Embed Size (px)
Citation preview

ORIGINAL PAPER
Plant regeneration in Stone pine (Pinus pinea L.) by somaticembryogenesis
E. Carneros Æ C. Celestino Æ K. Klimaszewska ÆY.-S. Park Æ M. Toribio Æ J. M. Bonga
Received: 21 May 2008 / Accepted: 13 May 2009
� Springer Science+Business Media B.V. 2009
Abstract Regeneration of plants by somatic embryo-
genesis (SE) was achieved in Stone pine (Pinus pinea), one
of the most characteristic tree species of the Mediterranean
ecosystem. The initial explants were megagametophytes
containing zygotic embryos from five selected half-sib
families collected at different dates over 2 consecutive
years. Rates of extrusion and initiation of SE differed in
both years. However, qualitative patterns were very simi-
lar: for most families, the responsive developmental win-
dow was from late cleavage polyembryony to early
cotyledonary stage. The highest overall mean frequencies
of extrusion and SE initiation (7 and 0.9%, respectively, for
the five families and the eight 2006 collections) were
obtained on a modified Litvay’s medium with 9 lM 2,4-D
and 4.5 lM BAP, supplemented with L-glutamine and
casein hydrolysate. Families showed large differences in
frequencies of SE initiation from year to year. Only seven
embryogenic lines were induced in 2005, representing
three of the five families tested, whereas 34 lines from all
the families were obtained in 2006. Proliferation of
embryonal masses (EM) was significantly improved when
they were subcultured after dispersing in liquid medium
and collected on filter paper disks, instead of being sub-
cultured as small clumps. This effect showed a significant
interaction with genotype. Several preconditioning treat-
ments and culture media combinations were tested for
embryo development and maturation. The high prolifera-
tion rate of EM hampered somatic embryo development.
However, up to 42 mature embryos from different lines of
three of the five families were obtained, 23 of them ger-
minated and seven converted into somatic seedlings.
Keywords Conifer � Embryonal mass � Genetic effect �Half-sib families � Somatic embryogenesis �Somatic seedling � Tree breeding
Abbreviations
2,4-D 2,4-Dichlorophenoxyacetic acid
ABA Abscisic acid
AC Activated charcoal
BAP Benzylaminopurine
EM Embryonal mass
FW Fresh weight
MVF Multi-varietal forestry
OP Open-pollinated
PGR Plant growth regulators
PPFD Photosynthetic photon flux density
SE Somatic embryogenesis
se Standard error
Introduction
Domestication of forest tree species is very recent com-
pared with agricultural crops. For centuries, forestry has
E. Carneros � C. Celestino � M. Toribio (&)
Instituto Madrileno de Investigacion y Desarrollo Rural, Agrario
y Alimentario (IMIDRA), Apartado 127, Alcala de Henares,
28800 Madrid, Spain
e-mail: [email protected]
K. Klimaszewska
Natural Resources Canada, Canadian Forest Service, Laurentian
Forestry Centre, P.O. Box 10380, Stn. Sainte-Foy, Quebec,
QC G1V 4C7, Canada
Y.-S. Park � J. M. Bonga
Natural Resources Canada, Canadian Forest Service, Canadian
Wood Fibre Centre, P.O. Box 4000, Fredericton, NB E3B 5P7,
Canada
123
Plant Cell Tiss Organ Cult
DOI 10.1007/s11240-009-9549-3

been more a ‘‘mining’’ than a ‘‘farming’’ activity because,
among various reasons, of the biological attributes of forest
trees—mainly their long generation times and, for most
species, poor ability to propagate vegetatively. This makes
forest genetic improvement programs much more difficult
than their agricultural counterparts. In particular, cloning,
which has been used for centuries in several woody crops
and fruit trees to capture all the components of genetic
variance, is routinely applied to only a few species of the
genus Cryptomeria, Populus, Salix, and Eucalyptus.
However, modern biotechnology is providing new tools in
addition to conventional plant breeding for forest tree
improvement (El-Kassaby 2004). Of these various tools,
somatic embryogenesis (SE) is considered the best vege-
tative propagation tissue culture method for most tree
species, both hardwood and softwood (Merkle and Nairn
2005; Nehra et al. 2005). This vegetative propagation
technique is enabling the development of multi-varietal
forestry (MVF), the use of tested tree varieties in forestry
(Park et al. 2006).
Plant regeneration by SE has been obtained in several
Pinus species (Zoglauer et al. 2003), but to our knowledge
it has not been reported for Stone pine (Pinus pinea L.).
Stone pine is one of the most important tree species in the
Mediterranean ecosystem, and is used for different pur-
poses, such as environmental restoration (afforestation of
coastal areas and continental dunes) and ornamental
planting. However, the most distinctive feature of this
species is the large size of its megagametophytes, which
are full of nutrients and therefore highly prized gastro-
nomically. In Spain, Stone pine covers around 360,000 ha,
which represents about 75% of the species’ total area.
Spain’s breeding program is mainly focused on improving
seed (pine nut) yield (Mutke et al. 2000). As the most
important economical use of this forest species is as a
‘‘fruit tree,’’ the development of clonal rootstocks to which
scions from trees with a high annual nut production can be
grafted (grafted varieties) is of great importance.
Vegetative propagation by rooting of cuttings in Stone
pine is difficult, and problems related to cyclophysis and
topophysis effects usually appear. Regarding micropropa-
gation, there are several reports describing the induction of
adventitious shoots from embryonic explants, and even
rooting of these microcuttings (Capuana and Giannini
1995; Diamantoglou et al. 1990; Garcıa-Ferriz et al. 1994;
Gonzalez et al. 1998; Oliveira et al. 2003; Valdes et al.
2001). However, the advantages of SE—particularly
maintenance of regeneration potential by cryopreservation
while the testing of clones is in progress (Park 2002)—
justify the development of SE protocols for this species.
Somatic embryogenesis is defined as a multi-step
regeneration process starting with formation of embryo-
suspensor masses, followed by embryo formation,
maturation, and plant regeneration. The induction of the
embryogenic response is influenced by several factors,
including basal medium components, exposure to exoge-
nous plant growth regulators (PGR), and culture condi-
tions. However, the features of the initial explant, mainly
the type and developmental stage as well as the genetic
make-up, largely determine the success of the embryogenic
response. Except in a few cases (Lelu et al. 1999), auxins
and cytokinins are necessary for inducing and sustaining
embryogenic tissue proliferation, whereas embryo early
development is triggered by the withdrawal of PGR. Fur-
ther growth of early somatic embryos to mature forms
usually requires the presence of abscisic acid (ABA) (von
Arnold et al. 2002).
For most coniferous species, especially those of the
genus Pinus, immature embryos are the optimal material
for induction of SE (see reviews by Gupta and Grob 1995;
Klimaszewska and Cyr 2002; Lelu-Walter et al. 2006;
Pullman and Johnson 2002; Tautorus et al. 1991; von
Arnold et al. 2002). Moreover, the embryogenic compe-
tence is often restricted to a short yearly time window, and,
therefore, identification of the proper developmental stage
of the immature zygotic embryo to initiate an embryogenic
line is of paramount importance. Nevertheless, somatic
embryos could also be induced sporadically from mature
zygotic embryos of a few pine species, increasing the
availability of suitable explants (Deb and Tandon 2002;
Garin et al. 1998; Radojevic et al. 1999; Tang et al. 2001).
Genotype plays a substantial role in morphogenic
responses. Many papers have reported large variation in SE
initiation frequencies among families of different conifer
species (Cheliak and Klimaszewska 1991; Garin et al.
1998; Miguel et al. 2004; Pullman and Johnson 2002).
Furthermore, detailed studies revealed a variable degree of
genetic control of the different stages of plant regeneration
by SE. Among them, initiation has been shown to be under
strong genetic control, mainly additive in both Picea gla-
uca and Pinus taeda (MacKay et al. 2006; Park et al.
1993).
The main objective of this work was to define methods
to achieve SE in Stone pine, by determining the develop-
mental window in seeds during which induction is feasible,
and considering the influence of culture medium and
genetic effects for the refinement of the SE process.
Materials and methods
Plant material
Open-pollinated (OP) green cones were collected from
several ramets of five selected clones in a Stone pine clonal
bank established at the National Forest Breeding Centre
Plant Cell Tiss Organ Cult
123
‘‘Puerta de Hierro’’ in Madrid (Spanish Ministry of Envi-
ronment). In order to induce somatic embryogenesis they
were collected at about weekly intervals from 23 June to 27
July 2005, and from 7 June to 27 July 2006. Whole cones
were submerged in 70% (v/v) ethanol for 3 min, followed
by immersion in a 10% (v/v) solution of commercial bleach
(sodium hypochlorite: 3.5% active chlorine) for 10 min,
and then air dried and stored in paper bags at 4�C for a
maximum of 2 weeks while all seeds were dissected. For
culture initiation, seeds from each of the five half-sib
families (11, 47, 58, 70, and 88) were removed from the
cones and disinfested in 10% (v/v) commercial bleach with
one drop of Tween� 20 for 15 min, and rinsed three to four
times with sterile water. Then, seed coats were removed.
In order to determine the developmental stage of zygotic
embryos in each collection date, a few zygotic embryos
were excised from randomly selected megagametophytes
collected weekly from the last week of May of both years.
For the first three harvests starting on 7 June of both years
when embryos were at a very early stage, slide samples
were prepared after staining with 1% (w/v) acetocarmine
and microscopically examined. Later collections of both
years were macroscopically examined.
Initiation of somatic embryogenesis
For initiation of embryogenic cultures, three culture media
were tested in 2005 and five culture media, two of which
were common to both years, were tested in 2006.
In the experiment carried out in 2005, cones from the
five families were sampled at six collection dates (23 June,
1, 6, 13, 20, and 27 July). The basal medium, labeled as
mLV2, consisted of a modified Litvay’s (Litvay et al. 1985)
nutrient medium with half-strength macroelements, and
full-strength Fe-EDTA and vitamins. Microelements were
modified as follow: boric acid and manganese and zinc
salts were at full strength; molybdenum, copper, and iodine
salts were at half strength; cobalt salt was reduced to
0.01 mg l-1, and 0.72 mg l-1 NiCl2 was added. Three
combinations of growth regulators at different concentra-
tions were tested: low, L-mLV2 (2.4 lM 2,4-D and 2.2 lM
BAP); medium, M-mLV2 (9 lM 2,4-D and 4.5 lM
BAP); and high, H-mLV2 (15 lM 2,4-D and 4.5 lM
BAP) (Table 1). All media contained sucrose at 10 g l-1.
Overall, 3,500 explants were cultured (six collection
dates 9 five half-sib families 9 three culture media 9 ten
seeds per replicate 9 three to four replicates per
treatment).
In the experiment carried out in 2006, cones from the
same five families were collected on eight collection dates
(7, 14, 23 June and 1, 6, 13, 20, and 27 July). Five culture
media were used; two of them were the same as described
above: M-mLV2 (9 lM 2,4-D and 4.5 lM BAP) and H-
mLV2 (15 lM 2,4-D and 4.5 lM BAP). The third culture
medium, labeled as ultra-high UH-mLV2, was the same
basal medium but supplemented with 20 lM 2,4-D and
4.5 lM BAP. Two additional media, in which the basal
medium was a modified Litvay’s medium with half-
strength macroelements and full-strength microelements,
Fe-EDTA and vitamins (mLV) (Klimaszewska et al. 2001),
were also assayed. They contained PGR at medium, M-
mLV (9.5 lM 2,4-D and 4.5 lM BAP) and high, H-mLV
(15 lM 2,4-D and 4.5 lM BAP) levels (Table 1). These
later two media contained sucrose at 20 g l-1. Overall,
7,200 explants were cultured (eight collection dates 9 five
half-sib families 9 five culture media 9 ten seeds per
replicate 9 three to four replicates per treatment).
Before autoclaving, 1 g l-1 casein hydrolysate (Duchefa
Biochimie B V., The Netherlands) was added to all media,
the pH was adjusted to 5.8, and they were solidified with
4 g l-1 Gelrite� (G1910, Sigma-Aldrich Co., USA). Also,
all media were supplemented with a filter-sterilized solu-
tion of 0.5 g l-1 L-glutamine (G8540, Sigma-Aldrich Co.,
USA) added to the cooled medium.
Megagametophytes with enclosed zygotic embryos were
placed on 90 9 16 mm Petri dishes, each contain-
ing approximately 25 ml of medium. Ten explants per
dish, sealed with Parafilm� (American Can Company,
Table 1 Micronutrient modifications, and plant growth regulators and sucrose content in the culture media used for initiation and maintenance
of embryogenic tissues
Label Salts (mg/l) PGR (lM) Sucrose (g/l)
NiCl2 CoCl2 CuSO4 Na2MoO4 KI 2,4D BAP
UH-mLV2 0.72 0.01 0.25 0.63 2.1 20 4.5 10
H-mLV2 0.72 0.01 0.25 0.63 2.1 15 4.5 10
M-mLV2 0.72 0.01 0.25 0.63 2.1 9 4.5 10
L-mLV2 0.72 0.01 0.25 0.63 2.1 2.4 2.2 10
H-mLV – 0.125 0.5 1.25 4.15 15 4.5 20
M-mLV – 0.125 0.5 1.25 4.15 9.5 4.5 20
Plant Cell Tiss Organ Cult
123

Greenwich, CT), were cultured and kept in darkness at
23 ± 1�C for the duration of each experiment. The cultures
were examined weekly for tissue growth. The megaga-
metophytes were not subcultured during the experiment,
which lasted up to 12 weeks. Three or four replicates (Petri
dishes) per each treatment were arranged, depending on the
availability of megagametophytes.
Extrusion was recorded when zygotic embryos pushed
out of the micropylar end of the megagametophyte, and
proliferated to form an initial embryonal mass (EM).
Explants were considered to have initiated SE when the
tissue showed continuous growth and produced amounts
sufficient for subculture. The number of established lines
was recorded after 4 months of culture under conditions
described below.
Proliferation and maintenance of EM
After 4–12 weeks on initiation media, proliferating EM
were separated from the megagametophyte, and subcul-
tured biweekly onto fresh maintenance medium in darkness
at 23 ± 1�C. The maintenance medium was the M-mLV
medium described above and was used for all embryogenic
lines, regardless of the culture medium used for initiation.
The effect of two procedures on EM proliferation was
studied. In the ‘‘clump procedure,’’ small pieces of tissue
collected at the EM periphery from different embryogenic
lines were subcultured under the maintenance conditions
described above. Initial explants were individual clumps of
about 100 mg of fresh weight. At the beginning of the
treatment, one clump was cultured per 90 mm diameter
Petri dish. Five dishes (replicates) per embryogenic line
were arranged. After each biweekly subculture, the indi-
vidual clumps were divided into four pieces and placed
again onto five new Petri dishes per line.
In the ‘‘dispersal procedure’’ (Klimaszewska and Smith
1997; Lelu-Walter et al. 2006), about 100 mg samples,
collected at the EM periphery, were suspended in 10 ml of
liquid M-mLV medium, vigorously shaken to break up the
tissue into a fine suspension, and poured onto a filter paper
disk (80 g/m2, 60–68 lm pore, Filter-Lab, ANOIA; Bar-
celona, Spain) in a Buchner funnel. Low-pressure pulse
was applied to drain the liquid, and the filter paper with
attached cells was placed on the surface of fresh M-mLV
semi-solid medium in a 90 mm diameter Petri dish. Five
filter paper disks with embryogenic tissue were arranged in
five dishes (replicates) per line and subcultured biweekly.
Five embryogenic lines chosen from different families
(7F11 and 2F70 initiated in 2005; 2F47, 9F58 and 6F88
initiated in 2006) underwent both procedures to study the
effect of procedure and genotype. Growth rate of tissue in
each replicate was recorded after 2, 4, and 6 weeks in
darkness at 23 ± 1�C. To calculate growth rate, fresh
weight of EM was determined at the beginning of each
subculture by subtracting the weight of a fresh medium-
containing dish from the weight of the same fresh medium-
containing dish but including the embryogenic tissue. The
dependent variable was the relative fresh weight increa-
se(s) after each period, defined as the fresh weight at the
end of that period minus the initial fresh weight, divided by
the initial fresh weight.
Maturation of somatic embryos
Several randomly chosen embryogenic cell lines from
different families were used to determine their capacity to
produce mature somatic embryos. The ages of the cultures
ranged from 1 to 10 months from the onset of initiation.
Two preconditioning procedures with several maturation
treatments were tested based on previous reports (Lelu
et al. 1999; Klimaszewska et al. 2001; Bonga 2004; Lelu-
Walter et al. 2006).
In the first experiment, carried out in 2006, four estab-
lished embryogenic lines (7F11, 13F58 and 2F70 initiated
in 2005; 5F47 initiated in 2006) were preconditioned by
subculturing EM on UL-mLV medium that was made up of
mLV medium with reduced PGR concentrations (0.24 lM
2,4-D and 0.22 lM BAP) with 20 g l-1 sucrose and
4 g l-1 Gelrite, for 1 month with biweekly subculture.
Then, a suspension of embryogenic tissue at a concentra-
tion of 10 g FW of EM per liter was prepared with the
same medium used for preconditioning but lacking gelling
agent, PGR, and glutamine, and supplemented with
10 g l-1 of activated charcoal (AC, C6289, Sigma-Aldrich
Co., USA). The dispersed tissue was collected by pouring
5 ml of the suspension onto filter paper disks as previously
described, and cultured on the different maturation media.
The maturation treatments were carried out according to a
complete factorial design in which the effects of nutrient
medium, AFC medium (Bonga 2004) vs. mLV; type of
sugar, maltose vs. sucrose both at 60 g l-1; gelling agent,
20 g l-1 Plantagar� (S1000, B&VS.R.L., Italy) vs. 10 g l-1
Gelrite; and ABA concentrations (80, 121, and 161 lM)
were tested. Therefore, considering the four genotypes, 96
treatments were included in the experiment. Cultures were
not transferred to fresh medium for the duration of the
experiment.
In the second experiment, carried out in 2006, seven
embryogenic lines (7F11, 13F58 and 2F70 initiated in
2005; 2F47, 5F47, 9F58 and 2F88 initiated in 2006) were
tested. The preconditioning of the EM was carried out as
described above, but all components of the culture med-
ium, except for PGR and Gelrite, were reduced by half.
Then, the suspension in liquid medium lacking PGR and
glutamine was prepared as above, but with the half-strength
mLV medium, and with or without 10 g l-1 of AC. Five
Plant Cell Tiss Organ Cult
123

milliliters of the suspensions (50 mg FW) were poured
onto filter paper disks and placed on the half-strength mLV
medium without PGR, solidified with 4 g l-1 Gelrite and
with or without 10 g l-1 of AC, for 1 week. Disks with
tissues from suspension prepared with AC were cultured on
medium without this compound, and vice versa. Matura-
tion was performed on mLV medium supplemented with
60 g l-1 sucrose and 10 g l-1 Gelrite. Two ways of sup-
plying ABA, continuous vs. pulse, at two concentrations,
80 and 121 lM, and 121 and 200 lM, respectively, were
tested. In total, 56 treatments were performed. Continuous
supply was accomplished by maintaining the paper disks
with EM on the ABA-containing media for the whole
length of the experiment, with monthly subcultures. In the
pulse treatments, cultures were on the ABA containing
media for 7 days, and then were transferred to PGR-free
medium for the whole length of the experiment, also with
monthly subcultures.
In both maturation procedures, cultures were kept in
darkness at 23 ± 1�C, and the treatments lasted 16 weeks.
Each filter paper disk with 50 mg FW of EM placed into a
90-mm diameter Petri dish with maturation medium was
considered a replicate, and five replicates per treatment
were arranged. The pH of the media was adjusted to
5.8 before autoclaving. Solutions of ABA and glutamine
were filter sterilized and added to the cooling autoclaved
medium.
Maturation ability of the different treatments was
recorded as the number of morphologically normal somatic
embryos (opaque, presence of cotyledons, embryonal root
caps, and smooth hypocotyls) obtained from 250 mg FW
of EM, per embryogenic line within each treatment.
Embryo germination and conversion to plant
Mature somatic embryos were picked from filter paper
disks after 4–6 months on maturation media and placed
horizontally, all with the cotyledons facing in one direc-
tion, on medium for germination. This medium consisted
of AFC nutrient medium without PGR, supplemented with
30 g l-1 sucrose and 6 g l-1 Plantagar. Petri dishes
(60 9 16 mm) were filled with 10 ml of this semisolid
medium. For pre-germination treatment, embryos were
stored in darkness at 4�C for 1 month. Then, cultures were
transferred to a growth chamber at 23 ± 1�C, with a 16-h
photoperiod and photosynthetic photon flux density
(PPFD) of 17–20 lmol m-2 s-1 provided by mixed Syl-
vania Gro-lux and Philips cool-white fluorescent tubes for
1 month. Petri dishes were tilted to a slanted position at an
angle of approximately 408, and embryos positioned with
their embryonal root caps pointing downward. When
somatic embryos started to germinate, they were asepti-
cally transplanted for further growth to culture tubes ‘‘De
Wit’’ (W1607, Duchefa Biochemie 2003) filled with 15 ml
liquid AFC medium without organic ammonium and PGR,
with 10 g l-1 sucrose and sterile perlite (2 mm particle
size) as support. The developing somatic seedlings were
placed in a growth chamber at 23 ± 1�C, and exposed to
PPFD of 120–180 lmol m-2 s-1 for a 16-h photoperiod.
After the plumule reached 1 cm, plantlets were trans-
planted ex vitro to 240 ml forest containers filled with
substrate (peat: perlite, 3:1, v/v) and the slow release fer-
tilizer Osmocote� at 3 g l-1 for acclimatization. They
were placed in a growth chamber at 23 ± 1�C, under 80%
relative humidity, and a 16-h photoperiod with PPDF of
120–180 lmol m-2 s-1.
Statistical analysis
Effects of crop year, collection date, family, culture med-
ium, genotype, and maintenance culture procedure were
evaluated by one- or two-way ANOVA, using Statistica for
Windows 5.1 software (StatSoft, Tulsa, OK). Data on fre-
quency of extrusion and initiation of embryogenic lines
were transformed by arcsin H(%). Relative growth data of
EM were transformed using the normal logarithm. Mean
values are shown with standard errors (se). Significant
differences between means were determined by Duncan’s
test at a significance level of 5%.
Results
Developmental stage of the zygotic embryos
Early observations showed that megagametophytes col-
lected in May of both years were translucent with visible
archegonia, but cell division of a developing proembryo
was not detected. We observed little variation in the
developmental stage of zygotic embryos for the same
collection dates between the 2 years. However, immature
embryos collected on 23 June 2005 were slightly more
advanced in development than on the same date in 2006,
although dominance was not detected yet.
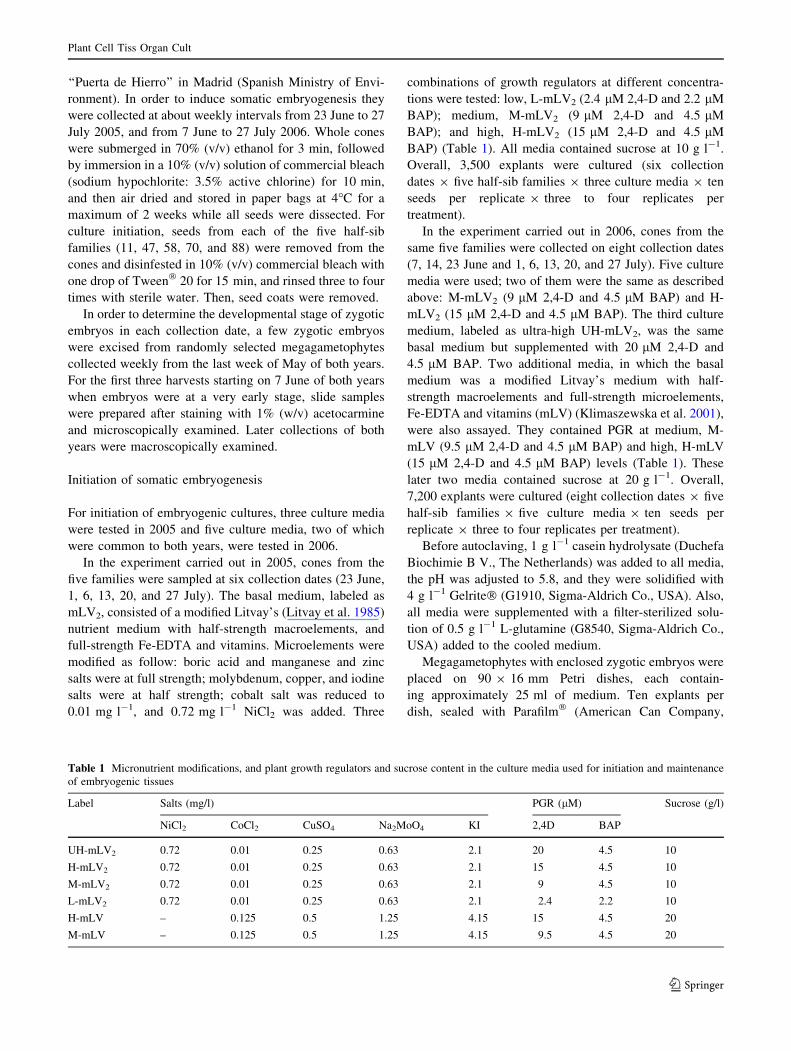
On 7 June of both years, megagametophytes were
slightly opaque (Fig. 1a) and early cell division of the
proembryo was noticed (pre-cleavage stage) (Fig. 1b). On
14 June, megagametophytes were opaque (Fig. 1c) and
most embryos were at the beginning of the cleavage stage
(Fig. 1d), with the terminal embryonal cells and subter-
minal suspensor cells placed deeper inside the female
gametophyte (Fig. 1c). On 23 June, cleavage polyembry-
ony was evident (Fig. 1e, f). At the first collection date in
July, embryos were at the post-cleavage stage, and the
beginning of dominance could be observed (Fig. 1g). On 6
July, the development of the dominant embryo occurred,
Plant Cell Tiss Organ Cult
123
showing the establishment of polarity, and the subordinate
embryos aborted (Fig. 1h). By mid-July, embryo domi-
nance was well established and the zygotic embryos
entered the precotyledonary stage, maintaining long sus-
pensors (Fig. 1i). At the last collection dates, on 20 and 27
July, completely developed embryos with elongating
cotyledons that were attached to remaining suspensor cells,
could be observed (Fig. 1j, k).
Extrusion and initiation of SE
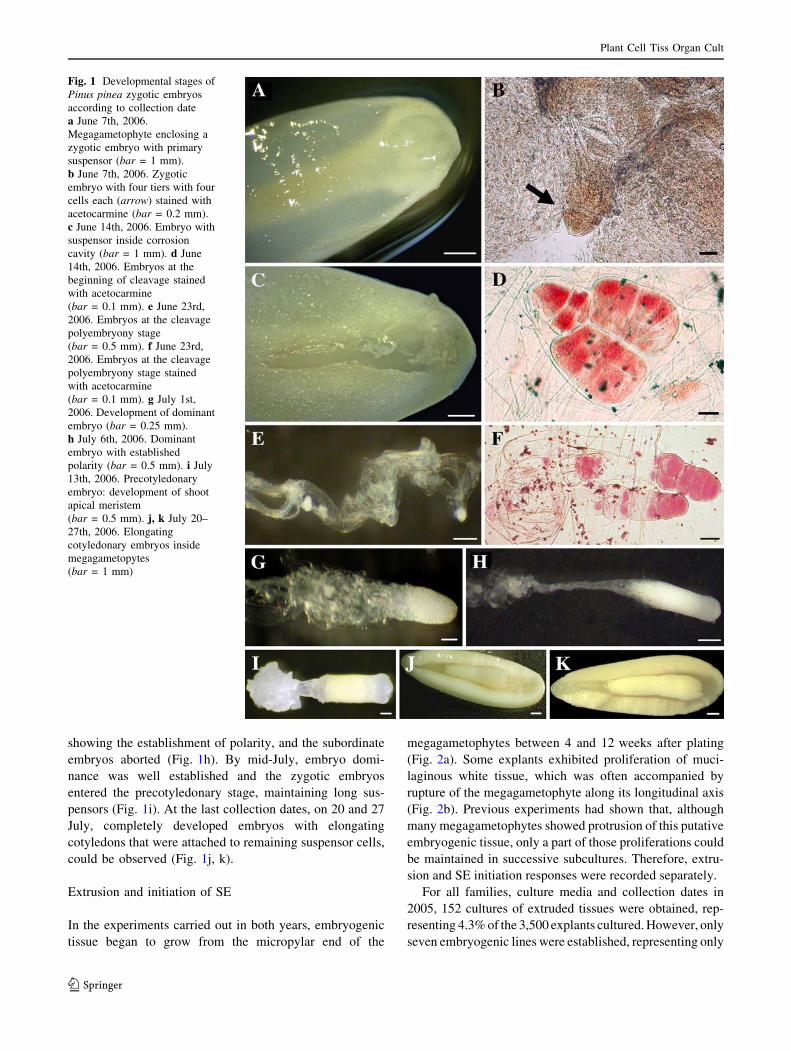
In the experiments carried out in both years, embryogenic
tissue began to grow from the micropylar end of the
megagametophytes between 4 and 12 weeks after plating
(Fig. 2a). Some explants exhibited proliferation of muci-
laginous white tissue, which was often accompanied by
rupture of the megagametophyte along its longitudinal axis
(Fig. 2b). Previous experiments had shown that, although
many megagametophytes showed protrusion of this putative
embryogenic tissue, only a part of those proliferations could
be maintained in successive subcultures. Therefore, extru-
sion and SE initiation responses were recorded separately.
For all families, culture media and collection dates in
2005, 152 cultures of extruded tissues were obtained, rep-
resenting 4.3% of the 3,500 explants cultured. However, only
seven embryogenic lines were established, representing only
Fig. 1 Developmental stages of
Pinus pinea zygotic embryos
according to collection date
a June 7th, 2006.
Megagametophyte enclosing a
zygotic embryo with primary
suspensor (bar = 1 mm).
b June 7th, 2006. Zygotic
embryo with four tiers with four
cells each (arrow) stained with
acetocarmine (bar = 0.2 mm).
c June 14th, 2006. Embryo with
suspensor inside corrosion
cavity (bar = 1 mm). d June
14th, 2006. Embryos at the
beginning of cleavage stained
with acetocarmine
(bar = 0.1 mm). e June 23rd,
2006. Embryos at the cleavage
polyembryony stage
(bar = 0.5 mm). f June 23rd,
2006. Embryos at the cleavage
polyembryony stage stained
with acetocarmine
(bar = 0.1 mm). g July 1st,
2006. Development of dominant
embryo (bar = 0.25 mm).
h July 6th, 2006. Dominant
embryo with established
polarity (bar = 0.5 mm). i July
13th, 2006. Precotyledonary
embryo: development of shoot
apical meristem
(bar = 0.5 mm). j, k July 20–
27th, 2006. Elongating
cotyledonary embryos inside
megagametopytes
(bar = 1 mm)
Plant Cell Tiss Organ Cult
123

0.2% of the initial explants. In 2006, 406 cultures of extruded
tissues were produced, representing 5.6% of the 7,200 cul-
tured explants, and 34 embryogenic lines were established,
representing 0.5% of the initial explants. Therefore, explants
were more responsive in 2006 than in 2005. When data from
media that were common to both years were considered, a
significant effect of year of collection was observed on both
extrusion (4.8 and 8.1% in 2005 and 2006, respectively,
P = 0.000) and SE initiation (0.3 and 0.6% in 2005 and
2006, respectively, P = 0.042).
Regarding the influence of culture medium, a significant
effect on the frequency of initiation was detected in the
2006 experiment (P = 0.046). This was mostly due to
basal medium composition, because mLV2 media rendered
frequencies that were three to four times higher than those
of mLV media (Table 2). However, 2,4-D concentrations
in the range of 9–20 lM did not affect either extrusion or
initiation in both years. Best results were observed with
M-mLV2 medium in 2006 that gave a frequency of initi-
ation nine times higher than obtained in 2005. However,
H-mLV2 medium with increased 2,4-D concentrations per-
formed equally in 2005 and 2006 experiments (Table 2).
The family 9 culture medium interaction was not
significant.
Fig. 2 Somatic embryogenesis in Pinus pinea. a Extrusion of putative
embryogenic tissues from a megagametophyte containing a developing
dominant zygotic embryo collected July 1st, 2006 (bar = 2 mm).
b Extrusion of putative embryogenic tissues from a cotyledonary zygotic
embryo collected July 20th, 2006 (bar = 2 mm). c Embryogenic tissue
growing on maintenance medium as clumps (bar = 4 mm). d Embryo-
genic tissue growing on maintenance medium dispersed on paper disks
(bar = 5 mm). e Early developmental stage of somatic embryos
growing in clumps (bar = 1 mm) f Early development of somatic
embryos growing dispersed on disks (bar = 0.1 mm). g Embryogenic
culture showing developing somatic embryos (bar = 0.5 mm).
h Maturation of somatic embryos in AFC medium with 121 lM ABA,
using activated charcoal in close contact with embryogenic tissue
(bar = 5 mm). i–k Different developmental stages of cotyledonary
somatic embryos (bar = 0.5 mm). l Germination of somatic embryo
(bar = 2 mm). m Somatic seedlings growing in perlite moistened with
liquid medium (Culture tube maximum diameter = 27 mm). n Accli-
matized somatic seedlings in forest containers (Container maximum
diameter = 52 mm)
Plant Cell Tiss Organ Cult
123
Although mLV2 medium was best suited for initiation,
preliminary trials showed that some embryogenic lines
gradually declined when they were subcultured in the same
initiation medium, producing slightly darker and less vig-
orously growing tissue. This decline was not observed when
the M-mLV medium was used. Therefore, all embryogenic
lines obtained from the several initiation media, were sub-
sequently subcultured on mLV medium supplemented with
the lower PGR concentrations for maintenance (9.5 lM
2.4D and 4.5 lM BAP). When the established embryogenic
lines were subcultured on growth regulator-free medium,
EM did not proliferate (data not shown).
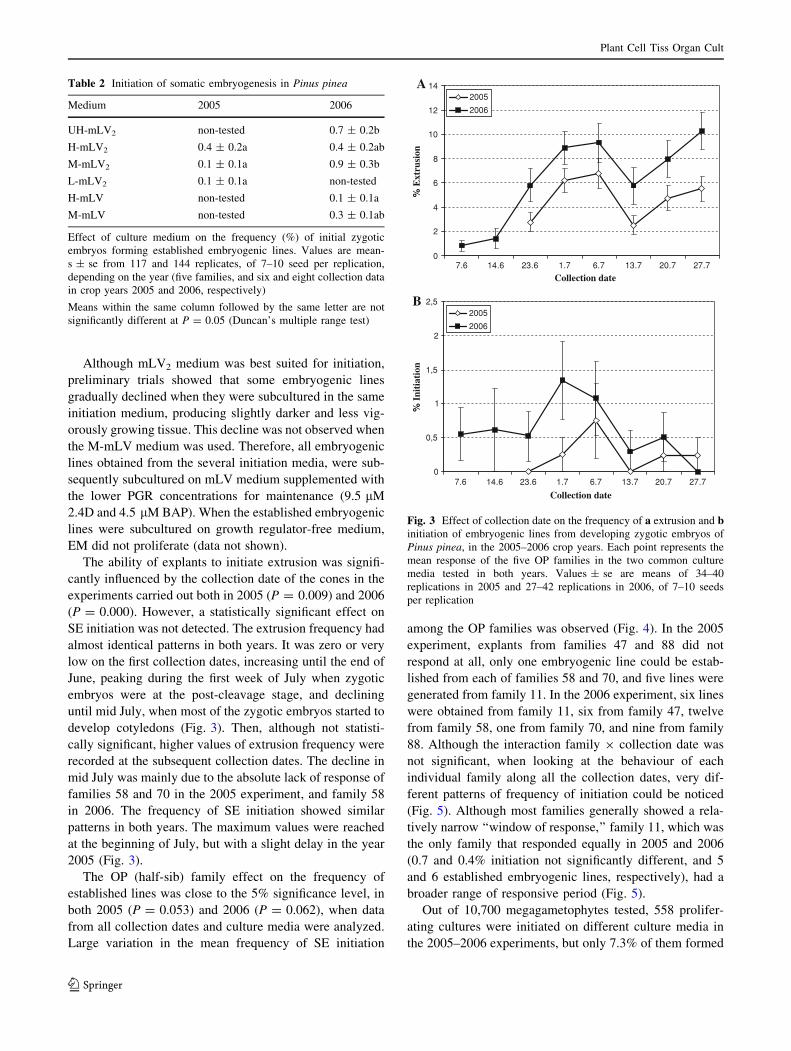
The ability of explants to initiate extrusion was signifi-
cantly influenced by the collection date of the cones in the
experiments carried out both in 2005 (P = 0.009) and 2006
(P = 0.000). However, a statistically significant effect on
SE initiation was not detected. The extrusion frequency had
almost identical patterns in both years. It was zero or very
low on the first collection dates, increasing until the end of
June, peaking during the first week of July when zygotic
embryos were at the post-cleavage stage, and declining
until mid July, when most of the zygotic embryos started to
develop cotyledons (Fig. 3). Then, although not statisti-
cally significant, higher values of extrusion frequency were
recorded at the subsequent collection dates. The decline in
mid July was mainly due to the absolute lack of response of
families 58 and 70 in the 2005 experiment, and family 58
in 2006. The frequency of SE initiation showed similar
patterns in both years. The maximum values were reached
at the beginning of July, but with a slight delay in the year
2005 (Fig. 3).
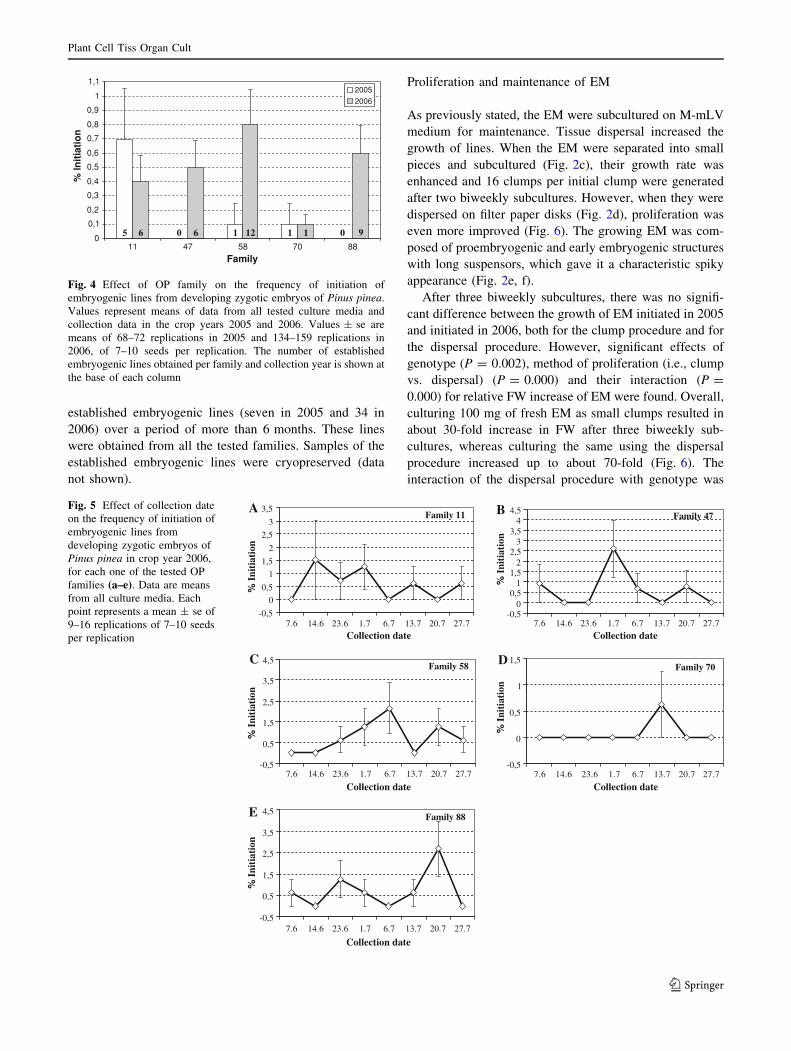
The OP (half-sib) family effect on the frequency of
established lines was close to the 5% significance level, in
both 2005 (P = 0.053) and 2006 (P = 0.062), when data
from all collection dates and culture media were analyzed.
Large variation in the mean frequency of SE initiation
among the OP families was observed (Fig. 4). In the 2005
experiment, explants from families 47 and 88 did not
respond at all, only one embryogenic line could be estab-
lished from each of families 58 and 70, and five lines were
generated from family 11. In the 2006 experiment, six lines
were obtained from family 11, six from family 47, twelve
from family 58, one from family 70, and nine from family
88. Although the interaction family 9 collection date was
not significant, when looking at the behaviour of each
individual family along all the collection dates, very dif-
ferent patterns of frequency of initiation could be noticed
(Fig. 5). Although most families generally showed a rela-
tively narrow ‘‘window of response,’’ family 11, which was
the only family that responded equally in 2005 and 2006
(0.7 and 0.4% initiation not significantly different, and 5
and 6 established embryogenic lines, respectively), had a
broader range of responsive period (Fig. 5).
Out of 10,700 megagametophytes tested, 558 prolifer-
ating cultures were initiated on different culture media in
the 2005–2006 experiments, but only 7.3% of them formed
Table 2 Initiation of somatic embryogenesis in Pinus pinea
Medium 2005 2006
UH-mLV2 non-tested 0.7 ± 0.2b
H-mLV2 0.4 ± 0.2a 0.4 ± 0.2ab
M-mLV2 0.1 ± 0.1a 0.9 ± 0.3b
L-mLV2 0.1 ± 0.1a non-tested
H-mLV non-tested 0.1 ± 0.1a
M-mLV non-tested 0.3 ± 0.1ab
Effect of culture medium on the frequency (%) of initial zygotic
embryos forming established embryogenic lines. Values are mean-
s ± se from 117 and 144 replicates, of 7–10 seed per replication,
depending on the year (five families, and six and eight collection data
in crop years 2005 and 2006, respectively)
Means within the same column followed by the same letter are not
significantly different at P = 0.05 (Duncan’s multiple range test)
0
2
4
6
8
10
12
14
Collection date
% E
xtru
sion
2005
2006
0
0,5
1
1,5
2
2,5
7.6 14.6 23.6 1.7 6.7 13.7 20.7 27.7
7.6 14.6 23.6 1.7 6.7 13.7 20.7 27.7
Collection date
% I
niti
atio
n
2005
2006
A
B
Fig. 3 Effect of collection date on the frequency of a extrusion and binitiation of embryogenic lines from developing zygotic embryos of
Pinus pinea, in the 2005–2006 crop years. Each point represents the
mean response of the five OP families in the two common culture
media tested in both years. Values ± se are means of 34–40
replications in 2005 and 27–42 replications in 2006, of 7–10 seeds
per replication
Plant Cell Tiss Organ Cult
123
established embryogenic lines (seven in 2005 and 34 in
2006) over a period of more than 6 months. These lines
were obtained from all the tested families. Samples of the
established embryogenic lines were cryopreserved (data
not shown).
Proliferation and maintenance of EM
As previously stated, the EM were subcultured on M-mLV
medium for maintenance. Tissue dispersal increased the
growth of lines. When the EM were separated into small
pieces and subcultured (Fig. 2c), their growth rate was
enhanced and 16 clumps per initial clump were generated
after two biweekly subcultures. However, when they were
dispersed on filter paper disks (Fig. 2d), proliferation was
even more improved (Fig. 6). The growing EM was com-
posed of proembryogenic and early embryogenic structures
with long suspensors, which gave it a characteristic spiky
appearance (Fig. 2e, f).
After three biweekly subcultures, there was no signifi-
cant difference between the growth of EM initiated in 2005
and initiated in 2006, both for the clump procedure and for
the dispersal procedure. However, significant effects of
genotype (P = 0.002), method of proliferation (i.e., clump
vs. dispersal) (P = 0.000) and their interaction (P =
0.000) for relative FW increase of EM were found. Overall,
culturing 100 mg of fresh EM as small clumps resulted in
about 30-fold increase in FW after three biweekly sub-
cultures, whereas culturing the same using the dispersal
procedure increased up to about 70-fold (Fig. 6). The
interaction of the dispersal procedure with genotype was
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
1,1
11 47 58 70 88
Family
% In
itia
tio
n2005
2006
0 0 5 1 16 6 9112
Fig. 4 Effect of OP family on the frequency of initiation of
embryogenic lines from developing zygotic embryos of Pinus pinea.
Values represent means of data from all tested culture media and
collection data in the crop years 2005 and 2006. Values ± se are
means of 68–72 replications in 2005 and 134–159 replications in
2006, of 7–10 seeds per replication. The number of established
embryogenic lines obtained per family and collection year is shown at
the base of each column
Family 11
-0,5
0
0,5
1
1,5
2
2,5
3
3,5
Collection date
% I
niti
atio
n
Family 47
-0,50
0,51
1,52
2,53
3,54
4,5
Collection date
% I
niti
atio
n
Family 58
-0,5
0,5
1,5
2,5
3,5
4,5
Collection date
% I
niti
atio
n
Family 70
-0,5
0
0,5
1
1,5
Collection date
% I
niti
atio
n
Family 88
-0,5
0,5
1,5
2,5
3,5
4,5
7.6 14.6 23.6 1.7 6.7 13.7 20.7 27.7 7.6 14.6 23.6 1.7 6.7 13.7 20.7 27.7
7.6 14.6 23.6 1.7 6.7 13.7 20.7 27.7 7.6 14.6 23.6 1.7 6.7 13.7 20.7 27.7
7.6 14.6 23.6 1.7 6.7 13.7 20.7 27.7
Collection date
% I
niti
atio
n
A B
C D
E
Fig. 5 Effect of collection date
on the frequency of initiation of
embryogenic lines from
developing zygotic embryos of
Pinus pinea in crop year 2006,
for each one of the tested OP
families (a–e). Data are means
from all culture media. Each
point represents a mean ± se of
9–16 replications of 7–10 seeds
per replication
Plant Cell Tiss Organ Cult
123
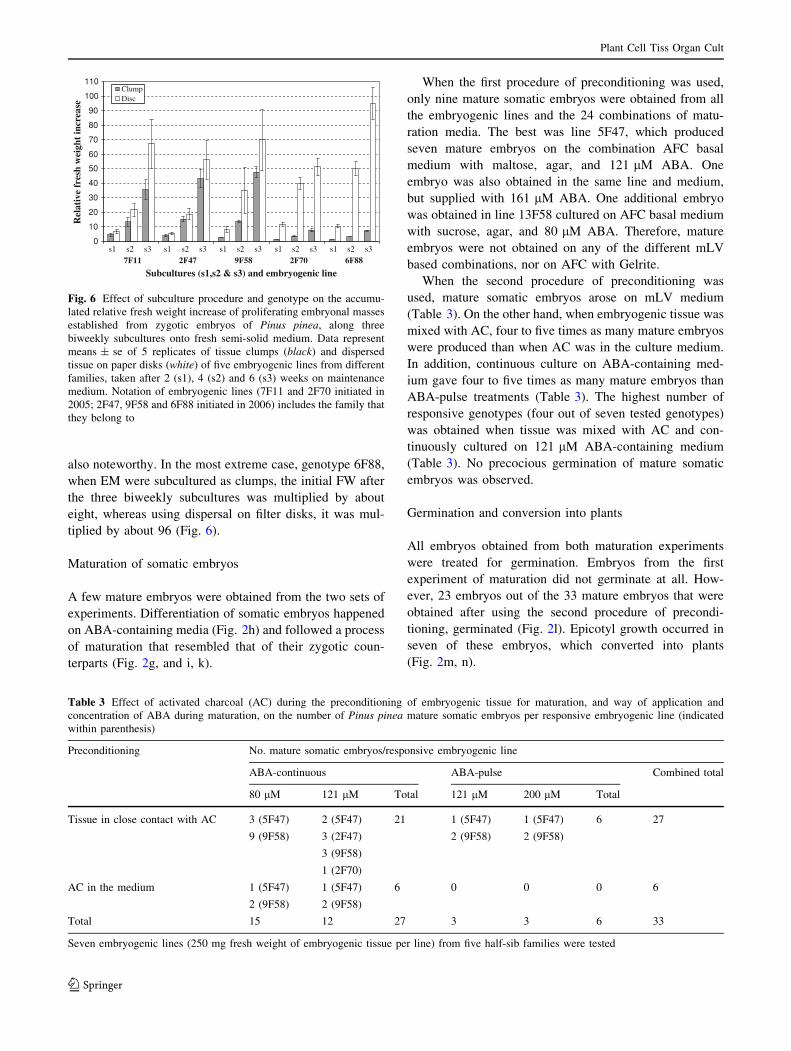
also noteworthy. In the most extreme case, genotype 6F88,
when EM were subcultured as clumps, the initial FW after
the three biweekly subcultures was multiplied by about
eight, whereas using dispersal on filter disks, it was mul-
tiplied by about 96 (Fig. 6).
Maturation of somatic embryos
A few mature embryos were obtained from the two sets of
experiments. Differentiation of somatic embryos happened
on ABA-containing media (Fig. 2h) and followed a process
of maturation that resembled that of their zygotic coun-
terparts (Fig. 2g, and i, k).
When the first procedure of preconditioning was used,
only nine mature somatic embryos were obtained from all
the embryogenic lines and the 24 combinations of matu-
ration media. The best was line 5F47, which produced
seven mature embryos on the combination AFC basal
medium with maltose, agar, and 121 lM ABA. One
embryo was also obtained in the same line and medium,
but supplied with 161 lM ABA. One additional embryo
was obtained in line 13F58 cultured on AFC basal medium
with sucrose, agar, and 80 lM ABA. Therefore, mature
embryos were not obtained on any of the different mLV
based combinations, nor on AFC with Gelrite.
When the second procedure of preconditioning was
used, mature somatic embryos arose on mLV medium
(Table 3). On the other hand, when embryogenic tissue was
mixed with AC, four to five times as many mature embryos
were produced than when AC was in the culture medium.
In addition, continuous culture on ABA-containing med-
ium gave four to five times as many mature embryos than
ABA-pulse treatments (Table 3). The highest number of
responsive genotypes (four out of seven tested genotypes)
was obtained when tissue was mixed with AC and con-
tinuously cultured on 121 lM ABA-containing medium
(Table 3). No precocious germination of mature somatic
embryos was observed.
Germination and conversion into plants
All embryos obtained from both maturation experiments
were treated for germination. Embryos from the first
experiment of maturation did not germinate at all. How-
ever, 23 embryos out of the 33 mature embryos that were
obtained after using the second procedure of precondi-
tioning, germinated (Fig. 2l). Epicotyl growth occurred in
seven of these embryos, which converted into plants
(Fig. 2m, n).
0
10
20
30
40
50
60
70
80
90
100
110
7F11 2F47 9F58 2F70 6F88
Subcultures (s1,s2 & s3) and embryogenic line
Rel
ativ
e fr
esh
wei
ght
incr
ease
Clump
Disc
s3s2 s3s2s1 s1s2 s3s1s2 s3s1s2 s3s1
Fig. 6 Effect of subculture procedure and genotype on the accumu-
lated relative fresh weight increase of proliferating embryonal masses
established from zygotic embryos of Pinus pinea, along three
biweekly subcultures onto fresh semi-solid medium. Data represent
means ± se of 5 replicates of tissue clumps (black) and dispersed
tissue on paper disks (white) of five embryogenic lines from different
families, taken after 2 (s1), 4 (s2) and 6 (s3) weeks on maintenance
medium. Notation of embryogenic lines (7F11 and 2F70 initiated in
2005; 2F47, 9F58 and 6F88 initiated in 2006) includes the family that
they belong to
Table 3 Effect of activated charcoal (AC) during the preconditioning of embryogenic tissue for maturation, and way of application and
concentration of ABA during maturation, on the number of Pinus pinea mature somatic embryos per responsive embryogenic line (indicated
within parenthesis)
Preconditioning No. mature somatic embryos/responsive embryogenic line
ABA-continuous ABA-pulse Combined total
80 lM 121 lM Total 121 lM 200 lM Total
Tissue in close contact with AC 3 (5F47)
9 (9F58)
2 (5F47) 21 1 (5F47)
2 (9F58)
1 (5F47)
2 (9F58)
6 27
3 (2F47)
3 (9F58)
1 (2F70)
AC in the medium 1 (5F47) 1 (5F47) 6 0 0 0 6
2 (9F58) 2 (9F58)
Total 15 12 27 3 3 6 33
Seven embryogenic lines (250 mg fresh weight of embryogenic tissue per line) from five half-sib families were tested
Plant Cell Tiss Organ Cult
123

Discussion
The initiation of SE from megagametophytes containing
zygotic embryos of Stone pine was achieved at very low
frequency, with an overall mean around 0.5%. Although
the first papers that reported conifer SE in other pines, such
as P. strobus (Finer et al. 1989), pointed out that initiation
was not a limiting factor, the frequencies of initiation for
most species described in these early studies were also very
low (Becwar et al. 1990; Percy et al. 2000). To avoid
problems of genetic erosion in breeding programs, it is
desirable to capture as many genotypes as possible. For
pine species, new protocols are currently being developed
to improve initiation frequencies (Pullman and Johnson
2002), and therefore increased efficiency of SE in Stone
pine is expected. For example, with an optimized protocol,
the initiation frequency of P. strobus was increased to an
overall frequency of 76% (Klimaszewska et al. 2001).
Although most of the extrusions from the megagame-
tophyte tissue could be established as embryogenic lines in
some species (Miguel et al. 2004; Salajova and Salaj 2005),
differences between frequencies of extrusion and initiation
of SE were observed in other species (Percy et al. 2000;
Pullman and Johnson 2002). This poses a problem for
application of SE in some pine species in multivarietal
forestry, because it limits the number of captured geno-
types. However, extrusion is necessary for SE initiation. In
P. taeda, one-fifth of the extrusions led to initiation of
embryogenic lines (MacKay et al. 2006). In the case of
Stone pine, only less than 5% of extrusions in the 2005
experiment, and 8% in the 2006 experiment became
established as embryogenic lines. However, these impor-
tant differences between proportions of different years of
collection indicated that some factors control the viability
of extruded tissues. Therefore, their identification and
manipulation could improve the initiation of SE. In
P. taeda, EM extrusion and initiation were partly under
different types of genetic control (MacKay et al. 2006).
The presence of the female megagametophyte in the cul-
ture may be related to this effect, as discussed below.
Significant differences have been found in initiation
frequencies between collection years. These differences
have also been reported for other pine species, such as P.
pinaster (Miguel et al. 2004) and P. nigra (Salajova and
Salaj 2005). Our results clearly indicated that differences
between both years were maintained on all collection dates.
Differences in initiation of EM in P. taeda have been
ascribed to stimulatory compounds produced by the female
gametophyte (Pullman and Johnson 2002). Some authors
have also pointed out that observed maternal effects on the
initiation of embryogenic lines could be linked to the
presence of the megagametophyte in culture (MacKay
et al. 2006). Stone pine has one of the biggest
megagametophytes among conifers and, therefore, its
possible influence deserves to be studied.
Experiments on media composition in this study were
not broad enough to draw conclusions about their effect on
the initiation of embryogenic lines. However, it seems that
concentrations of 2,4-D and BAP, supplied within the
range commonly used in other pine species, were less
important than basal medium formulation. A similar
response was observed in P. pinaster, in which basal
medium was more important than 2,4-D concentrations in
the range of 2.4–13.5 lM (Park et al. 2006). However the
effect of 2,4-D concentrations could be influenced by the
endogenous content of regulators, which could explain the
observed differences between years. Although some stud-
ies reported that nutrient media composition was not a
determinant to achieve SE in some conifers, Stone pine
seems to belong to the group of species that are nutrient
media-dependent (Li et al. 1998; Miguel et al. 2004; Park
et al. 2006). Preliminary tests with several culture media
formulations commonly used for conifers, failed with
Stone pine. Using a nickel-containing mLV medium with
reduced cobalt, which improved initiation in recalcitrant
species like P. banksiana (Park et al. 2006), and modified
micronutrients led to positive responses in Stone pine. This
medium was superior to the standard mLV medium (Kli-
maszewska et al. 2001) when both were compared in the
2006 experiments. Although the presence of nickel and a
reduced level of cobalt and other micronutrients is proba-
bly beneficial, the presence of sucrose at different con-
centrations cannot be ignored. Therefore, more research is
needed to determine the role of medium components to
further improve initiation frequencies. However, it is
known that the mineral content and PGR supplement of
media used for initiation is not be the best medium for
maintaining embryo cleavage in proliferating EM. This
was also the case in Stone pine, in which the standard mLV
medium with 9.5 lM 2,4-D, 4.5 lM BAP and 20 g/l
sucrose, was the best medium for maintaining embryogenic
lines.
Selection of competent explants is critical for inducing
SE in conifers (Tautorus et al. 1991). The highest fre-
quencies of SE induction in Stone pine occurred when
zygotic embryos were in the post-cleavage polyembryony
to cotyledonary stages. These developmental stages are
common to other pine species, in which the precotyledo-
nary stage was the most responsive (Klimaszewska et al.
2001; Lelu et al. 1999; Salajova and Salaj 2005). It is
noteworthy that the developmental window for embryo-
genic response in Stone pine was almost identical to that
reported for P. pinaster, another Mediterranean pine spe-
cies (Miguel et al. 2004). In both species, maximal fre-
quencies of SE initiation occurred between the end of June
and the first week of July. Interestingly, the decrease in the
Plant Cell Tiss Organ Cult
123

percentage of established lines from 6 to 13 July, followed
by a slight recovery, which was observed for both years of
this study, was also reported for P. pinaster (Miguel et al.
2004).
It is well established that the several phases of plant
regeneration by SE are under genetic control (Park et al.
1993, 1994). Significant differences in somatic embryo-
genic response among families of conifers such as Picea
mariana (Cheliak and Klimaszewska 1991), Pinus strobus
(Garin et al. 1998; Klimaszewska et al. 2001), P. monticola
(Percy et al. 2000), P. pinaster (Miguel et al. 2004) P.
taeda (MacKay et al. 2006), and P. sylvestris (Lelu-Walter
et al. 2008; Niskanen et al. 2004) have been reported. In
Stone pine, significant differences for extrusion, and nearly
significant differences for initiation, were found among the
families. Some of the families performed better than others,
which suggested an additive genetic control on the initia-
tion of SE in Stone pine, as was reported in other conifers
(MacKay et al. 2006; Park et al. 1993). Differences in SE
initiation among mother trees were recorded in this study:
family 58 almost doubled the overall mean value, whereas
family 70 was five times below the mean value with regard
to frequency of initiation. These data are similar to those
obtained in a broader experiment with 20 half-sib families
of P. pinaster, in which the best family produced three
times the overall mean and the worst was six times below
the average (Miguel et al. 2004). Although frequencies of
initiation in Stone pine were very low, the fact that
embryogenic tissue was obtained from all the five families
tested is encouraging.
Although the number of families tested in this study was
relatively low, the difference in their ability to achieve
initiation showed that proper selection of mother trees
could be used to increase the number of captured geno-
types. This study did not detect any significant effect of
interaction between families and collection dates, as
reported in P. strobus (Klimaszewska et al. 2001). How-
ever, as was observed in that species, different temporal
patterns for initiation were recorded for the different fam-
ilies in the present study, likely reflecting developmental
disparities, as has been reported for other species (Lelu
et al. 1999), but also showing a family with a wider range
of responses than others. It is interesting that this particular
family also performed equally in both crop years. The
search for and selection of these ‘‘broad spectrum geno-
types’’ for initiation ability as maternal trees for controlled
crosses could be useful for increasing the number of cap-
tured genotypes in a breeding program. Selection of the
most favorable maternal parent has been proposed for
improving SE initiation (MacKay et al. 2006; Niskanen
et al. 2004).
The proliferation of EM in Stone pine was achieved by
continuous formation of proembryogenic and early
embryonic structures, which assumed either a smooth or
spiky morphotype, similar to that of another pine species
(Ramarosandratana et al. 2001a). The development of these
morphotypes seemed to be genotype dependent, although
sometimes changes within a given genotype in an uncon-
trolled manner were observed. Growth of EM was more
favorable with subculturing after dispersion on filter disks
than when using tissue clumps. The culture technique
based on dispersing embryogenic tissue by suspending it in
liquid medium and collecting it on filter paper disks, which
was originally developed to rescue slow-growing EM of
eastern white pine (Klimaszewska and Smith 1997), was
particularly useful for proliferating Stone pine embryo-
genic tissue. It grew faster than when it was cultured as
small tissue clumps, multiplying the initial ± 100 mg FW
by 52–96 times, depending on genotype, after 6-week
period. As this effect was marked in the case of the slower-
growing genotypes, in which growth was up to more 12
times higher when using the dispersal instead of the clump
procedure, the culture of initial extrusions following this
procedure will likely increase the number of established
lines (Lelu-Walter et al. 2008).
The main objective of the maturation experiments in this
study was to demonstrate that differentiation and mature
embryos could be obtained from established embryogenic
lines of Stone pine. In the present study, preculture on
medium with reduced nutrients improved the number of
mature embryos. Vigorous proliferation of embryogenic
tissue hampered differentiation and maturation of somatic
embryos, as in other conifers (Breton et al. 2006). There-
fore, slower growth caused by the reduction of nutrients in
the preconditioning medium might be behind the improved
results obtained in this study. In addition, coating
embryogenic tissue with AC clearly improved maturation,
as was also shown in P. pinaster (Lelu-Walter et al. 2006),
probably by adsorbing endogenously produced substances,
because it was more effective when the tissue was coated
than when it was added to the culture medium. Mature
embryos were obtained in five out of seven tested
embryogenic lines, which is around 70% of the tested
genotypes. Although the number of somatic embryos
obtained in this first study was very low, the percentage of
responsive genotypes is comparable, or even higher, than
in other pine species (Miguel et al. 2004). However,
research that involves aspects such as the medium’s water
status (Klimaszewska and Smith 1997; Ramarosandratana
et al. 2001b), carbon sources and their interaction with
gelling agents (Ramarosandratana et al. 2001a), anti-auxins
(Find et al. 2002), ethylene modulators (Kong and Yeung
1994), or other factors, may improve the efficiency of SE in
P. pinea.
Although the number of mature embryos obtained from the
maturation experiments described in this study was too low to
Plant Cell Tiss Organ Cult
123

draw any conclusion about their germination and conversion
abilities, the high percentages of germination (70%) and
conversion (30%) that were obtained are encouraging.
Acknowledgments The authors gratefully thank N. Cleto and Y.
Vinuesa for their technical assistance. Funds were provided by pro-
jects AGL2002-00867 and AGL2005-07585, and IMIDRA and INIA
grants to E. Carneros. We wish to thank the National Forest Breeding
Centre ‘‘Puerta de Hierro’’ (Madrid) of the Spanish Ministry of
Environment, and Dr. Mutke for all their help in collecting plant
material. We thank the Canadian Forest Service for hosting E.
Carneros at its laboratory in Fredericton.
References
Becwar MR, Nagmani R, Wann SR (1990) Initiation of embryogenic
cultures and somatic embryo development in loblolly pine
(Pinus taeda). Can J Res 20:810–817. doi:10.1139/x90-107
Bonga JM (2004) The effect of various culture media on the formation
of embryo-like structures in cultures derived from explants taken
from mature Larix decidua. Plant Cell Tissue Organ Cult 77:43–
48. doi:10.1023/B:TICU.0000016488.79965.b7
Breton D, Harvengt L, Trontin J-F et al (2006) Long-term subculture
randomly affects morphology and subsequent maturation of
early somatic embryos in maritime pine. Plant Cell Tissue Organ
Cult 87:95–108. doi:10.1007/s11240-006-9144-9
Capuana M, Giannini R (1995) In vitro plantlet regeneration from
embryonic explants of Pinus pinea L. In Vitro Cell Dev Biol
Plant 31:202–206. doi:10.1007/BF02632022
Cheliak WM, Klimaszewska K (1991) Genetic variation in somatic
embryogenic response in open-pollinated families of black spruce.
Theor Appl Genet 82:185–190. doi:10.1007/BF00226211
Deb CR, Tandon P (2002) Somatic embryogenesis and plantlet
regeneration from mature zygotic embryos of Pinus kesiya (Royle
ex. Gord.). J Plant Biol 29:301–306
Diamantoglou S, Panagopoulos I, Munoz-Ferriz A et al (1990) In vitro
studies of embryo growth, callus formation and multiple bud
induction of Pinus pinea L. J Plant Physiol 137:58–63
Duchefa Biochemie BV (2003) Biochemicals, Plant cell and tissue
culture, Plant molecular biochemicals. Catalogue 2003–2005.
Haarlem, The Netherlands
El-Kassaby Y (2004) Feasibility and proposed outline of a global
review of forest biotechnology. Forest Genetic Resources Work-
ing Paper FGR/77E: Forest Resources Development Service,
Forest Resources Division. FAO, Rome
Find J, Grace L, Krogstrup P (2002) Effect of anti-auxins on maturation
of embryogenic tissue cultures of Nordmanns fir (Abies nordman-niana). Physiol Plant 116:231–237. doi:10.1034/j.1399-3054.
2002.1160213.x
Finer JJ, Kriebel HB, Becwar MR (1989) Initiation of embryogenic
callus and suspension cultures of eastern white pine (Pinusstrobus L). Plant Cell Rep 8:203–206. doi:10.1007/BF00778532
Garcıa-Ferriz L, Serrano L, Pardos JA (1994) In vitro shoot organo-
genesis from excised immature cotyledons and microcuttings
production in Stone Pine. Plant Cell Tissue Organ Cult 36:135–
140. doi:10.1007/BF00048324
Garin E, Isabel N, Plourde A (1998) Screening of large numbers of
seed families of Pinus strobus L. for somatic embryogenesis
from immature and mature zygotic embryos. Plant Cell Rep 18:
37–43. doi:10.1007/s002990050528
Gonzalez MV, Rey M, Tavazza R et al (1998) In vitro adventitious
shoot formation on cotyledons of Pinus pinea. HortScience 33:
749–750
Gupta PK, Grob JA (1995) Somatic embryogenesis in conifers. In:
Jain S, Gupta PK, Newton RJ (eds) Somatic embryogenesis in
woody plants, vol 1. Kluwer Academic, Dordrecht, pp 81–98
Klimaszewska K, Cyr DR (2002) Conifer somatic embryogenesis: I.
Development. Dendrobiology 48:31–39
Klimaszewska K, Smith D (1997) Maturation of somatic embryos of
Pinus strobus is promoted by a high concentration of gellan gum.
Physiol Plant 100:949–957. doi:10.1111/j.1399-3054.1997.tb00
022.x
Klimaszewska K, Park Y-S, Overton C et al (2001) Optimized
somatic embryogenesis in Pinus strobus L. In Vitro Cell Dev
Biol Plant 37:392–399. doi:10.1007/s11627-001-0069-z
Kong L, Yeung EC (1994) Effects of ethylene and ethylene inhibitors
on white spruce somatic embryo maturation. Plant Sci 104:71–80.
doi:10.1016/0168-9452(94)90192-9
Lelu MA, Bastien C, Drugeault A et al (1999) Somatic embryogen-
esis and plantlet development in Pinus sylvestris and Pinuspinaster on medium with and without growth regulators. Physiol
Plant 105:719–728. doi:10.1034/j.1399-3054.1999.105417.x
Lelu-Walter MA, Bernier-Cardou M, Klimaszewska K (2006)
Simplified and improved somatic embryogenesis for clonal
propagation of Pinus pinaster (Ait). Plant Cell Rep 25:767–776.
doi:10.1007/s00299-006-0115-8
Lelu-Walter MA, Bernier-Cardou M, Klimaszewska K (2008) Clonal
plant production from self- and cross-pollinated seed families of
Pinus sylvestris (L.) through somatic embryogenesis. Plant Cell
Tissue Organ Cult 92:31–45. doi:10.1007/s11240-007-9300-x
Li X, Huang F, Gbur E (1998) Effect of basal medium, growth
regulators and phytagel concentration of embryogenic cultures
from immature zygotic embryos of loblolly pine (Pinus taeda L.).
Plant Cell Rep 17:298–301. doi:10.1007/s002990050396
Litvay JD, Verma DC, Johnson MA (1985) Influence of loblolly pine
(Pinus taeda L.). Culture medium and its components on growth
and somatic embryogenesis of the wild carrot (Daucus carotaL.). Plant Cell Rep 4:325–328. doi:10.1007/BF00269890
MacKay JJ, Becwar MR, Park Y-S et al (2006) Genetic control of
somatic embryogenesis initiation in loblolly pine and implications
for breeding. Tree Genet Genomes 2:1–9. doi:10.1007/s11295-
005-0020-2
Merkle SA, Nairn CJ (2005) Hardwood tree biotechnology. In Vitro
Cell Dev Biol Plant 41:602–619. doi:10.1079/IVP2005687
Miguel C, Goncalves S, Tereso S et al (2004) Somatic embryogenesis from
20 open-pollinated families of Portuguese plus trees of maritime pine.
Plant Cell Tissue Organ Cult 76:121–130. doi:10.1023/B:TICU.
0000007253.91771.e3
Mutke S, Gordo J, Gil L (2000) The stone pine (Pinus pinea L.)
breeding programme in Castile-Leon (Central Spain). FAO-
CIHEAM NUCIS-Newsletter 9:50–55
Nehra NS, Becwar MR, Rottman WH et al (2005) Forest biotech-
nology: Innovative methods, emerging opportunities. In Vitro
Cell Dev Biol Plant 41:701–717. doi:10.1079/IVP2005691
Niskanen AM, Lu J, Seitz S et al (2004) Effect of parent genotype on
somatic embryogenesis in Scots pine (Pinus sylvestris). Tree
Physiol 24:1259–1265
Oliveira P, Barriga J, Cavaleiro C et al (2003) Sustained in vitro root
development obtained in Pinus pinea L inoculated with ectomy-
corrhizal fungi. Forestry 76:579–587. doi:10.1093/forestry/76.5.
579
Park Y-S (2002) Implementation of conifer somatic embryogenesis in
clonal forestry: technical requirements and deployment consid-
erations. Ann Sci 59:651–656. doi:10.1051/forest:2002051
Park Y-S, Pond SE, Bonga JM (1993) Initiation of somatic
embryogenesis in white spruce (Picea glauca): genetic control,
culture treatment effects, and implications for tree breeding.
Theor Appl Genet 86:427–436. doi:10.1007/BF00838557
Plant Cell Tiss Organ Cult
123

Park Y-S, Pond SE, Bonga JM (1994) Somatic embryogenesis in
white spruce (Picea glauca): genetic control in somatic embryos
exposed to storage, maturation treatments, germination, and
cryopreservation. Theor Appl Genet 89:742–750. doi:10.1007/
BF00223714
Park Y-S, Lelu-Walter MA, Harvengt L et al (2006) Initiation of
somatic embryogenesis in Pinus banksiana, P. strobus, P.
pinaster and P. sylvestris at three laboratories in Canada and
France. Plant Cell Tissue Organ Cult 86:87–101. doi:10.1007/
s11240-006-9101-7
Percy RE, Klimaszewska K, Cyr DR (2000) Evaluation of somatic
embryogenesis for clonal propagation of western white pine. Can
J Res 30:1867–1876. doi:10.1139/cjfr-30-12-1867
Pullman GS, Johnson S (2002) Somatic embryogenesis in loblolly
pine (Pinus taeda L.): improving culture initiation rates. Ann Sci
59:663–668. doi:10.1051/forest:2002053
Radojevic L, Alvarez C, Fraga MF et al (1999) Somatic embryogenic
tissue establishment from mature Pinus nigra Arn. Ssp.
Salzmannii embryos. In Vitro Cell Dev Biol Plant 35:206–209.
doi:10.1007/s11627-999-0078-x
Ramarosandratana A, Harvengt L, Bouvet A et al (2001a) Influence
of the embryonal-suspensor mass (ESM) sampling on develop-
ment and proliferation of maritime pine somatic embryos. Plant
Sci 160:473–479. doi:10.1016/S0168-9452(00)00410-6
Ramarosandratana A, Harvengt L, Bouvet A et al (2001b) Effects
of carbohydrate source, polyethylene glycol and gellan gum
concentration on embryonal-suspensor mass (ESM) proliferation
and maturation of maritime pine somatic embryos. In Vitro Cell Dev
Biol Plant 37:29–34. doi:10.1007/s11627-001-0006-1
Salajova T, Salaj J (2005) Somatic embryogenesis in Pinus nigra:
embryogenic tissue initiation, maturation and regeneration ability
of established cell lines. Biol Plant 49:333–339. doi:10.1007/
s10535-005-0003-z
Tang W, Guo Z, Ouyang F (2001) Plant regeneration from
embryogenic cultures initiated from mature loblolly pine zygotic
embryos. In Vitro Cell Dev Biol Plant 37:558–563. doi:10.1007/
s11627-001-0097-8
Tautorus TE, Fowke L, Dunstan DI (1991) Somatic embryogenesis in
conifers. Can J Bot 69:1873–1899. doi:10.1139/b91-237
Valdes AE, Ordas RJ, Fernandez B et al (2001) Relationships
between hormonal contents and the organogenic response in
Pinus pinea cotyledons. Plant Physiol Biochem 39:377–384. doi:
10.1016/S0981-9428(01)01253-0
von Arnold S, Sabala I, Bozhkov P et al (2002) Developmental
pathways of somatic embryogenesis. Plant Cell Tissue Organ Cult
69:233–249. doi:10.1023/A:1015673200621
Zoglauer K, Behrendt U, Rahmat A (2003) Somatic embryogenesis—
the gate to biotechnology in conifers. In: Laimer M, Rucker W et al
(eds) Plant tissue culture—100 years since Gottlieb Haberlandt.
Springer, Heidelberg, pp 175–202
Plant Cell Tiss Organ Cult
123





























