGGSFSEEEMVDFCLSLLVAGYETTSVLMTVAVKFLTETPAALAQLKEEHENMTKMKGENQPLEWSDYKSMNFTQCV 359 ..****: ** ** .**:**:*** ::**********.:**:********* **********:: **.:. ********* ***** AT5G05690 At-CYP90A1/CPD 336 VNETLRVANIIGGVFRRAMTDVEIKGYKIPKGWKVFSSFRAVHLDPNHFKDARTFNPWRWQSNSVTT--GPSNVFTPFGGGPRLCPGYEL 423 MLOC_10658 Hv-CPD 360 INETLRVANIISGVFRRANTDIHFKGYTIPKGCKIFASFRAVHLNNEHYENARTFDPWRWQSNNKLQNEVGANLFTPFGGGPRLCPGYEL 449 :**********.****** **:.:***.**** *:*:*******: :*:::****:*******. :*:**************** AT5G05690 At-CYP90A1/CPD 424 ARVALSVFLHRLVTGFSWVPAEQDKLVFFPTTRTQKRYPIFVKRRDFAT 472 MLOC_10658 Hv-CPD 450 ARVVISVFLHHLVMRFSWEAAEEDRLLFFPTTRTLKGYPINLTRRSKSV 498 ***.:*****:** *** **:*:*:******* * *** :.**. :.
J
AT5G38970 At-BR6ox1/CYP85A1 1 MGAMMVMMGLL---LIIVSLCSALLRWNQMRYTK-------NGLPPGTMGWPIFGETTEFLKQGPNFMRNQRLRYGSFFKSHLLGCPTLI 80 AT3G30180 At-BR6ox2/CYP85A2 1 MGIMMMILGLL---VIIVCLCTALLRWNQMRYSK-------KGLPPGTMGWPIFGETTEFLKQGPDFMKNQRLRYGSFFKSHILGCPTIV 80 AK372445 Hv-BRD 1 MAL--LLLALVVVVVGVVVASSLLLRWNEVRYGNGRRKHGDACLPPGTMGWPLFGETTEFLKQGPSFVKQRRLRYGRLFRTHILGCPTVV 88 MLOC_75134 Hv-DWARF 1 -MA--FLLVLVGVVVGVVLASSLLLRWNEVRYGNGRRKHGDGCLPPGTMGWPLFGETTEFLKQGPSFMKQRRLRYGRLFRTHILGCPTVV 87 .:: *: : :* .: *****::** : *********:************.*::::***** :*::*:*****:: AT5G38970 At-BR6ox1/CYP85A1 81 SMDSEVNRYI-LKNESKGLVPGYPQSMLDILGTCNMAAVHGSSHRLMRGSLLSLISSTMMRDHILPKVDHFMRSYLDQWNELEVIDIQDK 169 AT3G30180 At-BR6ox2/CYP85A2 81 SMDAELNRYI-LMNESKGLVAGYPQSMLDILGTCNIAAVHGPSHRLMRGSLLSLISPTMMKDHLLPKIDDFMRNYLCGWDDLETVDIQEK 169 AK372445 Hv-BRD 89 CMDPELNRRMLLQGEAGGMVPGYPQSMLDILGRNNIAAVHGPLHRVMRGAMLGLVRPAALRQSLLPKMDAFMRHHLAGWAG-TVVDVQAK 177 MLOC_75134 Hv-DWARF 88 CMDPELNRRMLLL--QGGLVPGYPQSMLDILGRNNIAAVHGPLHRVMRGAMLGLVCPTMLRQSLLPKIDAFMRDHLHGWAG-NIVDVQAK 174 .** *:** : * *:* *********** *:***** **:***::*.*: : ::: :***:* *** :* * :*:* * AT5G38970 At-BR6ox1/CYP85A1 170 TKHMAFLSSLTQIAGNLRKPFVEEFKTAFFKLVVGTLSVPIDLPGTNYRCGIQARNNIDRLLRELMQERRDSGETFTDMLGYLMKK--EG 257 AT3G30180 At-BR6ox2/CYP85A2 170 TKHMAFLSSLLQIAETLKKPEVEEYRTEFFKLVVGTLSVPIDIPGTNYRSGVQARNNIDRLLTELMQERKESGETFTDMLGYLMKK--ED 257 AK372445 Hv-BRD 178 TKEMALLSALRQIAGITAGPLSDALQTELYTLVLGTISLPINLPGTSYYQGFQARTKLVSMLEQMIAERRSSGEAHDDMLDALLRSGDDG 267 MLOC_75134 Hv-DWARF 175 TKEMALLSALRQIAGITAGPLSDALKTELYTLVLGTISLPINLPGTSYHQGFQARTKLVSMLEQMIAQRRSSAQAHDDMLDALLRSGDDG 264 **.**:**:* *** * : :* ::.**:**:*:**::***.* *.***.:: :* ::: :*:.*.::. *** *::. : AT5G38970 At-BR6ox1/CYP85A1 258 NRYPLTDEEIRDQVVTILYSGYETVSTTSMMALKYLHDHPKALQELRAEHLAFRERKRQDEPLGLEDVKSMKFTRAVIYETSRLATIVNG 347 AT3G30180 At-BR6ox2/CYP85A2 258 NRYLLTDKEIRDQVVTILYSGYETVSTTSMMALKYLHDHPKALEELRREHLAIRERKRPDEPLTLDDIKSMKFTRAVIFETSRLATIVNG 347 AK372445 Hv-BRD 268 TREKISDEQIIDLLIALIYSGYETMSTTSMMAVKYLSEHPRALQELRREHLDIRKGKLPEEAIGYDDFKSMSFTRAVIFETLRLATVVNG 357 MLOC_75134 Hv-DWARF 265 TREKLSDEQIIDLLIALIYSGYETMSTTSMMAVKYLSDHPRALEELRREHLDIRKGKSPEEAISYQDFKSMSFTRAIIFETLRLATVVNG 354 .* ::*::* * :::::******:*******:*** :**:**:*** *** :*: * :* : :*.***.****:*:** ****:*** AT5G38970 At-BR6ox1/CYP85A1 348 VLRKTTRDLEINGYLIPKGWRIYVYTREINYDANLYEDPLIFNPWRWMKKSLESQNSCFVFGGGTRLCPGKELGIVEISSFLHYFVTRYR 437 AT3G30180 At-BR6ox2/CYP85A2 348 VLRKTTHDLELNGYLIPKGWRIYVYTREINYDTSLYEDPMIFNPWRWMEKSLESKSYFLLFGGGVRLCPGKELGISEVSSFLHYFVTKYR 437 AK372445 Hv-BRD 358 LLRKTTQDVEMNGYVIPKGWRIYVYTREINYDPFMYPDPMTFNPWRWLEKNMESHPHFMLFGGGARMCPGKEVGTAEIATFLHYFVTRYR 447 MLOC_75134 Hv-DWARF 355 LLRKTTRDVEMNGYVIPKGWRIYVYTREINYDPFMYPDPMTFNPWRWLEKNMESHPHFMLFGGGSRMCPGKEVGMAEIATFLHRFVTRYR 444 :*****:*:*:***:***************** :* **: ******::*.:**: ::**** *:*****:* *:::*** ***:** AT5G38970 At-BR6ox1/CYP85A1 438 WEEIGGDELMVFPRVFAPKGFHLRISPY 465 AT3G30180 At-BR6ox2/CYP85A2 438 WEENGEDKLMVFPRVSAPKGYHLKCSPY 465 AK372445 Hv-BRD 448 WEEEGKNTILKFPRVEAPNGLLIRVRDC 475 MLOC_75134 Hv-DWARF 445 WEEEGKNTILKFPRVEAPNGLHIRVQDY 472 *** * : :: **** **:* ::

12
Supplemental Figure 4. Alignments of Brassinosteroid-Biosynthesis Enzymes from
Arabidopsis and Barley.
(A-J) Coding sequences of Arabidopsis genes (Vriet et al., 2012) and deduced polypeptide
sequences were used to identify barley orthologs via the IPK barley BLAST server
(http://webblast.ipk-gatersleben.de/). For CYP724B1/DWF11, rice sequences were used to
identify the barley ortholog (Vriet et al., 2012). Alignments between Arabidopsis and barley
polypeptide sequences were performed using the software Clustal Omega
(http://www.ebi.ac.uk/Tools/msa/clustalo/). For At-CYP90C1 and At-CYP90D1, which are both
able to catalyze the conversion of 6-deoxocathasterone to 6-deoxoteasterone (late C-6 oxidation
pathway) or cathasterone to teasterone (early C-6 oxidation pathway), we found only one barley
ortholog (MLOC_62829) in the currently available barley genome data set. The polypeptide of
MLOC_62829 had higher similarity to At-CYP90D1 than to At-CYP90C1 (58.4% and 47.9% of
identical amino acid residues, respectively), thus we named the barley ortholog Hv-CYP90D1.
The polypeptides of At-BR6ox 1 and 2 were most similar to those of AK372445 (Hv-BRD) and
MLOC_75134 (Hv-DWARF) (Gruszka et al., 2011). Both AK372445 and MLOC_75134 are
located on the same fingerprint contig_34437 in the telomeric region of chromosome 2HS.

13

14
B Haruna Nijo/Foma Haruna Nijo/Foma insertion proline rich M62V At-BR6ox1/CYP85A1 1 MGAMMVMMGLLL-IIVSLCSALLRWNQMRYTK-------NGL PPGTMGWPIFG ETTEFLKQGPNFMRNQRLRYGSFFKSHLLGCPTLISM 82 At-BR6ox2/CYP85A2 1 MGIMMMILGLLV-IIVCLCTALLRWNQMRYSK-------KGL PPGTMGWPIFG ETTEFLKQGPDFMKNQRLRYGSFFKSHILGCPTIVSM 82 Os-BRD1/DWF 1 MVLVAIGVVV--AAAVVVSSLLLRWNEVRYSRK------RGL PPGTMGWPLFG ETTEFLKQGPSFMKARRLRYGSVFRTHILGCPTVVCM 82 Bd-CYP85A 1 MVLAVLIGVL--VGIVLVSSLLLRWNEVRYSRK------QGL PPGTMGWPLFG ETTEFLKQGPSFMKARRLRYGSLFRTHILGCPTVVCM 82 Sb-CYP85A 1 MAVLLLLVAV--LGVVLASSLLLRWNELRYSRR------RGL PPGTMGWPLFG ETTEFLKQGPSFMKQRRLRYGSLFRTHILGCPTVVCM 82 Hv-BRD AK372445 1 MALLLLALVVVVVGVVVASSLLLRWNEVRYGNGRRKHGDACL PPGTMGWPLFG ETTEFLKQGPSFVKQRRLRYGRLFRTHILGCPTVVCM 90 Hv-BRD Bowman 1 MALLLLALV----GVVVASSLLLRWNEVRYGNGRRKHGDACL PPGTMGWPLFG ETTEFLKQGPSFMKQRRLRYGRLFRTHILGCPTVVCM 86 Hv-BRD Foma 1 MALLLLALVVV--GVVVASSLLLRWNEVRYGNGRRKHGDACL PPGTMGWPLFG ETTEFLKQGPSFVKQRRLRYGRLFRTHILGCPTVVCM 88 Hv-DWARF MLOC_75134 1 MAFLLVLVGVVV-GVVLASSLLLRWNEVRYGNGRRKHGDGCL PPGTMGWPLFG ETTEFLKQGPSFMKQRRLRYGRLFRTHILGCPTVVCM 89 * .: *****::** . * ********:** **********.*:: :***** .*::*:*****::.* SRS1 At-BR6ox1/CYP85A1 83 DSEVNRYILK-NESKGLVP GYPQSMLDILGTCNMAAVHGSSHRLMRG SLLSLISSTMMRDHILPKVDHFMRSYLDQWNEL---EVIDIQD 168 At-BR6ox2/CYP85A2 83 DAELNRYILM-NESKGLVA GYPQSMLDILGTCNIAAVHGPSHRLMRG SLLSLISPTMMKDHLLPKIDDFMRNYLCGWDDL---ETVDIQE 168 Os-BRD1/DWF 83 EAELNRRALA-SEGRGFVP GYPQSMLDILGRNNIAAVQGPLHRAMRG AMLSLVRPAMIRSSLLPKIDAFMRSHLAAWSSSSSSAVVDIQA 171 Bd-CYP85A 83 DPELNRQMLQQGEGRGFVP GYPQSMLDILGRNNIAAVHGPLHRAMRG SMLALVRPASIRSSLLPKIDAFMRSHLHGWAG----DLVDIQD 168 Sb-CYP85A 83 EPELNRRTLA-SDGAGFVP GYPQSMLDILGPNNIAAVHGPLHRAMRG AMLALTRAHMIRAALLPKIDAFMRAHLHGWAG----RRVDIQE 167 Hv-BRD AK372445 91 DPELNRRMLLQGEAGGMVP GYPQSMLDILGRNNIAAVHGPLHRVMRG AMLGLVRPAALRQSLLPKMDAFMRHHLAGWAG----TVVDVQA 176 Hv-BRD Bowman 87 DPELNRRMLLQGEAGGMVP GYPQSMLDILGRNNIAAVHGPLHRVMRG AMLGLVRPAALRQSLLPKMDAFMRHHLAGWAG----TVVDVQA 172 Hv-BRD Foma 89 DPELNRRMLLQGEAGGMVP GYPQSMLDILGRNNIAAVHGPLHRVMRG AMLGLVRPAALRQSLLPKMDAFMRHHLAGWAG----TVVDVQA 174 Hv-DWARF MLOC_75134 90 DPELNRRMLLL--QGGLVP GYPQSMLDILGRNNIAAVHGPLHRVMRG AMLGLVCPTMLRQSLLPKIDAFMRDHLHGWAG----NIVDVQA 173 : *:** * *:* *********** *:***:* ** *** ::*.* :: :***:* *** :* * :*:* ari-u.245(int) SRS2 SRS3 L257* At-BR6ox1/CYP85A1 169 KTKHMAFLSSLTQIAGNLRKPFVE EFKTAFFKLVVG TLSVPIDLPGTNYRCGIQAR NNIDRLLRELM QERRDSGETFTDMLGYLMK--KE 256 At-BR6ox2/CYP85A2 169 KTKHMAFLSSLLQIAETLKKPEVE EYRTEFFKLVVG TLSVPIDIPGTNYRSGVQAR NNIDRLLTELM QERKESGETFTDMLGYLMK--KE 256 Os-BRD1/DWF 172 KTKEMALLSALRQIAGVSAGPLSD ALKAELYTLVLG TISLPINLPGTNYYQGFKAR KKLVAMLEQMI AERRSSGQVHDDMLDALL-TGVE 260 Bd-CYP85A 169 KTKEMALLSALRQIAGITAGPLSD ALKTELYTLVLG TISLPINLPGTSYYQGFQAR KKLVSMLEKMI AERRSSGLVHNDMLDALL-SGND 257 Sb-CYP85A 168 MTKEMALLSALRQIAGISAGPLSD ALKAELYTLVLG TFSLPINIPGTNYSKGLQAR KKLVAMLRQMI ADRRSSGCAQDDMLDALL-SGNE 256 Hv-BRD AK372445 177 KTKEMALLSALRQIAGITAGPLSD ALQTELYTLVLG TISLPINLPGTSYYQGFQAR TKLVSMLEQMI AERRSSGEAHDDMLDALLRSGDD 266 Hv-BRD Bowman 173 KTKEMALLSALRQIAGITAGPLSD ALQTELYTLVLG TISLPINLPGTSYYQGFQAR TKLVSMLEQMI AERRSSGEAHDDMLDALLRSGDD 262 Hv-BRD Foma 175 KTKEMALLSALRQIAGITAGPLSD ALQTELYTLVLG TISLPINLPGTSYYQGFQAR TKLVSMLEQMI AERRSSGEAHDDMLDALLRSGDD 264 Hv-DWARF MLOC_75134 174 KTKEMALLSALRQIAGITAGPLSD ALKTELYTLVLG TISLPINLPGTSYHQGFQAR TKLVSMLEQMI AQRRSSAQAHDDMLDALLRSGDD 263 **.**:**:* *** * : :: ::.**:* *:*:**::***.* *.:** .:: :* ::: :*:.*. . *** *: : SRS4 SRS5 Glu-X-X-Arg At-BR6ox1/CYP85A1 257 GNRYPLTDEEIR DQVVTILYSGYETVS TTSMMALKYLHDHPKALQELRAEHLAFRERKRQDEPLGLEDVKSMKFTR AVIYETSRLATIVN 346 At-BR6ox2/CYP85A2 257 DNRYLLTDKEIR DQVVTILYSGYETVS TTSMMALKYLHDHPKALEELRREHLAIRERKRPDEPLTLDDIKSMKFTR AVIFETSRLATIVN 346 Os-BRD1/DWF 261 GTREKLTDEQII DLIITLIYSGYETMS TTSMMAVKYLSDHPKALEQLRKEHFDIRKGKAPEDAIDWNDFKSMTFTR AVIFETLRLATVVN 350 Bd-CYP85A 258 GTRERLSDEQII DLIITLIYSGYETMS TTSMMAVKYLSDHPKALEELRKEHFDIRKGKSPEEAIDYNDFKSMTFTR AVIFETLRLATVVN 347 Sb-CYP85A 257 GTRAKLTDDQII DLLITLIYSGYETVS TTSMMAVKYLSDNPKALEQIRKEHLDIRKAKSPDDALDWNDYKSMTFTK AVIYETLRLATVVN 346 Hv-BRD AK372445 267 GTREKISDEQII DLLIALIYSGYETMS TTSMMAVKYLSEHPRALQELRREHLDIRKGKLPEEAIGYDDFKSMSFTR AVIFETLRLATVVN 356 Hv-BRD Bowman 263 GTREKISDEQII DLLIALIYSGYETMS TTSMMAVKYLSEHPRALQELRREHLDIRKGKLPEEAIGYDDFKSMSFTR AVIFETLRLATVVN 352 Hv-BRD Foma 265 GTREKISDEQII DLLIALIYSGYETMS TTSMMAVKYLSEHPRALQELRREHLDIRKGKLPEEAIGYDDFKSMSFTR AVIFETLRLATVVN 354 Hv-DWARF MLOC_75134 264 GTREKLSDEQII DLLIALIYSGYETMS TTSMMAVKYLSDHPRALEELRREHLDIRKGKSPEEAISYQDFKSMSFTR AIIFETLRLATVVN 353 .* ::*.:* * :::::******:* ******:*** :.*:**:::* **: :*: * :: : :* ***.**: *:*:** ****:** brh3.y/ert-t.437(int) G353D dioxygen binding heme binding At-BR6ox1/CYP85A1 347 GVLRKTTRDLEINGYLIPKGWRIYVYTREINYDAN LYEDPLIFNPWRWMKKSLESQNS CFVFGGGTRLCPG KELGIVEISSFLHYFVTRY 436 At-BR6ox2/CYP85A2 347 GVLRKTTHDLELNGYLIPKGWRIYVYTREINYDTS LYEDPMIFNPWRWMEKSLESKSY FLLFGGGVRLCPG KELGISEVSSFLHYFVTKY 436 Os-BRD1/DWF 351 GLLRKTTQDVEMNGYVIPKGWRIYVYTREINYDPF LYPDPMTFNPWRWLEKNMESHPH FMLFGGGSRMCPG KEVGTVEIATFLHYFVTQY 440 Bd-CYP85A 348 GLLRKTTQDVEMNGYVIPKGWRIYVYTREINYDPC LYPDPMTFNPWRWLEKNMESHPH FMLFGGGGRMCPG KEVGTAEIATFLHYFVTRY 437 Sb-CYP85A 347 GLLRKTTQDVEMNGYVIPKGWRIYVYTREINYDPF LYPEPMVFNPWRWLETNLESHPH FMLFGGGARMCPG KEVGTVEIATFLHYFITRY 436 Hv-BRD AK372445 357 GLLRKTTQDVEMNGYVIPKGWRIYVYTREINYDPF MYPDPMTFNPWRWLEKNMESHPH FMLFGGGARMCPG KEVGTAEIATFLHYFVTRY 446 Hv-BRD Bowman 353 GLLRKTTQDVEMNGYVIPKGWRIYVYTREINYDPF MYPDPMTFNPWRWLEKNMESHPH FMLFGGGARMCPG KEVGTAEIATFLHYFVTRY 442 Hv-BRD Foma 355 GLLRKTTQDVEMNGYVIPKGWRIYVYTREINYDPF MYPDPMTFNPWRWLEKNMESHPH FMLFGGGARMCPG KEVGTAEIATFLHYFVTRY 444 Hv-DWARF MLOC_75134 354 GLLRKTTRDVEMNGYVIPKGWRIYVYTREINYDPF MYPDPMTFNPWRWLEKNMESHPH FMLFGGGSRMCPG KEVGMAEIATFLHRFVTRY 443 *:*****:*:*:***:***************** :* :*: ******::..:**: ::**** *:**** *:* *:::*** *:*:* brh3.g/brh3.h/brh3.i(int) ari-u.304(int) W444* K449* SRS6 At-BR6ox1/CYP85A1 437 RWEEIGGD ELMVFPRVF APKGFHLRISPY 465 At-BR6ox2/CYP85A2 437 RWEENGED KLMVFPRVS APKGYHLKCSPY 465 Os-BRD1/DWF 441 RWEEEGNN TILKFPRVE APNGLHIRVQDY 469 Bd-CYP85A 438 RWEEEGTN TILKFPRVE APNGLHIRVQNY 466 Sb-CYP85A 437 RWEEEGNN TISKFPRVA APNGLHIRVQDY 465 Hv-BRD AK372445 447 RWEEEGKN TILKFPRVE APNGLLIRVRDC 475 Hv-BRD Bowman 443 RWEEEGKN TILKFPRVE APNGLLIRVRDC 471 Hv-BRD Foma 446 RWEEEGKN TILKFPRVE APNGLLIRVRDC 473 Hv-DWARF MLOC_75134 445 RWEEEGKN TILKFPRVE APNGLHIRVQDY 472 **** * : : **** **:* ::
Supplemental Figure 5. In Silico Mapping and Detailed Analysis of Conserved Amino Acid
Residues of Hv-BRD (for Hordeum vulgare BRASSINOSTEROID-6-OXIDASE).
(A) The polypeptide encoded by barley Hv-BRD (AK372445) is most similar to the Arabidopsis
brassinosteroid-6-oxidase encoded by At-BR6ox1/CYP85A1 (Supplemental Figure 4 online). Hv-
BRD is located on fingerprint contig_34437 on chromosome 2H at position 2.27 cM between

15
SNP markers 1_0326 and 1_1059. In the barley genome zipper, the 4 cM interval contained 21
gene models but did not include the Hv-BRD ortholog of Brachypodium (Bradi1g15030), rice
(LOC_Os03g40540), and sorghum (Sb1g015040). In EnsemblPlant (http://plants.ensembl.org),
the same interval (contig_1675 to contig_43923) contained 68 gene models including Hv-BRD.
The location of Hv-BRD at position 2.27 cM overlaps with introgression regions (marked with
blue, dotted lines) of five near-isogenic lines with a brassinosteroid-mutant phenotype - BW031
(ari-u.245), BW091 (brh3.g), BW092 (brh3.h), BW094 (brh3.y), and BW324 (ert-t.55). The
combination of genetic and phenotypic data suggests that these mutations are located in the Hv-
BRD gene. BC, number of backcrosses to cultivar Bowman.
(B) Alignment of polypeptides similar to Hv-BRD with mutations indicated (light gray). The
sequence for cultivar Haruna Nijo was retrieved from GenBank (accession number AK372445).
SRS, substrate recognition site. Inferred positions of functional domains (Mori et al., 2002; Hong
et al., 2005; Kim et al., 2005; Sakamoto and Matsuoka, 2006) are indicated (red). Numbering of
modified amino-acid residues refers to the position in the polypeptide of the respective plant
species. The effect on plant phenotype caused by the mutations is indicated (mild,
int/intermediate, str/strong).

16

17
B At-CPD 1 -MAFTAFL---------LLLSSIAAGFLLL----------LRRTRYRRMGLPPGSLGLPLIGETFQLIGAYKTENPEPFIDERVARYG-S 69 Os-CPD1 1 -MAAAALLLL--AAAAAIVVVAMVLRWLLLLGGPAAGR------LGKRALMPPGSTGLPLIGETLRLISAYKTPNPEPFIDERVARHG-G 80 Os-CPD2 1 -MAAAALLLL--AAAAAAVVVAMALRWLLLLGGPAAGR------LGKRARMPPGSTGLPLIGETLRLISAYKTPNPEPFIDERVARHG-G 80 Sb-CPD 1 MDAGGSPPLFLLAAAATAVLAAAALGWLLLAGGRAAAAGRRRRSASSAPRLPPGSTGLPLIGETLRLIAAYKTPNPEPFIDERVARHGSG 90 Bd-CPD 1 MENAGALQLLSAAAAAVAVVVVVALRWLLAR--RAAGG------WKQRPRLPPGSTGLPLIGETLRLISAYKTPDPEPFIDERVARHG-G 81 Hv-CPD 1 -MDAGALQLAATAVA--IVAVALVCRWFLVA--CAARG------RKQRPRLPPGSTGLPLIGETLRLISAYKTPDPEPFIDERVARHG-G 78 : : . ::* :**** ********::**.**** :***********:* . At-CPD 70 VFMTHLFGEPTIFSADPETNRFVLQNEGKLFECSYPASICNLLGKHSLLLMKGSLHKRMHSLTMSFANSSIIKDHLMLDIDRLVRFNLDS 159 Os-CPD1 81 VFTTHVFGERTVFSADPAFNRLLLAAEGRAVHSSYPSSIATLLGARSLLLTRGAAHKRLHSLTFTRLGRP-ASPPLLAHIDRLVLATMRQ 169 Os-CPD2 81 VFTTHVFGERTVFSADPAFNRLLLAAEGRAVHSSYPSSIATLLGARSLLLTRGAAHKRLHSLTLTRLGRP-ASPPLLAHIDRLVLATMRQ 169 Sb-CPD 91 VFTTHVFGERTVLSADPAFNRLLLAAEGRAVSCSYPSSIATLLGPRSLLLTAGPAHKRLHSLTLTRLGRP-ASPPLLAHIDRLVLATMRD 179 Bd-CPD 82 VFTTHVFGERTVFSADPAFNRLLLAAEGRAVDCSYPSSITTLLGARSLLLTRGAAHKRLHSLTLTRLGRP-ASPPLLAHIDRLVLDTMRH 170 Hv-CPD 79 VFTTHIFGERTVFSADPAFNRLLLAAEGPAVNCSYPSSINTLLGARFLVLTRAPAHKRLHSLTLTRLGRP-ASQPLLAHIDRLVLATMRQ 167 ** **:*** *::**** **::* ** . .***:** .*** : *:* . ***:****:: . *: .***** .: At-CPD 160 WSS----RVLLMEEAKKITFELTVKQLMSFDPGEWSESLRKEYLLVIEGFFSLPLPLFS----TTYRKAIQARRKVAEALTVVVMKRREE 241 Os-CPD1 170 WEP--AATVRLMDEAKKITFNLTVKQLVSIEPGPWTESLRREYVKLIDGFFSIPFPLANLLPFTTYGQALKARKKVAGALREVIKKRMEE 257 Os-CPD2 170 WEP--AATVRLMDEAKKITFNLTVKQLVSIEPGPWTESLRREYVKLIDGFFSIPFPLAYFLPFTTYGQALKARKKVAGALREVIKKRMEE 257 Sb-CPD 180 WGRPGAAPVRLLDEAKKITFNLTVWQLVSIEPGPWTESVRREYVKLIDGFFSIPFPFAYLLPFTVYGQALKARKKVAGALREVIRKRMDE 269 Bd-CPD 171 WEP--AATVRLLDEAKKITFNLTVKQLVSIDPGPWTESLRREYVKLIDGFFSIPFPFASLLPFTTYGQALKSRKKVAGALREVIRKRMEE 258 Hv-CPD 168 WEP--TATVRLLDEAKKITFNLTVKQLVSIEPGPWTESLRCEYVKLIDGFFSIPFPFASFLPFTTYGQALKSRKKVAGALREVIRRRMEE 255 * * *::*******:*** **:*::** *:**:* **: :*:****:*:*: *.* :*:::*:*** ** *: :* :* dioxygen binding At-CPD 242 -----------EEEGAERKKDMLAALLAAD-DGFSDEEIVDFLV ALLVAGYETTSTIMTLA VKFLTETPLALAQLKEEHEKIRAMKSDSY 319 Os-CPD1 258 KAENGGSI--GDDEGKKEKKDMVEELLEAEGGSFSEEEMVDFCL SLLVAGYETTSMLMTLA VKFLTETPAALAELKEEHANIRDMKGKKQ 345 Os-CPD2 258 KAENGGSI--GDDEGKKEKKDMVEELLQAEGGSFSEEEMVDFCL SLLVAGYETTSVLMTLA VKFLTETPAALAELKEEHANIRDMKGKNQ 345 Sb-CPD 270 KAETGGASKNGEDEEKREKKDMVEELLEAEGGSFSVEEMVDFCL SLLVAGYETTSVLMTLA VKFLTETPAALAQLKEEHDNIRGIKGKDQ 359 Bd-CPD 259 RGEESGAT--EEAESKREKKDMVEELLQAEGGSFSEEEMVDFCL SLLVAGYETTSVLMTVA VKFLTETPAALAQLKEEHENMTNMKGENQ 346 Hv-CPD 256 RGEENGVK--EEMDGKREKKDMVEELLEADGGSFSEEEMVDFCL SLLVAGYETTSVLMTVA VKFLTETPAALAQLKEEHENMTKMKGENQ 343 : : ..****: ** *: .** **:*** : :********** :**:* ******** ***:***** :: :*... steroid binding At-CPD 320 SLEWSDYKS MPFTQCVVNETLR VANIIGGVFRRAMTDVEIKGYKIPKGWKVFSSFRAVHLDPNHFKDARTFNPWRWQSNSVTT— GPSNV 407 Os-CPD1 346 PLEWSDYKS MPFTQCVINETLR VGNIISGVFRRANTDIHYKDYTIPKGCKIFASFRAVHLNNEHYENARTFNPWRWQINNKLQNAVGANI 435 Os-CPD2 346 PLEWSDYKS MPFTQCVINETLR VGNIISGVFRRANTDIHYKDYTIPKGCKIFASFRAVHLNNEHYENARTFNPWRWQINNKLQNAVGANI 435 Sb-CPD 360 PLEWSDYKS MPFTQCVISETLR VANLIGGVFRRANTDIHFKDYVIPKGCKIFASFRAVHLNLEHYENARTFDPWRWQSKNKLQNAEGASL 449 Bd-CPD 347 PLEWSDYKS MTFTQCVINETLR VANIISGVFRRANTDIHFKGYTIPKGCKIFASFRAVHLNNDHYENARTFDPWRWQSNHKLQNEVGANL 436 Hv-CPD 344 PLEWSDYKS MNFTQCVINETLR VANIISGVFRRANTDIHFKGYTIPKGCKIFASFRAVHLNNEPYENARTFDPWRWQSNNKLQNEVGANL 433 ******** * *****:.**** *.*:*.****** **:. * * **** *:*:*******: : :::****:***** : :.: brh13.p(int) brh18.ac(mild-int) P445L P479S heme.binding . At-CPD 408 FT PFGGGPRLCPGYELARVALSVF LHRLVTGFSWVPAEQDKLVFFPTTRTQKRYPIFVKRRDFAT- 472 Os-CPD1 436 FT PFGGGPRLCPGYELARVVVSIF LHHLVTRFSWEETEEDRLVFFPTTRTLKGYPINLRLLSESIC 501 Os-CPD2 436 FT PFGGGPRLCPGYELARVVVSIF LHHLVTRFSWEETEEDRLVFFPTTRTLKGYPINLRLLSESIC 501 Sb-CPD 450 FT PFGGGPRLCPGYELARVVVSVF LHHLVTCFSWEEAEEDRIVFFPTTRTLKGYPINLRRRPDYA- 514 Bd-CPD 437 FT PFGGGPRLCPGYELARVVISVF LHHLVMRFSWEAAEEDRLVFFPTTRTLKGYPINLRQRS---- 498 Hv-CPD 434 FT PFGGGPRLCPGYELARVVISVF LHHLVMRFSWEAAEEDRLLFFPTTRTLKGYPINLTRRSKSV- 498 ** *****************.:*:* **:** *** :*:*:::******* * *** :
Supplemental Figure 6. In Silico Mapping and Detailed Analysis of Conserved Amino Acid
Residues of Hv-CPD (for Hordeum vulgare CONSTITUTIVE PHOTOMORPHOGENIC
DWARF).
(A) Hv-CPD (MLOC_10658), orthologous to Arabidopsis At-CYP90A1/CPD (AT5G05690), is
not anchored to a fingerprint contig in the currently available barley genome map (International
Barley Genome Sequencing Consortium, 2012). However, in the barley genome zipper (Mayer
et al., 2011), the ortholog of Hv-CPD in Brachypodium (Bradi4g43110) is located close to SNP
marker 3_0109 (AK355248 on fingerprint contig_38549 at 44.24 cM; ortholog of
Bradi4g43070), flanked by SNP markers 1_0661 (MLOC_33273, fingerprint contig_48625,
44.24 cM; ortholog of Bradi4g42940), 2_0737 (MLOC_5324, fingerprint contig_45234 at 44.51
cM; ortholog of Bradi4g44390), and 1_1128 (AK368749, fingerprint contig_44417, 45.17 cM;
ortholog of Bradi4g44460). The introgression region of BW084 (brh13.p) is bordered by
markers 2_1324 and 2_1121. In the barley genome zipper, this interval contained 1,380 gene

18
models including the Hv-CPD ortholog of Brachypodium, rice, and sorghum. In EnsemblPlant
(http://plants.ensembl.org), the same interval contained 808 gene models. BW089 (brh18.ac)
was not included in the marker analysis by Druka et al. (Druka et al., 2011) but brh18.ac has
previously been linked to brh13.p via the SSR marker Hv-LEU (Dahleen et al., 2005). BC,
number of backcrosses to cultivar Bowman.
(B) Alignment of polypeptides similar to Hv-CPD with the amino acid residues affected by the
mutations indicated (light gray). Inferred positions of functional domains (Mori et al., 2002;
Hong et al., 2005; Kim et al., 2005; Sakamoto and Matsuoka, 2006) are indicated (red).
Numbering of modified amino-acid residues refers to the position in the polypeptide of the
respective plant species. The effect on plant phenotype caused by the mutations is indicated
(mild, int/intermediate, str/strong).

19

20
B At-DIM 1 ----------------MSDL-----QTPLVRPKRKK-TWVDYFVKFRWIIVIFIVLPFSATFYFLIYLGDMWSESKSFEKRQKEHDENVK 68 Os-DIM 1 ----------------MADL-----QEPLVRPKRKK-VLVDYLVKFRWILVIFVVLPISALIYFNIYLGDVWSAMKSEKRRQKEHDDNVQ 68 Sb-DIM_1 1 MDSRLRQSLLEKHPPPMADV-----HEPLVRRKRKK-VLVDYLVQFRWILVIFVVLPISSLIYFNIFLGDMWSAMKSEKKRQKQHDENVQ 84 Sb-DIM_2 1 ----------------MADL-----QTPLVRPKRKKLVLVDYLVRLRWIPALFIALPISALIYFSVFVGNTWSAMKSEKRRQKEHEENMK 69 Bd-DIM_1 1 ----------------MADL-----QEPLVRGKRKK-VLVDYLVQFRWILVIFVVLPISSLIYFNIYLGDMWSAMKSEKKRQKEHDDNVQ 68 Bd-DIM_2 1 ----------------MADL-----QEPLVRGKRKK-VLVDYLVQFRWILVIFVVLPISSLIYFNIYLGDMWSAMKSEKKRQKEHDENVQ 68 Hv-DIM 1 ----------------MADL-----QTPLVRPKRKK-VLVDYLVQFRWILVIFVVLPASALIYFNIYLGDMWSAMKSEKKRQKEHDDNVQ 68 Seladin-1 1 ------------MEPAVSLAVCALLFLLWVRLKG----LEFVLIHQRWVFVCLFLLPLSLIFDIY-YYVRAWVVFKLSSA-PRLHEQRVR 72 :: ** * ::: **: . :. ** * : : : * * . : *::.:: ert-zd.159(mild-int) FAD binding E131V At-DIM 69 KVIKRLKG-RDASKDGLVCTARKPWIAVGMRNVDYKRARHFE VDLGEFRNILEINKEKMTARVEPLVNMGQISRATVPMNLSLAVVAELD 157 Os-DIM 69 KVVKRLKQ-RNPKKDGLVCTARKPWIAVGMRNVDYKRARHFE VDLSAFRNILEIDRERMVAKVEPLVNMGQITRATCPMNLALAVVAELD 157 Sb-DIM_1 85 KVVKRLKQ-RNPKKDGLVCTARKPWIAVGMRNVDYKRARHFE VDLSSFRNILEIDKERMVAKVEPLVNMGQITRATCPMNLALAVVAELD 173 Sb-DIM_2 70 RVVKRLKA-LFALL------GSHGWL---------------- --LACATNILEIDTERMVAKVEPLVSMGQITKATCPMNLSLAVAPEFD 134 Bd-DIM_1 69 KVVKRLKQ-RNPKKDGLVCTARKPWIAVGMRNVDYKRARHFE VDLSAFRNILEIDAERMVAKVEPLVNMGQISRATCPLNLSLAVVAELD 157 Bd-DIM_2 69 KVVKRLKQ-RNPKKDGLVCTARKPWIAVGMRNVDYKRARHFE VDLSAFRNILEIDTERMVAKVEPLVNMGQISRATCPMNVSLAVVAELD 157 Hv-DIM 69 KVVKRLKQ-RNPKKDGLVCTARKPWIAVGMRNVDYKRVRHFE VDLSAFRNILEIDAERMVAKVEPLVNMGQISRATCPMNLSLAVVAELD 157 Seladin-1 73 DIQKQVREWKEQGSKTFMCTGRPGWLTVSLRVGKYKKTHKN- -IMINLMDILEVDTKKQIVRVEPLVTMGQVTALLTSIGWTLPVLPELD 160 : *::: . *: : :***:: :: .:*****.***:: : :* * *:* brh14.q(mild-int) L164F FAD binding At-DIM 158 DLTVGGLINGYGIEGSSHIYGLFADTVEAYEIVLAGGELVRATRDNEYSDLYYAIPWSQGT -LGLLVAAEIRLIKVKEYMRLTYIPVKGD 246 Os-DIM 158 DLTVGGLINGYGIEGSSHLYGLFSDTVVAVEVVLADGRVVRATKDNEYSDLFYGIPWSQGT -LGFLVSAEIKLIPIKEYMRLTYTPVKGS 246 Sb-DIM_1 174 DLTVGGLINGYGIEGSSHLYGLFSDTVVAMEVVLADGRVVRATKDNEYSDLFYGIPWSQGT -LGFLVSAEIKLIPIKEYMKLTYIPVKGS 262 Sb-DIM_2 135 DLTVGGLINSYGISGGSHIYGLFTDTVVAMEVVLADGQVVRATMDNEHSDLFLWHAMVPRH DWAPCFSRDQAHSCQGIHEAHIYTPVRGT 224 Bd-DIM_1 158 DLTVGGLINGYGIEGSSHLYGLFSDTVVALEIVLADGKVVRATKDNEYSDLFYGVPWSQGT -IGFLVSAEIKLIPIKEYMRLTYTPVKGD 246 Bd-DIM_2 158 DLTVGGLINGYGIEGSSHLYGLFSDTVVALEIVLADGRVVRATKDNEYSDLFYGVPWSQGT -IGFLVSAEIKLIPIKEYMRLTYTPVKGD 246 Hv-DIM 158 DLTVGGLINGYGIEGSSHIYGLFSDTVVALEIVLADGRIVRATKDNEYSDLFYGVPWSQGT -LGFLVSAEIKLIPIKEYMRLTYTPVKGP 246 Seladin-1 161 DLTVGGLIMGTGIESSSHKYGLFQHICTAYELVLADGSFVRCTP-SENSDLFYAVPWSCGT -LGFLVAAEIRIIPAKKYVKLRFEPVRGL 248 ******** . **...** **** . * *:*** * .**.* .* ***: . .: : : : **:* At-DIM 247 LQALAQGYIDSFAPKDGDKSKIPDFVEGMVYNPTEGVMMVGTYASKEEAKKKGNKINNVGWWFKPWFYQHAQTALKK-GQFVEYIPTREY 335 Os-DIM 247 LKEIAQGYCDSFAPRDGDPAKVPDFVEGMVYTENEGVMMTGVYASKEEAKKKGNKINCVGWWFKPWFYQHAQTALKK-GEFVEYIPTREY 335 Sb-DIM_1 263 LKEIAQAYADSFAPRDGDPAKVPDFVEGMVYTESEGVMMTGVYASKEEAKKKGNKINCVGWWFKPWFYQHAQTALKR-GEFVEYIPTREY 351 Sb-DIM_2 225 LKEIAEAYADSFVPRDGDPAKVPDFVEGMVYSSSEGVTMTGVYASEEEAKKKGNRINRVGWWFKPWFYQYAETALKR-GEFVEYIPTREY 313 Bd-DIM_1 247 LKEIAQGYADSFAPRDGDPSKVPDFVEGMVYNPTEGVMMTGVYASKEEAKKKGNKINSVGWWFKPWFYQHAQTALKK-GEFVEYIPTRQY 335 Bd-DIM_2 247 LKEIGQAYADSFAPRDGDPSKVPDFVEGMVYTATESVMMTGVYASKEEAKKKGNKINSVGWWFKPWFYQHAQMALKK-GEFVEYIPTRQY 335 Hv-DIM 247 LKELAQAYADAVAPRDGDPAKVPDFVEGMVYNATEGVMMTGVYASKEEAKKKGNKINSVGWWFKPWFYQHAQTALKK-GEFVEYIPTREY 335 Seladin-1 249 EAICAK-FT------HESQRQENHFVEGLLYSLDEAVIMTGVMTDEAEP----SKLNSIGNYYKPWFFKHVENYLKTNREGLEYIPLRHY 327 .: : . . : .****::*. *.* *.*. :.: * .::* :* ::****:::.: ** : :**** *.* ari-o.40/ari-o.43/ari-o.143(mild-int) W345R At-DIM 336 YHRHTRCLYWEGKLILPFGDQFWFRYLLGWLMPPKVSLLKATQGEAIRNYYHDMHVIQDMLVPLYKVGDALEWVHREMEVYPIWLCPHKL 425 Os-DIM 336 YHRHTRCLYWEGKLILPFGDQFWFRFLLGWLMPPKVSLLKATQGESIRNYYHDNHVIQDMLVPLYKVGDALEFVHKEMEVYPLWLCPHRL 425 Sb-DIM_1 352 YHRHTRCLYWEGKLILPFGDQFWFRFLLGWLMPPKVSLLKATQGEAIRNYYHDNHVIQDMLVPLYKVGDALEFVHREMEVYPLWLCPHRL 441 Sb-DIM_2 314 YHRHTRSLYWEGKLIIPFGDQFWFRFLLGWLMPPKISLLKITQGEAIRNYYHDNHVIQDVLVPLHKVSDALEFAHRELEVYPVWLCPHRL 403 Bd-DIM_1 336 YHRHTRCLYWEGKLILPFGDQFWFRYLFGWLMPPKVSLLKATQGEAIRNYYHDNHVIQDMLVPLYKVGEALEFVHREMEVYPLWLCPHRL 425 Bd-DIM_2 336 YHRHTRCLYWEGKLILPFGDQFWFRYLFGWLMPPKVSLLKATQGEAIRNYYHDNHVIQDMLVPLYKVGDALEFVHREMEVYPLWLCPHRL 425 Hv-DIM 336 YHRHTRCLYWEGKLILPFGDQFWFRFLFGWLMPPKVSLLKATQGDAIRNYYHDNHVIQDMLVPLYKVGDALEFVHQEMEVYPLWLCPHRL 425 Seladin-1 328 YHRHTRSIFWELQDIIPFGNNPIFRYLFGWMVPPKISLLKLTQGETLRKLYEQHHVVQDMLVPMKCLQQALHTFQNDIHVYPIWLCPFIL 417 ******.::** : *:***:: **:*:**::***:**** ***:::*: *.: **:**:***: : :**. :.::.***:****. * At-DIM 426 FKQPIKGQIYPEPGFEYENRQGDTEDAQMYTDVGVYYAPGCVLRGEEFDGSEAVRRMEKWLIENHGFQPQYAVSE 500 Os-DIM 426 YKLPVKTMVYPEPGFEHHHRQGDTSYAQMFTDVGVYYAPGAVLRGEEFNGALAVHRLEQWLIENHSYQPQYAVSE 500 Sb-DIM_1 442 YKLPVKTMVYPEPGFEHQHRQGDTSYAQMFTDVGVYYAPGAVLRGEEFNGAEAVHRLEQWLIENHSYQPQYAVSE 516 Sb-DIM_2 404 YKLPVKTMVHPEPGFEQHRRKGDTSYAQMFTDVGFYYAPASVLRGEEFNGAEAVHRLEQWLIRNHGYQAQYAVSE 478 Bd-DIM_1 426 YKLPVKTMVYPEPGFELHQRQGDTSYAQMFTDVGVYYTPAFIFRGEEFNGVEAVKRLEQWLIENHSYQPQYAVSE 500 Bd-DIM_2 426 YKLPVKTMVYPEPGFELHQRQGDTSYAQMFTDVGVYYTPAFIFRGEEFNGVEAVRRLEQWLIENHSYQPQYAVSE 500 Hv-DIM 426 FKLPVKTMIYPEPGFEHHQRQGDTSYAQMFTDVGVYYTPAFIFRGEQFDGVEAVKRLEQWLIENHSYQPQYAVTE 500 Seladin-1 418 PSQP---------GLVHP----KGNEAELYIDIGAYGEPR----VKHFEARSCMRQLEKFVRSVHGFQMLYADCY 475 . * *: . . *::: *:* * * :.*:. .::::*::: *.:* ** brh16.v(mild-int) R508ACSTRRTTSTAGRSTAPWAPS* At-DIM 501 LDEKSFWRMFNGELYEECRKKYRAIGTFMSVYYKSKKGRKTEKEVREAEQAHLETAYAEAD 561 Os-DIM 501 LNEKDFWRMFDASHYEHCRQKYGAVGTFMSVYYKSKKGRKTEKEVQEAEAAILEPAYADEA 561 Sb-DIM_1 517 LNEKDFWRMFDASHYEHCRHKYGAVGTFMSVYYKSKKGRKTEKEVQEAEAAILEPAYADEA 577 Sb-DIM_2 479 LSEKDFWRMFDPSHYEHCRRKYGAVGSFMSAHYKSKKRQKERGGGARS------------- 526 Bd-DIM_1 501 LNEKDFWRMFDASHYEHCRQKYGAVGTFMSVYYKSKKGRKTEKEVQEAEAAILEPAYADEA 561 Bd-DIM_2 501 LNEKDFWRMFDASHYELCRQKYGAVGTFMSVYYKSKKGRKTEKEVQEAEAAILEPAYADEA 561 Hv-DIM 501 LNEKDFWRMFDASHYEHCRQKYGAVGTFMSVYYKSKKGRKSEKEVQEAEAAILEPAYADEA 561 Seladin-1 476 MNREEFWEMFDGSLYHKLREKLGCQDAFPEVYDKICKAARH-------------------- 516 :..:.**.**: . *. *.* . :* ..: * * :
Supplemental Figure 7. In Silico Mapping and Detailed Analysis of Conserved Amino Acid
Residues of Hv-DIM (for Hordeum vulgare DIMINUTO).
(A) Hv-DIM was mapped to the telomere of chromosome 7HL in the barley physical map. The
Hv-DIM unigene is identical to SNP 1_0547, which is located on fingerprint contig_44330 at

21
position 138.22 cM. In the barley genome zipper, the 2.4 cM interval [contig_44330 to
contig_48789; marked with blue dotted lines] contained three gene models including the Hv-
DIM ortholog of Brachypodium, rice, and sorghum. In EnsemblPlant (http://plants.ensembl.org),
the same interval contained 27 gene models including Hv-DIM. Six near-isogenic lines with a
brassinosteroid-mutant phenotype, BW053 (ari-o.40), BW072 (brh.af), BW085 (brh14.q),
BW087 (brh16.v), BW325 (ert-u.56), and BW333 (ert-zd.159) had an introgression region
overlapping the location of Hv-DIM (Dahleen et al., 2005; Druka et al., 2011; Franckowiak and
Lundqvist, 2012). The same was found for nine additional near-isogenic lines with a short-culm
phenotype, but without other distinct brassinosteroid-mutant characters (Dataset 1.1).
Sequencing of Hv-DIM from the latter nine lines revealed no mutations, which suggest that they
are not deficient in Hv-DIM. BC, number of backcrosses to cultivar Bowman.
(B) Alignment of polypeptides similar to Hv-DIM with the amino acid residues affected by the
mutations indicated (light gray). Inferred positions of functional domains (Mori et al., 2002;
Hong et al., 2005; Kim et al., 2005; Sakamoto and Matsuoka, 2006) are indicated (red).
Numbering of modified amino-acid residues refers to the position in the polypeptide of the
respective plant species. The effect on plant phenotype caused by the mutations is indicated
(mild, int/intermediate, str/strong).

22

23
B 3RGZ> uzu1.297(int) bri1-5(mild) bri1-5R1 | N-terminal cap F53S C69Y LRR1 G87E At-BRI1 1 MKTFSSFFLSVTTLFFFSFFSLSFQASPSQSLYREIHQLISFKDVLPDK---NLLPDWSSNKNPCTFDGVTCRDDK VTSIDLSSKPLNVGFSAVSSSLLS 97 Os-BRI1 1 ---MDSLWAAIAALFVAAAVVVRGAAAA-----DDAQLLEEFRQAVPNQ---AALKGWSGGDGACRFPGAGCRNGR LTSLSLAGVPLNAEFRAVAATLLQ 89 Sb-BRI1 1 ---MESPGLV---AVVALFVAVVAAVAA-----DDAQLLEQFKEAVPSQSQAADFRGWSASDGACKFPGAGCRGGR LTSLSLAAVPLNADFRAVEATLLQ 89 Bd-BRI1 1 ---MDSLRVAIAAALFVAAVAVAVAAAA-----DDAQLLEEFRAAVPNQ---ASLSGWKAADGACRFPGAACRAGR LTSLSLAGVPLNADFRAVAATLLQ 89 Hv-BRI1 1 ---MDCLRLAVAAAALL---LAALAAAA-----DDAQLLDEFRMALPSQ---APLEGWTAREGACRFPGAVCRGGR LTSLSLAAVTLNADFRAVANTLLQ 86 :.. . : : * .*: .:*.: : *.. . * * *. ** : :**:.*:. **. * ** :**. LRR2 LRR3 LRR4 At-BRI1 98 LTGLESLFLSNSHINGSVS---GFKC SASLTSLDLSRNS-LSGPVTTLTS-LGS CSGLKFLNVSSNTLDFPGKV---SGGLK 171 Os-BRI1 90 LGSVEVLSLRGANVSGALSAAGGARC GSKLQALDLSGNAALRGSVADVAALASA CGGLKTLNLSGDAVGAAKV--GGGGGPG 169 Sb-BRI1 90 LGSLETLSLRGANVSGALAA--VPRC GAKLQSLDLSGNAGLRGSVADVDALAAA CAGLSALNLSGCSVGGPRS--AGAVASG 167 Bd-BRI1 90 LSGVEALSLRGANVSGALAAAGGARC GGKLEALDLSGNAALRGSVADVAALADS CAGLKKLNLSGGAVGAAKA--GGGGGAG 169 Hv-BRI1 87 LSAVERLSLRGANVSGALAA---ARC GGKLEELDLSGNAALRGSVADVAALAGS CGALRTLNLSGDAVGAAKPAGGGGGGQG 165 * .:* * * :.:.*::: :* ...* **** *: * * *: : : : *..* **:*. :: . . LRR5 LRR6 LRR7 LRR8 At-BRI1 172 LNSLEVLDLSANSISGANVVGWVLSDG CGELKHLAISGNKISGDVDVSR CVNLEFLDVSSNNFSTGIPFLGD CSALQHLDISGNKLSGDFSR-AIST 267 Os-BRI1 170 FAGLDSLDLSNNKITDDSDLRWMVDAG ---------------------- VGAVRWLDLALNRISG-VPEFTN CSGLQYLDLSGNLIVGEVPGGALSD 243 Sb-BRI1 168 FARLDALDLSDNKISGDGDLRWMVGAG ---------------------- VGAVRRLDLSGNKISA-LPEFNN CSGLEYLDLSGNLIAGEVAGGILAD 241 Bd-BRI1 170 FAALDVLDLSNNKITGDAELRWMVGAG ---------------------- VGSVRWLDLAWNRISGELPDFTN CSGLQYLDLSGNLIDGDVAREALSG 244 Hv-BRI1 166 FAALDALDLSSNKIAGDADLRWMVGAG ---------------------- LGSVRWLDLAWNKISGGLSDFTN CSGLQYLDLSGNLIAGDVAAAALSG 240 : *: **** *.*: : *:: * :. **:: *.:* : : : **.*::**:*** : *:. :: uzu1.301(int) LRR9 LRR10 LRR11 LRR12 V282D At-BRI1 268 CTELKLLNISSNQFVGPIPPLP LKSLQYLSLAENKFTGEIPDFLSGA CDTLTGLDLSGNHFYGAVPPFFGS CSLLESLALSSNNFSGELPMDTLLK 363 Os-BRI1 244 ---------------------- ------------------------- CRGLKVLNLSFNHLAGVFPPDIAG LTSLNALNLSNNNFSGELPGEAFAK 292 Sb-BRI1 242 ---------------------- ------------------------- CRGLRTLNLSGNHLVGPFPPDVAA LTSLAALNLSNNNFSSELPADAFTE 290 Bd-BRI1 245 ---------------------- ------------------------- CRSLRALNLSSNHLAGAFPPNIAG LASLTALNLSNNNFSGEVPADAFTG 293 Hv-BRI1 241 ---------------------- ------------------------- CRSLRALNLSSNHLAGAFPPNIAG LTSLTALNLSNNNFSGEVPADAFTG 289 * * *:** **: * .** ... : * :* **.****.*:* ::: bri1-120(mild) LRR13 LRR14 S399F LRR15 LRR16 At-BRI1 364 MRGLKVLDLSFNEFSGELPESLTN LSASLLTLDLSSNNFSGPILPNLC QNPK NTLQELYLQNNGFTGKIPPTLSN CSELVSLHLSFNYLSGTIPSSLGS 462 Os-BRI1 293 LQQLTALSLSFNHFNGSIPDTVAS LP-ELQQLDLSSNTFSGTIPSSLC QDPN SKLHLLYLQNNYLTGGIPDAVSN CTSLVSLDLSLNYINGSIPASLGD 390 Sb-BRI1 291 LQQLKALSLSFNHFNGTIPDSLAA LP-ELDVLDLSSNSFSGTIPSSIC QGPN SSLRMLYLQNNYLSGAIPESISN CTRLQSLDLSLNNINGTLPASLGK 388 Bd-BRI1 294 LQQLKSLSLSFNHFTGSIPDSLAA LP-ELEVLDLSSNTFTGTIPSSIC QDPN SSLRVLYLQNNFLDGGIPEAISN CSNLVSLDLSLNYINGSIPESLGE 391 Hv-BRI1 290 LQQLQSLSLSFNHFSGSIPDSVAA LP-DLEVLDLSSNNFSGSIPDSLC QDPN SRLRVLYLQNNYLSGSIPEAVSN CTDLVSLDLSLNYINGSIPESLGE 387 :: * *.****.*.* :*:::: * .* ******.*:* * .:* * *: . *: ****** : * ** ::** *: * **.**:* :.*::* ***. d61-3(str) d61-5(str) d61-7(mild) LRR17 18 H420P N426Y LRR19 20 A467V At-BRI1 463 LSKLRDLKLWLNMLEGEIPQELMY VKTLETLILDFNDLTGEIPSGLSN CTNLNWISLSNNRLTGEIPKWIG RLENLAILKLSNNSFSGNIPAELGD 558 Os-BRI1 391 LGNLQDLILWQNELEGEIPASLSR IQGLEHLILDYNGLTGSIPPELAK CTKLNWISLASNRLSGPIPSWLG KLSYLAILKLSNNSFSGPIPPELGD 486 Sb-BRI1 389 LGELRDLILWQNLLVGEIPASLES LDKLEHLILDYNGLTGGIPPELSK CKDLNWISLASNQLSGPIPAWLG QLSNLAILKLSNNSFSGPIPAELGN 484 Bd-BRI1 392 LAHLQDLIMWQNSLEGEIPASLSR IRGLEHLILDYNGLSGSIPPDLAK CTQLNWISLASNRLSGPIPSWLG KLSNLAILKLSNNSFSGRVPPELGD 487 Hv-BRI1 388 LSRLQDLIMWQNLLEGEIPASLSS IPGLEHLILDYNGLTGSIPPELAK CKQLNWISLASNRLSGPIPSWLG KLSNLAILKLSNNSFTGKIPAELGD 483 *..*:** :* * * **** .* : ** ****:* *:* ** *:: *..******:.*:*:* ** *:* :*. ***********:* :* ***: bri1-113/d61-9(str/mild) sud1 d61-2(int) d61-8(mild) G611E/G539D G643E V491M G522E . bri1-7(mild) .bri1-6/119(mild) LRR21 island domain . G613S .G644D At-BRI1 559 CRSLIWLDLNTNLFNGTIPAAMFKQ SGKIAANFIAGKRYVYIKNDGMKKECHGAGNLLEFQGIRSEQLNRLSTRNPCNITSRVYGGHTSPTFDNNG 654 Os-BRI1 487 CQSLVWLDLNSNQLNGSIPKELAKQ SGKMNVGLIVGRPYVYLRNDELSSECRGKGSLLEFTSIRPDDLSRMPSKKLCNFT-RMYVGSTEYTFNKNG 581 Sb-BRI1 485 CQSLVWLDLNSNQLNGSIPAELAKQ SGKMNVGLVIGRPYVYLRNDELSSECHGKGSLLEFTSIRPEELSRMPSKKLCNFT-RVYMGSTEYTFNKNG 579 Bd-BRI1 488 CKSLVWLDLNNNQLNGSIPPELAEQ SGKMSVGLIIGRPYVYLRNDELSSQCRGKGSLLEFSSIRSEDLSRMPSKKLCNFT-RVYMGSTEYTFNKNG 582 Hv-BRI1 484 CKSLVWLDLNSNQLNGSIPPELAEQ SGKMTVGLIIGRPYVYLRNDELSSQCRGKGSLLEFSSIRSEDLSRMPSKKLCNFT-RMYMGSTEYTFNKNG 578 *:**:*****.* :**:** : :* ***: . :: *: ***::** :..:*:* *.**** .** ::*.*: ::: **:* *:* * *. **::** R564W T573K uzu1.256(mild) uzu1.b/ert-ii.79(int) bri1-9(mild) d61-11/12(int) LRR22 S622F LRR23 LRR24 G643E/R LRR25 At-BRI1 655 SMMFLDMSYNMLSGYIPKEIGS MPYLFILNLGHNDISGSIPDEVGD LRGLNILDLSSNKLDGRIPQAMSA LTMLTEIDLSNNNLSGPIPEMGQ 747 Os-BRI1 582 SMIFLDLSYNQLDSAIPGELGD MFYLMIMNLGHNLLSGTIPSRLAE AKKLAVLDLSYNQLEGPIPNSFSA LS-LSEINLSNNQLNGTIPELGS 673 Sb-BRI1 580 SMIFLDLSFNQLDSEIPKELGN MFYLMIMNLGHNLLSGVIPPELAG AKKLAVLDLSHNQLEGPIPNSFST LS-LSEINLSNNQLNGSIPELGS 671 Bd-BRI1 583 SMIFLDLSFNQLDSEIPKELGN MFYLMIMNLGHNLLSGPIPLELAG AKKLAVLDLSYNRLEGPIPSSFST LS-LSEINLSSNQLNGTIPELGS 674 Hv-BRI1 579 SMIFLDLSFNQLDSEIPKELGN MFYLMIMNLGHNLLSGAIPTELAG AKKLAVLDLSHNRLEGQIPSSFSS LS-LSEINLSSNQLNGTIPELGS 670 **:***:*:* *.. ** *:*. * **:*:***** :** ** .:. : * :**** *:*:* **.::*: *: *:**:**.*:*.* ***:*. uzu1.c(mild) bri1-102(str) <3RGZ R710K bri1(Y831F) T750I | . transmembrane domain Y831F At-BRI1 748 FETFPPAKFLNNPGLCGYPLPRCDPSNADGYAHHQRSHGRRPA SLAGSVAMGLLFSFVCIFGLILVG REMRKRRRKKEA---ELEMYAEGHGNSGDRTAN 844 Os-BRI1 674 LATFPKSQYENNTGLCGFPLPPCDHSS-PRSSNDHQ-SHRRQA SMASSIAMGLLFSLFCIIV-IIIA IGSKRRRLKNEEASTSRDIYIDSRSHSA--TM- 767 Sb-BRI1 672 LFTFPKISYENNSGLCGFPLLPCGHNAGSSSSNDRR-SHRNQA SLAGSVAMGLLFSLFCIVGIVIIA IECKKRKQINEEANTSRDIYIDSRSHSG--TM- 767 Bd-BRI1 675 LATFPKSQYENNSGLCGFPLPPCQAHAGQSASDGHQ-SHRRQA SLAGSVAMGLLFSLFCIFGLVIIA IESKKRRQKNEEASTSHDIYIDSRSHSG--TM- 770 Hv-BRI1 671 LATFPKSQYENNSGLCGFPLPPCESHTGQGSSNGGQ-SNRRKA SLAGSVAMGLLFSLFCIFGLVIIA IESKKRRQKNDEASTSRDIYIDSRSHSG--TM- 766 : *** .: ** ****:** * :. : *. * *:*.*:*******:.**. :::. ::*: :: . ::* :.:..*. *

24
4OAB> bri1(T872A) bri1-1(str) d54(str) d61-10(mild) uzu1.a(mild>str) | T872A A909T I834F T854I H857R At-BRI1 845 NTN-WKLT-GVKEALSINLAAFEKPLRKLTFADLLQATNGFHNDSLIGSGGFGDVYKAILKDGSAVAIKKLIHVSGQGDREFMAEMETIGKIKHRNLVPLLG 944 Os-BRI1 768 NSD-WRQNLSGTNLLSINLAAFEKPLQNLTLADLVEATNGFHIACQIGSGGFGDVYKAQLKDGKVVAIKKLIHVSGQGDREFTAEMETIGKIKHRNLVPLLG 868 Sb-BRI1 768 NSNNWR--LSGTNALSVNLAAFEKPLQKLTFNDLIVATNGFHNDSLIGSGGFGDVYKAQLKDGKVVAIKKLIHVSGQGDREFTAEMETIGRIKHRNLVPLLG 867 Bd-BRI1 771 NSN-WR--LSGTNALSINLAAFEKPLQKLTLGDLVEATNGFHNDSLIGSGGFGDVYKAQLKDGRIVAIKKLIHVSGQGDREFTAEMETIGKIKHRNLVPLLG 869 Hv-BRI1 767 NSN-WR--LSGTNALSINLAAFEKPLQKLTLGDLVEATNGFHNDSLIGSGGFGDVYKAQLKDGRVVAIKKLIHVSGQGDREFTAEMETIGKIKHRNLVPLLG 865 *:: *: . .: **:*********::**: **: ****** . ************ **** ***************** *******:*********** bri1-T1039A(int) bri1-S1042A(int) T1039A S1042A bri1-202(str) bri1-8/108-112(int) bri1-301(mild) bri1-D1027N(str) bri1-103(str) R952W R983N/R983Q G989I D1027N A1031T . . At-BRI1 945 YCKVGDERLLVYEFMKYGSLEDVLHDPKKAGVKLNWSTRRKIAIGSARGLAFLHHNCSPHIIHRDMKSSNVLLDENLEARVSDFGMARLMSAMDTHLSVSTL 1046 Os-BRI1 869 YCKAGEERLLVYDYMKFGSLEDVLHDRKKIGKKLNWEARRKIAVGAARGLAFLHHNCIPHIIHRDMKSSNVLIDEQLEARVSDFGMARLMSVVDTHLSVSTL 970 Sb-BRI1 868 YCKCGEERLLVYDYMSYGSLEDVLHDRKKVGIKLNWATRKKIAIGAARGLAYLHHNCIPHIIHRDMKSSNVLIDEQLEARVSDFGMARMMSVVDTHLSVSTL 969 Bd-BRI1 870 YCKIGEERLLMYDYMQFGSLEDVLHDRKKIGVKLNWPARRKIAIGAARGLAFLHHNCIPHIIHRDMKSSNVLVDENLEARVSDFGMARMMSVVDTHLSVSTL 971 Hv-BRI1 866 YCKIGEERLLMYDFMKYGSLEDVLHDRKKIGVRLNWAARRKIAIGAARGLAFLHHNCIPHIIHRDMKSSNVLVDENLEARVSDFGMARMMSVVDTHLSVSTL 967 *** *:****:*::*.:********* ** * :*** :*:***:*:*****:***** **************:**:************:**.:********* bri1-115(str) d61-1N/T(str) bri1-101(str) bri1-A1104Y bri1-L1106E bri1-117/118(str) G1048D T988I E1078K A1104Y L1106E D1139N At-BRI1 1047 AGTPGYVPPEYYQSFRCSTKGDVYSYGVVLLELLTGKRPTDSPDFGD-NNLVGWVKQHAKLRISDVFDPELMKEDPALEIELLQHLKVAVACLDDRAWRRPT 1147 Os-BRI1 971 AGTPGYVPPEYYQSFRCTTKGDVYSYGVVLLELLTGKPPTDSADFGEDNNLVGWVKQHTKLKITDVFDPELLKEDPSVELELLEHLKIACACLDDRPSRRPT 1072 Sb-BRI1 970 AGTPGYVPPEYYQSFRCTTKGDVYSYGVVLLELLTGKPPTDSTDFGEDNNLVGWVKQHSKSKVTDVFDPELVKEDPALEVELLEHLKIACLCLHDMPSKRPT 1071 Bd-BRI1 972 AGTPGYVPPEYYQSFRCTTKGDVYSYGVVLLELLTGKPPTDSADFGEDNNLVGWVKLHAKLKIIDVFDPELLKDDPSLELELLEHLKIACACLEDRPTRRPT 1073 Hv-BRI1 968 AGTPGYVPPEYYQSFRCTTKGDVYSYGVVLLELLTGKPPTDSTDFGEDHNLVGWVKMHTKLKITDVFDPELLKDDPTLELELLEHLKIACACLDDRPSRRPT 1069 *****************:******************* **** ***: .******* *:*.:: *******:*:**::*:***:***:* **.* :*** <4OAB | At-BRI1 1148 MVQVMAMFKEIQAGSGIDSQST---IRSIEDGGFSTIEMVDMSIKEVPEGKL 1196 Os-BRI1 1073 MLKVMAMFKEIQAGSTVDSKTSSAAAGSIDEGGYGVLDMP---LREAKEEKD 1121 Sb-BRI1 1072 MLKVMAMFKELQASSAVDSKTS-ECTGAMDDACFGDVEMTT--LKEDKEEKD 1120 Bd-BRI1 1074 MLKVMTMFKEIQAGSTVDSKTSSVATGLSDDVGFGVVDMT---LKEAKEEKD 1122 Hv-BRI1 1070 MLKVMTMFKEIQAGSTVDSKTSSVATGLSDDPGFGVMDMT---LKEAKEEKD 1118 *::**:****:**.* :**::: :: :. ::* ::* * *
Supplemental Figure 8. In Silico Mapping and Detailed Analysis of Conserved Amino Acid
Residues of Hv-BRI1 (for Hordeum vulgare BRASSINOSTEROID-INSENSITIVE 1).
(A) The previously identified uzu1.a mutation (Chono et al., 2003) is present in near-isogenic
lines BW885 (uzu1.a), BW860 (sld1.a + uzu1.a), and BW912 (wst1.c + uzu1.a). The three near-
isogenic lines delimit the inferred Hv-BRI1 position to an interval between SNP markers 1_0373
at 53.28 cM and 1_1394 at 59.77 cM on chromosome 3H (marked with blue, dotted lines). In the
barley genome zipper, the 6.5 cM interval contained 217 gene models including the BRI1
ortholog of Brachypodium, rice, and sorghum. In EnsemblPlant (http://plants.ensembl.org), the
same interval (fingerprint contig_46254 to 46159) contained 150 gene models. The interval
harbors Hv-BRI1 (MLOC_5176), which was mapped on fingerprint contig_48507 at 57.08 cM.
Two additional near-isogenic lines, BW033 (ari.256, renamed uzu1.256) and BW312 (ert-ii.79),
which like BW885 (uzu1.a) demonstrated a brassinosteroid signal-deficient phenotype in a leaf-
unrolling test (Figure 3E), had introgression regions overlapping with the position of Hv-BRI1.
BC, number of backcrosses to cultivar Bowman.
(B) Alignment of polypeptides similar to Hv-BRI1 with indication of amino acid residues
affected by the mutations (light gray). Shown mutation sites (light gray) were either reported for
Arabidopsis (acronym bri1) and rice (d54, d61) or were identified in barley. Reported gain-of-
function mutation sites in Arabidopsis are shown in dark gray; nonsense mutations were

25
excluded. Start and end of the BRI1 protein ectodomain and kinase domain resolved in X-ray
structure analyses (PDB-ID 3RGZ and 4OAB, respectively) are highlighted in yellow. Leucine-
rich repeats (LRR) and other structural elements (Kim and Wang, 2010; Jaillais et al., 2011) are
indicated (red font). The transmembrane domain was predicted using the software TMHMM
(Version 2.0) (Sonnhammer et al., 1998). Inferred positions of functional domains (Mori et al.,
2002; Hong et al., 2005; Kim et al., 2005; Sakamoto and Matsuoka, 2006) are indicated (red).
Numbering of modified amino-acid residues refers to the position in the polypeptide of the
respective plant species. The effect on plant phenotype caused by the mutations is indicated
(mild, int/intermediate, str/strong).

26
SI References
Bishop GJ, Yokota T (2001) Plants steroid hormones, brassinosteroids: current highlights of molecular aspects on their synthesis/metabolism, transport, perception and response. Plant Cell Physiol 42: 114-120
Chono M, Honda I, Zeniya H, Yoneyama K, Saisho D, Takeda K, Takatsuto S, Hoshino T, Watanabe Y (2003) A semidwarf phenotype of barley uzu results from a nucleotide substitution in the gene encoding a putative brassinosteroid receptor. Plant Physiol 133: 1209-1219
Dahleen LS, Vander Wal LJ, Franckowiak JD (2005) Characterization and molecular mapping of genes determining semidwarfism in barley. J Hered 96: 654-662
Druka A, Franckowiak J, Lundqvist U, Bonar N, Alexander J, Houston K, Radovic S, Shahinnia F, Vendramin V, Morgante M, Stein N, Waugh R (2011) Genetic dissection of barley morphology and development. Plant Physiol 155: 617-627
Franckowiak JD, Lundqvist U (2012) Descriptions of barley genetic stocks for 2012. Barley Genet Newsl 42: 36-792
Fujioka S, Noguchi T, Takatsuto S, Yoshida S (1998) Activity of brassinosteroids in the dwarf rice lamina inclination bioassay. Phytochemistry 49: 1841-1848
Gruszka D, Szarejko I, Maluszynski M (2011) Identification of barley DWARF gene involved in brassinosteroid synthesis. Plant Growth Regul 65: 343-358
Hong Z, Ueguchi-Tanaka M, Fujioka S, Takatsuto S, Yoshida S, Hasegawa Y, Ashikari M, Kitano H, Matsuoka M (2005) The rice brassinosteroid-deficient dwarf2 mutant, defective in the rice homolog of Arabidopsis DIMINUTO/DWARF1, is rescued by the endogenously accumulated alternative bioactive brassinosteroid, dolichosterone. Plant Cell 17: 2243-2254
Hong Z, Ueguchi-Tanaka M, Umemura K, Uozu S, Fujioka S, Takatsuto S, Yoshida S, Ashikari M, Kitano H, Matsuoka M (2003) A rice brassinosteroid-deficient mutant, ebisu dwarf (d2), is caused by a loss of function of a new member of cytochrome P450. Plant Cell 15: 2900-2910
International Barley Genome Sequencing Consortium (2012) A physical, genetic and functional sequence assembly of the barley genome. Nature 491: 711-716
Jaillais Y, Hothorn M, Belkhadir Y, Dabi T, Nimchuk ZL, Meyerowitz EM, Chory J (2011) Tyrosine phosphorylation controls brassinosteroid receptor activation by triggering membrane release of its kinase inhibitor. Genes Dev 25: 232-237
Kim BK, Fujioka S, Takatsuto S, Tsujimoto M, Choe S (2008) Castasterone is a likely end product of brassinosteroid biosynthetic pathway in rice. Biochem Biophys Res Commun 374: 614-619
Kim TW, Hwang JY, Kim YS, Joo SH, Chang SC, Lee JS, Takatsuto S, Kim SK (2005) Arabidopsis CYP85A2, a cytochrome P450, mediates the Baeyer-Villiger oxidation of castasterone to brassinolide in brassinosteroid biosynthesis. Plant Cell 17: 2397-2412

27
Kim TW, Wang ZY (2010) Brassinosteroid signal transduction from receptor kinases to transcription factors. Annu Rev Plant Biol 61: 681-704
Mayer KF, Martis M, Hedley PE, Simkova H, Liu H, Morris JA, Steuernagel B, Taudien S, Roessner S, Gundlach H, Kubalakova M, Suchankova P, Murat F, Felder M, Nussbaumer T, Graner A, Salse J, Endo T, Sakai H, Tanaka T, Itoh T, Sato K, Platzer M, Matsumoto T, Scholz U, Dolezel J, Waugh R, Stein N (2011) Unlocking the barley genome by chromosomal and comparative genomics. Plant Cell 23: 1249-1263
Mori M, Nomura T, Ooka H, Ishizaka M, Yokota T, Sugimoto K, Okabe K, Kajiwara H, Satoh K, Yamamoto K, Hirochika H, Kikuchi S (2002) Isolation and characterization of a rice dwarf mutant with a defect in brassinosteroid biosynthesis. Plant Physiol 130: 1152-1161
Ohnishi T, Godza B, Watanabe B, Fujioka S, Hategan L, Ide K, Shibata K, Yokota T, Szekeres M, Mizutani M (2012) CYP90A1/CPD, a brassinosteroid biosynthetic cytochrome P450 of Arabidopsis, catalyzes C-3 oxidation. J Biol Chem 287: 31551-31560
Sakamoto T, Matsuoka M (2006) Characterization of CONSTITUTIVE PHOTOMORPHOGENESIS AND DWARFISM homologs in rice (Oryza sativa L.). J Plant Growth Reg 25: 245-251
Sonnhammer EL, von Heijne G, Krogh A (1998) A hidden Markov model for predicting transmembrane helices in protein sequences. Proc Int Conf Intell Syst Mol Biol 6: 175-182
Vriet C, Russinova E, Reuzeau C (2012) Boosting crop yields with plant steroids. Plant Cell 24: 842-857





























